

Abstract
Transgenic plant-derived vaccines comprise a new type of bioreactor that combines plant genetic engineering technology with an organism's immunological response. This combination can be considered as a bioreactor that is produced by introducing foreign genes into plants that elicit special immunogenicity when introduced into animals or human beings. In comparison with traditional vaccines, plant vaccines have some significant advantages, such as low cost, greater safety, and greater effectiveness. In a number of recent studies, antigen-specific proteins have been successfully expressed in various plant tissues and have even been tested in animals and human beings. Therefore, edible vaccines of transgenic plants have a bright future. This review begins with a discussion of the immune mechanism and expression systems for transgenic plant vaccines. Then, current advances in different transgenic plant vaccines will be analyzed, including vaccines against pathogenic viruses, bacteria, and eukaryotic parasites. In view of the low expression levels for antigens in plants, high-level expression strategies of foreign protein in transgenic plants are recommended. Finally, the existing safety problems in transgenic plant vaccines were put forward will be discussed along with a number of appropriate solutions that will hopefully lead to future clinical application of edible plant vaccines.
Keywords: Transgenic, Plant, Vaccine, Safety
Introduction
Infectious diseases have become grave threats to human and animal health. However, the use of existing vaccines is an effective way of preventing the occurrence of these diseases. With the development of biotechnology, the type of vaccines has developed from the bacterial vaccine to subunit vaccine or DNA vaccine, but these products are difficult to market because of high production costs. Transgenic plant-based vaccines have some advantages over traditional injected vaccines, such as a wide-range possible expressed epitopes, large biomass of cultivation, low cost, and ease of transportation and preservation. Above all, vaccination can be directly carried out orally, rather than parenterally, and often without extraction and purification. The existence of the plant cell wall obviously has a sustained-release effect for the delivery of the antigen. Therefore, in recent years, the research and application of genetically modified (GM) plant vaccines has been a hot topic in vaccine research. This paper reviews current advances in different transgenic plant vaccines, such as vaccines against pathogenic viruses, bacteria, and eukaryotic parasites, as well as the existing safety problems in products obtained from transgenic plant vaccines.
Immune response to transgenic plant vaccines
Research on genetically modified plant vaccines has progressed for over 20 years. Curtiss and Cardineau 1990 first successfully expressed Streptococcus surface protein A (SPaA) gene in tobacco seeds, and the SPaA expressed in transgenic tobacco plants was shown to produce an immune reaction. Afterwards, Mason et al. (1992) produced Hepatitis B vaccine in transgenic tobacco, which demonstrated that proteins expressed in transgenic plants cannot only maintain their natural conformation, but they also retain their antigenic determinants by stimulating the immune response of B and T cells. Thus, the concept of an edible vaccine was presented for the first time. The idea of edible vaccine obtained from plants has received a great deal of attention from both academia and private enterprise. Kapusta et al. (1999) expressed HBsAg in transgenic lettuce and observed a preliminary immune effect via oral administration given to volunteers, which showed that edible vaccines from transgenic plants could induce potent specific immune responses in human beings. It has been demonstrated on the one hand that transgenic plant vaccine administered by an oral route could induce the body to produce both humoral immune and cellular immune responses, which ultimately can give rise to immunity (Rigano et al. 2006; Li and Xi 2004). On the other hand, both the stomach and intestinal tract have an independent mucosal immune system, which we consider in the following in a description of the mechanisms of activation of the immune system when plant virus vaccines are administered orally.
After human or animals eat transgenic plants (such as potatos or tomatoes) expressing the oral vaccine, the released antigens are identified and swallowed by ruffled cells (M cells) in intestinal mucosa associated with lymphoid tissue (such as Peyer's patch), and then are transferred to lower follicular tissue. After that, its antigen-presenting cells (APC cells) produce the antigen on the surface of APC cell and subsequently activate B lymphocytes with the help of Th cells. The activated B lymphocytes, which participate in mucosal immunity, are transferred to the mesentery lymph node and finally develop into plasma cells. The secreted IgA in plasma cells reaches membrane cavities through the epithelium and combines with membrane secretion to form secretory IgA (SIgA). Once they have been transferred to the membrane cavity, the SIgA will combine with specific pathogens (such as pathogenic microorganism toxin), and thereby produce certain immune protection. Subsequently, B lymphocytes activated in the mucosa, secrete serum type-specific antibodies into the blood, and generate certain immune protection. The cytokines in Th cells of the mucosal immune system also can activate cytotoxic T lymphocyte (CTL) and elicit a strong cellular immune response. Thus, transgenic plant vaccines can induce mucosal immunity and also cause humoral and cellular immunity. Studies have shown that mucosal immunity can produce immune protection against both gastrointestinal tract and non-gastrointestinal tract infections (Pearay et al. 2001; Zhu and Zhang 2006).
Plant expression system in plant-based vaccines
Exogenous target genes can be stably expressed and correctly assembled in plant cells via plant expression systems, which is key to successful transgenic plant vaccine production. Different methods of obtaining transgenic plant vaccines are presented in Fig 1. The current plant expression system has the following three principal aspects, including genome expression, chloroplast expression, and plant viral expression system.
Fig. 1.
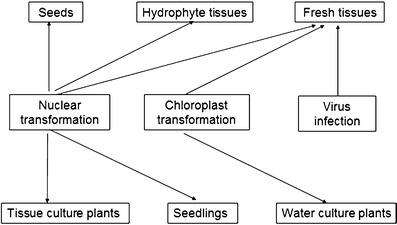
Different methods of acquiring transgenic plants vaccine
Plant cell genome expression system
The basic characteristic of the plant expression system is that it is easy to operate. Recombinant protein products expressed in plant seeds or tubers are easy to preserve and are especially well-suited for the production of polyvalent vaccines and poly-recombinant antibodies. Guerrero-Andrade et al. (2006) transformed the Newcastle disease virus fusion (F) protein gene into the maize, driven by the maize ubiquitin promoter. Fed with kernels containing the F protein, antibody production was induced in chickens. An exogenous target gene was integrated into the plant genome and transgenic seeds could be bred by traditional breeding technology, which means that they are easily stored, and ready for replanting. Plants are sufficiently versatile that multiple foreign genes may be stored in the same plant through the hybridization of different transgenic plants.
Chloroplast expression system
By gene gun bombardment, foreign genes are introduced to the plant chloroplasts and are likely to be integrated into chloroplast chromosomes. Although the chloroplast transformation system has only been applied to certain plants, the ability to introduce multiple copies of a gene would improve foreign protein expression in plants (Daniell et al. 2002). Moreover, the rigorous maternal inheritance pattern (namely a hereditary phenomenon controlled by the chromosomes outside the nucleus) of chloroplast genes may avoid genetic contamination during plant pollination and eliminate gene silencing in the process of transformation.
Plant viral expression system
This expression system is based on infection of a plant by a plant virus, which is competent to independently replicate, transcribe, and translate so as to produce many copies of a recombinant protein introduced into the plant viral genome. The advantages of this expression system lie in rapid research methods, higher yields, and smaller demands on the planting area. By improving plant virus expression systems, exogenous genes cannot only be expressed as soluble protein and distributed in the cytoplasm of the host plant cell, but they also can fuse with plant virus capsid protein genes and constantly be expressed when virus particles replicate and translate viral proteins in the infected state. The native cottontail rabbit papillomavirus (CRPV) L1 capsid protein gene was expressed transgenically via Agrobacterium tumefaciens transformation and transiently via a tobacco mosaic virus (TMV) vector in Nicotiana spp. L1 protein was detected in concentrated plant extracts at concentrations up to 0.4 mg/kg in TMV-infected plants (Kohl et al. 2006). A recombinant plant virus (CPMV-PARVO1) was inactivated by UV treatment to remove the possibility of the recombinant plant virus replication in a plant host after the manufacture of the VP2 capsid protein vaccine of canine parvovirus. The result showed that the inactivated canine parvovirus (CPV) was able to protect dogs from a lethal challenge with CPV, and all of the immunized dogs elicited high titers of peptide-specific antibody, which neutralized CPV in vitro (Langeveld et al. 2001). Yang et al. (2007) successfully expressed foot-and-mouth disease VP1 protein by Bamboo Mosaic virus carrier. In addition, other plant viruses are used for expression vectors, such as Tomato bushy stunt virus (TBSV) and Alfalfa mosaic virus (ALMV) (Yang et al. 2007).
Recent advances in transgenic plant vaccines
Over the past 10 years, since heavy-chain and light-chain immunoglobulins were successfully expressed in tobacco, and both of them were then assembled into a functioning antibody, plants have gradually become viable expression systems for the production of various types of antibodies. The number of articles published every year on transgenic plant vaccines in various databases (such as ScienceDirect, Springerlink, Wiley InterScience and Web of science) is shown in Fig 2. There were 835 articles in Elsevier, 841 in Springerlink, 808 in Wiley, and 313 in Web Of Science in 2011. Figure 2 shows that from 2000 to 2011, an increasing number of vaccines have been successfully expressed in various transgenic plants. At present, there are dozens of plants used for animal and human vaccines. The main immunogenic genes include: HBsAg gene, Escherichia coli heat-sensitive bowel toxin B subunit (LT-B) gene, rabies virus glycoprotein (G protein) gene, foot-and-mouth disease VP1 gene, rotavirus gene, and so on.
Fig. 2.
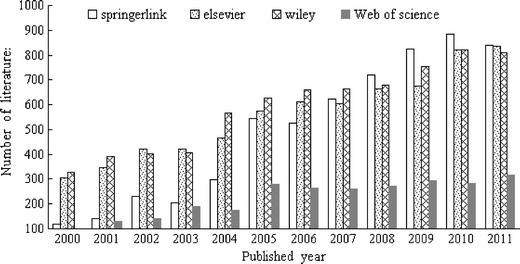
Number of published articles on transgenic plant vaccines in recent decades
Recent research has concentrated on the application of transgenic plant vaccines in humans and animals. Reported transgenic plant vaccines can be divided into the following four types: against bacteria, against viruses, against parasites, and immunocontraceptive vaccines.
Bacterial vaccine
The bacterial subunit vaccine products with great potential are the heat labile toxin (LT) of enterotoxigenic E. coli B subunit vaccine (LT-B) and the cholera toxin (CT) of Vibrio cholerae B subunit vaccine (CT-B). Recent applications of the two kinds of vaccines are shown in Table 1.
Table 1.
Advances of bacterial vaccine expressed in transgenic plants
| Pathogen | Antigen | Production system | Expression | Efficacy | References |
|---|---|---|---|---|---|
| Heat-labile toxin | LT-Ba | Tobacco leaf; potato | 0.001 %TSP/0.0002 % FW | Immunogenic by oral delivery to mice | (Haq et al. 1995) |
| Heat-labile toxin | LT-B | Tobacco leaf | 0.095 % of TSP | None tested | Wang et al. (2001) |
| Heat-labile toxin | LT-B | Potato tuber | 0.2 % TSP/0.001 % FW | Immunogenic and protective by oral delivery to mice | Mason et al. (1998) |
| Heat-labile toxin | LT-B | Potato tuber | None tested | Immunogenic and protective by oral delivery to humans | Tacket et al. (1998) |
| Heat-labile toxin | LT-B | Potato tuber | None tested | Immunogenic and protective by oral delivery to nice | Lauterslager et al. (2001) |
| Heat-labile toxin | LT-B | Maize seed | 1.8%TSP | None tested | Streatfield et al. (2001); Streatfield et al. (2002), (2003) |
| Heat-labile toxin | LT-B | Maize seed | 0.07 ~ 3.7 % TSP | Immunogenic and protective by oral delivery to mice | Chikwamba et al. (2002a, b); |
| Heat-labile toxin | LT-B | Maize seed | None tested | Immunogenic and protective by oral delivery to mice and swine | Lamphear et al. (2002) |
| Heat-labile toxin | LT-B | Chloroplasts of tobacco | 2.5 % TSP | None tested | Kang et al. (2003) |
| Heat-labile toxin | LT-B | Tobacco mosaic virus-infected Nicotiana benthamiana plants | 75 μg/g FW | Immunogenic by oral delivery to mice | Wagner et al. (2004) |
| Heat-labile toxin | LT-B | Transgenic corn | None tested | LT-B expressed in transgenic corn was well tolerated and immunogenic in humans. | Tacket et al. (2004) |
| Heat-labile toxin | LT-B | Tobacco | 3.3 % of TSP | None tested | Kang et al. (2005) |
| Heat-labile toxin | LT-B | Corn seeds | None tested | Systemic and mucosal antibody responses to LT-B in young and aged mice, and recall responses to oral administration and injection of LT-B in aged mice. | Karaman et al. (2006) |
| Heat-labile toxin | LT-B | Siberian ginseng | 0.36 % of TSP | None tested | Kang et al. (Kang et al. 2006a, b, c) |
| Heat-labile toxin | LT-B | Carrot | 0.3 % of TSP | None tested | Rosales-Mendoza et al. (2007a) |
| Heat-labile toxin | LT-B | Soybean seed | 2.4 % of the total seed protein | Induced mice both systemic IgG and IgA, and mucosal IgA antibody responses | Tomas et al. (2007) |
| Heat-labile toxin | LT-B | Lettuce leaf | 1.0–2.0 % of TSP | None tested; | Kim et al.(2007) |
| Heat-labile toxin | LT-B | Lettuce leaf | 0.05 % of TSP | Balb/c mice elicited specific and significant antibody responses in both serum and intestinal tissues | Martı´nez-Gonza´lez et al. (2011) |
| Heat-labile toxin; Chlamydophila psittaci | A DNA fragment encoding the MOMP gene of Chlamydophila psittaci was fused to LT-B gene (LTB-MOMP) | Rice leaves | 0.0033e0.0054 % of TSP | None tested | Zhang et al., (2008) |
| Heat-labile toxin | LT-B | Carrot | None tested | Balb/c mice elicited significant serum and intestinal antibody responses | Rosales-Mendoza et al. (2008) |
| Heat-labile toxin | The Escherichia coli heat-labile enterotoxin B subunit | Watercress | 1.3 % of TSP | None tested | Loc et al.(2011a) |
| Heat-labile toxin | Escherichia coli heat-labile enterotoxin B subunit | Peperomia pellucida | 0.75 % of TSP | None tested | Loc et al. (2010) |
| NDV Newcastle disease virus | Synthetic LT-B and Hemagglutinin–Neuraminidase-Neutralizing Epitope Fusion Protein | Tobacco chloroplasts | 0.5 % of TSP | None tested | Sim et al. (2009) |
| Heat-labile toxin | An Escherichia coli antigenic fusion protein comprising LT-B and the heat stable toxin | Tobacco plants | 2.3 % of TSP | Oral immunization of mice led to the induction of both serum and mucosal LTB-ST specific antibodies. | Rosales-Mendoza et al. (2007b) |
| Enteropathogenic Escherichia coli (EPEC) | Bundle-forming pilus structural subunit A | Tobacco leaf | 8 % TSP/1 % FW | Immunogenic by oral delivery to ice | da Silva et al. (2002) |
| Cholera toxin | CT-Bb | Potato tuber | 0.3 % TSP/0.002 % FW | Immunogenic and protective by oral delivery to mice | Arakawa et al. (1997) |
| Arakawa et al. (1998) | |||||
| Cholera toxin | CT-B | Tobacco leaf | 4 % TSP/0.5 % FW | No published data | Daniell et al. (2001) |
| Cholera toxin | CT-B | Tomato leaf and fruit | 0.04 % TSP/0.005 % FW | No published data | Jani et al. (2002) |
| Cholera toxin; rotavirus enterotoxin | CT-B fused to rotavirus enterotoxin NSP4 | Potato tuber | 0.0003 % FW | Immunogenic and protective (passive immunity) by oral delivery to mice | Yu and Langridge, (2001) |
| Cholera toxin | CT-B | Tomato fruit | 0.081 % of TSP | Gavage of mice induced both serum and mucosal CTB specific antibodies. | Jiang et al. (2007) |
| Cholera toxin | CT-B | Tomato fruit | 0.9 % of TSP | No published data | Loc et al. (2011b) |
| Cholera toxin | CT-B fused to an endoplasmic reticulum retention signal (SEKDEL) | TCarrot (Daucus carota L.) | 0.48 % of TSP | None tested | Kim et al. (2009) |
| Cholera toxin; porcine epidemic diarrhea virus | A synthetic CT-B was fused with a synthetic neutralizing epitope gene of the porcine epidemic diarrhea virus (sCTB–sCOE), | Lettuce (Lactuca sativa L.) | 0.0065 % of TSP | Induce efficient immune responses against porcine epidemic diarrhea virus infection by administration | Huy et al. (2011) |
| Cholera toxin | A cDNA encoding the simian–human immunodeWciency virus (SHIV 89.6p) Tat regulatory element protein was fused to the cterminus of CT-B (ctxB-tat) | Potato leaf | 0.005–0.007 % of TSP or 4.6 mg per 100 g potato tuber tissue | None tested | Kim et al. (2004a) |
| Cholera toxin; anthrax | A DNA encoding the 27-kDa domain I of anthrax lethal factor protein (LF), was linked to the carboxyl terminus of CT-B (CTB–LF). | Potato leaf | 0.002 % of TSP | None tested | Kim et al. (2004b) |
| Cholera toxin; simian immunodeficiency virus | A deoxyribonucleic acid (DNA) fragment encoding CT-B was linked 5' to the simian immunodeficiency virus (SIVmac) Gag p27 capsid gene (CTB–Gag) | Potato leaf | 0.016–0.022 % of TSP | None tested | Kim et al. (2004c) |
| Cholera toxin | CT-B | Tobacco plants | 1.5 % of TSP | None tested | Kang et al. (2004) |
| Cholera toxin; simian rotavirus | A gene encoding VP7 of simian rotavirus SA11 was fused to the carboxyl terminus of CT-B | Potato leaf | 0.01 % of the total soluble tuber protein | None tested | Choi et al. (2005) |
| Cholera toxin | CT-B | Lettuce plants | 0.24 % of TSP | None tested | Kim et al. (2006) |
| Cholera toxin | A synthetic CT-B | Tobacco plant | 1.8 % of TSP | None tested | Kang et al. (2006) |
| Cholera toxin | CT-B | Maize | 0.0197 % of TSP | Anti-CTB IgG and anti-CTB IgA were detected in the sera and fecal samples of the orally immunized mice | Karaman et al. (2012) |
| Cholera toxin | Fusion gene of CT-B and the B chain of human insulin | Tobacco plants | 0.11 % of TSP | None tested | Li et al. (2006a) |
| Cholera toxin | Fusing CT-B with allergen-specific T-cell epitopes | Rice seed | None tested | Mice suppressed allergen-specific IgE responses and pollen-induced clinical symptoms | Hidenori et al. (2008) |
| Cholera toxin; Vibrio cholerae | Accessory colonization factor subunit A (ACFA) of Vibrio cholerae and ACFA fused to CT-B | Tomato | 0.25 % and 0.08 % of TSP | None tested | Sharma et al. (2008a) |
| Cholera toxin | Synthetic CT-B gene | Rice endosperm | 2.1 % of TSP | None tested | Oszvald et al. (2008) |
| Cholera toxin; Vibrio cholerae | Toxin co-regulated pilus subunit A (TCPA) of Vibrio cholerae and its immunogenic epitopes fused to CT-B | Tomato | 0.17 and 0.096 % of TSP | None tested | Sharma et al. (2008b) |
| Cholera toxin; dengue virus | Cholera toxin B subunit-domain III of dengue virus envelope glycoprotein E fusion protein | Tobacco plant | 0.019 % of TSP | None tested | Kim et al. (2010) |
| Enterotoxin ricin; insulin | A fusion protein (INS–RTB)( the non-toxic B chain of the plant AB enterotoxin ricin (RTB) and linked to the coding region of the proinsulin gene (INS) | Potato plants | None tested | None tested | Carter et al. (2010) |
| Cholera toxin; Rabies | Rabies glycoprotein fused with CT-B | Tobacco plants | 0.4 % of TSP | None tested | Roy et al. (2010) |
aLT-B—heat labile toxin (LT) of enterotoxigenic E. coli (ETEC) B subunit
bCT-B—Cholera toxin B subunit
Enterotoxigenic strains of E. coli may produce a heat-labile holotoxin (LT), which will cause diarrhea. Karaman et al. (2006) engineered corn seeds to produce LT-B, the nontoxic subunit of LT, to serve as a plant-derived vaccine to traveler's diarrhea and as an adjuvant for co-administered proteins. Specific IgA and IgG antibodies were detectable in mice fed with transgenic corn. Tae-Jin Kang et al. (2003) reported a feasibility study for producing the nontoxic LT-B via chloroplast transformation of tobacco. The amount of LT-B protein detected in transplastomic tobacco leaf was approximately 2.5 % of the total soluble plant protein, approximately 250-fold higher than in plants generated via nuclear transformation.
Cholera is a highly epidemic diarrheal disease that continues to devastate many developing countries with poor socio-economic conditions, where the sanitation and public hygiene systems are rudimentary (Kaper et al. 1995). Cholera is caused by V. cholerae and disease severity is mediated by the potent action of the cholera toxin (CT), which stimulates the secretion of water and electrolytes into the intestine (Field et al. 1989). The CT has been reported as representative of adjuvants that can induce mucosal immunity efficiently (Elson 1989; McGhee et al. 1992). The generation of non-toxic CT derivatives that retain adjuvant activity could provide a safe alternative for the evaluation of these toxins as mucosal adjuvants in humans and animals. CT is composed of distinct A and B subunits. The pentameric B subunit (CT-B) contains five identical polypeptides, shows good immunogenicity, targets the glycosphingolipid receptors on eukaryotic cell surfaces, so consequently CT-B subunit is a well-characterized antigen against cholera. Jiang et al. (2007) developed transgenic tomato-expressing CT-B protein, especially in the ripening tomato fruit under the control of the tomato fruit-specific E8 promoter. Gavage of ripe transgenic tomato fruits induced both serum and mucosal CT-B-specific antibodies in mice. Kim et al. (2009) fused CT-B to an endoplasmic reticulum (ER) retention signal (SEKDEL) and expressed the protein in carrot roots. The produced sCT-B in transgenic carrot roots demonstrated strong affinity for GM1-ganglioside, suggesting that the sCT-B conserved the antigenic sites for binding and proper folding of the pentameric sCT-B structure. The expression level of sCT-B comprised approximately 0.48 % of total soluble protein (TSP) in root of transgenic carrot. A synthetic CTB was fused with a synthetic neutralizing epitope gene of the porcine epidemic diarrhea virus (sCTB–sCOE), and the sCTB–sCOE fusion gene was introduced into a plant expression vector under the control of the ubiquitin promoter. This plant expression vector was transformed into lettuce (Lactuca sativa L.) by the Agrobacterium-mediated transformation. The expression level of CTB–COE fusion proteins reached 0.0065 % of the total soluble protein in transgenic lettuce leaf tissues (Huy et al. 2011).
Viral vaccine
The most common applications of viral antigens in transgenic plants and current advances of research on the various viral vaccines are summarized in Table 2.
Table 2.
Advances of viral vaccine expressed in transgenic plants
| Disease | Pathogen | Antigen (protein or peptide types expressed) | Production system | Expression | Efficacy | Number of references |
|---|---|---|---|---|---|---|
| Food and mouth disease | FMDVa | Structural protein VP1 | Arabidopsis thaliana | None tested | Mice elicited specific antibody responses to synthetic peptides representing amino acid residues 135 to 160 of VP1 | Carrillo et al. (1998) |
| Food and mouth disease | FMDV | Structural protein VP1 | Alfalfa plants | None tested | Mice elicited a specific antibody response to a synthetic peptide representing amino acid residues 135–160 of VP1 | Wigdorovitz (1999) |
| Food and mouth disease | FMDV | Structural protein VP1 | Potatoes plants | None tested | The male BALB/c mice presented a FMDV VP1 specific antibody response and protection against the experimental challenge | Carrillo et al. (2001) |
| Food and mouth disease | FMDV | A highly immunogenic epitope from FMDV fused to glucuronidase (gus A) reporter gene | Alfalfa plants | None tested | Mice elicited a strong anti-FMDV antibody response against a synthetic peptide representing the region VP135–160 | Dus Santos et al. (2002) |
| Food and mouth disease | FMDV | Dominant epitopes of FMDV serotype O | Tobacco plant | None tested | Most guinea pigs were protected against FMDV challenge; Most of the suckling mice were also protected against FMDV challenge; indicating that antibodies produced in guinea pigs immunized with the TMVF11/TMVF14 mixture specifically neutralized FMDV | Wu et al. (2003) |
| Food and mouth disease | FMDV | Containing the genes encoding the polyprotein P1 and the protease 3C of FMDV | Alfalfa plants | None tested | Parenterally immunized mice developed a strong antibody response and were completely protected when challenged with the virulent virus | Dus Santos et al. (2005) |
| Food and mouth disease | FMDV | The epitope of t FMDV fused with a hepatitis B core protein | Tobacco (Nicotiana tabacum) plants | None tested | Mice, immunized intraperitoneally with a soluble crude extract of transgenic tobacco leaves, were found to produce specific antibody responses to both HBcAg and FMDV VP1 | Huang et al. (2005a) |
| Food and mouth disease | FMDV | Containing FMDV VP1 gene and the selective marker aadA gene | Tobacco chloroplast | 2–3 % of TSP | None tested | Li et al. (2006c) |
| Food and mouth disease | FMDV; Cholera toxin | pBI121CTBVP1, (FMDV protein VP1 fused with CT- B subunit) | Potato (Solanum tuberosum) | None tested | None tested | He et al. (2007) |
| Food and mouth disease | FMDV | Structural protein VP1 of FMDV | Stylosanthes guianensis cv. Reyan II. | 0.1–0.5 % of TSP | Mice developed a virus-specific immune response to the structural VP1 and intact FMDV particles | Wang et al. 2008a) |
| Food and mouth disease | FMDV | P1-2A3C, the structural polyprotein, P1-2A, and protease, 3C, from FMDV | Tomato plants | None tested | Guinea pigs developed a virus-specific antibody response against FMDV. Vaccinated guinea pigs were fully protected against a challenge infection | Pan et al. (2008) |
| Food and mouth disease | FMDV | A single chain variable antibody fragment (scFv) recognizing FMDV coat protein VP1 | Tobacco plants | 0.8 % of TSP | None tested | Joensuu et al. (2009) |
| Hepatitis B | HBVb | HBsAg (S protein) | Tobacco (Nicotiana tabacum cv. Samsun) | 16.5 ng/g FW | None tested | Mason et al. (1992) |
| Hepatitis B | HBV | HBsAg | Tobacco (Nicotiana tabacum cv. Samsun) | None tested | Mice elicited B- and T-cell responses | Thanavala et al. (1995) |
| Hepatitis B | HBV | pre-S2 + S gene (M protein) | Nicotiana benthamiana leaves | None tested | M protein provoked stronger serum antibody responses against HBsAg in mice and humans | Huang et al. (2005b) |
| Hepatitis B | HBV | HBsAg | Lupin (Lupinus luteus L.) | 11–150 ng/g FW | Mice developed significant levels of HBsAg-specific antibodies | Kapusta et al.(1999) |
| S protein | Potato tubers | 16 ug/g FW | A primary immune response was induced in mice fed transgenic tubes which could be greatly boosted by intraperitoneal delivery of a single subimmunogenic dose of commercial HBsAg vaccine. And mice elicited B-cell and T-cell responses | Richter et al. (2000) | ||
| Hepatitis B | HBV | S protein | Potato | 200–400 ng/g FW | None tested | Shulga et al.(2004) |
| Hepatitis B | HBV | S protein | Tomato | 40–160 ng/gFW | None tested | Ma et al.(2002) |
| Hepatitis B | HBV | S protein | Tomato | None tested | None tested | Carolina and Francisco (2004) |
| Hepatitis B | HBV | S protein | Tomato | None tested | None tested | Wang and Li (2008) |
| Hepatitis B | HBV | S protein | Cherry tomato | None tested | None tested | Hao et al. (2007) |
| Hepatitis B | HBV | S protein | Peanut Arachis hypogaea | 2.41 ug/gFW | None tested | Zhu et al.(2006) |
| Hepatitis B | HBV | S protein | Banana | 38 ng/g FW (pEFEHBS) 19.92 ng/g FW (pHER) | None tested | Sunil-Kumar et al. (2005) |
| Hepatitis B | HBV | S protein | Banana | None tested | None tested | Elkholy et al. (2009) |
| Hepatitis B | HBV | S protein | Kelf | 6.67 ug/gFW | None tested | Jiang et al. (2002) |
| Hepatitis B | HBV | S protein | Cherry tomatillo | 2–10 ug/gFW | Elicited an immediate high level recall antibody response | Gao et al. (2003) |
| Hepatitis B | HBV | S protein | Carrot | None tested | None tested | Zhao et al. (2002) |
| Hepatitis B | HBV | HBsAg (S protein) | Lettuce (Lactuca sativa L.) | 1-1.5 ng/g FW | Human volunteers developed specific serum-IgG response to plant-produced protein | Kapusta et al.(1999) |
| Hepatitis B | HBV | preS2–S protein | Carrot | 10 ng/g FW | None tested | Deineko et al. (2009) |
| Hepatitis B | HBV | L protein | Apple | 11.18 ng/gFW | None tested | Lou et al. (2005) |
| Porcine viral diarrhea disease | PEDVc | Neutralizing epitope of Korean PEDV (K-COE) | Tobacco plants | 0.1 % of TSP | None tested | Kang et al. (2005b) |
| Porcine viral diarrhea disease | PEDV | Neutralizing epitope of Korean PEDV (K-COE) | Potato plants | 0.1 % of TSP | None tested | Kim et al. (2005) |
| Porcine viral diarrhea disease | PEDV | The synthetic neutralizing epitope gene of PEDV | No-nicotine tobacco plants | 2.1 % of TSP | None tested | Kang et al. (2005a, b) |
| Porcine viral diarrhea disease | PEDV | Spike gene of PEDV | Sweet potato plants | None tested | None tested | Yang et al. (2005) |
| Porcine viral diarrhea disease | PEDV | A fusion protein consisting of the synthetic LTB and a synthetic neutralizing epitope of PEDV | Tobacco plants | None tested | None tested | Kang et al. (2006a, b, c) |
| Porcine viral diarrhea disease | PEDV | A fusion protein consisting of LTB and a synthetic core- neutralizing epitope (COE) of PEDV | Rice endosperm | None tested | None tested | Oszvald et al. (2007) |
| Porcine viral diarrhea disease | PEDV | A fusion gene encoding the synthetic enterotoxigenic LTB genetically fused with a synthetic neutralizing epitope of PEDV (sLTB-sCOE) | Lettuce cells (Lactuca sativa) | 0.026 ~ 0.048 % of TSP | None tested | Huy et al. (2009) |
| Porcine viral diarrhea disease | Rotavirus | Capsid protein (VP6) | Nicotiana benthamiana plants | None tested | None tested | O’Brien et al. (2000) |
| Porcine viral diarrhea disease | Bovine group A rotavirus | Capsid protein VP6 | Potato plants | None tested | Adult BALB/c mice after immunization showed the anti-VP6 response. | Matsumura et al. (2002) |
| Viral diarrhea disease | Murine rotavirus | Murine rotavirus gene six encoding the 41 kDa group specific capsid structural protein VP6 | Potato (Solanum tuberosum) | 0.01 % of TSP | Oral immunization of CD-1 mice generated measurable titers of both anti-VP6 serum IgG and intestinal IgA antibodies | Yu and Langridge (2003) |
| Rabies | Rabies virus | A gene for the rabies virus glycoprotein (G-protein) | Tomato plants | None tested | None tested | McGarvey et al. (1995) |
| Rabies | Rabies virus and HIV | Recombinant virus gene of rabies virus and HIV | Tobacco | None tested | Both antigens elicited specific virus-neutralizing antibodies in immunized mice | Yusibov et al. (1997) |
| Rabies | Rabies virus | Antigenic determinants from rabies virus glycoprotein (G protein) and nucleoprotein (N protein) | Nicotiana tabacum cv. Samsun NN plants; Nicotiana benthamiana and spinach (Spinacia oleracea) plants | None tested | Mice immunized with recombinant virus were protected against challenge infection | Yusibov et al. (2002) |
| Rabies | Rabies virus | A synthetic gene coding for the surface glycoprotein (G protein) of rabies virus | Tobacco | 0.38 % of TSP | Mice immunized intraperitoneally elicited high level of immune response. The plant-derived G protein induced complete protective immunity in mice against rabies virus | Ashraf et al. (2005) |
| Rabies | Rabies virus | A human anti-rabies virus monoclonal antibody | A Nicotiana tabacum cv. Xanthi cell culture | None tested | None tested | Girard et al. (2006) |
| Rabies | Rabies virus | A full-length nucleoprotein gene of rabies virus | Tomato; Nicotiana benthamiana | 1–5 % of TSP in tomato and 45 % in N. benthamiana. | Mice were immunized both intraperitoneally (i.p.) and orally with tomato protein extracts containing the N protein induced the production of antibodies. The antibody titer of mice immunized i.p., was at least four times higher than that of mice immunized orally. | Perea Arango et al. (2008) |
| Rabies; acute diarrhea | Rabies virus and cholera toxin | Rabies glycoprotein fused with B subunit of cholera toxin | Tobacco | 0.4 % of TSP | None tested | Roy et al. (2010) |
| SARS | SARS virus | A partial spike (S) protein of SARS-CoV | Tobacco plant cytosol and chloroplasts | None tested | None tested | Li (2006b) |
| AIDS | HIV | Tat protein of HIV-1 vaccine | Spinach | None tested | Mice having previously received oral Tat developed higher antibody titers to Tat than did the controls | Karasev (2005) |
| Newcastle disease | Newcastle disease virus | Fusion (F) protein of NDV | Maize | None tested | Feeding chickens with kernels containing the F protein induced the production of antibodies, which conferred protection against a viral challenge | Guerrero-Andrade et al. (2006) |
| Epidemic gastroenteritis | Norwalk virus | Norwalk virus capsid protein | Tomato | None tested | Four doses of 0.4 g freeze-dried tomato fruit containing 64 μg rNV (40 μg VLPs) induced NV-specific serum IgG and mucosal IgA in ≥ 80 % of mice, while doses of 0.8 g elicited systemic and mucosal antibody responses in all mice | Zhang (2006) |
| Epidemic gastroenteritis | Norwalk virus | The capsid protein of Norwalk virus | Tobacco and potato | None tested | Mice developed serum IgG specific for rNV | Mason et al. (1996) |
| Tuberculosis (TB) | Bacterium Mycobacterium tuberculosis | Tuberculosis (TB) ESAT6 antigen | Tobacco | 800 mg/kg of fresh leaves | None tested | Dorokhov et al. (2007) |
| Avian influenza | Pathogenic avian influenza A (H5N1) virus | Hemagglutinin (HA) | Barley grains | None tested | None tested | Bruchmüller et al. (2007) |
| Colorectal cancer | – | Colorectal cancer antigen EpCAM (pGA733) | Beta vulgaris var. cicla (Swiss chard) plants | None tested | Recombinant plant-derived antigen induced a humoral immune response in BALB/c mice | Brodzik et al. (2008) |
| Encephalitis | Encephalitis type B virus | Heavy-china variable domain of mAb against Encephalitis Type B virus | Tobacco | None tested | None tested | Wu and Xiong (1997) |
aFMDV—Food and mouth disease virus
bHBV—hepatitis B virus
cPEDV—porcine epidemic diarrhea virus
Foot-and-mouth disease virus vaccine
Foot-and-mouth disease (FMD) affects all domesticated cloven-hoofed animals (such as cows, pigs, and sheep) as well as wild ruminants. The causative foot-and-mouth disease virus (FMDV), an aphthovirus of the Picornaviridae family, is highly contagious. Thus, FMD is the most significant constraint to international trade in live animals and animal products today (Grubman and Baxt 2004). The FMDV structural protein VP1 plays a key role in the immunogenicity of the virion, which can elicit the production of specific antibodies against a major immunogenic site located between amino acids 140 and 160 (Meleon et al. 1979; Brown 1992).
The VP1 protein has been overexpressed in Arabidopsis thaliana, alfalfa, potato, and so on. Wigdorovitz et al. (1999) reported the development of transgenic plants of alfalfa expressing the structural protein VP1 of FMDV. Mice parenterally immunized by using leaf extracts from the transgenic plants developed a virus-specific immune response. Subsequently, Dus Santos et al. (2002) produced transgenic plants of alfalfa expressing the immunogenic site between amino acid residues 135–160 of structural protein VP1 (VP135–160) and the fused βGUS protein. After the FMDV epitope expressed in plants was highly immunogenic in mice, a strong anti-FMDV antibody response against a synthetic peptide representing the region VP135–160 was detectable. In 2005, Dus Santos et al. obtained the production of transgenic alfalfa plants containing the genes encoding the polyprotein P1 and the protease 3C of FMDV. Parenterally immunized mice developed a strong antibody response and were completely protected when challenged with the virulent virus. The transgenic tomato plants were also used to express the structural polyprotein P1-2A and protease 3C from FMDV. Guinea pigs immunized intramuscularly with foliar extracts from P1-2A3C-transgenic tomato plants were found to develop a virus-specific antibody response against FMDV (Pan et al. 2008). A single-chain variable antibody fragment (scFv) recognizing FMDV coat protein VP1 was expressed in transgenic tobacco plants. This scFv–ELP fusion accumulated up to 0.8 % of total soluble leaf protein in transgenic tobacco (Joensuu et al. 2009).
Hepatitis B virus vaccine
Hepatitis B virus (HBV) infection may lead to liver cirrhosis or hepatocellular carcinoma (Michel 2002). As a retrovirus, HBV is one of the smallest known eukaryotic DNA viruses. HBV negative strand has four ORFs, known as the C, P, S, and X gene, which respectively encode the four major viral proteins—core antigen (HBeAg/ HBcAg), HBV DNA polymerase (HBV DNA P), surface antigen (HBsAg), and X antigen (HBxAg). HBsAg, which is an outer membrane protein encoded by S zone of the HBV genome, is mainly related to the HBV vaccine.
Richter et al. (2000) developed transgenic potatoes expressing 1.1 μg g−1 HBsAg fresh tubers. By being fed three weekly doses of transgenic potato tubers plus 10 mg cholera toxin (CT), mice produced a primary serum antibody response that peaked at 73 mIU ml−1 3 weeks after the last dose. In addition, transgenic potato plants have been obtained by expressing HBsAg gene under the control of the double promoters of both 35S RNA of cauliflower mosaic virus (CaMV 35S) and the promoter of the patatin gene of potato tubers (Shulga et al. 2004). Huang et al. (2005) also constructed pMHB and transformed tobacco plants (Nicotiana benthamiana). The transgenic tobacco leaf extracts (with 1 μg of HBsAg) were administered to mice by intraperitoneal injection at weeks 0, 1, and 2. The serum anti-HBsAg antibody titers gradually increased and at the 12th week the immunized animals had an average titer of 1,165 mIU/ml.
Research has been reported on tomato transformation and regeneration containing HBsAg gene (Ma et al. 2002; Carolina and Francisco 2004; Wang and Li 2008). Our research group successfully trans-ferred HBsAg gene into cherry tomato (Lycopersivon esculentum Mill.) and peanut (a higher-expressing level of 2.41 μg/g FW) by Agrobacterium- mediated transformation (Zhu et al. 2006; Zhang et al. 2005; Guan et al. 2010).
Besides the plants noted above, an increasing number of transgenic plant systems are in the process of development, such as banana (Musa acuminate) (Sunil-Kumar et al. 2005; Elkholy et al. 2009), kelp (Thallus laminariae) (Jiang et al. 2002), cherry tomatillo (Physalis ixocarpa Brot) (Gao et al. 2003), carrot (Deineko et al. 2009), and soybean (Smith et al. 2002). Kapusta et al. (1999) took the lead in expressing HBsAg in lupin (Lupinus luteus) and lettuce (L. sativa) callus. These investigators found that the specific antibodies were produced in mice fed with the transformed callus. The HBV large-surface antigen gene PRS-S1S2S has also been expressed in transgenic apples (Lou et al. 2005).
Porcine viral diarrhea disease vaccine
Porcine viral diarrhea is a general term for some acute infectious diseases, which produce a water-like diarrhea and are caused by porcine transmissible gastroenteritis virus (TGEV), porcine epidemic diarrhea virus (PEDV), or porcine rotavirus (RTV).
Porcine transmissible gastroenteritis virus (TGEV) is the causative agent of acute diarrhea of newborn piglets, which provokes high mortality rates in affected farms. Protective immunity against this disease has to be developed in pregnant sows in order to confer passive protection to the piglets through colostrum and milk. Neutralizing antibodies against the virus are directed mainly to glycoprotein S, and relevant epitopes in neutralization have been mapped into the N-terminal domain of this protein (Garwes et al. 1978; Jiménez et al. 1986; Correa et al. 1988). Four major antigenic sites have been described in the globular part of glycoprotein S (gS), of which site A is considered to be immunodominant (De Diego et al. 1992, 1994; Sánchez et al. 1990). Transgenic potato plants were created to express the N-terminal domain of the glycoprotein S (N-gS) from TGEV. Extracts from transgenic potato tubers were inoculated intraperitoneally into mice, and the vaccinated mice developed serum IgG specific for TGEV (Gómez et al. 2000). In addition, Tuboly et al. (2000) generated three transgenic tobacco plant lines expressing the spike (S) protein of TGEV. The results showed that antigens from all these lines induced TGEV-specific immune responses in pigs, and the resultant antibody titers for all three constructs were similar.
Porcine epidemic diarrhea virus (PEDV) is an infectious, highly contagious virus of swine, which belongs to the Coronaviridae family (Pensaert and de Bouck 1978). PEDV destroys villus enterocytes and causes villus atrophy within the jejunum and ileum. It leads to enteritis in swine of all ages, and thus is often fatal in neonatal piglets (de Bouck and Pensaert 1980). Bae et al. (2003) developed transgenic tobacco plants that expressed the antigen protein corresponding to the neutralizing epitope of PEDV spike protein. Feeding the transgenic plants to mice induced both systemic and mucosal immune responses against the antigen. By using a tobacco mosaic virus (TMV)-based vector, the coding sequence of a core neutralizing epitope of PEDV (COE) gene was optimized based on the modification of codon usage in tobacco plant genes. The recombinant COE protein was up to 5.0 % of the total soluble protein in the leaves of tobacco plants infected with the TMV-based vector containing synthetic COE gene (Kang et al. 2004a). A fusion gene encoding the synthetic LT-B subunit genetically fused with a synthetic neutralizing epitope of porcine epidemic diarrhea virus (sLTB-sCOE) was introduced into lettuce cells (L. sativa) by Agrobacterium-mediated transformation methods. LTB-COE fusion protein comprised about 0.026–0.048 % of the total soluble protein in the transgenic lettuce leaf tissues (Huy et al. 2009).
Rotavirus is the leading cause of viral gastroenteritis in young children and animals worldwide. Rotavirus particles contain 11 dsDNA genomic segments surrounded by three concentric capsid protein layers: an inner layer VP2, an intermediate layer VP6, and an outer layer made up of capsid proteins VP7 and VP4 (Estes and Cohen 1989). Capsid protein VP6 is the major structural protein of rotavirus and makes up about 50 % of virion mass (Hsu et al. 1997). VP6 has been expressed in N. benthamiana plants either independently or as a fusion with the potato virus X coat protein. These plant-produced VP6 retained the ability to form trimers (O'Brien et al. 2000). Matsumura et al. (2002) reported expression of the major capsid protein VP6 of bovine group A rotavirus (GAR) in transgenic potato plants. Adult BALB/c mice were immunized intraperitoneally with concentrated transgenic potato extracts emulsified in Freund's adjuvant. Sera collected after immunization showed the anti-VP6 response. Subsequently, murine rotavirus gene six encoding VP6 was stably inserted into the Solanum tuberosum genome. The amount of VP6 protein in transgenic potato leaf and tuber was approximately 0.01 % of total soluble protein. Oral immunization of CD-1 mice generated measurable titers of both anti-VP6 serum IgG and intestinal IgA antibodies (Yu and Langridge 2003).
Rabies virus vaccine
Rabies virus, a rhabdovirus of the genus Lyssavirus, remains a significant threat to human and animal health throughout much of the world (Meslin et al. 1994). In the design of a rabies vaccine, representation of both the rabies virus glycoprotein (G protein) and nucleoprotein (N protein) antigens is desirable. The G protein is the major antigen responsible for the induction of protective immunity, while the N protein triggers rabies virus-specific T cells, facilitating the production of neutralizing antibodies and other immune mechanisms (Cox et al. 1977; Tollis et al. 1991).
A chimeric peptide containing antigenic determinants from rabies virus glycoprotein and nucleoprotein was cloned and expressed in tobacco and spinach plants. Mice immunized with recombinant virus were protected against challenge infection. Three of five human volunteers responded against the peptide antigen after ingesting infected spinach leaves (Yusibov et al. 2002). A Nicotiana tabacum cv. Xanthi cell culture was initiated from a transgenic plant expressing a human anti-rabies virus monoclonal antibody. Quantification of antibody production in plant cell suspension culture revealed 30 μg g−1 of cell dry weight for the highest-producing culture (0.5 mg/L), three times higher than from the original transgenic plant (Girard et al. 2006). Perea Arango et al. (2008) expressed a full-length nucleoprotein gene of rabies virus in transgenic tomato plants and also transiently expressed it in N. benthamiana plants by agroinfiltration. In both cases, the nucleoprotein was expressed at high levels, 1–5 % of total soluble protein in tomato and 45 % in N. benthamiana. In addition, Roy et al. (2010) reported the expression of a chimeric protein comprising the synthetic CT-B fused at its C-terminal with rabies surface glycoprotein (G protein) in tobacco plants. The approximately 80.3 kDa fusion polypeptide expressed at 0.4 % of the total soluble protein in leaves of the selected transgenic lines.
Other viral diseases
In recent years, there were other recombinant virus vaccines expressed in the transgenic plants, including: Tat protein of HIV-1 vaccine produced in spinach (Karasev et al. 2005), expression of the Newcastle disease virus (NDV) fusion protein in transgenic maize (Guerrero-Andrade et al. 2006), accumulation of recombinant SARS-CoV spike protein in tobacco plant cytosol and chloroplasts (Li et al. 2006b), Norwalk virus capsid protein expressed in tomato plant (Zhang et al. 2006), transgenic tobacco and potato (Mason et al. 1996), superexpression of tuberculosis antigens in tobacco leaves (Dorokhov et al. 2007), expression of influenza A (H5N1) vaccine in barley grains for oral bird immunization (Bruchmüller et al. 2007), and expression of EpCAM antigen in Beta vulgaris var. cicla (Swiss chard) plants (Brodzik et al. 2008).
Malaria vaccine
A gene encoding the C-terminal region of a major surface antigen of Plasmodium falciparum, referred to as PfMSP119, a major vaccine candidate for malaria, was cloned and transformed in tobacco leaves (Ghosh et al. 2002). Wang et al. (2008b) induced protective immunity against malaria infection in transgenic tobacco plants using Plasmodium yoelii merozoite surface protein 4/5 (PyMSP4/5) in a mouse model of malaria infection. PyMSP4/5 protein from the codon-optimized construct accumulated to 0.25 % of total soluble protein. In addition, tobacco-produced PyMSP4/5 protein was able to induce antigen-specific antibodies in mice.
The plant expression vector for the truncated fragments of SAG1 gene (t2SAG1) of Toxoplasma gondii was constructed and transformed into tomato plants by Agrobacterium tumefaciens under control of the both constitutively expressed cauliflower mosaic virus (CaMV) 35S promoter and the tomato fruit-specific E8 promoter (Zhou et al. 2004). In addition, there are relevant studies on the expression of a His-tagged truncated version of T. gondii dense granule 4 protein (Gra4163-345) in tobacco leaves (Ferraro et al. 2008), the protective antigenic gene FH3 of Fasciola hepatica in alfalfa plants (Li et al. 2003), and the ferritin gene of Schistosoma japonicum in rape plants (Yuan et al. 2008).
Immunocontraceptive vaccine
Fertility control may provide a non-lethal, more humane alternative for population control of vertebrate animals. A long-term strategy for it is to adopt immunocontraception, namely, that vaccination is used to induce antibodies against reproductive self-antigens, resulting in reduced fertility (Ferro 2002). In most cases, the zona pellucida (ZP) has been used as the vaccine antigen, which is composed of three sulfated glycoproteins; it can function in the fertilization of the oocyte by providing a substrate for sperm binding (Bleil and Wassarman 1980a; Munro and Pelham 1987). One of the three glycoproteins, ZP3, is the primary binding site for the sperm and has been investigated as a target for immune contraception (Bleil and Wassarman 1980b; Sacco 1979).
Marsupial-specific reproductive antigens expressed at high levels in edible transgenic plant tissue might provide a kind of safe, effective, and cheap oral delivery bait for immunocontraceptive control. As proof of concept, female possums vaccinated with immunocontraceptive antigens showed reduced fertility, and possums fed with potato-expressed LT-B had mucosal and systemic immune responses to the antigen. This demonstrated that immunocontraception was effective in possums and oral delivery in edible plant material might be possible (Polkinghorne et al. 2005).
High-level expression strategies of foreign protein in transgenic plants
In the early literature, expression levels of recombinant antigens in transformed plants were not high. Therefore, the main problem of transgenic plant vaccines was a high-level expression of foreign genes in the plants. Many scientists have launched research programs in order to improve the antigen expression level and to make plant vaccines produce effective immunological function with some significant achievements that will be described in the following.
Promoter optimization
The choice of a suitable plant promoter is an important way to enhance exogenous gene expression. At present, constitutive promoters are usually used in plant genetic engineering; for example, CaMV 35S promoter is used for dicotyledonous plants, and the actin promoter of rice and ubiquitin promoter of maize are used for monocotyledonous plants. However, constitutive promoters may cause a constant and continuous expression of target genes in plant tissues, leading to an excessive consumption of material and energy within the cell. Thus, constitutive expression may result in the inability to effectively regulate the expression of target genes both temporally and spatially. The consequence may be to interrupt the intrinsic metabolic balance in the plant, which is not conducive to the enhancement of the yield and the quality of the transgenic plant expression of target genes. Instead, exogenous genes should be expressed in particular periods and specific tissues or organs, rather than with a continuously high-expression level in receptor plants, in order to maximize production of plant-based vaccines or other medically relevant proteins. Therefore, the use of inducible or tissue-specific promoters may effectively improve the expression level of foreign proteins in a particular period or in specific organs. For example, tomato plants were transformed with the gene encoding CT-B along with an endoplasmic reticulum retention signal (SEKDEL) under the control of the CaMV 35S promoter via Agrobacterium-mediated transformation. Both tomato leaves and fruits expressed CTB at levels up to 0.02 % and 0.04 % of total soluble protein, respectively (Jani et al. 2002), but under the control of the tomato fruit-specific E8 promoter, transgenic tomato plants expressing CTB protein were developed, with the highest amount of CTB protein being 0.081 % of total soluble protein (Jiang et al. 2007).
Codon optimization
The difference in codon usage in plants, animals, and microbes is an issue that must be considered in any heterologous protein expression system. In order to optimize expression, the codons of foreign genes should be analyzed and replaced with appropriate codons preferred by plants. Codon optimization can significantly improve the expression level of the exogenous gene in plants. Mason et al. (1998) constructed a plant-optimized synthetic gene encoding LT-B for use in transgenic plants as an edible vaccine against enterotoxigenic E. coli. Expression of the synthetic LT-B gene in potato plants under the control of a constitutive promoter yielded increased accumulation (ranging from 5-to 40-fold) of LT-B in leaves and tubers, as compared to the bacterial LT-B gene. Dus Santos et al. (2005) transformed the genes encoding the polyprotein P1 and the protease 3C of FMDV by the codon preferred by A. thaliana into alfalfa plants. The highest accumulation of VP1 protein was found in transformed leaves at 0.5–1 mg/g fresh weight. Other research has shown that plant-optimized coding sequences can greatly enhance the expression of other bacterial genes in plants (Perlak et al. 1991; Adang et al. 1993).
Signal peptide utilization
The synthetic peptide in the cytoplasm and endoplasmic reticulum can further be located on different areas of the cell by the decision of a peptide signal sequence. The endoplasmic reticulum in plant cells provides an exogenous protein with a relatively stable environment for protein maturation. For this reason, the addition of an ER-retention signal at the C terminus has been suggested to enhance the accumulation of recombinant proteins in transgenic plants (Munro and Pelham 1987; Napier et al. 1998; Wandelt et al. 1992). Stoger et al. (2000) reported that protein accumulation was irrespective of the mRNA level, and suggested that the addition of KDEL at C terminus resulted in the accumulation of antibodies in plants. For example, the G protein of the rabies virus, which contained an endoplasmic reticulum retention signal in its C terminus, expressed in transgenic tobacco plant lines at 0.38 % of the total soluble leaf protein (Ashraf et al. 2005).
Addition of immune adjuvants
LT-B and CT-B have been shown to be effective carriers and adjuvants for genetically linked foreign proteins (Bagdasarian et al. 1999; Arakawa et al. 1998a; b). They can bind to ceramide-galactose sugar receptor molecules such as GM1 ganglioside (Holmgren et al. 1975). The performances of LTB carrier and adjuvant in animals immunized with the LTB-antigen fusion proteins are contingent on pentamer assembly. They can make their toxicity subunits (A) closely combine on the cell surface of intestinal mucosa so that fusion protein more easily react with gastrointestinal mucosa, thus stimulating mucosa to produce IgA and serum IgG and strengthening the immunity effect (Kim et al. 2007). Therefore, LTB and CTB can be used as immune adjuvants by fusing with antigenic determinant gene to enhance hapten immunogenicity.
Adoption of the chloroplast transformation system
Due to the high copy number of the chloroplast genome, chloroplast transformation can greatly improve the level of recombinant protein in transgenic plants. Integration of an unmodified CTB-coding sequence into chloroplast genomes (up to 10,000 copies per cell) resulted in the accumulation of up to 4.1 % of total soluble tobacco leaf protein as functional oligomers (410-fold higher expression levels than that of the unmodified LTB gene expressed via the nuclear genome) (Daniell et al. 2001). In order to resist the fungal pathogen Colletotrichum, that is so destructive to tobacco, the antimicrobial peptide MSI-99, an analog of magainin 2, was expressed via the chloroplast genome to obtain high levels of expression in transgenic tobacco (Nicotiana tabacum var. Petit Havana) plants (DeGray et al. 2001). The chloroplasts were estimated to express MSI-99 at 21.5 % to 43 % percent of total soluble protein (TSP). At present, a variety of vaccines (which can protect against Cholera, tetanus, anthrax, amebic disease, rotavirus, and so on) have had a successful expression in the plant chloroplasts (Verma and Daniell 2007). However, without the endoplasmic reticulum and golgi apparatus, the synthesis of protein in chloroplasts cannot be modified by glycosylation. Thus, chloroplasts are suitable only for expressing bacterium antigens and not for glycoproteins.
Safety issues and solutions for transgenic plant vaccines
The good qualities of GM plants are produced according to human needs, and the subsequently developed transgenic plants may create enormous economic and social benefits for mankind. However, as new species, once they are released to the natural environment, GM plants may bring mankind tremendous potential or real harm. In this regard, we have had profound lessons of history, such as the famous British Pusztai events, Cornell University butterflies event (Losey et al. 1999), Canada “superweeds” event (Orson 2001), Mexico corn event (Dalton 2001) and China Bt insect-resistant cotton event. All of these events are wake-up calls for the whole human race in all fields of research and application of GM organisms. Recently, safety problems in the environment and the role of transgenic plant vaccines in the human diet have become the focus of controversy.
Possible environmental safety problems
The effects of transgenic plants to non-objective organisms
The genes obtained by transformation technology will not only be spread to nature, possibly enter into the wild group through genetic drift, change genetic traits of the wild population, and then possibly cause adverse effects. Therefore, the release of genetically modified organisms into nature may contaminate the natural gene pool, break original ecological balance, and produce some unsuspected impact on the ecological environment (Ma et al. 2001). Meanwhile, beside pest and germ toxigenicity, insect or disease resistance by transgenic plants released into the environment may have direct or indirect negative effects on many beneficial organisms in the environment and even cause the death of some beneficial organisms (Zhuang and Cao 2008).
New environmental problems
Many genetically modified varieties contain the genes extracted from Bacillus, which may produce a kind of protein toxic to pests. If the anti-pest GM crop is planted for a long time in large areas, pests are likely to develop resistance and then pass it on to their offspring. So transgenic plants no longer have insect-resistance ability, the original pesticides will also no longer be effective, and eventually more intractable super pests will produce (Chen and Wang 2002). Meanwhile, transgenic plants released into the environment may generate gene transfer through pollination and allow some genes (such as genes of insect resistance, disease resistance, herbicide or environmental stresses tolerant) to become dominant over the related wild species or weeds (World Health Organization. 2004). Once obtaining adversity-resistant character of transgenic organisms, these weeds will become “superweeds”, severely endanger normal survival and growth of other creatures, and eventually disrupt the balance of the ecosystem.
The impacts on biodiversity
Hilbeck et al. (1998) fed transgenic Bacillus thuringiensis variety kurstaki (Berliner) corn plants to the European corn borer (lepidopterous target pest) and Spodoptera littoralis (Boisduval) (lepidopterous nontarget pest for B. thuringiensis). Mean total immature mortality for chrysopid larvae raised on B. thuringiensis-fed prey was 62 % compared with 37 % when raised on B. thuringiensis-free prey. Some biologists think that in order to keep its own stability and purity, nature's species strictly regulated the change of genetic material and genetic drift is limited to between the same species or allied species. A GM organism is obtained through an artificial method of mutual transfer between animals, plants, microbes and even human genes, so it breaks through the traditional concept of kingdom. GM organisms open a new door and span the intrinsic barrier among the species, thereby presenting many features and advantages that native species do not possess. If such species are released into the environment, the competitive relationship between species will change. The original natural ecological balance will be destroyed, the intrinsic complete food chain in ecological systems will also be damaged, and eventually species extinction and loss of biodiversity are all possibilities that must be considered (Yang et al. 2002).
Possible dietary safety problems
Toxicity
Food safety is a necessary prerequisite for diet, so the safety of bioengineered food products is currently of the greatest concern to people. The nutrient component changes and the appearance of anti-nutritive factors in some GM organism bodies (such as the changes of protease inhibitors or lipoxidase) are likely to produce some adverse effects on human health. Moreover, people know little about gene activity and worry that the sudden change of genes will lead to the production of some toxic substances (such as solanine and cucurbitacin). Furthermore, the transformed genes are not the ones of the original parent animals and plants, some of which even come from other creatures of different genera (including all sorts of bacteria, viruses, and organisms). Therefore, the safety of exogenous gene expression product is in doubt.
Irritability
The allergic substances may increase, and new allergens will also appear in the food supply due to gene transformation. Food anaphylaxis involves various abnormal reactions of human immune system to a certain protein. It has been discovered that peanut protein in soybean (Glycine max), Cry 9C poisonous protein in corn and 2S albumin in Brazil nuts may induce some people or animals' allergic reaction (Nordlee et al. 1996).
Antibiotic resistance
At present, most of the selected vectors in genetic engineering are antibiotic-resistant marker genes. The antibiotic resistance is introduced into the food chain through gene transfer. Therefore, it may produce drug-resistant bacteria or viruses within humans or animals, induce people to develop resistance to particular kinds of antibiotics, and eventually affect the effectiveness of antibiotic treatment (Cheng 2002). In 2002, a human DNA residual test of GM food was demonstrated in Britain. Seven volunteers, who underwent operations for colonic tissues excision, ate hamburgers with GM soybean. In another instance, a transgenic DNA residue was detectable inside small intestine intestinal flora (Chen and Yuan 2008). For transgenic soybean with marker antibiotics, it is thought that the antibiotic-containing food makes intestinal or oral bacteria produce a kind of resistance to antibiotics, but this has not been established.
Resolution of safety issues in transgenic organisms
Choice of biosafety marker genes
Marker genes in transgenic plants mainly include selected genes (enrichment of transformed cells, many of which are antibiotic-resistance genes) and reporter genes (easily detected expression products, such as green fluorescent protein (GFP) gene). In order to avoid some of the potential dangers of traditional gene selection (such as genetic drift, the development of immune tolerance in the human body, and damage to the ecological balance), marker genes should be eliminated or a positive screening system should be developed. The knock-out of marker genes has the drawbacks of low efficiency and complex operation; the most effective method at present is to screen for biological safety by using non-resistance marker genes. Although the transformed cell cannot be killed, the positive screening method may make the cell have a specific metabolic advantage or exploit a specific material by introducing specific genes, so as to develop vigorous growth and to achieve the screening purpose. The safe marker genes for the positive selection system are found to include mainly as follow: the genes coding enzymes that catalyze special sugars (pmi and xylA), the genes coding enzymes that interfere with amino acid metabolism (ak and dapA), green fluorescent protein (GFP) gene, beta–Glucuronidase (GUS) gene, ribitol operon(rtl) gene and chlorophyllous biosynthesis enzyme gene (hemL).
Reduction of unexpected side effects and enhancement of genetic stability
Due to the characteristics of transgenic technology, transformed plants can easily produce unexpected side effects. The integration of foreign DNA can also affect chromosome structures in the host plant so as to increase the mutation rate in the plant. The molecular detection methods used to detect such negative side effects are mainly proteomic analysis technology (such as two-dimensional electrophoresis) and gene expression analysis technology (such as gene chip technology), both of which have international general “real identity” principle as a guide to analyze the safety of transgenic plants. It is much more difficult to analyze chromosomal mutation in transgenic plants. However, it is remarkable that although mutation induced by change in chromosomal structure is an unfavorable factor in the commercialization of plant genetic engineering, it will provide the material basis for research on the transcription mechanisms of foreign genes and the influence of foreign genes on the genome of receptor plants.
The technique of the exogenous gene removal
According to a report in the Beijing Agricultural Journal, after 6 years of hard work, the laboratory research team led by Chinese scientist Li Yi made a great breakthrough in the research on the potential threats of transgenic plants to ecological environment and human health (Chui and Li 2007). They developed a technique to remove an exogenous gene and to insert the special gene in the target plants, which was controlled by a DNA regulatory fragment promoter. According to the intention of researchers, the special gene could spontaneously remove the exogenous gene at a specific time or a specific location in the plant. The pollen, sedes, and fruit of transgenic plants no longer contain foreign genes by using this method. Thus, a new method has been developed that permits non-transformed plants to be produced by using transgenic plants. This technique is expected to thoroughly resolve safety problems of genetically modified plants for application in food and medicine.
Other strategies
In addition to the above methods, there are other strategies, such as to improve the security management system of genetically modified organisms, to execute area-delimited management of production, to strengthen import–export control of transgenic agricultural products and to raise the level of safety consciousness of all people to transgenic foods.
Summary and perspectives
Transgenic plant vaccines have significant advantages compared with traditional vaccines. Future research should focus on producing safe, reliable, and efficient plant vaccines as pharmaceutical products. The low level of current expression systems is one of the major limitations to application of transgenic plants vaccines. However, recent publications show that expression levels can be improved by the chloroplast transformation, plant breeding, or food processing technology. Moreover, it is also been indicated that the application of carrier protein and auxiliary proteins or adjuvants may enhance the ability of the immune system to recognize antigens (Rigano and Walmsley 2005). This research has enabled the production of cheap and safe plant vaccines that can be administered orally to prevent the spread of disease. However, there is still a long way to go to industrialize transgenic plant vaccines. Especially some European countries continue to resist the use of transgenic plants, which poses difficulties for the development of transgenic plant vaccines. Consequently, much vaccine research is still in the experimental stage. Therefore, in order to promote the rapid development of transgenic plant vaccines, technological innovation should be intensified, and at the same time, the public should be informed about the advantages of transgenic plants to remove the fear. In 2006, the United States Department of Agriculture (USDA) approved the commercialization of transgenic plants vaccine against Newcastle disease. Although the vaccine has not yet reached the market, there is no doubt that this is a successful landmark example for transgenic plant vaccines. It is hoped that this example will play a significant role in promoting the development of other transgenic plant vaccines. With the establishment and optimization of GM technologies and corresponding means of detection, transgenic plant vaccines and antibodies have the prospect for vast applications in industrialized production.
Acknowledgments
This work was financially supported by National Natural ScienceFoundation of China (No. 31000144); Opening Foundation of Key Laboratory of Resource Biology and Biotechnology in Western China (Northwest University) (08JZ72); Specialized Foundation of Department of Education of Shaanxi Province, China (09JK746); Northwestern university graduate self-innovation project (10YZZ36); Yuncheng University Doctor Scientific research project (YQ-2011042). We are grateful to all members of Key Laboratory of Resource Biology and Biotechnology in Western China for numerous valuable discussions and advice.
Footnotes
Zheng-jun Guan and Bin Guo contributed equally to this paper.
Contributor Information
Zheng-jun Guan, Email: zhengjunguan@126.com.
Bin Guo, Email: guobin@nwu.edu.cn.
Ya-hui Wei, Phone: +86-029-88302199, FAX: +86-029-88302199, Email: weiyahui@nwu.edu.cn.
References
- Adang MJ, Brody MS, Cardineau G, Eagan N, Roush RT. The reconstruction and expression of a Bacillus thruingiensis cry/l/A gene in protoplasts and potato plants. Plant Mot Biof. 1993;21:1131–1145. doi: 10.1007/BF00023609. [DOI] [PubMed] [Google Scholar]
- Arakawa T, Chong DX, Merritt JL, Langridge WHR. Expression of cholera toxin B subunit oligomers in transgenic potato plants. Transgenic Res. 1997;6:403–413. doi: 10.1023/A:1018487401810. [DOI] [PubMed] [Google Scholar]
- Arakawa T, Chong DKX, Langridge WHR. Efficacy of a food plant-based oral cholera toxin B subunit vaccine. Nat Biotechnol. 1998;16:292–297. doi: 10.1038/nbt0398-292. [DOI] [PubMed] [Google Scholar]
- Arakawa T, Yu J, Chong DK, Hough J, Engen PC, Langridge WH. A plant-based cholera toxin B subunit-insulin fusion protein protects against the development of autoimmune diabetes. Nat Biotechnol. 1998;16:934–938. doi: 10.1038/nbt1098-934. [DOI] [PubMed] [Google Scholar]
- Ashraf S, Singh PK, Yadav DK, Shahnawaz M, Mishra S, Sawant SV, Tuli R. High level expression of surface glycoprotein of rabies virus in tobacco leaves and its immunoprotective activity in mice. J Biotechnol. 2005;119:1–14. doi: 10.1016/j.jbiotec.2005.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bae JL, Lee JG, Kang TJ, Jang HS, Jang YS, Yang MS. Induction of antigen-specific systemic and mucosal immune responses by feeding animals transgenic plants expressing the antigen. Vaccine. 2003;21:4052–4058. doi: 10.1016/S0264-410X(03)00360-8. [DOI] [PubMed] [Google Scholar]
- Bagdasarian MM, Nagai M, Frey J, Bagdasarian M. Immunigenicity of Actinobacillus ApxIA toxin epitopes fused to the E. coli heat-labile enterotoxin B subunit. Vaccine. 1999;17:441–447. doi: 10.1016/S0264-410X(98)00216-3. [DOI] [PubMed] [Google Scholar]
- Bleil JD, Wassarman PM. Mammalian sperm–egg interaction: identification of a glycoprotein in mouse egg zonae pellucidae possessing receptor activity for sperm. Cell. 1980;20:873. doi: 10.1016/0092-8674(80)90334-7. [DOI] [PubMed] [Google Scholar]
- Bleil JD, Wassarman PM. Structure and function of the zona pellucida: identification and characterization of the proteins of the mouse oocyte's zona pellucida. Dew Biol. 1980;76:185. doi: 10.1016/0012-1606(80)90371-1. [DOI] [PubMed] [Google Scholar]
- Brodzik R, Spitsin S, Golovkin M, Bandurska K, Portocarrero C, Steplewski Z, Koprowski H. Plant-derived EpCAM antigen induces protective anti-cancer response. Cancer Immunol Immunother. 2008;57:317–323. doi: 10.1007/s00262-007-0366-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown F. Vaccination against foot-and-mouth disease. Vaccine. 1992;10:1022–1026. doi: 10.1016/0264-410X(92)90111-V. [DOI] [PubMed] [Google Scholar]
- Bruchmüller A, Marthe C, Hensel G. Expression of influenza A (H5N1) vaccine in barley grains for oral bird immunization. J Verbraucherschutz Lebensmittelsicherheit. 2007;2(Suppl):118. doi: 10.1007/s00003-007-0280-x. [DOI] [Google Scholar]
- Carolina C, Francisco AC. Tomato transformation and transgenic plant production. Plant Cell Tissue Organ Cult. 2004;76:269–275. doi: 10.1023/B:TICU.0000009249.14051.77. [DOI] [Google Scholar]
- Carrillo C, Wigdorovitz A, Oliveros C. Protective immune response to foot and mouth disease virus with VP1 expressed in transgenic plants. J Virol. 1998;72:1688–1690. doi: 10.1128/jvi.72.2.1688-1690.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrillo C, Wigdorovitz A, Trono K. Induction of a virus-specific antibody response to foot-and-mouth disease virus using the structural protein VP1 expressed in transgenic potato plants. Virol Immunol. 2001;14:49–57. doi: 10.1089/08828240151061383. [DOI] [PubMed] [Google Scholar]
- Carter JE, Odumosu O, Langridge WHR. Expression of a ricin toxin b subunit: insulin fusion protein in edible plant tissues. Mol Biotechnol. 2010;44:90–100. doi: 10.1007/s12033-009-9217-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen YR, Wang H. Safety of gene engineering and transgenic foods. Food Science. 2002;23:145–149. [Google Scholar]
- Chen SL, Yuan Y. Genetically modified organisms and environment safety. China Tropical Medicine. 2008;8:67–71. [Google Scholar]
- Cheng ZQ. Safety evaluation of antibiotics genes in transgenic plants. Life Science. 2002;14:34–37. [Google Scholar]
- Chikwamba R, Cunnick J, Hathaway D, McMurray J, Mason H, Wang K. A functional antigen in a practical crop: LT-B producing maize protects mice against Escherichia coli heat labile enterotoxin (LT) and cholera toxin (CT) Transgenic Res. 2002;11:479–493. doi: 10.1023/A:1020393426750. [DOI] [PubMed] [Google Scholar]
- Chikwamba R, McMurray J, Shou H, Frame B, Pegg S-E, Scott P, Mason H, Wang K. Expression of a synthetic E. coli heatlabile enterotoxin B sub-unit (LT-B) in maize. Mol Breed. 2002;10:253–265. doi: 10.1023/A:1020509915672. [DOI] [Google Scholar]
- Choi NW, Estes MK, Langridge WHR. Synthesis and assembly of a cholera toxin b subunit-rotavirus vp7 fusion protein in transgenic potato. Mol Biotechnol. 2005;31:193–203. doi: 10.1385/MB:31:3:193. [DOI] [PubMed] [Google Scholar]
- Chui HC, Li Y. The solution of safety problem for genetically modified crops. International Talents Exchange. 2007;10:32–35. [Google Scholar]
- Correa I, Jiménez G, Suñé C, Bullido MJ, Enjuanes L. Antigenic structure of E2-glycoprotein of transmissible gastroenteritis coronavirus. Virus Res. 1988;10:77–94. doi: 10.1016/0168-1702(88)90059-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox JH, Dietzschold B, Schneider LG. Rabies virus glycoprotein. Part II. Biological and serological characterization. Infect Immun. 1977;16:754–759. doi: 10.1128/iai.16.3.754-759.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curtiss RI, Cardineau CA. Oral immunization by transgenic plants. World Plant Application. 1990;90:107–111. [Google Scholar]
- Da Silva JV, Garcia AB, Flores VMQ, de Macedo ZS, Medina-Acosta E. Phytosecretion of enteropathogenic Escherichia coli pilin subunit A in transgenic tobacco and its suitability for early life vaccinology. Vaccine. 2002;20:2091–2101. doi: 10.1016/S0264-410X(02)00057-9. [DOI] [PubMed] [Google Scholar]
- Dalton R. Transgenic corn found growing in Mexico. Nature. 2001;413:337. doi: 10.1038/35096714. [DOI] [PubMed] [Google Scholar]
- Daniell H, Lee SB, Panchai T, Wiebe PO. Expression of the native cholera toxin B subunit gene and assembly of functional oligomers in transgenic tobacco chloroplasts. J Mol Biol. 2001;311:1001–1009. doi: 10.1006/jmbi.2001.4921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daniell H, Khan MS, Allison L. Milestones in chloroplast genetic engineering: an environmentally friendly era in biotechnology. Trends Plant Sci. 2002;7:84–91. doi: 10.1016/S1360-1385(01)02193-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Bouck P, Pensaert M. Experimental infection of pigs with a new porcine enteric coronavirus, CV777. Am J Vet Res. 1980;1:219–223. [PubMed] [Google Scholar]
- De Diego M, Laviada MD, Enjuanes L, Escribano JM. Epitope specificity of protective lactogenic immunity against swine transmissible gastroenteritis virus. J Virol. 1992;66:6502–6508. doi: 10.1128/jvi.66.11.6502-6508.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Diego M, Rodríguez F, Alcaraz C, Gómez N, Alonso C, Escribano JM. Characterization of the IgA and subclass IgG responses to neutralizing epitopes after infection of pregnant sows with the transmissible gastroenteritis virus or the antigenically related porcine respiratory coronavirus. J Gen Virol. 1994;75:2585–2593. doi: 10.1099/0022-1317-75-10-2585. [DOI] [PubMed] [Google Scholar]
- DeGray G, Rajasekaran K, Smith F, Sanford J, Daniell H. Expression of an antimicrobial peptide via the chloroplast genome to control phytopathogenic bacteria and fungi. Plant Physiol. 2001;127:852–862. doi: 10.1104/pp.010233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deineko EV, Zagorskaya AA, Pozdnyakov SG, Filipenko EA, Permyakova NV, Sidorchuk YV. Comparative analysis of HBV M-antigen production in leaves of individual transgenic carrot plants. Doklady Biochem Biophys. 2009;425:76–79. doi: 10.1134/S1607672909020057. [DOI] [PubMed] [Google Scholar]
- Dorokhov YL, Sheveleva AA, Frolova OY, Komarova TV, Zvereva AS, Lvanov PA, Atabekov JG. Superexpression of tuberculosis antigens in plant leaves. Tuberculosis (Edinb) 2007;87:218–224. doi: 10.1016/j.tube.2006.10.001. [DOI] [PubMed] [Google Scholar]
- Dus Santos MJ, Wigdorovitz A, Trono K, Ríos RD, Franzone PM, Gil F, Moreno J, Carrillo C, Escribano JM, Borca MV. A novel methodology to develop a foot and mouth disease virus (FMDV) peptidebased vaccine in transgenic plants. Vaccine. 2002;20:1141–1147. doi: 10.1016/S0264-410X(01)00434-0. [DOI] [PubMed] [Google Scholar]
- Dus Santos MJ, Carrillo C, Ardila F, Ríos RD, Franzone PM, Piccone ME, Wigdorovitz A, Broca MV. Development of transgenic alfalfa plants containing the foot and mouth disease virus structural ployprotein gene P1 and its utilization as an experimental experimental immunogen. Vaccine. 2005;23:1838–1843. doi: 10.1016/j.vaccine.2004.11.014. [DOI] [PubMed] [Google Scholar]
- Elkholy SF, Ismail RM, Bahieldin A, Sadik AS, Madkour MA. Expression of hepatitis B surface antigen (HBsAg) gene in transgenic banana (Musa sp.) Arab J Biotechnol. 2009;12:291–302. [Google Scholar]
- Elson CO. Cholera toxin and its subunits as potential oral adjuvants. Curr Top Microbiol Immunol. 1989;146:29–33. doi: 10.1007/978-3-642-74529-4_3. [DOI] [PubMed] [Google Scholar]
- Estes MK, Cohen J. Rotavirus gene structure and function. Microbiol Rev. 1989;53:410–449. doi: 10.1128/mr.53.4.410-449.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferraro G, Becher ML, Angel SO, Zelada A, Mentaberry AN, Clemente M. Efficient expression of a Toxoplasma gondii dense granule Gra4 antigen in tobacco leaves. Exp Parasitol. 2008;120:118–122. doi: 10.1016/j.exppara.2008.06.002. [DOI] [PubMed] [Google Scholar]
- Ferro VA. Current advances in antifertility vaccines for fertility control and noncontraceptive applications. Expert Rev Anti Vacc. 2002;1:443–452. doi: 10.1586/14760584.1.4.443. [DOI] [PubMed] [Google Scholar]
- Field M, Rao MC, Chang EB. Intestinal electrolyte transport and diarrheal disease. N Engl J Med. 1989;321:800–806. doi: 10.1056/NEJM198909213211206. [DOI] [PubMed] [Google Scholar]
- Gao Y, Ma Y, Li M, Cheng T, Li SW, Zhang J, Xia NS. Oral immunization of animals with transgenic cherry tomatillo expressing HBsAg. World J Gastroenterol. 2003;9:996–1002. doi: 10.3748/wjg.v9.i5.996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garwes DJ, Lucas MH, Higgins DA, Pike BV, Cartwright SF. Antigenicity of structural components from porcine transmissible gastroenteritis virus. Vet Microbiol. 1978;3:179–190. doi: 10.1016/0378-1135(79)90034-8. [DOI] [Google Scholar]
- Ghosh S, Malhotra P, Lalitha PV, Guha-Mukherjee S, Chauhan VS. Expression of Plasmodium falciparum C-terminal region of merozoite surface protein (PfMSP119), a potential malaria vaccine candidate, in tobacco. Plant Science. 2002;162:335–343. doi: 10.1016/S0168-9452(01)00555-6. [DOI] [Google Scholar]
- Girard LS, Fabis MJ, Bastin M, Courtois D, Petiard V, Koprowski H. Expression of a human anti-rabies virus monoclonal antibody in tobacco cell culture. Biochem Biophys Res Commun. 2006;345:602–607. doi: 10.1016/j.bbrc.2006.03.219. [DOI] [PubMed] [Google Scholar]
- Gómez N, Wigdorovitz A, Castañón S, Gill F, Ordás R, Borca MV, Escribano JM. Oral immunogenicity of the plant derived spike protein from swine-transmissible gastroenteritis coronavirus. Arch Virol. 2000;145:1725–1732. doi: 10.1007/s007050070087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grubman MJ, Baxt B. Foot-and-mouth disease. Clin Microbiol Rev. 2004;17:465–493. doi: 10.1128/CMR.17.2.465-493.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guan ZJ, Guo B, Huo YL, Guan ZP, Wei YH. Overview of expression of hepatitis B surface antigen in transgenic plants. Vaccine. 2010;28:7351–7362. doi: 10.1016/j.vaccine.2010.08.100. [DOI] [PubMed] [Google Scholar]
- Guerrero-Andrade O, Loza-Rubio E, Olivera-Flores T, Fehérvári-Bone T, Gómez-Lim MA. Expression of the Newcastle disease virus fusion protein in transgenic maize and immunological studies. Transgenic Res. 2006;15:455–463. doi: 10.1007/s11248-006-0017-0. [DOI] [PubMed] [Google Scholar]
- Hao HY, Wei YH, Zhu JG. Expression of oral hepatitis B vaccine in transgenic tomato. Food Sci. 2007;28:201–204. [Google Scholar]
- Haq TA, Mason HS, Clements JD, Arntzen CJ. Oral immunization with a recombinant bacterial antigen produced in transgenic plants. Science. 1995;268:714–716. doi: 10.1126/science.7732379. [DOI] [PubMed] [Google Scholar]
- He DM, Qian KX, Shen GF, Li YN, Zhang ZF, Su ZL, Shao HB. Stable expression of foot-and-mouth disease virus protein VP1 fused with cholera toxin B subunit in the potato (Solanum tuberosum) Colloids Surf B: Biointerfaces. 2007;55:159–163. doi: 10.1016/j.colsurfb.2006.11.043. [DOI] [PubMed] [Google Scholar]
- Hidenori T, Takachika H, Li JY, Kaoru T, Ryotaro I, Hideyuki K, Fumio T. Efficient induction of oral tolerance by fusing cholera toxin B subunit with allergen-specific T-cell epitopes accumulated in rice seed. Vaccine. 2008;26:6027–6030. doi: 10.1016/j.vaccine.2008.09.019. [DOI] [PubMed] [Google Scholar]
- Hilbeck A, Baumgartner M, Fried P, Bigler F. Effects of transgenic Bacillus thuringiensis corn-fed prey on mortality and development time of immature Chrysoperla cornea (Neuroptera: Chrysopidae) Environmental Entomology. 1998;27:480–487. [Google Scholar]
- Holmgren J, Lonnroth I, Mansson J, Svennerholm L. Interaction of the cholera toxin and membrane Gm1 ganglioside of small intestine. Proc Nat Acad Sci USA. 1975;72:2520–2524. doi: 10.1073/pnas.72.7.2520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu GG, Bellamy AR, Yeager M. Projection structure of VP6, the rotavirus inner capsid protein, and comparison with bluetongue VP7. J Mol Biol. 1997;272:362–368. doi: 10.1006/jmbi.1997.1179. [DOI] [PubMed] [Google Scholar]
- Huang YH, Liang WQ, Wang YJ, Zhou ZA, Pan AH, Yang XH, Huang C, Chen JX, Zhang DB. Immunogenicity of the epitope of the foot-and-mouth disease virus fused with a hepatitis B core protein as expressed in transgenic tobacco. Viral Immunol. 2005;18:668–677. doi: 10.1089/vim.2005.18.668. [DOI] [PubMed] [Google Scholar]
- Huang Z, Elkin G, Maloney BJ. Virus-like particle expression and assembly in plants: hepatitis B and Norwalk viruses. Vaccine. 2005;23:1851–1858. doi: 10.1016/j.vaccine.2004.11.017. [DOI] [PubMed] [Google Scholar]
- Huy NX, Kim YS, Jun SC, Jin ZW, Park SM, Yang MS, Kim TG. Production of a heat-labile enterotoxin B subunit-porcine epidemic diarrhea virus-neutralizing epitope fusion protein in transgenic lettuce (Lactuca sativa) Biotechnol Bioprocess Eng. 2009;14:731–737. doi: 10.1007/s12257-009-3012-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huy NX, Yang MS, Kim TG. Expression of a cholera toxin B subunit-neutralizing epitope of the porcine epidemic diarrhea virus fusion gene in transgenic lettuce (Lactuca sativa L.) Mol Biotechnol. 2011;48:201–209. doi: 10.1007/s12033-010-9359-1. [DOI] [PubMed] [Google Scholar]
- Jani D, Singh ML, Mohammad RQ, Singh Y, Sharma AK, Tyagi AK. Expression of cholera toxin B subunit in transgenic tomato plants. Transgenic Res. 2002;11:447–454. doi: 10.1023/A:1020336332392. [DOI] [PubMed] [Google Scholar]
- Jiang P, Qin S, Tseng CK. Expression of hepatitis B surface antigen gene (HBsAg) in Laminaria japonica (Laminariales Phaeophyta) Chin Sci Bull. 2002;47:1438–1440. doi: 10.1360/02tb9317. [DOI] [Google Scholar]
- Jiang XL, He ZM, Peng ZQ, Qi Y, Chen Q, Yu SY. Cholera toxin B protein in transgenic tomato fruit induces systemic immune response in mice. Transgenic Res. 2007;16:169–175. doi: 10.1007/s11248-006-9023-5. [DOI] [PubMed] [Google Scholar]
- Jiménez G, Correa I, Melgosa MP, Bullido MJ, Enjuanes L. Critical epitopes in transmissible gastroenteritis virus neutralization. J Virol. 1986;60:131–139. doi: 10.1128/jvi.60.1.131-139.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joensuu JJ, Brown KD, Conley AJ, Clavijo A, Menassa JR, Brandle E. Expression and purification of an anti-foot-and-mouth disease virus single chain variable antibody fragment in tobacco plants. Transgenic Res. 2009;18:685–696. doi: 10.1007/s11248-009-9257-0. [DOI] [PubMed] [Google Scholar]
- Kang TJ, Loc NH, Jang MO, Jang YS, Kim YS, Seo JE, Yang MS. Expression of the B subunit of E. coli heat-labile enterotoxin in the chloroplasts of plants and its characterization. Transgenic Research. 2003;12:683–691. doi: 10.1023/B:TRAG.0000005114.23991.bc. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang TJ, Kang KH, Kim JA, Kwon TH, Jang YS, Yang MS. High-level expression of the neutralizing epitope of porcine epidemic diarrhea virus by a tobacco mosaic virus-based vector. Protein Expr Purif. 2004;38:129–135. doi: 10.1016/j.pep.2004.07.014. [DOI] [PubMed] [Google Scholar]
- Kang TJ, Loc NH, Jang MO, Yang MS. Modification of the cholera toxin B subunit coding sequence to enhance expression in plants. Mol Breed. 2004;13:143–153. doi: 10.1023/B:MOLB.0000018762.27841.7a. [DOI] [Google Scholar]
- Kang TJ, Kim YS, Jang YS, Yang MS. Expression of the synthetic neutralizing epitope gene of porcine epidemic diarrhea virus in tobacco plants without nicotine. Vaccine. 2005;23:2294–2297. doi: 10.1016/j.vaccine.2005.01.027. [DOI] [PubMed] [Google Scholar]
- Kang TJ, Seo JE, Kim DH, Kim TG, Jang YS, Yang MS. Cloning and sequence analysis of the Korean strain of spike gene of porcine epidemic diarrhea virus and expression of its neutralizing epitope in plants. Protein Expr Purif. 2005;41:378–383. doi: 10.1016/j.pep.2005.02.018. [DOI] [PubMed] [Google Scholar]
- Kang TJ, Han SC, Yang MS, Jang YS. Expression of synthetic neutralizing epitope of porcine epidemic diarrhea virus fused with synthetic B subunit of Escherichia coli heat-labile enterotoxin in tobacco plants. Protein Expr Purif. 2006;46:16–22. doi: 10.1016/j.pep.2005.07.026. [DOI] [PubMed] [Google Scholar]
- Kang TJ, Kim BG, Yang JY, Yang MS. Expression of a synthetic cholera toxin B subunit in tobacco using ubiquitin promoter and bar gene as a selectable marker. Mol Biotechnol. 2006;32:93–101. doi: 10.1385/MB:32:2:093. [DOI] [PubMed] [Google Scholar]
- Kang TJ, Lee WS, Choi EG, Kim JW, Kim BG, Yang MS. Mass production of somatic embryos expressing Escherichia coli heat-labile enterotoxin B subunit in Siberian ginseng. J Biotechnol. 2006;121:124–133. doi: 10.1016/j.jbiotec.2005.07.020. [DOI] [PubMed] [Google Scholar]
- Kaper JB, Morris JG, Levine M. Cholera. Clin Microbiol Rev. 1995;8:48–86. doi: 10.1128/cmr.8.1.48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kapusta J, Modelska A, Figlerowicz M. A plant-derived edible vaccine against hepatitis B virus. FASEB J. 1999;13:1796–1799. doi: 10.1096/fasebj.13.13.1796. [DOI] [PubMed] [Google Scholar]
- Karaman S, Cunnick J, Wang K. Analysis of immune response in young and aged mice vaccinated with corn-derived antigen against Escherichia coli heat-labile enterotoxin. Mol Biotechnol. 2006;32:31–42. doi: 10.1385/MB:32:1:031. [DOI] [PubMed] [Google Scholar]
- Karaman S, Cunnick J, Wang K. Expression of the cholera toxin B subunit (CT-B) in maize seeds and a combined mucosal treatment against cholera and traveler's diarrhea. Plant Cell Rep. 2012;31:527–537. doi: 10.1007/s00299-011-1146-3. [DOI] [PubMed] [Google Scholar]
- Karasev AV, Foulke S, Wellens C. Plant based HIV-1 vaccine candidate: Tat protein produced in spinach. Vaccine. 2005;23:1875–1880. doi: 10.1016/j.vaccine.2004.11.021. [DOI] [PubMed] [Google Scholar]
- Kim TG, Galloway DR, Langridge WHR. Synthesis and assembly of anthrax lethal factor-cholera toxin B-subunit fusion protein in transgenic potato. Mol Biotechnol. 2004;28:175–183. doi: 10.1385/MB:28:3:175. [DOI] [PubMed] [Google Scholar]
- Kim TG, Gruber A, Ruprecht RM, Langridge WHR. Synthesis and assembly of SIVmac gag p27 capsid protein cholera toxin B subunit fusion protein in transgenic potato. Mol Biotechnol. 2004;28:33–41. doi: 10.1385/MB:28:1:33. [DOI] [PubMed] [Google Scholar]
- Kim TG, Ruprecht R, Langridge William HR. Synthesis and assembly of a cholera toxin B subunit SHIV 89.6p Tat fusion protein in transgenic potato. Protein Expr Purif. 2004;35:313–319. doi: 10.1016/j.pep.2004.02.007. [DOI] [PubMed] [Google Scholar]
- Kim YS, Kang TJ, Jang YS, Yang MS. Expression of neutralizing epitope of porcine epidemic diarrhea virus in potato plants. Plant Cell Tiss Organ Cult. 2005;82:125–130. doi: 10.1007/s11240-004-7203-7. [DOI] [Google Scholar]
- Kim YS, Kim BG, Kim TG, Kang TJ, Yang MS. Expression of a cholera toxin B subunit in transgenic lettuce (Lactuca sativa L.) using Agrobacterium-mediated transformation system. Plant Cell Tiss Organ Cult. 2006;87:203–210. doi: 10.1007/s11240-006-9156-5. [DOI] [Google Scholar]
- Kim TG, Kim MY, Kim BG, Kang TJ, Kim YS, Jang YS, Arntzen CJ, Yang MS. Synthesis and assembly of Escherichia coli heat-labile enterotoxin B subunit in transgenic lettuce (Lactuca sativa L.) Protein Expr Purif. 2007;51:22–27. doi: 10.1016/j.pep.2006.05.024. [DOI] [PubMed] [Google Scholar]
- Kim YS, Kim MY, Kim TG, Yang MS. Expression and assembly of cholera toxin B subunit (CTB) in transgenic carrot (Daucus carota L.) Mol Biotechnol. 2009;41:8–14. doi: 10.1007/s12033-008-9086-z. [DOI] [PubMed] [Google Scholar]
- Kim TG, Kim MY, Yang MS. Cholera toxin B subunit-domain III of dengue virus envelope glycoprotein E fusion protein production in transgenic plants. Protein Expr Purif. 2010;74:236–241. doi: 10.1016/j.pep.2010.07.013. [DOI] [PubMed] [Google Scholar]
- Kohl T, Hitzeroth II, Stewart D, Varsani A, Govan VA, Christensen ND, Willianmson AL, Rybicki EP. Plant-produced cottontail rabbit papillomavirus L1 protein protects against tumor challenge: a proof-of-concept study. Clin Vaccine Immunol. 2006;13:845–853. doi: 10.1128/CVI.00072-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamphear BJ, Streatfield SJ, Jilka JM, Brooks CA, Barker DK, Turner DD, Delaney DE, Garcia M, Wiggins W, Woodard SL, Hood EE, Tizard IR, Lawhorn B, Howard JA. Delivery of subunit vaccines in maize seed. J Controlled Release. 2002;85:169–180. doi: 10.1016/S0168-3659(02)00282-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langeveld JP, Brennan FR, Martinez-Torrecuadrada JL, Jones TD, Boshuizen RS, Vela C, Casal JI, Kamstrup S, Dalsgaad K, Meloen R, Bendig MM, Hamilton WDO. Inactivated recombinant plant virus protects dogs from a lethal challenge with canine parvovirus. Vaccine. 2001;19:3661–3670. doi: 10.1016/S0264-410X(01)00083-4. [DOI] [PubMed] [Google Scholar]
- Lauterslager TGM, Florack DEA, vander Wal TJ, Molthoff JW, Langeveld JPM, Bosch D, Boersma WJA, Hilgers LAT. Oral immunization of naive and primed animals with transgenic potato tubers expressing LT-B. Vaccine. 2001;19:2749–2755. doi: 10.1016/S0264-410X(00)00513-2. [DOI] [PubMed] [Google Scholar]
- Li L, Xi JF. Advances in immune mechanism of transgenic plant oral vaccine. Foreign Medical Sciences Section of Immunology. 2004;27:354–357. [Google Scholar]
- Li WK, Chen YZ, Zhou Y, Xu H. Regeneration of F. hepatica protective antigenic gene transgenic alfalfa plants by Agrobacterium mediated gene transfer. Journal of Sichuan University (Natural Science Edition) 2003;40:144–147. [Google Scholar]
- Li D, O'Leary J, Huang Y, Huner Norman PA, Jevnikar Anthony M, Ma SW. Expression of cholera toxin B subunit and the B chain of human insulin as a fusion protein in transgenic tobacco plants. Plant Cell Rep. 2006;25:417–424. doi: 10.1007/s00299-005-0069-2. [DOI] [PubMed] [Google Scholar]
- Li HY, Ramalingam S, Chye ML. Accumulation of recombinant SARS-CoV spike protein in plant cytosol and chloroplasts indicatepotential for development of plant-derived oral vaccines. Exp Biol (MedMaywood) 2006;231:1346–1352. doi: 10.1177/153537020623100808. [DOI] [PubMed] [Google Scholar]
- Li YN, Sun M, Liu JX, Yang ZQ, Zhang ZF, Shen GF. High expression of foot-and mouth disease virus structural protein VP1 in tobacco chloroplasts. Plant Cell Rep. 2006;25:329–333. doi: 10.1007/s00299-005-0074-5. [DOI] [PubMed] [Google Scholar]
- Loc NH, Bach NH, Kim TG, Yang MS. Tissue culture and expression of Escherichia coli heat-labile enterotoxin B subunit in transgenic Peperomia pellucida. Protein Expr Purif. 2010;72:82–86. doi: 10.1016/j.pep.2010.02.010. [DOI] [PubMed] [Google Scholar]
- Loc NH, Song NV, Tien NQD, Minh TT, Nga PTQ, Kim TG, Yang MS. Expression of the Escherichia coli heat-labile enterotoxin B subunit in transgenic watercress (Nasturtium officinale L.) Plant Cell Tiss Organ Cult. 2011;105:39–45. doi: 10.1007/s11240-010-9835-0. [DOI] [Google Scholar]
- Loc NH, Thinh LT, Yang MS, Kim TG. Highly expressed cholera toxin B subunit in the fruit of a transgenic tomato (Lycopersicon esculentum L.) Biotechnol Bioprocess Eng. 2011;16:576–580. doi: 10.1007/s12257-010-0195-8. [DOI] [Google Scholar]
- Losey JE, Rayor LS, Carter ME. Transgenic pollen harms monarch larvae. Nature. 1999;399:214. doi: 10.1038/20338. [DOI] [PubMed] [Google Scholar]
- Lou XM, Zhang Z, Yao HQ, Xiong AS, Wang HK, Peng RH, Li X. Expression of human hepatitis B virus large surface antigen gene PRS-S1S2S in transgenic apples. J Fruit Sci. 2005;22:601–605. [Google Scholar]
- Ma SZ, Shi QH, Chen J. Farming transgenic organism safety management and its strategy of China. Environ Prot. 2001;12:10–12. [Google Scholar]
- Ma Y, Lin SQ, Gao Y, Zhang J, Lu LX, Xia NS. Transformation of HBsAg (hepatitis B virus surface antigen) into tomato plants. J Fujian Agric Forest Univ (Nat Sci Ed) 2002;31:223–228. [Google Scholar]
- Martı’nez-Gonza’lez L, Rosales-Mendoza S, Soria-Guerra RE, Moreno-Fierros L, Lo'pez-Revilla R, Korban SS, Guevara-Arauza JC, Alpuche-Soli's AG. Oral immunization with a lettuce-derived Escherichia coli heat-labile toxin B subunit induces neutralizing antibodies in mice. Plant Cell Tiss Organ Cult. 2011;107:441–449. doi: 10.1007/s11240-011-9994-7. [DOI] [Google Scholar]
- Mason HS, Lam DMK, Arntzen CJ. Expression of hepatitis B surface antigen in transgenic plants. Proc Natl Acad Sci USA. 1992;89:11745–11749. doi: 10.1073/pnas.89.24.11745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mason HS, Ball JM, Shi JJ, Jiang X, Estes MK, Arnten CJ. Expression of Norwalk virus capsid protein in transgenic tobacco and potato and its oral immunogenicity in mice. Proc Natl Acad Sci USA. 1996;93:5335–5340. doi: 10.1073/pnas.93.11.5335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mason HS, Haq TA, Clements JD, Arntzen CJ. Edible vaccine protects mice against Escherichia coli heat-labile enterotoxin (LT): potatoes expressing a synthetic LT-B gene. Vaccine. 1998;16:1336–1343. doi: 10.1016/S0264-410X(98)80020-0. [DOI] [PubMed] [Google Scholar]
- Matsumura T, Itchoda N, Tsunemitsu H. Production of immunogenic VP6 protein of bovine group A rotavirus in transgenic potato plants. Arch Virol. 2002;147:1263–1270. doi: 10.1007/s00705-002-0808-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGarvey PB, Hammond J, Dienelt MM, Hooper DC, Fu ZF, Dietzschold B, Koprowski H, Michaels FH. Expression of the rabies virus glycoprotein in transgenic tomatoes. Nature. 1995;13:1484–1487. doi: 10.1038/nbt1295-1484. [DOI] [PubMed] [Google Scholar]
- McGhee JR, Mestecky J, Dertzbaugh MT, Eldridge JH, Hirasawa M, Kiyono H. The mucosal immune system: from fundamental concepts to vaccine development. Vaccine. 1992;10:75–88. doi: 10.1016/0264-410X(92)90021-B. [DOI] [PubMed] [Google Scholar]
- Meleon RH, Rowlands DJ, Brown F. Comparison of the antibodies elicited by the individual structural polypeptides of foot-and-mouth disease and polioviruses. J Gen Virol. 1979;67:289–294. doi: 10.1099/0022-1317-67-2-289. [DOI] [PubMed] [Google Scholar]
- Meslin FX, Fishbein DB, Matter HC. Rationale and prospects for rabies elimination in developing countries. In: Rupprecht CE, Dietzschold B, Koprowski H, editors. Lyssaviruses. Berlin: Springer; 1994. pp. 1–26. [DOI] [PubMed] [Google Scholar]
- Michel ML. Towards immunotherapy for chronic hepatitis B virus infections. Vaccine. 2002;20:83–88. doi: 10.1016/S0264-410X(02)00393-6. [DOI] [PubMed] [Google Scholar]
- Munro S, Pelham HR. A C-terminal signal prevents secretion of luminal ER proteins. Cell. 1987;48:899–907. doi: 10.1016/0092-8674(87)90086-9. [DOI] [PubMed] [Google Scholar]
- Napier JA, Richard G, Shewry PR. Trafficking and stability of heterologous proteins in transgenic plants. Meth Biotechnol. 1998;3:189–202. doi: 10.1007/978-1-60327-260-5_15. [DOI] [Google Scholar]
- Nordlee J, Taylor S, Townsend J. Identification of a Brazil nut allergen in transgenic soybeans. New England J Medicine. 1996;334:688–692. doi: 10.1056/NEJM199603143341103. [DOI] [PubMed] [Google Scholar]
- O'Brien GJ, Bryant CJ, Voogd C, Greenberg HB, Gardner RC, Bellamy AR. Rotavirus VP6 expressed by PVX vectors in Nicotiana benthamiana coats PVX rods and also assembles into virus like particles. Virology. 2000;270:444–453. doi: 10.1006/viro.2000.0314. [DOI] [PubMed] [Google Scholar]
- Orson J (2001) Gene stacking in herbicide to lerant oilseed rape: lessons from the North American experience. English Nature Research Report, No. 443. English Nature: Peterborough.
- Oszvald M, Kang TJ, Tomoskozi S, Tamas C, Tamas L, Kim TG, Yang MS. Expression of a synthetic neutralizing epitope of porcine epidemic diarrhea virus fused with synthetic B subunit of Escherichia coli heat labile enterotoxin in rice endosperm. Mol Biotechnol. 2007;5:215–222. doi: 10.1007/BF02686007. [DOI] [PubMed] [Google Scholar]
- Oszvald M, Kang TJ, Tomoskozi S, Jenes B, Kim TG, Cha YS, Tamas L, Yang MS. Expression of cholera toxin B subunit in transgenic rice endosperm. Mol Biotechnol. 2008;40:261–226. doi: 10.1007/s12033-008-9083-2. [DOI] [PubMed] [Google Scholar]
- Pan L, Zhang Y, Wang Y, Wang B, Wang Y, Fang Y, Jiang S, Lv J, Wang W, Sun Y, Xie Q. Foliar extracts from transgenic tomato plants expressing the structural polyprotein, P1–2A, and protease, 3C, from foot-and- mouth disease virus elicit a protective response in guinea pigs. Vet Immunol Immunopathol. 2008;121:83–90. doi: 10.1016/j.vetimm.2007.08.010. [DOI] [PubMed] [Google Scholar]
- Pearay LO, Howard F, Robert CW. Vaccination strategies for mucosal immune responses. Clin Microbiol Rev. 2001;14:430–445. doi: 10.1128/CMR.14.2.430-445.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pensaert MB, de Bouck P. A new coronavirus-like particle associated with diarrhea in swine. Arch Virol. 1978;58:243–247. doi: 10.1007/BF01317606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perea Arango I, Loza Rubio E, Rojas Anaya E, Olivera Flores T, de la Vara LG, Gómez Lim MA (2008) Expression of the rabies virus nucleoprotein in plants at high-levels and evaluation of immune responses in mice. Plant Cell Rep 27:677–685 [DOI] [PubMed]
- Perlak FJ, Fuchs RL, Dean DA, McPherson SL, Fischhoff DA. Modification of the coding sequence enhances plant expression of insect control proteins. Proc Nat Acad Sci USA. 1991;88:3324–3328. doi: 10.1073/pnas.88.8.3324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polkinghorne I, Hamerli D, Cowan P, Duckworth J. Plant-based immunocontraceptive control of wildlife—“potentials, limitations, and possums”. Vaccine. 2005;23:1847–1850. doi: 10.1016/j.vaccine.2004.11.016. [DOI] [PubMed] [Google Scholar]
- Richter LJ, Thanavala Y, Arntzen GJ, Mason HS. Production of hepatitis B surface antigen in transgenic plants for oral immunization. Nat Biotechnol. 2000;18:1167–1171. doi: 10.1038/81153. [DOI] [PubMed] [Google Scholar]
- Rigano M, Walmsley AM. Expression systems and developments in plant-made vaccines. Immunol Cell Biol. 2005;83:271–277. doi: 10.1111/j.1440-1711.2005.01336.x. [DOI] [PubMed] [Google Scholar]
- Rigano MM, Dreitz S, Kipnis AP, Izzo AA, Walmsley AM. Oral immunogenicity of a plant-made subunit, tuberculosis vaccine. Vaccine. 2006;24:691–695. doi: 10.1016/j.vaccine.2005.08.009. [DOI] [PubMed] [Google Scholar]
- Rosales-Mendoza S, Soria-Guerra RE, Olivera-Flores MT, López-Revilla R, Argüello-Astorga GR, Jiménez-Bremont JF, García-de la Cruz RF, Loyola-Rodríguez JP, Alpuche-Solís AG (2007a) Expression of Escherichia coli heat-labile enterotoxin b subunit (LTB) in carrot (Daucus carota L.). Plant Cell Rep 26:969–976 [DOI] [PubMed]
- Rosales-Mendoza S, Alpuche-Solís AG, Soria-Guerra RE, Olivera-Flores MT, López-Revilla R, Argüello-Astorga GR, Jiménez-Bremont JF, García-de la Cruz RF, Loyola-Rodríguez JP. Expression of an Escherichia coli antigenic fusion protein comprising the heat labile toxin B subunit and the heat stable toxin, and its assembly as a functional oligomer in transplastomic tobacco plants. The Plant Journal. 2007b;57:45–54. doi: 10.1111/j.1365-313X.2008.03666.x. [DOI] [PubMed] [Google Scholar]
- Rosales-Mendoza S, Soria-Guerra RE, López-Revilla R, Moreno-Fierros L, Alpuche-Solís AG. Ingestion of transgenic carrots expressing the Escherichia coli heat-labile enterotoxin B subunit protects mice against cholera toxin challenge. Plant Cell Rep. 2008;27:79–84. doi: 10.1007/s00299-007-0439-z. [DOI] [PubMed] [Google Scholar]
- Roy S, Tyagi A, Tiwari S, Singh A, Sawant SV, Singh PK, Tuli R. Rabies glycoprotein fused with B subunit of cholera toxin expressed in tobacco plants folds into biologically active pentameric protein. Protein Expr Purif. 2010;70:184–190. doi: 10.1016/j.pep.2009.10.002. [DOI] [PubMed] [Google Scholar]
- Sacco AG. Circulating antibodies: contraceptive with no side effects. J Reprod Fert. 1979;56:533. doi: 10.1530/jrf.0.0560533. [DOI] [Google Scholar]
- Sánchez CM, Jiménez G, Laviada MD, Correa I, Suñé C, Bullido MJ, Gebauer F, Smerdou C, Callebau P, Escribano JM, Enjuanes L. Antigenic homology among coronaviruses related to transmissible gastroenteritis virus. Virology. 1990;174:410–417. doi: 10.1016/0042-6822(90)90094-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma MK, Jani D, Thungapathra M, Gautam JK, Meena LS, Singh Y, Ghosh A, Tyagi AK, Sharma AK. Expression of accessory colonization factor subunit A (ACFA) of Vibrio cholerae and ACFA fused to cholera toxin B subunit in transgenic tomato (Solanum lycopersicum) J Biotechnol. 2008;135:22–27. doi: 10.1016/j.jbiotec.2008.03.002. [DOI] [PubMed] [Google Scholar]
- Sharma MK, Singh NK, Jani D, Sisodia R, Thungapathra M, Gautam JK, Meena LS, Singh Y, Ghosh A, Tyagi AK, Sharma AK. Expression of toxin co-regulated pilus subunit A (TCPA) of Vibrio cholerae and its immunogenic epitopes fused to cholera toxin B subunit in transgenic tomato (Solanum lycopersicum) Plant Cell Rep. 2008;27:307–318. doi: 10.1007/s00299-007-0464-y. [DOI] [PubMed] [Google Scholar]
- Shulga NY, Rukavtsova EB, Krymsky MA, Borisova VN, Melnikov VA, Bykov VA. Expression and characterization of hepatitis B surface antigen in transgenic potato plants. Biochemistry (Mosc) 2004;69:1158–1164. doi: 10.1023/B:BIRY.0000046891.46282.c8. [DOI] [PubMed] [Google Scholar]
- Sim JS, Pak HK, Kim DS, Lee SB, Kim YH, Hahn BS. Expression and characterization of synthetic heat-labile enterotoxin B subunit and hemagglutinin–neuraminidase-neutralizing epitope fusion protein in Escherichia coli and tobacco chloroplasts. Plant Mol Biol Rep. 2009;27:388–399. doi: 10.1007/s11105-009-0114-3. [DOI] [Google Scholar]
- Smith ML, Mason HS, Shuler ML. Hepatitis surface (HBsAg) expression in plant cell culture: kinetics of antigen accumulation in batch culture and its intracellular form. Biotechnol Bioeng. 2002;80:812–822. doi: 10.1002/bit.10444. [DOI] [PubMed] [Google Scholar]
- Stoger E, Vaquero C, Torres E, Sack M, Nicholson L, Drossard J. Cereal crops as viable production and storage systems for pharmaceutical scFv antibodies. Plant Mol Biol. 2000;42:583–590. doi: 10.1023/A:1006301519427. [DOI] [PubMed] [Google Scholar]
- Streatfield SJ, Jilka JM, Hood EE, Turner DD, Bailey MR, Mayor JM, Woodard SL, Beifuss K, Horn ME, Delaney DE, Tizard IR, Howard JA. Plant-based vaccines: unique advantages. Vaccine. 2001;19:2742–2748. doi: 10.1016/S0264-410X(00)00512-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Streatfield SJ, Mayor JM, Barker DK, Brooks C, Lamphear BJ, Woodard SL, Beifuss KK, Vicuna DV, Massey LA, Horn ME, Delaney DD, Nikolov ZL, Hood EE, Jilka JM, Howard JA. Development of an edible subunit vaccine in corn against enterotoxigenic strains of Escherichia coli. In Vitro Cell Dev Biol Plant. 2002;38:11–17. doi: 10.1079/IVP2001247. [DOI] [Google Scholar]
- Streatfield SJ, Lane JR, Brooks CA, Barker DK, Poage ML, Mayor JM, Lamphear BJ, Drees CF, Jilka JM, Hood EE, Howard JA. Corn as a production system for human and animal vaccines. Vaccine. 2003;21:812–815. doi: 10.1016/S0264-410X(02)00605-9. [DOI] [PubMed] [Google Scholar]
- Sunil-Kumar GB, Ganapathi TR, Revathi CJ, Srinivas L, Bapat VA. Expression of hepatitis B surface antigen in transgenic banana plants. Planta. 2005;222:484–493. doi: 10.1007/s00425-005-1556-y. [DOI] [PubMed] [Google Scholar]
- Tacket C, Mason HS, Losonsky G, Clements JD, Levine MM, Arntzen CJ. Immunogenicity in humans of a recombinant bacterial antigen delivered in a transgenic potato. Nat Med. 1998;4:607–609. doi: 10.1038/nm0598-607. [DOI] [PubMed] [Google Scholar]
- Tacket CO, Pasetti MF, Edelman R, Howard JA, Streatfield S. Immunogenicity of recombinant LT-B delivered orally to humans in transgenic corn. Vaccine. 2004;22:4385–4389. doi: 10.1016/j.vaccine.2004.01.073. [DOI] [PubMed] [Google Scholar]
- Thanavala Y, Yang YF, Lyons P. Immunogenicity of transgenic plant-derived hepatitis B surface antigen. Proc Natl Acad Sci USA. 1995;92:3358–3361. doi: 10.1073/pnas.92.8.3358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tollis M, Dietzschold B, Viola CB, Koprowski H. Immunization of monkeys with rabies ribonucleoprotein (RNP) confers protective immunity against rabies. Vaccine. 1991;9:134–136. doi: 10.1016/0264-410X(91)90270-G. [DOI] [PubMed] [Google Scholar]
- Tomas M, Monica AS, Eliot MH, Terry WT. Production of Escherichia coli heat labile toxin (LT) B subunit in soybean seed and analysis of its immunogenicity as an oral vaccine. Vaccine. 2007;25:1647–1657. doi: 10.1016/j.vaccine.2006.11.010. [DOI] [PubMed] [Google Scholar]
- Tuboly T, Yub W, Bailey A, Degrandisc S, Dub S, Ericksonb L, Nagy E. Immunogenicity of porcine transmissible gastroenteritis virus spike protein expressed in plants. Vaccine. 2000;18:2023–2028. doi: 10.1016/S0264-410X(99)00525-3. [DOI] [PubMed] [Google Scholar]
- Verma D, Daniell H. Chloroplast vector systems for biotechnology applications. Plant Physiol. 2007;145:1129–1143. doi: 10.1104/pp.107.106690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wagner B, Hufnagl K, Radauer C, Wagner S, Baier K, Scheiner O, Wiedermann U, Breiteneder H. Expression of the B subunit of the heat-labile enterotoxin of Escherichia coli in tobacco mosaic virus-infected Nicotiana benthamiana plants and its characterization as mucosal immunogen and adjuvant. J Immunol Methods. 2004;287:203–215. doi: 10.1016/j.jim.2004.02.001. [DOI] [PubMed] [Google Scholar]
- Wandelt CI, Khan MRI, Craig S, Schroeder HE, Spencer D, Higgens TJV. Vicilin with carboxy-terminal KDEL is retained in the endoplasmic reticulum and accumulates to high levels in the leaves of transgenic plants. The Plant Journal. 1992;2:181–192. doi: 10.1046/j.1365-313x.1992.t01-41-00999.x. [DOI] [PubMed] [Google Scholar]
- Wang YQ, Li T. Transformation of HBsAg gene into tomato and production of transgenic tomato plants. J Southwest Univ (Nat Sci Ed) 2008;30:78–83. [Google Scholar]
- Wang XG, Zhang GH, Liu CX, Zhang YH, Xiao CZ, Fang RX. Purified cholera toxin B subunit from transgenic tobacco plants possesses authentic antigenicity. Biotechnol Bioeng. 2001;72:490–494. doi: 10.1002/1097-0290(20010220)72:4<490::AID-BIT1011>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- Wang DM, Zhu JB, Peng M, Zhou P. Induction of a protective antibody response to FMDV in mice following oral immunization with transgenic Stylosanthes spp. as a feedstuff additive. Transgenic Res. 2008;17:1163–1170. doi: 10.1007/s11248-008-9188-1. [DOI] [PubMed] [Google Scholar]
- Wang L, Webster DE, Campbell AE, Dry IB, Wesselingh SL, Coppel RL. Immunogenicity of Plasmodium yoelii merozoite surface protein 4/5 produced in transgenic plants. Int J Parasitol. 2008;38:103–110. doi: 10.1016/j.ijpara.2007.06.005. [DOI] [PubMed] [Google Scholar]
- Wigdorovitz A, Carrillo C, Dus Santos MJ, Trono K, Peralta A, Gómez MC, Ríos RD, Franzone PM, Sadir AM, Escribano JM, Borca MV. Induction of a protective antibody response to foot and mouth disease virus in mice following oral or parenteral immunization with alfalfa transgenic plants expressing the viral structural protein VP1. Virology. 1999;255:347–353. doi: 10.1006/viro.1998.9590. [DOI] [PubMed] [Google Scholar]
- World Health Organization. Laboratory biosafety manual. 3. Geneva: World Health Organization; 2004. [Google Scholar]
- Wu YQ, Xiong XM. Expression of heavy-china variable domain of mAb against Encephalitis Type B virus in transgenic tobacco. Chinese Science Bulletin. 1997;42:1038–1041. [Google Scholar]
- Wu LG, Jiang LB, Zhou ZA, Fan JH, Zhang QQ, Zhu HH, Han Q, Xu ZK. Expression of foot-and-mouth disease virus epitopes in tobacco by a tobacco mosaic virus-based vector. Vaccine. 2003;21:4390–4398. doi: 10.1016/S0264-410X(03)00428-6. [DOI] [PubMed] [Google Scholar]
- Yang CJ, Song L, Wang Z. The effects of transgenic soybean on biological diversity. Environ Prot. 2002;11:24–27. [Google Scholar]
- Yang KS, Kim HS, Lim S, Kwon S, Kwak S, Lee HS. Transgenic sweet potato (Ipomoea batatas) expressing spike gene of porcine epidemic diarrhea virus. Korean Journal of Plant Biotechnology. 2005;32:263–268. doi: 10.5010/JPB.2005.32.4.263. [DOI] [Google Scholar]
- Yang CD, Liao JT, Lai CY, Jong MH, Liang CM, Lin YL, Lin AS, Hsu YH, Liang SM. Induction of protective immunity in swine by recombinant bamboo mosaic virus expressing foot-and-mouth disease virus epitopes. BMC Biotechnol. 2007;7:62. doi: 10.1186/1472-6750-7-62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu J, Langridge WHR. A plant-based multicomponent vaccine protects mice from enteric diseases. Nat Biotechnol. 2001;19:548–552. doi: 10.1038/89297. [DOI] [PubMed] [Google Scholar]
- Yu J, Langridge W. Expression of rotavirus capsid protein VP6 in transgenic potato and its oral immuno- genicity in mice. Transgenic Research. 2003;12:163–169. doi: 10.1023/A:1022912130286. [DOI] [PubMed] [Google Scholar]
- Yuan X, Huang FS, Qiu Y, Hao ZY, Chen EM. Expression of Schistosoma japonicum ferritin in Brassia napus. Chinese Journal of Veterinary Science. 2008;28:926–929. [Google Scholar]
- Yusibov V, Modelska A, Steplewski K, Agadjanyan M, Weiner D, Hooper DC, Koprowski H. Antigens produced in plants by infection with chimeric plant viruses immunize against rabies virus and HIV-1. Proc Natl Acad Sci USA. 1997;94:5784–5788. doi: 10.1073/pnas.94.11.5784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yusibov V, Hooper DC, Spitsin SV, Fleysh N, Kean RB, Mikheeva T, Deka D, Karasev A, Cox S, Randall J, Koprowski H. Expression in plants and immunogenicity of plant virus-based experimental rabies vaccine. Vaccine. 2002;20:3155–3164. doi: 10.1016/S0264-410X(02)00260-8. [DOI] [PubMed] [Google Scholar]
- Zhang R, Wei YH, Liu K. Construction on plant expression vector with hepatitis B surface antigen gene. J Northwest Univ (Sci Online) 2005;3:1–6. [Google Scholar]
- Zhang X, Buehner NA, Hutson AM. Tomato is a highly effective vehicle for expression and oral immunization with Norwalk virus capsid protein. Plant Biotechnol J. 2006;4:419–432. doi: 10.1111/j.1467-7652.2006.00191.x. [DOI] [PubMed] [Google Scholar]
- Zhang XX, Yuan ZG, Guo XJ, Li JW, Li ZN, Wang QY. Expression of Chlamydophila psittaci MOMP heat-labile toxin B subunit fusion gene in transgenic rice. Biologicals. 2008;36:296–302. doi: 10.1016/j.biologicals.2008.04.002. [DOI] [PubMed] [Google Scholar]
- Zhao XY, Zhang XD, Zhang T, Yan HP, Luo CX, Yang FP. Cloning and expression of the hepatitis B surface antigen in carrots. Prog Microbiol Immunol. 2002;30:1–4. [Google Scholar]
- Zhou XH, Chen XG, Lu LQ, Li L, Wu Y, Xian P, Yan H, Wu K. Construction of the plant expression vector for the truncated fragments of SAG1 gene of Toxoplasma gondii. Chinese Journal of Zoonoses. 2004;20:662–665. [Google Scholar]
- Zhu S, Zhang SL. Advances of transgenic plant vaccine. Chinese Agricultural Science Bulletin. 2006;22:59–63. [Google Scholar]
- Zhu JG, Wei YH, Guo ZG. The transformation and expression of HBsAg gene in Arachis hypogaea. Chin Bull Bot. 2006;23L:665–669. [Google Scholar]
- Zhuang Y, Cao Y. Risks of transgenic organisms. Journal of Jilin university of agriculture sicence and technology. 2008;17:14–15. [Google Scholar]