

RÉSUMÉ
Le virus Ébola cause régulièrement depuis 1976 des petites épidémies meurtrières généralement maitrisées en quelques mois. Alors que seule l’Afrique Centrale en avait été victime jusqu’alors, une épidémie à virus Ébola d’une ampleur extraordinaire embrase dramatiquement plusieurs pays d’Afrique de l’Ouest depuis le mois de décembre 2013 principalement en raison des défaillances majeures dans la mise en œuvre des mesures visant à empêcher les transmissions interhumaines du virus. Après une période d’incubation d’environ une semaine, la maladie se manifeste par l’apparition soudaine d’une forte fièvre aboutissant in fine à des hémorragies multiples puis à la défaillance généralisée des organes. Plusieurs espèces de chauves-souris seraient les principaux réservoirs du virus Ébola. La contamination de l’homme se produirait soit directement auprès des chauves-souris, largement consommées par les populations locales, soit par l’intermédiaire d’espèces animales sensibles au virus, telles que les chimpanzés et les gorilles. À côté de ce « cycle naturel », l’hypothèse d’un « cycle épidémique » impliquant des espèces animales domestiques vivant dans les villages tels que les chiens ou les porcs, tend désormais à être sérieusement avancée. Ainsi, en fonction des animaux impliqués et de la forme clinique des infections développées, les modalités de la contamination de l’homme peuvent être multiples et sont donc encore largement méconnues. Dans un tel contexte, tous les efforts qui pourront être déployés pour percer le mystère de l’émergence du virus Ébola chez l’homme et clarifier les modalités de la transmission du virus, permettront peut-être de prédire voire d’anticiper l’apparition des épidémies. L’objectif de cette revue est de dresser un état des lieux exhaustif de l’écologie du virus Ébola et de mettre en lumière les évènements qui gouvernent la transmission du virus à l’homme tout en précisant les points encore nombreux qui demeurent non élucidés.
Mots-clés: Fièvre hémorragique à virus ébola, Réservoirs d’agents pathogènes, Chiroptera, Apoptose
Abstract
Since 1976 Ebola virus regularly has caused small deadly outbreaks in Central Africa, usually controlled in a few months. For the first time, an Ebola epidemic of exceptional magnitude dramatically engulfed several countries in West Africa since December 2013. Major failures of implementing measures to prevent human-to-human transmissions are the main cause of this large-scale Ebola outbreak. After about one-week incubation period, the Ebola virus disease is characterized by a sudden onset of high fever leading to multiple hemorrhages and to widespread organ failure.
Several bat species constitute the main reservoirs of Ebola viruses. Human contamination would occur either directly from bats, widely consumed by the local populations, or through animal species susceptible to Ebola infection, such as chimpanzees and gorillas. Alongside this “natural cycle”, an “epidemic cycle” involving domestic animals living in villages such as dogs or pigs, is seriously suggested. Thus, according to the diversity of concerned animals and their clinical infection form, modalities of human contamination can be multiple and are still largely unknown. In this context, all efforts that could be made to unravel the mystery of the Ebola virus emergence in humans and clarify modalities of the virus transmission, would allow for predicting or for anticipating the future occurrence of epidemics. This review aims to provide an exhaustive inventory of the Ebola ecology to highlight events governing the virus transmission to humans that still remain unsolved.
Key-words: Hemorrhagic fever, ebola; Disease reservoirs; Chiroptera; Apoptosis
INTRODUCTION
Une terrible et dramatique épidémie à virus Ébola (EBOV), d’une ampleur extraordinaire, sévit pour la première fois en Afrique de l’Ouest depuis le mois de décembre 2013 [1], [2]. Les analyses phylogénétiques ont identifié une souche virale similaire à celles responsables des épidémies qui ont sévi en Afrique Centrale depuis 1976, date de la découverte du virus Ébola [2], [3]. Apparue d’abord en Guinée Conakry, elle s’est propagée ensuite au Liberia, en Sierra Leone, puis au Nigeria. Alors que l’épidémie continue de se propager sans baisser de rythme et que de nombreux cas apparaissent chaque jour dans les pays touchés, l’OMS recense au 25/10/2014 un total de 12 164 cas dont 4439 décès, soit près de quatre fois plus que le nombre total de cas enregistrés au cours de toutes les épidémies survenues depuis 1976. Le nombre impressionnant de cas et de décès, qui est malheureusement loin d’être définitif, a contraint l’OMS à déclarer au mois d’août 2014 que la flambée épidémique à virus Ébola en Afrique de l’Ouest représentait « une urgence de santé publique de portée internationale et un risque de santé publique majeur pour les autres états » [4]. Et la situation ne fait qu’empirer. L’OMS a annoncé au mois d’octobre 2014 que cette épidémie représente désormais la plus grave crise sanitaire planétaire des 30 dernières années. La nouvelle dimension atteinte par cette épidémie d’Ébola bouleverse tous les paradigmes [5]. Minimisée et négligée par certains pendant près de 30 ans, principalement parce qu’elle ne faisait « pas assez de victimes », qu’elle ne touchait que des petits villages inconnus et perdus au cœur des forêts tropicales africaines et que son impact en santé publique était jugé ridicule comparé à d’autres maladies infectieuses comme le SIDA, le paludisme ou le choléra, la terrifiante épidémie qui sévit actuellement en Afrique de l’Ouest va indéniablement modifier les approches scientifique, sanitaire et politique de cette maladie voire des maladies infectieuses émergentes de manière générale. En effet, une crise sanitaire d’origine infectieuse doit certes être combattue dans l’urgence mais elle doit également être pensée et prédite sur la base de signes avant-coureurs réels comme c’est le cas du virus Ébola grâce aux enseignements qui auraient pu être déduits des épidémies précédentes. Car même si un processus d’émergence d’un virus est long et progressif, il peut déraper et s’emballer à tout moment, comme ce fut le cas du VIH SIDA et comme c’est le cas aujourd’hui du virus Ébola.
Cette revue a pour objectif de porter un éclairage sur le phénomène « Ébola » et de faire une synthèse des évènements qui ont conduit à la crise sanitaire actuelle depuis la première identification du virus en 1976. Cette revue dressera un état des lieux exhaustif de l’écologie du virus Ébola et insistera sur les différentes étapes et modalités de la circulation du virus qui l’a conduit de son hôte naturel, le réservoir animal, à l’Homme.
QUELQUES INFORMATIONS PRÉALABLES INDISPENSABLES
Le virus Ébola appartient à la famille virale des Filoviridae qui, avec les Rhabdoviridae (contenant par exemple le virus de la Rage), les Paramyxoviridae (contenant par exemple les virus de la Rougeole et des Oreillons) et les Bornaviridae, forment l’ordre des Mononegavirales, un groupe de virus possédant pour génome un seul brin d’ARN, linéaire, non segmenté et de polarité négative [6]. Les particules des virus appartenant à la famille des Filoviridae se présentent sous la forme caractéristique de filament, unique dans le monde de la virologie, d’environ 80 nm de diamètre et d’une longueur variant de quelques dizaines de nanomètres à 10–15 μm (Figure 1 ). D’autres formes peuvent être également parfois rencontrées principalement en culture cellulaire (Figure 1). Le génome comprend environ 19 000 nucléotides transcrits en sept ARN messagers codant sept protéines structurales (3′ leader, nucléoprotéine (NP), protéine virale (VP) 35, VP40, glycoprotéine (GP), VP30, VP24, et l’ARN polymérase (L) — 5′ trailer). La partie centrale du virion est occupée par un complexe ribonucléoprotéique (RNP) qui est constitué du brin d’ARN entouré par la NP elle-même liée aux protéines de matrice VP30 et VP35 et à l’ARN polymérase. L’enveloppe du virus est exclusivement constituée de la GP organisée en trimères.
Fig. 1.

Le virus Ébola. Morphologie et structure des particules virales.
La famille des Filoviridae contient trois genres distincts, Ebolavirus et Marburgvirus, tous deux associés à des épidémies meurtrières de fièvre hémorragique, et Cuevavirus, tout récemment identifié mais jamais détecté chez l’homme [7], [8]. Le genre Ebolavirus est lui-même subdivisé en cinq espèces virales, ne contenant chacune qu’un seul virus, associées chacune à un taux de létalité et à une localisation géographie plus ou moins spécifiques: L’espèce Zaïre ebolavirus, contenant le virus Ébola, sévit essentiellement en Afrique Centrale, désormais également en Afrique de l’Ouest, et est associé à un taux de mortalité d’environ 80%; L’espèce Sudan ebolavirus, contenant le virus Soudan, sévit dans les régions australes de l’Afrique et est associé à un taux de mortalité d’environ 50% [9], [10], [11]; L’espèce Bundigunyo ebolavirus, contenant le virus Bundigunyo, a été responsable de deux épidémies en Ouganda et en RDC, et est caractérisé par un taux de mortalité de 25% [12]; L’espèce Taï Forest ebolavirus, contenant le virus Taï Forest, n’a fait qu’un seul cas en 1995 en Côte-d’Ivoire [13], [14]; enfin l’espèce Reston ebolavirus, qui n’existe qu’en Asie, contenant le virus Reston, pathogène pour le singe macaque et le porc, pour lequel aucun cas n’a été détecté chez l’homme [15], [16].
En conséquence, malgré la légère confusion taxonomique entre les noms des virus et ceux des espèces virales, le virus Ébola, dont il sera question tout au long de ce document, ne correspond qu’à l’espèce Zaïre ebolavirus.
LA DÉCOUVERTE DU VIRUS ÉBOLA ET SES MANIFESTATIONS ULTÉRIEURES
Le virus Ébola a été identifié pour la première fois le 1 septembre 1976 dans le nord-est du Zaïre, désormais dénommé République Démocratique du Congo (RDC), lorsqu’il a été isolé chez plusieurs malades d’une épidémie qui fit 318 cas dont 284 morts [17]. La dénomination d’Ébola fut attribuée en référence à la rivière du même nom qui coule près du village Yambuku, épicentre de cette épidémie. Après un silence épidémiologique d’une vingtaine d’années au cours de laquelle aucun cas ne fut répertorié, le virus Ébola réapparut et fut responsable de quatre épidémies successives entre 1995 et 1997 (Figure 2 ). Cependant, malgré ces signaux, aussi forts que prédictifs, le feu médiatique et scientifique s’éteignit aussi rapidement qu’il ne s’était allumé. Le virus Ébola fit d’abord 256 victimes parmi 315 malades dans la ville de Kikwit et ses environs dans le sud-ouest de la RDC en 1995 [18]. Parallèlement, il tua quelques dizaines d’orpailleurs dans la zone de Mékouka au nord-est du Gabon où il récidiva quelques mois plus tard et un peu plus à l’Est sous la forme de deux épidémies rapprochées: la première au début de 1996 dans le village de Mayibout pour laquelle la majorité des 37 patients dont 21 décès étaient des enfants qui avaient participé au dépeçage d’un chimpanzé trouvé mort dans la forêt; la deuxième plus vers le sud dans la région de Booué entre octobre 1996 et mars 1997 [19]. Cette dernière épidémie, qui compta 60 cas dont 45 décès, généra pour la première fois un foyer secondaire de 15 cas dont 11 décès dans la capitale Libreville, et un cas en Afrique du sud, une infirmière contaminée par un patient gabonais en provenance de Libreville.
Fig. 2.
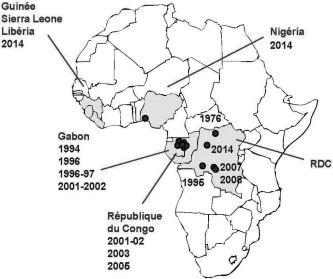
Inventaire et localisation des épidémies à virus Ébola.
Après de nouvelles années de répit, le virus Ébola surgit de nouveau en 2001. Tout d’abord, il causa quatre épidémies dans une seule petite région frontalière du Gabon et du Congo qui aboutirent à 312 victimes dont 263 décès entre 2001 et 2005 [20], [21], [22], [23]. Àcette même période,levirus Ébola contamina également les gorilles et les chimpanzés et provoqua des épidémies mortelles qui décimèrent environ 80% des populations qui peuplaient la région frontalière du nord-est du Gabon et du nord-ouest du Congo [21], [24], [25] ! Le virus Ébola tua de nouveau 187 personnes sur 264 en 2007 puis 15 parmi 32 autres personnes en 2008 dans une zone située autour du village de Luebo dans la province du Kasaï occidental en RDC [26]. Enfin, tout récemment le virus Ébola causa une nouvelle épidémie en RDC, heureusement rapidement contrôlée, qui aboutit à 70 cas dont 42 décès entre juillet et octobre 2014 [27].
Enfin, le virus Ébola surgit dans une partie du continent africain encore indemne, en Afrique de l’Ouest, et réussit à générer un chaos sanitaire caractérisé par 17 942 cas dont 6388 décès au 10 décembre 2014 (Figure 2).
À LA SOURCE DU VIRUS EBOLA, LE RÉSERVOIR ANIMAL
Essayons maintenant de nous pencher sur la source du virus Ébola, scientifiquement appelé réservoir. Comme tous les virus, Ébola ne peut pas se développer seul, il doit nécessairement utiliser la machinerie cellulaire d’un organisme, végétal ou animal, pour pouvoir se multiplier. N’étant pas adapté à l’homme, il est hébergé à l’intérieur d’un autre être vivant, longtemps resté inconnu et qui, encore aujourd’hui, n’est pas définitivement identifié. L’homme n’est en effet qu’un hôte accidentel qui se contamine lorsqu’il est en contact avec l’animal réservoir infecté. Depuis la première épidémie en 1976, les scientifiques ont tenté, dans plusieurs pays d’Afrique Centrale, d’identifier cet animal ou végétal mystérieux capable de vivre harmonieusement avec Ébola; il était même devenu un véritable fantasme pour l’homme.
Entre 1976 et 1997, de grandes quantités d’animaux, généralement capturés autour des foyers épidémiques, ont été sacrifiés pour cette fameuse quête. Au total, près de 7000 animaux vertébrés appartenant à de nombreuses espèces (singes, rongeurs sauvages, écureuils, chauves-souris, antilopes, porcs épics, potamochères, cochons domestiques, oiseaux, amphibiens, reptiles ...) et invertébrés (moustiques, punaises, puces, tiques, poux ...) ont été capturés près des habitations des malades lors des épidémies. Après de multiples études menées en RDC, au Soudan, en République centrafricaine, au Gabon et en Côte-d’Ivoire, les tentatives d’isolement et de détection d’anticorps spécifiques se révélèrent toutes infructueuses [17], [28], [29], [30], [31], [32]. À partir de 1998, les techniques de détection se sont considérablement améliorées avec l’introduction des outils de biologie moléculaire basés principalement sur la Polymerase Chain Reaction (PCR). Ainsi, de petites séquences génomiques du virus Ébola ont pu être amplifiées par PCR à partir des organes de six souris et d’une musaraigne [33]. Malheureusement, ces résultats n’ont pas été confirmés et aucune conclusion n’a pu être déduite. De même, les tentatives d’inoculations expérimentales de rongeurs, chauves-souris, oiseaux, reptiles, mollusques, arthropodes et de plantes ont toutes échoué. Toutefois, certaines chauves-souris appartenant aux genres Epomophorus et Tadarida ont développé une virémie transitoire d’environ quatre semaines, mais là encore, aucune preuve concluante sur le rôle des chauves-souris comme hôtes naturels du virus Ébola n’a pu être obtenue [34].
Le réservoir du virus Ébola a finalement pu être démasqué récemment. À la faveur des épidémies survenues entre 2001 et 2003 au Gabon, de nouvelles collectes ont été effectuées dans les zones atteintes par les épidémies. Les captures d’animaux se sont déroulées dans un rayon d’une dizaine de kilomètres autour des carcasses de gorilles et de chimpanzés infectées quelques jours seulement après leur découverte. Au total, 1030 animaux ont été analysés. Des IgG anti-ebola ont été détectées dans le sérum de 16 chauves-souris dont 4 de l’espèce Hypsignathus monstrosus, 8 de l’espèce Epomops franqueti et 4 de l’espèce Myonycteris torquata. De même, des séquences nucléotidiques virales d’Ébola ont été détectées dans les organes de 13 chauvessouris dont 3 spécimens de l’espèce Hypsignathus monstrosus, 5 de l’espèce Epomops franqueti et 5 de l’espèce Myonycteris torquata [35]. Ces résultats ont par la suite été confirmés par des enquêtes de séroprévalence menées sur 2147 animaux capturés au Gabon et au Congo Brazzaville entre 2003 et 2007 [36]. Des taux élevés et constants ont été obtenus tout au long des quatre années d’études chez les trois espèces, Hypsignathus monstrosus (7%), Epomops franqueti (4%) et Myonycteris torquata (3%).
Même si ces travaux n’ont pu aboutir à l’isolement du virus, les résultats obtenus indiquent qu’au moins trois espèces de chauves-souris frugivores représenteraient le réservoir du virus Ébola. Ces résultats complètent certains indices épidémiologiques recueillis lors des épidémies antérieures évoquant un lien entre l’apparition d’épidémie et la présence de chauves-souris, concordent avec les aires de vie de ces espèces qui s’étendent sur toutes les régions forestières de l’Afrique, du sud du Sénégal au nord de l’Afrique du Sud et à l’Est de la RDC en passant par tous les pays longeant l’Océan Atlantique et confirment enfin les études ayant mis en évidence une virémie transitoire chez certaines espèces de chauves-souris après infection expérimentale. Enfin, il est intéressant de constater que d’autres membres de la famille des Filoviridae ont pu être également détectés chez plusieurs espèces de chauves-souris, le virus de Marburg chez l’espèce Rousettus aegyptiacus d’où il a d’ailleurs pu être isolé, et le virus Lloviu chez plusieurs espèces de Miniopterus [37], [38], [39], [40]. De même, plusieurs virus appartenant à deux familles très proches des Filoviridae, les Paramyxoviridae (virus Hendra et Nipah) et les Rhabdoviridae (virus de la rage) ont pour réservoirs naturels certaines espèces de chauves-souris frugivores.
L’ensemble de ces résultats et de ces informations pointent certaines espèces de chauves-souris frugivores comme réservoirs probables des filovirus en général et du virus Ébola en particulier.
LACIRCULATIONDUVIRUSÉBOLADANSLAFAUNE:UNESTRATÉGIE GAGNANTE?
Lors des épisodes épidémiques survenus au Gabon et au Congo entre 2001 et 2005, de nombreux animaux morts ont été retrouvés dans la forêt des régions affectées [21], [41]. Au total, 44 carcasses ont été découvertes, autopsiées, prélevées puis analysées entre 2001 et 2005. Sur ces 44 carcasses, 16 animaux (12 gorilles, 3 chimpanzés et 1 céphalophe) se sont révélés positifs, ce qui indique que ces animaux ont été naturellement infectés par le virus Ébolaet qu’ils ont développé une infection probablement rapidement létale. Entre 2001 et 2003, une cinquantaine de carcasses de gorilles, 15 carcasses de chimpanzés et 14 carcasses de céphalophes ont été retrouvées et/ou signalées par les villageois dans la zone d’épidémie du Gabon et du Congo. Sachant qu’une carcasse disparaît complètement en moins de trois semaines en forêt tropicale, et que ces carcasses ont été découvertes à moins de 3–4 heures de marche des villages, il est probable que plusieurs centaines voire des milliers d’animaux ont péri pendant ces deux années dans l’ensemble de la région frontalière du nord-est du Gabon et du Congo. Par ailleurs, les calculs d’indices de présence des animaux (excréments, empreintes de pattes, végétaux brisés par le passage des animaux, présence de nids ...) ont révélé une hausse importante de la mortalité chez certaines espèces animales pendant les épidémies humaines. Les populations de gorilles et de céphalophes auraient ainsi chuté de 50% entre 2002 et 2003 et celles de chimpanzés de 88% [24], [25], [42]. Ces études démontrent dans leur ensemble que le virus Ébola se manifeste chez ces populations animales sous la forme de grandes épidémies meurtrières très localisées qui auraient été en grande partie responsables du déclin rapide et massif des populations de gorilles et de chimpanzés dans la région du nord-est du Gabon et du nord-ouest du Congo.
Les résultats obtenus à partir des carcasses animales indiquent également que les grands singes des forêts d’Afrique Centrale seraient fréquemment exposés au virus Ébola. Une enquête sérologique menée sur 790 prélèvements issus d’une vingtaine d’espèces de primates du Cameroun, du Gabon et de République du Congo confirme ces observations [43]. Cette étude montre que 12,9% des chimpanzés sauvages de ces trois pays possèdent des IgG anti-EBOV, certains des échantillons positifs ayant été prélevés avant les premières épidémies dans ces régions. Ces résultats suggèrent que ces animaux sont régulièrement en contact avec l’animal réservoir du virus, que certains d’entre eux développent des infections non létales et que le virus Ébola est vraisemblablement présent depuis très longtemps dans le bloc forestier d’Afrique Centrale. La présence d’anticorps spécifiques anti-EBOV chez d’autres espèces de primates (5 drills sur 34, 1 babouin sur 25, 1 mandrill sur 296 et 1 cercopithèque sur 107) suggère que la circulation du virus pourrait être beaucoup plus complexe que le simple passage du réservoir aux gorilles et aux chimpanzés. L’existence de plusieurs espèces réservoirs et la possibilité que d’autres espèces animales soient sensibles au virus et susceptibles de participer à la propagation du cycle naturel du virus dans la nature ne sont donc pas à exclure [44].
DERNIÈRE ÉTAPE: LA TRANSMISSION À L’HOMME
Dans la plupart des épidémies, la source de contamination de la première personne identifiée comme infectée (cas primaire) n’est pas connue. C’est le cas pour toutes les épidémies de la période 1976–1979, des épidémies de Mékouka (Gabon) en 1995, de Booué (Gabon) en 1996, et de Kikwit (RDC) en 1995.
Deux modes de contamination de l’homme sont proposés aujourd’hui (Figure 3 ): (i) une contamination indirecte par l’intermédiaire d’espèces animales sensibles au virus; (ii) une contamination directe auprès du réservoir.
Fig. 3.

Schéma synthétique des différents modes de contamination de l’homme par le virus Ébola conduisant à l’apparition des épidémies.
Un mode avéré, la contamination indirecte
Plusieurs études ont montré que la contamination de l’homme s’est parfois produite lors de la manipulation de carcasses infectées de gorilles, chimpanzés et de céphalophes [21], [41]. À titre d’exemple, des carcasses de gorilles seraient à l’origine des chaînes de Olloba 2001, Grand Etoumbi 2002, Entsiami 2002 et Yembelengoye 2002. Des carcasses de chimpanzés seraient à l’origine des chaînes d’Etakangaye 2001 et Olloba 2002. Enfin, des carcasses de céphalophes seraient à l’origine des chaînes de Mendemba 2001, Ekata 2001 et Mvoula 2003. Ainsi, l’infection des grands singes par le virus Ébola, rapidement mortelle, laisse des carcasses infectées au contact desquelles les humains, généralement des chasseurs, contractent le virus. Les grands singes se contamineraient eux-mêmes probablement directement auprès des chauves-souris, en particulier lorsque les animaux de ces espèces animales consomment en même temps les fruits d’un même arbre.
Afin d’élucider les modes de contamination des grands singes, des échantillons biologiques (muscle, peau ou organes) ont été systématiquement prélevés sur les carcasses de gorilles et de chimpanzés, puis la partie codante de la GP (gène le plus variable) caractérisée [45]. Ces analyses ont mis en évidence une séquence virale différente pour chaque carcasse. Des séquences virales différentes ont même été obtenues à partir de deux gorilles morts découverts à quelques mètres l’un de l’autre. Cette diversité génétique des souches virales observées chez les carcasses d’animaux morts exclut par conséquent une transmission du virus d’un individu à l’autre et suggère au contraire que l’infection des grands singes résulterait de contaminations simultanées mais indépendantes à partir de sources animales distinctes, probablement le réservoir naturel du virus Ébola [21], [45].
Comment les grands singes peuvent-ils se contaminer? Les épidémies survenant la plupart du temps à la fin des saisons sèches, le passage du virus du réservoir aux grands singes peut s’accomplir à la faveur de conditions environnementales particulières. En effet, les enquêtes épidémiologiques menées à cette période ont montré que les mortalités des grands singes surviennent principalement à la fin des saisons sèches, à une période de l’année où les ressources alimentaires se raréfient [21], [41]. L’appauvrissement en ressources alimentaires augmenterait la fréquence de la consommation simultanée de mêmes fruits par les chauves-souris et les grands singes. Ces rassemblements augmenteraient alors la probabilité de contact entre ces deux espèces. De plus, un certain nombre d’évènements comportementaux et physiologiques se produisent chez les espèces de chauves-souris incriminées pendant cette période, tels que les compétitions entre mâles, et les mises bas groupées des femelles. Ces évènements participeraient à modifier la nature des réponses immunitaires, propices à la reprise de la réplication virale dans les organes voire à l’apparition de virus dans la circulation sanguine. La contamination des grands singes interviendrait alors à la faveur de contacts sanguins directs avec le sang ou les tissus placentaires des femelles chauves-souris au moment de la parturition [46], ou alors dans une moindre mesure avec la salive, l’urine ou les fèces.
Un mode fortement suspecté mais non démontré, la contamination directe auprès du réservoir
Bien que la contamination directe des chauves-souris à l’homme n’ait jamais pu être prouvée de manière formelle, une étude, menée au cours de l’épidémie d’Ébola de 2007 en RDC, a mis en évidence pour la première fois un lien entre les chauvessouris et l’apparition d’une épidémie d’Ébola [47]. Les investigations épidémiologiques et écologiques ont en effet montré que l’épidémie était associée à l’arrivée massive de grandes colonies de chauves-souris près des villages touchés. Au cours d’une migration nord-sud, ces chauves-souris ont séjourné près des villages pendant environ deux mois, période pendant laquelle elles étaient massivement chassées et consommées par les villageois. La contamination se serait produite lors de la manipulation des animaux morts couverts de sang ou lors du dépeçage (Figure 3).
Dans l’espoir d’obtenir des informations sur l’implication des chauves-souris dans la propagation et le maintien du virus au sein de l’écosystème, la circulation du virus dans les populations humaines a été évaluée. Pour cela, les IgG anti-EBOV ont été recherchés à partir d’un échantillonnage stratifié par province [48]. Les populations d’étude ont été sélectionnées dans chaque province par sondage aléatoire des villages de moins de 500 habitants de manière à cibler préférentiellement les populations rurales. Au total, 4358 personnes ont été prélevées. De manière surprenante, près de 15% des personnes testées présentaient des IgG anti-EBOV. Cette prévalence atteint même 20% dans les régions forestières comparativement aux régions savanicoles où la prévalence n’est que de 3%. Ces résultats ont été confirmés par western-blot et, chez certaines personnes IgG +, par la mise en évidence d’une mémoire cellulaire spécifique exprimée par une augmentation significative des lymphocytes T CD3 + CD8 + IFNγ + à J2 et/ou J3 après la stimulation, comparativement à des personnes IgG négatives.
Ces résultats font état de prévalences étonnamment très élevées non compatibles avec les modes d’apparition du virus Ébola chez l’homme au Gabon tels qu’un faible nombre d’épidémies (4), des épidémies de faible envergure (moins de 100 cas), et une létalité élevée (80%) qui laissent peu de survivants après chaque épidémie. Ces taux élevés excluent donc l’hypothèse selon laquelle ces personnes IgG + seraient des survivants d’une infection par le virus Ébola [48]. Dans l’ensemble, ces résultats émettent au contraire l’hypothèse d’une exposition permanente au virus et l’existence de contacts fréquents entre les populations humaines et les sources animales. Les taux élevés suggèrent donc que ces sources animales sont probablement des animaux vivant en très grand nombre, pointant les chauves-souris comme candidats plausibles et excluant les grands singes et autres mammifères terrestres. Les contacts entre les habitants des villages et les chauves-souris se manifestent généralement lors de la consommation de fruits souillés par la salive ou par l’urine des chauves-souris qui peuplent par milliers voire par millions ces régions. Le développement des réponses IgG spécifiques pourrait alors résulter d’une infection rapidement avortée ou d’une simple stimulation antigénique par des particules virales non infectieuses. Bien que peu probable, l’hypothèse de la présence de souches virales non pathogènes hébergées chez les chauves-souris n’est pas à exclure.
Bien qu’exceptionnelle par son ampleur et sa dispersion géographique, l’apparition d’une épidémie d’Ébola en Afrique de l’Ouest, peuplée par les mêmes espèces de chauve-souris, ne surprend pourtant pas. À la faveur de conditions favorables, la sortie du virus de son hôte naturel est susceptible de se produire à n’importe quel endroit dans ces régions. La transmission à l’homme survient le plus souvent lors du dépeçage de ces animaux qui sont largement consommés par les populations peuplant ces régions.
Les facteurs qui gouvernent la transmission du virus à l’homme peuvent être subdivisés en trois groupes: — (i) les facteurs sociaux et comportementaux, c’est-à-dire tous les facteurs qui conduisent à l’exposition au virus; — (ii) les facteurs virologiques au sein de l’animal, c’est-à-dire la charge virale; — (iii) les facteurs virologiques dans les populations réservoirs, c’est-à-dire le taux d’animaux infectés au sein de la population toute entière. Alors que les raisons de la variabilité des facteurs des deux premiers groupes sont en partie connus (augmentation de la consommation de chauves-souris en cas de raréfaction des ressources alimentaires, augmentation de la charge virale chez les femelles en fin de gestation ou chez les individus affaiblis ou infectés par un autre agent pathogène), beaucoup reste à découvrir sur la variabilité des autres facteurs. L’allure globale des épidémies d’Ébola semble indiquer, à l’échelle continentale, une certaine cyclicité des épidémies avec des périodes basses et des périodes hautes, comme si à certains moments il y avait une augmentation significative du pourcentage d’animaux infectés sur l’ensemble du continent entrainant de facto une augmentation du risque de transmission à l’homme. Aujourd’hui, la survenue d’au moins deux épidémies différentes dans des régions éloignées l’une de l’autre (Guinée et RDC) tend à appuyer cette hypothèse.
Le rôle des animaux domestiques plus que jamais d’actualité
Par ailleurs, lors des dernières épidémies survenuesauGabon et au Congo, plusieurs chiens ont consommé des restes d’animaux infectés par le virus Ébola, sans pour autant présenter de signes cliniques visibles. Afin de vérifier si les chiens ont bien été en contact avec le virus, nous avons recherché la présence d’anticorps spécifiques du virus Ébola. Le pourcentage de chiens porteurs de tels anticorps croît de manière linéaire et significative à mesure que l’on s’approche des foyers épidémiques [49]. De 9% dans les deux grandes villes du Gabon, la prévalence passe à 15% dans la plus grande ville de la zone épidémique, puis à 25% dans les villages indemnes de cette même zone, pour atteindre 32% dans les villages où des cas humains ont pu être imputés à une source animale infectée. Les chiens pourraient donc être infectés par le virus Ébola, excréter transitoirement du virus, et devenir de ce fait une source potentielle d’infection pour l’homme. Ceci pourrait expliquer certaines contaminations humaines non élucidées. La corrélation linéaire qui existe entre la prévalence chez les chiens domestiques et le risque épidémique des villages dans lesquels ils vivent suggère que la prévalence chez ces animaux peut être utilisée comme indicateur de la présence du virus dans les régions où, hormis l’apparition de cas de mortalité animale et humaine, aucun signe ne peut indiquer la présence du virus Ébola.
L’AVENIR DU VIRUS ÉBOLA: SON INSTALLATION DANS L’ESPÈCE HUMAINE?
L’infection de l’homme par le virus Ébola se traduit par une maladie d’évolution rapide caractérisée par une forte fièvre et des hémorragies diffuses. Après une période d’incubation d’environ une semaine (moyenne 4–9 jours), la maladie survient brutalement par de la fièvre accompagnée de céphalées, nausées, douleurs musculaires et asthénie. Quelques jours plus tard apparaissent des signes cliniques diversifiés qui témoignent d’une diffusion généralisée du virus et d’une atteinte multiviscérale: rash érythémateux, symptômes digestifs violents (douleurs abdominales, vomissements, diarrhées profuses), troubles respiratoires (maux de gorge, toux) et signes neurologiques (prostration, confusion, délire) sont les symptômes les plus fréquemment observés. Les manifestations hémorragiques (méléna, hématémèse, injection conjonctivale, saignements de nez, saignements aux sites de ponction veineuse ...) ne surviennent que chez environ 1/3 des patients, généralement en phase terminale de la maladie, soit une à deux semaines après l’apparition des symptômes. Dans les formes mortelles, les symptômes et les hémorragies s’aggravent et se généralisent rapidement, la mort survient dans les 48 heures dans un état de choc. Dans les formes non mortelles, les symptômes s’atténuent rapidement, simultanément à la disparition du virus de la circulation sanguine. Même si une contamination peut théoriquement se produire pendant la période d’incubation ou après la guérison, les patients ne sont généralement contagieux que pendant la phase symptomatique. La convalescence est généralement longue et marquée d’accès fébriles épisodiques. Des signes d’orchite ou d’uvéite, liés à la persistance du virus dans les sites immunologiquement protégés, sont parfois observés pendant plusieurs semaines après la disparition des symptômes. Le virus a en effet pu être isolé près de deux mois après la fin des symptômes dans le sperme d’un patient.
La forme mortelle de l’infection Ébola est caractérisée par un profond dysfonctionnement de l’immunité innée (libération excessive de composants inflammatoires et absence des défenses antivirales) et un effondrement de l’immunité adaptative [50], [51], [52] (Figure 4 ). L’infection massive des monocytes et des cellules dendritiques conduit à une libération excessive de nombreux médiateurs pro-inflammatoires et de composants vasoactifs qui ont une implication majeure dans la physiopathologie [53], [54]. Cet orage inflammatoire, particulièrement important en phase terminale de la maladie, favorise en effet la fuite vasculaire, altère les fonctions des cellules endothéliales induisant in fine la coagulation intravasculaire disséminée [55], [56], [57]. D’autre part, certaines protéines virales sont capables d’inhiber les défenses antivirales des cellules [58]. Ainsi, la VP35 inhibe la synthèse des IFNa/ß en s’opposant à l’activation du facteur de régulation de l’interféron (IRF)-3 et IRF-7, interfère avec l’activation de la dsRNA-protéine kinase dépendante (PKR), et, comme les VP30 et VP40, inhibe le « RNA silencing » [59], [60]. La VP24 empêche l’accumulation nucléaire de la protéine STAT1 phosphorylée, neutralisant par conséquent la réponse aux IFNa/ß et IFNγ [61], [62]. L’effondrement de l’immunité adaptative se manifeste par une déplétion des cellules lymphoïdes dans les ganglions lymphatiques, la rate et le thymus, l’apoptose intravasculaire des lymphocytes T et B et des cellules NK, et l’absence de production d’IgG spécifiques. Les lymphocytes n’étant pas infectés par le virus, l’apoptose résulterait d’interactions avec des marqueurs de surface (Fas/FasL, TNF/TRAIL) et/ou de médiateurs solubles apoptogènes et/ou d’une activité superantigénique de certaines protéines virales [63], [64]. L’effondrement de l’immunité adaptative peut également résulter d’un défaut d’activation et de maturation des cellules dendritiques infectées alors incapables d’initier les réponses immunitaires.
Fig. 4.

Schéma synthétique des mécanismes physiopathologiques associées à l’infection par le virus Ébola.
La fièvre hémorragique Ébola est une zoonose transmise accidentellement à l’homme directement de son (ou de ses) réservoir(s), ou indirectement par l’intermédiaire d’espèces animales intermédiaires (chimpanzés, gorilles principalement). Chez son réservoir, le virus se réplique probablement très lentement au sein des organes cibles (rate, foie principalement), la charge virale est le plus souvent indétectable et l’animal reste la plupart du temps asymptomatique. Ainsi, l’évolution génomique du virus est très lente. La situation est totalement différente chez l’homme. Le virus trouve un terrain favorable à sa multiplication et atteint des charges virales extrêmement élevées, jusqu’à1 milliard de particules par mL de sang. La forte multiplication virale augmente donc considérablement la probabilité de mutations. Les études génomiques conduites à partir des souches isolées au cours des épidémies de 1976 à 2008 font état d’une très faible diversité génétique (autour de 2%). Cette relative stabilité du virus s’explique probablement par la faible envergure des épidémies survenues jusqu’ici. Or, l’ampleur exceptionnelle de l’épidémie qui affecte actuellement l’Afrique de l’Ouest pourrait modifier la situation et accélérer considérablement l’évolution génomique du virus. La multiplication du nombre de cas et donc du nombre de transmissions interhumaines va accroître le risque de mutations du virus qui pourrait être à l’ origine, à plus ou moins long terme, de modifications phénotypiques du virus telles qu’une modification du pouvoir pathogène du virus ou d’un changement du mode de transmission chez l’homme.
CONCLUSION
Diagnostic et riposte tardifs, insuffisance des structures sanitaires, manque de professionnalisme du personnel de santé, défaut d’adhésion d’une partie des populations aux méthodes d’isolement des patients principalement en raison de traditions culturelles parfois imperméables, déplacements incessants des personnes voire des malades eux-mêmes dus à un éparpillement extrême des familles résultant d’une culture nomade ancestrale et enfin contexte politique complexe issu d’un climat de guerre civile récent sont les principales raisons qui expliquent l’ampleur de l’épidémie actuelle en Afrique de l’Ouest. En quelque sorte, tous les ingrédients sont réunis pour que le virus Ébola puisse enfin se propager durablement au sein de l’espèce humaine à la faveur de modifications génomiques. Car, à l’instar de tous les êtres vivants, Ébola a besoin de se multiplier abondamment pour que des erreurs se produisent dans son génome, que des mutations surviennent quantitativement et qualitativement suffisamment pour qu’elles puissent s’accompagner de modifications phénotypiques telles qu’une diminution de la virulence, un allongement de la période d’incubation, une diminution et/ou un allongement de la durée des symptômes, et une modification du mode de transmission. Ces modifications, si elles venaient à se produire, auraient des conséquences dramatiques: elles augmenteraient la contagiosité et la transmissibilité du virus qui permettrait son installation durable voire définitive dans l’espèce humaine.
En raison de la multiplication des « sorties » du virus à partir de la faune animale, une réflexion sur les stratégies à mettre en place afin de prévoir la contamination de l’homme à partir de la faune animale et prévenir l’apparition des épidémies, doit être impérativement initiée le plus rapidement possible. Or, une telle démarche fait nécessairement appel à une connaissance parfaite de l’écologie du virus, et à l’identification complète des hôtes intermédiaires, des réservoirs et des modalités des transferts inter espèces. En particulier, la distribution géographique du virus doit permettre de déterminer de manière assez précise les zones à risque. De même, une certaine saisonnalité des épidémies a été observée à plusieurs reprises, ce qui nécessite l’intégration, dans les stratégies de prévention, d’une dimension temporelle dont les contours restent encore à définir.
La confirmation d’une cyclicité des épidémies à virus Ébola à l’échelle continentale, la recherche des facteurs conduisant à l’augmentation du taux global d’infection au sein des populations de chauves-souris pourrait être précieuse dans l’évaluation du risque d’émergence virale et la prédiction des épidémies. La détermination d’un seuil d’alerte pourrait s’avérer alors crucial dans la lutte contre les épidémies dans laquelle la rapidité d’intervention est fondamentale.
DISCUSSION
M. Pierre BÉGUÉ
Comment explique-t-on que les premières recherches de virus Ébola sur 10 000 animaux ont échoué et que des travaux récents ont démontré sans contestation que les chauves-souris en étaient porteuses? Le virus Ébola est-il fragile
Le virus Ébola est fragile dans le milieu extérieur. Il est rapidement inactivé.
Plusieurs raisons peuvent expliquer les échecs passés:
-
•
Les techniques anciennes de détection (isolement viral, sérologie par immunofluorscence ...) étaient très peu sensibles. Les technologies modernes, principalement basées sur la biologie moléculaire, ont considérablement amélioré les capacités de détection des agents infectieux.
-
•
Les recherches ont souvent été effectuées bien après l’émergence de l’épidémie, c’est ça dire après la date de sortie du virus de la faune vers la population humaine. Les chances de détecter un animal infecté plusieurs semaines voire plusieurs mois après le début de l’épidémie étaient donc minimes.
-
•
Faible prévalence d’infection au sein de l’espèce animale réservoir.
-
•
Faible charge virale dans l’individu infecté d’où une grande difficulté de détecter le virus avec les techniques anciennes.
Mme Monique ADOLPHE
Les chauves-souris réservoirs de virus Ébola existent-elles dans d’autres forêts équatoriales comme les forêts brésiliennes?
Des chauves-souris appartenant aux mêmes genres taxonomiques existent dans les autres continents. Seules les espèces sont différentes.
M. Pierre GODEAU
J’ai cru comprendre dans votre présentation que la contamination des grands singes par les chauves-souris se produisait préférentiellement en fin de saison sèche où les singes contraints de modifier leur alimentation consommaient des fruits contaminés par les chauves-souris, est-ce exact?
Ce ne sont que des présomptions. Il se produit un appauvrissement en ressources alimentaires pendant la saison sèche qui se manifeste par une diminution de fruits. La probabilité que les grands singes et les chauves-souris consomment les mêmes fruits aux mêmes moments est par conséquent plus élevée, ce qui augmente de facto le risque de contact. De plus, cette période correspond à la mise-bas des femelles. Les grands singes peuvent donc être aisément en contact avec du sang placentaire susceptible de s’écouler pendant la mise bas voire directement avec le placenta des femelles.
M. François BRICAIRE
A-t-on un début d’explication du fait que les chauves-souris sont des réservoirs de nombreux virus? Ébola mais aussi coronavirus, rage, etc.
Je pense que cette impression est purement factuelle, et est amplifiée par la couverture médiatique des maladies dues à certains virus dont les chauves-souris sont réservoirs (SARS, Ébola, Marburg ...). De plus, la découverte de ces virus chez les chauves-souris coïncide avec les avancées technologiques qui ont été déterminantes dans la détection de ces virus. De très nombreux autres virus sont hébergés par d’autres espèces animales, en particulier les rongeurs, comme par exemple la fièvre de Lassa en Afrique, responsable de centaines de milliers de morts chaque année.
M. André VACHERON
Étant allé pendant une dizaine d’années présider les thèses de Médecin dans les CHU de l’Afrique de l’Ouest, j’ai visité le CIRMF de Franceville au Gabon avec sa réserve de grands singes. Les pygmées de la forêt centrafricaine sont-ils atteints par le virus Ébola?
Oui, ils le sont. Les pygmées ont également payé un lourd tribut lors des épidémies d’Ébola qui ont sévi au Gabon et en République du Congo entre 1995 et 2005.
M. Marc GIRARD
A-t-on injecté du virus Ébola à des chauves-souris? Observe-t-on alors une infection aigüe, éventuellement transitoire? Ou au contraire une infection chronique d’où la crétion d’un réservoir?
Des infections expérimentales de chauves-souris ont été pratiquées vers la fin des années 90. Seules des virémies transitoires ont été observées. De nouvelles infections expérimentales sont actuellement tentées en Afrique du sud.
M. Jacques BATTIN
Avez-vous l’expérience de la borréliose, car on entend dire que la maladie de Lyme devient une pandémie dont le réservoir est constitué de rongeurs?
Nous n’avons pas d’expérience particulière de la Borréliose. C’est effectivement une problématique qu’il convient d’explorer en Afrique centrale.
M. André-Laurent PARODI
Dans d’autres régions d’Afrique, à l’Ouest (Rwanda, Burundi) vivent aussi de grands primates et des chauves-souris. Des schémas épidémiologiques semblables à celui que vous avez exposé ont-ils été observés?
Des schémas épidémiologiques similaires sont observés en Afrique de l’Ouest, en particulier s’agissant du virus de Marburg, autre membre appartenant à la famille virale des Filoviridae. Pour le virus de Marburg, il a été clairement démontré que la Roussette d’Égypte, une autre espèce de chauves-souris frugivore en était le réservoir. Le virus a même été isolé à partir de certains organes de plusieurs individus.
Footnotes
L’auteur déclare n’avoir aucun lien d’intérêt en relation avec le contenu de cet article.
Tirés-à-part: Professeur Éric Leroy
RÉFÉRENCES
- 1.Who Ebola Response Team Ebola virus disease in West Africa-the first 9 months of the epidemic and forward projections. N Engl J Med. 2014;371:1481–1495. doi: 10.1056/NEJMoa1411100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gire S.K., Goba A., Andersen K.G. Genomic surveillance elucidates Ebola virus origin and transmission during the 2014 outbreak. Science. 2014;345:1369–1372. doi: 10.1126/science.1259657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Baize S., Pannetier D., Oestereich L. Emergence of Zaire Ebola virus disease in Guinea. N Engl J Med. 2014;371:1418–1425. doi: 10.1056/NEJMoa1404505. [DOI] [PubMed] [Google Scholar]
- 4.Briand S., Bertherat E., Cox P. The international Ebola emergency. N Engl J Med. 2014;371:1180–1183. doi: 10.1056/NEJMp1409858. [DOI] [PubMed] [Google Scholar]
- 5.Leroy E.M., Labouba I., Maganga G.D. Ebola in West Africa: the outbreak able to change many things. Clin Microbiol Infect. 2014 doi: 10.1111/1469-0691.12781. [DOI] [PubMed] [Google Scholar]
- 6.Kuhn J.H. Filoviruses. A compendium of 40 years of epidemiological, clinical, and laboratory studies. Arch Virol Suppl. 2008;20:13–360. [PubMed] [Google Scholar]
- 7.Kuhn J.H., Bao Y., Bavari S. Virus nomenclature below the species level: a standardized nomenclature for natural variants of viruses assigned to the family Filoviridae. Arch Virol. 2013;158:301–311. doi: 10.1007/s00705-012-1454-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kuhn J.H., Andersen K.G., Bao Y. Filovirus RefSeq entries: evaluation and selection of filovirus type variants, type sequences. and names. Viruses. 2014;6:3663–3682. doi: 10.3390/v6093663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Smith D.I. Ebola haemorrhagic fever in Sudan. 1976. Bull. World Health Organ. 1978;56:247–270. [PMC free article] [PubMed] [Google Scholar]
- 10.Baron R.C., Mccormick J.B., Zubeir O.A. Ebola virus disease in southern Sudan: hospital dissemination and intrafamilial spread. Bull. World Health Organ. 1983;61:997–1003. [PMC free article] [PubMed] [Google Scholar]
- 11.Okware S.I., Omaswa F.G., Zaramba S. An outbreak of Ebola in Uganda. Trop. Med Int Health. 2002;7:1068–1075. doi: 10.1046/j.1365-3156.2002.00944.x. [DOI] [PubMed] [Google Scholar]
- 12.Towner J.S., Sealy T.K., Khristova M.L. Newly discovered ebola virus associated with hemorrhagic Fever outbreak in Uganda. PLoS Pathog. 2008;4:e1000212. doi: 10.1371/journal.ppat.1000212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Le Guenno B., Formenty P., Wyers M. Isolation and partial characterisation of a new strain of Ebola. The Lancet. 1995;345:1271–1274. doi: 10.1016/s0140-6736(95)90925-7. [DOI] [PubMed] [Google Scholar]
- 14.Formenty P., Hatz C., Le Guenno B. Human infection due to Ebola virus, subtype côte d’ivoire: clinical and biologic presentation. J Infect Dis. 1999;179:S48–S53. doi: 10.1086/514285. [DOI] [PubMed] [Google Scholar]
- 15.Rollin P.E., Williams R.J., Bressler D.S. Ebola (subtype Reston) virus among quarantined nonhuman primates recently imported from the Philippines to the United States. J Infect Dis. 1999;179:S108–S114. doi: 10.1086/514303. [DOI] [PubMed] [Google Scholar]
- 16.Barrette R.W., Metwally S.A., Rowland J.M. Discovery of swine as a host for the Reston ebolavirus. Science. 2009;325:204–206. doi: 10.1126/science.1172705. [DOI] [PubMed] [Google Scholar]
- 17.Johnson K.M. Ebola haemorrhagic fever in Zaire. 1976. Bull. World Health Organ. 1978;56:271–293. [PMC free article] [PubMed] [Google Scholar]
- 18.Khan A.S., Tshioko F.K., Heymann D.L. The reemergence of Ebola hemorrhagic fever, Democratic Republic of the Congo, 1995. J. Infect. Dis. 1999;179:S76–S86. doi: 10.1086/514306. [DOI] [PubMed] [Google Scholar]
- 19.Georges A.J., Leroy E.M., Renaut A.A. Ebola hemorrhagic fever outbreaks in Gabon, 1994–1997: Epidemiologic and Health control issues. J Infect Dis. 1999;179:S65–S75. doi: 10.1086/514290. [DOI] [PubMed] [Google Scholar]
- 20.Leroy E.M., Souquière S., Rouquet P. Re-emergence of Ebola haemorrhagic fever in Gabon. Lancet. 2002;359:712. doi: 10.1016/s0140-6736(02)07796-6. [DOI] [PubMed] [Google Scholar]
- 21.Leroy E.M., Rouquet P., Formenty P. Multiple Ebola virus transmission events and rapid decline of central african wildlife. Science. 2004;303:387–390. doi: 10.1126/science.1092528. [DOI] [PubMed] [Google Scholar]
- 22.Nkoghe D., Kone M.L., Yada A. A limited outbreak of Ebola haemorrhagic fever in Etoumbi, Republic of Congo. 2005. Trans R Soc Trop Med Hyg. 2011;105:466–472. doi: 10.1016/j.trstmh.2011.04.011. [DOI] [PubMed] [Google Scholar]
- 23.Formenty P., Leroy E.M., Epelboin A. Detection of Ebola virus in oral fluid specimens during outbreaks of Ebola virus hemorrhagic fever in the Republic of Congo. Clin Infect Dis. 2006;42:1521–1526. doi: 10.1086/503836. [DOI] [PubMed] [Google Scholar]
- 24.Walsh P.D., Abernethy K.A., Bermejo M. Catastrophic ape decline in western equatorial Africa. Nature. 2003;422:611–614. doi: 10.1038/nature01566. [DOI] [PubMed] [Google Scholar]
- 25.Bermejo M., Rodriguez-Teijeiro J.D., Illera G. Ebola outbreak killed 5000 gorillas. Science. 2006;314:1564. doi: 10.1126/science.1133105. [DOI] [PubMed] [Google Scholar]
- 26.Grard G., Biek R., Tamfum J.J. Emergence of divergent Zaire ebola virus strains in Democratic Republic of the Congo in 2007 and 2008. J Infect Dis. 2011;204(Suppl 3):S776–S784. doi: 10.1093/infdis/jir364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Maganga G.D., Kapetshi J., Berthet N. Ebola Virus Disease in the Democratic Republic of Congo. N Engl J Med. 2014 doi: 10.1056/NEJMoa1411099. [DOI] [PubMed] [Google Scholar]
- 28.Arata AA, Johnson B. Approaches towards studies on potential reservoirs of viral haemorrhagic fever in southern Sudan. 1977. [Google Scholar]
- 29.Heymann D.L., Weisfeld J.S., Webb P.A. Ebola hemorrhagic fever: Tandala, 1977–1978. J. Infect. Dis. 1980;142:372–376. doi: 10.1093/infdis/142.3.372. [DOI] [PubMed] [Google Scholar]
- 30.Breman J.G., Johnson K.M., van der Groen G. A search for Ebola virus in animals in the Democratic Republic of the Congo and Cameroon: Ecologic, virologic, and serologic surveys, 1979–1980. J. Infect. Dis. 1999;179:S139–S147. doi: 10.1086/514278. [DOI] [PubMed] [Google Scholar]
- 31.Reiter P., Turell M., Coleman R. Field investigations of an outbreak of Ebola hemorrhagic fever, Kikwit, Democratic Republic of the Congo, 1995: Arthropod studies. J Infect Dis. 1999;179:S148–S154. doi: 10.1086/514304. [DOI] [PubMed] [Google Scholar]
- 32.Leirs H., Mills J.N., Krebs J.W. Search for the Ebola virus reservoir in Kikwit, Democratic Republic of the Congo: Reflections on a vertebrate collection. J Infect Dis. 1999;179:S155–S163. doi: 10.1086/514299. [DOI] [PubMed] [Google Scholar]
- 33.Morvan J.M., Deubel V., Gounon P. Identification of Ebola virus sequences present as RNA or DNA in organs of terrestrial small mammals of the Central African Republic. Microbes Infect. 1999;1:1193–1201. doi: 10.1016/s1286-4579(99)00242-7. [DOI] [PubMed] [Google Scholar]
- 34.Swanepoel R., Leman P.A., Burt F.J. Experimental inoculation of plants and animals with Ebola virus. Emerg Infect Dis. 1996;2:321–325. doi: 10.3201/eid0204.960407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Leroy E.M., Kumulungui B., Pourrut X. Fruit bats as reservoirs of Ebola virus. Nature. 2005;438:575–576. doi: 10.1038/438575a. [DOI] [PubMed] [Google Scholar]
- 36.Pourrut X., Delicat A., Rollin P.E. Spatial and temporal patterns of Zaire ebolavirus antibody prevalence in the possible reservoir bat species. J Infect Dis. 2007;196(Suppl 2):S176–S183. doi: 10.1086/520541. [DOI] [PubMed] [Google Scholar]
- 37.Towner J.S., Pourrut X., Albarino C.G. Marburg virus infection detected in a common African bat. PLoS ONE. 2007;2:e764. doi: 10.1371/journal.pone.0000764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Towner J.S., Amman B.R., Sealy T.K. Isolation of genetically diverse Marburg viruses from Egyptian fruit bats. PLoS Pathog. 2009;5:e1000536. doi: 10.1371/journal.ppat.1000536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Swanepoel R., Smit S.B., Rollin P.E. Studies of reservoir hosts for Marburg virus. Emerg Infect Dis. 2007;13:1847–1851. doi: 10.3201/eid1312.071115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Negredo A., Palacios G., Vazquez-Moron S. Discovery of an ebolavirus-like filovirus in europe. PLoS Pathog. 2011;7:e1002304. doi: 10.1371/journal.ppat.1002304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rouquet P., Froment J.M., Bermejo M. Wild animal mortality monitoring and human Ebola outbreaks, Gabon and Republic of Congo, 2001–2003. Emerg Infect Dis. 2005;11:283–290. doi: 10.3201/eid1102.040533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Huijbregts B., de Wachter P., Ndong Obiang S. Ebola and the decline of gorilla Gorilla gorilla and chimpanzee Pan troglodytes populations in Minkebe forest, north-eastern Gabon. Oryx. 2003;37:437–443. [Google Scholar]
- 43.Leroy E.M., Telfer P., Kumulungui B. A serological survey of Ebola virus infection in central African nonhuman primates. J Infect Dis. 2004;190:1895–1899. doi: 10.1086/425421. [DOI] [PubMed] [Google Scholar]
- 44.Feldmann H., Wahl-Jensen V., Jones S.M. Ebola virus ecology: a continuing mystery. Trends Microbiol. 2004;12:433–437. doi: 10.1016/j.tim.2004.08.009. [DOI] [PubMed] [Google Scholar]
- 45.Wittmann T.J., Biek R., Hassanin A. Isolates of Zaire ebolavirus from wild apes reveal genetic lineage and recombinants. Proc. Natl. Acad. Sci USA. 2007;104:17123–17127. doi: 10.1073/pnas.0704076104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Leroy E., Pourrut X., Gonzalez J.P. Les chauves-souris, réservoirs du virus Ebola: le mystère se dissipe. Médecine/Sciences. 2006;22:78–79. doi: 10.1051/medsci/200622178. [DOI] [PubMed] [Google Scholar]
- 47.Leroy E.M., Epelboin A., Mondonge V. Human Ebola Outbreak Resulting from Direct Exposure to Fruit Bats in Luebo, Democratic Republic of Congo. 2007. Vector Borne Zoonotic Dis. 2009;6:723–728. doi: 10.1089/vbz.2008.0167. [DOI] [PubMed] [Google Scholar]
- 48.Becquart P., Wauquier N., Mahlakoiv T. High prevalence of both humoral and cellular immunity to Zaire ebolavirus among rural populations in Gabon. PLoS One. 2010;5 doi: 10.1371/journal.pone.0009126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Allela L., Boury O., Pouillot R. Ebola virus antibody prevalence in dogs and human risk. Emerg. Infect. Dis. 2005;11:385–390. doi: 10.3201/eid1103.040981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Baize S., Leroy E.M., Georges-Courbot M.C. Defective humoral responses and extensive intravascular apoptosis are associated with fatal outcome in Ebola virus-infected patients. Nature Med. 1999;5:423–426. doi: 10.1038/7422. [DOI] [PubMed] [Google Scholar]
- 51.Wauquier N., Becquart P., Padilla C. Human fatal zaire Ebola virus infection is associated with an aberrant innate immunity and massive lymphocyte apoptosis. PLoS Neg Trop Dis. 2010;e837:4. doi: 10.1371/journal.pntd.0000837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mahanty S., Bray M. Pathogenesis of filoviral haemorrhagic fevers. Lancet Infect. Dis. 2004;4:487–498. doi: 10.1016/S1473-3099(04)01103-X. [DOI] [PubMed] [Google Scholar]
- 53.Mahanty S., Hutchinson K., Agarwal S. Cutting edge: impairment of dendritic cells and adaptive immunity by Ebola and Lassa viruses. J Immunol. 2003;170:2797–2801. doi: 10.4049/jimmunol.170.6.2797. [DOI] [PubMed] [Google Scholar]
- 54.Geisbert T.W., Hensley L., Larsen T. Pathogenesis of Ebola Hemorrhagic fever in cynomolgus macaques: evidence that dendritic cells are early and sustained targets of infection. Am J Pathol. 2003;163:2347–2370. doi: 10.1016/S0002-9440(10)63591-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ströher U., West E., Bugany H. Infection and activation of monocytes by Marburg and Ebola viruses. J Virol. 2001;75:11025–11033. doi: 10.1128/JVI.75.22.11025-11033.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Gupta M., Mahanty S., Ahmed R. Monocyte-derived human macrophage and peripheral blood mononuclear cells infected with Ebola virus secrete MIP-1a and TNF-a and inhibit poly-IC-induced IFN-a in vitro. Virology. 2001;284:20–25. doi: 10.1006/viro.2001.0836. [DOI] [PubMed] [Google Scholar]
- 57.Baize S., Leroy E.M., Georges A.J. Inflammatory responses in Ebola virus-infected patients. Clin Exp Immunol. 2001;128:163–168. doi: 10.1046/j.1365-2249.2002.01800.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Leroy E.M., Gonzalez J.P., Ebola Baize S. Marburg haemorrhagic fever viruses: major scientific advances, but a relatively minor public health threat for Africa. Clin Microbiol Infect. 2011;17:964–976. doi: 10.1111/j.1469-0691.2011.03535.x. [DOI] [PubMed] [Google Scholar]
- 59.Feng Z., Cerveny M., Yan Z. The VP35 protein of Ebola virus inhibits the antiviral effect mediated by double-stranded RNA-dependent protein kinase PKR. J. Virol. 2007;81:182–192. doi: 10.1128/JVI.01006-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Fabozzi G., Nabel C.S., Dolan M.A. Ebolavirus proteins suppress the effects of small interfering RNA by direct interaction with the Mammalian RNA interference pathway. J Virol. 2011;85:2512–2523. doi: 10.1128/JVI.01160-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Reid S.P., Leung L.W., Hartman A.L. Ebola virus VP24 binds karyopherin alpha1 and blocks STAT1 nuclear accumulation. J. Virol. 2006;80:5156–5167. doi: 10.1128/JVI.02349-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Reid S.P., Valmas C., Martinez O. Ebola virus VP24 proteins inhibit the interaction of NP1-1 subfamily karyopherin alpha proteins with activated STAT1. J. Virol. 2007;81:13469–13477. doi: 10.1128/JVI.01097-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Leroy E.M., Becquart P., Wauquier N. Evidence for Ebola virus superantigen activity. J Virol. 2011;85:4041–4042. doi: 10.1128/JVI.00181-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Geisbert T.W., Hensley L.E., Gibb T.R. Apoptosis induced in vitro and in vivo during infection by Ebola and Marburg viruses. Lab. Invest. 2000;80:171–186. doi: 10.1038/labinvest.3780021. [DOI] [PubMed] [Google Scholar]