

Abstract
Type I interferons (IFN-α and -β) play an important role in the innate host defense against viral infection by inducing antiviral responses. In addition to direct antiviral activities, type I IFN serves as an important link between the innate and adaptive immune response through multiple mechanisms. Therefore, the outcome of a viral infection can be affected by IFN induction and the IFN sensitivity of a virus. North American porcine reproductive and respiratory syndrome virus (PRRSV) field isolates were studied with regard to IFN-α sensitivity and induction in order to understand the role of type I IFN in PRRSV pathogenesis. PRRSV isolates were differentially sensitive to porcine recombinant IFN-α (rIFN-α) and varied in their ability to induce IFN-α in porcine alveolar macrophages (PAM) cultures as measured by a porcine IFN-α specific ELISA on cell culture supernatants. Fifty-two plaques were purified from three PRRSV isolates (numbers 3, 7, and 12) and tested for IFN sensitivity and IFN induction. Plaque-derived populations were composed of heterogeneous populations in terms of IFN-inducing capacity and sensitivity to rIFN-α. When macrophages infected with isolates 3, 7, or 12 were treated with polycytidylic acid (polyI:C), IFN-α production was enhanced. Cells infected with isolate 3 and treated with polyI:C showed the most consistent and strongest enhancement of IFN-α production. It was demonstrated that the relatively low concentrations of IFN-α produced by isolate 3 contributed to the enhanced IFN-α synthesis in response to polyI:C. Isolates 7 and 12 significantly suppressed the enhanced IFN-α production by isolate 3 in polyI:C treated cells. To determine if suppression was at the level of IFN-α transcription, quantitative RT-PCR was performed for IFN-α mRNA and compared to GAPDH and cyclophilin mRNA quantification. However, the relative number of IFN-α transcript copies did not correlate with IFN-α protein levels, suggesting a post-transcriptional mechanism of suppression. In summary, these results demonstrate that PRRSV field isolates differ both in IFN-α sensitivity and induction. Furthermore, a PRRSV field isolate strongly enhance polyI:C-induced IFN-α production in PAM cultures and this priming effect was suppressed by other PRRSV isolates.
Abbreviations: PAM, porcine alveolar macrophages; polyI:C, polycytidylic acid; PRRSV, porcine reproductive and respiratory syndrome virus; rIFN-α, recombinant interferon-α; MOI, multiplicity of infection; p.i., post-infection; TCID50, 50% tissue culture infectious dose
Keywords: Porcine reproductive and respiratory syndrome virus, Alveolar macrophages, Interferon-α
1. Introduction
Porcine reproductive and respiratory syndrome (PRRS) is one of the most economically important diseases of swine. This disease was first detected in the U.S. in 1987 (Keffaber, 1989) and in Europe in 1990 (Wensvoort et al., 1991). The etiologic agent for this disease, PRRS virus (PRRSV) is an enveloped, positive-stranded RNA virus that is a member of the Arteriviridae family, order Nidovirales. The full-length genomic sequence has been determined for PRRSV isolates of both European and North American lineage (Allende et al., 1999, Meulenberg et al., 1993, Nelsen et al., 1999, Shen et al., 2000, Wootton et al., 2000). Molecular analysis of the prototype PRRSV VR-2332 and Lelystad strains (U.S. and European isolates, respectively) has suggested that divergently evolved strains emerged on two continents almost simultaneously, perhaps due to similar changes in swine management practices (Murtaugh et al., 1995, Nelsen et al., 1999). Analysis of partial genomic sequence data for hundreds of PRRSV strains reveals extensive diversity but also well-conserved regions (Andreyev et al., 1997, Meng et al., 1995; reviewed by Meng, 2000).
Disease caused by PRRSV is characterized by severe and sometimes fatal respiratory disease and reproductive failure. Infection with PRRSV also predisposes pigs to infection by bacterial pathogens as well as other viral pathogens (Benfield et al., 1992) and PRRSV is a key etiologic agent of the economically important porcine respiratory disease complex (PRDC). The most consistent pathological lesions caused by PRRSV during acute infection are interstitial pneumonia and mild lymphocytic encephalitis (Halbur et al., 1995, Plagemann, 1996, Rossow et al., 1995, Rossow et al., 1996). Tissue macrophages and monocytes are the major target cells during both acute and persistent infection (Molitor et al., 1997), although pneumocytes and epithelial germ cells of the testes have also been shown to be infected (Sur et al., 1996, Sur et al., 1997). It is important to note that clinical disease caused by PRRSV is highly variable, ranging from mild, subclinical infections to acute deaths of adult animals (Zimmerman et al., 1997). The differences in virulence have been attributed to numerous factors including host genetics, management practices, and virus strain heterogeneity (Halbur et al., 1995, Halbur et al., 1996, Halbur et al., 1998, Keffaber, 1989, Wensvoort, 1993). After the acute phase of PRRSV infection, which is typically characterized by viremia and clinical disease, many pigs fully recover yet carry a low-level viral infection for an extended period of time. Under experimental conditions, persistent infection with PRRSV has been well documented (Albina et al., 1994, Allende et al., 2000, Christopher-Hennings et al., 1995, Horter et al., 2002, Sur et al., 1996, Yoon et al., 1993, Wills et al., 2003). These “carrier” pigs are persistently infected with PRRSV and shed the virus, either intermittently or continuously, and may infect naïve pigs following direct or indirect contact. Most notably, infectious virus has been recovered for up to 157 days post-infection (Wills et al., 1997).
Producers often vaccinate swine against PRRSV with modified-live attenuated strains or killed virus vaccines. However, current vaccines do not provide satisfactory protection, possibly due both to strain variation and inadequate stimulation of the immune system. A protective immune response is possible since it has been demonstrated that previous exposure can provide protection when pigs are challenged with a homologous strain of PRRSV (Lager et al., 1999). However, protective immunity has never been consistently demonstrated for challenge with heterologous strains (Lager et al., 1999, Mengeling et al., 2003).
The ability of PRRSV to routinely establish a persistent infection in pigs coupled with experimental evidence of suboptimal humoral and cellular immunity to PRRSV suggests that the adaptive immune response to PRRSV is often ineffective (reviewed by Murtaugh et al., 2002). The innate immune response, of which type I IFN is a major component, plays a key role in establishment of an effective adaptive immune response. The type I IFN response serves as an important link between the innate and adaptive immune response through multiple mechanisms (reviewed by Biron, 1998). Type I IFN: (i) promotes the development of CD8+ T cell response (reviewed by Boehm et al., 1997), (ii) regulates the expression of many proteins responsible for generating antigenic peptides to be displayed in association with MHC class I (reviewed by York and Rock, 1996), (iii) enhances differentiation of dendritic antigen-presenting cells (Luft et al., 1998), (iv) stimulates the division of memory T cells by inducing IL-15 (Tough et al., 1999), (v) contributes to prolonging the lifespan of activated T cells (Marrack et al., 1999), and (vi) enhances IgG production and down-regulates IgE secretion in B cells (Finkelman et al., 1990, Finkelman et al., 1991).
In addition to a role in the establishment of an adaptive immune response, type I IFN also plays an important role in innate host defenses against viral infection. Type I IFN has been shown to activate or induce the synthesis of several proteins including 2,5-oligoadenylate synthase (OAS), double-stranded RNA-dependent protein kinase (PKR), Mx and ribonuclease L (RNase L) (Vilcek and Sen, 1996), all of which result in induction of an antiviral state (reviewed by Samuel, 2001). Not surprisingly, many viruses have evolved specific mechanisms to counteract the IFN response. For example, the hepatitis B virus ORF-C product and terminal protein (Whitten et al., 1991), the human papillomavirus (HPV)16 E6 and E7 proteins (Park et al., 2000, Ronco et al., 1998), and the influenza A virus NS1 protein (Talon et al., 2000) are viral proteins that have been shown to inhibit IFN synthesis. Some viral proteins, such as the adenoviral E1A (Zhang et al., 1996), Sendai virus C proteins (Komatsu et al., 2000) and Simian Virus 5 protein V (Didcock et al., 1999), block IFN signaling via the Jak-STAT pathways. Additionally, Herpes virus, SV40 and EMCV are known to block IFN-induction of the 2,5-OAS/RNase L system (Vilcek and Sen, 1996).
Better understanding of the type I IFN response following PRRSV infection will provide an important basis for understanding the adaptive immune response to this pathogen. Previous work (Albina et al., 1998a, Buddaert et al., 1998, van Reeth et al., 1999) has demonstrated that European PRRSV strains do not induce a strong IFN-α response, yet they are sensitive to the effects of IFN and can suppress a viral-induced IFN response. In this study, North American field isolates of PRRSV were evaluated with respect to induction, sensitivity and suppression of an in vitro type I IFN response. Differences in IFN phenotype were observed among North American PRRSV field isolates.
2. Materials and methods
2.1. Cell cultures
The PAM cultures were obtained by bronchoalveolar lavage of 2–3 week-old domestic piglets from a PRRSV seronegative herd. The lungs were removed from the piglets immediately after death and RPMI-1640 medium (Life Technologies, Grand Island, NY) was introduced through the main stem bronchi. Bronchoalveolar lavage fluid was centrifuged at 400 × g for 7 min. When cell pellets contained obvious contamination with red blood cells (RBC), PAM cultures were purified by Histopaque-1077 (Sigma Inc., St. Louis, MO) according to manufacturer's directions to remove red blood cell contamination. After centrifugation, cell pellets were resuspended in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS), 2 mM l-glutamine, 0.25 μg/ml fungizone, 100 U/ml penicillin, 10 μg/ml streptomycin sulfate and 5 μg/ml gentamicin (BioWhittaker, Walkersville, MD) and plated at a density of 2–4 × 105 cells/well in a 24-well Primaria plate (Becton Dickinson and Company, Franklin Lakes, NJ). The PAM cultures were confirmed to be PRRSV-negative by RT-PCR before use in subsequent experiments. PAM cultures were incubated for 24 h at 37 °C in a humidified 5% CO2 incubator and washed once with complete RPMI-1640 media before use.
The MARC-145 cell line is a clone of the African green monkey kidney cell line MA-104 which is highly permissive to PRRSV infection (Kim et al., 1993). Cells were cultured and maintained in Dulbecco's Modified Eagle medium (DMEM) supplemented with 10% FBS, 0.25 μg/ml fungizone, 100 U/ml penicillin, 10 μg/ml streptomycin sulfate and 5 μg/ml gentamicin (BioWhittaker Inc., Walkersville, MD) and then held at 37 °C in a humidified 5% CO2 incubator. Swine testicular (ST) cells were used to grow and titrate TGEV. ST cells were grown in DMEM supplemented with 5% heat-inactivated FBS, 2 mM l-glutamine, 0.25 μg/ml fungizone, 100 U/ml penicillin and 10 μg/ml streptomycin sulfate (BioWhittaker Inc., Walkersville, MD). All cells were maintained at 37 °C in a humidified 5% CO2 incubator.
2.2. Viruses
Fifteen PRRSV field isolates, numbered from 1 to 15, were obtained from clinical cases submitted to the University of Missouri's Veterinary Medicine Diagnostic Laboratory. Genomic sequences of open reading frames 2–7 of PRRSV isolates 3, 7, and 12 were submitted to GenBank and assigned accession numbers AY569972 (PRRSV 1530B), AY569974 (PRRSV 8981), and AY569973 (PRRSV 25544), respectively. Virus stocks of PRRSV isolates were prepared in PAM cultures. A low multiplicity of infection (MOI < 0.05) was used to prepare viral stocks, and the third to fifth passages were used for all experiments. Transmissible gastroenteritis virus (TGEV) (Purdue strain) was obtained from the University of Missouri's Veterinary Medicine Diagnostic Laboratory Serology Section.
To purify plaques from PRRSV isolates, confluent MARC-145 cells (Kim et al., 1993) in 6-well plates were infected with PRRSV at a very low MOI (<0.0001). After 2 h, cells were rinsed and overlayed with cell culture medium containing 0.5% agarose and incubated for 3–5 days until plaques were visible. Plaques were harvested by aspiration into a micropipette and diluted into a final volume of 0.2 ml of culture medium and used for experiments without further propagation. A total of 52 plaques were harvested, 16 from isolate 3, 16 from isolate 7 and 20 from isolate 12. The titers of original stocks of PRRSV plaques were analyzed by quantitative real-time RT-PCR using purified RNA from a titered virus stock to establish the standard curve. Titers from isolate 3, 7, and 12 plaques were 4.41 ± 0.099 TCID50/ml (mean ± S.E.M.), 4.81 ± 0.068 TCID50/ml, and 4.77 ± 0.077 TCID50/ml, respectively. The MOI used in experiments with plaque-purified stocks was approximately 0.01 for plaques from isolates 7 and 12, and 0.005 for isolate 3.
2.3. IFN sensitivity assay
PAM cultures in 24-well plates were pre-incubated with 1 or 100 U/ml rIFN-α (R&D Systems Inc., Minneapolis, MN) in cell culture media for 18 h before virus infection. Cells were washed with cell culture media and infected with 15 PRRSV field isolates at MOI = 0.01. After 48 h post-infection (p.i.), supernatants were harvested, frozen and thawed one time. Infectious virus titers in cell culture supernatants were determined by serial dilution in 96-well plates and calculated by the method of Reed and Muench (1938). For IFN-α sensitivity experiments of PRRSV isolates 7 and 12, rIFN-α (1, 10 or 100 U/ml) was added 18 h prior to PRRSV infection (MOI = 1) and virus growth curve experiments were performed. Cell culture supernatants were harvested at 6, 12, 24, 36, 48 h p.i. Infectious virus titers in cell culture supernatants were determined by serial 10-fold dilutions of viral stocks with 50% tissue culture infectious dose (TCID50) titers calculated by the method of Reed and Muench (1938).
2.4. Enzyme-linked immunosorbent assay (ELISA)
IFN-α was measured with a porcine IFN-α specific ELISA by using F17 monoclonal antibody (MAb) and K9 MAb (R&D Systems Inc., Minneapolis, MN) as previously described (Diaz de Arce et al., 1992). MAb K9 was conjugated with horseradish peroxidase (HRP) using a peroxidase labeling kit (Roche Molecular Biochemical, Indianapolis, IN). Flat-bottomed 96-well plates (Fisher Scientific, Houston, TX) were coated overnight at 4 °C with F17 at a concentration of 3 μg/plate in coating buffer (50 mM carbonate buffer, pH 9.5, Sigma Inc., St. Louis, MO). After blocking with 1% non-fat dried milk, 0.05% Tween 20 in phosphate buffered saline (PBS) for 1 h at 37 °C, the plates were washed five times with 0.05% Tween 20 in PBS. Samples (50 μl) were added into each well containing 50 μl of 1% non-fat dried milk, 0.05% Tween 20 in PBS and incubated for 2 h at 37 °C. Following five washes, 100 μl of peroxidase conjugated K9 was added to each well. After 1 h incubation and five washes, 100 μl of substrate solution, tetramethylbenzidine (Sigma Inc., St. Louis, MO), was added to each well. After 30 min, the reaction was stopped with 1N HCl and the optical density was measured at 450 nm by an ELISA plate reader. Quantified recombinant porcine IFN-α (rIFN-α, R&D Systems Inc., Minneapolis, MN) was used as a standard, and IFN-α concentrations were calculated based upon a standard curve. One unit/ml of rIFN-α is equivalent to 26 pg/ml.
2.5. PolyI:C experiments
PAM cultures were infected with PRRSV isolate 3, 7 or 12 at a MOI = 1 for 2 h. After 12 h p.i. the cell culture media was replaced with media containing polyI:C (Sigma Inc., St. Louis, MO) at 20 μg/ml. At 18 h after the initial PRRSV infection, supernatants were collected and the IFN-α concentration was measured by ELISA. For suppression experiments, isolate 3 was inoculated at 2 h p.i. into PAM cultures that were previously infected with either isolate 3, 7 or 12. After 10 h cell culture media was replaced with medium containing polyI:C at 20 μg/ml. Supernatants and cells were harvested at 18 h p.i and stored at −80 °C for subsequent analysis.
PRRSV isolates were UV-inactivated by exposing microcentrifuge tubes containing virus stocks to UV light (wavelength 312 nm) for 20 min. Stocks of UV-inactivated virus were demonstrated to be completely non-infectious for PAM cultures. PAM cultures were inoculated with either PRRSV or UV-inactivated PRRSV at MOI = 1, incubated for 2 h then washed with cell culture media.
Polyclonal antibody against porcine IFN-α (50 neutralizing units/ml, R&D Systems Inc., Minneapolis, MN) was added to cell culture media at the time of infection (2 h) and again after infection (10 h). At 12 h p.i. cell culture medium was replaced with media containing polyI:C (20 μg/ml). After an additional 6 h incubation, cell culture media was collected and analyzed for IFN-α production by using porcine specific ELISA.
2.6. Cell viability assays
At 12 h p.i. with isolate 3, 7 or 12 (MOI = 1), PAM cells were harvested and resuspended in PBS at a concentration of 106 cells/ml and assessed for cell viability. Propidium iodide (Sigma Inc., St. Louis, MO), which will positively stain dead cells with a disturbed cellular membrane, was added into 0.5 ml of cell suspension at 50 μg/ml and incubated at room temperature. After 30 min, cell viability was analyzed by flow cytometry. A second aliquot of infected cells with isolate 3, 7 or 12 was stained with 0.4% trypan blue (Bio-Whittaker, Walkersville, MD), and then quantified microscopically with a hemocytometer to determine the total number of cells and the number of dead cells, i.e., those retaining trypan blue. The CellTiter 96® AQueous One Solution Cell Proliferation Assay (Promega Corp., Madison, WI) was performed as described by the manufacturer. Briefly, 20 μl of the CellTiter 96 solution reagent was added into each well of 96-well plate and the plate was incubated for 3 h at 37 °C in a humidified, 5% CO2 incubator. Absorbance was measured at 490 nm. The OD measured at 490 nm represents the amount of tetrazolium dye (MTS)-to-formazan conversion, which represents the number of viable cells.
2.7. RNA extraction
Extraction of RNA from samples of IFN suppression experiments was performed using Trizol (Invitrogen, Carlsbad, CA) and the Nucleospin RNA II kit (BD Biosciences Inc., Palo Alto, CA) with DNAse I digestion performed directly on the spin column according to the manufacturer's instructions. Extractions of the purified competitor RNA were performed using the Qiagen RNAEasy kit (Qiagen Inc., Valencia, CA).
2.8. RNA quantification standards
Heterologous competitor RNA for quantification of swine IFN-α, cyclophilin or GADPH was synthesized using the respective real-time RT-PCR primer sequences in a methodology previous described (Kleiboeker, 2003). The concentration of purified competitor RNA was estimated by measuring the absorbance at 260 nm and the purity was assessed by determining the ratio of absorbance at 260 nm to the absorbance at 280 nm. Samples were considered to be relatively pure and suitable for use as quantification standards if the ratio was >2.0. Following purification the RNA was serially diluted in RNase-free dH2O and stored as aliquots at −80 °C. The number of molecules of competitor RNA/μl was estimated based on the RNA concentration and the molecular weight of the transcript.
2.9. Quantitative (TaqMan) RT-PCR
Amplification of 2 μl RNA was performed using the Qiagen QuantiTect Probe RT-PCR kit (Qiagen Inc., Valencia, CA) with thermocycling and detection performed in a Stratagene Mx4000 (Stratagene Inc., La Jolla, CA). Samples were analyzed in triplicate. Thermocycling conditions were: 50 °C (30 min), 95 °C (15 min), followed by 40 cycles of denaturation (94 °C, 15 s) and annealing/extension (60 °C, 60 s). GenBank accession numbers of sequences used for porcine IFN-α, porcine cyclophilin, and porcine GAPDH real-time RT-PCR assays were NM214393, AY008846, and AF141959, respectively. Primers used for 5′-exonuclease (TaqMan) amplification of swine IFN-α were (forward, position 195–212) 5′-TCTCATGCACCAGAGCCA-3′, (reverse, position 269–286) 5′-CCTGGACCACAGAAGGGA-3′, and for amplification of swine cyclophilin were (forward, position 189–206) 5′-ATGGCACTGGTGGCAAGT-3′, (reverse, position 251–268) 5′-GATGCCAGGACCCGTATG-3′. Primers used for 5′-exonuclease (TaqMan) amplification of for swine GAPDH were (forward, position 435–452) 5′-TGCCCAGAACATCATCCC-3′ and (reverse, position 476–493) 5′-GGATGACCTTGCCCACAG-3′. All oligonucleotide primers were used at a final concentration of 0.3 μM. The dual-labeled probe used for detection of IFN-α transcript was 5′-6-FAM-CTTGAGCCTTCTGGACCTGGTTGC-BHQ1-3′ (position 242–265), for detection of swine cyclophilin transcript was 5′-6-FAM-CATCTATGGAGAGAAATTTGATGATGAGA-BHQ1-3′ (position 207–236), and for detection of swine GAPDH transcript was 6-FAM-CTTCTACCGGCGCTGCCAAG-BHQ1-3′ (position 455–474). The dual-labeled probe used for detection of heterologous swine IFN-α, cyclophilin, or GAPDH competitor RNA was: 5′-HEX-TGTGCTGCAAGGCGATTAAGTTGGGT-BHQ2-3′. Each probe was used at a final concentration of 0.2 μM. All oligonucleotide primers and probes were synthesized by Integrated DNA Technologies Inc. (Coralville, IA). The relative copy numbers of IFN-α mRNA compared to cyclophilin or GAPDH were determined as previously described (Stordeur et al., 2002).
2.10. Statistical analysis
The Student's t-test was used for the statistical analyses. P-values of less than 0.05 were considered statistically significant.
3. Results
3.1. PRRSV field isolates differ in sensitivity to rIFN-α
To determine the sensitivity of North American PRRSV field isolates to exogenous rIFN-α, viral replication of 15 field isolates was assessed in PAM cultures that were pretreated with 1 or 100 U rIFN-α/ml cell culture media (Fig. 1 ). For most isolates, viral titers were either unchanged or slightly decreased by the addition of 1 U/ml of rIFN-α. However, replication of one isolate (number 15) was reduced by >1 log10 TCID50/ml at this dose. All isolates demonstrated greater declines in replication following pretreatment with 100 U/ml of rIFN-α, with titers for 5 of the 15 isolates declining to the detectable limits of the assay. This experiment was repeated twice independently and results were consistent between experiments.
Fig. 1.
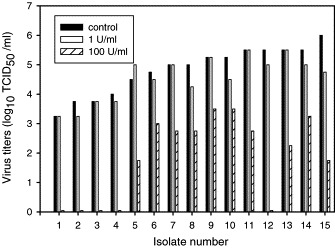
IFN-α sensitivity of PRRSV field isolates. PAM cultures were pretreated with 1 or 100 U/ml rIFN-α. After 18 h the macrophages were washed and infected with 15 PRRSV field isolates (MOI = 0.01). After 48 h p.i. supernatants were harvested, frozen and thawed one time and infectious virus titers were analyzed in PAM cultures. This experiment was repeated twice with consistent results.
To further characterize differences in sensitivity to rIFN-α, two isolates, numbers 7 and 12, were selected for use in a dose-response growth curve experiment. A representative experiment from 4 independent experiments is shown (Fig. 2 ). A dose-dependent reduction in viral titers was noted for both isolates. When PAM cultures were pretreated with rIFN-α at 1, 10 and 100 U/ml there was a significant (P < 0.05) reduction in viral titer from 12 to 36 h p.i. for isolate 12 whereas for isolate 7 only 10 and 100 U/ml of rIFN-α resulted in a significant reduction in viral yield. For PAM cultures that were pretreated with 1 U/ml of rIFN-α, titers of isolate 12 were 30-fold lower than isolate 7 at 12 h p.i. compared to the untreated controls for each isolate. At 12, 24, 36 and 48 h p.i. the titers of isolate 12 in 10 U/ml rIFN-α-treated cells were approximately 2.0–3.0 log10 50% tissue culture infectious dose (TCID50)/ml lower than in untreated cells. In contrast, the titers for isolate 7 were reduced by about 1.0–1.75 log10 TCID50/ml. When PAM cultures were pretreated with 100 U rIFN-α/ml, virus yields of isolate 12 were inhibited throughout the time course while virus yields of isolate 7 increased through the time course and reached titers of the untreated control at 48 h p.i. Taken together, these results demonstrate that North American field isolates have different sensitivities to exogenous rIFN-α in PAM cultures.
Fig. 2.
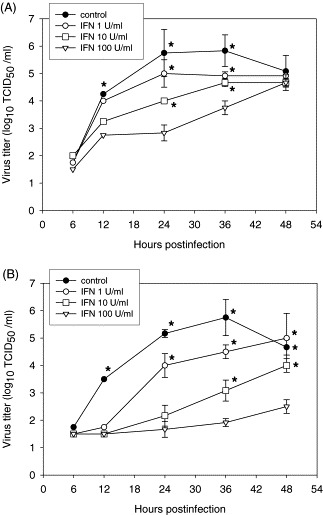
IFN-α sensitivity of PRRSV isolates 7 (A) and 12 (B). rIFN-α (1, 10 or 100 U/ml) was added 18 h prior to PRRSV infection (MOI = 1) and virus growth curve experiments were performed. Infectious virus titers were analyzed in PAM cultures. A representative example of four independent experiments is shown. *, Significant difference compared with control (P < 0.05).
3.2. Plaque-derived populations from field isolates differ in IFN-α sensitivity and induction
To investigate whether PRRSV isolates are composed of heterogeneous populations with measurable differences in rIFN-α sensitivity and IFN-α induction, plaque-derived populations of field isolates were prepared and tested. As shown in Fig. 3 , plaque-purified populations of all three isolates demonstrated a range in sensitivity to exogenous rIFN-α. For isolates 7 and 12, the range of reduction in viral titers was approximately 0.7–3.0 log10 TCID50/ml. For isolate 3, the range of reduction in viral titers was 0–3.0 log10 TCID50/ml. In preliminary experiments, isolate 3 was the most consistent IFN-α inducer compared to other field isolates tested. More than 2 U/ml of IFN-α was consistently synthesized in cells infected with isolate 3. In contrast IFN-α was typically undetectable in cultures infected with other isolates, although very low concentrations were detected in some experiments. Plaque-derived populations of field isolates were also characterized with respect to IFN-α induction (Fig. 4 ). While individual plaques from all three isolates induced relatively low concentrations of IFN-α compared to TGEV, IFN-α production varied among plaques from each isolate. IFN-α induced by plaque-derived populations from isolates 3, 7 and 12 were 2.31 ± 1.41, 0.3 ± 0.22, and 1.07 ± 0.57, respectively (mean ± S.D.). Plaques from isolate 3 induced a higher concentration of IFN-α than plaques of the other two isolates (P < 0.01).
Fig. 3.
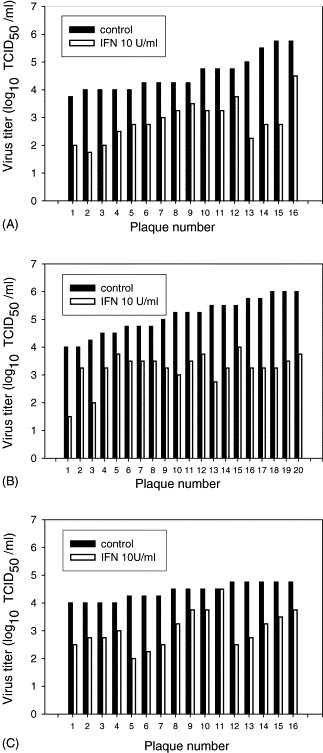
IFN-α sensitivity of PRRSV plaque-derived populations. Viral stocks of PRRSV field isolates 7 (A), 12 (B) and 3 (C) were plaque-purified on MARC-145 cells and 20, 16 and 16 plaques, respectively, were selected and used in experiments without further amplification in cell culture. rIFN-α (10 U/ml) pretreated PAM cultures were infected with PRRSV plaques (40 μl/well) and at 48 h p.i. supernatants were harvested and infectious virus titers were measured in PAM cultures.
Fig. 4.
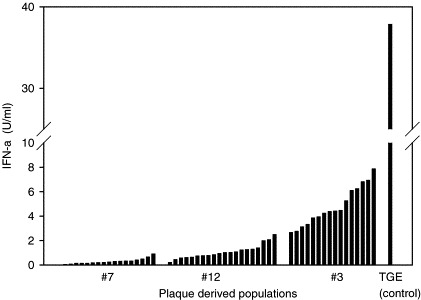
IFN-inducing capacity of PRRSV plaque-derived populations. PAM cultures were inoculated with 40 μl/well of each plaque and supernatants were harvested after 48 h and analyzed for IFN-α concentration using a porcine IFN-α-specific ELISA. Plaques from isolate 3 induced a higher mean concentration of IFN-α than plaques of the other two isolates (P < 0.01). One unit/ml of rIFN-α is equivalent to 26 pg/ml.
3.3. Effect of dsRNA on IFN production in PRRSV infected PAM cultures
To investigate the ability of PRRSV field isolates to suppress IFN-α induction by an IFN inducer, PAM cultures were infected with PRRSV then treated with polyI:C (20 μg/ml) after 12 h. In this experiment, no measurable amount of IFN-α was induced by PRRSV infection alone with any of the three PRRSV isolates (Fig. 5A). In the absence of PRRSV, polyI:C failed to induce IFN-α at 6 h, although IFN-α was induced at later time points. However, in the experiments shown samples from later time points were not used in order to avoid the influence of PRRSV-induced cytopathic effect on IFN-α production. When PRRSV-infected cells were treated with polyI:C, IFN-α production was consistently enhanced to the greatest extent for isolate 3. Therefore it was tested whether isolates 7 or 12 could suppress IFN-α induced by a combination of isolate 3 infection and polyI:C treatment of PAM cultures. Both isolates 7 and 12 significantly (P < 0.05) decreased IFN-α production by 53% and 40%, respectively. In contrast, IFN-α production was significantly (P < 0.05) increased by 37% in isolate 3 superinfected cells.
Fig. 5.
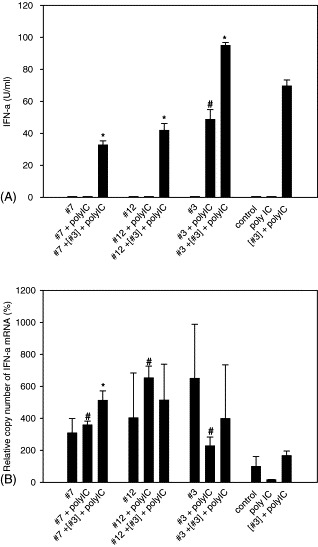
Effect of PRRSV on IFN-α induced by polyI:C treated PAM cultures. PAM cultures infected with each isolate were infected with isolate 3 at 2 h p.i. (expressed as [#3]) and then treated with polyI:C at 12 h p.i. Supernatants were collected at 18 h p.i. (A). IFN-α protein was measured by ELISA. One unit/ml of rIFN-α is equivalent to 26 pg/ml. (B) Quantitative real-time RT-PCR was performed by using total RNA extracted from PAM cultures. Results are expressed as relative copy numbers of IFN-α mRNA using cyclophilin as an internal control. Values are shown as the means ± S.D. from duplicate wells and represent at least two independent experiments. *, Significant difference compared with [#3] infected, polyI:C treated cells (P < 0.05). #, Significant difference compared with polyI:C only treated cells (P < 0.05). No significant differences were detected in the copy numbers of IFN-α mRNA among PRRSV isolate infected cells and control cells.
To demonstrate that suppression of IFN was not due to decreased cell viability, three cell viability assays (propidium iodide (PI) staining, trypan blue exclusion, and cell titer 96® AQueous cell proliferation and viability assay) were performed. When cell viability was analyzed by flow cytometry after PI staining, cell viability of PRRSV-infected cells at 12 h p.i. was 96.10%, 95.87%, and 95.52% for isolates 3, 7, and 12, respectively. Uninfected cells demonstrated 95.20% viability. This result was in excellent agreement with the other two independent assays used to assess cell viability.
To determine the effect of PRRSV on IFN-α mRNA synthesis, quantitative real-time RT-PCR was performed with samples from the suppression experiments. Normalization of IFN-α mRNA copy number was performed both with cyclophilin and GAPDH transcript, with equivalent results obtained. As shown in Fig. 5B, PRRSV isolates induced variable quantities of IFN-α mRNA in PAM cultures. The quantity of IFN-α mRNA did not correlate with the IFN-α protein level detected by ELISA (Fig. 5A). Consistently, IFN-α mRNA synthesis was increased in PRRSV-infected PAM cultures (P < 0.05), despite a lack of detectable IFN-α protein. As a control, real-time PCR without the RT step was performed to determine if levels of contaminating genomic DNA following DNAse I treatment affected real-time RT-PCR quantification. Relative amounts of DNA in each sample were always less than 2% of the total signal, demonstrating that the signal analyzed was from RNA rather than DNA.
UV-inactivated PRRSV stocks were tested to determine whether virus binding to the cells is responsible for enhanced IFN-α production by polyI:C in cells infected with isolate 3 (Fig. 6 ). UV-inactivation of virus reduced IFN-α in these cells to 11.7% of control values obtained using fully infectious virus (P < 0.05). Thus, virus binding to the cells alone was not sufficient to amplify IFN-α synthesis by polyI:C.
Fig. 6.
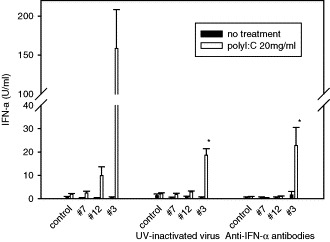
Effect of UV-inactivated virus and anti-IFN-α antibodies on IFN-α production. PAM cultures were infected with either PRRSV or UV-inactivated PRRSV at MOI = 1.0 for 2 h. Polyclonal antibody against porcine IFN-α (50 neutralizing units/ml) was added into cell cultures at the time of infection. At 12 h p.i. cell culture media was replaced with media containing polyI:C (20 μg/ml). After additional 6 h incubation, cell culture media was collected and analyzed for IFN-α production by using porcine specific ELISA. One unit/ml of rIFN-α is equivalent to 26 pg/ml. *, Significant difference compared to polyI:C treated and isolate 3 infected cells (P < 0.05).
To determine if IFN-α secreted after initial infection with isolate 3 was responsible for enhanced IFN-α production in respond to polyI:C, neutralizing antibody against porcine IFN-α was used (Fig. 6). Neutralizing antibody against pig IFN-α reduced IFN-α production by polyI:C in cells infected with isolate 3–14% of control values (P < 0.05) when added into cell cultures at a concentration of 50 neutralizing units/ml prior to polyI:C treatment. Pretreatment of macrophages with low concentration of rIFN-α (1 U/ml) also enhanced IFN-α production from 3 U/ml to more than 500 U/ml by polyI:C. Thus these experiments suggest that even low concentrations of IFN-α induced by isolate 3 are sufficient to strongly enhance the IFN response to polyI:C treatment.
4. Discussion
The type I IFN response plays an important role in host defenses against viral infection by performing immunoregulatory functions that link innate and adaptive immune responses by multiple mechanisms (reviewed by Biron, 1998) as well as through direct antiviral effects. In this study, North American PRRSV field isolates were evaluated with respect to induction, sensitivity and suppression of an in vitro IFN response in PAM cultures. Our results show that these isolates are sensitive to rIFN-α in a dose-dependent manner, but are generally poor inducers of IFN in vitro. Notably the extent of the sensitivity and induction of IFN not only differed between PRRSV isolates, but also showed variation among the plaque-derived populations within each isolate. Two of the field isolates studied were able to suppress the IFN-α production of cells dually-treated with polyI:C and a third North American PRRSV isolate. While considerable variation was noted for the levels of IFN-α measured between replicates, most likely due to the variability of alveolar macrophage function among pigs (du Manoir et al., 2002), the trends observed were consistent between experiments. Taken together, these results demonstrate that North American PRRSV isolates differ in the ability to induce or suppress IFN-α, which may contribute to both virulence differences commonly observed among PRRSV infections and the failure of PRRSV infection to consistently induce a rapid and robust sterilizing immune response.
The role of type I IFN in PRRSV pathogenesis has been addressed by previous research (Albina et al., 1998a, Buddaert et al., 1998, van Reeth et al., 1999), however this work was performed with two isolates (Lelystad and SDRPI) of the European PRRSV lineage, which is quite divergent (∼60% sequence identity) compared to North American strains. In the present study, it was demonstrated that North American PRRSV isolates were sensitive to rIFN-α in a dose-dependent manner and were poor IFN-α inducers in vitro. These results are consistent with previous studies using European PRRSV isolates (Albina et al., 1998a, Buddaert et al., 1998, van Reeth et al., 1999). Similar to results with type I IFN, North American PRRSV replication was also blocked by a type II IFN (IFN-γ) and virus replication was restored by the addition of 2-aminopurine, an inhibitor of PKR (Rowland et al., 2001).
Biological, antigenic, pathogenic, and genetic variation among PRRSV field isolates has been well documented (reviewed by Meng, 2000). The data presented herein demonstrated that North American PRRSV field isolates were differentially sensitive to the in vitro antiviral effects of rIFN-α and the use of plaque-derived populations from these field isolates demonstrated that closely related variants of PRRSV may differ in their IFN responses. Similarly, it has been shown that the sensitivity of reovirus, hepatitis C virus and lymphocytic choriomeningitis virus (LCMV) to type I IFN antiviral activity differs among viral strains (Enomoto et al., 1996, Moskophidis et al., 1994; reviewed by Samuel, 1998). A previous study suggested that significant differences of IFN-inducing capabilities could be a quasispecies marker of vesicular stomatitis virus (VSV) (Marcus et al., 1998). In contrast to the results presented herein, a study using the European PRRSV strains SDRPI and SDRPII found no strain differences in IFN-α sensitivity or induction, despite differences in clinical virulence (Albina et al., 1998a). Previous experimental work has demonstrated evidence of a relationship between IFN phenotypes and virulence for other viruses. For example, virulent measles virus induces lower levels of IFN than attenuated strains and IFN-resistant strains can establish persistent infections of the central nervous system (Carrigan and Knox, 1990). It has also been shown that the capability of non-cytopathic bovine viral diarrhea virus to establish persistent infections in the early fetus is related to its ability to suppress type I IFN synthesis (Charleston et al., 2001). Future in vivo studies will be needed to determine if the IFN phenotypes described in the present study are related to PRRSV virulence or persistence.
In the present study it was shown that polyI:C treatment of PRRSV-infected cells resulted in greatly enhanced IFN-α production, especially in PAM cultures infected with PRRSV isolate 3. Similar phenomenons have been reported by other studies. IFN-inducing Listeria monocytogenes (Havell, 1986) and type I IFN pretreatment (Rosztoczy and Megyeri, 1989) enhanced IFN production by polyI:C, Sendai virus, or endotoxin. A plausible mechanism of this enhancement is by positive feedback regulation of type I IFN synthesis, in which a weak IFN-α/β signaling contributes to the enhancement of IFN-α/β synthesis due to the accumulation of transcription factor IRF-7 (reviewed by Taniguchi and Takaoka, 2001). This positive feedback loop of IFN synthesis could be efficiently induced by a small amount of IFN-α produced during the first 12 h after isolate 3 infection, resulting in amplification of the IFN-α response to polyI:C. Consistent with this possibility was the observation that neutralizing antibody against IFN-α remarkably reduced IFN-α production by polyI:C in cells infected with isolate 3, and that 1 U/ml of rIFN-α added to poly I:C treated cells greatly increased the levels of IFN-α detected. Toll-like receptor 3 (TLR3) which recognizes polyI:C (Alexopoulou et al., 2001) could be also involved in the enhancement of IFN production. Measles virus strains with IFN-β inducing properties up-regulated the expression of TLR3 resulting in enhanced IFN-β production in response to polyI:C (Tanabe et al., 2003). Therefore IFN produced by isolate 3 may act through a similar mechanism to up-regulate TLR3 expression followed by increased IFN-α production after polyI:C treatment.
For viruses such as Sendai virus and VSV, low IFN-inducing strains have been shown to suppress IFN induction of high IFN-inducing strains of the same virus (Marcus et al., 1998, Mattana and Viscomi, 1998). In the present study, the levels of PRRSV IFN induction were typically too low to use in a similarly designed suppression experiment. Therefore the suppressive effect of PRRSV isolates 7 and 12 on IFN-α production was evaluated in cells infected with isolate 3 and treated with polyI:C, and significant suppression of IFN-α was demonstrated. European PRRSV was also shown to suppress IFN-α production by TGEV, both in vitro and in vivo (Albina et al., 1998a). However, this result was not corroborated in vivo by another group using the closely related porcine respiratory coronavirus (PRCV) (Buddaert et al., 1998). Microarray experiments have indicated that polyI:C, IFN and viruses induced different subsets of the same cellular genes by activating diverse signaling pathways (Geiss et al., 2001). Since IFN-α production was enhanced by the addition of polyI:C, it is likely that PRRSV isolates 7 and 12 suppress an IFN-α synthesis pathway activated by isolate 3 and polyI:C but not polyI:C alone. It has been shown that viruses which are resistant to the effects of IFN or can suppress IFN production have increased opportunities to spread before activation of the adaptive immune response (Moskophidis et al., 1994, Naniche et al., 2000). Mouse models of viral infection have clearly demonstrated that disrupting the IFN response leads to higher levels of viral replication. Mice deficient in an IFN receptor exhibited increased susceptibility to LCMV (van den Broek et al., 1995) and extensive spread of LCMV infection correlated with an isolate's relative resistance to IFN-α/β and IFN-γ (Moskophidis et al., 1994).
Viruses can suppress IFN-α synthesis both at the transcriptional and post-transcriptional level. Many viruses prevent mRNA synthesis of IFN-α through various mechanisms but VSV inhibits IFN-α synthesis by shutting down the host protein synthesis (Ahmed et al., 2003). The present study showed that PRRSV stimulated IFN-α mRNA synthesis but inhibited IFN-α protein expression. This result suggests that the mechanism by which PRRSV isolates suppress IFN-α production is mediated by generally inhibiting host protein expression or by specifically inhibiting IFN-α protein expression. Half-life times of IFN mRNA and protein could be different and that might be responsible for differences between mRNA and protein levels of IFN. However, increases in IFN-α mRNA expression for PRRSV infected PAM cultures were consistently detected at multiple times post-infection, yet for cells infected with isolate 7 or 12 very low to undetectable levels of IFN-α protein were typically observed at all times p.i. Thus, although the mechanisms of IFN-α suppression by PRRSV remain to be elucidated, data presented herein show that it is likely to be post-transcriptional in nature.
While the immune response against PRRSV is poorly understood, experimental work has demonstrated that the adaptive immune response of PRRSV-infected pigs is generally ineffective (Horter et al., 2002, Mengeling et al., 1996; reviewed by Murtaugh et al., 2002, Wills et al., 1997, Wills et al., 2003). Specific evidence of this includes a slow neutralizing antibody response, which is typically not detected until 3 weeks p.i. (Albina et al., 1998b) and does not reach maximum levels until 10–18 weeks p.i. (Nelson et al., 1994, Yoon et al., 1995). Wide variation has been shown in individual animals, with some studies demonstrating that many infected animals fail to develop a neutralizing antibody response (Loemba et al., 1996, Nelson et al., 1994). While the importance of a cell-mediated response for protection against PRRSV has not been debated, the effectiveness of this response during the early phases of disease also appears to be suboptimal (reviewed by Murtaugh et al., 2002). For example, the T-cell response to PRRSV is weak and transient and cannot be restimulated for more than 4 weeks post-challenge (Meier et al., 2000, Molitor et al., 1997). Additionally, IFN-γ responses of PRRSV-infected pigs were relatively weak and increased slowly in comparison to pseudorabies virus infected pigs (Meier et al., 2003). Although the precise mechanisms for the ineffective nature of the adaptive immune response to PRRSV is not known, PRRSV evasion of the innate immune responses, such as the type I IFN response, may set the stage for subsequent subversion of the adaptive immune response.
In conclusion, the data presented in this study demonstrates that North American PRRSV field isolates are distinctively different in their in vitro IFN phenotypes, a difference that could contribute to the variable clinical signs and pathology observed in PRRSV infections. Moreover, the lack of IFN response following infection with PRRSV could be involved in the establishment of persistent infections or secondary bacterial and viral infection, both of which are characteristic of PRRSV infections in swine. However, in vivo studies are required to fully define the role of type I IFN in PRRSV pathogenesis and virulence. The mechanisms involved in enhancement of IFN-α by PRRSV infected and polyI:C treated cells remains to be elucidated. Additionally, it will be important to determine whether PRRSV generally inhibits host protein synthesis or specifically down-regulates IFN expression and to identify the viral proteins that contribute to the different IFN response among PRRSV isolates. Knowledge about the role of IFN in PRRSV pathogenesis will provide new hypotheses for improved vaccines or methods to modulate the immune response in order to more effectively control PRRSV infection.
References
- Ahmed M., McKenzie M.O., Puckett S., Hojnacki M., Poliquin L., Lyles D.S. Ability of the matrix protein of vesicular stomatitis virus to suppress beta interferon gene expression is genetically correlated with the inhibition of host RNA and protein synthesis. J. Virol. 2003;77:4646–4657. doi: 10.1128/JVI.77.8.4646-4657.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albina E., Madec F., Cariolet R., Torrison J. Immune response and persistence of the porcine reproductive and respiratory syndrome virus in infected pigs and farm units. Vet. Rec. 1994;134:567–573. doi: 10.1136/vr.134.22.567. [DOI] [PubMed] [Google Scholar]
- Albina E., Carrat C., Charley B. Interferon-α response to swine arterivirus (PoAV), the porcine reproductive and respiratory syndrome virus. J. Interferon Cytokine Res. 1998;18:485–490. doi: 10.1089/jir.1998.18.485. [DOI] [PubMed] [Google Scholar]
- Albina E., Piriou L., Hutet E., Cariolet R., L’Hospitalier R. Immune responses in pigs infected with porcine reproductive and respiratory syndrome virus (PRRSV) Vet. Immunol. Immunopathol. 1998;61:49–66. doi: 10.1016/S0165-2427(97)00134-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexopoulou L., Holt A.C., Medzhitov R., Flavell R.A. Recognition of double-stranded RNA and activation of NF-kappaB by toll-like receptor 3. Nature. 2001;413:732–738. doi: 10.1038/35099560. [DOI] [PubMed] [Google Scholar]
- Allende R., Lewis T.L., Lu Z., Rock D.L., Kutish G.F., Ali A., Doster A.R., Osorio F.A. North American and European porcine reproductive and respiratory syndrome viruses differ in non-structural protein coding regions. J. Gen. Virol. 1999;80:307–315. doi: 10.1099/0022-1317-80-2-307. [DOI] [PubMed] [Google Scholar]
- Allende R., Laegreid W.W., Kutish G.F., Galeota J.A., Wills R.W., Osorio F.A. Porcine reproductive and respiratory syndrome virus: description of persistence in individual pigs upon experimental infection. J. Virol. 2000;74:10834–10837. doi: 10.1128/jvi.74.22.10834-10837.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andreyev V.G., Wesley R.D., Mengeling W.L., Vorwald A.C., Lager K.M. Genetic variation and phylogenetic relationships of 22 porcine reproductive and respiratory syndrome virus (PRRSV) field strains based on sequence analysis of open reading frame 5. Arch. Virol. 1997;142:993–1001. doi: 10.1007/s007050050134. [DOI] [PubMed] [Google Scholar]
- Benfield D.A., Nelson E., Collins J.E., Harris L., Goyal S.M., Robinson D., Christianson W.T., Morrison R.B., Gorcyca D., Chladek D. Characterization of swine infertility and respiratory syndrome (SIRS) virus isolate ATCC VR-2332. J. Vet. Diagn. Invest. 1992;4:127–133. doi: 10.1177/104063879200400202. [DOI] [PubMed] [Google Scholar]
- Biron C.A. Role of early cytokines, including alpha and beta interferons (IFN-alpha/beta), in innate and adaptive immune responses to viral infections. Semin. Immunol. 1998;10:383–390. doi: 10.1006/smim.1998.0138. [DOI] [PubMed] [Google Scholar]
- Boehm U., Klamp T., Groot M., Howard J.C. Cellular responses to interferon-gamma. Annu. Rev. Immunol. 1997;15:749–795. doi: 10.1146/annurev.immunol.15.1.749. [DOI] [PubMed] [Google Scholar]
- Buddaert W., Van Reeth K., Pensaert M. In vivo and in vitro interferon (IFN) studies with the porcine reproductive and respiratory syndrome virus (PRRSV) Adv. Exp. Med. Biol. 1998;440:461–467. doi: 10.1007/978-1-4615-5331-1_59. [DOI] [PubMed] [Google Scholar]
- Carrigan D.R., Knox K.K. Identification of interferon-resistant subpopulations in several strains of measles virus: positive selection by growth of the virus in brain tissue. J. Virol. 1990;64:1606–1615. doi: 10.1128/jvi.64.4.1606-1615.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charleston B., Fray M.D., Baigent S., Carr B.V., Morrison W.I. Establishment of persistent infection with non-cytopathic bovine viral diarrhoea virus in cattle is associated with a failure to induce type I interferon. J. Gen. Virol. 2001;82:1893–1897. doi: 10.1099/0022-1317-82-8-1893. [DOI] [PubMed] [Google Scholar]
- Christopher-Hennings J., Nelson E.A., Hines R.J., Nelson J.K., Swenson S.L., Zimmerman J.J., Chase C.C.L., Yaeger M.J., Benfield D.A. Persistence of porcine reproductive and respiratory syndrome virus in serum and semen of adult boars. J. Vet. Diagn. Invest. 1995;7:456–464. doi: 10.1177/104063879500700406. [DOI] [PubMed] [Google Scholar]
- Diaz de Arce H., Artursson K., L’Haridon R., Perers A., La Bonnardiere C., Alm G.V. A sensitive immunoassay for porcine interferon-alpha. Vet. Immunol. Immunopathol. 1992;30:319–327. doi: 10.1016/0165-2427(92)90102-v. [DOI] [PubMed] [Google Scholar]
- Didcock L., Young D.F., Goodbourn S., Randall R.E. The V protein of simian virus 5 inhibits interferon signaling by targeting STAT1 for proteasome-mediated degradation. J. Virol. 1999;73:9928–9933. doi: 10.1128/jvi.73.12.9928-9933.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- du Manoir J.M., Albright B.N., Stevenson G., Thompson S.H., Mitchell G.B., Clark M.E., Caswell J.L. Variability of neutrophil and pulmonary alveolar macrophage function in swine. Vet. Immunol. Immunopathol. 2002;89:175–186. doi: 10.1016/s0165-2427(02)00207-6. [DOI] [PubMed] [Google Scholar]
- Enomoto N., Sakuma I., Asahina Y., Kurosaki M., Murakami T., Yamamoto C., Ogura Y., Izumi N., Marumo F., Sato C. Mutations in the nonstructural protein 5A gene and response to interferon in patients with chronic hepatitis C virus 1b infection. N. Engl. J. Med. 1996;334:77–81. doi: 10.1056/NEJM199601113340203. [DOI] [PubMed] [Google Scholar]
- Finkelman F.D., Holmes J., Katona I.M., Urban J.F., Jr., Beckmann M.P., Park L.S., Schooley K.A., Coffman R.L., Mosmann T.R., Paul W.E. Lymphokine control of in vivo immunoglobulin isotype selection. Annu. Rev. Immunol. 1990;8:303–333. doi: 10.1146/annurev.iy.08.040190.001511. [DOI] [PubMed] [Google Scholar]
- Finkelman F.D., Svetic A., Gresser I., Snapper C., Holmes J., Trotta P.P., Katona I.M., Gause W.C. Regulation by interferon alpha of immunoglobulin isotype selection and lymphokine production in mice. J. Exp. Med. 1991;174:1179–1188. doi: 10.1084/jem.174.5.1179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geiss G., Jin G., Guo J., Bumgarner R., Katze M.G., Sen G.C. A comprehensive view of regulation of gene expression by double-stranded RNA-mediated cell signaling. J. Biol. Chem. 2001;276:30178–30182. doi: 10.1074/jbc.c100137200. [DOI] [PubMed] [Google Scholar]
- Halbur P., Paul P.S., Frey M.L., Landgraf J., Eernisse K., Meng X-J., Lum M.A., Andrews J.J., Rathje J.A. Comparison of the pathogenicity of two US Porcine reproductive and respiratory syndrome virus isolates with that of the Lelystad virus. Vet. Pathol. 1995;22:648–660. doi: 10.1177/030098589503200606. [DOI] [PubMed] [Google Scholar]
- Halbur P.G., Paul P.S., Meng X-J., Lum M.A., Rathje J.A. Comparative pathogenicity of nine US porcine reproductive and respiratory syndrome virus (PRRSV) isolates in a 5 week-old cesarian-derived colostrums-deprived pig model. J. Vet. Diagn. Invest. 1996;8:11–20. doi: 10.1177/104063879600800103. [DOI] [PubMed] [Google Scholar]
- Halbur P.G., Rothschild M.F., Thacker B.J., Meng X-J., Paul P.S., Bruna J.D. Differences in susceptibility of Duroc, Hampshire, and Meishan pigs to infection with a high virulence strain (VR2385) of porcine reproductive and respiratory syndrome virus (PRRSV) J. Anim. Breed. Genet. 1998;115:181–189. [Google Scholar]
- Havell E.A. Augmented induction of interferons during Listeria monocytogenes infection. J. Infect. Dis. 1986;153:960–969. doi: 10.1093/infdis/153.5.960. [DOI] [PubMed] [Google Scholar]
- Horter D.C., Pogranichniy R.M., Chang C.C., Evans R.B., Yoon K.J., Zimmerman J.J. Characterization of the carrier state in porcine reproductive and respiratory syndrome virus infection. Vet. Microbiol. 2002;86:213–228. doi: 10.1016/s0378-1135(02)00013-5. [DOI] [PubMed] [Google Scholar]
- Keffaber K.K. Reproductive failure of unknown etiology. Am. Assoc. Swine Prac. Newsl. 1989;1:1–9. [Google Scholar]
- Kleiboeker S.B. Applications of competitor RNA in diagnostic RT-PCR. J. Clin. Microbiol. 2003;41:2055–2061. doi: 10.1128/JCM.41.5.2055-2061.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim H.S., Kwang J., Yoon I.J., Joo H.S., Frey M.L. Enhanced replication of porcine reproductive and respiratory syndrome (PRRS) virus in a homogenous subpopulation of MA-104 cells. Arch. Virol. 1993;133:477–483. doi: 10.1007/BF01313785. [DOI] [PubMed] [Google Scholar]
- Komatsu T., Takeuchi K., Yokoo J., Tanaka Y., Gotoh B. Sendai virus blocks alpha interferon signaling to signal transducers and activators of transcription. J. Virol. 2000;74:2477–2480. doi: 10.1128/jvi.74.5.2477-2480.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lager K.M., Mengeling W.L., Brockmeier S.L. Evaluation of protective immunity in gilts inoculated with the NADC-8 isolate of porcine reproductive and respiratory syndrome virus (PRRSV) and challenge-exposed with an antigenically distinct PRRSV isolate. Am. J. Vet. Res. 1999;60:1022–1027. [PubMed] [Google Scholar]
- Loemba H.D., Mounir S., Mardassi H., Archambault D., Dea S. Kinetics of humoral immune response to the major structural proteins of the porcine reproductive and respiratory syndrome virus. Arch. Virol. 1996;141:751–761. doi: 10.1007/BF01718333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luft T., Pang K.C., Thomas E., Hertzog P., Hart D.N.J., Trapani J., Cebon J. Type I IFNs enhance the terminal differentiation of dendritic cells. J. Immunol. 1998;161:1947–1953. [PubMed] [Google Scholar]
- Marcus P.I., Rodriquez L.L., Sekellick M.J. Interferon induction as a quasispecies marker of vesicular stomatitis virus populations. J. Virol. 1998;72:542–549. doi: 10.1128/jvi.72.1.542-549.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marrack P., Kappler J., Mitchell T. Type I interferons keep activated T cells alive. J. Exp. Med. 1999;189:521–529. doi: 10.1084/jem.189.3.521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattana P., Viscomi G.C. Variations in the interferon-inducing capacity of Sendai virus subpopulations. J. Interferon Cytokine Res. 1998;18:399–405. doi: 10.1089/jir.1998.18.399. [DOI] [PubMed] [Google Scholar]
- Meier W., Wheeler J., Hussmann R.J., Osorio F., Zuckermann F.A. Characteristics of the immune response of pigs to PRRS virus. Vet. Res. (Paris) 2000;31:41. [Google Scholar]
- Meier W.A., Galeota J., Osorio F.A., Husmann R.J., Schnitzlein W.M., Zuckermann F.A. Gradual development of the interferon-gamma response of swine to porcine reproductive and respiratory syndrome virus infection or vaccination. Virology. 2003;309:18–31. doi: 10.1016/s0042-6822(03)00009-6. [DOI] [PubMed] [Google Scholar]
- Meng X.J., Paul P.S., Halbur P.G., Morozov I. Sequence comparison of open reading frames 2 to 5 of low and high virulence United States isolates of porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 1995;76:3181–3188. doi: 10.1099/0022-1317-76-12-3181. [DOI] [PubMed] [Google Scholar]
- Meng X.J. Heterogeneity of porcine reproductive and respiratory syndrome virus: implications for current vaccine efficacy and future vaccine development. Vet. Microbiol. 2000;74:309–329. doi: 10.1016/S0378-1135(00)00196-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mengeling W.L., Vorwald A.C., Lager K.M., Brockmeier S.L. Diagnosis of porcine reproductive and respiratory syndrome using infected alveolar macrophages collected from live pigs. Vet. Microbiol. 1996;49:105–115. doi: 10.1016/0378-1135(95)00173-5. [DOI] [PubMed] [Google Scholar]
- Mengeling W.L., Lager K.M., Vorwald A.C., Koehler K.J. Strain specificity of the immune response of pigs following vaccination with various strains of porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2003;93:13–24. doi: 10.1016/s0378-1135(02)00427-3. [DOI] [PubMed] [Google Scholar]
- Meulenberg J.J., Hulst M.M., de Meijer E.J., Moonen P.L., den Besten A., de Kluyver E.P., Wensvoort G., Moormann R.J. Lelystad virus, the causative agent of porcine epidemic abortion and respiratory syndrome (PEARS), is related to LDV and EAV. Virology. 1993;192:62–72. doi: 10.1006/viro.1993.1008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molitor T.W., Bautista E.M., Choi C.S. Immunity to PRRSV: double-edged sword. Vet. Microbiol. 1997;55:265–276. doi: 10.1016/S0378-1135(96)01327-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moskophidis D., Battegay M., Bruendler M.A., Laine E., Gresser I., Zinkernagel R.M. Resistance of lymphocytic choriomeningitis virus to alpha/beta interferon and to gamma interferon. J. Virol. 1994;68:1951–1955. doi: 10.1128/jvi.68.3.1951-1955.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murtaugh M.P., Elam M.R., Kakach L.T. Comparison of the structural protein coding sequences of the VR-2332 and Lelystad virus strains of the PRRS virus. Arch. Virol. 1995;140:1451–1460. doi: 10.1007/BF01322671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murtaugh M.P., Xiao Z., Zuckermann F. Immunological responses of swine to porcine reproductive and respiratory syndrome virus infection. Viral Immunol. 2002;15:533–547. doi: 10.1089/088282402320914485. [DOI] [PubMed] [Google Scholar]
- Naniche D., Yeh A., Eto D., Manchester M., Friedman P.M., Oldstone M.B. Evasion of host defenses by measles virus: wild-type measles virus infection interferes with induction of alpha/beta interferon production. J. Virol. 2000;74:7478–7484. doi: 10.1128/jvi.74.16.7478-7484.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson E.A., Christopher-Hennings J., Benfield D.A. Serum immune responses to the proteins of porcine reproductive and respiratory syndrome (PRRS) virus. J. Vet. Diagn. Invest. 1994;6:410–415. doi: 10.1177/104063879400600402. [DOI] [PubMed] [Google Scholar]
- Nelsen C.J., Murtaugh M.P., Faaberg K.S. Porcine reproductive and respiratory syndrome virus comparison: divergent evolution on two continents. J. Virol. 1999;73:270–280. doi: 10.1128/jvi.73.1.270-280.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park J.S., Kim E.J., Kwon H.J., Hwang E.S., Namkoong S.E., Um S.J. Inactivation of interferon regulatory factor-1 tumor suppressor protein by HPV E7 oncoprotein. Implication for the E7-mediated immune evasion mechanism in cervical carcinogenesis. J. Biol. Chem. 2000;275:6764–6769. doi: 10.1074/jbc.275.10.6764. [DOI] [PubMed] [Google Scholar]
- Plagemann P.G.W. Lactate dehyrogenase-elevating virus and related viruses. In: Fields B.N., Knipe M.D.M., Howley P.M., Chanock R.M., editors. Fields Virology. 3rd ed. Lippincott Williams & Wilkins; 1996. pp. 1105–1120. [Google Scholar]
- Reed L.J., Muench H.A. Simple method of determining fifty percent enpoints. Am. J. Hyg. 1938;27:494–497. [Google Scholar]
- Ronco L.V., Karpova A.Y., Vidal M., Howley P.M. Human papillomavirus 16 E6 oncoprotein binds to interferon regulatory factor-3 and inhibits its transcriptional activity. Genes Dev. 1998;12:2061–2072. doi: 10.1101/gad.12.13.2061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossow K.D., Collins J.E., Goyal S.M., Nelson E.A., Christopher-Hennings J., Benfield D.A. Pathogenesis of porcine reproductive and respiratory syndrome virus infection in gnotobiotic pigs. Vet. Pathol. 1995;32:361–373. doi: 10.1177/030098589503200404. [DOI] [PubMed] [Google Scholar]
- Rossow K.D., Laube K.L., Goyal S.M., Collins J.E. Fetal microscopic lesions in porcine reproductive and respiratory syndrome virus-induced abortion. Vet. Pathol. 1996;33:95–99. doi: 10.1177/030098589603300115. [DOI] [PubMed] [Google Scholar]
- Rosztoczy I., Megyeri K. Essentially pure murine interferon-alpha/beta primes poly rI:rC and Sendai virus-induced interferon production in mice. J. Biol. Regul. Homeost. Agents. 1989;3:35–38. [PubMed] [Google Scholar]
- Rowland R.R., Robinson B., Stefanick J., Kim T.S., Guanghua L., Lawson S.R., Benfield D.A. Inhibition of porcine reproductive and respiratory syndrome virus by interferon-gamma and recovery of virus replication with 2-aminopurine. Arch. Virol. 2001;146:539–555. doi: 10.1007/s007050170161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samuel C.E. Reoviruses and the interferon system. Curr. Top. Microbiol. Immunol. 1998;233:125–145. doi: 10.1007/978-3-642-72095-6_7. [DOI] [PubMed] [Google Scholar]
- Samuel C.E. Antiviral actions of interferons. Clin. Microbiol. Rev. 2001;14:778–809. doi: 10.1128/CMR.14.4.778-809.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen S., Kwang J., Liu W., Liu D.X. Determination of the complete nucleotide sequence of a vaccine strain of porcine reproductive and respiratory syndrome virus and identification of the Nsp2 gene with a unique insertion. Arch. Virol. 2000;145:871–883. doi: 10.1007/s007050050680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stordeur P., Poulin L.F., Craciun L., Zhou L., Schandene L., de Lavareille A., Goriely S., Goldman M. Cytokine mRNA quantification by real-time PCR. J. Immunol. Meth. 2002;259:55–64. doi: 10.1016/s0022-1759(01)00489-6. [DOI] [PubMed] [Google Scholar]
- Sur J.H., Cooper V.L., Galeota J.A., Hesse R.A., Doster A.R., Osorio F.A. In vivo detection of porcine reproductive and respiratory syndrome virus RNA by in situ hybridization at different times postinfection. J. Clin. Microbiol. 1996;34:2280–2286. doi: 10.1128/jcm.34.9.2280-2286.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sur J.H., Doster A.R., Christian J.S., Galeota J.A., Wills R.W., Zimmerman J.J., Osorio F.A. Porcine reproductive and respiratory syndrome virus replicates in testicular germ cells, alters spermatogenesis and induces germ cell death by apoptosis. J. Virol. 1997;71:9170–9179. doi: 10.1128/jvi.71.12.9170-9179.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talon J., Salvatore M., O’Neill R.E., Nakaya Y., Zheng H., Muster T., Garcia-Sastre A., Palese P. Influenza A and B viruses expressing altered NS1 proteins: a vaccine approach. Proc. Natl. Acad. Sci. U.S.A. 2000;97:4309–4314. doi: 10.1073/pnas.070525997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanabe M., Kurita-Taniguchi M., Takeuchi K., Takeda M., Ayata M., Ogura H., Matsumoto M., Seya T. Mechanism of up-regulation of human Toll-like receptor 3 secondary to infection of measles virus-attenuated strains. Biochem. Biophys. Res. Commun. 2003;311:39–48. doi: 10.1016/j.bbrc.2003.09.159. [DOI] [PubMed] [Google Scholar]
- Taniguchi T., Takaoka A. A weak signal for strong responses: interferon-alpha/beta revisited. Nat. Rev. Mol. Cell. Biol. 2001;2:378–386. doi: 10.1038/35073080. [DOI] [PubMed] [Google Scholar]
- Tough D.F., Sun S., Zhang X., Sprent J. Stimulation of naive and memory T cells by cytokines. Immunol. Rev. 1999;170:39–47. doi: 10.1111/j.1600-065x.1999.tb01327.x. [DOI] [PubMed] [Google Scholar]
- van den Broek M.F., Muller U., Huang S., Aguet M., Zinkernagel R.M. Antiviral defense in mice lacking both alpha/beta and gamma interferon receptors. J. Virol. 1995;69:4792–4796. doi: 10.1128/jvi.69.8.4792-4796.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Reeth K., Labarque G., Nauwynck H., Pensaert M. Differential production of proinflammatory cytokines in the pig lung during different respiratory virus infections: correlations with pathogenicity. Res. Vet. Sci. 1999;67:47–52. doi: 10.1053/rvsc.1998.0277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vilcek J., Sen G.C. Interferons and other cytokines. In: Fields B.N., Knipe M.D.M., Howley P.M., Chanock R.M., editors. Fields Virology. 3rd ed. Lippincott Williams & Wilkins; 1996. pp. 375–399. [Google Scholar]
- Wensvoort G., Terpstra C., Pol J.M.A., ter Laak E.A., Bloemrad M., de Kluyer E.P., Kragten C., van Buiten L., den Besten A., Wagenaar F., Broekhuijsen J.M., Moonen P.L.J.M., Zetstra T., de Boer E.A., Tibben H.J., de Jong M.F., van’t Veld P., Groenland G.J.R., van Gennep J.A., Voets M.T.H., Verheijden J.H.M., Braamskamp J. Mystery swine disease in The Netherlands: the isolation of Lelystad virus. Vet. Q. 1991;13:121–130. doi: 10.1080/01652176.1991.9694296. [DOI] [PubMed] [Google Scholar]
- Wensvoort G. Lelystad virus and the porcine epidemic abortion and respiratory syndrome. Vet. Res. 1993;24:117–124. [PubMed] [Google Scholar]
- Whitten T.M., Quets A.T., Schloemer R.H. Identification of the hepatitis B virus factor that inhibits expression of the beta interferon gene. J. Virol. 1991;65:4699–4704. doi: 10.1128/jvi.65.9.4699-4704.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wills R.W., Zimmerman J.J., Yoon K.J., Swenson S.L., McGinely M.J., Hill H.T., Platt K.B., Chistopher-Hennings J., Nelson E.A. PRRS virus: a persistent infection. Vet. Microbiol. 1997;55:231–240. doi: 10.1016/s0378-1135(96)01337-5. [DOI] [PubMed] [Google Scholar]
- Wills R.W., Doster A.R., Galeota J.A., Sur J.H., Osorio F.A. Duration of infection and proportion of pigs persistently infected with porcine reproductive and respiratory syndrome virus. J. Clin. Microbiol. 2003;41:58–62. doi: 10.1128/JCM.41.1.58-62.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wootton S., Yoo D., Rogan D. Full-length sequence of a Canadian porcine reproductive and respiratory syndrome virus (PRRSV) isolate. Arch. Virol. 2000;145:2297–2323. doi: 10.1007/s007050070022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoon I.J., Joo H.S., Christianson W.E., Morrison R.B., Dial G.D. Persistent and contact infection in nursery pigs experimentally infected with PRRS virus. Swine Health Prod. 1993;1:5–8. [Google Scholar]
- Yoon K.J., Zimmerman J.J., Swenson S.L., McGinley M.J., Eernisse K.A., Brevik A., Rhinehart L.L., Frey M.L., Hill H.T., Platt K.B. Characterization of the humoral immune response to porcine reproductive and respiratory syndrome (PRRS) virus infection. J. Vet. Diagn. Invest. 1995;7:305–312. doi: 10.1177/104063879500700302. [DOI] [PubMed] [Google Scholar]
- York I.A., Rock K.L. Antigen processing and presentation by the class I major histocompatibility complex. Annu. Rev. Immunol. 1996;14:369–396. doi: 10.1146/annurev.immunol.14.1.369. [DOI] [PubMed] [Google Scholar]
- Zhang J.J., Vinkemeier U., Gu W., Chakravarti D., Horvath C.M., Jr., Darnell J.R. Two contact regions between Stat1 and CBP/p300 in interferon gamma signaling. Proc. Natl. Acad. Sci. U.S.A. 1996;93:15092–15096. doi: 10.1073/pnas.93.26.15092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmerman J.J., Yoon K.J., Wills R.W., Swenson S.L. General overview of PRRSV: a perspective from the United States. Vet. Microbiol. 1997;55:187–196. doi: 10.1016/s0378-1135(96)01330-2. [DOI] [PubMed] [Google Scholar]