

Abstract
There is a close functional relationship between the male reproductive and immune systems. Immunological responses against sperm antigens or other elements of the reproductive tract can lead to androgen insufficiency, infertility, or chronic inflammation. Systemic or local immune activation and inflammation directly inhibit the hypothalamic-pituitary-Leydig cell axis, interfere with spermatogenic cell development, and may provoke sperm antibody formation. Traditional explanations for protection of spermatogenic cells, based on physical barriers or immune cell exclusion, are not consistent with either the organization of the reproductive tract or modern concepts of immunoregulation. Regulation of immune responses in the male reproductive tract involves immunoregulatory macrophages and lymphocytes, as well as active somatic cell suppression of antigen-specific immunity by anti-inflammatory and immunosuppressive factors, including cytokines and androgenic steroids. These restraints on antigen-specific immune mechanisms appear to be counterbalanced by enhanced innate immunity. Finally, inflammatory signaling pathways appear to play an important role in normal male reproductive function, and there is evidence that establishment of male reproduction during development is intimately linked to the maturation of the local immune environment.
Keywords: Autoimmunity, Epididymis, Immune privilege, Inflammation, Macrophage, Sertoli cell, Spermatogenesis, Steroidogenesis, Tolerance, Vas deferens
Introduction
The Male Reproductive Tract as a Unique Immunological Environment
The male reproductive tract, and the male gamete especially, present a unique challenge to the immune system. The human testis continuously produces highly differentiated sperm derived from a pool of spermatogonial stem cells at a level of productivity and complexity matched only by the hematopoietic system. In contrast to the hematopoietic tissues, however, differentiated sperm first appear at the time of sexual maturation, long after the maturation of the immune system and the establishment of systemic immune tolerance (Figure 19.1 ).1 In humans, the period between the editing of the lymphocyte repertoire and the first appearance of significant numbers of the earliest premeiotic germ cell (the spermatocytes) is generally more than 10 years. As a consequence, spermatogenic cells express many cell-specific proteins and other molecules that have the potential to be seen as “foreign” or “nonself” by the immune system.
FIGURE 19.1.
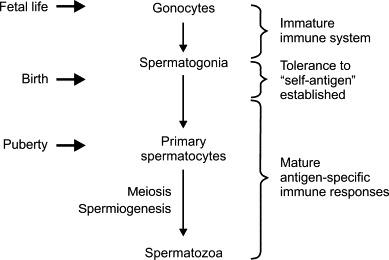
Developmental timeline of spermatogenesis and the maturation of the immune system.
The majority of spermatogenic cells do not appear in the testis until the initiation of meiosis at puberty, whereas tolerance to self-antigen is largely established by the time of birth. As the development of spermatozoa from the spermatogonial stem cell population involves complex processes of nuclear reorganization (meiosis) and cellular differentiation (spermiogenesis) that are unique to the testis, there exists enormous potential for antigens of spermatogenesis to evade conventional tolerance mechanisms.
We know that the immune system tends to view spermatogenic cells as foreign because of the relatively high incidence of autoimmune infertility among human populations. Even in developed countries with modern health care, sperm autoantibodies represent 5–10% of all male infertility,2., 3. while testicular biopsies from infertile men frequently display evidence of asymptomatic inflammatory reactions.4., 5. In other forms of autoimmune disease, such as type 1 diabetes or gastritis, the development of autoimmunity is due to disruption of the normal regulatory controls of the immune system, leading to reactions against antigens that normally are ignored. Only a limited number of antigens are involved and a specific dominant autoantigen usually has been identified.6 In the case of the male reproductive tract, by contrast, autoimmunity generally involves antigens that would not be edited out of the self-reactive repertoire in the first place. Hence, autoimmune infertility generally involves multiple antigens as well as different antigens from one individual to another.7., 8., 9. Moreover, infertility is not the only urological problem with an immunological basis. Chronic pelvic inflammatory disease in men, which may be accompanied by recurrent and even debilitating perineal or scrotal pain, is a serious and frequently intractable condition.10., 11., 12. Infections may represent the initial cause of the majority of such cases, but the underlying mechanisms almost certainly involve an autoimmune component.10
Unraveling the origin of these immunologically-based disturbances of reproductive function is an important clinical goal, but the converse, and equally important, scientific question is: What is it about the male reproductive tract that permits the continuous production of huge numbers of immunogenic cells expressing multiple autoantigens without apparent problems in the majority of individuals? In most respects, the immune system within the male reproductive tract appears relatively normal, with effective lymphatic drainage and relatively free access of immune cells.13., 14. Certainly, there are some unique structural characteristics of the male reproductive tract that may contribute. For example, the male gametes are held at a substantially lower temperature than the rest of the body in species with scrotal testes and epididymides. There also exists a very effective blood–testis barrier that sequesters most of the spermatogenic cells in the testis.15., 16., 17., 18. Nonetheless, these physical elements cannot account for all the manifestations of immunological protection in the male reproductive tract.
Instead, studies over many years have confirmed that the male reproductive tract, and the testis in particular, constitutes a unique immunoregulatory environment. There are communication and regulatory networks that are common to both male reproduction and the immune system, providing many striking overlaps between the control of spermatogenesis and the processes of inflammation and immune activation. Part of this network includes local immunoregulatory and immunosuppressive mechanisms, which exist to provide protection for the endogenous antigens of spermatogenesis, protection that also extends to antigens expressed by foreign grafts inserted into the testis environment.19 Immune cells that enter the environment of the male reproductive tract become functionally modified to restrict their proinflammatory activity and provide an immunologically constrained environment where antigen-specific immune responses are closely controlled. Balanced against this, of course, is the question of how the male reproductive tract is able to protect itself from recurrent infections and tumor development under these circumstances.
The observation that reproductive dysfunction is not only associated with local infection and its accompanying inflammation, but also with systemic disease, provides further evidence of an intimate relationship between the male reproductive tract and the immune system.20., 21. Many systemic illnesses are accompanied by a reduction in both serum androgen levels and sperm output, indicating that male sexual function and general well-being maintain a reciprocal relationship. It has been suggested that this represents a physiologically important mechanism, having evolved to limit reproductive activity during periods of illness. The possibility that acute inflammatory dysregulation of male reproductive function can lead to more permanent problems, such as autoimmune infertility or chronic inflammatory disease, also must be considered. It may even be the case that pre-existing hypogonadism predisposes men to inflammatory disease. At a fundamental level, the ability of the immune system and the male reproductive system to co-exist is no less essential to male reproductive success than the normal operation of the hypothalamic-pituitary unit or the critical interactions between somatic cells and spermatogenic cells. The use of the term immunophysiology in this context highlights the fact that the immunology and basic physiology of male reproduction cannot actually be separated.
Since the original publication of this chapter, nearly a decade ago, there have been considerable advances in our understanding of male reproductive immunophysiology. Most significantly, inflammation has moved to center stage in male infertility, with more testicular pathologies now being recognized as possessing inflammatory features,22., 23. and there is growing awareness that basic immunological mechanisms underlie normal reproductive tract function.24 There has been a considerable increase in knowledge of the mechanisms responsible for controlling testicular immune responses, as well as advances in understanding of the unique immunology of the epididymis and excurrent ducts. The principal aim of this chapter is to provide a comprehensive overview of the field, including how it has developed, its basic tenets and mechanisms, and a broad outline of the current state of knowledge. The overview is confined to consideration of mammalian species, as particularly relevant to the human.
Historical Aspects
The study of male reproductive tract immunophysiology extends back to the very beginnings of endocrinology and immunology. Inflammation of the testis and its association with mumps parotitis was known in ancient times, having been described in the writings of Hippocrates (c. 460–377 BC). Studies on the transplantation of testes in domestic chickens, generally considered to be the earliest studies in the field of endocrinology, were undertaken in the eighteenth century by Hunter and by Michaelis (cited by Setchell, 1990),25 although the first systematic experiments actually were reported by Berthold in 1849.26 “Successful” transplants of mammalian testes had to wait until some time later.27., 28., 29. In the early part of the last century prior to the discovery of the male sex hormone, testosterone, transplantation of male gonads, including transplants of animal gonads into humans, was undertaken by charlatans and serious researchers alike in the search for treatments to increase male health and virility.30., 31. It was quickly noted that, although the function of the interstitial tissue appeared to continue in these transplant experiments, tubule function was not preserved.32 While there is no doubt that temperature and revascularization issues were important limitations, it is now obvious that many of these experiments faced a crucial immunological impediment. Even today, grafts of testicular tissue generally are more successful in an immunologically compatible or immunocompromised host.33 The concept of immunoregulation entered the story with the discovery that allogeneic testicular grafts could be made to survive much more successfully if tolerance had been induced by injection of the donor allogeneic cells at the time of birth in the donor,34 or when transferred into the anterior chamber of the eye.35., 36. Sometime later, Billingham and others conducted a series of investigations into so-called immunologically privileged sites that led to recognition of the testis as a tissue that was particularly favorable for graft survival.37
Evidence that the spermatozoon itself is immunogenic to its autologous host dates back to the discovery of antibodies against sperm by Lansteiner38 and Metchnikoff.39 In the 1920s,40 Guyer was able to produce infertility in rabbits and guinea pigs by passive immunization with sperm specific antisera, while Kennedy reported degenerative changes in the testis following active immunization with autologous sperm.41 By the middle of the century, Voisin and colleagues were able to produce aspermatogenesis in guinea pigs by active immunization with testicular extracts.42 Eventually, an association between sperm autoantibody formation and infertility in humans was noted.43., 44. Further experimental evidence confirming the possibility of autoimmune reactions against the sperm led to a general acceptance that these are an important cause of infertility in men.2., 45. In time, this led to the broader concept that the testis environment must provide protection for the spermatogenic cells through specific regulatory mechanisms.
Biological sex exerts a significant influence on the immune system. Gender differences in health and the greater prevalence, earlier onset, and severity of autoimmune diseases, such as systemic lupus erythematosus and rheumatoid arthritis, among women had been recognized long before a real understanding of autoimmunity existed.46., 47. A specific effect of the testes on the immune system was reported as early as 1898 by Calzolari,48 when he found that castration of rabbits prior to puberty led to an increase or maintenance of the size of the thymus. This effect has been repeatedly confirmed and extended to include cell-mediated immunity and graft survival.49., 50. Many studies have shown that various immunological processes have gender-specific differences or can be affected by castration and/or sex steroid replacement. These data clearly established that products of the testis, and sex steroids in particular, regulate the immune system either directly or indirectly, setting the stage for the modern era of male reproductive immunophysiology.
Structure and Function of the Male Reproductive Tract Relevant to Immunophysiology
The anatomy and physiology of the testis and other components of the male reproductive tract are covered in detail elsewhere in this volume (see Chapters 3, 14, 16, 17, and 18Chapter 3Chapter 14Chapter 16Chapter 17Chapter 18). A brief outline highlighting the issues relevant to understanding the interface between the immune system and male reproductive tract is provided here.
The Testis
Structural Organization
Functionally and anatomically, the testis is separated into an avascular spermatogenic compartment, the seminiferous tubules, and a highly vascularized endocrine compartment, the interstitial tissue. The testis is enclosed by a fibrous capsule, but there is considerable species variation in the connective tissue of the testicular parenchyma. In the human, the testis is physically partitioned by connective tissue septa into discrete lobules containing the loops of the seminiferous tubules, which connect at both ends to the rete testis located along one pole of the testis.51 The products of the seminiferous tubules are collected by the rete testis and transferred to the adjacent epididymis, which is connected to the rete testis via a series of efferent ducts. The testes of rodent species, such as rat and mouse, comprise only very loose connective tissue with no distinct septa separating the seminiferous tubules.52 The interstitial tissue completely surrounds the seminiferous tubules (Figure 19.2 ), and contains the vasculature, lymphatic vessels, and nerves of the testis. The testicular blood supply arises from the abdominal aorta, and this produces a comparatively long and highly coiled spermatic artery that is particularly susceptible to physical insult and torsion in species with scrotal testes. The arterioles, capillaries, and venules of the testis thoroughly permeate the interstitial tissue surrounding the seminiferous tubules and rete testis. Consequently, these structures are close to an effective blood supply at all times in spite of the completely avascular nature of the spermatogenic compartment.
FIGURE 19.2.
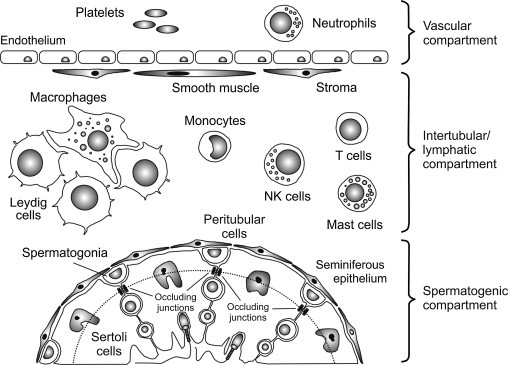
Immunological compartmentalization of the testis.
The mammalian testis comprises three immunologically distinct compartments: the vascular compartment and intertubular (or interstitial) compartment are separated by a layer of nonfenestrated endothelium, while the intertubular and spermatogenic compartments are separated by a layer of peritubular myoid cells and by occluding junctions between adjacent Sertoli cells. These junctions constitute the blood–testis barrier, which further divides the seminiferous epithelium into a basal and an adluminal region (dotted line). The adluminal region contains the meiotic germ cells within a highly specialized microenvironment. Under normal conditions, monocytes, macrophages, T cells, NK cells and, in some species, mast cells and/or eosinophils have relatively free access to the intertubular compartment, but are entirely excluded from the adluminal region of the seminiferous epithelium. Neutrophils are confined to the vascular compartment except during specific immunological events.
Unlike the capillaries of other endocrine glands, the majority of testicular capillaries are not fenestrated,53 and the mechanisms whereby molecules enter and exit the testis via this route still await resolution.54 There appears to be very little functional restriction on the exchange of even large molecules across this barrier and the interstitial fluid is very similar in its overall composition to that of the circulating blood.55 The venous drainage of the testis via the spermatic veins is closely associated with the arterial supply, which together form a very effective countercurrent heat and solute exchange structure, called the pampiniform plexus.56
The organization of the testicular lymphatics varies between species, ranging from irregular channels or sinusoids that are incompletely bounded by endothelial cells in rodents, to large discrete lymphatic vessels in humans, to very small, rapid flow lymphatics in porcine species.52., 57. These lymphatics completely invest the entire interstitial tissue and pass without restriction to local draining lymph nodes, principally the lumbar or para-aortic lymph nodes.58., 59. In the laboratory rat, drainage to the iliac and renal nodes predominates, but there may be some lymphatic drainage directly to the thoracic ducts without passing through regional lymph nodes.13
The most prominent cell type present in the interstitium is the Leydig cell,60 which produces androgens.61 Macrophages are commonly observed in the interstitium of most, if not all, species and many testes also contain variable numbers of mast cells and/or eosinophils.62., 63. Less numerous, but ubiquitous nonetheless, are the intratesticular lymphocytes: T cells and natural killer (NK) cells.64., 65., 66., 67., 68., 69., 70.
The seminiferous tubules are bounded by a circumferential layer of peritubular cells and the basal lamina, which together form the limiting structure on which rests the Sertoli cells and the spermatogonia (Figure 19.2). Immune cells are occasionally seen within this boundary layer.71 The Sertoli cell provides the structural framework for the organization of the seminiferous epithelium, but also plays a crucial role in supporting and directing the development of the spermatogenic cells. Adjacent Sertoli cells and spermatogenic cells maintain intimate contact at all times, with junctional and membrane specializations providing physical contact and communication.72 At the time of puberty, cohorts of mitotically dividing spermatogonia begin to enter meiosis at regular intervals, moving away from the periphery of the tubule and becoming spermatocytes. Meiosis produces haploid round spermatids, which subsequently undergo considerable structural differentiation to become mature or elongated spermatids. Once these cells are released by the Sertoli cell into the tubule lumen they are called spermatozoa and fluid secreted by the Sertoli cells sweeps the released spermatozoa toward the rete testis.
The seminiferous epithelium is highly organized. As each cohort of spermatogonia becomes committed to the spermatogenic process, they displace earlier cohorts toward the lumen of the tubule, so that multiple generations of developing germ cells co-exist in each segment of the seminiferous epithelium. As a consequence of the regular intervals of spermatogonial commitment and the constant rate of the spermatogenic process itself, the generations form specific associations of germ cells at different levels of maturity within the epithelium. These associations constitute the stages of a recurring developmental sequence, called the “cycle of the seminiferous epithelium,” which is also promulgated along the entire length of the tubule (see Chapters 16).51 This organized complexity implies a high degree of communication and regulation across the generations as well as between spermatogenic cells and supporting Sertoli cells.
Elaborate occluding junctions between adjacent Sertoli cells form an intercellular barrier that is completely impermeable even to small molecules.15., 16. This constitutes the main component of the blood–testis barrier and separates the premeiotic and early meiotic cells in the basal region of the seminiferous epithelium from the adluminal spermatocytes and spermatids (Figure 19.2). In this way, a large majority of the developing germ cells are sequestered within a highly specialized environment and effectively isolated from the vasculature and immune system. In contrast, the rete testis epithelium lacks both Sertoli cells and their highly specialized junctional specializations. The epithelial barrier restricting movement from the blood into the rete testis appears to be substantially less effective than that of the seminiferous epithelium, with the result that immunoglobulins and possibly even immune cells are able to cross the epithelium.64., 73.
Endocrine Regulation
Male reproduction is maintained by pulsatile secretion of gonadotropin releasing hormone (GnRH) by the hypothalamus, which stimulates concordant pulses of luteinizing hormone (LH) and follicle-stimulating hormone (FSH) from the anterior pituitary.74 In the testis, LH binds to specific G-coupled receptors on the surface of the Leydig cells, thereby stimulating adenylate cyclase to produce the intracellular second messenger, cAMP, and activating the cAMP-dependent protein kinase A (Figure 19.3 ).61 This activation mobilizes cholesterol from intracellular stores, extracellular lipoprotein sources, or de novo synthesis from acetate, and stimulates the transfer of the cholesterol to the inner-mitochondrial membrane through the action of the steroidogenic acute regulatory protein (STAR).75 Ongoing maintenance of steroidogenic enzyme expression is also under LH/cAMP control.76 Once cholesterol enters the mitochondrion, it is metabolized to pregnenolone via the action of the cytochrome P450 cholesterol side-chain cleavage enzyme (CYP11A) residing on the inside face of the inner matrix membrane. Pregnenolone diffuses out of the mitochondrion to the smooth endoplasmic reticulum, where it may be converted to progesterone by 3β-hydroxysteroid dehydrogenase/Δ4-Δ5 isomerase (HSD3β). Pregnenolone and progesterone are first metabolized to their 17α-hydroxy forms and then to the weak androgens, dehydroepiandrosterone and androstenedione, respectively, by the action of steroid 17α-hydroxylase/17,20 lyase (CYP17A). Finally, androstenedione is converted to testosterone by the action of hydroxysteroid (17β) dehydrogenase (HSD17β), and dehydroepiandrosterone is converted to androstenediol and then testosterone, by the sequential actions of HSD17β and HSD3β. Testosterone is secreted from the Leydig cell and serves as the principal androgen in both the testis and circulation.
FIGURE 19.3.
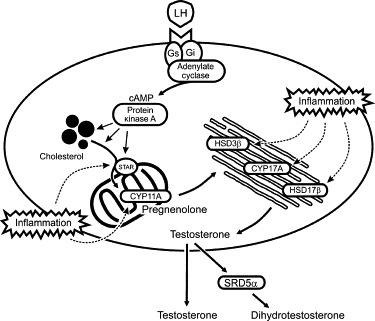
Regulation of testosterone biosynthesis in Leydig cells and sites of inhibition during inflammation.
The gonadotropin, LH, binds to a G protein-coupled receptor on the cell surface, thereby activating adenylate cyclase, production of cAMP and protein kinase A activity. This stimulates the transfer of cholesterol from intracellular stores into the mitochondria through the action of the steroidogenic acute regulatory protein (STAR), where the cholesterol side-chain cleavage enzyme (CYP11A) converts the cholesterol to pregnenolone. Pregnenolone is converted to testosterone in the smooth endoplasmic reticulum by the enzymes, 3β-hydroxysteroid dehydrogenase/Δ4-Δ5 isomerase (HSD3β), steroid 17α-hydroxylase/17,20 lyase (CYP17A) and hydroxysteroid (17β) dehydrogenase (HSD17β). Testosterone is reduced by the action of the 5α-reductase enzyme (SRD5α) to the more potent androgen, dihydrotestosterone. Inflammation inhibits the activity of STAR and all the main enzymes of the steroidogenic pathway.
Both testosterone and FSH bind to specific Sertoli cell receptors to regulate spermatogenesis and Sertoli cell functions, including secretion of the protein hormone, inhibin.77 In turn, testosterone and inhibin operate via a negative feedback loop to regulate LH and FSH synthesis and secretion at the pituitary and hypothalamic levels.78 Withdrawal of androgens leads to rapid cessation of spermatogenesis, although the levels of intratesticular testosterone required to maintain qualitatively normal spermatogenesis are considerably lower than the intratesticular concentrations that normally exist.79., 80. Consequently, spermatogenesis can tolerate even relatively large declines in testicular androgen production with relatively minor losses of efficiency. In contrast, peripheral levels of androgens are critical; even small reductions can have profound effects on many androgen-dependent functions, including accessory gland function, secondary sex characteristics, and libido.81 Peripheral androgen levels are dependent upon both Leydig cell production and testicular vascular function, so that interference with the vasculature of the testis can alter circulating testosterone levels quite significantly.82 Conversion of testosterone and androstenedione to estrogens by the cytochrome P450 enzyme aromatase (CYP19A) in the Leydig cell and Sertoli cell is also required for normal development and function of the efferent ducts and epididymis.83
The Epididymis, Vas Deferens, and Accessory Glands
The epididymis comprises a long single, highly coiled epididymal duct lined primarily by columnar principal cells with extensive apical stereocilia. Testicular fluid secreted by the Sertoli cells is largely reabsorbed by the epithelial cells of the efferent ducts and the proximal regions (caput) of the epididymis.84 Sperm maturation occurs during transit through the epididymal duct and sperm are stored prior to ejaculation in the distal (cauda) region of the epididymis.85., 86. The cauda epididymis is connected to the vas deferens, a highly muscularized duct that drives the epididymal contents toward the urethra at the time of ejaculation. The testicular and epididymal secretions constitute only about 10% of the ejaculate, with the remaining 90% of the semen coming from the accessory glands: the seminal vesicles and prostate, in particular.87 All the posttesticular ductal structures of the male tract and the accessory glands are dependent upon androgens for normal development and maintenance of function.81 However, in contrast to the testis, conversion of testosterone to the more potent androgen, 5α-dihydrotestosterone (DHT), by the action of steroid 5α-reductase (SRD5α) is usually necessary in most other androgen-responsive tissues.88
It might be assumed that sperm spend the majority of their time in the testis and epididymis and appear only briefly within the vas deferens and urethra during ejaculation. Critically, however, some sperm may be retained within the tract for much longer periods, as spermatozoa continue to appear in the ejaculates of vasectomized men for several months even after a successful procedure.89 In fact, the presence of intact sperm even has been noted in human prostate glands collected following prostatic surgery or postmortem, suggesting that such ectopic sperm may play a role in the etiology of prostatic inflammation and possibly even sperm autoimmunity.90
There is a blood–epididymis barrier restricting movement of molecules across the epididymal epithelium, although evidence suggests that this barrier is not as elaborate or effective as the blood–testis barrier.72., 86., 91., 92., 93. In normal adults, circulating immunoglobulin appears to be restricted, although perhaps not entirely excluded, from passage into the epididymal fluid.91 The most striking contrast with the seminiferous epithelium is the presence of macrophages and lymphocytes within the epididymal epithelium.65., 94., 95., 96., 97., 98., 99., 100., 101., 102. The presence of these cells suggests the existence of a very different immunological environment compared with that of the testis. Epithelial-type barriers, similar to that present in the epididymis, are found throughout the remainder of the reproductive tract.72., 93., 103.
The Immune System and Its Endocrine Control
General Principles
Fundamentally, the immune system provides protection for more complex animals from invading organisms that seek to exploit vulnerabilities or other opportunities to infect their host. This protection involves a complex suite of cells and molecules that allows the animal to identify and then eliminate the invading pathogen. In vertebrates, the immune system comprises an innate immune system, which generally recognizes uniquely conserved molecular patterns expressed by various pathogens, and the adaptive (or acquired) immune system, which specifically recognizes molecular patterns that are foreign to the host. Neither system operates in isolation and many of the cellular and molecular mechanisms overlap (Figure 19.4 ). The functions of both innate and adaptive immunity impact upon the male reproductive tract at multiple levels and have profound consequences for male reproduction.
FIGURE 19.4.
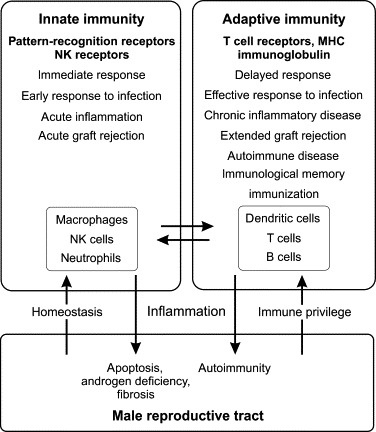
Components and properties of the innate and adaptive immune systems, and their interaction with the male reproductive tract.
Innate immunity is the immediate response to infection and other external threats, subsequently activating the more delayed adaptive immune response. The male reproductive tract regulates innate and adaptive immunity in order to maintain protection against infections, while at the same time suppressing antigen-specific immunity to protect the spermatogenic cells (immune privilege). Activation of the innate and adaptive immune systems, due to infection or other inflammatory stimulus, has detrimental effects on male reproductive tract function, resulting in loss of androgens and spermatogenic disruption, and can lead to tissue damage, ongoing autoimmunity and infertility.
The cellular components of the immune system are the leukocytes, or white blood cells. These cells and their products circulate continuously through the blood, lymph, and tissues in both surveillance and effector modes. The innate immune system is comprised principally of the mononuclear phagocytes (monocytes and macrophages) and granulocytes or polymorphonuclear cells (neutrophils, eosinophils, basophils, and mast cells), but also involves cells more closely aligned with the adaptive responses (NK cells and dendritic cells). The cellular components of the adaptive immune system are the lymphocytes (T cells, B cells, and NK cells), and the “professional” antigen-presenting cells (dendritic cells and macrophages). In modern immunology, the cells of the immune system and their various functional subsets are primarily identified and even defined by expression of specific antigens, called cluster designation (CD) markers, recognized by well-characterized monoclonal antibodies (Table 19.1 ).104
TABLE 19.1.
Cluster Designation (CD) Markers Relevant to the Male Reproductive Tracta
| Marker | Gene Name, Common or Superseded Designation(s) | Function(s) |
|---|---|---|
| CD1 | Ly-38, R3 (CD1D) | Nonclassical MHC; presentation of lipid and glycolipid antigens |
| CD3 | T3, Leu 4 | Signaling component of the TCR complex |
| CD4 | T4, Leu 3 | Co-receptor for recognition of MHC class II; component of the TCR complex |
| CD8 | Ly-2, Ly-3, T8, Leu 2 | Co-receptor for recognition of MHC class I; component of the TCR complex |
| CD11a CD11b CD11c |
ITGAL, LFA-1, Ly-15, Ly-21 ITGAM, Mac-1, Ly-40 ITGAX, Leu M5 |
Integrin α chains; adhesion molecules, expressed on leukocytes |
| CD14 | LPS-R | Lipopolysaccharide-binding protein complex co-receptor; TLR4 co-receptor |
| CD16 | FCGR3, FcγRIII, Ly-17 | Receptor for Fc fragment of immunoglobulin G; expressed on subset of NK cells |
| CD18 | ITGB2, LCAMB | Integrin β2; adhesion molecule, pairs with CD11 |
| CD25 | IL2RA, Ly-43 | IL2 receptor α chain; marker for activated and Treg cells |
| CD28 | T44 | Receptor for CD80 and CD86; expressed on activated T cells, NK cells |
| CD30 | TNFRSF8 | TNF receptor superfamily, member 8; expressed on activated T cells, B cells |
| CD40 | TNFRSF5 | TNF receptor superfamily, member 5; co-stimulatory receptor; expressed on antigen-presenting cells |
| CD45 | PTPRC, LCA, Ly-5, T200 | Protein tyrosine phosphatase, receptor type, C; leukocyte common antigen; expressed on all leukocytes |
| CD46 | MCP | Membrane cofactor protein; complement regulatory protein |
| CD52 | CAMPATH-1 antigen | Complement regulatory protein; Treg co-stimulatory co-receptor |
| CD54 | ICAM1, Ly-47 | Intercellular cell adhesion molecule-1; expressed on activated endothelial cells |
| CD55 | DAF | Decay accelerating factor for complement; complement regulatory protein |
| CD56 | NCAM1 | Neural cell adhesion molecule-1; specific variant expressed on NK cells |
| CD59 | MAC-IP | Membrane attack complex inhibition factor; complement regulatory protein |
| CD68 | Macrosialin | Lysosomal membrane glycoprotein; expressed by dendritic cells, monocytes, some macrophages |
| CD80 | B7-1, B7/BB1, Ly-53 | Co-stimulatory co-receptor; ligand for CD28; expressed on antigen-presenting cells |
| CD86 | B7-2, Ly-58 | Co-stimulatory co-receptor; ligand for CD28; expressed on antigen-presenting cells |
| CD95 | FAS, APO-1 | Receptor for CD95 ligand; mediates apoptosis; expressed on activated lymphocytes |
| CD106 | VCAM1 | Vascular cell adhesion molecule-1; expressed on activated endothelial cells |
| CD126 | IL6R | Interleukin-6 receptor subunit; pairs with interleukin-6 signal transducer |
| CD130 | IL6ST, gp130 | Interleukin-6 signal transducer |
| CD152 | CTLA4 | Cytotoxic T-lymphocyte-associated antigen 4; inhibitory receptor for CD80 and CD86; expressed on T cells |
| CD154 | CD40LG | Ligand for CD40; co-stimulatory co-receptor; expressed on T cells |
| CD163 | M130 | Scavenger receptor for hemoglobin-haptoglobin complex; expressed on macrophage subset (M2) |
| CD206 | MRC1 | Macrophage mannose receptor 1; expressed on macrophage subset (M2) |
Refer to text for full details. Note that some designations represent multiple protein members.
The Innate Immune Response
The innate immune system provides the first line of defense against external threats through an inherent ability to recognize and rapidly respond to a broad range of pathogens and other immunogens, and by promoting the process of inflammation. Innate immunity plays a fundamental role in the response of the male reproductive tract to infections, but it also exhibits a much wider role in male reproduction because a number of regulatory mechanisms are shared by the innate immune and the reproductive systems.
Pattern Recognition Receptors and Activation of Innate Immunity
Activation of the innate immune response involves pattern-recognition receptors, which recognize specific motifs, or pathogen-associated molecular patterns (PAMPs), produced by bacterial, viral, fungal, and protozoan pathogens.105 Unlike classical ligand receptors, these receptors are able to respond to multiple ligands that possess related, rather than identical, structures. The canonical pattern-recognition receptors are a family of transmembrane receptors called the Toll-like receptors (TLR), which are expressed on the cell surface and on intracellular endosomes.106 There are several families of intracytoplasmic pattern-recognition receptors: the nucleotide binding and oligomerization domain (NOD)-like receptors (NLR), the retinoic acid-inducible gene (RIG)-like receptors (RLR), and the C-type lectin receptors (CLR).107 Importantly, many of these receptors can also interact with endogenous molecules released by cell damage, called danger-associated molecular patterns (DAMPs), which include high-mobility group box 1 protein (HMGB1), heat shock proteins, extracellular matrix components, and nucleic acids.108
The TLRs are highly expressed by myeloid-lineage cells (monocytes, macrophages, and dendritic cells), but are also found on other leukocytes, epithelial cells, and stromal cells. There are 10 TLRs (numbered TLR1–10) in the human, but the laboratory rodents (rats and mice) possess an additional three TLRs (TLR11–13).106., 109. These receptors detect unique ligands of bacterial, viral, and fungal origin, such as bacterial and viral nucleic acids, bacterial lipopeptides, peptidoglycans, and lipopolysaccharides (LPS). LPS is a component of the cell wall of gram-negative bacteria, such as Escherichia coli, and the receptor for LPS is TLR4, which requires a co-receptor called MD2 (myeloid differentiation 2 protein), and the LPS-binding protein CD14 for full activation.110., 111.
The ligand-binding region of the TLRs is characterized by multiple N-terminal leucine-rich repeats, which facilitate detection of specific molecular patterns. These receptors are functionally related to the interleukin-1 (IL1) receptor (IL1R), with which they share a conserved cytoplasmic domain called the Toll/IL1R (TIR) domain.106., 112. Activation of the TLRs involves receptor dimerization and interaction of the TIR domain with an intracellular TIR domain-containing adaptor protein (Figure 19.5 ). In the case of all the TLRs, except TLR3, the adaptor protein is myeloid differentiation primary-response protein 88 (MYD88), which signals via IL1 receptor-associated kinase (IRAK) and tumor necrosis factor (TNF) receptor-associated factor 6 (TRAF6). This leads to activation of the p38 mitogen-activated protein kinase (MAPK14) and Jun N-terminal kinase (MAPK8), and nuclear translocation of the transcription factors, nuclear factor kappa B (NFκB), and activated protein-1 (AP-1).113., 114. This, in turn, induces expression of genes encoding the key pro-inflammatory cytokines and mediators, including both IL1α and β forms (IL1α and IL1β), TNF, IL6, IL8 (C-X-C motif ligand 8; CXCL8), IL12, inducible nitric oxide synthase (NOS2) and prostaglandin-endoperoxide synthase 2 (PTGS2; cyclooxygenase 2; Table 19.2 ).115., 116. In addition, TLR3 and TLR4 interact with the adaptor protein, TIR domain-containing adaptor molecule 1 (TICAM1), to activate TRAF3 and the transcription factor, interferon regulatory factor 3 (IRF3), resulting in production of the type 1 interferons (IFNα and IFNβ).115 Some of the NLRs, which detect various bacterial PAMPs within the cytosol, likewise exert their actions via activation of NFκB and the MAP kinases, but a subset of the NLRs work by induction of the cysteine protease, caspase-1 (CASP1; interleukin-1β converting enzyme), through assembly of a large intracellular protein complex called the inflammasome.117., 118. Inflammasomes are generically composed of a pattern-recognition domain-containing protein, an adaptor molecule bearing a caspase activation and recruitment domain (CARD), and CASP1 itself, which activates the key pro-inflammatory cytokines, IL1β and IL18, by processing their inactive precursors (Figure 19.5).118 These complexes are activated by various PAMPs and DAMPs, including bacterial toxins, viral RNA, and particulates, such as silica and uric acid crystals. Activation of the pattern-recognition receptors leads to functional activation of immune cells and sets the inflammatory response in train.
FIGURE 19.5.
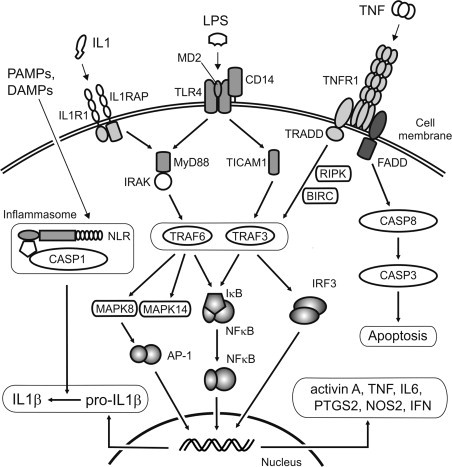
Schematic of inflammatory signaling pathways.
Binding of bacterial LPS to Toll-like receptor 4 (TLR4), which also comprises co-receptor proteins MD2 and CD14, or binding of IL1 to the IL1 receptor (IL1R1) and engagement of the ILR acceptor protein (IL1RAP), leads to interaction with the adaptor molecules, MYD88 or TICAM1. Signaling via MYD88 occurs through the IL1 receptor-associated kinase (IRAK) and TNFR-associated factor 6 (TRAF6), leading to degradation of the NFκB repressor protein IκB, and activation of mitogen-activated protein kinases (MAPK8 and MAPK14). These kinases activate multiple downstream events, including production of the transcription factor AP-1. Signaling via TICAM1 occurs through TRAF3 and activates the interferon-induced transcription factor, IRF3. However, there is considerable potential for overlap between signaling pathways. Depending upon which adaptor molecule is engaged, binding of TNF to its receptor (TNFR1) can lead to activation of TRAFs via the TNFR-associated death domain protein (TRADD)-mediated pathway, via receptor interacting serine–threonine kinase (RIPK) or baculoviral IAP repeat containing protein (BIRC), or to the caspase activation cascade and apoptosis through the FAS-associated death domain protein (FADD). The transcription factors, NFκB, AP-1 and IRF3, translocate to the nucleus to induce transcription of inflammatory genes. Various pathogen-associated and endogenous molecules (PAMPs and DAMPs) bind to intracellular pattern-recognition receptors associated with the inflammasome, such as the NOD-like receptors (NLR), and activate caspase-1 (CASP1), which is required to process pro-IL1β into the active pro-inflammatory protein. Note that the pathways shown are highly simplified – not all intermediates or potential interactions are depicted.
TABLE 19.2.
Key Enzymes Involved in Inflammation and Immunity in the Male Reproductive Tract
| Enzyme Name | Gene Name | Common Names | Properties/Function |
|---|---|---|---|
| Nitric oxide synthase, type I | NOS1 | Neuronal NOS (nNOS) | Ca/calmodulin regulated |
| Nitric oxide synthase, type II | NOS2 | Inducible NOS (iNOS) | Transcriptionally and translationally regulated |
| Nitric oxide synthase, type III | NOS3 | Endothelial NOS (eNOS) | Ca/calmodulin regulated |
| Phospholipase A2 | Multiple genes | Hydrolysis of free fatty acid from membrane phospholipids | |
| Prostaglandin-endoperoxide synthase 1 | PTGS1 | Cyclooxygenase 1 (COX1) | Constitutive, converts arachidonic acid to prostaglandin G/H |
| Prostaglandin-endoperoxide synthase 2 | PTGS2 | Cyclooxygenase 2 (COX2) | Inducible, converts arachidonic acid to prostaglandin G/H |
| Prostaglandin E synthase | PTGES | Converts prostaglandin H to prostaglandin E | |
| Caspase 1 | CASP1 | Interleukin-1β converting enzyme (ICE) | Activation of IL1β and IL18 |
| Caspase 3 | CASP3 | Mediates apoptotic pathway | |
| Caspase 8 | CASP8 | Mediates apoptotic pathway | |
| Indoleamine 2,3-dioxygenase | IDO | Metabolism of tryptophan, immunoregulation | |
| Interleukin-1 receptor-associated kinase | IRAK | Activation of TRAF6, 3 | |
| IκB kinase | IKBK | Activation of NFκB | |
| Mitogen-activated protein kinase 14 | MAPK14 | p38 MAP kinase | Stress/inflammatory signaling |
| Mitogen-activated protein kinase 8 | MAPK8 | JUN N-terminal kinase (JNK) | Stress/inflammatory signaling, activation of AP-1 |
| Mitogen-activated protein kinase 3 | MAPK3 | Extracellular signal-regulated kinase 1 (ERK1) | Stress/inflammatory signaling |
| Mitogen-activated protein kinase 1 | MAPK1 | Extracellular signal-regulated kinase 1 (ERK2) | Stress/inflammatory signaling |
| Akt protein kinase | AKT | Protein kinase B | Cell survival, stress/inflammatory signaling |
| Tyrosine-protein kinase Src | SRC | Immunoregulation | |
| Janus kinase | JAK | Activation of STAT | |
| Phosphatidylinositol-4,5-biphosphate 3-kinase | PIK3 | Phosphoinositide-3-kinase (PI3K) | Stress/inflammatory signaling |
| Protein kinase, cAMP-dependent | PRKA | Protein kinase A | Protein phosphorylation, signaling |
| Protein kinase C | PRKC | Protein phosphorylation, signaling |
Inflammation
Inflammation is the immune system’s initial response to a pathological challenge and involves major changes in homeostasis: fever, activation of immune cells, increased blood flow at the site of inflammation, and enhanced pain sensitivity.119 Apart from specific pathogens, the inflammatory cascade can be triggered by tissue injury, insoluble particulates, activation of the plasma complement and clotting/fibrinolytic protease pathways, opsonized (antibody or complement-coated) particles or immune complexes, and intracellular components released by tissue damage. Monocytes and macrophages are by far the most effective promoters of the inflammatory response. Mast cells regulate vascular permeability, vasodilation, and leukocyte recruitment and play a critical role in allergic inflammation.120 Neutrophils provide rapid and effective clearance of extracellular pathogens.121 These immune cell types also exert a regulatory influence over the subsequent immune response.
In addition to the pro-inflammatory cytokines, the main mediators of the inflammatory process are pro-inflammatory prostaglandins and leukotrienes, neuropeptides such as substance P, vasoactive amines produced by basophils and mast cells (histamine and serotonin) and acute phase proteins from the liver, such as C-reactive protein and serum amyloid A. Local production of chemoattractive cytokines (chemokines), such as IL8 and chemokine (C–C motif) ligand 2 (CCL2; monocyte chemoattractant protein-1), together with upregulation of specific adhesion molecules on the endothelium (selectin E, selectin P, intercellular adhesion molecule-1 (ICAM1), vascular adhesion molecule-1 (VCAM1)) and on the leukocytes (selectin L, integrins), allow circulating neutrophils, monocytes and lymphocytes to specifically target and enter the affected tissues.122 Resolution of the inflammatory response involves production of anti-inflammatory immunoregulatory cytokines, such as transforming growth factor-β (TGFβ) and IL10, some prostaglandins, late-acute-phase proteins, and activation of the hypothalamic-pituitary-adrenal axis to produce anti-inflammatory corticosteroids.123., 124., 125. Long-term consequences of inflammation include increased tissue fibrosis and persistent alterations in the number, type, and activity of leukocytes within the affected tissue.126
The main effectors of the innate immune system are hydrolytic enzymes (e.g., lysozyme, serprocidins), antimicrobial proteins (e.g., complement, defensins), cytotoxic reactive oxygen species (ROS), NO, cytotoxic cytokines such as TNF, and the antiviral interferons.119 Moreover, activation of the innate immune system leads to recruitment of the adaptive immune system.
The Adaptive Immune Response
The adaptive immune system involves complex cellular interactions that promote the functional maturation and expansion of regulatory and effector lymphocytes, thereby providing efficacy, specificity, and memory to the immune response, but requiring time to become effective. It operates through the ability to recognize and respond to molecular motifs (antigens), usually associated with proteins, that are not part of the normal host repertoire, and may indicate the presence of an external threat. Adaptive immunity is responsible for autoimmune reactions in the male reproductive tract that can lead to infertility and chronic inflammatory conditions. These reactions are mediated, but also regulated, by T cells, B cells, and NK cells, which have access to the male reproductive tract.
T-Cell and B-Cell Regulation and Functions
The adaptive immune system is dependent upon the unique ability of lymphocytes to generate a vast repertoire of cell surface receptors that can bind to almost any conceivable molecular surface, without ever having encountered the molecule before. On B cells, these receptors are surface-bound immunoglobulins, from which are derived the circulating antibodies. The core proteins of the T-cell receptor (TCR) are structurally related to the immunoglobulins, but the TCR itself is a complex of interacting surface proteins.127 The diversity of these receptors involves extensive rearrangements of the basic immunoglobulin and TCR gene regions, which produce randomly reassembled genes encoding proteins, each with a very specific and unique topography.128., 129. Each precursor T cell and B cell expresses a surface receptor that is specific for a unique antigenic determinant and all their offspring (clones) will express the same receptor and specificity. B cells interact more or less directly with the antigenic molecule in situ. However, more precise regulation of the immune response involving T cells is determined by proteins of the highly polymorphic major histocompatibility complex (MHC), expressed on the surface of antigen-presenting cells.130 Almost all cells in the body can act as antigen-presenting cells by proteolytically converting intracellular proteins, of either endogenous or infectious (e.g. viral) origin, into short antigenic peptides, which are then incorporated into a structural groove on the extracellular surface of the MHC protein complex during its assembly in the endoplasmic reticulum.131 Some antigen-presenting cells (dendritic cells and macrophages) are able to phagocytose exogenous proteins, typically proteins of pathogenic origin, but also proteins derived from endogenous sources including the spermatogenic cells, and process these proteins for antigen-MHC complex formation. The TCR subsequently binds to the antigen-MHC complex on the surface of the antigen-presenting cell leading to the activation and proliferation of the T cell (Figure 19.6 ).
FIGURE 19.6.
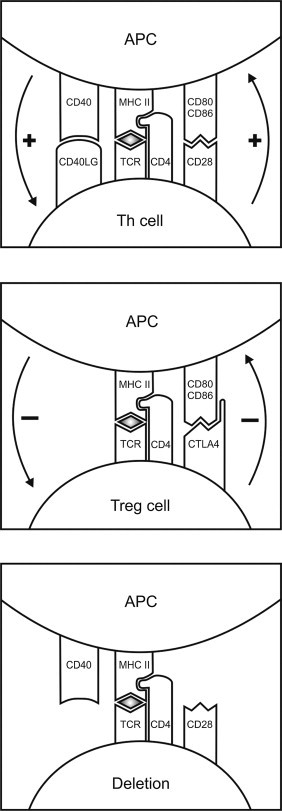
The antigen-presenting cell (APC)–T-cell synapse and the adaptive immune response.
Recognition of the MHC class II-peptide antigen complex by the T-cell receptor (TCR) of a naïve Th cell together with engagement of the CD28:CD80/CD86 and CD40/CD40LG receptor/co-receptor pairs can lead to generation of Th1 cells, if type 1 cytokines (IL12 and IFNγ) are present. If interleukin-6 (IL6) and type 2 cytokines (IL4, IL5 and IL13) are present, Th2 cells are produced, and Th17 cells are produced when IL6 and transforming growth factor-β (TGFβ) are present. Engagement between the APC and T cell via the CTLA4 receptor produces an inhibitory response, as occurs in Treg cell interactions. Engagement of the APC and T cell in the absence of adequate co-stimulation or cytokine activity results in deletion or inactivation (anergy) of the Th cell.
Typically, circulating T cells express one of the co-receptor proteins, CD4 and CD8, as part of their TCR, which permit them to recognize antigens associated with MHC class II or MHC class I molecules, respectively.132 Antigens are presented to CD4+ T cells by the professional antigen-presenting cells that express MHC class II antigens (dendritic cells, macrophages, and B cells).133 On the other hand, CD8+ T cells are recognized by MHC class I antigens, which are ubiquitously expressed. Activation of the T cell requires physical interaction between co-stimulatory ligand-receptor pairs, particularly CD28:B7 (CD80/CD86) and CD40:CD40 ligand (CD40LG), and production of either type 1 cytokines [IL2, IL12 and interferon-γ (IFNγ)] or type 2 cytokines (IL4, IL5, IL10 and IL13; Figure 19.6).134., 135. As a result of this complexity, T-cell activation can produce different outcomes depending upon the co-stimulatory molecules engaged and cytokines produced. Accordingly, naïve CD4+ T cells may become type 1 helper (Th1) cells, which produce Th1 cytokines, direct the development of the cellular immune response involving cytotoxic CD8+ T cells, and are associated with graft rejection, or they may become type 2 helper (Th2) cells, which produce Th2 cytokines, promote B cell development and antibody responses, and regulate allergic responses (Figure 19.7 ).134., 135.
FIGURE 19.7.
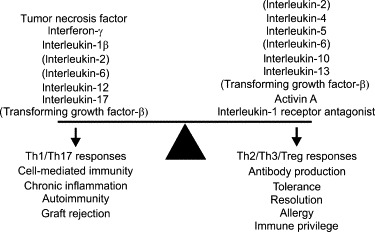
The cytokine balance and immune deviation.
Cytokines can be designated either pro-inflammatory or anti-inflammatory/immunoregulatory, depending upon their predominant activities. The former group is associated with Th1 or Th17 type responses (cell-mediated immunity, autoimmunity) and the latter with Th2, Th3, or Treg type responses (antibody production, allergy, tolerance). It is important to note that several cytokines, such as IL6 and TGFβ, possess both pro- and anti-inflammatory properties, and fall within both groups. It is the combination of cytokines present that determines the T-cell response outcome.
In addition to the Th1 and Th2 cell subsets, which are primarily responsible for dictating the nature of the subsequent immune response to a particular antigen, there are a number of other T-cell subsets with specialized functions, which have been more recently described. The Th17 cell subset is functionally related to the Th1 subset but characteristically produces IL17, IL21, and IL22.136 The Th17 cell is generated by exposure to IL6 and TGFβ and regulates protection against extracellular pathogens, including recruitment and activation of neutrophils, differentiation of B cells, and the inflammatory activity of epithelial cells.137 However, these cells also have been implicated in the development of autoimmune disease and in allergy.136., 138. Absence of appropriate co-stimulatory molecule interactions and/or the presence of anti-inflammatory or immunosuppressive cytokines may lead to T-cell inactivation (anergy) and deletion or the generation of regulatory or suppressor T cells (Figure 19.6).139., 140. These T-cell subsets are responsible for regulating antigen-specific immunity and maintaining peripheral immune tolerance.
The development of B cells into antibody-secreting plasma cells following interaction with antigen requires specific Th2 cell help.141 Once activated, these cells initially secrete multivalent IgM, but the cells gradually mature to produce high affinity IgG with the same antigenic specificity.128 Finally, following the resolution of the immune response, at least some activated T and B cells clones persist as memory cells, with the result that lymphocyte responses to antigens generally develop much faster upon second exposure to the antigen.142., 143.
NK Cell and NK T-Cell Regulation and Functions
NK cells are lymphocytes that span the interface between the innate and adaptive immune systems.144 These cells are developmentally related to cytotoxic CD8+ T cells but are capable of recognizing and destroying transformed cells, such as virally-infected or tumor cells, without the need for prior sensitization by exposure to antigen. They can be activated by pro-inflammatory cytokines, notably IL12 and antiviral type 1 interferons.145 They interact with their targets via a complex of stimulatory and inhibitory surface receptors, which allow them to recognize ligands uniquely expressed by transformed cells and to detect the lack of MHC class I expression, a characteristic of transformed cells.144., 146. Accordingly, NK cells are able to rapidly mobilize against pathogenic challenges long before T- and B-cell responses can develop. However, activated NK cells also participate in the adaptive immune response. They possess antigen-specific receptors, similar to cytotoxic CD8+ T cells, and have been shown to regulate dendritic cell responses in a positive or negative manner, for example, by inducing dendritic cell death through contact-mediated lysis or by producing immunoregulatory cytokines, such as IFNγ, IL4, and TGFβ.144., 147.
NK T cells are a distinct cytotoxic lymphocyte subset that is defined by responsiveness to bacterial and mammalian glycolipid antigens presented by the nonclassical MHC class I molecule, designated CD1D.148 These cells are considered to be T cells with NK activity, rather than NK cells per se, but they also appear to play an important role in immune regulation through production of either IFNγ or IL4/IL10.149., 150. Accordingly, both NK T cells and NK cells promote and mediate immunity to bacteria, viruses, and tumors, but are also capable of suppressing cell-mediated autoimmunity and graft rejection responses.
Immunological Tolerance and Regulatory Lymphocyte Subsets
The ability of randomly-generated, antigen-specific T and B cell clones to ignore antigens expressed by the host organism relies upon an effective cell-editing process, collectively referred to as tolerance. The tolerogenic process involves central and peripheral mechanisms, both of which play a crucial role in protecting the spermatogenic cells and other antigenic components of the male reproductive tract from the immune system. Central tolerance occurs primarily in the thymus, during normal development, when expression of antigens by the thymic epithelium leads to functional deletion of self-reactive T cells by inactivation and apoptosis.1 A similar mechanism mediates B-cell editing in the bone marrow.151 It is now recognized that this process involves the promiscuous expression of many tissue-specific antigens in the thymic epithelium induced by the transcription factor, autoimmune regulator (AIRE).152 Even sperm-specific antigens have been found to be expressed in the thymus under this mechanism.153 However, this mechanism is not completely efficacious because autoreactive lymphocytes persist and may expand or become activated later in life, resulting in autoimmune disease.154., 155.
The mechanisms of peripheral tolerance, which need to operate effectively throughout life, are more complex. Peripheral tolerance involves functional deletion of autoreactive lymphocytes in the peripheral (secondary) lymphoid tissues due to weak antigen-stimulation in the absence of appropriate co-stimulation, production of blocking and anti-idiotypic antibodies, and development of antigen-specific regulatory and suppressor lymphocytes.139., 156. This involves an ongoing process of low-dose exposure of these cells to their antigen, accompanied by either modified co-stimulatory or specific immunoregulatory signals from antigen-presenting cells, regulatory lymphocytes, or other cell types in the vicinity.157 This means, for example, that engagement of the peptide-MHC class II complex with the TCR in the absence of linkage of CD80/CD86 to CD28, in the presence of immunoregulatory cytokines, such as IL10 or TGFβ, or engagement by CD80/86 of the inhibitory T cell receptor, CTLA4, in place of CD28, leads to T-cell deletion by inactivation or apoptosis, and/or induction of regulatory/suppressor T-cell activity (Figure 19.6).140., 158. The best characterized, and arguably the most important, of these regulatory lymphocytes is the CD4+CD25+ regulatory T cell (Treg) subset, which expresses the transcription factor, FOXP3.140 These cells can be induced by TGFβ and IL2, and selectively produce immunoregulatory TGFβ and IL10, but also appear to act via direct contact with antigen-presenting cells and other T cell subsets.159., 160. Both NK cells and NK T cells are also capable of antigen-recognition and immunosuppressive activity. Other T cell subsets implicated in immunosuppression through their ability to produce TGFβ and IL10 include Th3 cells, Tr1 (T regulatory 1) cells and γδ T cells.161., 162., 163.
Autoimmunity and Rejection Responses in Immunity
Autoimmune disease generally represents the failure of tolerance. Somatic mutation of the antigen-receptor expressed by a T cell or B cell may result in the creation of new self-reactive clones, thereby subverting central tolerance.164 Autoimmunity may result from an inflammatory response to an infection that damages or overwhelms normal mechanisms of self-tolerance, or where the infection involves organisms that express antigens that may cross-react with self-antigens (molecular mimicry).165 Studies of disease models in humans and experimental rodents have also established that autoimmunity may have a genetic basis, as in the case of polyglandular autoimmune (PGA) syndromes. In humans, type 1 PGA is associated with a mutation in the AIRE transcription factor,166 while type 2 PGA is related to a defect in regulatory T cell function.167
Failure of tolerance results in autoimmunity, but failure to induce tolerance in the first place lies behind transplantation rejection responses. The leukocytes of both the graft recipient and the donor tissue react toward their respective antigens, leading to the rejection of tissues that are not antigenically matched, in a process called graft-versus-host disease.168 The response is said to be allogeneic if it occurs across genetic boundaries within the same species (allograft), while xenogeneic responses involve a graft and host belonging to different species (xenograft). The extensive polymorphism of the MHC, which is also called the human leukocyte antigen (HLA) complex in the human, is a major contributor to the allogeneic rejection response. Rejection responses typically involve the classical MHC class 1a (HLA-A, HLA-B and HLA-C) and MHC class II (HLA-D) antigens responsible for antigen presentation during T-cell activation.131 On the other hand, nonclassical MHC antigens that are able to inhibit T cell and NK cell activity, such as HLA-G and HLA-E, are associated with suppression of the adaptive immune response and maintaining peripheral tolerance.169 Consequently, both classical and nonclassical MHC are involved in regulating autoimmune responses, including autoimmunity in the male reproductive tract.
Initiation and Control of the Immune Response and Immunity
Antigen-specific immune responses are initiated in the secondary lymphoid tissues, particularly within the follicles and germinal centers of the draining lymph nodes and spleen or lymphoepithelial aggregates of mucosal tissues, where antigen-presenting cells come into contact with large numbers of T and B cells.170 This implies that antigens normally must travel via the lymphatics to one of these organs, either by simple diffusion or carried by an antigen-presenting cell. Generally, immunologists do not consider that primary immune activation can occur within nonlymphoid tissues or assume that any response generated there is likely to be vigorous enough to lead to full participation by the immune system. However, there is some evidence that primary activation can occur outside the lymphoid tissues, especially during graft rejection.171., 172.
Moreover, an alternative immune activation model has been proposed that combines elements of both the innate and adaptive immune system. This is called the danger hypothesis, which proposes that antigen-presenting cells responds to substances that cause or signal damage, rather than to those that are simply unrecognized.173 These danger signals include CD40LG, the early pro-inflammatory cytokines IL1β and TNF, interferons, and heat-shock proteins, as well as substances that are normally only found inside cells (e.g. nucleotides, unmethylated CpG sequences in mammalian double-stranded DNA) and hyaluron breakdown products.173 In this model, activation of the immune system occurs as a response to evidence of an extant threat rather than toward a specific feature of the threat itself. This mechanism may serve to explain the onset of certain autoimmune diseases.
Inflammation and immunity are damaging to the tissues in which they occur. Apart from the normal mechanisms of tolerance, the immune system needs to limit inflammation and immune responses to minimize such damage. Accordingly, inflammation triggers secretion from the adrenal gland of glucocorticosteroids, which repress expression of NFκB, inhibit the production of pro-inflammatory cytokines and mediators while stimulating the anti-inflammatory cytokines, IL4 and IL10, reduce expression of leukocyte adhesion molecules, and induce lymphocyte apoptosis.124., 174., 175., 176., 177. The immune cells themselves produce other anti-inflammatory and immunosuppressive molecules, including prostaglandins D and J and lipoxins.178., 179. Moreover, activated lymphocytes have a limited lifespan, undergoing a process of activation-induced cell death following upregulation of the extrinsic apoptotic signal mediated by interaction of the FAS receptor with its ligand (FASL), eventually leaving behind a relatively small population of long-lived memory cells.143., 180. There are also specific mechanisms to inhibit antibody activity and promote antibody clearance, such as the induction of anti-idiotype (antitopographical) antibodies.181
Mucosal Immunity
Muscosal immunity concerns immunity at the interface between the external and internal environments. Mechanisms involved in mucosal immune responses are very similar to those of other tissues and lymphoid organs. Discrete lymphoepithelial aggregates (mucosa-associated lymphoepithelial tissue or MALT) within mucosal surface tissues allow interaction between antigen-presenting cells and lymphocytes to facilitate local immune responses.182 Reactions to inhaled or digested antigens are controlled through a process called mucosal tolerance,162., 182. and failure of tolerance in the gastrointestinal and respiratory tracts may result in allergies. The mechanisms of mucosal tolerance involve clonal T-cell deletion or anergy and active suppression by regulatory/suppressor T cells.183 Although CD8+ T cells predominate in the mucosal epithelium, it is the CD4+ T cells that are responsible for mucosal tolerance, producing IL4 and IL10 (Th2 cells), and TGFβ (Th3 cells).162., 183., 184. Mucosal intraepithelial lymphocytes also express CD1D constitutively and can activate immunoregulatory NK T cells in the epithelium.185
A key feature of mucosal immunity is the isotype of the antibodies normally involved at mucosal surfaces. Tight junctions between the epithelial cells restrict passage of both antigen and antibody, so that antibody responses are dominated by secretory IgA, which lacks the ability to activate complement and possesses strong anti-inflammatory properties.186 In order to cross epithelial barriers, a unique mechanism for transport of dimers of IgA through the epithelial cells involving a protein called secretory component is necessary.187 Thus, IgA is generally the major immunoglobulin in secretions of mucosal surfaces. In mucosa, the epithelial cells also play an important role in host defense by producing many molecules associated with innate and adaptive immunity, including defensins, regulatory cytokines, TLRs, MHC antigens, leukocyte adhesion molecules, and co-stimulatory molecules.188., 189.
There is strong evidence that the male reproductive tract is a constituent of the mucosal immune system.91., 190. As far as the male reproductive tract is concerned, both IgA and serum-derived IgG appear in semen and bound to the sperm of men with autoimmune infertility.3., 45., 191. It would be expected that the epithelial cells of the male urogenital tract might play a similar active role in regulating local immune responses. Although discrete MALTs have never been described within the male tract, the existence of similar or analogous structures cannot be entirely excluded.
Regulation of Immunity by Hormones and Gender
Corticosteroid Control of Inflammation and Immunity
Corticosteroids are essential physiological regulators of the immune response, acting through the ubiquitously expressed glucocorticoid receptor to exert negative feedback on the inflammatory response and control innate and acquired immunity at multiple levels.124., 174., 175., 176., 177. Clinically, they are widely used in the treatment of autoimmune and inflammatory diseases. The particular relevance of corticosteroids for male reproductive function is the fact that they have inhibitory effects on testicular development and suppress Leydig cell steroidogenesis at all levels of the hypothalamic-pituitary-testicular axis with the result that activation of the hypothalamic-pituitary-adrenal axis during inflammation negatively impacts upon male reproductive function (Figure 19.8 ).192
FIGURE 19.8.
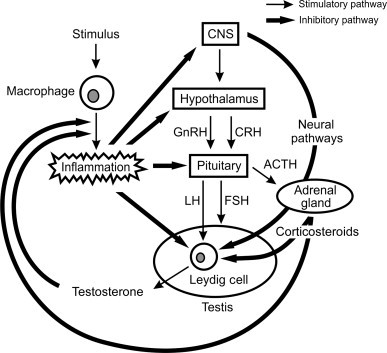
The interface between the hypothalamic-pituitary-gonadal-adrenal axis and inflammation.
Regulation of the testes and adrenal glands is under the control of the central nervous system (CNS), which integrates information from the periphery and modulates secretion of the gonadotropin-releasing hormone (GnRH) and corticotropin-releasing hormone (CRH) by the hypothalamus. These hypothalamic peptides stimulate secretion of the gonadotropins (LH and FSH) and adrenocorticotropic hormone (ACTH), respectively. LH acts directly on the Leydig cells to stimulate steroidogenesis, while FSH controls the development and activity of the Sertoli cells. ACTH stimulates secretion of corticosteroids by the adrenals, which generally exert direct inhibitory effects on Leydig cell steroidogenic activity. Activation of macrophage function by an inflammatory stimulus triggers a cascade of events and secretions, which interact with the hypothalamic-pituitary-gonadal axis at all levels, inhibiting gonadotropin secretion and steroidogenesis and stimulating the hypothalamic-pituitary-adrenal axis. There is evidence that inhibitory regulation of steroidogenesis in response to inflammation also involves direct neural pathways from the CNS. As a consequence, inflammation profoundly inhibits the ability of the Leydig cell to produce testosterone. Eventually, steroids produced by the adrenals and testis exert feedback inhibitory effects on the inflammatory process, bringing about the resolution of the inflammation and recovery of testicular testosterone production.
Sex-Specific Regulation of Immune Function
The male reproductive system exerts a profound inhibitory effect on the development and function of the immune system. Compared with females, males have lower serum immunoglobulin levels, reduced cellular immunity and less effective responses to antigenic challenge, while the incidence of autoimmune diseases are generally far less common and less severe in males than in females.193 This can be attributed, in part, to genetic and epigenetic differences between the sexes, as indicated by studies in sex-reversed mice, which established that XX-bearing mice displayed greater susceptibility to autoimmune disease than XY-bearing mice, independent of gonad type or hormones.194 However, testis ablation and male sex steroid replacement studies have also established a critical role for products secreted by the testis in this divergence. Such studies demonstrated that androgens inhibit many immune parameters, particularly the size of the thymus and other immune tissues, lymphocyte number and activity, and antibody production, but also graft rejection, autoimmune responses, and resistance to infection.195., 196., 197. These findings have been supported in more recent years by studies in transgenic animals lacking the androgen receptor,198., 199. and by clinical data from hypogonadotropic men.200., 201. Studies on macrophages and other androgen-responsive cell types in vitro have indicated that androgens are able to inhibit NFκB and expression of inflammatory genes, such as TLR4, IL1, and TNF.202., 203., 204. On the other hand, androgens appear to stimulate neutrophil proliferation growth and function through activation of the extracellular signal-regulated (ERK) kinases (mitogen-activated kinase 3/mitogen-activated protein kinase 1; MAPK3/MAPK1) and production of the neutrophil growth factor, granulocyte colony stimulating factor.198., 205.
The classical androgen receptor is a cytoplasmic protein that binds androgens with high affinity, and translocates to the nucleus, where it acts as a transcription factor by binding to androgen-response elements in the promoter of responsive genes.206 Studies by early researchers established that functional androgen receptors were primarily expressed on the stromal and epithelial cells of the immune tissues,207., 208. suggesting that the effects of androgens on immunity were exerted indirectly at the tissue level, rather than by direct effects on the circulating lymphocytes. In fact, the expression and relative importance of the classical androgen receptor on lymphocytes remains somewhat equivocal.209., 210. However, it is now clear that steroids can also interact with membrane-bound G protein-coupled receptors to trigger nongenomic responses in target cells.211., 212. Studies have shown that androgens can alter [Ca] fluxes in lymphocytes and macrophages via such membrane-mediated interactions,213., 214. and that this signaling affects gene expression and function in the target cells.215
Obviously, many questions remain concerning the detailed cellular mechanisms that mediate the actions of androgens on immune cells, but it is clear that these steroids have profound effects on immune responses and are capable of directly modulating these functions within the male reproductive tract and adjacent draining lymph nodes. Furthermore, this regulation may also involve other steroids produced by the testis, such as progesterone and the estrogens, which also have direct effects on immune cell function.196., 216. Estrogens, in particular, have been implicated in regulating the differentiation and maturation of dendritic cells, and inhibiting their co-stimulatory activity.217., 218. There is even evidence that LH and FSH themselves directly regulate macrophage and lymphocyte growth and cytokine production acting through specific receptors expressed by these cells.219., 220.
Protein hormones and neuropeptides produced by the testis have direct effects on immunity. These include the pro-opiomelanocortin gene-derived peptides β-endorphin and α-melanocyte-stimulating hormone (α-MSH), which are produced by the Leydig cells and macrophages of the testis, and testicular GnRH.221., 222., 223. The testis is a significant source of cytokines with immunoregulatory activity, specifically, members of the highly immunosuppressive/anti-inflammatory TGFβ family.224., 225. There is no doubt that exposure of circulating immune cells to these molecules within the testis, and/or their secretion into the blood and lymph, plays a significant role in local immunoregulation and sexual dimorphism of immune function.
Immune Cells of the Male Reproductive Tract
Far from being a site where the immune system is restricted entry, macrophages, lymphocytes and granulocytes are characteristic features of the male reproductive tract (Table 19.3 ). There are substantial differences in the number and type of these cells within the different tissues and from species to species, which have important implications for understanding the immunophysiology of the male tract.
TABLE 19.3.
Quantification of Immune Cells in the Normal Testis of Adult Rats and Humansa
| Cell Type | Rat Testis (× 106/g tissue) | Human Testis (× 106/g tissue) |
|---|---|---|
| Macrophages | 5–10 | 10–25b |
| Dendritic cells | 0.2–0.3 | Present |
| T cells | 1–2 | 1.4–2.4 |
| CD8+ T cells | 0.6–1.8 | Present |
| CD4+ T cells | 0.2–0.3 | Present |
| NK cells | 0.6–1.0 | 1.0–2.8 |
| Mast cells | Capsule region only | Present |
Estimates based on data from stereological analysis of testes from Sprague–Dawley rats and from adult human testes with normal spermatogenesis (Hedger MP and Hayes RD, unpublished data).69., 70., 226., 227. The study of Vergouwen and colleagues228 indicates that CBA/P mouse testes contain approximately 2–4 × 106 macrophages/g tissue, but there are no definitive quantitative studies of other leukocyte subsets in the mouse testis.
Upper limit calculated from data obtained by Frungieri and colleagues229 using a well-characterized monoclonal antibody against CD68. The observation that macrophage numbers in the normal human testis are at least as large, if not larger, than those found in the either the rat or mouse testis is consistent with nonquantitative observations using several macrophage markers.67., 230., 231., 232.
Macrophages and Dendritic Cells in the Testis
Macrophages and dendritic cells are found in every tissue of the body. Although some dendritic cells have a lymphoid origin,233 these cell types are chiefly derived from the circulating monocyte pool and are directed along tissue-specific developmental pathways by the influence of the local immunological environment.234 As a consequence, macrophages, in particular, are extremely heterogeneous in appearance and function.235., 236. Superficially, the microglial cells of the brain and the Kupffer cells of the liver have little in common, but both cell types are macrophages and share common features: they express markers of the mononuclear phagocyte lineage, are actively mobile, phagocytic, cytotoxic, and are involved in tissue restructuring and antigen-presentation to CD4+ T cells.
Distribution and Properties of the Testicular Macrophage Population
Early interest in testicular macrophages arose from studies on the accumulation of nondigestible tracers and radionuclides in the testicular interstitial tissue.237., 238. In spite of some early speculation that these cells might have a nonhematopoietic origin, studies by Miller and colleagues,237., 239., 240. and subsequently by Hutson and colleagues,241., 242., 243., 244. established that the testicular macrophages share the classical characteristics of resident macrophages or tissue-fixed macrophages. They display the characteristic nuclear and cytoplasmic morphology of the mononuclear phagocyte lineage, are actively phagocytic, bactericidal and adherent in culture, and they express macrophage-specific enzymes, cytokine receptors, and surface antigen markers.69., 239., 240., 243., 245., 246., 247.
There are substantial populations of resident macrophages in both the rat and mouse testes.69., 228., 248., 249., 250. Other species with large numbers of testicular macrophages include the guinea pig, hamster, horse, bull, and human.229., 251., 252., 253. These cells are almost entirely confined to the interstitial tissue under normal (noninflamed) conditions. In the boar, which has a relatively sparse testicular interstitial connective tissue and very large numbers of Leydig cells, macrophages appear to represent a smaller proportion of total interstitial cells.254 Curiously, the ram testis appears to possess only small numbers of recognizable macrophages, in spite of the general similarity of the testicular interstitial tissue and lymphatic organization in this species with that of the bull or human.52., 68. Testicular macrophages have been most intensively studied in the rat, with less extensive investigation in the mouse, and relatively limited investigations in other species. It generally has been assumed that the rat testicular macrophages are representative, but data suggest that there are significant functional differences even between rat and mouse testis macrophages. Further study on the macrophages of other species, particularly the human and primates, is clearly necessary, and discussion of this complex cell type in the testis necessarily reflects the rather narrow available knowledge base.
In the rat and mouse, the ratio of macrophages to Leydig cells appears to be relatively fixed at approximately one macrophage to every four or five Leydig cells,245., 248., 249., 250. and macrophages display a very close physical and functional relationship with the Leydig cells. Ultrastructural studies have established the existence of highly-specialized cytoplasmic interdigitations linking the two cell types, indicating the potential for direct exchange of information and material,60., 240., 244., 255. while macrophages and Leydig cells undergo parallel alterations in morphology and cytoplasmic volume in experimental models of cryptorchidism and vasectomy in the adult rat.256., 257. During aging, testicular macrophages retain their morphological association with Leydig cells, but the cytoplasmic interdigitations are lost.258 Aging testicular macrophages also acquire lipofuscin granules similar to those observed in aged Leydig cells.258 Most surprising, however, was the discovery that macrophages play an essential role in normal Leydig cell development and in maintaining Leydig cell steroidogenic function in the adult.208., 259., 260., 261., 262., 263., 264., 265. It appears that the testicular macrophages have an integral role in testicular biology that goes beyond the typical functions of tissue-fixed macrophages in phagocytosis of cellular debris, connective tissue remodeling, maintenance of innate immunity, and regulation of adaptive immunity.
The immunological functions of the testicular macrophage population have been investigated in a number of studies, but the data are by no means comprehensive and many questions still remain. Although several studies have shown that the majority of rat macrophages express MHC class II molecules under normal conditions, indicating a capacity for antigen-presentation to helper and regulatory T cell subsets,68., 246., 266. Tung and colleagues reported that strong class II expression in the normal mouse testis was restricted to cells located around the rete testis and was only upregulated in the interstitial tissue following immunization with testicular antigens in the presence of adjuvant.267., 268. This seems to indicate a fundamental difference in baseline activation status and antigen-presenting activity between the testicular macrophages of the rat and mouse. Human testicular macrophages appear to be more like those of the rat, with significant MHC class II expression found throughout interstitial tissue samples from normal men.67., 230. Studies of the ability of rat testicular macrophages to provide help for T cell activation in vitro indicate that these cells are deficient in co-stimulatory activity.269., 270. In the mouse testis, a lack of expression of the essential co-stimulatory molecules for T cell activation, CD80 and CD86, has been reported,271 but this does not appear to be true for the rat.272 With respect to inflammation, selective depletion of testicular macrophages in the rat with a macrophage-specific toxin exacerbated the testicular inflammatory response induced by hyperstimulation of the Leydig cells with pathological doses of the LH agonist, human chorionic gonadotropin (hCG).273 Significantly, rat testicular macrophages display relatively poor expression of critical proinflammatory genes and proteins, including IL1β, TNF, IL6, IL12, NOS2, and macrophage migration inhibitory factor (MIF), in response to activation by bacterial LPS or regulatory cytokines269., 274., 275., 276., 277., 278.; however, these cells express constitutively high levels of the anti-inflammatory/immunosuppressive cytokine, IL10.270 Examination of murine testicular macrophages indicates a similarly reduced pro-inflammatory capacity (Hedger MP and Winnall WR, unpublished data).
The accumulated data confirm that macrophages from both rat and mouse testes, although different in some functional respects, have reduced inflammatory and co-stimulatory activities. Macrophages displaying such properties are called alternatively activated or M2 macrophages (analogous to Th2 cells); they are characteristic of tumors and late-stage inflammation.236., 279. During inflammatory and immune responses, M2 macrophages serve to regulate the activity of pro-inflammatory or classical M1 macrophages to moderate cell-mediated immunity, and thereby play important roles in reducing autoimmune and rejection responses.
Heterogeneity of the Testicular Macrophage Population
Although testicular macrophages collectively display an alternatively activated phenotype, quantitative studies of macrophage numbers using specific antigenic markers in the rat testis indicate that testicular macrophages do not represent a homogenous population.69., 226., 276. The majority of testicular macrophages in the rat express a molecule that is specific to tissue-fixed macrophages in most nonlymphoid tissues, recognized by the monoclonal antibody, ED2.280 This molecule has been identified as CD163, a member of the group B scavenger receptor cysteine-rich protein superfamily.281 It has been implicated in the stimulation of IL10 production by monocytes and macrophages following the endocytosis of hemoglobin:haptoglobin complexes, and is highly expressed by M2 macrophages.282., 283. However, a significant subset of testicular macrophages in the rat (about 15–20% of the total) do not express this marker. These macrophages can be identified by expression of the lysosomal antigen, CD68, recognized by antibody ED1, which suggests they may be recently recruited or even transient macrophages.69., 268., 276., 284. Moreover, about half of the CD163+ subset appears to have lost CD68 expression. These data indicate the existence of several populations of macrophages in the normal rat testis, putatively representing different stages of development and/or functional states (Figure 19.9 ).284
FIGURE 19.9.
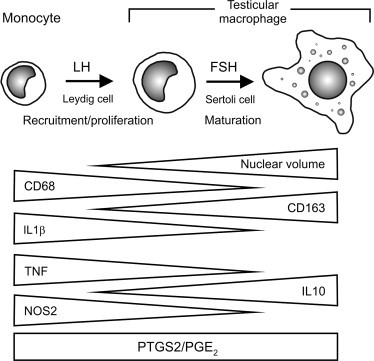
Maturation of the resident macrophage population of the rat testis.
Macrophages in the rat testis are heterogeneous, corresponding to different stages of maturation from circulating monocytes through to a distinct testicular resident macrophage phenotype. This testicular phenotype is characterized by an increased nuclear and cytoplasmic volume, loss of the CD68 marker recognized by antibody ED1, upregulation of the resident macrophage surface marker ED2 (CD163), loss of ability to produce pro-inflammatory mediators and constitutive production of the immunoregulatory cytokine, interleukin-10 (IL10). Expression of the inducible form of prostaglandin-endoperoxide synthase (PTGS2) and the ability to produce prostaglandins, such as PGE2, does not appear to change. Recruitment of macrophages to the testis and/or their proliferation is under the control of LH, acting via the Leydig cells, while maturation to the mature testicular phenotype appears to be FSH-dependent, indicating regulation by the Sertoli cells.
This heterogeneity may have functional correlates. Compared with CD68− macrophages, CD68+ testicular macrophages possesses a distinctly smaller nuclear diameter and displayed overall higher levels of expression of NOS2 and CCL2 in normal and LPS-stimulated rats, based on immunohistochemical studies.276., 285. Expression of IL1β protein after LPS treatment appears to be confined to a relatively small subset of testicular macrophages.274 Since the CD68+CD163− macrophage subset increases in number in the testis after LPS treatment, it may be assumed that this subpopulation represents newly-arrived pro-inflammatory macrophages. However, when macrophages were isolated from noninflamed rat testes on the basis of CD163 expression by flow cytometery, the CD163− subpopulation displayed very poor production of either proinflammatory cytokines or IL10 in response to LPS in vitro.270 Altogether, these data suggest that the CD68+CD163− subpopulation present in the normal testis comprises recently recruited or transient macrophages, and it is functionally distinct from the proinflammatory CD68+CD163− macrophages that invade the rat testis following an LPS challenge in vivo.276., 286.
There is evidence for macrophage heterogeneity in mouse and human testes, as well. Studies have shown that macrophages isolated from the mouse testis are heterogeneous in their ability to produce TGFβ in vitro,287 and approximately 20% express high levels of the alternative-activation marker, Ym1.288 In the human testis, Frungieri and colleagues found that CD68+ macrophages displayed substantial variation in expression of CD163.229 All these observations are consistent with the hypothesis that the testis contains subsets of macrophages with varying pro-inflammatory capacities, although it appears that an alternatively activated, anti-inflammatory/immunosuppressive phenotype predominates under normal conditions.
Intratesticular Dendritic Cells
Dendritic cells, which are distinguishable from macrophages by their distinctive morphology and expression of specific functional markers, such as integrin αx (ITGAX; CD11c) and integrin αE2, have been observed in the rat, mouse, and human testis.230., 266., 289., 290., 291. Dendritic cells are found throughout the interstitium, but they are much less prominent than the testicular macrophages. In spite of the common lineage, dendritic cells lack the efficient phagocytic and cell-killing capabilities of macrophages, although they are much more effective as antigen-presenting cells.133 Crucially, dendritic cells are able to promote either cell-mediated immunity or tolerance to specific antigens, depending upon their maturity and activation status.156 These cells are involved in directing immune responses within the testis and adjacent lymph nodes, and studies suggest that immature dendritic cells play a role in regulating tolerance in the normal testis.271., 272., 292.
Recruitment and Regulation of Testicular Macrophages
At birth, there are relatively few macrophages in the testis, but macrophage numbers expand dramatically during testicular development, coinciding with proliferation of the adult Leydig cell population and the appearance of the meiotic spermatocytes.228., 248., 249., 293. Indeed, the development of both the testicular macrophage and the adult Leydig cell populations appears to be interdependent. There is considerable evidence that macrophages are required for adult Leydig cell development and function.208., 259., 261., 262., 264. Conversely, studies from a number of groups have shown that the accumulation of macrophages during testicular development and maintenance of their numbers in the adult are dependent upon the action of pituitary LH and, more specifically, the Leydig cells themselves (Figure 19.9).69., 226., 293., 294., 295., 296., 297. Depletion of the Leydig cells or suppression of their activity in the adult testis by various means causes a progressive decline in the number of testicular macrophages.69., 226., 294., 295., 296., 297., 298. Although there is evidence that macrophages in the testis possess LH receptors,299., 300. the balance of the data suggest that the Leydig cell is responsible, rather than a direct action of LH itself. Testicular macrophage depletion occurs in models in which serum LH is increased (e.g. following treatment with the Leydig cell toxin, ethane dimethane sulfonate (EDS))69 or decreased (e.g. hypophysectomy, GnRH immunization, suppression of endogenous androgen with subcutaneous testosterone implants).294., 295., 296., 297., 298.
The precise mechanisms involved in maintaining the testicular macrophage population are not known. Complete depletion of spermatogenic cells by cryptorchidism has absolutely no effect on testicular macrophage numbers in the rat,226 suggesting that their regulation does not involve the seminiferous epithelium. Moreover, it does not appear that androgens are directly involved.226., 293., 295. However, it is likely that direct contact with the Leydig cell membrane and/or nonandrogenic products of the Leydig cells may be responsible.240., 244. Studies have clearly indicated that normal development of the testicular macrophage population, as in other tissues, involves the macrophage growth factor, colony stimulating factor-1 (CSF1).247., 264., 285., 301. The chemoattractant cytokine, MIF, which is constitutively expressed by the Leydig cells, likewise may be involved.226., 278., 302. Furthermore, intratesticular production of the chemokines, chemokine (C-X3-C motif) ligand 1 (CX3CL1; fractalkine) and CCL2, is implicated in the recruitment of circulating monocytes under normal conditions and during inflammation, respectively.285., 286., 303.
It remains an area of ongoing discussion whether resident macrophages found in most tissues under noninflammatory conditions are largely derived and sustained from the circulating monocytes or by proliferation within the tissue itself.235., 304. The significant population of macrophages expressing CD68, but not CD163, in the normal testis suggests that there may be constant recruitment of these cells from the monocyte pool.284 On the other hand, there is evidence that testicular resident macrophages may undergo active proliferation by mitosis, at least under certain circumstances: during the early inflammatory phase following destruction of the Leydig cells by EDS305 and during testis development in rats and mice.293., 306. Consequently, it is not absolutely clear whether the resident macrophage population of the testis, once established, is maintained by recruitment of new monocytes or by local proliferation. In fact, it is most likely that both processes are involved. Based on the extended persistence of radionuclides in rodent, canine, and human testicular macrophages, on the other hand, it would appear that most resident macrophages do not escape the testis alive.237., 238.
While initial evidence that testicular macrophages respond directly to FSH has been shown to be due to an experimental artifact,307 a stereological examination of macrophage recruitment to the testis in GnRH-immunized rats given recombinant FSH replacement demonstrated that FSH stimulates an increase in macrophage nuclear volume.294 This indicates an effect on macrophage activity in the testis, which is almost certainly mediated via the Sertoli cell, the only testicular cell type able to respond directly to FSH. It appears that, while Leydig cells are responsible for recruiting and maintaining the testicular macrophage population, the Sertoli cell may play a role in directing at least some of the testis-specific functions of these cells (Figure 19.9). These two somatic cells act together to recruit and modify the function of the testicular macrophages, thereby bringing about the unique resident macrophage phenotype found in this organ.
Lymphocytes in the Testis
It is a common misconception that lymphocytes are not found within the normal testis. In fact, lymphocytes are relatively prominent within and adjacent to the epithelium of the rete testis and are sparsely distributed throughout the testicular interstitial tissue in all species so far studied.65., 66., 67., 68., 69., 70., 231. In the rat and human testes, lymphocytes actually represent about 10–20% of the total leukocyte population, although the proportion of these cells in the normal mouse testis appears to be somewhat lower (Table 19.3) (Hedger MP, unpublished data).69., 227. It is worth noting that considerable differences may exist among different strains of animals and between animals raised under different degrees of exposure to environmental pathogens, as lymphocyte populations are very sensitive to differences in genetic background and past immunological events. Functional characterization of these cells has received relatively little attention: T cells and NK cells, but not B cells, have been described in the normal rat, mouse and human testis.65., 67., 68., 69., 70., 227. Most evidence suggests that the intratesticular lymphocyte population is skewed toward major histocompatibility complex (MHC) class I restricted (CD8+) cells and cells expressing NK cell markers (Hedger MP, unpublished data).65., 69., 70., 227.
Typically, T cells circulate through tissues as part of their surveillance function, and it is widely believed that activated T cells tend to recirculate in tissues where they initially encountered an antigen.308., 309. The phenotype of the majority of the T cells of the rat testis is certainly consistent with activated or memory T cells.70 There is clear evidence that lymphocyte numbers increase in the testes of men with infertility and sperm autoimmunity,66., 232. and lymphocyte numbers in the rat testis appear to gradually increase with age while macrophage numbers remain relatively constant (Hedger MP, unpublished data). These data suggest that at least some of the T cells in the testis are specific for testicular autoantigens. Alternatively, some of these cells may represent T cells that have been exposed to exogenous antigens within the testis environs during past infections.
The observation that there are substantial numbers of cells expressing NK markers in the testes of rats, mice, and humans is particularly significant (Hedger MP and Aridi DZ, unpublished data).70 Following isolation from the normal rat and mouse testis, it was confirmed that these cells have specific NK activity against transformed cells in vitro, and that a significant proportion of these cells are actually NK T cells (Hedger MP and Aridi DZ, unpublished data). In addition, CD4+CD25+FOXP3+ Treg cells are present in both rodent species, but are considerably more prominent in the mouse testis than in the rat. Immunohistochemistry has recently localized Treg cells to the human testis, as well.310 The T cells and NK T cells from the rat testis were found to produce IL10 under both unstimulated and PMA/ionomycin-stimulated conditions, although the T cells displayed a very high proportion of anergic and apoptotic cells compared with T cells from other tissues (Hedger MP and Aridi DZ, unpublished data). These observations indicate that the lymphocytes found in the normal testis comprise cells involved in cell-killing and innate immunity, that immunoregulatory subsets (NK, NK T and Treg cells) are highly represented, and that intratesticular T-cell responses to activation are significantly abrogated.
In normal testes, lymphocytes are rarely, if ever, observed within the epithelium or lumen of the seminiferous tubules, although lymphocytes appear to be able to cross the epithelium of the rete testis.64., 65., 66. Nonetheless, the number of lymphocytes and macrophages increases dramatically within the testicular interstitial tissue during testicular infection, experimental autoimmune orchitis, leukemic relapse, and testicular tumorigenesis.311., 312., 313., 314., 315., 316. This infiltration is generally associated with failure of spermatogenesis, and the immune cells may eventually breach the tight junctions of the blood–testis barrier and invade the seminiferous epithelium. Moreover, human infertility is often associated with increased numbers of intratesticular lymphocytes and macrophages, as evidence of ongoing inflammation even in the absence of an overt immune event.4., 5. It remains to be determined whether this is a cause or a response to testicular damage or failure, but there tends to be a relationship between the number of intratesticular lymphocytes and the extent of testicular disruption.
Mast Cells and Eosinophils in the Testis
Mast cells and eosinophils are specialized mediators and regulators of inflammation, particularly in the context of allergic responses.120., 121. Mast cells, in particular, are found in almost all tissues, including those of the male reproductive tract, but their distribution in the testis displays very distinctive, species-specific differences.62 In the testes of the rat, mouse, dog, cat, bull, and deer, these cells are largely absent from the testicular parenchyma but are frequently associated with blood vessels in the testicular capsule. In contrast, mast cells are found throughout the interstitial tissue in equine and human testes,62., 71., 317. while both mast cells and eosinophils are relatively abundant in porcine species.62 The functional significance of these cells in the testis is not known, but it is reasonable to assume that they play a role in local innate immunity and possibly a role in the fine control of testicular blood flow. In humans, testicular mast cell numbers decline with advancing age.317 However, increased numbers of intratesticular mast cells and increased mast cell expression of PTGS2 are associated with various forms of testicular failure, and they have been found to correlate with the severity of damage and increased NOS2 expression in the Leydig cells.22., 318., 319., 320. In contrast to mast cells and eosinophils, neutrophils are only found in the testis in conditions of testicular inflammation or damage.276., 314., 321., 322.
In the adult rat, mast cells are normally confined to the subcapsular region, but they proliferate dramatically throughout the testicular parenchyma following ablation of the Leydig cells by EDS.69., 323., 324., 325. The degree of proliferation appears to be under the control of the Leydig cells, suggesting that these cells produce an inhibitor of mast cell activity.69., 325. However, the increased mast cell numbers persist even after the Leydig cells recover and testis function returns to normal following EDS treatment. The species distribution of mast cells and eosinophils suggests some relationship with the level of aromatase activity of the Leydig cells,326., 327. and neonatal estrogen treatment increases mast cell numbers in the rat testis.325., 328. Stem cell factor (SCF), which is growth factor for both mast cells and Leydig cells that is produced by the Sertoli cell, also may be involved.329., 330. While eosinophils are not a feature of the normal rat or mouse testis, these cells were occasionally observed in the testes of GnRH-immunized rats.294 It appears that the regulatory mechanisms for the two different granulocyte subsets in the testis may be quite separate and distinct.
Immune Cells in the Epididymis
In contrast to the testis, macrophages and lymphocytes in the epididymis are frequently observed within the epithelium, where they are commonly identified as halo cells, as well as the interstitial tissue.64., 65., 66., 94., 95., 96., 97., 98., 99., 100., 331., 332., 333., 334. Published immune cell subset data for the epididymis have been complicated by the fact that some studies have not clearly differentiated between lymphocytes and macrophages because the CD4 and CD8 antigens are also expressed by rat and human monocytes and macrophages.335., 336. Studies in the mouse, where these antigens are more restricted to lymphocyte subsets, indicate that macrophages are the major epididymal leukocytes, located chiefly in the interstitial and peritubular regions, and that there appears to be a slight preponderance of MHC class II restricted CD4+ T cells (helper and regulatory T cell subsets) over the CD8+ T cell subset in the interstitial tissue, typical of blood and most other tissues.95., 96., 99. Conversely, the intraepithelial lymphocytes are predominantly CD8+ T cells, which is a common feature of mucosal epithelia.65., 96., 98., 334. This distribution of T cell subsets is consistent with the observation that the interstitial tissue macrophages express MHC class II antigens, whereas macrophages within the epididymal epithelium mostly do not.96., 99. As in the testis, development of the epididymal macrophages is dependent upon CSF1.337 Studies have suggested that the basal cells, located adjacent to the basal lamina of the epididymis, exhibit structural and antigenic properties typical of macrophages.94., 338. The numbers of the basal cells expressing macrophage-specific markers in the mouse are increased by the presence of damaged sperm, and it has been speculated that these cells are actually a type of resident macrophages, which could play a role in regulating immunity in the epididymis.338., 339.
Recently, dendritic cells have been identified, using specific reporter-labeled fluorescence imaging, as a major component of the epididymal epithelium in the mouse.102 These cells form a dense network in the basal region of the epithelium and extend their processes between the epithelial cells. They express characteristic dendritic cell and antigen-presenting surface markers, including ITGAX, CX3CR1, MHC class II antigens and CD80/86, and possess effective antigen-presenting activity in vitro. These cells are completely distinct in morphology and functional properties from the conventional intraepithelial macrophages or the basal cells.
Immune cells are found in all regions of the epididymis, although there is a tendency toward larger numbers and activity of all leukocyte subsets within the peritubular zone and epithelium of the caput, compared with the cauda.96., 97., 98., 99., 338. The number of intraepithelial macrophage and CD8+ T cells increases preferentially in the more proximal regions of the epithelium during aging and increased spermatogenic disturbance in rats, as well.100 Moreover, the intraepithelial dendritic cells appear to be particularly active within the proximal caput, such that their processes can extend all the way through the epithelium; they presumably can sample antigens within the epididymal lumen and present them to CD4+ T cells in the epididymal stroma and local lymph nodes.102 The lymphatics of the epididymis have been studied in detail in the rat and mouse; they show a distinct regional distribution favoring the cauda, but otherwise appear to be unremarkable.340., 341., 342.
The very different distribution of leukocytes in the epididymis compared with the testis indicates the existence of a different immunological environment, where separation between the antigenic sperm and the local immune cells is less robust, and more dynamic interactions are possible. Obviously, separation of antigens and immune cells is not the principal mechanism protecting sperm from immunological attack in this organ. In fact, it has been speculated that the macrophages within the epididymal duct may even be responsible for phagocytosis of senescent and excess sperm, although the evidence for this is equivocal.333., 343., 344., 345. The immunophysiological functions of the macrophages and lymphocytes within the epididymis are only just being uncovered, and they represent a fertile area for future research.
Immune Cells in the Vas Deferens, Accessory Glands, and Urethra
The organization of the vas deferens is very different from that of the epididymis: it is not coiled but has an extensively convoluted epithelium and multiple layers of smooth muscle. Sperm are not stored in the vas, and the majority of sperm pass through the duct only transiently at the time of ejaculation, but this structure plays a special role in our understanding of autoimmunity in the male tract because of the widespread use of vasectomy as a contraceptive method. Approximately 70% of all vasectomized men develop sperm antibodies; in many of these men, the antibodies are persistent, thereby frustrating later attempts to restore fertility.346., 347. Sperm antibodies are likewise associated with obstructive azoospermia and congenital absence of the vas (CAV) in men,348 and similar antibody responses occur in experimental animal models.332., 349., 350., 351., 352. These observations clearly indicate that obstruction or damage to the vas deferens is a potent inducer of sperm autoimmunity.
Although less prominent than in the epididymis, intraepithelial lymphocytes, principally CD8+ T cells, are found throughout the vas deferens, seminal vesicles, prostate, and urethra.64., 66., 353., 354., 355., 356., 357. Macrophages appear to be relatively rare in the epithelia, but they are found in the stroma of these tissues even under noninflammatory conditions, where they are dependent upon CSF1 for their development.337., 353. Relatively little is known about the immunological properties of these cells, but a predominance of cytotoxic CD8+ T cells is a common characteristic of all mucosal epithelia. While an immunoregulatory role of these lymphocytes in the reproductive epithelium remains to be determined, these cells are almost certainly involved in mediating cellular immunity at these sites.
It is self-evident that the leukocytes found in semen, even in men with no obvious infection or other inflammatory condition, must originate from the epithelium of the ducts and organs of the male reproductive tract.358 Logically, immune cells pass with relative ease across the epithelium in certain areas, thereby coming into physical contact with the sperm at the time of ejaculation. It is nonetheless curious that, while macrophages and lymphocytes are typically present, the leukocytes most commonly found in the semen are neutrophils, which are not a major leukocyte subset within either the stroma or epithelia of the male reproductive tract.358., 359., 360., 361., 362., 363., 364. Because vasectomy dramatically reduces leukocyte numbers in the ejaculate, the epididymis and proximal vas deferens appear to be a major source of these cells, at least under normal conditions, but the accessory glands (seminal vesicles and prostate) are also implicated as a source of the seminal neutrophils.364., 365., 366.
Immunological and Inflammatory Mediators in the Testis
Inflammation and activation of the immune response has immediate and mostly negative effects on male reproduction and fertility.20., 21., 367. It is widely assumed that this reproductive disruption is related to fever and the effects of raised body temperature on spermatogenesis in febrile patients, but there is actually very little experimental evidence to support this assumption. In fact, there is an enormous body of research to indicate that other inflammatory mechanisms can affect male reproductive function and are much more likely to disrupt reproduction.
More than 25 years ago, it was reported that rat testicular extracts contained a protein with properties similar to IL1, which eventually turned out to be IL1, produced by Sertoli cells rather than by intratesticular leukocytes.368., 369. Although generic molecules with inflammation-regulating activity, such as the prostaglandins, were already known to be produced by the reproductive tract, this discovery provided the first indication that an inflammatory molecule belonging to the immune system could be produced by a somatic cell of the male reproductive system. In subsequent years, it became increasingly obvious that many inflammatory and immunoregulatory cytokines could be produced by somatic and spermatogenic cells of the testis in response to an inflammatory stimulus, including IL1, IL6, TNF, and TGFβ family members and interferons. Moreover, once it became apparent that their production was constitutive, in the absence of any evidence of cryptic inflammation, or could be stimulated by physiological stimuli, such as hormones or cell–cell interactions, a role in normal reproductive physiology was anticipated. It is now very evident that inflammatory and immunoregulatory molecules produced by the testis are involved in regulating both immunological processes within the testis and many aspects of normal testicular function associated with fertility. The production and actions of the most prominent of these molecules are briefly outlined in this section of the review. Integration of their actions and the consequences for testicular immune responses and fertility are covered in later sections.
The Interleukin-1 Family and the TLRs
These molecules are related by the fact that the IL1 receptor and the TLRs belong to a family of dimerizing receptors that contain the conserved cytoplasmic TIR domain.106., 112. Both IL1 and TLR ligands act through similar signaling pathways to activate intracellular MAP kinases and the pro-inflammatory transcription factor, NFκB, and induce the expression of early pro-inflammatory genes, including IL1 (Figure 19.5).
Interleukin-1
The archetypical pro-inflammatory cytokine, IL1 is represented by two related proteins (IL1α and IL1β) capable of binding to the same receptor to exert an almost identical range of effects.370 These proteins are produced by many cells types, but activated monocytes and macrophage are the major source of secreted IL1.370 Both IL1α and IL1β are single chain proteins, with only about 25% homology at the mature protein level. They are synthesized as 31–33 kDa precursor proteins that are cleaved enzymatically to active 17-kDa forms (Figure 19.10 ). Both the precursor and mature forms of IL1α are biologically active, but the IL1β precursor is inactive. Consequently, production of active IL1β is a two-step process involving NFκB-induced transcription and translation of the IL1β precursor gene, followed by cleavage of the precursor to the mature cytokine by CASP1.117., 118. This processing of IL1β is linked to its secretion into the extracellular space. Conversion of IL1α does not involve CASP1, but the precursor can be cleaved by calcium-dependent membrane-associated cysteine proteases, or calpains, and extracellular proteases.371 In contrast to IL1β, the majority of IL1α tends to remain within the cell or associated with the cell membrane, although it may be found in secretions as well.370 It is generally believed to act as an autocrine growth factor or as a mediator of direct cell-to-cell communication, but the mechanisms involved are poorly defined. Both IL1α and IL1β differ from most cytokines in that they lack a signal sequence and their mechanisms of secretion remains to be elucidated.
FIGURE 19.10.
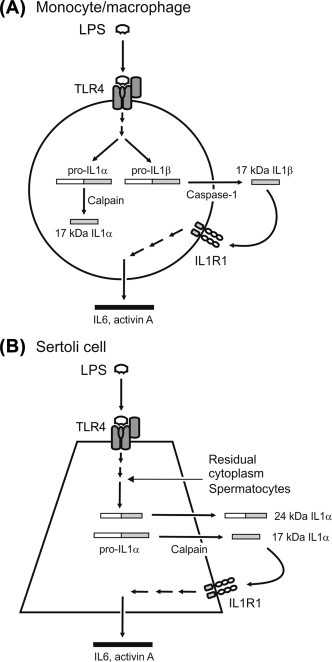
Comparison of the regulation and production of interleukin-1 (IL1), IL6 and activin A in the monocyte/macrophage and Sertoli cell.
(A) Activation of the TLR4 complex on the surface of the macrophage by bacterial lipopolysaccharide (LPS) upregulates expression of both IL1α and IL1β, which subsequently are processed to their mature bioactive 17-kDa forms by the action of calpain and caspase-1, respectively. IL1α tends to remain associated with the cell, but IL1β is secreted upon cleavage and binds to the IL1 receptor (IL1R1) on the cell surface to stimulate production of other inflammatory cytokines, such as IL6 and activin A. (B) The Sertoli responds to stimulation by LPS, spermatocytes and the residual cytoplasm of released spermatozoa by producing IL1α, preferentially. Two alternate transcripts of IL1α are produced by the Sertoli cell, including a transcript lacking the calpain cleavage site domain, which encodes a 24-kDa form of IL1α with reduced bioactivity. Both forms appear to be secreted by the adult Sertoli cell and both are able to signal via the IL1 receptor to stimulate the production of IL6 and activin A.
Intratesticular Production of Interleukin-1
Intratesticular IL1α and IL1β have been observed in many species, including the human,372 but have been most extensively studied in the rat. Endogenous production of IL1α appears in the rat testis at around 20 days of age, more or less coinciding with progression of the first wave of spermatogenesis.369., 373., 374., 375., 376. This appears to be almost entirely due to the Sertoli cells, which synthesize and secrete both the mature 17-kDa IL1α molecule and a slightly larger 24-kDa testis-specific form of IL1α—the latter being the product of an altered mRNA transcript lacking the calpain cleavage site necessary for processing of the precursor (Figure 19.10).377., 378., 379. Adult rat seminiferous tubule extracts also contain an immunoreactive 45-kDa IL1α protein that has yet to be fully characterized, but may be another variant of the precursor mRNA with one or more introns retained. These testis-specific transcripts are biologically active, although less potent than mature 17-kDa IL1α.377., 380. There is some evidence that rat spermatocytes and spermatids also express IL1α constitutively.381
Production of IL1α by the Sertoli is under control of the spermatogenic cells and shows a distinct cyclical variation that is related to the changes in spermatogenic cell associations as determined by the cycle of the seminiferous epithelium.373., 382., 383., 384. It is strongly stimulated by phagocytosis of the residual cytoplasm cast off by the spermatids when they are released into the tubule lumen.375., 376., 385. Sertoli cell production and secretion of IL1α in vitro is induced by various inflammatory stimuli, including LPS and IL1 itself, but not by FSH.376., 385., 386., 387., 388. Studies indicate that Sertoli cells from prepubertal rats produce, but do not secrete, IL1α, whereas Sertoli cells from adult rats are able to secrete biologically active IL1α.369., 379., 389. The mechanism underlying this maturational change in ability of the Sertoli cell to secrete IL1α is unknown.
In contrast to IL1α, IL1β appears not to be produced in significant amounts in the rat testis under normal conditions, but is upregulated in a subset of testicular macrophages and the Leydig cells during inflammation.274., 369., 373. Curiously, the majority of this secreted testicular IL1β is present as the precursor protein.274 Studies have shown that Leydig cells express the mRNA for both IL1α and IL1β in vitro in response to stimulation by LPS or exogenous IL1,390., 391., 392. and whole testes and isolated macrophages collected from LPS-treated rats or mice express IL1β mRNA,393., 394., 395. and the immunoreactive protein.396 Thus, while IL1β expression in the normal testis is relatively low, it increases during inflammation, possibly due to production by interstitial cells rather than by the Sertoli cells. The response of testicular IL1α to inflammation in vivo is less clear: although production by the Sertoli cell is upregulated by inflammatory stimuli in vitro, recent in vivo studies report either no response or reduction during systemic inflammation.396., 397.
Intratesticular Actions of Interleukin-1
Numerous studies have established that IL1, whether produced intratesticularly or external to the testis, is an effective regulator of Leydig cell steroidogenesis, spermatogonial and spermatocyte development, and various Sertoli cell functions associated with supporting spermatogenesis.
The actions of IL1 are mediated by the IL1 type I receptor (IL1R1), which belongs to the immunoglobulin superfamily of receptors, but also bears an intracellular TIR domain (Figure 19.5).106., 112. Following ligand binding, a second TIR domain-containing subunit, called the IL1R acceptor protein (IL1RAP), is recruited to form a complex that binds the MYD88 adaptor protein and activates IRAK, leading to NFκB nuclear translocation and MAP kinase activation. Moreover, IL1 is able to activate other signaling pathways through induction of phosphatidylinositol-4,5-bisphosphate 3-kinase (PIK3)/Akt protein kinase (AKT) and tyrosine-protein kinase Src (SRC), hydrolysis of GTP, phosphatidylcholine, phosphatidylserine or phosphatidylethanolamine, release of ceramide by neutral sphingomyelinase, and release of arachidonic acid by phospholipase A2 (PLA2).370., 398. There is an IL1 type II receptor (IL1R2), which lacks a TIR domain and can act as a decoy or inhibitory receptor.399
In cultures of Leydig cells from immature rats (10–20 days of age), IL1β caused a dose-dependent increase in DNA synthesis, and IL1α was much less potent than IL1β.400 The effect of IL1β was not observed in Leydig cells isolated from older animals, suggesting that macrophage-derived IL1β could play a role in Leydig cell proliferation during prepubertal development.400 The possible involvement of prostaglandins in this developmental regulation is indicated by the observations that IL1β stimulates the expression of PTGS2, both IL1 isoforms and IL6 in rat progenitor Leydig cells, and that prostaglandins act as intermediates in this stimulatory pathway.401
A review of the literature dealing with the direct effects of IL1 on testosterone production by mature Leydig cells in vitro reveals reports of stimulation, inhibition or even of no effect.395., 402., 403., 404., 405., 406. Presumably, these apparent discrepancies arise from differences in the experimental conditions or methodologies employed. On balance, studies suggest that IL1 is either stimulatory or has no effect on basal testosterone production by adult Leydig cells,406., 407., 408. and there is general agreement that IL1 inhibits LH/hCG and/or cAMP stimulated testosterone production.395., 402., 403., 406., 407., 409. In the mouse, the major site of inhibition is at the level of CYP17A expression (Figure 19.3).403., 410. In the rat, IL1 appears to regulate Leydig cell steroidogenesis at the level of CYP11A, while STAR gene expression and protein synthesis are unaffected.404., 411. The effects of testicular IL1α on Leydig cell steroidogenesis may be dependent on both the variant of IL1α involved and on the stage of development of the Leydig cell. The 32-kDa IL1α precursor and the 17-kDa mature IL1α are both inhibitory to gonadotropin-stimulated or cAMP-stimulated steroidogenesis in mature Leydig cells, whereas the testis-specific 24-kDa IL1α splice variant has no effect.380 In contrast, all three forms of IL1α stimulated testosterone production by immature rat Leydig cells. When IL1β was injected into the immature rat testis, Leydig cell steroidogenesis and serum testosterone levels were stimulated within 24 h, although Leydig cell activity had declined several days later.412 No similar effect was observed in the adult testis. The signaling mechanisms mediating IL1 effects on the Leydig cells involve PTGS2 enzyme expression and PGE2 acting as an intermediate.402., 413. Studies using Leydig cells from immature rats have also implicated the MAP kinases, MAPK14 and MAPK3/MAPK1, in the regulation of STAR and cholesterol mobilization.380., 414.
The IL1 receptor (IL1R1), along with the decoy receptor (IL1R2), has been localized to most testicular cell types in rat, mouse and/or human testis, including the spermatogenic cells and Sertoli cells.415 It has been shown that IL1α is able to stimulate mitotic DNA synthesis in intermediate and B type spermatogonia and meiotic DNA synthesis in preleptotene spermatocytes.382., 383., 384., 416. In vitro, IL1 stimulates proliferation of prepubertal Sertoli cells417 and regulates a number of activities of more mature Sertoli cell that support or control spermatogenesis, including aromatase activity,418 production of glucose transporters, lactate and transferrin,419., 420., 421. and production of IL1α, IL6 and the IL1 receptor.375., 385. IL1 also regulates production by the Sertoli cell of the inhibin B heterodimer and the homologous β-subunit homodimer, activin A, in an inverse relationship with FSH.389., 422. Considered more broadly, IL1 opposes the mostly stimulatory effects of FSH on Sertoli cell activity. Moreover, IL1 regulates the ability of the Sertoli cell to maintain contact with other Sertoli cells and developing spermatogenic cells, by altering the Sertoli cell cytoskeleton, and stimulates opening of the occluding junctions that make up the blood–testis barrier.423 Regulation of Sertoli cell function by exogenous IL1 has been found to involve multiple pathway intermediates, including MAPK14, MAPK8 and MAPK3/MAPK1, PIK3/AKT, PTGS2/PGE2 and NOS2/NO, although the more predictable participation of MYD88 and NFκB still awaits investigation.417., 424., 425., 426., 427.
Interleukin-18 and Other Interleukin-1 Family Cytokines
The IL1 family comprises several structurally related proteins, which appear to have arisen by gene duplication.428 The most closely-related to IL1α and IL1β is IL18, which is normally processed to its mature form by the inflammasome and the action of CASP1, like IL1β, but acts via a distinct IL18 receptor.429 Both IL18 and its receptor have been found in the seminiferous epithelium of the rat, with IL18 mRNA and protein localized to spermatocytes and round spermatids.430 Murine Leydig cells produce IL18, CASP1, and the IL18 receptor in response to stimulation by LPS,431 and IL18 has been detected by immunohistochemistry in human testis, where its expression is reduced in impaired spermatogenesis.432 Recombinant IL18 stimulates spermatogonial DNA synthesis in cultures of rat seminiferous tubules, without influencing germ cell apoptosis.430 As is the case for IL1β, the majority of IL18 produced within the rat testis is in the precursor form,430 indicating that the processing of these cytokines by the inflammasome/CASP1 in the testis is an area that merits more investigation.
The IL1 receptor antagonist (IL1RN) binds to the IL1 receptors but lacks the ability to transduce a signal, and so acts as a competitive antagonist of IL1 action.433 It has been shown to be produced by mouse Sertoli cells, and its production is stimulated by FSH, LPS and IL1.434 Male mice lacking IL1RN have higher intratesticular levels of IL1α and IL1β, along with reduced fertility and litter sizes and increased morphological abnormalities and premature activation of their sperm, although sperm number and motility is not affected.435
The TLRs
Studies have revealed that TLRs are widely expressed in the male reproductive tract (Table 19.4 ).24., 101. The other pattern-recognition receptor families have yet to receive much attention in this context, but several NLR and RLR family members have already been identified in the testis.436., 445., 446.
TABLE 19.4.
Toll-like Receptor Expression in the Epithelium of the Male Reproductive Tracta
| Receptor | Principal Ligands | Principal Pathogens | Cellular Location | Sertoli Cells | Epididymis | Vas Deferens |
|---|---|---|---|---|---|---|
| TLR1 | Triacyl lipopeptides | Bacteria, mycobacteria | Cell surface | +++ | ++ | +++ |
| TLR2 | Lipoproteins, peptidoglycans | Bacteria, mycobacteria, viruses | Cell surface | ++++ | ++ | + |
| TLR3 | dsRNA | Viruses | Endosomes | ++++ | +++ | ++ |
| TLR4 | Lipopolysaccharides | Bacteria, viruses | Cell surface | ++++ | ++ | + |
| TLR5 | Flagellin | Bacteria | Cell surface | +++ | +++ | +++ |
| TLR6 | Diacyl lipopeptides, zymosan | Bacteria, fungi | Cell surface | +++ | +++ | + |
| TLR7 | ssRNA | Viruses | Endosomes | +/− | + | + |
| TLR8b | ssRNA | Viruses | Endosomes | − | +/− | − |
| TLR9 | CpG DNA | Bacteria, viruses, protists | Endosomes | − | ++ | ++ |
| TLR10c | Unknown | Bacteria | Cell surface | + | + | − |
| TLR11d | Profilin | Bacteria | Endosomes | + | +++ | +++ |
| TLR12d | Profilin | Bacteria | Endosomes | − | ND | ND |
| TLR13d | Ribosomal RNA | Bacteria | Endosomes | + | ND | ND |
Studies in the rat and mouse have established that Sertoli cells express TLR1-6 at significant levels, with relatively low expression of TLR7 and 13 in the mouse and of TLR 10 and 11 in the rat.388., 436., 437., 438., 439., 440. Quantitative examination of the TLR4 complex in rat Sertoli cells confirmed that the level of expression of TLR4 and its co-receptor, MD2, was similar in the Sertoli cell and testicular macrophage, but that Sertoli cells displayed low basal or LPS-induced expression of the TLR4 accessory protein, CD14.388 Cytokine production responses to TLR1-6 ligands have been confirmed in both rat and mouse cells. The Sertoli cells produce appropriate cytokines, such as IL1α, IL6, and CCL2, in response to TLR4 activation by LPS,379., 388., 436., 437. and type 1 interferons in response to TLR3 or TLR4 activation438., 439., 440.; however, the precise roles of specific signaling pathways in these responses requires further clarification, particularly with respect to the importance of the adaptor protein MYD88 and the transcription factor NFκB.436., 437., 438., 439., 440.
The majority of studies on the effects of TLR ligands on Sertoli cells have employed LPS, which induces inflammatory gene responses in the Sertoli cells that are similar to those observed in macrophages.24 However, LPS obtained from different bacterial strains can have quite different chemical composition and is frequently contaminated by other TLR ligands (e.g. bacterial lipoproteins and peptidoglycans).447 This means that many studies in the literature using LPS actually describe responses involving multiple TLRs (usually TLR2 and TLR4). When highly purified LPS was used, rat Sertoli cells were more than 10-fold less sensitive to LPS than testicular macrophages, but they expressed similar levels of IL1α and IL6 and much greater levels of activin A when maximally stimulated.388 These Sertoli cells also responded to the synthetic lipopeptide Pam3Cys (a specific TLR2 ligand) with a more prolonged pattern of gene expression. The need for relatively high doses of LPS to stimulate the Sertoli cell is probably related to the relatively low level of expression of the accessory protein, CD14, which serves to amplify the response to LPS in macrophages.110 These data indicate that Sertoli cells respond to bacterial ligands acting through both TLR2 and TLR4, although they are less sensitive to these ligands in comparison with local macrophages and display a Sertoli cell-specific pattern of gene expression in response.
There have been few studies on the effects of TLR ligands on noninflammatory responses in the Sertoli cell: exposure of Sertoli cells to LPS in vitro directly inhibited lactate production and plasminogen activator activity, which are important functions for supporting spermatogenic cell development.448 In other studies, LPS induced oxidative stress in Sertoli cells by increasing ROS production and reducing antioxidant activity,449 while activation of TLR3, a receptor for viral double-stranded RNA, stimulated scavenger receptor expression and phagocytosis of apoptotic spermatogenic cells by Sertoli cells in culture.439
In rat and/or mouse studies, mRNA for TLR2, 3, 4, 7, 9, 10, and 12, along with low levels of MD2 and CD14, have been observed in Leydig cells; TLR2, 3, 4, 6, and 11 have been observed in peritubular cells; and TLR2, 3, and 4 have been observed in spermatogenic cells (Hedger MP and Winnall WR, unpublished data).388., 436., 450., 451. These cells also respond to TLR ligands in vitro. Leydig cells produce pro-inflammatory cytokines and type 1 interferons, but they reduce their sterodogenic activity when stimulated with LPS or a TLR3 agonist.390., 392., 450. Peritubular cells respond to LPS by producing the macrophage chemoattractant, CCL2.303 Most remarkably, activation of TLR3 has been reported to induce the inflammatory transcription factors, NFκB and IRF3, and a range of genes involved in inflammation and antiviral responses in mouse spermatogonia and spermatocytes.451
In the immune system, the TLRs and IL1 are fundamental regulators of NFκB activity. The regulation and functional role of this pro-inflammatory transcription factor in normal spermatogenesis is not fully understood at this time, but existing data suggest that there is an interaction between endogenously-expressed inflammatory molecules and NFκB in controlling seminiferous epithelial function. NFκB is constitutively present and active in the nucleus of cultured rat Sertoli cells and displays a cyclical localization within the nuclei of the Sertoli cells and spermatogenic cells throughout the cycle of the seminiferous epithelium.452 Induced expression of NFκB stimulates androgen receptor expression, and therefore androgen responsiveness, in rat Sertoli cells,453 but activation of NFκB in Sertoli and/or spermatogenic cells has also been implicated as an apoptosis-inducing signal in damage models, involving exposure to toxins, ischemia-reperfusion injury, and cryptorchidism.454., 455., 456., 457.
The TNF Family
Tumor Necrosis Factor
Members of the TNF family of cytokines are commonly membrane-bound homotrimers that can be released as soluble ligands by proteolytic cleavage. The canonical member of this family, TNF is a 17-kDa glycosylated polypeptide that is secreted principally by activated monocytes and macrophages, and binds to receptors that are present on most cells in the body.458 This cytokine plays a central role in the initiation of the inflammatory response, and can exert pro-inflammatory or cytotoxic effects depending upon the receptor subtype engaged and the expression of specific adaptor proteins within the target cells (Figure 19.5). There are two families of TNF receptors (TNFR).459., 460. The TNFR type 1 family of receptors induce cell death through a motif in their cytoplasmic regions called the death domain (DD), and includes TNFR1, FAS and the death receptors (DR)3-6. When TNF binds to TNFR1, there is recruitment to the DD of the intracellular adaptor proteins TRADD (TNFR-associated death domain) and FADD (Fas-associated death domain), leading to activation of the cell death caspase pathway. However, the binding of TRADD can also lead to recruitment of the cellular inhibitor of apoptosis (baculoviral IAP repeat containing; BIRC) protein or receptor interacting serine–threonine kinase (RIPK), enabling the binding of TRAF2. This complex mediates activation of the NFκB pathway and the MAP kinases, MAPK14 and MAPK8. The TNFR type 2 receptors, which include TNFR2, TNFR superfamily member 8 (TNFRSF8, CD30) and 5 (TNFRSF5, CD40), and the lymphotoxin receptor, do not contain a DD in their intracellular domains, and instead associate with TRAFs leading to the activation of cell signaling events.
Intratesticular Production and Actions of TNF
In common with IL1, TNF regulates Leydig cell steroidogenesis and the spermatogenesis-supporting functions of the Sertoli cell, but TNF appears to act as a regulator of spermatogenic cell survival, rather than of proliferation or development. In situ hybridization studies in mice first identified the presence of TNF mRNA in round spermatids, pachytene spermatocytes, and testicular interstitial macrophages.461 Bioactive TNF was produced by the round spermatids in vitro and mRNA for the corresponding receptor was located on Sertoli and Leydig cells. Similarly, in the seminiferous epithelium of the rat, TNF has been localized to the spermatogenic cell compartment, and TNF receptors primarily localized to the Sertoli cells.462 Treatment of isolated rat or mouse testicular macrophages with LPS also induces TNF secretion.463., 464.
The majority of studies performed using a variety of systems, including intact animal studies,465., 466. isolated primary cultures of Leydig cells,410., 467., 468., 469. and MA-10 tumor Leydig cells transfected with CYP17A-reporter constructs,470 report inhibitory effects of TNF on Leydig cell steroidogenesis. Inhibition of LH/hCG binding, STAR expression, and cholesterol mobilization has been observed,466., 471. but these studies also indicate that inhibition by TNF occurs downstream of cAMP production at the level of steroidogenic gene expression (Figure 19.3). In cultured mouse Leydig cells, TNF had no effect on basal expression of CYP11A, but it suppressed basal expression of HSD3β and the expression of CYP11A, CYP17A, and HSD3β induced by analogs of cAMP.410., 468. The inhibitory effect of TNF on CYP17A gene expression was found to be mediated through protein kinase C, possibly by activation of MAPK8, MAPK3/MAPK1, and the transcription factors, NFκB and AP-1.467., 470., 472. Several other studies have implicated the sphingomyelin/ceramide-dependent pathway in TNF-mediated inhibition of Leydig cell testosterone secretion.466., 469. Although the levels of testosterone and the mRNA levels of steroidogenesis-related genes were significantly lower after puberty in TNF knockout mouse testes than in wild-type testes, this may be attributed to increased levels of anti-Müllerian hormone in these animals.473
Within the seminiferous epithelium, TNF appears to play a complex role in the regulation of Sertoli cell and peritubular cell function, which is probably attributable to the ability of TNF to stimulate both cell-killing and cell signaling pathways. Many of the effects of TNF have been shown to involve the activation of NFκB and MAP kinases in the Sertoli cell.453., 474., 475., 476., 477., 478. Similar to IL1, TNF stimulates basal lactate production by cultured Sertoli cells, but generally antagonizes the actions of FSH on Sertoli cell function, including the stimulation of lactate production, aromatase activity and inhibin.479., 480., 481. Nevertheless, TNF receptor subunit protein expression has been shown to be stimulated by FSH in porcine Sertoli cells.482 TNF stimulates the expression of inflammatory cytokines, chemokines, and leukocyte adhesion molecules expression in both Sertoli cells and peritubular cells.303., 387., 477., 478. Furthermore, TNF stimulates plasminogen activator inhibitor expression in rat testicular peritubular cells, indicating that it plays a key role in controlling testicular protease activity.483 In Sertoli cell monolayer cultures, TNF disrupts inter-Sertoli cell tight-junction assembly by inhibiting production of the junction protein, occludin, and by regulating matrix metalloprotease and protease inhibitor activity.484 Delfino and colleagues have shown that TNF stimulates androgen receptor expression in Sertoli cells via upregulation of NFκB, which binds to several enhancer motifs in the androgen receptor promoter.453
Single nucleotide polymorphisms in the TNF gene cluster producing elevated TNF levels have been shown to be associated with reduced sperm count and sperm motility in infertile men,485 but the outcomes for spermatogenesis of TNF action are not always so clear-cut. Paradoxically, TNF reduces spontaneous spermatogenic cell degeneration in cultured human and rat seminiferous tubules, apparently by regulating FASL and/or NFκB levels in the Sertoli cell, indicative of a germ cell survival role for TNF mediated through the Sertoli cell.475., 476. At first glance, this would appear to run counter to the observation that activation of NFκB in the Sertoli cell or germ cells induces spermatogenic cell apoptosis in various damage models,454., 455., 456., 457. and the studies that have identified a role for germ cell-secreted TNF in the disruption of spermatogenesis in response to cytotoxic injury of the Sertoli cell.486 However, NFκB is also able to regulate and interact with pro-survival genes, such as the inhibitor of apoptosis (IAP) and BCL2 families.459., 487. The interactions between TNF, NFκB and different apoptotic regulators produces complicated outcomes for spermatogenic cell survival in different situations, and the responsible mechanisms invite further investigation.
FAS and FAS Ligand
The death receptor FAS and its ligand (FASL) most commonly act as membrane-bound trimeric ligand–receptor pairs mediating cell–cell interactions, and FAS–FASL binding on activated T cells is essential for moderating the immune response.180 Typically, the DD in the cytoplasmic region of FAS recruits the FADD adaptor protein and induces T cell death via caspase-dependent apoptosis.488 In the testis, FAS and FASL have been implicated in regulating spermatogenic cell apoptosis during testicular damage and, more controversially, in maintaining immune privilege.489., 490. Studies intended to localize FASL in the testis under normal conditions have produced conflicting results, which may be attributed to differences in detection methods, limitations of the reagents employed, and the fact that these molecules are readily inducible.491., 492. Thus, FASL has been described as being present in rat, mouse, porcine, and human Sertoli cells, and absent in most germ cells,489., 490., 493., 494. but others have reported that FASL expression in the rat seminiferous epithelium is confined to the germ cells.492., 495. FAS has been identified on isolated mouse Sertoli cells,496 but in intact testes it has been localized to spermatogonia and spermatocytes from the pubertal period onwards.490., 495., 497.
Nonetheless, FAS expression is clearly associated with spermatogenic cells that are undergoing apoptosis.475., 486., 490., 497. Furthermore, FAS can be induced in the Sertoli cell by the action of TNF and IFNγ.474., 496. Expression of both FAS and FASL is upregulated in various models of seminiferous epithelium damage, indicating that this mechanism is important in regulating germ cell apoptosis in cases of physical and toxicological insult.490., 498. Induction of FAS and FASL is also implicated in the testicular response to inflammation.499
Interleukin-6
In contrast to either IL1 or TNF, IL6 is a cytokine with both pro-inflammatory and anti-inflammatory actions (Figure 19.7); for example, IL6 stimulates B cell growth and differentiation and promotes neutrophil activity and survival, but inhibits the effects of IL1 and TNF, and induces IL1RN and IL10 expression.125., 500. In the late inflammatory phase, it stimulates the acute-phase response and promotes fibrosis. Outside the immune system, IL6 plays crucial roles in hematopoiesis, liver and neuronal function, energy metabolism, and cancer. Cytokines of the IL6 type, which include leukemia inhibitory factor (LIF) and IL11, exert their action via binding to specific receptors that can form complexes with a common, ubiquitously-expressed membrane signal transducer, glycoprotein 130 (interleukin-6 signal transducer; IL6ST).501 In lymphocytes and hepatocytes, IL6 binds to its specific membrane-bound receptor (IL6R; CD126), which has no intrinsic kinase activity and dimerization with IL6ST leads to activation of the Janus kinase (JAK)/signal transducer and activator of transcription (STAT) and MAP kinase signaling pathways (classic IL6 signaling). However, all cells that express IL6ST are able to respond to IL6 bound to soluble IL6R, which may be produced from an alternatively-spliced mRNA transcript or released by proteolytic cleavage of membrane-bound IL6R, a process called trans-signaling.
In vitro, IL6 is constitutively produced by Sertoli cells and Leydig cells.375., 386., 390., 502., 503., 504. Production of IL6 in rat or mouse Sertoli cells is stimulated by a range of agents, including FSH, testosterone, IL1, TNF, and LPS, and is inhibited by INFG.387., 496. Phagocytosis of residual cytoplasm shed by late spermatids at the time of spermiation triggers Sertoli cell IL1α production, which stimulates IL6 secretion in an autocrine manner, by activating leukotriene production via the lipoxygenase pathway (Figure 19.10).375., 387. Leydig cells are strong producers of IL6 after stimulation by LH, LPS, or IL1 in vitro, and may be the major source of this cytokine in the testis.390., 503., 504. Human peritubular cells express IL6 mRNA, but possibly not the protein, when stimulated by TNF,478 and isolated spermatogenic cells from adult mice express IL6 mRNA and protein when stimulated with a TLR3 ligand, poly(I:C).451
Although not as intensively studied in the testis as either IL1 or TNF, it appears that IL6 regulates the function of somatic, spermatogenic, and immune cells. Both IL6R and IL6ST mRNA are expressed in rat Sertoli cells and are stimulated by IL1 and by IL6, but only the IL6R subunit is stimulated by FSH.505 In the Sertoli cell, IL6 increases basal and FSH-induced transferrin and cyclic GMP secretion,506., 507. but reduces κ opioid receptor mRNA levels.508 IL6 has been shown to act as an inhibitor of meiotic DNA synthesis in preleptotene spermatocytes.509 An effect of IL6 on testicular steroidogenesis at the level of both the Leydig cell and the hypothalamic-pituitary levels is indicated by the observation that subcutaneous IL6 administration in men produced prolonged suppression of serum testosterone levels, without apparent changes in gonadotropins.510
The Transforming Growth Factor-β Family
Cytokines of the TGFβ family, similar to IL6, possess both pro-inflammatory/pro-fibrotic and anti-inflammatory/immunosuppressive properties (Figure 19.7), but also regulate growth and differentiation at a fundamental level throughout the entire body. They are mostly homo- or heterodimers that act by binding two transmembrane receptor kinases, denoted as type I and type II, and assembling them into an active heterotetramer.511., 512. This triggers a transphosphorylation cascade that begins with the activation of the type I subunit kinase by the type II subunit kinase, and is propagated by the canonical SMAD (small for body size/mothers against decapentaplegic homolog) protein signaling pathway, which in the case of TGFβ and activin involves SMADs 2, 3 and 4, and to non-SMAD-mediated signaling pathways.511., 513. These receptors are expressed in most cells and tissues.
Transforming Growth Factor-β
The three forms of TGFβ (TGFβ1, TGFβ2, and TGFβ3) found in mammals are disulfide-linked dimers of approximately 25 kDa in mass, derived from three structurally-similar subunits possessing intrastrand disulfide bonds that form a cysteine knot folding motif (Figure 19.11 ).514 Heterodimers of the subunits can also occur but are less abundant. All three forms are differentially expressed by Sertoli cells, peritubular cells, and Leydig cells, particularly in the fetal and immature testis, although production appears to decline considerably during the establishment of complete spermatogenesis.515., 516. In the postpubertal testis, they also have been localized to the spermatogenic cells in a development-specific pattern of expression.225., 517., 518.
FIGURE 19.11.
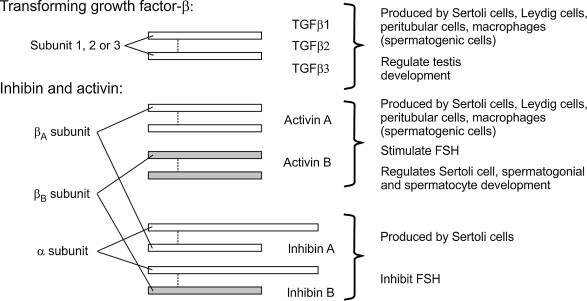
The transforming growth factor-β (TGFβ) and activin/inhibin family of cytokines.
These cytokines are dimers of disulfide-linked subunits with considerable sequence and structural homology, and have diverse actions within the reproductive and immune systems. Three distinct subunits lead to production of homodimers (or less frequently heterodimers) of TGFβ1, 2, or 3. Dimerization of two homologous β-subunits lead to either activin A, activin B, or less commonly activin AB (not shown). Dimerization of the activin subunits with the closely related α-subunit produces the inhibins, which are produced almost exclusively by the Sertoli cells. Activins derived from the βC-subunit (not shown), appear to play regulatory roles through formation of nonactivating homodimers and heterodimers.
The receptors for TGFβ are found in both somatic and spermatogenic cells.225., 517., 519. In the fetal and immature testis, TGFβ has been implicated in controlling the development and apoptosis of gonocytes and spermatogonia, seminiferous tubule formation and Leydig cell differentiation, and Sertoli cell-peritubular cell interactions.516., 520., 521., 522. In the adult testis, TGFβ stimulates apoptosis and inhibits meiosis during spermatogenesis,523., 524., 525. and inhibits Leydig cell steroidogenesis at the level of LH receptor number and signaling, as well as distal to cAMP production at the level of CYP17A expression.526., 527. Crucially, TGFβ2 and TGFβ3 regulate Sertoli cell tight junction dynamics, via the MAPK14 pathway, indicating a role for TGFβ-mediated signaling pathways in regulating the permeability of the blood–testis barrier.528 Finally, TGFβ is a potent regulator of inflammation and the activity of immune cells, particularly of Th17 and Treg cells,529., 530. suggesting a role in controlling intratesticular immune responses.
Activin
The activins display a high degree of sequence and structural homology with TGFβ, operate through a similar receptor system, and signal via common pathways; nonetheless, they have their own characteristic range of biological actions.531 There are four forms of activin subunits in mammals (βA, βB, βC and βE), but only the first three appear to be particularly relevant to the male reproductive tract.224 Activins A and B are dimers of the β-subunits of inhibin A and B, which are themselves heterodimers of one of two β-subunits (βA and βB) with a common α-subunit (Figure 19.11).532 Consequently, activin A (βAβA), activin B (βBβB), or activin AB (βAβB) forms exist, although most studies to date have concentrated on the activin A homodimer. Activin A is commonly believed to be the most biologically active form.533 Whereas the chief physiological role of inhibin is to inhibit FSH at the anterior pituitary level, activin A and B stimulate FSH production.534 Although both inhibin and the activins circulate in the blood, production of inhibin is almost exclusively confined to the Sertoli cell,535., 536. while the activins are produced in many different cell types and tissues, including the anterior pituitary, where they act as local regulators of FSH .535., 537., 538., 539.
Activins bind to one of two specific type II activin receptors (ACVR2A or ACVR2B), which are able to dimerize with a type I activin receptor serine/threonine kinase (activin receptor-like kinase, ALK).512., 540. As is the case for TGFβ, postreceptor signaling occurs via SMAD2/3/4, and several alternative signaling pathways, including inflammatory pathways involving MAPK14, MAPK8, and MAPK3/MAPK1.512., 513., 541. Inhibin acts as a competitive inhibitor of activin because it can bind to the type II activin receptors but cannot bring about receptor subunit multimerization: this interaction of inhibin with the activin receptor is facilitated by a specific co-receptor protein called TGFβ receptor type III (TGFBR3), or betaglycan.542 Activin C is a dimer of the structurally-related βC-subunit, which does not appear to dimerize with the inhibin α-subunit and is unable to facilitate activin receptor signaling.543., 544. It is most highly expressed in the liver but is present in the testis.545 Homodimers and heterodimers comprising the βB- and βC-subunits appear to act as weak competitive agonists or antagonists of activin A.533., 544., 546., 547. Moreover, activin bioactivity can be effectively neutralized in the circulation and in tissues by an endogenous, high-affinity activin binding protein, follistatin.548
In the hematopoietic and immune systems, activin A is produced by activated monocytes, macrophages and dendritic cells, Th2 cells, bone marrow stromal cells, mast cells and neutrophils, and is stimulated by IL1, TLR ligands, and TNF, acting through MYD88/TRAF signaling, MAPK14 and MAPK8.531 The βA-subunit gene promoter comprises one or more AP-1 binding sites, but there do not appear to be any typical consensus NFκB sites within the proximal promoter.549., 550. Expression is stimulated in a synergistic manner by the Th2 cell transcription factor c-MAF and NFAT, a transcription factor that is expressed in immune cells.551 Transcription of the βA-subunit in response to cAMP probably involves the cAMP-responsive transcription factor, AP-2.552 The presence of multiple phorbol ester-responsive elements (AP-1 and AP-2) in the promoter, and the stimulation of activin A production by phorbol esters in several cell types, also implicate protein kinase C in its regulation.550., 552. Multiple AP-1 and AP-2 sites have been identified in the promoter of the βB-subunit, suggesting that activin B may be, at least in part, regulated in a similar manner to that of activin A.552., 553. In the murine liver, expression of the βB-subunit is stimulated by systemic administration of LPS, implicating TLR signaling in its regulation.554
A large body of evidence suggests that activin A acts to promote inflammation and activation of macrophages and dendritic cells during the initial phase of the immune response, but suppresses immune responses once inflammation is established and activin A concentrations increase.531 In cells of the monocyte/macrophage lineage that had not been activated, activin A induced nuclear translocation of NFκB, and production of pro-inflammatory mediators, such as IL1β, TNF, IL6, NO, and prostanoids.555., 556. However, in activated monocyte/macrophages, activin A inhibited processing of the IL1β precursor and production of TLR4, CD14, and pro-inflammatory mediators and increased production of IL1RN.557., 558. Similarly, activin A stimulated the recruitment and development of monocyte-derived dendritic cells,559., 560. but inhibited the ability of dendritic cells to mature and stimulate T cell activation.561., 562. Inhibitory effects of activin A on the antigen-presenting activity of macrophages have been observed as well,563 and activin A stimulates the development of antigen-specific Treg cells.564 Furthermore, activin A inhibits many of the actions regulated by IL1, IL6, and IFNγ, including T-cell and B-cell proliferation, monocyte phagocytosis, and production of acute-phase proteins.557., 565., 566., 567. Together with the fact that activin A is produced by Th2 cells and has been implicated in promoting various Th2-regulated immune responses, such as asthma and atopy,568 mast cell recruitment, maturation, and activity,569 and B cell immunoglobulin production,555., 570. these observations suggest that activin A may be a crucial Th2 cytokine. Likewise, the effects of activin A on macrophage inflammatory responses indicate an ability to switch pro-inflammatory M1 macrophages to an alternatively-activated, anti-inflammatory M2 phenotype. Finally, activin A has been shown to be both a fundamental regulator and an important intermediary in the process leading from inflammation to fibrosis, which is also associated with the M2 macrophage phenotype.279., 571.
In the adult testis, βA-subunit mRNA or activin A protein is found in the Sertoli cells, Leydig cells, peritubular cells, resident macrophages, mast cells, and most spermatogenic cells, except spermatogonia.224., 572., 573. In the adult rat, intratesticular fluid levels of activin A are 5–10 times higher than normal circulating concentrations.274 In cultured Sertoli cells, activin A is stimulated by IL1 and TLR ligands (Figure 19.10), and inhibited by FSH and the cAMP/protein kinase A signaling pathway.379., 388., 389., 422., 481., 573. Immediately following spermiation, there is a spike in activin A production by the Sertoli cells, possibly driven by the surge of ILα produced by the Sertoli cell at this time.572., 573. Testicular macrophages, Leydig cells, and Sertoli cells respond to LPS by increasing βA-subunit expression in vitro, although the Sertoli cells display a much more robust response and higher basal levels of production.224., 388. However, the actual role of activin A in testicular inflammation remains to be determined because the high intratesticular levels of activin A in the adult rat were not acutely affected by LPS treatment in vivo.274 Significant production of activin B in the testis has yet to be confirmed: the βB-subunit is produced by the Sertoli cell, but it is not clear whether any of this goes to form activin B homodimers, as inhibin B heterodimers are by far the major form of inhibin in the testes of most species, including the human.422., 574. The activin βC-subunit has been immunolocalized to Sertoli cells, peritubular cells and Leydig cells, as well as spermatogonia, primary spermatocytes, and elongating spermatids.545., 575.
Activin receptors are expressed by most, if not all, of the somatic and spermatogenic cells of the testis.572., 576., 577. Activin, together with TGFβ, exerts complex regulation of spermatogenic cell, Sertoli cell and Leydig cell development and function, and disorders of activin/TGFβ signaling are implicated in the onset of testicular cancer.578 Depending upon the culture system employed, activin A exerts both stimulatory and inhibitory effects on spermatogonial cell proliferation,579., 580., 581. and is able to maintain the condensed mitochondrial morphology found in spermatogenic cells beyond the leptotene stage of the first meiotic prophase.582 Activin A regulates mature Sertoli cell function by disrupting tight junction formation and blood–testis-barrier function, and stimulating their proliferative activity.583 Activin A has been shown to regulate steroidogenesis by immature and adult Leydig cells in culture and by the MA-10 Leydig cell line,584., 585., 586. although these effects were only evident after extended culture under non-physiological conditions. In mice, deletion of the βC-subunit had no overt effect on testis function,587 but overexpression of the βC-subunit resulted in progressive hypospermatogenesis, which was proposed to be due to antagonism of activin A (and possibly activin B) in the testis.547 Like TGFβ, activin is expected to be involved in controlling inflammation and immunity in the testis, by regulation of the maturation and activity of the intratesticular macrophages and lymphocytes.
The Interferons
The interferons are functionally-related cytokines with antiviral, antiproliferative, and immunomodulatory actions, comprising three main groups (α, β, and γ), based on their structural relationships and major cellular sources.115., 588. The type I interferons (IFNα and β) are produced by a broad range of cell types: typically, multiple forms of IFNα are produced by monocytes and macrophages, while two forms of IFNβ are produced by fibroblasts and epithelial cells, but they all exert their antiviral and antiproliferative effects via the IFNα receptor (IFNAR). The single type II interferon, IFNγ, is produced by NK and NK T cells, activated T cells and, under certain conditions, by macrophages and dendritic cells.589., 590. It acts through the IFNγ receptor (IFNGR), and regulates the activity of antigen-presenting cells as part of the adaptive immune response, in addition to its antiviral functions. In other words, IFNγ is a type II interferon, but a type 1 (Th1) cytokine. Production of type I interferons is normally stimulated through TLR3, which detects double-stranded viral RNA, and through TLR4, the receptor for bacterial LPS.115 These TLRs upregulate the interferon-inducing transcription factor, IRF3, by engagement of the TICAM1 adaptor protein and interaction with TRAF3 (Figure 19.5).106., 115. Induction of IFNγ involves IL12 and IL18, type 1 interferons, JAK/STAT signaling, and the transcription factors, NFκB and AP-1, in a cell-specific manner.589., 590.
Viral infections stimulate type 1 interferon and IFNγ production by Sertoli cells, peritubular cells, Leydig cells, and testicular macrophages, leading to production of canonical antiviral proteins in the testis.438., 591., 592., 593. In vitro, type 1 interferon expression was induced by activation of TLR3 or TLR4 in murine Sertoli cells,438., 439. and by activation of TLR3 in murine spermatogenic cells.451 Production of IFNγ in the testis has been implicated in both the regulation of immune privilege and onset of autoimmune orchitis.594., 595. Moreover, IFNγ stimulates Sertoli cell production of FAS and CASP1, and is implicated as the mediator of Sertoli cell and germ cell apoptosis under various conditions.496., 596. In a transgenic mouse model, overexpression of IFNβ in the seminiferous tubules caused spermatogenic cell loss and sterility.597 These disparate observations are indicative of a complex relationship between the interferons, the local immune system, and spermatogenesis.
In a study on normal, healthy men, treatment with human IFNα significantly decreased serum testosterone levels.598 This was most likely due to inhibition of steroidogenesis at the Leydig cell and hypothalamic-pituitary levels, because serum gonadotropin levels were unaffected. Both IFNα and IFNγ can inhibit testosterone production in primary cultures of porcine Leydig cells, and studies indicated that IFNγ inhibits cholesterol transport into the mitochondria, and inhibits expression of the STAR protein, CYP11A, and CYP17A (Figure 19.3).599., 600. In other experimental studies, human and murine IFNα inhibited steroidogenesis in rat Leydig cells.601 These data indicate that interferons may contribute to the overall decline in steroidogenic function that is commonly observed in viral infections.21., 602., 603. However, Dejucq and colleagues592 have shown that Sertoli and Leydig cells in the rat testis strongly expressed IFNα and IFNγ during infection with Sendai virus, and that elevation in expression was associated with an increase in testosterone production by Leydig cells infected with Sendai virus in vitro. This result highlights the fact that responses to some infections may exert a stimulatory effect on steroidogenesis within the testis, even though factors that are inhibitory to steroidogenesis are induced.604
The Eicosanoids
The eicosanoids are formed by oxidation of the 20-carbon fatty acid, arachidonic acid, and encompass the prostaglandins (PGD, PGE, and PGF), prostacyclins (PGI), thromboxanes (Tx), lipoxins, and leukotrienes (LT) (Figure 19.12 ). These lipids are signaling molecules, fundamentally involved in physiological processes, including inflammation and immunity. They arise from hydrolysis of arachidonic acid from membrane glycerophospholipids, through the action of phospholipase A2 (PLA2).605 The rate-limiting step in the conversion of arachidonic acid to the prostanoids is cyclooxygenation, catalyzed by one of two closely related enzymes, PTGS1 and PTGS2 (Table 19.2). While PTGS1 is constitutively expressed, PTGS2 expression is inducible during inflammation, through the IL1/TLR and TNF signaling pathways.606 Production of the specific prostanoid classes, however, requires the subsequent actions of specific prostaglandin synthases and thromboxane synthase.607 In turn, the prostaglandins and thromboxanes interact with their respective membrane-bound G protein-coupled receptors to regulate cell growth, vascular smooth muscle tone, vascular permeability, and immune cell activity.608 Prostanoids exert both pro-inflammatory and immunosuppressive actions through these receptor interactions, or through metabolism to other active immunoregulatory metabolites, such as the immunosuppressive ligand for peroxisome proliferator-activated receptor-γ (PPARγ), 15deoxy-PGJ2.609., 610. An alternative synthetic pathway, catalyzed by the lipoxygenases, converts arachidonic acid to the closely-related leukotrienes and lipoxins, which likewise possess diverse metabolic, vascular, and inflammation-regulating activities.178., 611.
FIGURE 19.12.
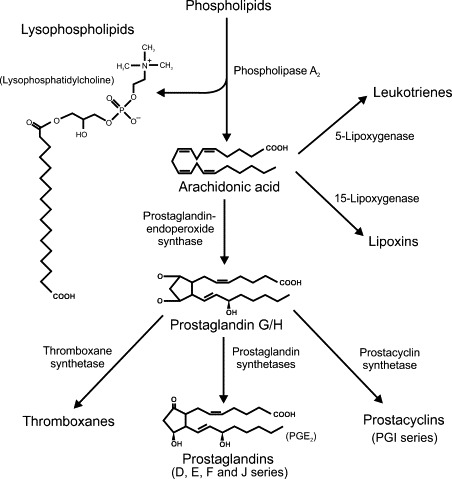
Synthesis of bioactive lipids.
Cleavage of membrane-bound phospholipids by the action of phospholipase A2 leads to production of arachidonic acid and lysophospholipids, such as the lysophosphatidylcholines. Arachidonic acid may be converted to prostaglandins, prostacyclins or thromboxanes via the intermediates prostaglandin G and H by the action of the rate-limiting prostaglandin-endoperoxide synthase (PTGS) enzymes. Alternatively, lipoxygenases may catalyze the conversion of arachidonic acid to leukotrienes or lipoxins. Most of the products in these complex pathways have specific regulatory activities, including profound effects on immune responses and the vasculature.
The enzymes responsible for prostanoid biosynthesis are expressed throughout the male reproductive tract.612., 613., 614., 615. Most testicular cells express both forms of PTGS and possess the capacity to produce prostaglandins in vitro,615 although there are significant cell type-specific and species-specific differences in the relative levels of expression.616., 617., 618. In the normal rat testis, PTGS2 is responsible for the majority of prostaglandin production, and chronic treatment with a specific PTGS2 inhibitor, celecoxib, considerably reduces intratesticular PGE2 levels.615., 619. On the other hand, intratesticular PGE2 levels are barely affected by acute LPS treatment.615 This may be due to the fact that macrophages in the rat testis express PTGS2 at very low levels, relative to other testicular cells, but they are the only testis cell type to respond to an inflammatory stimulus with a significant increase in PTGS2 activity.615 These data indicate a maintenance role for the so-called inducible PTGS2 enzyme in testicular function.
Prostaglandins, particularly PGE2 and PGF2α, are clearly implicated in the control of Leydig cell development in the immature testis, production of pro-inflammatory cytokines by the Leydig cell, autoregulation of steroidogenesis in the adult, and the decline in Leydig cell function that occurs during aging.401., 620., 621. However, the actual importance of prostanoids in regulating testicular function remains uncertain: male mice lacking either PTGS2 or PTGS1 are considered to be fertile,622 and studies on the effects of prostaglandins or various PTGS inhibitors on spermatogenesis and fertility have produced conflicting outcomes.618., 619., 623., 624. Products of the lipoxygenase pathway, most notably LTB4, have been implicated in the regulation of Leydig cell steroidogenesis by LH.625., 626. Furthermore, production of PGE2 and PGF2α induced by PTGS2 mediates the effects of IL1 on protein and lipid regulation by the Sertoli cell involved in supporting spermatogenesis and inflammation in the testis.424., 425. A relationship between expression levels of IL1β and PTGS2 in the testes of infertile men also has been reported.627 These observations suggest that PTGS2 and prostanoid production mediate at least some of the effects of inflammation on testicular function.
Oxidative Enzymes, Nitric Oxide Synthases, and Reactive Oxygen Species
Reactive oxygen species are normal products of metabolism. They are produced continuously as byproducts of mitochondrial and microsomal electron transport reactions and other metabolic processes.628 Cytochrome P450 enzymes, such as those that constitute the steroid synthetic pathways, produce ROS as a by-product of their catalytic mechanism.629., 630. However, rapid production of ROS as part of a respiratory burst by macrophages and neutrophils is also an important component of the early response to infection.121., 631.
The most important ROS are the superoxide anion (O2 −), hydrogen peroxide (H2O2), hydroxyl radical (HO•), nitric oxide (NO•), and peroxynitrite anion (ONOO−)(Figure 19.13 ). During inflammation, their rapid production is stimulated by the activity of the enzymes NADPH oxidase and NOS2.631., 632. These small molecules are cytotoxic to microorganisms, although some play a more complex role in cell signaling processes, as well.633 They also interact with vital intracellular macromolecules, such as proteins, lipids and DNA, to cause oxidative damage.633., 634. Cellular antioxidant systems that normally protect cells from oxidative damage include mitochondrial superoxide dismutase, catalase and glutathione peroxidases.635 Excess production of ROS during oxidative stress, such as occurs during ischemia-reperfusion injury or excessive inflammation, can overwhelm these protective mechanisms, resulting in cell damage, and apoptosis or necrosis of affected cells, thereby contributing to the pathology of many diseases.636
FIGURE 19.13.
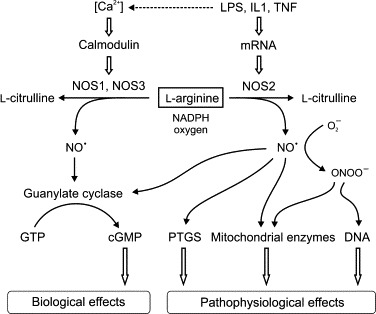
Regulation and actions of nitric oxide synthase (NOS) and NO production in normal physiology and pathophysiology.
NO is produced by conversion of l-arginine to l-citrulline by NOS, and the freely diffusible NO radical (NO•) exerts a variety of physiological functions through regulation of cGMP-dependent protein kinases. The activity of the constitutively expressed isoforms, endothelial NOS (NOS3), and neuronal NOS (NOS1) are regulated through calcium-calmodulin. Synthesis of a constitutively active form of NOS (NOS2) is induced by inflammatory mediators and is responsible for the large upregulation of NO production during inflammation. High levels of NO• can be detrimental due to direct reaction with the heme group of prostaglandin-endoperoxide synthase (PTGS), mitochondrial enzymes and DNA, effects which can be potentiated by interaction with superoxide to produce the extremely reactive peroxynitrite anion. Peroxynitrite is the reaction product of NO and superoxide, and is one of the major cytotoxic agents produced during sepsis, inflammation and ischemia/reperfusion. Peroxynitrite decomposes to produce the hydroxyl radicals, which are potent oxidants.
At high levels, NO is extremely cytotoxic, especially as a precursor of the peroxynitrite anion, but at low levels NO acts as an intracellular and extracellular signaling molecule and has potent vasodilatory effects.637 It can be produced by one of three related enzymes, neuronal NOS (NOS1), NOS2, and endothelial NOS (NOS3), which catalyze the conversion of l-arginine to l-citrulline and NO.638 These enzymes are homodimeric proteins encoded by three separate genes. Both NOS1 and NOS3 are constitutively expressed enzymes whose activity is regulated through a calcium-calmodulin mediated mechanism, whereas NOS2 is a constitutively activated enzyme that is regulated at the transcriptional and translational levels by inflammatory mediators, including LPS, IL1, and TNF.637
Studies have reported the presence of NOS in Sertoli, Leydig, and peritubular cells, spermatogenic cells, testicular macrophages, and vascular endothelial cells of various species.639., 640., 641., 642. The NOS3 gene also produces a testis-specific isoform, TnNOS, which has been localized to Leydig cells.643 In the normal rat seminiferous epithelium, NOS2 is expressed by elongating spermatids and pachytene spermatocytes, particularly during the stages immediately following sperm release, with relatively lower levels of expression in Sertoli cells and peritubular cells throughout the entire cycle.641 Moreover, NOS2 is upregulated by LPS treatment in Leydig, Sertoli, peritubular, and spermatogenic cells, as well as a subset of the testicular macrophages.276., 641., 644., 645. This macrophage expression of NOS2 appears to be preferentially expressed by the infiltrating CD163- macrophages of the testis and is not detectable in the majority of resident macrophages.276., 641. Testicular cell NOS2 expression and NO production are also increased by inflammatory events induced by testicular ischemia-reperfusion injury or heat.646., 647., 648., 649.
Nitric oxide is an important regulator of spermatogenesis. Localized production of NO, by spermatogenic cells in particular, is implicated in control of the formation and disassembly of the Sertoli cell junctions that constitute the blood–testis barrier, as well as the junctional complexes involved in Sertoli-germ cell adhesion.640 In transgenic mice lacking NOS2, pachytene and round spermatid apoptosis is significantly reduced, leading to an increase in daily sperm output and indicating a key role for NOS2 in limiting germ cell survival and/or the carrying capacity of the Sertoli cells.647 Sertoli cell and Leydig cell numbers are also increased in these mice, indicating that developmental effects of NOS2 and NO are involved.650 On the other hand, spermatogenic damage caused by testicular torsion or cryptorchidism can be ameliorated by reducing NOS or NO levels.649., 651. Increased NOS2 is related to severity of testicular failure and mast cell numbers in infertile men.320
Treatment of murine Leydig cell primary cultures or MA-10 Leydig tumor cells with hydrogen peroxide causes oxidative damage, resulting in a marked reduction of the mitochondrial electrochemical gradient (ΔΨm) and decreased STAR and HSD3β protein levels.652., 653., 654. Elevation of endogenous ROS, particularly the superoxide anion and hydrogen peroxide, inhibits the activity of cytochrome P450 enzymes in the steroidogenic pathway, and plays a critical role in Leydig cell desensitization to hyperstimulation by LH or exogenous hCG,629., 630. which is used to treat delayed testicular descent in young boys.655 In a similar manner, NO inhibits Leydig cell steroidogenesis directly, and treatment with NOS inhibitors counteracts the decrease in testosterone associated with inflammation or stress.656., 657., 658., 659. While this may involve oxidative damage via generation of reactive nitrogen species such as the peroxynitrite anion, it has been shown that NO inhibits steroid biosynthesis by binding reversibly to the heme group of cytochrome P450 enzymes.656., 660. Increased testosterone in NOS2-deficient mice may account for the developmental effects observed in these mice.650 Moreover, NO is a potent vasodilator, and plays a role in the endogenous control of testicular blood flow and formation of the interstitial fluid, which also can impact upon Leydig cell function and spermatogenesis.641., 658.
Inflammatory Signaling Pathways and Testicular Function
It is now clear that inflammatory and immunoregulatory factors and pathways are constitutively active in the normal (noninflamed) testis, and that these factors and pathways have consequences for spermatogenesis and steroidogenesis. Clinically, there is a growing recognition that testicular failure, irrespective of etiology, correlates with increased inflammatory gene expression and immune cell infiltrates in infertile men.4., 5., 22., 23., 229., 318., 320., 372., 432., 627. What is the actual physiological relevance of these experimental and clinical observations, and what might this mean for understanding the effects of infection, inflammation and immune responses on male fertility? The physiological implications may be broadly separated into outcomes for effects on testicular steroidogenesis, which could lead to androgen deficiency problems, and more direct effects on spermatogenesis and sperm output.
Steroidogenesis
In terms of normal Leydig cell steroidogenesis, it may be anticipated that local production of cytokines, and small molecule inflammatory mediators, such as NO and prostanoids, will influence intratesticular androgen levels. These molecules may play a role in facilitating communication between cells of the seminiferous epithelium—the Sertoli cells, peritubular cells and spermatogenic cells—and the Leydig cell. Such paracrine actions have long been associated with local control of steroidogenesis, as a mechanism for modulating the response of the Leydig cell to stimulation by LH, in response to changing requirements of the seminiferous epithelium.61., 661. Moreover, since Leydig cells themselves produce these inflammatory regulators, they could be involved in autocrine regulation of steroidogenesis as well. Certainly, production of NO and other ROS, either by the Leydig cell itself or through its close physical relationship with the resident macrophages, is implicated in Leydig cell desensitization to stimulation by LH.394., 409., 629., 630. In terms of pathology, increased levels of inflammatory regulators, arising from either the circulation or the testis, during infection and inflammation, should similarly impact upon steroidogenesis.
Spermatogenesis and the Cycle of the Seminiferous Epithelium
Studies in the rat and mouse indicate that certain cytokines and inflammatory mediators are expressed in a dynamic manner throughout the spermatogenic process, in the complete absence of external drivers of inflammation. Specifically, there is a surge of production of ILα, IL6 and activin A by the Sertoli cell at the time of the release of mature spermatids from the seminiferous epithelium (spermiation), which is reflected in a concomitant increase in nuclear translocation of NFκB and production of TNF and NOS2 in the meiotic and postmeiotic spermatogenic cells.373., 375., 384., 452., 461., 509., 572., 573., 641. This is a time when a number of critical physiological events occur within the seminiferous epithelium, including the phagocytosis of spermatid residual cytoplasm by the Sertoli cells, an increase in DNA synthesis associated with spermatogonial mitosis and the entry of preleptotene spermatocytes into meiosis, and the transition of these early spermatocytes through the tight junctions that normally constitute the blood–testis barrier.18 The coincidence of these events is particularly significant because it has been established that ILα, IL6, and activin A are regulators of spermatogonial proliferation and differentiation,382., 383., 384., 416., 509., 579., 580., 581. while IL1α, activin A, TNF, and NO have all been shown to stimulate the opening of the blood–testis barrier by modifying the Sertoli cell cytoskeleton, inhibiting production of proteins involved in Sertoli–Sertoli and Sertoli-spermatogenic cell junctions and/or regulating protease and protease inhibitor activity.423., 484., 583., 640.
These observations suggest the existence of a fundamental mechanism, whereby the spermatogenic cells themselves drive inflammatory pathways within the seminiferous epithelium (Figure 19.14 ).24., 224. In this model, phagocytosis of the residual cytoplasm at the time of spermiation stimulates inflammatory signaling and cytokine production in the Sertoli cells,375., 376., 385., 387., 573. which subsequently induce NFκB nuclear translocation and inflammatory gene expression in the spermatogenic cells.451., 452., 641. These induced inflammatory mediators, in turn, regulate the proliferation and differentiation of the spermatogonia and spermatocytes, and regulate the integrity of the tight junctions to allow the germ cells to pass through the blood–testis barrier. At the same time, these cytokines regulate the activity of the Sertoli cell and modulate the response of the cell to hormonal stimulation. This represents a feasible mechanism whereby the most recent generation of mature spermatozoa is able to communicate with and coordinate the activity of subsequent generations of spermatogenic cells.
FIGURE 19.14.
Hypothetical roles of inflammatory signaling pathways in control of spermatogenesis.
Spermatogonia enter meiosis to become spermatocytes and pass through the tight junctions between adjacent Sertoli cells. At the end of meiosis, the resulting haploid spermatids undergo major structural differentiation and are released from the seminiferous epithelium as spermatozoa (spermiation), leaving behind most their (residual) cytoplasm, which is phagocytosed by the Sertoli cells. Spermiation coincides with a surge of production of inflammatory mediators, such as interleukin-1α (IL1α), IL6, and activin A by the Sertoli cells, and tumor necrosis factor (TNF) and NO by the spermatogenic cells, which regulate the proliferation and maturation of the nearby spermatogonia and spermatocytes, and reorganization of the tight junctions to allow spermatocytes to pass into the luminal compartment. This localized inflammatory response may be mediated through activation of pattern-recognition receptors (PRR) in the Sertoli cell by endogenous spermatogenic molecules. Degenerating spermatogenic cells may also drive these pathways at other stages of the spermatogenic cycle, and inflammation caused by infection will disrupt these processes. Inflammatory mediators also regulate the supportive functions of the Sertoli cells, such as the production of transferrin, lactate, stem cell factor (SCF), plasminogen activator (PLA), and inhibin B, which are stimulated by androgens and by FSH acting via cAMP. Although these separate signaling pathways control some functional outcomes in common, there is clear evidence for reciprocal inhibitory regulation between the signaling pathways as well.
In immunology, this type of inflammatory pathway activation is called sterile or non-pathogen-mediated inflammation, and similar mechanisms have been proposed for controlling activities in the epithelia of the intestine and lung, and in the vascular endothelium.662., 663., 664. The actual trigger for this response in the Sertoli cell may involve the physical process of phagocytosis of the residual cytoplasm and activation of pattern-recognition receptors (PRR in Figure 19.14), such as the TLRs and the inflammasome.108., 117. These receptors are capable of detecting and responding to various endogenous intracellular ligands, including the nuclear protein, HMGB1, heat shock proteins, CpG-rich DNA and RNA, all of which are found in spermatogenic cells.7., 108., 291., 665.
Furthermore, some endogenously-produced inflammatory mediators, most notably IL1α, remain elevated within the Sertoli cell long after the spermiation event, which comprises only a brief period within the process of spermatogenesis.373., 375., 384., 452., 461., 502., 509., 641. This may be attributable to ongoing contact between the spermatogenic cells and Sertoli cell, mediated by pattern-recognition receptors on the cell surface, and/or by phagocytosis of degenerating cells, since many spermatogenic cells do not survive to complete spermatogenesis and are lost through apoptosis.490., 666. In support of this possibility, Zhang and colleagues have recently reported that damaged spermatogenic cells induce inflammatory gene expression, including IL1β, TNF, and IL6, in mouse Sertoli cells through activation of TLR2 and 4.667 Such interactions may be driving other cyclical functions of the Sertoli cell, including the responsiveness of the Sertoli cell to FSH and androgen,668., 669. and the expression of regulators of cytokine activity, such as follistatin.572., 670.
The Significance of Inflammatory Signaling Pathways for Male Fertility
Studies on inflammatory processes in normal testicular function, or male reproductive function in general, should be tempered by two considerations. Firstly, because the mediators of inflammation and their signaling pathways are exquisitely sensitive to stimulation by microbial contamination, stress, or even the very act of cell isolation, it should not be assumed that expression levels are always indicative of a role in normal physiology. Many early studies of these processes may have been compromised by failure to eliminate endotoxin (endogenous LPS) contamination from the experiments, or the extreme sensitivity and nonquantitative nature of the detection methods used, such as RT-PCR. It is only when molecules have been repeatedly shown to be constitutively and substantially expressed in the absence of exogenous inflammatory stimuli, and to have consistent and coherent effects on reproductive function in diverse studies, that we may assume that these molecules play a role over and above the usual response to infection or other immune activation events.
The second consideration lies in the fact that mice with transgenic deletions of many of these inflammatory and immunoregulatory factors, their synthetic enzymes or receptors, appear to be fertile.671., 672., 673., 674. However, this assessment may fail to take into account the fact that the reproductive phenotype is subtle, or even enhances fertility, as occurs in the NOS2-deficient mouse.647., 650. Sometimes, more careful examination of the reproductive tract and fertility reveals reproductive defects in male mice that are, nonetheless, capable of producing offspring.264., 435. Moreover, given the extraordinary complexity, diversity and redundancy of the immune system, effects may only manifest after compound deletions of multiple functions, or on certain genetic backgrounds. Some effects may not be evident under normal animal husbandry conditions, and may only become evident in older animals or in animals living in the wild exposed to environmental stresses, dietary deficiency or specific pathogens. Some effects may only become evident when other physiological processes are also compromised, such as occurs in male mice infected with Taenia crassiceps.672 Wild-type mice with this infection display increased IL6 expression and become feminized due to excessive aromatase activity. This steroidogenic deviation is completely prevented in IL6 deficient mice, which otherwise have similar reproductive functions to those of wild-type mice. This observation is consistent with studies implicating IL6 as a regulator of aromatase activity,675 but also provides an example of a reproductive knockout phenotype that only manifests itself following specific immunological events.
Effects of Immune Cells and Inflammation on Male Reproduction
The Role of Macrophages in Leydig Cell Development and Steroidogenesis
Macrophages play an important role in Leydig cell development. During puberty, there is a close temporal link between the maturation of the adult Leydig cell population and the increasing number of testicular resident macrophages.228., 249., 293. Moreover, specific depletion of the intratesticular macrophages inhibits the development of Leydig cells in immature rats, as well as the recovery of Leydig cells after EDS-treatment in adult rats.259., 261., 262. The op/op mouse, which has an inactivating mutation in the CSF1 gene and consequently has reduced numbers of macrophages, also displays very poor fertility and reduced testosterone production due to developmental and steroidogenic abnormalities of the Leydig cells.264., 301. The influence of the testicular macrophages on Leydig cell growth and differentiation may involve specific macrophage-derived cytokines, such as IL1,400 or the intercytoplasmic specializations between the two cell types, which appear very early in adult testicular development in the rat.244
Studies on the effects of resident macrophages on Leydig cell steroidogenesis under non-inflammatory conditions have produced inconsistent results. Stimulation of macrophage phagocytosis with an intratesticular injection of latex beads increased the steroidogenic capacity of Leydig cells in adult rats.676 However, in macrophage-depleted adult rat testes, both a decline263., 273. and an increase260., 265. in testosterone production have been reported by different groups using essentially the same model. Moreover, medium collected from supposedly unstimulated testicular macrophages in culture have been shown to both stimulate677., 678. and inhibit405., 408., 678. Leydig cell steroidogenesis in vitro. Similar discrepancies were observed in Leydig-macrophage co-cultures. The most likely explanation for these differences is that partial activation of the macrophages during isolation, or possibly endotoxin contamination, may have affected the results. Neuroendocrine influences also may be important; for example, studies of the bank vole revealed that testosterone production by Leydig cells from long photoperiod animals was more sensitive to stimulation by testicular macrophage conditioned medium and more sensitive to IL1α-mediated inhibition in comparison with short photoperiod animals.679 On balance, however, the published data seem to indicate that resting testicular macrophages have a positive or trophic effect on Leydig cell steroidogenesis.
Testicular macrophages have been shown to express the cholesterol 25-hydroxylase enzyme and produce 25-hydroxycholesterol.677 It has been proposed that testicular macrophages, due to their close association with the Leydig cells, provide 25-hydroxycholesterol as a substrate for testosterone biosynthesis, by-passing the need for STAR and supporting basal steroidogenesis. Interestingly, the 25-hydroxylase enzyme is negatively regulated by testosterone suggesting that there may be a feedback loop between the macrophages and Leydig cells.680., 681. Furthermore, 25-hydroxycholesterol has been suggested to be one of the factors which macrophages secrete in support of Leydig cell proliferation and development.682
Effects of Inflammation on the Leydig Cell and Steroidogenesis
Reduced testosterone production is an important mechanism underlying the inhibition of male fertility associated with inflammatory disease, regardless of whether or not infection is involved. This can involve inhibitory effects at multiple levels of the hypothalamic-pituitary-testicular axis. Men with critical illness, burn trauma, sepsis, and rheumatoid arthritis have elevated serum TNF and IL1 levels and reduced testosterone.367., 683., 684., 685. Similar decreases in gonadal function have been reproduced in experimental animal models of chronic inflammation and systemic immune activation. For example, experimental adjuvant-induced arthritis causes a dramatic reduction in serum testosterone levels in rats,686., 687. and conditioned medium from testicular macrophages isolated from adjuvant-induced arthritic rats is inhibitory to Leydig cell testosterone production in vitro.687
The literature contains numerous experimental models that have been used to study the effects of inflammation on testicular function, although interpretation of the results can often be problematic. In general, studies employing bacterial or viral infections are complicated by the difficulties in distinguishing the effects of inflammation from the actions of the pathogen itself, or the subsequent immune response to the pathogen. Furthermore, although inflammation is characteristically inhibitory to Leydig cells steroidogenesis, increases in androgen production may be observed in some models of infection.592., 604. Likewise, experimental models involving active immunization, spontaneous autoimmunity, vasectomy, or tumors suffer from similar complications, where it is difficult to separate the inflammatory response from the immune response. More precise information can be obtained from studies using LPS, other TLR ligands, or inflammatory cytokines, to induce inflammation in the absence of a specific pathogen or antigen.
Lipopolysaccharide-Induced and Cytokine-Induced Inflammation Models
Intraperitoneal or intravenous injection of LPS in rats, mice, and rams or induction of sepsis with cecal matter in rats is followed by a significant decrease in intratesticular and serum testosterone levels.322., 658., 688., 689., 690., 691. There have been distinct differences reported in the dynamics and severity of this inhibition, from study to study and between different species. Responses in mice generally appear to be more rapid, larger, and more prolonged than those of rats, which showed a biphasic response at approximately 6 and 24 h after treatment with a brief recovery period in between.322., 393., 689., 690., 692. Serum testosterone levels actually appear to rise in the boar following endotoxin treatment, as a result of increases in pulsatile LH secretion.693., 694., 695. However, one should not conclude that all differences in experimental results obtained using LPS are simply due to differences between animal models. The LPS obtained from different bacterial strains, and even from different batches of the same strain, can have quite different chemical composition and potency, and is usually contaminated to varying degrees by other TLR ligands.447
What is evident is that systemic administration of LPS exerts predominantly inhibitory effects on Leydig cell steroidogenesis at the testicular level and at the hypothalamic-pituitary level, and inhibition may also involve peripheral responses to inflammation, such as corticosteroid production (Figure 19.8). In the rat model, suppression of steroidogenesis by LPS across a very broad range of doses was found to be due to inhibition of both LH secretion and a reduced capacity of the Leydig cells to produce testosterone in response to LH-stimulation.322., 393., 696. Furthermore, the data indicated that the early phase of suppression of testosterone was due primarily to direct inhibition of Leydig cell steroidogenesis, whereas the later phase involved extratesticular mechanisms, including glucocorticoid-induced inhibition.322., 393.
Similar inhibitory effects to those caused by LPS on testicular steroidogenesis and serum testosterone levels can be induced in male rats by infusion of TNF alone.465., 466. In the human, Van der Poll and colleagues injected six healthy men with recombinant TNF and measured serum concentrations of gonadotropins, testosterone, and sex hormone-binding globulin (SHBG).697 The TNF induced an early and transient increase in serum LH, whereas the concentrations of FSH remained unchanged. The increase in LH concentrations was followed by a transient decrease in serum testosterone levels after 4 h. LH had returned to control values when the testosterone levels reached a nadir, and SHBG was not affected. The results indicate that TNF affects the hypothalamic-pituitary-testicular axis at multiple levels in normal men.697 In a similar study, serum testosterone was decreased without significant effects on LH levels in men treated with high doses of IL6.698 In yet another study, male patients treated with high doses of IL2 as therapy for metastatic cancer were found to have significantly reduced serum testosterone levels.699 Finally, multiple inflammatory markers, including IL1β, TNF, and IL6, are elevated in obese men and men with type 2 diabetes,700., 701. conditions known to be associated with hypogonadism and decreased circulating testosterone levels. Collectively, these experimental and clinical observations predict that in conditions characterized by elevated inflammatory mediators, in the absence of infection, there is a concomitant decrease in serum testosterone levels.
Effects of Inflammation on Leydig Cell Steroidogenesis at the Testicular Level
Most evidence suggests that the effects of LPS-induced inflammation on the Leydig cell itself are mediated through exposure to increased levels of pro-inflammatory molecules that inhibit steroidogenesis, most notably IL1, TNF, NO, and other ROS, and PGE2, as outlined earlier (Figure 19.3). Although isolated Leydig cells express the LPS receptor subunits at relatively low levels,388 and there is evidence that they respond to LPS by increased expression of inflammatory cytokines, including IL1β and TNF,431., 450. it is extremely difficult to completely eliminate macrophages from Leydig cell preparations. Consequently, it remains unclear whether LPS can regulate Leydig cell steroidogenesis by direct action. Regardless of this uncertainty, there are several significant sources of pro-inflammatory molecules within the testis. The testicular macrophages produce IL1, TNF, NO, and other ROS, and PGE2 when stimulated by LPS, although their production capacity is reduced compared with macrophages from other sites.243., 269., 270., 274., 276., 277., 394., 395., 464., 615., 645., 702. Sertoli cells, peritubular cells, and spermatogenic cells are also potential sources of these pro-inflammatory molecules.369., 381., 461., 615., 641. The role of NO and other ROS may be particularly important: treatment of adult mice with LPS causes oxidative damage to Leydig cells, which manifests as a marked reduction of the mitochondrial electrochemical gradient, decreased STAR and HSD3β protein levels, and a fall in serum testosterone, similar to the effects of oxidation by hydrogen peroxide in cultured Leydig cells.652., 653., 654. Moreover, treatment of rodents with NOS inhibitors counteracts the decrease in serum testosterone levels caused by stress and sepsis.657., 658. Curiously, pretreatment of adult rats with the PTGS2 inhibitor, celecoxib, reduced endogenous intratesticular PGE2 levels, and partially reversed the inhibition of testosterone in response to LPS, without blocking the increased expression of IL1β, TNF, or NOS2 in the testis.619
An increase in peripheral levels of cytokines from the circulation due to activation of macrophages and other immune cells in the blood and tissues may also be involved in inhibiting Leydig cell steroidogenesis during LPS-induced inflammation. This may involve the same pro-inflammatory molecules that are produced in the testis following LPS treatment, but potentially other cytokines as well. For example, IL2, which is an autocrine T cell growth factor, inhibits gonadotropin-stimulated testosterone production by rat Leydig cells at the level of CYP17A,703 but IL2 also stimulates IL1, TNF, and IFNγ.704., 705.
The Neural-Immuno-Endocrine Axis in Control of Testicular Steroidogenesis
In addition to direct effects of inflammatory mediators on the Leydig cells, testicular steroidogenesis is modulated during inflammation by neuroendocrine and neuroimmunological regulatory networks (Figure 19.8). These include the hypothalamic-pituitary-adrenal axis, central control of gonadotropin secretion from the anterior pituitary and neural inputs into the testis that regulate Leydig cell function by direct action or through changes in the testicular vasculature.
Activation of the hypothalamic-pituitary-adrenal axis and resulting production of corticosteroids are the key elements of the normal stress response that, among other things, modulates and limits the severity of the inflammatory response.124., 174., 175., 176., 177. Moreover, corticosteroids inhibit LH and FSH secretion from the pituitary,706 but also regulate steroidogenesis directly via specific receptors expressed on the Leydig cell surface,255., 707. acting principally through suppression of the critical steroidogenic enzymes, CYP11A, HSD3β and CYP17A, and induction of Leydig cell apoptosis.192., 708., 709. The response of the Leydig cells to corticosteroids is modulated by the actions of corticosteroid 11β-dehydrogenase (HSD11β), and inhibition of Leydig cell function occurs when glucocorticoid levels exceed the capacity of this enzyme to metabolize them.192 These mechanisms are activated, and impact upon testicular steroidogenesis, in LPS-induced inflammation.393 Hypothalamic corticotropin-releasing hormone (CRH) also inhibits gonadotropin secretion during stress and inflammatory responses.710
Inflammatory mediators themselves can directly act upon the hypothalamic-pituitary unit to inhibit reproduction. Several studies have shown that LPS, IL1, IL6, and TNF can inhibit GnRH release and/or LH secretion at the level of the hypothalamus and pituitary,711., 712., 713., 714. although NO appears to have the opposite effect.715., 716. Furthermore, the observation that IL1 administered intracerebrally can reduce Leydig cell testosterone responses in the absence of decreased LH secretion, for example, in rats pretreated with a GnRH antagonist and therefore lacking LH secretion, led Rivier and colleagues to postulate that there is a direct neuronal connection between the brain and the testis that is activated by cytokines and other stressors within the central nervous system.717., 718. Injection of pseudorabies virus, a transganglionic retrograde tracer, into the testes labeled the spinal cord, the brain stem, and the hypothalamus, while spinal cord injury significantly reduced this staining, and abolished the ability of either IL1β or CRH administered centrally to inhibit testosterone responses to hCG.719., 720. Studies indicate that these agents stimulate a neural pathway within the brain, possibly involving central catecholamine pathways and NO, which suppresses Leydig cell function by inhibiting cholesterol transport protein levels, independent of effects on the pituitary.720., 721.
Furthermore, there is considerable evidence for the control of steroidogenesis by conventional neural pathways acting either directly on the Leydig cells or on the testicular vasculature. Leydig cell steroidogenesis and circulating androgen levels are extremely sensitive to reductions in testicular blood flow, as may occur during inflammation.658., 722. Moreover, Leydig cells can be directly inhibited by a number of neurotransmitters, including serotonin and the catecholamines and the testis is well-supplied by both catecholinergic and serotonergic innervation.723., 724., 725., 726. Activation of these neural pathways during inflammation, or alterations in other neural pathways supplying the testis, may contribute to the overall suppression of Leydig cell steroidogenesis.
Vascular Inflammation and Leydig Cell Function
While changes in testicular blood flow affect Leydig cell function and steroidogenic output, disruption of blood flow can be a source of inflammation as well. Torsion of the spermatic cord, or experimental clamping of the testicular artery in laboratory rodents, renders the testis ischemic, and subsequent reperfusion causes an increase in oxidative stress and intratesticular inflammation, as indicated by increased intratesticular IL1β and TNF expression, activation of NFκB and stress-related kinase signaling pathways in the Sertoli cells, and neutrophil recruitment, resulting in decreased testosterone production and spermatogenesis.82., 457., 727., 728. Local increases in reactive oxygen and nitrogen species are also implicated in the suppression of Leydig cell steroidogenesis in this model.648., 729., 730. The degree of damage (ischemia-reperfusion injury) is related to the duration of the torsion and resulting stress, and Leydig cell function is compromised in the long term.82 Similar effects on Leydig cell steroidogenesis can be observed after treatment of rats with other vasoactive inflammatory mediators, such as serotonin and histamine.731., 732.
Effects of Inflammation on Spermatogenesis
Spermatogenesis is affected by inflammation through direct interactions with the inflammatory process and indirectly through alterations in hormone levels and the testicular vasculature. Specifically, this section is concerned with the effects of innate immune responses on Sertoli cell function and spermatogenesis, as distinct from antigen-specific reactions to the cells of the seminiferous epithelium.
Lipopolysaccharide-Induced Inflammation and Spermatogenesis
Acute systemic administration of LPS has been found to affect spermatogenesis in a number of species, resulting in apoptosis and progressive loss of spermatogenesis in the testis within days of administration, and reduced sperm concentration, motility, and viability for extended periods afterward.322., 499., 688., 693., 694., 696., 733. In the adult rat, administration of a single dose of LPS across a very broad range of sublethal concentrations inhibited the formation of testicular interstitial fluid and suppressed intratesticular and circulating levels of testosterone, by inhibiting LH secretion from the anterior pituitary and reducing the capacity of the Leydig cells to produce testosterone in response to stimulation.24., 322., 393., 696. At the highest doses of LPS, this was followed by apoptosis of spermatogonia and spermatocytes, and premature release of spermatocytes and early round spermatids from the epithelium several days later.322 Detailed stereological analysis established that the effects on spermatogenesis were quite specific.696 At 24 h after treatment, there was a significant delay of spermatocyte development at the leptotene/zygotene phase of meiosis, followed within 6 days by premature release of these cells and the adjacent, but more luminally located, generation of round spermatids. There was also an increase in apoptosis of nearby spermatocytes and spermatids within the same timeframe.696 These effects occurred more rapidly and were quite different from the effects of androgen withdrawal, which inhibits the release of mature spermatids into the tubule lumen and the integrity of the junctions between the Sertoli cells and midphase round spermatids.734., 735. In fact, lower doses of LPS caused a similar degree of inhibition of intratesticular testosterone levels to that caused by high dose LPS, without causing significant spermatogenic damage.24., 322. Moreover, even very high doses of LPS did not reduce intratesticular levels of testosterone in the rat much below 30% of control, well above the threshold necessary to sustain spermatogenesis in this species.79., 80., 734.
Although there are elements of the LPS-mediated response that resemble damage caused by testicular heating, in fact, rodents have a very poor fever response to LPS and the doses of LPS that affect spermatogenesis in rats actually caused a fall in body temperature.24., 322., 736. Nor was the pattern of spermatogenic degeneration consistent with vascular disruption. Administration of LPS did appear to alter blood flow through the rat testis, possibly as a result of vasodilation of the testicular arteries due to increased testicular NOS2 expression,322., 641. but there was no evidence of ischemia or ischemic damage.276., 322. Paradoxically, while LPS treatment caused an increase in vascular endothelial cell leakage in the testis, the inflammation was not accompanied by edema; in fact, interstitial fluid volume in the testis fell quite dramatically in this model.322
A primary cause of damage to spermatogenesis in LPS-induced inflammation could be the action of pro-inflammatory regulators on the seminiferous epithelium itself. This might be the result of increased levels of circulating molecules as well as their local production by intratesticular immune cells and somatic cells. In addition to the resident macrophage population, LPS stimulates an increase in intratesticular monocytes in the rat,276., 285. and neutrophils in the boar testis interstitium.694., 695. In vivo, LPS treatment upregulates testicular expression of pro-inflammatory genes, including CCL2, IL1β, TNF, IL6, and NOS2.274., 276., 285., 322., 393., 396., 619., 641., 691., 737., 738., 739. These molecules are constitutively produced by testicular macrophages, Sertoli cells, peritubular cells, Leydig cells, and/or spermatogenic cells, and their production can be stimulated by LPS in most, if not all, of these cells in vitro. Most importantly, these molecules are involved in normal spermatogenic function, with direct and complex effects on spermatogonial and spermatocyte development, the tight junctions of the blood–testis barrier, and local immune cell activity, mediated via inflammatory signaling pathways within the seminiferous epithelium (Figure 19.14). Moreover, FAS and FASL, which have been implicated in regulating spermatogenic cell apoptosis under normal and pathological conditions, are also increased in the seminiferous epithelium of the LPS-treated mouse.499 Overexpression of these regulators and universal activation of inflammatory signaling pathways in the testis, induced by LPS, would disrupt the normal regulatory processes underlying spermatogenesis. Critically, as most of the normal functions of the seminiferous epithelium are dependent upon androgen support, these effects of inflammatory disturbances could be exacerbated by the concomitant reduction in testosterone levels within the testis.
The severity of the LPS-induced inflammation appears to influence the resulting pattern of spermatogenic damage. In some studies in rats, high doses of LPS are associated with rapid and pronounced epithelial damage, and focal spermatogonial/spermatocyte apoptosis, within 3 days after LPS administration.24., 322. Studies have demonstrated an association between these earlier, more severe, damage events and testicular oxidative stress responses,691., 740. and anti-oxidants can have a protective effect on testicular damage responses to LPS in vitro and in vivo.738., 741., 742. Responses include the induction of the stress proteins, heat shock protein 60 (HSP-60), HMGB1 and HMGB2, as well as increased lipid peroxidation, reduced antioxidant activities, mitochondrial dysfunction and spermatogenic cell apoptosis. Several of these induced molecules can activate TLR signaling as well,24., 108. potentially mediating additional damage.
In summary, the damaging effects of LPS-induced inflammation on spermatogenesis involve multiple mechanisms related to the severity of the stimulus and inflammatory response. These can include direct or indirect inhibition of Leydig cell function, vascular disruption and oxidative stress, as well as interference with the endogenous inflammatory pathways involved in normal regulation of the seminiferous epithelium (Figure 19.14). Differences in damage susceptibility may be attributable to actual differences in sensitivity to LPS at the cellular level, or the probability that circulating LPS may have variable access to the various compartments of the testis. For example, Sertoli cells are much less sensitive to stimulation by LPS than are testicular macrophages, possibly due to differences in expression of the TLR4 accessory protein, CD14.388 Moreover, higher concentrations may be required to permit sufficient LPS to traverse the vascular and lymphatic endothelium of the interstitial tissue as well as the peritubular basement membrane and cell layers to directly affect seminiferous epithelial function. As would be expected, direct injection of LPS into the mouse seminiferous tubules, via the efferent ducts, caused extensive spermatogenic disruption,743 but there have been no quantitative, comparative studies of the effects of systemic and intratesticular routes of administration to date.
Vascular Inflammation Models
Whereas LPS causes inflammation by direct activation, there are a number of clinical and experimental models of testicular inflammation that are secondary to vascular disruption; these include ischemia-reperfusion injury, vascular malformations (e.g. varicocele), heat or cryptorchidism, and hyperstimulation of the Leydig cells with LH/hCG. These models display similarities, but also some significant differences, to LPS-induced inflammation.
Experimental ischemia-reperfusion injury causes rapid apoptosis of the spermatogenic cells entering mitosis—spermatogonia and the very early spermatocytes—followed by progressive depopulation of the seminiferous epithelium.744., 745. Damage is proportional to the duration of ischemia and, when the procedure is performed unilaterally, the contralateral testis is also affected, which has been attributed to an autoimmune response or to reflex vasoconstriction via neural pathways.746., 747., 748., 749., 750. Although Leydig cell function is compromised, this is not the primary cause of damage in this model.82 Intratesticular inflammation and oxidative stress are implicated, however, because there is activation of NFκB, increased MAP kinase activity and production of IL1β and TNF in the affected testis, together with increased local ROS production and an increase in intratesticular immune cells.457., 727., 729., 751., 752. Accordingly, treatment with antioxidants and anti-inflammatories can reduce the resulting spermatogenic damage.753., 754. The role of NO is less clear, because this molecule has both protective and damaging effects in ischemia-reperfusion injury models, and its impact on the testis may depend on the severity of the ischemic insult.648., 649., 755. The most characteristic feature of experimental ischemia-reperfusion injury is a large increase in neutrophils in the testicular subcapsular venules of both the treated and untreated testis, and testicular damage can be reduced or prevented by blocking neutrophil recruitment or activity.727., 729., 756., 757., 758. Thus, ischemia-reperfusion injury to spermatogenesis is an inflammatory event mediated primarily by infiltrating neutrophils, with altered blood flow and oxidative stress leading to apoptosis of early spermatogenic cells, in particular.
A varicocele is a dilation of the testicular veins and swelling of the pampiniform venous plexus, and is a cause of altered testicular blood flow, frequently observed in male infertility patients.759 In both humans and experimental animals, varicocele is associated with increased intratesticular NO levels and oxidative stress, an increase in intratesticular inflammatory markers, including IL1, and reduced spermatogenesis.760., 761., 762., 763., 764. These observations clearly indicate that this apparently minor vascular anomaly can be a cause of inflammation and damage in the testis, although its clinical significance and the benefits of treatment are a source of ongoing discussion.
Administration of high doses of hCG is used in the treatment of delayed testicular descent in young boys and its impact on testicular function has been extensively studied in adult rats.655 This treatment causes a hyperstimulation syndrome comprising a transient decrease in testicular blood flow, immediately followed by increased testicular blood flow and pressure, opening of the vascular endothelial cell junctions, and an increase in testicular interstitial fluid volume some 16–24 h later.765 The syndrome is accompanied by accumulation of intravascular and interstitial neutrophils in the testis,766., 767. an increase in intratesticular expression of IL1β and other pro-inflammatory cytokines,768 failure of mature sperm release, vacuolation of the seminiferous epithelium and apoptosis, and loss of spermatogonia and primary spermatocytes.676., 769. In rats, the response to hCG can be eliminated by depletion of the Leydig cells with EDS,770 or depletion of neutrophils with a specific antiserum,766 and exogenous IL1β is able to replicate most of the effects of hCG.391., 771. These observations suggest that this is an inflammatory response, possibly mediated via IL1β secreted by the Leydig cells. Interestingly, the vascular response is quite different to that of LPS-induced inflammation, which causes a massive reduction in interstitial fluid volume in the testis,322 although some of the germ cell damage seen is similar in both models. It is significant that disruption of the seminiferous epithelium in both the hCG-hyperstimulation rat model and the ischemia-reperfusion model appear to be dependent upon recruitment of neutrophils to the testicular vasculature or interstitial tissue. Equally significant is the observation that both depletion of the testicular macrophages using liposome-encapsulated dichloromethylene diphosphonate273 or stimulation of the macrophages with latex beads676 exacerbate the effects of hCG on the testis, suggesting that the resident macrophages normally play a role in limiting the inflammatory response.
In general, there is a close relationship between vascular disturbance in the testis and inflammation, production of NO, and oxidative stress, leading to spermatogenic damage. These same processes are also implicated in other models of testicular damage, such as the response to heat or cryptorchidism, irradiation, and posttesticular obstruction events.646., 651., 772., 773. While Leydig cell function is frequently compromised in models of testicular inflammation, reduction of androgens is not the primary cause of spermatogenic failure. It is clear that inflammation has direct inhibitory effects on one or more stages of spermatogenic cell development. It also should be pointed out that, as in most other tissues, inflammatory responses in the testis do not appear to automatically produce autoimmune complications. However, inflammation can lead to increased fibrosis in the testis, presumably through increased production of pro-fibrotic cytokines, such as TNF, TGFβ and activin A, and increased testicular fibrosis is associated with spermatogenic failure and infertility.318., 774., 775.
Inflammation in the Epididymis, Vas Deferens, and Accessory Glands
In comparison with the testis, there have been relatively few studies on inflammatory pathways or the effects of inflammation within the remainder of the male reproductive tract. These tissues display a high degree of regional, or segmental, specialization, corresponding to differential gene expression, leukocyte populations, and physiological functions.
Production and Actions of Inflammatory Mediators
The TLRs are found throughout the epithelium, and in immune cells, of the posttesticular reproductive tract.109., 441., 442., 443., 444., 776. Studies in the rat and mouse have produced some conflicting data, but overall indicate that expression levels of TLR1-6 in the epididymis are similar to those found in the testis, with expression progressively declining toward the vas deferens. On the other hand, expression of TLRs 7, 9, and 11 tends to be higher in the epididymis and vas deferens than in the testis (Table 19.4). This may have an influence on bacterial and viral pathogenicity in the different regions of the tract. The crucial TLR adaptor proteins, MYD88 and TICAM1, have been detected in all segments of the rat epididymis and vas deferens,441 and the TLR4 co-receptor CD14 has been localized to the epididymis in the rat and human, where it appears to be more highly expressed than in the testis.443., 777.
All 10 human TLRs, as well as MYD88, TICAM1, and CD14, have been detected in human prostate samples.778 The TLR2 and 4 proteins were localized to epithelial cells in the rat prostate,779., 780., 781. and TLR2 and 9 have been shown to be expressed by human prostate cancer cell lines.782., 783. In the human prostate, TLR3 and 8 immunoreactivity was observed on the apical surface of luminal epithelial cells, and epithelial cells exhibited TLR9 in the penile urethra.784 In rat seminal vesicles, mRNA for TLR2, 5, 6, 7, 8, 9, 10, and 11 were highly expressed, with low levels of the remaining TLRs.776 Notably, expression of CD14 was higher in the prostate and seminal vesicles of the rat than in any other tissue of the male reproductive tract.443 Studies have shown that sequence variants in TLR4 and the TLR1-TLR6-TLR10 gene cluster in the human are associated with an increased risk of prostate cancer, suggesting a link between susceptibility to inflammation and tumorigenesis in this organ.785., 786.
Roles for TLR signaling in controlling functions of the epididymis and vas deferens, similar to those proposed for the testis, can be anticipated. The epithelium of the caput epididymis comprises numerous functionally active immune cells, particularly dendritic cells and macrophages, but also CD8+ T cells.96., 97., 98., 102. This is consistent with the relatively high expression of TLRs in this region of the epididymis. In the regions of the epididymis closest to the testis, moreover, epithelial cell apoptosis is inhibited by repression of NFκB and STAT signaling and activation of the MAPK3/MAPK1 pathway, under the control of androgens, cytokines, or other secretions from the testis.787., 788. These pathways are normally regulated by pathways involved in TLR signaling, and the relationship between the TLRs and testicular factors in their control requires elucidation. Recently, it was discovered that activin A is highly expressed in the epithelium of the caput epididymis.789., 790., 791. This expression is likely to be driven by TLR signaling, and because activin A is a critical regulator of macrophage, dendritic cell and T cell functions, one can assume that this cytokine influences immune cell activity and inflammation in the proximal epididymis. However, other functions of activin A also may be anticipated. In transgenic mice that overexpress the activin-binding protein, follistatin, fluid resorption and sperm transport in the efferent ducts and proximal epididymis is impaired, suggesting a role for endogenous activin in the control of fluid dynamics.792 Furthermore, activin A has been shown to induce SRD5α in genital fibroblasts,793 and the caput epididymis is the site of highest expression of this enzyme, indicating that activin A may regulate androgen activity in the epididymis.
In the more distal epididymis (corpus and cauda) and passing into the vas deferens, the pattern of cytokine and inflammatory regulator activity is different to that found in the caput epididymis (Figure 19.15 ). Activin A production progressively declines, while local production of follistatin increases to the point where follistatin expression levels are higher in the murine vas deferens than in any other tissue examined.537., 789., 791., 794. These observations suggest that controlling activin activity in the distal epididymis and vas deferens is functionally important. Significantly, the progressive reduction in activin bioactivity parallels the reduction in immune cell number and activity in the epithelium. Expression of TGFβ1 also appears to be greatest in the caput of the marmoset and mouse,794., 795. but displays a more distal distribution in the rat.794., 796., 797. Furthermore, TGFβ3 was most highly expressed in the corpus of the rat epididymis, but TGFβ2 was not detected.797 Removal of the testis or inhibition of SRD5α activity increased expression of both TGFβ1 and TGFβ3, particularly in the caput.796., 798. These studies indicate species-specific differences in TGFβ expression that are, most likely, androgen-regulated. Functional roles for TGFβ in the epididymis or vas deferens have yet to be established, but a potential role for TGFβ1 in regulating tight junction formation between epithelial cells from the vas deferens has been demonstrated.799 In direct contrast with activin A and TGFβ, NOS enzyme activity and PTGS2 expression are much greater in the cauda epididymis and vas deferens than in the more proximal regions of the epididymis.614., 616., 800., 801.
FIGURE 19.15.
Region-specific immunity in the epididymis and vas deferens.
Activin A and its binding protein, follistatin, are inversely distributed in a distinctive proximal-to-distal pattern, with highest levels of activin A expression in the caput, and highest follistatin expression in the distal vas. Elevated production of activin A coincides with increased numbers and apparent activity of intra-epithelial immune cells, most notably dendritic cells, but also macrophages and CD8+ T cells. The immunoregulatory enzyme, indoleamine 2,3-dioxygenase (IDO), is highly expressed in the caput, with the exception of the region directly adjacent to the testis (initial segment). Susceptibility to sperm antibody formation following damage to the reproductive tract increases in a proximal-to-distal pattern, indicating a progressive decline in local immunoregulatory mechanisms.
The data indicate that sperm are exposed to a dynamically changing immunoregulatory environment regulated by local inflammatory signaling pathways in the epithelial cells of the male reproductive tract. Moreover, there is evidence that the sperm themselves respond directly to inflammatory stimuli. Human sperm express TLR receptor proteins and respond to bacterial TLR2 and 4 ligands with reduced motility and increased apoptosis.442., 802., 803. TNF has been shown to regulate sperm membrane lipid peroxidation and NO production, and inhibit sperm viability and/or motility,804., 805., 806. and MAP kinase signaling pathways in sperm are implicated in controlling motility, capacitation, and the acrosome reaction.807
Effects of Inflammation
There have been limited studies on the effects of inflammation, as distinct from infection, on cellular and molecular responses of the epididymis, vas deferens, or accessory glands. In rats, systemic administration of LPS induced activation of NFκB in the epididymis and increased several inflammation-responsive genes, including IL1β, although it was not clear that this was a response of the epididymal epithelium, since TLR4 expression was also localized to macrophages in the stroma.443 In a similar study, no immediate morphological changes to the epididymis, at least at the light microscope level, were observed.322 Injection of LPS directly into the caput epididymis of the rat, however, caused an increase in IL1β expression, as well as hyperemia, edema, and accumulation of immune cells in the interstitium, along with inflammatory damage to the epididymal epithelium, reduced sperm motility and decreased expression of several β-defensins.808 Similarly, retrograde inoculation via the vas deferens using E. coli as a source of LPS caused a neutrophilic infiltration and edema in the cauda epididymis and proximal vas deferens, and elevated expression of IL1α, IL1β and IL4, although several other cytokines, including IL6 and TNF, were not significantly altered three days later.809 The IL1 receptor has been detected in the murine epididymal epithelium, indicating the ability to respond directly to this fundamental pro-inflammatory cytokine.415 An experimental left varicocele in the rat significantly reduced epididymal weight and diameter of the duct, together with increased apoptosis of principal cells, proportional to the duration of the varicocele.810 Chronic treatment of mice with the anti-inflammatory PTGS2 inhibitor, nimesulide, caused epithelial cell damage and apoptosis in the vas deferens, and a large reduction in sperm motility, but not sperm count or viability.811
Treatment with LPS of the murine caput epididymal epithelial cell line PC1, which expresses TLR4, induced NFκB activation and expression of the pattern-recognition receptor for bacterial peptidoglycan, nucleotide-binding oligomerization domain 2 (NOD2).812 Sequential stimulation with LPS and the peptidoglycan muramyl dipeptide enhanced TNF mRNA expression in these cells. Infection of rat cauda epididymal epithelial cells in culture with Staphylococcus aureus activated NFκB and MAPK14, production of IL1β, TNF and NO, and expression of mRNA for TLR2 and NOS2.444 Similarly, treatment with LPS and IFNγ together stimulated NOS2 expression and NO production by cultured rat epididymal epithelial cells.813 The epithelial cells of the prostate and seminal vesicles also respond to inflammation induced by LPS or bacteria, with activation of NFκB, increased TLR receptor expression, production of inflammatory mediators, such as NO, and various chemokines, resulting in immune cell infiltration in vivo.103., 779., 781., 814., 815.
Overall, these studies provide clear evidence that inflammation both regulates and disrupts cellular responses throughout the male reproductive tract, with direct consequences for function and fertility. There are obvious parallels with the testis, but the underlying mechanisms in these tissues remain to be fully elucidated, and more investigation is warranted. This is particularly important when considering syndromes of chronic, antibiotic-resistant pelvic or perineal pain, such as chronic prostatitis, which are suspected to have an inflammatory basis, but lack a coherent etiology or pathology.10., 11., 12.
Inflammation, Semen, and Seminal Leukocytes
In humans, seminal plasma is generally the only window available for investigating the immunology of the male reproductive tract. More invasive procedures such as biopsies are rarely performed, produce limited and selective samples, and have the potential to cause inflammatory reactions themselves. Much is made, therefore, of the relationship between measurements of leukocytes, antibodies and cytokines in seminal plasma, and fertility. However, while there is little doubt that sperm antibodies contribute to infertility, and that elevated cytokines are an indicator of ongoing infection, inflammation or other immune events, the diagnostic significance of cytokines and, especially, leukocytes in the semen is a subject of ongoing debate.
Many pro-inflammatory and immunoregulatory cytokines are found in samples of seminal plasma, even those from men with apparently normal fertility.816 The list also includes molecules that modulate the activity of specific cytokines, such as the soluble IL2 and IL6 receptors, and follistatin.817., 818. This is not particularly surprising because these cytokines are present in most biological fluids and are modulated by the general health status of the individual. Elevated cytokine concentrations, both pro- and anti-inflammatory, in seminal plasma are commonly associated with inflammation, infection and increased leukocyte numbers in the male tract, but specific positive and negative associations with infertility of a noninflammatory nature have proven less consistent.819., 820., 821. In a recent evaluation of the extensive literature on this topic, Pilatz and colleagues concluded that, while seminal cytokine profiles might be useful for diagnosis and monitoring of urogenital infections and inflammation, there was no obvious evidence for any associations with poor semen quality or infertility.822 However, a relationship has been reported between NO levels and oxidative stress in seminal plasma and infertility due to varicocele and other vascular anomalies, or prostatic inflammation/autoimmunity.823., 824.
An elevated number of leukocytes in the semen is usually considered to be an indication of infection, and the World Health Organization (WHO) has set an arbitrary level of 1 × 106/ml as the threshold of normality. In fact, leukocytes are present even in semen of fully fertile men, with many studies agreeing that 104–105/ml is a relatively normal value, and even in men with urogenital infections leukocyte numbers do not always reach the WHO level.358., 825. Indeed, leukocyte numbers alone may be a very unreliable indicator of the presence or absence of infection. The origin of these cells is also somewhat obscure. Evidence suggest that the epididymis or vas may be a major source,364., 365., 366. but the primary leukocyte subset present in most semen samples are neutrophils.358., 359., 360., 361., 362., 363., 364. Neutrophils are not a normal feature of the tissues of the tract, but may enter the semen via the accessory glands. This may involve the production of neutrophil-attracting chemokines, such as CXCL6 and CXCL9, by epithelial cells in the epididymis and prostate.826., 827.
Finally, the impact of these cells on fertility is poorly understood. Some studies have shown a relationship between leukocytospermia and impaired sperm function,828., 829., 830. and data suggest that these cells might be a major source of ROS or other molecules causing sperm damage.358., 831., 832. Other studies have failed to confirm a clear link between seminal leukocytes and infertility or sperm antibody formation.358., 362., 833. It even has been suggested that they might play a beneficial role by the removal by phagocytosis of abnormal sperm or assisting in sperm capacitation.343., 362. It is fair to say that the role of leukocytes in semen is more complicated than might be expected, and many questions still remain.358., 364., 833.
Autoimmunity in the Male Reproductive Tract
Sperm Antibodies
Clinically, the most common immunological dysfunction of the male reproductive tract is the formation of sperm antibodies, which may inhibit sperm motility through agglutination reactions, target the sperm for immunological destruction in the male or female tract, or block essential surface receptors and molecules required for fertilization (Figure 19.16 ).834., 835. There is no single dominant antigen associated with sperm autoimmunity, and multiple antigens are usually involved.7., 8., 9., 291. These antigens have been extensively studied in experimental animals and in infertile patients, and include numerous sperm surface, as well as intracellular proteins, glycoproteins, and glycolipids.
FIGURE 19.16.
Sperm antibody binding sites and effects.
The principal sequelae of antibody binding to spermatozoa are: (1) activation of immune cells against the sperm via complement or through interaction with immunoglobulin Fc receptors on phagocytes leading to cytotoxicity; (2) blocking or interfering with surface recognition molecules, in particular the egg binding receptors in the postacrosomal region; and (3) agglutination caused by cross-linking of sperm by multivalent antibody which may impede the ability of sperm to swim freely in the female tract.
The Origins of Sperm Antibody Formation
Physical damage to the male reproductive tract can lead to sperm antibody formation. This is best exemplified by the high incidence of sperm antibodies associated with vasectomy, where up to 70% of patients develop antibodies after the procedure.346., 347., 348. Blunt trauma and testicular biopsy may cause sperm antibodies to form as well.836., 837. Sperm antibodies are associated with obstructive azoospermia and congenital absence of the vas deferens,348., 838. and with reproductive tract infection involving orchitis and/or epididymitis, although there is evidence that inflammation alone may not be sufficient to cause antibody formation.839., 840. There is a definite relationship between sperm antibodies and the existence of other autoantibodies, suggesting an important role for genetic factors in predisposition to sperm antibody formation.841., 842. A separate subset of antibodies directed against steroidogenesis-specific antigens expressed by the Leydig cells is associated with polyglandular autoimmunity, although sperm antibodies also may be found in patients with these conditions.843., 844.
In animal models, sperm antibodies can be induced by active immunization with spermatogenic cells or whole sperm, or with testicular or epididymal extracts. This procedure usually involves the use of adjuvants to boost the immune response, but protocols leading to antibody formation without adjuvants also have been reported.845 As in humans, antibodies may develop following experimental vasectomy or vasal obstruction in rats, guinea pigs, mice, rabbits, and monkeys.97., 332., 349., 351., 352., 846., 847. The strength of the response and its outcomes are very much strain-dependent. In some of these experimental models, the formation of antibodies is associated with the development of orchitis and variable degrees of damage to the seminiferous epithelium, an outcome that is rarely, if ever, observed in humans under the same conditions.
Measurement and Clinical Significance of Sperm Antibodies
There has been considerable debate regarding the clinical significance of sperm antibodies or, perhaps more precisely, the tests used to measure them. Although antibody levels in seminal plasma or even blood are frequently quantified, it has been suggested that only antibodies that are actually bound to the sperm, and more specifically the anterior portion of the sperm, should be considered to be of real significance, and then only when a majority of the sperm are affected.2., 835., 848., 849. Hence, studies that use tests that detect sperm bound antibodies (e.g. agglutination and immunobead binding tests) are most likely to provide reliable results.834., 850., 851. Although IgG can cross the reproductive tract epithelium at sites such as the rete testis to bind to sperm,852 it also has been argued that only IgA antibodies are evidence of a specific mucosal immune response and should be considered of greater prognostic value in semen samples. The most rigorous approach to this issue involves confirmation that the antibody binding has functional consequences using a sperm function test, such as the mucus penetration assay, before accepting the pathological significance of the antibodies.2., 835., 849.
Even allowing for the variation in methods employed, it appears that populations vary widely in the incidence of sperm antibodies. This may be due in part to genetic differences in susceptibility to antibody formation, although the prevalence of reproductive tract infections in the population and a lack of suitable treatment options are probably a stronger determinant. In developed nations, the incidence of sperm antibodies is around 5–10% of all infertility cases.2., 3. On the other hand, in parts of central Africa where treatment for reproductive tract infection is limited, or there is a reliance on self-medication and alternative therapies, the incidence of sperm antibodies among infertile populations is much higher.853 In considering these figures, it also should be borne in mind that sperm antibodies may be present in the semen or blood of men of proven normal fertility.854 Moreover, in the absence of any other underlying cause being evident, the presence of sperm antibodies tends to be assumed to be the cause of the infertility. It is quite possible that the incidence of infertility due primarily to sperm antibody reactions is lower than these values might suggest. While treatment with high dose corticosteroids has been used with limited success in the past to treat sperm antibodies, sperm washing coupled with intrauterine insemination, in vitro fertilization or intracytoplasmic sperm injection are the current methods of choice.834., 837.
Equally serious for the infertile couple is the development of sperm antibodies in the female tract. While largely outside the scope of this review, it is important to recognize that histocompatibility differences between partners may create potential problems for sperm survival and successful fertilization.855., 856., 857.
Autoimmune Orchitis
The Etiology of Orchitis
In humans, isolated orchitis is usually associated with viral infections, most commonly postpubertal mumps.858., 859., 860., 861., 862., 863. Nonviral causes include mycobacteria (e.g. Mycobacterium leprae and M. tuberculosis) and parasites, such as Filariasis and Malaria. Bacterial orchitis is usually caused by retrograde ascent of urethral pathogens, such as E. coli, Chlamydia trachomatis, and Neisseria gonorrhoeae, and is almost invariably associated with inflammation of the epididymis (epididymo-orchitis). In both clinical and experimental models of infectious orchitis,313., 603., 864. the immunological response to the infection may cause collateral damage to testicular cells, causing obstruction, fibrosis, and displacement or ischemia, and increases the risk of subsequent autoimmune reactions against testicular antigens. Autoimmune orchitis may also develop as a consequence of neoplasia, in response to an external antigenic challenge (e.g. vasectomy or active immunization), or may arise spontaneously, in the sense that no obvious cause can be discerned.859., 860., 865. Whereas infectious orchitis resolves once the infection has been cleared, autoimmunity may lead to chronic inflammatory disease. In contrast with the incidence of sperm autoantibodies, however, autoimmune reactions against spermatogenic cells within the testis appear to be comparatively rare among most human populations.
While there are several animal models of spontaneous autoimmune orchitis, generally attributable to genetic or epigenetic causes, in the mink,866 rats,867 and dogs,868 the incidence of such events have been difficult to establish in humans, given that the autoimmune event and its diagnosis might be separated by many years. There is evidence for immune complex formation in the basement membrane of the seminiferous epithelium,869., 870., 871. and a high prevalence of asymptomatic testicular inflammatory lesions,4., 5. among infertile men. The problem lies in establishing whether these lesions are evidence of a past autoimmune reaction or secondary to inflammation from other causes. The testis is one of the organs that is affected in PGA syndromes, which are caused by failure of normal mechanisms of tolerance,166., 167., 872. suggesting that other genetic causes for autoimmune orchitis may exist. There is some evidence of an association of orchitis with other autoimmune inflammatory diseases, particularly vasculitis (inflammation of the blood vessels) in men.873 A genetic predisposition for development of autoimmunity may require a secondary trigger, in the form of a precipitating infection, physical trauma to the reproductive tract or other inflammatory event, in order to proceed to actual autoimmune orchitis.
Experimental Autoimmune Orchitis
Several experimental animal models of autoimmune orchitis have been extensively studied by a number of groups, seeking to understand testicular immunophysiology and the causes of autoimmune infertility.155., 311., 313., 314., 315., 331., 845., 874., 875. Most of these models involve immunization with testicular antigens, including spermatogenic cell, somatic cell, and extracellular matrix antigens, using complete adjuvants (containing active or inactivated microbial agents), or adoptive transfer of lymphocytes from animals with on-going autoimmune orchitis. However, at least one model of induced orchitis involving subcutaneous injection of syngeneic (genetically identical) spermatogenic cells into a susceptible strain of mice has been described.876., 877. A particularly interesting model of orchitis development occurs subsequent to vasectomy in some animals. Whereas vasectomy leads to sperm antibody formation alone in most strains of rats and mice, vasectomy results in a rapid progression to full-blown orchitis in the Lewis rat,349., 350., 351. and SWR/J mouse.878 Similar responses to vasectomy or epididymal obstruction have been observed in the rabbit,879 guinea pig,880 and rhesus monkey.881 Finally, removal of the thymus gland around postnatal day three in mice causes polyglandular autoimmunity, which can include autoimmune orchitis similar to that found in humans,155., 874., 875. indicating that immunoregulation by T cells is involved in the normal protection of at least some testis-specific antigens. These models have been crucial to the development of our concepts of the control of immune responses in the testis.
The majority of research has been focused upon autoimmune reactions to the germ cells, but testicular autoimmune disease also may involve the somatic cells. Autoimmune polyglandular diseases, most notably Addison’s disease, are associated with specific immune reactions to steroidogenic cells throughout the body, including Leydig cells, thereby leading to hypogonadism.843., 882. This autoimmunity primarily involves reactions against the steroidogenic enzymes and hormone receptors,883., 884. although spermatogenic cell autoantibodies are commonly observed, as well.844
Although the models of experimental autoimmune orchitis differ in respect of the method of induction, there are common features in the progression of disease, which are summarized here. The onset of autoimmune orchitis in animals generally involves the appearance of testicular antibodies and immune complexes, followed by infiltration and activation of macrophages, dendritic cells and effector lymphocytes, invasion of the seminiferous tubules, sloughing of germ cells and focal necrosis, accumulation of neutrophils and/or eosinophils, and finally aspermatogenesis.311., 314., 331., 885., 886., 887. The epididymis and vas deferens are frequently involved. In adoptive transfer studies in mice, it was shown that CD4+ T cells, but not antibody, can transfer orchitis from one animal to another, clearly implicating a cellular rather than humorally-mediated mechanism of disease development.267., 268., 864. In adoptive transfer, moreover, the early disease almost invariably occurs in the region where the seminiferous tubules connect to the rete testis,267., 268. consistent with the fact that this is where access to the antigens of spermatogenesis is likely to be greatest,64., 73., 267. and there is a greater concentration of MHC class II positive macrophages.267., 268., 314., 331., 888. In the adjuvant-free model of induced orchitis, a similar pattern of disease initiation has been observed in this region.889 Active immunization protocols using adjuvant, on the other hand, cause a more widespread induction of disease throughout the testis.267., 311.
Progression of autoimmune orchitis is marked by expansion and activation of the testicular dendritic cell and macrophage populations, which remain elevated throughout the course of the disease.292., 887. Increases in IL17-producing CD4+ Th17 cells and CD8+ T cells, and even Treg cells have been observed during the onset of autoimmune orchitis in the rat, and persist through the chronic phase, indicating that interaction between these cell subsets may determine the course of the disease.885., 886. Several immunodominant antigens have been identified in the rat, and include antigens that are not specific to the testis.291 The pro-inflammatory cytokines, TNF and IFNγ, FAS and FASL, and NOS2 have been identified as potential causative factors in the development of orchitis, principally by stimulating antigen-presentation activity and spermatogenic cell apoptosis.595., 890., 891., 892., 893., 894. Although IL6 production by testicular macrophages increased during experimental autoimmune orchitis in rats,895 and was implicated in the disruption of the inter-Sertoli cell tight junctions,896 administration of IL6 has been shown to inhibit the progression of orchitis in LPS-resistant mice.897 The progression of orchitis may also be inhibited by IL10 and by androgens.898., 899. The crucial requirement in the development of orchitis is to overcome the normal mechanism of systemic tolerance to testicular antigens, as well as the local immunoregulatory mechanisms.
Typically, immunological events induced in one testis can lead to damage in the contralateral testis, a condition sometimes called sympathetic orchiopathia. Such responses have been observed following a unilateral physical or toxic insult,900., 901. infection with E. coli or other pathogens,902., 903. and ischemia or ischemia-reperfusion injury.746., 747., 748., 904. This is usually attributable to generation of autoreactive lymphocytes and antibodies caused by events in one testis traveling to the other testis to initiate disease, in a manner comparable to adoptive transfer of autoimmunity from one animal to another.
The precise reasons why some animal strains are more susceptible to the development of autoimmune orchitis remain to be fully explained, but it is obvious that susceptibility to autoimmune reactions to sperm and testis antigens is genetically determined.905., 906. Clinically, there is an association between sperm antibodies and other autoimmune antibodies,841., 842. and the development of sperm antibodies following vasectomy is strongly associated with the MHC.907 In mice, specific orchitis susceptibility genes have been mapped to both MHC and non-MHC regions, and the loci linked to development of orchitis also have been shown to govern susceptibility to other autoimmune diseases, such as encephalomyelitis and diabetes.908., 909.
Autoimmunity in the Epididymis, Vas Deferens, and Prostate
Clinically, epididymitis is the most common intrascrotal inflammation, manifesting as pain, nodules, edema, urinary difficulties, fever, urethral discharge, and/or infertility, although it also can be asymptomatic. Acute epididymitis is most commonly due to retrograde ascent of urethral pathogens and sexually-transmitted bacterial infections—most notably C. trachomatis and N. gonorrhoeae, but also Ureaplasma urealyticum, E. coli, Enterococcus faecalis, and Pseudomonas aeruginosa.860 Less frequently, systemic bacterial infections, such as M. tuberculosis, Haemophilus influenzae type B, and Salmonella, or nonbacterial infections, such as cytomegalovirus and filariasis, may be responsible, especially in children or immunocompromised individuals. Epididymitis also may have noninfectious origins in various medical procedures, the arrhythmia drug amiodarone, trauma, physical stress, vasectomy, urinary reflux, neoplasia, postinfectious and systemic inflammation, vasculitis, and systemic autoimmunity.11., 12., 860., 910., 911. Vasitis is relatively rare and is usually caused by injury, bacterial infection, and following surgical manipulation, postvasectomy or following prostatectomy.912 It may present as pain or be asymptomatic. If the inflammatory response in the epididymis or vas deferens is severe enough, this can lead to damage to the mucosal epithelium and/or obstructive lesions that may inhibit fertility. Prostatitis and vesiculitis are most commonly caused by bacterial infections, although antibiotic-resistant chronic prostatitis/chronic pelvic pain has suspected inflammatory, probably autoimmune, origins.10., 860., 910., 911.
Epididymitis is much more frequent than isolated orchitis, which may be a reflection of closer proximity to the external environment. However, different immunoregulatory mechanisms in the two organs may also be a contributing factor. Experimental evidence suggests that the epididymis may be more susceptible to inflammation and subsequent leukocytic infiltration than is the testis. For example, infiltration of lymphocytes and neutrophils into the interstitial tissue are frequently observed in the epididymis and vas deferens, but rarely in the testis, of aging mice.913 In mice injected intravenously with Bordetella pertussis and adjuvant, neutrophils invade the stroma of the epididymis, vas deferens and accessory glands, but the testis is not affected.914 In the alymphoplastic (aly) mouse, eosinophils and macrophages spontaneously accumulate in the stroma of the epididymis and vas deferens, but not the testis.915
Experimentally, epididymitis can be induced by retrograde inoculation with bacteria, such as E. coli, via the vas deferens, which leads to epididymitis, vasitis and orchitis, but is complicated by the presence of the bacteria itself.95., 809., 916., 917., 918. Noninfectious models of epididymitis involve active immunization, neonatal thymectomy, and vasectomy. Epididymitis (actually epididymo-orchitis) and vasitis can occur following active immunization with testicular extracts in the presence of suitable adjuvants in rats, mice and guinea-pigs.267., 314., 331. Passive transfer of T cells from mice immunized in this manner also leads to epididymitis, with testicular involvement initially confined to the region around the rete testis and the efferent ducts.267., 268., 890. Subcutaneous injection of syngeneic testicular spermatogenic cells into mice in the absence of adjuvant causes orchitis without epididymitis,876 but passive transfer of the lymphocytes from these mice after stimulation in vitro, favors the induction of epididymitis.919., 920. In other words, the transfer of T cells activated specifically against spermatogenic antigens selectively induces epididymitis in nonimmunized mice.889 Removal of the thymus at 3 days of age in mice, which abrogates peripheral tolerance mediated by regulatory T cells,921 initially causes epididymovasitis in the postpubertal period, followed by an increasing, but always smaller, incidence of orchitis many weeks later.874., 922. Likewise, mice with a targeted deletion of the immunoregulatory AIRE transcription factor developed sperm antibodies and leukocytic infiltrates in the epididymis, without orchitis.923 Vasectomized A/J mice that have been immunized with syngeneic testicular spermatogenic cells develop epididymitis independent of orchitis,924., 925. suggesting that the epididymis can be the primary site of immune reactions following vasectomy, as well. The epididymis is also more susceptible than the testis to spermatic granuloma formation in humans and in experimental mice.926., 927. Taken altogether, these observations highlight a greater susceptibility of the epididymis to inflammation, relative to the testis, when responses to spermatogenic antigens occur. Accordingly, detailed studies in mice have established that the immunogenetics underlying susceptibility to autoimmune orchitis and epididymitis are quite distinct.906
Infection and inflammation in the epididymis is characterized by infiltration of macrophages, lymphocytes and neutrophils into the interstitial and peritubular zones, as well as the epididymal epithelium.95., 99., 331., 332., 845., 874., 922., 925., 928. The consequences include reduced sperm productivity and function, increased sperm antibody formation, as well as damage to the epithelium and potential obstruction of the epididymal duct. Similar responses in the vas deferens and accessory glands may interfere with seminal fluid production and sperm transport. As in the testis, these disruptions of function may be due to the actions of the activated immune cells, but may also be due to altered epithelial cell or stromal cell function, through the activation of TLR signaling pathways, pro-inflammatory cytokines and other inflammatory mediators. Importantly, ongoing inflammation in these tissues, regardless of the cause, may be a source of chronic pelvic or perineal discomfort or pain.
Immune Privilege in the Male Reproductive Tract
Based on the data as outlined in previous sections, it is evident that the testis is an immunological target, and that autoimmune responses to spermatogenic antigens will occur given the right conditions, prompting the question: What are the normal systems that exist within the testis to prevent or regulate development of autoimmunity there? In other words, what is the functional and physiological basis of “immune privilege” in the testis? The concept of immunological privilege has generally been used by immunologists to describe tissues that are not readily accessible to antigen-presenting cells and circulating lymphocytes or that have deficient lymphatic drainage to local lymph nodes, such as the brain, the anterior chamber of the eye and the placenta.37., 873., 929. Neither of these properties applies to the testis, and the most common misconception centers on the role of the blood–testis barrier.
The Immunological Role of the Blood–Testis Barrier
The actual location of the blood–testis barrier lies in tight occluding junctional specializations (zonulae occludens) between adjacent Sertoli cells in the seminiferous epithelium (Figure 19.2).16., 17. These elaborate junctions are quite distinct in organization from the normal adhering junctions and gap junctions found in most epithelia and are assembled at the time of puberty to form a complete seal separating the basal and adluminal compartments of the epithelium.15., 866. Their primary role is to create a unique physiological environment for the meiotic and postmeiotic germ cells, and all spermatogenic cells beyond the leptotene or zygotene spermatocyte stage are completely surrounded by the cytoplasm of adjacent Sertoli cells and are entirely dependent upon these cells for their support and regulation.72., 77., 930., 931. The process of movement of the spermatogenic cells themselves through the junctions during spermatogenesis does not involve a breach of junctional integrity.72., 931. Indeed, loss of barrier function is a precipitating factor in many forms of testicular failure, including spermatogenic damage during inflammation and infection.
A broader definition of the blood–testis barrier beyond the concept of a physiological barrier incorporates protection of the spermatogenic cells from harmful influences. Some definitions of the blood–testis barrier, therefore, also include the efflux-pump barrier system involving drug-transport proteins, such as p-glycoprotein and the multidrug resistance associated protein-1, on the capillary endothelium, peritubular myoid cells, and the basal aspect of the Sertoli cells.932., 933. While these elements may certainly be important for protection of the testis against toxic agents, their contribution to immunoregulation is likely to be marginal.
There are several lines of evidence that the blood–testis barrier does not account for all the manifestations of immune privilege in the testis. Studies have shown that spermatogenic cell autoantigens are not confined behind the Sertoli cell tight junctions, and are expressed by spermatogonia and the early spermatocytes.934 Moreover, there is clear evidence that the barrier is incomplete in the epithelium of the rete testis. Significantly, orchitis can be passively transferred to naïve mice by lymphocytes from mice with active autoimmune orchitis.267., 268., 919. In many seasonally breeding species, annual regression of both the later spermatogenic cells and blood–testis barrier occurs without inducing overt inflammation or autoimmunity.72., 866. Finally, the blood–testis barrier cannot explain the enhanced survival of grafts within the interstitial tissue.14., 266. On balance, the data lead to the conclusion that the blood–testis barrier does not prevent exposure of germ cell antigens to the immune system, indeed antibodies and lymphocytes specific for spermatogenic antigens may be a normal feature of the circulating immune repertoire.854 Although the sequestration of a large proportion of the antigenic burden behind the blood–testis barrier no doubt plays a role in this process, it cannot be the primary explanation for functional immune privilege in the testis.
Evidence from Transplantation Studies
Perhaps the most intriguing aspect of immune responses in the male tract is the observation that allografts and even xenografts into the testicular interstitial tissue outside the blood–testis barrier are preserved for extended periods of time, possibly even indefinitely.14., 266., 935., 936., 937. This enhanced survival is not simply due to the reduced ambient temperature of the testis, as grafts to the equally hypothermic skin of the ear are not preserved,266 while grafts continue to survive in testes that have been translocated to the abdominal cavity.266., 936., 937. Moreover, the efferent lymphatics of the testis do not show any evidence of functional deficiency.13 However, intratesticular parathyroid allografts fail to survive in rats sensitized against the donor antigens, while long-established intratesticular allografts are rapidly rejected following active immunization of the recipient with donor tissue.14 These observations led to the hypothesis that local immunoregulation mechanisms are responsible for graft survival, and, more specifically, that the inductive phase of the immune response is suppressed within the testis and/or its draining lymph nodes. In other words, the immune system may be unable to recognize and/or respond to foreign antigens within the testicular environment.
It should be noted that studies on graft survival in the testes of laboratory rodents have been very successful, but the data in other species are less consistent. Similar transplant studies in the ram and cynomolgus monkey have proven unsuccessful.938., 939. Moreover, the type of graft also may be a factor. Selawry and colleagues were able to demonstrate survival of pancreatic islet allografts or xenografts in abdominally-located testes, but not in scrotal testes, of the rat.937 Whether these differences in outcome are due to differences in testicular architecture, differences in lymphatic organization or vascularization, the size, health, and type of graft, the underlying immunogenetics of the donor and host, effectiveness of local immunoregulatory mechanisms, species differences in systemic immunity, or even the surgical procedures employed, remains to be answered.
Evidence suggests that testicular tissue and some testicular cells, in particular, have inherent characteristics that may make them more amenable to transplantation.30., 940., 941. However, just as intratesticular grafts produce different outcomes in different studies, studies on transplantation of testicular tissue have met with variable degrees of success. Early observations were that fetal and postnatal testis tissues are viable as grafts under the kidney capsule of outbred rats, but that adult testis tissue is quickly rejected.941., 942. Perhaps surprisingly, transplantation of fragments of intact testicular tissue from numerous species under the skin of immunodeficient mice tends to be extremely successful, with vascularization, normal steroidogenesis and even complete spermatogenesis becoming established, although there is no exit for the spermatozoa that are produced.943 The ability of spermatogenesis to occur in such grafts is probably related to the reduced temperature of the skin, but immunocompromised recipient animals are generally required to avoid rejection responses. On the other hand, adult testis allografts have been observed to survive under the kidney capsule in immunocompetent mice,489 and allogeneic transplantation of spermatogenic cells, which were subsequently able to undergo normal spermatogenic development, have been successfully performed in several domestic species with intact immune systems.944., 945., 946. Kimmel and colleagues observed that human testis xenografts to the murine kidney failed to survive in intact mice, but survival was possible in recipient mice with an inactivating deletion of MHC class II expression, thereby implicating a CD4+ T cell-mediated rejection process.947 Discovering the reasons why different models produce such widely different outcomes may provide a better understanding of the fundamental requirements for immunological privilege in the testis.
Immune Tolerance and Immunoregulation
It has become increasingly apparent that central and peripheral immune tolerance contributes to testicular immune privilege. This was first indicated by the fact that hypogonadism and sperm autoimmunity are frequently associated with the PGA syndromes, type 1 and 2, which are due to a mutation in the AIRE transcription factor that controls thymic antigen expression during immune development,166 and a defect in regulatory T cell function,167 respectively. Mice with a deletion of AIRE develop sperm antibodies and inflammatory lesions in the epididymis (O’Bryan MK, personal communication).923 Exposure to testicular antigens during maturation of the immune system has been shown to reduce the severity of induced autoimmune orchitis in adult genetically-immunodeficient mice that have had their immune system re-constituted.948 Moreover, the cluster of autoimmune endocrinopathies induced by thymectomy at 3 days of age in mice and rats frequently includes orchitis.155., 872., 874., 875., 922. These disease models in humans and rodents establish a link between testicular autoimmunity and failure of tolerance, more precisely, a shift in the balance between autoreactive T cells and specific regulatory T cells.
Suppressor or regulatory T cells have experienced a major revival in interest in recent times, with most attention directed toward the CD4+CD25+FOXP3+ Treg cell subset, but there is also evidence for CD8+ regulatory T cell populations.949., 950. As would be expected, Treg cells can be found in both the normal and inflamed testis,310., 885. even if their actual role in the testis has yet to be firmly established. Nonetheless, several studies indicate that exposure of T cells to the testis environment induces an immune deviation response, that is, a switch from cell-mediated (Th1) type immunity to a Th2/Treg response that is predominantly immunoregulatory and tolerogenic (Figure 19.7). Injection of soluble antigen into the testis produces specific suppression of T cell-mediated responses against the injected antigens.951., 952., 953. Studies on pancreatic islet cell allografts in the mouse testis indicate that activated or memory T cells directed against graft antigens are destroyed when they enter the testis environment and that graft antigen-specific Treg cells are preferentially produced.954., 955. Isolation of a CD4+ T cell line that was able to downregulate the development of adoptive transfer of autoimmune orchitis in mice has been reported,956 and a crucial role for Treg cells in controlling the development of orchitis in response to vasectomy also has been demonstrated in mice.957 A substantial increase in evidence linking Treg cells and other regulatory T cell subsets to the control of testicular immune responses can be anticipated in the near future.
FAS Ligand
In 1995, Bellgrau and colleagues489 reported that expression of FASL was expressed by mouse Sertoli cells, and that mice deficient in either FASL or its receptor, FAS, did not show evidence of testicular immune privilege. In this study, FAS-FASL interaction was implicated in the prevention of antigen-specific responses within the testis, and subsequently, in other immune-privileged or immune-deficient sites.958 It was proposed that T cells entering the testis and becoming activated by exposure to their respective antigen are immediately killed by interaction between FAS on their surface and FASL, either expressed on, or secreted by, the Sertoli cells. While this was an attractive hypothesis, a number of concerns regarding the FASL expression hypothesis as an overarching explanation for immune privilege arose. Several groups found that expression of FASL did not confer immunoprotection in their own studies in the testis or in other tissues, and in fact caused quite virulent inflammatory reactions in some cases.959., 960. FASL also appeared to be expressed at relatively high levels in epithelia of human tissues not generally thought of as immunologically privileged, such as the esophagus and lung.961 Finally, Kimmel and colleagues reported being unable to detect FASL expression at all in normal human testes which had been flushed of all peripheral blood cell contamination prior to collection of RNA.947
The role of FASL in normal testis function or immunophysiology still remains unresolved. Several studies have suggested that, although FASL mRNA can be detected in Sertoli cells using very sensitive methods such as RT-PCR, the protein is not significantly expressed by the Sertoli cells in the normal adult testis, and in fact it may only be the spermatogenic cells that express the ligand constitutively.492., 959., 962. In the immature rat and porcine testis FASL protein has been detected in Sertoli cells by immunohistochemistry,490., 963. but the specificity of FASL antisera used for this purpose has been challenged in at least one major review of the issue.491 While it appears certain that modulation of FAS and FASL in the seminiferous epithelium plays an important role in regulating spermatogenic cell apoptosis, particularly in various testicular damage models,490., 772., 964. a crucial role in maintaining immune privilege in the testis remains doubtful on the weight of evidence.
Major Histocompatibility Complex Expression in the Testis
Most studies have reported a characteristic absence of expression of both MHC class I and class II proteins on the cells of the seminiferous epithelium under normal conditions.67., 68., 230., 232., 266., 267., 315., 965., 966., 967. This suggests that spermatogenic cells may be able to avoid direct recognition by CD4+ and CD8+ T cells, which might be important for reducing the potential for antigen-specific immune responses in the seminiferous epithelium. On the other hand, there are some data to indicate that mRNA and protein for both MHC class I and class II molecules are expressed in human spermatozoa,968., 969., 970. suggesting that mRNA present in the spermatogenic cells may be translated into protein at some time after they are released from the testis. This delayed expression of MHC molecules may play a role in protection of the sperm against infection or immune cells in the reproductive tract.
In contrast to the seminiferous epithelium, both MHC class I and II proteins are expressed in the testicular interstitial tissue. As would be expected, MHC class I expression is found on most interstitial cells, including the Leydig cells.230., 965. In the rat and human, testicular macrophages and dendritic cells express MHC class II throughout the interstitium,239., 246., 266., 965. but studies on the mouse testis indicate that expression of MHC class II is concentrated on cells in regions adjacent to the rete testis.267., 268. A greatly reduced number of MHC class II positive cells in the ram testis is consistent with the relatively low number of resident macrophages in this species.68 In light of all the observations, however, it appears unlikely that a lack of MHC class II-positive antigen-presenting cells is a contributing factor in testicular immune privilege. Differences in the distribution of such cells, however, may be reflected in differences in susceptibility and development of autoimmune reactions between species, or possibly even between different strains. During the development of experimental autoimmune orchitis, which requires overcoming peripheral tolerance to intratesticular antigens, MHC class II expression and the cells bearing these molecules—dendritic cells and macrophages—are increased within the testis and in the draining lymph nodes.292., 887.
The nonclassical MHC class I genes, HLA-G and HLA-E, are associated with suppression of the adaptive immune response.169., 971. The soluble form of the HLA-G molecule has been localized by immunohistochemistry to Sertoli cells, spermatocytes, spermatids and some interstitial cells in the rhesus monkey testis,972., 973. and expression of HLA-G and HLA-E has been detected in isolated human spermatogenic cells.970 More recently, HLA-G has been detected in the seminiferous tubules and efferent duct epithelium of the human testis.974 This molecule is associated with the placenta in protecting fetal alloantigens from the maternal immune system,169., 972. and its soluble form has been implicated in apoptosis of alloreactive CD8+ T cells.971 Its significance to the testis is entirely unknown at this time.
Immunological Responses in the Testis
It is evident that maintenance of peripheral tolerance is important for testicular immune privilege, and that loss of tolerance to testicular antigens leads to autoimmunity and testicular damage. Obviously, the MHC class II-expressing antigen-presenting cells of the testis must play a crucial role in this process, by creating a tolerogenic environment. The balance of data suggest that the majority of dendritic cells and macrophages in the mouse display reduced expression of MHC class II molecule and the regulatory CD80/CD86 co-receptors,267., 271. suggesting that they may be more likely to promote tolerance, but this does not appear to be true for the rat.266., 272. However, expression of other co-regulatory ligands and receptors, for example CD40LG or inhibitory variants of CD80/CD86,975 have yet to be investigated in these cells. On the other hand, it is well-established that the macrophages of the rat and mouse display the anti-inflammatory/immunosuppressive M2 phenotype, manifesting as preferential production of immunosuppressive cytokines, such as IL10, and a reduced capacity for T cell activation.269., 270., 275., 287., 288. This suggests that the predominantly immunosuppressive phenotype of the testicular macrophages is a critical element in maintaining testicular immune privilege, and that deviation of the macrophage and dendritic cell phenotype toward support of cell-mediated immunity is an essential event leading to autoimmune orchitis.272., 292., 887., 893.
Based on the existing evidence, it can be proposed that circulating CD4+ effector T cells entering the testis are inactivated, deleted, or made tolerogenic by the resident antigen-presenting cell population. This model of tolerance in the testis is consistent with the presence in the testicular interstitial tissue of Treg cells,310., 885. but also of NK and NK T cells (Hedger MP and Aridi DZ, unpublished data).70 Cells of the NK lineage are able to modulate dendritic cell activity to control adaptive immune responses.976., 977., 978. These cells play a key role in promoting graft survival,149., 168. and have been implicated in the generation of CD8+ regulatory/suppressor T cells in another immune privileged site, the anterior chamber of the eye.150., 979., 980. Consequently, it is anticipated that intratesticular NK and NK T cells are involved in production of regulatory/suppressor T cells specific to the testis. Finally, a role for γδ T cells, a minor subset of T cells that possess an alternate TCR structure, has been invoked in suppressing autoimmune reactions in bacterial-induced autoimmune orchitis in the mouse, in part through production of IL10 and TGFβ.313., 903. This lymphocyte subset also appears to be involved in maintaining immune privilege in the eye.981
Testicular Immunoregulatory Mechanisms
Immune privilege, in the classical sense of providing extended graft protection, is restricted to a small number of tissues, so there must be some specific properties of the testis that create an environment where adaptive immunity is so closely regulated. Historically, the spermatogenic cells and Leydig cells have been implicated,982., 983. as they are cell types with functions that are unique to the testis. Early studies showed that spermatogenic cells can inhibit lymphocyte activity in vitro and in vivo.983., 984. More recently, Cheng and colleagues reported that the CD80/CD86 variant T cell regulator, programmed death-1 ligand-1 (PD-L1 or B7-H1), is expressed on murine spermatocytes and spermatids, and supports the survival of intratesticular pancreatic islet allografts.985 However, contact between T cells and these cells behind the blood–testis barrier is unlikely, and complete disruption of spermatogenesis by translocation of the testis to the abdominal cavity does not appear to abrogate survival of intratesticular allografts or xenografts.266., 937., 986. Nonetheless, even if the spermatogenic cells do not play a direct role in immunoregulation, it remains possible that these cells play a role by stimulation of immunoregulatory functions of the Leydig cells, and more importantly, the Sertoli cells.
The Role of the Sertoli Cell
There is little doubt that the Sertoli cell possesses unique immunoregulatory properties. Sertoli cells from immature rat, murine or porcine testes display extended survival as allografts or xenografts, and co-transplantation of Sertoli cells or testis cell mixtures containing these cells confers increased survival on a number of cell and tissue allografts and xenografts, including neural cells, pancreatic islets and skin.987., 988., 989., 990. Such procedures have even been used for protection of islet transplants in humans, with some success and controversy.991 Although the immunoprotective properties of the Sertoli cells appear to be dependent upon their number and ability to form intercellular tight junctions and tubule-like structures, survival of co-transplanted cells adjacent to, but outside, these structures indicates that the protective mechanism does not require the creation of a physical barrier.992 Protection appears to be an inherent property of the Sertoli cell, involving factors expressed on the surface or secreted from the cell (Figure 19.17 ).
FIGURE 19.17.
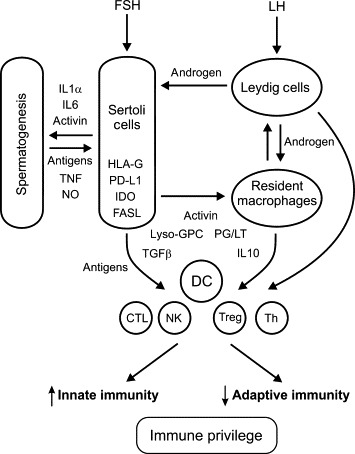
Intercompartmental interactions regulating steroidogenesis, spermatogenesis and immune responses in the adult testis.
Within the seminiferous epithelium, the Sertoli cell supports and regulates spermatogenesis, through production of cytokines, including interleukin-1α (IL1α), IL6, and activin. Production of these cytokines is stimulated by the presence of the spermatogenic cells. Spermatogenic cells also produce tumor necrosis factor (TNF) and NO, which regulate Sertoli cell functions, such as the maintenance of the blood–testis barrier. Sertoli cells possess a number of immunosuppressive activities, which include expression of indoleamine 2,3-dioxygenase (IDO), HLA-G, programmed death-1 ligand-1 (PD-L1), and FAS ligand (FASL). Spermatogenic antigens released from the seminiferous epithelium, together with local production of immunoregulatory molecules, including IL10, transforming growth factor-β (TGFβ), activin, prostaglandins/leukotrienes (PG/LT), and lyso-glycerophosphatidylcholines (lyso-GPCs), create an environment that promotes tolerogenic responses (alternatively activated resident macrophages, Treg cells) and innate immunity (NK cells, cytotoxic T cell), while inhibiting cell-mediated immunity (helper T cells). Leydig cells are responsible for recruiting macrophages into the testis and may have further immunomodulatory actions through production of androgens, and other molecules with immunoregulatory actions, such as prostaglandins (PGs). DC, dendritic cells; NK, NK cells: CTL, cytotoxic lymphocytes; Th, helper T cells; Treg, regulatory T cells.
Sertoli cells express comparatively low levels of classical MHC antigens, and produce the nonclassical MHC class I antigen, HLA-G, which would enhance their potential to avoid detection by T cells and subsequent immune activation.230., 967., 972., 993. On the other hand, expression of ICAM1 and VCAM1 by the Sertoli cells suggests that these cells can selectively bind and interact directly with circulating lymphocytes.477., 994. Mouse Sertoli cells increase expression of PD-L1, but not positive co-stimulatory molecules, in response to IFNγ in vitro.594 Human Sertoli cells were found to suppress MHC class II and inflammatory gene expression in human endothelial cell cultures stimulated with INFγ or TNF, and to inhibit the ability of these cells to stimulate spleen lymphocyte proliferation.995 Finally, Sertoli cells are similar to macrophages in that they possess an enormous capacity for phagocytosis of senescent cells, cell debris, and other potentially antigenic complexes. Together with the inherent ability of the Sertoli cell to provide a highly supportive environment for cell growth and differentiation,996 these characteristics no doubt all contribute to the unique graft-protecting abilities of the Sertoli cell.
Sertoli cells express and secrete a wide range of immunoregulatory molecules, several of which have been implicated in graft protection or immune privilege.997 Studies have shown that Sertoli cells secrete lymphocyte-inhibiting activity in culture,998., 999. and are major sites of production of both TGFβ and activin A.224., 225. Production of TGFβ1 by co-transplanted Sertoli cells has been implicated in protection of syngeneic transplants of pancreatic islets in streptozotocin-induced diabetic rats and nonobese diabetic mice.990., 1000. There is evidence that FASL expression on the Sertoli cell may enhance graft survival, as well.988., 1000., 1001. Sertoli cells produce several complement inhibitors, as well as inhibitors of granzyme B, which is a lytic molecule produced by cytotoxic lymphocytes.1002., 1003., 1004. Furthermore, IL6, which is secreted by the Sertoli cell under hormonal control, has a number of immunoregulatory properties, stimulating the production of anti-inflammatory cytokines by T cells,125 and inhibiting the maturation of dendritic cells from circulating monocytes.234 Cultured murine Sertoli cells progressively produce indoleamine 2,3-dioxygenase (IDO), an enzyme which catalyzes the metabolism of tryptophan, and which has been shown to stimulate dendritic cell-initiated tolerance,1005 and to stimulate the development of Treg cells in tumors and in pregnancy.1006., 1007. Silencing of IDO with si-mRNA inhibited the ability of these cells to reduce diabetes in non-obese diabetic mice.1006 Finally, mice with a Sertoli cell-specific deletion of the androgen receptor have a defective blood–testis barrier, the testis becomes infiltrated by macrophages, neutrophils, plasma cells, and eosinophils, and they develop antibodies against spermatogenic cells.1008 This suggests that loss of androgen regulation in the Sertoli cell leads to a loss of testicular immune privilege.
The Role of the Leydig Cell and Androgens
It seems likely that Leydig cells actually promote inflammation-related responses of the testis, as these cells produce several inflammatory mediators constitutively (MIF, NOS2) or in response to inflammatory stimuli (IL1, IL6, and CCL2).226., 276., 278., 285., 390., 391., 392., 503., 504., 641., 1009. These cells also play a central role in the regulation of interstitial fluid formation in both hCG-hyperstimulation and LPS-induced inflammation through control of endothelial cell permeability.397., 1010., 1011. Conversely, early studies established that mouse Leydig cells, but not other testicular cells, bound to lymphocytes, macrophages and eosinophils specifically in vitro,1012., 1013. and that the presence of rat or mouse Leydig cells could inhibit lymphocyte proliferation responses in vitro.982 This binding may have been due, at least in part, to a variant of VCAM1 expressed on the Leydig cell surface,1014 while inhibition of lymphocytes by Leydig cells in vitro appears to involve both direct contact and secretion of soluble inhibitory factors.1015 The Leydig cell-secreted cytokine, MIF, is a potent inhibitor of cytotoxic T cell and NK cell activity,1016., 1017., 1018. and it may be that a function of this cytokine in the testis is to restrain cell killing in the absence of an overt immunological challenge. However, the most significant role of the Leydig cells in mediating immune control of the testis may lie in their ability to facilitate recruitment of resident macrophages in the testis (Figure 19.17).69., 226., 293., 296. Junctional specializations between Leydig cells and resident macrophages mediate close physical attachment of these cells in the normal testis,240., 244. and the immunoregulatory significance of these interactions has yet to be explored in any detail.
The principal endocrine function of the Leydig cell is the production of testosterone, and concentrations of this androgen are extremely high within the testicular interstitial tissue.79., 80. There is some evidence in vitro that androgens are able to regulate lymphocyte-mediated immunity by stimulating the production of IL10 by CD4+ T cells, and stimulating the induction or proliferation of Treg cells.197., 210., 213., 214., 899. A more general effect of androgens on the Th1/Th2 cytokine balance was demonstrated by the fact that administration of testosterone to hypogonadal men stimulated baseline IL10 levels and inhibited IL1β and TNF levels in serum.1019 However, in vivo evidence for local immunoregulation by androgens in the testis is conflicting. Androgens do not appear to directly regulate macrophage or lymphocyte numbers,226., 227., 295. and manipulation of Leydig cell function and androgen production by a number of methods had no effect on survival of parathyroid or pancreatic cell allografts in the rat testis.935., 936., 986., 1020. On the other hand, treatment with exogenous testosterone suppressed the progression of autoimmune orchitis in rats, by reducing the intratesticular accumulation of macrophages and CD4+ T cells, and pro-inflammatory/Th1 cytokine expression, while increasing the number of Treg cells.899
Treatment with estrogen of the recipient rats prior to placement of parathyroid allografts was found to abrogate graft survival in the normal scrotal testis.266 The problem with interpreting this finding is that estrogen inhibits androgen production by the Leydig cell, but probably has direct effects on immune responses and graft rejection in the testis as well. Estrogens regulate dendritic cell differentiation and activity,217., 218. and overexpression of CYP19A, leading to an elevated estrogen-to-androgen ratio causes progressively severe inflammation in the mouse testis, with massive proliferation and activation of intratesticular macrophages and testicular damage.1021 Overall, it remains difficult to say what important role androgens and estrogens may play in regulating immune responses in the testis. Moreover, it should be noted that in addition to steroids, Leydig cells produce several other factors with lymphocyte regulating activity. These include the antiproliferative pro-opiomelanocortin peptides,223., 1022. and MIF, which notwithstanding its pro-inflammatory functions, inhibits the cell killing activity of cytotoxic T cells and NK cells.1016., 1017., 1018.
Soluble Immunosuppressive Activities and Bioactive Lipids
Rat testicular interstitial fluid is a potent inhibitor of T cell activation responses in vitro, in spite of the presence of substantial levels of locally-produced IL1, clearly indicating that soluble immunosuppressive factors are a principal influence on lymphocytes circulating through the interstitial tissue.965., 1023. Similar inhibitory effects on lymphocytes in vitro have been observed using whole testis extracts from mice.1024 Early investigations established that this inhibition could not be attributed to androgens,1023., 1025. or to other candidate molecules known to be present in testicular interstitial fluid, such as PGE2,1026 TGFβ family members,274., 1027. or IL10.1028
Purification of this activity established that the lymphocyte inhibition was due to the presence of several lyso-glycerophosphatidylcholines (lyso-GPCs) in the interstitial fluid.1029 These molecules are produced by the cleavage of a single fatty acid chain from plasma membrane phosphocholine-containing phospholipids via the action of PLA2, and are known to possess potent anti-inflammatory and immunoregulatory functions (Figure 19.12).1030 The actual mechanisms have proven difficult to study because lysophospholipids are cytotoxic at high concentrations.1031 It still remains to be established whether the effects of lyso-GPCs on T-cell activity at physiological concentrations are mediated by a specific receptor, a direct physical interaction with the cell membrane, or both. However, it has been demonstrated that these lyso-GPCs are specific ligands for CD1D-restricted T cells, suggesting a role for these molecules in controlling NK T-cell responses and activity in the testis.1032 Other lysophospholipds, such as lysophosphatidic acid and sphingosine-1-phosphate, for which G protein-coupled receptors are already known, have been implicated in regulating spermatogenesis and spermatogenic cell survival.1033 The production, regulation, and activity of these molecules within the testis certainly deserve further study.
Prostanoids, which are derived from arachidonic acid cleaved from plasma membrane phospholipids by the action of PLA2, are widely produced throughout the testis due to the promiscuous expression of PTGS2 and PTGS1.615 In culture, Sertoli and/or Leydig cells can produce all the classes of prostanoids: PGE2, PGF2α, PGD2, PGI2 and TxA.425., 617., 1034., 1035. In the adult rat testis, PGE2 and PGF2α predominate, with PGD2 detected at much lower levels; there is no comparable data for TxA or PGI2.1036 Direct evidence for the role of prostanoids in regulating testicular inflammation and immunity, however, is slight.
In macrophages from other tissues, PGE2 has been shown to reduce pro-inflammatory activity and promote an alternatively activated (M2) macrophage functional phenotype.1037., 1038., 1039. Studies by Kern and colleagues confirmed that the addition of the broad PTGS inhibitor, indomethacin, to cultures of rat testicular macrophages stimulated their production of IL1β, IL6 and TNF in response to LPS-stimulation and blocked their inhibitory effect on peripheral blood lymphocyte proliferation in vitro.269., 277. It was suggested that PGE2 might be responsible for the testicular macrophage phenotype that favors immune privilege. Working against this hypothesis, testicular macrophages produce little PGE2 under normal conditions, and blocking PTGS2 activity in vivo or in vitro did not affect the production of pro-inflammatory cytokines in the testis in response to LPS.276., 615., 619. However, chronic inhibition of PTGS2 did inhibit interstitial fluid formation in rat normal testes, even though this treatment ameliorated the loss of fluid that usually occurs during LPS-induced inflammation.619 These data indicate that prostanoids are involved in control of vascular tone in the normal and inflamed testis. This suggests that the use of anti-inflammatories by men with marginal fertility might need to be considered as a factor when undertaking assisted reproductive therapies.
Spermatogenic Cell and Testis Transplantation
Since the pioneering work of Brinster and colleagues,1040 the restoration of fertility in mice with congenital or chemically-induced absence of spermatogenesis, through transfer of isolated spermatogonial stem cells from fertile donors is now routine. This procedure usually involves injection of spermatogonia directly into the rete testis or efferent ducts of the recipient. Cross-species transplantation has only been fully successful in the case of rat spermatogonia to the mouse testis,1041 although spermatogonial proliferation and even partial spermatogenesis in the murine testis has been observed for a range of species, including the human.1042., 1043. Spermatogenic development of transplanted stem cells in the testis of species other than the mouse also has been achieved, including rat into rat,1044 and mouse into rat, bovine or monkey testes.1045., 1046. Successful transplantation of Sertoli cells1047., 1048. and Leydig cell stem cells1049 have been reported in mice, using similar protocols.
Studies on spermatogenic stem cell transplantation generally employ a histocompatible or immunodeficient recipient. Allogeneic transplantation has been performed in a number of domestic animals and in monkeys with intact immune systems, causing minor inflammatory complications, but with some degree of success.1050., 1051., 1052. Transplantation of rat spermatogenic stem cells into immunocompetent mice, producing epididymal spermatozoa in a small number of animals, has also been reported.1053 A characteristic feature of transplantation in immunologically intact animals appears to be an increase in peritubular macrophages and macrophage invasion into the seminiferous epithelium and tubule lumen.1054., 1055. Whether these cells may be serving an immunoprotective function, or are simply responding to an inflammatory stimulus from degenerating spermatogenic cells, is unclear. In a limited study of the immunological parameters affecting spermatogonial transplantation,33 Kanatsu-Shinohara and colleagues found that allogeneic transplantation of mouse spermatogonia was only partially successful, with arrest at the round spermatid stage in highly disorganized tubules, mononuclear cell infiltration in the interstitial tissue and eventual rejection after several months. Mature spermatozoa and successful natural mating was able to be achieved by the use of donor stem cells from immature testes and treatment of the recipient animal with anti-CD4 and anti-CD8 antibodies or the immunosuppressive agent, rapamycin, to inhibit T cell activity.
The available data suggest that spermatogonial stem cell transplantation can be performed across allogeneic and xenogeneic barriers without rejection, but that the preparation of the donor cells and recipient and the transfer procedure itself may affect the outcome. This appears comparable to the variable success experienced in studies on engraftment into the interstitial tissue. It is not yet clear what the critical factors may be, but it should be noted that the spermatogonial cells normally are injected via the rete testis or efferent ducts, and into testes that lack mature germ cells. The rete testis is the most susceptible region of the testis for initiation of autoimmune reactions.267., 268., 888. It is possible that the cytotoxic treatments used to deplete the endogenous spermatogenic cells may increase the risk of graft failure and rejection, or that the lack of endogenous spermatogenic cells may contribute to an absence or failure of tolerance to these cells, or alterations in the local immunoprotective environment. Possibly the most important factor that requires investigation is the composition of the spermatogonial cell preparation itself, and the degree to which it might be “contaminated” with resident testicular macrophages or Sertoli cells, as these have potent immunoregulatory properties.
The Definition and Significance of Testicular Immune Privilege
The original definition of immune privilege specifically applied to tissues where normal rejection responses against foreign tissue grafts, specifically allografts or xenografts, are reduced or prevented,37., 929. but the term is most commonly used to refer to a site where lymphatic drainage is deficient or access by immune cell is restricted. Sequestration of antigens of the central nervous system behind the blood–brain barrier is usually considered to be the best example of this.1056 However, these are principally functional definitions, which emerged prior to discovery of the cellular and molecular mechanisms underlying immunological tolerance and the regulation of immunity. A more contemporary definition of immune privilege, which encompasses both of these traditional concepts as well as more recent immunophysiological principles, can be stated as “the extended survival of cells expressing antigens that under normal circumstances should provoke an immune response, as well as the mechanisms that contribute to this survival”.1057
It appears that testicular immune privilege is dependent upon a number of overlapping mechanisms: (1) maintenance of central and peripheral tolerance, (2) the blood–testis barrier, (3) reduced immunogenicity of the testicular cells, (4) the anti-inflammatory phenotype of the resident macrophages, (5) immunosuppressive properties of the Sertoli cells and Leydig cells, and (6) local production of immunoregulatory/immunosuppressive cytokines, steroids and other bioactive lipids. The existence of multiple mechanisms may help to explain why orchitis and testicular autoimmunity affects only a minor subset of patients with PGA syndromes, and are not universally induced by day 3 postthymectomy in rats and mice. Moreover, based on the evidence from transplantation studies, it would appear that the ability of antigens within the testis to evade the immune system is not an absolute property of the testis or its draining lymph nodes, but is provisional upon certain criteria being met, such as the absence of testicular damage or other precipitating events. Tissues traditionally identified as being immunologically privileged, such as the testis and anterior chamber of the eye, may simply lie at one extreme of a range, as many of the mechanisms implicated in testicular immune privilege also exist to a more or lesser extent in most tissues. It might be most appropriate to think of immune privilege as simply a manifestation of a particularly effective immunoregulatory environment. The need for enhanced immunoregulation in tissues such as the testis or the eye may be related to the burden of the autoantigens present, and/or the need to prevent active inflammation causing damage to particularly susceptible tissues. While it is debatable whether autoimmunity to spermatogenic cells is more catastrophic to the organism than autoimmune gastritis, diabetes or thyroiditis, there must have been considerable evolutionary selection pressure applied to ensure that the immune system was restrained from attacking the post-meiotic germ cells the moment that they appeared during sexual maturation.
Immunology of the Epididymis and Vas Deferens
In contrast to the testis, the remainder of the male reproductive tract does not appear to be immunologically privileged, at least not in terms of extended graft survival.1058., 1059. This raises the question as to how sperm are able to continue to evade the immune system once they leave the testis environment. It is evident that the epithelial tight junctions of the epididymis and vas deferens are not as elaborate or effective as those of the blood–testis barrier.72., 86., 91., 92., 93. Moreover, leukocytes—dendritic cells, macrophages and CD8+ T cells, in particular—are commonly found within the epithelium of these tissues and appear to be able to have contact with the luminal contents.65., 96., 98., 102., 334. Nonetheless, the fact that autoantigenic sperm can survive within the epididymis for considerable periods of time without eliciting an autoimmune response indicates that effective immunoregulatory mechanisms must be in operation.101
In general, the epididymis and vas deferens appear to be significantly more susceptible to inflammation and autoimmunity than the testis. Immune cell infiltrates are much more likely to appear in the epididymis than the testis in aging mice, in mice injected with B. pertussis and adjuvant, in mice with the alymphoplasia mutation, and in mice with a deletion of the AIRE transcription factor.913., 914., 915., 923. In addition, epididymitis or epididymovasitis precedes orchitis in the day 3 thymectomized mouse model, and following transfer of lymphocytes from mice immunized against spermatogenic cell antigens.267., 874., 876. These observations suggest that the epididymis and vas deferens may be much more reliant on maintenance of tolerance to protect the sperm from immunological damage, compared with the testis.
Investigation of the incidence of sperm antibodies in men with obstructive azoospermia, congenital absence of the vas or following vasectomy indicated that lesions in the caput epididymis are associated with negligible antibody formation, whereas 66–100% of patients with obstructions in the cauda epididymis or postvasectomy had sperm antibodies (Figure 19.15).348 This clearly indicates that obstruction or damage to the epididymis and vas deferens is a potent inducer of sperm autoimmunity, but that proximity to the testis reduces the severity of the response. Expression of inflammatory and immunoregulatory molecules and the distribution of immune cells in the epididymis and vas deferens is highly regionalized. Specifically, there is a much greater number of dendritic cells, macrophages and T cells in the proximal epididymis, than in the more distal epididymis and vas deferens, and the dendritic cells appear to be more active.96., 98., 102. Recently, it has been discovered that the immunoregulatory enzyme, IDO, is highly expressed in this region of the epididymis, and pro-inflammatory cytokine expression is higher than normal in the caput epididymis of IDO-deficient mice.1060., 1061. Moreover, IDO is regulated through the SMAD2/3/4 signaling pathway that is activated by activin A,1005 and activin A levels are also highest in this region of the epididymis.789 It can be proposed that activin A may drive IDO expression in the proximal epididymis, and together these immunoregulatory proteins induce a tolerogenic program in the intraepithelial dendritic cell and T cell populations. Although this hypothesis still remains to be confirmed, this would provide an effective mechanism for promoting tolerance to sperm antigens as they emerge from the immune privileged environment of the testis. The activity of Treg cells has been shown to control the pattern and severity of autoimmune responses to vasectomy in mice.957
Within the wider mucosal immune system, there is evidence that epithelial cells themselves can present antigen to activate immunosuppressive/regulatory T cell function,188., 1062., 1063. and in this regard it is relevant that the principal cells of the epididymis are constantly absorbing large quantities of luminal fluid and its contents, including spermatozoa.1064., 1065. There is evidence that these cells also produce IL10.809., 1066. Moreover, epithelial cells of the epididymis and vas deferens express the glycosylphosphatidylinositol–anchored protein, CD52,1067 which is a co-stimulatory molecule for Treg cell development,1068 but may also play a role in protecting sperm against complement-mediated damage.1069., 1070. Consequently, removal of defective sperm from the epididymal lumen by the principal cells and presentation of their antigens to the intraepithelial lymphocytes might present an additional mechanism for controlling sperm autoimmunity in the epididymis, and possibly the vas deferens as well.
In addition to the tolerogenic mechanisms shared with the common mucosal system, there is evidence of other immunoregulatory mechanisms that may be more specific to the male reproductive tract. For example, immunosuppressive factors produced by the testis may diffuse into the epididymal fluid, and contribute to the unique immunoregulatory environment of the caput epididymis.348 Modifications of the sperm surface membrane by epididymal secretions may act to obscure sperm antigens,85 and expression of immunoregulatory molecules on the surface of the sperm itself, which include classical and nonclassical MHC antigens,968., 970. a CD4-like MHC ligand,1071., 1072. bacterial and viral TLRs,451., 802. and FASL,492 also may play a role in evading immune responses in the epididymis and vas deferens.
Immunoregulation by Seminal Plasma
Seminal plasma is profoundly immunosuppressive, as defined by the ability to inhibit various T cell and NK cell activities in vitro.1073., 1074. This immunosuppressive activity has been proposed to play a role in preventing lymphocyte responses against sperm autoantigens in the male and female reproductive tracts,856., 1075. and, more recently, to prime the female immune system to tolerate paternal antigens on the developing fetus.1076 The activity can been attributed to a number of specific and nonspecific factors, including prostasomes,1077., 1078. oxidized polyamines,1079 prostaglandins of the E series,1074., 1080. nonspecific lymphocyte-suppressing proteins,1081., 1082. and immunoregulatory cytokines.817., 818., 1083., 1084., 1085.
Prostasomes are multilaminar vesicles secreted by the normal prostate, and are a major component of human semen.1078 Pure preparations of prostasomes inhibit mitogen-induced T cell proliferation and inhibit macrophage phagocytic activity in vitro.1077 The complement inhibitors, CD46, CD55 and CD59, have been found on the surface of prostasomes.1086., 1087. Seminal plasma also contains very high concentrations of the polyamines, spermine and spermidine.1079 These polyamines are not immunosuppressive themselves, but are converted to their oxidized forms that are inhibitory of cell growth by the action of polyamine oxidase, an enzyme found in serum used in culture media.1088 Oxidized polyamines are unstable and rapidly metabolized to the cytotoxic molecules, acrolein and putrescine.1089 Prostasomes and polyamines are responsible for much of the apparent immunosuppressive activity of the ejaculate measured using lymphocyte cultures, but whether these factors have any physiological significance in terms of controlling immune responses in vivo remains speculative.1090 On the other hand, following removal of the prostasomes and inactivation of polyamine activity in human seminal plasma samples from infertility clinic patients an inverse relationship between T cell inhibitory activity and the incidence autoimmune infertility associated with sperm antibodies has been observed.1091
Human seminal plasma contains extraordinarily high concentrations of PGE2, PGE1 and their 19-hydroxylated forms.1092., 1093. Aside from their well-characterized effects on vascular permeability and smooth muscle contractility, these hormones inhibit T cell proliferation, NK cell cytotoxicity and the pro-inflammatory activities of macrophages and T cells.609., 1094. They are produced throughout the male reproductive tract: the prostaglandins in seminal plasma largely come from the seminal vesicles, although the vas deferens also may be a major source because this tissue expresses extremely high levels of COX2 under normal conditions.614., 801. Following removal of the prostasomes and inhibition of polyamine oxidation, prostaglandins appear to be responsible for most of the immunosuppressive activity of human seminal plasma.284., 818. Seminal plasma PGE2 has been implicated in the promotion of tolerogenic responses in dendritic cells and the production of Treg cells.1076., 1095.
The cytokines with immunosuppressive/immunoregulatory activity that have been positively identified in human seminal plasma are TGFβ1 and TGFβ2,1083., 1085., 1096. IL10,817., 1084., 1097. and the activins, A and B.1098., 1099. The TGFβ in seminal plasma is derived from the distal tract, comprising the seminal vesicles and/or the prostate gland.1096 IL10 is a product of monocyte/macrophages and T cells, but epithelial cells of the male tract are also a potential source of this cytokine.809., 1066. Although activin A subunit mRNA and protein is expressed in the human and mouse prostate,537., 1100. activin A levels in human seminal plasma are effectively eliminated by vasectomy, indicating a principally testicular or, more likely, epididymal origin.789., 1098. The most abundant and effective of the immunosuppressive cytokines in seminal plasma is TGFβ1, by a considerable margin.818., 1091., 1101.
A positive relationship between the level of immunosuppressive activity in human seminal plasma and the presence of sperm antibodies has been observed,818 but the actual relationship between immunosuppressive molecules in seminal plasma and immunological infertility in men remains to be established. It also should be borne in mind that these molecules have direct effects on the epithelia, stroma and leukocytes of the female genital tract.1101., 1102. Animal studies show an intense but transient inflammatory response in the endometrium at mating stimulated by seminal plasma and one consequence of this inflammatory response is the induction of a state of hyporesponsiveness to paternal MHC class I antigens.1103 Insemination is causally linked to the activation and expansion of populations of lymphocytes mediating active immune tolerance at the embryo implantation site, specifically Treg cells, a process in which seminal plasma TGFβ1 and PGE2 have been particularly implicated.857., 1101., 1103. Semen may therefore play a critical role in providing the antigenic and environmental signals necessary to initiate an appropriate maternal immune response to the conceptus during pregnancy.
Other immunosuppressive/immunoregulatory molecules that are neither prostaglandins nor cytokines, have been detected in seminal plasma or accessory gland secretions from various species.1081., 1082. Some of these molecules have been identified, including the protein clusterin and a prostatic steroid binding protein in rats,1104., 1105. IgA,816 and soluble HLA-G.974 The actual significance of these molecules in immune regulation and autoimmune fertility requires further functional characterization.
Innate Immunity in the Male Reproductive Tract: The Response to Infections and Tumors
If the male reproductive tract, and the testis in particular, is a site of reduced antigen-specific immune responses, then the question must be asked: How does the genital tract avoid recurrent infections or the development of tumors? Interest in the question has been stimulated by the fact that the male reproductive tract is a major site of transmission of human immunodeficiency virus (HIV),1106 and by the observation that relapsing lymphoblastic leukemia in the testis following treatment is a frequent problem in male patients.1107 While the progression of HIV in the male reproductive tract is very poorly defined, destruction of the spermatogenic cells is a characteristic feature of HIV infection in men.1108 The testis is also suspected to be a sanctuary where HIV may be able to take refuge during therapy, possibly behind the blood–testis barrier, thereby subverting therapeutic effectiveness.1109., 1110. Studies by Jahnukainen and colleagues in rats have suggested that testicular relapse of leukemia may be due to the unique immunoregulatory environment of the testis and, specifically, the ability of Leydig cells to bind lymphocytes.1111 Moreover, it appears that the immunological protection in the testis is extended not just to spermatogenic cell antigens and graft antigens, but also to tumor-specific antigens that would induce immune reactions elsewhere in the body.1112
As a result of these and other observations, it has been suggested that virally or tumorigenically transformed cells might be able to evade both the immune system and cytotoxic drugs by ‘hiding-out’ in the testis. The reality is, however, that infection of the testis is relatively rare compared with the remainder of the genital tract,860 and testicular tumors are no more frequent than tumors in other parts of the body.1113 Moreover, when they do occur, testicular tumors are accompanied by the expected mononuclear cell infiltrates, which are related to the size, progression and type of the tumor.312., 316. The effective deficiency in adaptive immune responses within the testis, therefore, implies that innate (pathogen-specific) immunity might have increased importance for dealing with tumors and infected cells at this site. Hypothetically, relying principally on mechanisms of innate immunity to provide protection against infections and tumors reduces the risk that inflammation will lead to antigen-specific immune responses and subsequent autoimmune damage to fertility. There is some justification for this assumption, based on studies of the male reproductive tract and from analogy with the rest of the common mucosal system. The mechanisms responsible broadly fall into cell-mediated responses and secreted molecules.
Cellular Responses
Although the environment of the male reproductive tract tends toward suppression of antigen-specific immunity and maintaining tolerance, these tissues are densely populated by effector cells involved in innate immunity, specifically macrophages and lymphocytes expressing CD8, which is expressed by both cytotoxic T cells and NK cells.65., 66., 70., 96., 98., 227., 334., 354., 357. Significantly, NK cells are able to recognize and destroy infected or transformed cells without prior immunization, and so can act independently of the adaptive immune response. In addition to the testicular macrophages, both the Sertoli cell and Leydig cell are able to recognize and respond to bacterial pathogens directly,376., 386., 387., 390., 392., 994. through expression of specific TLRs on their surface. The importance of the TLRs in protecting against infection in the male reproductive tract is indicated by the fact that pathogens can increase their virulence by subverting the function of these molecules, for example, in the case of uropathogenic strains of E. coli.275., 436. Suppression of TLR expression throughout the male reproductive tract also appears to be a strategy utilized by HIV to evade immunity.784
All mucosa protect themselves through a number of nonimmunological mechanisms that support the integrity of the epithelial barrier. These include: tight junctions that restrict passage between cells, secretion of mucins and other antigenically inert molecules, control of local pH, recruitment of phagocytes (macrophages and neutrophils) to the epithelium, and production of lysozyme, ROS, proteases, complement components and antimicrobial cytokines, and peptides.162., 182., 190., 1063. It could be productive to investigate the role of these processes in resisting infection in the male reproductive tract.
Secreted Factors
Immunoglobulins
Immunoglobulin A is found in the fluid of all tissues of the male reproductive tract, but as vasectomy does not substantially alter IgA levels in semen, it appears that most comes from the urethra and accessory glands; more specifically, from the prostate gland.1114., 1115., 1116. Plasma cells secreting IgA have been found in the urethral gland,354., 1117. but most IgA in the reproductive tract is believed to be derived initially from the circulation.190., 355. Production of secretory component by the prostatic epithelium, which is required for transport of IgA across the mucosa, is androgen regulated.1116 In men, bacterial infection of the reproductive tract is associated with a large increase in secretion of IgA in prostatic fluid.1114 This anti-inflammatory immunoglobulin presumably plays an important first line of defense against infection in the male reproductive tract.
Interferons
These molecules possess a range of immunoregulatory and antiviral actions, including suppression of viral proliferation, cytotoxicity and, in the case of the type 2 interferon, IFNγ, stimulation of MHC class I and class II expression.115., 1118. The viral pathogen receptor, TLR3, which acts via the TICAM1 adaptor protein to induce production of type 1 interferons, is expressed by testicular macrophages, Sertoli cells, Leydig cells, peritubular cells and spermatogenic cells, and by epithelial cells of the epididymis and vas deferens.438., 441., 442., 450., 451. In the testis, activation of TLR3 increases interferon production and interferon-inducible genes involved in viral protection in Sertoli cells, Leydig cells, and even spermatogenic cells.438., 439., 440., 450., 451., 597. Interferon production may also be induced through activation of TLR4. Testicular macrophages produce type 1 interferons constitutively, but viral infections upregulate their expression in both macrophages and somatic cells of the testis.591., 592. The strongest interferon response to infection occurs in the Leydig and Sertoli cells, but only the Leydig cells appear to express IFNγ under such conditions.
Defensins
Defensins are small (3–4 kDa) positively-charged antimicrobial peptides, which are able to disrupt bacteria, fungi, parasites, and some enveloped viruses by forming multimeric pores in the cell membranes of these pathogens.1119 The defensins belong to one of two families, the α- and β-defensins (DEFA and DEFB).1120 The β-defensins are produced by most mucosal epithelial tissues, but appear to be preferentially expressed in the urogenital tract, particularly in the testis and epididymis in the male, and their production is stimulated by TLR ligands and cytokines.1121., 1122. Genes encoding these peptides are expressed in discrete clusters along the length of the male reproductive tract,794., 1123. and a number of epididymal-specific β-defensins have been identified in the mouse and the rat.1124., 1125., 1126. Several of the β-defensins appear to be required for normal sperm maturation or for maintaining sperm motility in the epididymis, so that changes in their expression during infection also may contribute to sperm dysfunction.808., 1124., 1127.
Implications, Applications, and the Next 10 Years
This review has concentrated almost exclusively upon inflammation and immunity in the male reproductive tract as a means to understanding immune-based infertility and the impact of infection and inflammation on male reproduction. However, these processes have broader implications relevant to other areas of research and, in this last section, applications to clinical issues of contraceptive development, transplantation medicine and chronic pain are briefly highlighted.
Immunocontraception
Conceptually, immunocontraception involving a vaccine that targets sperm antigens or reproductive hormones has the advantages of a potentially high degree of specificity and convincing proof-of-principle from the many patients with pre-existing autoimmune infertility. Although vaccines against gonadotropins actually have been trialed,1128 it is unlikely that such an approach would be readily applicable to contraception in men. Other contraceptive vaccine studies have concentrated on antigens involved in critical sperm functions.1129., 1130. Such approaches raise concerns related to safety, efficacy and reversibility, but progress has been impeded largely by the issue of variability of immune responses after vaccination.1131 Genetic differences in immune response genes in the human population and the complexity of the immune system itself might make it difficult to develop a single vaccine that works effectively enough in all men to compete with currently available hormone-based approaches to contraception. Another concern would be the risk of inducing more widespread autoimmune disease, since there is an established relationship between sperm autoimmunity and autoimmunity in general.841., 842. Furthermore, it may be difficult to develop a contraceptive vaccine that is entirely reversible. Nonetheless, the targeting of critical reproductive antigens to control fertility is a very attractive idea, which will most likely continue to engage reproductive immunologists. Most benefit from such studies may come from the identification and characterization of molecules with crucial functions in fertility which might be used as molecular targets for other drug agents that are not nonimmunologically-based.
Transplantation Medicine and Immunotherapy
One of the most obvious potential benefits from understanding the principles of immunoregulation as they pertain to the male reproductive tract is the possibility that these mechanisms might be applicable to controlling immunity at other sites. If there are immunoregulatory mechanisms that are more or less unique to the testis, for example, where extended graft survival has already been shown to be feasible, then elucidation of these mechanisms has the potential to lead to entirely novel methods of reducing graft rejection in general. The current methods used in transplantation medicine involve broad spectrum immunosuppressives, principally cyclic peptides of fungal origin that interfere with T cell signaling and prevent proliferation of these immune cells in a nonselective manner.1132., 1133. These drugs have considerable harmful side-effects, such as generalized immune suppression, nephrotoxicity and inherent tumorigenicity,1134., 1135. and require close monitoring and adjustment for the remainder of life. They also have damaging effects on testicular steroidogenesis and spermatogenesis.1136., 1137. Real alternatives or adjuncts to these agents, based on more physiological regulatory principles, would definitely be desirable. In particular, the unique immunological properties of the Sertoli cell raise hopes of an exciting new therapeutic opportunity in cell-based gene therapy.989., 1138. Such applications could include, not only the use of ectopic Sertoli cells to support pancreatic islet engraftment, for example, but also the employment of genetically-engineered Sertoli cell lines for delivery of therapeutic proteins. Time will tell if such a cell-based gene therapy approach is clinically viable.
Sterile Inflammation and Chronic Pain
An important concept that has arisen from studies on inflammatory pathways and immunity in normal male reproduction is the realization that dysregulation of these processes may be a cause of sterile, antibiotic-resistant inflammation and chronic pain. Chronic pelvic and perineal pain is normally associated with epididymo-orchitis, vasitis and/or prostatitis.10., 11., 12., 860., 910., 912. These conditions are usually due to infection or physical damage, but the ongoing production of endogenous ligands and danger signals, autoimmunity or alterations in regulatory signaling may cause inflammation to persist long after the infection has cleared or the trauma has been repaired.10., 12., 1139. Prostatic hyperplasia and tumors also may be a cause, and inflammation may arise spontaneously in a susceptible subpopulation of patients. These chronic inflammatory pain conditions are poorly understood, and frequently difficult to treat.12 Treatment alternatives are limited to standard anti-inflammatory and pain-alleviating measures, and the actual mechanisms involved are not well-characterized. Several prostanoids, including PGE2 and PGI2, have pain-inducing properties,1140 and there is an association between chronic pelvic pain and production of the pro-inflammatory cytokines, IL1β, TNF, IL8 and IFNγ, in particular,1139., 1141. but research in this area is still in need of a rational hypothetical background. Given that these conditions are so poorly understood, extremely debilitating and frequently untreatable, they provide a real opportunity for effective future investigations.
Conclusion
In summary, the need for a close functional relationship between the male reproductive and immune systems is self-evident. Spermatogenic cells are susceptible to immunological responses and activation of immunity against sperm or other elements of the reproductive tract can lead to androgen insufficiency, infertility or chronic inflammation. Inflammation and immune activation directly inhibit the hypothalamic-pituitary-Leydig cell axis at several levels, interfere with essential interactions between the Sertoli cells and spermatogenic cells and increase the potential for sperm antibody formation, a major cause of infertility in men. Nonetheless, spermatogenic cells normally are ignored by the immune system, as are grafts of foreign tissue placed within the testicular capsule. Traditional explanations for the protection of these cells, based on ‘immune privilege’ of the testis maintained by the blood–testis barrier or by exclusion of immune cells, are not consistent with either the histological organization of the reproductive tract or modern concepts of immunoregulation. A more realistic understanding of the control of immune responses in the male reproductive tract encompasses the activity of immunoregulatory macrophages and lymphocytes and peripheral tolerance, as well as active suppression of antigen-specific immunity by somatic cells involving regulatory cytokines, androgenic steroids and other anti-inflammatory and immunosuppressive factors. Dysregulation of this normal environment caused by infection, local or systemic inflammation, toxic insult, active immunization or deletion of regulatory T cells may activate the circulating immune cells, leading to a range of effects from temporary disturbance of spermatogenesis and steroidogenesis, all the way through to the creation of testis-reactive T cells and autoimmunity. Equally important for maintaining fertility, it appears that the restraints on antigen-specific immunity in the male reproductive tract are counterbalanced by enhanced local innate immune mechanisms and conventional mucosal immunity. In fact, it is increasingly evident that inflammatory signaling pathways are fundamental to the control of normal functions of the seminiferous epithelium, and possibly other functions throughout the male reproductive tract. Studies also suggest that establishment of male reproductive function is linked to the normal development of the local immune environment.
This review is intended to establish the concept that normal male reproductive function and the response to disease represent different facets of the same regulatory environment, involving complex interactions between somatic cells, resident immune cells and the circulating cellular elements of the immune system. The use of the term immunophysiology is intended to reflect this conflation of immunology and organ function. Unraveling these processes has obvious importance for issues related to male reproduction, such as autoimmune infertility, germ cell and testis transplantation, reproductive tract infection and chronic inflammatory pain. There is still a long way to go in order to understand these interactions completely, and how they impact upon health and physiology. Many unanswered questions remain, including:
-
•
What are the local factors that regulate recruitment and functional modulation of macrophages and dendritic cells in the testis and downstream reproductive tract?
-
•
What are the specific roles of T cell subsets in protecting intratesticular and sperm antigens?
-
•
What is the relative contribution of local immunosuppressive mechanisms, such as the lyso-GPCs, toward this protection?
-
•
What are the molecular details of inflammatory signaling in the seminiferous epithelium, and how do these impact upon reproductive health?
-
•
What is the real clinical significance of leukocytes, cytokines and antibodies in semen?
-
•
What are the mechanisms that regulate passage through the various testicular compartments, including the testicular capillaries and blood–testis barrier?
It is certain that the dramatic increase in interest, awareness and research activity in this area of male reproductive biology clearly means that there will be considerable new discoveries in the near future, with exciting and perhaps even totally unexpected implications and benefits. Indeed, such studies are crucial, in light of the increasing incidence of idiopathic male reproductive disorders,1142 established and emerging infections with reproductive tract involvement (severe acute respiratory syndrome, resistant strains of tuberculosis),863 and the resurgence of preventable diseases, such as mumps orchitis,858 that threaten male fertility and reproductive health.
References
- 1.Guerau-de-Arellano M., Martinic M., Benoist C., Mathis D. Neonatal tolerance revisited: a perinatal window for Aire control of autoimmunity. J Exp Med. 2009;206:1245–1252. doi: 10.1084/jem.20090300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Baker H.W., Clarke G.N., Hudson B., McBain J.C., McGowan M.P., Pepperell R.J. Treatment of sperm autoimmunity in men. Clin Reprod Fertil. 1983;2:55–71. [PubMed] [Google Scholar]
- 3.Lenzi A., Gandini L., Lombardo F., Rago R., Paoli D., Dondero F. Antisperm antibody detection: 2. Clinical, biological, and statistical correlation between methods. Am J Reprod Immunol. 1997;38:224–230. doi: 10.1111/j.1600-0897.1997.tb00303.x. [DOI] [PubMed] [Google Scholar]
- 4.Schuppe H.C., Meinhardt A., Allam J.P., Bergmann M., Weidner W., Haidl G. Chronic orchitis: a neglected cause of male infertility? Andrologia. 2008;40:84–91. doi: 10.1111/j.1439-0272.2008.00837.x. [DOI] [PubMed] [Google Scholar]
- 5.Suominen J., Soderstrom K.O. Lymphocyte infiltration in human testicular biopsies. Int J Androl. 1982;5:461–466. doi: 10.1111/j.1365-2605.1982.tb00277.x. [DOI] [PubMed] [Google Scholar]
- 6.Toh B.H., Sentry J.W., Alderuccio F. The causative H+/K+ ATPase antigen in the pathogenesis of autoimmune gastritis. Immunol Today. 2000;21:348–354. doi: 10.1016/s0167-5699(00)01653-4. [DOI] [PubMed] [Google Scholar]
- 7.Bohring C., Krause E., Habermann B., Krause W. Isolation and identification of sperm membrane antigens recognized by antisperm antibodies, and their possible role in immunological infertility disease. Mol Hum Reprod. 2001;7:113–118. doi: 10.1093/molehr/7.2.113. [DOI] [PubMed] [Google Scholar]
- 8.Shetty J., Naaby-Hansen S., Shibahara H., Bronson R., Flickinger C.J., Herr J.C. Human sperm proteome: immunodominant sperm surface antigens identified with sera from infertile men and women. Biol Reprod. 1999;61:61–69. doi: 10.1095/biolreprod61.1.61. [DOI] [PubMed] [Google Scholar]
- 9.Aitken R.J., Hulme M.J., Henderson C.J., Hargreave T.B., Ross A. Analysis of the surface labelling characteristics of human spermatozoa and the interaction with anti-sperm antibodies. J Reprod Fertil. 1987;80:473–485. doi: 10.1530/jrf.0.0800473. [DOI] [PubMed] [Google Scholar]
- 10.Rivero V.E., Motrich R.D., Maccioni M., Riera C.M. Autoimmune etiology in chronic prostatitis syndrome: an advance in the understanding of this pathology. Crit Rev Immunol. 2007;27:33–46. doi: 10.1615/critrevimmunol.v27.i1.30. [DOI] [PubMed] [Google Scholar]
- 11.Chan P.T., Schlegel P.N. Inflammatory conditions of the male excurrent ductal system. Part I. J Androl. 2002;23:453–460. [PubMed] [Google Scholar]
- 12.Strebel R.T., Schmidt C., Beatrice J., Sulser T. Chronic scrotal pain syndrome (CSPS): the widespread use of antibiotics is not justified. Andrology. 2013;1:155–159. doi: 10.1111/j.2047-2927.2012.00017.x. [DOI] [PubMed] [Google Scholar]
- 13.Head J.R., Neaves W.B., Billingham R.E. Reconsideration of the lymphatic drainage of the rat testis. Transplantation. 1983;35:91–95. doi: 10.1097/00007890-198301000-00017. [DOI] [PubMed] [Google Scholar]
- 14.Head J.R., Neaves W.B., Billingham R.E. Immune privilege in the testis. I. Basic parameters of allograft survival. Transplantation. 1983;36:423–431. doi: 10.1097/00007890-198310000-00014. [DOI] [PubMed] [Google Scholar]
- 15.Lie P.P.Y., Cheng C.Y., Mruk D.D. Signalling pathways regulating the blood-testis barrier. Int J Biochem Cell Biol. 2013;45:621–625. doi: 10.1016/j.biocel.2012.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dym M., Fawcett D.W. The blood-testis barrier in the rat and the physiological compartmentation of the seminiferous epithelium. Biol Reprod. 1970;3:308–326. doi: 10.1093/biolreprod/3.3.308. [DOI] [PubMed] [Google Scholar]
- 17.Setchell B.P., Voglmayr J.K., Waites G.M. A blood-testis barrier restricting passage from blood into rete testis fluid but not into lymph. J Physiol. 1969;200:73–85. doi: 10.1113/jphysiol.1969.sp008682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Russell L. Movement of spermatocytes from the basal to the adluminal compartment of the rat testis. Am J Anat. 1977;148:313–328. doi: 10.1002/aja.1001480303. [DOI] [PubMed] [Google Scholar]
- 19.Meinhardt A., Hedger M.P. Immunological, paracrine and endocrine aspects of testicular immune privilege. Mol Cell Endocrinol. 2011;335:60–68. doi: 10.1016/j.mce.2010.03.022. [DOI] [PubMed] [Google Scholar]
- 20.Andrade-Rocha F.T. Temporary impairment of semen quality following recent acute fever. Ann Clin Lab Sci. 2013;43:94–97. [PubMed] [Google Scholar]
- 21.Adamopoulos D.A., Lawrence D.M., Vassilopoulos P., Contoyiannis P.A., Swyer G.I. Pituitary–testicular interrelationships in mumps orchitis and other viral infections. Br Med J. 1978;1:1177–1180. doi: 10.1136/bmj.1.6121.1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Spiess A.N., Feig C., Schulze W. Cross-platform gene expression signature of human spermatogenic failure reveals inflammatory-like response. Hum Reprod. 2007;22:2936–2946. doi: 10.1093/humrep/dem292. [DOI] [PubMed] [Google Scholar]
- 23.Lardenois A., Chalmel F., Barrionuevo F., Demougin P., Scherer G., Primig M. Profiling spermatogenic failure in adult testes bearing Sox9-deficient Sertoli cells identifies genes involved in feminization, inflammation and stress. Reprod Biol Endocrinol. 2010;8:154. doi: 10.1186/1477-7827-8-154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hedger M.P. Toll-like receptors and signalling in spermatogenesis and testicular responses to inflammation – a perspective. J Reprod Immunol. 2011;88:130–141. doi: 10.1016/j.jri.2011.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Setchell B.P. The testis and tissue transplantation: historical aspects. J Reprod Immunol. 1990;18:1–8. doi: 10.1016/0165-0378(90)90020-7. [DOI] [PubMed] [Google Scholar]
- 26.Berthold A.A. Über die Transplantation der Hoden. Arch Anat Physiol Wiss Med. 1849;16:42–46. [Google Scholar]
- 27.Sand K. Experiments on the internal secretion of the sexual glands, especially on experimental hermaphroditism. J Physiol. 1919;53:257–263. doi: 10.1113/jphysiol.1919.sp001875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Steinach E. Geschelchtstrieb und echtsekundäre Geschelchtsmerkmale als folgender innersekretorischen Funktion der Keimdrüsen. Zentralbl Physiol. 1923;24:551–566. [Google Scholar]
- 29.Cevelotto G. Über Verpflanzungen und Gefuerungen der Hoden. Frankf Z Pathol. 1909;3:331–337. [Google Scholar]
- 30.Barten E.J., Newling D.W. Transplantation of the testis; from the past to the present. Int J Androl. 1996;19:205–211. doi: 10.1111/j.1365-2605.1996.tb00464.x. [DOI] [PubMed] [Google Scholar]
- 31.Sengoopta C. ‘Dr Steinach coming to make old young!’: sex glands, vasectomy and the quest for rejuvenation in the roaring twenties. Endeavour. 2003;27:122–126. doi: 10.1016/s0160-9327(03)00102-9. [DOI] [PubMed] [Google Scholar]
- 32.Moore C.R. On the properties of the gonads as controllers of somatic and physical characteristics. IX. Testis graft reactions in different environments (rat) Am J Anat. 1926;37:351–416. [Google Scholar]
- 33.Kanatsu-Shinohara M., Ogonuki N., Inoue K. Allogeneic offspring produced by male germ line stem cell transplantation into infertile mouse testis. Biol Reprod. 2003;68:167–173. doi: 10.1095/biolreprod.102.008516. [DOI] [PubMed] [Google Scholar]
- 34.Leckband E. Testicular homografts in tolerant mice. Transplantation. 1964;2:522–533. doi: 10.1097/00007890-196407000-00010. [DOI] [PubMed] [Google Scholar]
- 35.Turner C.D. Intra-ocular homotransplantation of prepuberal testes in the rat. Am J Anat. 1938;63:101–159. [Google Scholar]
- 36.Dameron J.T. The anterior chamber of the eye for investigative purposes. A site for transplantation of fetal endocrine tissues and cancer, and for the study of tissue reaction. Surgery. 1951;30:787–799. [PubMed] [Google Scholar]
- 37.Barker C.F., Billingham R.E. Immunologically privileged sites. Adv Immunol. 1977;25:1–54. [PubMed] [Google Scholar]
- 38.Lansteiner K. Zur Kenntnis der spezifisch auf Blutkörperchen wirkenden Sera. Zentralbl Bacteriol. 1899;25:546–549. [Google Scholar]
- 39.Metchinikoff E.L. Recherches sur l’influence de l’organisme sur les toxines. Sur la spermotoxine et l’antispermotoxine. Ann Inst Pasteur. 1900;14:1–12. [Google Scholar]
- 40.Guyer M.F. Studies of cytolysins. III. Experiments with spermatotoxins. J Exp Zool. 1922;35:207–223. [Google Scholar]
- 41.Kennedy W.P. The production of spermatoxins. Q J Exp Physiol. 1924;14:279–283. [Google Scholar]
- 42.Voisin G., Delaunay A., Barber M. Sur les lésions testiculaires provoqueés chez le cobaye par iso- et autosensibilisation. Ann Inst Pasteur. 1951;81:48–63. [PubMed] [Google Scholar]
- 43.Rümke P. The presence of sperm antibodies in the serum of two patients with oligospermia. Vox Sang. 1954;4:135–140. [Google Scholar]
- 44.Wilson L. Sperm agglutinins in human semen, blood. Proc Soc Exp Biol Med. 1954;85:652–655. doi: 10.3181/00379727-85-20982. [DOI] [PubMed] [Google Scholar]
- 45.Stedronska J., Hendry W.F. The value of the mixed antiglobulin reaction (MAR test) as an addition to routine seminal analysis in the evaluation of the subfertile couple. Am J Reprod Immunol. 1983;3:89–91. doi: 10.1111/j.1600-0897.1983.tb00221.x. [DOI] [PubMed] [Google Scholar]
- 46.Michaels R.M., Rogers K.D. A sex difference in immunologic responsiveness. Pediatrics. 1971;47:120–123. [PubMed] [Google Scholar]
- 47.Inman R.D. Immunologic sex differences and the female predominance in systemic lupus erythematosus. Arthritis Rheum. 1978;21:849–852. doi: 10.1002/art.1780210718. [DOI] [PubMed] [Google Scholar]
- 48.Calzolari A. Recherches experimentales sur un rapport probable entre la fonction du thymus et celle des testicules. Arch Ital Biol. 1898;30:71–77. [Google Scholar]
- 49.Chiodi H. The relationship between the thymus and the sexual organs. Endocrinology. 1940;26:107–116. [Google Scholar]
- 50.Castro J.E., Hamilton D.N. Adrenalectomy and orchidectomy as immunopotentiating procedures. Transplantation. 1972;13:615–616. [PubMed] [Google Scholar]
- 51.Kerr J.B. The cytology of the human testis. In: Burger H., de Kretser D.M., editors. The testis. 2nd ed. Raven Press; New York: 1989. pp. 197–229. [Google Scholar]
- 52.Fawcett D.W., Neaves W.B., Flores M.N. Comparative observations on intertubular lymphatics and the organization of the interstitial tissue of the mammalian testis. Biol Reprod. 1973;9:500–532. doi: 10.1093/biolreprod/9.5.500. [DOI] [PubMed] [Google Scholar]
- 53.Ergün S., Davidoff M., Holstein A.F. Capillaries in the lamina propria of human seminiferous tubules are partly fenestrated. Cell Tissue Res. 1996;286:93–102. doi: 10.1007/s004410050678. [DOI] [PubMed] [Google Scholar]
- 54.Turner T.T., Rhoades C.P. Testicular capillary permeability: the movement of luteinizing hormone from the vascular to the interstitial compartment. J Androl. 1995;16:417–423. [PubMed] [Google Scholar]
- 55.Maddocks S., Setchell B.P. The composition of extracellular interstitial fluid collected with a push-pull cannula from the testes of adult rats. J Physiol. 1988;407:363–372. doi: 10.1113/jphysiol.1988.sp017420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hees H., Leiser R., Kohler T., Wrobel K.H. Vascular morphology of the bovine spermatic cord and testis. I. Light- and scanning electron-microscopic studies on the testicular artery and pampiniform plexus. Cell Tissue Res. 1984;237:31–38. doi: 10.1007/BF00229196. [DOI] [PubMed] [Google Scholar]
- 57.Setchell B.P. The flow and composition of lymph from the testes of pigs with some observations on the effect of raised venous pressure. Comp Biochem Physiol A. 1982;73:201–205. doi: 10.1016/0300-9629(82)90056-1. [DOI] [PubMed] [Google Scholar]
- 58.Moller R. Arrangement and fine structure of lymphatic vessels in the human spermatic cord. Andrologia. 1980;12:564–576. doi: 10.1111/j.1439-0272.1980.tb01353.x. [DOI] [PubMed] [Google Scholar]
- 59.Itoh M., Li X.Q., Yano A., Xie Q., Takeuchi Y. Patterns of efferent lymphatics of the mouse testis. J Androl. 1998;19:466–472. [PubMed] [Google Scholar]
- 60.Christensen A.K., Gillim S.W. The correlation of fine structure and function in steroid-secreting cells, with emphasis on those of the gonad. In: McKearns K.W., editor. The gonads. Appleton-Century-Crofts; New York: 1969. pp. 415–488. [Google Scholar]
- 61.Hedger M.P., de Kretser D.M. Leydig cell function and its regulation. Results Probl Cell Differ. 2000;28:69–110. doi: 10.1007/978-3-540-48461-5_4. [DOI] [PubMed] [Google Scholar]
- 62.Anton F., Morales C., Aguilar R., Bellido C., Aguilar E., Gaytan F. A comparative study of mast cells and eosinophil leukocytes in the mammalian testis. Zentralbl Veterinarmed A. 1998;45:209–218. doi: 10.1111/j.1439-0442.1998.tb00819.x. [DOI] [PubMed] [Google Scholar]
- 63.Hutson J.C. Testicular macrophages. Int Rev Cytol. 1994;149:99–143. doi: 10.1016/s0074-7696(08)62087-2. [DOI] [PubMed] [Google Scholar]
- 64.Dym M., Romrell L.J. Intraepithelial lymphocytes in the male reproductive tract of rats and rhesus monkeys. J Reprod Fertil. 1975;42:1–7. doi: 10.1530/jrf.0.0420001. [DOI] [PubMed] [Google Scholar]
- 65.Ritchie A.W., Hargreave T.B., James K., Chisholm G.D. Intra-epithelial lymphocytes in the normal epididymis. A mechanism for tolerance to sperm auto-antigens? Br J Urol. 1984;56:79–83. doi: 10.1111/j.1464-410x.1984.tb07169.x. [DOI] [PubMed] [Google Scholar]
- 66.el-Demiry M.I., Hargreave T.B., Busuttil A., James K., Ritchie A.W., Chisholm G.D. Lymphocyte sub-populations in the male genital tract. Br J Urol. 1985;57:769–774. doi: 10.1111/j.1464-410x.1985.tb07051.x. [DOI] [PubMed] [Google Scholar]
- 67.Pöllänen P., Niemi M. Immunohistochemical identification of macrophages, lymphoid cells and HLA antigens in the human testis. Int J Androl. 1987;10:37–42. doi: 10.1111/j.1365-2605.1987.tb00163.x. [DOI] [PubMed] [Google Scholar]
- 68.Pöllänen P., Maddocks S. Macrophages, lymphocytes and MHC II antigen in the ram and the rat testis. J Reprod Fertil. 1988;82:437–445. doi: 10.1530/jrf.0.0820437. [DOI] [PubMed] [Google Scholar]
- 69.Wang J., Wreford N.G., Lan H.Y., Atkins R., Hedger M.P. Leukocyte populations of the adult rat testis following removal of the Leydig cells by treatment with ethane dimethane sulfonate and subcutaneous testosterone implants. Biol Reprod. 1994;51:551–561. doi: 10.1095/biolreprod51.3.551. [DOI] [PubMed] [Google Scholar]
- 70.Tompkins A.B., Hutchinson P., de Kretser D.M., Hedger M.P. Characterization of lymphocytes in the adult rat testis by flow cytometry: effects of activin and transforming growth factor β on lymphocyte subsets in vitro. Biol Reprod. 1998;58:943–951. doi: 10.1095/biolreprod58.4.943. [DOI] [PubMed] [Google Scholar]
- 71.Hermo L., Lalli M. Monocytes and mast cells in the limiting membrane of human seminiferous tubules. Biol Reprod. 1978;19:92–100. doi: 10.1095/biolreprod19.1.92. [DOI] [PubMed] [Google Scholar]
- 72.Pelletier R.M. The tight junctions in the testis, epididymis and vas deferens. In: Cereijido M., Anderson J., editors. Tight junctions. 2nd ed. CRC Press; Boca Raton: 2001. pp. 599–628. [Google Scholar]
- 73.Koskimies A.I., Kormano M. The proteins in fluids from the seminiferous tubules and rete testis of the rat. J Reprod Fertil. 1973;34:433–434. doi: 10.1530/jrf.0.0340433. [DOI] [PubMed] [Google Scholar]
- 74.Belchetz P.E., Plant T.M., Nakai Y., Keogh E.J., Knobil E. Hypophysial responses to continuous and intermittent delivery of hypothalamic gonadotropin-releasing hormone. Science. 1978;202:631–633. doi: 10.1126/science.100883. [DOI] [PubMed] [Google Scholar]
- 75.Clark B.J., Wells J., King S.R., Stocco D.M. The purification, cloning, and expression of a novel luteinizing hormone-induced mitochondrial protein in MA-10 mouse Leydig tumor cells. Characterization of the steroidogenic acute regulatory protein (StAR) J Biol Chem. 1994;269:28314–28322. [PubMed] [Google Scholar]
- 76.Payne A.H., Hales D.B. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr Rev. 2004;25:947–970. doi: 10.1210/er.2003-0030. [DOI] [PubMed] [Google Scholar]
- 77.Jégou B. The Sertoli cell. In: de Kretser D.M., editor. The testis. Baillière Tindall; London: 1992. pp. 273–311. [Google Scholar]
- 78.Plant T.M., Winters S.J., Attardi B.J., Majumdar S.S. The follicle stimulating hormone-inhibin feedback loop in male primates. Hum Reprod. 1993;8(Suppl. 2):41–44. doi: 10.1093/humrep/8.suppl_2.41. [DOI] [PubMed] [Google Scholar]
- 79.Sharpe R.M., Donachie K., Cooper I. Re-evaluation of the intratesticular level of testosterone required for quantitative maintenance of spermatogenesis in the rat. J Endocrinol. 1988;117:19–26. doi: 10.1677/joe.0.1170019. [DOI] [PubMed] [Google Scholar]
- 80.Zirkin B.R., Santulli R., Awoniyi C.A., Ewing L.L. Maintenance of advanced spermatogenic cells in the adult rat testis: quantitative relationship to testosterone concentration within the testis. Endocrinology. 1989;124:3043–3049. doi: 10.1210/endo-124-6-3043. [DOI] [PubMed] [Google Scholar]
- 81.Mooradian A.D., Morley J.E., Korenman S.G. Biological actions of androgens. Endocr Rev. 1987;8:1–28. doi: 10.1210/edrv-8-1-1. [DOI] [PubMed] [Google Scholar]
- 82.Turner T.T., Bang H.J., Lysiak J.J. Experimental testicular torsion: reperfusion blood flow and subsequent testicular venous plasma testosterone concentrations. Urology. 2005;65:390–394. doi: 10.1016/j.urology.2004.09.033. [DOI] [PubMed] [Google Scholar]
- 83.Hess R.A., Fernandes S.A., Gomes G.R., Oliveira C.A., Lazari M.F., Porto C.S. Estrogen and its receptors in efferent ductules and epididymis. J Androl. 2011;32:600–613. doi: 10.2164/jandrol.110.012872. [DOI] [PubMed] [Google Scholar]
- 84.Clulow J., Jones R.C., Hansen L.A., Man S.Y. Fluid and electrolyte reabsorption in the ductuli efferentes testis. J Reprod Fertil Suppl. 1998;53:1–14. [PubMed] [Google Scholar]
- 85.Eddy E.M., Vernon R.B., Muller C.H., Hahnel A.C., Fenderson B.A. Immunodissection of sperm surface modifications during epididymal maturation. Am J Anat. 1985;174:225–237. doi: 10.1002/aja.1001740305. [DOI] [PubMed] [Google Scholar]
- 86.Hinton B.T., Palladino M.A. Epididymal epithelium: its contribution to the formation of a luminal fluid microenvironment. Microsc Res Tech. 1995;30:67–81. doi: 10.1002/jemt.1070300106. [DOI] [PubMed] [Google Scholar]
- 87.de Kretser D.M., Temple-Smith P.D., Kerr J.B. Anatomical and functional aspects of the male reproductive organs. In: Bandhauer K., Frick J., editors. Disturbances of male fertility. Springer-Verlag; Berlin: 1982. pp. 1–131. [Google Scholar]
- 88.Russell D.W., Wilson J.D. Steroid 5α-reductase: two genes/two enzymes. Annu Rev Biochem. 1994;63:25–61. doi: 10.1146/annurev.bi.63.070194.000325. [DOI] [PubMed] [Google Scholar]
- 89.Marwood R.P., Beral V. Disappearance of spermatozoa from ejaculate after vasectomy. Br Med J. 1979;1:87. doi: 10.1136/bmj.1.6156.87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.McClinton S., Eremin O., Miller I.D. Inflammatory infiltrate in prostatic hyperplasia – evidence of a host response to intraprostatic spermatozoa? Br J Urol. 1990;65:606–610. doi: 10.1111/j.1464-410x.1990.tb14828.x. [DOI] [PubMed] [Google Scholar]
- 91.Beagley K.W., Wu Z.L., Pomering M., Jones R.C. Immune responses in the epididymis: implications for immunocontraception. J Reprod Fertil Suppl. 1998;53:235–245. [PubMed] [Google Scholar]
- 92.Cyr D.G., Robaire B., Hermo L. Structure and turnover of junctional complexes between principal cells of the rat epididymis. Microsc Res Tech. 1995;30:54–66. doi: 10.1002/jemt.1070300105. [DOI] [PubMed] [Google Scholar]
- 93.Suzuki F., Nagano T. Regional differentiation of cell junctions in the excurrent duct epithelium of the rat testis as revealed by freeze-fracture. Anat Rec. 1978;191:503–519. doi: 10.1002/ar.1091910409. [DOI] [PubMed] [Google Scholar]
- 94.Yeung C.H., Nashan D., Sorg C. Basal cells of the human epididymis–antigenic and ultrastructural similarities to tissue-fixed macrophages. Biol Reprod. 1994;50:917–926. doi: 10.1095/biolreprod50.4.917. [DOI] [PubMed] [Google Scholar]
- 95.Nashan D., Jantos C., Ahlers D. Immuno-competent cells in the murine epididymis following infection with Escherichia coli. Int J Androl. 1993;16:47–52. doi: 10.1111/j.1365-2605.1993.tb01152.x. [DOI] [PubMed] [Google Scholar]
- 96.Nashan D., Malorny U., Sorg C., Cooper T., Nieschlag E. Immuno-competent cells in the murine epididymis. Int J Androl. 1989;12:85–94. doi: 10.1111/j.1365-2605.1989.tb01289.x. [DOI] [PubMed] [Google Scholar]
- 97.Hooper P., Smythe E., Richards R.C., Howard C.V., Lynch R.V., Lewis-Jones D.I. Total number of immunocompetent cells in the normal rat epididymis and after vasectomy. J Reprod Fertil. 1995;104:193–198. doi: 10.1530/jrf.0.1040193. [DOI] [PubMed] [Google Scholar]
- 98.Flickinger C.J., Bush L.A., Howards S.S., Herr J.C. Distribution of leukocytes in the epithelium and interstitium of four regions of the Lewis rat epididymis. Anat Rec. 1997;248:380–390. doi: 10.1002/(SICI)1097-0185(199707)248:3<380::AID-AR11>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- 99.Nashan D., Cooper T.G., Knuth U.A., Schubeus P., Sorg C., Nieschlag E. Presence and distribution of leucocyte subsets in the murine epididymis after vasectomy. Int J Androl. 1990;13:39–49. doi: 10.1111/j.1365-2605.1990.tb00958.x. [DOI] [PubMed] [Google Scholar]
- 100.Serre V., Robaire B. Distribution of immune cells in the epididymis of the aging Brown Norway rat is segment-specific and related to the luminal content. Biol Reprod. 1999;61:705–714. doi: 10.1095/biolreprod61.3.705. [DOI] [PubMed] [Google Scholar]
- 101.Hedger M.P. Immunophysiology and pathology of inflammation in the testis and epididymis. J Androl. 2011;32:625–640. doi: 10.2164/jandrol.111.012989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Da Silva N., Cortez-Retamozo V., Reinecker H.C. A dense network of dendritic cells populates the murine epididymis. Reproduction. 2011;141:653–663. doi: 10.1530/REP-10-0493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Fulmer B.R., Turner T.T. A blood-prostate barrier restricts cell and molecular movement across the rat ventral prostate epithelium. J Urol. 2000;163:1591–1594. [PubMed] [Google Scholar]
- 104.Mason D., Simmons D., Buckley C. Oxford University Press; Oxford: 2002. Leucocyte typing VII. [Google Scholar]
- 105.Akira S., Uematsu S., Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. doi: 10.1016/j.cell.2006.02.015. [DOI] [PubMed] [Google Scholar]
- 106.O’Neill L.A., Bowie A.G. The family of five: TIR-domain-containing adaptors in Toll-like receptor signalling. Nat Rev Immunol. 2007;7:353–364. doi: 10.1038/nri2079. [DOI] [PubMed] [Google Scholar]
- 107.Takeuchi O., Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140:805–820. doi: 10.1016/j.cell.2010.01.022. [DOI] [PubMed] [Google Scholar]
- 108.Tang D., Kang R., Coyne C.B., Zeh H.J., Lotze M.T. PAMPs and DAMPs: signal 0s that spur autophagy and immunity. Immunol Rev. 2012;249:158–175. doi: 10.1111/j.1600-065X.2012.01146.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Zhang D., Zhang G., Hayden M.S. A toll-like receptor that prevents infection by uropathogenic bacteria. Science. 2004;303:1522–1526. doi: 10.1126/science.1094351. [DOI] [PubMed] [Google Scholar]
- 110.Haziot A., Ferrero E., Kontgen F. Resistance to endotoxin shock and reduced dissemination of gram-negative bacteria in CD14-deficient mice. Immunity. 1996;4:407–414. doi: 10.1016/s1074-7613(00)80254-x. [DOI] [PubMed] [Google Scholar]
- 111.Schromm A.B., Lien E., Henneke P. Molecular genetic analysis of an endotoxin nonresponder mutant cell line: a point mutation in a conserved region of MD-2 abolishes endotoxin-induced signaling. J Exp Med. 2001;194:79–88. doi: 10.1084/jem.194.1.79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.O’Neill L.A., Fitzgerald K.A., Bowie A.G. The Toll-IL-1 receptor adaptor family grows to five members. Trends Immunol. 2003;24:286–290. doi: 10.1016/s1471-4906(03)00115-7. [DOI] [PubMed] [Google Scholar]
- 113.Foletta V.C., Segal D.H., Cohen D.R. Transcriptional regulation in the immune system: all roads lead to AP-1. J Leuk Biol. 1998;63:139–152. doi: 10.1002/jlb.63.2.139. [DOI] [PubMed] [Google Scholar]
- 114.Tang G., Minemoto Y., Dibling B. Inhibition of JNK activation through NF-κB target genes. Nature. 2001;414:313–317. doi: 10.1038/35104568. [DOI] [PubMed] [Google Scholar]
- 115.Hertzog P.J., O’Neill L.A., Hamilton J.A. The interferon in TLR signaling: more than just antiviral. Trends Immunol. 2003;24:534–539. doi: 10.1016/j.it.2003.08.006. [DOI] [PubMed] [Google Scholar]
- 116.Cook D.N., Pisetsky D.S., Schwartz D.A. Toll-like receptors in the pathogenesis of human disease. Nat Immunol. 2004;5:975–979. doi: 10.1038/ni1116. [DOI] [PubMed] [Google Scholar]
- 117.Deretic V. Autophagy: an emerging immunological paradigm. J Immunol. 2012;189:15–20. doi: 10.4049/jimmunol.1102108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Davis B.K., Wen H., Ting J.P. The inflammasome NLRs in immunity, inflammation, and associated diseases. Annu Rev Immunol. 2011;29:707–735. doi: 10.1146/annurev-immunol-031210-101405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Rosenberg H.F., Gallin J.I. Inflammation. In: Paul W.E., editor. Fundamental immunology. 5th ed. Lippincott, Williams and Wilkins; Philadelphia: 2003. pp. 1151–1169. [Google Scholar]
- 120.Galli S.J., Nakae S., Tsai M. Mast cells in the development of adaptive immune responses. Nat Immunol. 2005;6:135–142. doi: 10.1038/ni1158. [DOI] [PubMed] [Google Scholar]
- 121.Mantovani A., Cassatella M.A., Costantini C., Jaillon S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat Rev Immunol. 2011;11:519–531. doi: 10.1038/nri3024. [DOI] [PubMed] [Google Scholar]
- 122.Moser B., Loetscher P. Lymphocyte traffic control by chemokines. Nat Immunol. 2001;2:123–128. doi: 10.1038/84219. [DOI] [PubMed] [Google Scholar]
- 123.Kulkarni A.B., Huh C.G., Becker D. Transforming growth factor β1 null mutation in mice causes excessive inflammatory response and early death. Proc Natl Acad Sci USA. 1993;90:770–774. doi: 10.1073/pnas.90.2.770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Kapcala L.P., Chautard T., Eskay R.L. The protective role of the hypothalamic-pituitary-adrenal axis against lethality produced by immune, infectious, and inflammatory stress. Ann NY Acad Sci. 1995;771:419–437. doi: 10.1111/j.1749-6632.1995.tb44699.x. [DOI] [PubMed] [Google Scholar]
- 125.Tilg H., Dinarello C.A., Mier J.W. IL-6 and APPs: anti-inflammatory and immunosuppressive mediators. Immunol Today. 1997;18:428–432. doi: 10.1016/s0167-5699(97)01103-1. [DOI] [PubMed] [Google Scholar]
- 126.Branton M.H., Kopp J.B. TGF-β and fibrosis. Microbes Infect. 1999;1:1349–1365. doi: 10.1016/s1286-4579(99)00250-6. [DOI] [PubMed] [Google Scholar]
- 127.Smith-Garvin J.E., Koretzky G.A., Jordan M.S. T cell activation. Annu Rev Immunol. 2009;27:591–619. doi: 10.1146/annurev.immunol.021908.132706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Tonegawa S. Somatic generation of antibody diversity. Nature. 1983;302:575–581. doi: 10.1038/302575a0. [DOI] [PubMed] [Google Scholar]
- 129.Hood L., Kronenberg M., Hunkapiller T. T cell antigen receptors and the immunoglobulin supergene family. Cell. 1985;40:225–229. doi: 10.1016/0092-8674(85)90133-3. [DOI] [PubMed] [Google Scholar]
- 130.Zinkernagel R.M., Doherty P.C. MHC-restricted cytotoxic T cells: studies on the biological role of polymorphic major transplantation antigens determining T-cell restriction-specificity, function, and responsiveness. Adv Immunol. 1979;27:51–177. doi: 10.1016/s0065-2776(08)60262-x. [DOI] [PubMed] [Google Scholar]
- 131.Fooksman D.R., Vardhana S., Vasiliver-Shamis G. Functional anatomy of T cell activation and synapse formation. Annu Rev Immunol. 2010;28:79–105. doi: 10.1146/annurev-immunol-030409-101308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Janeway C.A., Jr. The T cell receptor as a multicomponent signalling machine: CD4/CD8 coreceptors and CD45 in T cell activation. Annu Rev Immunol. 1992;10:645–674. doi: 10.1146/annurev.iy.10.040192.003241. [DOI] [PubMed] [Google Scholar]
- 133.Banchereau J., Steinman R.M. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. doi: 10.1038/32588. [DOI] [PubMed] [Google Scholar]
- 134.Constant S.L., Bottomly K. Induction of Th1 and Th2 CD4+ T cell responses: the alternative approaches. Annu Rev Immunol. 1997;15:297–322. doi: 10.1146/annurev.immunol.15.1.297. [DOI] [PubMed] [Google Scholar]
- 135.Moser M., Murphy K.M. Dendritic cell regulation of TH1-TH2 development. Nat Immunol. 2000;1:199–205. doi: 10.1038/79734. [DOI] [PubMed] [Google Scholar]
- 136.Ouyang W., Kolls J.K., Zheng Y. The biological functions of T helper 17 cell effector cytokines in inflammation. Immunity. 2008;28:454–467. doi: 10.1016/j.immuni.2008.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Ghoreschi K., Laurence A., Yang X.P., Hirahara K., O’Shea J.J. T helper 17 cell heterogeneity and pathogenicity in autoimmune disease. Trends Immunol. 2011;32:395–401. doi: 10.1016/j.it.2011.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Oboki K., Ohno T., Saito H., Nakae S. Th17 and allergy. Allerg Int. 2008;57:121–134. doi: 10.2332/allergolint.R-07-160. [DOI] [PubMed] [Google Scholar]
- 139.Gilliet M., Liu Y.J. Generation of human CD8 T regulatory cells by CD40 ligand-activated plasmacytoid dendritic cells. J Exp Med. 2002;195:695–704. doi: 10.1084/jem.20011603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Josefowicz S.Z., Lu L.F., Rudensky A.Y. Regulatory T cells: mechanisms of differentiation and function. Annu Rev Immunol. 2012;30:531–564. doi: 10.1146/annurev.immunol.25.022106.141623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.DeFranco A.L. Molecular aspects of B-lymphocyte activation. Annu Rev Cell Biol. 1987;3:143–178. doi: 10.1146/annurev.cb.03.110187.001043. [DOI] [PubMed] [Google Scholar]
- 142.Schittek B., Rajewsky K. Maintenance of B-cell memory by long-lived cells generated from proliferating precursors. Nature. 1990;346:749–751. doi: 10.1038/346749a0. [DOI] [PubMed] [Google Scholar]
- 143.Dutton R.W., Bradley L.M., Swain S.L. T cell memory. Annu Rev Immunol. 1998;16:201–223. doi: 10.1146/annurev.immunol.16.1.201. [DOI] [PubMed] [Google Scholar]
- 144.Sun J.C., Lanier L.L. NK cell development, homeostasis and function: parallels with CD8+ T cells. Nat Rev Immunol. 2011;11:645–657. doi: 10.1038/nri3044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Loza M.J., Perussia B. Final steps of natural killer cell maturation: a model for type 1–type 2 differentiation? Nat Immunol. 2001;2:917–924. doi: 10.1038/ni1001-917. [DOI] [PubMed] [Google Scholar]
- 146.Kraus E., Lambracht D., Wonigeit K., Hünig T. Negative regulation of rat natural killer cell activity by major histocompatibility complex class I recognition. Eur J Immunol. 1996;26:2582–2586. doi: 10.1002/eji.1830261107. [DOI] [PubMed] [Google Scholar]
- 147.Laouar Y., Sutterwala F.S., Gorelik L., Flavell R.A. Transforming growth factor-β controls T helper type 1 cell development through regulation of natural killer cell interferon-γ. Nat Immunol. 2005;6:600–607. doi: 10.1038/ni1197. [DOI] [PubMed] [Google Scholar]
- 148.Godfrey D.I., Stankovic S., Baxter A.G. Raising the NKT cell family. Nat Immunol. 2010;11:197–206. doi: 10.1038/ni.1841. [DOI] [PubMed] [Google Scholar]
- 149.Baker J., Verneris M.R., Ito M., Shizuru J.A., Negrin R.S. Expansion of cytolytic CD8+ natural killer T cells with limited capacity for graft-versus-host disease induction due to interferon γ production. Blood. 2001;97:2923–2931. doi: 10.1182/blood.v97.10.2923. [DOI] [PubMed] [Google Scholar]
- 150.Sonoda K.H., Faunce D.E., Taniguchi M., Exley M., Balk S., Stein-Streilein J. NK T cell-derived IL-10 is essential for the differentiation of antigen-specific T regulatory cells in systemic tolerance. J Immunol. 2001;166:42–50. doi: 10.4049/jimmunol.166.1.42. [DOI] [PubMed] [Google Scholar]
- 151.Nossal G.J. Negative selection of lymphocytes. Cell. 1994;76:229–239. doi: 10.1016/0092-8674(94)90331-x. [DOI] [PubMed] [Google Scholar]
- 152.Metzger T.C., Anderson M.S. Control of central and peripheral tolerance by Aire. Immunol Rev. 2011;241:89–103. doi: 10.1111/j.1600-065X.2011.01008.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Gotter J., Brors B., Hergenhahn M., Kyewski B. Medullary epithelial cells of the human thymus express a highly diverse selection of tissue-specific genes colocalized in chromosomal clusters. J Exp Med. 2004;199:155–166. doi: 10.1084/jem.20031677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Ermann J., Fathman C.G. Autoimmune diseases: genes, bugs and failed regulation. Nat Immunol. 2001;2:759–761. doi: 10.1038/ni0901-759. [DOI] [PubMed] [Google Scholar]
- 155.Sakaguchi S., Sakaguchi N. Thymus and autoimmunity: capacity of the normal thymus to produce pathogenic self-reactive T cells and conditions required for their induction of autoimmune disease. J Exp Med. 1990;172:537–545. doi: 10.1084/jem.172.2.537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Mueller D.L. Mechanisms maintaining peripheral tolerance. Nat Immunol. 2010;11:21–27. doi: 10.1038/ni.1817. [DOI] [PubMed] [Google Scholar]
- 157.Bluestone J.A. Mechanisms of tolerance. Immunol Rev. 2011;241:5–19. doi: 10.1111/j.1600-065X.2011.01019.x. [DOI] [PubMed] [Google Scholar]
- 158.von Boehmer H. Mechanisms of suppression by suppressor T cells. Nat Immunol. 2005;6:338–344. doi: 10.1038/ni1180. [DOI] [PubMed] [Google Scholar]
- 159.Piccirillo C.A., Letterio J.J., Thornton A.M. CD4+CD25+ regulatory T cells can mediate suppressor function in the absence of transforming growth factor β1 production and responsiveness. J Exp Med. 2002;196:237–246. doi: 10.1084/jem.20020590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Nakamura K., Kitani A., Strober W. Cell contact-dependent immunosuppression by CD4+CD25+ regulatory T cells is mediated by cell surface-bound transforming growth factor β. J Exp Med. 2001;194:629–644. doi: 10.1084/jem.194.5.629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Kuhnlein P., Park J.H., Herrmann T., Elbe A., Hunig T. Identification and characterization of rat gamma/delta T lymphocytes in peripheral lymphoid organs, small intestine, and skin with a monoclonal antibody to a constant determinant of the gamma/delta T cell receptor. J Immunol. 1994;153:979–986. [PubMed] [Google Scholar]
- 162.MacDonald T.T. T cell immunity to oral allergens. Curr Opin Immunol. 1998;10:620–627. doi: 10.1016/s0952-7915(98)80079-4. [DOI] [PubMed] [Google Scholar]
- 163.Seo N., Hayakawa S., Takigawa M., Tokura Y. Interleukin-10 expressed at early tumour sites induces subsequent generation of CD4+ T-regulatory cells and systemic collapse of antitumour immunity. Immunology. 2001;103:449–457. doi: 10.1046/j.1365-2567.2001.01279.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Burrows P.D., Suematsu S., Watanabe T. Activation of self-reactive B cells and autoimmune diseases. Rev Immunogen. 2000;2:38–51. [PubMed] [Google Scholar]
- 165.Merkler D., Horvath E., Bruck W., Zinkernagel R.M., de la Torre J.C., Pinschewer D.D. “Viral déjà vu” elicits organ-specific immune disease independent of reactivity to self. J Clin Invest. 2006;116:1254–1263. doi: 10.1172/JCI27372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Pitkanen J., Peterson P. Autoimmune regulator: from loss of function to autoimmunity. Genes Immun. 2003;4:12–21. doi: 10.1038/sj.gene.6363929. [DOI] [PubMed] [Google Scholar]
- 167.Kriegel M.A., Lohmann T., Gabler C., Blank N., Kalden J.R., Lorenz H.M. Defective suppressor function of human CD4+ CD25+ regulatory T cells in autoimmune polyglandular syndrome type II. J Exp Med. 2004;199:1285–1291. doi: 10.1084/jem.20032158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Baker M.B., Altman N.H., Podack E.R., Levy R.B. The role of cell-mediated cytotoxicity in acute GVHD after MHC-matched allogeneic bone marrow transplantation in mice. J Exp Med. 1996;183:2645–2656. doi: 10.1084/jem.183.6.2645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Hviid T.V. HLA-G in human reproduction: aspects of genetics, function and pregnancy complications. Hum Reprod Update. 2006;12:209–232. doi: 10.1093/humupd/dmi048. [DOI] [PubMed] [Google Scholar]
- 170.Romani N., Ratzinger G., Pfaller K. Migration of dendritic cells into lymphatics-the Langerhans cell example: routes, regulation, and relevance. Int Rev Cytol. 2001;207:237–270. doi: 10.1016/s0074-7696(01)07007-3. [DOI] [PubMed] [Google Scholar]
- 171.Pedersen N.C., Morris B. The role of the lymphatic system in the rejection of homografts: a study of lymph from renal transplants. J Exp Med. 1970;131:936–969. doi: 10.1084/jem.131.5.936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Gruber S.A. The case for local immunosuppression. Transplantation. 1992;54:1–11. doi: 10.1097/00007890-199207000-00001. [DOI] [PubMed] [Google Scholar]
- 173.Gallucci S., Matzinger P. Danger signals: SOS to the immune system. Curr Opin Immunol. 2001;13:114–119. doi: 10.1016/s0952-7915(00)00191-6. [DOI] [PubMed] [Google Scholar]
- 174.McKay L.I., Cidlowski J.A. CBP (CREB binding protein) integrates NF-kappaB (nuclear factor-kappaB) and glucocorticoid receptor physical interactions and antagonism. Mol Endocrinol. 2000;14:1222–1234. doi: 10.1210/mend.14.8.0506. [DOI] [PubMed] [Google Scholar]
- 175.Cutolo M., Straub R.H. Insights into endocrine-immunological disturbances in autoimmunity and their impact on treatment. Arthritis Res Ther. 2009;11:218. doi: 10.1186/ar2630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Liberman A.C., Druker J., Perone M.J., Arzt E. Glucocorticoids in the regulation of transcription factors that control cytokine synthesis. Cytokine Growth Factor Rev. 2007;18:45–56. doi: 10.1016/j.cytogfr.2007.01.005. [DOI] [PubMed] [Google Scholar]
- 177.Tait A.S., Butts C.L., Sternberg E.M. The role of glucocorticoids and progestins in inflammatory, autoimmune, and infectious disease. J Leuk Biol. 2008;84:924–931. doi: 10.1189/jlb.0208104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Levy B.D., Clish C.B., Schmidt B., Gronert K., Serhan C.N. Lipid mediator class switching during acute inflammation: signals in resolution. Nat Immunol. 2001;2:612–619. doi: 10.1038/89759. [DOI] [PubMed] [Google Scholar]
- 179.Herlong J.L., Scott T.R. Positioning prostanoids of the D and J series in the immunopathogenic scheme. Immunol Lett. 2006;102:121–131. doi: 10.1016/j.imlet.2005.10.004. [DOI] [PubMed] [Google Scholar]
- 180.Ju S.T., Panka D.J., Cui H. Fas(CD95)/FasL interactions required for programmed cell death after T-cell activation. Nature. 1995;373:444–448. doi: 10.1038/373444a0. [DOI] [PubMed] [Google Scholar]
- 181.Isaacs J.D. The antiglobulin response to therapeutic antibodies. Sem Immunol. 1990;2:449–456. [PubMed] [Google Scholar]
- 182.Czerkinsky C., Anjuere F., McGhee J.R. Mucosal immunity and tolerance: relevance to vaccine development. Immunol Rev. 1999;170:197–222. doi: 10.1111/j.1600-065X.1999.tb01339.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Thorstenson K.M., Khoruts A. Generation of anergic and potentially immunoregulatory CD25+CD4 T cells in vivo after induction of peripheral tolerance with intravenous or oral antigen. J Immunol. 2001;167:188–195. doi: 10.4049/jimmunol.167.1.188. [DOI] [PubMed] [Google Scholar]
- 184.Khoo U.Y., Proctor I.E., Macpherson A.J. CD4+ T cell down-regulation in human intestinal mucosa. Evidence for intestinal tolerance to luminal bacterial antigens. J Immunol. 1997;158:3626–3634. [PubMed] [Google Scholar]
- 185.Matsuda J.L., Naidenko O.V., Gapin L. Tracking the response of natural killer T cells to a glycolipid antigen using CD1d tetramers. J Exp Med. 2000;192:741–754. doi: 10.1084/jem.192.5.741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Russell M.W., Sibley D.A., Nikolova E.B., Tomana M., Mestecky J. IgA antibody as a non-inflammatory regulator of immunity. Biochem Soc Trans. 1997;25:466–470. doi: 10.1042/bst0250466. [DOI] [PubMed] [Google Scholar]
- 187.Hendrickson B.A., Rindisbacher L., Corthesy B. Lack of association of secretory component with IgA in J chain-deficient mice. J Immunol. 1996;157:750–754. [PubMed] [Google Scholar]
- 188.Hershberg R.M., Cho D.H., Youakim A. Highly polarized HLA class II antigen processing and presentation by human intestinal epithelial cells. J Clin Invest. 1998;102:792–803. doi: 10.1172/JCI3201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Jung H.C., Eckmann L., Yang S.K. A distinct array of proinflammatory cytokines is expressed in human colon epithelial cells in response to bacterial invasion. J Clin Invest. 1995;95:55–65. doi: 10.1172/JCI117676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Clifton V.L., Husband A.J., Kay D.J. Local immunity in the male reproductive tract. Immunol Cell Biol. 1992;70:301–307. doi: 10.1038/icb.1992.38. [DOI] [PubMed] [Google Scholar]
- 191.Pattinson H.A., Mortimer D. Prevalence of sperm surface antibodies in the male partners of infertile couples as determined by immunobead screening. Fertil Steril. 1987;48:466–469. doi: 10.1016/s0015-0282(16)59420-5. [DOI] [PubMed] [Google Scholar]
- 192.Hardy M.P., Gao H.B., Dong Q. Stress hormone and male reproductive function. Cell Tissue Res. 2005;322:147–153. doi: 10.1007/s00441-005-0006-2. [DOI] [PubMed] [Google Scholar]
- 193.Lockshin M.D. Sex differences in autoimmune disease. Lupus. 2006;15:753–756. doi: 10.1177/0961203306069353. [DOI] [PubMed] [Google Scholar]
- 194.Smith-Bouvier D.L., Divekar A.A., Sasidhar M. A role for sex chromosome complement in the female bias in autoimmune disease. J Exp Med. 2008;205:1099–1108. doi: 10.1084/jem.20070850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Sakiani S., Olsen N.J., Kovacs W.J. Gonadal steroids and humoral immunity. Nat Rev Endocrinol. 2013;9:56–62. doi: 10.1038/nrendo.2012.206. [DOI] [PubMed] [Google Scholar]
- 196.Hince M., Sakkal S., Vlahos K., Dudakov J., Boyd R., Chidgey A. The role of sex steroids and gonadectomy in the control of thymic involution. Cell Immunol. 2008;252:122–138. doi: 10.1016/j.cellimm.2007.10.007. [DOI] [PubMed] [Google Scholar]
- 197.Zhang M.A., Rego D., Moshkova M. Peroxisome proliferator-activated receptor (PPAR)α and -γ regulate IFNγ and IL-17A production by human T cells in a sex-specific way. Proc Natl Acad Sci USA. 2012;109:9505–9510. doi: 10.1073/pnas.1118458109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Chuang K.H., Altuwaijri S., Li G. Neutropenia with impaired host defense against microbial infection in mice lacking androgen receptor. J Exp Med. 2009;206:1181–1199. doi: 10.1084/jem.20082521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Lai J.J., Lai K.P., Zeng W.P., Chuang K.H., Altuwaijri S., Chang C.S. Androgen receptor influences on body defense system via modulation of innate and adaptive immune systems. Lessons from conditional AR knockout mice. Am J Pathol. 2012;181:1504–1512. doi: 10.1016/j.ajpath.2012.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Corrales J.J., Almeida M., Cordero M. Enhanced immunological response by dendritic cells in male hypogonadism. Eur J Clin Invest. 2012;42:1205–1212. doi: 10.1111/j.1365-2362.2012.02712.x. [DOI] [PubMed] [Google Scholar]
- 201.Yesilova Z., Ozata M., Kocar I.H. The effects of gonadotropin treatment on the immunological features of male patients with idiopathic hypogonadotropic hypogonadism. J Clin Endocrinol Metab. 2000;85:66–70. doi: 10.1210/jcem.85.1.6226. [DOI] [PubMed] [Google Scholar]
- 202.Xu J., Itoh Y., Hayashi H., Takii T., Miyazawa K., Onozaki K. Dihydrotestosterone inhibits interleukin-1α or tumor necrosis factor α-induced proinflammatory cytokine production via androgen receptor-dependent inhibition of nuclear factor-κB activation in rheumatoid fibroblast-like synovial cell line. Biol Pharm Bull. 2011;34:1724–1730. doi: 10.1248/bpb.34.1724. [DOI] [PubMed] [Google Scholar]
- 203.Rettew J.A., Huet-Hudson Y.M., Marriott I. Testosterone reduces macrophage expression in the mouse of Toll-like receptor 4, a trigger for inflammation and innate immunity. Biol Reprod. 2008;78:432–437. doi: 10.1095/biolreprod.107.063545. [DOI] [PubMed] [Google Scholar]
- 204.Norata G.D., Cattaneo P., Poletti A., Catapano A.L. The androgen derivative 5α-androstane-3β,17β-diol inhibits tumor necrosis factor α and lipopolysaccharide induced inflammatory response in human endothelial cells and in mice aorta. Atherosclerosis. 2010;212:100–106. doi: 10.1016/j.atherosclerosis.2010.05.015. [DOI] [PubMed] [Google Scholar]
- 205.Pergola C., Dodt G., Rossi A. ERK-mediated regulation of leukotriene biosynthesis by androgens: a molecular basis for gender differences in inflammation and asthma. Proc Natl Acad Sci USA. 2008;105:19881–19886. doi: 10.1073/pnas.0809120105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.MacLean H.E., Warne G.L., Zajac J.D. Defects of androgen receptor function: from sex reversal to motor neurone disease. Mol Cell Endocrinol. 1995;112:133–141. doi: 10.1016/0303-7207(95)03608-a. [DOI] [PubMed] [Google Scholar]
- 207.McCruden A.B., Stimson W.H. Androgen binding cytosol receptors in the rat thymus: physicochemical properties, specificity and localisation. Thymus. 1981;3:105–117. [PubMed] [Google Scholar]
- 208.Cohen J.H., Danel L., Cordier G., Saez S., Revillard J.P. Sex steroid receptors in peripheral T cells: absence of androgen receptors and restriction of estrogen receptors to OKT8-positive cells. J Immunol. 1983;131:2767–2771. [PubMed] [Google Scholar]
- 209.Mantalaris A., Panoskaltsis N., Sakai Y. Localization of androgen receptor expression in human bone marrow. J Pathol. 2001;193:361–366. doi: 10.1002/1096-9896(0000)9999:9999<::AID-PATH803>3.0.CO;2-W. [DOI] [PubMed] [Google Scholar]
- 210.Liva S.M., Voskuhl R.R. Testosterone acts directly on CD4+ T lymphocytes to increase IL-10 production. J Immunol. 2001;167:2060–2067. doi: 10.4049/jimmunol.167.4.2060. [DOI] [PubMed] [Google Scholar]
- 211.Braun A.M., Thomas P. Biochemical characterization of a membrane androgen receptor in the ovary of the atlantic croaker (Micropogonias undulatus) Biol Reprod. 2004;71:146–155. doi: 10.1095/biolreprod.103.025825. [DOI] [PubMed] [Google Scholar]
- 212.Zhu Y., Bond J., Thomas P. Identification, classification, and partial characterization of genes in humans and other vertebrates homologous to a fish membrane progestin receptor. Proc Natl Acad Sci USA. 2003;100:2237–2242. doi: 10.1073/pnas.0436133100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Benten W.P., Lieberherr M., Sekeris C.E., Wunderlich F. Testosterone induces Ca2+ influx via non-genomic surface receptors in activated T cells. FEBS Lett. 1997;407:211–214. doi: 10.1016/s0014-5793(97)00346-3. [DOI] [PubMed] [Google Scholar]
- 214.Benten W.P., Lieberherr M., Stamm O., Wrehlke C., Guo Z., Wunderlich F. Testosterone signaling through internalizable surface receptors in androgen receptor-free macrophages. Mol Biol Cell. 1999;10:3113–3123. doi: 10.1091/mbc.10.10.3113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Benten W.P., Guo Z., Krucken J., Wunderlich F. Rapid effects of androgens in macrophages. Steroids. 2004;69:585–590. doi: 10.1016/j.steroids.2004.05.007. [DOI] [PubMed] [Google Scholar]
- 216.van den Broek H.H., Damoiseaux J.G., De Baets M.H., Hupperts R.M. The influence of sex hormones on cytokines in multiple sclerosis and experimental autoimmune encephalomyelitis: a review. Mult Scler. 2005;11:349–359. doi: 10.1191/1352458505ms1174rr. [DOI] [PubMed] [Google Scholar]
- 217.Kovats S., Carreras E. Regulation of dendritic cell differentiation and function by estrogen receptor ligands. Cell Immunol. 2008;252:81–90. doi: 10.1016/j.cellimm.2007.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Segerer S.E., Muller N., van den Brandt J. Impact of female sex hormones on the maturation and function of human dendritic cells. Am J Reprod Immunol. 2009;62:165–173. doi: 10.1111/j.1600-0897.2009.00726.x. [DOI] [PubMed] [Google Scholar]
- 219.Robinson L.J., Tourkova I., Wang Y. FSH-receptor isoforms and FSH-dependent gene transcription in human monocytes and osteoclasts. Biochem Biophys Res Commun. 2010;394:12–17. doi: 10.1016/j.bbrc.2010.02.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Carbone F., Procaccini C., De Rosa V. Divergent immunomodulatory effects of recombinant and urinary-derived FSH, LH, and hCG on human CD4+ T cells. J Reprod Immunol. 2010;85:172–179. doi: 10.1016/j.jri.2010.02.009. [DOI] [PubMed] [Google Scholar]
- 221.He X.L., Hedger M.P., Clements J.A., Risbridger G.P. Localization of immunoreactive β-endorphin and adrenocorticotropic hormone and pro-opiomelanocortin mRNA to rat testicular interstitial tissue macrophages. Biol Reprod. 1991;45:282–289. doi: 10.1095/biolreprod45.2.282. [DOI] [PubMed] [Google Scholar]
- 222.Hedger M.P., Robertson D.M., Browne C.A., de Kretser D.M. Studies on the identification of a LHRH-like peptide in the rat testis. Ann NY Acad Sci. 1984;438:371–381. doi: 10.1111/j.1749-6632.1984.tb38298.x. [DOI] [PubMed] [Google Scholar]
- 223.Chen C.L., Mather J.P., Morris P.L., Bardin C.W. Expression of pro-opiomelanocortin-like gene in the testis and epididymis. Proc Natl Acad Sci USA. 1984;81:5672–5675. doi: 10.1073/pnas.81.18.5672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Hedger M.P., Winnall W.R. Regulation of activin and inhibin in the adult testis and the evidence for functional roles in spermatogenesis and immunoregulation. Mol Cell Endocrinol. 2012;359:30–42. doi: 10.1016/j.mce.2011.09.031. [DOI] [PubMed] [Google Scholar]
- 225.Caussanel V., Tabone E., Hendrick J.C., Dacheux F., Benahmed M. Cellular distribution of transforming growth factor betas 1, 2, and 3 and their types I and II receptors during postnatal development and spermatogenesis in the boar testis. Biol Reprod. 1997;56:357–367. doi: 10.1095/biolreprod56.2.357. [DOI] [PubMed] [Google Scholar]
- 226.Meinhardt A., Bacher M., Metz C. Local regulation of macrophage subsets in the adult rat testis: examination of the roles of the seminiferous tubules, testosterone, and macrophage-migration inhibitory factor. Biol Reprod. 1998;59:371–378. doi: 10.1095/biolreprod59.2.371. [DOI] [PubMed] [Google Scholar]
- 227.Hedger M.P., Meinhardt A. Local regulation of T cell numbers and lymphocyte-inhibiting activity in the interstitial tissue of the adult rat testis. J Reprod Immunol. 2000;48:69–80. doi: 10.1016/s0165-0378(00)00071-1. [DOI] [PubMed] [Google Scholar]
- 228.Vergouwen R.P., Huiskamp R., Bas R.J., Roepers-Gajadien H.L., Davids J.A., de Rooij D.G. Postnatal development of testicular cell populations in mice. J Reprod Fertil. 1993;99:479–485. doi: 10.1530/jrf.0.0990479. [DOI] [PubMed] [Google Scholar]
- 229.Frungieri M.B., Calandra R.S., Lustig L. Number, distribution pattern, and identification of macrophages in the testes of infertile men. Fertil Steril. 2002;78:298–306. doi: 10.1016/s0015-0282(02)03206-5. [DOI] [PubMed] [Google Scholar]
- 230.Haas G.G., Jr., D’Cruz O.J., De Bault L.E. Distribution of human leukocyte antigen-ABC and -D/DR antigens in the unfixed human testis. Am J Reprod Immunol Microbiol. 1988;18:47–51. doi: 10.1111/j.1600-0897.1988.tb00234.x. [DOI] [PubMed] [Google Scholar]
- 231.el-Demiry M., James K. Lymphocyte subsets and macrophages in the male genital tract in health and disease. A monoclonal antibody-based study. Eur Urol. 1988;14:226–235. doi: 10.1159/000472944. [DOI] [PubMed] [Google Scholar]
- 232.el-Demiry M.I., Hargreave T.B., Busuttil A., Elton R., James K., Chisholm G.D. Immunocompetent cells in human testis in health and disease. Fertil Steril. 1987;48:470–479. doi: 10.1016/s0015-0282(16)59421-7. [DOI] [PubMed] [Google Scholar]
- 233.Briere F., Bendriss-Vermare N., Delale T. Origin and filiation of human plasmacytoid dendritic cells. Hum Immunol. 2002;63:1081–1093. doi: 10.1016/s0198-8859(02)00746-2. [DOI] [PubMed] [Google Scholar]
- 234.Chomarat P., Banchereau J., Davoust J., Palucka A.K. IL-6 switches the differentiation of monocytes from dendritic cells to macrophages. Nat Immunol. 2000;1:510–514. doi: 10.1038/82763. [DOI] [PubMed] [Google Scholar]
- 235.van Furth R. Phagocytic cells: development and distribution of mononuclear phagocytes in normal steady state and inflammation. In: Gallin J.I., Goldstein I.M., Snyderman R., editors. Inflammation: basic principles and clinical correlates. Raven Press; New York: 1988. pp. 281–295. [Google Scholar]
- 236.Mantovani A., Sica A. Macrophages, innate immunity and cancer: balance, tolerance, and diversity. Curr Opin Immunol. 2010;22:231–237. doi: 10.1016/j.coi.2010.01.009. [DOI] [PubMed] [Google Scholar]
- 237.Miller S.C. Localization of plutonium-241 in the testis. An interspecies comparison using light and electron microscope autoradiography. Int J Radiat Biol Relat Stud Phys Chem Med. 1982;41:633–643. doi: 10.1080/09553008214550731. [DOI] [PubMed] [Google Scholar]
- 238.Russell J.J., Lindenbaum A. One-year study of nonuniformly distributed plutonium in mouse testis as related to spermatogonial irradiation. Health Phys. 1979;36:153–157. doi: 10.1097/00004032-197902000-00006. [DOI] [PubMed] [Google Scholar]
- 239.Miller S.C., Bowman B.M., Roberts L.K. Identification and characterization of mononuclear phagocytes isolated from rat testicular interstitial tissues. J Leuk Biol. 1984;36:679–687. doi: 10.1002/jlb.36.6.679. [DOI] [PubMed] [Google Scholar]
- 240.Miller S.C., Bowman B.M., Rowland H.G. Structure, cytochemistry, endocytic activity, and immunoglobulin (Fc) receptors of rat testicular interstitial-tissue macrophages. Am J Anat. 1983;168:1–13. doi: 10.1002/aja.1001680102. [DOI] [PubMed] [Google Scholar]
- 241.Yee J.B., Hutson J.C. Testicular macrophages: isolation, characterization and hormonal responsiveness. Biol Reprod. 1983;29:1319–1326. doi: 10.1095/biolreprod29.5.1319. [DOI] [PubMed] [Google Scholar]
- 242.Yee J.B., Hutson J.C. Effects of testicular macrophage-conditioned medium on Leydig cells in culture. Endocrinology. 1985;116:2682–2684. doi: 10.1210/endo-116-6-2682. [DOI] [PubMed] [Google Scholar]
- 243.Wei R.Q., Yee J.B., Straus D.C., Hutson J.C. Bactericidal activity of testicular macrophages. Biol Reprod. 1988;38:830–835. doi: 10.1095/biolreprod38.4.830. [DOI] [PubMed] [Google Scholar]
- 244.Hutson J.C. Development of cytoplasmic digitations between Leydig cells and testicular macrophages of the rat. Cell Tissue Res. 1992;267:385–389. doi: 10.1007/BF00302977. [DOI] [PubMed] [Google Scholar]
- 245.Niemi M., Sharpe R.M., Brown W.R. Macrophages in the interstitial tissue of the rat testis. Cell Tissue Res. 1986;243:337–344. doi: 10.1007/BF00251049. [DOI] [PubMed] [Google Scholar]
- 246.Hedger M.P., Eddy E.M. The heterogeneity of isolated adult rat Leydig cells separated on Percoll density gradients: an immunological, cytochemical, and functional analysis. Endocrinology. 1987;121:1824–1838. doi: 10.1210/endo-121-5-1824. [DOI] [PubMed] [Google Scholar]
- 247.Sasmono R.T., Oceandy D., Pollard J.W. A macrophage colony-stimulating factor receptor-green fluorescent protein transgene is expressed throughout the mononuclear phagocyte system of the mouse. Blood. 2003;101:1155–1163. doi: 10.1182/blood-2002-02-0569. [DOI] [PubMed] [Google Scholar]
- 248.Mendis-Handagama S.M., Risbridger G.P., de Kretser D.M. Morphometric analysis of the components of the neonatal and the adult rat testis interstitium. Int J Androl. 1987;10:525–534. doi: 10.1111/j.1365-2605.1987.tb00352.x. [DOI] [PubMed] [Google Scholar]
- 249.Hardy M.P., Zirkin B.R., Ewing L.L. Kinetic studies on the development of the adult population of Leydig cells in testes of the pubertal rat. Endocrinology. 1989;124:762–770. doi: 10.1210/endo-124-2-762. [DOI] [PubMed] [Google Scholar]
- 250.Hume D.A., Halpin D., Charlton H., Gordon S. The mononuclear phagocyte system of the mouse defined by immunohistochemical localization of antigen F4/80: macrophages of endocrine organs. Proc Natl Acad Sci USA. 1984;81:4174–4177. doi: 10.1073/pnas.81.13.4174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Wrobel K.H., Dostal S., Schimmel M. Postnatal development of the tubular lamina propria and the intertubular tissue in the bovine testis. Cell Tissue Res. 1988;252:639–653. doi: 10.1007/BF00216652. [DOI] [PubMed] [Google Scholar]
- 252.Mendis-Handagama S.M., Zirkin B.R., Ewing L.L. Comparison of components of the testis interstitium with testosterone secretion in hamster, rat, and guinea pig testes perfused in vitro. Am J Anat. 1988;181:12–22. doi: 10.1002/aja.1001810103. [DOI] [PubMed] [Google Scholar]
- 253.Clemmons A.J., Thompson D.L., Jr., Johnson L. Local initiation of spermatogenesis in the horse. Biol Reprod. 1995;52:1258–1267. doi: 10.1095/biolreprod52.6.1258. [DOI] [PubMed] [Google Scholar]
- 254.Sur J.H., Doster A.R., Christian J.S. Porcine reproductive and respiratory syndrome virus replicates in testicular germ cells, alters spermatogenesis, and induces germ cell death by apoptosis. J Virol. 1997;71:9170–9179. doi: 10.1128/jvi.71.12.9170-9179.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Bambino T.H., Hsueh A.J. Direct inhibitory effect of glucocorticoids upon testicular luteinizing hormone receptor and steroidogenesis in vivo and in vitro. Endocrinology. 1981;108:2142–2148. doi: 10.1210/endo-108-6-2142. [DOI] [PubMed] [Google Scholar]
- 256.Bergh A. Treatment with hCG increases the size of Leydig cells and testicular macrophages in unilaterally cryptorchid rats. Int J Androl. 1987;10:765–772. doi: 10.1111/j.1365-2605.1987.tb00380.x. [DOI] [PubMed] [Google Scholar]
- 257.Geierhaas B., Bornstein S.R., Jarry H., Scherbaum W.A., Herrmann M., Pfeiffer E.F. Morphological and hormonal changes following vasectomy in rats, suggesting a functional role for Leydig-cell associated macrophages. Horm Metab Res. 1991;23:373–378. doi: 10.1055/s-2007-1003704. [DOI] [PubMed] [Google Scholar]
- 258.Giannessi F., Giambelluca M.A., Scavuzzo M.C., Ruffoli R. Ultrastructure of testicular macrophages in aging mice. J Morphol. 2005;263:39–46. doi: 10.1002/jmor.10287. [DOI] [PubMed] [Google Scholar]
- 259.Gaytan F., Bellido C., Aguilar E., van Rooijen N. Requirement for testicular macrophages in Leydig cell proliferation and differentiation during prepubertal development in rats. J Reprod Fertil. 1994;102:393–399. doi: 10.1530/jrf.0.1020393. [DOI] [PubMed] [Google Scholar]
- 260.Gaytan F., Bellido C., Aguilar E., van Rooijen N. Pituitary-testicular axis in rats lacking testicular macrophages. Eur J Endocrinol. 1995;132:218–222. doi: 10.1530/eje.0.1320218. [DOI] [PubMed] [Google Scholar]
- 261.Gaytan F., Bellido C., Morales C., Reymundo C., Aguilar E., Van Rooijen N. Effects of macrophage depletion at different times after treatment with ethylene dimethane sulfonate (EDS) on the regeneration of Leydig cells in the adult rat. J Androl. 1994;15:558–564. [PubMed] [Google Scholar]
- 262.Gaytan F., Bellido C., Morales C., Reymundo C., Aguilar E., van Rooijen N. Selective depletion of testicular macrophages and prevention of Leydig cell repopulation after treatment with ethylene dimethane sulfonate in rats. J Reprod Fertil. 1994;101:175–182. doi: 10.1530/jrf.0.1010175. [DOI] [PubMed] [Google Scholar]
- 263.Bergh A., Damber J.E., van Rooijen N. Liposome-mediated macrophage depletion: an experimental approach to study the role of testicular macrophages in the rat. J Endocrinol. 1993;136:407–413. doi: 10.1677/joe.0.1360407. [DOI] [PubMed] [Google Scholar]
- 264.Cohen P.E., Chisholm O., Arceci R.J., Stanley E.R., Pollard J.W. Absence of colony-stimulating factor-1 in osteopetrotic (csfmop/csfmop) mice results in male fertility defects. Biol Reprod. 1996;55:310–317. doi: 10.1095/biolreprod55.2.310. [DOI] [PubMed] [Google Scholar]
- 265.Gaytan F., Bellido C., Morales C., Garcia M., van Rooijen N., Aguilar E. In vivo manipulation (depletion versus activation) of testicular macrophages: central and local effects. J Endocrinol. 1996;150:57–65. doi: 10.1677/joe.0.1500057. [DOI] [PubMed] [Google Scholar]
- 266.Head J.R., Billingham R.E. Immune privilege in the testis. II. Evaluation of potential local factors. Transplantation. 1985;40:269–275. doi: 10.1097/00007890-198509000-00010. [DOI] [PubMed] [Google Scholar]
- 267.Tung K.S., Yule T.D., Mahi-Brown C.A., Listrom M.B. Distribution of histopathology and Ia positive cells in actively induced and passively transferred experimental autoimmune orchitis. J Immunol. 1987;138:752–759. [PubMed] [Google Scholar]
- 268.Mahi-Brown C.A., Yule T.D., Tung K.S. Adoptive transfer of murine autoimmune orchitis to naive recipients with immune lymphocytes. Cell Immunol. 1987;106:408–419. doi: 10.1016/0008-8749(87)90183-3. [DOI] [PubMed] [Google Scholar]
- 269.Kern S., Maddocks S. Indomethacin blocks the immunosuppressive activity of rat testicular macrophages cultured in vitro. J Reprod Immunol. 1995;28:189–201. doi: 10.1016/0165-0378(95)91391-q. [DOI] [PubMed] [Google Scholar]
- 270.Winnall W.R., Muir J.A., Hedger M.P. Rat resident testicular macrophages have an alternatively activated phenotype and constitutively produce interleukin-10 in vitro. J Leuk Biol. 2011;90:133–143. doi: 10.1189/jlb.1010557. [DOI] [PubMed] [Google Scholar]
- 271.Sainio-Pöllänen S., Saari T., Simell O., Pöllänen P. CD28-CD80/CD86 interactions in testicular immunoregulation. J Reprod Immunol. 1996;31:145–163. doi: 10.1016/0165-0378(96)00983-7. [DOI] [PubMed] [Google Scholar]
- 272.Rival C., Guazzone V.A., von Wulffen W. Expression of co-stimulatory molecules, chemokine receptors and proinflammatory cytokines in dendritic cells from normal and chronically inflamed rat testis. Mol Hum Reprod. 2007;13:853–861. doi: 10.1093/molehr/gam067. [DOI] [PubMed] [Google Scholar]
- 273.Bergh A., Damber J.E., van Rooijen N. The human chorionic gonadotrophin-induced inflammation-like response is enhanced in macrophage-depleted rat testes. J Endocrinol. 1993;136:415–420. doi: 10.1677/joe.0.1360415. [DOI] [PubMed] [Google Scholar]
- 274.O’Bryan M.K., Gerdprasert O., Nikolic-Paterson D.J. Cytokine profiles in the testes of rats treated with lipopolysaccharide reveal localized suppression of inflammatory responses. Am J Physiol Regul Integr Comp Physiol. 2005;288:R1744–R1755. doi: 10.1152/ajpregu.00651.2004. [DOI] [PubMed] [Google Scholar]
- 275.Bhushan S., Hossain H., Lu Y. Uropathogenic E. coli induce different immune response in testicular and peritoneal macrophages: implications for testicular immune privilege. PLoS One. 2011;6:e28452. doi: 10.1371/journal.pone.0028452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 276.Gerdprasert O., O’Bryan M.K., Muir J.A. The response of testicular leukocytes to lipopolysaccharide-induced inflammation: further evidence for heterogeneity of the testicular macrophage population. Cell Tissue Res. 2002;308:277–285. doi: 10.1007/s00441-002-0547-6. [DOI] [PubMed] [Google Scholar]
- 277.Kern S., Robertson S.A., Mau V.J., Maddocks S. Cytokine secretion by macrophages in the rat testis. Biol Reprod. 1995;53:1407–1416. doi: 10.1095/biolreprod53.6.1407. [DOI] [PubMed] [Google Scholar]
- 278.Meinhardt A., Bacher M., McFarlane J.R. Macrophage migration inhibitory factor production by Leydig cells: evidence for a role in the regulation of testicular function. Endocrinology. 1996;137:5090–5095. doi: 10.1210/endo.137.11.8895383. [DOI] [PubMed] [Google Scholar]
- 279.Murray P.J., Wynn T.A. Protective and pathogenic functions of macrophage subsets. Nat Rev Immunol. 2011;11:723–737. doi: 10.1038/nri3073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280.Barbé E., Damoiseaux J.G., Döpp E.A., Dijkstra C.D. Characterization and expression of the antigen present on resident rat macrophages recognized by monoclonal antibody ED2. Immunobiology. 1990;182:88–99. doi: 10.1016/S0171-2985(11)80586-3. [DOI] [PubMed] [Google Scholar]
- 281.van den Berg T.K., Döpp E.A., Dijkstra C.D. Rat macrophages: membrane glycoproteins in differentiation and function. Immunol Rev. 2001;184:45–57. doi: 10.1034/j.1600-065x.2001.1840105.x. [DOI] [PubMed] [Google Scholar]
- 282.Philippidis P., Mason J.C., Evans B.J. Hemoglobin scavenger receptor CD163 mediates interleukin-10 release and heme oxygenase-1 synthesis: antiinflammatory monocyte-macrophage responses in vitro, in resolving skin blisters in vivo, and after cardiopulmonary bypass surgery. Circ Res. 2004;94:119–126. doi: 10.1161/01.RES.0000109414.78907.F9. [DOI] [PubMed] [Google Scholar]
- 283.Sierra-Filardi E., Vega M.A., Sánchez-Mateos P., Corbí A.L., Puig-Kröger A. Heme Oxygenase-1 expression in M-CSF-polarized M2 macrophages contributes to LPS-induced IL-10 release. Immunobiology. 2010;215:788–795. doi: 10.1016/j.imbio.2010.05.020. [DOI] [PubMed] [Google Scholar]
- 284.Hedger M.P. Macrophages and the immune responsiveness of the testis. J Reprod Immunol. 2002;57:19–34. doi: 10.1016/s0165-0378(02)00016-5. [DOI] [PubMed] [Google Scholar]
- 285.Gerdprasert O., O’Bryan M.K., Nikolic-Paterson D.J., Sebire K., de Kretser D.M., Hedger M.P. Expression of monocyte chemoattractant protein-1 and macrophage colony-stimulating factor in normal and inflamed rat testis. Mol Hum Reprod. 2002;8:518–524. doi: 10.1093/molehr/8.6.518. [DOI] [PubMed] [Google Scholar]
- 286.Winnall W.R., Hedger M.P. Phenotypic and functional heterogeneity of the testicular macrophage population: a new regulatory model. J Reprod Immunol. 2013;97:147–158. doi: 10.1016/j.jri.2013.01.001. [DOI] [PubMed] [Google Scholar]
- 287.Bryniarski K., Szczepanik M., Maresz K., Ptak M., Ptak W. Subpopulations of mouse testicular macrophages and their immunoregulatory function. Am J Reprod Immunol. 2004;52:27–35. doi: 10.1111/j.1600-0897.2004.00178.x. [DOI] [PubMed] [Google Scholar]
- 288.Maresz K., Ponomarev E.D., Barteneva N., Tan Y., Mann M.K., Dittel B.N. IL-13 induces the expression of the alternative activation marker Ym1 in a subset of testicular macrophages. J Reprod Immunol. 2008;78:140–148. doi: 10.1016/j.jri.2008.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289.Hoek A., Allaerts W., Leenen P.J., Schoemaker J., Drexhage H.A. Dendritic cells and macrophages in the pituitary and the gonads. Evidence for their role in the fine regulation of the reproductive endocrine response. Eur J Endocrinol. 1997;136:8–24. doi: 10.1530/eje.0.1360008. [DOI] [PubMed] [Google Scholar]
- 290.Rival C., Lustig L., Iosub R. Identification of a dendritic cell population in normal testis and in chronically inflamed testis of rats with autoimmune orchitis. Cell Tissue Res. 2006;324:311–318. doi: 10.1007/s00441-005-0129-5. [DOI] [PubMed] [Google Scholar]
- 291.Fijak M., Iosub R., Schneider E. Identification of immunodominant autoantigens in rat autoimmune orchitis. J Pathol. 2005;207:127–138. doi: 10.1002/path.1828. [DOI] [PubMed] [Google Scholar]
- 292.Guazzone V.A., Hollwegs S., Mardirosian M. Characterization of dendritic cells in testicular draining lymph nodes in a rat model of experimental autoimmune orchitis. Int J Androl. 2011;34:276–289. doi: 10.1111/j.1365-2605.2010.01082.x. [DOI] [PubMed] [Google Scholar]
- 293.Raburn D.J., Coquelin A., Reinhart A.J., Hutson J.C. Regulation of the macrophage population in postnatal rat testis. J Reprod Immunol. 1993;24:139–151. doi: 10.1016/0165-0378(93)90016-b. [DOI] [PubMed] [Google Scholar]
- 294.Duckett R.J., Hedger M.P., McLachlan R.I., Wreford N.G. The effects of gonadotropin-releasing hormone immunization and recombinant follicle-stimulating hormone on the Leydig cell and macrophage populations of the adult rat testis. J Androl. 1997;18:417–423. [PubMed] [Google Scholar]
- 295.Duckett R.J., Wreford N.G., Meachem S.J., McLachlan R.I., Hedger M.P. Effect of chorionic gonadotropin and flutamide on Leydig cell and macrophage populations in the testosterone-estradiol-implanted adult rat. J Androl. 1997;18:656–662. [PubMed] [Google Scholar]
- 296.Gaytan F., Bellido C., Romero J.L., Morales C., Reymundo C., Aguilar E. Decreased number and size and the defective function of testicular macrophages in long-term hypophysectomized rats are reversed by treatment with human gonadotrophins. J Endocrinol. 1994;140:399–407. doi: 10.1677/joe.0.1400399. [DOI] [PubMed] [Google Scholar]
- 297.Gaytan F., Romero J.L., Bellido C., Morales C., Reymundo C., Aguilar E. Effects of growth hormone and prolactin on testicular macrophages in long-term hypophysectomized rats. J Reprod Immunol. 1994;27:73–84. doi: 10.1016/0165-0378(94)90016-7. [DOI] [PubMed] [Google Scholar]
- 298.Dombrowicz D., Sente B., Closset J., Hennen G. Dose-dependent effects of human prolactin on the immature hypophysectomized rat testis. Endocrinology. 1992;130:695–700. doi: 10.1210/endo.130.2.1733717. [DOI] [PubMed] [Google Scholar]
- 299.Jackson A.E., de Kretser D.M. Ultrastructural immunoperoxidase investigations of hCG binding to isolated testicular intertubular cells. Histochem J. 1991;23:517–528. doi: 10.1007/BF01041178. [DOI] [PubMed] [Google Scholar]
- 300.Zhang Y.M., Rao Ch V., Lei Z.M. Macrophages in human reproductive tissues contain luteinizing hormone/chorionic gonadotropin receptors. Am J Reprod Immunol. 2003;49:93–100. doi: 10.1034/j.1600-0897.2003.00013.x. [DOI] [PubMed] [Google Scholar]
- 301.Cohen P.E., Hardy M.P., Pollard J.W. Colony-stimulating factor-1 plays a major role in the development of reproductive function in male mice. Mol Endocrinol. 1997;11:1636–1650. doi: 10.1210/mend.11.11.0009. [DOI] [PubMed] [Google Scholar]
- 302.Meinhardt A., Bacher M., O’Bryan M.K. A switch in the cellular localization of macrophage migration inhibitory factor in the rat testis after ethane dimethane sulfonate treatment. J Cell Sci. 1999;112:1337–1344. doi: 10.1242/jcs.112.9.1337. [DOI] [PubMed] [Google Scholar]
- 303.Aubry F., Habasque C., Satie A.P., Jégou B., Samson M. Expression and regulation of the CC-chemokine monocyte chemoattractant protein-1 in rat testicular cells in primary culture. Biol Reprod. 2000;62:1427–1435. doi: 10.1095/biolreprod62.5.1427. [DOI] [PubMed] [Google Scholar]
- 304.Hashimoto D., Chow A., Noizat C. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity. 2013;38:792–804. doi: 10.1016/j.immuni.2013.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305.Schlatt S., de Kretser D.M., Hedger M.P. Mitosis of resident macrophages in the adult rat testis. J Reprod Fertil. 1999;116:223–228. doi: 10.1530/jrf.0.1160223. [DOI] [PubMed] [Google Scholar]
- 306.Vergouwen R.P., Jacobs S.G., Huiskamp R., Davids J.A., de Rooij D.G. Proliferative activity of gonocytes, Sertoli cells and interstitial cells during testicular development in mice. J Reprod Fertil. 1991;93:233–243. doi: 10.1530/jrf.0.0930233. [DOI] [PubMed] [Google Scholar]
- 307.Carpenter A.M., Lukyanenko Y.O., Lee V.H., Hutson J.C. FSH does not directly influence testicular macrophages. J Androl. 1998;19:420–427. [PubMed] [Google Scholar]
- 308.Picker L.J., Butcher E.C. Physiological and molecular mechanisms of lymphocyte homing. Annu Rev Immunol. 1992;10:561–591. doi: 10.1146/annurev.iy.10.040192.003021. [DOI] [PubMed] [Google Scholar]
- 309.Kantele A., Zivny J., Hakkinen M., Elson C.O., Mestecky J. Differential homing commitments of antigen-specific T cells after oral or parenteral immunization in humans. J Immunol. 1999;162:5173–5177. [PubMed] [Google Scholar]
- 310.Duan Y.G., Yu C.F., Novak N. Immunodeviation towards a Th17 immune response associated with testicular damage in azoospermic men. Int J Androl. 2011;34:e536–e545. doi: 10.1111/j.1365-2605.2010.01137.x. [DOI] [PubMed] [Google Scholar]
- 311.Doncel G.F., Di Paola J.A., Lustig L. Sequential study of the histopathology and cellular and humoral immune response during the development of an autoimmune orchitis in Wistar rats. Am J Reprod Immunol. 1989;20:44–51. doi: 10.1111/j.1600-0897.1989.tb00638.x. [DOI] [PubMed] [Google Scholar]
- 312.Jahnukainen K., Jørgensen N., Pöllänen P., Giwercman A., Skakkebæk N.E. Incidence of testicular mononuclear cell infiltrates in normal human males and in patients with germ cell neoplasia. Int J Androl. 1995;18:313–320. doi: 10.1111/j.1365-2605.1995.tb00568.x. [DOI] [PubMed] [Google Scholar]
- 313.Mukasa A., Hiromatsu K., Matsuzaki G., O’Brien R., Born W., Nomoto K. Bacterial infection of the testis leading to autoaggressive immunity triggers apparently opposed responses of αβ and γδ T cells. J Immunol. 1995;155:2047–2056. [PubMed] [Google Scholar]
- 314.Kohno S., Munoz J.A., Williams T.M., Teuscher C., Bernard C.C., Tung K.S. Immunopathology of murine experimental allergic orchitis. J Immunol. 1983;130:2675–2682. [PubMed] [Google Scholar]
- 315.Lustig L., Lourtau L., Perez R., Doncel G.F. Phenotypic characterization of lymphocytic cell infiltrates into the testes of rats undergoing autoimmune orchitis. Int J Androl. 1993;16:279–284. doi: 10.1111/j.1365-2605.1993.tb01192.x. [DOI] [PubMed] [Google Scholar]
- 316.Torres A., Casanova J.F., Nistal M., Regadera J. Quantification of immunocompetent cells in testicular germ cell tumours. Histopathology. 1997;30:23–30. doi: 10.1046/j.1365-2559.1997.d01-560.x. [DOI] [PubMed] [Google Scholar]
- 317.Nistal M., Santamaria L., Paniagua R. Mast cells in the human testis and epididymis from birth to adulthood. Acta Anat. 1984;119:155–160. doi: 10.1159/000145878. [DOI] [PubMed] [Google Scholar]
- 318.Meineke V., Frungieri M.B., Jessberger B., Vogt H., Mayerhofer A. Human testicular mast cells contain tryptase: increased mast cell number and altered distribution in the testes of infertile men. Fertil Steril. 2000;74:239–244. doi: 10.1016/s0015-0282(00)00626-9. [DOI] [PubMed] [Google Scholar]
- 319.Welter H., Kohn F.M., Mayerhofer A. Mast cells in human testicular biopsies from patients with mixed atrophy: increased numbers, heterogeneity, and expression of cyclooxygenase 2 and prostaglandin D2 synthase. Fertil Steril. 2011;96:309–313. doi: 10.1016/j.fertnstert.2011.05.035. [DOI] [PubMed] [Google Scholar]
- 320.Sezer C., Koksal I.T., Usta M.F. Relationship between mast cell and iNOS expression in testicular tissue associated with infertility. Arch Androl. 2005;51:149–158. doi: 10.1080/014850190518161. [DOI] [PubMed] [Google Scholar]
- 321.Nistal M., Santana A., Paniaqua R., Palacios J. Testicular toxoplasmosis in two men with the acquired immunodeficiency syndrome (AIDS) Arch Pathol Lab Med. 1986;110:744–746. [PubMed] [Google Scholar]
- 322.O’Bryan M.K., Schlatt S., Phillips D.J., de Kretser D.M., Hedger M.P. Bacterial lipopolysaccharide-induced inflammation compromises testicular function at multiple levels in vivo. Endocrinology. 2000;141:238–246. doi: 10.1210/endo.141.1.7240. [DOI] [PubMed] [Google Scholar]
- 323.Jackson A.E., O’Leary P.C., Ayers M.M., de Kretser D.M. The effects of ethylene dimethane sulphonate (EDS) on rat Leydig cells: evidence to support a connective tissue origin of Leydig cells. Biol Reprod. 1986;35:425–437. doi: 10.1095/biolreprod35.2.425. [DOI] [PubMed] [Google Scholar]
- 324.Gaytan F., Aceitero J., Lucena C. Simultaneous proliferation and differentiation of mast cells and Leydig cells in the rat testis. Are common regulatory factors involved? J Androl. 1992;13:387–397. [PubMed] [Google Scholar]
- 325.Gaytan F., Bellido C., Aceitero J., Aguilar E., Sanchez-Criado J.E. Leydig cell involvement in the paracrine regulation of mast cells in the testicular interstitium of the rat. Biol Reprod. 1990;43:665–671. doi: 10.1095/biolreprod43.4.665. [DOI] [PubMed] [Google Scholar]
- 326.Raeside J.I., Renaud R.L. Estrogen and androgen production by purified Leydig cells of mature boars. Biol Reprod. 1983;28:727–733. doi: 10.1095/biolreprod28.3.727. [DOI] [PubMed] [Google Scholar]
- 327.Eisenhauer K.M., McCue P.M., Nayden D.K., Osawa Y., Roser J.F. Localization of aromatase in equine Leydig cells. Domest Anim Endocrinol. 1994;11:291–298. doi: 10.1016/0739-7240(94)90020-5. [DOI] [PubMed] [Google Scholar]
- 328.Gaytan F., Bellido C., Carrera G., Aguilar E. Differentiation of mast cells during postnatal development of neonatally estrogen-treated rats. Cell Tissue Res. 1990;259:25–31. doi: 10.1007/BF00571426. [DOI] [PubMed] [Google Scholar]
- 329.Maurer M., Echtenacher B., Hültner L. The c-kit ligand, stem cell factor, can enhance innate immunity through effects on mast cells. J Exp Med. 1998;188:2343–2348. doi: 10.1084/jem.188.12.2343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 330.Yan W., Kero J., Huhtaniemi I., Toppari J. Stem cell factor functions as a survival factor for mature Leydig cells and a growth factor for precursor Leydig cells after ethylene dimethane sulfonate treatment: implication of a role of the stem cell factor/c-Kit system in Leydig cell development. Dev Biol. 2000;227:169–182. doi: 10.1006/dbio.2000.9885. [DOI] [PubMed] [Google Scholar]
- 331.Zhou Z.Z., Zheng Y., Steenstra R., Hickey W.F., Teuscher C. Actively-induced experimental allergic orchitis (EAO) in Lewis/NCR rats: sequential histo- and immunopathologic analysis. Autoimmunity. 1989;3:125–134. doi: 10.3109/08916938909019961. [DOI] [PubMed] [Google Scholar]
- 332.Flickinger C.J., Herr J.C., Caloras D., Sisak J.R., Howards S.S. Inflammatory changes in the epididymis after vasectomy in the Lewis rat. Biol Reprod. 1990;43:34–45. doi: 10.1095/biolreprod43.1.34. [DOI] [PubMed] [Google Scholar]
- 333.Barratt C.L., Cohen J. Quantitation of sperm disposal and phagocytic cells in the tract of short- and long-term vasectomized mice. J Reprod Fertil. 1987;81:377–384. doi: 10.1530/jrf.0.0810377. [DOI] [PubMed] [Google Scholar]
- 334.Yakirevich E., Yanai O., Sova Y. Cytotoxic phenotype of intra-epithelial lymphocytes in normal and cryptorchid human testicular excurrent ducts. Hum Reprod. 2002;17:275–283. doi: 10.1093/humrep/17.2.275. [DOI] [PubMed] [Google Scholar]
- 335.Crocker P.R., Jefferies W.A., Clark S.J., Chung L.P., Gordon S. Species heterogeneity in macrophage expression of the CD4 antigen. J Exp Med. 1987;166:613–618. doi: 10.1084/jem.166.2.613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 336.Gibbings D.J., Marcet-Palacios M., Sekar Y., Ng M.C., Befus A.D. CD8α is expressed by human monocytes and enhances FcγR-dependent responses. BMC Immunol. 2007;8:12. doi: 10.1186/1471-2172-8-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 337.Pollard J.W., Dominguez M.G., Mocci S., Cohen P.E., Stanley E.R. Effect of the colony-stimulating factor-1 null mutation, osteopetrotic (csfmop), on the distribution of macrophages in the male mouse reproductive tract. Biol Reprod. 1997;56:1290–1300. doi: 10.1095/biolreprod56.5.1290. [DOI] [PubMed] [Google Scholar]
- 338.Seiler P., Cooper T.G., Yeung C.H., Nieschlag E. Regional variation in macrophage antigen expression by murine epididymal basal cells and their regulation by testicular factors. J Androl. 1999;20:738–746. [PubMed] [Google Scholar]
- 339.Seiler P., Cooper T.G., Nieschlag E. Sperm number and condition affect the number of basal cells and their expression of macrophage antigen in the murine epididymis. Int J Androl. 2000;23:65–76. doi: 10.1046/j.1365-2605.2000.00210.x. [DOI] [PubMed] [Google Scholar]
- 340.Perez-Clavier R., Harrison R.G., Macmillian E.W. The pattern of the lymphatic drainage of the rat epididymis. J Anat. 1982;134:667–675. [PMC free article] [PubMed] [Google Scholar]
- 341.McDonald S.W., Scothorne R.J. The lymphatic drainage of the epididymis and of the ductus deferens of the rat, with reference to the immune response to vasectomy. J Anat. 1988;158:57–64. [PMC free article] [PubMed] [Google Scholar]
- 342.Hirai S., Naito M., Terayama H. Difference in abundance of blood and lymphatic capillaries in the murine epididymis. Med Mol Morphol. 2010;43:37–42. doi: 10.1007/s00795-009-0473-8. [DOI] [PubMed] [Google Scholar]
- 343.Tomlinson M.J., White A., Barratt C.L., Bolton A.E., Cooke I.D. The removal of morphologically abnormal sperm forms by phagocytes: a positive role for seminal leukocytes? Hum Reprod. 1992;7:517–522. doi: 10.1093/oxfordjournals.humrep.a137682. [DOI] [PubMed] [Google Scholar]
- 344.Jones R. Sperm survival versus degradation in the mammalian epididymis: a hypothesis. Biol Reprod. 2004;71:1405–1411. doi: 10.1095/biolreprod.104.031252. [DOI] [PubMed] [Google Scholar]
- 345.Cooper T.G., Yeung C.H., Jones R., Orgebin-Crist M.C., Robaire B. Rebuttal of a role for the epididymis in sperm quality control by phagocytosis of defective sperm. J Cell Sci. 2002;115:5–7. doi: 10.1242/jcs.115.1.5. [DOI] [PubMed] [Google Scholar]
- 346.Hellema H.W., Samuel T., Rümke P. Sperm autoantibodies as a consequence of vasectomy. II. Long-term follow-up studies. Clin Exp Immunol. 1979;38:31–36. [PMC free article] [PubMed] [Google Scholar]
- 347.Alexander N.J., Anderson D.J. Vasectomy: consequences of autoimmunity to sperm antigens. Fertil Steril. 1979;32:253–260. doi: 10.1016/s0015-0282(16)44228-7. [DOI] [PubMed] [Google Scholar]
- 348.de Kretser D.M., Huidobro C., Southwick G.J., Temple-Smith P.D. The role of the epididymis in human infertility. J Reprod Fertil Suppl. 1998;53:271–275. [PubMed] [Google Scholar]
- 349.Herr J.C., Flickinger C.J., Howards S.S. The relation between antisperm antibodies and testicular alterations after vasectomy and vasovasostomy in Lewis rats. Biol Reprod. 1987;37:1297–1305. doi: 10.1095/biolreprod37.5.1297. [DOI] [PubMed] [Google Scholar]
- 350.Flickinger C.J., Herr J.C., Howards S.S. Early testicular changes after vasectomy and vasovasostomy in Lewis rats. Anat Rec. 1990;227:37–46. doi: 10.1002/ar.1092270106. [DOI] [PubMed] [Google Scholar]
- 351.Bigazzi P.E., Kosuda L.L., Harnick L.L. Sperm autoantibodies in vasectomized rats of different inbred strains. Science. 1977;197:1282–1283. doi: 10.1126/science.897668. [DOI] [PubMed] [Google Scholar]
- 352.Flickinger C.J., Herr J.C., Baran M.L., Howards S.S. Temporal appearance of antisperm antibodies during sexual maturation of rats after obstruction of the vas deferens. J Androl. 1995;16:75–79. [PubMed] [Google Scholar]
- 353.Theyer G., Kramer G., Assmann I. Phenotypic characterization of infiltrating leukocytes in benign prostatic hyperplasia. Lab Invest. 1992;66:96–107. [PubMed] [Google Scholar]
- 354.Pudney J., Anderson D.J. Immunobiology of the human penile urethra. Am J Pathol. 1995;147:155–165. [PMC free article] [PubMed] [Google Scholar]
- 355.Husband A.J., Clifton V.L. Role of intestinal immunization in urinary tract defence. Immunol Cell Biol. 1989;67:371–376. doi: 10.1038/icb.1989.53. [DOI] [PubMed] [Google Scholar]
- 356.Quayle A.J., Pudney J., Munoz D.E., Anderson D.J. Characterization of T lymphocytes and antigen-presenting cells in the murine male urethra. Biol Reprod. 1994;51:809–820. doi: 10.1095/biolreprod51.5.809. [DOI] [PubMed] [Google Scholar]
- 357.Bostwick D.G., de la Roza G., Dundore P., Corica F.A., Iczkowski K.A. Intraepithelial and stromal lymphocytes in the normal human prostate. Prostate. 2003;55:187–193. doi: 10.1002/pros.10224. [DOI] [PubMed] [Google Scholar]
- 358.Aitken R.J., Baker H.W. Seminal leukocytes: passengers, terrorists or good samaritans? Hum Reprod. 1995;10:1736–1739. doi: 10.1093/oxfordjournals.humrep.a136165. [DOI] [PubMed] [Google Scholar]
- 359.el-Demiry M.I., Young H., Elton R.A., Hargreave T.B., James K., Chisholm G.D. Leucocytes in the ejaculate from fertile and infertile men. Br J Urol. 1986;58:715–720. doi: 10.1111/j.1464-410x.1986.tb05919.x. [DOI] [PubMed] [Google Scholar]
- 360.el-Demiry M.I., Hargreave T.B., Busuttil A., James K., Chisholm G.D. Identifying leucocytes and leucocyte subpopulations in semen using monoclonal antibody probes. Urology. 1986;28:492–496. doi: 10.1016/0090-4295(86)90150-0. [DOI] [PubMed] [Google Scholar]
- 361.Wolff H., Anderson D.J. Immunohistologic characterization and quantitation of leukocyte subpopulations in human semen. Fertil Steril. 1988;49:497–504. [PubMed] [Google Scholar]
- 362.Tomlinson M.J., Barratt C.L., Cooke I.D. Prospective study of leukocytes and leukocyte subpopulations in semen suggests they are not a cause of male infertility. Fertil Steril. 1993;60:1069–1075. doi: 10.1016/s0015-0282(16)56412-7. [DOI] [PubMed] [Google Scholar]
- 363.Denny T.N., Scolpino A., Garcia A. Evaluation of T-lymphocyte subsets present in semen and peripheral blood of healthy donors: a report from the heterosexual transmission study. Cytometry. 1995;20:349–355. doi: 10.1002/cyto.990200411. [DOI] [PubMed] [Google Scholar]
- 364.Wolff H. The biologic significance of white blood cells in semen. Fertil Steril. 1995;63:1143–1157. doi: 10.1016/s0015-0282(16)57588-8. [DOI] [PubMed] [Google Scholar]
- 365.Anderson D.J., Politch J.A., Martinez A., Van Voorhis B.J., Padian N.S., O’Brien T.R. White blood cells and HIV-1 in semen from vasectomised seropositive men. Lancet. 1991;338:573–574. doi: 10.1016/0140-6736(91)91139-l. [DOI] [PubMed] [Google Scholar]
- 366.Schwartz G.G. Can vasectomy reduce the sexual transmission of HIV? J Clin Epidemiol. 1990;43:1433. doi: 10.1016/0895-4356(90)90115-6. [DOI] [PubMed] [Google Scholar]
- 367.Handelsman D.J. Testicular dysfunction in systemic disease. Endocrinol Metab Clin North Am. 1994;23:839–856. [PubMed] [Google Scholar]
- 368.Khan S.A., Söder O., Syed V., Gustafsson K., Lindh M., Ritzén E.M. The rat testis produces large amounts of an interleukin-1-like factor. Int J Androl. 1987;10:495–503. doi: 10.1111/j.1365-2605.1987.tb00224.x. [DOI] [PubMed] [Google Scholar]
- 369.Gérard N., Syed V., Bardin W., Genetet N., Jégou B. Sertoli cells are the site of interleukin-1α synthesis in rat testis. Mol Cell Endocrinol. 1991;82:R13–R16. doi: 10.1016/0303-7207(91)90019-o. [DOI] [PubMed] [Google Scholar]
- 370.Dinarello C.A. Biologic basis for interleukin-1 in disease. Blood. 1996;87:2095–2147. [PubMed] [Google Scholar]
- 371.Watanabe N., Kobayashi Y. Selective release of a processed form of interleukin 1α. Cytokine. 1994;6:597–601. doi: 10.1016/1043-4666(94)90046-9. [DOI] [PubMed] [Google Scholar]
- 372.Abu Elheija M., Dyomin V., Ganaiem M., Lunenfeld E., Vardy N.S., Huleihel M. Distinct expression of interleukin-1α, interleukin-1β, and interleukin-1 receptor antagonist in testicular tissues and cells from human biopsies with normal and abnormal histology. J Interferon Cytokine Res. 2011;31:401–408. doi: 10.1089/jir.2010.0059. [DOI] [PubMed] [Google Scholar]
- 373.Jonsson C.K., Zetterström R.H., Holst M., Parvinen M., Söder O. Constitutive expression of interleukin-1α messenger ribonucleic acid in rat Sertoli cells is dependent upon interaction with germ cells. Endocrinology. 1999;140:3755–3761. doi: 10.1210/endo.140.8.6900. [DOI] [PubMed] [Google Scholar]
- 374.Syed V., Söder O., Arver S., Lindh M., Khan S., Ritzén E.M. Ontogeny and cellular origin of an interleukin-1-like factor in the reproductive tract of the male rat. Int J Androl. 1988;11:437–447. doi: 10.1111/j.1365-2605.1988.tb01016.x. [DOI] [PubMed] [Google Scholar]
- 375.Syed V., Stéphan J.P., Gérard N. Residual bodies activate Sertoli cell interleukin-1α (IL-1α) release, which triggers IL-6 production by an autocrine mechanism, through the lipoxygenase pathway. Endocrinology. 1995;136:3070–3078. doi: 10.1210/endo.136.7.7789334. [DOI] [PubMed] [Google Scholar]
- 376.Gérard N., Syed V., Jégou B. Lipopolysaccharide, latex beads and residual bodies are potent activators of Sertoli cell interleukin-1α production. Biochem Biophys Res Commun. 1992;185:154–161. doi: 10.1016/s0006-291x(05)80969-6. [DOI] [PubMed] [Google Scholar]
- 377.Sultana T., Svechnikov K., Weber G., Söder O. Molecular cloning and expression of a functionally different alternative splice variant of prointerleukin-1α from the rat testis. Endocrinology. 2000;141:4413–4418. doi: 10.1210/endo.141.12.7824. [DOI] [PubMed] [Google Scholar]
- 378.Sultana T., Wahab-Wahlgren A., Assmus M., Parvinen M., Weber G., Söder O. Expression and regulation of the prointerleukin-1α processing enzymes calpain I and II in the rat testis. Int J Androl. 2003;26:37–45. doi: 10.1046/j.1365-2605.2003.00386.x. [DOI] [PubMed] [Google Scholar]
- 379.Winnall W.R., Okuma Y., Saito K., Muir J.A., Hedger M.P. Regulation of interleukin 1α, activin and inhibin by lipopolysaccharide in Sertoli cells from prepubertal rats. Mol Cell Endocrinol. 2009;307:169–175. doi: 10.1016/j.mce.2009.02.007. [DOI] [PubMed] [Google Scholar]
- 380.Svechnikov K.V., Sultana T., Söder O. Age-dependent stimulation of Leydig cell steroidogenesis by interleukin-1 isoforms. Mol Cell Endocrinol. 2001;182:193–201. doi: 10.1016/s0303-7207(01)00554-8. [DOI] [PubMed] [Google Scholar]
- 381.Haugen T.B., Landmark B.F., Josefsen G.M., Hansson V., Högset A. The mature form of interleukin-1α is constitutively expressed in immature male germ cells from rat. Mol Cell Endocrinol. 1994;105:R19–R23. doi: 10.1016/0303-7207(94)90177-5. [DOI] [PubMed] [Google Scholar]
- 382.Hakovirta H., Pentitilä T.L., Pöllänen P., Fröysa B., Söder O., Parvinen M. Interleukin-1 bioactivity and DNA synthesis in X-irradiated rat testes. Int J Androl. 1993;16:159–164. doi: 10.1111/j.1365-2605.1993.tb01170.x. [DOI] [PubMed] [Google Scholar]
- 383.Parvinen M., Söder O., Mali P., Fröysa B., Ritzén E.M. In vitro stimulation of stage-specific deoxyribonucleic acid synthesis in rat seminiferous tubule segments by interleukin-1α. Endocrinology. 1991;129:1614–1620. doi: 10.1210/endo-129-3-1614. [DOI] [PubMed] [Google Scholar]
- 384.Söder O., Syed V., Callard G.V. Production and secretion of an interleukin-1-like factor is stage-dependent and correlates with spermatogonial DNA synthesis in the rat seminiferous epithelium. Int J Androl. 1991;14:223–231. doi: 10.1111/j.1365-2605.1991.tb01084.x. [DOI] [PubMed] [Google Scholar]
- 385.Wang J.E., Josefsen G.M., Hansson V., Haugen T.B. Residual bodies and IL-1α stimulate expression of mRNA for IL-1α and IL-1 receptor type I in cultured rat Sertoli cells. Mol Cell Endocrinol. 1998;137:139–144. doi: 10.1016/s0303-7207(97)00246-3. [DOI] [PubMed] [Google Scholar]
- 386.Cudicini C., Kercret H., Touzalin A.M., Ballet F., Jégou B. Vectorial production of interleukin 1 and interleukin 6 by rat Sertoli cells cultured in a dual culture compartment system. Endocrinology. 1997;138:2863–2870. doi: 10.1210/endo.138.7.5289. [DOI] [PubMed] [Google Scholar]
- 387.Stéphan J.P., Syed V., Jégou B. Regulation of Sertoli cell IL-1 and IL-6 production in vitro. Mol Cell Endocrinol. 1997;134:109–118. doi: 10.1016/s0303-7207(97)00172-x. [DOI] [PubMed] [Google Scholar]
- 388.Winnall W.R., Muir J.A., Hedger M.P. Differential responses of epithelial Sertoli cells of the rat testis to Toll-like receptor 2 and 4 ligands: implications for studies of testicular inflammation using bacterial lipopolysaccharides. Innate Immun. 2011;17:123–136. doi: 10.1177/1753425909354764. [DOI] [PubMed] [Google Scholar]
- 389.Okuma Y., O’Connor A.E., Muir J.A., Stanton P.G., de Kretser D.M., Hedger M.P. Regulation of activin A and inhibin B secretion by inflammatory mediators in adult rat Sertoli cell cultures. J Endocrinol. 2005;187:125–134. doi: 10.1677/joe.1.06266. [DOI] [PubMed] [Google Scholar]
- 390.Cudicini C., Lejeune H., Gomez E. Human Leydig cells and Sertoli cells are producers of interleukins-1 and -6. J Clin Endocrinol Metab. 1997;82:1426–1433. doi: 10.1210/jcem.82.5.3938. [DOI] [PubMed] [Google Scholar]
- 391.Lin T., Wang D., Nagpal M.L. Human chorionic gonadotropin induces interleukin-1 gene expression in rat Leydig cells in vivo. Mol Cell Endocrinol. 1993;95:139–145. doi: 10.1016/0303-7207(93)90039-m. [DOI] [PubMed] [Google Scholar]
- 392.Wang D.L., Nagpal M.L., Calkins J.H., Chang W.W., Sigel M.M., Lin T. Interleukin-1β induces interleukin-1α messenger ribonucleic acid expression in primary cultures of Leydig cells. Endocrinology. 1991;129:2862–2866. doi: 10.1210/endo-129-6-2862. [DOI] [PubMed] [Google Scholar]
- 393.Gow R.M., O’Bryan M.K., Canny B.J., Ooi G.T., Hedger M.P. Differential effects of dexamethasone treatment on lipopolysaccharide-induced testicular inflammation and reproductive hormone inhibition in adult rats. J Endocrinol. 2001;168:193–201. doi: 10.1677/joe.0.1680193. [DOI] [PubMed] [Google Scholar]
- 394.Xiong Y., Hales D.B. Immune-endocrine interactions in the mouse testis: cytokine-mediated inhibition of Leydig cell steroidogenesis. Endocr J. 1994;2:223–228. [Google Scholar]
- 395.Hales D.B., Xiong Y., Tur-Kaspa I. The role of cytokines in the regulation of Leydig cell P450c17 gene expression. J Steroid Biochem Mol Biol. 1992;43:907–914. doi: 10.1016/0960-0760(92)90318-D. [DOI] [PubMed] [Google Scholar]
- 396.Jonsson C.K., Setchell B.P., Martinelle N., Svechnikov K., Söder O. Endotoxin-induced interleukin 1 expression in testicular macrophages is accompanied by downregulation of the constitutive expression in Sertoli cells. Cytokine. 2001;14:283–288. doi: 10.1006/cyto.2001.0878. [DOI] [PubMed] [Google Scholar]
- 397.Hedger M., Klug J., Fröhlich S., Müller R., Meinhardt A. Regulatory cytokine expression and interstitial fluid formation in the normal and inflamed rat testis are under Leydig cell control. J Androl. 2005;26:379–386. doi: 10.2164/jandrol.04149. [DOI] [PubMed] [Google Scholar]
- 398.Kenny E.F., O’Neill L.A. Signalling adaptors used by Toll-like receptors: an update. Cytokine. 2008;43:342–349. doi: 10.1016/j.cyto.2008.07.010. [DOI] [PubMed] [Google Scholar]
- 399.Sims J.E., Gayle M.A., Slack J.L. Interleukin 1 signaling occurs exclusively via the type I receptor. Proc Natl Acad Sci USA. 1993;90:6155–6159. doi: 10.1073/pnas.90.13.6155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 400.Khan S.A., Khan S.J., Dorrington J.H. Interleukin-1 stimulates deoxyribonucleic acid synthesis in immature rat Leydig cells in vitro. Endocrinology. 1992;131:1853–1857. doi: 10.1210/endo.131.4.1396331. [DOI] [PubMed] [Google Scholar]
- 401.Walch L., Morris P.L. Cyclooxygenase 2 pathway mediates IL-1β regulation of IL-1α, -1β, and IL-6 mRNA levels in Leydig cell progenitors. Endocrinology. 2002;143:3276–3283. doi: 10.1210/en.2002-220410. [DOI] [PubMed] [Google Scholar]
- 402.Calkins J.H., Sigel M.M., Nankin H.R., Lin T. Interleukin-1 inhibits Leydig cell steroidogenesis in primary culture. Endocrinology. 1988;123:1605–1610. doi: 10.1210/endo-123-3-1605. [DOI] [PubMed] [Google Scholar]
- 403.Hales D.B. Interleukin-1 inhibits Leydig cell steroidogenesis primarily by decreasing 17α-hydroxylase/C17-20 lyase cytochrome P450 expression. Endocrinology. 1992;131:2165–2172. doi: 10.1210/endo.131.5.1425417. [DOI] [PubMed] [Google Scholar]
- 404.Lin T., Wang T.L., Nagpal M.L., Calkins J.H., Chang W.W., Chi R. Interleukin-1 inhibits cholesterol side-chain cleavage cytochrome P450 expression in primary cultures of Leydig cells. Endocrinology. 1991;129:1305–1311. doi: 10.1210/endo-129-3-1305. [DOI] [PubMed] [Google Scholar]
- 405.Sun X.R., Hedger M.P., Risbridger G.P. The effect of testicular macrophages and interleukin-1 on testosterone production by purified adult rat Leydig cells cultured under in vitro maintenance conditions. Endocrinology. 1993;132:186–192. doi: 10.1210/endo.132.1.8419122. [DOI] [PubMed] [Google Scholar]
- 406.Verhoeven G., Cailleau J., Van Damme J., Billiau A. Interleukin-1 stimulates steroidogenesis in cultured rat Leydig cells. Mol Cell Endocrinol. 1988;57:51–60. doi: 10.1016/0303-7207(88)90031-7. [DOI] [PubMed] [Google Scholar]
- 407.Moore C., Moger W.H. Interleukin-1α-induced changes in androgen and cyclic adenosine 3′,5′-monophosphate release in adult rat Leydig cells in culture. J Endocrinol. 1991;129:381–390. doi: 10.1677/joe.0.1290381. [DOI] [PubMed] [Google Scholar]
- 408.Watson M.E., Newman R.J., Payne A.M., Abdelrahim M., Francis G.L. The effect of macrophage conditioned media on Leydig cell function. Ann Clin Lab Sci. 1994;24:84–95. [PubMed] [Google Scholar]
- 409.Hales D.B. Testicular macrophage modulation of Leydig cell steroidogenesis. J Reprod Immunol. 2002;57:3–18. doi: 10.1016/s0165-0378(02)00020-7. [DOI] [PubMed] [Google Scholar]
- 410.Xiong Y., Hales D.B. Differential effects of tumor necrosis factor-α and interleukin-1 on 3 β-hydroxysteroid dehydrogenase/Δ5 --> Δ4 isomerase expression in mouse Leydig cells. Endocrine. 1997;7:295–301. doi: 10.1007/BF02801322. [DOI] [PubMed] [Google Scholar]
- 411.Lin T., Wang D., Stocco D.M. Interleukin-1 inhibits Leydig cell steroidogenesis without affecting steroidogenic acute regulatory protein messenger ribonucleic acid or protein levels. J Endocrinol. 1998;156:461–467. doi: 10.1677/joe.0.1560461. [DOI] [PubMed] [Google Scholar]
- 412.Gerendai I., Banczerowski P., Csernus V. Interleukin 1-β injected into the testis acutely stimulates and later attenuates testicular steroidogenesis of the immature rat. Endocrine. 2005;28:165–170. doi: 10.1385/ENDO:28:2:165. [DOI] [PubMed] [Google Scholar]
- 413.Wang X., Dyson M.T., Jo Y., Stocco D.M. Inhibition of cyclooxygenase-2 activity enhances steroidogenesis and steroidogenic acute regulatory gene expression in MA-10 mouse Leydig cells. Endocrinology. 2003;144:3368–3375. doi: 10.1210/en.2002-0081. [DOI] [PubMed] [Google Scholar]
- 414.Renlund N., Jo Y., Svechnikova I. Induction of steroidogenesis in immature rat Leydig cells by interleukin-1α is dependent on extracellular signal-regulated kinases. J Mol Endocrinol. 2006;36:327–336. doi: 10.1677/jme.1.01963. [DOI] [PubMed] [Google Scholar]
- 415.Gomez E., Morel G., Cavalier A. Type I and type II interleukin-1 receptor expression in rat, mouse, and human testes. Biol Reprod. 1997;56:1513–1526. doi: 10.1095/biolreprod56.6.1513. [DOI] [PubMed] [Google Scholar]
- 416.Pöllänen P., Söder O., Parvinen M. Interleukin-1α stimulation of spermatogonial proliferation in vivo. Reprod Fertil Dev. 1989;1:85–87. doi: 10.1071/rd9890085. [DOI] [PubMed] [Google Scholar]
- 417.Petersen C., Svechnikov K., Fröysa B., Söder O. The p38 MAPK pathway mediates interleukin-1-induced Sertoli cell proliferation. Cytokine. 2005;32:51–59. doi: 10.1016/j.cyto.2005.07.014. [DOI] [PubMed] [Google Scholar]
- 418.Khan S.A., Nieschlag E. Interleukin-1 inhibits follitropin-induced aromatase activity in immature rat Sertoli cells in vitro. Mol Cell Endocrinol. 1991;75:1–7. doi: 10.1016/0303-7207(91)90238-n. [DOI] [PubMed] [Google Scholar]
- 419.Nehar D., Mauduit C., Boussouar F., Benahmed M. Interleukin 1α stimulates lactate dehydrogenase A expression and lactate production in cultured porcine Sertoli cells. Biol Reprod. 1998;59:1425–1432. doi: 10.1095/biolreprod59.6.1425. [DOI] [PubMed] [Google Scholar]
- 420.Hoeben E., Van Damme J., Put W., Swinnen J.V., Verhoeven G. Cytokines derived from activated human mononuclear cells markedly stimulate transferrin secretion by cultured Sertoli cells. Endocrinology. 1996;137:514–521. doi: 10.1210/endo.137.2.8593797. [DOI] [PubMed] [Google Scholar]
- 421.Galardo M.N., Riera M.F., Pellizzari E.H. Regulation of expression of Sertoli cell glucose transporters 1 and 3 by FSH, IL1β, and bFGF at two different time-points in pubertal development. Cell Tissue Res. 2008;334:295–304. doi: 10.1007/s00441-008-0656-y. [DOI] [PubMed] [Google Scholar]
- 422.Okuma Y., Saito K., O’Connor A.E., Phillips D.J., de Kretser D.M., Hedger M.P. Reciprocal regulation of activin A and inhibin B by interleukin-1 (IL-1) and follicle-stimulating hormone (FSH) in rat Sertoli cells in vitro. J Endocrinol. 2005;185:99–110. doi: 10.1677/joe.1.06053. [DOI] [PubMed] [Google Scholar]
- 423.Lie P.P., Cheng C.Y., Mruk D.D. Interleukin-1α is a regulator of the blood-testis barrier. FASEB J. 2011;25:1244–1253. doi: 10.1096/fj.10-169995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 424.Ishikawa T., Hwang K., Lazzarino D., Morris P.L. Sertoli cell expression of steroidogenic acute regulatory protein-related lipid transfer 1 and 5 domain-containing proteins and sterol regulatory element binding protein-1 are interleukin-1β regulated by activation of c-Jun N-terminal kinase and cyclooxygenase-2 and cytokine induction. Endocrinology. 2005;146:5100–5111. doi: 10.1210/en.2005-0567. [DOI] [PubMed] [Google Scholar]
- 425.Ishikawa T., Morris P.L. A multistep kinase-based Sertoli cell autocrine-amplifying loop regulates prostaglandins, their receptors, and cytokines. Endocrinology. 2006;147:1706–1716. doi: 10.1210/en.2005-1576. [DOI] [PubMed] [Google Scholar]
- 426.Ishikawa T., Morris P.L. Interleukin-1β signals through a c-Jun N-terminal kinase-dependent inducible nitric oxide synthase and nitric oxide production pathway in Sertoli epithelial cells. Endocrinology. 2006;147:5424–5430. doi: 10.1210/en.2006-0643. [DOI] [PubMed] [Google Scholar]
- 427.Riera M.F., Galardo M.N., Pellizzari E.H., Meroni S.B., Cigorraga S.B. Participation of phosphatidyl inositol 3-kinase/protein kinase B and ERK1/2 pathways in interleukin-1β stimulation of lactate production in Sertoli cells. Reproduction. 2007;133:763–773. doi: 10.1530/rep.1.01091. [DOI] [PubMed] [Google Scholar]
- 428.Dunn E., Sims J.E., Nicklin M.J., O’Neill L.A. Annotating genes with potential roles in the immune system: six new members of the IL-1 family. Trends Immunol. 2001;22:533–536. doi: 10.1016/s1471-4906(01)02034-8. [DOI] [PubMed] [Google Scholar]
- 429.Gracie J.A., Robertson S.E., McInnes I.B. Interleukin-18. J Leuk Biol. 2003;73:213–224. doi: 10.1189/jlb.0602313. [DOI] [PubMed] [Google Scholar]
- 430.Strand M.L., Wahlgren A., Svechnikov K., Zetterström C., Setchell B.P., Söder O. Interleukin-18 is expressed in rat testis and may promote germ cell growth. Mol Cell Endocrinol. 2005;240:64–73. doi: 10.1016/j.mce.2005.05.011. [DOI] [PubMed] [Google Scholar]
- 431.Abu Elhija M., Lunenfeld E., Eldar-Geva T., Huleihel M. Lipopolysaccharide increased the expression levels of IL-18, ICE and IL-18 R in murine Leydig cells. Am J Reprod Immunol. 2008;60:151–159. doi: 10.1111/j.1600-0897.2008.00607.x. [DOI] [PubMed] [Google Scholar]
- 432.Komsky A., Huleihel M., Ganaiem M. Presence of IL-18 in testicular tissue of fertile and infertile men. Andrologia. 2012;44:1–8. doi: 10.1111/j.1439-0272.2010.01090.x. [DOI] [PubMed] [Google Scholar]
- 433.Arend W.P. Interleukin-1 receptor antagonist. Adv Immunol. 1993;54:167–227. doi: 10.1016/s0065-2776(08)60535-0. [DOI] [PubMed] [Google Scholar]
- 434.Zeyse D., Lunenfeld E., Beck M., Prinsloo I., Huleihel M. Interleukin-1 receptor antagonist is produced by Sertoli cells in vitro. Endocrinology. 2000;141:1521–1527. doi: 10.1210/endo.141.4.7401. [DOI] [PubMed] [Google Scholar]
- 435.Ganaiem M., AbuElhija M., Lunenfeld E. Effect of interleukin-1 receptor antagonist gene deletion on male mouse fertility. Endocrinology. 2009;150:295–303. doi: 10.1210/en.2008-0848. [DOI] [PubMed] [Google Scholar]
- 436.Bhushan S., Tchatalbachev S., Klug J. Uropathogenic Escherichia coli block MyD88-dependent and activate MyD88-independent signaling pathways in rat testicular cells. J Immunol. 2008;180:5537–5547. doi: 10.4049/jimmunol.180.8.5537. [DOI] [PubMed] [Google Scholar]
- 437.Riccioli A., Starace D., Galli R. Sertoli cells initiate testicular innate immune responses through TLR activation. J Immunol. 2006;177:7122–7130. doi: 10.4049/jimmunol.177.10.7122. [DOI] [PubMed] [Google Scholar]
- 438.Starace D., Galli R., Paone A. Toll-like receptor 3 activation induces antiviral immune responses in mouse Sertoli cells. Biol Reprod. 2008;79:766–775. doi: 10.1095/biolreprod.108.068619. [DOI] [PubMed] [Google Scholar]
- 439.Wu H., Wang H., Xiong W., Chen S., Tang H., Han D. Expression patterns and functions of Toll-like receptors in mouse Sertoli cells. Endocrinology. 2008;149:4402–4412. doi: 10.1210/en.2007-1776. [DOI] [PubMed] [Google Scholar]
- 440.Sun B., Qi N., Shang T., Wu H., Deng T., Han D. Sertoli cell-initiated testicular innate immune response through toll-like receptor-3 activation is negatively regulated by Tyro3, Axl, and mer receptors. Endocrinology. 2010;151:2886–2897. doi: 10.1210/en.2009-1498. [DOI] [PubMed] [Google Scholar]
- 441.Palladino M.A., Johnson T.A., Gupta R., Chapman J.L., Ojha P. Members of the Toll-like receptor family of innate immunity pattern-recognition receptors are abundant in the male rat reproductive tract. Biol Reprod. 2007;76:958–964. doi: 10.1095/biolreprod.106.059410. [DOI] [PubMed] [Google Scholar]
- 442.Palladino M.A., Savarese M.A., Chapman J.L., Dughi M.K., Plaska D. Localization of Toll-like receptors on epididymal epithelial cells and spermatozoa. Am J Reprod Immunol. 2008;60:541–555. doi: 10.1111/j.1600-0897.2008.00654.x. [DOI] [PubMed] [Google Scholar]
- 443.Rodrigues A., Queiróz D.B., Honda L., Silva E.J., Hall S.H., Avellar M.C. Activation of toll-like receptor 4 (TLR4) by in vivo and in vitro exposure of rat epididymis to lipopolysaccharide from Escherichia coli. Biol Reprod. 2008;79:1135–1147. doi: 10.1095/biolreprod.108.069930. [DOI] [PubMed] [Google Scholar]
- 444.Zhao Y.T., Guo J.H., Wu Z.L., Xiong Y., Zhou W.L. Innate immune responses of epididymal epithelial cells to Staphylococcus aureus infection. Immunol Lett. 2008;119:84–90. doi: 10.1016/j.imlet.2008.05.002. [DOI] [PubMed] [Google Scholar]
- 445.Kummer J.A., Broekhuizen R., Everett H. Inflammasome components NALP 1 and 3 show distinct but separate expression profiles in human tissues suggesting a site-specific role in the inflammatory response. J Histochem Cytochem. 2007;55:443–452. doi: 10.1369/jhc.6A7101.2006. [DOI] [PubMed] [Google Scholar]
- 446.Lech M., Avila-Ferrufino A., Skuginna V., Susanti H.E., Anders H.J. Quantitative expression of RIG-like helicase, NOD-like receptor and inflammasome-related mRNAs in humans and mice. Int Immunol. 2010;22:717–728. doi: 10.1093/intimm/dxq058. [DOI] [PubMed] [Google Scholar]
- 447.Hirschfeld M., Ma Y., Weis J.H., Vogel S.N., Weis J.J. Cutting edge: repurification of lipopolysaccharide eliminates signaling through both human and murine Toll-like receptor 2. J Immunol. 2000;165:618–622. doi: 10.4049/jimmunol.165.2.618. [DOI] [PubMed] [Google Scholar]
- 448.Bourdon V., Defamie N., Fenichel P., Pointis G. Regulation of tissue-type plasminogen activator and its inhibitor (PAI-1) by lipopolysaccharide-induced phagocytosis in a Sertoli cell line. Exp Cell Res. 1999;247:367–372. doi: 10.1006/excr.1998.4369. [DOI] [PubMed] [Google Scholar]
- 449.Aly H.A., Lightfoot D.A., El-Shemy H.A. Bacterial lipopolysaccharide-induced oxidative stress in adult rat Sertoli cells in vitro. Toxicol In Vitro. 2010;24:1266–1272. doi: 10.1016/j.tiv.2010.01.009. [DOI] [PubMed] [Google Scholar]
- 450.Shang T., Zhang X., Wang T., Sun B., Deng T., Han D. Toll-like receptor-initiated testicular innate immune responses in mouse Leydig cells. Endocrinology. 2011;152:2827–2836. doi: 10.1210/en.2011-0031. [DOI] [PubMed] [Google Scholar]
- 451.Wang T., Zhang X., Chen Q. Toll-like receptor 3-initiated antiviral responses in mouse male germ cells in vitro. Biol Reprod. 2012;86:106. doi: 10.1095/biolreprod.111.096719. [DOI] [PubMed] [Google Scholar]
- 452.Delfino F., Walker W.H. Stage-specific nuclear expression of NF-κB in mammalian testis. Mol Endocrinol. 1998;12:1696–1707. doi: 10.1210/mend.12.11.0194. [DOI] [PubMed] [Google Scholar]
- 453.Delfino F.J., Boustead J.N., Fix C., Walker W.H. NF-κB and TNF-α stimulate androgen receptor expression in Sertoli cells. Mol Cell Endocrinol. 2003;201:1–12. doi: 10.1016/s0303-7207(03)00005-4. [DOI] [PubMed] [Google Scholar]
- 454.Mizuno K., Hayashi Y., Kojima Y., Nakane A., Tozawa K., Kohri K. Activation of NF-κB associated with germ cell apoptosis in testes of experimentally induced cryptorchid rat model. Urology. 2009;73:389–393. doi: 10.1016/j.urology.2008.09.019. [DOI] [PubMed] [Google Scholar]
- 455.Rasoulpour R.J., Boekelheide K. NF-κB is activated in the rat testis following exposure to mono-(2-ethylhexyl) phthalate. Biol Reprod. 2005;72:479–486. doi: 10.1095/biolreprod.104.034363. [DOI] [PubMed] [Google Scholar]
- 456.Pentikäinen V., Suomalainen L., Erkkilä K. Nuclear factor-κB activation in human testicular apoptosis. Am J Pathol. 2002;160:205–218. doi: 10.1016/s0002-9440(10)64364-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 457.Lysiak J.J., Bang H.J., Nguyen Q.A., Turner T.T. Activation of the nuclear factor kappa B pathway following ischemia-reperfusion of the murine testis. J Androl. 2005;26:129–135. [PubMed] [Google Scholar]
- 458.Bradley J.R. TNF-mediated inflammatory disease. J Pathol. 2008;214:149–160. doi: 10.1002/path.2287. [DOI] [PubMed] [Google Scholar]
- 459.Mak T.W., Yeh W.C. Signaling for survival and apoptosis in the immune system. Arthritis Res. 2002;4(Suppl. 3):S243–S252. doi: 10.1186/ar569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 460.Chung J.Y., Park Y.C., Ye H., Wu H. All TRAFs are not created equal: common and distinct molecular mechanisms of TRAF-mediated signal transduction. J Cell Sci. 2002;115:679–688. doi: 10.1242/jcs.115.4.679. [DOI] [PubMed] [Google Scholar]
- 461.De S.K., Chen H.L., Pace J.L., Hunt J.S., Terranova P.F., Enders G.C. Expression of tumor necrosis factor-α in mouse spermatogenic cells. Endocrinology. 1993;133:389–396. doi: 10.1210/endo.133.1.8319585. [DOI] [PubMed] [Google Scholar]
- 462.Li M.W., Xia W., Mruk D.D. Tumor necrosis factor α reversibly disrupts the blood-testis barrier and impairs Sertoli-germ cell adhesion in the seminiferous epithelium of adult rat testes. J Endocrinol. 2006;190:313–329. doi: 10.1677/joe.1.06781. [DOI] [PubMed] [Google Scholar]
- 463.Moore C., Hutson J.C. Physiological relevance of tumor necrosis factor in mediating macrophage-Leydig cell interactions. Endocrinology. 1994;134:63–69. doi: 10.1210/endo.134.1.8275970. [DOI] [PubMed] [Google Scholar]
- 464.Xiong Y., Hales D.B. Expression, regulation, and production of tumor necrosis factor-α in mouse testicular interstitial macrophages in vitro. Endocrinology. 1993;133:2568–2573. doi: 10.1210/endo.133.6.8243279. [DOI] [PubMed] [Google Scholar]
- 465.Mealy K., Robinson B., Millette C.F., Majzoub J., Wilmore D.W. The testicular effects of tumor necrosis factor. Ann Surg. 1990;211:470–475. doi: 10.1097/00000658-199004000-00014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 466.Morales V., Santana P., Diaz R. Intratesticular delivery of tumor necrosis factor-α and ceramide directly abrogates steroidogenic acute regulatory protein expression and Leydig cell steroidogenesis in adult rats. Endocrinology. 2003;144:4763–4772. doi: 10.1210/en.2003-0569. [DOI] [PubMed] [Google Scholar]
- 467.Hong C.Y., Park J.H., Ahn R.S. Molecular mechanism of suppression of testicular steroidogenesis by proinflammatory cytokine tumor necrosis factor α. Mol Cell Biol. 2004;24:2593–2604. doi: 10.1128/MCB.24.7.2593-2604.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 468.Xiong Y., Hales D.B. The role of tumor necrosis factor-α in the regulation of mouse Leydig cell steroidogenesis. Endocrinology. 1993;132:2438–2444. doi: 10.1210/endo.132.6.8504748. [DOI] [PubMed] [Google Scholar]
- 469.Meroni S.B., Pellizzari E.H., Cánepa D.F., Cigorraga S.B. Possible involvement of ceramide in the regulation of rat Leydig cell function. J Steroid Biochem Mol Biol. 2000;75:307–313. doi: 10.1016/s0960-0760(00)00188-6. [DOI] [PubMed] [Google Scholar]
- 470.Li X., Youngblood G.L., Payne A.H., Hales D.B. Tumor necrosis factor-α inhibition of 17α-hydroxylase/C17-20 lyase gene (Cyp17) expression. Endocrinology. 1995;136:3519–3526. doi: 10.1210/endo.136.8.7628389. [DOI] [PubMed] [Google Scholar]
- 471.Mauduit C., Gasnier F., Rey C. Tumor necrosis factor-α inhibits leydig cell steroidogenesis through a decrease in steroidogenic acute regulatory protein expression. Endocrinology. 1998;139:2863–2868. doi: 10.1210/endo.139.6.6077. [DOI] [PubMed] [Google Scholar]
- 472.Li X., Hales K.H., Watanabe G., Lee R.J., Pestell R.G., Hales D.B. The effect of tumor necrosis factor-α and cAMP on induction of AP-1 activity in MA-10 tumor Leydig cells. Endocrine. 1997;6:317–324. doi: 10.1007/BF02820509. [DOI] [PubMed] [Google Scholar]
- 473.Suh J.H., Gong E.Y., Hong C.Y. Reduced testicular steroidogenesis in tumor necrosis factor-α knockout mice. J Steroid Biochem Mol Biol. 2008;112:117–121. doi: 10.1016/j.jsbmb.2008.09.003. [DOI] [PubMed] [Google Scholar]
- 474.Starace D., Riccioli A., D’Alessio A. Characterization of signaling pathways leading to Fas expression induced by TNF-α: pivotal role of NF-κB. FASEB J. 2005;19:473–475. doi: 10.1096/fj.04-2726fje. [DOI] [PubMed] [Google Scholar]
- 475.Pentikäinen V., Erkkilä K., Suomalainen L. TNFα down-regulates the Fas ligand and inhibits germ cell apoptosis in the human testis. J Clin Endocrinol Metab. 2001;86:4480–4488. doi: 10.1210/jcem.86.9.7861. [DOI] [PubMed] [Google Scholar]
- 476.Suominen J.S., Wang Y., Kaipia A., Toppari J. Tumor necrosis factor-alpha (TNF-α) promotes cell survival during spermatogenesis, and this effect can be blocked by infliximab, a TNF-α antagonist. Eur J Endocrinol. 2004;151:629–640. doi: 10.1530/eje.0.1510629. [DOI] [PubMed] [Google Scholar]
- 477.De Cesaris P., Starace D., Riccioli A., Padula F., Filippini A., Ziparo E. Tumor necrosis factor-α induces interleukin-6 production and integrin ligand expression by distinct transduction pathways. J Biol Chem. 1998;273:7566–7571. doi: 10.1074/jbc.273.13.7566. [DOI] [PubMed] [Google Scholar]
- 478.Schell C., Albrecht M., Mayer C., Schwarzer J.U., Frungieri M.B., Mayerhofer A. Exploring human testicular peritubular cells: identification of secretory products and regulation by tumor necrosis factor-α. Endocrinology. 2008;149:1678–1686. doi: 10.1210/en.2007-1064. [DOI] [PubMed] [Google Scholar]
- 479.Nehar D., Mauduit C., Boussouar F., Benahmed M. Tumor necrosis factor-α-stimulated lactate production is linked to lactate dehydrogenase A expression and activity increase in porcine cultured Sertoli cells. Endocrinology. 1997;138:1964–1971. doi: 10.1210/endo.138.5.5098. [DOI] [PubMed] [Google Scholar]
- 480.Riera M.F., Meroni S.B., Gómez G.E., Schteingart H.F., Pellizzari E.H., Cigorraga S.B. Regulation of lactate production by FSH, IL1β, and TNFα in rat Sertoli cells. Gen Comp Endocrinol. 2001;122:88–97. doi: 10.1006/gcen.2001.7619. [DOI] [PubMed] [Google Scholar]
- 481.Saito K., Winnall W.R., Muir J.A., Hedger M.P. Regulation of Sertoli cell activin A and inhibin B by tumour necrosis factor α and interleukin 1α: interaction with follicle-stimulating hormone/adenosine 3′,5′-cyclic phosphate signalling. Mol Cell Endocrinol. 2011;335:195–203. doi: 10.1016/j.mce.2011.01.014. [DOI] [PubMed] [Google Scholar]
- 482.Mauduit C., Besset V., Caussanel V., Benahmed M. Tumor necrosis factor α receptor p55 is under hormonal (follicle-stimulating hormone) control in testicular Sertoli cells. Biochem Biophys Res Commun. 1996;224:631–637. doi: 10.1006/bbrc.1996.1077. [DOI] [PubMed] [Google Scholar]
- 483.Le Magueresse-Battistoni B., Pernod G., Kolodié L., Morera A.M., Benahmed M. Tumor necrosis factor-α regulates plasminogen activator inhibitor-1 in rat testicular peritubular cells. Endocrinology. 1997;138:1097–1105. doi: 10.1210/endo.138.3.4963. [DOI] [PubMed] [Google Scholar]
- 484.Siu M.K., Lee W.M., Cheng C.Y. The interplay of collagen IV, tumor necrosis factor-α, gelatinase B (matrix metalloprotease-9), and tissue inhibitor of metalloproteases-1 in the basal lamina regulates Sertoli cell-tight junction dynamics in the rat testis. Endocrinology. 2003;144:371–387. doi: 10.1210/en.2002-220786. [DOI] [PubMed] [Google Scholar]
- 485.Tronchon V., Vialard F., El Sirkasi M. Tumor necrosis factor-alpha -308 polymorphism in infertile men with altered sperm production or motility. Hum Reprod. 2008;23:2858–2866. doi: 10.1093/humrep/den277. [DOI] [PubMed] [Google Scholar]
- 486.Yao P.L., Lin Y.C., Sawhney P., Richburg J.H. Transcriptional regulation of FasL expression and participation of sTNF-α in response to Sertoli cell injury. J Biol Chem. 2007;282:5420–5431. doi: 10.1074/jbc.M609068200. [DOI] [PubMed] [Google Scholar]
- 487.Varfolomeev E., Vucic D. (Un)expected roles of c-IAPs in apoptotic and NFκB signaling pathways. Cell Cycle. 2008;7:1511–1521. doi: 10.4161/cc.7.11.5959. [DOI] [PubMed] [Google Scholar]
- 488.Schütze S., Tchikov V., Schneider-Brachert W. Regulation of TNFR1 and CD95 signalling by receptor compartmentalization. Nat Rev Mol Cell Biol. 2008;9:655–662. doi: 10.1038/nrm2430. [DOI] [PubMed] [Google Scholar]
- 489.Bellgrau D., Gold D., Selawry H., Moore J., Franzusoff A., Duke R.C. A role for CD95 ligand in preventing graft rejection. Nature. 1995;377:630–632. doi: 10.1038/377630a0. [DOI] [PubMed] [Google Scholar]
- 490.Lee J., Richburg J.H., Younkin S.C., Boekelheide K. The Fas system is a key regulator of germ cell apoptosis in the testis. Endocrinology. 1997;138:2081–2088. doi: 10.1210/endo.138.5.5110. [DOI] [PubMed] [Google Scholar]
- 491.Restifo N.P. Not so Fas: re-evaluating the mechanisms of immune privilege and tumor escape. Nat Med. 2000;6:493–495. doi: 10.1038/74955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 492.D’Alessio A., Riccioli A., Lauretti P. Testicular FasL is expressed by sperm cells. Proc Natl Acad Sci USA. 2001;98:3316–3321. doi: 10.1073/pnas.051566098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 493.Brændstrup O., Bols B., Jensen L. Fas and Fas-ligand expression in seminomatous testes. APMIS. 1999;107:431–436. doi: 10.1111/j.1699-0463.1999.tb01577.x. [DOI] [PubMed] [Google Scholar]
- 494.D’Abrizio P., Baldini E., Russo P.F. Ontogenesis and cell specific localization of Fas ligand expression in the rat testis. Int J Androl. 2004;27:304–310. doi: 10.1111/j.1365-2605.2004.00483.x. [DOI] [PubMed] [Google Scholar]
- 495.Celik-Ozenci C., Sahin Z., Ustunel I. The Fas system may have a role in male reproduction. Fertil Steril. 2006;85(Suppl. 1):1168–1178. doi: 10.1016/j.fertnstert.2005.08.058. [DOI] [PubMed] [Google Scholar]
- 496.Riccioli A., Starace D., D’Alessio A. TNF-α and IFN-γ regulate expression and function of the Fas system in the seminiferous epithelium. J Immunol. 2000;165:743–749. doi: 10.4049/jimmunol.165.2.743. [DOI] [PubMed] [Google Scholar]
- 497.Lizama C., Alfaro I., Reyes J.G., Moreno R.D. Up-regulation of CD95 (Apo-1/Fas) is associated with spermatocyte apoptosis during the first round of spermatogenesis in the rat. Apoptosis. 2007;12:499–512. doi: 10.1007/s10495-006-0012-1. [DOI] [PubMed] [Google Scholar]
- 498.Ogi S., Tanji N., Yokoyama M., Takeuchi M., Terada N. Involvement of Fas in the apoptosis of mouse germ cells induced by experimental cryptorchidism. Urol Res. 1998;26:17–21. doi: 10.1007/s002400050017. [DOI] [PubMed] [Google Scholar]
- 499.Kajihara T., Okagaki R., Ishihara O. LPS-induced transient testicular dysfunction accompanied by apoptosis of testicular germ cells in mice. Med Mol Morphol. 2006;39:203–208. doi: 10.1007/s00795-006-0334-7. [DOI] [PubMed] [Google Scholar]
- 500.O’Reilly S., Ciechomska M., Cant R., Hügle T., van Laar J.M. Interleukin-6, its role in fibrosing conditions. Cytokine Growth Factor Rev. 2012;23:99–107. doi: 10.1016/j.cytogfr.2012.04.003. [DOI] [PubMed] [Google Scholar]
- 501.Heinrich P.C., Behrmann I., Haan S., Hermanns H.M., Muller-Newen G., Schaper F. Principles of interleukin (IL)-6-type cytokine signalling and its regulation. Biochem J. 2003;374:1–20. doi: 10.1042/BJ20030407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 502.Syed V., Gerard N., Kaipia A., Bardin C.W., Parvinen M., Jegou B. Identification, ontogeny, and regulation of an interleukin-6-like factor in the rat seminiferous tubule. Endocrinology. 1993;132:293–299. doi: 10.1210/endo.132.1.8380379. [DOI] [PubMed] [Google Scholar]
- 503.Okuda Y., Sun X.R., Morris P.L. Interleukin-6 (IL-6) mRNAs expressed in Leydig and Sertoli cells are regulated by cytokines, gonadotropins and neuropeptides. Endocr J. 1994;2:617–624. [Google Scholar]
- 504.Boockfor F.R., Wang D., Lin T., Nagpal M.L., Spangelo B.L. Interleukin-6 secretion from rat Leydig cells in culture. Endocrinology. 1994;134:2150–2155. doi: 10.1210/endo.134.5.8156916. [DOI] [PubMed] [Google Scholar]
- 505.Fujisawa M., Okuda Y., Fujioka H., Kamidono S. Expression and regulation of gp130 messenger ribonucleic acid in cultured immature rat Sertoli cells. Endocr Res. 2002;28:1–8. doi: 10.1081/erc-120004532. [DOI] [PubMed] [Google Scholar]
- 506.Hoeben E., Wuyts A., Proost P., Van Damme J., Verhoeven G. Identification of IL-6 as one of the important cytokines responsible for the ability of mononuclear cells to stimulate Sertoli cell functions. Mol Cell Endocrinol. 1997;132:149–160. doi: 10.1016/s0303-7207(97)00132-9. [DOI] [PubMed] [Google Scholar]
- 507.Boockfor F.R., Schwarz L.K. Effects of interleukin-6, interleukin-2, and tumor necrosis factor alpha on transferrin release from Sertoli cells in culture. Endocrinology. 1991;129:256–262. doi: 10.1210/endo-129-1-256. [DOI] [PubMed] [Google Scholar]
- 508.Jenab S., Morris P.L. Interleukin-6 regulation of κ opioid receptor gene expression in primary Sertoli cells. Endocrine. 2000;13:11–15. doi: 10.1385/ENDO:13:1:11. [DOI] [PubMed] [Google Scholar]
- 509.Hakovirta H., Syed V., Jégou B., Parvinen M. Function of interleukin-6 as an inhibitor of meiotic DNA synthesis in the rat seminiferous epithelium. Mol Cell Endocrinol. 1995;108:193–198. doi: 10.1016/0303-7207(95)03475-m. [DOI] [PubMed] [Google Scholar]
- 510.Bornstein S.R., Rutkowski H., Vrezas I. Cytokines and steroidogenesis. Mol Cell Endocrinol. 2004;215:135–141. doi: 10.1016/j.mce.2003.11.022. [DOI] [PubMed] [Google Scholar]
- 511.Hinck A.P. Structural studies of the TGF-βs and their receptors – insights into evolution of the TGF-β superfamily. FEBS Lett. 2012;586:1860–1870. doi: 10.1016/j.febslet.2012.05.028. [DOI] [PubMed] [Google Scholar]
- 512.de Caestecker M. The transforming growth factor-β superfamily of receptors. Cytokine Growth Factor Rev. 2004;15:1–11. doi: 10.1016/j.cytogfr.2003.10.004. [DOI] [PubMed] [Google Scholar]
- 513.Heldin C.H., Landström M., Moustakas A. Mechanism of TGF-β signaling to growth arrest, apoptosis, and epithelial-mesenchymal transition. Curr Opin Cell Biol. 2009;21:166–176. doi: 10.1016/j.ceb.2009.01.021. [DOI] [PubMed] [Google Scholar]
- 514.Venkataraman G., Sasisekharan V., Cooney C.L., Langer R., Sasisekharan R. Complex flexibility of the transforming growth factor β superfamily. Proc Natl Acad Sci USA. 1995;92:5406–5410. doi: 10.1073/pnas.92.12.5406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 515.Mullaney B.P., Skinner M.K. Transforming growth factor-β (β1, β2, and β3) gene expression and action during pubertal development of the seminiferous tubule: potential role at the onset of spermatogenesis. Mol Endocrinol. 1993;7:67–76. doi: 10.1210/mend.7.1.8446109. [DOI] [PubMed] [Google Scholar]
- 516.Konrad L., Albrecht M., Renneberg H., Aumüller G. Transforming growth factor-β2 mediates mesenchymal–epithelial interactions of testicular somatic cells. Endocrinology. 2000;141:3679–3686. doi: 10.1210/endo.141.10.7728. [DOI] [PubMed] [Google Scholar]
- 517.Gonzalez C.R., Matzkin M.E., Frungieri M.B. Expression of the TGF-beta1 system in human testicular pathologies. Reprod Biol Endocrinol. 2010;8:148. doi: 10.1186/1477-7827-8-148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 518.Teerds K.J., Dorrington J.H. Localization of transforming growth factor β1 and β2 during testicular development in the rat. Biol Reprod. 1993;48:40–45. doi: 10.1095/biolreprod48.1.40. [DOI] [PubMed] [Google Scholar]
- 519.Le Magueresse-Battistoni B., Morera A.M., Goddard I., Benahmed M. Expression of mRNAs for transforming growth factor-β receptors in the rat testis. Endocrinology. 1995;136:2788–2791. doi: 10.1210/endo.136.6.7750505. [DOI] [PubMed] [Google Scholar]
- 520.Dickson C., Webster D.R., Johnson H., Cecilia Millena A., Khan S.A. Transforming growth factor-β effects on morphology of immature rat Leydig cells. Mol Cell Endocrinol. 2002;195:65–77. doi: 10.1016/s0303-7207(02)00216-2. [DOI] [PubMed] [Google Scholar]
- 521.Olaso R., Pairault C., Boulogne B., Durand P., Habert R. Transforming growth factor β1 and β2 reduce the number of gonocytes by increasing apoptosis. Endocrinology. 1998;139:733–740. doi: 10.1210/endo.139.2.5765. [DOI] [PubMed] [Google Scholar]
- 522.Gonzalez C.R., Gonzalez B., Rulli S.B. TGF-β1 system in Leydig cells. Part II: TGF-β1 and progesterone, through Smad1/5, are involved in the hyperplasia/hypertrophy of Leydig cells. J Reprod Dev. 2010;56:400–404. doi: 10.1262/jrd.09-167n. [DOI] [PubMed] [Google Scholar]
- 523.Maire M., Florin A., Kaszas K. Alteration of transforming growth factor-β signaling system expression in adult rat germ cells with a chronic apoptotic cell death process after fetal androgen disruption. Endocrinology. 2005;146:5135–5143. doi: 10.1210/en.2005-0592. [DOI] [PubMed] [Google Scholar]
- 524.Perrard M.H., Durand P. Redundancy of the effect of TGFβ1 and β-NGF on the second meiotic division of rat spermatocytes. Microsc Res Tech. 2009;72:596–602. doi: 10.1002/jemt.20706. [DOI] [PubMed] [Google Scholar]
- 525.Moreno S.G., Attali M., Allemand I. TGFβ signaling in male germ cells regulates gonocyte quiescence and fertility in mice. Dev Biol. 2010;342:74–84. doi: 10.1016/j.ydbio.2010.03.007. [DOI] [PubMed] [Google Scholar]
- 526.Avallet O., Vigier M., Perrard-Sapori M.H., Saez J.M. Transforming growth factor β inhibits Leydig cell functions. Biochem Biophys Res Commun. 1987;146:575–581. doi: 10.1016/0006-291x(87)90567-5. [DOI] [PubMed] [Google Scholar]
- 527.Chuzel F., Clark A.M., Avallet O., Saez J.M. Transcriptional regulation of the lutropin/human choriogonadotropin receptor and three enzymes of steroidogenesis by growth factors in cultured pig Leydig cells. Eur J Biochem. 1996;239:8–16. doi: 10.1111/j.1432-1033.1996.0008u.x. [DOI] [PubMed] [Google Scholar]
- 528.Lui W.Y., Wong C.H., Mruk D.D., Cheng C.Y. TGF-β3 regulates the blood-testis barrier dynamics via the p38 mitogen activated protein (MAP) kinase pathway: an in vivo study. Endocrinology. 2003;144:1139–1142. doi: 10.1210/en.2002-0211. [DOI] [PubMed] [Google Scholar]
- 529.Schmidt-Weber C.B., Blaser K. Regulation and role of transforming growth factor-β in immune tolerance induction and inflammation. Curr Opin Immunol. 2004;16:709–716. doi: 10.1016/j.coi.2004.09.008. [DOI] [PubMed] [Google Scholar]
- 530.Tran D.Q. TGF-β: the sword, the wand, and the shield of FOXP3+ regulatory T cells. J Mol Cell Biol. 2012;4:29–37. doi: 10.1093/jmcb/mjr033. [DOI] [PubMed] [Google Scholar]
- 531.Hedger M.P., Winnall W.R., Phillips D.J., de Kretser D.M. The regulation and functions of activin and follistatin in inflammation and immunity. Vitamin Horm. 2011;85:255–297. doi: 10.1016/B978-0-12-385961-7.00013-5. [DOI] [PubMed] [Google Scholar]
- 532.de Kretser D.M., Robertson D.M. The isolation and physiology of inhibin and related proteins. Biol Reprod. 1989;40:33–47. doi: 10.1095/biolreprod40.1.33. [DOI] [PubMed] [Google Scholar]
- 533.Brown C.W., Houston-Hawkins D.E., Woodruff T.K., Matzuk M.M. Insertion of Inhbb into the Inhba locus rescues the Inhba-null phenotype and reveals new activin functions. Nat Genet. 2000;25:453–457. doi: 10.1038/78161. [DOI] [PubMed] [Google Scholar]
- 534.Ling N., Ying S.Y., Ueno N. Pituitary FSH is released by a heterodimer of the β-subunits from the two forms of inhibin. Nature. 1986;321:779–782. doi: 10.1038/321779a0. [DOI] [PubMed] [Google Scholar]
- 535.Meunier H., Rivier C., Evans R.M., Vale W. Gonadal and extragonadal expression of inhibin α, βA, and βB subunits in various tissues predicts diverse functions. Proc Natl Acad Sci USA. 1988;85:247–251. doi: 10.1073/pnas.85.1.247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 536.de Kretser D.M., Buzzard J.J., Okuma Y. The role of activin, follistatin and inhibin in testicular physiology. Mol Cell Endocrinol. 2004;225:57–64. doi: 10.1016/j.mce.2004.07.008. [DOI] [PubMed] [Google Scholar]
- 537.Wu H., Chen Y., Winnall W.R., Phillips D.J., Hedger M.P. Acute regulation of activin A and its binding protein, follistatin, in serum and tissues following lipopolysaccharide treatment of adult male mice. Am J Physiol Regul Integr Comp Physiol. 2012;303:R665–R675. doi: 10.1152/ajpregu.00478.2011. [DOI] [PubMed] [Google Scholar]
- 538.Schneider O., Nau R., Michel U. Comparative analysis of follistatin-, activin beta A- and activin beta B-mRNA steady-state levels in diverse porcine tissues by multiplex S1 nuclease analysis. Eur J Endocrinol. 2000;142:537–544. doi: 10.1530/eje.0.1420537. [DOI] [PubMed] [Google Scholar]
- 539.Tuuri T., Erämaa M., Hildén K., Ritvos O. The tissue distribution of activin βA- and βB-subunit and follistatin messenger ribonucleic acids suggests multiple sites of action for the activin-follistatin system during human development. J Clin Endocrinol Metab. 1994;78:1521–1524. doi: 10.1210/jcem.78.6.8200957. [DOI] [PubMed] [Google Scholar]
- 540.Tsuchida K., Nakatani M., Uezumi A., Murakami T., Cui X. Signal transduction pathway through activin receptors as a therapeutic target of musculoskeletal diseases and cancer. Endocr J. 2008;55:11–21. doi: 10.1507/endocrj.kr-110. [DOI] [PubMed] [Google Scholar]
- 541.Huang H.M., Chiou H.Y., Chang J.L. Activin A induces erythroid gene expressions and inhibits mitogenic cytokine-mediated K562 colony formation by activating p38 MAPK. J Cell Biochem. 2006;98:789–797. doi: 10.1002/jcb.20713. [DOI] [PubMed] [Google Scholar]
- 542.Makanji Y., Walton K.L., Wilce M.C., Chan K.L., Robertson D.M., Harrison C.A. Suppression of inhibin A biological activity by alterations in the binding site for betaglycan. J Biol Chem. 2008;283:16743–16751. doi: 10.1074/jbc.M801045200. [DOI] [PubMed] [Google Scholar]
- 543.Muenster U., Harrison C.A., Donaldson C., Vale W., Fischer W.H. An activin-A/C chimera exhibits activin and myostatin antagonistic properties. J Biol Chem. 2005;280:36626–36632. doi: 10.1074/jbc.M507236200. [DOI] [PubMed] [Google Scholar]
- 544.Mellor S.L., Ball E.M., O’Connor A.E. Activin βC-subunit heterodimers provide a new mechanism of regulating activin levels in the prostate. Endocrinology. 2003;144:4410–4419. doi: 10.1210/en.2003-0225. [DOI] [PubMed] [Google Scholar]
- 545.Gold E.J., O’Bryan M.K., Mellor S.L. Cell-specific expression of βC-activin in the rat reproductive tract, adrenal and liver. Mol Cell Endocrinol. 2004;222:61–69. doi: 10.1016/j.mce.2004.05.007. [DOI] [PubMed] [Google Scholar]
- 546.Thompson T.B., Cook R.W., Chapman S.C., Jardetzky T.S., Woodruff T.K. Beta A versus beta B: is it merely a matter of expression? Mol Cell Endocrinol. 2004;225:9–17. doi: 10.1016/j.mce.2004.02.007. [DOI] [PubMed] [Google Scholar]
- 547.Gold E., Jetly N., O’Bryan M.K. Activin C antagonizes activin A in vitro and overexpression leads to pathologies in vivo. Am J Pathol. 2009;174:184–195. doi: 10.2353/ajpath.2009.080296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 548.Nakamura T., Sugino K., Titani K., Sugino H. Follistatin, an activin-binding protein, associates with heparan sulfate chains of proteoglycans on follicular granulosa cells. J Biol Chem. 1991;266:19432–19437. [PubMed] [Google Scholar]
- 549.Tanimoto K., Yoshida E., Mita S., Nibu Y., Murakami K., Fukamizu A. Human activin βA gene. Identification of novel 5′ exon, functional promoter, and enhancers. J Biol Chem. 1996;271:32760–32769. doi: 10.1074/jbc.271.51.32760. [DOI] [PubMed] [Google Scholar]
- 550.Ardekani A.M., Romanelli J.C., Mayo K.E. Structure of the rat inhibin and activin βA-subunit gene and regulation in an ovarian granulosa cell line. Endocrinology. 1998;139:3271–3279. doi: 10.1210/endo.139.7.6116. [DOI] [PubMed] [Google Scholar]
- 551.Ogawa K., Funaba M., Chen Y., Tsujimoto M. Activin A functions as a Th2 cytokine in the promotion of the alternative activation of macrophages. J Immunol. 2006;177:6787–6794. doi: 10.4049/jimmunol.177.10.6787. [DOI] [PubMed] [Google Scholar]
- 552.Thompson D.A., Cronin C.N., Martin F. Genomic cloning and sequence analyses of the bovine α-, βA- and βB-inhibin/activin genes. Identification of transcription factor AP-2-binding sites in the 5′-flanking regions by DNase I footprinting. Eur J Biochem. 1994;226:751–764. doi: 10.1111/j.1432-1033.1994.00751.x. [DOI] [PubMed] [Google Scholar]
- 553.Dykema J.C., Mayo K.E. Two messenger ribonucleic acids encoding the common βB-chain of inhibin and activin have distinct 5′-initiation sites and are differentially regulated in rat granulosa cells. Endocrinology. 1994;135:702–711. doi: 10.1210/endo.135.2.8033818. [DOI] [PubMed] [Google Scholar]
- 554.Jones K.L., Mansell A., Patella S. Activin A is a critical component of the inflammatory response, and its binding protein, follistatin, reduces mortality in endotoxemia. Proc Natl Acad Sci USA. 2007;104:16239–16244. doi: 10.1073/pnas.0705971104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 555.Yamashita N., Nakajima T., Takahashi H., Kaneoka H., Mizushima Y., Sakane T. Effects of activin A on IgE synthesis and cytokine production by human peripheral mononuclear cells. Clin Exp Immunol. 1993;94:214–219. doi: 10.1111/j.1365-2249.1993.tb06003.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 556.Sugatani T., Alvarez U.M., Hruska K.A. Activin A stimulates IκB-α/NFκB and RANK expression for osteoclast differentiation, but not AKT survival pathway in osteoclast precursors. J Cell Biochem. 2003;90:59–67. doi: 10.1002/jcb.10613. [DOI] [PubMed] [Google Scholar]
- 557.Ohguchi M., Yamato K., Ishihara Y. Activin A regulates the production of mature interleukin-1β and interleukin-1 receptor antagonist in human monocytic cells. J Interferon Cytokine Res. 1998;18:491–498. doi: 10.1089/jir.1998.18.491. [DOI] [PubMed] [Google Scholar]
- 558.Wang S.Y., Tai G.X., Zhang P.Y., Mu D.P., Zhang X.J., Liu Z.H. Inhibitory effect of activin A on activation of lipopolysaccharide-stimulated mouse macrophage RAW264.7 cells. Cytokine. 2008;42:85–91. doi: 10.1016/j.cyto.2008.01.010. [DOI] [PubMed] [Google Scholar]
- 559.Salogni L., Musso T., Bosisio D. Activin A induces dendritic cell migration through the polarized release of CXC chemokine ligands 12 and 14. Blood. 2009;113:5848–5856. doi: 10.1182/blood-2008-12-194597. [DOI] [PubMed] [Google Scholar]
- 560.Scutera S., Riboldi E., Daniele R. Production and function of activin A in human dendritic cells. Eur Cytokine Netw. 2008;19:60–68. doi: 10.1684/ecn.2008.0121. [DOI] [PubMed] [Google Scholar]
- 561.Robson N.C., Phillips D.J., McAlpine T. Activin-A: a novel dendritic cell-derived cytokine that potently attenuates CD40 ligand-specific cytokine and chemokine production. Blood. 2008;111:2733–2743. doi: 10.1182/blood-2007-03-080994. [DOI] [PubMed] [Google Scholar]
- 562.Robson N.C., Wei H., McAlpine T., Kirkpatrick N., Cebon J., Maraskovsky E. Activin-A attenuates several human natural killer cell functions. Blood. 2009;113:3218–3225. doi: 10.1182/blood-2008-07-166926. [DOI] [PubMed] [Google Scholar]
- 563.Zhou J., Tai G., Liu H. Activin A down-regulates the phagocytosis of lipopolysaccharide-activated mouse peritoneal macrophages in vitro and in vivo. Cell Immunol. 2009;255:69–75. doi: 10.1016/j.cellimm.2008.11.001. [DOI] [PubMed] [Google Scholar]
- 564.Huber S., Stahl F.R., Schrader J. Activin a promotes the TGF-β-induced conversion of CD4+CD25− T cells into Foxp3+ induced regulatory T cells. J Immunol. 2009;182:4633–4640. doi: 10.4049/jimmunol.0803143. [DOI] [PubMed] [Google Scholar]
- 565.Russell C.E., Hedger M.P., Brauman J.N., de Kretser D.M., Phillips D.J. Activin A regulates growth and acute phase proteins in the human liver cell line, HepG2. Mol Cell Endocrinol. 1999;148:129–136. doi: 10.1016/s0303-7207(98)00226-3. [DOI] [PubMed] [Google Scholar]
- 566.Hedger M.P., Drummond A.E., Robertson D.M., Risbridger G.P., de Kretser D.M. Inhibin and activin regulate [3H]thymidine uptake by rat thymocytes and 3T3 cells in vitro. Mol Cell Endocrinol. 1989;61:133–138. doi: 10.1016/0303-7207(89)90198-6. [DOI] [PubMed] [Google Scholar]
- 567.Phillips D.J., Brauman J.N., Mason A.J., de Kretser D.M., Hedger M.P. A sensitive and specific in vitro bioassay for activin using a mouse plasmacytoma cell line, MPC-11. J Endocrinol. 1999;162:111–116. doi: 10.1677/joe.0.1620111. [DOI] [PubMed] [Google Scholar]
- 568.Jones C.P., Gregory L.G., Causton B., Campbell G.A., Lloyd C.M. Activin A and TGF-β promote TH9 cell-mediated pulmonary allergic pathology. J Allergy Clin Immunol. 2012;129:1000–1010. doi: 10.1016/j.jaci.2011.12.965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 569.Funaba M., Ikeda T., Ogawa K., Murakami M., Abe M. Role of activin A in murine mast cells: modulation of cell growth, differentiation, and migration. J Leuk Biol. 2003;73:793–801. doi: 10.1189/jlb.0103012. [DOI] [PubMed] [Google Scholar]
- 570.Ogawa K., Funaba M., Tsujimoto M. A dual role of activin A in regulating immunoglobulin production of B cells. J Leuk Biol. 2008;83:1451–1458. doi: 10.1189/jlb.1007710. [DOI] [PubMed] [Google Scholar]
- 571.Aoki F., Kojima I. Therapeutic potential of follistatin to promote tissue regeneration and prevent tissue fibrosis. Endocr J. 2007;54:849–854. doi: 10.1507/endocrj.kr07e-001. [DOI] [PubMed] [Google Scholar]
- 572.Kaipia A., Penttila T.L., Shimasaki S., Ling N., Parvinen M., Toppari J. Expression of inhibin βA and βB, follistatin and activin-A receptor messenger ribonucleic acids in the rat seminiferous epithelium. Endocrinology. 1992;131:2703–2710. doi: 10.1210/endo.131.6.1332846. [DOI] [PubMed] [Google Scholar]
- 573.Okuma Y., O’Connor A.E., Hayashi T., Loveland K.L., de Kretser D.M., Hedger M.P. Regulated production of activin A and inhibin B throughout the cycle of the seminiferous epithelium in the rat. J Endocrinol. 2006;190:331–340. doi: 10.1677/joe.1.06706. [DOI] [PubMed] [Google Scholar]
- 574.Illingworth P.J., Groome N.P., Byrd W. Inhibin-B: a likely candidate for the physiologically important form of inhibin in men. J Clin Endocrinol Metab. 1996;81:1321–1325. doi: 10.1210/jcem.81.4.8636325. [DOI] [PubMed] [Google Scholar]
- 575.Barakat B., O’Connor A.E., Gold E., de Kretser D.M., Loveland K.L. Inhibin, activin, follistatin and FSH serum levels and testicular production are highly modulated during the first spermatogenic wave in mice. Reproduction. 2008;136:345–359. doi: 10.1530/REP-08-0140. [DOI] [PubMed] [Google Scholar]
- 576.Cameron V.A., Nishimura E., Mathews L.S., Lewis K.A., Sawchenko P.E., Vale W.W. Hybridization histochemical localization of activin receptor subtypes in rat brain, pituitary, ovary, and testis. Endocrinology. 1994;134:799–808. doi: 10.1210/endo.134.2.8299574. [DOI] [PubMed] [Google Scholar]
- 577.Dias V., Meachem S., Rajpert-De Meyts E., McLachlan R., Manuelpillai U., Loveland K.L. Activin receptor subunits in normal and dysfunctional adult human testis. Hum Reprod. 2008;23:412–420. doi: 10.1093/humrep/dem343. [DOI] [PubMed] [Google Scholar]
- 578.Loveland K.L., Dias V., Meachem S., Rajpert-De Meyts E. The transforming growth factor-β superfamily in early spermatogenesis: potential relevance to testicular dysgenesis. Int J Androl. 2007;30:377–384. doi: 10.1111/j.1365-2605.2007.00785.x. [DOI] [PubMed] [Google Scholar]
- 579.Mather J.P., Attie K.M., Woodruff T.K., Rice G.C., Phillips D.M. Activin stimulates spermatogonial proliferation in germ-Sertoli cell cocultures from immature rat testis. Endocrinology. 1990;127:3206–3214. doi: 10.1210/endo-127-6-3206. [DOI] [PubMed] [Google Scholar]
- 580.Hakovirta H., Kaipia A., Söder O., Parvinen M. Effects of activin-A, inhibin-A, and transforming growth factor-β1 on stage-specific deoxyribonucleic acid synthesis during rat seminiferous epithelial cycle. Endocrinology. 1993;133:1664–1668. doi: 10.1210/endo.133.4.8404607. [DOI] [PubMed] [Google Scholar]
- 581.Meehan T., Schlatt S., O’Bryan M.K., de Kretser D.M., Loveland K.L. Regulation of germ cell and Sertoli cell development by activin, follistatin, and FSH. Dev Biol. 2000;220:225–237. doi: 10.1006/dbio.2000.9625. [DOI] [PubMed] [Google Scholar]
- 582.Meinhardt A., McFarlane J.R., Seitz J., de Kretser D.M. Activin maintains the condensed type of mitochondria in germ cells. Mol Cell Endocrinol. 2000;168:111–117. doi: 10.1016/s0303-7207(00)00308-7. [DOI] [PubMed] [Google Scholar]
- 583.Nicholls P.K., Stanton P.G., Chen J.L. Activin signaling regulates Sertoli cell differentiation and function. Endocrinology. 2012;153:6065–6077. doi: 10.1210/en.2012-1821. [DOI] [PubMed] [Google Scholar]
- 584.Renlund N., O’Neill F.H., Zhang L., Sidis Y., Teixeira J. Activin receptor-like kinase-2 inhibits activin signaling by blocking the binding of activin to its type II receptor. J Endocrinol. 2007;195:95–103. doi: 10.1677/JOE-07-0281. [DOI] [PubMed] [Google Scholar]
- 585.Lejeune H., Chuzel F., Sanchez P., Durand P., Mather J.P., Saez J.M. Stimulating effect of both human recombinant inhibin A and activin A on immature porcine Leydig cell functions in vitro. Endocrinology. 1997;138:4783–4791. doi: 10.1210/endo.138.11.5542. [DOI] [PubMed] [Google Scholar]
- 586.Lin T., Calkins J.K., Morris P.L., Vale W., Bardin C.W. Regulation of Leydig cell function in primary culture by inhibin and activin. Endocrinology. 1989;125:2134–2140. doi: 10.1210/endo-125-4-2134. [DOI] [PubMed] [Google Scholar]
- 587.Lau A.L., Kumar T.R., Nishimori K., Bonadio J., Matzuk M.M. Activin βC and βE genes are not essential for mouse liver growth, differentiation, and regeneration. Mol Cell Biol. 2000;20:6127–6137. doi: 10.1128/mcb.20.16.6127-6137.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 588.Malmgaard L. Induction and regulation of IFNs during viral infections. J Interferon Cytokine Res. 2004;24:439–454. doi: 10.1089/1079990041689665. [DOI] [PubMed] [Google Scholar]
- 589.Varma T.K., Lin C.Y., Toliver-Kinsky T.E., Sherwood E.R. Endotoxin-induced gamma interferon production: contributing cell types and key regulatory factors. Clin Diagn Lab Immunol. 2002;9:530–543. doi: 10.1128/CDLI.9.3.530-543.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 590.Schoenborn J.R., Wilson C.B. Regulation of interferon-gamma during innate and adaptive immune responses. Adv Immunol. 2007;96:41–101. doi: 10.1016/S0065-2776(07)96002-2. [DOI] [PubMed] [Google Scholar]
- 591.Dejucq N., Liénard M.O., Jégou B. Interferons and interferon-induced antiviral proteins in the testis. J Reprod Immunol. 1998;41:291–300. doi: 10.1016/s0165-0378(98)00065-5. [DOI] [PubMed] [Google Scholar]
- 592.Dejucq N., Lienard M.O., Guillaume E., Dorval I., Jégou B. Expression of interferons-α and -γ in testicular interstitial tissue and spermatogonia of the rat. Endocrinology. 1998;139:3081–3087. doi: 10.1210/endo.139.7.6083. [DOI] [PubMed] [Google Scholar]
- 593.Hu J., You S., Li W. Expression and regulation of interferon-γ-inducible protein 10 gene in rat Leydig cells. Endocrinology. 1998;139:3637–3645. doi: 10.1210/endo.139.8.6143. [DOI] [PubMed] [Google Scholar]
- 594.Dal Secco V., Riccioli A., Padula F., Ziparo E., Filippini A. Mouse Sertoli cells display phenotypical and functional traits of antigen-presenting cells in response to interferon gamma. Biol Reprod. 2008;78:234–242. doi: 10.1095/biolreprod.107.063578. [DOI] [PubMed] [Google Scholar]
- 595.Itoh M., Yano A., Xie Q. Essential pathogenic role for endogenous interferon-gamma (IFN-γ) during disease onset phase of murine experimental autoimmune orchitis. I. In vivo studies. Clin Exp Immunol. 1998;111:513–520. doi: 10.1046/j.1365-2249.1998.00527.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 596.Kanzaki M., Morris P.L. Identification and regulation of testicular interferon-γ (IFNγ) receptor subunits: IFNγ enhances interferon regulatory factor-1 and interleukin-1β converting enzyme expression. Endocrinology. 1998;139:2636–2644. doi: 10.1210/endo.139.5.5975. [DOI] [PubMed] [Google Scholar]
- 597.Satie A.P., Mazaud-Guittot S., Seif I. Excess type I interferon signaling in the mouse seminiferous tubules leads to germ cell loss and sterility. J Biol Chem. 2011;286:23280–23295. doi: 10.1074/jbc.M111.229120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 598.Orava M., Cantell K., Vihko R. Treatment with preparations of human leukocyte interferon decreases serum testosterone concentrations in men. Int J Cancer. 1986;38:295–296. doi: 10.1002/ijc.2910380222. [DOI] [PubMed] [Google Scholar]
- 599.Orava M., Cantell K., Vihko R. Human leukocyte interferon inhibits human chorionic gonadotropin stimulated testosterone production by porcine Leydig cells in culture. Biochem Biophys Res Commun. 1985;127:809–815. doi: 10.1016/s0006-291x(85)80015-2. [DOI] [PubMed] [Google Scholar]
- 600.Orava M., Voutilainen R., Vihko R. Interferon-γ inhibits steroidogenesis and accumulation of mRNA of the steroidogenic enzymes, P450SCC and P450C17 in cultured porcine Leydig cells. Mol Endocrinol. 1989;3:887–894. doi: 10.1210/mend-3-6-887. [DOI] [PubMed] [Google Scholar]
- 601.Montor J.M., Mendoza M.E., Romano M.C. Effect of human and murine interferon-α on steroid production by rat ovarian cells. Life Sci. 1998;62:1733–1744. doi: 10.1016/s0024-3205(98)00135-0. [DOI] [PubMed] [Google Scholar]
- 602.Dobs A.S., Dempsey M.A., Ladenson P.W., Polk B.F. Endocrine disorders in men infected with human immunodeficiency virus. Am J Med. 1988;84:611–616. doi: 10.1016/0002-9343(88)90144-1. [DOI] [PubMed] [Google Scholar]
- 603.Fountain S., Holland M.K., Hinds L.A., Janssens P.A., Kerr P.J. Interstitial orchitis with impaired steroidogenesis and spermatogenesis in the testes of rabbits infected with an attenuated strain of myxoma virus. J Reprod Fertil. 1997;110:161–169. doi: 10.1530/jrf.0.1100161. [DOI] [PubMed] [Google Scholar]
- 604.Lim A., Kumar V., Dass S.A.H., Vyas A. Toxoplasma gondii infection enhances testicular steroidogenesis in rats. Mol Ecol. 2013;22:102–110. doi: 10.1111/mec.12042. [DOI] [PubMed] [Google Scholar]
- 605.Schaloske R.H., Dennis E.A. The phospholipase A2 superfamily and its group numbering system. Biochim Biophys Acta. 2006;1761:1246–1259. doi: 10.1016/j.bbalip.2006.07.011. [DOI] [PubMed] [Google Scholar]
- 606.Smith W.L., DeWitt D.L., Garavito R.M. Cyclooxygenases: structural, cellular, and molecular biology. Annu Rev Biochem. 2000;69:145–182. doi: 10.1146/annurev.biochem.69.1.145. [DOI] [PubMed] [Google Scholar]
- 607.Ueno N., Takegoshi Y., Kamei D., Kudo I., Murakami M. Coupling between cyclooxygenases and terminal prostanoid synthases. Biochem Biophys Res Commun. 2005;338:70–76. doi: 10.1016/j.bbrc.2005.08.152. [DOI] [PubMed] [Google Scholar]
- 608.Hata A.N., Breyer R.M. Pharmacology and signaling of prostaglandin receptors: multiple roles in inflammation and immune modulation. Pharmacol Ther. 2004;103:147–166. doi: 10.1016/j.pharmthera.2004.06.003. [DOI] [PubMed] [Google Scholar]
- 609.Harris S.G., Padilla J., Koumas L., Ray D., Phipps R.P. Prostaglandins as modulators of immunity. Trends Immunol. 2002;23:144–150. doi: 10.1016/s1471-4906(01)02154-8. [DOI] [PubMed] [Google Scholar]
- 610.Jiang C., Ting A.T., Seed B. PPAR-γ agonists inhibit production of monocyte inflammatory cytokines. Nature. 1998;391:82–86. doi: 10.1038/34184. [DOI] [PubMed] [Google Scholar]
- 611.Funk C.D. Prostaglandins and leukotrienes: advances in eicosanoid biology. Science. 2001;294:1871–1875. doi: 10.1126/science.294.5548.1871. [DOI] [PubMed] [Google Scholar]
- 612.Takahashi N., Takeuchi K., Abe T., Murakami K., Yamaguchi M., Abe K. Immunohistochemical localization of thromboxane receptor and thromboxane synthase in rat testis. Endocrinology. 1995;136:4143–4146. doi: 10.1210/endo.136.9.7649122. [DOI] [PubMed] [Google Scholar]
- 613.Gerena R.L., Eguchi N., Urade Y., Killian G.J. Stage and region-specific localization of lipocalin-type prostaglandin D synthase in the adult murine testis and epididymis. J Androl. 2000;21:848–854. [PubMed] [Google Scholar]
- 614.Lazarus M., Munday C.J., Eguchi N. Immunohistochemical localization of microsomal PGE synthase-1 and cyclooxygenases in male mouse reproductive organs. Endocrinology. 2002;143:2410–2419. doi: 10.1210/endo.143.6.8872. [DOI] [PubMed] [Google Scholar]
- 615.Winnall W.R., Ali U., O’Bryan M.K. Constitutive expression of prostaglandin-endoperoxide synthase 2 by somatic and spermatogenic cells is responsible for prostaglandin E2 production in the adult rat testis. Biol Reprod. 2007;76:759–768. doi: 10.1095/biolreprod.106.053124. [DOI] [PubMed] [Google Scholar]
- 616.Lazarus M., Eguchi N., Matsumoto S. Species-specific expression of microsomal prostaglandin E synthase-1 and cyclooxygenases in male monkey reproductive organs. Prostagland Leukot Essent Fatty Acids. 2004;71:233–240. doi: 10.1016/j.plefa.2004.03.018. [DOI] [PubMed] [Google Scholar]
- 617.Frungieri M.B., Gonzalez-Calvar S.I., Parborell F., Albrecht M., Mayerhofer A., Calandra R.S. Cyclooxygenase-2 and prostaglandin F2α in Syrian hamster Leydig cells: inhibitory role on luteinizing hormone/human chorionic gonadotropin-stimulated testosterone production. Endocrinology. 2006;147:4476–4485. doi: 10.1210/en.2006-0090. [DOI] [PubMed] [Google Scholar]
- 618.Balaji T., Ramanathan M., Padmanabhan Menon V. Localization of cyclooxygenase-2 in mice testis and assessment of its possible role through suppressing its expression using nimesulide: a preferential cyclooxygenase-2 inhibitor. Prostagland Leukot Essent Fatty Acids. 2007;76:341–348. doi: 10.1016/j.plefa.2007.04.006. [DOI] [PubMed] [Google Scholar]
- 619.Winnall W.R., Muir J.A., Liew S., Hirst J.J., Meachem S.J., Hedger M.P. Effects of chronic celecoxib on testicular function in normal and lipopolysaccharide-treated rats. Int J Androl. 2009;32:542–555. doi: 10.1111/j.1365-2605.2008.00895.x. [DOI] [PubMed] [Google Scholar]
- 620.Baker P.J., O’Shaughnessy P.J. Expression of prostaglandin D synthetase during development in the mouse testis. Reproduction. 2001;122:553–559. doi: 10.1530/rep.0.1220553. [DOI] [PubMed] [Google Scholar]
- 621.Wang X., Shen C.L., Dyson M.T. Cyclooxygenase-2 regulation of the age-related decline in testosterone biosynthesis. Endocrinology. 2005;146:4202–4208. doi: 10.1210/en.2005-0298. [DOI] [PubMed] [Google Scholar]
- 622.Langenbach R., Loftin C., Lee C., Tiano H. Cyclooxygenase knockout mice: models for elucidating isoform-specific functions. Biochem Pharmacol. 1999;58:1237–1246. doi: 10.1016/s0006-2952(99)00158-6. [DOI] [PubMed] [Google Scholar]
- 623.Abbatiello E.R., Kaminsky M., Weisbroth S. The effect of prostaglandins and prostaglandin inhibitors on spermatogenesis. Int J Fertil. 1975;20:177–182. [PubMed] [Google Scholar]
- 624.Kubota H., Sasaki S., Kubota Y. Cyclooxygenase-2 protects germ cells against spermatogenesis disturbance in experimental cryptorchidism model mice. J Androl. 2011;32:77–85. doi: 10.2164/jandrol.109.008888. [DOI] [PubMed] [Google Scholar]
- 625.Sullivan M.H., Cooke B.A. Control and production of leukotriene B4 in rat tumour and testicular Leydig cells. Biochem J. 1985;230:821–824. doi: 10.1042/bj2300821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 626.Papadopoulos V., Carreau S., Drosdowsky M.A. Effect of leukotriene B4 on purified rat Leydig cell function. Med Sci Res. 1987;15:11–12. [Google Scholar]
- 627.Matzkin M.E., Mayerhofer A., Rossi S.P. Cyclooxygenase-2 in testes of infertile men: evidence for the induction of prostaglandin synthesis by interleukin-1β. Fertil Steril. 2010;94:1933–1936. doi: 10.1016/j.fertnstert.2010.01.039. [DOI] [PubMed] [Google Scholar]
- 628.Stuart J.A., Brown M.F. Mitochondrial DNA maintenance and bioenergetics. Biochim Biophys Acta. 2006;1757:79–89. doi: 10.1016/j.bbabio.2006.01.003. [DOI] [PubMed] [Google Scholar]
- 629.Georgiou M., Perkins L.M., Payne A.H. Steroid synthesis-dependent, oxygen-mediated damage of mitochondrial and microsomal cytochrome P-450 enzymes in rat Leydig cell cultures. Endocrinology. 1987;121:1390–1399. doi: 10.1210/endo-121-4-1390. [DOI] [PubMed] [Google Scholar]
- 630.Quinn P.G., Payne A.H. Steroid product-induced, oxygen-mediated damage of microsomal cytochrome P-450 enzymes in Leydig cell cultures. Relationship to desensitization. J Biol Chem. 1985;260:2092–2099. [PubMed] [Google Scholar]
- 631.Forman H.J., Torres M. Reactive oxygen species and cell signaling: respiratory burst in macrophage signaling. Am J Respir Crit Care Med. 2002;166:S4–S8. doi: 10.1164/rccm.2206007. [DOI] [PubMed] [Google Scholar]
- 632.Nathan C. Role of iNOS in human host defense. Science. 2006;312:1874–1875. doi: 10.1126/science.312.5782.1874b. [DOI] [PubMed] [Google Scholar]
- 633.Droge W. Free radicals in the physiological control of cell function. Physiol Rev. 2002;82:47–95. doi: 10.1152/physrev.00018.2001. [DOI] [PubMed] [Google Scholar]
- 634.Niki E., Yoshida Y., Saito Y., Noguchi N. Lipid peroxidation: mechanisms, inhibition, and biological effects. Biochem Biophys Res Commun. 2005;338:668–676. doi: 10.1016/j.bbrc.2005.08.072. [DOI] [PubMed] [Google Scholar]
- 635.Sies H. Strategies of antioxidant defense. Eur J Biochem. 1993;215:213–219. doi: 10.1111/j.1432-1033.1993.tb18025.x. [DOI] [PubMed] [Google Scholar]
- 636.Simonian N.A., Coyle J.T. Oxidative stress in neurodegenerative diseases. Annu Rev Pharmacol Toxicol. 1996;36:83–106. doi: 10.1146/annurev.pa.36.040196.000503. [DOI] [PubMed] [Google Scholar]
- 637.Bogdan C. Nitric oxide and the immune response. Nat Immunol. 2001;2:907–916. doi: 10.1038/ni1001-907. [DOI] [PubMed] [Google Scholar]
- 638.Alderton W.K., Cooper C.E., Knowles R.G. Nitric oxide synthases: structure, function and inhibition. Biochem J. 2001;357:593–615. doi: 10.1042/0264-6021:3570593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 639.Davidoff M.S., Middendorff R., Mayer B., Holstein A.F. Nitric oxide synthase (NOS-I) in Leydig cells of the human testis. Arch Histol Cytol. 1995;58:17–30. doi: 10.1679/aohc.58.17. [DOI] [PubMed] [Google Scholar]
- 640.Lee N.P., Mruk D.D., Wong C.H., Cheng C.Y. Regulation of Sertoli-germ cell adherens junction dynamics in the testis via the nitric oxide synthase (NOS)/cGMP/protein kinase G (PRKG)/β-catenin (CATNB) signaling pathway: an in vitro and in vivo study. Biol Reprod. 2005;73:458–471. doi: 10.1095/biolreprod.105.040766. [DOI] [PubMed] [Google Scholar]
- 641.O’Bryan M.K., Schlatt S., Gerdprasert O., Phillips D.J., de Kretser D.M., Hedger M.P. Inducible nitric oxide synthase in the rat testis: evidence for potential roles in both normal function and inflammation-mediated infertility. Biol Reprod. 2000;63:1285–1293. doi: 10.1095/biolreprod63.5.1285. [DOI] [PubMed] [Google Scholar]
- 642.Zini A., O’Bryan M.K., Magid M.S., Schlegel P.N. Immunohistochemical localization of endothelial nitric oxide synthase in human testis, epididymis, and vas deferens suggests a possible role for nitric oxide in spermatogenesis, sperm maturation, and programmed cell death. Biol Reprod. 1996;55:935–941. doi: 10.1095/biolreprod55.5.935. [DOI] [PubMed] [Google Scholar]
- 643.Wang Y., Goligorsky M.S., Lin M., Wilcox J.N., Marsden P.A. A novel, testis-specific mRNA transcript encoding an NH2-terminal truncated nitric-oxide synthase. J Biol Chem. 1997;272:11392–11401. [PubMed] [Google Scholar]
- 644.Bauché F., Stéphan J.P., Touzalin A.M., Jégou B. In vitro regulation of an inducible-type NO synthase in the rat seminiferous tubule cells. Biol Reprod. 1998;58:431–438. doi: 10.1095/biolreprod58.2.431. [DOI] [PubMed] [Google Scholar]
- 645.Weissman B.A., Niu E., Ge R. Paracrine modulation of androgen synthesis in rat leydig cells by nitric oxide. J Androl. 2005;26:369–378. doi: 10.2164/jandrol.04178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 646.Guo J., Jia Y., Tao S.X. Expression of nitric oxide synthase during germ cell apoptosis in testis of cynomolgus monkey after testosterone and heat treatment. J Androl. 2009;30:190–199. doi: 10.2164/jandrol.108.005538. [DOI] [PubMed] [Google Scholar]
- 647.Lue Y., Sinha Hikim A.P., Wang C., Leung A., Swerdloff R.S. Functional role of inducible nitric oxide synthase in the induction of male germ cell apoptosis, regulation of sperm number, and determination of testes size: evidence from null mutant mice. Endocrinology. 2003;144:3092–3100. doi: 10.1210/en.2002-0142. [DOI] [PubMed] [Google Scholar]
- 648.Moon C., Ahn M., Kim S., Yasuzumi F., Shin T. Increased expression of both constitutive and inducible forms of nitric oxide synthase in the delayed phase of acute experimental testicular torsion. J Vet Med Sci. 2005;67:453–456. doi: 10.1292/jvms.67.453. [DOI] [PubMed] [Google Scholar]
- 649.Shiraishi K., Naito K., Yoshida K. Nitric oxide promotes germ cell necrosis in the delayed phase after experimental testicular torsion of rat. Biol Reprod. 2001;65:514–521. doi: 10.1095/biolreprod65.2.514. [DOI] [PubMed] [Google Scholar]
- 650.Auharek S.A., Lara N.L., Avelar G.F., Sharpe R.M., França L.R. Effects of inducible nitric oxide synthase (iNOS) deficiency in mice on Sertoli cell proliferation and perinatal testis development. Int J Androl. 2012;35:741–751. doi: 10.1111/j.1365-2605.2012.01264.x. [DOI] [PubMed] [Google Scholar]
- 651.Kondo Y., Ishikawa T., Yamaguchi K., Yada T., Fujisawa M. Oral administration of tetrahydrobiopterin attenuates testicular damage by reducing nitric oxide synthase activity in a cryptorchid mouse model. J Androl. 2008;29:153–163. doi: 10.2164/jandrol.107.002758. [DOI] [PubMed] [Google Scholar]
- 652.Allen J.A., Diemer T., Janus P., Hales K.H., Hales D.B. Bacterial endotoxin lipopolysaccharide and reactive oxygen species inhibit Leydig cell steroidogenesis via perturbation of mitochondria. Endocrine. 2004;25:265–275. doi: 10.1385/ENDO:25:3:265. [DOI] [PubMed] [Google Scholar]
- 653.Diemer T., Allen J.A., Hales K.H., Hales D.B. Reactive oxygen disrupts mitochondria in MA-10 tumor Leydig cells and inhibits steroidogenic acute regulatory (StAR) protein and steroidogenesis. Endocrinology. 2003;144:2882–2891. doi: 10.1210/en.2002-0090. [DOI] [PubMed] [Google Scholar]
- 654.Stocco D.M., Wells J., Clark B.J. The effects of hydrogen peroxide on steroidogenesis in mouse Leydig tumor cells. Endocrinology. 1993;133:2827–2832. doi: 10.1210/endo.133.6.8243310. [DOI] [PubMed] [Google Scholar]
- 655.Hjertkvist M., Läckgren G., Plöen L., Bergh A. Does HCG treatment induce inflammation-like changes in undescended testes in boys? J Pediatr Surg. 1993;28:254–258. doi: 10.1016/s0022-3468(05)80288-x. [DOI] [PubMed] [Google Scholar]
- 656.Del Punta K., Charreau E.H., Pignataro O.P. Nitric oxide inhibits Leydig cell steroidogenesis. Endocrinology. 1996;137:5337–5343. doi: 10.1210/endo.137.12.8940355. [DOI] [PubMed] [Google Scholar]
- 657.Kostic T.S., Andric S.A., Maric D., Kovacevic R.Z. Inhibitory effects of stress-activated nitric oxide on antioxidant enzymes and testicular steroidogenesis. J Steroid Biochem Mol Biol. 2000;75:299–306. doi: 10.1016/s0960-0760(00)00185-0. [DOI] [PubMed] [Google Scholar]
- 658.Sharma A.C., Sam A.D., 2nd, Lee L.Y. Effect of NG-nitro-L-arginine methyl ester on testicular blood flow and serum steroid hormones during sepsis. Shock. 1998;9:416–421. doi: 10.1097/00024382-199806000-00005. [DOI] [PubMed] [Google Scholar]
- 659.Welch C., Watson M.E., Poth M., Hong T., Francis G.L. Evidence to suggest nitric oxide is an interstitial regulator of Leydig cell steroidogenesis. Metab Clin Exper. 1995;44:234–238. doi: 10.1016/0026-0495(95)90271-6. [DOI] [PubMed] [Google Scholar]
- 660.Cymeryng C.B., Dada L.A., Podesta E.J. Effect of nitric oxide on rat adrenal zona fasciculata steroidogenesis. J Endocrinol. 1998;158:197–203. doi: 10.1677/joe.0.1580197. [DOI] [PubMed] [Google Scholar]
- 661.Sharpe R.M. Local control of testicular function. Q J Exp Physiol. 1983;68:265–287. doi: 10.1113/expphysiol.1983.sp002723. [DOI] [PubMed] [Google Scholar]
- 662.Björkbacka H., Kunjathoor V.V., Moore K.J. Reduced atherosclerosis in MyD88-null mice links elevated serum cholesterol levels to activation of innate immunity signaling pathways. Nat Med. 2004;10:416–421. doi: 10.1038/nm1008. [DOI] [PubMed] [Google Scholar]
- 663.Jiang D., Liang J., Fan J. Regulation of lung injury and repair by Toll-like receptors and hyaluronan. Nat Med. 2005;11:1173–1179. doi: 10.1038/nm1315. [DOI] [PubMed] [Google Scholar]
- 664.Rakoff-Nahoum S., Paglino J., Eslami-Varzaneh F., Edberg S., Medzhitov R. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell. 2004;118:229–241. doi: 10.1016/j.cell.2004.07.002. [DOI] [PubMed] [Google Scholar]
- 665.Zetterström C.K., Strand M.L., Söder O. The high mobility group box chromosomal protein 1 is expressed in the human and rat testis where it may function as an antibacterial factor. Hum Reprod. 2006;21:2801–2809. doi: 10.1093/humrep/del256. [DOI] [PubMed] [Google Scholar]
- 666.Oldereid N.B., De Angelis P., Wiger R., Clausen O.P. Expression of Bcl-2 family proteins and spontaneous apoptosis in normal human testis. Mol Hum Reprod. 2001;7:403–408. doi: 10.1093/molehr/7.5.403. [DOI] [PubMed] [Google Scholar]
- 667.Zhang X., Wang T., Deng T. Damaged spermatogenic cells induce inflammatory gene expression in mouse Sertoli cells through the activation of Toll-like receptors 2 and 4. Mol Cell Endocrinol. 2013;365:162–173. doi: 10.1016/j.mce.2012.10.016. [DOI] [PubMed] [Google Scholar]
- 668.Kangasniemi M., Kaipia A., Mali P., Toppari J., Huhtaniemi I., Parvinen M. Modulation of basal and FSH-dependent cyclic AMP production in rat seminiferous tubules staged by an improved transillumination technique. Anat Rec. 1990;227:62–76. doi: 10.1002/ar.1092270108. [DOI] [PubMed] [Google Scholar]
- 669.O’Donnell L., Pratis K., Wagenfeld A. Transcriptional profiling of the hormone-responsive stages of spermatogenesis reveals cell-, stage-, and hormone-specific events. Endocrinology. 2009;150:5074–5084. doi: 10.1210/en.2009-0755. [DOI] [PubMed] [Google Scholar]
- 670.Meinhardt A., O’Bryan M.K., McFarlane J.R. Localization of follistatin in the rat testis. J Reprod Fertil. 1998;112:233–241. doi: 10.1530/jrf.0.1120233. [DOI] [PubMed] [Google Scholar]
- 671.Cohen P.E., Pollard J.W. Normal sexual function in male mice lacking a functional type I interleukin-1 (IL-1) receptor. Endocrinology. 1998;139:815–818. doi: 10.1210/endo.139.2.5914. [DOI] [PubMed] [Google Scholar]
- 672.Morales-Montor J., Baig S., Mitchell R., Deway K., Hallal-Calleros C., Damian R.T. Immunoendocrine interactions during chronic cysticercosis determine male mouse feminization: role of IL-6. J Immunol. 2001;167:4527–4533. doi: 10.4049/jimmunol.167.8.4527. [DOI] [PubMed] [Google Scholar]
- 673.Pasparakis M., Alexopoulou L., Episkopou V., Kollias G. Immune and inflammatory responses in TNFα-deficient mice: a critical requirement for TNFα in the formation of primary B cell follicles, follicular dendritic cell networks and germinal centers, and in the maturation of the humoral immune response. J Exp Med. 1996;184:1397–1411. doi: 10.1084/jem.184.4.1397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 674.Fattori E., Cappelletti M., Costa P. Defective inflammatory response in interleukin 6-deficient mice. J Exp Med. 1994;180:1243–1250. doi: 10.1084/jem.180.4.1243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 675.Reed M.J., Coldham N.G., Patel S.R., Ghilchik M.W., James V.H. Interleukin-1 and interleukin-6 in breast cyst fluid: their role in regulating aromatase activity in breast cancer cells. J Endocrinol. 1992;132:R5–R8. doi: 10.1677/joe.0.132r005. [DOI] [PubMed] [Google Scholar]
- 676.Kerr J.B., Sharpe R.M. Macrophage activation enhances the human chorionic gonadotrophin-induced disruption of spermatogenesis in the rat. J Endocrinol. 1989;121:285–292. doi: 10.1677/joe.0.1210285. [DOI] [PubMed] [Google Scholar]
- 677.Nes W.D., Lukyanenko Y.O., Jia Z.H. Identification of the lipophilic factor produced by macrophages that stimulates steroidogenesis. Endocrinology. 2000;141:953–958. doi: 10.1210/endo.141.3.7350. [DOI] [PubMed] [Google Scholar]
- 678.Suescun M.O., Calandra R.S., Lustig L. Effect of testicular macrophage conditioned media from rats with autoimmune orchitis on Leydig cell function. Am J Reprod Immunol. 2000;43:116–123. doi: 10.1111/j.8755-8920.2000.430208.x. [DOI] [PubMed] [Google Scholar]
- 679.Kmicikiewicz I., Wojtusiak A., Bilinska B. The effect of testicular macrophages, macrophage-conditioned medium and interleukin-1α on bank vole Leydig cell steroidogenesis. Exp Clin Endocrinol Diabetes. 1999;107:262–271. doi: 10.1055/s-0029-1212110. [DOI] [PubMed] [Google Scholar]
- 680.Chen J.J., Lukyanenko Y., Hutson J.C. 25-hydroxycholesterol is produced by testicular macrophages during the early postnatal period and influences differentiation of Leydig cells in vitro. Biol Reprod. 2002;66:1336–1341. doi: 10.1095/biolreprod66.5.1336. [DOI] [PubMed] [Google Scholar]
- 681.Lukyanenko Y., Chen J.J., Hutson J.C. Testosterone regulates 25-hydroxycholesterol production in testicular macrophages. Biol Reprod. 2002;67:1435–1438. doi: 10.1095/biolreprod.102.007575. [DOI] [PubMed] [Google Scholar]
- 682.Haider S.G. Cell biology of Leydig cells in the testis. Int Rev Cytol. 2004;233:181–241. doi: 10.1016/S0074-7696(04)33005-6. [DOI] [PubMed] [Google Scholar]
- 683.Martens H.F., Sheets P.K., Tenover J.S., Dugowson C.E., Bremner W.J., Starkebaum G. Decreased testosterone levels in men with rheumatoid arthritis: effect of low dose prednisone therapy. J Rheumatol. 1994;21:1427–1431. [PubMed] [Google Scholar]
- 684.Spratt D.I., Cox P., Orav J., Moloney J., Bigos T. Reproductive axis suppression in acute illness is related to disease severity. J Clin Endocrinol Metab. 1993;76:1548–1554. doi: 10.1210/jcem.76.6.8501163. [DOI] [PubMed] [Google Scholar]
- 685.Calandra T., Bochud P.Y., Heumann D. Cytokines in septic shock. Curr Clin Top Infect Dis. 2002;22:1–23. [PubMed] [Google Scholar]
- 686.Bruot B.C., Clemens J.W. Regulation of testosterone production in the adjuvant-induced arthritic rat. J Androl. 1992;13:87–92. [PubMed] [Google Scholar]
- 687.Clemens J.W., Bruot B.C. Testicular dysfunction in the adjuvant-induced arthritic rat. J Androl. 1989;10:419–424. doi: 10.1002/j.1939-4640.1989.tb00130.x. [DOI] [PubMed] [Google Scholar]
- 688.Wallgren M., Kindahl H., Larsson K. Clinical, endocrinological and spermatological studies after endotoxin in the ram. Zentralbl Veterinarmed A. 1989;36:90–103. doi: 10.1111/j.1439-0442.1989.tb00708.x. [DOI] [PubMed] [Google Scholar]
- 689.Hales K.H., Diemer T., Ginde S. Diametric effects of bacterial endotoxin lipopolysaccharide on adrenal and Leydig cell steroidogenic acute regulatory protein. Endocrinology. 2000;141:4000–4012. doi: 10.1210/endo.141.11.7780. [DOI] [PubMed] [Google Scholar]
- 690.Bosmann H.B., Hales K.H., Li X., Liu Z., Stocco D.M., Hales D.B. Acute in vivo inhibition of testosterone by endotoxin parallels loss of steroidogenic acute regulatory (StAR) protein in Leydig cells. Endocrinology. 1996;137:4522–4525. doi: 10.1210/endo.137.10.8828518. [DOI] [PubMed] [Google Scholar]
- 691.Reddy M.M., Mahipal S.V., Subhashini J. Bacterial lipopolysaccharide-induced oxidative stress in the impairment of steroidogenesis and spermatogenesis in rats. Reprod Toxicol. 2006;22:493–500. doi: 10.1016/j.reprotox.2006.03.003. [DOI] [PubMed] [Google Scholar]
- 692.Sewer M.B., Morgan E.T. Down-regulation of the expression of three major rat liver cytochrome P450S by endotoxin in vivo occurs independently of nitric oxide production. J Pharmacol Exp Ther. 1998;287:352–358. [PubMed] [Google Scholar]
- 693.Wallgren M. Clinical, endocrinological and spermatological studies after endotoxin injection in the boar. Zentralbl Veterinarmed A. 1989;36:664–675. doi: 10.1111/j.1439-0442.1989.tb00778.x. [DOI] [PubMed] [Google Scholar]
- 694.Wallgren M., Kindahl H., Rodriguez-Martinez H. Alterations in testicular function after endotoxin injection in the boar. Int J Androl. 1993;16:235–243. doi: 10.1111/j.1365-2605.1993.tb01185.x. [DOI] [PubMed] [Google Scholar]
- 695.Wallgren M., Kindahl H., Rodriguez-Martinez H. Modulation of endotoxin-induced andrological alterations by flunixin meglumine in the boar. Zentralbl Veterinarmed A. 1995;42:357–369. doi: 10.1111/j.1439-0442.1995.tb00388.x. [DOI] [PubMed] [Google Scholar]
- 696.Liew S.H., Meachem S.J., Hedger M.P. A stereological analysis of the response of spermatogenesis to an acute inflammatory episode in adult rats. J Androl. 2007;28:176–185. doi: 10.2164/jandrol.106.000752. [DOI] [PubMed] [Google Scholar]
- 697.van der Poll T., Romijn J.A., Endert E., Sauerwein H.P. Effects of tumor necrosis factor on the hypothalamic-pituitary-testicular axis in healthy men. Metabolism. 1993;42:303–307. doi: 10.1016/0026-0495(93)90078-3. [DOI] [PubMed] [Google Scholar]
- 698.Tsigos C., Papanicolaou D.A., Kyrou I., Raptis S.A., Chrousos G.P. Dose-dependent effects of recombinant human interleukin-6 on the pituitary-testicular axis. J Interferon Cytokine Res. 1999;19:1271–1276. doi: 10.1089/107999099312948. [DOI] [PubMed] [Google Scholar]
- 699.Meikle A.W., Cardoso de Sousa J.C., Ward J.H., Woodward M., Samlowski W.E. Reduction of testosterone synthesis after high dose interleukin-2 therapy of metastatic cancer. J Clin Endocrinol Metab. 1991;73:931–935. doi: 10.1210/jcem-73-5-931. [DOI] [PubMed] [Google Scholar]
- 700.Tsigos C., Kyrou I., Chala E. Circulating tumor necrosis factor α concentrations are higher in abdominal versus peripheral obesity. Metabolism. 1999;48:1332–1335. doi: 10.1016/s0026-0495(99)90277-9. [DOI] [PubMed] [Google Scholar]
- 701.Donath M.Y., Shoelson S.E. Type 2 diabetes as an inflammatory disease. Nat Rev Immunol. 2011;11:98–107. doi: 10.1038/nri2925. [DOI] [PubMed] [Google Scholar]
- 702.Hutson J.C. Secretion of tumor necrosis factor alpha by testicular macrophages. J Reprod Immunol. 1993;23:63–72. doi: 10.1016/0165-0378(93)90027-f. [DOI] [PubMed] [Google Scholar]
- 703.Guo H., Calkins J.H., Sigel M.M., Lin T. Interleukin-2 is a potent inhibitor of Leydig cell steroidogenesis. Endocrinology. 1990;127:1234–1239. doi: 10.1210/endo-127-3-1234. [DOI] [PubMed] [Google Scholar]
- 704.Nedwin G.E., Svedersky L.P., Bringman T.S., Palladino M.A., Jr., Goeddel D.V. Effect of interleukin 2, interferon-γ, and mitogens on the production of tumor necrosis factors α and β. J Immunol. 1985;135:2492–2497. [PubMed] [Google Scholar]
- 705.Kasahara T., Hooks J.J., Dougherty S.F., Oppenheim J.J. Interleukin-2-mediated immune interferon (IFN-γ) production by human T cells and T cell subsets. J Immunol. 1983;130:1784–1789. [PubMed] [Google Scholar]
- 706.Sowers J.R., Rice B.F., Blanchard S. Effect of dexamethsone on luteinizing hormone and follicle stimulating hormone responses to LHRH and to clomiphene in the follicular phase of women with normal menstrual cycles. Horm Metab Res. 1979;11:478–480. doi: 10.1055/s-0028-1092765. [DOI] [PubMed] [Google Scholar]
- 707.Stalker A., Hermo L., Antakly T. Covalent affinity labeling, radioautography, and immunocytochemistry localize the glucocorticoid receptor in rat testicular Leydig cells. Am J Anat. 1989;186:369–377. doi: 10.1002/aja.1001860406. [DOI] [PubMed] [Google Scholar]
- 708.Hales D.B., Payne A.H. Glucocorticoid-mediated repression of P450scc mRNA and de novo synthesis in cultured Leydig cells. Endocrinology. 1989;124:2099–2104. doi: 10.1210/endo-124-5-2099. [DOI] [PubMed] [Google Scholar]
- 709.Sapolsky R.M. Stress-induced suppression of testicular function in the wild baboon: role of glucocorticoids. Endocrinology. 1985;116:2273–2278. doi: 10.1210/endo-116-6-2273. [DOI] [PubMed] [Google Scholar]
- 710.Rivier C., Rivier J., Vale W. Stress-induced inhibition of reproductive functions: role of endogenous corticotropin-releasing factor. Science. 1986;231:607–609. doi: 10.1126/science.3003907. [DOI] [PubMed] [Google Scholar]
- 711.Battaglia D.F., Bowen J.M., Krasa H.B., Thrun L.A., Viguie C., Karsch F.J. Endotoxin inhibits the reproductive neuroendocrine axis while stimulating adrenal steroids: a simultaneous view from hypophyseal portal and peripheral blood. Endocrinology. 1997;138:4273–4281. doi: 10.1210/endo.138.10.5449. [DOI] [PubMed] [Google Scholar]
- 712.Refojo D., Arias P., Moguilevsky J.A., Feleder C. Effect of bacterial endotoxin on in vivo pulsatile gonadotropin secretion in adult male rats. Neuroendocrinology. 1998;67:275–281. doi: 10.1159/000054323. [DOI] [PubMed] [Google Scholar]
- 713.Rivest S., Rivier C. Centrally injected interleukin-1β inhibits the hypothalamic LHRH secretion and circulating LH levels via prostaglandins in rats. J Neuroendocrinol. 1993;5:445–450. doi: 10.1111/j.1365-2826.1993.tb00506.x. [DOI] [PubMed] [Google Scholar]
- 714.Russell S.H., Small C.J., Stanley S.A., Franks S., Ghatei M.A., Bloom S.R. The in vitro role of tumour necrosis factor-alpha and interleukin-6 in the hypothalamic-pituitary gonadal axis. J Neuroendocrinol. 2001;13:296–301. doi: 10.1046/j.1365-2826.2001.00632.x. [DOI] [PubMed] [Google Scholar]
- 715.Yu W.H., Walczewska A., Karanth S., McCann S.M. Nitric oxide mediates leptin-induced luteinizing hormone-releasing hormone (LHRH) and LHRH and leptin-induced LH release from the pituitary gland. Endocrinology. 1997;138:5055–5058. doi: 10.1210/endo.138.11.5649. [DOI] [PubMed] [Google Scholar]
- 716.Gaytán F., Bellido C., Aguilar R., Morales C., van Rooijen N., Aguilar E. Role of the testis in the response of the pituitary-testicular axis to nitric oxide-related agents. Eur J Endocrinol. 1997;137:301–308. doi: 10.1530/eje.0.1370301. [DOI] [PubMed] [Google Scholar]
- 717.Ogilvie K.M., Held Hales K., Roberts M.E., Hales D.B., Rivier C. The inhibitory effect of intracerebroventricularly injected interleukin 1β on testosterone secretion in the rat: role of steroidogenic acute regulatory protein. Biol Reprod. 1999;60:527–533. doi: 10.1095/biolreprod60.2.527. [DOI] [PubMed] [Google Scholar]
- 718.Turnbull A.V., Rivier C. Inhibition of gonadotropin-induced testosterone secretion by the intracerebroventricular injection of interleukin-1β in the male rat. Endocrinology. 1997;138:1008–1013. doi: 10.1210/endo.138.3.5019. [DOI] [PubMed] [Google Scholar]
- 719.Lee S., Miselis R., Rivier C. Anatomical and functional evidence for a neural hypothalamic-testicular pathway that is independent of the pituitary. Endocrinology. 2002;143:4447–4454. doi: 10.1210/en.2002-220392. [DOI] [PubMed] [Google Scholar]
- 720.Selvage D.J., Lee S.Y., Parsons L.H., Seo D.O., Rivier C.L. A hypothalamic-testicular neural pathway is influenced by brain catecholamines, but not testicular blood flow. Endocrinology. 2004;145:1750–1759. doi: 10.1210/en.2003-1441. [DOI] [PubMed] [Google Scholar]
- 721.Herman M., Rivier C. Activation of a neural brain-testicular pathway rapidly lowers Leydig cell levels of the steroidogenic acute regulatory protein and the peripheral-type benzodiazepine receptor while increasing levels of neuronal nitric oxide synthase. Endocrinology. 2006;147:624–633. doi: 10.1210/en.2005-0879. [DOI] [PubMed] [Google Scholar]
- 722.Setchell B.P., Galil K.A. Limitations imposed by testicular blood flow on the function of Leydig cells in rats in vivo. Aust J Biol Sci. 1983;36:285–293. doi: 10.1071/bi9830285. [DOI] [PubMed] [Google Scholar]
- 723.Anakwe O.O., Moger W.H. Catecholamine stimulation of androgen production by rat Leydig cells. Interactions with luteinizing hormone and luteinizing hormone-releasing hormone. Biol Reprod. 1986;35:806–814. doi: 10.1095/biolreprod35.4.806. [DOI] [PubMed] [Google Scholar]
- 724.Middendorff R., Davidoff M., Holstein A.F. Neuroendocrine marker substances in human Leydig cells–changes by disturbances of testicular function. Andrologia. 1993;25:257–262. doi: 10.1111/j.1439-0272.1993.tb02722.x. [DOI] [PubMed] [Google Scholar]
- 725.Tinajero J.C., Fabbri A., Dufau M.L. Serotonergic inhibition of rat Leydig cell function by propranolol. Endocrinology. 1993;133:257–264. doi: 10.1210/endo.133.1.8391422. [DOI] [PubMed] [Google Scholar]
- 726.Mayerhofer A., Frungieri M.B., Bulling A., Fritz S. Sources and function of neuronal signalling molecules in the gonads. Medicina. 1999;59:542–545. [PubMed] [Google Scholar]
- 727.Lysiak J.J., Nguyen Q.A., Kirby J.L., Turner T.T. Ischemia-reperfusion of the murine testis stimulates the expression of proinflammatory cytokines and activation of c-jun N-terminal kinase in a pathway to E-selectin expression. Biol Reprod. 2003;69:202–210. doi: 10.1095/biolreprod.102.013318. [DOI] [PubMed] [Google Scholar]
- 728.Minutoli L., Antonuccio P., Polito F. Involvement of mitogen-activated protein kinases (MAPKs) during testicular ischemia-reperfusion injury in nuclear factor-κB knock-out mice. Life Sci. 2007;81:413–422. doi: 10.1016/j.lfs.2007.06.016. [DOI] [PubMed] [Google Scholar]
- 729.Turner T.T., Bang H.J., Lysiak J.L. The molecular pathology of experimental testicular torsion suggests adjunct therapy to surgical repair. J Urol. 2004;172:2574–2578. doi: 10.1097/01.ju.0000144203.30718.19. [DOI] [PubMed] [Google Scholar]
- 730.De Stefani S., Silingardi V., Micali S. Experimental varicocele in the rat: early evaluation of the nitric oxide levels and histological alterations in the testicular tissue. Andrologia. 2005;37:115–118. doi: 10.1111/j.1439-0272.2005.00660.x. [DOI] [PubMed] [Google Scholar]
- 731.Mondillo C., Pagotto R.M., Piotrkowski B. Involvement of nitric oxide synthase in the mechanism of histamine-induced inhibition of Leydig cell steroidogenesis via histamine receptor subtypes in Sprague-Dawley rats. Biol Reprod. 2009;80:144–152. doi: 10.1095/biolreprod.108.069484. [DOI] [PubMed] [Google Scholar]
- 732.Hedger M.P., Khatab S., Gonzales G., de Kretser D.M. Acute and short-term actions of serotonin administration on the pituitary-testicular axis in the adult rat. Reprod Fertil Dev. 1995;7:1101–1109. doi: 10.1071/rd9951101. [DOI] [PubMed] [Google Scholar]
- 733.Collodel G., Castellini C., Del Vecchio M. Effect of a bacterial lipopolysaccharide treatment on rabbit testis and ejaculated sperm. Reprod Domest Anim. 2012;47:372–378. doi: 10.1111/j.1439-0531.2011.01882.x. [DOI] [PubMed] [Google Scholar]
- 734.McLachlan R.I., Wreford N.G., O’Donnell L., de Kretser D.M., Robertson D.M. The endocrine regulation of spermatogenesis: independent roles for testosterone and FSH. J Endocrinol. 1996;148:1–9. doi: 10.1677/joe.0.1480001. [DOI] [PubMed] [Google Scholar]
- 735.O’Donnell L., McLachlan R.I., Wreford N.G., de Kretser D.M., Robertson D.M. Testosterone withdrawal promotes stage-specific detachment of round spermatids from the rat seminiferous epithelium. Biol Reprod. 1996;55:895–901. doi: 10.1095/biolreprod55.4.895. [DOI] [PubMed] [Google Scholar]
- 736.Tulassay Z., Viczián M., Böjthe L., Czeizel A. Quantitative histological studies on the injury of spermatogenesis induced by endotoxin in rats. J Reprod Fertil. 1970;22:161–164. doi: 10.1530/jrf.0.0220161. [DOI] [PubMed] [Google Scholar]
- 737.Takao T., Culp S.G., De Souza E.B. Reciprocal modulation of interleukin-1β (IL-1β) and IL-1 receptors by lipopolysaccharide (endotoxin) treatment in the mouse brain-endocrine-immune axis. Endocrinology. 1993;132:1497–1504. doi: 10.1210/endo.132.4.8462448. [DOI] [PubMed] [Google Scholar]
- 738.Li L., Ma P., Liu Y. Intermedin attenuates LPS-induced inflammation in the rat testis. PLoS One. 2013;8:e65278. doi: 10.1371/journal.pone.0065278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 739.Abu Elhija M., Potashnik H., Lunenfeld E. Testicular interleukin-6 response to systemic inflammation. Eur Cytokine Netw. 2005;16:167–172. [PubMed] [Google Scholar]
- 740.Metukuri M.R., Reddy C.M., Reddy P.R., Reddanna P. Bacterial LPS mediated acute inflammation-induced spermatogenic failure in rats: role of stress response proteins and mitochondrial dysfunction. Inflammation. 2010;33:235–243. doi: 10.1007/s10753-009-9177-4. [DOI] [PubMed] [Google Scholar]
- 741.Aly H.A., Lightfoot D.A., El-Shemy H.A. Modulatory role of lipoic acid on lipopolysaccharide-induced oxidative stress in adult rat Sertoli cells in vitro. Chem Biol Interact. 2009;182:112–118. doi: 10.1016/j.cbi.2009.08.013. [DOI] [PubMed] [Google Scholar]
- 742.Ashour A.E., Abdel-Hamied H.E., Korashy H.M., Al-Shabanah O.A., Abd-Allah A.R. Alpha-lipoic acid rebalances redox and immune-testicular milieu in septic rats. Chem Biol Interact. 2011;189:198–205. doi: 10.1016/j.cbi.2010.12.023. [DOI] [PubMed] [Google Scholar]
- 743.Nagaosa K., Nakashima C., Kishimoto A., Nakanishi Y. Immune response to bacteria in seminiferous epithelium. Reproduction. 2009;137:879–888. doi: 10.1530/REP-08-0460. [DOI] [PubMed] [Google Scholar]
- 744.Turner T.T., Brown K.J. Spermatic cord torsion: loss of spermatogenesis despite return of blood flow. Biol Reprod. 1993;49:401–407. doi: 10.1095/biolreprod49.2.401. [DOI] [PubMed] [Google Scholar]
- 745.Bergh A., Collin O., Lissbrant E. Effects of acute graded reductions in testicular blood flow on testicular morphology in the adult rat. Biol Reprod. 2001;64:13–20. doi: 10.1095/biolreprod64.1.13. [DOI] [PubMed] [Google Scholar]
- 746.Lewis-Jones D.I., Moreno de Marval M., Harrison R.G. Impairment of rat spermatogenesis following unilateral experimental ischemia. Fertil Steril. 1982;38:482–490. doi: 10.1016/s0015-0282(16)46585-4. [DOI] [PubMed] [Google Scholar]
- 747.Sukhotnik I., Miselevich I., Lurie M., Nativ O., Coran A.G., Mogilner J.G. The time relationship between ipsilateral testicular ischemia and germ cell apoptosis in the contralateral testis in rat. Pediatr Surg Int. 2005;21:512–516. doi: 10.1007/s00383-005-1477-7. [DOI] [PubMed] [Google Scholar]
- 748.Schanaider A., Aiex C.A., Errico G. Immunological effects of acute testicular torsion on the contralateral testis in rats. Eur J Pediatr Surg. 2011;21:370–374. doi: 10.1055/s-0031-1285841. [DOI] [PubMed] [Google Scholar]
- 749.Kolettis P.N., Stowe N.T., Inman S.R., Thomas A.J., Jr. Acute spermatic cord torsion alters the microcirculation of the contralateral testis. J Urol. 1996;155:350–354. [PubMed] [Google Scholar]
- 750.Altavilla D., Romeo C., Squadrito F. Molecular pathways involved in the early and late damage induced by testis ischemia: evidence for a rational pharmacological modulation. Curr Med Chem. 2012;19:1219–1224. doi: 10.2174/092986712799320538. [DOI] [PubMed] [Google Scholar]
- 751.Minutoli L., Antonuccio P., Polito F. Mitogen-activated protein kinase 3/mitogen-activated protein kinase 1 activates apoptosis during testicular ischemia-reperfusion injury in a nuclear factor-κB-independent manner. Eur J Pharmacol. 2009;604:27–35. doi: 10.1016/j.ejphar.2008.12.028. [DOI] [PubMed] [Google Scholar]
- 752.Rodriguez M.G., Rival C., Theas M.S., Lustig L. Immunohistopathology of the contralateral testis of rats undergoing experimental torsion of the spermatic cord. Asian J Androl. 2006;8:576–583. doi: 10.1111/j.1745-7262.2006.00146.x. [DOI] [PubMed] [Google Scholar]
- 753.Koc A., Narci A., Duru M., Gergerlioglu H.S., Akaydin Y., Sogut S. The protective role of erdosteine on testicular tissue after testicular torsion and detorsion. Mol Cell Biochem. 2005;280:193–199. doi: 10.1007/s11010-005-8911-y. [DOI] [PubMed] [Google Scholar]
- 754.Mogilner J.G., Elenberg Y., Lurie M., Shiloni E., Coran A.G., Sukhotnik I. Effect of dexamethasone on germ cell apoptosis in the contralateral testis after testicular ischemia-reperfusion injury in the rat. Fertil Steril. 2006;85(Suppl. 1):1111–1117. doi: 10.1016/j.fertnstert.2005.10.021. [DOI] [PubMed] [Google Scholar]
- 755.Üstün H., Akgül K.T., Ayyildiz A. Effect of phospodiesterase 5 inhibitors on apoptosis and nitric oxide synthases in testis torsion: an experimental study. Pediatr Surg Int. 2008;24:205–211. doi: 10.1007/s00383-007-2058-8. [DOI] [PubMed] [Google Scholar]
- 756.Sukhotnik I., Greenblatt R., Voskoboinik K., Lurie M., Coran A.G., Mogilner J.G. Relationship between time of reperfusion and E-selectin expression, neutrophil recruitment, and germ cell apoptosis after testicular ischemia in a rat model. Fertil Steril. 2008;90:1517–1522. doi: 10.1016/j.fertnstert.2007.07.1334. [DOI] [PubMed] [Google Scholar]
- 757.Tsounapi P., Saito M., Dimitriadis F. Protective effect of sivelestat, a neutrophil elastase inhibitor, on ipsilateral and contralateral testes after unilateral testicular ischaemia-reperfusion injury in rats. BJU Int. 2011;107:329–336. doi: 10.1111/j.1464-410X.2010.09481.x. [DOI] [PubMed] [Google Scholar]
- 758.Lysiak J.J., Turner S.D., Nguyen Q.A., Singbartl K., Ley K., Turner T.T. Essential role of neutrophils in germ cell-specific apoptosis following ischemia/reperfusion injury of the mouse testis. Biol Reprod. 2001;65:718–725. doi: 10.1095/biolreprod65.3.718. [DOI] [PubMed] [Google Scholar]
- 759.Lee J., Binsaleh S., Lo K., Jarvi K. Varicoceles: the diagnostic dilemma. J Androl. 2008;29:143–146. doi: 10.2164/jandrol.107.003467. [DOI] [PubMed] [Google Scholar]
- 760.Sahin Z., Celik-Ozenci C., Akkoyunlu G. Increased expression of interleukin-1α and interleukin-1β is associated with experimental varicocele. Fertil Steril. 2006;85(Suppl. 1):1265–1275. doi: 10.1016/j.fertnstert.2005.10.025. [DOI] [PubMed] [Google Scholar]
- 761.Shiraishi K., Naito K. Nitric oxide produced in the testis is involved in dilatation of the internal spermatic vein that compromises spermatogenesis in infertile men with varicocele. BJU Int. 2007;99:1086–1090. doi: 10.1111/j.1464-410X.2007.06800.x. [DOI] [PubMed] [Google Scholar]
- 762.Koksal I.T., Ishak Y., Usta M. Varicocele-induced testicular dysfunction may be associated with disruption of blood-testis barrier. Arch Androl. 2007;53:43–48. doi: 10.1080/01485010600822606. [DOI] [PubMed] [Google Scholar]
- 763.Celik O., Kutlu O., Tekcan M., Celik-Ozenci C., Koksal I.T. Role of TNF-related apoptosis-inducing ligand (TRAIL) in the pathogenesis of varicocele-induced testicular dysfunction. Asian J Androl. 2013;15:269–274. doi: 10.1038/aja.2012.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 764.Agarwal A., Hamada A., Esteves S.C. Insight into oxidative stress in varicocele-associated male infertility: part 1. Nat Rev Urol. 2012;9:678–690. doi: 10.1038/nrurol.2012.197. [DOI] [PubMed] [Google Scholar]
- 765.Setchell B.P., Sharpe R.M. Effect of injected human chorionic gonadotrophin on capillary permeability, extracellular fluid volume and the flow of lymph and blood in the testes of rats. J Endocrinol. 1981;91:245–254. doi: 10.1677/joe.0.0910245. [DOI] [PubMed] [Google Scholar]
- 766.Widmark A., Bergh A., Damber J.E., Smedegård G. Leucocytes mediate the hCG-induced increase in testicular venular permeability. Mol Cell Endocrinol. 1987;53:25–31. doi: 10.1016/0303-7207(87)90188-2. [DOI] [PubMed] [Google Scholar]
- 767.Bergh A., Widmark A., Damber J.E., Cajander S. Are leukocytes involved in the human chorionic gonadotropin-induced increase in testicular vascular permeability? Endocrinology. 1986;119:586–590. doi: 10.1210/endo-119-2-586. [DOI] [PubMed] [Google Scholar]
- 768.Assmus M., Svechnikov K., von Euler M. Single subcutaneous administration of chorionic gonadotropin to rats induces a rapid and transient increase in testicular expression of pro-inflammatory cytokines. Pediatr Res. 2005;57:896–901. doi: 10.1203/01.PDR.0000161410.30145.15. [DOI] [PubMed] [Google Scholar]
- 769.Kerr J.B., Sharpe R.M. Focal disruption of spermatogenesis in the testis of adult rats after a single administration of human chorionic gonadotrophin. Cell Tissue Res. 1989;257:163–169. doi: 10.1007/BF00221647. [DOI] [PubMed] [Google Scholar]
- 770.Setchell B.P., Rommerts F.F. The importance of the Leydig cells in the vascular response to hCG in the rat testis. Int J Androl. 1985;8:436–440. doi: 10.1111/j.1365-2605.1985.tb00857.x. [DOI] [PubMed] [Google Scholar]
- 771.Bergh A., Damber J.E., Hjertkvist M. Human chorionic gonadotrophin-induced testicular inflammation may be related to increased sensitivity to interleukin-1. Int J Androl. 1996;19:229–236. doi: 10.1111/j.1365-2605.1996.tb00467.x. [DOI] [PubMed] [Google Scholar]
- 772.Ohta Y., Nishikawa A., Fukazawa Y. Apoptosis in adult mouse testis induced by experimental cryptorchidism. Acta Anat. 1996;157:195–204. doi: 10.1159/000147881. [DOI] [PubMed] [Google Scholar]
- 773.Oktem G., Altay B., Turna B. Determination of nitric oxide synthase activity and apoptosis of germ cells in different obstruction models. Acta Histochem. 2009;111:119–126. doi: 10.1016/j.acthis.2007.01.005. [DOI] [PubMed] [Google Scholar]
- 774.Sato Y., Nozawa S., Iwamoto T. Study of spermatogenesis and thickening of lamina propria in the human seminiferous tubules. Fertil Steril. 2008;90:1310–1312. doi: 10.1016/j.fertnstert.2007.10.019. [DOI] [PubMed] [Google Scholar]
- 775.Suskind A., Hayner-Buchan A., Feustel P.J., Kogan B.A. Fibrosis correlates with detailed histological analysis of human undescended testes. BJU Int. 2008;101:1441–1445. doi: 10.1111/j.1464-410X.2007.07406.x. [DOI] [PubMed] [Google Scholar]
- 776.Biswas B., Narmadha G., Choudhary M., French F.S., Hall S.H., Yenugu S. Identification of Toll-like receptors in the rat (Rattus norvegicus): messenger RNA expression in the male reproductive tract under conditions of androgen variation. Am J Reprod Immunol. 2009;62:243–252. doi: 10.1111/j.1600-0897.2009.00732.x. [DOI] [PubMed] [Google Scholar]
- 777.Malm J., Nordahl E.A., Bjartell A. Lipopolysaccharide-binding protein is produced in the epididymis and associated with spermatozoa and prostasomes. J Reprod Immunol. 2005;66:33–43. doi: 10.1016/j.jri.2005.01.005. [DOI] [PubMed] [Google Scholar]
- 778.Nishimura M., Naito S. Tissue-specific mRNA expression profiles of human toll-like receptors and related genes. Biol Pharm Bull. 2005;28:886–892. doi: 10.1248/bpb.28.886. [DOI] [PubMed] [Google Scholar]
- 779.Mackern-Oberti J.P., Maccioni M., Cuffini C., Gatti G., Rivero V.E. Susceptibility of prostate epithelial cells to Chlamydia muridarum infection and their role in innate immunity by recruitment of intracellular Toll-like receptors 4 and 2 and MyD88 to the inclusion. Infect Immun. 2006;74:6973–6981. doi: 10.1128/IAI.00593-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 780.Quintar A.A., Roth F.D., De Paul A.L., Aoki A., Maldonado C.A. Toll-like receptor 4 in rat prostate: modulation by testosterone and acute bacterial infection in epithelial and stromal cells. Biol Reprod. 2006;75:664–672. doi: 10.1095/biolreprod.106.053967. [DOI] [PubMed] [Google Scholar]
- 781.Gatti G., Rivero V., Motrich R.D., Maccioni M. Prostate epithelial cells can act as early sensors of infection by up-regulating TLR4 expression and proinflammatory mediators upon LPS stimulation. J Leuk Biol. 2006;79:989–998. doi: 10.1189/jlb.1005597. [DOI] [PubMed] [Google Scholar]
- 782.Ilvesaro J.M., Merrell M.A., Swain T.M. Toll like receptor-9 agonists stimulate prostate cancer invasion in vitro. Prostate. 2007;67:774–781. doi: 10.1002/pros.20562. [DOI] [PubMed] [Google Scholar]
- 783.Takeyama K., Mitsuzawa H., Shimizu T. Prostate cell lines secrete IL-8 in response to Mycoplasma hominis through Toll-like receptor 2-mediated mechanism. Prostate. 2006;66:386–391. doi: 10.1002/pros.20358. [DOI] [PubMed] [Google Scholar]
- 784.Pudney J., Anderson D.J. Expression of Toll-like receptors in genital tract tissues from normal and HIV-infected men. Am J Reprod Immunol. 2010;65:28–43. doi: 10.1111/j.1600-0897.2010.00877.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 785.Chen Y.C., Giovannucci E., Lazarus R., Kraft P., Ketkar S., Hunter D.J. Sequence variants of Toll-like receptor 4 and susceptibility to prostate cancer. Cancer Res. 2005;65:11771–11778. doi: 10.1158/0008-5472.CAN-05-2078. [DOI] [PubMed] [Google Scholar]
- 786.Sun J., Wiklund F., Zheng S.L. Sequence variants in Toll-like receptor gene cluster (TLR6-TLR1-TLR10) and prostate cancer risk. J Natl Cancer Inst. 2005;97:525–532. doi: 10.1093/jnci/dji070. [DOI] [PubMed] [Google Scholar]
- 787.Xu B., Abdel-Fattah R., Yang L., Crenshaw S.A., Black M.B., Hinton B.T. Testicular lumicrine factors regulate ERK, STAT, and NFKB pathways in the initial segment of the rat epididymis to prevent apoptosis. Biol Reprod. 2011;84:1282–1291. doi: 10.1095/biolreprod.110.090324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 788.Turner T.T., Johnston D.S., Jelinsky S.A., Tomsig J.L., Finger J.N. Segment boundaries of the adult rat epididymis limit interstitial signaling by potential paracrine factors and segments lose differential gene expression after efferent duct ligation. Asian J Androl. 2007;9:565–573. doi: 10.1111/j.1745-7262.2007.00302.x. [DOI] [PubMed] [Google Scholar]
- 789.Winnall W.R., Wu H., Sarraj M.A. Expression patterns of activin, inhibin and follistatin variants in the adult male mouse reproductive tract suggest important roles in the epididymis and vas deferens. Reprod Fertil Dev. 2013;25:570–580. doi: 10.1071/RD11287. [DOI] [PubMed] [Google Scholar]
- 790.Bahathiq A.O., Stewart R.L., Baxter L., Wells M., Moore H.D., Ledger W.L. Tissue immunoexpression and messenger ribonucleic acid localization of inhibin/activin subunit in human epididymis. Fertil Steril. 2005;83:78–85. doi: 10.1016/j.fertnstert.2004.05.104. [DOI] [PubMed] [Google Scholar]
- 791.Zhang T., Guo C.X., Hu Z.Y., Liu Y.X. Localization of plasminogen activator and inhibitor, LH and androgen receptors and inhibin subunits in monkey epididymis. Mol Hum Reprod. 1997;3:945–952. doi: 10.1093/molehr/3.11.945. [DOI] [PubMed] [Google Scholar]
- 792.Seachrist D.D., Johnson E., Magee C. Overexpression of follistatin in the mouse epididymis disrupts fluid resorption and sperm transit in testicular excurrent ducts. Biol Reprod. 2012;87:41. doi: 10.1095/biolreprod.111.097527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 793.Antonipillai I., Wahe M., Yamamoto J., Horton R. Activin and inhibin have opposite effects on steroid 5α-reductase activity in genital skin fibroblasts. Mol Cell Endocrinol. 1995;107:99–104. doi: 10.1016/0303-7207(94)03430-2. [DOI] [PubMed] [Google Scholar]
- 794.Jelinsky S.A., Turner T.T., Bang H.J. The rat epididymal transcriptome: comparison of segmental gene expression in the rat and mouse epididymides. Biol Reprod. 2007;76:561–570. doi: 10.1095/biolreprod.106.057323. [DOI] [PubMed] [Google Scholar]
- 795.Bomgardner D., Wehrenberg U., Rune G.M. TGF-β could be involved in paracrine actions in the epididymis of the marmoset monkey (Callithrix jacchus) J Androl. 1999;20:375–383. [PubMed] [Google Scholar]
- 796.Henderson N.A., Cooke G.M., Robaire B. Region-specific expression of androgen and growth factor pathway genes in the rat epididymis and the effects of dual 5α-reductase inhibition. J Endocrinol. 2006;190:779–791. doi: 10.1677/joe.1.06862. [DOI] [PubMed] [Google Scholar]
- 797.Desai K.V., Flanders K.C., Kondaiah P. Expression of transforming growth factor-β isoforms in the rat male accessory sex organs and epididymis. Cell Tissue Res. 1998;294:271–277. doi: 10.1007/s004410051177. [DOI] [PubMed] [Google Scholar]
- 798.Desai K.V., Kondaiah P. Androgen ablation results in differential regulation of transforming growth factor-β isoforms in rat male accessory sex organs and epididymis. J Mol Endocrinol. 2000;24:253–260. doi: 10.1677/jme.0.0240253. [DOI] [PubMed] [Google Scholar]
- 799.Pierucci-Alves F., Yi S., Schultz B.D. Transforming growth factor beta 1 induces tight junction disruptions and loss of transepithelial resistance across porcine vas deferens epithelial cells. Biol Reprod. 2012;86:36. doi: 10.1095/biolreprod.111.092262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 800.Burnett A.L., Ricker D.D., Chamness S.L. Localization of nitric oxide synthase in the reproductive organs of the male rat. Biol Reprod. 1995;52:1–7. doi: 10.1095/biolreprod52.1.1. [DOI] [PubMed] [Google Scholar]
- 801.McKanna J.A., Zhang M.Z., Wang J.L., Cheng H., Harris R.C. Constitutive expression of cyclooxygenase-2 in rat vas deferens. Am J Physiol. 1998;275:R227–R233. doi: 10.1152/ajpregu.1998.275.1.R227. [DOI] [PubMed] [Google Scholar]
- 802.Fujita Y., Mihara T., Okazaki T. Toll-like receptors (TLR) 2 and 4 on human sperm recognize bacterial endotoxins and mediate apoptosis. Hum Reprod. 2011;26:2799–2806. doi: 10.1093/humrep/der234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 803.Sikka S.C., Champion H.C., Bivalacqua T.J. Role of genitourinary inflammation in infertility: synergistic effect of lipopolysaccharide and interferon-γ on human spermatozoa. Int J Androl. 2001;24:136–141. doi: 10.1046/j.1365-2605.2001.00279.x. [DOI] [PubMed] [Google Scholar]
- 804.Martínez P., Proverbio F., Camejo M.I. Sperm lipid peroxidation and pro-inflammatory cytokines. Asian J Androl. 2007;9:102–107. doi: 10.1111/j.1745-7262.2007.00238.x. [DOI] [PubMed] [Google Scholar]
- 805.Lampiao F., du Plessis S.S. TNF-α and IL-6 affect human sperm function by elevating nitric oxide production. Reprod Biomed Online. 2008;17:628–631. doi: 10.1016/s1472-6483(10)60309-4. [DOI] [PubMed] [Google Scholar]
- 806.Perdichizzi A., Nicoletti F., La Vignera S. Effects of tumour necrosis factor-α on human sperm motility and apoptosis. J Clin Immunol. 2007;27:152–162. doi: 10.1007/s10875-007-9071-5. [DOI] [PubMed] [Google Scholar]
- 807.Almog T., Naor Z. Mitogen activated protein kinases (MAPKs) as regulators of spermatogenesis and spermatozoa functions. Mol Cell Endocrinol. 2008;282:39–44. doi: 10.1016/j.mce.2007.11.011. [DOI] [PubMed] [Google Scholar]
- 808.Cao D., Li Y., Yang R. Lipopolysaccharide-induced epididymitis disrupts epididymal beta-defensin expression and inhibits sperm motility in rats. Biol Reprod. 2010;83:1064–1070. doi: 10.1095/biolreprod.109.082180. [DOI] [PubMed] [Google Scholar]
- 809.Turner T.T., Mammen T., Kavoussi P., Lysiak J.J., Costabile R.A. Cytokine responses to E. coli-induced epididymitis in the rat: blockade by vasectomy. Urology. 2011;77:1507.e9–1507.e14. doi: 10.1016/j.urology.2011.02.037. [DOI] [PubMed] [Google Scholar]
- 810.Ozturk U., Kefeli M., Asci R., Akpolat I., Buyukalpelli R., Sarikaya S. The effects of experimental left varicocele on the epididymis. Syst Biol Reprod Med. 2008;54:177–184. doi: 10.1080/19396360802415752. [DOI] [PubMed] [Google Scholar]
- 811.Balaji T., Ramanathan M., Menon V.P. Localization of cyclooxygenase-2 in mice vas deferens and its effects on fertility upon suppression using nimesulide – a preferential cyclooxygenase-2 inhibitor. Toxicology. 2007;234:135–144. doi: 10.1016/j.tox.2007.02.011. [DOI] [PubMed] [Google Scholar]
- 812.Mühlbauer M., Cheely A.W., Yenugu S., Jobin C. Regulation and functional impact of lipopolysaccharide induced Nod2 gene expression in the murine epididymal epithelial cell line PC1. Immunology. 2008;124:256–264. doi: 10.1111/j.1365-2567.2007.02763.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 813.Wiszniewska B., Kurzawa R., Ciechanowicz A., Machalinski B. Inducible nitric oxide synthase in the epithelial epididymal cells of the rat. Reprod Fertil Dev. 1997;9:789–794. doi: 10.1071/r97063. [DOI] [PubMed] [Google Scholar]
- 814.Mackern-Oberti J.P., Maccioni M., Breser M.L., Eley A., Miethke T., Rivero V.E. Innate immunity in the male genital tract: Chlamydia trachomatis induces keratinocyte-derived chemokine production in prostate, seminal vesicle and epididymis/vas deferens primary cultures. J Med Microbiol. 2011;60:307–316. doi: 10.1099/jmm.0.024877-0. [DOI] [PubMed] [Google Scholar]
- 815.Biswas B., Yenugu S. Antimicrobial responses in the male reproductive tract of lipopolysaccharide challenged rats. Am J Reprod Immunol. 2011;65:557–568. doi: 10.1111/j.1600-0897.2010.00937.x. [DOI] [PubMed] [Google Scholar]
- 816.Politch J.A., Tucker L., Bowman F.P., Anderson D.J. Concentrations and significance of cytokines and other immunologic factors in semen of healthy fertile men. Hum Reprod. 2007;22:2928–2935. doi: 10.1093/humrep/dem281. [DOI] [PubMed] [Google Scholar]
- 817.Huleihel M., Lunenfeld E., Horowitz S. Expression of IL-12, IL-10, PGE2, sIL-2R and sIL-6R in seminal plasma of fertile and infertile men. Andrologia. 1999;31:283–288. doi: 10.1046/j.1439-0272.1999.00289.x. [DOI] [PubMed] [Google Scholar]
- 818.Ochsenkühn R., O’Connor A.E., Hirst J.J., Baker H.W.G., de Kretser D.M., Hedger M.P. The relationship between immunosuppressive activity and immunoregulatory cytokines in seminal plasma: influence of sperm autoimmunity and seminal leukocytes. J Reprod Immunol. 2006;71:57–74. doi: 10.1016/j.jri.2006.01.002. [DOI] [PubMed] [Google Scholar]
- 819.Eggert-Kruse W., Kiefer I., Beck C., Demirakca T., Strowitzki T. Role for tumor necrosis factor alpha (TNF-α) and interleukin 1-beta (IL-1β) determination in seminal plasma during infertility investigation. Fertil Steril. 2007;87:810–823. doi: 10.1016/j.fertnstert.2006.08.103. [DOI] [PubMed] [Google Scholar]
- 820.Moretti E., Cosci I., Spreafico A., Serchi T., Cuppone A.M., Collodel G. Semen characteristics and inflammatory mediators in infertile men with different clinical diagnoses. Int J Androl. 2009;32:637–646. doi: 10.1111/j.1365-2605.2008.00911.x. [DOI] [PubMed] [Google Scholar]
- 821.Matalliotakis I., Arici A., Goumenou A. Distinct expression pattern of cytokines in semen of men with genital infection and oligo-terato-asthenozoospermia. Am J Reprod Immunol. 2002;48:170–175. doi: 10.1034/j.1600-0897.2002.01086.x. [DOI] [PubMed] [Google Scholar]
- 822.Pilatz A., Hudemann C., Wagenlehner F. Seminal cytokines. Is quantification useful in urogenital disorders? Urologe. 2013;52:359. doi: 10.1007/s00120-013-3141-5. [DOI] [PubMed] [Google Scholar]
- 823.Battaglia C., Giulini S., Regnani G. Seminal plasma nitrite/nitrate and intratesticular Doppler flow in fertile and infertile subjects. Hum Reprod. 2000;15:2554–2558. doi: 10.1093/humrep/15.12.2554. [DOI] [PubMed] [Google Scholar]
- 824.Sakamoto Y., Ishikawa T., Kondo Y., Yamaguchi K., Fujisawa M. The assessment of oxidative stress in infertile patients with varicocele. BJU Int. 2008;101:1547–1552. doi: 10.1111/j.1464-410X.2008.07517.x. [DOI] [PubMed] [Google Scholar]
- 825.Barratt C.L., Harrison P.E., Robinson A., Kessopoulou E., Cooke I.D. Seminal white blood cells in men with urethral tract infection. A monoclonal antibody study. Br J Urol. 1991;68:531–536. doi: 10.1111/j.1464-410x.1991.tb15399.x. [DOI] [PubMed] [Google Scholar]
- 826.Collin M., Linge H.M., Bjartell A., Giwercman A., Malm J., Egesten A. Constitutive expression of the antibacterial CXC chemokine GCP-2/CXCL6 by epithelial cells of the male reproductive tract. J Reprod Immunol. 2008;79:37–43. doi: 10.1016/j.jri.2008.08.003. [DOI] [PubMed] [Google Scholar]
- 827.Linge H.M., Collin M., Giwercman A., Malm J., Bjartell A., Egesten A. The antibacterial chemokine MIG/CXCL9 is constitutively expressed in epithelial cells of the male urogenital tract and is present in seminal plasma. J Interferon Cytokine Res. 2008;28:191–196. doi: 10.1089/jir.2007.0100. [DOI] [PubMed] [Google Scholar]
- 828.Auroux M. Nonspermatozoal cells in human sperm: a study of 1243 subfertile and 253 fertile men. Arch Androl. 1984;12:197–201. doi: 10.3109/01485018409161176. [DOI] [PubMed] [Google Scholar]
- 829.Aziz N., Agarwal A., Lewis-Jones I., Sharma R.K., Thomas A.J., Jr. Novel associations between specific sperm morphological defects and leukocytospermia. Fertil Steril. 2004;82:621–627. doi: 10.1016/j.fertnstert.2004.02.112. [DOI] [PubMed] [Google Scholar]
- 830.Wolff H., Politch J.A., Martinez A., Haimovici F., Hill J.A., Anderson D.J. Leukocytospermia is associated with poor semen quality. Fertil Steril. 1990;53:528–536. [PubMed] [Google Scholar]
- 831.Hill J.A., Haimovici F., Politch J.A., Anderson D.J. Effects of soluble products of activated lymphocytes and macrophages (lymphokines and monokines) on human sperm motion parameters. Fertil Steril. 1987;47:460–465. doi: 10.1016/s0015-0282(16)59056-6. [DOI] [PubMed] [Google Scholar]
- 832.Tomlinson M.J., East S.J., Barratt C.L., Bolton A.E., Cooke I.D. Preliminary communication: possible role of reactive nitrogen intermediates in leucocyte-mediated sperm dysfunction. Am J Reprod Immunol. 1992;27:89–92. doi: 10.1111/j.1600-0897.1992.tb00730.x. [DOI] [PubMed] [Google Scholar]
- 833.Barratt C.L., Harrison P.E., Robinson A., Cooke I.D. Antisperm antibodies and lymphocyte subsets in semen – not a simple relationship. Int J Androl. 1990;13:50–58. doi: 10.1111/j.1365-2605.1990.tb00959.x. [DOI] [PubMed] [Google Scholar]
- 834.Hendry W.F. Detection and treatment of antispermatozoal antibodies in men. Reprod Fertil Dev. 1989;1:205–220. doi: 10.1071/rd9890205. discussion 220–2. [DOI] [PubMed] [Google Scholar]
- 835.Kremer J., Jager S. The significance of antisperm antibodies for sperm-cervical mucus interaction. Hum Reprod. 1992;7:781–784. doi: 10.1093/oxfordjournals.humrep.a137737. [DOI] [PubMed] [Google Scholar]
- 836.Lin W.W., Kim E.D., Quesada E.T., Lipshultz L.I., Coburn M. Unilateral testicular injury from external trauma: evaluation of semen quality and endocrine parameters. J Urol. 1998;159:841–843. doi: 10.1016/s0022-5347(01)63748-0. [DOI] [PubMed] [Google Scholar]
- 837.McLachlan R.I. Basis, diagnosis and treatment of immunological infertility in men. J Reprod Immunol. 2002;57:35–45. doi: 10.1016/s0165-0378(02)00014-1. [DOI] [PubMed] [Google Scholar]
- 838.Patrizio P., Silber S.J., Ord T., Moretti-Rojas I., Asch R.H. Relationship of epididymal sperm antibodies to their in vitro fertilization capacity in men with congenital absence of the vas deferens. Fertil Steril. 1992;58:1006–1010. doi: 10.1016/s0015-0282(16)55451-x. [DOI] [PubMed] [Google Scholar]
- 839.Eggert-Kruse W., Probst S., Rohr G., Tilgen W., Runnebaum B. Induction of immunoresponse by subclinical male genital tract infection? Fertil Steril. 1996;65:1202–1209. [PubMed] [Google Scholar]
- 840.Ness R.B., Markovic N., Carlson C.L., Coughlin M.T. Do men become infertile after having sexually transmitted urethritis? An epidemiologic examination. Fertil Steril. 1997;68:205–213. doi: 10.1016/s0015-0282(97)81502-6. [DOI] [PubMed] [Google Scholar]
- 841.Paschke R., Bertelsbeck D.S., Tsalimalma K., Nieschlag E. Association of sperm antibodies with other autoantibodies in infertile men. Am J Reprod Immunol. 1994;32:88–94. doi: 10.1111/j.1600-0897.1994.tb01098.x. [DOI] [PubMed] [Google Scholar]
- 842.Baker H.W., Clarke G.N., McGowan M.P., Koh S.H., Cauchi M.N. Increased frequency of autoantibodies in men with sperm antibodies. Fertil Steril. 1985;43:438–441. doi: 10.1016/s0015-0282(16)48445-1. [DOI] [PubMed] [Google Scholar]
- 843.Elder M., Maclaren N., Riley W. Gonadal autoantibodies in patients with hypogonadism and/or Addison’s disease. J Clin Endocrinol Metab. 1981;52:1137–1142. doi: 10.1210/jcem-52-6-1137. [DOI] [PubMed] [Google Scholar]
- 844.Tsatsoulis A., Shalet S.M. Antisperm antibodies in the polyglandular autoimmune (PGA) syndrome type I: response to cyclical steroid therapy. Clin Endocrinol. 1991;35:299–303. doi: 10.1111/j.1365-2265.1991.tb03540.x. [DOI] [PubMed] [Google Scholar]
- 845.Itoh M., Hiramine C., Tokunaga Y., Mukasa A., Hojo K. A new murine model of autoimmune orchitis induced by immunization with viable syngeneic testicular germ cells alone. II. Immunohistochemical findings of fully-developed inflammatory lesion. Autoimmunity. 1991;10:89–97. doi: 10.3109/08916939109004812. [DOI] [PubMed] [Google Scholar]
- 846.Alexander N.J., Fulgham D.L. Antibodies to spermatozoa in male monkeys: mode of action. Fertil Steril. 1978;30:334–342. [PubMed] [Google Scholar]
- 847.Flickinger C.J., Bush L.A., Williams M.V., Naaby-Hansen S., Howards S.S., Herr J.C. Post-obstruction rat sperm autoantigens identified by two-dimensional gel electrophoresis and western blotting. J Reprod Immunol. 1999;43:35–53. doi: 10.1016/s0165-0378(98)00090-4. [DOI] [PubMed] [Google Scholar]
- 848.Barratt C.L., Dunphy B.C., McLeod I., Cooke I.D. The poor prognostic value of low to moderate levels of sperm surface-bound antibodies. Hum Reprod. 1992;7:95–98. doi: 10.1093/oxfordjournals.humrep.a137567. [DOI] [PubMed] [Google Scholar]
- 849.Eggert-Kruse W., Hofsäß A., Haury E., Tilgen W., Gerhard I., Runnebaum B. Relationship between local anti-sperm antibodies and sperm-mucus interaction in vitro and in vivo. Hum Reprod. 1991;6:267–276. doi: 10.1093/oxfordjournals.humrep.a137320. [DOI] [PubMed] [Google Scholar]
- 850.Clarke G.N. Lack of correlation between the immunobead test and the enzyme-linked immunosorbent assay for sperm antibody detection. Am J Reprod Immunol Microbiol. 1988;18:44–46. doi: 10.1111/j.1600-0897.1988.tb00233.x. [DOI] [PubMed] [Google Scholar]
- 851.Mettler L., Czuppon A.B., Alexander N. Antibodies to spermatozoa and seminal plasma antigens detected by various enzyme-linked immunosorbent (ELISA) assays. J Reprod Immunol. 1985;8:301–312. doi: 10.1016/0165-0378(85)90004-x. [DOI] [PubMed] [Google Scholar]
- 852.Knee R.A., Hickey D.K., Beagley K.W., Jones R.C. Transport of IgG across the blood-luminal barrier of the male reproductive tract of the rat and the effect of estradiol administration on reabsorption of fluid and IgG by the epididymal ducts. Biol Reprod. 2005;73:688–694. doi: 10.1095/biolreprod.105.041079. [DOI] [PubMed] [Google Scholar]
- 853.Ekwere P.D. Immunological infertility among Nigerian men: incidence of circulating antisperm auto-antibodies and some clinical observations: a preliminary report. Br J Urol. 1995;76:366–370. doi: 10.1111/j.1464-410x.1995.tb07716.x. [DOI] [PubMed] [Google Scholar]
- 854.Turek P.J., Lipshultz L.I. Immunologic infertility. Urol Clin North Am. 1994;21:447–468. [PubMed] [Google Scholar]
- 855.Witkin S.S. Failure of sperm-induced immunosuppression: association with antisperm antibodies in women. Am J Obstet Gynecol. 1989;160:1166–1168. doi: 10.1016/0002-9378(89)90181-6. [DOI] [PubMed] [Google Scholar]
- 856.Alexander N.J., Anderson D.J. Immunology of semen. Fertil Steril. 1987;47:192–205. doi: 10.1016/s0015-0282(16)49990-5. [DOI] [PubMed] [Google Scholar]
- 857.Robertson S.A., Bromfield J.J., Tremellen K.P. Seminal ‘priming’ for protection from pre-eclampsia-a unifying hypothesis. J Reprod Immunol. 2003;59:253–265. doi: 10.1016/s0165-0378(03)00052-4. [DOI] [PubMed] [Google Scholar]
- 858.Davis N.F., McGuire B.B., Mahon J.A., Smyth A.E., O’Malley K.J., Fitzpatrick J.M. The increasing incidence of mumps orchitis: a comprehensive review. BJU Int. 2010 doi: 10.1111/j.1464-410X.2009.09148.x. [DOI] [PubMed] [Google Scholar]
- 859.Roy S., Hooda S., Parwani A.V. Idiopathic granulomatous orchitis. Pathol Res Pract. 2011;207:275–278. doi: 10.1016/j.prp.2011.02.005. [DOI] [PubMed] [Google Scholar]
- 860.Krieger J.N. Epididymitis, orchitis, and related conditions. Sex Transm Dis. 1984;11:173–181. doi: 10.1097/00007435-198407000-00012. [DOI] [PubMed] [Google Scholar]
- 861.Jenkin G.A., Choo M., Hosking P., Johnson P.D. Candidal epididymo-orchitis: case report and review. Clin Infect Dis. 1998;26:942–945. doi: 10.1086/513937. [DOI] [PubMed] [Google Scholar]
- 862.Wall J.R., Wright D.J. Antibodies against testicular germinal cells in lepromatous leprosy. Clin Exp Immunol. 1974;17:51–59. [PMC free article] [PubMed] [Google Scholar]
- 863.Xu J., Qi L., Chi X. Orchitis: a complication of severe acute respiratory syndrome (SARS) Biol Reprod. 2006;74:410–416. doi: 10.1095/biolreprod.105.044776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 864.Matsuzaki G., Sonoda K.H., Mukasa A. The characterization of testicular cell (TC)-specific T-cell clones induced by intratesticular Listeria monocytogenes infection: TC-specific T cells with atypical cytokine profile transfer orchitis. Immunology. 1997;91:520–528. doi: 10.1046/j.1365-2567.1997.00306.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 865.Silva C.A., Cocuzza M., Borba E.F., Bonfa E. Cutting-edge issues in autoimmune orchitis. Clin Rev Allergy Immunol. 2012;42:256–263. doi: 10.1007/s12016-011-8281-z. [DOI] [PubMed] [Google Scholar]
- 866.Pelletier R.M. Cyclic formation and decay of the blood-testis barrier in the mink (Mustela vison), a seasonal breeder. Am J Anat. 1986;175:91–117. doi: 10.1002/aja.1001750109. [DOI] [PubMed] [Google Scholar]
- 867.Furbeth C., Hubner G., Thoenes G.H. Spontaneous immune complex orchitis in brown Norway rats. Virchows Arch B Cell Pathol. 1989;57:37–45. doi: 10.1007/BF02899063. [DOI] [PubMed] [Google Scholar]
- 868.Fritz T.E., Lombard S.A., Tyler S.A., Norris W.P. Pathology and familial incidence of orchitis and its relation to thyroiditis in a closed beagle colony. Exp Mol Pathol. 1976;24:142–158. doi: 10.1016/0014-4800(76)90002-2. [DOI] [PubMed] [Google Scholar]
- 869.Lehmann D., Emmons L.R. Immunological phenomena observed in the testis and their possible role in infertility. Am J Reprod Immunol. 1989;19:43–52. doi: 10.1111/j.1600-0897.1989.tb00547.x. [DOI] [PubMed] [Google Scholar]
- 870.Rabin B.S., Nankin H.R., Troen P. Immunologic studies of patients with idiopathic oligospermia. In: Troen P., Nankin H.R., editors. The testis in normal and infertile men. Raven Press; New York: 1977. pp. 435–444. [Google Scholar]
- 871.Salomon F., Hedinger C.E. Histopathology of immunologic lesions of the human testis. In: Bigazzi P.E., editor. Immunology of the male reproductive system. Marcel Dekker; New York: 1987. pp. 203–231. [Google Scholar]
- 872.Maclaren N., Chen Q.Y., Kukreja A., Marker J., Zhang C.H., Sun Z.S. Autoimmune hypogonadism as part of an autoimmune polyglandular syndrome. J Soc Gynecol Invest. 2001;8:S52–S54. doi: 10.1016/s1071-5576(00)00109-x. [DOI] [PubMed] [Google Scholar]
- 873.Pannek J., Haupt G. Orchitis due to vasculitis in autoimmune diseases. Scand J Rheumatol. 1997;26:151–154. doi: 10.3109/03009749709065674. [DOI] [PubMed] [Google Scholar]
- 874.Tung K.S., Smith S., Teuscher C., Cook C., Anderson R.E. Murine autoimmune oophoritis, epididymoorchitis, and gastritis induced by day 3 thymectomy. Immunopathology. Am J Pathol. 1987;126:293–302. [PMC free article] [PubMed] [Google Scholar]
- 875.Tung K.S., Smith S., Matzner P. Murine autoimmune oophoritis, epididymoorchitis, and gastritis induced by day 3 thymectomy. Autoantibodies. Am J Pathol. 1987;126:303–314. [PMC free article] [PubMed] [Google Scholar]
- 876.Itoh M., Hiramine C., Hojo K. A new murine model of autoimmune orchitis induced by immunization with viable syngeneic testicular germ cells alone. I. Immunological and histological studies. Clin Exp Immunol. 1991;83:137–142. doi: 10.1111/j.1365-2249.1991.tb05603.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 877.Naito M., Hirai S., Terayama H. Postinflammation stage of autoimmune orchitis induced by immunization with syngeneic testicular germ cells alone in mice. Med Mol Morphol. 2012;45:35–44. doi: 10.1007/s00795-011-0539-2. [DOI] [PubMed] [Google Scholar]
- 878.Kojima A., Spencer C.A. Genetic susceptibility to testicular autoimmunity: comparison between postthymectomy and postvasectomy models in mice. Biol Reprod. 1983;29:195–205. doi: 10.1095/biolreprod29.1.195. [DOI] [PubMed] [Google Scholar]
- 879.Tumboh-Oeri A.G., Roberts T.K. Immunological and morphological consequences of vasectomy in the rabbit. Experientia. 1979;35:675–676. doi: 10.1007/BF01960393. [DOI] [PubMed] [Google Scholar]
- 880.Tung K.S. Allergic orchitis lesions are adoptively transferred from vasoligated guinea pigs to syngeneic recipients. Science. 1978;201:833–835. doi: 10.1126/science.684410. [DOI] [PubMed] [Google Scholar]
- 881.Tung K.S., Alexander N.J. Monocytic orchitis and aspermatogenesis in normal and vasectomized rhesus macaques (Macaca mulatta) Am J Pathol. 1980;101:17–30. [PMC free article] [PubMed] [Google Scholar]
- 882.Neufeld M., Maclaren N.K., Blizzard R.M. Two types of autoimmune Addison’s disease associated with different polyglandular autoimmune (PGA) syndromes. Medicine. 1981;60:355–362. doi: 10.1097/00005792-198109000-00003. [DOI] [PubMed] [Google Scholar]
- 883.Uibo R., Aavik E., Peterson P. Autoantibodies to cytochrome P450 enzymes P450scc, P450c17, and P450c21 in autoimmune polyglandular disease types I and II and in isolated Addison’s disease. J Clin Endocrinol Metab. 1994;78:323–328. doi: 10.1210/jcem.78.2.8106620. [DOI] [PubMed] [Google Scholar]
- 884.Arif S., Vallian S., Farzaneh F. Identification of 3 β-hydroxysteroid dehydrogenase as a novel target of steroid cell autoantibodies: association of autoantibodies with endocrine autoimmune disease. J Clin Endocrinol Metab. 1996;81:4439–4445. doi: 10.1210/jcem.81.12.8954056. [DOI] [PubMed] [Google Scholar]
- 885.Jacobo P., Guazzone V.A., Jarazo-Dietrich S., Theas M.S., Lustig L. Differential changes in CD4+ and CD8+ effector and regulatory T lymphocyte subsets in the testis of rats undergoing autoimmune orchitis. J Reprod Immunol. 2009;81:44–54. doi: 10.1016/j.jri.2009.04.005. [DOI] [PubMed] [Google Scholar]
- 886.Jacobo P., Perez C.V., Theas M.S., Guazzone V.A., Lustig L. CD4+ and CD8+ T cells producing Th1 and Th17 cytokines are involved in the pathogenesis of autoimmune orchitis. Reproduction. 2011;141:249–258. doi: 10.1530/REP-10-0362. [DOI] [PubMed] [Google Scholar]
- 887.Rival C., Theas M.S., Suescun M.O. Functional and phenotypic characteristics of testicular macrophages in experimental autoimmune orchitis. J Pathol. 2008;215:108–117. doi: 10.1002/path.2328. [DOI] [PubMed] [Google Scholar]
- 888.Itoh M., De-Rooij D., Takeuchi Y. Mode of inflammatory cell infiltration in testes of mice injected with syngeneic testicular germ cells without adjuvant. J Anat. 1995;187:671–679. [PMC free article] [PubMed] [Google Scholar]
- 889.Naito M., Itoh M. Patterns of infiltration of lymphocytes into the testis under normal and pathological conditions in mice. Am J Reprod Immunol. 2008;59:55–61. doi: 10.1111/j.1600-0897.2007.00556.x. [DOI] [PubMed] [Google Scholar]
- 890.Yule T.D., Tung K.S. Experimental autoimmune orchitis induced by testis and sperm antigen-specific T cell clones: an important pathogenic cytokine is tumor necrosis factor. Endocrinology. 1993;133:1098–1107. doi: 10.1210/endo.133.3.8103448. [DOI] [PubMed] [Google Scholar]
- 891.Theas M.S., Rival C., Jarazo-Dietrich S., Jacobo P., Guazzone V.A., Lustig L. Tumour necrosis factor-α released by testicular macrophages induces apoptosis of germ cells in autoimmune orchitis. Hum Reprod. 2008;23:1865–1872. doi: 10.1093/humrep/den240. [DOI] [PubMed] [Google Scholar]
- 892.Jacobo P.V., Fass M., Perez C.V., Jarazo-Dietrich S., Lustig L., Theas M.S. Involvement of soluble Fas Ligand in germ cell apoptosis in testis of rats undergoing autoimmune orchitis. Cytokine. 2012;60:385–392. doi: 10.1016/j.cyto.2012.07.020. [DOI] [PubMed] [Google Scholar]
- 893.Jarazo-Dietrich S., Jacobo P., Pérez C.V., Guazzone V.A., Lustig L., Theas M.S. Up regulation of nitric oxide synthase-nitric oxide system in the testis of rats undergoing autoimmune orchitis. Immunobiol. 2012;217:778–787. doi: 10.1016/j.imbio.2012.04.007. [DOI] [PubMed] [Google Scholar]
- 894.Kuerban M., Naito M., Hirai S. Involvement of Fas/Fas-L and Bax/Bcl-2 systems in germ cell death following immunization with syngeneic testicular germ cells in mice. J Androl. 2012;33:824–831. doi: 10.2164/jandrol.111.015529. [DOI] [PubMed] [Google Scholar]
- 895.Rival C., Theas M.S., Guazzone V.A., Lustig L. Interleukin-6 and IL-6 receptor cell expression in testis of rats with autoimmune orchitis. J Reprod Immunol. 2006;70:43–58. doi: 10.1016/j.jri.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 896.Pérez C.V., Sobarzo C.M., Jacobo P.V. Loss of occludin expression and impairment of blood-testis barrier permeability in rats with autoimmune orchitis: effect of interleukin 6 on Sertoli cell tight junctions. Biol Reprod. 2012;87:122. doi: 10.1095/biolreprod.112.101709. [DOI] [PubMed] [Google Scholar]
- 897.Li L., Itoh M., Ablake M., Macri B., Bendtzen K., Nicoletti F. Prevention of murine experimental autoimmune orchitis by recombinant human interleukin-6. Clin Immunol. 2002;102:135–137. doi: 10.1006/clim.2001.5155. [DOI] [PubMed] [Google Scholar]
- 898.Watanabe M., Kashiwakura Y., Kusumi N. Adeno-associated virus-mediated human IL-10 gene transfer suppresses the development of experimental autoimmune orchitis. Gene Ther. 2005;12:1126–1132. doi: 10.1038/sj.gt.3302463. [DOI] [PubMed] [Google Scholar]
- 899.Fijak M., Schneider E., Klug J. Testosterone replacement effectively inhibits the development of experimental autoimmune orchitis in rats: evidence for a direct role of testosterone on regulatory T cell expansion. J Immunol. 2011;186:5162–5172. doi: 10.4049/jimmunol.1001958. [DOI] [PubMed] [Google Scholar]
- 900.Boughton B., Spector W.G. “Auto-immune” testicular lesions induced by injury to the contralateral testis and intradermal injection of adjuvant. J Pathol Bacteriol. 1963;86:69–74. doi: 10.1002/path.1700860108. [DOI] [PubMed] [Google Scholar]
- 901.Suominen J.J. Sympathetic auto-immune orchitis. Andrologia. 1995;27:213–216. [PubMed] [Google Scholar]
- 902.Demir A., Türker P., Önol F.F., Sirvanci S., Findik A., Tarcan T. Effect of experimentally induced Escherichia coli epididymo-orchitis and ciprofloxacin treatment on rat spermatogenesis. Int J Urol. 2007;14:268–272. doi: 10.1111/j.1442-2042.2007.01682.x. [DOI] [PubMed] [Google Scholar]
- 903.Mukasa A., Yoshida H., Kobayashi N., Matsuzaki G., Nomoto K. γδ T cells in infection-induced and autoimmune-induced testicular inflammation. Immunology. 1998;95:395–401. doi: 10.1046/j.1365-2567.1998.00585.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 904.Lewis-Jones D.I., Lynch R.V., Kerrigan D.D., Davies I. Long-term study of the immuno-pathological consequences of sympathetic orchiopathia in the rat. J Reprod Fertil. 1987;80:641–647. doi: 10.1530/jrf.0.0800641. [DOI] [PubMed] [Google Scholar]
- 905.del Rio R., McAllister R.D., Meeker N.D. Identification of Orch3, a locus controlling dominant resistance to autoimmune orchitis, as kinesin family member 1C. PLoS Genet. 2012;8:e1003140. doi: 10.1371/journal.pgen.1003140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 906.Roper R.J., Doerge R.W., Call S.B., Tung K.S., Hickey W.F., Teuscher C. Autoimmune orchitis, epididymitis, and vasitis are immunogenetically distinct lesions. Am J Pathol. 1998;152:1337–1345. [PMC free article] [PubMed] [Google Scholar]
- 907.Law H.Y., Bodmer W.F., Mathews J.D., Skegg D.C. The immune response to vasectomy and its relation to the HLA system. Tissue Antigens. 1979;14:115–139. doi: 10.1111/j.1399-0039.1979.tb00830.x. [DOI] [PubMed] [Google Scholar]
- 908.Butterfield R.J., Sudweeks J.D., Blankenhorn E.P. New genetic loci that control susceptibility and symptoms of experimental allergic encephalomyelitis in inbred mice. J Immunol. 1998;161:1860–1867. [PubMed] [Google Scholar]
- 909.Meeker N.D., Hickey W.F., Korngold R. Multiple loci govern the bone marrow-derived immunoregulatory mechanism controlling dominant resistance to autoimmune orchitis. Proc Natl Acad Sci USA. 1995;92:5684–5688. doi: 10.1073/pnas.92.12.5684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 910.Trojian T.H., Lishnak T.S., Heiman D. Epididymitis and orchitis: an overview. Am Fam Physician. 2009;79:583–587. [PubMed] [Google Scholar]
- 911.La Vignera S., Vicari E., Condorelli R.A., D’Agata R., Calogero A.E. Male accessory gland infection and sperm parameters (review) Int J Androl. 2011;34:e330–e347. doi: 10.1111/j.1365-2605.2011.01200.x. [DOI] [PubMed] [Google Scholar]
- 912.Chan P.T., Schlegel P.N. Inflammatory conditions of the male excurrent ductal system. Part II. J Androl. 2002;23:461–469. [PubMed] [Google Scholar]
- 913.Itoh M., Miyamoto K., Ohga T., Takeuchi Y. Spontaneous occurrence of vasculitis-like lesions in male reproductive tissues in mice: a histological study. Arch Androl. 1999;42:151–159. doi: 10.1080/014850199262814. [DOI] [PubMed] [Google Scholar]
- 914.Itoh M., Chen X.H., Takeuchi Y., Miki T. Morphological demonstration of the immune privilege in the testis using adjuvants: tissue responses of male reproductive organs in mice injected with Bordetella pertussigens. Arch Histol Cytol. 1995;58:575–579. doi: 10.1679/aohc.58.575. [DOI] [PubMed] [Google Scholar]
- 915.Itoh M., Miyamoto K., Ooga T., Iwahashi K., Takeuchi Y. Spontaneous accumulation of eosinophils and macrophages throughout the stroma of the epididymis and vas deferens in alymphoplasia (aly) mutant mice: I. A histological study. Am J Reprod Immunol. 1999;42:246–253. doi: 10.1111/j.1600-0897.1999.tb00098.x. [DOI] [PubMed] [Google Scholar]
- 916.Ludwig M., Jantos C.A., Wolf S. Tissue penetration of sparfloxacin in a rat model of experimental Escherichia coli epididymitis. Infection. 1997;25:178–184. doi: 10.1007/BF02113609. [DOI] [PubMed] [Google Scholar]
- 917.Kaya M., Boleken M.E., Zeyrek F. Oxidative and antioxidative status in the testes of rats with acute epididymitis. Urol Int. 2006;76:353–358. doi: 10.1159/000092063. [DOI] [PubMed] [Google Scholar]
- 918.Lucchetta R., Clavert A., Meyer J.M., Bollack C. Acute experimental E. coli epididymitis in the rat and its consequences on spermatogenesis. Urol Res. 1983;11:117–120. doi: 10.1007/BF00257715. [DOI] [PubMed] [Google Scholar]
- 919.Itoh M., Mukasa A., Tokunaga Y., Hiramine C., Hojo K. New experimental model for adoptive transfer of murine autoimmune orchitis. Andrologia. 1991;23:415–420. doi: 10.1111/j.1439-0272.1991.tb02590.x. [DOI] [PubMed] [Google Scholar]
- 920.Itoh M., Hiramine C., Mukasa A. Establishment of an experimental model of autoimmune epididymo-orchitis induced by the transfer of a T-cell line in mice. Int J Androl. 1992;15:170–181. [PubMed] [Google Scholar]
- 921.Thornton A.M., Shevach E.M. CD4+CD25+ immunoregulatory T cells suppress polyclonal T cell activation in vitro by inhibiting interleukin 2 production. J Exp Med. 1998;188:287–296. doi: 10.1084/jem.188.2.287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 922.Taguchi O., Nishizuka Y. Experimental autoimmune orchitis after neonatal thymectomy in the mouse. Clin Exp Immunol. 1981;46:425–434. [PMC free article] [PubMed] [Google Scholar]
- 923.Hubert F.X., Kinkel S.A., Crewther P.E. Aire-deficient C57BL/6 mice mimicking the common human 13-base pair deletion mutation present with only a mild autoimmune phenotype. J Immunol. 2009;182:3902–3918. doi: 10.4049/jimmunol.0802124. [DOI] [PubMed] [Google Scholar]
- 924.Qu N., Naito M., Terayama H. Developmental ontogeny of autoantigens associated with localized autoimmunity in murine testis and epididymis. J Reprod Immunol. 2010;87:45–51. doi: 10.1016/j.jri.2010.05.005. [DOI] [PubMed] [Google Scholar]
- 925.Qu N., Terayama H., Naito M. Caput epididymitis but not orchitis was induced by vasectomy in a murine model of experimental autoimmune orchitis. Reproduction. 2008;135:859–866. doi: 10.1530/REP-08-0018. [DOI] [PubMed] [Google Scholar]
- 926.Itoh M., Xie Q., Miyamoto K., Takeuchi Y. Major differences between the testis and epididymis in the induction of granulomas in response to extravasated germ cells. I. A light microscopical study in mice. Int J Androl. 1999;22:316–323. doi: 10.1046/j.1365-2605.1999.00186.x. [DOI] [PubMed] [Google Scholar]
- 927.Nistal M., Mate A., Paniagua R. Granulomatous epididymal lesion of possible ischemic origin. Am J Surg Pathol. 1997;21:951–956. doi: 10.1097/00000478-199708000-00010. [DOI] [PubMed] [Google Scholar]
- 928.Itoh M., Terayama H., Naito M., Ogawa Y., Tainosho S. Tissue microcircumstances for leukocytic infiltration into the testis and epididymis in mice. J Reprod Immunol. 2005;67:57–67. doi: 10.1016/j.jri.2005.06.007. [DOI] [PubMed] [Google Scholar]
- 929.Head J.R., Billingham R.E. Immunologically privileged sites in transplantation immunology and oncology. Perspect Biol Med. 1985;29:115–131. doi: 10.1353/pbm.1985.0038. [DOI] [PubMed] [Google Scholar]
- 930.Griswold M.D. Protein secretions of Sertoli cells. Int Rev Cytol. 1988;110:133–156. doi: 10.1016/s0074-7696(08)61849-5. [DOI] [PubMed] [Google Scholar]
- 931.Cheng C.Y., Mruk D.D. The blood-testis barrier and its implications for male contraception. Pharmacol Rev. 2012;64:16–64. doi: 10.1124/pr.110.002790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 932.Bart J., Hollema H., Groen H.J. The distribution of drug-efflux pumps, P-gp, BCRP, MRP1 and MRP2, in the normal blood-testis barrier and in primary testicular tumours. Eur J Cancer. 2004;40:2064–2070. doi: 10.1016/j.ejca.2004.05.010. [DOI] [PubMed] [Google Scholar]
- 933.Su L., Mruk D.D., Lui W.Y., Lee W.M., Cheng C.Y. P-glycoprotein regulates blood-testis barrier dynamics via its effects on the occludin/zonula occludens 1 (ZO-1) protein complex mediated by focal adhesion kinase (FAK) Proc Natl Acad Sci USA. 2011;108:19623–19628. doi: 10.1073/pnas.1111414108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 934.Yule T.D., Montoya G.D., Russell L.D., Williams T.M., Tung K.S. Autoantigenic germ cells exist outside the blood testis barrier. J Immunol. 1988;141:1161–1167. [PubMed] [Google Scholar]
- 935.Whitmore W.F., III, Gittes R.F. Intratesticular grafts: the testis as an exceptional immunologically privileged site. Trans Am Assoc Genitourin Surg. 1978;70:76–80. [PubMed] [Google Scholar]
- 936.Whitmore W.F., III, Karsh L., Gittes R.F. The role of germinal epithelium and spermatogenesis in the privileged survival of intratesticular grafts. J Urol. 1985;134:782–786. doi: 10.1016/s0022-5347(17)47438-6. [DOI] [PubMed] [Google Scholar]
- 937.Selawry H.P., Whittington K. Extended allograft survival of islets grafted into intra-abdominally placed testis. Diabetes. 1984;33:405–406. doi: 10.2337/diab.33.4.405. [DOI] [PubMed] [Google Scholar]
- 938.Maddocks S., Setchell B.P. The rejection of thyroid allografts in the ovine testis. Immunol Cell Biol. 1988;66:1–8. doi: 10.1038/icb.1988.1. [DOI] [PubMed] [Google Scholar]
- 939.Setchell B.P., Granholm T., Ritzén E.M. Failure of thyroid allografts to function in the testes of cynomolgous monkeys. J Reprod Immunol. 1995;28:75–80. doi: 10.1016/0165-0378(94)00897-g. [DOI] [PubMed] [Google Scholar]
- 940.Neaves W.B., Billingham R.E. Transplantation of the testis. Transplantation. 1979;28:163–165. doi: 10.1097/00007890-197909000-00001. [DOI] [PubMed] [Google Scholar]
- 941.Statter M.B., Foglia R.P., Parks D.E., Donahoe P.K. Fetal and postnatal testis shows immunoprivilege as donor tissue. J Urol. 1988;139:204–210. doi: 10.1016/s0022-5347(17)42354-8. [DOI] [PubMed] [Google Scholar]
- 942.Barksdale E.M., Jr., McGenis T.G., Donahoe P.K. Gonadotropins moderate rejection of trophic-specific congenic testes grafts. J Pediatr Surg. 1991;26:886–892. doi: 10.1016/0022-3468(91)90831-d. [DOI] [PubMed] [Google Scholar]
- 943.Rodriguez-Sosa J.R., Dobrinski I. Recent developments in testis tissue xenografting. Reproduction. 2009;138:187–194. doi: 10.1530/REP-09-0012. [DOI] [PubMed] [Google Scholar]
- 944.Dobrinski I. Germ cell transplantation and testis tissue xenografting in domestic animals. Anim Reprod Sci. 2005;89:137–145. doi: 10.1016/j.anireprosci.2005.06.020. [DOI] [PubMed] [Google Scholar]
- 945.Herrid M., Vignarajan S., Davey R., Dobrinski I., Hill J.R. Successful transplantation of bovine testicular cells to heterologous recipients. Reproduction. 2006;132:617–624. doi: 10.1530/rep.1.01125. [DOI] [PubMed] [Google Scholar]
- 946.Kim Y., Turner D., Nelson J., Dobrinski I., McEntee M., Travis A.J. Production of donor-derived sperm after spermatogonial stem cell transplantation in the dog. Reproduction. 2008;136:823–831. doi: 10.1530/REP-08-0226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 947.Kimmel S.G., Ohbatake M., Kushida M., Merguerian P., Clarke I.D., Kim P.C. Murine xenogeneic immune responses to the human testis: a presumed immune-privileged tissue. Transplantation. 2000;69:1075–1084. doi: 10.1097/00007890-200003270-00010. [DOI] [PubMed] [Google Scholar]
- 948.Wakabayashi A., Eishi Y., Nakamura K. Regulation of experimental autoimmune orchitis by the presence or absence of testicular antigens during immunological development in SCID mice reconstituted with fetal liver cells. Immunology. 1997;92:84–90. doi: 10.1046/j.1365-2567.1997.00316.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 949.Chang C.C., Ciubotariu R., Manavalan J.S. Tolerization of dendritic cells by T(S) cells: the crucial role of inhibitory receptors ILT3 and ILT4. Nat Immunol. 2002;3:237–243. doi: 10.1038/ni760. [DOI] [PubMed] [Google Scholar]
- 950.Liu Z., Tugulea S., Cortesini R., Suciu-Foca N. Specific suppression of T helper alloreactivity by allo-MHC class I-restricted CD8+CD28− T cells. Int Immunol. 1998;10:775–783. doi: 10.1093/intimm/10.6.775. [DOI] [PubMed] [Google Scholar]
- 951.Ditzian-Kadanoff R. Testicular-associated immune deviation and prevention of adjuvant-induced arthritis by three tolerization methods. Scand J Immunol. 1999;50:150–158. doi: 10.1046/j.1365-3083.1999.00567.x. [DOI] [PubMed] [Google Scholar]
- 952.Li H., Ren J., Dhabuwala C.B., Shichi H. Immunotolerance induced by intratesticular antigen priming: expression of TGF-β, Fas and Fas ligand. Ocul Immunol Inflamm. 1997;5:75–84. doi: 10.3109/09273949709085055. [DOI] [PubMed] [Google Scholar]
- 953.Veräjänkorva E., Setälä N., Teros T., Salmi A.A., Pöllänen P. Testicular-associated immune deviation: flushing of the testicular lymph sinusoids induces immunosuppression and inhibits formation of EAE in SJL mice. Scand J Immunol. 2002;55:478–483. doi: 10.1046/j.1365-3083.2002.01083.x. [DOI] [PubMed] [Google Scholar]
- 954.Dai Z., Nasr I.W., Reel M. Impaired recall of CD8 memory T cells in immunologically privileged tissue. J Immunol. 2005;174:1165–1170. doi: 10.4049/jimmunol.174.3.1165. [DOI] [PubMed] [Google Scholar]
- 955.Nasr I.W., Wang Y., Gao G. Testicular immune privilege promotes transplantation tolerance by altering the balance between memory and regulatory T cells. J Immunol. 2005;174:6161–6168. doi: 10.4049/jimmunol.174.10.6161. [DOI] [PubMed] [Google Scholar]
- 956.Itoh M., Mukasa A., Tokunaga Y., Hiramine C., Hojo K. Suppression of efferent limb of testicular autoimmune response by a regulatory CD4+ T cell line in mice. Clin Exp Immunol. 1992;87:455–460. doi: 10.1111/j.1365-2249.1992.tb03019.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 957.Wheeler K., Tardif S., Rival C. Regulatory T cells control tolerogenic versus autoimmune response to sperm in vasectomy. Proc Natl Acad Sci USA. 2011;108:7511–7516. doi: 10.1073/pnas.1017615108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 958.Griffith T.S., Ferguson T.A. The role of FasL-induced apoptosis in immune privilege. Immunol Today. 1997;18:240–244. doi: 10.1016/s0167-5699(97)81663-5. [DOI] [PubMed] [Google Scholar]
- 959.Allison J., Georgiou H.M., Strasser A., Vaux D.L. Transgenic expression of CD95 ligand on islet beta cells induces a granulocytic infiltration but does not confer immune privilege upon islet allografts. Proc Natl Acad Sci USA. 1997;94:3943–3947. doi: 10.1073/pnas.94.8.3943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 960.Kang S.M., Schneider D.B., Lin Z. Fas ligand expression in islets of Langerhans does not confer immune privilege and instead targets them for rapid destruction. Nat Med. 1997;3:738–743. doi: 10.1038/nm0797-738. [DOI] [PubMed] [Google Scholar]
- 961.Xerri L., Devilard E., Hassoun J., Mawas C., Birg F. Fas ligand is not only expressed in immune privileged human organs but is also coexpressed with Fas in various epithelial tissues. Mol Pathol. 1997;50:87–91. doi: 10.1136/mp.50.2.87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 962.French L.E., Hahne M., Viard I. Fas and Fas ligand in embryos and adult mice: ligand expression in several immune-privileged tissues and coexpression in adult tissues characterized by apoptotic cell turnover. J Cell Biol. 1996;133:335–343. doi: 10.1083/jcb.133.2.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 963.Sanberg P.R., Saporta S., Borlongan C.V., Othberg A.I., Allen R.C., Cameron D.F. The testis-derived cultured Sertoli cell as a natural Fas-L secreting cell for immunosuppressive cellular therapy. Cell Transplant. 1997;6:191–193. doi: 10.1177/096368979700600213. [DOI] [PubMed] [Google Scholar]
- 964.Nandi S., Banerjee P.P., Zirkin B.R. Germ cell apoptosis in the testes of Sprague Dawley rats following testosterone withdrawal by ethane 1,2-dimethanesulfonate administration: relationship to Fas? Biol Reprod. 1999;61:70–75. doi: 10.1095/biolreprod61.1.70. [DOI] [PubMed] [Google Scholar]
- 965.Pöllänen P., Jahnukainen K., Punnonen J., Sainio-Pöllänen S. Ontogeny of immunosuppressive activity, MHC antigens and leukocytes in the rat testis. J Reprod Immunol. 1992;21:257–274. doi: 10.1016/0165-0378(92)90030-8. [DOI] [PubMed] [Google Scholar]
- 966.Anderson D.J., Narayan P., DeWolf W.C. Major histocompatibility antigens are not detectable on post-meiotic human testicular germ cells. J Immunol. 1984;133:1962–1965. [PubMed] [Google Scholar]
- 967.Kohno S., Ziparo E., Marek L.F., Tung K.S. Murine Sertoli cells: major histocompatibility antigens and glycoconjugates. J Reprod Immunol. 1983;5:339–350. doi: 10.1016/0165-0378(83)90243-7. [DOI] [PubMed] [Google Scholar]
- 968.Martin-Villa J.M., Longas J., Arnaiz-Villena A. Cyclic expression of HLA class I and II molecules on the surface of purified human spermatozoa and their control by serum inhibin B levels. Biol Reprod. 1999;61:1381–1386. doi: 10.1095/biolreprod61.6.1381. [DOI] [PubMed] [Google Scholar]
- 969.Jassim A., Ollier W., Payne A., Biro A., Oliver R.T., Festenstein H. Analysis of HLA antigens on germ cells in human semen. Eur J Immunol. 1989;19:1215–1220. doi: 10.1002/eji.1830190710. [DOI] [PubMed] [Google Scholar]
- 970.Fiszer D., Ulbrecht M., Fernandez N., Johnson J.P., Weiss E.H., Kurpisz M. Analysis of HLA class Ib gene expression in male gametogenic cells. Eur J Immunol. 1997;27:1691–1695. doi: 10.1002/eji.1830270715. [DOI] [PubMed] [Google Scholar]
- 971.Fournel S., Aguerre-Girr M., Huc X. Cutting edge: soluble HLA-G1 triggers CD95/CD95 ligand-mediated apoptosis in activated CD8+ cells by interacting with CD8. J Immunol. 2000;164:6100–6104. doi: 10.4049/jimmunol.164.12.6100. [DOI] [PubMed] [Google Scholar]
- 972.Ryan A.F., Grendell R.L., Geraghty D.E., Golos T.G. A soluble isoform of the rhesus monkey nonclassical MHC class I molecule Mamu-AG is expressed in the placenta and the testis. J Immunol. 2002;169:673–683. doi: 10.4049/jimmunol.169.2.673. [DOI] [PubMed] [Google Scholar]
- 973.Slukvin, Watkins D.I., Golos T.G. Tissue distribution of the mRNA for a rhesus monkey major histocompatibility class Ib molecule, Mamu-AG. Tissue Antigens. 1999;53:282–291. doi: 10.1034/j.1399-0039.1999.530309.x. [DOI] [PubMed] [Google Scholar]
- 974.Larsen M.H., Bzorek M., Pass M.B. Human leukocyte antigen-G in the male reproductive system and in seminal plasma. Mol Hum Reprod. 2011;17:727–738. doi: 10.1093/molehr/gar052. [DOI] [PubMed] [Google Scholar]
- 975.Magnus T., Schreiner B., Korn T. Microglial expression of the B7 family member B7 homolog 1 confers strong immune inhibition: implications for immune responses and autoimmunity in the CNS. J Neurosci. 2005;25:2537–2546. doi: 10.1523/JNEUROSCI.4794-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 976.Piccioli D., Sbrana S., Melandri E., Valiante N.M. Contact-dependent stimulation and inhibition of dendritic cells by natural killer cells. J Exp Med. 2002;195:335–341. doi: 10.1084/jem.20010934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 977.Andrews D.M., Scalzo A.A., Yokoyama W.M., Smyth M.J., Degli-Esposti M.A. Functional interactions between dendritic cells and NK cells during viral infection. Nat Immunol. 2003;4:175–181. doi: 10.1038/ni880. [DOI] [PubMed] [Google Scholar]
- 978.Mocikat R., Braumuller H., Gumy A. Natural killer cells activated by MHC class Ilow targets prime dendritic cells to induce protective CD8 T cell responses. Immunity. 2003;19:561–569. doi: 10.1016/s1074-7613(03)00264-4. [DOI] [PubMed] [Google Scholar]
- 979.Nakamura T., Sonoda K.H., Faunce D.E. CD4+ NKT cells, but not conventional CD4+ T cells, are required to generate efferent CD8+ T regulatory cells following antigen inoculation in an immune-privileged site. J Immunol. 2003;171:1266–1271. doi: 10.4049/jimmunol.171.3.1266. [DOI] [PubMed] [Google Scholar]
- 980.Sonoda K.H., Stein-Streilein J. Ocular immune privilege and CD1d-reactive natural killer T cells. Cornea. 2002;21:S33–S38. doi: 10.1097/00003226-200203001-00008. [DOI] [PubMed] [Google Scholar]
- 981.Skelsey M.E., Mellon J., Niederkorn J.Y. γδ T cells are needed for ocular immune privilege and corneal graft survival. J Immunol. 2001;166:4327–4333. doi: 10.4049/jimmunol.166.7.4327. [DOI] [PubMed] [Google Scholar]
- 982.Born W., Wekerle H. Leydig cells nonspecifically suppress lymphoproliferation in vitro: implications for the testis as an immunologically privileged site. Am J Reprod Immunol. 1982;2:291–295. doi: 10.1111/j.1600-0897.1982.tb00195.x. [DOI] [PubMed] [Google Scholar]
- 983.Hurtenbach U., Morgenstern F., Bennett D. Induction of tolerance in vitro by autologous murine testicular cells. J Exp Med. 1980;151:827–838. doi: 10.1084/jem.151.4.827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 984.Hurtenbach U., Shearer G.M. Germ cell-induced immune suppression in mice. Effect of inoculation of syngeneic spermatozoa on cell-mediated immune responses. J Exp Med. 1982;155:1719–1729. doi: 10.1084/jem.155.6.1719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 985.Cheng X., Dai H., Wan N., Moore Y., Vankayalapati R., Dai Z. Interaction of programmed death-1 and programmed death-1 ligand-1 contributes to testicular immune privilege. Transplantation. 2009;87:1778–1786. doi: 10.1097/TP.0b013e3181a75633. [DOI] [PubMed] [Google Scholar]
- 986.Selawry H.P., Whittington K.B. Prolonged intratesticular islet allograft survival is not dependent on local steroidogenesis. Horm Metab Res. 1988;20:562–565. doi: 10.1055/s-2007-1010885. [DOI] [PubMed] [Google Scholar]
- 987.Bistoni G., Calvitti M., Mancuso F. Prolongation of skin allograft survival in rats by the transplantation of microencapsulated xenogeneic neonatal porcine Sertoli cells. Biomaterials. 2012;33:5333–5340. doi: 10.1016/j.biomaterials.2012.04.020. [DOI] [PubMed] [Google Scholar]
- 988.Korbutt G.S., Suarez-Pinzon W.L., Power R.F., Rajotte R.V., Rabinovitch A. Testicular Sertoli cells exert both protective and destructive effects on syngeneic islet grafts in non-obese diabetic mice. Diabetologia. 2000;43:474–480. doi: 10.1007/s001250051331. [DOI] [PubMed] [Google Scholar]
- 989.Mital P., Kaur G., Dufour J.M. Immunoprotective Sertoli cells: making allogeneic and xenogeneic transplantation feasible. Reproduction. 2010;139:495–504. doi: 10.1530/REP-09-0384. [DOI] [PubMed] [Google Scholar]
- 990.Suarez-Pinzon W., Korbutt G.S., Power R., Hooton J., Rajotte R.V., Rabinovitch A. Testicular Sertoli cells protect islet β-cells from autoimmune destruction in NOD mice by a transforming growth factor-β1-dependent mechanism. Diabetes. 2000;49:1810–1818. doi: 10.2337/diabetes.49.11.1810. [DOI] [PubMed] [Google Scholar]
- 991.Valdés-González R.A., Dorantes L.M., Garibay G.N. Xenotransplantation of porcine neonatal islets of Langerhans and Sertoli cells: a 4-year study. Eur J Endocrinol. 2005;153:419–427. doi: 10.1530/eje.1.01982. [DOI] [PubMed] [Google Scholar]
- 992.Dufour J.M., Lord S.J., Kin T. Comparison of successful and unsuccessful islet/Sertoli cell cotransplant grafts in streptozotocin-induced diabetic mice. Cell Transplant. 2008;16:1029–1038. [PubMed] [Google Scholar]
- 993.Turek P.J., Malkowicz S.B., Tomaszewski J.E., Wein A.J., Peehl D. The role of the Sertoli cell in active immunosuppression in the human testis. Br J Urol. 1996;77:891–895. doi: 10.1046/j.1464-410x.1996.00322.x. [DOI] [PubMed] [Google Scholar]
- 994.Riccioli A., Filippini A., De Cesaris P. Inflammatory mediators increase surface expression of integrin ligands, adhesion to lymphocytes, and secretion of interleukin 6 in mouse Sertoli cells. Proc Natl Acad Sci USA. 1995;92:5808–5812. doi: 10.1073/pnas.92.13.5808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 995.Fan P., He L., Pu D. Testicular Sertoli cells influence the proliferation and immunogenicity of co-cultured endothelial cells. Biochem Biophys Res Comm. 2011;404:829–833. doi: 10.1016/j.bbrc.2010.12.068. [DOI] [PubMed] [Google Scholar]
- 996.Teng Y., Xue W.J., Ding X.M. Isolation and culture of adult Sertoli cells and their effects on the function of co-cultured allogeneic islets in vitro. Chin Med J. 2005;118:1857–1862. [PubMed] [Google Scholar]
- 997.Doyle T.J., Kaur G., Putrevu S.M. Immunoprotective properties of primary Sertoli cells in mice: potential functional pathways that confer immune privilege. Biol Reprod. 2012;86:6. doi: 10.1095/biolreprod.110.089425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 998.Nikolova D.B., Kancheva L.S., Surneva M.D., Martinova Y.S. Species-specific effect of proteins secreted by cultured pre-pubertal rat Sertoli cells on natural killer cell activity. Immunopharmacology. 1992;23:15–20. doi: 10.1016/0162-3109(92)90004-v. [DOI] [PubMed] [Google Scholar]
- 999.Yin Z.Z., Xie L., Zeng M.H. Sertoli cells induce xenolymphocyte apoptosis in vitro. Transplant Proc. 2006;38:3309–3311. doi: 10.1016/j.transproceed.2006.10.126. [DOI] [PubMed] [Google Scholar]
- 1000.Li Y., Xue W.J., Tian X.H. Study on systemic immune tolerance induction in rat islet transplantation by intravenous infusion of Sertoli cells. Transplantation. 2010;89:1430–1437. doi: 10.1097/TP.0b013e3181da607e. [DOI] [PubMed] [Google Scholar]
- 1001.Takeda Y., Gotoh M., Dono K. Protection of islet allografts transplanted together with Fas ligand expressing testicular allografts. Diabetologia. 1998;41:315–321. doi: 10.1007/s001250050909. [DOI] [PubMed] [Google Scholar]
- 1002.Lee H.M., Oh B.C., Lim D.P. Role of complement regulatory proteins in the survival of murine allo-transplanted Sertoli cells. J Korean Med Sci. 2007;22:277–282. doi: 10.3346/jkms.2007.22.2.277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1003.Sipione S., Simmen K.C., Lord S.J. Identification of a novel human granzyme B inhibitor secreted by cultured Sertoli cells. J Immunol. 2006;177:5051–5058. doi: 10.4049/jimmunol.177.8.5051. [DOI] [PubMed] [Google Scholar]
- 1004.Yin Z., Wang L., Xiang Y. Resistance of neonatal porcine Sertoli cells to human xenoantibody and complement-mediated lysis is associated with low expression of α-Gal and high production of clusterin and CD59. Xenotransplantation. 2010;17:215–223. doi: 10.1111/j.1399-3089.2010.00581.x. [DOI] [PubMed] [Google Scholar]
- 1005.Pallotta M.T., Orabona C., Volpi C. Indoleamine 2,3-dioxygenase is a signaling protein in long-term tolerance by dendritic cells. Nat Immunol. 2011;12:870–878. doi: 10.1038/ni.2077. [DOI] [PubMed] [Google Scholar]
- 1006.Fallarino F., Luca G., Calvitti M. Therapy of experimental type 1 diabetes by isolated Sertoli cell xenografts alone. J Exp Med. 2009;206:2511–2526. doi: 10.1084/jem.20090134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1007.Munn D.H., Mellor A.L. Indoleamine 2,3-dioxygenase and tumor-induced tolerance. J Clin Invest. 2007;117:1147–1154. doi: 10.1172/JCI31178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1008.Meng J., Greenlee A.R., Taub C.J., Braun R.E. Sertoli cell-specific deletion of the androgen receptor compromises testicular immune privilege in mice. Biol Reprod. 2011;85:254–260. doi: 10.1095/biolreprod.110.090621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1009.Guazzone V.A., Rival C., Denduchis B., Lustig L. Monocyte chemoattractant protein-1 (MCP-1/CCL2) in experimental autoimmune orchitis. J Reprod Immunol. 2003;60:143–157. doi: 10.1016/j.jri.2003.08.001. [DOI] [PubMed] [Google Scholar]
- 1010.Collin O., Bergh A. Leydig cells secrete factors which increase vascular permeability and endothelial cell proliferation. Int J Androl. 1996;19:221–228. doi: 10.1111/j.1365-2605.1996.tb00466.x. [DOI] [PubMed] [Google Scholar]
- 1011.Maddocks S., Sharpe R.M. Interstitial fluid volume in the rat testis: androgen-dependent regulation by the seminiferous tubules? J Endocrinol. 1989;120:215–222. doi: 10.1677/joe.0.1200215. [DOI] [PubMed] [Google Scholar]
- 1012.Rivenson A., Ohmori T., Hamazaki M., Madden R. Cell surface recognition: spontaneous identification of mouse Leydig cells by lymphocytes, macrophages and eosinophils. Cell Mol Biol. 1981;27:49–56. [PubMed] [Google Scholar]
- 1013.Born W., Wekerle H. Selective, immunologically nonspecific adherence of lymphoid and myeloid cells to Leydig cells. Eur J Cell Biol. 1981;25:76–81. [PubMed] [Google Scholar]
- 1014.Sainio-Pöllänen S., Sundström J., Erkkilä S. CD106 (VCAM-1) in testicular immunoregulation. J Reprod Immunol. 1997;33:221–238. doi: 10.1016/s0165-0378(97)00024-7. [DOI] [PubMed] [Google Scholar]
- 1015.Hedger M.P., Qin J.X., Robertson D.M., de Kretser D.M. Intragonadal regulation of immune system functions. Reprod Fertil Dev. 1990;2:263–280. doi: 10.1071/rd9900263. [DOI] [PubMed] [Google Scholar]
- 1016.Abe R., Peng T., Sailors J., Bucala R., Metz C.N. Regulation of the CTL response by macrophage migration inhibitory factor. J Immunol. 2001;166:747–753. doi: 10.4049/jimmunol.166.2.747. [DOI] [PubMed] [Google Scholar]
- 1017.Apte R.S., Sinha D., Mayhew E., Wistow G.J., Niederkorn J.Y. Role of macrophage migration inhibitory factor in inhibiting NK cell activity and preserving immune privilege. J Immunol. 1998;160:5693–5696. [PubMed] [Google Scholar]
- 1018.Repp A.C., Mayhew E.S., Apte S., Niederkorn J.Y. Human uveal melanoma cells produce macrophage migration-inhibitory factor to prevent lysis by NK cells. J Immunol. 2000;165:710–715. doi: 10.4049/jimmunol.165.2.710. [DOI] [PubMed] [Google Scholar]
- 1019.Malkin C.J., Pugh P.J., Jones R.D., Kapoor D., Channer K.S., Jones T.H. The effect of testosterone replacement on endogenous inflammatory cytokines and lipid profiles in hypogonadal men. J Clin Endocrinol Metab. 2004;89:3313–3318. doi: 10.1210/jc.2003-031069. [DOI] [PubMed] [Google Scholar]
- 1020.Cameron D.F., Whittington K., Schultz R.E., Selawry H.P. Successful islet/abdominal testis transplantation does not require Leydig cells. Transplantation. 1990;50:649–653. doi: 10.1097/00007890-199010000-00024. [DOI] [PubMed] [Google Scholar]
- 1021.Li X., Strauss L., Kaatrasalo A. Transgenic mice expressing P450 aromatase as a model for male infertility associated with chronic inflammation in the testis. Endocrinology. 2006;147:1271–1277. doi: 10.1210/en.2005-0654. [DOI] [PubMed] [Google Scholar]
- 1022.Bhardwaj R.S., Schwarz A., Becher E. Pro-opiomelanocortin-derived peptides induce IL-10 production in human monocytes. J Immunol. 1996;156:2517–2521. [PubMed] [Google Scholar]
- 1023.Hedger M.P., Nikolic-Paterson D.J., Hutchinson P., Atkins R.C., de Kretser D.M. Immunoregulatory activity in adult rat testicular interstitial fluid: roles of interleukin-1 and transforming growth factor β. Biol Reprod. 1998;58:927–934. doi: 10.1095/biolreprod58.4.927. [DOI] [PubMed] [Google Scholar]
- 1024.Emoto M., Nishikawa F., Oku D., Hamuro A., Kita E., Kashiba S. Suppressive effect of a mouse testicular extract on lymphocyte activation. Int J Androl. 1991;14:291–302. doi: 10.1111/j.1365-2605.1991.tb01095.x. [DOI] [PubMed] [Google Scholar]
- 1025.Hedger M.P., Wang J., Lan H.Y., Atkins R.C., Wreford N.G. Immunoregulatory activity in adult rat testicular interstitial fluid: relationship with intratesticular CD8+ lymphocytes following treatment with ethane dimethane sulfonate and testosterone implants. Biol Reprod. 1998;58:935–942. doi: 10.1095/biolreprod58.4.935. [DOI] [PubMed] [Google Scholar]
- 1026.Abayasekara D.R., Kurlak L.O., Jeremy J.Y., Dandona P., Sharpe R.M., Cooke B.A. The levels and possible involvement of leukotriene B4 and prostaglandin F2α in the control of interstitial fluid volume in the rat testis. Int J Androl. 1990;13:408–418. doi: 10.1111/j.1365-2605.1990.tb01049.x. [DOI] [PubMed] [Google Scholar]
- 1027.Pöllänen P., von Euler M., Jahnukainen K. Role of transforming growth factor β in testicular immunosuppression. J Reprod Immunol. 1993;24:123–137. doi: 10.1016/0165-0378(93)90015-a. [DOI] [PubMed] [Google Scholar]
- 1028.Veräjänkorva E., Martikainen M., Pöllänen P. Cytokines in the BALB/c mouse testis in various conditions. Asian J Androl. 2001;3:9–19. [PubMed] [Google Scholar]
- 1029.Foulds L.M., Boysen R.I., Crane M. Molecular identification of lyso-glycerophosphocholines as endogenous immunosuppressives in bovine and rat gonadal fluids. Biol Reprod. 2008;79:525–536. doi: 10.1095/biolreprod.107.064386. [DOI] [PubMed] [Google Scholar]
- 1030.Kabarowski J.H., Xu Y., Witte O.N. Lysophosphatidylcholine as a ligand for immunoregulation. Biochem Pharmacol. 2002;64:161–167. doi: 10.1016/s0006-2952(02)01179-6. [DOI] [PubMed] [Google Scholar]
- 1031.Im Y.J., Lee Y.K., Chung H.Y., Im D.S. Multiple actions of lysophosphatidylcholine in human Jurkat T cells. Acta Pharmacol Sin. 2006;27:700–707. doi: 10.1111/j.1745-7254.2006.00339.x. [DOI] [PubMed] [Google Scholar]
- 1032.Chang D.H., Deng H., Matthews P. Inflammation-associated lysophospholipids as ligands for CD1d-restricted T cells in human cancer. Blood. 2008;112:1308–1316. doi: 10.1182/blood-2008-04-149831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1033.Ye X. Lysophospholipid signaling in the function and pathology of the reproductive system. Hum Reprod Update. 2008;14:519–536. doi: 10.1093/humupd/dmn023. [DOI] [PubMed] [Google Scholar]
- 1034.Schell C., Frungieri M.B., Albrecht M. A prostaglandin D2 system in the human testis. Fertil Steril. 2007;88:233–236. doi: 10.1016/j.fertnstert.2006.11.100. [DOI] [PubMed] [Google Scholar]
- 1035.Jannini E.A., Ulisse S., Cecconi S. Follicle-stimulating hormone-induced phospholipase A2 activity and eicosanoid generation in rat Sertoli cells. Biol Reprod. 1994;51:140–145. doi: 10.1095/biolreprod51.1.140. [DOI] [PubMed] [Google Scholar]
- 1036.Reddy G.P., Prasad M., Sailesh S., Kumar Y.V., Reddanna P. The production of arachidonic acid metabolites in rat testis. Prostaglandins. 1992;44:497–507. doi: 10.1016/0090-6980(92)90020-t. [DOI] [PubMed] [Google Scholar]
- 1037.Hart P.H., Whitty G.A., Piccoli D.S., Hamilton J.A. Control by IFN-γ and PGE2 of TNFα and IL-1 production by human monocytes. Immunology. 1989;66:376–383. [PMC free article] [PubMed] [Google Scholar]
- 1038.Kunkel S.L., Spengler M., May M.A., Spengler R., Larrick J., Remick D. Prostaglandin E2 regulates macrophage-derived tumor necrosis factor gene expression. J Biol Chem. 1988;263:5380–5384. [PubMed] [Google Scholar]
- 1039.Scales W.E., Chensue S.W., Otterness I., Kunkel S.L. Regulation of monokine gene expression: prostaglandin E2 suppresses tumor necrosis factor but not interleukin-1α or β-mRNA and cell-associated bioactivity. J Leuk Biol. 1989;45:416–421. [PubMed] [Google Scholar]
- 1040.Brinster R.L., Zimmermann J.W. Spermatogenesis following male germ-cell transplantation. Proc Natl Acad Sci USA. 1994;91:11298–11302. doi: 10.1073/pnas.91.24.11298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1041.Shinohara T., Kato M., Takehashi M. Rats produced by interspecies spermatogonial transplantation in mice and in vitro microinsemination. Proc Natl Acad Sci USA. 2006;103:13624–13628. doi: 10.1073/pnas.0604205103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1042.Dobrinski I., Avarbock M.R., Brinster R.L. Germ cell transplantation from large domestic animals into mouse testes. Mol Reprod Dev. 2000;57:270–279. doi: 10.1002/1098-2795(200011)57:3<270::AID-MRD9>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- 1043.Nagano M., Patrizio P., Brinster R.L. Long-term survival of human spermatogonial stem cells in mouse testes. Fertil Steril. 2002;78:1225–1233. doi: 10.1016/s0015-0282(02)04345-5. [DOI] [PubMed] [Google Scholar]
- 1044.Jiang F.X., Short R.V. Male germ cell transplantation in rats: apparent synchronization of spermatogenesis between host and donor seminiferous epithelia. Int J Androl. 1995;18:326–330. doi: 10.1111/j.1365-2605.1995.tb00570.x. [DOI] [PubMed] [Google Scholar]
- 1045.Ogawa T., Dobrinski I., Brinster R.L. Recipient preparation is critical for spermatogonial transplantation in the rat. Tissue Cell. 1999;31:461–472. doi: 10.1054/tice.1999.0060. [DOI] [PubMed] [Google Scholar]
- 1046.Schlatt S., Rosiepen G., Weinbauer G.F., Rolf C., Brook P.F., Nieschlag E. Germ cell transfer into rat, bovine, monkey and human testes. Hum Reprod. 1999;14:144–150. doi: 10.1093/humrep/14.1.144. [DOI] [PubMed] [Google Scholar]
- 1047.Shinohara T., Orwig K.E., Avarbock M.R., Brinster R.L. Restoration of spermatogenesis in infertile mice by Sertoli cell transplantation. Biol Reprod. 2003;68:1064–1071. doi: 10.1095/biolreprod.102.009977. [DOI] [PubMed] [Google Scholar]
- 1048.Zhang Z., Shao S., Shetty G., Meistrich M.L. Donor Sertoli cells transplanted into irradiated rat testes stimulate partial recovery of endogenous spermatogenesis. Reproduction. 2009;137:497–508. doi: 10.1530/REP-08-0120. [DOI] [PubMed] [Google Scholar]
- 1049.Lo K.C., Lei Z., Rao Ch V., Beck J., Lamb D.J. De novo testosterone production in luteinizing hormone receptor knockout mice after transplantation of Leydig stem cells. Endocrinology. 2004;145:4011–4015. doi: 10.1210/en.2003-1729. [DOI] [PubMed] [Google Scholar]
- 1050.Hill J.R., Dobrinski I. Male germ cell transplantation in livestock. Reprod Fertil Dev. 2006;18:13–18. doi: 10.1071/rd05123. [DOI] [PubMed] [Google Scholar]
- 1051.Dobrinski I., Travis A.J. Germ cell transplantation for the propagation of companion animals, non-domestic and endangered species. Reprod Fertil Dev. 2007;19:732–739. doi: 10.1071/rd07036. [DOI] [PubMed] [Google Scholar]
- 1052.Hermann B.P., Sukhwani M., Winkler F. Spermatogonial stem cell transplantation into Rhesus testes regenerates spermatogenesis producing functional sperm. Cell Stem Cell. 2012;11:715–726. doi: 10.1016/j.stem.2012.07.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1053.Qu N., Naito M., Li J., Terayama H., Hirai S., Itoh M. Xenogeneic and endogenous spermatogenesis following transplantation of rat germ cells into testes of immunocompetent mice. Reprod Fertil Dev. 2012;24:337–343. doi: 10.1071/RD10349. [DOI] [PubMed] [Google Scholar]
- 1054.Parreira G.G., Ogawa T., Avarbock M.R., França L.R., Brinster R.L., Russell L.D. Development of germ cell transplants in mice. Biol Reprod. 1998;59:1360–1370. doi: 10.1095/biolreprod59.6.1360. [DOI] [PubMed] [Google Scholar]
- 1055.Dias F.F., Chiarini-Garcia H., Parreira G.G., Melo R.C. Mice spermatogonial stem cells transplantation induces macrophage migration into the seminiferous epithelium and lipid body formation: high-resolution light microscopy and ultrastructural studies. Microsc Microanal. 2011;17:1002–1014. doi: 10.1017/S1431927611012098. [DOI] [PubMed] [Google Scholar]
- 1056.Carson M.J., Doose J.M., Melchior B., Schmid C.D., Ploix C.C. CNS immune privilege: hiding in plain sight. Immunol Rev. 2006;213:48–65. doi: 10.1111/j.1600-065X.2006.00441.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1057.Hedger M.P. Immunologically privileged environments. In: Halberstadt C.R., Emerich D.F., editors. Cellular transplantation from laboratory to clinic. Elsevier; Amsterdam: 2007. pp. 567–590. [Google Scholar]
- 1058.Kazeem A.A. A critical consideration of the rat epididymis as an immunologically privileged site. Scand J Immunol. 1988;27:149–156. doi: 10.1111/j.1365-3083.1988.tb02333.x. [DOI] [PubMed] [Google Scholar]
- 1059.Neaves W.B., Billingham R.E. The lymphatic drainage of the rat prostate and its status as an immunologically privileged site. Transplantation. 1979;27:127–132. doi: 10.1097/00007890-197902000-00011. [DOI] [PubMed] [Google Scholar]
- 1060.Britan A., Maffre V., Tone S., Drevet J.R. Quantitative and spatial differences in the expression of tryptophan-metabolizing enzymes in mouse epididymis. Cell Tissue Res. 2006;324:301–310. doi: 10.1007/s00441-005-0151-7. [DOI] [PubMed] [Google Scholar]
- 1061.Jrad-Lamine A., Henry-Berger J., Gourbeyre P. Deficient tryptophan catabolism along the kynurenine pathway reveals that the epididymis is in a unique tolerogenic state. J Biol Chem. 2011;286:8030–8042. doi: 10.1074/jbc.M110.172114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1062.Kalb T.H., Chuang M.T., Marom Z., Mayer L. Evidence for accessory cell function by class II MHC antigen-expressing airway epithelial cells. Am J Respir Cell Mol Biol. 1991;4:320–329. doi: 10.1165/ajrcmb/4.4.320. [DOI] [PubMed] [Google Scholar]
- 1063.Campbell N., Yio X.Y., So L.P., Li Y., Mayer L. The intestinal epithelial cell: processing and presentation of antigen to the mucosal immune system. Immunol Rev. 1999;172:315–324. doi: 10.1111/j.1600-065x.1999.tb01375.x. [DOI] [PubMed] [Google Scholar]
- 1064.Moore H.D., Bedford J.M. The differential absorptive activity of epithelial cells of the rat epididymus before and after castration. Anat Rec. 1979;193:313–327. doi: 10.1002/ar.1091930210. [DOI] [PubMed] [Google Scholar]
- 1065.Quintar A.A., Nishioka S., Torres A.I., Maldonado C.A., Aoki A. The initial segment of the rat epididymis is able to uptake immature germ cells shed by testicular damage. Cells Tissues Organs. 2011;193:170–183. doi: 10.1159/000317127. [DOI] [PubMed] [Google Scholar]
- 1066.Veräjänkorva E., Pöllänen P., Hänninen A., Martikainen M., Sundström J., Antola H. IL-10 is highly expressed in the cryptorchid cryptepididymal epithelium: a probable mechanism preventing immune responses against autoantigenic spermatozoa in the epididymal tubule. Int J Androl. 2002;25:129–133. doi: 10.1046/j.1365-2605.2002.00336.x. [DOI] [PubMed] [Google Scholar]
- 1067.Koyama K., Hasegawa A., Komori S. Functional aspects of CD52 in reproduction. J Reprod Immunol. 2009;83:56–59. doi: 10.1016/j.jri.2009.06.263. [DOI] [PubMed] [Google Scholar]
- 1068.Watanabe T., Masuyama J., Sohma Y. CD52 is a novel costimulatory molecule for induction of CD4+ regulatory T cells. Clin Immunol. 2006;120:247–259. doi: 10.1016/j.clim.2006.05.006. [DOI] [PubMed] [Google Scholar]
- 1069.Ito K., Hasegawa A., Komori S., Koyama K. Biochemical property and immunogenicity of mouse male reproductive tract CD52 (mrt-CD52) J Reprod Immunol. 2007;75:32–39. doi: 10.1016/j.jri.2007.03.001. [DOI] [PubMed] [Google Scholar]
- 1070.Hardiyanto L., Hasegawa A., Komori S. The N-linked carbohydrate moiety of male reproductive tract CD52 (mrt-CD52) interferes with the complement system via binding to C1q. J Reprod Immunol. 2012;94:142–150. doi: 10.1016/j.jri.2012.01.002. [DOI] [PubMed] [Google Scholar]
- 1071.Ashida E.R., Scofield V.L. Lymphocyte major histocompatibility complex-encoded class II structures may act as sperm receptors. Proc Natl Acad Sci USA. 1987;84:3395–3399. doi: 10.1073/pnas.84.10.3395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1072.Scofield V.L., Clisham R., Bandyopadhyay L., Gladstone P., Zamboni L., Raghupathy R. Binding of sperm to somatic cells via HLA-DR. Modulation by sulfated carbohydrates. J Immunol. 1992;148:1718–1724. [PubMed] [Google Scholar]
- 1073.Petersen B.H., Lammel C.J., Stites D.P., Brooks G.F. Human seminal plasma inhibition of complement. J Lab Clin Med. 1980;96:582–591. [PubMed] [Google Scholar]
- 1074.Vallely P.J., Sharrard R.M., Rees R.C. The identification of factors in seminal plasma responsible for suppression of natural killer cell activity. Immunology. 1988;63:451–456. [PMC free article] [PubMed] [Google Scholar]
- 1075.James K., Hargreave T.B. Immunosuppression by seminal plasma and its possible clinical significance. Immunol Today. 1984;5:353–363. doi: 10.1016/0167-5699(84)90079-3. [DOI] [PubMed] [Google Scholar]
- 1076.Robertson S.A., Prins J.R., Sharkey D.J., Moldenhauer L.M. Seminal fluid and the generation of regulatory T cells for embryo implantation. Am J Reprod Immunol. 2013;69:315–330. doi: 10.1111/aji.12107. [DOI] [PubMed] [Google Scholar]
- 1077.Kelly R.W., Holland P., Skibinski G. Extracellular organelles (prostasomes) are immunosuppressive components of human semen. Clin Exp Immunol. 1991;86:550–556. doi: 10.1111/j.1365-2249.1991.tb02968.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1078.Ronquist G., Brody I. The prostasome: its secretion and function in man. Biochim Biophys Acta. 1985;822:203–218. doi: 10.1016/0304-4157(85)90008-5. [DOI] [PubMed] [Google Scholar]
- 1079.Allen R.D., Roberts T.K. The relationship between the immunosuppressive and cytotoxic effects of human seminal plasma. Am J Reprod Immunol Microbiol. 1986;11:59–64. doi: 10.1111/j.1600-0897.1986.tb00030.x. [DOI] [PubMed] [Google Scholar]
- 1080.Quayle A.J., Kelly R.W., Hargreave T.B., James K. Immunosuppression by seminal prostaglandins. Clin Exp Immunol. 1989;75:387–391. [PMC free article] [PubMed] [Google Scholar]
- 1081.Fahmi H.A., Hunter A.G., Markham R.J., Seguin B.E. Identification of an immunosuppressive protein in bovine seminal plasma with activity against bovine lymphocytes. J Dairy Sci. 1985;68:2322–2328. doi: 10.3168/jds.S0022-0302(85)81106-1. [DOI] [PubMed] [Google Scholar]
- 1082.Veselský L., Dostál J., Kraus M. Reverse effect of indomethacin on the immunosuppressive activity of boar seminal immunosuppressive fraction. Anim Reprod Sci. 2002;71:111–123. doi: 10.1016/s0378-4320(02)00015-5. [DOI] [PubMed] [Google Scholar]
- 1083.Loras B., Vételé F., El Malki A., Rollet J., Soufir J.C., Benahmed M. Seminal transforming growth factor-β in normal and infertile men. Hum Reprod. 1999;14:1534–1539. doi: 10.1093/humrep/14.6.1534. [DOI] [PubMed] [Google Scholar]
- 1084.Miller L.J., Fischer K.A., Goralnick S.J. Interleukin-10 levels in seminal plasma: implications for chronic prostatitis-chronic pelvic pain syndrome. J Urol. 2002;167:753–756. doi: 10.1016/S0022-5347(01)69139-0. [DOI] [PubMed] [Google Scholar]
- 1085.Nocera M., Chu T.M. Transforming growth factor β as an immunosuppressive protein in human seminal plasma. Am J Reprod Immunol. 1993;30:1–8. doi: 10.1111/j.1600-0897.1993.tb00594.x. [DOI] [PubMed] [Google Scholar]
- 1086.Harris C.L., Mizuno M., Morgan B.P. Complement and complement regulators in the male reproductive system. Mol Immunol. 2006;43:57–67. doi: 10.1016/j.molimm.2005.06.026. [DOI] [PubMed] [Google Scholar]
- 1087.Mizuno M., Harris C.L., Morgan B.P. Spermatogenic cells distal to the blood-testis barrier in rats lack C3 convertase regulators and may be at risk of complement-mediated injury. J Reprod Immunol. 2006;69:23–34. doi: 10.1016/j.jri.2005.11.002. [DOI] [PubMed] [Google Scholar]
- 1088.Allen J.C., Smith C.J., Hussain J.I., Thomas J.M., Gaugas J.M. Inhibition of lymphocyte proliferation by polyamines requires ruminant-plasma polyamine oxidase. Eur J Biochem. 1979;102:153–158. doi: 10.1111/j.1432-1033.1979.tb06275.x. [DOI] [PubMed] [Google Scholar]
- 1089.Kimes B.W., Morris D.R. Preparation and stability of oxidized polyamines. Biochim Biophys Acta. 1971;228:223–234. doi: 10.1016/0005-2787(71)90562-4. [DOI] [PubMed] [Google Scholar]
- 1090.Maayan R., Zukerman Z., Shohat B. Oxidation of polyamines in human seminal plasma: a possible role in immunological infertility. Arch Androl. 1995;34:95–99. doi: 10.3109/01485019508987836. [DOI] [PubMed] [Google Scholar]
- 1091.Imade G.E., Baker H.W., de Kretser D.M., Hedger M.P. Immunosuppressive activities in the seminal plasma of infertile men: relationship to sperm antibodies and autoimmunity. Hum Reprod. 1997;12:256–262. doi: 10.1093/humrep/12.2.256. [DOI] [PubMed] [Google Scholar]
- 1092.Kelly R.W., Taylor P.L., Hearn J.P., Short R.V., Martin D.E., Marston J.H. 19-Hydroxyprostaglandin E1 as a major component of the semen of primates. Nature. 1976;260:544–545. doi: 10.1038/260544a0. [DOI] [PubMed] [Google Scholar]
- 1093.Kelly R.W., Quayle A.J., Wallace E.M., Wu F.C., Hargreave T.B., James K. Immunosuppression by seminal plasma from fertile and infertile men: inhibition of natural killer cell function correlates with seminal PG concentration. Prostagland Leukot Essent Fatty Acids. 1991;42:257–260. doi: 10.1016/0952-3278(91)90092-j. [DOI] [PubMed] [Google Scholar]
- 1094.Morelli A.E., Thomson A.W. Dendritic cells under the spell of prostaglandins. Trends Immunol. 2003;24:108–111. doi: 10.1016/s1471-4906(03)00023-1. [DOI] [PubMed] [Google Scholar]
- 1095.Lenicov F.R., Rodrigues C.R., Sabatté J. Semen promotes the differentiation of tolerogenic dendritic cells. J Immunol. 2012;189:4777–4786. doi: 10.4049/jimmunol.1202089. [DOI] [PubMed] [Google Scholar]
- 1096.Nocera M., Chu T.M. Characterization of latent transforming growth factor-β from human seminal plasma. Am J Reprod Immunol. 1995;33:282–291. doi: 10.1111/j.1600-0897.1995.tb00897.x. [DOI] [PubMed] [Google Scholar]
- 1097.Shoskes D.A., Albakri Q., Thomas K., Cook D. Cytokine polymorphisms in men with chronic prostatitis/chronic pelvic pain syndrome: association with diagnosis and treatment response. J Urol. 2002;168:331–335. [PubMed] [Google Scholar]
- 1098.Anderson R.A., Evans L.W., Irvine D.S., McIntyre M.A., Groome N.P., Riley S.C. Follistatin and activin A production by the male reproductive tract. Hum Reprod. 1998;13:3319–3325. doi: 10.1093/humrep/13.12.3319. [DOI] [PubMed] [Google Scholar]
- 1099.Ludlow H., Phillips D.J., Myers M. A new ‘total’ activin B enzyme-linked immunosorbent assay (ELISA): development and validation for human samples. Clin Endocrinol. 2009;71:867–873. doi: 10.1111/j.1365-2265.2009.03567.x. [DOI] [PubMed] [Google Scholar]
- 1100.Thomas T.Z., Chapman S.M., Hong W. Inhibins, activins, and follistatins: expression of mRNAs and cellular localization in tissues from men with benign prostatic hyperplasia. Prostate. 1998;34:34–43. doi: 10.1002/(sici)1097-0045(19980101)34:1<34::aid-pros5>3.0.co;2-k. [DOI] [PubMed] [Google Scholar]
- 1101.Robertson S.A., Ingman W.V., O’Leary S., Sharkey D.J., Tremellen K.P. Transforming growth factor β – a mediator of immune deviation in seminal plasma. J Reprod Immunol. 2002;57:109–128. doi: 10.1016/s0165-0378(02)00015-3. [DOI] [PubMed] [Google Scholar]
- 1102.Sharkey D.J., Tremellen K.P., Jasper M.J., Gemzell-Danielsson K., Robertson S.A. Seminal fluid induces leukocyte recruitment and cytokine and chemokine mRNA expression in the human cervix after coitus. J Immunol. 2012;188:2445–2454. doi: 10.4049/jimmunol.1102736. [DOI] [PubMed] [Google Scholar]
- 1103.Robertson S.A., Mau V.J., Hudson S.N., Tremellen K.P. Cytokine-leukocyte networks and the establishment of pregnancy. Am J Reprod Immunol. 1997;37:438–442. doi: 10.1111/j.1600-0897.1997.tb00257.x. [DOI] [PubMed] [Google Scholar]
- 1104.O’Bryan M.K., Baker H.W., Saunders J.R. Human seminal clusterin (SP-40,40). Isolation and characterization. J Clin Invest. 1990;85:1477–1486. doi: 10.1172/JCI114594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1105.Maccioni M., Riera C.M., Rivero V.E. Identification of rat prostatic steroid binding protein (PSBP) as an immunosuppressive factor. J Reprod Immunol. 2001;50:133–149. doi: 10.1016/s0165-0378(01)00060-2. [DOI] [PubMed] [Google Scholar]
- 1106.Shattock R.J., Moore J.P. Inhibiting sexual transmission of HIV-1 infection. Nat Rev Microbiol. 2003;1:25–34. doi: 10.1038/nrmicro729. [DOI] [PubMed] [Google Scholar]
- 1107.Brecher M.L. Treatment of acute lymphoid leukemia in children: current regimens and future prospects. N Y State J Med. 1986;86:188–196. [PubMed] [Google Scholar]
- 1108.Shevchuk M.M., Pigato J.B., Khalife G., Armenakas N.A., Fracchia J.A. Changing testicular histology in AIDS: its implication for sexual transmission of HIV. Urology. 1999;53:203–208. doi: 10.1016/s0090-4295(98)00463-4. [DOI] [PubMed] [Google Scholar]
- 1109.Costiniuk C.T., Kovacs C., Routy J.P. Human immunodeficiency virus rebound in blood and seminal plasma following discontinuation of antiretroviral therapy. AIDS Res Hum Retroviruses. 2013;29:266–269. doi: 10.1089/aid.2011.0343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1110.Shehu-Xhilaga M., de Kretser D., Dejucq-Rainsford N., Hedger M. Standing in the way of eradication: HIV-1 infection and treatment in the male genital tract. Curr HIV Res. 2005;3:345–359. doi: 10.2174/157016205774370375. [DOI] [PubMed] [Google Scholar]
- 1111.Jahnukainen K., Saari T., Salmi T.T., Pöllänen P., Pelliniemi L.J. Reactions of Leydig cells and blood vessels to lymphoblastic leukemia in the rat testis. Leukemia. 1995;9:908–914. [PubMed] [Google Scholar]
- 1112.Uyttenhove C., Godfraind C., Lethé B. The expression of mouse gene P1A in testis does not prevent safe induction of cytolytic T cells against a P1A-encoded tumor antigen. Int J Cancer. 1997;70:349–356. doi: 10.1002/(sici)1097-0215(19970127)70:3<349::aid-ijc17>3.0.co;2-a. [DOI] [PubMed] [Google Scholar]
- 1113.Giwercman A., Carlsen E., Keiding N., Skakkebaek N.E. Evidence for increasing incidence of abnormalities of the human testis: a review. Environ Health Perspect. 1993;101(Suppl. 2):65–71. doi: 10.1289/ehp.93101s265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1114.Fowler J.E., Jr., Kaiser D.L., Mariano M. Immunologic response of the prostate to bacteriuria and bacterial prostatitis. I. Immunoglobulin concentrations in prostatic fluid. J Urol. 1982;128:158–164. doi: 10.1016/s0022-5347(17)52810-4. [DOI] [PubMed] [Google Scholar]
- 1115.Fowler J.E., Jr., Mariano M. Immunoglobulin in seminal fluid of fertile, infertile, vasectomy and vasectomy reversal patients. J Urol. 1983;129:869–872. doi: 10.1016/s0022-5347(17)52403-9. [DOI] [PubMed] [Google Scholar]
- 1116.Stern J.E., Gardner S., Quirk D., Wira C.R. Secretory immune system of the male reproductive tract: effects of dihydrotestosterone and estradiol on IgA and secretory component levels. J Reprod Immunol. 1992;22:73–85. doi: 10.1016/0165-0378(92)90007-q. [DOI] [PubMed] [Google Scholar]
- 1117.Parr M.B., Parr E.L. Immunohistochemical localization of secretory component and immunoglobulin A in the urogenital tract of the male rodent. J Reprod Fertil. 1989;85:115–124. doi: 10.1530/jrf.0.0850115. [DOI] [PubMed] [Google Scholar]
- 1118.Billiau A. Interferon-γ in autoimmunity. Cytokine Growth Factor Rev. 1996;7:25–34. doi: 10.1016/1359-6101(96)00004-4. [DOI] [PubMed] [Google Scholar]
- 1119.Boman H.G. Peptide antibiotics and their role in innate immunity. Annu Rev Immunol. 1995;13:61–92. doi: 10.1146/annurev.iy.13.040195.000425. [DOI] [PubMed] [Google Scholar]
- 1120.Hancock R.E., Diamond G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000;8:402–410. doi: 10.1016/s0966-842x(00)01823-0. [DOI] [PubMed] [Google Scholar]
- 1121.Com E., Bourgeon F., Evrard B. Expression of antimicrobial defensins in the male reproductive tract of rats, mice, and humans. Biol Reprod. 2003;68:95–104. doi: 10.1095/biolreprod.102.005389. [DOI] [PubMed] [Google Scholar]
- 1122.Patil A.A., Cai Y., Sang Y., Blecha F., Zhang G. Cross-species analysis of the mammalian beta-defensin gene family: presence of syntenic gene clusters and preferential expression in the male reproductive tract. Physiol Genomics. 2005;23:5–17. doi: 10.1152/physiolgenomics.00104.2005. [DOI] [PubMed] [Google Scholar]
- 1123.Johnston D.S., Jelinsky S.A., Bang H.J. The mouse epididymal transcriptome: transcriptional profiling of segmental gene expression in the epididymis. Biol Reprod. 2005;73:404–413. doi: 10.1095/biolreprod.105.039719. [DOI] [PubMed] [Google Scholar]
- 1124.Zhou C.X., Zhang Y.L., Xiao L. An epididymis-specific β-defensin is important for the initiation of sperm maturation. Nat Cell Biol. 2004;6:458–464. doi: 10.1038/ncb1127. [DOI] [PubMed] [Google Scholar]
- 1125.Jalkanen J., Huhtaniemi I., Poutanen M. Discovery and characterization of new epididymis-specific beta-defensins in mice. Biochim Biophys Acta. 2005;1730:22–30. doi: 10.1016/j.bbaexp.2005.05.010. [DOI] [PubMed] [Google Scholar]
- 1126.Hall S.H., Yenugu S., Radhakrishnan Y., Avellar M.C., Petrusz P., French F.S. Characterization and functions of beta defensins in the epididymis. Asian J Androl. 2007;9:453–462. doi: 10.1111/j.1745-7262.2007.00298.x. [DOI] [PubMed] [Google Scholar]
- 1127.Zhao Y., Diao H., Ni Z., Hu S., Yu H., Zhang Y. The epididymis-specific antimicrobial peptide β-defensin 15 is required for sperm motility and male fertility in the rat (Rattus norvegicus) Cell Mol Life Sci. 2010;68:697–708. doi: 10.1007/s00018-010-0478-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1128.Gupta S.K. Status of immunodiagnosis and immunocontraceptive vaccines in India. Adv Biochem Eng Biotechnol. 2003;85:181–214. doi: 10.1007/3-540-36466-8_6. [DOI] [PubMed] [Google Scholar]
- 1129.Diekman A.B., Herr J.C. Sperm antigens and their use in the development of an immunocontraceptive. Am J Reprod Immunol. 1997;37:111–117. doi: 10.1111/j.1600-0897.1997.tb00199.x. [DOI] [PubMed] [Google Scholar]
- 1130.O’Hern P.A., Bambra C.S., Isahakia M., Goldberg E. Reversible contraception in female baboons immunized with a synthetic epitope of sperm-specific lactate dehydrogenase. Biol Reprod. 1995;52:331–339. doi: 10.1095/biolreprod52.2.331. [DOI] [PubMed] [Google Scholar]
- 1131.Naz R.K. Development of genetically engineered human sperm immunocontraceptives. J Reprod Immunol. 2009;83:145–150. doi: 10.1016/j.jri.2009.06.260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1132.Rovira P., Mascarell L., Truffa-Bachi P. The impact of immunosuppressive drugs on the analysis of T cell activation. Curr Med Chem. 2000;7:673–692. doi: 10.2174/0929867003374778. [DOI] [PubMed] [Google Scholar]
- 1133.Liu J. FK506 and cyclosporin, molecular probes for studying intracellular signal transduction. Immunol Today. 1993;14:290–295. doi: 10.1016/0167-5699(93)90048-P. [DOI] [PubMed] [Google Scholar]
- 1134.Pulla B., Barri Y.M., Anaissie E. Acute renal failure following bone marrow transplantation. Ren Fail. 1998;20:421–435. doi: 10.3109/08860229809045131. [DOI] [PubMed] [Google Scholar]
- 1135.Ryffel B. The carcinogenicity of ciclosporin. Toxicology. 1992;73:1–22. doi: 10.1016/0300-483x(92)90166-c. [DOI] [PubMed] [Google Scholar]
- 1136.Caneguim B.H., Cerri P.S., Spolidório L.C., Miraglia S.M., Sasso-Cerri E. Structural alterations in the seminiferous tubules of rats treated with immunosuppressor tacrolimus. Reprod Biol Endocrinol. 2009;7:19. doi: 10.1186/1477-7827-7-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1137.Ben Ali R., Klouz A., Boubaker S., Lakhal M., Belkahia C. An animal model of testicular toxicity by cyclosporine: evaluation and protection. Fundam Clin Pharmacol. 2009;23:241–246. doi: 10.1111/j.1472-8206.2009.00680.x. [DOI] [PubMed] [Google Scholar]
- 1138.Kaur G., Long C.R., Dufour J.M. Genetically engineered immune privileged Sertoli cells: a new road to cell based gene therapy. Spermatogenesis. 2012;2:23–31. doi: 10.4161/spmg.19119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 1139.Penna G., Mondaini N., Amuchastegui S. Seminal plasma cytokines and chemokines in prostate inflammation: interleukin 8 as a predictive biomarker in chronic prostatitis/chronic pelvic pain syndrome and benign prostatic hyperplasia. Eur Urol. 2007;51:524–533. doi: 10.1016/j.eururo.2006.07.016. [DOI] [PubMed] [Google Scholar]
- 1140.Malan T.P., Jr., Porreca F. Lipid mediators regulating pain sensitivity. Prostagland Lipid Mediat. 2005;77:123–130. doi: 10.1016/j.prostaglandins.2004.09.008. [DOI] [PubMed] [Google Scholar]
- 1141.Motrich R.D., van Etten E., Baeke F., Riera C.M., Mathieu C., Rivero V.E. Crucial role of interferon-γ in experimental autoimmune prostatitis. J Urol. 2010;183:1213–1220. doi: 10.1016/j.juro.2009.11.008. [DOI] [PubMed] [Google Scholar]
- 1142.Povey A.C., Stocks S.J. Epidemiology and trends in male subfertility. Hum Fertil. 2010;13:182–188. doi: 10.3109/14647273.2010.515700. [DOI] [PubMed] [Google Scholar]