

Introduction
A successful immune response is critically necessary to eradicate infectious challenges and prevent dissemination of the infection in the host. However, if inflammation is not limited and becomes generalized, it can result in the constellation of signs and symptoms of a systemic inflammatory response syndrome (SIRS). If the infection is not contained, the spread of the pathogen from its local origin through the blood may result in systemic endothelial activation and precipitate sepsis, severe sepsis, and septic shock. Progression of sepsis to shock may lead to multiple organ dysfunction syndrome (MODS) and ultimately death.
Host immunity is divided into innate and adaptive immune systems for purposes of discussion and teaching but there is a great deal of interaction between the two systems. Innate immunity is rapid, largely nonspecific, and composed of barriers, phagocytic cells, the complement system, and other soluble components of inflammation. After breech of a barrier, cellular elements of the innate immune response are the first line of defense against the development and progression of infection. Adaptive immunity, which is antigen specific, is long lived, and often takes several days to develop, provides immunologic specificity and memory. These systems work together to protect the host from pathogenic challenge but may also precipitate host injury through aberrant responses. The outcome of infection is dependent on at least four major factors: (1) the pathogen, (2) the pathogen load, (3) the site of infection, and (4) the host response. Less is known about the host response in neonates compared with adults for a number of reasons, the principal one being a highly variable definition of disease.
Our understanding of the pathophysiology of sepsis is largely from investigations in adult populations, including both humans and animals. There is clear evidence from both preclinical models of sepsis and humans that neonates manifest different host immune responses as compared with adults.1, 2, 3, 4 Even in comparison with children, neonates manifest a unique host immune response to septic shock.5 Thus neonatal-specific clinical investigations, particularly in very preterm infants, are required to improve both survival and long-term outcomes for these populations. A better understanding of the pathophysiology will uncover new opportunities for interventional studies ultimately aimed at improving outcomes. To this end, in this chapter we explore the pathophysiology of sepsis in the neonate, with special attention paid to the immunobiology of sepsis.
Definition of Sepsis
Adult and pediatric intensivists currently use generally accepted definitions for sepsis for goal-based therapeutic interventions.6, 7, 8, 9 These definitions are critical to facilitate epidemiologic studies, to accurately determine disease prevalence, to select patients for clinical trials, and ultimately to improve the delivery of care. The generally accepted pediatric definition for sepsis, established in 2005, was intended for all children (<18 years old), including term neonates (≥37 weeks' completed gestation).6 Preterm neonates (<37 weeks' completed gestation) were specifically excluded from the pediatric generally accepted definitions, and neonatal-perinatal subspecialists were not represented among the pediatric consensus experts. To investigate whether the pediatric generally accepted definitions for SIRS and sepsis applied to term infants, Hofer and colleagues10 retrospectively examined 476 term neonates and found that the generally accepted definitions applied to only 53% of cases of culture-positive early-onset sepsis. Neonatal sepsis has been inconsistently defined on the basis of a variety of clinical and laboratory criteria, which makes the study of this condition very difficult.11 Diagnostic challenges and uncertain disease epidemiology necessarily result from a variable definition of disease. The lack of a generally accepted definition for neonatal sepsis remains a significant hindrance towards improving outcomes and accurately describing disease pathophysiology. Thus working definitions for the sepsis continuum, specific for preterm and term neonates, are needed to provide a uniform basis for clinicians and researchers to study and diagnose severe sepsis. The addition of immune biomarker–based staging of disease to clinical sign staging is highly likely to increase the accuracy of patient classification for future multicenter clinical trials that will test novel interventions.
Epidemiology and Risk Factors for Development of Neonatal Sepsis
Sepsis or serious infection within the first 4 weeks of life kills more than 1 million newborns globally every year.12, 13 The incidence of neonatal sepsis is variable (from less than 1% to more than 35% of live births) on the basis of gestational age and time of onset (early-onset sepsis [<72 hours after birth] or late-onset sepsis [≥72 hours after birth]).14, 15, 16, 17, 18, 19, 20 Preterm neonates have the greatest sepsis incidence and mortality rates among all age-groups21, 22, 23, 24, 25, 26 (Figure 152-1 ).
Figure 152-1.
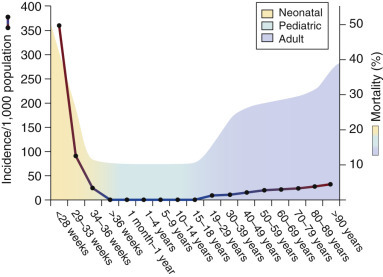
Sepsis incidence and mortality in humans across developmental age-groups.
Risk factors for developing sepsis in neonates, particularly the very premature, have been well described.14, 15, 27, 28, 29, 30, 31, 32, 33 Prematurity, low birth weight (especially infants weighing less than 1,000 g), male sex, a maternal vaginal culture positive for group B streptococcus (GBS), prolonged rupture of membranes, maternal intrapartum fever, and chorioamnionitis are strongly associated with an increased risk for early-onset sepsis.30 Chorioamnionitis is associated with the greatest risk for subsequent clinical or culture-proven sepsis.29 Recent studies demonstrate the risk for sepsis in newborn infants born to women with clinical chorioamnionitis is strongly dependent on gestational age, with minimal risk in neonates aged 35 weeks or older and greater risk with increasing degrees of prematurity.34, 35, 36, 37, 38, 39, 40, 41 The risk for neonatal sepsis conferred by maternal GBS colonization29 is significantly reduced with adequate intrapartum antibiotic prophylaxis.42 Despite the efficacy of this intervention, the incidence of invasive GBS disease in African American neonates is still more than twice that in white babies,43 and the incidence of Escherichia coli sepsis may be rising in very-low-birth-weight (VLBW) neonates.44 Vaginal delivery in the presence of maternal active primary herpes simplex virus significantly increases the risk for a neonatal herpes simplex virus infection, which has a fulminant course and high mortality.45, 46, 47 Preexisting maternal immunodeficiency or sepsis also increases the risk for sepsis in the neonate.48 In addition, care practices after birth, such as intubation, mechanical ventilation, and placement of central venous lines, increase the risk for the development of sepsis.49
Microbiology of Sepsis in Neonates
A number of pathogens have been associated with sepsis in the neonatal period. The predominant cause is bacterial; however, certain viral infections are associated with a fulminant course and significant mortality.50, 51, 52 In a large (n = 104,676), multicenter study of VLBW infants (<1500 g), gram-positive organisms accounted for 34% of pathogens causing early-onset sepsis and 61% of those causing late-onset sepsis. In contrast, gram-negative organisms were responsible for 58% of early-onset sepsis and 26% of late-onset sepsis.53 Candida species accounted for 3% of cases of early-onset sepsis and 11% of cases of late-onset sepsis. Infection by gram-negative organisms, particularly Pseudomonas species, carries a higher risk for fulminant course and death than infection by other pathogen groups.14, 15, 49, 54, 55, 56 Gram-positive causes of sepsis are dominated by GBS and coagulase-negative staphylococci (CoNS).15, 57 Although the high mortality rate for GBS has been well described (especially among infants born prematurely), mortality rates associated with CoNS are significantly lower.15, 16 Fungi may also be associated with fulminant neonatal sepsis and predominantly affect VLBW infants.15, 58, 59 Independent predictors of in-hospital neonatal mortality after late-onset sepsis were Pseudomonas infection (adjusted odds ratio [OR], 14.31; 95% confidence interval [CI], 3.87% to 53.0%) and fungemia (OR, 5.69; 95% CI, 2.48% to 13.01%).60 The limited sensitivity of current methods to identify causative organisms is partially due to an inability to take a large sample of blood from newborn infants with suspected sepsis.61 Blood culture–negative (“clinical”) sepsis is estimated to occur at a nearly 10-fold greater rate than blood culture–positive sepsis.62 In some of these infants, sepsis may also be due to novel viral pathogens associated with sepsis-like syndromes (e.g., echovirus, enterovirus, parechovirus, Coxsackie virus, adenovirus, parainfluenza virus, rhinovirus, and coronavirus).50, 63, 64, 65
The Role of Barrier Defenses in Neonatal Sepsis
Physical barriers, including skin and mucosal surfaces, are the first point of contact between the host and potential pathogens. Thus a successful immune defense in addition to epithelial barrier function is critical to prevent the development of local infection. Multiple immune elements are present to prevent attachment and propagation of pathogens while simultaneously permitting the presence of commensal organisms required for homeostasis. Vernix enhances skin barrier function in late-preterm and term neonates. Vernix is a complex material comprising water (80.5%), lipids (10.3%), and proteins (9.1%) produced by fetal sebaceous glands during the last trimester66 and is largely absent in preterm neonates born before 28 weeks' gestation. Vernix provides a barrier to water loss, improves temperature control, and serves as a shield containing antioxidants and innate immune factors such as antimicrobial proteins and peptides (APPs).67 The APPs on the surface of the newborn's skin (and replete in the amniotic fluid68, 69, 70) are capable of killing/inactivating common neonatal pathogens, including GBS, E. coli, and Candida species.71 Erythema toxicum is an immune-mediated manifestation that results from bacterial colonization of the skin occurring shortly after birth.72, 73 This common cutaneous immune response is less common in preterm infants than in term infants, highlighting the impact of developmental age on host immune capabilities.74 In contrast to the moist mucosal surfaces of the respiratory and gastrointestinal (GI) tracts, the skin is arid, which further reduces the chances for microbial invasion.
The outermost layer of the skin, the stratum corneum, prevents microbial invasion, maintains temperature, and reduces the risk for dehydration through prevention of transcutaneous water loss.75 The immature and incompletely developed stratum corneum of preterm newborns takes at least 1 to 2 weeks after birth to become fully functional76 and may take up to 8 weeks in the extremely preterm neonate, significantly increasing the risk for barrier dysfunction.77 Disruption of the cutaneous barrier by trauma (e.g., placement of an intravenous catheter or heel stick) or chemical burn allows microorganisms to enter the subcutaneous tissue, increasing the likelihood of their establishing a local infection (Figure 152-2 ). The likelihood of a microbial breach of the cutaneous barrier rises in the presence of intravenous catheters, which are essential for critical care. Emollients, aimed at enhancing the barrier function of preterm newborn skin, increase the risk for nosocomial infection and their use is not recommended.78
Figure 152-2.
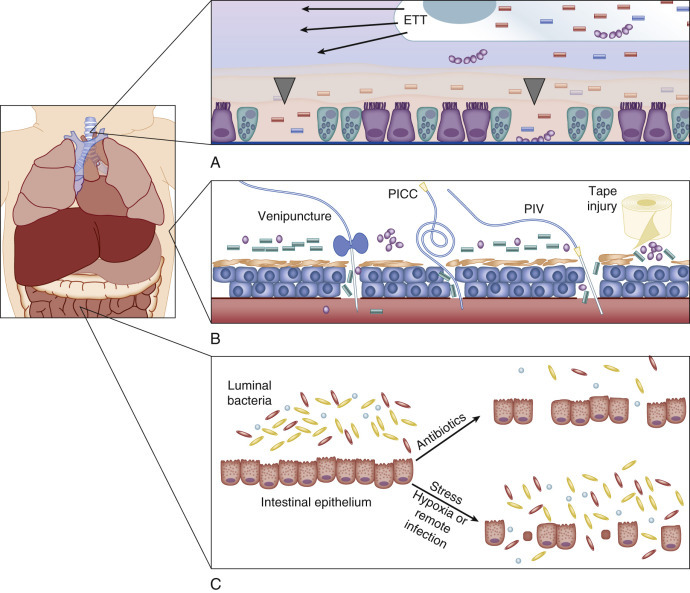
Physical barriers.
A, Respiratory mucosa. A foreign body (an endotracheal tube, ETT) and/or positive pressure can irritate and injure the respiratory epithelium (ciliated cells; gray arrowheads denote denuded areas). Increased numbers of goblet cells (blue cells with inclusions) with decreased mucociliary clearance of the airway further increase the likelihood of infection (bacteria represented by purple spherical chains and blue/pink rods). B, Skin. Disruptions associated with trauma (venipuncture or heel stick), a peripherally inserted central catheter (PICC), a peripheral intravenous line (PIV), or tape-related abrasions compromise the skin barrier (bacteria represented by clusters of purple spheres and green rods). C, Gastrointestinal mucosa. Luminal bacteria (microbiota) are a valuable component of the mucosal barrier. The interaction between intestinal bacteria and intestinal epithelium is necessary for homeostasis and normal function of repair mechanisms. Disruption of this interaction, through the use of antibiotics or via stress to the organism (e.g., hypoxia or remote infection such as sepsis or pneumonia), results in loss of homeostasis and degradation of the intestinal boundaries with subsequent microbial translocation.
Mucosal barriers contain multiple components that serve to prevent infection, including acidic pH, mucus, cilia, proteolytic enzymes, APPs, opsonins such as surfactant proteins, sentinel immune cells such as macrophages, dendritic cells, polymorphonuclear neutrophils (PMNs), and T cells, as well as commensal organisms.79 Like the skin, the GI mucosa is quickly colonized after birth and contains a significant repository of microorganisms.80, 81, 82 GI barrier integrity, paramount for prevention of spread of microorganisms out of the intestinal compartment, is dependent on the interaction between commensal organisms and host epithelium. Interleukin (IL)-17, produced by type 3 intestinal innate lymphoid cells in the presence of the microbiota, drives granulocytosis and may protect the neonatal host from infectious challenge.83 A loss of intestinal barrier integrity likely plays a role in the development of necrotizing enterocolitis (NEC) and late-onset sepsis.84, 85 Prolonged antibiotic treatment, hypoxia, and remote infection are factors known to disrupt or injure the neonatal intestinal barrier (see Figure 152-2).86, 87, 88 Under these circumstances, the gut may become the motor of systemic inflammation.89 Mechanistically, Paneth cells and intestinal lymphoid cells may release excessive amounts of IL-17, which, in turn, plays a critical role in the development of SIRS.90 Many interventions aimed at reducing the frequency of sepsis in neonates via enhancement of mucosal barrier integrity have been evaluated. Neither probiotics nor glutamine supplementation has reduced the incidence of neonatal sepsis.91 In contrast, human milk feeding is associated with a reduction in the risk for sepsis92 and NEC93, 94 and is strongly encouraged, especially in preterm infants.
Respiratory mucosa is defended in utero by amniotic fluid and pulmonary APPs, surfactant proteins A and D, alveolar macrophages, and PMNs, among other immune elements. The surface and submucosal gland epithelium of the conducting airways is a constitutive primary participant in innate immunity through the production of mucus and mucociliary clearance of pathogens and debris.95 Premature neonates have relatively more goblet cells than do maturer neonates, leading to a decrease in mucociliary clearance. Respiratory mucosal function can be impaired by surfactant and saliva deficiency, altered mucus production, and mechanical ventilation. Ventilation is associated with decreased mucociliary clearance, airway irritation, and parenchymal lung injury (see Figure 152-2). Intubation is also associated with the progressive accumulation of colonizing bacteria and bacterial endotoxin in respiratory fluids, with concomitant mobilization of endotoxin-modulating APPs to the airway.96 Neonates with surfactant deficiency lack APPs such as surfactant proteins A and D, which are also absent in commercially available surfactant preparations.97 There is an age-dependent maturation in the ability of respiratory epithelium to elaborate APPs (cathelicidin and β-defensins), such that the respiratory epithelium of preterm newborns mounts a deficient APP response.98 These deficiencies as well as those related to cellular function in combination with invasive procedures lead to a reduction in respiratory barrier function that increases the risk for sepsis.
Molecular Events During Early Infection
Pathogen Recognition
Once the local barrier function has been compromised, pathogen recognition by local immune sentinel cells is the first step towards the development of an immune response (Figure 152-3 ). Elegant sensing mechanisms have evolved to facilitate detection of potentially pathogenic microorganisms. Multiple classes of pathogen recognition receptors (PRRs) have been discovered that serve as detectors of pathogen-associated molecular patterns (PAMPs), including cell wall and membrane components, flagellum, nucleic acids, and carbohydrates.99 A litany of PRR classes have been discovered, including the Toll-like receptors (TLRs), NOD-like receptors (NLRs), retinoic acid–inducible protein I like receptors (RLRs), peptidoglycan recognition proteins, β2-integrins, and C-type lectin receptors. The TLRs, β2-integrins, and C-type lectin receptors detect pathogens both on the cell surface and in endosomes, whereas RLRs and NLRs detect pathogens only intracellularly. The discovery that TLR4 was integral for a robust lipopolysaccharide (LPS)-mediated inflammatory response after gram-negative sepsis may be why TLRs have been more thoroughly investigated in the setting of sepsis than other PRRs.100 Each of the 10 known TLRs in humans, present on and within multiple cell types, recognizes extracellular and intracellular pathogens via specific PAMPs.101, 102 Multiple TLRs may be activated in concert by intact or partial microorganisms and in turn activate multiple second-messenger pathways simultaneously.102, 103
Figure 152-3.
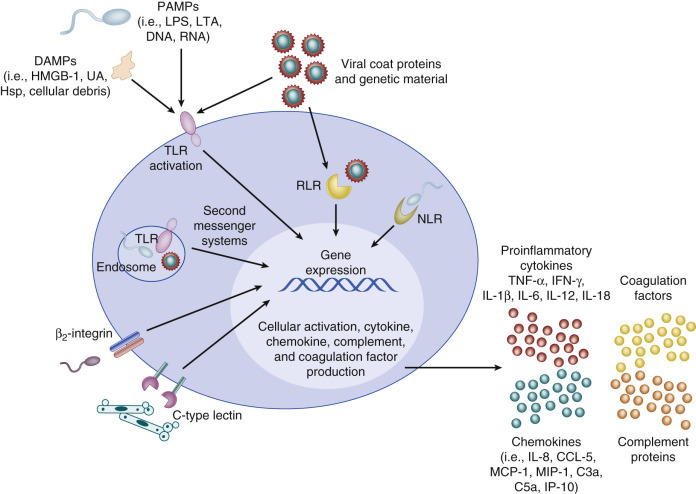
Activation of sentinel immune cells.
Sentinel cells (e.g., monocytes, macrophages) sense pathogens via pathogen-associated molecular patterns (PAMPs) or damage associated molecular patterns (DAMPs) binding to pathogen recognition receptors. Pathogen recognition receptors include Toll-like receptors, retinoic acid–inducible protein I like receptors, NOD-like receptors, C-type lectin receptors, and β2-integrins. PAMPs include lipopolysaccharide (LPS), lipoteichoic acid (LTA), DNA, and RNA. DAMPs can also be sensed through Toll-like receptors and include uric acid (UA), heat shock proteins (Hsp), and high-mobility group box 1 (HMGB-1). Signaling occurs through a series of second messengers and results in transcription and translation of cytokines and chemokines that amplify the immune response. IFN, Interferon; IL, interleukin; MCP, monocyte chemoattractant protein; MIP, macrophage inflammatory protein; NLR, NOD-like receptors; RLR, retinoic acid–inducible protein I like receptor; TNF, tumor necrosis factor; TLR, Toll-like receptor.
(From Wynn JL, Wong HR: Pathophysiology and treatment of septic shock in neonates. Clin Perinatol 37(2):439–479, 2010.)
LPS is the prototypic mediator of systemic inflammation and generates many of the clinical findings of sepsis and septic shock, including MODS and death.104 LPS signals through TLR4 in conjunction with the adaptor proteins CD14 and myeloid differentiation factor 2.105 In adults a reduction in mortality and improvement in hemodynamics were demonstrated when the level of serum LPS was reduced.106 The level of LPS is elevated in blood from infected neonates and those with NEC even in the absence of gram-negative bacteremia.84 High levels of circulating endotoxin found during sepsis and NEC are associated with multiorgan failure, thrombocytopenia, neutropenia, and death.84 Administration of anti-LPS antibodies to a small number of neonates with sepsis (n = 16) with serum endotoxin present reduced the time to recovery but not mortality as compared with the values in placebo-treated neonates.107 Reduction of serum LPS levels by exchange transfusion in infected neonates (n = 10) was associated with improved survival.108
Bacterial cell wall components (such as lipoteichoic acid) signal primarily through TLR1, TLR2, and TLR6, flagellin signals through TLR5, and CpG double-stranded DNA signals through TLR9. Common viral PAMPs such as double-stranded RNA or single-stranded RNA signal through TLR3, and TLR7 and TLR8, respectively. Agonist-TLR binding results in a signaling cascade of intracellular second-messenger proteins ultimately leading to production of cytokines and chemokines, as well as activation of other antimicrobial effector mechanisms.101 Signaling through TLRs typically leads to the production of nuclear factor κB (NF-κB)-dependent inflammatory cytokines and chemokines, whereas signaling through Toll/IL-1 receptor–domain-containing adapter inducing interferon (IFN)-β (TRIF) induces production of type I IFNs, as well as NF-κB-related inflammatory cytokines. In neonates of all gestational ages, up-regulation of TLR2 and TLR4 messenger RNA (mRNA) occurs during gram-positive and gram-negative infection, respectively.109 Dysregulation or overexpression of TLR4 is involved in the development of NEC in experimental animal models,110 implicating the importance of TLRs in the initial immune response to pathogens and their role in neonatal sepsis.
NOD-Like Receptors, Retinoic Acid–Inducible Protein I-Like Receptors, C-Type Lectin Receptors, and β2-Integrins
Other important intracellular PRRs include NLRs and RLRs. For NLRs, multiple cytosolic proteins are able to act as PAMP sensors (e.g., NLRP1, NLRP3, and NLRC4) and coalesce with adaptor proteins and procaspase 1 to form a multimeric protein complex termed the inflammasome. 111 The formation of the inflammasome results in the conversion of procaspase to active caspase 1, which cleaves the inactive precursor proteins IL-1β and IL-18 to their active forms.111 RLRs are cytoplasmic RNA helicases that, like TLR3, sense double-stranded RNA of viral origin and induce type I IFN production and NF-κB activation.102 To date, the impact of RLR and NLR signaling has not been specifically examined in neonates with sepsis.
In addition to its roles in leukocyte function (adhesion, phagocytosis, migration, and activation) and complement binding, complement receptor 3 (CR3, also known as MAC-1 and CD11b-CD18) functions as a pathogen sensor on the surface of phagocytes. CR3 binds LPS, as well as a broad range of other microbial products, in cooperation with or independently of CD14, leading to up-regulation of inducible nitric oxide (NO) synthase and NO production.112 Diminished expression of L-selectin and CR3 on stimulated neonatal PMNs impairs activation and accumulation at sites of inflammation.99, 113, 114 Decreased expression of L-selectin and CR3 persists for at least the first month of life in term infants, possibly contributing to an increased risk for infection.115 The expression of CR3 (CD11b) may be reduced further in preterm neonates as compared with term neonates.116 In umbilical cord blood from neonates of less than 30 weeks' gestation, PMN CR3 content was similar to levels found in patients with type 1 leukocyte adhesion deficiency (failure to express CD18).113, 114 Thus decreased leukocyte CR3 surface expression increases the likelihood of suboptimal pathogen detection and cellular activation, particularly in the preterm neonate.
C-type lectin receptors are PRRs that recognize bacterial, viral, fungal, and parasitic carbohydrate moieties. C-type lectin receptors may be expressed on the cell surface (e.g., macrophage mannose receptor, mincle receptor, dectin 1, and dectin 2) or secreted as soluble proteins (e.g., mannose-binding lectin [MBL], (which is also named mannan-binding protein or mannan-binding lectin) as one of the acute-phase reactants. Once bound to its carbohydrate ligand, MBL initiates activation of complement via the lectin pathway to promote opsonization and phagocytic clearance of pathogens. Plasma MBL concentrations are low at birth (especially in preterm infants) but rise steadily throughout infancy and childhood.117 Low levels of MBL are associated with the increased incidence of sepsis in neonates.118, 119, 120 In addition to decreased concentrations at birth, certain genetic polymorphisms of MBL (namely, MBL2), have been associated with an increased risk for infection in some,121 but not all, studies.122, 123, 124 M-ficolin activates the complement system in a manner similar to MBL and its level is elevated in neonates with sepsis.125
The Role of Inflammation
PRR stimulation results in rapid inflammatory mediator transcription and translation directed at cellular activation and clearance of pathogenic organisms126 (see Figure 152-3). During sepsis and septic shock, multiple proinflammatory cytokines have been identified, including IL-1β, IL-6, IL-8 (CXCL8), IL-12, IL-18, IFN-γ, and tumor necrosis factor (TNF)-α.127 Compared with adults with sepsis, neonates with sepsis produce less IL-1β, TNF-α, IFN-γ, and IL-12.128, 129, 130, 131, 132, 133 The decreased cytokine production is due in part to decreased production of important intracellular mediators of TLR signaling, including myeloid differentiation factor 88, IFN regulatory factor 5, and p38, which exhibit gestational age–specific decrements.134 Recent studies have demonstrated impaired inflammasome activation and mature IL-1β production by neonatal mononuclear cells.121, 135 In a comprehensive study (>140 analytes) of serum from neonates evaluated for late-onset sepsis, IL-18 emerged as a predictive biomarker to differentiate infected neonates from uninfected neonates.136 IL-18 reduces PMN apoptosis,137 drives IFN-γ production,138 and induces production of TNF-α, IL-1β, and CXCL8.139 IL-18 primes PMNs for degranulation with production of reactive oxygen intermediates on subsequent stimulation.140 Dysregulation of many of these functions linked to IL-18 are seen in sepsis and septic shock. Increased IL-18 levels have been demonstrated in premature neonates with brain injury141 and also in an experimental model of NEC,142, 143, 144 highlighting activation pathways common with those in ischemia and inflammation. Excessive levels of IL-1β, TNF-α, IL-6, CXCL8, IL-10, and IL-18, such as those seen with advanced-stage NEC, severe sepsis, or septic shock, correlate with poor survival.84, 145, 146, 147, 148 Altered cytokine levels (increased IL-10 and IL-6 levels and decreased CCL5 levels) may identify those neonates at highest risk for the development of sepsis-associated disseminated intravascular coagulation (DIC).149
Proinflammatory cytokine production leads to activation of endothelial cells, including increased expression of cell adhesion molecules that facilitate leukocyte recruitment and diapedesis (Figure 152-4 ). Up-regulation of cell adhesion molecules (soluble intercellular adhesion molecule, vascular cell adhesion molecule, L-selectin, P-selectin, E-selectins, and CD11b-CD18) during sepsis facilitates rolling and extravascular migration of leukocytes.150, 151, 152, 153 Decreased production of L-selectin and expression of C3 in PMNs and monocytes derived from neonates may impair accumulation at sites of inflammation.113, 114
Figure 152-4.
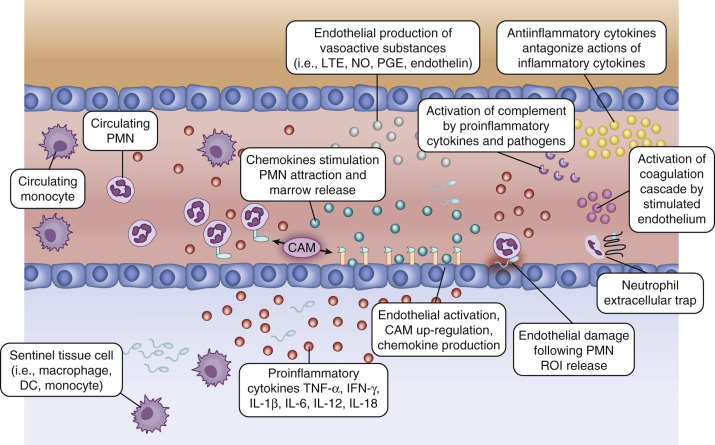
Cellular recruitment and endothelial activation following pathogen detection.
Pathogen-stimulated tissue/blood monocytes, dendritic cells, and macrophages release proinflammatory cytokines that activate the surrounding endothelium. Endothelial activation results in up-regulation of cell adhesion molecules, production of chemokines and vasoactive substances, activation of complement, and development of a procoagulant state. Recruitment of polymorphonuclear neutrophils (PMNs) occurs along the chemokine gradient surrounding the area of inflammation. Antiinflammatory cytokines counter the actions of proinflammatory cytokines to prevent excessive cellular activation and recruitment that can result in tissue damage and systemic inflammation. Endothelium can be damaged when PMNs release reactive oxygen intermediates or from neutrophil extracellular traps. CAM, Cell adhesion molecule; DC, dendritic cell; IFN, Interferon; IL, interleukin; LTE, leukotriene; NO, nitric oxide; PGE, prostaglandin E; ROI, reactive oxygen intermediate; TNF, tumor necrosis factor.
(From Wynn JL, Wong HR: Pathophysiology and treatment of septic shock in neonates. Clin Perinatol 37(2):439–479, 2010.)
Chemokine gradients produced by endothelial cells and local macrophages are necessary for effective and specific leukocyte attraction and accumulation (see Figure 152-4). Without adequate leukocyte recruitment, there is increased risk for propagation from a localized to a systemic infection. Although poor cellular chemotaxis in the neonate has been observed, it is not likely a result of reduced serum concentrations of chemokines as baseline levels are similar in preterm and term neonates as compared with adults.154 Suboptimal cellular chemotaxis may be related to other mechanisms, such as poor complement receptor up-regulation after stimulation,99 deficiencies in another downstream signaling process,155 or inhibition by bacterial products.156 The levels of a wide variety of chemokines are increased during sepsis, including CXCL10 (IP-10), CCL5 (RANTES), CCL2 (monocyte chemoattractant protein 1), CCL3 (macrophage inflammatory protein 1α), and CXCL8.157 The levels of other chemoattractive molecules also increase with sepsis, including complement proteins C3a and C5a, APPs, including cathelicidins and defensins, and components of invading bacteria themselves.127, 136 The importance of chemoattractive substances in the pathogenesis of severe sepsis is highlighted by studies showing that CXCL8 can be used as a stratifying factor for survival in children,158 and C5a is implicated in sepsis-associated organ dysfunction in adults.104 Chemokine investigations in infected neonates revealed that CXCL10 is a sensitive early marker of infection,157 and low CCL5 levels may predict development of DIC.149
Damage-associated molecular patterns (or alarmins), such as intracellular proteins or mediators released by dying or damaged cells, may also active PRRs. For example, the damage-associated molecular pattern high-mobility group box 1 (HMGB-1) is involved in the progression of sepsis to septic shock in adults.104, 159 Macrophages or endothelial cells stimulated with LPS or TNF-α produce HMGB-1, which signals through TLR2, TLR4, and receptor for advanced glycation end products (RAGE).160 HMGB-1 results in cytokine production, activation of coagulation, and PMN recruitment.159, 161 HMGB-1 mediates disruption of epithelial junctions within the gut via the induction of reactive nitrogen intermediates, leading to increased bacterial translocation.162 The role of HMGB-1 and RAGE signaling in human neonates with sepsis has not been well characterized but has been shown to be involved in the pathophysiology of NEC in an experimental model.163 Significantly lower soluble RAGE levels were found in human fetuses that mounted robust inflammatory responses and HMGB-1 levels correlated significantly with the levels of IL-6 and S100β calcium-binding protein in the fetal circulation.164
Other specific damage-associated molecular patterns, including heat shock proteins and uric acid, may also stimulate TLRs, regulate PMN function, and serve as immune adjuvants. Heat shock protein production in infected neonates has not been evaluated but polymorphisms in heat shock proteins increase the risk for acute renal failure in preterm neonates.165 The levels of heat shock proteins are significantly elevated in infected adults and children.166 Elevated heat shock protein 60 and heat shock protein 70 level measured within 24 hours of pediatric intensive care unit admission were associated with septic shock and there was a strong trend towards increased mortality.167, 168 Uric acid can increase cytokine production, PMN recruitment, and dendritic cell stimulation169 and may also serve as an antioxidant.170 The level of uric acid is reduced in the serum of neonates with sepsis as compared with control neonates.171
In addition to facilitating leukocyte attraction, proinflammatory stimuli result in production of vasoactive substances that decrease or increase vascular tone and alter vascular permeability (see Figure 152-4). These include platelet-activating factor, thromboxane, leukotrienes, NO, histamine, bradykinin, and prostaglandins.172, 173 These substances are produced predominantly by host endothelium and mast cells. Activated PMNs produce phospholipase A2 (PLA2), the level of which is increased in the serum of neonates with sepsis174 and leads to generation of vasoactive substances, including prostaglandins and leukotriene. Thromboxane produced by activated platelets and endothelin 1 produced by activated endothelium175 are potent vasoconstrictors that participate in the development of pulmonary hypertension.176, 177, 178, 179 Overproduction of cytokines and vasoactive substances is associated with circulatory alterations and organ failure seen in severe sepsis and septic shock (Figure 152-5 ).6, 180, 181, 182, 183
Figure 152-5.
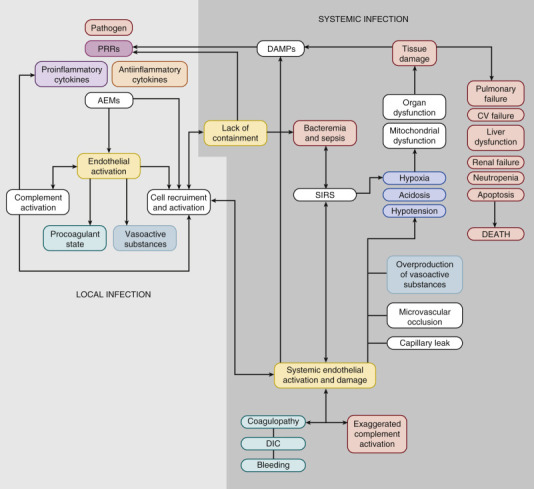
Pathophysiology of neonatal sepsis and septic shock. AEMs, Antimicrobial effector mechanisms; CV, cardiovascular; DAMPs, damage-associated molecular patterns; DIC, disseminated intravascular coagulation; PRRs, pattern recognition receptors; SIRS, systemic inflammatory response syndrome.
(From Wynn JL, Wong HR: Pathophysiology and treatment of septic shock in neonates. Clin Perinatol 37(2):439–479, 2010.)
The Antiinflammatory Response
If the pathogen is not contained locally and inflammatory homeostasis is not restored, SIRS may develop, and lead to MODS and death (see Figure 152-5).184 The traditional paradigm for understanding the host response to sepsis consists of an intense proinflammatory response, or SIRS, temporally followed by a compensatory antiinflammatory response syndrome. This paradigm has been challenged by the failure of multiple antiinflammatory strategies to improve sepsis outcomes in adults.185 New data in adults and children demonstrate simultaneous proinflammatory/antiinflammatory responses where the magnitude of either response may determine outcome.186, 187 Near simultaneous increases in antiinflammatory cytokine production (transforming growth factor β, IL-4, IL-10, IL-11, and IL-13) occur in neonates during infection, countering the actions of proinflammatory cytokines.127, 188, 189 These mediators blunt the activation and recruitment of phagocytic cells, reduce fever, modify coagulation factor expression, and decrease production of reactive oxygen and nitrogen intermediates, NO, and other vasoactive mediators.190, 191, 192, 193, 194, 195
Soluble cytokine and receptor antagonists produced during sepsis also modulate proinflammatory mediator action. Elevation of the levels of TNF receptor 2 (which regulates the concentration of TNF-α), soluble IL-6 receptor, soluble IL-2, and IL-1 receptor antagonist have been documented in neonatal sepsis with resolution after effective treatment.189, 196, 197 The role of these regulatory cytokine inhibitors in the immune response to neonatal sepsis and septic shock has been incompletely characterized. Soluble RAGE competes with cell-bound RAGE for the binding of HMGB-1 and other RAGE ligands.198 Soluble RAGE has antiinflammatory effects and its level is elevated in adults during sepsis.199 Furthermore, soluble RAGE improved survival and reduced inflammation when given to infected adult rodents.200 Serum soluble triggering receptor expressed on myeloid cells 1 may reduce inflammatory signaling for triggering receptor expressed on myeloid cells 1, and predict mortality in preterm neonates.201
MicroRNAs may regulate inflammation at the level of gene expression via several putative mechanisms.202 Several pilot studies in rodents and humans have demonstrated regulatory functions for microRNA in neonates.203, 204, 205, 206, 207, 208 The impact of regulatory microRNAs and their effects on the host inflammatory response in neonates with sepsis are unclear.
Endogenous cortisol is induced by proinflammatory cytokines and attenuates the intensity of SIRS associated with severe sepsis and septic shock.209 The use of cortisol in adults with sepsis has been controversial.210, 211 Cortisol production in newborn infants is significantly increased early in shock.212 However, very preterm neonates may have relative adrenal insufficiency that may contribute to hemodynamic instability and hypotension. Cortisol replacement may be critical in these infants and deserves further study.213 It is important to note, however, that in children with septic shock, adjunctive corticosteroid therapy is associated with repression of gene programs corresponding to the adaptive immune system.214
The Impact of Genetics in Sepsis-Associated Inflammatory Signaling
A twin study which assessed the frequency of infections among monoygotic and dizygotic prematurely born twins concluded that 49.0% (p = .002) of the variance in susceptibility to late-onset sepsis was due to genetic factors alone.215 The impact of genetics in the host response is also underscored by the increased risk for death from infection seen with African American race or male sex among low-birth-weight infants.216 An ethnically unique single nucleotide polymorphism in the TLR4 promoter region was significantly associated with gram-negative bacterial infections in preterm infants.217 Several recent studies in newborn infants have demonstrated an association between small variations in DNA, specifically single nucleotide polymorphisms, and infection development and outcomes.122, 218, 219, 220, 221, 222
Because TLRs play an essential role in recognition and response to pathogens, alterations in their expression, structure, signaling pathways, and function can have consequences for host defense. Polymorphisms or mutations in TLRs are associated with increased risk for infection in adults223, 224, 225, 226 and children210, 227, 228 but are less well characterized in neonates. After confounders had been controlled for, the presence of a TLR4 single nucleotide polymorphism was associated with a three-fold increase in the risk for gram-negative infections in VLBW infants.222 Polymorphisms in the TLR2, TLR5, IL10, and PLA2G2A (which encodes PLA2) genes were associated with the development of neonatal sepsis.218
Modifications in expression or function of costimulatory molecules necessary for TLR activation are also associated with an increased risk for infection. For example, the levels of LPS-binding protein (LBP; which binds intravascular LPS) and the LPS coreceptor CD14 are both increased during neonatal sepsis.211, 229, 230 Furthermore, genetic variations in these proteins have been associated with increased risk for sepsis in adults.231, 232, 233 Genetic polymorphisms in myeloid differentiation factor 2, a small protein involved in LPS signaling through TLR4, increase the risk for organ dysfunction and sepsis in adults234 but the significance in neonates is unknown. Polymorphisms in post-TLR activation intracellular signaling molecules, including myeloid differentiation factor 88,235 IL-1 receptor–associated kinase 4,236 and NF-κB essential modulator,237 are associated with invasive bacterial infection in older populations. Additional genetic polymorphisms in intracellular second-messenger inflammatory signaling systems with impact on neonatal sepsis risk and progression are likely to be uncovered with the implementation of biobanking and mining of stored samples.
Mutations have been identified in NLRs that are involved in the pathogenesis of Crohn's disease (NOD2)238 and neonatal-onset multisystem inflammatory disease (NLRP3).239 RLR mutations have been identified but have unknown clinical significance.240 No mutations in specific domains of NLRs have been found in neonates with sepsis or NEC.220, 231, 232, 233, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252 The importance of NLRs in Listeria monocytogenes infections in neonates is unknown.
Inflammatory Response Proteins
Complement
Complement is an important component of early innate immunity that facilitates killing of bacteria through opsonization and direct microbicidal activity. Complement components also possess chemotactic or anaphylactic activity that increases leukocyte aggregation and local vascular permeability. Furthermore, complement reciprocally activates a number of other important processes, such as coagulation, proinflammatory cytokine production, and leukocyte activation.104 Contrary to its name, the alternative pathway is the primary mechanism of amplification of complement activation after C3 convertase assembly (which cleaves C3 to C3a and C3b). Dysregulation of complement activation may contribute to adverse effects in individuals with severe sepsis or septic shock. Neonates, particularly the very premature, exhibit decreased basal levels of complement proteins and function for both the alternative pathway and the classical pathway.253, 254 Moreover, as compared with adults, neonates exhibit gestational age–related degrees of depressed complement-mediated opsonic capabilities.255 As such, complement-mediated opsonization is poor in premature neonates and limited in term neonates.255, 256
Complement-mediated activation of leukocytes during sepsis occurs via up-regulated cell surface receptors (complement receptor 1 [CD35] and CR3).257, 258 C3b and C5a facilitate opsonization (primarily C3b), redistribute blood flow, and increase inflammation, platelet aggregation, and release of reactive oxygen intermediates (primarily C5a).259, 260 C5a-mediated local leukocyte activation also results in increased cytokine production with subsequent up-regulation of adhesion molecules on vascular endothelium and increased cell recruitment to the site of infection.261 Data in adults link elevated C5a levels with multiple facets of sepsis-associated disease, such as DIC, cardiac dysfunction, increased proinflammatory cytokine levels, SIRS, apoptosis of adrenal medullary cells leading to adrenal insufficiency, and PMN dysfunction.104 Septic shock in adult humans was associated with extensive complement activation, C-reactive protein–dependent loss of C5a receptor on neutrophils, and the appearance of circulating C5a receptor in serum, which correlated with a poor outcome.262 Deficiencies in C5a receptor found in term neonates as compared with adults may limit the ability to respond to C5a and therefore increase the likelihood of infection.240 The expression of C5a receptor on preterm PMNs is unknown. The extent to which C5a or other complement proteins play a role in the development of disease in septic neonates remains to be determined.
Complement regulatory proteins modify the effects of complement and prevent potential damage due to overactivation. In particular, CD59 blocks C9 polymerization and target lysis, CD55 destabilizes CD35 and C3 and C5 convertases, and CD35 accelerates the deactivation of C3b.263 Dysregulation of complement activation can lead to a vicious activation cycle that results in excessive cellular stimulation, cytokine production, endothelial cell activation, and local tissue damage promulgating SIRS and septic shock (see Figure 152-5).264
Acute-Phase Reactants
In addition to the initial inflammatory response including complement activation, molecular detection of PAMPs promotes IL-1β and IL-6 production, which in turn increases the production of multiple other innate proteins that possess valuable immune function and serve to reduce pathogen load.265 Acute-phase reactant proteins, produced predominantly in the liver, include C-reactive protein (opsonin), serum amyloid A (cellular recruitment), lactoferrin (reduces the level of available iron/antimicrobial peptide lactoferricin), procalcitonin (unknown function), haptoglobin, fibronectin (opsonic function), pentraxin 3 (binds C1q and activates the classical complement pathway), MBL, and LPS-binding protein.127, 211, 229, 265, 266, 267, 268, 269, 270 Acute-phase reactant proteins have been studied in neonates with sepsis primarily to assess them for diagnostic utility rather than immunologic function. In particular, elevated plasma concentrations of C-reactive protein and LPS-binding protein are often associated with early-onset sepsis.229, 271 The levels of IL-10 and C-reactive protein were significantly higher in preterm infants who did not survive sepsis, pneumonia, or NEC.272 A lack of sustained increase in the production of C-reactive protein and serum amyloid A during sepsis has also been associated with a fulminant course.273
Passive Immunoglobulin
The fetus receives antibodies from the mother via active placental transfer, with a significant increase beginning around 20 weeks' gestation. As a result of a shorter period of gestation, preterm neonates have lower IgG subclass levels as compared with term neonates, particularly IgG1 and IgG2 subclasses.274 Preterm neonates (24 to 32 weeks' gestation) with low IgG levels (serum total IgG levels below 400 mg/dL at birth) were at increased risk for development of late-onset sepsis but not death compared to those with levels above 400 mg/dL. However, IgG titers and opsonic activity to CoNS were not predictive of late-onset CoNS sepsis.275 Reliance on other means of innate immune defense likely provides the premature neonate with alternative microbial control mechanisms. Despite the presence of maternally derived immunoglobulin and acute-phase reactant proteins, neonates exhibit impaired opsonizing activity compared with adults, which likely increases the risk for progression of infection.276 Complement plays a critical role in immunoglobulin-mediated opsonization and effector cell phagocytosis.277 Although immunoglobulin has many putative beneficial immunologic functions, most of these have not been demonstrated or examined in preterm infants.278 The dependence on complement for effective immunoglobulin-based opsonization and pathogen clearance may help explain the lack of efficacy of intravenous immunoglobulin to prevent sepsis or death from sepsis in neonates.279, 280, 281, 282, 283, 284, 285
Antimicrobial Proteins and Peptides
APPs are the most phylogenetically ancient means of innate immune defense against microbial invasion. Present in nearly every organism, including bacteria, plants, insects, nonmammalian vertebrates, and mammals, these small, often cationic peptides are capable of killing microbes of multiple types, including viruses, bacteria, parasites, and fungi, largely by disruption of the pathogen membrane.286 Constitutive expression of APPs occurs in humans on barrier areas with consistent microbial exposure such as skin and mucosa. After microbial stimulation, both release of preformed APPs and inducible expression are thought to contribute to early host defense.287 Importantly, there is no evidence for the development of microbial resistance to APPs that target fundamental components of the microbial cell wall. Some APPs can bind and neutralize microbial components such as endotoxin, precluding engagement with TLRs and other PRRs, and diminish inflammation. Many APPs can potentially reduce the intensity of the inflammatory response associated with the presence of bacterial toxins.288, 289, 290 Because endotoxemia is an important contributor to neonatal MODS and death with sepsis and NEC,84 LPS-binding/blocking strategies, including use of synthetic APPs, may have a significant positive impact on outcomes.288, 291
Bactericidal/permeability-increasing protein (BPI) is a 55-kDa protein present in the respiratory tract, PMN primary granules, and plasma. BPI exerts selective cytotoxic, antiendotoxic, and opsonic activity against gram-negative bacteria.288 Plasma BPI concentrations were higher in critically ill children with sepsis syndrome or organ system failure than in critically ill children without sepsis syndrome or organ system failure, and BPI levels positively correlated with the pediatric risk for death score.265 PMNs from term neonates are deficient in BPI, potentially contributing to the increased risk for infection.292 Whereas term neonates demonstrate up-regulation of plasma BPI during infection, premature neonates showed a decreased ability to mobilize BPI on stimulation,293 which may contribute to their risk for infection with gram-negative bacteria. Polymorphisms in BPI increase the risk for gram-negative sepsis in children, but the impact of these polymorphisms in neonates is unknown.294 Compared with PMNs from adults, PMNs from term neonates produce similar quantities of defensins but reduced quantities of BPI and elastase.292, 295, 296 Recombinant BPI (rBPI21) treatment was associated with improved functional outcome, reduced amputation, but no difference in mortality in a multicenter study of children with severe systemic meningococcal disease.297
Lactoferrin is the major whey protein in mammalian milk (in particularly high concentrations in colostrum) and is important in innate immune host defenses. Lactoferrin is present in tears and saliva and has antimicrobial activity both via binding iron and by direct membrane disruption activity via a portion of its amino-terminal lactoferricin.298 Lactoferrin is also an alarmin (e.g., HMGB-1 or IL-33), capable of activating leukocytes, binding endotoxin, and modifying the host response by acting as a transcription factor that regulates mRNA decay.299, 300 Bovine lactoferrin has been shown to reduce the incidence of bacterial and fungal sepsis301, 302 and NEC in preterm infants.303
Lysozyme is present in tears, tracheal aspirates, skin, and PMN primary and secondary granules and contributes to degradation of peptidoglycan in bacterial cell walls. Secretory PLA2 can destroy gram-positive bacteria through hydrolysis of their membrane lipids.174 PMN elastase is a serine protease released by activated PMNs with microbicidal function and is believed to play a role in the inflammatory damage seen with PMN recruitment, particularly in the lung.116, 136 Cathelicidin and the defensins are other APPs that possess antimicrobial properties.304 Cathelicidin is present in the amniotic fluid, vernix, skin, saliva, respiratory tract, and leukocytes. α-Defensins are cysteine-rich 4-kDa peptides found in amniotic fluid, vernix, spleen, cornea, thymus, Paneth cells, and leukocytes. β-Defensins are found in the skin, GI tract, urinary system, reproductive organs (placenta, uterus, testes, kidney), respiratory tract, breast milk, mammary gland, and thymus.
In addition to microbicidal action, APPs have a wide range of immunomodulatory effects on multiple cell types from both the innate immune system and the adaptive immune system.287, 305, 306 These immunomodulatory effects include altered cytokine and chemokine production, improved cellular chemotaxis and recruitment, improved cell function (maturation, activation, phagocytosis, reactive oxygen intermediate production), enhancement of wound healing (neovascularization, mitogenesis), and decreased apoptosis.
The cytosolic granules of PMN are rich in APPs, including α-defensins, lactoferrin, lysozyme, cathelicidin, soluble PLA2, and BPI. Gestational age–related decreases in the umbilical cord blood concentration of several APPs (cathelicidin, BPI, calprotectin, soluble PLA2, α-defensins) in comparison with maternal serum levels have been drescibed.307 Plasma APP deficiencies may contribute to the increased risk for infection associated with prematurity, and their absence may increase the risk for endotoxemia. Compared with term neonates, preterm neonates showed lower human β-defensin 2 levels in umbilical cord blood.308 Up-regulation of APPs (defensins) occurs in blood of infected adults309 and children (defensins, lactoferrin).310 The effect of sepsis on the production of plasma APPs in neonates has not been investigated in detail.
Coagulation
The development of a procoagulant state in the surrounding microvasculature allows the trapping of invading pathogens and prevents further dissemination (see Figure 152-5). In general, the intrinsic pathway amplifies coagulation after initiation by the extrinsic pathway.311 Reduced levels of vitamin K–dependent factors (factors II, VII, IX, and X), reduced thrombin generation, reduced consumption of platelets with formation of microthrombi, and reduced levels of counterregulatory elements (inhibitors) increase the risk for bleeding in infants and children.312 During sepsis, a microvascular procoagulant state develops via stimulation of phagocytes, platelets, and endothelium, resulting in expression of tissue factor.313, 314 Tissue factor–mediated activation of the coagulation cascade results in activation of thrombin-antithrombin complex, plasminogen activator inhibitor type 1, and plasmin–α2-antiplasmin complex,315 as well as inactivation of protein S and depletion of the anticoagulant proteins antithrombin III and protein C.316, 317, 318 Decreased activated protein C levels were associated with increased risk for death from sepsis in preterm neonates.319 A randomized controlled trial of activated protein C revealed no change in mortality among pediatric patients with sepsis, but term infants younger than 60 days old experienced increased bleeding.320
The coagulation cascade is intimately tied to inflammation and complement activation.104 Cytokine production increases expression of endothelial tissue plasminogen activator inhibitor type 1. Plasminogen activator inhibitor type 1 inhibits fibrinolysis by inhibiting the conversion of plasminogen to plasmin, which in turn is important for the breakdown of fibrin. Deposition of fibrin in small vessels leads to inadequate tissue perfusion and organ failure.321 Increased plasminogen activator inhibitor type 1 levels are associated with increased IL-6, nitrite, and nitrate levels (metabolites of NO production), the development of organ failure, and increased mortality.321 Sepsis is associated with thrombocytopenia in neonates,322 which is attributed to reduced megakaryopoiesis in the setting of consumption with clot formation.323 Decreased platelet function in preterm neonates with sepsis further increases the risk for bleeding.324 In extremely low-birth-weight infants, platelets are hyporeactive for the first few days after birth, complicating the ability of the immune system to contain a microbiologic threat and increasing the risk for hemorrhage.325 Clotting can lead to propagation of inflammation via thrombin-induced production of platelet-activating factor. PMNs activated by platelet-activating factor or platelet TLR4 may then contribute to further endothelial injury and dysfunction, leading to the development of a vicious clotting-inflammation-clotting cycle. Activated platelets may be consumed in clot formation and/or may also be removed from the circulation by the liver,326 potentially resulting in thrombocytopenia, particularly during gram-negative and fungal infections.196, 322, 327
Systemic activation of coagulation is associated with consumption of clotting factors and increased risk for bleeding, prolonged proinflammatory responses, and DIC.128, 149, 328 This finding is consistent with the elevated serum levels of IL-652 and high frequency of DIC seen with disseminated herpes simplex virus infection.329 In adult mice, protease-activated receptor 1 plays a major role in orchestrating the interplay between coagulation and inflammation.330 Protease-activated receptor 1 may modify the endothelial response during neonatal sepsis and thus is a target for therapeutic intervention.
Role of Vascular Endothelium
Recent studies have shown the critical importance of vascular endothelial activation in the early recognition and containment of microbial invasion. In transgenic mice, it was shown that pulmonary endothelial cells sense blood-borne bacteria and their products,156 whereas alveolar macrophages patrol the air spaces.331 These data illustrate the role of endothelium and help to explain in part the occurrence of acute respiratory distress syndrome (ARDS) and persistent pulmonary hypertension of the newborn associated with severe sepsis in the absence of a primary pulmonary infectious focus. Expression of TLRs allows endothelium to become activated in the presence of microbial components, leading to production of cytokines, chemokines, and adhesion molecules (e.g., vascular cell adhesion molecule, intercellular cell adhesion molecule, L-selectin, P-selectin, and E-selectin). These substances are all necessary to attract immune cells (primarily PMNs) to the site of infection and to facilitate pathogen containment.150, 151, 152, 153, 156 Vasoactive substances released from activated leukocytes, platelets, and endothelial cells include platelet-activating factor, thromboxanes, leukotrienes, NO, histamine, bradykinin, and prostaglandins.172, 173 The balance of NO and endothelin 1, a vasoconstrictor, may be disrupted with endothelial damage, favoring the constrictive effects of endothelin 1 and leading to ischemia and injury.175 This phenomenon may explain in part why NO inhibitors increased mortality in adults with septic shock.301 Stimulated endothelium can be a double-edged sword, however, because excessive activation can lead to systemic overproduction of cytokines and vasoactive substances (including NO). Endothelial cell apoptosis, detachment from the lamina, and alterations in vascular tone combine to promote capillary leak, leading to hypovolemia, shock, and organ failure156, 302, 303 (see Figure 152-5). Release of myeloperoxidase from PMNs may also injure surrounding endothelium.332 Activated or damaged endothelium establishes a prothrombotic environment that can result in local microvascular occlusion314 or progress to DIC.333
The glucocorticoid receptor is the target for cortisol, the primary endogenous glucocorticoid in humans, produced in the zona fasciculata of the adrenal glands. Endothelial glucocorticoid receptor is a critical negative regulator of inducible NO synthase expression and NF-κB activation,334 demonstrating a protective role of the endothelium during sepsis. Studies have revealed a potential role of plasma angiopoietin during pediatric septic shock.335 The level of angiopoietin 1, which protects against vascular leak, was reduced, whereas the level of angiopoietin 2, which promotes vascular permeability, was elevated, highlighting a novel potential therapeutic opportunity to reduce end-organ injury. The roles for endothelial glucocorticoid receptor and angiopoietin 1 in neonatal sepsis are unknown.
The role of endothelium activation during sepsis and septic shock in neonates, particularly in premature neonates, has been less well investigated. Toxins from GBS have been shown to damage pulmonary endothelium336 and likely participate in pulmonary complications associated with GBS pneumonia such as ARDS and the development of persistent pulmonary hypertension of the newborn.337 The levels of the adhesion molecules E-selectin and P-selectin, expressed and secreted by activated endothelium, are increased in the serum of neonates with sepsis136 and likely reflect significant endothelial activation. Endothelial TLR4 activation impaired intestinal perfusion in an experimental model of NEC, via endothelial NO synthase–nitrite–NO signaling.338
Innate Immune Cellular Contributions
The PMN is the primary effector of innate immune cellular defense. Endothelial cells produce activating cytokines and chemokine gradients that recruit circulating PMNs to the site of infection. Expression of cell adhesion molecules by PMNs and endothelium allows cells to roll and extravasate into surrounding tissues. Activated PMNs phagocytose and kill pathogens via oxygen-dependent and oxygen-independent mechanisms. IL-1β is produced by activated PMNs largely via an NLRP3-ASC-caspase 1–dependent* mechanism that amplifies the recruitment of additional PMNs from the bone marrow to the site of infection.339
Activated PMNs may release reactive nitrogen species, reactive oxygen species, and proteolytic enzymes via activation of membrane-associated NADPH oxidase. These reactive intermediates and enzymes can lead to destruction of nonphagocytosed bacteria but can also cause local tissue destruction, including neonatal endothelial and lung injury, as well as surfactant inactivation,116, 340 and thus play a role in progression from sepsis to MODS.
Neonatal PMNs exhibit quantitative and qualitative deficits as compared with adult PMNs.295, 341 Respiratory burst activity is suppressed in PMNs during neonatal sepsis and may contribute to poor microbicidal activity.342, 343, 344 Compared with adult PMNs, neonatal PMNs exhibit delayed apoptosis,345, 346 as well as sustained capacity for activation (CD11b up-regulation) and cytotoxic function (reactive oxygen intermediate production) that contributes to tissue damage.347 Reduced apoptosis with prolonged survival of PMNs may result in improved bacterial clearance but may also paradoxically increase the risk for sustained PMN-mediated tissue damage. Increased serum PMN elastase, urokinase plasminogen activator, and urokinase plasminogen activator receptor levels are found at the time of presentation in infected neonates.136
With PMN death, DNA (chromatin), histones, and APPs are expelled into the environment and serve to trap bacteria (neutrophil extracellular traps [NETs]).348 The formation of NETs can occur after activation of platelet TLR4349 and may lead to excessive local inflammation and tissue damage.350 High early levels of circulating free PMN-derived DNA produced by NETs are associated with MODS and death.351 NETs contain destructive proteases capable of killing bacteria even after the PMN has died.352 Formation of NETs is reduced in PMNs from preterm neonates and nearly absent in term neonates353 but may occur with sustained cellular stimulation.354 NET formation may result in collateral damage to surrounding tissues when the target microbe is too large to be effectively phagocytosed (e.g., fungal hyphae).355 The contribution of NET production to detrimental outcomes in infected neonates is unknown but excessive NET formation with collateral tissue injury may contribute to the poor outcomes seen in preterm neonates with fungal infections.356
Rapid depletion of bone marrow PMN reserves during infection, particularly in neonates,357 can lead to neutropenia, with consequent impaired antimicrobial defenses and significantly increased risk for death.358 In a multivariate analysis, neutropenia and metabolic acidosis were associated with fatal neonatal sepsis.359 Neutropenia is particularly common in gram-negative sepsis in neonates.360 Release of immature PMN forms (bands), which exhibit greater dysfunction than mature PMNs,361 may further predispose to adverse outcomes. Murine neonates with experimental sepsis exhibit delayed emergency myelopoiesis (a process by which the host repopulates peripheral myeloid cells lost early during sepsis), that is independent of TRIF and myeloid differentiation factor 88.362 Interventions aimed at addressing reduced PMN numbers in neonates have included provision of mature PMNs363 and prophylaxis or treatment with colony-stimulating factors (granulocyte colony-stimulating factor and granulocyte-macrophage colony-stimulating factor). Despite strong biologic plausibility, these interventions have been unsuccessful at reducing the neonatal infectious burden.364, 365, 366 In a metaanalysis, treatment with colony-stimulating factor therapy (granulocyte colony-stimulating factor, granulocyte-macrophage colony-stimulating factor) in a subgroup (n = 97) of neutropenic neonates (absolute neutrophil count less than 1700/µL) with culture-positive sepsis (largely gram-negative and GBS) significantly reduced the risk for death (relative risk, 0.34; 95% CI, 0.12 to 0.92).365 Therefore, stimulation of granulopoiesis may be beneficial under these specific circumstances, although further studies focused on this subpopulation and outcomes are needed.
Irreversible aggregation and accumulation of newborn PMNs in the vascular space after stimulation leads to decreased diapedesis, rapid depletion of bone marrow reserves, vascular crowding,367 and increased likelihood of microvascular occlusion.368 Neonatal PMN deformation compared with adult PMN deformation is reduced at the baseline, which increases the risk for occlusion.367 Furthermore, low blood pressure/flow states seen during septic shock further exacerbate existing microvascular ischemia.295 In combination, these deficiencies increase the propensity for systemic spread of infection, and set the stage for microvascular occlusion.
Other Innate Cellular Contributions
Many other cells besides PMNs are involved in the development of an immune response to infection. Monocytes, macrophages, and dendritic cells amplify cellular recruitment through production of inflammatory mediators, activation of endothelium, phagocytosis and killing of pathogens, and antigen presentation to T and B cells of the adaptive immune system. The primary functions of monocytes are the synthesis of crucial inflammatory proteins369 and antigen presentation to naïve CD4+ T cells.370
The patterns of cytokine production can promote the differentiation of naïve CD4+ T cells into distinct subtypes of T cells that serve important roles in the clearance of pathogens. For example, T-helper 1 (TH1) cells are produced from naïve CD4+ T cells after exposure to IFN-γ and IL-12, and support cell-mediated immunity against intracellular pathogens through production of IFN-γ, TNF-α, and lymphotoxin. T-helper 2(TH2) cells arise in the presence of IL-2 and IL-4, produce IL-4, IL-5, and IL-13, down-regulate TH1 responses, and support humoral immunity, as well as defense against extracellular parasites. A third subset of TH cells, T-helper 17 cells, are generated in the presence of transforming growth factor β, IL-6, IL-21, and IL-23. These cells produce IL-17 and IL-22, which are important for defense against extracellular bacteria and fungi. Neonatal mononuclear cells exhibit a bias away from TH1 cell–polarizing activity because of increased IL-6 and low TNF-α production.371 This may be beneficial because of mobilization of antiinfective proteins/peptides that serve to protect the newborn during microbial colonization266 and development of immune tolerance.341 The adverse consequence is a reduced ability to respond to infection with microorganisms; particularly intracellular pathogens such as Listeria sp.372 and mycobacteria.373 Preterm infants (<30 weeks) may have greater attenuation of TNF-α and IL-6 secretion compared with term infants and adults.277
There is decreased monocytic recruitment to areas of inflammation during sepsis because of decreased chemotactic ability.374 Although the levels of peripheral monocytes decrease early during sepsis (between 60 nd 120 hours), secondary to extravasation and differentiation into macrophages, sepsis-related elevation of macrophage colony-stimulating factor375 results in a late increase in the number of peripheral monocytes (>120 hours).376 In addition to altered cytokine production and suboptimal recruitment, monocyte phagocytic function is reduced during sepsis.377 Antigen presentation to naïve CD4+ T cells is an important immune function performed by monocytes. The decreased antigen-presenting function in monocytes from newborn infants is in part due to decreased MHC class 2 molecule expression378 and decreased expression of costimulatory molecules, including CD86 and CD40.379
Monocytes leave the bloodstream, enter the tissues, and differentiate into macrophages and dendritic cells. Monocytes and macrophages are closely related to PMNs (common myeloid progenitor) and can kill pathogens by similar means. Circulating monocytes differentiate into macrophages after exposure to maturing cytokines, and exit the bloodstream into tissues. Important substances produced by stimulated monocytes/macrophages include complement components, cytokines (both proinflammatory and antiinflammatory), coagulation factors, and extracellular matrix proteins.369 Located just below epithelial borders, macrophages encounter pathogens immediately after entry. Macrophages are avidly phagocytic and generate APPs to reduce bacterial burden, such as lactoferrin, defensins, transferrin, and lysozyme. In addition, macrophages play an important role in the amplification of the immune response through the production of cytokines and chemokines, as well as in antigen presentation to naïve CD4+ T cells. Macrophages are poorly responsive to several TLR agonists.380
Dendritic cells are antigen-presenting cells that function as a liaison between the innate immune system and the adaptive immune system through induction of antigen-specific T cell–mediated immunity. Dendritic cells from newborn infants exhibit a reduced antigen-presenting function when compared with adult cells379 and require increased stimulation for activation.381 Evaluations of neonatal dendritic cell function suggest a tendency towards poor up-regulation of costimulatory molecules (CD80/CD86) and activation markers (CD83), poor stimulation of T-cell proliferation, and a tendency towards the induction of immune tolerance.382 Although preterm and term infants and adults have similar numbers of “plasmacytoid” dendritic cells in their blood, the capacity to produce IFN-α on TLR9 challenge was significantly decreased in preterm neonates and may increase susceptibility to viral infections.383 Dendritic cells in umbilical cord blood can effectively induce cytotoxic lymphocyte responses.384 Depletion of dendritic cells has been reported in adult animals385 and adult patients386 with sepsis; their role in the immune response to neonatal sepsis is not well characterized.
Eosinophils phagocytose antigen-antibody complexes and release cytokines, chemokines, cytotoxic molecules, APPs, and other substances (prostaglandins, thromboxanes, leukotrienes) when stimulated.387 Eosinophilia is commonly observed in neonates with sepsis due to Candida sp.388 and bacteria,389 and is seen in infants with NEC.387 In infants of less than 26 weeks' gestation, eosinophilia (absolute eosinophil count more than 1000/mm3) may predict bacterial sepsis.389 Eosinophilia in premature infants is not associated with production of IgE.390 Studies have demonstrated an integral role for eosinophils in adult intestinal integrity and revealed a novel innate bactericidal nonphagocytic function via extracellular catapulting of mitochondrial DNA nets with associated bound toxic proteins.391 The precise role of eosinophils in the neonatal immune response to sepsis and in maintenance of intestinal integrity has yet to be determined.
Mast cells play a role in the response to pathogen invasion as a part of the innate cellular immune system via production of histamines (which promote vasodilation and up-regulation of P-selectin), cytokines, PMN recruitment, bacterial phagocytosis, and antigen presentation.392, 393 Mast cell involvement was demonstrated in infants with erythema toxicum, where mast cell recruitment, degranulation, and expression of APPs occurs.394 Adult rodents deficient in mast cells exhibit impaired PMN influx,395 impaired clearance of enteric organisms, and decreased sepsis survival.396 Mast cell production of histamine likely contributes to the vasodilation associated with sepsis and septic shock. Like eosinophils and PMNs, mast cells are capable of killing bacteria via generation of extracellular traps in adults.397 This means of immune protection has not been investigated in neonates. Mast cells may also alter adaptive immune function by patterning the TH2 immunophenotype seen in the neonate and therefore contribute to the increased risk for infection. Immature dendritic cells exposed to histamine during maturation (with LPS) exhibit altered T-cell polarizing activity with predominance towards the TH2 phenotype via increased production of IL-10 and decreased production of IL-2.398 Furthermore, mast cells from neonates were shown to secrete significantly more histamine after stimulation as compared with adults,399 which may contribute to the development of shock.400
The role of natural killer (NK) cells in neonatal bacterial sepsis is incompletely defined. NK cell numbers increase with increasing gestational age,401 Furthermore, a reduced percentage of NK cells present at birth may be a risk factor for late-onset sepsis in preterm infants.402 It is noteworthy that the numbers of circulating NK cells are not significantly different in neonates with or without infection370, 403; however, the numbers of circulating NK cells are decreased in newborn infants with shock.404 The mechanisms used by NK cells to destroy bacteria include secretion of APPs (defensins), direct contact and lysis, antibody-dependent cellular cytotoxicity, and IFN-γ production.405 In neonates with bacterial sepsis, NK cells are activated, as evidenced by up-regulation of CD69.2, 406 Despite activation, NK cell cytotoxicity is deficient in infants with sepsis and recurrent infections.370, 405 Although neonatal macrophages exhibit impaired baseline activation in response to IFN-γ,341 NK cell–mediated production of IFN-γ can enhance their phagocytic capability. Further studies are necessary to more clearly define the role of NK cells in neonatal bacterial sepsis.
CD71+Ter119+ (erythroid) cells may contribute to the increased susceptibility of the neonate to infection by reducing the inflammatory response associated with bacterial colonization of the gut. For example, ex vivo TNF-α production by stimulated adult effector cells was reduced in the presence of murine neonatal splenic CD71+ erythroid cells via an arginase 2–dependnent mechanism.407 The CD71+ erythroid population represents a large portion of murine fetal liver, neonatal spleen/bone marrow, and adult bone marrow.408, 409, 410 Furthermore, the murine neonatal spleen contains large numbers of colony-forming progenitor cells for 2 to 3 weeks after birth.411 Of note and in stark contrast to the lymphoid and reticuloendothelial system roles of the spleen in the healthy adult, the spleen is normally a major site of erythropoiesis during fetal and neonatal life, to support rapid fetal and postnatal growth in the setting of significantly reduced erythroid reservoirs as compared with the adult reservoirs.410, 412, 413 A lack of effect on neonatal murine survival to polymicrobial sepsis after adoptive transfer or diminution of CD71+ erythroid splenocytes suggests that the impact of these cells on neonatal infection risk and progression may be limited.414
Contributions of the Adaptive Immune System
The contribution of the adaptive immune system in the neonatal host response to sepsis is uncertain. The 5- to 7-day interval required for development of an adaptive immune response—namely, the selection and amplification of specific clones of lymphocytes (B cells and T cells) that results in immunologic memory—argues against a central role for adaptive immunity in the protective response to early neonatal bacterial sepsis. As a result, the neonate is thought to largely depend on innate immunity for protection from infection during the first days of life. In adults, absence or dysfunction of the adaptive immune system has a profound impact on survival in preclinical models.415 B cells (and in particular B-cell cytokine production) and not T cells were shown to be important in the early host response to experimental sepsis.416 Studies using neonatal mice lacking an adaptive immune system showed no difference in polymicrobial sepsis survival as compared with survival of wild-type mice with an intact adaptive immune system.3 Furthermore, there are many quantitative and qualitative differences in lymphocytes from neonates compared with lymphocytes from adults,417 each with a respective proposed clinical impact.87 As these findings illustrate, the contribution of adaptive immunity for protection and response against sepsis, and in particular which components are protective, is unclear in the most immature and requires further investigation.
Peripheral blood examination has yielded inconsistent changes in the percentage, number, and type of circulating lymphocytes during sepsis.403, 418, 419, 420, 421, 422, 423 Moreover, changes related to the timing of sepsis onset (early-onset or late-onset sepsis) and prematurity have been incompletely characterized. T regulatory cells are abundant and potent at birth, facilitating inhibition of TH1 cell immunity,424 and perhaps mediating a state of immunologic tolerance.425 Although the numbers of splenic T regulatory cells are increased in murine neonates and adults with sepsis, depletion of T regulatory cells had no effect on survival of murine adults.2, 426 Alterations in the number or function of T regulatory cells in human neonatal sepsis have not been reported.
Examination of peripheral blood to identify markers of sepsis has yielded a number of lymphocyte cell-surface molecules whose levels increase during sepsis. Activation of neonatal T cells is evidenced by increased CD45RO expression (present on T cells after antigenic stimulation) at the time of sepsis diagnosis,419, 427, 428 and with congenital infection,429 although changes in number may take several days to occur after stimulation.430 Other markers of lymphocyte activation may be found at different time points during the course of infection. For example, expression of the activation marker CD69 is increased on T cells (CD4+) early in the infectious process, whereas CD25 and CD45RO expression persists for several days.406 Increased expression of CD4+ T-cell carcinoembryonic antigen–related cell adhesion molecule 1 (CD66a) in preterm infants with late-onset sepsis may contribute to sepsis-associated immune suppression.431 HLA-DR expression is increased on multiple cell types during neonatal sepsis.406 In contrast to adults, a large portion of neonatal T cells produce CXCL8, which activates PMN and γδ T cells.432 These data show that neonatal T cells are activated and are capable of playing a role in the host response to bacterial sepsis in vivo.
Neonatal lymphocyte function is skewed towards TH2 responses, setting the stage for immune tolerance (TH2) rather than immune priming for infection (TH1).417 Newborn infants must overcome that immune modulation in order to mount effective responses to specific infectious challenges and respond to vaccination. Examples of the impact of this immunopolarization include decreased IFN-γ production by CD4+ and CD8+ T cells as compared with production in children and adults.433, 434 The likely significance of decreased IFN-γ production is a reduction in activation of other immune cells, such as macrophages.
Reports of lymphocyte function in infected newborns are very limited. Expansion of lymphocytes after antigenic stimulation is important for development of sustained immunity. Decreased lymphocyte proliferative responses have been shown during the first 8 weeks of life in VLBW neonates,421 and may predispose the premature neonate to development of late-onset sepsis. For example, T-lymphocyte function was depressed in infected newborn infants, and especially in those with multiorgan failure, versus healthy term or growing preterm infants.435 Similarly, production of lymphocyte-associated cytokines after stimulation of umbilical cord blood peripheral blood mononuclear cells with GBS was significantly deficient in preterm and term infants compared with adults.277 Cytomegalovirus infection in utero leads to the expansion and differentiation of mature cytomegalovirus-specific CD8+ T cells, which have characteristics similar to adult CD8+ T cells.436 These cells showed potent perforin-dependent cytolytic activity and produce antiviral cytokines, highlighting the potential for adult-like immunocompetence of neonatal T cells under specific circumstances.
An important location for effective lymphocytic function during systemic bacterial infection is the spleen. The marginal zone of the spleen facilitates the clearance of bacteria, particularly encapsulated organisms, from the bloodstream. These functions are accomplished via the interaction of multiple leukocytes, including macrophages, dendritic cells, B cells, and T cells, within follicles of the spleen. The neonatal splenic marginal zone is immature, owing to a lack of antecedent antigen exposure and is virtually devoid of CD21+ B cells.412 As a result of this functional asplenia, there is decreased clearance of pathogens from the blood and potential for a more fulminant course with bacteremia.437, 438
B cells are critically important in the adult host response to sepsis. Data suggest antibody-independent and antibody-dependent roles for B cells in the outcome of sepsis.416 Studies deciphering the role of B cells in neonatal sepsis are very limited, and thus the role B cells play in the neonatal host response is unclear. After GBS meningitis, the level of IgM was increased, suggesting B cells from neonates can respond to pathogenic challenge.439 Premature neonates with perinatal infection or nosocomial infection may show signs of humoral immunoparalysis, manifested by decreased IgM/IgG production ex vivo as compared with production in their healthy age-matched counterparts.440 Sepsis in early life did not reduce serum antibody titers in preterm infants after heptavalent pneumococcal conjugate vaccine exposure but was associated with a reduced opsonization titer to a single serotype, suggesting the capacity to respond to vaccination or other immune challenge may be altered.441
In the setting of reduced classic adaptive immune function seen in early life as compared with the function in adults, innate lymphoid populations (which lack B cell receptor and T cell receptor) may play a significant role in protecting the neonate from infectious challenge.442, 443, 444, 445, 446, 447, 448 Examples of innate lymphoid cells include γδ T cells, intestinal lymphoid cells, invariant NK T cells, mucosa-associated invariant T cells, and B1 cells. Mechanistic investigations that fully explore the role of these newly discovered populations in the neonatal host response to sepsis are likely to uncover novel therapeutic opportunities.
Impact of Systems Biology Approaches
Systems biology and the use of “omic” approaches have the potential to produce significant insights into the pathogenesis of sepsis. Genomic and proteomic approaches have yielded important data associated with septic shock in older populations.449, 450, 451, 452, 453, 454, 455, 456, 457 The use of these modern techniques in the study of neonatal inflammation and response to pathogen challenge has only just begun.136, 198, 458, 459 The ability to profile genome-wide expression has significantly enhanced our understanding of the complexity of the host immune response to sepsis in children.5, 449, 450, 453, 460 For example, genome-wide expression profiling revealed zinc homeostasis as an important feature of pediatric sepsis.450, 453, 461 Prophylactic zinc supplementation reduced bacterial load and mortality in a murine model of peritoneal sepsis.462 However, oral zinc supplementation did not alter mortality in neonates with probable sepsis.463
In a study of pediatric patients who met the criteria for septic shock,6 a unique whole-blood transcriptomic response was found in neonates as compared with infants, toddlers, and school-age children. Neonates manifested the largest number of uniquely regulated genes, representing both innate and adaptive immune system pathways, and showed a predominance of down-regulated transcripts representing the adaptive immune system.5 The number of up-regulated genes increased in proportion with developmental age. Investigation of the murine circulating leukocyte transcriptome revealed significant differences in the host immune response to sepsis across the age spectrum (neonate, young adult, elderly), despite similar increases in mortality among the neonates and elderly mice as compared with young adult mice.1 These data underscore the impact of developmental age on the host immune response and suggest that therapeutics, which may have efficacy in older populations, may be ineffective in or possibly detrimental to neonates.
Because the transition to extrauterine life is associated with dramatic changes in physiology, the whole-blood transcriptome is likely to be quite different between both uninfected infants and in the host response to sepsis by timing after birth. Indeed, an unsupervised analysis of the whole-blood genome-wide transcriptome on prospectively collected peripheral blood samples from infants evaluated for sepsis revealed the major node of separation between groups (infected or uninfected) was the timing of evaluation relative to birth (early, lass than 3 days or late, more than 3 days).464 Principal component analyses revealed significant differences between patients with early or late sepsis despite the presence of similar key immunologic pathway aberrations in both groups. This study highlights both the uninfected state and the host responses to sepsis are significantly affected by timing relative to birth.
A study of VLBW infants with blood culture–proven late-onset sepsis (59% CoNS) identified a 554-gene signature associated with sepsis, with increased expression of the TNF-α network, including matrix metalloproteinase 8 and CD177 among the most commonly up-regulated genes.465 Elevated matrix metalloproteinase 8 mRNA expression and activity in septic shock correlated with decreased survival and increased organ failure in pediatric patients. Matrix metalloproteinase 8 is a direct activator of NF-κB.466 Inhibition (genetic or pharmacologic) of matrix metalloproteinase 8 leads to improved survival and a blunted inflammatory profile in a murine model of sepsis. Most recently, a 52-gene network was uncovered and validated that accurately identified infected infants, who exhibited increased expression of genes for innate immune and metabolic pathways with decreased levels of adaptive immune transcripts.467 Using a proteomics approach, Ng and colleagues468 identified pro-apolipoprotein CII and a desarginine variant of serum amyloid A as promising biomarkers for late-onset sepsis and NEC in preterm infants.468 It is very likely that implementation of unbiased “omic” approaches will reveal critical age-appropriate pathways and opportunities for therapeutic interventions aimed at improving neonatal sepsis outcomes.
Sepsis-Associated Organ Failure
Sepsis that leads to shock and organ failure carries the worst prognosis. SIRS contributes to the development of organ failure in neonates (see Figure 152-5).52, 148, 181, 469 Persistent decreases in capillary perfusion are associated with MODS and death in adults.470 Lethargy, shock, and birth weight less than 1500 g were independent predictors of sepsis-related death.471 In neonates, impairment of the cardiovascular system, manifested by poor perfusion or hypotension, is invariably associated with septic shock. Sustained poor organ perfusion in neonatal sepsis and septic shock due to cardiovascular dysfunction is associated with MODS affecting the kidney,472, 473 liver,474 gut,475 and central nervous system476 (see Figure 152-5). The mechanism of organ failure may be decreased oxygen utilization associated with mitochondrial dysfunction rather than poor oxygen delivery to tissue.477, 478 On the basis of available evidence, it has been speculated that the prolonged SIRS associated with severe sepsis and shock leads to organ failure via a cessation of energy-consuming processes.479, 480 Development of severe NEC is also associated with severe sepsis, shock, MODS, and death.84, 481 The need for intubation or initiation of vasoactive medications, and hypoglycemia, thrombocytopenia, increased prothrombin time, or excessive bleeding as presenting laboratory signs of sepsis are risk factors for sepsis-related death.359, 475, 482 Independent predictors of in-hospital late-onset sepsis death during the birth hospitalization were the presence of congenital anomalies (OR, 4.12; 95% CI, 1.60 to 10.60), neuromuscular comorbidities (OR, 3.34; 95% CI, 1.66 to 6.73), and secondary pulmonary hypertension with/without cor pulmonale (OR, 23.48; 95% CI, 5.96 to 92.49),60 underscoring the impact of organ-level comorbidities that increase neonatal sepsis mortality.
Cardiovascular System
The most common organ dysfunction associated with sepsis is cardiovascular dysfunction. Cardiovascular dysfunction associated with sepsis may lead to shock that is a composite of hypovolemic, cardiogenic, and distributive shock. Distributive shock is related to endothelial NO production that leads to excessive vasodilation. Cardiogenic shock may be related to mitochondrial death (induced by reactive nitrogen and reactive oxygen intermediates) with subsequent myocardial dysfunction. Abnormalities in peripheral vasoregulation and myocardial dysfunction may play a larger role in hemodynamic derangements in pediatric patients, especially infants and neonates.
In children, a non-hyperdynamic state with reduced cardiac output and increased systemic vascular resistance is most commonly observed in the setting of sepsis.483, 484, 485, 486, 487 The hemodynamic presentation in neonates is much more variable484 and is complicated by an unclear association between a normal blood pressure and adequate systemic blood flow.488, 489 Microcirculatory flow is impaired in term neonates even with mild to moderate severity of infection.490 Preterm neonates with sepsis have relatively high left and right cardiac outputs and low systemic vascular resistances. However, a decrease in right or left ventricular output of more than 50% has been associated with increased mortality in neonatal sepsis.491 An elevated left ventricular output but normal ejection fraction in preterm neonates with septic shock suggests that septic shock in preterm neonates is predominantly due to vasoregulatory failure. Neonatal sepsis may or may not be associated with left ventricular diastolic dysfunction; however, cardiac injury as manifested by elevated levels of cardiac troponin T may complicate the clinical picture.492, 493 Abnormal peripheral vasoregulation with or without myocardial dysfunction is the primary mechanism for the hypotension accompanying septic shock in the neonate.494 Therefore, infected neonates may present with hypotension and adequate perfusion (warm shock) or inadequate perfusion (cold shock). Myocardial dysfunction can lead to ventricular wall stretch that in turn elevates B-type natriuretic peptide levels. B-type natriuretic peptide levels are elevated in children with septic shock,495 and increased levels have utility as prognostic indicators of death.496 Plasma NO level is elevated in neonates with sepsis and shock compared wit those with shock alone.182 Elevated serum lactate level (>3 mmol/L) distinguished nonsurvivors from survivors in a pediatric study that included neonates.497
Immune System
After severe sepsis or septic shock, there is an increased risk for subsequent infection and death in children and adults. This phenomenon is termed immunoparalysis and is associated with reduced MHC class 2 expression and TNF-α production by mononuclear cells after endotoxin stimulation. In addition to altered monocytic responses, there is significant loss of lymphoid CD4+ T and B cells via caspase-dependent apoptotic pathways.415, 498 Whether by clonal selection, apoptosis, or elevated endogenous glucocorticoid levels,499, 500, 501 lymphocyte loss may lead to a state of immune compromise after the acute phase of sepsis.412, 499, 501, 502, 503, 504, 505 New data suggests that IL-7 may play an important role in promoting T-cell activation and prevention of apoptosis.506 The importance of immunoparalysis has been convincingly demonstrated in infected adults507, 508, 509, 510 and children.511 However, the clinical impact in the preterm neonate in whom adaptive immune function is less well developed is uncertain.512, 513
In examinations of peripheral blood and postmortem spleens from infected adults, there is significant loss of B and CD4+ T lymphocytes and dendritic cells,386, 498 resulting in decreased antigen presentation, antibody production, and macrophage activation.514 Circulating peripheral absolute lymphocyte counts can drop significantly in adults with sepsis but this phenomena is also seen in critically ill adults who are not infected.477 Sustained lymphopenia significantly increases the risk for secondary infection, MODS, and death in children.505 Extensive loss of lymphocytes (both B and T lymphocytes) has been described in postmortem specimens from the thymus and spleen in infected preterm and term infants.412, 499, 501, 502, 503, 504 The number and the size of the follicles in the spleen decreased significantly and the total number of cells decreased by more than three times; similar changes were found in lymph nodes.478 However, these histopathologic splenic findings are in contradiction to earlier reports where no differences were described in infected and uninfected infants.412 Splenomegaly may occur in infants with late-onset sepsis and may be due to splenic congestion in the absence of hyperplasia of white pulp.502
The mechanisms responsible for immune alterations after sepsis are beginning to emerge. The intensity of the inflammatory response may be modified by neural-based mechanisms.515 T cell–secreted acetylcholine acts on macrophages to reduce production of TNF, IL-1, IL-18, HMGB-1, and other cytokines.516 The role of vagal tone in the neonatal host response to sepsis is unclear.
Discovery and characterization of the impact of epigenetic-mediated immune system functional alterations after sepsis is an area of intense research. DNA methylation and posttranslational modification of histone proteins (methylation, acetylation, phosphorylation, ubiquitination, SUMOylation) may occur after sepsis.126, 517 These DNA alterations may modify transcription factor access of gene-specific promoter regions, ultimately leading to short-term and long-term changes in gene expression and immune function. The DNA methylation pattern in the promoter region of the CALCA gene varied in different types of bacterial sepsis in preterm infants, suggesting its potential use as an epigenetic biomarker.518
Trained immunity, the term coined to describe an adaptive innate immune response, may also be a positive or negative consequence of sepsis in early life.519 Mechanisms that underlie trained immunity are beginning to emerge, and include DNA methylation and modification of energy utilization pathways.520, 521 Nonspecific vaccine benefits and resistance to subsequent pathogen challenge after innate immune priming or previous infection are likely manifestations of trained immunity in neonates.3, 513, 522 The cell types, extent, and duration of trained immunity-based modifications in neonates with sepsis have not been studied. Myeloid suppressor cells manifest immunosuppressive activity with sepsis523 and were recently described in neonates. Myeloid suppressor cells are present at high frequency at birth and decline in number with postnatal age. They inhibit T-cell proliferative responses and IFN-γ production.524 Reactivation of viral infection that may contribute to morbidity and mortality has been demonstrated in infected adults.525 The impact of this phenomenon in neonates is unknown.
Pulmonary System
Acute hypoxic respiratory failure, ARDS, and acute lung injury are common pulmonary complications associated with severe sepsis. Destruction of the alveolar capillary membrane leads to refractory hypoxemia. After direct or indirect insults to the lung, alveolar macrophages produce chemokines that mitigate PMN influx to lung parenchyma. Activated PMNs release reactive oxygen and reactive nitrogen intermediates that damage endothelial and epithelial barriers, leading to leakage of protein-rich edema fluid into the air spaces. Other pulmonary complications with severe sepsis may include secondary surfactant deficiency,526 primary or secondary pneumonia,527 and reactive pulmonary hypertension.528, 529 Infants with sepsis and persistent pulmonary hypertension of the newborn may require inhaled NO in addition to optimized ventilation strategies such as high-frequency oscillatory ventilation.530 If oxygenation or tissue perfusion remains severely compromised despite optimal medical management, extracorporeal membrane oxygenation should be considered in neonates weighing more than 2 kg without contraindications.531, 532
Central Nervous System
The detrimental neurodevelopmental long-term impact of sepsis has been demonstrated in multiple studies and has been reviewed in detail.533, 534, 535, 536, 537 Central nervous system injury is predominantly white-matter injury (loss of pre-oligodendrocytes), manifested by focal cystic periventricular leukomalacia, diffuse necrosis, or a combination of these entities.538, 539 Central nervous system injury is, in part, mediated by inflammation with or without direct pathogen invasion.141, 540, 541 The impact of sepsis on central nervous system injury is intensified with lower gestational ages, highlighting the detrimental effects of sepsis on the developing brain.539 The importance of evaluating the preterm infant for disseminated infection that may include meningitis cannot be overemphasized. A complete evaluation, including cultures of blood, urine and cerebrospinal fluid, is uncommon,356 although one third of the cases of culture-positive meningitis in VLBW infants are associated with negative concurrent blood cultures.542 Clinically apparent seizures may occur in 25% of VLBW preterm infants with meningitis.542 Low-voltage background pattern, sleep-wake cycling, and seizure activity on the amplitude-integrated electroencephalogram may be helpful to predict neurologic outcome in infants with sepsis or meningitis.543 Significantly lower resistance, vasodilatation, and higher blood flow were noted in all the cerebral arteries of infants with sepsis. Increase in cerebral blood flow velocity was correlated with elevated IL-6 concentrations.544 Alterations in blood flow in preterm infants, in addition to factors associated with sepsis, such as respiratory distress, hypercarbia, hypotension, and patent ductus arteriosus, contribute to the risk for intracerebral hemorrhage.
Other Organ System Contributions
Endocrine abnormalities may include altered thyroid function545 and adrenal insufficiency associated with refractory hypotension.546 Inadequate adrenocortical responses are associated with increased mortality.547, 548 Cortisol production in the neonate is significantly increased early in septic shock.212 However, very preterm neonates can have relative adrenal insufficiency that may contribute to hemodynamic instability and hypotension. Hydrocortisone has not been evaluated in large prospective randomized clinical trials for the treatment of septic shock in the neonate but it has been shown to increase blood pressure, decrease heart rate, and decrease vasoactive medication requirements in preterm and term neonates in addition to its cytokine-suppressing effects.549, 550, 551 If hydrocortisone treatment is considered, the obtaining of a pretreatment serum cortisol level is prudent in order to differentiate contributing causes of hypotension.
Sepsis was the most common cause (78%) of acute kidney injury in term neonates and was associated with high mortality (37%) (n = 49).552 The frequency of acute renal failure (defined as a blood urea nitrogen level greater than 20mg/dL) in infected neonates was 26% and oliguria occurred in 15% of acute kidney failure cases.553 Acute kidney injury in preterm neonates is associated with high mortality.554 Hepatic injury and dysfunction are frequent associations with severe sepsis. The mechanisms include reduced hepatic perfusion associated with septic shock and mitochondrial energy failure. Reductions in coagulation and complement factors, acute-phase reactant proteins, and increases in the levels of transaminases and bilirubin are commonly seen, especially in association with decreased perfusion states. Energy expenditure and oxygen consumption are increased during sepsis,555 and decreased mitochondrial oxidative function precipitated by hypoxia and the presence of reactive oxygen intermediates may lead to impaired growth, caloric deficiency, and energy failure.556, 557
Multiorgan Failure/Dysfunction
Sepsis that leads to MODS carries a dismal prognosis. Inadequate cardiac output and microcirculatory failure, which may be combined with formation of microthrombi and DIC, can lead to poor perfusion to the kidney,472, 473 liver,474 gut,475 and central nervous system.52, 148, 181, 469, 476 Recent studies suggest that the mechanism of organ failure in sepsis may relate to decreased oxygen utilization associated with mitochondrial dysfunction rather than poor oxygen delivery to tissues.479, 480 Mitochondrial dysfunction can initiate activation of cell death pathways, including apoptosis, pyroptosis, necrosis, and NETosis (i.e. cell death mediated by NETs). Damage-associated molecular patterns (including nucleosomes and microparticles) created by activation of these cell death programs further amplify the host inflammatory response.
Free radicals play an important role in the inflammatory process of sepsis.558 In a sepsis model in neonatal piglets, edaravone, a novel free radical scavenger, increased mean arterial pressure and cardiac output, lowered heart rate, reduced hydroperoxide, nitrite, and nitrate levels, delayed the TNF-α surge, prevented HMGB-1 level elevation, and was associated with longer survival times.559 Increased plasma nitrite and nitrate concentrations are associated with the development of multiple organ failure in pediatric patients with sepsis560, 561 but have not been investigated in neonates.
Future Considerations
The incidence of neonatal sepsis remains high and outcomes remain poor despite considerable technologic advances in the field of neonatology. Much remains to be learned about the impact of developmental age on the host response to sepsis and what facets are critically important. Important considerations for future investigations include the development and implementation of a generally accepted definition for neonatal sepsis, the use of homogeneous systems (only neonatal components) for human ex vivo studies, and transgenic approaches in preclinical models, alongside observational studies in humans to ensure meaningful findings.
 Complete reference list is available at
www.ExpertConsult.com
.
Complete reference list is available at
www.ExpertConsult.com
.
Footnotes
ASC, Apoptosis-associated speck-like protein containing a carboxy-terminal CARD.
References
- 1.Gentile LF, Nacionales DC, Lopez MC. Protective immunity and defects in the neonatal and elderly immune response to sepsis. J Immunol. 2014;192(7):3156–3165. doi: 10.4049/jimmunol.1301726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wynn JL, Scumpia PO, Delano MJ. Increased mortality and altered immunity in neonatal sepsis produced by generalized peritonitis. Shock. 2007;28(6):675–683. doi: 10.1097/SHK.0b013e3180556d09. [DOI] [PubMed] [Google Scholar]
- 3.Wynn JL, Scumpia PO, Winfield RD. Defective innate immunity predisposes murine neonates to poor sepsis outcome but is reversed by TLR agonists. Blood. 2008;112(5):1750–1758. doi: 10.1182/blood-2008-01-130500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wynn J, Cornell TT, Wong HR. The host response to sepsis and developmental impact. Pediatrics. 2010;125(5):1031–1041. doi: 10.1542/peds.2009-3301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wynn JL, Cvijanovich NZ, Allen GL. The influence of developmental age on the early transcriptomic response of children with septic shock. Mol Med. 2011;17(11–12):1146–1156. doi: 10.2119/molmed.2011.00169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Goldstein B, Giroir B, Randolph A. International pediatric sepsis consensus conference: definitions for sepsis and organ dysfunction in pediatrics. Pediatr Crit Care Med. 2005;6(1):2–8. doi: 10.1097/01.PCC.0000149131.72248.E6. [DOI] [PubMed] [Google Scholar]
- 7.Bone RC, Balk RA, Cerra FB. Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. The ACCP/SCCM Consensus Conference Committee. American College of Chest Physicians/Society of Critical Care Medicine. Chest. 1992;101(6):1644–1655. doi: 10.1378/chest.101.6.1644. [DOI] [PubMed] [Google Scholar]
- 8.Brierley J, Carcillo JA, Choong K. Clinical practice parameters for hemodynamic support of pediatric and neonatal septic shock: 2007 update from the American College of Critical Care Medicine. Crit Care Med. 2009;37(2):666–688. doi: 10.1097/CCM.0b013e31819323c6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dellinger RP, Levy MM, Carlet JM. Surviving Sepsis Campaign: international guidelines for management of severe sepsis and septic shock: 2008. Intensive Care Med. 2008;34(1):17–60. doi: 10.1007/s00134-007-0934-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hofer N, Zacharias E, Muller W, Resch B. Performance of the definitions of the systemic inflammatory response syndrome and sepsis in neonates. J Perinat Med. 2012;40(5):587–590. doi: 10.1515/jpm-2011-0308. [DOI] [PubMed] [Google Scholar]
- 11.Wynn JL, Wong HR, Shanley TP. Time for a neonatal-specific consensus definition for sepsis. Pediatr Crit Care Med. 2014;15(6):523–528. doi: 10.1097/PCC.0000000000000157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Liu L, Johnson HL, Cousens S. Global, regional, and national causes of child mortality: an updated systematic analysis for 2010 with time trends since 2000. Lancet. 2012;379(9832):2151–2161. doi: 10.1016/S0140-6736(12)60560-1. [DOI] [PubMed] [Google Scholar]
- 13.Lawn JE, Cousens S, Zupan J. 4 million neonatal deaths: when? Where? Why? Lancet. 2005;365(9462):891–900. doi: 10.1016/S0140-6736(05)71048-5. [DOI] [PubMed] [Google Scholar]
- 14.Stoll BJ, Hansen N, Fanaroff AA. Changes in pathogens causing early-onset sepsis in very-low-birth-weight infants. N Engl J Med. 2002;347(4):240–247. doi: 10.1056/NEJMoa012657. [DOI] [PubMed] [Google Scholar]
- 15.Stoll BJ, Hansen N, Fanaroff AA. Late-onset sepsis in very low birth weight neonates: the experience of the NICHD Neonatal Research Network. Pediatrics. 2002;110(2 Pt 1):285–291. doi: 10.1542/peds.110.2.285. [DOI] [PubMed] [Google Scholar]
- 16.Stoll BJ, Hansen NI, Higgins RD. Very low birth weight preterm infants with early onset neonatal sepsis: the predominance of gram-negative infections continues in the National Institute of Child Health and Human Development Neonatal Research Network, 2002-2003. Pediatr Infect Dis J. 2005;24(7):635–639. doi: 10.1097/01.inf.0000168749.82105.64. [DOI] [PubMed] [Google Scholar]
- 17.Haque KN, Khan MA, Kerry S. Pattern of culture-proven neonatal sepsis in a district general hospital in the United Kingdom. Infect Control Hosp Epidemiol. 2004;25(9):759–764. doi: 10.1086/502473. [DOI] [PubMed] [Google Scholar]
- 18.Klinger G, Levy I, Sirota L. Outcome of early-onset sepsis in a national cohort of very low birth weight infants. Pediatrics. 2010;125(4):e736–e740. doi: 10.1542/peds.2009-2017. [DOI] [PubMed] [Google Scholar]
- 19.Boghossian NS, Page GP, Bell EF. Late-onset sepsis in very low birth weight infants from singleton and multiple-gestation births. J Pediatr. 2013;162(6):1120–1124.e1. doi: 10.1016/j.jpeds.2012.11.089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Stoll BJ, Hansen NI, Sanchez PJ. Early onset neonatal sepsis: the burden of group B streptococcal and E. coli disease continues. Pediatrics. 2011;127(5):817–826. doi: 10.1542/peds.2010-2217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Barton L, Hodgman JE, Pavlova Z. Causes of death in the extremely low birth weight infant. Pediatrics. 1999;103(2):446–451. doi: 10.1542/peds.103.2.446. [DOI] [PubMed] [Google Scholar]
- 22.Cohen-Wolkowiez M, Moran C, Benjamin DK. Early and late onset sepsis in late preterm infants. Pediatr Infect Dis J. 2009;28(12):1052–1056. doi: 10.1097/inf.0b013e3181acf6bd. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Girard TD, Opal SM, Ely EW. Insights into severe sepsis in older patients: from epidemiology to evidence-based management. Clin Infect Dis. 2005;40(5):719–727. doi: 10.1086/427876. [DOI] [PubMed] [Google Scholar]
- 24.Martin GS, Mannino DM, Moss M. The effect of age on the development and outcome of adult sepsis. Crit Care Med. 2006;34(1):15–21. doi: 10.1097/01.ccm.0000194535.82812.ba. [DOI] [PubMed] [Google Scholar]
- 25.Stoll BJ, Hansen NI, Bell EF. Neonatal outcomes of extremely preterm infants from the NICHD Neonatal Research Network. Pediatrics. 2010;126(3):443–456. doi: 10.1542/peds.2009-2959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Watson RS, Carcillo JA, Linde-Zwirble WT. The epidemiology of severe sepsis in children in the United States. Am J Respir Crit Care Med. 2003;167(5):695–6701. doi: 10.1164/rccm.200207-682OC. [DOI] [PubMed] [Google Scholar]
- 27.Shah GS, Budhathoki S, Das BK, Mandal RN. Risk factors in early neonatal sepsis. Kathmandu Univ Med J. 2006;4(2):187–191. [PubMed] [Google Scholar]
- 28.Salem SY, Sheiner E, Zmora E. Risk factors for early neonatal sepsis. Arch Gynecol Obstet. 2006;274(4):198–202. doi: 10.1007/s00404-006-0135-1. [DOI] [PubMed] [Google Scholar]
- 29.Yancey MK, Duff P, Kubilis P. Risk factors for neonatal sepsis. Obstet Gynecol. 1996;87(2):188–194. doi: 10.1016/0029-7844(95)00402-5. [DOI] [PubMed] [Google Scholar]
- 30.Benitz WE, Gould JB, Druzin ML. Risk factors for early-onset group B streptococcal sepsis: estimation of odds ratios by critical literature review. Pediatrics. 1999;103(6):e77. doi: 10.1542/peds.103.6.e77. [DOI] [PubMed] [Google Scholar]
- 31.Fanaroff AA, Korones SB, Wright LL. Incidence, presenting features, risk factors and significance of late onset septicemia in very low birth weight infants. Pediatr Infect Dis J. 1998;17(7):593–598. doi: 10.1097/00006454-199807000-00004. [DOI] [PubMed] [Google Scholar]
- 32.Schuchat A, Zywicki SS, Dinsmoor MJ. Risk factors and opportunities for prevention of early-onset neonatal sepsis: a multicenter case-control study. Pediatrics. 2000;105(1 Pt 1):21–26. doi: 10.1542/peds.105.1.21. [DOI] [PubMed] [Google Scholar]
- 33.Escobar GJ, Li DK, Armstrong MA. Neonatal sepsis workups in infants >/=2000 grams at birth: a population-based study. Pediatrics. 2000;106(2 Pt 1):256–263. doi: 10.1542/peds.106.2.256. [DOI] [PubMed] [Google Scholar]
- 34.Gagliardi L, Rusconi F, Bellu R. Association of maternal hypertension and chorioamnionitis with preterm outcomes. Pediatrics. 2014;134(1):e154–e161. doi: 10.1542/peds.2013-3898. [DOI] [PubMed] [Google Scholar]
- 35.Garcia-Munoz Rodrigo F, Galan Henriquez G, Figueras Aloy J, Garcia-Alix Perez A. Outcomes of very-low-birth-weight infants exposed to maternal clinical chorioamnionitis: a multicentre study. Neonatology. 2014;106(3):229–234. doi: 10.1159/000363127. [DOI] [PubMed] [Google Scholar]
- 36.Jackson GL, Engle WD, Sendelbach DM. Are complete blood cell counts useful in the evaluation of asymptomatic neonates exposed to suspected chorioamnionitis? Pediatrics. 2004;113(5):1173–1180. doi: 10.1542/peds.113.5.1173. [DOI] [PubMed] [Google Scholar]
- 37.Jackson GL, Rawiki P, Sendelbach D. Hospital course and short-term outcomes of term and late preterm neonates following exposure to prolonged rupture of membranes and/or chorioamnionitis. Pediatr Infect Dis J. 2012;31(1):89–90. doi: 10.1097/INF.0b013e31822fb15d. [DOI] [PubMed] [Google Scholar]
- 38.Kiser C, Nawab U, McKenna K, Aghai ZH. Role of guidelines on length of therapy in chorioamnionitis and neonatal sepsis. Pediatrics. 2014;133(6):992–998. doi: 10.1542/peds.2013-2927. [DOI] [PubMed] [Google Scholar]
- 39.Pappas A, Kendrick DE, Shankaran S. Chorioamnionitis and early childhood outcomes among extremely low-gestational-age neonates. JAMA Pediatr. 2014;168(2):137–147. doi: 10.1001/jamapediatrics.2013.4248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Soraisham AS, Singhal N, McMillan DD. A multicenter study on the clinical outcome of chorioamnionitis in preterm infants. Am J Obstet Gynecol. 2009;200(4):372.e1–372.e6. doi: 10.1016/j.ajog.2008.11.034. [DOI] [PubMed] [Google Scholar]
- 41.Garcia-Munoz Rodrigo F, Galan Henriquez GM, Ospina CG. Morbidity and mortality among very-low-birth-weight infants born to mothers with clinical chorioamnionitis. Pediatr Neonatol. 2014;55(5):381–386. doi: 10.1016/j.pedneo.2013.12.007. [DOI] [PubMed] [Google Scholar]
- 42.Schrag SJ, Zell ER, Lynfield R. A population-based comparison of strategies to prevent early-onset group B streptococcal disease in neonates. N Engl J Med. 2002;347(4):233–239. doi: 10.1056/NEJMoa020205. [DOI] [PubMed] [Google Scholar]
- 43.Centers for Disease Control and Prevention Active Bacterial Core surveillance (ABCs) report: emerging infections program network: group B streptococcus, 2013 provisional. 2014. http://www.cdc.gov/abcs/reports-findings/survreports/gbs13.pdf Accessed July 29, 2014.
- 44.Bizzarro MJ, Dembry LM, Baltimore RS, Gallagher PG. Changing patterns in neonatal Escherichia coli sepsis and ampicillin resistance in the era of intrapartum antibiotic prophylaxis. Pediatrics. 2008;121(4):689–696. doi: 10.1542/peds.2007-2171. [DOI] [PubMed] [Google Scholar]
- 45.Whitley R, Arvin A, Prober C. Predictors of morbidity and mortality in neonates with herpes simplex virus infections. N Engl J Med. 1991;324(7):450–454. doi: 10.1056/NEJM199102143240704. [DOI] [PubMed] [Google Scholar]
- 46.Brown ZA, Benedetti J, Ashley R. Neonatal herpes simplex virus infection in relation to asymptomatic maternal infection at the time of labor. N Engl J Med. 1991;324(18):1247–1252. doi: 10.1056/NEJM199105023241804. [DOI] [PubMed] [Google Scholar]
- 47.ACOG Practice Bulletin Clinical management guidelines for obstetrician–gynecologists. Number 82, June 2007. Management of herpes in pregnancy. Obstet Gynecol. 2007;109(6):1489–1498. doi: 10.1097/01.aog.0000263902.31953.3e. [DOI] [PubMed] [Google Scholar]
- 48.Schaffer FM, Newton JA. Intravenous gamma globulin administration to common variable immunodeficient women during pregnancy: case report and review of the literature. J Perinatol. 1994;14(2):114–117. [PubMed] [Google Scholar]
- 49.Kermorvant-Duchemin E, Laborie S, Rabilloud M. Outcome and prognostic factors in neonates with septic shock. Pediatr Crit Care Med. 2008;9(2):186–191. doi: 10.1097/PCC.0b013e31816689a8. [DOI] [PubMed] [Google Scholar]
- 50.Verboon-Maciolek MA, Krediet TG, Gerards LJ. Clinical and epidemiologic characteristics of viral infections in a neonatal intensive care unit during a 12-year period. Pediatr Infect Dis J. 2005;24(10):901–904. doi: 10.1097/01.inf.0000180471.03702.7f. [DOI] [PubMed] [Google Scholar]
References
- 1.Gentile LF, Nacionales DC, Lopez MC. Protective immunity and defects in the neonatal and elderly immune response to sepsis. J Immunol. 2014;192(7):3156–3165. doi: 10.4049/jimmunol.1301726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wynn JL, Scumpia PO, Delano MJ. Increased mortality and altered immunity in neonatal sepsis produced by generalized peritonitis. Shock. 2007;28(6):675–683. doi: 10.1097/SHK.0b013e3180556d09. [DOI] [PubMed] [Google Scholar]
- 3.Wynn JL, Scumpia PO, Winfield RD. Defective innate immunity predisposes murine neonates to poor sepsis outcome but is reversed by TLR agonists. Blood. 2008;112(5):1750–1758. doi: 10.1182/blood-2008-01-130500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wynn J, Cornell TT, Wong HR. The host response to sepsis and developmental impact. Pediatrics. 2010;125(5):1031–1041. doi: 10.1542/peds.2009-3301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wynn JL, Cvijanovich NZ, Allen GL. The influence of developmental age on the early transcriptomic response of children with septic shock. Mol Med. 2011;17(11–12):1146–1156. doi: 10.2119/molmed.2011.00169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Goldstein B, Giroir B, Randolph A. International pediatric sepsis consensus conference: definitions for sepsis and organ dysfunction in pediatrics. Pediatr Crit Care Med. 2005;6(1):2–8. doi: 10.1097/01.PCC.0000149131.72248.E6. [DOI] [PubMed] [Google Scholar]
- 7.Bone RC, Balk RA, Cerra FB. Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. The ACCP/SCCM Consensus Conference Committee. American College of Chest Physicians/Society of Critical Care Medicine. Chest. 1992;101(6):1644–1655. doi: 10.1378/chest.101.6.1644. [DOI] [PubMed] [Google Scholar]
- 8.Brierley J, Carcillo JA, Choong K. Clinical practice parameters for hemodynamic support of pediatric and neonatal septic shock: 2007 update from the American College of Critical Care Medicine. Crit Care Med. 2009;37(2):666–688. doi: 10.1097/CCM.0b013e31819323c6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dellinger RP, Levy MM, Carlet JM. Surviving Sepsis Campaign: international guidelines for management of severe sepsis and septic shock: 2008. Intensive Care Med. 2008;34(1):17–60. doi: 10.1007/s00134-007-0934-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hofer N, Zacharias E, Muller W, Resch B. Performance of the definitions of the systemic inflammatory response syndrome and sepsis in neonates. J Perinat Med. 2012;40(5):587–590. doi: 10.1515/jpm-2011-0308. [DOI] [PubMed] [Google Scholar]
- 11.Wynn JL, Wong HR, Shanley TP. Time for a neonatal-specific consensus definition for sepsis. Pediatr Crit Care Med. 2014;15(6):523–528. doi: 10.1097/PCC.0000000000000157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Liu L, Johnson HL, Cousens S. Global, regional, and national causes of child mortality: an updated systematic analysis for 2010 with time trends since 2000. Lancet. 2012;379(9832):2151–2161. doi: 10.1016/S0140-6736(12)60560-1. [DOI] [PubMed] [Google Scholar]
- 13.Lawn JE, Cousens S, Zupan J. 4 million neonatal deaths: when? Where? Why? Lancet. 2005;365(9462):891–900. doi: 10.1016/S0140-6736(05)71048-5. [DOI] [PubMed] [Google Scholar]
- 14.Stoll BJ, Hansen N, Fanaroff AA. Changes in pathogens causing early-onset sepsis in very-low-birth-weight infants. N Engl J Med. 2002;347(4):240–247. doi: 10.1056/NEJMoa012657. [DOI] [PubMed] [Google Scholar]
- 15.Stoll BJ, Hansen N, Fanaroff AA. Late-onset sepsis in very low birth weight neonates: the experience of the NICHD Neonatal Research Network. Pediatrics. 2002;110(2 Pt 1):285–291. doi: 10.1542/peds.110.2.285. [DOI] [PubMed] [Google Scholar]
- 16.Stoll BJ, Hansen NI, Higgins RD. Very low birth weight preterm infants with early onset neonatal sepsis: the predominance of gram-negative infections continues in the National Institute of Child Health and Human Development Neonatal Research Network, 2002-2003. Pediatr Infect Dis J. 2005;24(7):635–639. doi: 10.1097/01.inf.0000168749.82105.64. [DOI] [PubMed] [Google Scholar]
- 17.Haque KN, Khan MA, Kerry S. Pattern of culture-proven neonatal sepsis in a district general hospital in the United Kingdom. Infect Control Hosp Epidemiol. 2004;25(9):759–764. doi: 10.1086/502473. [DOI] [PubMed] [Google Scholar]
- 18.Klinger G, Levy I, Sirota L. Outcome of early-onset sepsis in a national cohort of very low birth weight infants. Pediatrics. 2010;125(4):e736–e740. doi: 10.1542/peds.2009-2017. [DOI] [PubMed] [Google Scholar]
- 19.Boghossian NS, Page GP, Bell EF. Late-onset sepsis in very low birth weight infants from singleton and multiple-gestation births. J Pediatr. 2013;162(6):1120–1124.e1. doi: 10.1016/j.jpeds.2012.11.089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Stoll BJ, Hansen NI, Sanchez PJ. Early onset neonatal sepsis: the burden of group B streptococcal and E. coli disease continues. Pediatrics. 2011;127(5):817–826. doi: 10.1542/peds.2010-2217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Barton L, Hodgman JE, Pavlova Z. Causes of death in the extremely low birth weight infant. Pediatrics. 1999;103(2):446–451. doi: 10.1542/peds.103.2.446. [DOI] [PubMed] [Google Scholar]
- 22.Cohen-Wolkowiez M, Moran C, Benjamin DK. Early and late onset sepsis in late preterm infants. Pediatr Infect Dis J. 2009;28(12):1052–1056. doi: 10.1097/inf.0b013e3181acf6bd. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Girard TD, Opal SM, Ely EW. Insights into severe sepsis in older patients: from epidemiology to evidence-based management. Clin Infect Dis. 2005;40(5):719–727. doi: 10.1086/427876. [DOI] [PubMed] [Google Scholar]
- 24.Martin GS, Mannino DM, Moss M. The effect of age on the development and outcome of adult sepsis. Crit Care Med. 2006;34(1):15–21. doi: 10.1097/01.ccm.0000194535.82812.ba. [DOI] [PubMed] [Google Scholar]
- 25.Stoll BJ, Hansen NI, Bell EF. Neonatal outcomes of extremely preterm infants from the NICHD Neonatal Research Network. Pediatrics. 2010;126(3):443–456. doi: 10.1542/peds.2009-2959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Watson RS, Carcillo JA, Linde-Zwirble WT. The epidemiology of severe sepsis in children in the United States. Am J Respir Crit Care Med. 2003;167(5):695–6701. doi: 10.1164/rccm.200207-682OC. [DOI] [PubMed] [Google Scholar]
- 27.Shah GS, Budhathoki S, Das BK, Mandal RN. Risk factors in early neonatal sepsis. Kathmandu Univ Med J. 2006;4(2):187–191. [PubMed] [Google Scholar]
- 28.Salem SY, Sheiner E, Zmora E. Risk factors for early neonatal sepsis. Arch Gynecol Obstet. 2006;274(4):198–202. doi: 10.1007/s00404-006-0135-1. [DOI] [PubMed] [Google Scholar]
- 29.Yancey MK, Duff P, Kubilis P. Risk factors for neonatal sepsis. Obstet Gynecol. 1996;87(2):188–194. doi: 10.1016/0029-7844(95)00402-5. [DOI] [PubMed] [Google Scholar]
- 30.Benitz WE, Gould JB, Druzin ML. Risk factors for early-onset group B streptococcal sepsis: estimation of odds ratios by critical literature review. Pediatrics. 1999;103(6):e77. doi: 10.1542/peds.103.6.e77. [DOI] [PubMed] [Google Scholar]
- 31.Fanaroff AA, Korones SB, Wright LL. Incidence, presenting features, risk factors and significance of late onset septicemia in very low birth weight infants. Pediatr Infect Dis J. 1998;17(7):593–598. doi: 10.1097/00006454-199807000-00004. [DOI] [PubMed] [Google Scholar]
- 32.Schuchat A, Zywicki SS, Dinsmoor MJ. Risk factors and opportunities for prevention of early-onset neonatal sepsis: a multicenter case-control study. Pediatrics. 2000;105(1 Pt 1):21–26. doi: 10.1542/peds.105.1.21. [DOI] [PubMed] [Google Scholar]
- 33.Escobar GJ, Li DK, Armstrong MA. Neonatal sepsis workups in infants >/=2000 grams at birth: a population-based study. Pediatrics. 2000;106(2 Pt 1):256–263. doi: 10.1542/peds.106.2.256. [DOI] [PubMed] [Google Scholar]
- 34.Gagliardi L, Rusconi F, Bellu R. Association of maternal hypertension and chorioamnionitis with preterm outcomes. Pediatrics. 2014;134(1):e154–e161. doi: 10.1542/peds.2013-3898. [DOI] [PubMed] [Google Scholar]
- 35.Garcia-Munoz Rodrigo F, Galan Henriquez G, Figueras Aloy J, Garcia-Alix Perez A. Outcomes of very-low-birth-weight infants exposed to maternal clinical chorioamnionitis: a multicentre study. Neonatology. 2014;106(3):229–234. doi: 10.1159/000363127. [DOI] [PubMed] [Google Scholar]
- 36.Jackson GL, Engle WD, Sendelbach DM. Are complete blood cell counts useful in the evaluation of asymptomatic neonates exposed to suspected chorioamnionitis? Pediatrics. 2004;113(5):1173–1180. doi: 10.1542/peds.113.5.1173. [DOI] [PubMed] [Google Scholar]
- 37.Jackson GL, Rawiki P, Sendelbach D. Hospital course and short-term outcomes of term and late preterm neonates following exposure to prolonged rupture of membranes and/or chorioamnionitis. Pediatr Infect Dis J. 2012;31(1):89–90. doi: 10.1097/INF.0b013e31822fb15d. [DOI] [PubMed] [Google Scholar]
- 38.Kiser C, Nawab U, McKenna K, Aghai ZH. Role of guidelines on length of therapy in chorioamnionitis and neonatal sepsis. Pediatrics. 2014;133(6):992–998. doi: 10.1542/peds.2013-2927. [DOI] [PubMed] [Google Scholar]
- 39.Pappas A, Kendrick DE, Shankaran S. Chorioamnionitis and early childhood outcomes among extremely low-gestational-age neonates. JAMA Pediatr. 2014;168(2):137–147. doi: 10.1001/jamapediatrics.2013.4248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Soraisham AS, Singhal N, McMillan DD. A multicenter study on the clinical outcome of chorioamnionitis in preterm infants. Am J Obstet Gynecol. 2009;200(4):372.e1–372.e6. doi: 10.1016/j.ajog.2008.11.034. [DOI] [PubMed] [Google Scholar]
- 41.Garcia-Munoz Rodrigo F, Galan Henriquez GM, Ospina CG. Morbidity and mortality among very-low-birth-weight infants born to mothers with clinical chorioamnionitis. Pediatr Neonatol. 2014;55(5):381–386. doi: 10.1016/j.pedneo.2013.12.007. [DOI] [PubMed] [Google Scholar]
- 42.Schrag SJ, Zell ER, Lynfield R. A population-based comparison of strategies to prevent early-onset group B streptococcal disease in neonates. N Engl J Med. 2002;347(4):233–239. doi: 10.1056/NEJMoa020205. [DOI] [PubMed] [Google Scholar]
- 43.Centers for Disease Control and Prevention Active Bacterial Core surveillance (ABCs) report: emerging infections program network: group B streptococcus, 2013 provisional. 2014. http://www.cdc.gov/abcs/reports-findings/survreports/gbs13.pdf Accessed July 29, 2014.
- 44.Bizzarro MJ, Dembry LM, Baltimore RS, Gallagher PG. Changing patterns in neonatal Escherichia coli sepsis and ampicillin resistance in the era of intrapartum antibiotic prophylaxis. Pediatrics. 2008;121(4):689–696. doi: 10.1542/peds.2007-2171. [DOI] [PubMed] [Google Scholar]
- 45.Whitley R, Arvin A, Prober C. Predictors of morbidity and mortality in neonates with herpes simplex virus infections. N Engl J Med. 1991;324(7):450–454. doi: 10.1056/NEJM199102143240704. [DOI] [PubMed] [Google Scholar]
- 46.Brown ZA, Benedetti J, Ashley R. Neonatal herpes simplex virus infection in relation to asymptomatic maternal infection at the time of labor. N Engl J Med. 1991;324(18):1247–1252. doi: 10.1056/NEJM199105023241804. [DOI] [PubMed] [Google Scholar]
- 47.ACOG Practice Bulletin Clinical management guidelines for obstetrician–gynecologists. Number 82, June 2007. Management of herpes in pregnancy. Obstet Gynecol. 2007;109(6):1489–1498. doi: 10.1097/01.aog.0000263902.31953.3e. [DOI] [PubMed] [Google Scholar]
- 48.Schaffer FM, Newton JA. Intravenous gamma globulin administration to common variable immunodeficient women during pregnancy: case report and review of the literature. J Perinatol. 1994;14(2):114–117. [PubMed] [Google Scholar]
- 49.Kermorvant-Duchemin E, Laborie S, Rabilloud M. Outcome and prognostic factors in neonates with septic shock. Pediatr Crit Care Med. 2008;9(2):186–191. doi: 10.1097/PCC.0b013e31816689a8. [DOI] [PubMed] [Google Scholar]
- 50.Verboon-Maciolek MA, Krediet TG, Gerards LJ. Clinical and epidemiologic characteristics of viral infections in a neonatal intensive care unit during a 12-year period. Pediatr Infect Dis J. 2005;24(10):901–904. doi: 10.1097/01.inf.0000180471.03702.7f. [DOI] [PubMed] [Google Scholar]
- 51.Verboon-Maciolek MA, Krediet TG, Gerards LJ. Severe neonatal parechovirus infection and similarity with enterovirus infection. Pediatr Infect Dis J. 2008;27(3):241–245. doi: 10.1097/INF.0b013e31815c1b07. [DOI] [PubMed] [Google Scholar]
- 52.Kawada J, Kimura H, Ito Y. Evaluation of systemic inflammatory responses in neonates with herpes simplex virus infection. J Infect Dis. 2004;190(3):494–498. doi: 10.1086/422325. [DOI] [PubMed] [Google Scholar]
- 53.Hornik CP, Fort P, Clark RH. Early and late onset sepsis in very-low-birth-weight infants from a large group of neonatal intensive care units. Early Hum Dev. 2012;88(Suppl 2):S69–S74. doi: 10.1016/S0378-3782(12)70019-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Karlowicz MG, Buescher ES, Surka AE. Fulminant late-onset sepsis in a neonatal intensive care unit, 1988-1997, and the impact of avoiding empiric vancomycin therapy. Pediatrics. 2000;106(6):1387–1390. doi: 10.1542/peds.106.6.1387. [DOI] [PubMed] [Google Scholar]
- 55.Joshi SG, Ghole VS, Niphadkar KB. Neonatal gram-negative bacteremia. Indian J Pediatr. 2000;67(1):27–32. doi: 10.1007/BF02802632. [DOI] [PubMed] [Google Scholar]
- 56.Orsi GB, d'Ettorre G, Panero A. Hospital-acquired infection surveillance in a neonatal intensive care unit. Am J Infect Control. 2009;37(3):201–203. doi: 10.1016/j.ajic.2008.05.009. [DOI] [PubMed] [Google Scholar]
- 57.Hyde TB, Hilger TM, Reingold A. Trends in incidence and antimicrobial resistance of early-onset sepsis: population-based surveillance in San Francisco and Atlanta. Pediatrics. 2002;110(4):690–695. doi: 10.1542/peds.110.4.690. [DOI] [PubMed] [Google Scholar]
- 58.Benjamin DK, DeLong E, Cotten CM. Mortality following blood culture in premature infants: increased with Gram-negative bacteremia and candidemia, but not Gram-positive bacteremia. J Perinatol. 2004;24(3):175–180. doi: 10.1038/sj.jp.7211068. [DOI] [PubMed] [Google Scholar]
- 59.Benjamin DK, Jr, Stoll BJ, Fanaroff AA. Neonatal candidiasis among extremely low birth weight infants: risk factors, mortality rates, and neurodevelopmental outcomes at 18 to 22 months. Pediatrics. 2006;117(1):84–92. doi: 10.1542/peds.2004-2292. [DOI] [PubMed] [Google Scholar]
- 60.Tsai MH, Hsu JF, Chu SM. Incidence, clinical characteristics and risk factors for adverse outcome in neonates with late-onset sepsis. Pediatr Infect Dis J. 2014;33(1):e7–e13. doi: 10.1097/INF.0b013e3182a72ee0. [DOI] [PubMed] [Google Scholar]
- 61.Schelonka RL, Chai MK, Yoder BA. Volume of blood required to detect common neonatal pathogens. J Pediatr. 1996;129(2):275–278. doi: 10.1016/s0022-3476(96)70254-8. [DOI] [PubMed] [Google Scholar]
- 62.Lukacs SL, Schrag SJ. Clinical sepsis in neonates and young infants, United States, 1988-2006. J Pediatr. 2012;160(6):960–965.e1. doi: 10.1016/j.jpeds.2011.12.023. [DOI] [PubMed] [Google Scholar]
- 63.Smit PM, Pronk SM, Kaandorp JC. RT-PCR detection of respiratory pathogens in newborn children admitted to a neonatal medium care unit. Pediatr Res. 2013;73(3):355–361. doi: 10.1038/pr.2012.176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kusuhara K, Saito M, Sasaki Y. An echovirus type 18 outbreak in a neonatal intensive care unit. Eur J Pediatr. 2008;167(5):587–589. doi: 10.1007/s00431-007-0516-x. [DOI] [PubMed] [Google Scholar]
- 65.Civardi E, Tzialla C, Baldanti F. Viral outbreaks in neonatal intensive care units: what we do not know. Am J Infect Control. 2013;41(10):854–856. doi: 10.1016/j.ajic.2013.01.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Tollin M, Bergsson G, Kai-Larsen Y. Vernix caseosa as a multi-component defence system based on polypeptides, lipids and their interactions. Cell Mol Life Sci. 2005;62(19–20):2390–2399. doi: 10.1007/s00018-005-5260-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Visscher MO, Narendran V, Pickens WL. Vernix caseosa in neonatal adaptation. J Perinatol. 2005;25(7):440–446. doi: 10.1038/sj.jp.7211305. [DOI] [PubMed] [Google Scholar]
- 68.Erez O, Romero R, Tarca AL. Differential expression pattern of genes encoding for anti-microbial peptides in the fetal membranes of patients with spontaneous preterm labor and intact membranes and those with preterm prelabor rupture of the membranes. J Matern Fetal Neonatal Med. 2009:1–13. doi: 10.3109/14767050902994796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.King AE, Kelly RW, Sallenave JM. Innate immune defences in the human uterus during pregnancy. Placenta. 2007;28(11–12):1099–1106. doi: 10.1016/j.placenta.2007.06.002. [DOI] [PubMed] [Google Scholar]
- 70.Leth-Larsen R, Floridon C, Nielsen O, Holmskov U. Surfactant protein D in the female genital tract. Mol Hum Reprod. 2004;10(3):149–154. doi: 10.1093/molehr/gah022. [DOI] [PubMed] [Google Scholar]
- 71.Dorschner RA, Lin KH, Murakami M, Gallo RL. Neonatal skin in mice and humans expresses increased levels of antimicrobial peptides: innate immunity during development of the adaptive response. Pediatr Res. 2003;53(4):566–572. doi: 10.1203/01.PDR.0000057205.64451.B7. [DOI] [PubMed] [Google Scholar]
- 72.Marchini G, Lindow S, Brismar H. The newborn infant is protected by an innate antimicrobial barrier: peptide antibiotics are present in the skin and vernix caseosa. Br J Dermatol. 2002;147(6):1127–1134. doi: 10.1046/j.1365-2133.2002.05014.x. [DOI] [PubMed] [Google Scholar]
- 73.Marchini G, Nelson A, Edner J. Erythema toxicum neonatorum is an innate immune response to commensal microbes penetrated into the skin of the newborn infant. Pediatr Res. 2005;58(3):613–616. doi: 10.1203/01.pdr.0000176836.27156.32. [DOI] [PubMed] [Google Scholar]
- 74.Carr JA, Hodgman JE, Freedman RI, Levan NE. Relationship between toxic erythema and infant maturity. Am J Dis Child. 1966;112(2):129–134. doi: 10.1001/archpedi.1966.02090110073006. [DOI] [PubMed] [Google Scholar]
- 75.Rutter N. Clinical consequences of an immature barrier. Semin Neonatol. 2000;5(4):281–287. doi: 10.1053/siny.2000.0014. [DOI] [PubMed] [Google Scholar]
- 76.Evans NJ, Rutter N. Development of the epidermis in the newborn. Biol Neonate. 1986;49(2):74–80. doi: 10.1159/000242513. [DOI] [PubMed] [Google Scholar]
- 77.Kalia YN, Nonato LB, Lund CH, Guy RH. Development of skin barrier function in premature infants. J Invest Dermatol. 1998;111(2):320–326. doi: 10.1046/j.1523-1747.1998.00289.x. [DOI] [PubMed] [Google Scholar]
- 78.Conner JM, Soll RF, Edwards WH. Topical ointment for preventing infection in preterm infants. Cochrane Database Syst Rev. 2004;(1) doi: 10.1002/14651858.CD001150.pub2. CD001150. [DOI] [PubMed] [Google Scholar]
- 79.Lewis D, Wilson C. Developmental immunology and role of host defenses in fetal and neonatal susceptibility to infection. In: Remington JS, Klein JO, Wilson CB, Baker CJ, editors. Infectious diseases of the fetus and newborn infant. ed 6. Saunders; Philadelphia: 2006. pp. 87–210. [Google Scholar]
- 80.Martin CR, Walker WA. Probiotics: role in pathophysiology and prevention in necrotizing enterocolitis. Semin Perinatol. 2008;32(2):127–137. doi: 10.1053/j.semperi.2008.01.006. [DOI] [PubMed] [Google Scholar]
- 81.La Rosa PS, Warner BB, Zhou Y. Patterned progression of bacterial populations in the premature infant gut. Proc Natl Acad Sci U S A. 2014;111(34):12522–12527. doi: 10.1073/pnas.1409497111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Holgerson PL, Vestman NR, Claesson R. Oral microbial profile discriminates breast-fed from formula-fed infants. J Pediatr Gastroenterol Nutr. 2013;56(2):127–136. doi: 10.1097/MPG.0b013e31826f2bc6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Deshmukh HS, Liu Y, Menkiti OR. The microbiota regulates neutrophil homeostasis and host resistance to Escherichia coli K1 sepsis in neonatal mice. Nat Med. 2014;20(5):524–530. doi: 10.1038/nm.3542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Sharma R, Tepas JJ, 3rd, Hudak ML. Neonatal gut barrier and multiple organ failure: role of endotoxin and proinflammatory cytokines in sepsis and necrotizing enterocolitis. J Pediatr Surg. 2007;42(3):454–461. doi: 10.1016/j.jpedsurg.2006.10.038. [DOI] [PubMed] [Google Scholar]
- 85.Cotten CM, Taylor S, Stoll B. Prolonged duration of initial empirical antibiotic treatment is associated with increased rates of necrotizing enterocolitis and death for extremely low birth weight infants. Pediatrics. 2009;123(1):58–66. doi: 10.1542/peds.2007-3423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Neu J. Gastrointestinal maturation and implications for infant feeding. Early Hum Dev. 2007;83(12):767–775. doi: 10.1016/j.earlhumdev.2007.09.009. [DOI] [PubMed] [Google Scholar]
- 87.Wynn JL, Neu J, Moldawer LL, Levy O. Potential of immunomodulatory agents for prevention and treatment of neonatal sepsis. J Perinatol. 2009;29(2):79–88. doi: 10.1038/jp.2008.132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Glasgow TS, Young PC, Wallin J. Association of intrapartum antibiotic exposure and late-onset serious bacterial infections in infants. Pediatrics. 2005;116(3):696–702. doi: 10.1542/peds.2004-2421. [DOI] [PubMed] [Google Scholar]
- 89.Carrico CJ, Meakins JL, Marshall JC. Multiple-organ-failure syndrome. Arch Surg. 1986;121(2):196–208. doi: 10.1001/archsurg.1986.01400020082010. [DOI] [PubMed] [Google Scholar]
- 90.Takahashi N, Vanlaere I, de Rycke R. IL-17 produced by Paneth cells drives TNF-induced shock. J Exp Med. 2008;205(8):1755–1761. doi: 10.1084/jem.20080588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Poindexter BB, Ehrenkranz RA, Stoll BJ. Parenteral glutamine supplementation does not reduce the risk of mortality or late-onset sepsis in extremely low birth weight infants. Pediatrics. 2004;113(5):1209–1215. doi: 10.1542/peds.113.5.1209. [DOI] [PubMed] [Google Scholar]
- 92.Patel AL, Johnson TJ, Engstrom JL. Impact of early human milk on sepsis and health-care costs in very low birth weight infants. J Perinatol. 2013;33(7):514–519. doi: 10.1038/jp.2013.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Meinzen-Derr J, Poindexter B, Wrage L. Role of human milk in extremely low birth weight infants' risk of necrotizing enterocolitis or death. J Perinatol. 2009;29(1):57–62. doi: 10.1038/jp.2008.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Lucas A, Cole TJ. Breast milk and neonatal necrotising enterocolitis. Lancet. 1990;336(8730):1519–1523. doi: 10.1016/0140-6736(90)93304-8. [DOI] [PubMed] [Google Scholar]
- 95.Bartlett JA, Fischer AJ, McCray PB., Jr Innate immune functions of the airway epithelium. Contrib Microbiol. 2008;15:147–163. doi: 10.1159/000136349. [DOI] [PubMed] [Google Scholar]
- 96.Nathe KE, Parad R, Van Marter LJ. Endotoxin-directed innate immunity in tracheal aspirates of mechanically ventilated human neonates. Pediatr Res. 2009;66(2):191–196. doi: 10.1203/PDR.0b013e3181aa33d7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Pfister RH, Soll RF. New synthetic surfactants: the next generation? Biol Neonate. 2005;87(4):338–344. doi: 10.1159/000084882. [DOI] [PubMed] [Google Scholar]
- 98.Starner TD, Agerberth B, Gudmundsson GH, McCray PB., Jr Expression and activity of beta-defensins and LL-37 in the developing human lung. J Immunol. 2005;174(3):1608–1615. doi: 10.4049/jimmunol.174.3.1608. [DOI] [PubMed] [Google Scholar]
- 99.Anderson DC, Rothlein R, Marlin SD. Impaired transendothelial migration by neonatal neutrophils: abnormalities of Mac-1 (CD11b/CD18)-dependent adherence reactions. Blood. 1990;76(12):2613–2621. [PubMed] [Google Scholar]
- 100.Poltorak A, He X, Smirnova I. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science. 1998;282(5396):2085–2088. doi: 10.1126/science.282.5396.2085. [DOI] [PubMed] [Google Scholar]
- 101.Kumagai Y, Takeuchi O, Akira S. Pathogen recognition by innate receptors. J Infect Chemother. 2008;14(2):86–92. doi: 10.1007/s10156-008-0596-1. [DOI] [PubMed] [Google Scholar]
- 102.Trinchieri G, Sher A. Cooperation of Toll-like receptor signals in innate immune defence. Nat Rev Immunol. 2007;7(3):179–190. doi: 10.1038/nri2038. [DOI] [PubMed] [Google Scholar]
- 103.Krumbiegel D, Zepp F, Meyer CU. Combined Toll-like receptor agonists synergistically increase production of inflammatory cytokines in human neonatal dendritic cells. Hum Immunol. 2007;68(10):813–822. doi: 10.1016/j.humimm.2007.08.001. [DOI] [PubMed] [Google Scholar]
- 104.Rittirsch D, Flierl MA, Ward PA. Harmful molecular mechanisms in sepsis. Nat Rev Immunol. 2008;8(10):776–787. doi: 10.1038/nri2402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Kawai T, Akira S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int Immunol. 2009;21(4):317–337. doi: 10.1093/intimm/dxp017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Cruz DN, Antonelli M, Fumagalli R. Early use of polymyxin B hemoperfusion in abdominal septic shock: the EUPHAS randomized controlled trial. JAMA. 2009;301(23):2445–2452. doi: 10.1001/jama.2009.856. [DOI] [PubMed] [Google Scholar]
- 107.Adhikari M, Coovadia HM, Gaffin SL. Septicaemic low birthweight neonates treated with human antibodies to endotoxin. Arch Dis Child. 1985;60(4):382–384. doi: 10.1136/adc.60.4.382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Togari H, Mikawa M, Iwanaga T. Endotoxin clearance by exchange blood transfusion in septic shock neonates. Acta Paediatr Scand. 1983;72(1):87–91. doi: 10.1111/j.1651-2227.1983.tb09669.x. [DOI] [PubMed] [Google Scholar]
- 109.Zhang JP, Chen C, Yang Y. [Changes and clinical significance of Toll-like receptor 2 and 4 expression in neonatal infections] Zhonghua Er Ke Za Zhi. 2007;45(2):130–133. [PubMed] [Google Scholar]
- 110.Leaphart CL, Cavallo J, Gribar SC. A critical role for TLR4 in the pathogenesis of necrotizing enterocolitis by modulating intestinal injury and repair. J Immunol. 2007;179(7):4808–4820. doi: 10.4049/jimmunol.179.7.4808. [DOI] [PubMed] [Google Scholar]
- 111.Latz E, Xiao TS, Stutz A. Activation and regulation of the inflammasomes. Nat Rev Immunol. 2013;13(6):397–411. doi: 10.1038/nri3452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Ehlers MR. CR3: a general purpose adhesion-recognition receptor essential for innate immunity. Microbes Infect. 2000;2(3):289–294. doi: 10.1016/s1286-4579(00)00299-9. [DOI] [PubMed] [Google Scholar]
- 113.McEvoy LT, Zakem-Cloud H, Tosi MF. Total cell content of CR3 (CD11b/CD18) and LFA-1 (CD11a/CD18) in neonatal neutrophils: relationship to gestational age. Blood. 1996;87(9):3929–3933. [PubMed] [Google Scholar]
- 114.Buhrer C, Graulich J, Stibenz D. L-selectin is down-regulated in umbilical cord blood granulocytes and monocytes of newborn infants with acute bacterial infection. Pediatr Res. 1994;36(6):799–804. doi: 10.1203/00006450-199412000-00020. [DOI] [PubMed] [Google Scholar]
- 115.Kim SK, Keeney SE, Alpard SK, Schmalstieg FC. Comparison of L-selectin and CD11b on neutrophils of adults and neonates during the first month of life. Pediatr Res. 2003;53(1):132–136. doi: 10.1203/00006450-200301000-00022. [DOI] [PubMed] [Google Scholar]
- 116.Nupponen I, Pesonen E, Andersson S. Neutrophil activation in preterm infants who have respiratory distress syndrome. Pediatrics. 2002;110(1 Pt 1):36–41. doi: 10.1542/peds.110.1.36. [DOI] [PubMed] [Google Scholar]
- 117.Auriti C, Fiscarelli E, Ronchetti MP. Procalcitonin in detecting neonatal nosocomial sepsis. Arch Dis Child Fetal Neonatal Ed. 2012;97(5):F368–F370. doi: 10.1136/fetalneonatal-2010-194100. [DOI] [PubMed] [Google Scholar]
- 118.Dzwonek AB, Neth OW, Thiebaut R. The role of mannose-binding lectin in susceptibility to infection in preterm neonates. Pediatr Res. 2008;63(6):680–685. doi: 10.1203/PDR.0b013e31816fdbff. [DOI] [PubMed] [Google Scholar]
- 119.Frakking FN, Brouwer N, van Eijkelenburg NK. Low mannose-binding lectin (MBL) levels in neonates with pneumonia and sepsis. Clin Exp Immunol. 2007;150(2):255–262. doi: 10.1111/j.1365-2249.2007.03479.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Frakking FN, Brouwer N, Zweers D. High prevalence of mannose-binding lectin (MBL) deficiency in premature neonates. Clin Exp Immunol. 2006;145(1):5–12. doi: 10.1111/j.1365-2249.2006.03093.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Sharma AA, Jen R, Kan B. Impaired NLRP3 inflammasome activity during fetal development regulates IL-1β production in human monocytes. Eur J Immunol. 2015;45(1):238–249. doi: 10.1002/eji.201444707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.van der Zwet WC, Catsburg A, van Elburg RM. Mannose-binding lectin (MBL) genotype in relation to risk of nosocomial infection in pre-term neonates in the neonatal intensive care unit. Clin Microbiol Infect. 2008;14(2):130–135. doi: 10.1111/j.1469-0691.2007.01886.x. [DOI] [PubMed] [Google Scholar]
- 123.Auriti C, Prencipe G, Inglese R. Role of mannose-binding lectin in nosocomial sepsis in critically ill neonates. Hum Immunol. 2010;71(11):1084–1088. doi: 10.1016/j.humimm.2010.08.012. [DOI] [PubMed] [Google Scholar]
- 124.Szala A, Paradowska E, Nowakowska D. Mannan-binding lectin-2 (MBL2) gene polymorphisms in prenatal and perinatal cytomegalovirus infections. Mol Immunol. 2011;48(15–16):2203–2206. doi: 10.1016/j.molimm.2011.06.220. [DOI] [PubMed] [Google Scholar]
- 125.Schlapbach LJ, Kjaer TR, Thiel S. M-ficolin concentrations in cord blood are related to circulating phagocytes and to early-onset sepsis. Pediatr Res. 2012;71(4 Pt 1):368–374. doi: 10.1038/pr.2011.71. [DOI] [PubMed] [Google Scholar]
- 126.Cornell TT, Wynn J, Shanley TP. Mechanisms and regulation of the gene-expression response to sepsis. Pediatrics. 2010;125(6):1248–1258. doi: 10.1542/peds.2009-3274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Ng PC. Diagnostic markers of infection in neonates. Arch Dis Child Fetal Neonatal Ed. 2004;89(3):F229–F235. doi: 10.1136/adc.2002.023838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Ng PC, Li K, Wong RP. Proinflammatory and anti-inflammatory cytokine responses in preterm infants with systemic infections. Arch Dis Child Fetal Neonatal Ed. 2003;88(3):F209–F213. doi: 10.1136/fn.88.3.F209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Levy O, Zarember KA, Roy RM. Selective impairment of TLR-mediated innate immunity in human newborns: neonatal blood plasma reduces monocyte TNF-alpha induction by bacterial lipopeptides, lipopolysaccharide, and imiquimod, but preserves the response to R-848. J Immunol. 2004;173(7):4627–4634. doi: 10.4049/jimmunol.173.7.4627. [DOI] [PubMed] [Google Scholar]
- 130.Bozza FA, Salluh JI, Japiassu AM. Cytokine profiles as markers of disease severity in sepsis: a multiplex analysis. Crit Care. 2007;11(2):R49. doi: 10.1186/cc5783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Hodge G, Hodge S, Haslam R. Rapid simultaneous measurement of multiple cytokines using 100 microl sample volumes—association with neonatal sepsis. Clin Exp Immunol. 2004;137(2):402–407. doi: 10.1111/j.1365-2249.2004.02529.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Heper Y, Akalin EH, Mistik R. Evaluation of serum C-reactive protein, procalcitonin, tumor necrosis factor alpha, and interleukin-10 levels as diagnostic and prognostic parameters in patients with community-acquired sepsis, severe sepsis, and septic shock. Eur J Clin Microbiol Infect Dis. 2006;25(8):481–491. doi: 10.1007/s10096-006-0168-1. [DOI] [PubMed] [Google Scholar]
- 133.Atici A, Satar M, Cetiner S, Yaman A. Serum tumor necrosis factor-alpha in neonatal sepsis. Am J Perinatol. 1997;14(7):401–404. doi: 10.1055/s-2007-994168. [DOI] [PubMed] [Google Scholar]
- 134.Sadeghi K, Berger A, Langgartner M. Immaturity of infection control in preterm and term newborns is associated with impaired toll-like receptor signaling. J Infect Dis. 2007;195(2):296–302. doi: 10.1086/509892. [DOI] [PubMed] [Google Scholar]
- 135.Philbin VJ, Dowling DJ, Gallington LC. Imidazoquinoline Toll-like receptor 8 agonists activate human newborn monocytes and dendritic cells through adenosine-refractory and caspase-1-dependent pathways. J Allergy Clin Immunol. 2012;130(1):195–204.e9. doi: 10.1016/j.jaci.2012.02.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Kingsmore SF, Kennedy N, Halliday HL. Identification of diagnostic biomarkers for infection in premature neonates. Mol Cell Proteomics. 2008;7(10):1863–1875. doi: 10.1074/mcp.M800175-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Hirata J, Kotani J, Aoyama M. A role for IL-18 in human neutrophil apoptosis. Shock. 2008;30(6):628–633. doi: 10.1097/SHK.0b013e31817c0c69. [DOI] [PubMed] [Google Scholar]
- 138.Cusumano V, Midiri A, Cusumano VV. Interleukin-18 is an essential element in host resistance to experimental group B streptococcal disease in neonates. Infect Immun. 2004;72(1):295–300. doi: 10.1128/IAI.72.1.295-300.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Puren AJ, Fantuzzi G, Gu Y. Interleukin-18 (IFNgamma-inducing factor) induces IL-8 and IL-1beta via TNFalpha production from non-CD14+ human blood mononuclear cells. J Clin Invest. 1998;101(3):711–721. doi: 10.1172/JCI1379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Elbim C, Guichard C, Dang PM. Interleukin-18 primes the oxidative burst of neutrophils in response to formyl-peptides: role of cytochrome b558 translocation and N-formyl peptide receptor endocytosis. Clin Diagn Lab Immunol. 2005;12(3):436–446. doi: 10.1128/CDLI.12.3.436-446.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Hagberg H, Mallard C, Jacobsson B. Role of cytokines in preterm labour and brain injury. BJOG. 2005;112(Suppl 1):16–18. doi: 10.1111/j.1471-0528.2005.00578.x. [DOI] [PubMed] [Google Scholar]
- 142.Halpern MD, Holubec H, Dominguez JA. Up-regulation of IL-18 and IL-12 in the ileum of neonatal rats with necrotizing enterocolitis. Pediatr Res. 2002;51(6):733–739. doi: 10.1203/00006450-200206000-00012. [DOI] [PubMed] [Google Scholar]
- 143.Heninger E, Treszl A, Kocsis I. Genetic variants of the interleukin-18 promoter region (-607) influence the course of necrotising enterocolitis in very low birth weight neonates. Eur J Pediatr. 2002;161(7):410–411. doi: 10.1007/s00431-002-0968-y. [DOI] [PubMed] [Google Scholar]
- 144.Halpern MD, Khailova L, Molla-Hosseini D. Decreased development of necrotizing enterocolitis in IL-18-deficient mice. Am J Physiol Gastrointest Liver Physiol. 2008;294(1):G20–G26. doi: 10.1152/ajpgi.00168.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Miller LC, Isa S, LoPreste G. Neonatal interleukin-1 beta, interleukin-6, and tumor necrosis factor: cord blood levels and cellular production. J Pediatr. 1990;117(6):961–965. doi: 10.1016/s0022-3476(05)80145-3. [DOI] [PubMed] [Google Scholar]
- 146.Harris MC, Costarino AT, Jr, Sullivan JS. Cytokine elevations in critically ill infants with sepsis and necrotizing enterocolitis. J Pediatr. 1994;124(1):105–111. doi: 10.1016/s0022-3476(94)70264-0. [DOI] [PubMed] [Google Scholar]
- 147.Girardin EP, Berner ME, Grau GE. Serum tumour necrosis factor in newborns at risk for infections. Eur J Pediatr. 1990;149(9):645–647. doi: 10.1007/BF02034754. [DOI] [PubMed] [Google Scholar]
- 148.Harris MC, D'Angio CT, Gallagher PR. Cytokine elaboration in critically ill infants with bacterial sepsis, necrotizing entercolitis, or sepsis syndrome: correlation with clinical parameters of inflammation and mortality. J Pediatr. 2005;147(4):462–468. doi: 10.1016/j.jpeds.2005.04.037. [DOI] [PubMed] [Google Scholar]
- 149.Ng PC, Li K, Leung TF. Early prediction of sepsis-induced disseminated intravascular coagulation with interleukin-10, interleukin-6, and RANTES in preterm infants. Clin Chem. 2006;52(6):1181–1189. doi: 10.1373/clinchem.2005.062075. [DOI] [PubMed] [Google Scholar]
- 150.Dollner H, Vatten L, Austgulen R. Early diagnostic markers for neonatal sepsis: comparing C-reactive protein, interleukin-6, soluble tumour necrosis factor receptors and soluble adhesion molecules. J Clin Epidemiol. 2001;54(12):1251–1257. doi: 10.1016/s0895-4356(01)00400-0. [DOI] [PubMed] [Google Scholar]
- 151.Turunen R, Andersson S, Nupponen I. Increased CD11b-density on circulating phagocytes as an early sign of late-onset sepsis in extremely low-birth-weight infants. Pediatr Res. 2005;57(2):270–275. doi: 10.1203/01.PDR.0000148717.59861.2C. [DOI] [PubMed] [Google Scholar]
- 152.Figueras-Aloy J, Gomez-Lopez L, Rodriguez-Miguelez JM. Serum soluble ICAM-1, VCAM-1, L-selectin, and P-selectin levels as markers of infection and their relation to clinical severity in neonatal sepsis. Am J Perinatol. 2007;24(6):331–338. doi: 10.1055/s-2007-981851. [DOI] [PubMed] [Google Scholar]
- 153.Kourtis AP, Lee FK, Stoll BJ. Soluble L-selectin, a marker of immune activation, in neonatal infection. Clin Immunol. 2003;109(2):224–228. doi: 10.1016/s1521-6616(03)00209-2. [DOI] [PubMed] [Google Scholar]
- 154.Sullivan SE, Staba SL, Gersting JA. Circulating concentrations of chemokines in cord blood, neonates, and adults. Pediatr Res. 2002;51(5):653–657. doi: 10.1203/00006450-200205000-00018. [DOI] [PubMed] [Google Scholar]
- 155.Meade VM, Barese CN, Kim C. Rac2 concentrations in umbilical cord neutrophils. Biol Neonate. 2006;90(3):156–159. doi: 10.1159/000092451. [DOI] [PubMed] [Google Scholar]
- 156.Hickey MJ, Kubes P. Intravascular immunity: the host-pathogen encounter in blood vessels. Nat Rev Immunol. 2009;9(5):364–375. doi: 10.1038/nri2532. [DOI] [PubMed] [Google Scholar]
- 157.Ng PC, Li K, Chui KM. IP-10 is an early diagnostic marker for identification of late-onset bacterial infection in preterm infants. Pediatr Res. 2007;61(1):93–98. doi: 10.1203/01.pdr.0000250207.95723.96. [DOI] [PubMed] [Google Scholar]
- 158.Wong HR, Cvijanovich N, Wheeler DS. Interleukin-8 as a stratification tool for interventional trials involving pediatric septic shock. Am J Respir Crit Care Med. 2008;178(3):276–282. doi: 10.1164/rccm.200801-131OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Lotze MT, Tracey KJ. High-mobility group box 1 protein (HMGB1): nuclear weapon in the immune arsenal. Nat Rev Immunol. 2005;5(4):331–342. doi: 10.1038/nri1594. [DOI] [PubMed] [Google Scholar]
- 160.Mullins GE, Sunden-Cullberg J, Johansson AS. Activation of human umbilical vein endothelial cells leads to relocation and release of high-mobility group box chromosomal protein 1. Scand J Immunol. 2004;60(6):566–573. doi: 10.1111/j.0300-9475.2004.01518.x. [DOI] [PubMed] [Google Scholar]
- 161.van Zoelen MA, Yang H, Florquin S. Role of toll-like receptors 2 and 4, and the receptor for advanced glycation end products in high-mobility group box 1-induced inflammation in vivo. Shock. 2009;31(3):280–284. doi: 10.1097/SHK.0b013e318186262d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Sappington PL, Yang R, Yang H. HMGB1 B box increases the permeability of Caco-2 enterocytic monolayers and impairs intestinal barrier function in mice. Gastroenterology. 2002;123(3):790–802. doi: 10.1053/gast.2002.35391. [DOI] [PubMed] [Google Scholar]
- 163.Zamora R, Grishin A, Wong C. High-mobility group box 1 protein is an inflammatory mediator in necrotizing enterocolitis: protective effect of the macrophage deactivator semapimod. Am J Physiol Gastrointest Liver Physiol. 2005;289(4):G643–G652. doi: 10.1152/ajpgi.00067.2005. [DOI] [PubMed] [Google Scholar]
- 164.Buhimschi CS, Baumbusch MA, Dulay AT. Characterization of RAGE, HMGB1, and S100beta in inflammation-induced preterm birth and fetal tissue injury. Am J Pathol. 2009;175(3):958–975. doi: 10.2353/ajpath.2009.090156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Vasarhelyi B, Toth-Heyn P, Treszl A, Tulassay T. Genetic polymorphisms and risk for acute renal failure in preterm neonates. Pediatr Nephrol. 2005;20(2):132–135. doi: 10.1007/s00467-004-1711-x. [DOI] [PubMed] [Google Scholar]
- 166.Pack CD, Kumaraguru U, Suvas S, Rouse BT. Heat-shock protein 70 acts as an effective adjuvant in neonatal mice and confers protection against challenge with herpes simplex virus. Vaccine. 2005;23(27):3526–3534. doi: 10.1016/j.vaccine.2005.01.152. [DOI] [PubMed] [Google Scholar]
- 167.Wheeler DS, Lahni P, Odoms K. Extracellular heat shock protein 60 (Hsp60) levels in children with septic shock. Inflamm Res. 2007;56(5):216–219. doi: 10.1007/s00011-007-6108-4. [DOI] [PubMed] [Google Scholar]
- 168.Wheeler DS, Fisher LE, Jr, Catravas JD. Extracellular hsp70 levels in children with septic shock. Pediatr Crit Care Med. 2005;6(3):308–311. doi: 10.1097/01.PCC.0000161075.97355.2E. [DOI] [PubMed] [Google Scholar]
- 169.Kono H, Rock KL. How dying cells alert the immune system to danger. Nat Rev Immunol. 2008;8(4):279–289. doi: 10.1038/nri2215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Batra S, Kumar R, Kapoor AK, Ray G. Alterations in antioxidant status during neonatal sepsis. Ann Trop Paediatr. 2000;20(1):27–33. doi: 10.1080/02724930092039. [DOI] [PubMed] [Google Scholar]
- 171.Kapoor K, Basu S, Das BK, Bhatia BD. Lipid peroxidation and antioxidants in neonatal septicemia. J Trop Pediatr. 2006;52(5):372–375. doi: 10.1093/tropej/fml013. [DOI] [PubMed] [Google Scholar]
- 172.Marshall JC. Such stuff as dreams are made on: mediator-directed therapy in sepsis. Nat Rev Drug Discov. 2003;2(5):391–405. doi: 10.1038/nrd1084. [DOI] [PubMed] [Google Scholar]
- 173.Marom D, Yuhas Y, Sirota L. Nitric oxide levels in preterm and term infants and in premature infants with bacteremia. Biol Neonate. 2004;86(3):160–164. doi: 10.1159/000079375. [DOI] [PubMed] [Google Scholar]
- 174.Schrama AJ, de Beaufort AJ, Poorthuis BJ. Secretory phospholipase A2 in newborn infants with sepsis. J Perinatol. 2008;28(4):291–296. doi: 10.1038/sj.jp.7211929. [DOI] [PubMed] [Google Scholar]
- 175.Siauw C, Kobsar A, Dornieden C. Group B streptococcus isolates from septic patients and healthy carriers differentially activate platelet signaling cascades. Thromb Haemost. 2006;95(5):836–849. doi: 10.1160/th05-08-0534. [DOI] [PubMed] [Google Scholar]
- 176.Figueras-Aloy J, Gomez-Lopez L, Rodriguez-Miguelez JM. Plasma endothelin-1 and clinical manifestations of neonatal sepsis. J Perinat Med. 2004;32(6):522–526. doi: 10.1515/JPM.2004.126. [DOI] [PubMed] [Google Scholar]
- 177.Kuhl PG, Cotton RB, Schweer H, Seyberth HW. Endogenous formation of prostanoids in neonates with persistent pulmonary hypertension. Arch Dis Child. 1989;64(7 Spec No):949–952. doi: 10.1136/adc.64.7_spec_no.949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Rosenberg AA, Kennaugh J, Koppenhafer SL. Elevated immunoreactive endothelin-1 levels in newborn infants with persistent pulmonary hypertension. J Pediatr. 1993;123(1):109–114. doi: 10.1016/s0022-3476(05)81552-5. [DOI] [PubMed] [Google Scholar]
- 179.Rossi P, Persson B, Boels PJ. Endotoxemic pulmonary hypertension is largely mediated by endothelin-induced venous constriction. Intensive Care Med. 2008;34(5):873–880. doi: 10.1007/s00134-007-0980-9. [DOI] [PubMed] [Google Scholar]
- 180.Bulger EM, Maier RV. Lipid mediators in the pathophysiology of critical illness. Crit Care Med. 2000;28(4 Suppl):N27–N36. doi: 10.1097/00003246-200004001-00004. [DOI] [PubMed] [Google Scholar]
- 181.Gotsch F, Romero R, Kusanovic JP. The fetal inflammatory response syndrome. Clin Obstet Gynecol. 2007;50(3):652–683. doi: 10.1097/GRF.0b013e31811ebef6. [DOI] [PubMed] [Google Scholar]
- 182.Shi Y, Li HQ, Shen CK. Plasma nitric oxide levels in newborn infants with sepsis. J Pediatr. 1993;123(3):435–438. doi: 10.1016/s0022-3476(05)81753-6. [DOI] [PubMed] [Google Scholar]
- 183.Kultursay N, Kantar M, Akisu M. Platelet-activating factor concentrations in healthy and septic neonates. Eur J Pediatr. 1999;158(9):740–741. doi: 10.1007/s004310051191. [DOI] [PubMed] [Google Scholar]
- 184.Sriskandan S, Altmann DM. The immunology of sepsis. J Pathol. 2008;214(2):211–223. doi: 10.1002/path.2274. [DOI] [PubMed] [Google Scholar]
- 185.Marshall JC. Why have clinical trials in sepsis failed? Trends Mol Med. 2014;20(4):195–203. doi: 10.1016/j.molmed.2014.01.007. [DOI] [PubMed] [Google Scholar]
- 186.Xiao W, Mindrinos MN, Seok J. A genomic storm in critically injured humans. J Exp Med. 2011;208(13):2581–2590. doi: 10.1084/jem.20111354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Wong HR, Wheeler DS, Tegtmeyer K. Toward a clinically feasible gene expression-based subclassification strategy for septic shock: proof of concept. Crit Care Med. 2010;38(10):1955–1961. doi: 10.1097/CCM.0b013e3181eb924f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Hartel C, Osthues I, Rupp J. Characterisation of the host inflammatory response to Staphylococcus epidermidis in neonatal whole blood. Arch Dis Child Fetal Neonatal Ed. 2008;93(2):F140–F145. doi: 10.1136/adc.2007.124685. [DOI] [PubMed] [Google Scholar]
- 189.Sikora JP, Chlebna-Sokol D, Krzyzanska-Oberbek A. Proinflammatory cytokines (IL-6, IL-8), cytokine inhibitors (IL-6sR, sTNFRII) and anti-inflammatory cytokines (IL-10, IL-13) in the pathogenesis of sepsis in newborns and infants. Arch Immunol Ther Exp (Warsz) 2001;49(5):399–404. [PubMed] [Google Scholar]
- 190.Opal SM, Esmon CT. Bench-to-bedside review: functional relationships between coagulation and the innate immune response and their respective roles in the pathogenesis of sepsis. Crit Care. 2003;7(1):23–38. doi: 10.1186/cc1854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Wang P, Wu P, Siegel MI. Interleukin (IL)-10 inhibits nuclear factor κB (NFκB) activation in human monocytes. IL-10 and IL-4 suppress cytokine synthesis by different mechanisms. J Biol Chem. 1995;270(16):9558–9563. doi: 10.1074/jbc.270.16.9558. [DOI] [PubMed] [Google Scholar]
- 192.Brubaker JO, Montaner LJ. Role of interleukin-13 in innate and adaptive immunity. Cell Mol Biol (Noisy-Le-Grand) 2001;47(4):637–651. [PubMed] [Google Scholar]
- 193.Trepicchio WL, Bozza M, Pedneault G, Dorner AJ. Recombinant human IL-11 attenuates the inflammatory response through down-regulation of proinflammatory cytokine release and nitric oxide production. J Immunol. 1996;157(8):3627–3634. [PubMed] [Google Scholar]
- 194.Koj A. Termination of acute-phase response: role of some cytokines and anti-inflammatory drugs. Gen Pharmacol. 1998;31(1):9–18. doi: 10.1016/s0306-3623(97)00435-7. [DOI] [PubMed] [Google Scholar]
- 195.Andrade EB, Alves J, Madureira P. TLR2-induced IL-10 production impairs neutrophil recruitment to infected tissues during neonatal bacterial sepsis. J Immunol. 2013;191(9):4759–4768. doi: 10.4049/jimmunol.1301752. [DOI] [PubMed] [Google Scholar]
- 196.Spear ML, Stefano JL, Fawcett P, Proujansky R. Soluble interleukin-2 receptor as a predictor of neonatal sepsis. J Pediatr. 1995;126(6):982–985. doi: 10.1016/s0022-3476(95)70228-8. [DOI] [PubMed] [Google Scholar]
- 197.Dollner H, Vatten L, Linnebo I. Inflammatory mediators in umbilical plasma from neonates who develop early-onset sepsis. Biol Neonate. 2001;80(1):41–47. doi: 10.1159/000047118. [DOI] [PubMed] [Google Scholar]
- 198.Buhimschi CS, Bhandari V, Han YW. Using proteomics in perinatal and neonatal sepsis: hopes and challenges for the future. Curr Opin Infect Dis. 2009;22(3):235–243. doi: 10.1097/QCO.0b013e32832a5963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Bopp C, Hofer S, Weitz J. sRAGE is elevated in septic patients and associated with patients outcome. J Surg Res. 2008;147(1):79–83. doi: 10.1016/j.jss.2007.07.014. [DOI] [PubMed] [Google Scholar]
- 200.Liliensiek B, Weigand MA, Bierhaus A. Receptor for advanced glycation end products (RAGE) regulates sepsis but not the adaptive immune response. J Clin Invest. 2004;113(11):1641–1650. doi: 10.1172/JCI18704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Adly AA, Ismail EA, Andrawes NG, El-Saadany MA. Circulating soluble triggering receptor expressed on myeloid cells-1 (sTREM-1) as diagnostic and prognostic marker in neonatal sepsis. Cytokine. 2014;65(2):184–191. doi: 10.1016/j.cyto.2013.11.004. [DOI] [PubMed] [Google Scholar]
- 202.Ameres SL, Zamore PD. Diversifying microRNA sequence and function. Nat Rev Mol Cell Biol. 2013;14(8):475–488. doi: 10.1038/nrm3611. [DOI] [PubMed] [Google Scholar]
- 203.Palin AC, Ramachandran V, Acharya S, Lewis DB. Human neonatal naive CD4+ T cells have enhanced activation-dependent signaling regulated by the microRNA miR-181a. J Immunol. 2013;190(6):2682–2691. doi: 10.4049/jimmunol.1202534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Bhaskaran M, Xi D, Wang Y. Identification of microRNAs changed in the neonatal lungs in response to hyperoxia exposure. Physiol Genomics. 2012;44(20):970–980. doi: 10.1152/physiolgenomics.00145.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Huang HC, Yu HR, Huang LT. miRNA-125b regulates TNF-alpha production in CD14+ neonatal monocytes via post-transcriptional regulation. J Leukoc Biol. 2012;92(1):171–182. doi: 10.1189/jlb.1211593. [DOI] [PubMed] [Google Scholar]
- 206.Lederhuber H, Baer K, Altiok I. MicroRNA-146: tiny player in neonatal innate immunity? Neonatology. 2011;99(1):51–56. doi: 10.1159/000301938. [DOI] [PubMed] [Google Scholar]
- 207.Chassin C, Kocur M, Pott J. miR-146a mediates protective innate immune tolerance in the neonate intestine. Cell Host Microbe. 2010;8(4):358–368. doi: 10.1016/j.chom.2010.09.005. [DOI] [PubMed] [Google Scholar]
- 208.Chen RF, Huang HC, Ou CY. MicroRNA-21 expression in neonatal blood associated with antenatal immunoglobulin E production and development of allergic rhinitis. Clin Exp Allergy. 2010;40(10):1482–1490. doi: 10.1111/j.1365-2222.2010.03592.x. [DOI] [PubMed] [Google Scholar]
- 209.Annane D, Bellissant E, Bollaert PE. Corticosteroids for severe sepsis and septic shock: a systematic review and meta-analysis. BMJ. 2004;329(7464):480. doi: 10.1136/bmj.38181.482222.55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Faber J, Meyer CU, Gemmer C. Human toll-like receptor 4 mutations are associated with susceptibility to invasive meningococcal disease in infancy. Pediatr Infect Dis J. 2006;25(1):80–81. doi: 10.1097/01.inf.0000195595.22547.fe. [DOI] [PubMed] [Google Scholar]
- 211.Behrendt D, Dembinski J, Heep A, Bartmann P. Lipopolysaccharide binding protein in preterm infants. Arch Dis Child Fetal Neonatal Ed. 2004;89(6):F551–F554. doi: 10.1136/adc.2003.030049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Togari H, Sugiyama S, Ogino T. Interactions of endotoxin with cortisol and acute phase proteins in septic shock neonates. Acta Paediatr Scand. 1986;75(1):69–74. doi: 10.1111/j.1651-2227.1986.tb10159.x. [DOI] [PubMed] [Google Scholar]
- 213.Fernandez EF, Watterberg KL. Relative adrenal insufficiency in the preterm and term infant. J Perinatol. 2009;29(Suppl 2):S44–S49. doi: 10.1038/jp.2009.24. [DOI] [PubMed] [Google Scholar]
- 214.Wong HR, Cvijanovich NZ, Allen GL. Corticosteroids are associated with repression of adaptive immunity gene programs in pediatric septic shock. Am J Respir Crit Care Med. 2014;189(8):940–946. doi: 10.1164/rccm.201401-0171OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Bizzarro MJ, Jiang Y, Hussain N. The impact of environmental and genetic factors on neonatal late-onset sepsis. J Pediatr. 2011;158(2):234–238.e1. doi: 10.1016/j.jpeds.2010.07.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Person MK, Esposito DH, Holman RC. Risk factors for infectious disease death among infants in the United States. Pediatr Infect Dis J. 2014;33(11):e280–e285. doi: 10.1097/INF.0000000000000414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Zhou JG, Zhang JP, Lee BH. Toll-like receptor 4 polymorphisms in gram-negative bacterial infections of Han Chinese neonates. Am J Perinatol. 2015;32(4):363–370. doi: 10.1055/s-0034-1387929. [DOI] [PubMed] [Google Scholar]
- 218.Abu-Maziad A, Schaa K, Bell EF. Role of polymorphic variants as genetic modulators of infection in neonatal sepsis. Pediatr Res. 2010;68(4):323–329. doi: 10.1203/PDR.0b013e3181e6a068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Koroglu OA, Onay H, Erdemir G. Mannose-binding lectin gene polymorphism and early neonatal outcome in preterm infants. Neonatology. 2010;98(4):305–312. doi: 10.1159/000291487. [DOI] [PubMed] [Google Scholar]
- 220.Baier RJ, Loggins J, Yanamandra K. IL-10, IL-6 and CD14 polymorphisms and sepsis outcome in ventilated very low birth weight infants. BMC Med. 2006;4:10. doi: 10.1186/1741-7015-4-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Hedberg CL, Adcock K, Martin J. Tumor necrosis factor alpha−308 polymorphism associated with increased sepsis mortality in ventilated very low birth weight infants. Pediatr Infect Dis J. 2004;23(5):424–428. doi: 10.1097/01.inf.0000122607.73324.20. [DOI] [PubMed] [Google Scholar]
- 222.Sampath V, Mulrooney NP, Garland JS. Toll-like receptor genetic variants are associated with Gram-negative infections in VLBW infants. J Perinatol. 2013;33(10):772–777. doi: 10.1038/jp.2013.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Bochud PY, Chien JW, Marr KA. Toll-like receptor 4 polymorphisms and aspergillosis in stem-cell transplantation. N Engl J Med. 2008;359(17):1766–1777. doi: 10.1056/NEJMoa0802629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Wurfel MM, Gordon AC, Holden TD. Toll-like receptor 1 polymorphisms affect innate immune responses and outcomes in sepsis. Am J Respir Crit Care Med. 2008;178(7):710–720. doi: 10.1164/rccm.200803-462OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Agnese DM, Calvano JE, Hahm SJ. Human toll-like receptor 4 mutations but not CD14 polymorphisms are associated with an increased risk of gram-negative infections. J Infect Dis. 2002;186(10):1522–1525. doi: 10.1086/344893. [DOI] [PubMed] [Google Scholar]
- 226.Lorenz E, Mira JP, Cornish KL. A novel polymorphism in the toll-like receptor 2 gene and its potential association with staphylococcal infection. Infect Immun. 2000;68(11):6398–6401. doi: 10.1128/iai.68.11.6398-6401.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Zhang SY, Jouanguy E, Ugolini S. TLR3 deficiency in patients with herpes simplex encephalitis. Science. 2007;317(5844):1522–1527. doi: 10.1126/science.1139522. [DOI] [PubMed] [Google Scholar]
- 228.Mockenhaupt FP, Cramer JP, Hamann L. Toll-like receptor (TLR) polymorphisms in African children: common TLR-4 variants predispose to severe malaria. J Commun Dis. 2006;38(3):230–245. [PubMed] [Google Scholar]
- 229.Berner R, Furll B, Stelter F. Elevated levels of lipopolysaccharide-binding protein and soluble CD14 in plasma in neonatal early-onset sepsis. Clin Diagn Lab Immunol. 2002;9(2):440–445. doi: 10.1128/CDLI.9.2.440-445.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Blanco A, Solis G, Arranz E. Serum levels of CD14 in neonatal sepsis by Gram-positive and Gram-negative bacteria. Acta Paediatr. 1996;85(6):728–732. doi: 10.1111/j.1651-2227.1996.tb14135.x. [DOI] [PubMed] [Google Scholar]
- 231.Hartel C, Rupp J, Hoegemann A. 159C>T CD14 genotype–functional effects on innate immune responses in term neonates. Hum Immunol. 2008;69(6):338–343. doi: 10.1016/j.humimm.2008.04.011. [DOI] [PubMed] [Google Scholar]
- 232.Mollen KP, Gribar SC, Anand RJ. Increased expression and internalization of the endotoxin coreceptor CD14 in enterocytes occur as an early event in the development of experimental necrotizing enterocolitis. J Pediatr Surg. 2008;43(6):1175–1181. doi: 10.1016/j.jpedsurg.2008.02.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Hubacek JA, Stuber F, Frohlich D. Gene variants of the bactericidal/permeability increasing protein and lipopolysaccharide binding protein in sepsis patients: gender-specific genetic predisposition to sepsis. Crit Care Med. 2001;29(3):557–561. doi: 10.1097/00003246-200103000-00015. [DOI] [PubMed] [Google Scholar]
- 234.Gu W, Shan YA, Zhou J. Functional significance of gene polymorphisms in the promoter of myeloid differentiation-2. Ann Surg. 2007;246(1):151–158. doi: 10.1097/01.sla.0000262788.67171.3f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.von Bernuth H, Picard C, Jin Z. Pyogenic bacterial infections in humans with MyD88 deficiency. Science. 2008;321(5889):691–696. doi: 10.1126/science.1158298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Picard C, Puel A, Bonnet M. Pyogenic bacterial infections in humans with IRAK-4 deficiency. Science. 2003;299(5615):2076–2079. doi: 10.1126/science.1081902. [DOI] [PubMed] [Google Scholar]
- 237.Ku CL, Picard C, Erdos M. IRAK4 and NEMO mutations in otherwise healthy children with recurrent invasive pneumococcal disease. J Med Genet. 2007;44(1):16–23. doi: 10.1136/jmg.2006.044446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Ogura Y, Bonen DK, Inohara N. A frameshift mutation in NOD2 associated with susceptibility to Crohn's disease. Nature. 2001;411(6837):603–606. doi: 10.1038/35079114. [DOI] [PubMed] [Google Scholar]
- 239.Goldbach-Mansky R, Dailey NJ, Canna SW. Neonatal-onset multisystem inflammatory disease responsive to interleukin-1β inhibition. N Engl J Med. 2006;355(6):581–592. doi: 10.1056/NEJMoa055137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Nybo M, Sorensen O, Leslie R, Wang P. Reduced expression of C5a receptors on neutrophils from cord blood. Arch Dis Child Fetal Neonatal Ed. 1998;78(2):F129–F132. doi: 10.1136/fn.78.2.f129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Ahrens P, Kattner E, Kohler B. Mutations of genes involved in the innate immune system as predictors of sepsis in very low birth weight infants. Pediatr Res. 2004;55(4):652–656. doi: 10.1203/01.PDR.0000112100.61253.85. [DOI] [PubMed] [Google Scholar]
- 242.Chauhan M, McGuire W. Interleukin-6 (-174C) polymorphism and the risk of sepsis in very low birth weight infants: meta-analysis. Arch Dis Child Fetal Neonatal Ed. 2008;93(6):F427–F429. doi: 10.1136/adc.2007.134205. [DOI] [PubMed] [Google Scholar]
- 243.Dzwonek AB, Neth OW, Thiebaut R. The role of mannose-binding lectin in susceptibility to infection in preterm neonates. Pediatr Res. 2008;63(6):680–685. doi: 10.1203/PDR.0b013e31816fdbff. [DOI] [PubMed] [Google Scholar]
- 244.Gopel W, Hartel C, Ahrens P. Interleukin-6-174-genotype, sepsis and cerebral injury in very low birth weight infants. Genes Immun. 2006;7(1):65–68. doi: 10.1038/sj.gene.6364264. [DOI] [PubMed] [Google Scholar]
- 245.Hartel C, Schultz C, Herting E, Gopel W. Genetic association studies in VLBW infants exemplifying susceptibility to sepsis—recent findings and implications for future research. Acta Paediatr. 2007;96(2):158–165. doi: 10.1111/j.1651-2227.2007.00128.x. [DOI] [PubMed] [Google Scholar]
- 246.Reiman M, Kujari H, Ekholm E. Interleukin-6 polymorphism is associated with chorioamnionitis and neonatal infections in preterm infants. J Pediatr. 2008;153(1):19–24. doi: 10.1016/j.jpeds.2008.02.009. [DOI] [PubMed] [Google Scholar]
- 247.Schueller AC, Heep A, Kattner E. Prevalence of two tumor necrosis factor gene polymorphisms in premature infants with early onset sepsis. Biol Neonate. 2006;90(4):229–232. doi: 10.1159/000093605. [DOI] [PubMed] [Google Scholar]
- 248.Treszl A, Heninger E, Kalman A. Lower prevalence of IL-4 receptor alpha-chain gene G variant in very-low-birth-weight infants with necrotizing enterocolitis. J Pediatr Surg. 2003;38(9):1374–1378. doi: 10.1016/s0022-3468(03)00399-3. [DOI] [PubMed] [Google Scholar]
- 249.Treszl A, Kocsis I, Szathmari M. Genetic variants of TNF-α, IL-1β, IL-4 receptor α-chain, IL-6 and IL-10 genes are not risk factors for sepsis in low-birth-weight infants. Biol Neonate. 2003;83(4):241–245. doi: 10.1159/000069484. [DOI] [PubMed] [Google Scholar]
- 250.Treszl A, Kocsis I, Szathmari M. Genetic variants of the tumour necrosis factor-alpha promoter gene do not influence the development of necrotizing enterocolitis. Acta Paediatr. 2001;90(10):1182–1185. doi: 10.1111/j.1651-2227.2001.tb03251.x. [DOI] [PubMed] [Google Scholar]
- 251.Treszl A, Tulassay T, Vasarhelyi B. Genetic basis for necrotizing enterocolitis—risk factors and their relations to genetic polymorphisms. Front Biosci. 2006;11:570–580. doi: 10.2741/1819. [DOI] [PubMed] [Google Scholar]
- 252.Carvalho JK, Moore DB, Luz RA. Prediction of sepsis-related outcomes in neonates through systematic genotyping of polymorphisms in genes for innate immunity and inflammation: a narrative review and critical perspective. Sao Paulo Med J. 2013;131(5):338–350. doi: 10.1590/1516-3180.2013.1315519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253.Wolach B, Dolfin T, Regev R. The development of the complement system after 28 weeks' gestation. Acta Paediatr. 1997;86(5):523–527. doi: 10.1111/j.1651-2227.1997.tb08924.x. [DOI] [PubMed] [Google Scholar]
- 254.Notarangelo LD, Chirico G, Chiara A. Activity of classical and alternative pathways of complement in preterm and small for gestational age infants. Pediatr Res. 1984;18(3):281–285. doi: 10.1203/00006450-198403000-00014. [DOI] [PubMed] [Google Scholar]
- 255.Drossou V, Kanakoudi F, Diamanti E. Concentrations of main serum opsonins in early infancy. Arch Dis Child Fetal Neonatal Ed. 1995;72(3):F172–F175. doi: 10.1136/fn.72.3.f172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Miller ME, Stiehm ER. Phagocytic, opsonic and immunoglobulin studies in newborns. Calif Med. 1973;119(2):43–63. [PMC free article] [PubMed] [Google Scholar]
- 257.Nupponen I, Andersson S, Jarvenpaa AL. Neutrophil CD11b expression and circulating interleukin-8 as diagnostic markers for early-onset neonatal sepsis. Pediatrics. 2001;108(1):E12. doi: 10.1542/peds.108.1.e12. [DOI] [PubMed] [Google Scholar]
- 258.Berger M, O'Shea J, Cross AS. Human neutrophils increase expression of C3bi as well as C3b receptors upon activation. J Clin Invest. 1984;74(5):1566–15671. doi: 10.1172/JCI111572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259.Snyderman R, Goetzl EJ. Molecular and cellular mechanisms of leukocyte chemotaxis. Science. 1981;213(4510):830–837. doi: 10.1126/science.6266014. [DOI] [PubMed] [Google Scholar]
- 260.Vogt W. Anaphylatoxins: possible roles in disease. Complement. 1986;3(3):177–188. doi: 10.1159/000467894. [DOI] [PubMed] [Google Scholar]
- 261.Markiewski MM, Lambris JD. The role of complement in inflammatory diseases from behind the scenes into the spotlight. Am J Pathol. 2007;171(3):715–727. doi: 10.2353/ajpath.2007.070166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Unnewehr H, Rittirsch D, Sarma JV. Changes and regulation of the C5a receptor on neutrophils during septic shock in humans. J Immunol. 2013;190(8):4215–4225. doi: 10.4049/jimmunol.1200534. [DOI] [PubMed] [Google Scholar]
- 263.Lothian C, Dahlgren C, Lagercrantz H, Lundahl J. Different expression and mobilisation of the complement regulatory proteins CD35, CD55 and CD59 in neonatal and adult neutrophils. Biol Neonate. 1997;72(1):15–21. doi: 10.1159/000244461. [DOI] [PubMed] [Google Scholar]
- 264.Castellheim A, Lindenskov PH, Pharo A. Meconium aspiration syndrome induces complement-associated systemic inflammatory response in newborn piglets. Scand J Immunol. 2005;61(3):217–225. doi: 10.1111/j.1365-3083.2005.01532.x. [DOI] [PubMed] [Google Scholar]
- 265.Wong HR, Doughty LA, Wedel N. Plasma bactericidal/permeability-increasing protein concentrations in critically ill children with the sepsis syndrome. Pediatr Infect Dis J. 1995;14(12):1087–1091. doi: 10.1097/00006454-199512000-00011. [DOI] [PubMed] [Google Scholar]
- 266.Levy O. Innate immunity of the newborn: basic mechanisms and clinical correlates. Nat Rev Immunol. 2007;7(5):379–390. doi: 10.1038/nri2075. [DOI] [PubMed] [Google Scholar]
- 267.Romeo MG, Tina LG, Sciacca A. [Decreased plasma fibronectin (pFN) level in preterm infants with infections] Pediatr Med Chir. 1995;17(6):563–566. [PubMed] [Google Scholar]
- 268.Kalayci AG, Adam B, Yilmazer F. The value of immunoglobulin and complement levels in the early diagnosis of neonatal sepsis. Acta Paediatr. 1997;86(9):999–1002. doi: 10.1111/j.1651-2227.1997.tb15187.x. [DOI] [PubMed] [Google Scholar]
- 269.Dyke MP, Forsyth KD. Decreased plasma fibronectin concentrations in preterm infants with septicaemia. Arch Dis Child. 1993;68(5 Spec No):557–560. doi: 10.1136/adc.68.5_spec_no.557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 270.Buhimschi CS, Bhandari V, Dulay AT. Proteomics mapping of cord blood identifies haptoglobin “switch-on” pattern as biomarker of early-onset neonatal sepsis in preterm newborns. PLoS ONE. 2011;6(10):e26111. doi: 10.1371/journal.pone.0026111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Benitz WE, Han MY, Madan A, Ramachandra P. Serial serum C-reactive protein levels in the diagnosis of neonatal infection. Pediatrics. 1998;102(4):E41. doi: 10.1542/peds.102.4.e41. [DOI] [PubMed] [Google Scholar]
- 272.Romagnoli C, Frezza S, Cingolani A. Plasma levels of interleukin-6 and interleukin-10 in preterm neonates evaluated for sepsis. Eur J Pediatr. 2001;160(6):345–350. doi: 10.1007/pl00008445. [DOI] [PubMed] [Google Scholar]
- 273.Arnon S, Litmanovitz I, Regev R. The prognostic virtue of inflammatory markers during late-onset sepsis in preterm infants. J Perinat Med. 2004;32(2):176–180. doi: 10.1515/JPM.2004.032. [DOI] [PubMed] [Google Scholar]
- 274.Malek A, Sager R, Schneider H. Maternal-fetal transport of immunoglobulin G and its subclasses during the third trimester of human pregnancy. Am J Reprod Immunol. 1994;32(1):8–14. doi: 10.1111/j.1600-0897.1994.tb00873.x. [DOI] [PubMed] [Google Scholar]
- 275.Krediet TG, Beurskens FJ, van Dijk H. Antibody responses and opsonic activity in sera of preterm neonates with coagulase-negative staphylococcal septicemia and the effect of the administration of fresh frozen plasma. Pediatr Res. 1998;43(5):645–651. doi: 10.1203/00006450-199805000-00013. [DOI] [PubMed] [Google Scholar]
- 276.Madden NP, Levinsky RJ, Bayston R. Surgery, sepsis, and nonspecific immune function in neonates. J Pediatr Surg. 1989;24(6):562–566. doi: 10.1016/s0022-3468(89)80506-8. [DOI] [PubMed] [Google Scholar]
- 277.Currie AJ, Curtis S, Strunk T. Preterm infants have deficient monocyte and lymphocyte cytokine responses to group B streptococcus. Infect Immun. 2011;79(4):1588–1596. doi: 10.1128/IAI.00535-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 278.Wynn JL, Seed PC, Cotten CM. Does IVIg administration yield improved immune function in very premature neonates? J Perinatol. 2010;30(10):635–642. doi: 10.1038/jp.2009.197. [DOI] [PubMed] [Google Scholar]
- 279.Benjamin DK, Jr, Schelonka R, White R. A blinded, randomized, multicenter study of an intravenous Staphylococcus aureus immune globulin. J Perinatol. 2006;26(5):290. doi: 10.1038/sj.jp.7211496. [DOI] [PubMed] [Google Scholar]
- 280.Brocklehurst P, Farrell B, King A. Treatment of neonatal sepsis with intravenous immune globulin. N Engl J Med. 2011;365(13):1201–1211. doi: 10.1056/NEJMoa1100441. [DOI] [PubMed] [Google Scholar]
- 281.DeJonge M, Burchfield D, Bloom B. Clinical trial of safety and efficacy of INH-A21 for the prevention of nosocomial staphylococcal bloodstream infection in premature infants. J Pediatr. 2007;151(3):260–265.e1. doi: 10.1016/j.jpeds.2007.04.060. [DOI] [PubMed] [Google Scholar]
- 282.Weisman LE, Thackray HM, Garcia-Prats JA. Phase 1/2 double-blind, placebo-controlled, dose escalation, safety, and pharmacokinetic study of pagibaximab (BSYX-A110), an antistaphylococcal monoclonal antibody for the prevention of staphylococcal bloodstream infections, in very-low-birth-weight neonates. Antimicrob Agents Chemother. 2009;53(7):2879–2886. doi: 10.1128/AAC.01565-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 283.Akdag A, Dilmen U, Haque K. Role of pentoxifylline and/or IgM-enriched intravenous immunoglobulin in the management of neonatal sepsis. Am J Perinatol. 2014;31(10):905–912. doi: 10.1055/s-0033-1363771. [DOI] [PubMed] [Google Scholar]
- 284.Ohlsson A, Lacy JB. Intravenous immunoglobulin for preventing infection in preterm and/or low birth weight infants. Cochrane Database Syst Rev. 2013;(7) doi: 10.1002/14651858.CD000361.pub3. CD000361. [DOI] [PubMed] [Google Scholar]
- 285.Ohlsson A, Lacy JB. Intravenous immunoglobulin for suspected or proven infection in neonates. Cochrane Database Syst Rev. 2013;(7) doi: 10.1002/14651858.CD001239.pub4. CD001239. [DOI] [PubMed] [Google Scholar]
- 286.Ganz T. Defensins: antimicrobial peptides of innate immunity. Nat Rev Immunol. 2003;3(9):710–720. doi: 10.1038/nri1180. [DOI] [PubMed] [Google Scholar]
- 287.Yoshio H, Lagercrantz H, Gudmundsson GH, Agerberth B. First line of defense in early human life. Semin Perinatol. 2004;28(4):304–311. doi: 10.1053/j.semperi.2004.08.008. [DOI] [PubMed] [Google Scholar]
- 288.Levy O, Sisson RB, Kenyon J. Enhancement of neonatal innate defense: effects of adding an N-terminal recombinant fragment of bactericidal/permeability-increasing protein on growth and tumor necrosis factor-inducing activity of gram-negative bacteria tested in neonatal cord blood ex vivo. Infect Immun. 2000;68(9):5120–5125. doi: 10.1128/iai.68.9.5120-5125.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289.Motzkus D, Schulz-Maronde S, Heitland A. The novel beta-defensin DEFB123 prevents lipopolysaccharide-mediated effects in vitro and in vivo. FASEB J. 2006;20(10):1701–1702. doi: 10.1096/fj.05-4970fje. [DOI] [PubMed] [Google Scholar]
- 290.Nagaoka I, Hirota S, Niyonsaba F. Augmentation of the lipopolysaccharide-neutralizing activities of human cathelicidin CAP18/LL-37-derived antimicrobial peptides by replacement with hydrophobic and cationic amino acid residues. Clin Diagn Lab Immunol. 2002;9(5):972–982. doi: 10.1128/CDLI.9.5.972-982.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Reinhart K, Gluck T, Ligtenberg J. CD14 receptor occupancy in severe sepsis: results of a phase I clinical trial with a recombinant chimeric CD14 monoclonal antibody (IC14) Crit Care Med. 2004;32(5):1100–1108. doi: 10.1097/01.ccm.0000124870.42312.c4. [DOI] [PubMed] [Google Scholar]
- 292.Levy O, Martin S, Eichenwald E. Impaired innate immunity in the newborn: newborn neutrophils are deficient in bactericidal/permeability-increasing protein. Pediatrics. 1999;104(6):1327–1333. doi: 10.1542/peds.104.6.1327. [DOI] [PubMed] [Google Scholar]
- 293.Nupponen I, Turunen R, Nevalainen T. Extracellular release of bactericidal/permeability-increasing protein in newborn infants. Pediatr Res. 2002;51(6):670–674. doi: 10.1203/00006450-200206000-00002. [DOI] [PubMed] [Google Scholar]
- 294.Michalek J, Svetlikova P, Fedora M. Bactericidal permeability increasing protein gene variants in children with sepsis. Intensive Care Med. 2007;33(12):2158–2164. doi: 10.1007/s00134-007-0860-3. [DOI] [PubMed] [Google Scholar]
- 295.Urlichs F, Speer CP. Neutrophil function in preterm and term infants. NeoReviews. 2004;5:e417–e430. [Google Scholar]
- 296.Kjeldsen L, Sengelov H, Lollike K, Borregaard N. Granules and secretory vesicles in human neonatal neutrophils. Pediatr Res. 1996;40(1):120–129. doi: 10.1203/00006450-199607000-00021. [DOI] [PubMed] [Google Scholar]
- 297.Giroir BP, Scannon PJ, Levin M. Bactericidal/permeability-increasing protein—lessons learned from the phase III, randomized, clinical trial of rBPI21 for adjunctive treatment of children with severe meningococcemia. Crit Care Med. 2001;29(7 Suppl):S130–S135. doi: 10.1097/00003246-200107001-00039. [DOI] [PubMed] [Google Scholar]
- 298.Yamauchi K, Tomita M, Giehl TJ, Ellison RT., 3rd. Antibacterial activity of lactoferrin and a pepsin-derived lactoferrin peptide fragment. Infect Immun. 1993;61(2):719–728. doi: 10.1128/iai.61.2.719-728.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 299.Vogel HJ. Lactoferrin, a bird's eye view. Biochem Cell Biol. 2012;90(3):233–244. doi: 10.1139/o2012-016. [DOI] [PubMed] [Google Scholar]
- 300.Mariller C, Hardiville S, Hoedt E. Proteomic approach to the identification of novel delta-lactoferrin target genes: characterization of DcpS, an mRNA scavenger decapping enzyme. Biochimie. 2009;91(1):109–122. doi: 10.1016/j.biochi.2008.07.009. [DOI] [PubMed] [Google Scholar]
- 301.Lopez A, Lorente JA, Steingrub J. Multiple-center, randomized, placebo-controlled, double-blind study of the nitric oxide synthase inhibitor 546C88: effect on survival in patients with septic shock. Crit Care Med. 2004;32(1):21–30. doi: 10.1097/01.CCM.0000105581.01815.C6. [DOI] [PubMed] [Google Scholar]
- 302.Peters K, Unger RE, Brunner J, Kirkpatrick CJ. Molecular basis of endothelial dysfunction in sepsis. Cardiovasc Res. 2003;60(1):49–57. doi: 10.1016/s0008-6363(03)00397-3. [DOI] [PubMed] [Google Scholar]
- 303.Cribbs SK, Martin GS, Rojas M. Monitoring of endothelial dysfunction in critically ill patients: the role of endothelial progenitor cells. Curr Opin Crit Care. 2008;14(3):354–360. doi: 10.1097/MCC.0b013e3282fc216d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 304.Ayabe T, Ashida T, Kohgo Y, Kono T. The role of Paneth cells and their antimicrobial peptides in innate host defense. Trends Microbiol. 2004;12(8):394–398. doi: 10.1016/j.tim.2004.06.007. [DOI] [PubMed] [Google Scholar]
- 305.Holzl MA, Hofer J, Steinberger P. Host antimicrobial proteins as endogenous immunomodulators. Immunol Lett. 2008;119(1–2):4–11. doi: 10.1016/j.imlet.2008.05.003. [DOI] [PubMed] [Google Scholar]
- 306.Selsted ME, Ouellette AJ. Mammalian defensins in the antimicrobial immune response. Nat Immunol. 2005;6(6):551–557. doi: 10.1038/ni1206. [DOI] [PubMed] [Google Scholar]
- 307.Strunk T, Doherty D, Richmond P. Reduced levels of antimicrobial proteins and peptides in human cord blood plasma. Arch Dis Child Fetal Neonatal Ed. 2009;94(3):F230–F231. doi: 10.1136/adc.2008.143438. [DOI] [PubMed] [Google Scholar]
- 308.Olbrich P, Pavon A, Rosso ML. Association of human beta-defensin-2 serum levels and sepsis in preterm neonates*. Pediatr Crit Care Med. 2013;14(8):796–800. doi: 10.1097/PCC.0b013e3182975e0f. [DOI] [PubMed] [Google Scholar]
- 309.Ihi T, Nakazato M, Mukae H, Matsukura S. Elevated concentrations of human neutrophil peptides in plasma, blood, and body fluids from patients with infections. Clin Infect Dis. 1997;25(5):1134–1140. doi: 10.1086/516075. [DOI] [PubMed] [Google Scholar]
- 310.Thomas NJ, Carcillo JA, Doughty LA. Plasma concentrations of defensins and lactoferrin in children with severe sepsis. Pediatr Infect Dis J. 2002;21(1):34–38. doi: 10.1097/00006454-200201000-00008. [DOI] [PubMed] [Google Scholar]
- 311.Aird WC. Vascular bed-specific hemostasis: role of endothelium in sepsis pathogenesis. Crit Care Med. 2001;29(7 Suppl):S28–S34. doi: 10.1097/00003246-200107001-00013. discussion S35. [DOI] [PubMed] [Google Scholar]
- 312.Kuhle S, Male C, Mitchell L. Developmental hemostasis: pro- and anticoagulant systems during childhood. Semin Thromb Hemost. 2003;29(4):329–338. doi: 10.1055/s-2003-42584. [DOI] [PubMed] [Google Scholar]
- 313.Rivers RP, Cattermole HE, Wright I. The expression of surface tissue factor apoprotein by blood monocytes in the course of infections in early infancy. Pediatr Res. 1992;31(6):567–573. doi: 10.1203/00006450-199206000-00006. [DOI] [PubMed] [Google Scholar]
- 314.Markiewski MM, Nilsson B, Ekdahl KN. Complement and coagulation: strangers or partners in crime? Trends Immunol. 2007;28(4):184–192. doi: 10.1016/j.it.2007.02.006. [DOI] [PubMed] [Google Scholar]
- 315.Aronis S, Platokouki H, Photopoulos S. Indications of coagulation and/or fibrinolytic system activation in healthy and sick very-low-birth-weight neonates. Biol Neonate. 1998;74(5):337–344. doi: 10.1159/000014051. [DOI] [PubMed] [Google Scholar]
- 316.Roman J, Velasco F, Fernandez F. Coagulation, fibrinolytic and kallikrein systems in neonates with uncomplicated sepsis and septic shock. Haemostasis. 1993;23(3):142–148. doi: 10.1159/000216867. [DOI] [PubMed] [Google Scholar]
- 317.Lauterbach R, Pawlik D, Radziszewska R. Plasma antithrombin III and protein C levels in early recognition of late-onset sepsis in newborns. Eur J Pediatr. 2006;165(9):585–589. doi: 10.1007/s00431-006-0139-7. [DOI] [PubMed] [Google Scholar]
- 318.El Beshlawy A, Alaraby I, Abou Hussein H. Study of protein C, protein S, and antithrombin III in newborns with sepsis. Pediatr Crit Care Med. 2010;11(1):52–59. doi: 10.1097/PCC.0b013e3181c59032. [DOI] [PubMed] [Google Scholar]
- 319.Venkataseshan S, Dutta S, Ahluwalia J, Narang A. Low plasma protein C values predict mortality in low birth weight neonates with septicemia. Pediatr Infect Dis J. 2007;26(8):684–688. doi: 10.1097/INF.0b013e3180f616f0. [DOI] [PubMed] [Google Scholar]
- 320.Nadel S, Goldstein B, Williams MD. Drotrecogin alfa (activated) in children with severe sepsis: a multicentre phase III randomised controlled trial. Lancet. 2007;369(9564):836–843. doi: 10.1016/S0140-6736(07)60411-5. [DOI] [PubMed] [Google Scholar]
- 321.Green J, Doughty L, Kaplan SS. The tissue factor and plasminogen activator inhibitor type-1 response in pediatric sepsis-induced multiple organ failure. Thromb Haemost. 2002;87(2):218–223. [PubMed] [Google Scholar]
- 322.Sola MC, Del Vecchio A, Rimsza LM. Evaluation and treatment of thrombocytopenia in the neonatal intensive care unit. Clin Perinatol. 2000;27(3):655–679. doi: 10.1016/s0095-5108(05)70044-0. [DOI] [PubMed] [Google Scholar]
- 323.Cremer M, Weimann A, Szekessy D. Low immature platelet fraction suggests decreased megakaryopoiesis in neonates with sepsis or necrotizing enterocolitis. J Perinatol. 2013;33(8):622–626. doi: 10.1038/jp.2013.21. [DOI] [PubMed] [Google Scholar]
- 324.Finkelstein Y, Shenkman B, Sirota L. Whole blood platelet deposition on extracellular matrix under flow conditions in preterm neonatal sepsis. Eur J Pediatr. 2002;161(5):270–274. doi: 10.1007/s00431-002-0938-4. [DOI] [PubMed] [Google Scholar]
- 325.Bednarek FJ, Bean S, Barnard MR. The platelet hyporeactivity of extremely low birth weight neonates is age-dependent. Thromb Res. 2009;124(1):42–45. doi: 10.1016/j.thromres.2008.10.004. [DOI] [PubMed] [Google Scholar]
- 326.Grewal PK, Uchiyama S, Ditto D. The Ashwell receptor mitigates the lethal coagulopathy of sepsis. Nat Med. 2008;14(6):648–655. doi: 10.1038/nm1760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327.Guida JD, Kunig AM, Leef KH. Platelet count and sepsis in very low birth weight neonates: is there an organism-specific response? Pediatrics. 2003;111(6 Pt 1):1411–1415. doi: 10.1542/peds.111.6.1411. [DOI] [PubMed] [Google Scholar]
- 328.Hathaway WE, Mull MM, Pechet GS. Disseminated intravascular coagulation in the newborn. Pediatrics. 1969;43(2):233–240. [PubMed] [Google Scholar]
- 329.Kimberlin DW, Lin CY, Jacobs RF. Natural history of neonatal herpes simplex virus infections in the acyclovir era. Pediatrics. 2001;108(2):223–229. doi: 10.1542/peds.108.2.223. [DOI] [PubMed] [Google Scholar]
- 330.Asehnoune K, Moine P. Protease-activated receptor-1: key player in the sepsis coagulation-inflammation crosstalk. Crit Care. 2013;17(1):119. doi: 10.1186/cc12502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 331.Arai H, Matsuda T, Goto R, Takada G. Increased numbers of macrophages in tracheal aspirates in premature infants with funisitis. Pediatr Int. 2008;50(2):184–188. doi: 10.1111/j.1442-200X.2008.02558.x. [DOI] [PubMed] [Google Scholar]
- 332.Pitanga TN, de Aragao Franca L, Rocha VC. Neutrophil-derived microparticles induce myeloperoxidase-mediated damage of vascular endothelial cells. BMC Cell Biol. 2014;15:21. doi: 10.1186/1471-2121-15-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 333.Nimah M, Brilli RJ. Coagulation dysfunction in sepsis and multiple organ system failure. Crit Care Clin. 2003;19(3):441–458. doi: 10.1016/s0749-0704(03)00008-3. [DOI] [PubMed] [Google Scholar]
- 334.Goodwin JE, Feng Y, Velazquez H, Sessa WC. Endothelial glucocorticoid receptor is required for protection against sepsis. Proc Natl Acad Sci U S A. 2013;110(1):306–311. doi: 10.1073/pnas.1210200110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 335.Giuliano JS, Jr, Lahni PM, Harmon K. Admission angiopoietin levels in children with septic shock. Shock. 2007;28(6):650–654. [PMC free article] [PubMed] [Google Scholar]
- 336.Gibson RL, Nizet V, Rubens CE. Group B streptococcal beta-hemolysin promotes injury of lung microvascular endothelial cells. Pediatr Res. 1999;45(5 Pt 1):626–634. doi: 10.1203/00006450-199905010-00003. [DOI] [PubMed] [Google Scholar]
- 337.Dakshinamurti S. Pathophysiologic mechanisms of persistent pulmonary hypertension of the newborn. Pediatr Pulmonol. 2005;39(6):492–503. doi: 10.1002/ppul.20201. [DOI] [PubMed] [Google Scholar]
- 338.Yazji I, Sodhi CP, Lee EK. Endothelial TLR4 activation impairs intestinal microcirculatory perfusion in necrotizing enterocolitis via eNOS-NO-nitrite signaling. Proc Natl Acad Sci U S A. 2013;110(23):9451–9456. doi: 10.1073/pnas.1219997110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 339.Cho JS, Guo Y, Ramos RI. Neutrophil-derived IL-1β is sufficient for abscess formation in immunity against Staphylococcus aureus in mice. PLoS Pathog. 2012;8(11):e1003047. doi: 10.1371/journal.ppat.1003047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 340.Ohman L, Tullus K, Katouli M. Correlation between susceptibility of infants to infections and interaction with neutrophils of Escherichia coli strains causing neonatal and infantile septicemia. J Infect Dis. 1995;171(1):128–133. doi: 10.1093/infdis/171.1.128. [DOI] [PubMed] [Google Scholar]
- 341.Marodi L. Innate cellular immune responses in newborns. Clin Immunol. 2006;118(2–3):137–144. doi: 10.1016/j.clim.2005.10.012. [DOI] [PubMed] [Google Scholar]
- 342.Drossou V, Kanakoudi F, Tzimouli V. Impact of prematurity, stress and sepsis on the neutrophil respiratory burst activity of neonates. Biol Neonate. 1997;72(4):201–209. doi: 10.1159/000244485. [DOI] [PubMed] [Google Scholar]
- 343.Shigeoka AO, Santos JI, Hill HR. Functional analysis of neutrophil granulocytes from healthy, infected, and stressed neonates. J Pediatr. 1979;95(3):454–460. doi: 10.1016/s0022-3476(79)80535-1. [DOI] [PubMed] [Google Scholar]
- 344.Wright WC, Jr, Ank BJ, Herbert J, Stiehm ER. Decreased bactericidal activity of leukocytes of stressed newborn infants. Pediatrics. 1975;56(4):579–584. [PubMed] [Google Scholar]
- 345.Allgaier B, Shi M, Luo D, Koenig JM. Spontaneous and Fas-mediated apoptosis are diminished in umbilical cord blood neutrophils compared with adult neutrophils. J Leukoc Biol. 1998;64(3):331–336. doi: 10.1002/jlb.64.3.331. [DOI] [PubMed] [Google Scholar]
- 346.Hanna N, Vasquez P, Pham P. Mechanisms underlying reduced apoptosis in neonatal neutrophils. Pediatr Res. 2005;57(1):56–62. doi: 10.1203/01.PDR.0000147568.14392.F0. [DOI] [PubMed] [Google Scholar]
- 347.Koenig JM, Stegner JJ, Schmeck AC. Neonatal neutrophils with prolonged survival exhibit enhanced inflammatory and cytotoxic responsiveness. Pediatr Res. 2005;57(3):424–429. doi: 10.1203/01.PDR.0000153945.49022.96. [DOI] [PubMed] [Google Scholar]
- 348.Brinkmann V, Reichard U, Goosmann C. Neutrophil extracellular traps kill bacteria. Science. 2004;303(5663):1532–1535. doi: 10.1126/science.1092385. [DOI] [PubMed] [Google Scholar]
- 349.Clark SR, Ma AC, Tavener SA. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat Med. 2007;13(4):463–469. doi: 10.1038/nm1565. [DOI] [PubMed] [Google Scholar]
- 350.Villanueva E, Yalavarthi S, Berthier CC. Netting neutrophils induce endothelial damage, infiltrate tissues, and expose immunostimulatory molecules in systemic lupus erythematosus. J Immunol. 2011;187(1):538–552. doi: 10.4049/jimmunol.1100450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 351.Margraf S, Logters T, Reipen J. Neutrophil-derived circulating free DNA (cf-DNA/NETs): a potential prognostic marker for posttraumatic development of inflammatory second hit and sepsis. Shock. 2008;30(4):352–358. doi: 10.1097/SHK.0b013e31816a6bb1. [DOI] [PubMed] [Google Scholar]
- 352.Fuchs TA, Abed U, Goosmann C. Novel cell death program leads to neutrophil extracellular traps. J Cell Biol. 2007;176(2):231–241. doi: 10.1083/jcb.200606027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 353.Yost CC, Cody MJ, Harris ES. Impaired neutrophil extracellular trap (NET) formation: a novel innate immune deficiency of human neonates. Blood. 2009;113(25):6419–6427. doi: 10.1182/blood-2008-07-171629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 354.Marcos V, Nussbaum C, Vitkov L. Delayed but functional neutrophil extracellular trap formation in neonates. Blood. 2009;114(23):4908–4911. doi: 10.1182/blood-2009-09-242388. author reply 4911–4912. [DOI] [PubMed] [Google Scholar]
- 355.Branzk N, Lubojemska A, Hardison SE. Neutrophils sense microbe size and selectively release neutrophil extracellular traps in response to large pathogens. Nat Immunol. 2014;15(11):1017–1025. doi: 10.1038/ni.2987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 356.Wynn JL, Tan S, Gantz MG. Outcomes following candiduria in extremely low birth weight infants. Clin Infect Dis. 2012;54(3):331–339. doi: 10.1093/cid/cir800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 357.Christensen RD, Rothstein G. Exhaustion of mature marrow neutrophils in neonates with sepsis. J Pediatr. 1980;96(2):316–318. doi: 10.1016/s0022-3476(80)80837-7. [DOI] [PubMed] [Google Scholar]
- 358.Engle WA, McGuire WA, Schreiner RL, Yu PL. Neutrophil storage pool depletion in neonates with sepsis and neutropenia. J Pediatr. 1988;113(4):747–749. doi: 10.1016/s0022-3476(88)80394-9. [DOI] [PubMed] [Google Scholar]
- 359.Mathur NB, Singh A, Sharma VK, Satyanarayana L. Evaluation of risk factors for fatal neonatal sepsis. Indian Pediatr. 1996;33(10):817–822. [PubMed] [Google Scholar]
- 360.Sarkar S, Bhagat I, Hieber S, Donn SM. Can neutrophil responses in very low birth weight infants predict the organisms responsible for late-onset bacterial or fungal sepsis? J Perinatol. 2006;26(8):501–505. doi: 10.1038/sj.jp.7211554. [DOI] [PubMed] [Google Scholar]
- 361.Christensen RD, Bradley PP, Rothstein G. The leukocyte left shift in clinical and experimental neonatal sepsis. J Pediatr. 1981;98(1):101–105. doi: 10.1016/s0022-3476(81)80553-7. [DOI] [PubMed] [Google Scholar]
- 362.Cuenca AG, Cuenca AL, Gentile LF. Delayed emergency myelopoiesis following polymicrobial sepsis in neonates. Innate Immun. 2015;21(4):386–391. doi: 10.1177/1753425914542445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 363.Mohan P, Brocklehurst P. Granulocyte transfusions for neonates with confirmed or suspected sepsis and neutropaenia. Cochrane Database Syst Rev. 2003;(4) doi: 10.1002/14651858.CD003956. CD003956. [DOI] [PubMed] [Google Scholar]
- 364.Carr R, Brocklehurst P, Dore CJ, Modi N. Granulocyte-macrophage colony stimulating factor administered as prophylaxis for reduction of sepsis in extremely preterm, small for gestational age neonates (the PROGRAMS trial): a single-blind, multicentre, randomised controlled trial. Lancet. 2009;373(9659):226–233. doi: 10.1016/S0140-6736(09)60071-4. [DOI] [PubMed] [Google Scholar]
- 365.Carr R, Modi N, Dore C. G-CSF and GM-CSF for treating or preventing neonatal infections. Cochrane Database Syst Rev. 2003;(3) doi: 10.1002/14651858.CD003066. CD003066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 366.Kuhn P, Messer J, Paupe A. A multicenter, randomized, placebo-controlled trial of prophylactic recombinant granulocyte-colony stimulating factor in preterm neonates with neutropenia. J Pediatr. 2009;155(3):324–330.e1. doi: 10.1016/j.jpeds.2009.03.019. [DOI] [PubMed] [Google Scholar]
- 367.Linderkamp O, Ruef P, Brenner B. Passive deformability of mature, immature, and active neutrophils in healthy and septicemic neonates. Pediatr Res. 1998;44(6):946–950. doi: 10.1203/00006450-199812000-00021. [DOI] [PubMed] [Google Scholar]
- 368.Mease AD, Burgess DP, Thomas PJ. Irreversible neutrophil aggregation. A mechanism of decreased newborn neutrophil chemotactic response. Am J Pathol. 1981;104(1):98–102. [PMC free article] [PubMed] [Google Scholar]
- 369.Nathan CF. Secretory products of macrophages. J Clin Invest. 1987;79(2):319–326. doi: 10.1172/JCI112815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 370.el-Sameea ER, Metwally SS, Mashhour E. Evaluation of natural killer cells as diagnostic markers of early onset neonatal sepsis: comparison with C-reactive protein and interleukin-8. Egypt J Immunol. 2004;11(1):91–102. [PubMed] [Google Scholar]
- 371.Angelone DF, Wessels MR, Coughlin M. Innate immunity of the human newborn is polarized toward a high ratio of IL-6/TNF-alpha production in vitro and in vivo. Pediatr Res. 2006;60(2):205–209. doi: 10.1203/01.pdr.0000228319.10481.ea. [DOI] [PubMed] [Google Scholar]
- 372.Marodi L. Down-regulation of Th1 responses in human neonates. Clin Exp Immunol. 2002;128(1):1–2. doi: 10.1046/j.1365-2249.2002.01873.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 373.Ottenhoff TH, De Boer T, van Dissel JT, Verreck FA. Human deficiencies in type-1 cytokine receptors reveal the essential role of type-1 cytokines in immunity to intracellular bacteria. Adv Exp Med Biol. 2003;531:279–294. doi: 10.1007/978-1-4615-0059-9_24. [DOI] [PubMed] [Google Scholar]
- 374.Klein RB, Fischer TJ, Gard SE. Decreased mononuclear and polymorphonuclear chemotaxis in human newborns, infants, and young children. Pediatrics. 1977;60(4):467–472. [PubMed] [Google Scholar]
- 375.Oren H, Duman N, Abacioglu H. Association between serum macrophage colony-stimulating factor levels and monocyte and thrombocyte counts in healthy, hypoxic, and septic term neonates. Pediatrics. 2001;108(2):329–332. doi: 10.1542/peds.108.2.329. [DOI] [PubMed] [Google Scholar]
- 376.Weinberg AG, Rosenfeld CR, Manroe BL, Browne R. Neonatal blood cell count in health and disease. II. Values for lymphocytes, monocytes, and eosinophils. J Pediatr. 1985;106(3):462–466. doi: 10.1016/s0022-3476(85)80681-8. [DOI] [PubMed] [Google Scholar]
- 377.Hallwirth U, Pomberger G, Zaknun D. Monocyte phagocytosis as a reliable parameter for predicting early-onset sepsis in very low birthweight infants. Early Hum Dev. 2002;67(1–2):1–9. doi: 10.1016/s0378-3782(01)00245-6. [DOI] [PubMed] [Google Scholar]
- 378.Jones CA, Holloway JA, Warner JO. Phenotype of fetal monocytes and B lymphocytes during the third trimester of pregnancy. J Reprod Immunol. 2002;56(1–2):45–60. doi: 10.1016/s0165-0378(02)00022-0. [DOI] [PubMed] [Google Scholar]
- 379.Velilla PA, Rugeles MT, Chougnet CA. Defective antigen-presenting cell function in human neonates. Clin Immunol. 2006;121(3):251–259. doi: 10.1016/j.clim.2006.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 380.Chelvarajan L, Popa D, Liu Y. Molecular mechanisms underlying anti-inflammatory phenotype of neonatal splenic macrophages. J Leukoc Biol. 2007;82(2):403–416. doi: 10.1189/jlb.0107071. [DOI] [PubMed] [Google Scholar]
- 381.Krumbiegel D, Rohr J, Schmidtke P. Efficient maturation and cytokine production of neonatal DCs requires combined proinflammatory signals. Clin Dev Immunol. 2005;12(2):99–105. doi: 10.1080/17402520500116772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 382.Wong OH, Huang FP, Chiang AK. Differential responses of cord and adult blood-derived dendritic cells to dying cells. Immunology. 2005;116(1):13–20. doi: 10.1111/j.1365-2567.2005.02191.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 383.Schuller SS, Sadeghi K, Wisgrill L. Preterm neonates display altered plasmacytoid dendritic cell function and morphology. J Leukoc Biol. 2013;93(5):781–788. doi: 10.1189/jlb.1011525. [DOI] [PubMed] [Google Scholar]
- 384.Salio M, Dulphy N, Renneson J. Efficient priming of antigen-specific cytotoxic T lymphocytes by human cord blood dendritic cells. Int Immunol. 2003;15(10):1265–1273. doi: 10.1093/intimm/dxg123. [DOI] [PubMed] [Google Scholar]
- 385.Scumpia PO, McAuliffe PF, O'Malley KA. CD11c+ dendritic cells are required for survival in murine polymicrobial sepsis. J Immunol. 2005;175(5):3282–3286. doi: 10.4049/jimmunol.175.5.3282. [DOI] [PubMed] [Google Scholar]
- 386.Hotchkiss RS, Tinsley KW, Swanson PE. Depletion of dendritic cells, but not macrophages, in patients with sepsis. J Immunol. 2002;168(5):2493–2500. doi: 10.4049/jimmunol.168.5.2493. [DOI] [PubMed] [Google Scholar]
- 387.Juul SE, Haynes JW, McPherson RJ. Evaluation of eosinophilia in hospitalized preterm infants. J Perinatol. 2005;25(3):182–188. doi: 10.1038/sj.jp.7211226. [DOI] [PubMed] [Google Scholar]
- 388.Wolach B, Bogger-Goren S, Whyte R. Perinatal hematological profile of newborn infants with candida antenatal infections. Biol Neonate. 1991;59(1):5–12. doi: 10.1159/000243316. [DOI] [PubMed] [Google Scholar]
- 389.Patel L, Garvey B, Arnon S, Roberts IA. Eosinophilia in newborn infants. Acta Paediatr. 1994;83(8):797–801. doi: 10.1111/j.1651-2227.1994.tb13146.x. [DOI] [PubMed] [Google Scholar]
- 390.Rothberg AD, Cohn RJ, Argent AC. Eosinophilia in premature neonates. Phase 2 of a biphasic granulopoietic response. S Afr Med J. 1983;64(14):539–541. [PubMed] [Google Scholar]
- 391.Yousefi S, Gold JA, Andina N. Catapult-like release of mitochondrial DNA by eosinophils contributes to antibacterial defense. Nat Med. 2008;14(9):949–953. doi: 10.1038/nm.1855. [DOI] [PubMed] [Google Scholar]
- 392.Marshall JS, Jawdat DM. Mast cells in innate immunity. J Allergy Clin Immunol. 2004;114(1):21–27. doi: 10.1016/j.jaci.2004.04.045. [DOI] [PubMed] [Google Scholar]
- 393.Lin AM, Rubin CJ, Khandpur R. Mast cells and neutrophils release IL-17 through extracellular trap formation in psoriasis. J Immunol. 2011;187(1):490–500. doi: 10.4049/jimmunol.1100123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 394.Nelson A, Ulfgren AK, Edner J. Urticaria Neonatorum: accumulation of tryptase-expressing mast cells in the skin lesions of newborns with Erythema Toxicum. Pediatr Allergy Immunol. 2007;18(8):652–658. doi: 10.1111/j.1399-3038.2007.00568.x. [DOI] [PubMed] [Google Scholar]
- 395.Malaviya R, Ikeda T, Ross E, Abraham SN. Mast cell modulation of neutrophil influx and bacterial clearance at sites of infection through TNF-alpha. Nature. 1996;381(6577):77–80. doi: 10.1038/381077a0. [DOI] [PubMed] [Google Scholar]
- 396.Echtenacher B, Mannel DN, Hultner L. Critical protective role of mast cells in a model of acute septic peritonitis. Nature. 1996;381(6577):75–77. doi: 10.1038/381075a0. [DOI] [PubMed] [Google Scholar]
- 397.von Kockritz-Blickwede M, Goldmann O, Thulin P. Phagocytosis-independent antimicrobial activity of mast cells by means of extracellular trap formation. Blood. 2008;111(6):3070–3080. doi: 10.1182/blood-2007-07-104018. [DOI] [PubMed] [Google Scholar]
- 398.Mazzoni A, Young HA, Spitzer JH. Histamine regulates cytokine production in maturing dendritic cells, resulting in altered T cell polarization. J Clin Invest. 2001;108(12):1865–1873. doi: 10.1172/JCI13930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 399.Damsgaard TE, Nielsen BW, Henriques U. Histamine releasing cells of the newborn. Mast cells from the umbilical cord matrix and basophils from cord blood. Pediatr Allergy Immunol. 1996;7(2):83–90. doi: 10.1111/j.1399-3038.1996.tb00111.x. [DOI] [PubMed] [Google Scholar]
- 400.Schneider LA, Schlenner SM, Feyerabend TB. Molecular mechanism of mast cell mediated innate defense against endothelin and snake venom sarafotoxin. J Exp Med. 2007;204(11):2629–2639. doi: 10.1084/jem.20071262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 401.Perez A, Gurbindo MD, Resino S. NK cell increase in neonates from the preterm to the full-term period of gestation. Neonatology. 2007;92(3):158–163. doi: 10.1159/000101567. [DOI] [PubMed] [Google Scholar]
- 402.Ma L, Chen R, Liu F. Reduced NK cell percentage at birth is associated with late onset infection in very preterm neonates. Scand J Immunol. 2014;80(1):50–56. doi: 10.1111/sji.12181. [DOI] [PubMed] [Google Scholar]
- 403.Juretic E, Juretic A, Uzarevic B, Petrovecki M. Alterations in lymphocyte phenotype of infected preterm newborns. Biol Neonate. 2001;80(3):223–227. doi: 10.1159/000047147. [DOI] [PubMed] [Google Scholar]
- 404.Mazur B, Godula-Stuglik U, Domarecki A, Stojewska M. [The influence of severe infections on the natural killer cells in neonates with vary gestational age] Ginekol Pol. 2000;71(6):542–549. [PubMed] [Google Scholar]
- 405.Georgeson GD, Szony BJ, Streitman K. Natural killer cell cytotoxicity is deficient in newborns with sepsis and recurrent infections. Eur J Pediatr. 2001;160(8):478–482. doi: 10.1007/s004310100773. [DOI] [PubMed] [Google Scholar]
- 406.Hodge G, Hodge S, Han P, Haslam R. Multiple leucocyte activation markers to detect neonatal infection. Clin Exp Immunol. 2004;135(1):125–129. doi: 10.1111/j.1365-2249.2004.02346.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 407.Elahi S, Ertelt JM, Kinder JM. Immunosuppressive CD71+ erythroid cells compromise neonatal host defence against infection. Nature. 2013;504(7478):158–162. doi: 10.1038/nature12675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 408.Ned RM, Swat W, Andrews NC. Transferrin receptor 1 is differentially required in lymphocyte development. Blood. 2003;102(10):3711–3718. doi: 10.1182/blood-2003-04-1086. [DOI] [PubMed] [Google Scholar]
- 409.Li H, Rybicki AC, Suzuka SM. Transferrin therapy ameliorates disease in beta-thalassemic mice. Nat Med. 2010;16(2):177–182. doi: 10.1038/nm.2073. [DOI] [PubMed] [Google Scholar]
- 410.Socolovsky M, Nam H, Fleming MD. Ineffective erythropoiesis in Stat5a−/−5b−/− mice due to decreased survival of early erythroblasts. Blood. 2001;98(12):3261–3273. doi: 10.1182/blood.v98.12.3261. [DOI] [PubMed] [Google Scholar]
- 411.Wolber FM, Leonard E, Michael S. Roles of spleen and liver in development of the murine hematopoietic system. Exp Hematol. 2002;30(9):1010–1019. doi: 10.1016/s0301-472x(02)00881-0. [DOI] [PubMed] [Google Scholar]
- 412.Timens W, Boes A, Rozeboom-Uiterwijk T, Poppema S. Immaturity of the human splenic marginal zone in infancy. Possible contribution to the deficient infant immune response. J Immunol. 1989;143(10):3200–3206. [PubMed] [Google Scholar]
- 413.Timens W, Rozeboom T, Poppema S. Fetal and neonatal development of human spleen: an immunohistological study. Immunology. 1987;60(4):603–609. [PMC free article] [PubMed] [Google Scholar]
- 414.Wynn JL, Scumpia PO, Stocks BT. Neonatal CD71+ erythroid cells do not modify murine sepsis mortality. J Immunol. 2015;195(3):1064–1070. doi: 10.4049/jimmunol.1500771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 415.Hotchkiss RS, Chang KC, Swanson PE. Caspase inhibitors improve survival in sepsis: a critical role of the lymphocyte. Nat Immunol. 2000;1(6):496–501. doi: 10.1038/82741. [DOI] [PubMed] [Google Scholar]
- 416.Kelly-Scumpia KM, Scumpia PO, Weinstein JS. B cells enhance early innate immune responses during bacterial sepsis. J Exp Med. 2011;208(8):1673–1682. doi: 10.1084/jem.20101715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 417.Adkins B, Leclerc C, Marshall-Clarke S. Neonatal adaptive immunity comes of age. Nat Rev Immunol. 2004;4(7):553–564. doi: 10.1038/nri1394. [DOI] [PubMed] [Google Scholar]
- 418.Godula-Stuglik U, Mazur B, Mikusz G. Lymphocyte subpopulations in full-term septic neonates. Pediatr Int. 1999;41(5):500–505. doi: 10.1046/j.1442-200x.1999.01120.x. [DOI] [PubMed] [Google Scholar]
- 419.Aygun AD, Citak Kurt AN, Godekmerdan A. Neonates with culture proven sepsis have lower amounts and percentage of CD45RA+ T cells. Inflammation. 2008;31(4):222–226. doi: 10.1007/s10753-008-9068-0. [DOI] [PubMed] [Google Scholar]
- 420.Hotoura E, Giapros V, Kostoula A. Pre-inflammatory mediators and lymphocyte subpopulations in preterm neonates with sepsis. Inflammation. 2012;35(3):1094–1101. doi: 10.1007/s10753-011-9416-3. [DOI] [PubMed] [Google Scholar]
- 421.Bussel JB, Cunningham-Rundles S, LaGamma EF, Shellabarger M. Analysis of lymphocyte proliferative response subpopulations in very low birth weight infants and during the first 8 weeks of life. Pediatr Res. 1988;23(5):457–462. doi: 10.1203/00006450-198805000-00003. [DOI] [PubMed] [Google Scholar]
- 422.Behrendt J, Karpe J, Mazur B. [Quantitative changes of CD3+, CD4+, CD8+, HLA-DR+, T lymphocytes and CD25+ cells in eutrophic full-term neonates with staphylococcal septicemia] Przegl Lek. 2002;59(Suppl 1):50–53. [PubMed] [Google Scholar]
- 423.Godula-Stuglik U, Mazur B, Behrendt J. Peripheral blood cells with expression of the interleukin-2 receptor (CD25+) in severely infected full-term neonates. Acta Paediatr. 2001;90(7):819–820. [PubMed] [Google Scholar]
- 424.Godfrey WR, Spoden DJ, Ge YG. Cord blood CD4+CD25+-derived T regulatory cell lines express FoxP3 protein and manifest potent suppressor function. Blood. 2005;105(2):750–758. doi: 10.1182/blood-2004-06-2467. [DOI] [PubMed] [Google Scholar]
- 425.Aluvihare VR, Kallikourdis M, Betz AG. Regulatory T cells mediate maternal tolerance to the fetus. Nat Immunol. 2004;5(3):266–271. doi: 10.1038/ni1037. [DOI] [PubMed] [Google Scholar]
- 426.Scumpia PO, Delano MJ, Kelly KM. Increased natural CD4+CD25+ regulatory T cells and their suppressor activity do not contribute to mortality in murine polymicrobial sepsis. J Immunol. 2006;177(11):7943–7949. doi: 10.4049/jimmunol.177.11.7943. [DOI] [PubMed] [Google Scholar]
- 427.Tezuka T, Sugita K, Mizobe N. Transient increase in CD45RO expression on T lymphocytes in infected newborns. Pediatr Res. 1998;43(2):283–290. doi: 10.1203/00006450-199802000-00021. [DOI] [PubMed] [Google Scholar]
- 428.Hodge S, Hodge G, Flower R, Han P. Surface activation markers of T lymphocytes: role in the detection of infection in neonates. Clin Exp Immunol. 1998;113(1):33–38. doi: 10.1046/j.1365-2249.1998.00626.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 429.Michie C, Harvey D. Can expression of CD45RO, a T-cell surface molecule, be used to detect congenital infection? Lancet. 1994;343(8908):1259–1260. doi: 10.1016/s0140-6736(94)92153-9. [DOI] [PubMed] [Google Scholar]
- 430.Bruning T, Daiminger A, Enders G. Diagnostic value of CD45RO expression on circulating T lymphocytes of fetuses and newborn infants with pre-, peri- or early post-natal infections. Clin Exp Immunol. 1997;107(2):306–311. doi: 10.1111/j.1365-2249.1997.268-ce1165.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 431.van der Flier M, Sharma DB, Estevao S. Increased CD4+ T cell co-inhibitory immune receptor CEACAM1 in neonatal sepsis and soluble-CEACAM1 in meningococcal sepsis: a role in sepsis-associated immune suppression? PLoS ONE. 2013;8(7):e68294. doi: 10.1371/journal.pone.0068294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 432.Gibbons D, Fleming P, Virasami A. Interleukin-8 (CXCL8) production is a signatory T cell effector function of human newborn infants. Nat Med. 2014;20(10):1206–1210. doi: 10.1038/nm.3670. [DOI] [PubMed] [Google Scholar]
- 433.Matsuoka T, Matsubara T, Katayama K. Increase of cord blood cytokine-producing T cells in intrauterine infection. Pediatr Int. 2001;43(5):453–457. doi: 10.1046/j.1442-200x.2001.01445.x. [DOI] [PubMed] [Google Scholar]
- 434.Gasparoni A, Ciardelli L, Avanzini A. Age-related changes in intracellular TH1/TH2 cytokine production, immunoproliferative T lymphocyte response and natural killer cell activity in newborns, children and adults. Biol Neonate. 2003;84(4):297–303. doi: 10.1159/000073638. [DOI] [PubMed] [Google Scholar]
- 435.Galal N, Badawy A, Khalaf-Allah O, Youssef A. Identification of T-lymphocyte function in healthy vs. septic preterms and its relation to candidal infections in the hospital setting. Indian J Pediatr. 2012;79(3):348–352. doi: 10.1007/s12098-011-0602-y. [DOI] [PubMed] [Google Scholar]
- 436.Marchant A, Appay V, Van Der Sande M. Mature CD8+ T lymphocyte response to viral infection during fetal life. J Clin Invest. 2003;111(11):1747–1755. doi: 10.1172/JCI17470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 437.Hansen K, Singer DB. Asplenic-hyposplenic overwhelming sepsis: postsplenectomy sepsis revisited. Pediatr Dev Pathol. 2001;4(2):105–121. doi: 10.1007/s100240010145. [DOI] [PubMed] [Google Scholar]
- 438.William BM, Corazza GR. Hyposplenism: a comprehensive review. Part I: basic concepts and causes. Hematology. 2007;12(1):1–13. doi: 10.1080/10245330600938422. [DOI] [PubMed] [Google Scholar]
- 439.Vallejo JG, Baker CJ, Edwards MS. Demonstration of circulating group B streptococcal immune complexes in neonates with meningitis. J Clin Microbiol. 1994;32(9):2041–2045. doi: 10.1128/jcm.32.9.2041-2045.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 440.Cukrowska B, Lodinova-Zadnikova R, Sokol D, Tlaskalova-Hogenova H. In vitro immunoglobulin response of fetal B-cells is influenced by perinatal infections and antibiotic treatment: a study in preterm infants. Eur J Pediatr. 1999;158(6):463–468. doi: 10.1007/s004310051121. [DOI] [PubMed] [Google Scholar]
- 441.Wynn JL, Li L, Cotten CM. Blood stream infection is associated with altered heptavalent pneumococcal conjugate vaccine immune responses in very low birth weight infants. J Perinatol. 2013;33(8):613–618. doi: 10.1038/jp.2013.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 442.Lanier LL. Shades of grey–the blurring view of innate and adaptive immunity. Nat Rev Immunol. 2013;13(2):73–74. doi: 10.1038/nri3389. [DOI] [PubMed] [Google Scholar]
- 443.Emoto M, Miyamoto M, Emoto Y. A critical role of T-cell receptor gamma/delta cells in antibacterial protection in mice early in life. Hepatology. 2001;33(4):887–893. doi: 10.1053/jhep.2001.23504. [DOI] [PubMed] [Google Scholar]
- 444.Jouen-Beades F, Paris E, Dieulois C. In vivo and in vitro activation and expansion of gammadelta T cells during Listeria monocytogenes infection in humans. Infect Immun. 1997;65(10):4267–4272. doi: 10.1128/iai.65.10.4267-4272.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 445.Hedges JF, Lubick KJ, Jutila MA. Gamma delta T cells respond directly to pathogen-associated molecular patterns. J Immunol. 2005;174(10):6045–6053. doi: 10.4049/jimmunol.174.10.6045. [DOI] [PubMed] [Google Scholar]
- 446.Krolak-Olejnik B, Mazur B. Influence of perinatal risk factors on CD3+/TCR αβ and CD3+/TCR γδ lymphocytes in cord blood of preterm neonates. Pediatr Int. 2004;46(4):403–408. doi: 10.1111/j.1442-200x.2004.01926.x. [DOI] [PubMed] [Google Scholar]
- 447.Morita CT, Parker CM, Brenner MB, Band H. TCR usage and functional capabilities of human gamma delta T cells at birth. J Immunol. 1994;153(9):3979–3988. [PubMed] [Google Scholar]
- 448.Chen L, He Z, Slinger E. IL-23 activates innate lymphoid cells to promote neonatal intestinal pathology. Mucosal Immunol. 2014;8:390–402. doi: 10.1038/mi.2014.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 449.Wong HR, Cvijanovich N, Lin R. Identification of pediatric septic shock subclasses based on genome-wide expression profiling. BMC Med. 2009;7:34. doi: 10.1186/1741-7015-7-34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 450.Wong HR, Cvijanovich N, Allen GL. Genomic expression profiling across the pediatric systemic inflammatory response syndrome, sepsis, and septic shock spectrum. Crit Care Med. 2009;37(5):1558–1566. doi: 10.1097/CCM.0b013e31819fcc08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 451.Wong HR, Odoms K, Sakthivel B. Divergence of canonical danger signals: the genome-level expression patterns of human mononuclear cells subjected to heat shock or lipopolysaccharide. BMC Immunol. 2008;9:24. doi: 10.1186/1471-2172-9-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 452.Cvijanovich N, Shanley TP, Lin R. Validating the genomic signature of pediatric septic shock. Physiol Genomics. 2008;34(1):127–134. doi: 10.1152/physiolgenomics.00025.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 453.Shanley TP, Cvijanovich N, Lin R. Genome-level longitudinal expression of signaling pathways and gene networks in pediatric septic shock. Mol Med. 2007;13(9–10):495–508. doi: 10.2119/2007-00065.Shanley. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 454.Tang BM, McLean AS, Dawes IW. Gene-expression profiling of gram-positive and gram-negative sepsis in critically ill patients. Crit Care Med. 2008;36(4):1125–1128. doi: 10.1097/CCM.0b013e3181692c0b. [DOI] [PubMed] [Google Scholar]
- 455.Tang BM, McLean AS, Dawes IW. The use of gene-expression profiling to identify candidate genes in human sepsis. Am J Respir Crit Care Med. 2007;176(7):676–684. doi: 10.1164/rccm.200612-1819OC. [DOI] [PubMed] [Google Scholar]
- 456.Tang BM, McLean AS, Dawes IW. Gene-expression profiling of peripheral blood mononuclear cells in sepsis. Crit Care Med. 2009;37(3):882–888. doi: 10.1097/CCM.0b013e31819b52fd. [DOI] [PubMed] [Google Scholar]
- 457.Karvunidis T, Mares J, Thongboonkerd V, Matejovic M. Recent progress of proteomics in critical illness. Shock. 2009;31(6):545–552. doi: 10.1097/SHK.0b013e3181986eab. [DOI] [PubMed] [Google Scholar]
- 458.Buhimschi CS, Bhandari V, Hamar BD. Proteomic profiling of the amniotic fluid to detect inflammation, infection, and neonatal sepsis. PLoS Med. 2007;4(1):e18. doi: 10.1371/journal.pmed.0040018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 459.Ghantous A, Saffery R, Cros MP. Optimized DNA extraction from neonatal dried blood spots: application in methylome profiling. BMC Biotechnol. 2014;14:60. doi: 10.1186/1472-6750-14-60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 460.Wong HR, Cvijanovich NZ, Allen GL. Validation of a gene expression-based subclassification strategy for pediatric septic shock. Crit Care Med. 2011;39(11):2511–2517. doi: 10.1097/CCM.0b013e3182257675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 461.Wong HR, Shanley TP, Sakthivel B. Genome-level expression profiles in pediatric septic shock indicate a role for altered zinc homeostasis in poor outcome. Physiol Genomics. 2007;30(2):146–155. doi: 10.1152/physiolgenomics.00024.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 462.Nowak JE, Harmon K, Caldwell CC, Wong HR. Prophylactic zinc supplementation reduces bacterial load and improves survival in a murine model of sepsis. Pediatr Crit Care Med. 2012;13(5):e323–e329. doi: 10.1097/PCC.0b013e31824fbd90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 463.Mehta K, Bhatta NK, Majhi S. Oral zinc supplementation for reducing mortality in probable neonatal sepsis: a double blind randomized placebo controlled trial. Indian Pediatr. 2013;50(4):390–393. doi: 10.1007/s13312-013-0120-2. [DOI] [PubMed] [Google Scholar]
- 464.Wynn JL, Guthrie SO, Wong HR. Post-natal age is a critical determinant of the neonatal host response to sepsis. Mol Med. 2015;21(1):496–504. doi: 10.2119/molmed.2015.00064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 465.Cernada M, Serna E, Bauerl C. Genome-wide expression profiles in very low birth weight infants with neonatal sepsis. Pediatrics. 2014;133(5):e1203–e1211. doi: 10.1542/peds.2013-2552. [DOI] [PubMed] [Google Scholar]
- 466.Solan PD, Dunsmore KE, Denenberg AG. A novel role for matrix metalloproteinase-8 in sepsis. Crit Care Med. 2012;40(2):379–387. doi: 10.1097/CCM.0b013e318232e404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 467.Smith CL, Dickinson P, Forster T. Identification of a human neonatal immune-metabolic network associated with bacterial infection. Nat Commun. 2014;5:4649. doi: 10.1038/ncomms5649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 468.Ng PC, Ang IL, Chiu RW. Host-response biomarkers for diagnosis of late-onset septicemia and necrotizing enterocolitis in preterm infants. J Clin Invest. 2010;120(8):2989–3000. doi: 10.1172/JCI40196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 469.Di Naro E, Cromi A, Ghezzi F. Fetal thymic involution: a sonographic marker of the fetal inflammatory response syndrome. Am J Obstet Gynecol. 2006;194(1):153–159. doi: 10.1016/j.ajog.2005.05.036. [DOI] [PubMed] [Google Scholar]
- 470.Sakr Y, Dubois MJ, De Backer D. Persistent microcirculatory alterations are associated with organ failure and death in patients with septic shock. Crit Care Med. 2004;32(9):1825–1831. doi: 10.1097/01.ccm.0000138558.16257.3f. [DOI] [PubMed] [Google Scholar]
- 471.Zakariya BP, Bhat BV, Harish BN. Risk factors and predictors of mortality in culture proven neonatal sepsis. Indian J Pediatr. 2012;79(3):358–361. doi: 10.1007/s12098-011-0584-9. [DOI] [PubMed] [Google Scholar]
- 472.Cochat P, Bourgeois J, Gilly J. [Anatomical study of the kidneys of newborn infants dying after a septic state] Pediatrie. 1986;41(1):7–15. [PubMed] [Google Scholar]
- 473.Csaicsich D, Russo-Schlaff N, Messerschmidt A. Renal failure, comorbidity and mortality in preterm infants. Wien Klin Wochenschr. 2008;120(5–6):153–157. doi: 10.1007/s00508-008-0941-5. [DOI] [PubMed] [Google Scholar]
- 474.Spapen H. Liver perfusion in sepsis, septic shock, and multiorgan failure. Anat Rec (Hoboken) 2008;291(6):714–720. doi: 10.1002/ar.20646. [DOI] [PubMed] [Google Scholar]
- 475.Bhutta ZA, Yusuf K. Neonatal sepsis in Karachi: factors determining outcome and mortality. J Trop Pediatr. 1997;43(2):65–70. doi: 10.1093/tropej/43.2.65. [DOI] [PubMed] [Google Scholar]
- 476.Faix RG, Donn SM. Association of septic shock caused by early-onset group B streptococcal sepsis and periventricular leukomalacia in the preterm infant. Pediatrics. 1985;76(3):415–419. [PubMed] [Google Scholar]
- 477.Hotchkiss RS, Osmon SB, Chang KC. Accelerated lymphocyte death in sepsis occurs by both the death receptor and mitochondrial pathways. J Immunol. 2005;174(8):5110–5118. doi: 10.4049/jimmunol.174.8.5110. [DOI] [PubMed] [Google Scholar]
- 478.Gurevich P, Ben-Hur H, Czernobilsky B. Pathology of lymphoid organs in low birth weight infants subjected to antigen-related diseases: a morphological and morphometric study. Pathology. 1995;27(2):121–126. doi: 10.1080/00313029500169702. [DOI] [PubMed] [Google Scholar]
- 479.Cinel I, Dellinger RP. Advances in pathogenesis and management of sepsis. Curr Opin Infect Dis. 2007;20(4):345–352. doi: 10.1097/QCO.0b013e32818be70a. [DOI] [PubMed] [Google Scholar]
- 480.Cinel I, Opal SM. Molecular biology of inflammation and sepsis: a primer. Crit Care Med. 2009;37(1):291–304. doi: 10.1097/CCM.0b013e31819267fb. [DOI] [PubMed] [Google Scholar]
- 481.Sonntag J, Wagner MH, Waldschmidt J. Multisystem organ failure and capillary leak syndrome in severe necrotizing enterocolitis of very low birth weight infants. J Pediatr Surg. 1998;33(3):481–484. doi: 10.1016/s0022-3468(98)90092-6. [DOI] [PubMed] [Google Scholar]
- 482.Levit O, Bhandari V, Li FY. Clinical and laboratory factors that predict death in very low birth weight infants presenting with late-onset sepsis. Pediatr Infect Dis J. 2014;33(2):143–146. doi: 10.1097/INF.0000000000000024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 483.Tabbutt S. Heart failure in pediatric septic shock: utilizing inotropic support. Crit Care Med. 2001;29(10 Suppl):S231–S236. doi: 10.1097/00003246-200110001-00005. [DOI] [PubMed] [Google Scholar]
- 484.McKiernan CA, Lieberman SA. Circulatory shock in children: an overview. Pediatr Rev. 2005;26(12):451–460. doi: 10.1542/pir.26-12-451. [DOI] [PubMed] [Google Scholar]
- 485.Ceneviva G, Paschall JA, Maffei F, Carcillo JA. Hemodynamic support in fluid-refractory pediatric septic shock. Pediatrics. 1998;102(2):e19. doi: 10.1542/peds.102.2.e19. [DOI] [PubMed] [Google Scholar]
- 486.Carcillo JA, Fields AI. Clinical practice parameters for hemodynamic support of pediatric and neonatal patients in septic shock. Crit Care Med. 2002;30(6):1365–1378. doi: 10.1097/00003246-200206000-00040. [DOI] [PubMed] [Google Scholar]
- 487.Brierley J, Peters MJ. Distinct hemodynamic patterns of septic shock at presentation to pediatric intensive care. Pediatrics. 2008;122(4):752–759. doi: 10.1542/peds.2007-1979. [DOI] [PubMed] [Google Scholar]
- 488.Evans N. Which inotrope for which baby? Arch Dis Child Fetal Neonatal Ed. 2006;91(3):F213–F220. doi: 10.1136/adc.2005.071829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 489.Kluckow M, Evans N. Relationship between blood pressure and cardiac output in preterm infants requiring mechanical ventilation. J Pediatr. 1996;129(4):506–512. doi: 10.1016/s0022-3476(96)70114-2. [DOI] [PubMed] [Google Scholar]
- 490.Alba-Alejandre I, Hiedl S, Genzel-Boroviczeny O. Microcirculatory changes in term newborns with suspected infection: an observational prospective study. Int J Pediatr. 2013;2013:768784. doi: 10.1155/2013/768784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 491.de Waal K, Evans N. Hemodynamics in preterm infants with late-onset sepsis. J Pediatr. 2010;156(6):918–922 e1. doi: 10.1016/j.jpeds.2009.12.026. [DOI] [PubMed] [Google Scholar]
- 492.Tomerak RH, El-Badawy AA, Hussein G. Echocardiogram done early in neonatal sepsis: what does it add? J Investig Med. 2012;60(4):680–684. doi: 10.2310/JIM.0b013e318249fc95. [DOI] [PubMed] [Google Scholar]
- 493.Abdel-Hady HE, Matter MK, El-Arman MM. Myocardial dysfunction in neonatal sepsis: a tissue Doppler imaging study. Pediatr Crit Care Med. 2012;13(3):318–323. doi: 10.1097/PCC.0b013e3182257b6b. [DOI] [PubMed] [Google Scholar]
- 494.Seri I, Noori S. Diagnosis and treatment of neonatal hypotension outside the transitional period. Early Hum Dev. 2005;81(5):405–411. doi: 10.1016/j.earlhumdev.2005.03.008. [DOI] [PubMed] [Google Scholar]
- 495.Domico M, Liao P, Anas N, Mink RB. Elevation of brain natriuretic peptide levels in children with septic shock. Pediatr Crit Care Med. 2008;9(5):478–483. doi: 10.1097/PCC.0b013e3181849b99. [DOI] [PubMed] [Google Scholar]
- 496.Wang F, Wu Y, Tang L. Brain natriuretic peptide for prediction of mortality in patients with sepsis: a systematic review and meta-analysis. Crit Care. 2012;16(3):R74. doi: 10.1186/cc11331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 497.Duke TD, Butt W, South M. Predictors of mortality and multiple organ failure in children with sepsis. Intensive Care Med. 1997;23(6):684–692. doi: 10.1007/s001340050394. [DOI] [PubMed] [Google Scholar]
- 498.Hotchkiss RS, Tinsley KW, Swanson PE. Sepsis-induced apoptosis causes progressive profound depletion of B and CD4+ T lymphocytes in humans. J Immunol. 2001;166(11):6952–6963. doi: 10.4049/jimmunol.166.11.6952. [DOI] [PubMed] [Google Scholar]
- 499.van Baarlen J, Schuurman HJ, Huber J. Acute thymus involution in infancy and childhood: a reliable marker for duration of acute illness. Hum Pathol. 1988;19(10):1155–1160. doi: 10.1016/s0046-8177(88)80146-1. [DOI] [PubMed] [Google Scholar]
- 500.Chen CM, Yu KY, Lin HC. Thymus size and its relationship to perinatal events. Acta Paediatr. 2000;89(8):975–978. doi: 10.1080/080352500750043468. [DOI] [PubMed] [Google Scholar]
- 501.Glavina-Durdov M, Springer O, Capkun V. The grade of acute thymus involution in neonates correlates with the duration of acute illness and with the percentage of lymphocytes in peripheral blood smear. Pathological study. Biol Neonate. 2003;83(4):229–234. doi: 10.1159/000069481. [DOI] [PubMed] [Google Scholar]
- 502.Itoh K, Aihara H, Takada S. Clinicopathological differences between early-onset and late-onset sepsis and pneumonia in very low birth weight infants. Pediatr Pathol. 1990;10(5):757–768. doi: 10.3109/15513819009064710. [DOI] [PubMed] [Google Scholar]
- 503.Toti P, De Felice C, Occhini R. Spleen depletion in neonatal sepsis and chorioamnionitis. Am J Clin Pathol. 2004;122(5):765–771. doi: 10.1309/RV6E-9BMC-9954-A2WU. [DOI] [PubMed] [Google Scholar]
- 504.Toti P, De Felice C, Stumpo M. Acute thymic involution in fetuses and neonates with chorioamnionitis. Hum Pathol. 2000;31(9):1121–1128. doi: 10.1053/hupa.2000.16676. [DOI] [PubMed] [Google Scholar]
- 505.Felmet KA, Hall MW, Clark RS. Prolonged lymphopenia, lymphoid depletion, and hypoprolactinemia in children with nosocomial sepsis and multiple organ failure. J Immunol. 2005;174(6):3765–3772. doi: 10.4049/jimmunol.174.6.3765. [DOI] [PubMed] [Google Scholar]
- 506.Hotchkiss RS, Monneret G, Payen D. Immunosuppression in sepsis: a novel understanding of the disorder and a new therapeutic approach. Lancet Infect Dis. 2013;13(3):260–268. doi: 10.1016/S1473-3099(13)70001-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 507.Hotchkiss RS, Coopersmith CM, Karl IE. Prevention of lymphocyte apoptosis—a potential treatment of sepsis? Clin Infect Dis. 2005;41(Suppl 7):S465–S469. doi: 10.1086/431998. [DOI] [PubMed] [Google Scholar]
- 508.Kalman L, Lindegren ML, Kobrynski L. Mutations in genes required for T-cell development: IL7R, CD45, IL2RG, JAK3, RAG1, RAG2, ARTEMIS, and ADA and severe combined immunodeficiency: HuGE review. Genet Med. 2004;6(1):16–26. doi: 10.1097/01.GIM.0000105752.80592.A3. [DOI] [PubMed] [Google Scholar]
- 509.Le Tulzo Y, Pangault C, Gacouin A. Early circulating lymphocyte apoptosis in human septic shock is associated with poor outcome. Shock. 2002;18(6):487–494. doi: 10.1097/00024382-200212000-00001. [DOI] [PubMed] [Google Scholar]
- 510.Boomer JS, To K, Chang KC. Immunosuppression in patients who die of sepsis and multiple organ failure. JAMA. 2011;306(23):2594–2605. doi: 10.1001/jama.2011.1829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 511.Frazier WJ, Hall MW. Immunoparalysis and adverse outcomes from critical illness. Pediatr Clin North Am. 2008;55(3):647–668. doi: 10.1016/j.pcl.2008.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 512.Lin CB, Hornik CP, Clark R. Very low birth weight neonates who survive early-onset sepsis do not have an increased risk of developing late-onset sepsis. Early Hum Dev. 2012;88(11):905–909. doi: 10.1016/j.earlhumdev.2012.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 513.Wynn JL, Hansen NI, Das A. Early sepsis does not increase the risk of late sepsis in very low birth weight neonates. J Pediatr. 2013;162(5):942–948.e3. doi: 10.1016/j.jpeds.2012.11.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 514.Hotchkiss RS, Karl IE. The pathophysiology and treatment of sepsis. N Engl J Med. 2003;348(2):138–150. doi: 10.1056/NEJMra021333. [DOI] [PubMed] [Google Scholar]
- 515.Wang H, Yu M, Ochani M. Nicotinic acetylcholine receptor α7 subunit is an essential regulator of inflammation. Nature. 2003;421(6921):384–388. doi: 10.1038/nature01339. [DOI] [PubMed] [Google Scholar]
- 516.Andersson U, Tracey KJ. Neural reflexes in inflammation and immunity. J Exp Med. 2012;209(6):1057–1068. doi: 10.1084/jem.20120571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 517.Carson WF, Cavassani KA, Dou Y, Kunkel SL. Epigenetic regulation of immune cell functions during post-septic immunosuppression. Epigenetics. 2011;6(3):273–283. doi: 10.4161/epi.6.3.14017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 518.Tendl KA, Schulz SM, Mechtler TP. DNA methylation pattern of CALCA in preterm neonates with bacterial sepsis as a putative epigenetic biomarker. Epigenetics. 2013;8(12):1261–1267. doi: 10.4161/epi.26645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 519.Levy O, Wynn JL. A prime time for trained immunity: innate immune memory in newborns and infants. Neonatology. 2014;105(2):136–141. doi: 10.1159/000356035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 520.Saeed S, Quintin J, Kerstens HH. Epigenetic programming of monocyte-to-macrophage differentiation and trained innate immunity. Science. 2014;345(6204):1251086. doi: 10.1126/science.1251086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 521.Cheng SC, Quintin J, Cramer RA. mTOR- and HIF-1alpha-mediated aerobic glycolysis as metabolic basis for trained immunity. Science. 2014;345(6204):1250684. doi: 10.1126/science.1250684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 522.Aaby P, Kollmann TR, Benn CS. Nonspecific effects of neonatal and infant vaccination: public-health, immunological and conceptual challenges. Nat Immunol. 2014;15(10):895–899. doi: 10.1038/ni.2961. [DOI] [PubMed] [Google Scholar]
- 523.Delano MJ, Scumpia PO, Weinstein JS. MyD88-dependent expansion of an immature GR-1+CD11b+ population induces T cell suppression and Th2 polarization in sepsis. J Exp Med. 2007;204(6):1463–1474. doi: 10.1084/jem.20062602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 524.Gervassi A, Lejarcegui N, Dross S. Myeloid derived suppressor cells are present at high frequency in neonates and suppress in vitro T cell responses. PLoS ONE. 2014;9(9):e107816. doi: 10.1371/journal.pone.0107816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 525.Walton AH, Muenzer JT, Rasche D. Reactivation of multiple viruses in patients with sepsis. PLoS ONE. 2014;9(2):e98819. doi: 10.1371/journal.pone.0098819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 526.Engle WA. Surfactant-replacement therapy for respiratory distress in the preterm and term neonate. Pediatrics. 2008;121(2):419–432. doi: 10.1542/peds.2007-3283. [DOI] [PubMed] [Google Scholar]
- 527.Aikio O, Vuopala K, Pokela ML, Hallman M. Diminished inducible nitric oxide synthase expression in fulminant early-onset neonatal pneumonia. Pediatrics. 2000;105(5):1013–1019. doi: 10.1542/peds.105.5.1013. [DOI] [PubMed] [Google Scholar]
- 528.Pfenninger J, Tschaeppeler H, Wagner BP. The paradox of adult respiratory distress syndrome in neonates. Pediatr Pulmonol. 1991;10(1):18–24. doi: 10.1002/ppul.1950100105. [DOI] [PubMed] [Google Scholar]
- 529.Luce WA, Hoffman TM, Bauer JA. Bench-to-bedside review: Developmental influences on the mechanisms, treatment and outcomes of cardiovascular dysfunction in neonatal versus adult sepsis. Crit Care. 2007;11(5):228. doi: 10.1186/cc6091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 530.Roberts JD, Jr, Fineman JR, Morin FC., 3rd Inhaled nitric oxide and persistent pulmonary hypertension of the newborn. N Engl J Med. 1997;336(9):605–610. doi: 10.1056/NEJM199702273360902. [DOI] [PubMed] [Google Scholar]
- 531.Farrow KN, Fliman P, Steinhorn RH. The diseases treated with ECMO: focus on PPHN. Semin Perinatol. 2005;29(1):8–14. doi: 10.1053/j.semperi.2005.02.003. [DOI] [PubMed] [Google Scholar]
- 532.Maclaren G, Butt W. Extracorporeal membrane oxygenation and sepsis. Crit Care Resusc. 2007;9(1):76–80. [PubMed] [Google Scholar]
- 533.Martin CR, Dammann O, Allred EN. Neurodevelopment of extremely preterm infants who had necrotizing enterocolitis with or without late bacteremia. J Pediatr. 2010;157(5):751–756.e1. doi: 10.1016/j.jpeds.2010.05.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 534.Mitha A, Foix-L'Helias L, Arnaud C. Neonatal infection and 5-year neurodevelopmental outcome of very preterm infants. Pediatrics. 2013;132(2):e372–e380. doi: 10.1542/peds.2012-3979. [DOI] [PubMed] [Google Scholar]
- 535.Schlapbach LJ, Aebischer M, Adams M. Impact of sepsis on neurodevelopmental outcome in a Swiss national cohort of extremely premature infants. Pediatrics. 2011;128(2):e348–e357. doi: 10.1542/peds.2010-3338. [DOI] [PubMed] [Google Scholar]
- 536.Strunk T, Inder T, Wang X. Infection-induced inflammation and cerebral injury in preterm infants. Lancet Infect Dis. 2014;14(8):751–762. doi: 10.1016/S1473-3099(14)70710-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 537.Volpe JJ. Postnatal sepsis, necrotizing entercolitis, and the critical role of systemic inflammation in white matter injury in premature infants. J Pediatr. 2008;153(2):160–163. doi: 10.1016/j.jpeds.2008.04.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 538.Chau V, Brant R, Poskitt KJ. Postnatal infection is associated with widespread abnormalities of brain development in premature newborns. Pediatr Res. 2012;71(3):274–279. doi: 10.1038/pr.2011.40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 539.Shah DK, Doyle LW, Anderson PJ. Adverse neurodevelopment in preterm infants with postnatal sepsis or necrotizing enterocolitis is mediated by white matter abnormalities on magnetic resonance imaging at term. J Pediatr. 2008;153(2):170–175.e1. doi: 10.1016/j.jpeds.2008.02.033. [DOI] [PubMed] [Google Scholar]
- 540.Procianoy RS, Silveira RC. Association between high cytokine levels with white matter injury in preterm infants with sepsis. Pediatr Crit Care Med. 2012;13(2):183–187. doi: 10.1097/PCC.0b013e3182231074. [DOI] [PubMed] [Google Scholar]
- 541.Ellison VJ, Mocatta TJ, Winterbourn CC. The relationship of CSF and plasma cytokine levels to cerebral white matter injury in the premature newborn. Pediatr Res. 2005;57(2):282–286. doi: 10.1203/01.PDR.0000148286.53572.95. [DOI] [PubMed] [Google Scholar]
- 542.Stoll BJ, Hansen N, Fanaroff AA. To tap or not to tap: high likelihood of meningitis without sepsis among very low birth weight infants. Pediatrics. 2004;113(5):1181–1186. doi: 10.1542/peds.113.5.1181. [DOI] [PubMed] [Google Scholar]
- 543.ter Horst HJ, van Olffen M, Remmelts HJ. The prognostic value of amplitude integrated EEG in neonatal sepsis and/or meningitis. Acta Paediatr. 2010;99(2):194–200. doi: 10.1111/j.1651-2227.2009.01567.x. [DOI] [PubMed] [Google Scholar]
- 544.Basu S, Dewangan S, Shukla RC. Cerebral blood flow velocity in early-onset neonatal sepsis and its clinical significance. Eur J Pediatr. 2012;171(6):901–909. doi: 10.1007/s00431-011-1643-y. [DOI] [PubMed] [Google Scholar]
- 545.Das BK, Agarwal P, Agarwal JK, Mishra OP. Serum cortisol and thyroid hormone levels in neonates with sepsis. Indian J Pediatr. 2002;69(8):663–665. doi: 10.1007/BF02722699. [DOI] [PubMed] [Google Scholar]
- 546.Ng PC, Lam CW, Fok TF. Refractory hypotension in preterm infants with adrenocortical insufficiency. Arch Dis Child Fetal Neonatal Ed. 2001;84(2):F122–F124. doi: 10.1136/fn.84.2.F122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 547.Baker CF, Barks JD, Engmann C. Hydrocortisone administration for the treatment of refractory hypotension in critically ill newborns. J Perinatol. 2008;28(6):412–419. doi: 10.1038/jp.2008.16. [DOI] [PubMed] [Google Scholar]
- 548.Soliman AT, Taman KH, Rizk MM. Circulating adrenocorticotropic hormone (ACTH) and cortisol concentrations in normal, appropriate-for-gestational-age newborns versus those with sepsis and respiratory distress: cortisol response to low-dose and standard-dose ACTH tests. Metabolism. 2004;53(2):209–214. doi: 10.1016/j.metabol.2003.09.005. [DOI] [PubMed] [Google Scholar]
- 549.Noori S, Friedlich P, Wong P. Hemodynamic changes after low-dosage hydrocortisone administration in vasopressor-treated preterm and term neonates. Pediatrics. 2006;118(4):1456–1466. doi: 10.1542/peds.2006-0661. [DOI] [PubMed] [Google Scholar]
- 550.Seri I, Tan R, Evans J. Cardiovascular effects of hydrocortisone in preterm infants with pressor-resistant hypotension. Pediatrics. 2001;107(5):1070–1074. doi: 10.1542/peds.107.5.1070. [DOI] [PubMed] [Google Scholar]
- 551.Irakam A, Miskolci V, Vancurova I, Davidson D. Dose-related inhibition of proinflammatory cytokine release from neutrophils of the newborn by dexamethasone, betamethasone, and hydrocortisone. Biol Neonate. 2002;82(2):89–95. doi: 10.1159/000063094. [DOI] [PubMed] [Google Scholar]
- 552.Momtaz HE, Sabzehei MK, Rasuli B, Torabian S. The main etiologies of acute kidney injury in the newborns hospitalized in the neonatal intensive care unit. J Clin Neonatol. 2014;3(2):99–102. doi: 10.4103/2249-4847.134691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 553.Mathur NB, Agarwal HS, Maria A. Acute renal failure in neonatal sepsis. Indian J Pediatr. 2006;73(6):499–502. doi: 10.1007/BF02759894. [DOI] [PubMed] [Google Scholar]
- 554.Stojanovic V, Barisic N, Milanovic B, Doronjski A. Acute kidney injury in preterm infants admitted to a neonatal intensive care unit. Pediatr Nephrol. 2014;29(11):2213–2220. doi: 10.1007/s00467-014-2837-0. [DOI] [PubMed] [Google Scholar]
- 555.Bauer J, Hentschel R, Linderkamp O. Effect of sepsis syndrome on neonatal oxygen consumption and energy expenditure. Pediatrics. 2002;110(6):e69. doi: 10.1542/peds.110.6.e69. [DOI] [PubMed] [Google Scholar]
- 556.Romeo C, Eaton S, Spitz L, Pierro A. Nitric oxide inhibits neonatal hepatocyte oxidative metabolism. J Pediatr Surg. 2000;35(1):44–48. doi: 10.1016/s0022-3468(00)80011-1. [DOI] [PubMed] [Google Scholar]
- 557.Eaton S. Impaired energy metabolism during neonatal sepsis: the effects of glutamine. Proc Nutr Soc. 2003;62(3):745–751. doi: 10.1079/PNS2003284. [DOI] [PubMed] [Google Scholar]
- 558.Spasojevic I, Obradovic B, Spasic S. Bench-to-bedside review: neonatal sepsis - redox processes in pathogenesis. Crit Care. 2012;16(3):221. doi: 10.1186/cc11183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 559.Kato S, Hussein MH, Kakita H. Edaravone, a novel free radical scavenger, reduces high-mobility group box 1 and prolongs survival in a neonatal sepsis model. Shock. 2009;32(6):586–592. doi: 10.1097/SHK.0b013e3181a2b886. [DOI] [PubMed] [Google Scholar]
- 560.Doughty LA, Kaplan SS, Carcillo JA. Inflammatory cytokine and nitric oxide responses in pediatric sepsis and organ failure. Crit Care Med. 1996;24(7):1137–1143. doi: 10.1097/00003246-199607000-00012. [DOI] [PubMed] [Google Scholar]
- 561.Doughty L, Carcillo JA, Kaplan S, Janosky J. Plasma nitrite and nitrate concentrations and multiple organ failure in pediatric sepsis. Crit Care Med. 1998;26(1):157–162. doi: 10.1097/00003246-199801000-00032. [DOI] [PubMed] [Google Scholar]