

Abstract
Bacteria and viruses pose serious challenges for humans because they evolve continuously. Despite ongoing efforts, antiviral drugs to treat many of the most troubling viruses have not been approved yet. The recent launch of new antimicrobials is generating hope as more and more pathogens around the world become resistant to available drugs. But extra effort is still needed. One of the current strategies for antiviral and antibiotic drug development is the search for host cellular pathways used by many different pathogens. For example, many viruses and bacteria alter lipid synthesis and transport to build their own organelles inside infected cells. The characterization of these interactions will be fundamental to identify new targets for antiviral and antibiotic drug development. This review discusses how viruses and bacteria subvert cell machineries for lipid synthesis and transport and summarises the most promising compounds that interfere with these pathways.
1. INTRODUCTION
Emerging viruses and multidrug‐resistant bacteria constitute a serious public health problem, which will likely become a global safety challenge over the next decades. Faced with this situation, the need for searching new broad‐spectrum antivirals and antibiotics has emerged. Most antibiotics in late‐stage clinical development belong to existing antibiotic classes, and a few are new compounds directed against novel targets (Fernandes & Martens, 2017). Blocking cellular pathways used by pathogens in combined therapies against several targets is an interesting strategy to treat viral and bacterial infections while avoiding antiviral or antibiotic resistance.
Classically, the study of pathogen–host interactions had been focused on proteins and nucleic acids from the pathogen and the host. However, in recent years, lipidomics has gained importance in virology and bacteriology. Lipids are much more than structural components of membranes or molecules for energy storage, and they also play important roles in cellular signalling and regulatory processes in both healthy and infected cells.
The lipid composition of membranes affects their morphology and biophysical properties such as charge, curvature, width, packing defects, and fluidity. The study of changes in lipid biosynthesis pathways and transport in infected cells is a promising research field because many viruses and bacteria manipulate host membranes and lipid flows (Altan‐Bonnet, 2017; Toledo & Benach, 2015; van der Schaar, Dorobantu, Albulescu, Strating, & van Kuppeveld, 2016). In this review, we summarise the main pathways of lipid synthesis and transport hijacked by viruses and bacteria and the drugs that interfere with these general processes.
2. VIRUSES
Viruses are obligate intracellular pathogens that require the host cell machinery to replicate. Both DNA and RNA viruses usurp and take advantage of cellular membranes at different steps of their replication cycle. Viruses are able to induce the synthesis of new membranes and reorganise membranes of cell compartments to generate new organelles known as replication organelles or viral factories, which support viral genome replication and assembly of virus progeny. The use of membranes from organelles of the secretory pathway is quite widespread among RNA viruses. For instance, flaviviruses such as dengue virus (DENV), West Nile virus (WNV), and hepatitis C virus (HCV; Gillespie, Hoenen, Morgan, & Mackenzie, 2010; Romero‐Brey et al., 2012; Welsch et al., 2009), reoviruses (Tenorio et al., 2018), and severe acute respiratory syndrome coronavirus (Knoops et al., 2008) exploit endoplasmic reticulum (ER) membranes. The Golgi apparatus is used by Bunyaviruses (Fontana, Lopez‐Montero, Elliott, Fernandez, & Risco, 2008) and enteroviruses (Limpens et al., 2011). Flock House virus (FHV) builds factories in mitochondria (Kopek, Perkins, Miller, Ellisman, & Ahlquist, 2007), tomato bushy stunt virus (TBSV) uses peroxisomes (Fernandez de Castro, Fernandez, Barajas, Nagy, & Risco, 2017), rubella virus uses lysosomes (Fontana et al., 2010), and Sindbis virus assembles replication complexes at the plasma membrane, endosomes, and lysosomes (Frolova, Gorchakov, Pereboeva, Atasheva, & Frolov, 2010).
Viral genome replication in cell membranes has important advantages (Altan‐Bonnet, 2017): (a) restricting movement on the two‐dimensional plane concentrates viral replicases and saves time during replication; (b) activation of viral polymerases that depend on binding to specific membrane lipids; and (c) the assembly of viral replication complexes (VRCs) in pocket‐like structures protects the viral machinery from detection by the host cell innate immune defences.
Membrane‐associated viral replication is closely related to lipid synthesis and transport. Viruses generally subvert lipid flows by three different mechanisms: the translocation and/or regulation of lipid biosynthesis enzymes, the interference with lipid‐mediated signalling through the regulation of phosphatidylinositol (PI) kinases, and the regulation of lipid trafficking at membrane contact sites (MCSs) between adjacent organelles. Here, we present the current knowledge on these pathways and a list of drugs that interfere with them and have been identified to exhibit antiviral capacity (summarised in Figures 1 and 2).
Figure 1.

Schematic summary of host lipid synthesis and transport pathways used by viruses and bacteria. Compounds that interfere with these pathways and have antiviral or antibiotic activity are indicated. (1) Disruption of lipid rafts affects West Nile virus, Newcastle disease virus, human immunodeficiency virus‐1, and influenza A, bovine parainfluenza, and syncytial viruses and Salmonella typhimurium, Shigella, Mycobacterium avium, Pseudomonas aeruginosa, and Chlamydia trachomatis infection. (2) Inhibition of phospholipase PLA2G4A impairs hepatitis C virus (HCV) replication. (3) Inhibition of 3‐hydroxy‐3‐methyl‐glutaryl‐coenzyme A (HMG‐CoA) reductase by statins interferes with infection of many viruses and bacteria such as West Nile virus (WNV), dengue virus (DENV), tomato bushy stunt virus, Chlamydia pneumoniae, Staphylococcus aureus, Staphylococcus pneumoniae, Mycobacterium tuberculosis, Klebsiella pneumoniae, Escherichia coli, and Coxiella burnetii. (4) Inhibition of acyl‐CoA:cholesterol (Ch) acyltransferase (ACAT) blocks infection of HCV and C. trachomatis. (5 and 6) Inhibition of two different steps of triglyceride (TG) synthesis impairs HCV assembly. (7) When fatty acid (FA) synthase (FASN) is inhibited by C75 or cerulenin, replication of WNV, Usutu virus, DENV, and Flock House virus is blocked. (8) Inhibition of FA‐desaturase stearoyl‐CoA desaturase 1 (SCD1) decreases HCV and DENV infectivity. (9) Blockage of ceramide (Cer) synthesis at two different steps impairs replication of viruses such as WNV, HCV, and hepatitis B virus. (10) Inhibition of the lipid transfer protein Cer transfer protein (CERT) impairs HCV particle release and C. trachomatis infection. (11) Inhibition of sphingomyelin (SM) synthase alters HCV viral cycle. (12) Inhibition of sphingomyelinase (Asm) acts as an antibiotic against S. aureus, P. aeruginosa, Neisseria gonorrhoeae, M. tuberculosis, Mycobacterium smegmatis, E. coli, and C. burnetii. (13) HCV replication decreases when treating with uridine diphosphate–glucose Cer synthase (UGCG) synthetic inhibitors. (14) Inhibition of oxysterol‐binding protein (OSBP) impairs replication of enterovirus, encephalomyocarditis virus, and HCV. (15) Inhibition of Niemann–Pick type C1 protein (NPC1) impedes the entry of Ebola virus and other filoviruses because NPC1 is the specific cell receptor for these viruses; inhibition of NPC1‐mediated Ch transport impairs the replication of DENV, HEV, and HCV viruses and reduces the infection of C. trachomatis, Anaplasma phagocytophilum, and C. burnetii. (16) Inhibition of scramblase phospholipid scramblase‐1 (PLSCR1) reduces herpes simplex virus infection. Biosynthetic processes are compartmentalised in different subcellular locations or organelles. EFE: elder fruit extract; MβCD: methyl‐β‐cyclodextrin; PS: phosphatidylserine; PE: phosphatidylethanolamine; PC: phosphatidylcholine; TCA: tricarboxylic acid cycle; ACC: acetyl‐CoA carboxylase; ChE: cholesterol ester; DGAT1: diacylglycerol acyltransferase 1; DGAT2: diacylglycerol acyltransferase 2; LC‐FACS: long‐chain acyl‐CoA synthetase; MUFAs: monounsaturated fatty acids; PUFAs: polyunsaturated fatty acids; SMS: sphingomyelin synthase; CerS: ceramide synthase; SPT: serine palmitoyltransferase; FAPP2: four‐phosphate adaptor protein 2; GlcCer: glucosylceramide; GSLs: glycosphingolipids; ITZ: itraconazole; PI4P: phosphatidylinositol 4‐phosphate
Figure 2.
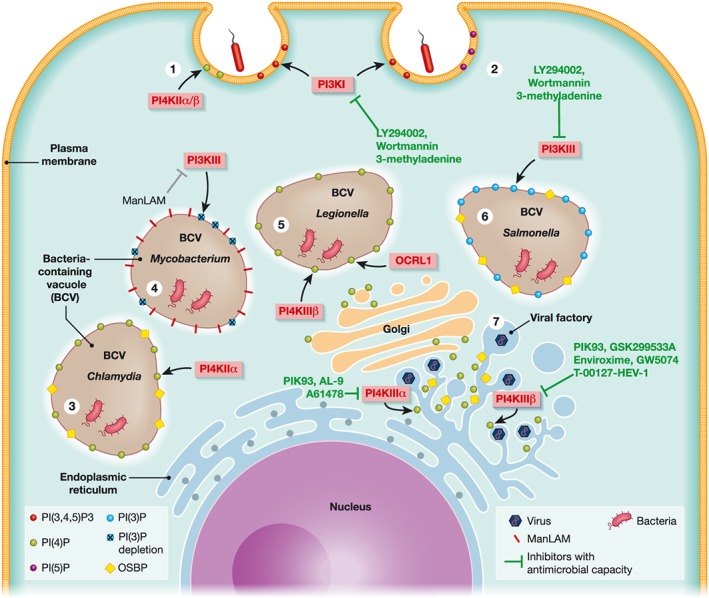
Schematic summary of phosphatidylinositol (PI)‐derived signalling pathways triggered by bacteria and viruses. Compounds that interfere with these pathways and have antiviral or antibiotic activity are indicated. (1) Listeria uses the InIB effector, which engages with host receptor Met and stimulates the recruitment of PI3KI to the bacterial entry sites. PI3K activation produces PI(3,4,5)P3, resulting in the reorganisation of cytoskeleton favouring the entry of Listeria. Both Listeria and Chlamydia additionally recruit PI4KIIα/β to the plasma membrane, where it produces PI(4)P also promoting uptake. (2) Brucella and Salmonella require PI3KI recruitment and activity for entering the cell. Alternately, Anaplasma phagocytophilum and Shigella need the subsequent PI3K–AKT pathway to delay apoptosis. Moreover, Shigella's entry site requires an enrichment of PI(5)P stimulated by IpgD. (3) After entry, Chlamydia recruits PI4KIIα to the inclusion, generating PI(4)P and leading to the recruitment of the lipid transfer protein oxysterol‐binding protein (OSBP). (4) Mycobacterium produces the mannose‐capped lipoarabinomannan (ManLAM) to interfere with PI3KIII activity. Disruption of PI3KIII reduces the amounts of PI(3)P on the inclusion surface and avoids phagosomal maturation. (5) Recruitment of PI4KIIIβ and OCRL1 together with the hijacking of the secretory pathway enriches Legionella inclusion in PI(4)P. (6) The Salmonella effector SopB recruits PI3KIII to the inclusion surface to maintain high levels of PI(3)P. The SseL effector binds to OSBP, which is recruited to the inclusion periphery. (7) Many different viruses recruit and exploit host PI4KIII kinases to build viral factories within reorganised cell membranes. Hepatitis C virus and encephalomyocarditis virus remodel endoplasmic reticulum membranes to build replication organelles with the assistance of PI4KIIIα that generates PI(4)P at the replication sites. In a similar manner, PI4KIIIβ is used by other viruses such as picornaviruses, enteroviruses, Coxsackievirus B3, and human rhinoviruses
2.1. Translocation and regulation of lipid biosynthesis enzymes
The development of metabolomics and lipidomics based on liquid chromatography and mass spectrometry techniques has contributed to characterize and monitor changes in the amount of different membrane and signalling lipids during viral infections. For instance, HCV infection has been discovered to increase cellular levels of cholesterol and sphingolipids (SLs) and to modify the phospholipid metabolism leading to an accumulation of phosphatidylcholine (PC) and triglyceride with longer and polyunsaturated fatty acid (FA) chains (Diamond et al., 2010; Gullberg et al., 2018; Hofmann et al., 2018; Roe, Kensicki, Mohney, & Hall, 2011). Lipidomics analysis of cells infected with other related flaviviruses such as DENV and WNV has revealed a marked increase in FAs, glycerophospholipids (GPLs), and SL content (Chotiwan et al., 2018; Perera et al., 2012) and has allowed to identify specific markers of infection, for example, lysophosphatidylcholine (Liebscher et al., 2018).
These techniques have also allowed the characterization of the lipidome of enveloped viruses. By this way, human immunodeficiency virus‐1 lipid envelope has been observed to be different from the producer cell plasma membrane, suggesting that viruses bud from specialised membrane subdomains, which are enriched in particular lipids (Lorizate et al., 2012).
Taken together, all these data prove the complex remodelling of the host lipid metabolism induced by viral infection.
2.1.1. Regulation of glycerophospholipid synthesis
GPLs are the major components of biological membranes. Their structure is based on a polar group head, which defines their charge and shape properties and allows the interaction with water and molecules near the external leaflet of the membrane, and a pair of FA hydrophobic tails.
The rate‐limiting step of GPLs biosynthesis relies on FA availability. In order to generate a larger pool of FA, HCV directly induces the overexpression of FA synthase (FASN), the key multienzyme complex for FA synthesis and therefore GPLs. This is a possible mechanism by which HCV infection alters the cellular lipid profile in liver and causes diseases such as steatosis (Yang et al., 2008). DENV nonstructural protein 3 (NS3) is responsible for FASN recruitment to replication factories (Heaton et al., 2010). After infection, cellular membranes showed a FASN‐dependent enrichment in unsaturated phospholipids and lipid signalling molecules such as sphingomyelin (SM; Perera et al., 2012). Two related flaviviruses, WNV and Usutu virus (USUV; Martin‐Acebes, Blazquez, Jimenez de Oya, Escribano‐Romero, & Saiz, 2011), and the nonrelated alphanodavirus FHV (Kampmueller & Miller, 2005) also showed an RNA replication dependency on FA synthesis. Thus, modulating FA synthesis can be an effective antiviral strategy.
DENV, WNV, USUV, and FHV viruses are sensitive to the inhibitory effect of cerulenin and C75, two specific inhibitors of FASN. Cerulenin is a natural product of the fungus Cephalosporium caerulens, which only inactivates the β‐ketoacyl synthase activity of FASN. C75 is a synthetic inhibitor that also inactivates the enoyl reductase and thioesterase partial activities of FASN with about the same rates as for inactivation of the β‐ketoacyl synthase (Rendina & Cheng, 2005). C75 was designed to be less reactive and safer than the classical inhibitor, cerulenin.
Appropriate membrane packing and fluidity are essential for viral RNA replication. These membrane properties mostly depend on the saturation grade and length of the FA hydrophobic tales. The stearoyl‐CoA desaturase 1 (SCD1) is a rate‐limiting enzyme in the biosynthesis of monounsaturated FAs from their saturated FA precursors. It catalyses the formation of a cis‐double bond at the Δ9 position of stearoyl and palmitoyl‐CoA. Brome mosaic virus (BMV) requires unsaturated FAs, and mutations in the host FA desaturases inhibit BMV replication (Lee, Ishikawa, & Ahlquist, 2001). Synthetic piperidine–aryl urea‐based inhibitors such as A939572 have been developed to specifically block SCD1, and they have been proven to impair HCV and DENV infection in cell culture (Gullberg et al., 2018; Nguyen et al., 2014). SCD1 inhibitors are claimed to be promising candidates to treat various diseases including cancer, Alzheimer's disease, and viral infections, so recently, novel SCD1 inhibitors have been developed and reported (reviewed by Uto, 2016).
PC and phosphatidylethanolamine (PE) are the most abundant GPLs in the cell. They are synthesized by two different metabolic routes, from a choline or ethanolamine polar head and a diacylglycerol backbone through the so‐called Kennedy pathways, which are the most exploited ways by most mammalian cells, and through the CDP–DAG pathway that is the main PC source for yeast. This alternative pathway consists of a lineal route by which PE suffers three consecutive methylations mediated by the choline‐requiring protein 2 (cho2) to become PC (reviewed by Henry, Kohlwein, & Carman, 2012).
Global PC synthesis is significantly enhanced during infection of many (+)ssRNA viruses such as DENV, FHV, poliovirus, and HCV (Castorena, Stapleford, & Miller, 2010; Vance, Trip, & Paddon, 1980; Zhang et al., 2016). Moreover, BMV replication protein 1a interacts with and recruits cho2 to the site of viral replication. In CHO2 deletion yeast mutants, BMV replication was inhibited up to 30‐fold (Zhang et al., 2016).
On the other hand, the plant tombusviruses TBSV, cucumber necrosis virus, and Carnation Italian ringspot virus and the unrelated insect Nodamura virus require high levels of PE at the viral replication sites. All these viruses take advantage of the increased PE level in cho2Δ yeast cells, whereas depletion of the PE levels reduces viral replication (Xu & Nagy, 2015). As PE has a small head resulting in a conical shape, it has been proposed to contribute to the induction of the characteristic negative curvature of single‐membrane vesicles or spherules with VRCs (Altan‐Bonnet, 2017). In addition, at the replication complex, the TBSV RNA‐dependent RNA polymerase (RdRp) is stimulated upon PE binding (Xu & Nagy, 2015) but inhibited by phosphatidylglycerol.
Although further investigation is needed to identify the particular PC and PE synthesis pathways used by different viruses, the fact that one pathway could supply the physiological function of the other provides the opportunity to develop antivirals targeting the enzymes subverted by viruses, for example, cho2.
Enteroviruses and other enveloped viruses accumulate phosphatidylserine (PS) at VRC subdomains (Amara & Mercer, 2015). PS accumulation has been usually associated to PS receptor‐dependent uptake or “apoptotic mimicry,” as we discuss below.
2.1.2. Regulation of lipid rafts and cholesterol synthesis
The cholesterol and SL content in biological membranes defines important physical and biological properties. These lipids do not homogeneously distribute in membranes but together constitute detergent‐resistant domains or lipid rafts, which are involved in membrane trafficking and cell signalling.
Cholesterol is a critical membrane component that determines membrane fluidity, helps to stabilise the curvature and packing defects of membranes, and regulates the formation and function of membrane‐bound complexes of lipids and proteins. Viruses take advantage of these properties to make specialised membrane platforms for RNA replication and virus assembly.
Lipid rafts have been proven to participate in key steps of the life of viruses, such as entry, RNA translation, virus progeny assembly, and egress (Shahsavandi, Ebrahimi, & Hasaninejad Farahani, 2017). For some arboviruses, cholesterol plays an important role in the process of virus entry into cells. For example, the Gc structural protein of Rift Valley fever virus directly recognises and binds to GPLs at the plasma membrane. Insertion of Gc is followed by accumulation of cholesterol and local membrane reorganisation (Guardado‐Calvo et al., 2017). Depletion of cholesterol and consequent disruption of lipid rafts by methyl‐β‐cyclodextrin (MβCD) decreased the infectivity of Newcastle disease virus, WNV, HCV, and bovine parainfluenza virus, mainly through blocking virus entry into cells (Kapadia, Barth, Baumert, McKeating, & Chisari, 2007; Li, Yu, & Hou, 2017; Mackenzie, Khromykh, & Parton, 2007; Martin, Holguera, Sanchez‐Felipe, Villar, & Munoz‐Barroso, 2012). During influenza A and syncitial virus infections, a similar effect has been observed after treatment with gemfribrozil (Bajimaya, Frankl, Hayashi, & Takimoto, 2017) and with a natural elder fruit extract (Shahsavandi et al., 2017).
Cholesterol, PI 4‐phosphate (PI4P), and PE concentrate also in membranes of VRCs. VRCs of poliovirus, rhinovirus, and Coxsackievirus B3 accumulate cholesterol (Hsu et al., 2010; Ilnytska et al., 2013; Roulin et al., 2014). To facilitate viral genome replication, WNV modulates host cholesterol homeostasis by redistributing cholesterol‐synthesizing enzymes and cellular cholesterol to replication sites (Mackenzie et al., 2007). Moreover, characteristic highly curved nanoperiodic cubic membranes imaged by electron microscopy in cells infected by several (+)ssRNA viruses belonging to the Coronaviridae, Picornaviridae, Togaviridae, and Flaviviridae families are thought to be linked to changes in cell cholesterol metabolism through activation of cellular 3‐hydroxy‐3‐methyl‐glutaryl‐coenzyme A reductase (HMG‐CoA reductase or HMGCR; Deng, Almsherqi, Ng, & Kohlwein, 2010).
In fact, many viruses are sensitive to inhibitors of HMGCR, the rate‐limiting enzyme of cholesterol biosynthesis. This is the case of statins used for treating hypercholesterolaemia in humans. Statins act by competitively inhibiting HMGCR. Because their molecular structure is similar to HMG‐CoA, statins fit into the catalytic active site of the enzyme and compete with the native substrate. This competition reduces the rate by which HMGCR produces mevalonate, the next molecule in the cascade that eventually generates cholesterol (Stancu & Sima, 2001). Thus, statins deplete cholesterol and disrupt lipid rafts.
For example, WNV, DENV, and TBSV are inhibited by lovastatin (Mackenzie et al., 2007; Rothwell et al., 2009; Sharma, Sasvari, & Nagy, 2010; Soto‐Acosta, Bautista‐Carbajal, Cervantes‐Salazar, Angel‐Ambrocio, & Del Angel, 2017), and DENV and human immunodeficiency virus‐1 are inhibited by simvastatin (Bryan‐Marrugo et al., 2016; DeLucia, Rinaldo, & Rappocciolo, 2018). HCV is also sensitive to different statins (Amemiya et al., 2008; Kim et al., 2007; Rao & Pandya, 2011).
2.1.3. Regulation of sphingolipid synthesis
Lipidomics studies have shown an increase in the content of ceramide (Cer) and SM in flavivirus‐infected cells. Cer has specifically been associated with WNV replication and viral particle biogenesis. SM is enriched in VRC membranes of viruses phylogenetically related to WNV, such as DENV and HCV (Martin‐Acebes et al., 2016).
HCV RdRp is activated by SM binding. Twenty SM molecules interact with a specific binding domain in the structure of HCV RdRp and change the conformation of its catalytic pocket leading to RNA recruitment and replication (Weng et al., 2010).
Inhibition of different enzymes that mediate SL biosynthesis decreases infectivity of different viruses. De novo synthesis of SLs initiates at the ER with the synthesis of sphinganine from palmitoyl‐CoA and serine by the enzyme serine palmitoyltransferase. Three inhibitors of serine palmitoyltransferase have been described to present antiviral activity against HCV, hepatitis B virus, and WNV: ISP‐1, myriocin, and NA255 (Aizaki et al., 2008; Aktepe, Pham, & Mackenzie, 2015; Amemiya et al., 2008; Sakamoto et al., 2005; Tatematsu, Tanaka, Sugiyama, Sudoh, & Mizokami, 2011; Weng et al., 2010). Then, sphinganine is acylated and transformed into Cer by the Cer synthase. Cer is then delivered to the Golgi complex, where it is converted into SM by the SM synthase or into complex glycosphingolipids by the uridine diphosphate–glucose Cer synthase. Pharmacological inhibition of SM synthase with D609, SPK‐601, and MS‐209 or inhibition of uridine diphosphate–glucose Cer synthase by N‐Butyldeoxynojirimycin (NB‐DMJ) and D,L‐threo‐1‐phenyl‐2‐decanoylamino‐3‐morpholino‐1‐propanol (PDMP), respectively, reduced the infectivity of HCV and WNV (Khan et al., 2014; Martin‐Acebes et al., 2016). Inhibition of Cer synthase by fumonisin B1 also impairs WNV replication (Aktepe et al., 2015). As observed, viruses of the Flaviviridae have been found to be the most sensitive to SL biosynthesis blockage.
Other lipid biosynthetic enzymes have been associated with viral infections. For example, HCV takes advantage of three lipid synthesis enzymes, which are related to the biogenesis and maintenance of cellular lipid droplets (LDs). LDs are dynamic organelles enriched in triglycerides and cholesterol esters that regulate the storage and hydrolysis of neutral lipids and participate in lipid energy balance and signalling, cell immunity, protein storage, and trafficking (Welte, 2015). After replication, HCV proteins are targeted to LD, where virus assembly takes place. Pharmacological inhibition of long‐chain acyl‐CoA synthetase, diacylglycerol acyltransferase 1, and acyl‐CoA:cholesterol acyltransferase results in the blockade of virion assembly and egress (Herker et al., 2010; Liefhebber, Hague, Zhang, Wakelam, & McLauchlan, 2014). Targeting synthesis of lipids stored at LDs might be an alternative for therapeutic treatment of chronic HCV infection. Other host factor involved in the recruitment of HCV to LD is the lipid‐modifying phospholipase PLA2G4A (Menzel, 2012). Picornaviruses have also been shown to utilise a different A2 phospholipase, the PLA2G16. In this case, the enzyme assists in genome delivery from virions into the cytoplasm after entry in cells (Staring et al., 2017).
2.2. Interfering with lipid‐mediated signalling
PI‐related lipids have recently emerged as signalling molecules involved in nearly all aspects of cellular regulation. The production of PI4P has been described to play a key regulatory role for a wide range of cellular activities, including membrane trafficking and biogenesis, via the recruitment of effector proteins related to lipid biosynthesis and transport.
The major pool of PI4P is synthesized in Golgi membranes by PI 4‐kinases type 3 (PI4KIII), distinguishing two different isoforms PI4PIIIα and PI4PIIIβ. Many different viruses exploit host PI4KIII kinases. For example, HCV, some enteroviruses, and picornaviruses stimulate PI4 kinases to generate PI4P in situ at replication sites. This local increase of PI4P has significant impact on membrane remodelling, assembly of VRCs, and viral genome replication. HCV nonstructural protein NS5A and encephalomyocarditis virus (EMCV) nonstructural protein 3A recruit and activate PI4KIIIα (Dorobantu et al., 2015; Reiss et al., 2011). However, enteroviruses use their 3A nonstructural protein to recruit PI4KIIIβ to VRCs (Hsu et al., 2010).
PI4KIIIβ is a specific target of protein kinase inhibitors such as enviroxime‐like compounds. Enviroxime, GW5074, and T‐00127‐HEV are used as antipoliovirus compounds (Arita et al., 2011), and other synthetic enviroxime‐related compounds as GSK2998533A and BF738735 (Arita et al., 2011; Roulin et al., 2014) are broad‐spectrum antivirals against different enteroviruses.
On the other hand, pharmacological inhibition of PI4KIIIα by AL‐9 decreases infectivity of Saffold virus 3, EMCV, and HCV (Dorobantu et al., 2015). Tyrphostin AG1478, a known epidermal growth factor receptor inhibitor and an anticancer chemotherapy agent, also inhibits PI4KIIIα activity in both in vitro and cell cultures and impairs replication of EMCV and HCV (Dorobantu et al., 2016).
In contrast, PIK93 inhibits both PI4KIIIβ and PI4PIIIα isoforms. This compound is efficient against enteroviruses, including poliovirus and Coxsackievirus B3, human rhinoviruses (Arita, 2014; Arita et al., 2011; Roulin et al., 2014), and HCV (Hsu et al., 2010). The PIK93 lack of specificity for PI4KIII isoforms and the fact that it inhibits PI3Ks at high concentrations raise concern about its use as an antiviral agent (Bishe, Syed, & Siddiqui, 2012). Although in vitro studies have not found significant problems with cellular viability, some in vivo studies have shown that animals treated with PIK93 and other related small PI4Ks inhibitors suffered from adverse effects. However, given that PI4KIII kinases are used by many different viruses, they are attractive targets for the development of broad‐spectrum antivirals. PI4KIII inhibitors with less toxicity and cross‐reactivity with other cellular kinases are currently under consideration (Rutaganira et al., 2016; van der Schaar et al., 2013).
2.3. Interfering with lipid trafficking at membrane contact sites
In most instances, proteins are delivered to their final destination in particular membranes by vesicular shuttles that originate at the ER and follow the secretory pathway. However, for many lipids, there are nonvesicular modes of transport. These are faster than vesicular trafficking and produce local lipid enrichment and considerable asymmetries in the lipid composition of membranes. Nonvesicular transport also ensures the supply of lipids to organelles out of the secretory pathway, such as mitochondria. Because lipids are insoluble in water, during nonvesicular transport, they are shielded from the water‐rich cytoplasm by proteins that hold them in an internal hydrophobic pocket. Such lipid transfer proteins (LTPs) often act in regions in which one organelle comes ~30 nm away from another membrane, the so‐called MCSs (Wong, Copic, & Levine, 2017). Plants induce LTPs' expression and secretion as a defence mechanism against pathogens (Kushwaha, Singh, Basu, & Chakraborty, 2015; Ooi et al., 2008; Park et al., 2002; Sohal, Pallas, & Jenkins, 1999). On the other hand, animal viruses hijack LTPs that mobilise cholesterol and SLs to build their replication neo‐organelles. The best documented cases are described in the next sections.
2.3.1. Oxysterol‐binding protein
Oxysterol‐binding protein (OSBP) exchanges PI4P and cholesterol at MCSs between Golgi apparatus and ER. Transport of cholesterol by OSBP is highly dependent on PI4KIII activity and PI4P levels at Golgi membranes. PI4KIIIs synthesize PI4P on the surface of Golgi membranes, and PI4P lipids serve as an anchor for OSBP, which drives transport and accumulation of cholesterol.
Different viruses belonging to the Picornaviridae family such as enterovirus, poliovirus, and EMCV, as well as HCV, another positive stranded nonrelated virus, use the so‐called PI4KIII–PI4P–OSBP pathway (Arita, 2014; van der Schaar et al., 2016).
Due to the observation that deleting OSBP does not compromise cellular viability, most probably because other redundant LTPs could make up for its function, the interest in studying OSBP as a target for broad‐spectrum antivirals has grown. To date, three different drugs have been developed for specifically inhibiting OSBP. Itraconazole, OSW‐1, and TTP‐8307 bind to the OSBP hydrophobic pocket and compete with cholesterol. They have been successfully tested against enterovirus, EMCV, and HCV (Albulescu et al., 2015; Albulescu et al., 2017; Strating et al., 2015; Wang et al., 2014).
2.3.2. Ceramide transfer protein and four‐phosphate adaptor protein 2
Both Cer transfer protein (CERT) and four‐phosphate adaptor protein 2 (FAPP2) mediate the traffic of SL precursors. Because these LTPs accumulate SLs at the replication sites, they are good targets to interfere with flaviviruses. CERT is responsible for transferring Cer from ER to Golgi where synthesis of complex SLs takes place (Hanada et al., 2003). CERT is necessary for HCV egress (Bishe et al., 2012), and its pharmacological inhibition with the synthetic compound HPA‐12 dramatically impairs HCV release (Aizaki et al., 2008; Yasuda et al., 2001). Moreover, CERT and OSBP are regulated by protein kinase D, so that phosphorylated forms of both LTPs are inactive (Amako, Syed, & Siddiqui, 2011). Drugs interfering with host protein kinase D such as CRT0066101, a PKD‐specific small‐molecule inhibitor (Harikumar et al., 2010), might become efficient antivirals.
The FAPP2 docks the Golgi through PI4P and shuttles glucosylceramide (Godi et al., 2004), a substrate of a subset of complex glycosphingolipids. FAPP2 localizes to HCV replication sites and mediates the accumulation of lactosylceramide at the VRCs. FAPP2 depletion disrupts HCV replication complexes and impairs HCV replication (Khan et al., 2014).
2.3.3. Niemann–Pick type C1 protein
Niemann–Pick type C1 protein (NPC1) was discovered while studying a human neurodegenerative disease characterized by the disruption of subcellular cholesterol transport. NPC1 is a glycoprotein with 13 transmembrane domains that localizes to late endosomes and transiently in lysosomes and trans‐Golgi network. NPC1 acts in tandem with NPC2 to carry out the transfer of extracellular and recycled free cholesterol from the endosomal/lysosomal system to trans‐Golgi network and ER (Ioannou, 2000). NPC1 is the cell receptor of Ebola virus, Marburg virus, and other filoviruses (Carette et al., 2011). U18666A has been developed as a specific inhibitor of NPC1 (Lu et al., 2015). The administration of U18666A and other multiple cationic amphiphiles induces the accumulation of cholesterol in endosomes and lysosomes and inhibits Ebola entry and infection (Shoemaker et al., 2013).
Targeting NPC1 also impairs the replication of other RNA viruses such as DENV (Poh et al., 2012) and hepatitis E virus (Yin, Ambardekar, Lu, & Feng, 2016). NPC1 has not been found to be the cellular receptor for these viruses, so the adverse effects on viral replication after treatment with U18666A or NPC1 depletion must be due to effects at a postentry step.
In the case of HCV, which takes advantage of the cellular endosomal and exosomal pathways to egress from the host cell, U18666A blocks the release of viral particles by inhibiting NPC1 and therefore the transport and functionality of multivesicular bodies. This leads to an accumulation of mature viral particles in exosomes. Moreover, U18666A leads to the formation of large dysfunctional lysosome‐like structures that mediate the degradation of these viral particles (Elgner et al., 2016).
2.4. Viral apoptosis mimicry
PS is an anionic phospholipid usually oriented to the cytosolic side of the plasma membrane. Under pathological and cellular stress situations, PS molecules are flipped and exposed on the cell surface, serving like an alert flag for detection by the immune system. PS exposure has been linked to apoptosis, infections, and cancer (Birge et al., 2016).
PS anionic lipids are present in the membranes of many enveloped RNA viruses, including DENV, WNV, chikungunya, Ebola, Lassa fever virus, human immunodeficiency virus, and DNA viruses such as vaccinia. PS‐enriched envelopes of viruses are recognised by cell surface PS receptors that mediate virus uptake in a process known as viral apoptosis mimicry (Amara & Mercer, 2015).
Exposed anionic phospholipids on infected cells may provide a new target for the generation of antiviral agents against multiple classes of viruses (Soares, King, & Thorpe, 2008). Bavituximab, a PS‐targeting monoclonal antibody, successfully blocks infectivity of enveloped viruses such as Ebola and Lassa fever viruses in in vitro assays and in small animal models (Dowall et al., 2015; Soares et al., 2008).
The molecular mechanism by which PS is accumulated into viral budding sites in not yet well understood. Recently, the OSBP‐related proteins ORP5 and ORP8 have been shown to facilitate the PS/PI4P exchange at ER–plasma membrane MCSs (Chung et al., 2015). Another study suggests that the phospholipid scramblase‐1, a calcium‐responsive enzyme that flips PS between membrane leaflets, might participate in PS extracellular exposure in cells infected with herpes virus. Inhibition of phospholipid scramblase‐1 with its antagonist R5421 prevents the virus‐induced exposure of PS on the cell surface and reduces virus infection (Cheshenko, Pierce, & Herold, 2018).
3. BACTERIA
Bacteria have developed a number of strategies to modify host lipids during infection. The nature and effects of these changes are not fully understood, but it is known that some bacteria manipulate lipids to survive inside eukaryotic cells or to spread by triggering cell death (Toledo & Benach, 2015). Signalling cascades related to cholesterol, SLs, and phosphoinositides are particularly relevant during bacterial infections. Many different bacteria need cholesterol for entering cells or building neo‐organelles (Stehr, Elamin, & Singh, 2012). However, bacteria cannot synthetize cholesterol and need to obtain it from the infected cell. Acid sphingomyelinase (Asm) is one of the hydrolases that generates Cer from SM. Asm activation produces Cer‐enriched domains in the plasma membrane that are used by bacteria for entering cells or to trigger apoptosis. On the other hand, Asm is also crucially involved in the host response against pathogenic bacteria. Finally, phosphoinositides (PtdIns) and related enzymes control many processes including vesicular trafficking, actin polymerisation, and different signalling cascades that can modify membranes during phagocytosis or activate the endocytic pathway. Bacteria hijack all these pathways together with LTPs.
Here, we summarise the strategies used by different pathogenic bacteria to subvert lipid synthesis and transport. Targeting these pathways can generate new efficient antibiotics (summarised in Figures 1 and 2).
3.1. Regulation of lipid biosynthesis
3.1.1. Cholesterol and lipid rafts
De novo cholesterol biosynthesis occurs in the ER, with the conversion of HMG‐CoA to mevalonate by HMGCR. Cellular cholesterol levels can also be increased by the uptake of cholesterol bound to low‐density lipoprotein (LDL) via the LDL receptor. When cellular cholesterol levels are high, it can be transported out of the cell, broken down into bile acids or steroids, or esterified and stored in LDs. In eukaryotic cells, cholesterol and SLs are the main components of membrane microdomains known as lipid rafts. These microdomains that contain tightly packed receptor clusters and other signalling proteins are involved in many signal transduction pathways.
Anaplasma phagocytophilum and Brucella species enter cells in a lipid raft‐dependent manner (Naroeni & Porte, 2002; Xiong, Lin, & Rikihisa, 2009). Pharmacological disruption of cholesterol flows impairs the assembly of lipid rafts. Thus, treatment with U18666A (Xiong & Rikihisa, 2012), ketoconazole, or filipin III (Watarai et al., 2002; Watarai, Makino, Fujii, Okamoto, & Shirahata, 2002) makes these bacteria incapable of infecting cells. Chlamydia has developed different strategies to incorporate lipids via vesicular and nonvesicular transport (Hackstadt, Scidmore, & Rockey, 1995; Hatch & McClarty, 1998; Wylie, Hatch, & McClarty, 1997). Cholesterol depletion with MβCD (Jutras, Abrami, & Dautry‐Varsat, 2003; Korhonen et al., 2012), simvastatin (Erkkila et al., 2005), U18666A (Beatty, 2006), and an acyl‐CoA:cholesterol acyltransferase inhibitor (Peters & Byrne, 2015) have shown that elementary bodies, the extracellular infectious stage of Chlamydia, attaches to cholesterol‐rich sites at the plasma membrane. However, the importance of lipid rafts in Chlamydia entry is a matter of controversy because some studies have shown that Chlamydia can enter cells in a way that does not depend on lipid rafts (Gabel, Elwell, van Ijzendoorn, & Engel, 2004; Norkin, Wolfrom, & Stuart, 2001). Contradictory results can be attributed to studies with various bacterial isolates and different host cells.
After entering cells, A. phagocytophilum, Brucella, Mycobacterium, Salmonella, Chlamydia, and Coxiella burnetii establish their replicative inclusions in membrane‐bound compartments derived from phagosomes or organelles of the endocytic pathway (Alpuche‐Aranda, Berthiaume, Mock, Swanson, & Miller, 1995; Heinzen, Scidmore, Rockey, & Hackstadt, 1996; Niu, Yamaguchi, & Rikihisa, 2008; Oh et al., 1996). Although these bacteria lack the genes for cholesterol biosynthesis, cholesterol content increases in infected cells. In particular, Chlamydia spp., C. burnetii, and A. phagocytophilum modulate host cholesterol homeostasis at the level of gene transcription, such as the LDL receptor or the HMGCR (Gilk, 2012; Lin & Rikihisa, 2003). Chlamydia downregulates cholesterol efflux and requires de novo cholesterol synthesis (Bashmakov, Zigangirova, Pashko, Kapotina, & Petyaev, 2010), whereas A. phagocytophilum alters LDL uptake by infected cells and causes a twofold increase of their cholesterol content (Xiong et al., 2009). C. burnetii increases the cholesterol content of infected cells by 70% (Czyz et al., 2014; Howe & Heinzen, 2006). During early stages of infection, the inclusion membrane of C. burnetii has low levels of cholesterol, but at later stages, the amount of cholesterol increases, which suggests a temporal regulation of cholesterol biosynthesis and uptake. C. burnetii has been shown to be sensitive to the treatment with statins (Howe & Heinzen, 2006). Mycobacterium and Salmonella do not need lipid rafts for entry, but intracellular cholesterol is important for infection. As mentioned above, Mycobacterium cannot synthesize cholesterol and needs to obtain cholesterol from the host through alternative means such as the mce4 gene cluster and the ABC‐like‐ATP‐dependent system that import cholesterol into bacteria. Disrupting cholesterol metabolism diminishes Mycobacterium entry (Kaul, Anand, & Verma, 2004), replication (de Chastellier & Thilo, 2006; Miner, Chang, Pandey, Sassetti, & Sherman, 2009; Mohn et al., 2008; Parihar et al., 2014; Russell, Cardona, Kim, Allain, & Altare, 2009), and subsequent stages of infection (Nesbitt et al., 2010; Pandey & Sassetti, 2008). Salmonella uses the pathogenicity island 1 and 2 type III secretion systems that localize in the lumen of the inclusion, where they mediate the esterification of cholesterol (Haraga, Ohlson, & Miller, 2008; Nawabi, Catron, & Haldar, 2008). Pharmacological disruption of cholesterol with statins (Catron et al., 2004) or MβCD (Huang, 2011) reduces intracellular Salmonella growth. Other bacteria such as Pseudomonas aeruginosa (Kowalski & Pier, 2004), Staphylococcus pneumoniae (Bergman et al., 2011), Staphylococcus aureus (Jerwood & Cohen, 2008), Escherichia coli, and Klebsiella pneumoniae (Al‐Kuraishy, Al‐Gareeb, & Al‐Buhadily, 2018) are also susceptible to the treatment with statins. Pharmacological inhibition of cholesterol and genetic depletion experiments confirmed that targeting cholesterol can lead to efficient strategies against all these bacteria by reducing entry and replication in host cells.
3.1.2. Sphingolipid and phospholipid synthesis
Although incorporation of Cer to lipid rafts is essential for the initiation of cell defence against pathogens such as Salmonella (Huang, 2016; McCollister, Myers, Jones‐Carson, Voelker, & Vazquez‐Torres, 2007), this process is used by some bacteria to infect cells. During Mycobacterium infection, Asm contributes to the fusion of the Mycobacterium‐containing vacuole with the phagosome, a process that mediates the elimination of bacteria (Vazquez et al., 2016). Mycobacterium avoids this host defence via PI signalling. Thus, Mycobacterium tuberculosis synthesizes glycosylated phosphoinositides such as mannose‐capped lipoarabinomannan (ManLAM; Beatty et al., 2000; Fratti, Chua, Vergne, & Deretic, 2003). Furthermore, the phosphatase activity of SapM, MptpB, and MptpA reduces the amount of PtdIns(3)P on the Mycobacterium‐containing vacuole surface (Chua & Deretic, 2004). Both ManLAM and phosphatase effectors interfere with phagosomal maturation and degradation of bacteria (Castandet et al., 2005; Singh et al., 2003). Interestingly, Asm is also important for Mycobacterium infection. Pharmacological inhibition of Asm with imipramine (Godbole et al., 2015) and infection in Asm‐deficient mice (Utermohlen, Herz, Schramm, & Kronke, 2008) showed that the growth of M. tuberculosis, Mycobacterium smegmatis, and Mycobacterium avium was blocked. Moreover, Asm‐deficient mice were more resistant to Mycobacterium lethal infection than wild‐type mice. Additionally, both Neisseria and Pseudomonas trigger Asm activation, Cer release, and the assembly of Cer‐enriched rafts that are essential for infection (Grassme et al., 1997; Grassme et al., 2003; Hauck et al., 2000). Cer‐enriched platforms serve as scaffold for the recruitment of cellular receptors such as the cystic fibrosis transmembrane conductance regulator, used by Pseudomonas to enter cells (Schroeder et al., 2002). Deficiency of Asm and specific pharmacological inhibition blocked Neisseria (Grassme et al., 1997; Hauck et al., 2000; Simonis, Hebling, Gulbins, Schneider‐Schaulies, & Schubert‐Unkmeir, 2014) and Pseudomonas (Becker et al., 2010; Cremesti et al., 2001; Riethmuller et al., 2009) entry and infection. Asm inhibitors were also effective against other bacteria such as S. aureus (Peng et al., 2015) and E. coli (Falcone et al., 2004), demonstrating their potential as broad‐spectrum antibiotics.
Chlamydia incorporates lipid storage organelles into the inclusion lumen (Cocchiaro, Kumar, Fischer, Hackstadt, & Valdivia, 2008), and it has its own machinery to synthetize GPLs such as PE, phosphatidylglycerol, and PS but needs to hijack other host‐derived lipids such as FAs. Chlamydia also needs to transport cholesterol and SLs to the bacterial cell wall (Yao, Cherian, Frank, & Rock, 2015; Yao, Dodson, Frank, & Rock, 2015). There is a close association between Golgi apparatus, where SM is synthetized, and Chlamydia inclusions. Chlamydia intercepts the exocytic transport of SM‐containing vesicles and redirects them to the inclusions (Wolf & Hackstadt, 2001). For this purpose, Chlamydia also uses the CERT whose role in infection will be discussed below.
3.2. Interfering with lipid‐mediated signalling
PI‐related lipids are signalling molecules involved in a wide range of cellular processes such as eukaryotic signal transduction, cytoskeleton rearrangement, and membrane dynamics. The plasma membrane is enriched in PI(4,5)P2 and PI(4)P, phagosomes and early endosomes are enriched in PI(3)P, and multivesicular bodies and late endosomes contain PI(3)P and PI(3,5)P2. Bacteria subvert PtdIns flows by encoding enzymes, principally phosphatases, and by altering the expression or localization of host proteins that modulate PI intracellular levels. The presence of different species of lipids on the surface of bacterial inclusions creates an optimal micro‐environment for recruiting specific proteins, shooting signalling cascades, and avoiding cellular defences.
Two mayor signalling cascades have been proposed for Listeria invasion (Mengaud et al., 1996; Shen, Naujokas, Park, & Ireton, 2000). These two pathways depend on virulence factors InIA or InIB. InIB interaction with its natural receptor Met (Marino, Braun, Cossart, & Ghosh, 1999; Shen et al., 2000) triggers the activation and translocation of PI3KIA to Listeria monocytogenes entry site. The subsequent signalling cascade produces an increase in lipid products, in particular PI(3,4,5)P3 (Ireton et al., 1996; Ireton, Payrastre, & Cossart, 1999) that recruits effectors for actin reorganisation. PI3K cascade and cytoskeletal rearrangements are critical for L. monocytogenes invasion, and that is why PI3K inhibitors such as wortmannin and LY294002 block infection (Ireton et al., 1996). During Shigella infection, the sites of internalisation colocalize with PI(5)P, which is produced with the participation of the bacterial phosphatase IpgD. The accumulation of PI5P at the plasma membrane reduces tether forces and favours entry. When PI3KI–AKT pathway is activated, the regulation of antiapoptotic effectors is triggered. However, blocking PI3K with wortmannin does not affect Shigella entry, suggesting that PI3KI activity is not involved in invasion but in later steps of infection (Lafont, Tran Van Nhieu, Hanada, Sansonetti, & van der Goot, 2002; Pendaries et al., 2006). A. phagocytophilum also takes advantage of the PI3K–AKT pathway to delay apoptosis, something observed in experiments of PI3K pharmacological inhibition with LY294002 (Khanal, Sultana, Catravas, Carlyon, & Neelakanta, 2017; Sarkar et al., 2012). Brucella also requires both PI3KI and PI3KII activity for entering the cell and replicate (Guzman‐Verri et al., 2001). Salmonella can modulate PI‐derived signalling at different stages of infection via the bacterial phosphatase effector SopB. By a not well‐understood SopB‐dependent process, PI3KI converts PI(4,5)P2 to PI(3,4,5)P3 at the plasma membrane and produces a double effect: First, the reduction of interactions between membrane and cytoskeleton leads to bacterial invasion (Terebiznik et al., 2002), and second, the recruitment of AKT activates cell survival pathways that avoid apoptosis. SopB also reduces PI(4,5)P2 and PS levels on the Salmonella vacuole surface, where charge diminishes avoiding the fusion with lysosomes (Bakowski et al., 2010). Another essential SopB function for Salmonella replication is the recruitment of Rab5 and PI3KIII to the inclusion surface, which is required to maintain high levels of PI(3)P (Bujny et al., 2008; Hernandez, Hueffer, Wenk, & Galan, 2004). Two studies (Auweter et al., 2011; Auweter, Yu, Arena, Guttman, & Finlay, 2012) showed that SseL, another Salmonella effector, interacts with the lipid transfer protein OSBP and enhances Salmonella replication, probably by modifying the lipid composition of the replicative vacuole. In contrast, Mycobacterium alters PI(3)P levels by a dual strategy. Mycobacterium encodes different phosphatase effectors to arrest phagosomal maturation and avoid its elimination by reducing the amount of PI(3)P on the Mycobacterium inclusion surface (Castandet et al., 2005; Singh et al., 2003). Mycobacterium also expresses a glycosylated PI analogue, the ManLAM, that inhibits the activation of PI3KIII, the kinase involved in PI(3)P production.
Chlamydia activates phosphoinositide‐dependent signalling pathways such as PDPK1‐MYC (Al‐Zeer et al., 2017) and PI3K‐AKT (Zou et al., 2018) to favour infection and survival. PI4KII has been shown to be important for Chlamydia and Listeria infection. In addition to PI3K, the activation of PI4KII is required for Listeria invasion, and its blockade impairs entry without perturbing the plasma membrane content in PI (4,5)P2 and PI(3,4,5)P3 (Pizarro‐Cerda et al., 2007). Although the participation of PI4K in Listeria infection is not well understood, these results suggest a postentry signalling role. Legionella modifies PI flows with a number of effectors that interfere with endocytic and secretory pathways and with communication with the ER. Legionella pneumophila hijacks PC and diacylglycerol on the Legionella‐containing vacuole membrane (Viner, Chetrit, Ehrlich, & Segal, 2012). Furthermore, it is known that L. pneumophila enriches its inclusion membrane with PI(4)P in a PI4KIII‐dependent manner (Brombacher et al., 2009; Weber, Ragaz, Reus, Nyfeler, & Hilbi, 2006). Additionally, treatment with the PI4KIII inhibitor PIK93 reduces L. pneumophila replication (Hubber et al., 2014), confirming the importance of the PI4Ks in Legionella infection. However, the requirement of PI3K in L. pneumophila entry and replication has been a matter of debate (Charpentier et al., 2009; Khelef, Shuman, & Maxfield, 2001; Tachado, Samrakandi, & Cirillo, 2008; Weber et al., 2006). Chlamydia recruits PI4KIIα, has high levels of PI4P in the membrane inclusion, and recruits the lipid transfer protein OSBP in a PI4P‐dependent way. Although its role is still unclear, OSBP probably participates in inclusion membrane remodelling by modifying its lipid composition (Moorhead, Jung, Smirnov, Kaufer, & Scidmore, 2010), something that could make Chlamydia sensitive to inhibitors of OSBP.
3.3. Interfering with lipid trafficking
Although little is known about how bacteria interfere with lipid trafficking, it has been reported that some bacteria hijack LTPs. The best documented cases are described below.
3.3.1. Niemann–Pick type C1 protein
Pharmacological inhibition and genetic deficiency studies showed that NPC1 is used by A. phagocytophilum and Brucella. Recruitment of NPC1 produces an accumulation of cholesterol in the inclusions of A. phagocytophilum. Moreover, treatment with the NPC1 inhibitor U18666A blocks the transport of cholesterol to the bacterial inclusion and stops bacterial growth, evidencing the essential role of this LTP in infection (Xiong et al., 2009; Xiong & Rikihisa, 2012). In the case of Brucella, lipid rafts are important for the entry of bacteria in the host cell (Watarai, Makino, Michikawa, et al., 2002). Because NPC1 is important for the maintenance of cholesterol‐rich lipid rafts, targeting NPC1 blocks Brucella infection. Therefore, NPC1‐deficient mice are resistant to Brucella infection (Watarai, Makino, Fujii, et al., 2002). U18666A is efficient also against C. burnetii and Chlamydia trachomatis (Beatty, 2006; Howe & Heinzen, 2006), but the role of NPC1 in infection is still unknown.
3.3.2. Ceramide transfer protein
As mentioned before, CERT transports Cer from the ER membrane to the Golgi apparatus where the SM synthase 1 transforms it into SM. Chlamydia cannot synthesize SLs that are essential for the biogenesis of bacterial inclusions and replication of bacteria (van Ooij et al., 2000). Cryo‐electron microscopy and immunogold labelling showed that CERT is recruited to bacterial inclusions. Chlamydia uses CERT to transport Cer from the ER to the bacterial inclusion for SM synthesis (Derre, Swiss, & Agaisse, 2011; Elwell et al., 2011). SM then incorporates into the inclusion membrane and the bacterial cell wall. The compound HPA‐12 has been reported as a specific CERT inhibitor. However, in CERT‐knockout cells, SL transport still works, and HPA‐12 strongly impairs this transport (Carabeo, Mead, & Hackstadt, 2003; Elwell et al., 2011; Koch‐Edelmann et al., 2017). These results suggest that HPA‐12 is an inhibitor of Cer transporters other than CERT.
3.3.3. Oxysterol‐binding protein‐related protein 1
Oxysterol‐binding protein‐related protein 1 (ORP1L) belongs to the OSBP‐related protein family. ORP1L acts as a cholesterol sensor that modulates the formation of MCSs between the late endosomes and the ER. It has been recently reported that Coxiella recruits ORP1L to MCSs formed between the bacterial inclusion and the ER, which is followed by changes in the membrane properties of the inclusion vacuole (Justis et al., 2017; Wallqvist et al., 2017). ORP1L attaches to the Coxiella‐containing vacuole membrane through its N‐terminal ankyrin repeats that are involved in protein–protein interactions, but the specific function of ORP1L in the infection is still unclear.
4. CONCLUSIONS
Therapeutic approaches based on specific factors of host–pathogen interactions are a promising avenue to overcome treatment failure in infectious diseases. Lipids are gaining importance as targets for antiviral and antibiotic development. Because many viruses and bacteria use lipids to build neo‐organelles for replication and persistence, inhibitors of the synthesis of cholesterol, Cer, SLs, FAs, and phospholipids interfere with growth of many pathogens. Besides, drugs against protein kinases target signalling pathways needed for infection. Figures 1 and 2 summarise current knowledge on host lipid synthesis, transport, and signalling pathways used by viruses and bacteria. Compounds that interfere with these pathways may become efficient antivirals or antibiotics. Moreover, drugs tested against viruses might be efficient also against bacteria and vice versa. Notably, targeting lipid transport may produce less toxic antivirals and antibiotics as intracellular pathogens depend on specific LTPs for making their neo‐organelles, although cells may have more than one LTP for the same function. Within this context, high‐throughput screening of clinically tested compounds will provide a rapid means to identify undiscovered, antiviral, and antibiotic functions for well‐characterized molecules that target lipid metabolism and flows.
CONFLICTS OF INTEREST
The authors declare to have no conflicts of interest.
ACKNOWLEDGEMENTS
This work has been funded by Contract BES‐2016‐078317 for predoctoral fellows (to A. F.‐ O.) and Research Grant BIO2015‐68758‐R (to C. R.) from the Ministry of Science, Innovation and Universities of Spain. The authors thank Dr R. Tenorio for critically revising the manuscript.
Fernández‐Oliva A, Ortega‐González P, Risco C. Targeting host lipid flows: Exploring new antiviral and antibiotic strategies. Cellular Microbiology. 2019;21:e12996 10.1111/cmi.12996
REFERENCES
- Aizaki, H. , Morikawa, K. , Fukasawa, M. , Hara, H. , Inoue, Y. , Tani, H. , … Suzuki, T. (2008). Critical role of virion‐associated cholesterol and sphingolipid in hepatitis C virus infection. Journal of Virology, 82, 5715–5724. 10.1128/JVI.02530-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aktepe, T. E. , Pham, H. , & Mackenzie, J. M. (2015). Differential utilisation of ceramide during replication of the flaviviruses West Nile and dengue virus. Virology, 484, 241–250. 10.1016/j.virol.2015.06.015 [DOI] [PubMed] [Google Scholar]
- Albulescu, L. , Bigay, J. , Biswas, B. , Weber‐Boyvat, M. , Dorobantu, C. M. , Delang, L. , … Strating, J. R. P. M. (2017). Uncovering oxysterol‐binding protein (OSBP) as a target of the anti‐enteroviral compound TTP‐8307. Antiviral Research, 140, 37–44. 10.1016/j.antiviral.2017.01.008 [DOI] [PubMed] [Google Scholar]
- Albulescu, L. , Strating, J. R. , Thibaut, H. J. , van der Linden, L. , Shair, M. D. , Neyts, J. , & van Kuppeveld, F. J. (2015). Broad‐range inhibition of enterovirus replication by OSW‐1, a natural compound targeting OSBP. Antiviral Research, 117, 110–114. 10.1016/j.antiviral.2015.02.013 [DOI] [PubMed] [Google Scholar]
- Al‐Kuraishy, H. M. , Al‐Gareeb, A. I. , & Al‐Buhadily, A. K. (2018). Rosuvastatin as forthcoming antibiotic or as adjuvant additive agent: In vitro novel antibacterial study. Journal of Laboratory Physicians, 10, 271–275. 10.4103/JLP.JLP_170_17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alpuche‐Aranda, C. M. , Berthiaume, E. P. , Mock, B. , Swanson, J. A. , & Miller, S. I. (1995). Spacious phagosome formation within mouse macrophages correlates with Salmonella serotype pathogenicity and host susceptibility. Infection and Immunity, 63, 4456–4462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altan‐Bonnet, N. (2017). Lipid tales of viral replication and transmission. Trends in Cell Biology, 27, 201–213. 10.1016/j.tcb.2016.09.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al‐Zeer, M. A. , Xavier, A. , Abu Lubad, M. , Sigulla, J. , Kessler, M. , Hurwitz, R. , & Meyer, T. F. (2017). Chlamydia trachomatis prevents apoptosis via activation of PDPK1‐MYC and enhanced mitochondrial binding of hexokinase II. eBioMedicine, 23, 100–110. 10.1016/j.ebiom.2017.08.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amako, Y. , Syed, G. H. , & Siddiqui, A. (2011). Protein kinase D negatively regulates hepatitis C virus secretion through phosphorylation of oxysterol‐binding protein and ceramide transfer protein. The Journal of Biological Chemistry, 286, 11265–11274. 10.1074/jbc.M110.182097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amara, A. , & Mercer, J. (2015). Viral apoptotic mimicry. Nature Reviews. Microbiology, 13, 461–469. 10.1038/nrmicro3469 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amemiya, F. , Maekawa, S. , Itakura, Y. , Kanayama, A. , Matsui, A. , Takano, S. , … Enomoto, N. (2008). Targeting lipid metabolism in the treatment of hepatitis C virus infection. The Journal of Infectious Diseases, 197(3), 361–370. 10.1086/525287 [DOI] [PubMed] [Google Scholar]
- Arita, M. (2014). Phosphatidylinositol‐4 kinase III beta and oxysterol‐binding protein accumulate unesterified cholesterol on poliovirus‐induced membrane structure. Microbiology and Immunology, 58, 239–256. 10.1111/1348-0421.12144 [DOI] [PubMed] [Google Scholar]
- Arita, M. , Kojima, H. , Nagano, T. , Okabe, T. , Wakita, T. , & Shimizu, H. (2011). Phosphatidylinositol 4‐kinase III beta is a target of enviroxime‐like compounds for antipoliovirus activity. Journal of Virology, 85, 2364–2372. 10.1128/JVI.02249-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Auweter, S. D. , Bhavsar, A. P. , de Hoog, C. L. , Li, Y. , Chan, Y. A. , van der Heijden, J. , … Finlay, B. B. (2011). Quantitative mass spectrometry catalogues Salmonella pathogenicity island‐2 effectors and identifies their cognate host binding partners. The Journal of Biological Chemistry, 286, 24023–24035. 10.1074/jbc.M111.224600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Auweter, S. D. , Yu, H. B. , Arena, E. T. , Guttman, J. A. , & Finlay, B. B. (2012). Oxysterol‐binding protein (OSBP) enhances replication of intracellular Salmonella and binds the Salmonella SPI‐2 effector SseL via its N‐terminus. Microbes and Infection, 14, 148–154. 10.1016/j.micinf.2011.09.003 [DOI] [PubMed] [Google Scholar]
- Bajimaya, S. , Frankl, T. , Hayashi, T. , & Takimoto, T. (2017). Cholesterol is required for stability and infectivity of influenza A and respiratory syncytial viruses. Virology, 510, 234–241. 10.1016/j.virol.2017.07.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bakowski, M. A. , Braun, V. , Lam, G. Y. , Yeung, T. , Heo, W. D. , Meyer, T. , et al. (2010). The phosphoinositide phosphatase SopB manipulates membrane surface charge and trafficking of the Salmonella‐containing vacuole. Cell Host & Microbe, 7, 453–462. 10.1016/j.chom.2010.05.011 [DOI] [PubMed] [Google Scholar]
- Bashmakov, Y. K. , Zigangirova, N. A. , Pashko, Y. P. , Kapotina, L. N. , & Petyaev, I. M. (2010). Chlamydia trachomatis growth inhibition and restoration of LDL‐receptor level in HepG2 cells treated with mevastatin. Comparative Hepatology, 9, 3 10.1186/1476-5926-9-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beatty, W. L. (2006). Trafficking from CD63‐positive late endocytic multivesicular bodies is essential for intracellular development of Chlamydia trachomatis . Journal of Cell Science, 119, 350–359. 10.1242/jcs.02733 [DOI] [PubMed] [Google Scholar]
- Beatty, W. L. , Rhoades, E. R. , Ullrich, H. J. , Chatterjee, D. , Heuser, J. E. , & Russell, D. G. (2000). Trafficking and release of mycobacterial lipids from infected macrophages. Traffic, 1, 235–247. 10.1034/j.1600-0854.2000.010306.x [DOI] [PubMed] [Google Scholar]
- Becker, K. A. , Riethmuller, J. , Luth, A. , Doring, G. , Kleuser, B. , & Gulbins, E. (2010). Acid sphingomyelinase inhibitors normalize pulmonary ceramide and inflammation in cystic fibrosis. American Journal of Respiratory Cell and Molecular Biology, 42, 716–724. 10.1165/rcmb.2009-0174OC [DOI] [PubMed] [Google Scholar]
- Bergman, P. , Linde, C. , Putsep, K. , Pohanka, A. , Normark, S. , Henriques‐Normark, B. , et al. (2011). Studies on the antibacterial effects of statins—In vitro and in vivo. PLoS One, 6, e24394 10.1371/journal.pone.0024394 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birge, R. B. , Boeltz, S. , Kumar, S. , Carlson, J. , Wanderley, J. , Calianese, D. , … Herrmann, M. (2016). Phosphatidylserine is a global immunosuppressive signal in efferocytosis, infectious disease, and cancer. Cell Death and Differentiation, 23, 962–978. 10.1038/cdd.2016.11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bishe, B. , Syed, G. , & Siddiqui, A. (2012). Phosphoinositides in the hepatitis C virus life cycle. Viruses, 4, 2340–2358. 10.3390/v4102340 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brombacher, E. , Urwyler, S. , Ragaz, C. , Weber, S. S. , Kami, K. , Overduin, M. , & Hilbi, H. (2009). Rab1 guanine nucleotide exchange factor SidM is a major phosphatidylinositol 4‐phosphate‐binding effector protein of Legionella pneumophila . The Journal of Biological Chemistry, 284, 4846–4856. 10.1074/jbc.M807505200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bryan‐Marrugo, O. L. , Arellanos‐Soto, D. , Rojas‐Martinez, A. , Barrera‐Saldana, H. , Ramos‐Jimenez, J. , Vidaltamayo, R. , & Rivas‐Estilla, A. M. (2016). The antidengue virus properties of statins may be associated with alterations in the cellular antiviral profile expression. Molecular Medicine Reports, 14, 2155–2163. 10.3892/mmr.2016.5519 [DOI] [PubMed] [Google Scholar]
- Bujny, M. V. , Ewels, P. A. , Humphrey, S. , Attar, N. , Jepson, M. A. , & Cullen, P. J. (2008). Sorting nexin‐1 defines an early phase of Salmonella‐containing vacuole‐remodeling during Salmonella infection. Journal of Cell Science, 121, 2027–2036. 10.1242/jcs.018432 [DOI] [PubMed] [Google Scholar]
- Carabeo, R. A. , Mead, D. J. , & Hackstadt, T. (2003). Golgi‐dependent transport of cholesterol to the Chlamydia trachomatis inclusion. Proceedings of the National Academy of Sciences of the United States of America, 100, 6771–6776. 10.1073/pnas.1131289100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carette, J. E. , Raaben, M. , Wong, A. C. , Herbert, A. S. , Obernosterer, G. , Mulherkar, N. , … Brummelkamp, T. R. (2011). Ebola virus entry requires the cholesterol transporter Niemann–Pick C1. Nature, 477, 340–343. 10.1038/nature10348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castandet, J. , Prost, J. F. , Peyron, P. , Astarie‐Dequeker, C. , Anes, E. , Cozzone, A. J. , … Maridonneau‐Parini, I. (2005). Tyrosine phosphatase MptpA of Mycobacterium tuberculosis inhibits phagocytosis and increases actin polymerization in macrophages. Research in Microbiology, 156, 1005–1013. 10.1016/j.resmic.2005.05.013 [DOI] [PubMed] [Google Scholar]
- Castorena, K. M. , Stapleford, K. A. , & Miller, D. J. (2010). Complementary transcriptomic, lipidomic, and targeted functional genetic analyses in cultured Drosophila cells highlight the role of glycerophospholipid metabolism in Flock House virus RNA replication. BMC Genomics, 11, 183 10.1186/1471-2164-11-183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Catron, D. M. , Lange, Y. , Borensztajn, J. , Sylvester, M. D. , Jones, B. D. , & Haldar, K. (2004). Salmonella enterica serovar Typhimurium requires nonsterol precursors of the cholesterol biosynthetic pathway for intracellular proliferation. Infection and Immunity, 72, 1036–1042. 10.1128/IAI.72.2.1036-1042.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charpentier, X. , Gabay, J. E. , Reyes, M. , Zhu, J. W. , Weiss, A. , & Shuman, H. A. (2009). Chemical genetics reveals bacterial and host cell functions critical for type IV effector translocation by Legionella pneumophila . PLoS Pathogens, 5, e1000501 10.1371/journal.ppat.1000501 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheshenko, N. , Pierce, C. , & Herold, B. C. (2018). Herpes simplex viruses activate phospholipid scramblase to redistribute phosphatidylserines and Akt to the outer leaflet of the plasma membrane and promote viral entry. PLoS Pathogens, 14, e1006766 10.1371/journal.ppat.1006766 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chotiwan, N. , Andre, B. G. , Sanchez‐Vargas, I. , Islam, M. N. , Grabowski, J. M. , Hopf‐Jannasch, A. , … Perera, R. (2018). Dynamic remodeling of lipids coincides with dengue virus replication in the midgut of Aedes aegypti mosquitoes. PLoS Pathogens, 14(2), e1006853 10.1371/journal.ppat.1006853 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chua, J. , & Deretic, V. (2004). Mycobacterium tuberculosis reprograms waves of phosphatidylinositol 3‐phosphate on phagosomal organelles. The Journal of Biological Chemistry, 279, 36982–36992. 10.1074/jbc.M405082200 [DOI] [PubMed] [Google Scholar]
- Chung, J. , Torta, F. , Masai, K. , Lucast, L. , Czapla, H. , Tanner, L. B. , … de Camilli, P. (2015). INTRACELLULAR TRANSPORT. PI4P/phosphatidylserine countertransport at ORP5‐ and ORP8‐mediated ER–plasma membrane contacts. Science, 349, 428–432. 10.1126/science.aab1370 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cocchiaro, J. L. , Kumar, Y. , Fischer, E. R. , Hackstadt, T. , & Valdivia, R. H. (2008). Cytoplasmic lipid droplets are translocated into the lumen of the Chlamydia trachomatis parasitophorous vacuole. Proceedings of the National Academy of Sciences of the United States of America, 105, 9379–9384. 10.1073/pnas.0712241105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cremesti, A. , Paris, F. , Grassme, H. , Holler, N. , Tschopp, J. , Fuks, Z. , et al. (2001). Ceramide enables fas to cap and kill. The Journal of Biological Chemistry, 276, 23954–23961. 10.1074/jbc.M101866200 [DOI] [PubMed] [Google Scholar]
- Czyz, D. M. , Potluri, L. P. , Jain‐Gupta, N. , Riley, S. P. , Martinez, J. J. , Steck, T. L. , et al. (2014). Host‐directed antimicrobial drugs with broad‐spectrum efficacy against intracellular bacterial pathogens. MBio, 5, e01534–e01514. 10.1128/mBio.01534-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Chastellier, C. , & Thilo, L. (2006). Cholesterol depletion in Mycobacterium avium‐infected macrophages overcomes the block in phagosome maturation and leads to the reversible sequestration of viable mycobacteria in phagolysosome‐derived autophagic vacuoles. Cellular Microbiology, 8, 242–256. 10.1111/j.1462-5822.2005.00617.x [DOI] [PubMed] [Google Scholar]
- DeLucia, D. C. , Rinaldo, C. R. , & Rappocciolo, G. (2018). Inefficient HIV‐1 trans infection of CD4+ T cells by macrophages from HIV‐1 nonprogressors is associated with altered membrane cholesterol and DC‐SIGN. Journal of Virology, 92 10.1128/JVI.00092-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng, Y. , Almsherqi, Z. A. , Ng, M. M. , & Kohlwein, S. D. (2010). Do viruses subvert cholesterol homeostasis to induce host cubic membranes? Trends in Cell Biology, 20, 371–379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derre, I. , Swiss, R. , & Agaisse, H. (2011). The lipid transfer protein CERT interacts with the Chlamydia inclusion protein IncD and participates to ER–Chlamydia inclusion membrane contact sites. PLoS Pathogens, 7, e1002092 10.1371/journal.ppat.1002092 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diamond, D. L. , Syder, A. J. , Jacobs, J. M. , Sorensen, C. M. , Walters, K. A. , Proll, S. C. , … Katze, M. G. (2010). Temporal proteome and lipidome profiles reveal hepatitis C virus‐associated reprogramming of hepatocellular metabolism and bioenergetics. PLoS Pathogens, 6(1), e1000719 10.1371/journal.ppat.1000719 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dorobantu, C. M. , Albulescu, L. , Harak, C. , Feng, Q. , van Kampen, M. , Strating, J. R. , et al. (2015). Modulation of the host lipid landscape to promote RNA virus replication: The picornavirus encephalomyocarditis virus converges on the pathway used by hepatitis C virus. PLoS Pathogens, 11, e1005185 10.1371/journal.ppat.1005185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dorobantu, C. M. , Harak, C. , Klein, R. , van der Linden, L. , Strating, J. R. , van der Schaar, H. M. , et al. (2016). Tyrphostin AG1478 inhibits encephalomyocarditis virus and hepatitis C virus by targeting phosphatidylinositol 4‐kinase IIIα. Antimicrobial Agents and Chemotherapy, 60, 6402–6406. 10.1128/AAC.01331-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dowall, S. D. , Graham, V. A. , Corbin‐Lickfett, K. , Empig, C. , Schlunegger, K. , Bruce, C. B. , … Hewson, R. (2015). Effective binding of a phosphatidylserine‐targeting antibody to Ebola virus infected cells and purified virions. Journal of Immunology Research, 2015(347903), 1–9. 10.1155/2015/347903 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elgner, F. , Ren, H. , Medvedev, R. , Ploen, D. , Himmelsbach, K. , Boller, K. , & Hildt, E. (2016). The intracellular cholesterol transport inhibitor U18666A inhibits the exosome‐dependent release of mature hepatitis C virus. Journal of Virology, 90(24), 11181–11196. 10.1128/JVI.01053-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elwell, C. A. , Jiang, S. , Kim, J. H. , Lee, A. , Wittmann, T. , Hanada, K. , … Engel, J. N. (2011). Chlamydia trachomatis co‐opts GBF1 and CERT to acquire host sphingomyelin for distinct roles during intracellular development. PLoS Pathogens, 7, e1002198 10.1371/journal.ppat.1002198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erkkila, L. , Jauhiainen, M. , Laitinen, K. , Haasio, K. , Tiirola, T. , Saikku, P. , & Leinonen, M. (2005). Effect of simvastatin, an established lipid‐lowering drug, on pulmonary Chlamydia pneumoniae infection in mice. Antimicrobial Agents and Chemotherapy, 49, 3959–3962. 10.1128/AAC.49.9.3959-3962.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falcone, S. , Perrotta, C. , De Palma, C. , Pisconti, A. , Sciorati, C. , Capobianco, A. , et al. (2004). Activation of acid sphingomyelinase and its inhibition by the nitric oxide/cyclic guanosine 3′,5′‐monophosphate pathway: Key events in Escherichia coli‐elicited apoptosis of dendritic cells. Journal of Immunology, 173, 4452–4463. 10.4049/jimmunol.173.7.4452 [DOI] [PubMed] [Google Scholar]
- Fernandes, P. , & Martens, E. (2017). Antibiotics in late clinical development. Biochemical Pharmacology, 133, 152–163. 10.1016/j.bcp.2016.09.025 [DOI] [PubMed] [Google Scholar]
- Fernandez de Castro, I. , Fernandez, J. J. , Barajas, D. , Nagy, P. D. , & Risco, C. (2017). Three‐dimensional imaging of the intracellular assembly of a functional viral RNA replicase complex. Journal of Cell Science, 130, 260–268. 10.1242/jcs.181586 [DOI] [PubMed] [Google Scholar]
- Fontana, J. , Lopez‐Iglesias, C. , Tzeng, W. P. , Frey, T. K. , Fernandez, J. J. , & Risco, C. (2010). Three‐dimensional structure of rubella virus factories. Virology, 405(2), 579–591. 10.1016/j.virol.2010.06.043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fontana, J. , Lopez‐Montero, N. , Elliott, R. M. , Fernandez, J. J. , & Risco, C. (2008). The unique architecture of Bunyamwera virus factories around the Golgi complex. Cellular Microbiology, 10, 2012–2028. 10.1111/j.1462-5822.2008.01184.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fratti, R. A. , Chua, J. , Vergne, I. , & Deretic, V. (2003). Mycobacterium tuberculosis glycosylated phosphatidylinositol causes phagosome maturation arrest. Proceedings of the National Academy of Sciences of the United States of America, 100, 5437–5442. 10.1073/pnas.0737613100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frolova, E. I. , Gorchakov, R. , Pereboeva, L. , Atasheva, S. , & Frolov, I. (2010). Functional Sindbis virus replicative complexes are formed at the plasma membrane. Journal of Virology, 84, 11679–11695. 10.1128/JVI.01441-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gabel, B. R. , Elwell, C. , van Ijzendoorn, S. C. , & Engel, J. N. (2004). Lipid raft‐mediated entry is not required for Chlamydia trachomatis infection of cultured epithelial cells. Infection and Immunity, 72, 7367–7373. 10.1128/IAI.72.12.7367-7373.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilk, S. D. (2012). Role of lipids in Coxiella burnetii infection. Advances in Experimental Medicine and Biology, 984, 199–213. 10.1007/978-94-007-4315-1_10 [DOI] [PubMed] [Google Scholar]
- Gillespie, L. K. , Hoenen, A. , Morgan, G. , & Mackenzie, J. M. (2010). The endoplasmic reticulum provides the membrane platform for biogenesis of the flavivirus replication complex. Journal of Virology, 84, 10438–10447. 10.1128/JVI.00986-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Godbole, A. A. , Ahmed, W. , Bhat, R. S. , Bradley, E. K. , Ekins, S. , & Nagaraja, V. (2015). Targeting Mycobacterium tuberculosis topoisomerase I by small‐molecule inhibitors. Antimicrobial Agents and Chemotherapy, 59, 1549–1557. 10.1128/AAC.04516-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Godi, A. , Di Campli, A. , Konstantakopoulos, A. , Di Tullio, G. , Alessi, D. R. , Kular, G. S. , et al. (2004). FAPPs control Golgi‐to‐cell‐surface membrane traffic by binding to ARF and PtdIns(4)P. Nature Cell Biology, 6, 393–404. 10.1038/ncb1119 [DOI] [PubMed] [Google Scholar]
- Grassme, H. , Gulbins, E. , Brenner, B. , Ferlinz, K. , Sandhoff, K. , Harzer, K. , et al. (1997). Acidic sphingomyelinase mediates entry of N. gonorrhoeae into nonphagocytic cells. Cell, 91, 605–615. [DOI] [PubMed] [Google Scholar]
- Grassme, H. , Jendrossek, V. , Riehle, A. , von Kurthy, G. , Berger, J. , Schwarz, H. , et al. (2003). Host defense against Pseudomonas aeruginosa requires ceramide‐rich membrane rafts. Nature Medicine, 9, 322–330. 10.1038/nm823 [DOI] [PubMed] [Google Scholar]
- Guardado‐Calvo, P. , Atkovska, K. , Jeffers, S. A. , Grau, N. , Backovic, M. , Perez‐Vargas, J. , et al. (2017). A glycerophospholipid‐specific pocket in the RVFV class II fusion protein drives target membrane insertion. Science, 358, 663–667. 10.1126/science.aal2712 [DOI] [PubMed] [Google Scholar]
- Gullberg, R. C. , Steel, J. J. , Pujari, V. , Rovnak, J. , Crick, D. C. , & Perera, R. (2018). Stearoly‐CoA desaturase 1 differentiates early and advanced dengue virus infections and determines virus particle infectivity. PLoS Pathogens, 14(8), e1007261 10.1371/journal.ppat.1007261 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guzman‐Verri, C. , Chaves‐Olarte, E. , von Eichel‐Streiber, C. , Lopez‐Goni, I. , Thelestam, M. , Arvidson, S. , et al. (2001). GTPases of the Rho subfamily are required for Brucella abortus internalization in nonprofessional phagocytes: Direct activation of Cdc42. The Journal of Biological Chemistry, 276, 44435–44443. 10.1074/jbc.M105606200 [DOI] [PubMed] [Google Scholar]
- Hackstadt, T. , Scidmore, M. A. , & Rockey, D. D. (1995). Lipid metabolism in Chlamydia trachomatis‐infected cells: Directed trafficking of Golgi‐derived sphingolipids to the chlamydial inclusion. Proceedings of the National Academy of Sciences of the United States of America, 92, 4877–4881. 10.1073/pnas.92.11.4877 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanada, K. , Kumagai, K. , Yasuda, S. , Miura, Y. , Kawano, M. , Fukasawa, M. , & Nishijima, M. (2003). Molecular machinery for non‐vesicular trafficking of ceramide. Nature, 426, 803–809. 10.1038/nature02188 [DOI] [PubMed] [Google Scholar]
- Haraga, A. , Ohlson, M. B. , & Miller, S. I. (2008). Salmonellae interplay with host cells. Nature Reviews. Microbiology, 6, 53–66. 10.1038/nrmicro1788 [DOI] [PubMed] [Google Scholar]
- Harikumar, K. B. , Kunnumakkara, A. B. , Ochi, N. , Tong, Z. , Deorukhkar, A. , Sung, B. , … Guha, S. (2010). A novel small‐molecule inhibitor of protein kinase D blocks pancreatic cancer growth in vitro and in vivo. Molecular Cancer Therapeutics, 9, 1136–1146. 10.1158/1535-7163.MCT-09-1145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hatch, G. M. , & McClarty, G. (1998). Phospholipid composition of purified Chlamydia trachomatis mimics that of the eucaryotic host cell. Infection and Immunity, 66, 3727–3735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauck, C. R. , Grassme, H. , Bock, J. , Jendrossek, V. , Ferlinz, K. , Meyer, T. F. , & Gulbins, E. (2000). Acid sphingomyelinase is involved in CEACAM receptor‐mediated phagocytosis of Neisseria gonorrhoeae . FEBS Letters, 478, 260–266. 10.1016/S0014-5793(00)01851-2 [DOI] [PubMed] [Google Scholar]
- Heaton, N. S. , Perera, R. , Berger, K. L. , Khadka, S. , Lacount, D. J. , Kuhn, R. J. , & Randall, G. (2010). Dengue virus nonstructural protein 3 redistributes fatty acid synthase to sites of viral replication and increases cellular fatty acid synthesis. Proceedings of the National Academy of Sciences of the United States of America, 107, 17345–17350. 10.1073/pnas.1010811107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heinzen, R. A. , Scidmore, M. A. , Rockey, D. D. , & Hackstadt, T. (1996). Differential interaction with endocytic and exocytic pathways distinguish parasitophorous vacuoles of Coxiella burnetii and Chlamydia trachomatis . Infection and Immunity, 64, 796–809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henry, S. A. , Kohlwein, S. D. , & Carman, G. M. (2012). Metabolism and regulation of glycerolipids in the yeast Saccharomyces cerevisiae . Genetics, 190, 317–349. 10.1534/genetics.111.130286 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herker, E. , Harris, C. , Hernandez, C. , Carpentier, A. , Kaehlcke, K. , Rosenberg, A. R. , … Ott, M. (2010). Efficient hepatitis C virus particle formation requieres diacylglycerol acyltransferase‐1. Nature Medicine, 16(11), 1295–1298. 10.1038/nm.2238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernandez, L. D. , Hueffer, K. , Wenk, M. R. , & Galan, J. E. (2004). Salmonella modulates vesicular traffic by altering phosphoinositide metabolism. Science, 304, 1805–1807. 10.1126/science.1098188 [DOI] [PubMed] [Google Scholar]
- Hofmann, S. , Krajewski, M. , Scherer, C. , Scholz, V. , Mordhorst, V. , Truschow, P. , … Herker, E. (2018). Complex lipid metabolic remodeling is required for efficient hepatitis C virus replication. Molecular and Cell Biology of Lipids, 1863, 1041–1056. 10.1016/j.bbalip.2018.06.002 [DOI] [PubMed] [Google Scholar]
- van der Schaar, H. M. , Dorobantu, C. M. , Albulescu, L. , Strating, J. , & van Kuppeveld, F. J. M. (2016). Fat(al) attraction: Picornaviruses usurp lipid transfer at membrane contact sites to create replication organelles. Trends in Microbiology, 24, 535–546. 10.1016/j.tim.2016.02.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Schaar, H. M. , Leyssen, P. , Thibaut, H. J. , de Palma, A. , van der Linden, L. , Lanke, K. H. , et al. (2013). A novel, broad‐spectrum inhibitor of enterovirus replication that targets host cell factor phosphatidylinositol 4‐kinase IIIβ. Antimicrobial Agents and Chemotherapy, 57, 4971–4981. 10.1128/AAC.01175-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howe, D. , & Heinzen, R. A. (2006). Coxiella burnetii inhabits a cholesterol‐rich vacuole and influences cellular cholesterol metabolism. Cellular Microbiology, 8, 496–507. 10.1111/j.1462-5822.2005.00641.x [DOI] [PubMed] [Google Scholar]
- Hsu, N. Y. , Ilnytska, O. , Belov, G. , Santiana, M. , Chen, Y. H. , Takvorian, P. M. , … Altan‐Bonnet, N. (2010). Viral reorganization of the secretory pathway generates distinct organelles for RNA replication. Cell, 141, 799–811. 10.1016/j.cell.2010.03.050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang, F. C. (2011). Plasma membrane cholesterol plays a critical role in the Salmonella‐induced anti‐inflammatory response in intestinal epithelial cells. Cellular Immunology, 271, 480–487. 10.1016/j.cellimm.2011.08.018 [DOI] [PubMed] [Google Scholar]
- Huang, F. C. (2016). De novo sphingolipid synthesis is essential for Salmonella‐induced autophagy and human beta‐defensin 2 expression in intestinal epithelial cells. Gut Pathogens, 8, 5 10.1186/s13099-016-0088-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hubber, A. , Arasaki, K. , Nakatsu, F. , Hardiman, C. , Lambright, D. , De Camilli, P. , et al. (2014). The machinery at endoplasmic reticulum–plasma membrane contact sites contributes to spatial regulation of multiple Legionella effector proteins. PLoS Pathogens, 10, e1004222 10.1371/journal.ppat.1004222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ilnytska, O. , Santiana, M. , Hsu, N. Y. , Du, W. L. , Chen, Y. H. , Viktorova, E. G. , et al. (2013). Enteroviruses harness the cellular endocytic machinery to remodel the host cell cholesterol landscape for effective viral replication. Cell Host & Microbe, 14, 281–293. 10.1016/j.chom.2013.08.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ioannou, Y. A. (2000). The structure and function of the Niemann–Pick C1 protein. Molecular Genetics and Metabolism, 71, 175–181. 10.1006/mgme.2000.3061 [DOI] [PubMed] [Google Scholar]
- Ireton, K. , Payrastre, B. , Chap, H. , Ogawa, W. , Sakaue, H. , Kasuga, M. , & Cossart, P. (1996). A role for phosphoinositide 3‐kinase in bacterial invasion. Science, 274, 780–782. 10.1126/science.274.5288.780 [DOI] [PubMed] [Google Scholar]
- Ireton, K. , Payrastre, B. , & Cossart, P. (1999). The Listeria monocytogenes protein InlB is an agonist of mammalian phosphoinositide 3‐kinase. The Journal of Biological Chemistry, 274, 17025–17032. 10.1074/jbc.274.24.17025 [DOI] [PubMed] [Google Scholar]
- Jerwood, S. , & Cohen, J. (2008). Unexpected antimicrobial effect of statins. The Journal of Antimicrobial Chemotherapy, 61, 362–364. 10.1093/jac/dkm496 [DOI] [PubMed] [Google Scholar]
- Justis, A. V. , Hansen, B. , Beare, P. A. , King, K. B. , Heinzen, R. A. , & Gilk, S. D. (2017). Interactions between the Coxiella burnetii parasitophorous vacuole and the endoplasmic reticulum involve the host protein ORP1L. Cellular Microbiology, 19, e12637 10.1111/cmi.12637 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jutras, I. , Abrami, L. , & Dautry‐Varsat, A. (2003). Entry of the lymphogranuloma venereum strain of Chlamydia trachomatis into host cells involves cholesterol‐rich membrane domains. Infection and Immunity, 71, 260–266. 10.1128/IAI.71.1.260-266.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kampmueller, K. M. , & Miller, D. J. (2005). The cellular chaperone heat shock protein 90 facilitates Flock House virus RNA replication in Drosophila cells. Journal of Virology, 79, 6827–6837. 10.1128/JVI.79.11.6827-6837.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kapadia, S. B. , Barth, H. , Baumert, T. , McKeating, J. A. , & Chisari, F. V. (2007). Initiation of hepatitis C virus infection is dependent on cholesterol and cooperativity between CD81 and scavenger receptor B type I. Journal of Virology, 81(1), 374–383. 10.1128/JVI.01134-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaul, D. , Anand, P. K. , & Verma, I. (2004). Cholesterol‐sensor initiates M. tuberculosis entry into human macrophages. Molecular and Cellular Biochemistry, 258, 219–222. 10.1023/B:MCBI.0000012851.42642.be [DOI] [PubMed] [Google Scholar]
- Khan, I. , Katikaneni, D. S. , Han, Q. , Sanchez‐Felipe, L. , Hanada, K. , Ambrose, R. L. , … Konan, K. V. (2014). Modulation of hepatitis C virus genome replication by glycosphingolipids and four‐phosphate adaptor protein 2. Journal of Virology, 88, 12276–12295. 10.1128/JVI.00970-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khanal, S. , Sultana, H. , Catravas, J. D. , Carlyon, J. A. , & Neelakanta, G. (2017). Anaplasma phagocytophilum infection modulates expression of megakaryocyte cell cycle genes through phosphatidylinositol‐3‐kinase signaling. PLoS One, 12, e0182898 10.1371/journal.pone.0182898 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khelef, N. , Shuman, H. A. , & Maxfield, F. R. (2001). Phagocytosis of wild‐type Legionella pneumophila occurs through a wortmannin‐insensitive pathway. Infection and Immunity, 69, 5157–5161. 10.1128/IAI.69.8.5157-5161.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, S. S. , Peng, L. F. , Lin, W. , Choe, W. H. , Sakamoto, N. , Schreiber, S. L. , & Chung, R. T. (2007). A cell‐based, high‐throughtput screen for small molecule regulators of hepatits C virus. Gastroenterology, 132(1), 311–320. 10.1053/j.gastro.2006.10.032 [DOI] [PubMed] [Google Scholar]
- Knoops, K. , Kikkert, M. , Worm, S. H. , Zevenhoven‐Dobbe, J. C. , van der Meer, Y. , Koster, A. J. , et al. (2008). SARS‐coronavirus replication is supported by a reticulovesicular network of modified endoplasmic reticulum. PLoS Biology, 6, e226 10.1371/journal.pbio.0060226 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch‐Edelmann, S. , Banhart, S. , Saied, E. M. , Rose, L. , Aeberhard, L. , Laue, M. , … Heuer, D. (2017). The cellular ceramide transport protein CERT promotes Chlamydia psittaci infection and controls bacterial sphingolipid uptake. Cellular Microbiology, 19 10.1111/cmi.12752 [DOI] [PubMed] [Google Scholar]
- Kopek, B. G. , Perkins, G. , Miller, D. J. , Ellisman, M. H. , & Ahlquist, P. (2007). Three‐dimensional analysis of a viral RNA replication complex reveals a virus‐induced mini‐organelle. PLoS Biology, 5, e220 10.1371/journal.pbio.0050220 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korhonen, J. T. , Puolakkainen, M. , Haveri, A. , Tammiruusu, A. , Sarvas, M. , & Lahesmaa, R. (2012). Chlamydia pneumoniae entry into epithelial cells by clathrin‐independent endocytosis. Microbial Pathogenesis, 52, 157–164. 10.1016/j.micpath.2011.12.002 [DOI] [PubMed] [Google Scholar]
- Kowalski, M. P. , & Pier, G. B. (2004). Localization of cystic fibrosis transmembrane conductance regulator to lipid rafts of epithelial cells is required for Pseudomonas aeruginosa‐induced cellular activation. Journal of Immunology, 172, 418–425. 10.4049/jimmunol.172.1.418 [DOI] [PubMed] [Google Scholar]
- Kushwaha, N. , Singh, A. K. , Basu, S. , & Chakraborty, S. (2015). Differential response of diverse solanaceous hosts to tomato leaf curl New Delhi virus infection indicates coordinated action of NBS‐LRR and RNAi‐mediated host defense. Archives of Virology, 160, 1499–1509. 10.1007/s00705-015-2399-x [DOI] [PubMed] [Google Scholar]
- Lafont, F. , Tran Van Nhieu, G. , Hanada, K. , Sansonetti, P. , & van der Goot, F. G. (2002). Initial steps of Shigella infection depend on the cholesterol/sphingolipid raft‐mediated CD44–IpaB interaction. The EMBO Journal, 21, 4449–4457. 10.1093/emboj/cdf457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, W. M. , Ishikawa, M. , & Ahlquist, P. (2001). Mutation of host Δ9 fatty acid desaturase inhibits brome mosaic virus RNA replication between template recognition and RNA synthesis. Journal of Virology, 75, 2097–2106. 10.1128/JVI.75.5.2097-2106.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, L. , Yu, L. , & Hou, X. (2017). Cholesterol‐rich lipid rafts play a critical role in bovine parainfluenza virus type 3 (BPIV3) infection. Research in Veterinary Science, 114, 341–347. 10.1016/j.rvsc.2017.04.009 [DOI] [PubMed] [Google Scholar]
- Liebscher, S. , Ambrose, R. L. , Aktepe, T. E. , Mikulasova, A. , Prier, J. E. , Gillespie, L. K. , … Mackenzie, J. M. (2018). Phospholipase A2 activity during the replication cycle of the flavivirus West Nile virus. PLoS Pathogens, 14(4), e1007029 10.1371/journal.ppat.1007029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liefhebber, J. M. P. , Hague, C. V. , Zhang, Q. , Wakelam, M. J. O. , & McLauchlan, J. (2014). Modulation of triglyceride and cholesterol ester synthesis impairs assembly of infectious hepatitis C virus. The Journal of Biological Chemistry, 289(31), 21276–21288. 10.1074/jbc.M114.582999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Limpens, R. W. , van der Schaar, H. M. , Kumar, D. , Koster, A. J. , Snijder, E. J. , van Kuppeveld, F. J. , & Barcena, M. (2011). The transformation of enterovirus replication structures: A three‐dimensional study of single‐ and double‐membrane compartments. MBio, 2 10.1128/mBio.00166-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin, M. , & Rikihisa, Y. (2003). Ehrlichia chaffeensis and Anaplasma phagocytophilum lack genes for lipid A biosynthesis and incorporate cholesterol for their survival. Infection and Immunity, 71, 5324–5331. 10.1128/IAI.71.9.5324-5331.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lorizate, M. , Sachsenheimer, T. , Glass, B. , Habermann, A. , Gerl, M. J. , Kräusslich, H. G. , & Brügger, B. (2012). Comparative lipidomics analysis of HIV‐1 particles and their producer cell membrane in different cell lines. Cellular Microbiology, 15(2), 292–304. [DOI] [PubMed] [Google Scholar]
- Lu, F. , Liang, Q. , Abi‐Mosleh, L. , Das, A. , De Brabander, J. K. , Goldstein, J. L. , & Brown, M. S. (2015). Identification of NPC1 as the target of U18666A, an inhibitor of lysosomal cholesterol export and Ebola infection. eLife, 4 10.7554/eLife.12177 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackenzie, J. M. , Khromykh, A. A. , & Parton, R. G. (2007). Cholesterol manipulation by West Nile virus perturbs the cellular immune response. Cell Host & Microbe, 2, 229–239. 10.1016/j.chom.2007.09.003 [DOI] [PubMed] [Google Scholar]
- Marino, M. , Braun, L. , Cossart, P. , & Ghosh, P. (1999). Structure of the lnlB leucine‐rich repeats, a domain that triggers host cell invasion by the bacterial pathogen L. monocytogenes . Molecular Cell, 4, 1063–1072. 10.1016/S1097-2765(00)80234-8 [DOI] [PubMed] [Google Scholar]
- Martin, J. J. , Holguera, J. , Sanchez‐Felipe, L. , Villar, E. , & Munoz‐Barroso, I. (2012). Cholesterol dependence of Newcastle disease virus entry. Biochimica et Biophysica Acta, 1818, 753–761. 10.1016/j.bbamem.2011.12.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin‐Acebes, M. A. , Blazquez, A. B. , Jimenez de Oya, N. , Escribano‐Romero, E. , & Saiz, J. C. (2011). West Nile virus replication requires fatty acid synthesis but is independent on phosphatidylinositol‐4‐phosphate lipids. PLoS One, 6, e24970 10.1371/journal.pone.0024970 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin‐Acebes, M. A. , Gabande‐Rodriguez, E. , Garcia‐Cabrero, A. M. , Sanchez, M. P. , Ledesma, M. D. , Sobrino, F. , & Saiz, J. C. (2016). Host sphingomyelin increases West Nile virus infection in vivo. Journal of Lipid Research, 57, 422–432. 10.1194/jlr.M064212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCollister, B. D. , Myers, J. T. , Jones‐Carson, J. , Voelker, D. R. , & Vazquez‐Torres, A. (2007). Constitutive acid sphingomyelinase enhances early and late macrophage killing of Salmonella enterica serovar Typhimurium . Infection and Immunity, 75, 5346–5352. 10.1128/IAI.00689-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mengaud, J. , Lecuit, M. , Lebrun, M. , Nato, F. , Mazie, J. C. , & Cossart, P. (1996). Antibodies to the leucine‐rich repeat region of internalin block entry of Listeria monocytogenes into cells expressing E‐cadherin. Infection and Immunity, 64, 5430–5433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menzel, N. (2012). MAP‐kinase regulated cytosolic phospholipase A2 activity is essential for production of infectious hepatitis C virus particles. PLoS Pathogens, 8(7), e1002829 10.1371/journal.ppat.1002829 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miner, M. D. , Chang, J. C. , Pandey, A. K. , Sassetti, C. M. , & Sherman, D. R. (2009). Role of cholesterol in Mycobacterium tuberculosis infection. Indian Journal of Experimental Biology, 47, 407–411. [PubMed] [Google Scholar]
- Mohn, W. W. , van der Geize, R. , Stewart, G. R. , Okamoto, S. , Liu, J. , Dijkhuizen, L. , & Eltis, L. D. (2008). The actinobacterial mce4 locus encodes a steroid transporter. The Journal of Biological Chemistry, 283, 35368–35374. 10.1074/jbc.M805496200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moorhead, A. M. , Jung, J. Y. , Smirnov, A. , Kaufer, S. , & Scidmore, M. A. (2010). Multiple host proteins that function in phosphatidylinositol‐4‐phosphate metabolism are recruited to the chlamydial inclusion. Infection and Immunity, 78, 1990–2007. 10.1128/IAI.01340-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Naroeni, A. , & Porte, F. (2002). Role of cholesterol and the ganglioside GM(1) in entry and short‐term survival of Brucella suis in murine macrophages. Infection and Immunity, 70, 1640–1644. 10.1128/IAI.70.3.1640-1644.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nawabi, P. , Catron, D. M. , & Haldar, K. (2008). Esterification of cholesterol by a type III secretion effector during intracellular Salmonella infection. Molecular Microbiology, 68, 173–185. 10.1111/j.1365-2958.2008.06142.x [DOI] [PubMed] [Google Scholar]
- Nesbitt, N. M. , Yang, X. , Fontan, P. , Kolesnikova, I. , Smith, I. , Sampson, N. S. , & Dubnau, E. (2010). A thiolase of Mycobacterium tuberculosis is required for virulence and production of androstenedione and androstadienedione from cholesterol. Infection and Immunity, 78, 275–282. 10.1128/IAI.00893-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen, L. N. , Lim, Y. S. , Pham, L. V. , Shin, H. Y. , Kim, Y. S. , & Hwang, S. B. (2014). Stearoyl coenzyme A desaturase I is associated with hepatitis C virus replication complex and regulates viral replication. Journal of Virology, 88(21), 12311–12325. 10.1128/JVI.01678-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niu, H. , Yamaguchi, M. , & Rikihisa, Y. (2008). Subversion of cellular autophagy by Anaplasma phagocytophilum . Cellular Microbiology, 10, 593–605. 10.1111/j.1462-5822.2007.01068.x [DOI] [PubMed] [Google Scholar]
- Norkin, L. C. , Wolfrom, S. A. , & Stuart, E. S. (2001). Association of caveolin with Chlamydia trachomatis inclusions at early and late stages of infection. Experimental Cell Research, 266, 229–238. 10.1006/excr.2001.5202 [DOI] [PubMed] [Google Scholar]
- Oh, Y. K. , Alpuche‐Aranda, C. , Berthiaume, E. , Jinks, T. , Miller, S. I. , & Swanson, J. A. (1996). Rapid and complete fusion of macrophage lysosomes with phagosomes containing Salmonella typhimurium . Infection and Immunity, 64, 3877–3883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ooi, L. S. , Tian, L. , Su, M. , Ho, W. S. , Sun, S. S. , Chung, H. Y. , et al. (2008). Isolation, characterization, molecular cloning and modeling of a new lipid transfer protein with antiviral and antiproliferative activities from Narcissus tazetta . Peptides, 29, 2101–2109. 10.1016/j.peptides.2008.08.020 [DOI] [PubMed] [Google Scholar]
- van Ooij, C. , Kalman, L. , van, I. , Nishijima, M. , Hanada, K. , Mostov, K. , & Engel, J. N. (2000). Host cell‐derived sphingolipids are required for the intracellular growth of Chlamydia trachomatis . Cellular Microbiology, 2, 627–637. 10.1046/j.1462-5822.2000.00077.x [DOI] [PubMed] [Google Scholar]
- Pandey, A. K. , & Sassetti, C. M. (2008). Mycobacterial persistence requires the utilization of host cholesterol. Proceedings of the National Academy of Sciences of the United States of America, 105, 4376–4380. 10.1073/pnas.0711159105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parihar, S. P. , Guler, R. , Khutlang, R. , Lang, D. M. , Hurdayal, R. , Mhlanga, M. M. , … Brombacher, F. (2014). Statin therapy reduces the Mycobacterium tuberculosis burden in human macrophages and in mice by enhancing autophagy and phagosome maturation. The Journal of Infectious Diseases, 209, 754–763. 10.1093/infdis/jit550 [DOI] [PubMed] [Google Scholar]
- Park, C. J. , Shin, R. , Park, J. M. , Lee, G. J. , You, J. S. , & Paek, K. H. (2002). Induction of pepper cDNA encoding a lipid transfer protein during the resistance response to tobacco mosaic virus. Plant Molecular Biology, 48, 243–254. 10.1023/A:1013383329361 [DOI] [PubMed] [Google Scholar]
- Pendaries, C. , Tronchere, H. , Arbibe, L. , Mounier, J. , Gozani, O. , Cantley, L. , et al. (2006). PtdIns5P activates the host cell PI3‐kinase/Akt pathway during Shigella flexneri infection. The EMBO Journal, 25, 1024–1034. 10.1038/sj.emboj.7601001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng, H. , Li, C. , Kadow, S. , Henry, B. D. , Steinmann, J. , Becker, K. A. , … Grassmé, H. (2015). Acid sphingomyelinase inhibition protects mice from lung edema and lethal Staphylococcus aureus sepsis. J Mol Med (Berl), 93, 675–689. 10.1007/s00109-014-1246-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perera, R. , Riley, C. , Isaac, G. , Hopf‐Jannasch, A. S. , Moore, R. J. , Weitz, K. W. , … Kuhn, R. J. (2012). Dengue virus infection perturbs lipid homeostasis in infected mosquito cells. PLoS Pathogens, 8, e1002584 10.1371/journal.ppat.1002584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters, J. , & Byrne, G. I. (2015). Chlamydia trachomatis growth depends on eukaryotic cholesterol esterification and is affected by acyl‐CoA:cholesterol acyltransferase inhibition. Pathogens and Disease, 73(6), ftv028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pizarro‐Cerda, J. , Payrastre, B. , Wang, Y. J. , Veiga, E. , Yin, H. L. , & Cossart, P. (2007). Type II phosphatidylinositol 4‐kinases promote Listeria monocytogenes entry into target cells. Cellular Microbiology, 9, 2381–2390. 10.1111/j.1462-5822.2007.00967.x [DOI] [PubMed] [Google Scholar]
- Poh, M. K. , Shui, G. , Xie, X. , Shi, P. Y. , Wenk, M. R. , & Gu, F. (2012). U18666A, an intra‐cellular cholesterol transport inhibitor, inhibits dengue virus entry and replication. Antiviral Research, 93, 191–198. 10.1016/j.antiviral.2011.11.014 [DOI] [PubMed] [Google Scholar]
- Rao, G. A. , & Pandya, P. K. (2011). Statin therapy improves sustained virologic response among diabetic patients with chronic hepatitis C. Gastroenterology, 140(1), 144–152. 10.1053/j.gastro.2010.08.055 [DOI] [PubMed] [Google Scholar]
- Reiss, S. , Rebhan, I. , Backes, P. , Romero‐Brey, I. , Erfle, H. , Matula, P. , … Bartenschlager, R. (2011). Recruitment and activation of a lipid kinase by hepatitis C virus NS5A is essential for integrity of the membranous replication compartment. Cell Host & Microbe, 9, 32–45. 10.1016/j.chom.2010.12.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rendina, A. R. , & Cheng, D. (2005). Characterization of the inactivation of rat fatty acid synthase by C75: Inhibition of partial reactions and protection by substrates. The Biochemical Journal, 388, 895–903. 10.1042/BJ20041963 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riethmuller, J. , Anthonysamy, J. , Serra, E. , Schwab, M. , Doring, G. , & Gulbins, E. (2009). Therapeutic efficacy and safety of amitriptyline in patients with cystic fibrosis. Cellular Physiology and Biochemistry: International Journal of Experimental Cellular Physiology, Biochemistry, and Pharmacology, 24, 65–72. 10.1159/000227814 [DOI] [PubMed] [Google Scholar]
- Roe, B. , Kensicki, E. , Mohney, T. , & Hall, W. W. (2011). Metabolomic profile of hepatitis C virus‐infected hepatocytes. PLoS One, 6, e23641 10.1371/journal.pone.0023641 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romero‐Brey, I. , Merz, A. , Chiramel, A. , Lee, J. Y. , Chlanda, P. , Haselman, U. , … Bartenschlager, R. (2012). Three‐dimensional architecture and biogenesis of membrane structures associated with hepatitis C virus replication. PLoS Pathogens, 8, e1003056 10.1371/journal.ppat.1003056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rothwell, C. , Lebreton, A. , Young Ng, C. , Lim, J. Y. , Liu, W. , Vasudevan, S. , et al. (2009). Cholesterol biosynthesis modulation regulates dengue viral replication. Virology, 389, 8–19. 10.1016/j.virol.2009.03.025 [DOI] [PubMed] [Google Scholar]
- Roulin, P. S. , Lotzerich, M. , Torta, F. , Tanner, L. B. , van Kuppeveld, F. J. , Wenk, M. R. , & Greber, U. F. (2014). Rhinovirus uses a phosphatidylinositol 4‐phosphate/cholesterol counter‐current for the formation of replication compartments at the ER–Golgi interface. Cell Host & Microbe, 16, 677–690. 10.1016/j.chom.2014.10.003 [DOI] [PubMed] [Google Scholar]
- Russell, D. G. , Cardona, P. J. , Kim, M. J. , Allain, S. , & Altare, F. (2009). Foamy macrophages and the progression of the human tuberculosis granuloma. Nature Immunology, 10, 943–948. 10.1038/ni.1781 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rutaganira, F. U. , Fowler, M. L. , McPhail, J. A. , Gelman, M. A. , Nguyen, K. , Xiong, A. , … Burke, J. E. (2016). Design and structural characterization of potent and selective inhibitors of phosphatidylinositol 4 kinase IIIβ. Journal of Medicinal Chemistry, 59, 1830–1839. 10.1021/acs.jmedchem.5b01311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakamoto, H. , Okamoto, K. , Aoki, M. , Kato, H. , Katsume, A. , Ohta, A. , … Sudoh, M. (2005). Host sphingolipid biosynthesis as a target for hepatitis C virus therapy. Nature Chemical Biology, 1, 333–337. 10.1038/nchembio742 [DOI] [PubMed] [Google Scholar]
- Sarkar, A. , Hellberg, L. , Bhattacharyya, A. , Behnen, M. , Wang, K. , Lord, J. M. , … Laskay, T. (2012). Infection with Anaplasma phagocytophilum activates the phosphatidylinositol 3‐kinase/Akt and NF‐κB survival pathways in neutrophil granulocytes. Infection and Immunity, 80, 1615–1623. 10.1128/IAI.05219-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroeder, T. H. , Lee, M. M. , Yacono, P. W. , Cannon, C. L. , Gerceker, A. A. , Golan, D. E. , & Pier, G. B. (2002). CFTR is a pattern recognition molecule that extracts Pseudomonas aeruginosa LPS from the outer membrane into epithelial cells and activates NF‐κB translocation. Proceedings of the National Academy of Sciences of the United States of America, 99, 6907–6912. 10.1073/pnas.092160899 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahsavandi, S. , Ebrahimi, M. M. , & Hasaninejad Farahani, A. (2017). Interfering with lipid raft association: A mechanism to control influenza virus infection by Sambucus nigra . Iranian Journal of Pharmaceutical Research: IJPR, 16, 1147–1154. [PMC free article] [PubMed] [Google Scholar]
- Sharma, M. , Sasvari, Z. , & Nagy, P. D. (2010). Inhibition of sterol biosynthesis reduces tombusvirus replication in yeast and plants. Journal of Virology, 84, 2270–2281. 10.1128/JVI.02003-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen, Y. , Naujokas, M. , Park, M. , & Ireton, K. (2000). InIB‐dependent internalization of Listeria is mediated by the Met receptor tyrosine kinase. Cell, 103, 501–510. 10.1016/S0092-8674(00)00141-0 [DOI] [PubMed] [Google Scholar]
- Shoemaker, C. J. , Schornberg, K. L. , Delos, S. E. , Scully, C. , Pajouhesh, H. , Olinger, G. G. , … White, J. M. (2013). Multiple cationic amphiphiles induce a Niemann–Pick C phenotype and inhibit Ebola virus entry and infection. PLoS One, 8, e56265 10.1371/journal.pone.0056265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simonis, A. , Hebling, S. , Gulbins, E. , Schneider‐Schaulies, S. , & Schubert‐Unkmeir, A. (2014). Differential activation of acid sphingomyelinase and ceramide release determines invasiveness of Neisseria meningitidis into brain endothelial cells. PLoS Pathogens, 10, e1004160 10.1371/journal.ppat.1004160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh, R. , Rao, V. , Shakila, H. , Gupta, R. , Khera, A. , Dhar, N. , … Tyagi, A. K. (2003). Disruption of mptpB impairs the ability of Mycobacterium tuberculosis to survive in guinea pigs. Molecular Microbiology, 50, 751–762. 10.1046/j.1365-2958.2003.03712.x [DOI] [PubMed] [Google Scholar]
- Soares, M. M. , King, S. W. , & Thorpe, P. E. (2008). Targeting inside‐out phosphatidylserine as a therapeutic strategy for viral diseases. Nature Medicine, 14, 1357–1362. 10.1038/nm.1885 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sohal, A. K. , Pallas, J. A. , & Jenkins, G. I. (1999). The promoter of a Brassica napus lipid transfer protein gene is active in a range of tissues and stimulated by light and viral infection in transgenic Arabidopsis . Plant Molecular Biology, 41, 75–87. 10.1023/A:1006232700835 [DOI] [PubMed] [Google Scholar]
- Soto‐Acosta, R. , Bautista‐Carbajal, P. , Cervantes‐Salazar, M. , Angel‐Ambrocio, A. H. , & Del Angel, R. M. (2017). DENV up‐regulates the HMG‐CoA reductase activity through the impairment of AMPK phosphorylation: A potential antiviral target. PLoS Pathogens, 13, e1006257 10.1371/journal.ppat.1006257 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stancu, C. , & Sima, A. (2001). Statins: Mechanism of action and effects. Journal of Cellular and Molecular Medicine, 5, 378–387. 10.1111/j.1582-4934.2001.tb00172.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Staring, J. , von Castelmur, E. , Blomen, V. A. , van den Hengel, L. G. , Brockmann, M. , Baggen, J. , … Brummelkamp, T. R. (2017). PLA2G16 represents a switch between entry and clearance of Picornaviridae. Nature, 541, 412–416. 10.1038/nature21032 [DOI] [PubMed] [Google Scholar]
- Stehr, M. , Elamin, A. A. , & Singh, M. (2012). Cytosolic lipid inclusions formed during infection by viral and bacterial pathogens. Microbes and Infection, 14, 1227–1237. 10.1016/j.micinf.2012.08.001 [DOI] [PubMed] [Google Scholar]
- Strating, J. R. , van der Linden, L. , Albulescu, L. , Bigay, J. , Arita, M. , Delang, L. , et al. (2015). Itraconazole inhibits enterovirus replication by targeting the oxysterol‐binding protein. Cell Reports, 10, 600–615. 10.1016/j.celrep.2014.12.054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tachado, S. D. , Samrakandi, M. M. , & Cirillo, J. D. (2008). Non‐opsonic phagocytosis of Legionella pneumophila by macrophages is mediated by phosphatidylinositol 3‐kinase. PLoS One, 3, e3324 10.1371/journal.pone.0003324 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tatematsu, K. , Tanaka, Y. , Sugiyama, M. , Sudoh, M. , & Mizokami, M. (2011). Host sphingolipid biosynthesis is a promising therapeutic target for the inhibition of hepatitis B virus replication. Journal of Medical Virology, 83, 587–593. 10.1002/jmv.21970 [DOI] [PubMed] [Google Scholar]
- Tenorio, R. , Fernandez de Castro, I. , Knowlton, J. J. , Zamora, P. F. , Lee, C. H. , Mainou, B. A. , … Risco, C. (2018). Reovirus σNS and μNS proteins remodel the endoplasmic reticulum to build replication neo‐organelles. MBio, 9(4), e01253–e01218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terebiznik, M. R. , Vieira, O. V. , Marcus, S. L. , Slade, A. , Yip, C. M. , Trimble, W. S. , … Grinstein, S. (2002). Elimination of host cell PtdIns(4,5)P(2) by bacterial SigD promotes membrane fission during invasion by Salmonella . Nature Cell Biology, 4, 766–773. 10.1038/ncb854 [DOI] [PubMed] [Google Scholar]
- Toledo, A. , & Benach, J. L. (2015). Hijacking and use of host lipids by intracellular pathogens. Microbiology Spectrum, 3(6). 10.1128/microbiolspec.VMBF-0001-2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Utermohlen, O. , Herz, J. , Schramm, M. , & Kronke, M. (2008). Fusogenicity of membranes: The impact of acid sphingomyelinase on innate immune responses. Immunobiology, 213, 307–314. 10.1016/j.imbio.2007.10.016 [DOI] [PubMed] [Google Scholar]
- Uto, Y. (2016). Recent progress in the discovery and development of stearoyl CoA desaturase inhibitors. Chemistry and Physics of Lipids, 197, 3–12. 10.1016/j.chemphyslip.2015.08.018 [DOI] [PubMed] [Google Scholar]
- Vance, D. E. , Trip, E. M. , & Paddon, H. B. (1980). Poliovirus increases phosphatidylcholine biosynthesis in HeLa cells by stimulation of the rate‐limiting reaction catalyzed by CTP: Phosphocholine cytidylyltransferase. The Journal of Biological Chemistry, 255, 1064–1069. [PubMed] [Google Scholar]
- Vazquez, C. L. , Rodgers, A. , Herbst, S. , Coade, S. , Gronow, A. , Guzman, C. A. , et al. (2016). The proneurotrophin receptor sortilin is required for Mycobacterium tuberculosis control by macrophages. Scientific Reports, 6, 29332 10.1038/srep29332 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Viner, R. , Chetrit, D. , Ehrlich, M. , & Segal, G. (2012). Identification of two Legionella pneumophila effectors that manipulate host phospholipids biosynthesis. PLoS Pathogens, 8, e1002988 10.1371/journal.ppat.1002988 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallqvist, A. , Wang, H. , Zavaljevski, N. , Memisevic, V. , Kwon, K. , Pieper, R. , et al. (2017). Mechanisms of action of Coxiella burnetii effectors inferred from host–pathogen protein interactions. PLoS One, 12, e0188071 10.1371/journal.pone.0188071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, H. , Perry, J. W. , Lauring, A. S. , Neddermann, P. , De Francesco, R. , & Tai, A. W. (2014). Oxysterol‐binding protein is a phosphatidylinositol 4‐kinase effector required for HCV replication membrane integrity and cholesterol trafficking. Gastroenterology, 146(1373‐1385), e1371–e1311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watarai, M. , Makino, S. , Fujii, Y. , Okamoto, K. , & Shirahata, T. (2002). Modulation of Brucella‐induced macropinocytosis by lipid rafts mediates intracellular replication. Cellular Microbiology, 4, 341–355. 10.1046/j.1462-5822.2002.00195.x [DOI] [PubMed] [Google Scholar]
- Watarai, M. , Makino, S. , Michikawa, M. , Yanagisawa, K. , Murakami, S. , & Shirahata, T. (2002). Macrophage plasma membrane cholesterol contributes to Brucella abortus infection of mice. Infection and Immunity, 70, 4818–4825. 10.1128/IAI.70.9.4818-4825.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber, S. S. , Ragaz, C. , Reus, K. , Nyfeler, Y. , & Hilbi, H. (2006). Legionella pneumophila exploits PI(4)P to anchor secreted effector proteins to the replicative vacuole. PLoS Pathogens, 2, e46 10.1371/journal.ppat.0020046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welsch, S. , Miller, S. , Romero‐Brey, I. , Merz, A. , Bleck, C. K. , Walther, P. , et al. (2009). Composition and three‐dimensional architecture of the dengue virus replication and assembly sites. Cell Host & Microbe, 5, 365–375. 10.1016/j.chom.2009.03.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welte, M. A. (2015). Expanding roles for lipid droplets. Current Biology, 25, R470–R481. 10.1016/j.cub.2015.04.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weng, L. , Hirata, Y. , Arai, M. , Kohara, M. , Wakita, T. , Watashi, K. , … Toyoda, T. (2010). Sphingomyelin activates hepatitis C virus RNA polymerase in a genotype‐specific manner. Journal of Virology, 84, 11761–11770. 10.1128/JVI.00638-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolf, K. , & Hackstadt, T. (2001). Sphingomyelin trafficking in Chlamydia pneumoniae‐infected cells. Cellular Microbiology, 3, 145–152. 10.1046/j.1462-5822.2001.00098.x [DOI] [PubMed] [Google Scholar]
- Wong, L. H. , Copic, A. , & Levine, T. P. (2017). Advances on the transfer of lipids by lipid transfer proteins. Trends in Biochemical Sciences, 42, 516–530. 10.1016/j.tibs.2017.05.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wylie, J. L. , Hatch, G. M. , & McClarty, G. (1997). Host cell phospholipids are trafficked to and then modified by Chlamydia trachomatis . Journal of Bacteriology, 179, 7233–7242. 10.1128/jb.179.23.7233-7242.1997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiong, Q. , Lin, M. , & Rikihisa, Y. (2009). Cholesterol‐dependent Anaplasma phagocytophilum exploits the low‐density lipoprotein uptake pathway. PLoS Pathogens, 5, e1000329 10.1371/journal.ppat.1000329 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiong, Q. , & Rikihisa, Y. (2012). Subversion of NPC1 pathway of cholesterol transport by Anaplasma phagocytophilum . Cellular Microbiology, 14, 560–576. 10.1111/j.1462-5822.2011.01742.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu, K. , & Nagy, P. D. (2015). RNA virus replication depends on enrichment of phosphatidylethanolamine at replication sites in subcellular membranes. Proceedings of the National Academy of Sciences of the United States of America, 112, E1782–E1791. 10.1073/pnas.1418971112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, W. , Hood, B. L. , Chadwick, S. L. , Liu, S. , Watkins, S. C. , Luo, G. , … Wang, T. (2008). Fatty acid synthase is up‐regulated during hepatitis C virus infection and regulates hepatitis C virus entry and production. Hepatology, 48, 1396–1403. 10.1002/hep.22508 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao, J. , Cherian, P. T. , Frank, M. W. , & Rock, C. O. (2015). Chlamydia trachomatis relies on autonomous phospholipid synthesis for membrane biogenesis. The Journal of Biological Chemistry, 290, 18874–18888. 10.1074/jbc.M115.657148 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao, J. , Dodson, V. J. , Frank, M. W. , & Rock, C. O. (2015). Chlamydia trachomatis scavenges host fatty acids for phospholipid synthesis via an acyl–acyl carrier protein synthetase. The Journal of Biological Chemistry, 290, 22163–22173. 10.1074/jbc.M115.671008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yasuda, S. , Kitagawa, H. , Ueno, M. , Ishitani, H. , Fukasawa, M. , Nishijima, M. , … Hanada, K. (2001). A novel inhibitor of ceramide trafficking from the endoplasmic reticulum to the site of sphingomyelin synthesis. The Journal of Biological Chemistry, 276, 43994–44002. 10.1074/jbc.M104884200 [DOI] [PubMed] [Google Scholar]
- Yin, X. , Ambardekar, C. , Lu, Y. , & Feng, Z. (2016). Distinct entry mechanisms for nonenveloped and quasi‐enveloped hepatitis E viruses. Journal of Virology, 90, 4232–4242. 10.1128/JVI.02804-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, J. , Zhang, Z. , Chukkapalli, V. , Nchoutmboube, J. A. , Li, J. , Randall, G. , … Wang, X. (2016). Positive‐strand RNA viruses stimulate host phosphatidylcholine synthesis at viral replication sites. Proceedings of the National Academy of Sciences of the United States of America, 113, E1064–E1073. 10.1073/pnas.1519730113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zou, Y. , Lei, W. , Su, S. , Bu, J. , Zhu, S. , Huang, Q. , & Li, Z. (2018). Chlamydia trachomatis plasmid‐encoded protein Pgp3 inhibits apoptosis via the PI3K–AKT‐mediated MDM2–p53 axis. Molecular and Cellular Biochemistry. 10.1007/s11010-018-3422-9 [DOI] [PubMed] [Google Scholar]