

Abstract
Background and Objectives
Allogeneic hematopoietic stem cell transplantation (HSCT) is a routine clinical procedure performed to treat patients with haematological malignancies, primary immune deficiencies or metabolic disorders. Infections during lymphopenia after allogeneic HSCT are associated with high mortality and morbidity. Typical infectious agents are Epstein–Barr virus, cytomegalovirus, herpes simplex virus, varicella‐zoster virus and fungi. The study aim was to evaluate whether measurement of the responses of antigen‐specific T‐cells, recognizing infectious pathogens would correlate to protective functions in the stem cell recipient post‐transplant.
Materials and Methods
Twenty‐one grafts were analysed by flow cytometry and cells were stimulated in vitro with relevant infectious antigens, followed by evaluation of T‐cell proliferation and cytokine production. Results were compared to the recipients’ clinical records 1‐year post‐transplantation.
Results
We show that an extensive repertoire of transferred antigen‐specific T‐cells from allogeneic donor grafts against infectious agents, involved in post‐transplant infections, are linked to an absence of infectious complications for the recipient up‐to 1‐year post‐transplant. The protective effect was associated with antigen‐specific T‐cell proliferation and IL‐1β secretion.
Conclusion
Our results suggest that assaying T‐cell function before HSCT could determine individual risks for infectious complications and thus aid in clinical decision‐making regarding prophylactic and pre‐emptive anti‐infective therapy.
Keywords: allogeneic hematopoietic stem cell transplantation, FASCIA, infectious complications, stem cell products, transferred immunity
Introduction
Allogeneic hematopoietic stem cell transplantation (HSCT) is a routine clinical procedure performed on more than 20 000 patients annually, mostly to treat patients with haematological malignancies but also primary immune or metabolic deficiencies 1. The source of stem cells can be bone marrow (BM), umbilical cord blood or peripheral blood stem cells (PBSC) 2. In order to prevent rejection of the new graft and to reduce the number of malignant cells, patients are conditioned before transplantation with either myeloablative, containing high dose cytotoxic drugs with or without whole body irradiation or reduced intensity conditioning (RIC) 3. Stem cell products used for allogeneic HSCT contain CD34+ hematopoietic cells and a number of additional leucocytes, that is granulocytes, monocytes, T‐, B‐ and NK‐cells 4. The quality of the allogeneic stem cell graft at the time of transplantation is of importance for clinical outcome. It was recently shown, that poor graft viability was related to increased incidence of acute GVHD and transplant related mortality 5, indicating that the stem cell graft composition with respect to antigen specific T‐ and B‐cells is of importance for immune reconstitution and clinical outcome 6.
Allogeneic HSCT is associated with a number of serious complications. The transplanted graft can induce graft‐versus‐host disease (GVHD), a potentially life threatening condition where donor T‐cells in the graft attack recipient tissues. Recently, graft manipulations before transplantation have been utilized to prevent GVHD, for example through T‐ and B‐cells depletion from haploidentical stem cell grafts or through depletion of T‐cells expressing the T‐cell receptor (TCR) αβ, leaving the TCRγδ+ T‐cells in the graft 7, 8. There are conflicting data regarding the benefits of TCRγδ+ T‐cells in allogeneic HSCT. Recent studies suggest a favourable clinical outcome for patients with elevated levels of TCRγδ+ T‐cells after transplantation 2, 9, 10.
Infections during the lymphopenic state after stem cell transplantation is another complication associated with high mortality and morbidity. Some of the most common agents causing post‐transplant infections are Epstein–Barr virus (EBV), cytomegalovirus (CMV), herpes simplex virus (HSV), varicella‐zoster virus (VZV) as well as fungal infections caused by candida or aspergillus 3, 11. A considerable effort is dedicated to reducing the risk of CMV and EBV reactivation as well as to prevent fungal infections. Prophylactic antimicrobial drugs are administered and patient and donor pairs are screened to prevent pairing of CMV and EBV sero‐negative donors with sero‐positive recipients in combination with early microbiological diagnostics 12, 13. Moreover, adoptive transfer of virus‐specific T‐cell clones can be used as pre‐emptive and antiviral treatment 14, 15, 16, 17.
Here we show a possible protective function by means of evaluating proliferation and cytokine production of antigen‐specific T‐ and B‐cells in 21 stem cell products used for allogeneic HSCT. The donor cell products were stimulated with infectious antigens commonly involved in transplant associated infectious complications, and the results were correlated to the clinical outcome 1‐year post‐transplantation. Moreover, an extensive repertoire of transferred effector functions of antigen‐specific T‐cells against HSV, EBV, CMV, VZV, adenovirus and Candida albicans, in allogeneic donor stem cell grafts is associated with an absence of infectious complications within 1‐year post‐transplant. The protective effect was not associated with graft cell‐composition or the transfused cell dose.
Materials and methods
Study participants and infectious complications
Approximately 50 × 106 cells from stem cell products (5 BM and 17 PBSC) from healthy donors used for allogeneic HSCT were obtained from the stem cell laboratory at Karolinska University Hospital. One patient died 5 days post‐transplantation and data from the corresponding stem cell donor was excluded from the study, except in the comparisons between donor serology and the cellular responses measured by FASCIA to HSV, EBV, CMV or VZV.
Viral reactivation of CMV was defined as a post‐transplant‐positive blood sample with >10 000 CMV copies/ml as assessed by PCR or evidence of CMV induced pathology in biopsy specimens from the affected organ. Viral reactivation of EBV was defined as clinically symptomatic disease coupled to a post‐transplant‐positive blood sample with >1000 EBV copies/ml as assessed by PCR or evidence of EBV‐induced pathology in biopsy specimens from the affected organ. VZV reactivation was defined as post‐transplant clinically symptomatic VZV disease with positive VZV PCR in a sample from the affected body site. HSV1 and HSV2 reactivation was defined as post‐transplant clinically symptomatic disease with positive HSV1 or HSV2 PCR in a sample from the affected body site. Coronavirus pneumonia was defined as post‐transplant clinically symptomatic disease accompanied by a chest CT showing typical infiltrates and coronavirus‐positive respiratory specimens as assessed by PCR. Oral candidiasis was defined as post‐transplant clinically symptomatic disease accompanied by positive oral fungal cultures. Bacterial septicaemia was defined as blood culture‐positive post‐transplant clinical sepsis. Patient and donor information is displayed in Table 1. The regional ethical board granted ethical approval for the study.
Table 1.
Recipient and donor clinical data
| Product | Age R (yrs) | Age D (yrs) | Source | MUD/Sib | Engraftment (day) | TNC/kg bw (108/kg) | CMV serology (D) | CMV serology (R) | EBV serology (D) | EBV serology (R) | Diagnosis | Infectious complications within 1 yr after HSCT | ATG Yes/no | Other clinical information |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 66 | 25 | PB | MUD | 25 | 5·8 | Pos | Pos | Neg | Posb | ALL Ph+ | CMV, HSV‐2 HSV‐1 | Y | Relapse |
| 2 | 47 | 41 | BM | MUD | 19 | 2·7 | Pos | Pos | Pos | Pos | AML | Y | ||
| 3 | 48 | 46 | PB | Sib | 11 | 23·2 | Neg | Posa | Pos | Pos | MDS | CMV, Staph + E‐coli sepsis, HSV‐1 | N | GVHD grade III, Deceased |
| 4 | 28 | 24 | PB | MUD | 15 | 16·7 | Pos | Pos | Pos | Pos | Pre‐B ALL | Y | Chronic GVHD grade I | |
| 5 | 1 | 29 | BM | MUD | 20 | 11·4 | Neg | Neg | Pos | Negb | CGD (X‐linked) | EBV, CMV | Y | |
| 6 | 16 | 28 | BM | MUD | 14 | 3·9 | Neg | Posa | Pos | Pos | Pre‐B ALL | CMV, Oral Candidiasis | Y | |
| 7 | 53 | 26 | PB | MUD | 14 | 8 | Neg | Posa | Pos | Pos | MDS | Y | ||
| 8 | 35 | 28 | PB | MUD | 22 | 12 | Neg | Posa | Pos | Pos | Pre‐B ALL | CMV, HSV‐1, EBV | Y | GVHD grade II, CMV CTL |
| 9 | 68 | 38 | PB | MUD | 18 | 11·3 | Neg | Posa | Pos | Pos | MDS | VZV, CMV | Y | Relapse |
| 10 | 10 | 7 | BM | Sib | 17 | 3·9 | Pos | Pos | Neg | Posb | Pre‐B ALL | VZV | N | Chronic GVHD grade II |
| 11 | 67 | 33 | PB | MUD | 32 | 14·2 | Pos | Pos | Pos | Pos | AML | Y | Pneumonia, Deceased | |
| 12 | 66 | 29 | PB | MUD | 16 | 7·9 | Neg | Neg | Pos | Pos | AML | VZV | Y | |
| 13 | 9 | 27 | PB | MUD | 15 | 25·4 | Pos | Pos | Pos | Pos | AML Ph+ | Y | ||
| 15 | 44 | 43 | PB | MUD | 18 | 4·5 | Neg | Posa | Pos | Pos | AML | CMV | N | |
| 16 | 49 | 34 | PB | MUD | 17 | 7 | Neg | Posa | Pos | Negb | Myeloma | EBV, CMV, Salmonella+Staph abscess, sepsis. | Y | Relapse |
| 17 | 46 | 34 | PB | MUD | 31 | 14·3 | Pos | Pos | Pos | Pos | AML | N | Relapse, Deceased | |
| 18 | 48 | 24 | PB | MUD | 19 | 12·3 | Pos | Pos | Neg | Neg | AML | CMV, VZV | Y | Relapse |
| 19 | 49 | 46 | PB | Sib | 18 | 4·1 | Pos | Pos | Pos | Pos | AML/myelofibrosis | N | Relapse, Deceased | |
| 20 | 62 | 26 | PB | MUD | 16 | 6·8 | Pos | Pos | Pos | Pos | MDS | Y | ||
| 21 | 68 | 27 | PB | MUD | 13 | 13·2 | Neg | Neg | Pos | Pos | AML | Y | ||
| 22 | 15 | 19 | BM | Sib | 19 | 3·3 | Neg | Neg | Neg | Neg | T‐cell lymphoma | VZV, Oral Candidiasis, Corona virus pneumonia | N | GVHD grade II |
|
Median 48 (1–68) |
Median 28 (7–46) |
16 PB/5 BM | 17 MUD/4 Sib |
Median 18 (11–32) |
Median 8 (2·7–25·4) | 10 P/11 N | 17 P/4 N | 17 P/4 N | 16 P/5 N | 15/6 Y/N |
D, Donor, R: recipient, PB, peripheral blood stem cells, BM, bone marrow, MUD, matched unrelated donor, Sib, sibling, TNC, Total nucleated cells, ALL, acute lymphocytic leukaemia, AML, acute myeloid leukaemia, MDS, myelodysplastic syndrome, CGD, chronic granulomatous disease, CMV, Cytomegalovirus, HSV, Herpes simplex virus 1 and 2, EBV, Epstein–Barr virus, VZV, Varicella‐zoster virus, ATG, antithymocyte immuno‐globuline, GVHD, Graft versus host disease.
CMV mismatch.
EBV mismatch.
This article is being made freely available through PubMed Central as part of the COVID-19 public health emergency response. It can be used for unrestricted research re-use and analysis in any form or by any means with acknowledgement of the original source, for the duration of the public health emergency.
Antigens and pharmaceutical drugs
All mitogens and antigens were titrated for optimal responses. The following concentrations were used in the FASCIA (flow cytometric assay for specific cell‐mediated immune‐response in activated whole blood) experiments: 5 μg/ml pokeweed mitogen (PWM), 100 ng/ml of both staphylococcal enterotoxin A and B (SEA + SEB) (all from Sigma Aldrich, MO, USA), 10 μg/ml tuberculin purified protein derivative (PPD), 4 IU/ml tetanus toxin (TT) (both from Statens Serum Institute, Copenhagen, DK) and 20 μg/ml Candida albicans (Greer Laboratories Inc., NC, USA). Influenza vaccine ‘Fluarix’, varicella zoster virus vaccine (VZV) ‘Varilrix’ (both from GlaxoSmithKline AB, Middlesex, UK) were diluted 1:100. 40 μg/ml adenovirus grade 2 antigen, 50 μg/ml P3H3 cell extract (EBV), 40 μg/ml HSV type 1 antigen and 40 μg/ml CMV grade 2 antigen (Microbix Biosystems Inc., Mississauga, ON, Canada).
FASCIA‐functional proliferation assay
The functional responses to infectious antigens were assessed by the clinically used flow cytometry based FASCIA‐method 18. To remove citrate from the stem cell products cells were washed twice and resuspended in FASCIA culture media containing RPMI 1640 (Gibco) supplemented with 10% human AB‐serum, supplemented 100 IU/ml penicillin, 100 IU/ml streptomycin (all from Gibco, Paisley, UK), 2 mm L‐glutamine (Invitrogen, MD, USA) and 2·0 μmol/ml Ca2+ (Calcium‐Sandoz, Sandoz, NJ, USA). The stem cell products (5 BM and 17 PB) were stimulated with tetanus toxin, PWM, SEA + SEB, influenza‐, VZV‐, adenovirus‐, CMV‐, EBV‐, HSV antigen, candida or left unstimulated according to a revised FASCIA‐protocol where 600 000 cells were cultured in 1 ml of culture media containing Ca2+ and incubated for 7 days in 37° C, 5% CO2 and 95% humidity. The cell supernatants were removed on day 7 and stored at −80° C until cytokine/chemokine analysis was performed. Cells were stained with CD3‐FITC/CD4‐PE Simultest mix and CD25‐APC (BD Biosciences, CA, USA). The background, PWM‐ and EBV‐stimulated FASCIA culture tubes were also stained with CD19‐PC7 (Beckman Coulter, Marseille, France). The tubes were incubated 10 min at room temperature (RT) in the dark, followed by erythrocytes lysing with 1× IO Test lysing solution (Beckman Coulter) and washing. The cell pellets were resuspended in 450 μl PBS. Blast numbers were acquired during 80 seconds with a Navios flow cytometer (Beckman Coulter, CA, USA). A stimulation index (SI) (the number of stimulated CD3+ or CD19+ blasts divided by the number of blasts in the unstimulated tubes) was calculated to compare the proliferative responses against infectious antigens. A stimulation index greater than 4 was considered a positive cellular response. The cut‐off was based on the median antigen‐specific T‐cell response from 14 healthy blood donors stimulated with influenza vaccine and TT (SI ranged from 0·37 to 31·6, data not shown).
Lineage markers and T‐cell subsets
Lineage markers and T‐cell subtypes in the donor stem cell products were analysed by flow cytometry. The cells were stained with the following antibodies: CD34‐FITC, TCRαβ‐FITC, CD16‐PE, CD56‐PE, CD25‐APC (all from BD Biosciences), CD45‐ECD, CD4‐PC7, CD19‐PC7, CD3‐Pacific blue (all from Beckman Coulter), TCRγδ‐PE (Miltenyi Biotec) and CD8‐PC5 (Dako). Acquisition was performed on a Navios flow cytometry instrument and analysed using kaluza software (Beckman Coulter).
Cytokine detection
Cytokines and chemokines in culture supernatants from unstimulated FASCIA tubes and tubes stimulated with PWM, CMV, EBV and HSV from all patients were measured with a 5‐plex Milliplex human cytokine⁄chemokine kit, containing monoclonal antibodies to detect IFNγ, IL‐1b, IL‐4, IL‐10 and IL‐17A according to the manufacturer's instructions (EMD Millipore corporation, MA, USA). Sample analysis was performed on a Magpix instrument using xponent software (Luminex, TX, USA). TGFβ‐1 was measured with ELISA, according to the manufacturer's instruction (R&D Systems, MN, USA).
Statistics
One‐way anova and followed by Bonferroni′s multiple comparison test were used to determine differences between groups defined by infectious complications as well as comparisons on cell frequency. Fisher′s exact test was used for nominal variables, comparing FASCIA responses with reactivation of EBV or CMV. Finally, a composite analysis was performed where a positive response against any of HSV, EBV, CMV, VZV, candida or adenovirus rendered 1 point with a maximum of 6 points. Students’ t‐test was used for statistical calculations. In all statistical analysis a P‐value <0·05 was considered statistically significant. For the statistical analysis graph pad prism version 6 (GraphPad Software Inc., CA, USA) was used.
Results
Allogeneic stem cell donors and patients
A majority, 17 of 21 donors, was matched unrelated donors. The rest consisted of matched siblings. The median age of the recipients was 48 years (range 1–68). All patients, except one with X‐linked chronic granulomatous disease (CGD), suffered from haematological malignancies. ATG were administered to 15 of 21 patients as part of their conditioning regime. The stem cell doses of mononuclear cells ranged from 2·7 to 25·4 × 108/kg bodyweight (median 9·6 × 108/kg). It took an average of 18 days (range 11–32 days) for engraftment, defined as an absolute neutrophil count of at least 0·5 × 109/L, to occur. At clinical follow‐up 1‐year post‐transplant, 12 of the 21 patients had experienced either reactivation of one or more latent viruses, that is CMV, HSV, EBV or VZV or suffered from candidiasis. Among the seven cases where CMV sero‐positive recipients received stem cell products from sero‐negative donors, six of the recipients had experienced CMV infection. The two EBV sero‐positive recipients receiving sero‐negative stem cell products had not experienced EBV reactivation at the 1‐year follow‐up. Five patients were diagnosed with GVHD (grade I–III), six relapsed and four patients were deceased at the 1‐year follow‐up (Table 1).
Cellular responses in stem cell products against infectious antigens measured by FASCIA
Sixteen stem cell products obtained from peripheral blood and five from BM were evaluated for their cellular responses against infectious antigens prior to transplant. Using a modified version of the FASCIA method, cells were left unstimulated or stimulated with PWM, SEA/SEB, CMV‐, VZV‐, HSV‐, EBV‐, influenza‐, adenovirus‐antigens, candida or TT. Positive controls; PWM (SI mean 45) and SEA/SEB (SI mean 33) elicited positive T‐cell proliferation in all stem cell products. The mean frequency of cell products with positive antigen‐specific T‐cells response, was 23% against CMV, 32% against VZV, 23% against HSV, 81% against EBV, 50% against TT, 68% against candida, 59% against influenza and 45% against adenovirus antigen. When B‐cell proliferation in the stem cell products were analysed, 71% showed positive response against PWM and 91% of were positive against EBV (Fig. 1a).
Figure 1.
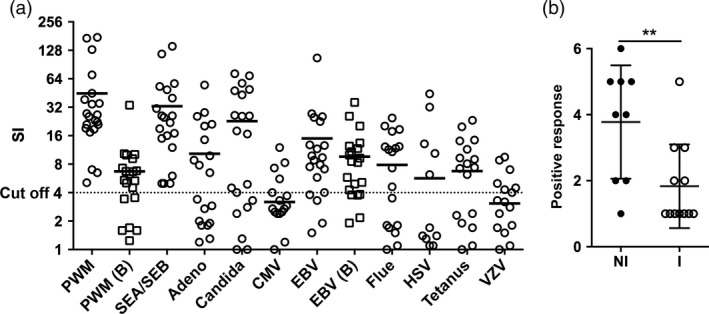
Functional analysis of T‐and B‐cells in stem cells products. Proliferation of T‐and B‐cells in donor stem cell products was analysed using the FASCIA method. B‐cell proliferation was only measured upon stimulation with PWM and EBV, depicted PMW (b) and EBV (b) in 1a. Cells were unstimulated or stimulated with PWM, SEA/SEB, CMV, VZV, HSV, EBV, tetanus toxoid, candida, influenza antigen or adenovirus antigen. Proliferation was expressed as stimulation indices (SI) with an SI > 4 considered positive. A: Proliferation against antigens, horizontal line represents mean SI and the dotted line represents cut‐off. B: Stem cell products were divided into two groups, ‘infection’ (I, n = 12) and ‘no infection’ (NI, n = 9) based on patients’ clinical outcome. A composite analysis was performed where a positive response against any of HSV, EBV, CMV, VZV, candida or adenovirus rendered 1 point with a maximum of 6 points. **P‐value < 0·01.
Since all stem cell donors for allogeneic HSCT are tested for humoral responses, that is, serum IgG, against HSV, EBV, CMV or VZV, these could be compared to the proliferative responses against the same antigens by the FASCIA method. There was a large concurrence of serologic positivity and the presence of positive proliferative responses to the same antigen. The greatest concordance was observed for EBV with 14 of 17 (83%) EBV‐sero‐positive donors showing positive proliferative responses against EBV antigens by the FASCIA method (Table 2).
Table 2.
Serology and FASCIA response for donor stem cells products
| HSV serology | HSV FASCIA | EBV serology | EBV FASCIA | CMV serology | CMV FASCIA | VZV serology | VZV FASCIA | |
|---|---|---|---|---|---|---|---|---|
| 1 | − | N | − | N | + | N | + | N |
| 2 | + | N | + | N | + | P | + | N |
| 3 | + | N | + | P | − | N | + | N |
| 4 | + | P | + | P | + | N | + | P |
| 5 | − | N | + | N | − | N | + | N |
| 6 | + | N | + | P | − | N | + | N |
| 7 | + | P | + | P | − | N | + | P |
| 8 | − | N | + | P | − | N | + | N |
| 9 | − | N | + | N | − | N | + | N |
| 10 | − | N | − | N | + | P | + | N |
| 11 | + | N | + | P | + | N | + | N |
| 12 | − | N | + | P | − | N | + | N |
| 13 | − | N | + | P | + | N | + | N |
| 14 | − | N | − | P | + | N | + | P |
| 15 | + | P | + | P | − | N | + | P |
| 16 | − | N | + | P | − | N | + | N |
| 17 | + | P | + | P | + | P | + | N |
| 18 | − | N | − | P | + | N | + | N |
| 19 | + | P | + | P | + | P | + | P |
| 20 | − | N | + | P | + | P | + | P |
| 21 | − | N | + | P | − | N | + | P |
| 22 | − | N | − | P | − | N | + | P |
| % Positive | 41 | 55a | 77 | 82a | 50 | 45a | 100 | 36a |
−, negative donor serology, +, positive donor serology, N, negative FASCIA response, P, positive FASCIA response.
Frequency‐positive serology corresponding with positive FASCIA response.
This article is being made freely available through PubMed Central as part of the COVID-19 public health emergency response. It can be used for unrestricted research re-use and analysis in any form or by any means with acknowledgement of the original source, for the duration of the public health emergency.
Next, we investigated whether donor T‐cell specificity to infectious agents could be protective for the recipient after stem cell transplantation. We analysed the FASCIA results for candida, adenovirus HSV, EBV, CMV or VZV and compared them to the patients’ clinical records 1‐year post‐transplant. Based on the individual patients’ infectious complications to candida, adenovirus HSV, EBV, CMV or VZV (Table 1, defined in the Material and Method section), the corresponding FASCIA results were divided into two groups, where one group represented patients without the aforementioned infectious complications (NI, n = 9) and the other group represented patients experiencing infectious complications (I, n = 12). When FASCIA data for individual infectious agents were compared between the NI or the I groups, there were no statistically significant differences (not shown). However, a composite analysis, grouping proliferative responses by the FASCIA method against all of the tested antigens, clearly showed a strong correlation between NI status and the presence of proliferative responses in the SCPs against multiple antigens (P‐value 0·007) (Fig. 1b). This indicated a transferred protective function. Furthermore, 1‐year post‐transplant, nine patients had experienced episodes of CMV‐reactivation and three patients of EBV‐reactivation (Table 1). The clinical data were compared to the corresponding FASCIA T‐cell response after CMV or EBV stimulation. In patients with reactivation of CMV infection, there was no positive FASCIA responses against CMV in any of the nine stem cell products. In patients that did not experience viral reactivation within 1 year post‐transplantation, 5 out of 12 (42%) showed a positive response to CMV (P‐value Fisher's exact test, 0·045). Of the three patients with EBV‐reactivation, two had donor products positive for EBV in FASCIA and one was negative, 14 out of 18 (78%) showed a positive response to EBV‐stimulation (Table 3).
Table 3.
Viral reactivation 1‐year post‐transplant
| No reactivation | Reactivation | P‐value | |
|---|---|---|---|
| CMV | |||
| Pos FASCIA | 5 | 0 | |
| Neg FASCIA | 7 | 9 | |
| % pos FASCIA | 42 | 0 | 0·045 |
| EBV | |||
| Pos FASCIA | 14 | 2 | |
| Neg FASCIA | 4 | 1 | |
| % pos FASCIA | 78 | 67 | ns |
ns, not significant.
This article is being made freely available through PubMed Central as part of the COVID-19 public health emergency response. It can be used for unrestricted research re-use and analysis in any form or by any means with acknowledgement of the original source, for the duration of the public health emergency.
Leucocyte content in the SCPs
In addition to CD34+ hematopoietic stem cells, the donor stem cell products contain other leucocytes 5. The cellular content in the stem cell products were assessed by flow cytometry and analysed using forward or side scatter properties or through antibodies against several lineage markers. The two patient groups, infection (I) and no infection (NI), were compared to evaluate whether the leucocyte composition in the stem cell products and T‐cell differentiation status were associated with infectious complications 1‐year post‐transplant. The frequency of granulocytes, monocytes, lymphocytes, T‐cells, NK‐cells, B‐cells or CD34+ stem cells in the products was similar between groups (Fig. 2a). Similarly, when T‐cells were further analysed in differentiated lineage subsets, no differences in the frequency of CD3+ TCRγδ+ or CD3+ TCR‐αβ+ T‐cell were detected between groups (Fig. 2b,c).
Figure 2.
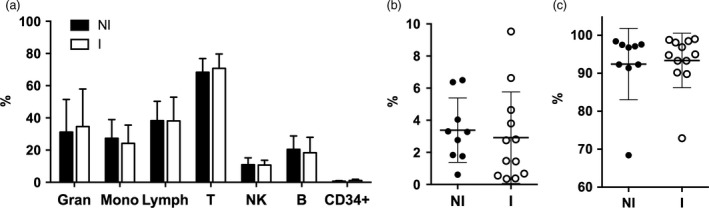
Leucocyte content in the stem cell products. Donor stem cell products were divided into two groups, infection (I, n = 12) and no infection (NI, n = 9) and the cellular content in the products were assessed by flow cytometry with subsequent analysis on forward or side scatter properties or through antibodies against lineage markers. (a) Frequency (%) of granulocytes, monocytes, lymphocytes and CD34‐positive stem cells out of total leucocytes as well as T‐cells, NK‐cells and B‐cells out of total lymphocytes. Frequency (%) of T‐cells expressing TCRγδ (b) or TCRαβ (c) out of total CD3+ cells. Mean and error bars, representing standard deviation, are depicted in all figures.
Cytokine production
We then investigated the cytokine/chemokine effector function, in response to various viral antigens, of cells in the FASCIA cultures. IL‐1β is mainly secreted by monocytes and macrophages of the innate immune system, while IFNγ, IL‐4, IL‐10, IL‐17 and TGFβ are secreted from different T‐cell subsets and represent important effector functions in adoptive immune responses against infectious pathogens 19, 20, 21. FASCIA culture supernatants from unstimulated cells or cells stimulated with PWM, CMV, EBV or HSV from all stem cell products were harvested on day 7 of incubation and analysed using a multiplex kit or ELISA. Again, the results were divided according to the patients’ infectious complications yielding two groups; patients with infectious complications (I) and patients without infection (NI). There was no difference between groups in concentration of IL‐4 or TGFβ in supernatants from cultures stimulated with EBV, CMV or HSV. Furthermore, the concentrations were low and there was no difference between background and stimulation with positive control PWM (not shown).
Compared to unstimulated FASCIA cultures PWM stimulated increased secretion of IL‐1β, IFNγ, IL‐10 and IL‐17 in all FASCIA products. There were no differences in secretion of IL‐4, IFNγ, IL‐10 and IL‐17 between the NI and the I groups. However, FASCIA supernatants from the NI group, stimulated with PWM (mean 94·2 ng/ml), contained significantly more IL‐1β (P‐value 0·012), compared to supernatants from the I group (mean 66·2 ng/ml). There were no differences between groups in secretion of IL‐4, IL‐10 or IL‐17 upon stimulation with EBV, CMV or HSV, nor did the concentration of IL‐1β and IFNγ differ between groups in cultures stimulated with EBV. Moreover, in CMV and HSV‐stimulated cell culture supernatants the concentration of IL‐1β was significantly higher (P‐values 0·022 and 0·026, respectively) in the SCP NI group (mean 23·2 ng/ml for CMV and 8·35 ng/ml for HSV), compared to supernatants from the infection group (mean 0·97 ng/ml for CMV and 1·0 pg/ml for HSV). A similar but not significant trend (P‐value 0·052) was seen for CMV‐stimulated IFNγ production, where the mean of for the non‐infection group was 10·1 ng/ml, whereas the mean concentration was 0·74 ng/ml for the infection group (Fig. 3a–c).
Figure 3.
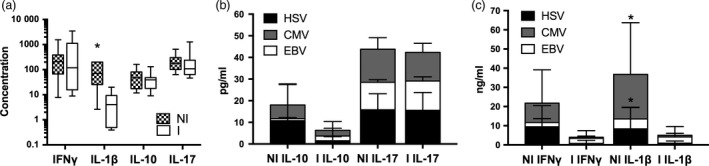
Cytokine secretion in FASCIA cultures stimulated with EBV, CMV, HCV or PWM. Cytokines were analysed at day 7 in FASCIA culture supernatants. Results were divided based on presence of infection (I, n = 12) or no infection (NI, n = 9) in patients at the 1‐year follow‐up. (a) Box and whiskers plot showing PWM stimulated concentration of IFNγ and IL‐1β (in ng/ml) as well as IL‐10 and IL‐17 (in pg/ml) for stem cell products in the non‐infection group (NI, checked boxes) and infection group (I, white boxes). Concentration of IL‐10 and IL‐17 (b) as well as IFNγ and IL‐1β (c) in supernatants from cultures stimulated with HSV (black bars), CMV (grey bars) or EBV (white bars) divided based on absence of infectious complications (NI) or infectious complications (I). *P‐value <0·05.
Discussion
Infectious complications due to poor T‐cell reconstitution or lymphocytopenia in general are major causes of morbidity and mortality after allogeneic HSCT 12. Here we evaluated whether antigen‐specific T‐cells in allogeneic stem cell grafts are able to reduce post‐transplant infectious complications up to 1 year after transplantation. Cells in stem cell products obtained from donors for allogeneic HSCT were stimulated with antigens associated with infectious complications post‐HSCT, that is, CMV, EBV, HSV and VZV, candida and adenovirus. Antigen‐specificity function was evaluated by means of T‐cell proliferation with a flow cytometry based assay 18, as well as by cytokine production. T‐cells from all stem cell products proliferated against the positive control, indicating that the T‐cells in the grafts were functional and viable pretransplant. The frequency of cell products with positive T‐cell responses to infectious antigens varied. Only about 25% showed a positive response against CMV and HSV, about 33% against VZV, while more than 80% of the stem cell products were positive against EBV. Taking donor age in consideration, these data reflect the seroprevalence in Sweden since the 1960s 22. Moreover, the measured FASCIA T‐cell responses against CMV, EBV, HSV and VZV were concurrent with donor serology for the same pathogens. Proliferative responses towards EBV‐antigens were detected in the vast majority of EBV sero‐positive donors (14/17) (Table 2). This is in agreement with previous studies showing that EBV specific T‐cells make up 0·5–1–5% of circulating T‐cells in EBV sero‐positive individuals 23, 24. A smaller fraction of CMV, HSV and VZV sero‐positive donors showed detectable proliferative T‐cell responses likely reflecting a greater inter‐individual variability in circulating antigen‐specific T‐cells for these pathogens. Furthermore, among the nine patients with CMV‐reactivation at the time of clinical follow‐up post‐transplant, there were no positive T‐cell responses measured by FASCIA against CMV in the corresponding stem cell products (Table 3). These data show an impending role for a pretransplant graft T‐cell proliferation assay in providing additional information regarding donor cells’ pathogen‐specific immunological competence. Six of the seven CMV sero‐positive patients receiving stem cells from CMV sero‐negative donors had experienced viral reactivation within 1‐year post‐transplant. This is in line with previous data showing that CMV seropositivity is an independent risk factor for CMV reactivation 25.
One‐year post‐HSCT the patients’ clinical records were assessed for infectious complications and the patients’ corresponding donor cells were divided into two groups. One group consisted of stem cell products transplanted to patients with infectious complications post‐transplantation (I) and those transplanted to patients without infectious complications (NI). When T‐cell responses to CMV, EBV, HSV and VZV, candida and adenovirus were analysed in composite and compared between these two groups, T‐cells from the group that was not associated with infectious complications post‐transplant proliferated more and to a wider range compared to T‐cells given to patients who experienced infectious complications. There was no difference in the concentration of IL‐4, IL‐10, IL‐17 or TGFβ between the two patient groups, suggesting that neither regulatory T‐cells, nor Th17 cells were involved in the protective effect seen in patients without infection at the 1‐year follow‐up.
However, IL‐1β and IFNγ production upon stimulation with CMV and HSV was higher in the group without infectious complications (NI) compared to the group with infectious complications (I), indicating that the NI donor products contained T‐cells with a pronounced ability to recognize and execute effector functions against the tested pathogens, compared to T‐cells in the other group (I). IL‐1β is a ‘master’ pro‐inflammatory cytokine of the IL‐1 cytokine family. It is important for autophagy and host defence against pathogens, but has also been shown to be involved in the pathogenesis of auto‐inflammatory diseases. Binding of IL‐1β to its receptor results in transcription of pro‐inflammatory genes and secretion of other IL‐1 cytokine family members, such as IL‐18 and IL‐36 26. Cytokines of the IL‐1 family induces secretion of IFNγ by T helper cells 27, 28. Moreover, it was recently shown that T‐helper cells induce optimal secretion of IFNγ through an autocrine IL‐1β activity 29. With respect to our data, this proposes that IL‐1β is also a master regulator for activation and proliferation of protective antigen‐specific T‐cells in an allogeneic transplantation setting. Additionally, allogeneic stimulation per se has been shown to be beneficial in an in vitro study demonstrating that stimulation with viral antigens, binding EBV‐ and CMV‐ specific T‐cells, in combination with allogeneic stimulation increased the number of antigen‐specific T‐cells and their effector function 30.
Complications after HSCT are multifactorial. Age, malignancy, pre‐HSCT treatment, conditioning, GVHD and T‐cells depletion strategies all affect T‐cell reconstitution and the patient's ability to fight infections 3. Phenotypic analysis of the donor stem cell products by flow cytometry showed that there was no difference in the frequency of granulocytes, monocytes, lymphocytes, T‐, NK‐, B‐cells, CD34+ cells or T‐cells expressing CD3+ TCRαβ+ or CD3+ TCRγδ+ between the two groups with or without infectious complications presented here (Fig. 2). Nor did the absolute mononuclear cell dose administered during transplantation differ between groups (Table 1). Yet, increased overall survival has been associated with a higher dose of transferred CD8+ T‐cells in stem cell products for allogeneic HSCT. The authors did not evaluate T‐cell clonal diversity in the stem cell products 31. In light of the results presented here, perhaps the improved clinical outcome is not due to the T‐cell dose per se, but rather an effect of the cell composition in regard to antigen‐specificity. It seems that donor grafts with a wider repertoire of antigen‐specific T‐cells against common post‐transplant infectious complications with a more pronounced cytokine production can be protective for the patient within 1‐year post‐transplantation and possibly longer. Donor‐derived CD4+ T‐cell clones, specific for tetanus toxin, have been shown to persist in the recipient up to 5 years after bone marrow transplantation 32.
The stem cell products in this study were not manipulated prior to transplantation. However, the vast majority of recipients receiving cells derived from peripheral blood were given antithymocyte immuno‐globuline (ATG) (15/16) a risk factor for post‐transplant infections 25, 33. Of the patients receiving ATG, seven belonged to the group without infectious complications and eight to the group with infectious complications, indicating that ATG administration as such did not determine the frequency of patients suffering from the tested infectious complications.
Our study indicates that monitoring of T‐cell functionality; against pathogens involved in transplant related infectious complications are of importance prior to allogeneic HSCT. Routinely assaying T‐cell function in allogeneic donor grafts against certain viruses and fungi causing illness after HSCT could assist gauging individual risks for post‐transplant infectious complications and aid in decision‐making regarding prophylactic and pre‐emptive anti‐infective therapy.
Acknowledgements
We thank lab technicians at the Stem cell laboratory unit at Karolinska university hospital in Stockholm as well as Hans Grönlund for technical assistance. The study was supported by Karolinska University Hospital and Swedish Foundation for Strategic Research. All authors contributed to either the planning of the study, the laboratory work or writing of the paper. There are no financial or commercial interests.
References
- 1. Pavletic SZ, Fowler DH: Are we making progress in GVHD prophylaxis and treatment? Hematology Am Soc Hematol Educ Program 2012; 2012:251–264 [DOI] [PubMed] [Google Scholar]
- 2. Scheper W, van Dorp S, Kersting S, et al: gammadeltaT cells elicited by CMV reactivation after allo‐SCT cross‐recognize CMV and leukemia. Leukemia 2013; 27:1328–1338 [DOI] [PubMed] [Google Scholar]
- 3. Welniak LA, Blazar BR, Murphy WJ: Immunobiology of allogeneic hematopoietic stem cell transplantation. Annu Rev Immunol 2007; 25:139–170 [DOI] [PubMed] [Google Scholar]
- 4. Wikell H, Ponandai‐Srinivasan S, Mattsson J, et al: Cord blood graft composition impacts the clinical outcome of allogeneic stem cell transplantation. Transpl Infect Dis 2014; 16:203–212 [DOI] [PubMed] [Google Scholar]
- 5. Watz E, Remberger M, Ringden O, et al: Quality of the hematopoietic stem cell graft affects the clinical outcome of allogeneic stem cell transplantation. Transfusion 2015; 55:2339–2350 [DOI] [PubMed] [Google Scholar]
- 6. Maury S, Mary JY, Rabian C, et al: Prolonged immune deficiency following allogeneic stem cell transplantation: risk factors and complications in adult patients. Br J Haematol 2001; 115:630–641 [DOI] [PubMed] [Google Scholar]
- 7. Radestad E, Wikell H, Engstrom M, et al: Alpha/beta T‐cell depleted grafts as an immunological booster to treat graft failure after hematopoietic stem cell transplantation with HLA‐matched related and unrelated donors. J Immunol Res 2014; 2014:578741 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Marek A, Stern M, Chalandon Y, et al: The impact of T‐cell depletion techniques on the outcome after haploidentical hematopoietic SCT. Bone Marrow Transplant 2014; 49:55–61 [DOI] [PubMed] [Google Scholar]
- 9. Minculescu L, Sengelov H: The role of gamma delta T cells in haematopoietic stem cell transplantation. Scand J Immunol 2015; 81:459–468 [DOI] [PubMed] [Google Scholar]
- 10. Airoldi I, Bertaina A, Prigione I, et al: gammadelta T‐cell reconstitution after HLA‐haploidentical hematopoietic transplantation depleted of TCR‐alphabeta+/CD19+ lymphocytes. Blood 2015; 125:2349–2358 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Shiley K, Blumberg E: Herpes viruses in transplant recipients: HSV, VZV, human herpes viruses, and EBV. Infect Dis Clin North Am 2010; 24:373–393 [DOI] [PubMed] [Google Scholar]
- 12. Arnaout K, Patel N, Jain M, et al: Complications of allogeneic hematopoietic stem cell transplantation. Cancer Invest 2014; 32:349–362 [DOI] [PubMed] [Google Scholar]
- 13. Lin R, Liu Q: Diagnosis and treatment of viral diseases in recipients of allogeneic hematopoietic stem cell transplantation. J Hematol Oncol 2013; 6:94 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Walter EA, Greenberg PD, Gilbert MJ, et al: Reconstitution of cellular immunity against cytomegalovirus in recipients of allogeneic bone marrow by transfer of T‐cell clones from the donor. N Engl J Med 1995; 333:1038–1044 [DOI] [PubMed] [Google Scholar]
- 15. Rooney CM, Smith CA, Ng CY, et al: Use of gene‐modified virus‐specific T lymphocytes to control Epstein‐Barr‐virus‐related lymphoproliferation. Lancet 1995; 345:9–13 [DOI] [PubMed] [Google Scholar]
- 16. Uhlin M, Gertow J, Uzunel M, et al: Rapid salvage treatment with virus‐specific T cells for therapy‐resistant disease. Clin Infect Dis 2012; 55:1064–1073 [DOI] [PubMed] [Google Scholar]
- 17. Einsele H, Roosnek E, Rufer N, et al: Infusion of cytomegalovirus (CMV)‐specific T cells for the treatment of CMV infection not responding to antiviral chemotherapy. Blood 2002; 99:3916–3922 [DOI] [PubMed] [Google Scholar]
- 18. Marits P, Wikstrom AC, Popadic D, et al: Evaluation of T and B lymphocyte function in clinical practice using a flow cytometry based proliferation assay. Clin Immunol 2014; 153:332–342 [DOI] [PubMed] [Google Scholar]
- 19. Akdis M, Burgler S, Crameri R, et al: Interleukins, from 1 to 37, and interferon‐gamma: receptors, functions, and roles in diseases. J Allergy Clin Immunol 2011; 127:701–721 e1–70 [DOI] [PubMed] [Google Scholar]
- 20. Marques RE, Guabiraba R, Russo RC, et al: Targeting CCL5 in inflammation. Expert Opin Ther Targets 2013; 17:1439–1460 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Dinarello CA: Biologic basis for interleukin‐1 in disease. Blood 1996; 87:2095–2147 [PubMed] [Google Scholar]
- 22. Svahn A, Berggren J, Parke A, et al: Changes in seroprevalence to four herpesviruses over 30 years in Swedish children aged 9–12 years. J Clin Virol 2006; 37:118–123 [DOI] [PubMed] [Google Scholar]
- 23. Kuzushima K, Hayashi N, Kudoh A, et al: Tetramer‐assisted identification and characterization of epitopes recognized by HLA A*2402‐restricted Epstein‐Barr virus‐specific CD8+ T cells. Blood 2003; 101:1460–1468 [DOI] [PubMed] [Google Scholar]
- 24. Kuzushima K, Hoshino Y, Fujii K, et al: Rapid determination of Epstein‐Barr virus‐specific CD8(+) T‐cell frequencies by flow cytometry. Blood 1999; 94:3094–3100 [PubMed] [Google Scholar]
- 25. Schonberger S, Meisel R, Adams O, et al: Prospective, comprehensive, and effective viral monitoring in children undergoing allogeneic hematopoietic stem cell transplantation. Biol Blood Marrow Transplant 2010; 16:1428–1435 [DOI] [PubMed] [Google Scholar]
- 26. Joosten LA, Netea MG, Dinarello CA: Interleukin‐1beta in innate inflammation, autophagy and immunity. Semin Immunol 2013; 25:416–424 [DOI] [PubMed] [Google Scholar]
- 27. Tominaga K, Yoshimoto T, Torigoe K, et al: IL‐12 synergizes with IL‐18 or IL‐1beta for IFN‐gamma production from human T cells. Int Immunol 2000; 12:151–160 [DOI] [PubMed] [Google Scholar]
- 28. Yoshimoto T, Takeda K, Tanaka T, et al: IL‐12 up‐regulates IL‐18 receptor expression on T cells, Th1 cells, and B cells: synergism with IL‐18 for IFN‐gamma production. J Immunol 1998; 161:3400–3407 [PubMed] [Google Scholar]
- 29. Arbore G, West EE, Spolski R, et al: T helper 1 immunity requires complement‐driven NLRP3 inflammasome activity in CD4(+) T cells. Science 2016; 352:aad1210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. D'Orsogna LJ, van den Heuvel H, van der Meer‐Prins EM, et al: Stimulation of human EBV‐ and CMV‐specific cytolytic effector function using allogeneic HLA molecules. J Immunol 2012; 189:4825–4831 [DOI] [PubMed] [Google Scholar]
- 31. Cao TM, Shizuru JA, Wong RM, et al: Engraftment and survival following reduced‐intensity allogeneic peripheral blood hematopoietic cell transplantation is affected by CD8+ T‐cell dose. Blood 2005; 105:2300–2306 [DOI] [PubMed] [Google Scholar]
- 32. Vavassori M, Maccario R, Moretta A, et al: Restricted TCR repertoire and long‐term persistence of donor‐derived antigen‐experienced CD4+ T cells in allogeneic bone marrow transplantation recipients. J Immunol 1996; 157:5739–5747 [PubMed] [Google Scholar]
- 33. Binkert L, Medinger M, Halter JP, et al: Lower dose anti‐thymocyte globulin for GvHD prophylaxis results in improved survival after allogeneic stem cell transplantation. Bone Marrow Transplant 2015; 50:1331–1336 [DOI] [PubMed] [Google Scholar]