

Abstract
Chemosensory epithelial cells (EpCs) are specialized cells that promote innate type 2 immunity and protective neurally-mediated reflexes in the airway. Their effector programs and modes of activation are not fully understood. Here we define the transcriptional signature of two choline acetyl transferase (ChAT)-expressing nasal EpC populations. They are found in the respiratory and olfactory mucosa, and express key chemosensory cell genes including the transcription factor Pou2f3, the cation channel Trpm5, and the type 2 cytokine Il25. Moreover, these cells share a core transcriptional signature with chemosensory cells from intestine, trachea and thymus, and cluster with tracheal brush cells (BrCs) independently from other respiratory EpC, indicating they are part of the brush/tuft cell family. Both nasal BrC subsets express high levels of transcripts encoding cysteinyl leukotriene (CysLT) biosynthetic enzymes. In response to ionophore, unfractionated nasal BrCs generate CysLTs at levels exceeding that of the adjacent hematopoietic cells isolated from naïve mucosa. Among activating receptors, BrCs express the purinergic receptor P2Y2. Accordingly, the epithelial stress signal ATP and aeroallergens that elicit ATP release trigger BrC CysLT generation, which is mediated by the P2Y2 receptor. ATP- and aeroallergen-elicited CysLT generation in the nasal lavage is reduced in mice lacking Pou2f3, a requisite transcription factor for BrC development. Finally, aeroallergen-induced airway eosinophilia is reduced in BrC-deficient mice. These results identify a previously undescribed BrC sensor and effector pathway leading to generation of lipid mediators in response to luminal signals. Further, they suggest that BrC sensing of local damage may provide an important sentinel immune function.
One Sentence Summary:
Aeroallergens and ATP elicit cysteinyl leukotrienes from nasal brush cells through the purinergic receptor P2Y2.
Introduction
Epithelial cells (EpCs) play a critical role in mucosal host defense through a diverse array of innate effector programs. In the airway, they produce interferons and antimicrobial peptides, generate proinflammatory cytokines, release mucins, and activate sensory neurons (1). Recent work in the human airway at the resolution of a single cell has demonstrated a remarkable subspecialization of these epithelial functions and defined novel cell types (2–4), but the pathways connecting EpC recognition of danger to specific effector functions are still poorly defined.
Chemosensory EpCs are detected in the airway, urinary and gut mucosa (5). They are referred to as tuft cells in the intestinal mucosa (6–8), brush cells (BrCs) in the trachea (9), and solitary chemosensory cells (SCCs) in the nasal respiratory mucosa (10). They were first characterized by their unique morphology – an amphora or teardrop shape and a “tuft” of short brush-like projections extending into the mucosal lumen (11, 12). Later studies defined some chemosensory cells by their expression of bitter taste receptors and proteins associated with the canonical taste transduction system, including the G protein α-gustducin (10, 13), phospholipase C-β (PLCβ) (9, 14, 15) and the calcium-activated cation channel transient receptor potential melastatin 5 (TRPM5) (14–17) . These cells are specified by the transcription factor Pou2f3 which is required for their development in the anterior nasal cavity (18), the trachea (19), and the small intestine (8). Microvillus cells (MVCs), a distinct EpC population detected in the mouse olfactory mucosa express TRPM5 (20–22) and phospholipase C-β (PLCβ) (23) and depend on Pou2f3 (24). Although MVCs are noted to express taste receptors and Gnat3 (encoding α-gustducin) at lower levels (23), an unsupervised analysis comparing this population to chemosensory EpC subsets has not been reported.
More recent work on this EpC family has elucidated shared functions. For example, chemosensory EpCs are the dominant source of IL-25 in the naïve airway and intestinal mucosa. In the intestine, tuft cell IL-25 generation activates IL-13-producing group 2 innate lymphoid cells (ILC2s) and elicits goblet cell metaplasia, which is essential for immunity to helminths and parasites (6–8, 25, 26). In the lung, BrC generation of IL-25 promotes eosinophilic type 2 inflammation in response to aeroallergens (27). Thus, chemosensory EpCs appear to play an essential upstream role in driving type 2 inflammation elicited by diverse pathogens across mucosal tissues.
In the airways, chemosensory EpCs are noted to have protective functions in some contexts. Tracheal BrCs and SCCs regulate protective respiratory reflexes such as apnea or sneezing in response to activation of canonical taste receptors by bitter compounds, odorants and bacterial quorum-sensing molecules (9, 28, 29). This protective function has been most closely linked to their generation of acetylcholine, leading to activation of peptidergic sensory nerve fibers (9, 28). MVCs also generate acetylcholine, but use it to modulate the activity of olfactory neurons and maintain homeostasis in the olfactory epithelium (22, 30, 31). Thus, this second effector pathway endows EpCs with a broader functional capacity to elicit immediate coordinated protective responses at an organismal level.
A common and essential feature of both chemosensory EpCs and MVCs is their ability to rapidly respond to relevant luminal stimuli with changes in intracellular calcium levels (14, 17, 22), a flux required for TRPM5 activation (32) and subsequent acetylcholine release (33). Notably, calcium flux is also essential for the generation of eicosanoids such as cysteinyl leukotrienes (CysLTs), potent proinflammatory lipids. These have been implicated as possible chemosensory EpC mediators based on the detection of high levels of transcripts encoding CysLT biosynthetic enzymes in intestinal tuft cells (15, 25, 34) and tracheal BrCs (2, 3, 27). Furthermore, we recently reported that CysLT generation in the respiratory tract occurs in concert with BrC activation (27), but whether BrCs actually generate CysLTs and the mechanism by which that may occur has not been explored.
CysLTs (LTC4, LTD4, and LTE4) are derived from arachidonic acid through the 5-lipoxygenase (5-LO) pathway. On cell activation, group IV cytosolic phospholipase A2 (cPLA2α) cleaves phospholipids at the outer nuclear membrane to generate free arachidonate. 5-LO then oxidizes arachidonic acid in the presence of 5-LO activating protein (FLAP) to generate leukotriene A4 (LTA4), which is subsequently converted to LTC4 by leukotriene C4 synthase (LTC4S)(35). After export from the cell, LTC4 is converted to LTD4 and LTE4 by γ-glutamyl transpeptidases and dipeptidases (36). Both cPLA2α and 5-LO require calcium for translocation to the nuclear membrane (37, 38). Accordingly, Syk-dependent leukocyte receptors that elicit sustained fluxes in intracellular calcium such as FcεR1 (39, 40), and the fungal receptors Dectin-1 (41) and Dectin-2 (42) elicit robust generation of CysLTs, whereas Toll-like receptor signaling does not (43).
While no EpC receptors are reported to mediate CysLT generation, we previously found that airway inhalation of the fungal species Alternaria alternata (Alternaria) elicits important CysLT-dependent epithelial defenses including the rapid release of goblet cell mucin in the nasal mucosa over minutes (44) and the expansion of IL-25-producing BrCs in the trachea over days (27). Thus, we hypothesized that airway BrCs may generate significant quantities of CysLTs and that we might use this function to further define the molecular signaling regulating BrC sensing.
Here we establish flow cytometric criteria to preferentially enrich for choline acetyl transferase positive MVCs or ChAT+ SCCs in the nasal mucosa, and perform transcriptional profiling to understand their sensing capacity and effector programs. As expected, each population expresses Pou2f3 and Trpm5. These nasal MVCs and SCCs are enriched for 64 or 63 of the 88 genes previously reported to be the transcriptional signature of brush/tuft cells (25), and each population clusters with tracheal BrCs independently from other airway EpCs, indicating that they are in the BrC family. We find that CysLT generating enzymes are among the most highly expressed transcripts that distinguish these airway BrC subsets from surrounding EpCs. Ex vivo stimulation of unfractionated nasal BrCs with calcium ionophore elicits robust generation of CysLTs that exceed the levels generated by adjacent CD45+ leukocytes. Furthermore, we find that the damage-associated molecules adenosine triphosphate (ATP) and uridine triphosphate (UTP) and complex aeroallergens, including the house dust mite Dermatophagoides pteronyssinus (Dp) and the mold Alternaria, elicit robust BrC generation of CysLTs. Inhibition of the nucleotide receptor P2Y2 and genetic deletion in P2ry2−/− mice abolished CysLT generation elicited by ATP. Remarkably, P2Y2 receptor inhibition also significantly reduced Alternaria-elicited CysLTs establishing P2Y2 as a novel BrC receptor involved in sensing a common environmental microbe. CysLT generation was absent in the nasal lavage of ATP-challenged P2ry2−/− mice and ATP- or aeroallergen-challenged Pou2f3−/− BrC-deficient mice, demonstrating that this novel BrC effector pathway can be elicited by aeroallergens and damage-associated molecular patterns in the airway. Finally, Alternaria-triggered airway eosinophilia at early and later timepoints was selectively reduced in BrC-deficient Pou2f3−/− mice, demonstrating the importance of BrCs in previously established models of innate and adaptive type 2 inflammation.
Results
Two populations of nasal ChAT-eGFP+ EpCs share the BrC signature
To define the shared and unique features of ChAT-enhanced green fluorescent protein positive (eGFP+) EpCs in the upper and lower airways, we first isolated ChAT-eGFP+ EpCs from the nasal mucosa of naïve ChAT(BAC)-eGFP mice (45) and compared them to tracheal BrCs from our previous study (27). Nasal ChAT-eGFP+ EpCs were similar to tracheal BrCs in their expression of high levels of EpCAM and ChAT-eGFP. However, in the nose, the ChAT-eGFP+ EpCs accounted for a much higher portion of EpCAM+ EpCs, at 4-5% of live CD45low/− cells and 2-3% of all live cells (Fig. 1A–B), compared to 0.5% as reported in the lower airways (9, 27) and intestine (6, 7, 25). Thus, >18,000 ChAT-eGFP+ EpCs could be recovered per mouse nose. Fluorescent-activated cell sorting (FACS) analysis of nasal ChAT-eGFP+ BrCs demonstrated a distinct profile. Only a small portion of ChAT-eGFP+ EpCs in the nose had high forward scatter (FSC) and high side scatter (SSC) characteristics similar to tracheal BrCs (27) (Fig. S1A). These FSChiSSChi cells accounted for 5% of ChAT-eGFP+ EpCs in the nose. The majority (95%) of nasal ChAT-eGFP+ EpCs were FSC low and SSC low suggesting they are smaller and agranular. ChAT-eGFP+ EpCs have been previously reported in two locations in the nasal mucosa. In the respiratory mucosa and vomeronasal organ, ChAT expressing EpCs are rare, large and morphologically complex cells with processes extending to both the luminal and basal surface of the epithelium and are referred to as SCCs (28, 46). By contrast, in the olfactory mucosa, ChAT-eGFP+ EpCs are smaller, more abundant, and have a more uniform structure and are termed MVCs (22). Thus, their relative frequency and flow cytometric parameters suggest that the FSChiSSChi ChAT-eGFP+ EpCs may be classical SCCs which derive from the respiratory mucosa, while the FSClowSSClow ChAT-eGFP+ EpCs may be MVCs which derive from the olfactory mucosa.
Fig. 1. Two populations of nasal ChAT-eGFP+ EpCs belong to the BrC/tuft cell family. (A-D).

EpCAM+CD45−/loweGFP+ and EpCAM+CD45−/loweGFP− EpCs were isolated by FACS sorting from naïve nasal mucosa of ChAT-eGFP mice. (A) FACS gating strategy demonstrating two populations of nasal ChAT-eGFP+ EpCs. (B) ChAT-eGFP+ EpCs recovered as a percent of all live cells (left) and total number recovered per mouse (middle). Frequency of FSChiSSChi and FSClowSSClow ChAT-eGFP+ EpCs as a percent of all ChAT-eGFP+ EpCs (right). (C, E). Normalized counts of the indicated genes derived from RNA-seq analysis using DeSeq2. (D) Hierarchical clustering of the top 50 most variably expressed genes among the ChAT-eGFP+ EpCs in the trachea and nose. (F) Whole mount of nasal septum of a ChAT-eGFP mouse, eGFP fluorescence was enhanced with an anti-eGFP antibody (green). (G) Whole nasal septum staining for DCLK1 (red), ChAT-eGFP (green) and EpCAM (grey). (H) Cross-section of the nasal cavity and staining of paraffin embedded slides for ChAT-eGFP (green) , Gα-gustducin (red) and Hoechst (blue).
To more clearly define these populations, we sorted each separately and compared them to tracheal ChAT-eGFP+ BrCs from our previously published RNA sequencing analysis (27). Like tracheal BrCs (also ChAT-eGFP+), both populations of nasal ChAT-eGFP+ cells express high levels of Chat, encoding choline acetyltransferase, Pou2f3, the transcription factor required for the development of chemosensory cells in the airway and intestine (8, 18, 19, 24), and Trpm5 encoding the calcium-activated, non-selective cation channel TRPM5 specific to taste-transducing cells (47). Other BrC-specific markers (Plcb2, Avil, Sox9) were also expressed in all airway ChAT-eGFP+ cells (Fig. 1C). To further define the possible functional heterogeneity between the three populations of airway ChAT-eGFP+ EpCs, we performed hierarchical clustering of the 50 most highly variable genes among nasal and tracheal ChAT-eGFP+ EpCs (Fig. 1D). This analysis suggested that tracheal BrCs were more similar to FSChiSSChi ChAT-eGFP+ EpCs than to FSClowSSClow ChAT-eGFP+ EpCs. Canonical taste receptors were more highly expressed on FSChiSSChi ChAT-eGFP+ cells and tracheal BrCs than on FSClowSSClow ChAT-eGFP+ cells but some were expressed in all subsets (Fig. 1D and Fig. S1B). On the other hand, the transcripts for the chemosensory cell-specific cytokine IL-25 and its receptor IL-17RB were more highly expressed on FSClowSSClow cells (Fig. 1D–E), suggesting differential capacity to respond to the environment. Interestingly, FSClowSSClow ChAT-eGFP+ EpCs differentially expressed the zinc factor transcription factor Egr2 involved in both immune and peripheral nervous cell maturation (48–50). Finally, in contrast to tracheal BrCs, both FSClowSSClow and FSChiSSChi ChAT-eGFP+ nasal EpCs express low levels of the Wnt pathway-associated stem cell G protein-coupled receptor (GPCR) Lgr5(51), suggesting that some of the genes marking these populations are specific to their location in the upper vs. lower airways. Notably, the chemosensory cell markers Gnat3, encoding the taste transduction protein Gα-gustducin and Dclk1, encoding the chemosensory cell-specific cytoskeletal protein DCLK1, were expressed in tracheal BrCs and FSChiFSChi ChAT-eGFP+ EpCs at much higher levels than in FSClowSSClow ChAT-eGFP+ EpCs (Fig. 1E). We also compared the two nasal ChAT-eGFP+ EpC populations to the shared brush/tuft cell signature of intestinal, tracheal, thymus, and gallbladder tuft cells (25). FSClowSSClow ChAT-eGFP+ EpCs demonstrated 2-fold enrichment for 64 out of the 88 genes identified as a pan-tuft cell signature, while FSChiSSChi ChAT-eGFP+ EpCs demonstrated 2-fold enrichment for 63 out of the 88 genes identified as pan-tuft cell signature (Fig. S1C) (25), indicating that both nasal ChAT-eGFP+ EpC populations were in the BrC/tuft cell family.
To confirm that our sorted FSClowSSClow and FSChiSSChi ChAT-eGFP+ EpCs represented the previously reported olfactory mucosal MVCs and respiratory mucosal SCCs respectively, we assessed them in situ. As expected, ChAT-eGFP+ BrCs were rare in the nasal respiratory epithelium, and highly concentrated in the main olfactory epithelium (Fig. 1F). Cells found in the two locations varied in size and morphology, with smaller uniform cells in the olfactory epithelium and larger morphologically heterogeneous BrCs in the respiratory epithelium, consistent with the numbers and FSC and SSC characteristics of our FSClowSSClow and FSChiFSChi ChAT-eGFP+ EpCs, respectively. To validate these assignments, we assessed DCLK1 and Gα-gustducin, each of which was expressed at higher levels in the transcriptome of FSChiSSChi ChAT-eGFP+ EpCs, compared to FSClowSSClow ChAT-eGFP+ EpCs. While ChAT-eGFP+ EpCs in the respiratory mucosa expressed DCLK1 (Fig. 1G) and Gα-gustducin (Fig. 1H), we did not detect either marker by histology in the ChAT-eGFP+ EpCs in the olfactory epithelium, consistent with previous reports (10, 21, 24, 52). This suggests that the rare, large FSChiSSChi ChAT-eGFP+ EpCs, expressing Dclk1 and Gnat3, correspond to the BrCs in the respiratory epithelium, while the abundant, small FSClowSSClow ChAT-eGFP+ EpCs correspond to the homogeneous smaller, DCLK1- and Gα-gustducin-negative BrCs in the olfactory epithelium. Hereafter we refer to them as respiratory and olfactory BrCs.
Thus, taken together we find that two subsets of nasal ChAT-eGFP+ EpCs are unique, but share a large number of genes with chemosensory EpCs in the trachea, intestine, gallbladder and thymus (25, 27). Both ChAT-expressing EpCs subsets are part of the BrC/tuft cell family. Notably, nasal respiratory and olfactory BrCs (corresponding to SCCs and MVCs, respectively) express both IL-25 and ChAT, indicating that they are each poised to generate diverse effector molecules and elicit both immune and neural effector functions. Furthermore, we find that ChAT-eGFP+ EpCs are sufficiently abundant in the naïve nasal mucosa of mice to allow for isolation and ex vivo functional experiments.
Nasal BrCs are the dominant source of CysLTs in the naïve nasal airway mucosa
To define the unique functions of airway BrCs, we next compared the transcriptional profile of nasal and tracheal EpCAM+eGFP+ BrCs to nasal and tracheal EpCAM+eGFP− EpCs. Principal component analysis demonstrated that the three populations of airway BrCs grouped quite distinctly from other EpCs but also separately from each other (Fig. 2A). The degree of similarity between tracheal BrCs and the two subsets of nasal BrCs was further highlighted by their closeness when we performed Euclidean distance measurements. This analysis demonstrates that the three subsets of BrCs are distinct but notably closer to each other than to either nasal or tracheal EpCs (Fig. 2B). To determine the features that distinguish airway BrCs from the rest of airway EpCs, we evaluated the 100 most highly variable genes, defining a core signature shared between tracheal, nasal respiratory and nasal olfactory BrCs (Fig. 2C). As expected, among the shared genes were the pan-tuft cell marker advillin (Avil), the tuft cell-specific cytokine Il25 and the taste transduction machinery genes Trpm5 and Gnat3, as well as the bitter taste receptor Tas2r108. The nasal EpCs were a mixture of cell types as evidenced by their high differential expression of genes in the ciliated, goblet, club and basal airway epithelial signature defined by single cell sequencing of mouse tracheas (Fig. S2A)(2). Notably, among the most highly differentially expressed genes conserved across all BrCs were transcripts encoding three proteins critical to CysLT biosynthesis – Alox5ap (5-LO activating protein, FLAP) Alox5 (5-LO), and Ltc4s (LTC4S) (Fig. 2C and S2B). Additionally, BrCs had conserved expression of genes linked to calcium signaling in the phosphatidylinositol – phospholipase signaling pathway. Among them was Pik3r5, encoding the regulatory subunit of phosphatidylinositol 3-kinase and Pik3cg, encoding phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit gamma. Additional signaling proteins that were highly expressed in airway BrCs included the genes related to chemosensory signaling Plcb2, Plcg2, and Itpr2 (Fig. S2C). The transcripts Alox5, Alox5ap and Ltc4s, as well as Pla2g4a (cPLA2α) were expressed at remarkably high levels in all airway BrCs (Fig. 2D). BrCs also expressed Abcc1, encoding multidrug resistance-associated transporter 1 that allows for LTC4 export to the extracellular space, where it can be rapidly converted to LTD4 and LTE4 (36). In contrast, Lta4h encoding the enzyme that converts LTA4 to LTB4 was not differentially expressed in BrCs. Collectively, this data suggests that airway BrCs are poised to respond to luminal signals with a robust calcium signaling and CysLT generation.
Fig. 2. Airway BrCs are potent producers of CysLTs.
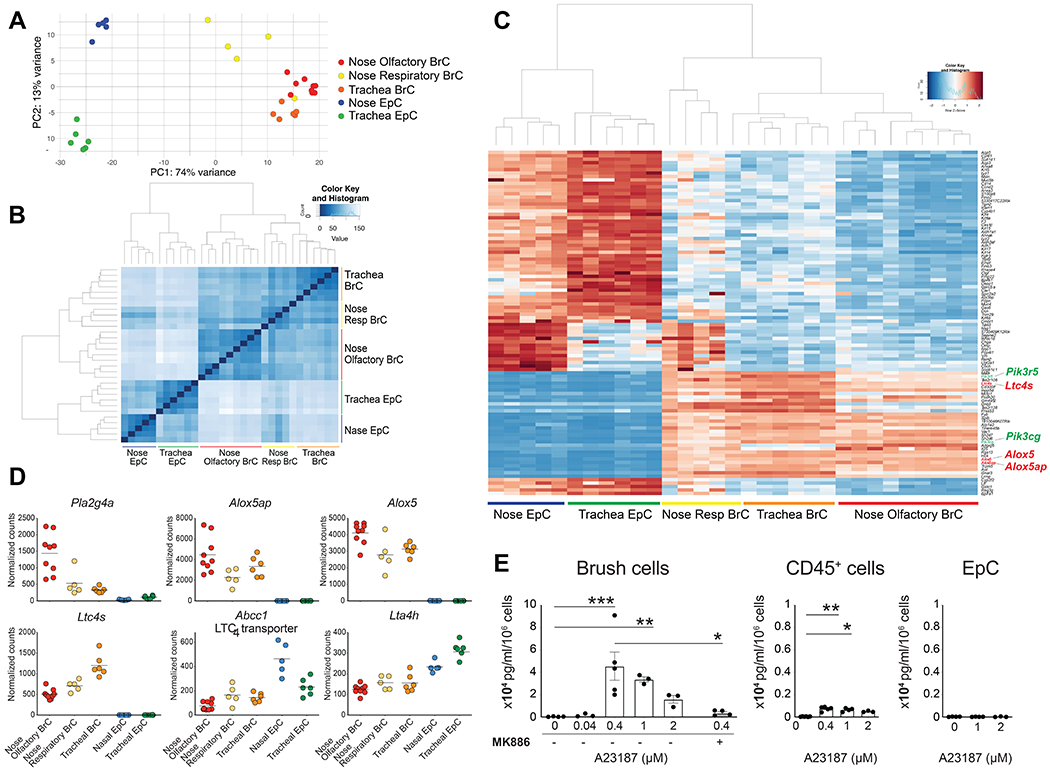
(A) Principal component analysis of ChAT-eGFP+ BrCs and ChAT-eGFP− EpCs from the nose and trachea using the top 100 most variable transcripts. Numbers indicate frequency of transcripts described by each principal component. (B) Euclidean distance matrix of the rlog transformed values derived from RNA sequencing. (C) Hierarchical clustering of the top 100 most variable genes derived from DeSeq2 analysis. Transcripts of enzymes in the CysLT biosynthetic pathway are highlighted in red. Transcripts of proteins in the phosphatidylinositol pathway are highlighted in green. (D) Normalized counts of transcripts encoding CysLT biosynthetic enzymes and transporters. (E) BrCs (EpCAM+CD45low/−eGFP+), CD45+ cells and EpCs (EpCAM+CD45−eGFP−) were isolated from the nasal mucosa and stimulated ex vivo with the indicated doses of calcium ionophore (A23187). Where indicated, cells were pre-treated for 15 min with the FLAP inhibitor MK-886. The concentration of CysLTs in the supernatants was measured by ELISA at 30 min. Data are means ± SEM, from at least three independent experiments, each dot represents a separate biological replicate, * p<0.05, ** p<0.01.
CysLT generation is best characterized in myeloid cells including mast cells, macrophages and dendritic cells (40, 42). To confirm that CysLTs are generated by BrCs at appreciable levels, we developed an ex vivo protocol for BrC stimulation. Olfactory and respiratory nasal BrCs were not distinguished in these experiments as each population expresses high levels of CysLT-generating enzymes and the respiratory BrCs are less abundant. We isolated nasal BrCs, rested them for 18 hours, stimulated them with calcium ionophore A23187, and compared their CysLT-generating capacity to that of sorted CD45+ hematopoietic cells and ChAT-eGFP− EpCs in the nasal mucosa. Isolated nasal BrCs stimulated with calcium ionophore generated high levels of CysLTs (Fig. 2E). MK-886, an inhibitor of FLAP, reduced CysLT generation, indicating activity of the conventional CysLT biosynthetic pathway. While ChAT-eGFP− nasal EpCs did not generate any appreciable levels of CysLTs (Fig. 2E), CD45+ hematopoietic cells isolated from the nasal mucosa of the same mice generated CysLTs, as expected (Fig. 2E). The pool included CD11c+MHC+ dendritic cells and CD11b+CD11c+SiglecF+ monocyte/macrophages (Fig. S3A). Among the other populations known to produce CysLTs, CD11b+CD11c−SiglecF+ eosinophils (Fig. S3A) and FcεR1+CD117+ mast cells (Fig. S3B) were extremely rare in the digested naïve nasal mucosa, consistent with our previous extensive histologic evaluation (44). Surprisingly, on a per cell basis, BrC CysLT generation exceeded that elicited from CD45+ cells even when normalized for cell count. Thus, although 10% of the CD45+ cell group was DCs, the level of CysLTs generated from the CD45+ pool was less than 10% of that generated by BrCs. To consider the potential contribution of BrCs to the total CysLT-generating capacity of the naïve nasal mucosa, we multiplied the CysLTs generated on a per cell basis by the number of cells recovered per mouse. Nasal BrCs accounted for >80% of the total CysLT-generating capacity of the naïve nasal mucosa (Fig. S3C), suggesting that this cell type is a significant source of CysLTs. Collectively, our data indicate that a core feature of airway BrCs is their ability to respond to activating stimuli with robust CysLT generation.
P2Y2 receptor activation elicits BrC generation of CysLTs
Having established a biologically relevant ex vivo assay for BrC function, we sought to identify the receptors that may trigger CysLT generation. We first focused on GPCRs, some of which are highly expressed in chemosensory EpCs and elicit calcium flux by coupling to Gq. Since olfactory BrCs account for >95% of BrCs in the nose and likely contribute to the bulk of CysLT production, we performed a pairwise comparison between olfactory BrCs and nasal EpCs across 632 GPCRs from the GPCR databank (https://gpcrdb.org). Several taste receptor genes including Tas1r3, Tas2r126, Tas2r108 and Tas2r138 were more highly expressed in respiratory and olfactory BrCs as compared to non-cholinergic airway EpC (Fig. 3A, Fig. S4A and S4B). Consistent with their closer relationship to tracheal BrCs, respiratory BrCs expressed a wider array of taste receptor transcripts at high levels (Fig. S4B). Notably, the transcript for the succinate receptor Sucnr1, an activating receptor on intestinal chemosensory tuft cells (25, 26, 53), was expressed in tracheal BrCs at much higher levels than nasal respiratory BrCs and was undetectable in nasal olfactory BrCs (Fig. S4C). Among the differentially expressed transcripts in olfactory BrCs was P2ry2, the gene encoding the Gq-coupled receptor P2Y2 (Fig. 3A). Further assessment of the purinergic receptor family demonstrated that P2Y2 and P2X4 receptors were highly expressed in all three airway BrC populations while the rest of the purinergic receptors were expressed at low or undetectable levels (Fig. 3B). Taken together, these results highlight the heterogeneity of activating receptors expressed by chemosensory EpCs that might reflect the local requirement for innate responses to pathogens, allergens and metabolites.
Fig. 3. P2Y2 receptor activation elicits CysLTs from airway BrCs.
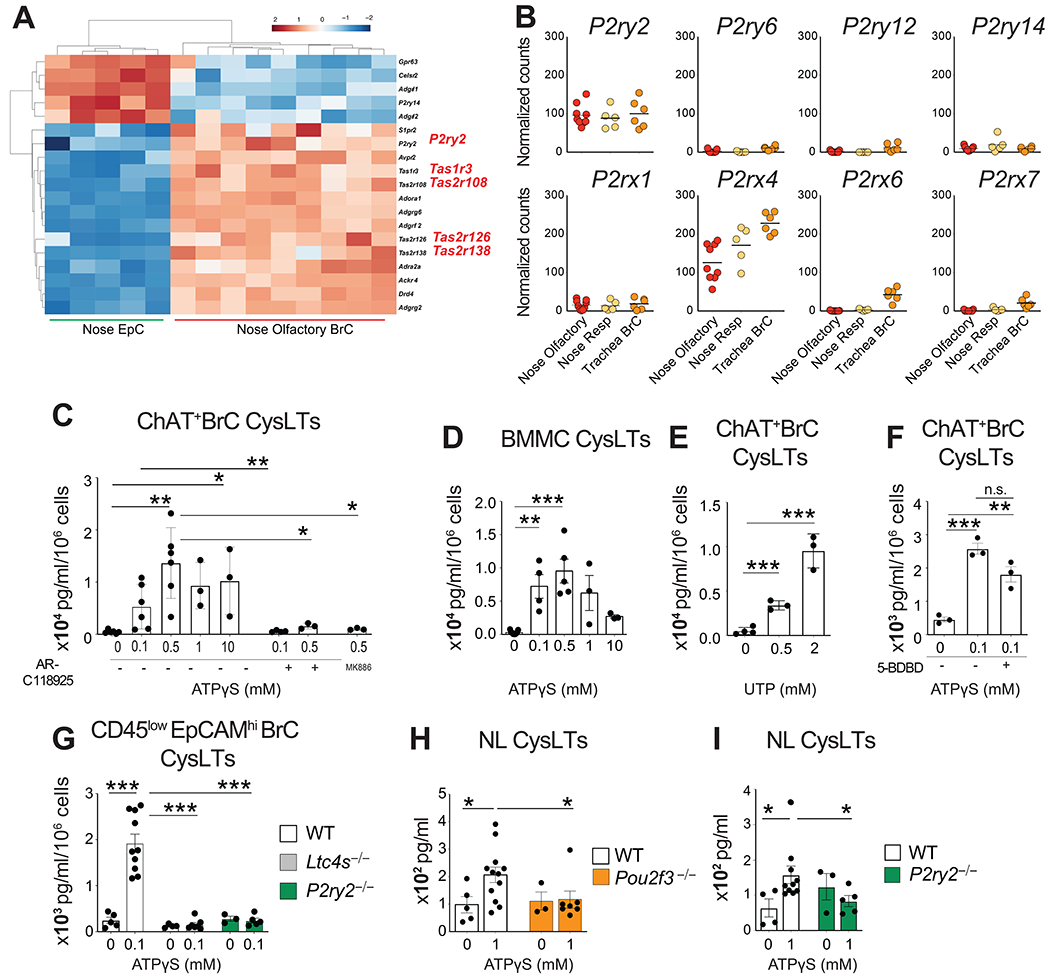
(A) Transcripts of differentially expressed GPCRs in olfactory nasal BrCs compared to nasal EpCs using criteria of log2 fold change of 2 or greater expression in one relative to the other with a p adjusted value of <0.01. (B) Transcripts for genes encoding purinergic receptors expressed at levels of 5 or above among nasal BrCs. (C, E) BrCs (EpCAM+CD45low/−eGFP+) were isolated from the nasal mucosa (nose) of ChAT-eGFP mice and were stimulated ex vivo with the indicated doses of ATPγS (C) or UTP (E) for 30 min. Where indicated, BrCs were pre-treated for 15 min with the P2RY2-specific inhibitor AR-C118925, the FLAP inhibitor MK-886 or HBSS and subsequently stimulated with ATPγS. (D) Mouse BMMCs were stimulated with ATPγS for 30 min. The concentrations of CysLTs in the supernatants were measured by enzyme immunoassay at 30 min. (F) ChAT-eGFP+ BrCs were pre-treated for 15 min with the P2X4-specific inhibitor 5-BDBD and subsequently stimulated with ATPγS. CysLTs were measured in the supernatant at 30 min. (G) CD45lowEpCAMhi BrCs from WT, Ltc4s−/− and P2ry2−/− mice were stimulated ex vivo with 0.1 mM of ATPγS. The concentration of CysLTs was measured by enzyme immunoassay. (H, I) ATP at the indicated dose was administered intranasally to naïve WT, Pou2f3−/− (H) and P2ry2−/− (I) mice and nasal lavage (NL) was obtained at 30 min. The concentration of CysLTs were measured by ELISA after acetone precipitation. Data are means ± SEM, each dot is a separate biological replicate, data are pooled from at least 2 independent experiments * p<0.05, ** p<0.01, *** p<0.001.
The purinergic receptors have both overlapping and distinct affinities for the nucleotides ATP, ADP and UTP (54–58). Both ATP and UTP are recognized by P2Y2 receptor, while the P2X4 receptor is selectively activated by ATP (59). To determine if the purinergic ligand ATP activates BrCs to produce CysLTs, we first stimulated BrCs isolated from ChAT-eGFP mice with increasing concentrations of ATP and measured CysLTs in the supernatant. The stable analogue ATPγS elicited CysLTs from BrCs in a dose-dependent fashion that was detected at 100 μM and peaked at 500 μM (Fig. 3C). This dose response and peak quantity of CysLT generation was similar to that of ATPγS-stimulated bone marrow-derived mast cells (BMMC) (Fig. 3D), which express several purinergic receptors (60, 61). CysLT generation was blunted in cells treated with the FLAP inhibitor MK-886, confirming the specificity of the assay (Fig. 3C). BrC viability both an hour after sorting and after overnight rest was preserved, as demonstrated by minimal lactate dehydrogenase (LDH) release (Fig. S4D). Consistent with that, ATP-mediated CysLT generation was not different in paired samples of BrCs stimulated with ATPγS at 3 h and 18 h after isolation (Fig. S4E).
Pre-treatment of BrCs with the P2Y2 receptor inhibitor AR-C118925 also significantly reduced BrC CysLTs elicited by both 100 and 500 μM of ATPγS (Fig. 3C), suggesting that P2Y2 is a BrC receptor mediating ATP-elicited generation of CysLTs. Accordingly, UTP stimulation of BrCs also triggered CysLT generation (Fig. 3E) and was inhibited by AR-C118925 (Fig. S4F). Whereas no ATPγS-elicited CysLT generation was detected in the presence of the P2Y2 receptor inhibitor AR-C118925 (Fig. 3C), there was significant induction of CysLT generation in the presence of the P2X4 receptor inhibitor 5-(3-bromophenyl)-1,3-dihydro-2H-benzofuro[3,2-e]-1,4-diazepin--2one (5-BDBD)(62) (Fig. 3F). Together, our pharmacologic studies strongly suggested that ATP is a potent signal for BrC activation for CysLT generation through the P2Y2 receptor.
To confirm the importance of P2Y2 receptor signaling in mice with genetic deletion of P2Y2, we first sought to identify BrCs in the naïve nasal mucosa of WT mice without fluorescent reporter expression. Two recent studies (2, 34) have noted that both intestinal tuft and tracheal brush cells express appreciable levels of Ptprc, the transcript encoding the hematopoetic receptor CD45. We found that Ptprc was also expressed in nasal BrCs (Fig. S5A) and confirmed that low levels of CD45 are expressed in nasal ChAT-eGFP+ EpCs by FACS (Fig. S5B). Backgating on CD45lowEpCAMhi cells in the ChAT-eGFP mice showed that this population was greatly enriched for BrCs, as 90% percent of this population was ChAT-eGPF+ (Fig. S5C). Notably however, this cell isolation strategy was not sensitive to capture all ChAT-eGFP+ cells, given the CD45 median fluorescence intensity (MFI), only 2 logs higher than the negative population. When compared to WT mice, the CD45lowEpCAMhi cells were nearly absent in Pou2f3−/− mice confirming that CD45lowEpCAMhi cells are dominantly BrCs (Fig. S5D). Pooled CD45+ cells and pooled EpCAM+ positive EpC numbers were unchanged in the Pou2f3−/− mice (Fig. S5E). We confirmed that these CD45lowEpCAMhi cells isolated from WT mice generate CysLTs in response to stimulation with ATP (Fig. S5F). When compared to CD45−EpCAMhi cells, CD45lowEpCAMhi cells generated higher levels of CysLTs on a per cell basis (Fig. S5F). The detection of CysLTs in the supernatants of CD45−EpCAMhi EpCs, a population of mixed CD45−BrCs and EpCs, suggests that both BrCs with higher and lower expression of CD45 are capable of generating CysLTs. We isolated CD45lowEpCAMhi BrCs from WT, Ltc4s−/− and P2ry2−/− mice and measured the levels of CysLTs in the supernatant after ex vivo ATPγS stimulation. CysLTs were detectable in the supernatants of ATPγS -stimulated CD45lowEpCAMhi BrCs from WT mice but were absent in those isolated from Ltc4s−/− mice confirming that CysLTs generated in response to ATPγS are a product of the classical CysLT biosynthetic cascade (Fig. 3G). Importantly, CysLT generation was absent in the supernatants of CD45lowEpCAMhi BrCs isolated from P2ry2−/− mice (Fig. 3G) defining P2Y2 as the specific BrC receptor triggered by ATP for CysLT generation.
To determine whether this alarmin recognition system that we characterized on isolated BrCs was operative in vivo, we instilled ATPγS intranasally and measured CysLTs in the acetone-precipitated nasal lavage 30 min later. ATP elicited dose-dependent CysLT generation in naïve WT mice (Fig. S4G). By contrast, there was no response to intranasal ATP in BrC-deficient Pou2f3−/− mice (Fig. 3H) or in mice lacking the P2Y2 receptor (Fig. 3I), indicating that airway BrCs respond to the alarmin ATP with robust generation of CysLTs in the nasal mucosa mediated through the P2Y2 receptor.
Aeroallergens elicit P2Y2-dependent BrC CysLT generation and BrC-dependent airway eosinophilia
Several common airborne aeroallergens have been reported to trigger the calcium-dependent exocytosis of ATP from human airway EpCs (63, 64). To determine whether aeroallergens trigger BrC CysLT generation and whether it may be mediated by P2Y2, we incubated BrCs ex vivo with increasing doses of the mold aeroallergen Alternaria or the house dust mite aeroallergen Dermatophagoides pteronyssinus (Dp). Both Alternaria and Dp elicited CysLTs from BrCs at doses of 400 and 800 μg/ml (Fig. 4A). CysLT generation from Alternaria- or Dp-stimulated CD45+ cells and Alternaria- stimulated EpCs isolated from the same mice was negligible (Fig. 4B, C). Consistent with that, CysLTs were absent in the supernatants of Alternaria-stimulated CD45lowEpCAMhi BrCs isolated from Ltc4s−/− mice (Fig. 4D), confirming the specificity of CysLT generation in response to this complex allergen extract. Pre-treatment of BrCs with the P2Y2 receptor inhibitor AR-C118925 significantly reduced Alternaria-elicited CysLTs in the supernatant (Fig. 4E). Thus, while ATP is a direct activator of P2Y2 (58), allergens such as Alternaria and Dp elicit CysLTs in part through the indirect release of ATP from BrCs (63). These results demonstrate that P2Y2 receptor-mediated CysLT generation is a BrC effector pathway relevant to airborne exposures.
Fig. 4. Allergen-elicited BrC activation triggers airway CysLT generation.
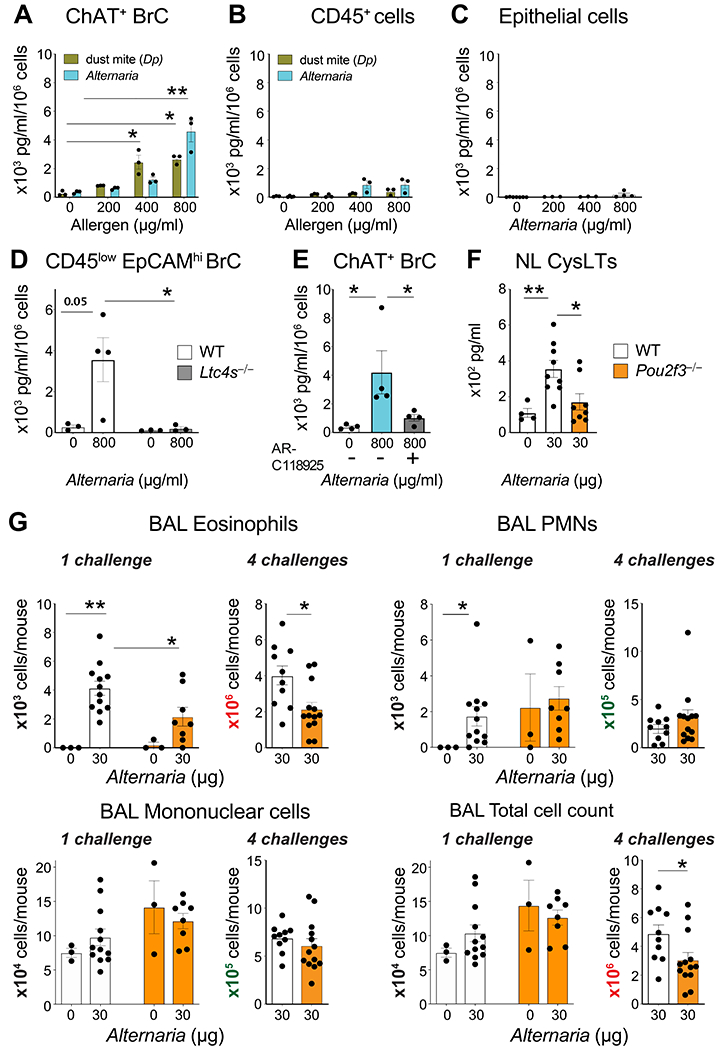
(A-C) ChAT-eGFP+ BrCs, CD45+ cells and EpC were isolated from the nasal mucosa of ChAT-eGFP mice and were stimulated ex vivo with the indicated doses of Alternaria or Dp. (D) CD45lowEpCAMhi BrCs from WT and Ltc4s−/− were stimulated ex vivo with Alternaria. (E) ChAT-eGFP+ BrCs were pre-treated with HBSS with or without the P2Y2 inhibitor AR-C118925 for 15 min and were subsequently stimulated with Alternaria. CysLTs were measured in the supernatant at 30 min by enzyme immunoassay. (F) Alternaria or HBSS was administered intranasally to naïve WT and Pou2f3−/− mice and nasal lavage (NL) was obtained at 30 min. The concentration of CysLTs were measured by ELISA after acetone precipitation. (G) WT and Pou2f3−/− mice were given one (1 challenge) or four (4 challenges) doses of Alternaria intranasally and the BAL lavage was evaluated 24 hour or 48 hours after the last challenge, respectively. The total number cells was counted and differential subsets were evaluated using Diff-Quick. Data are means ± SEM pooled from 3 independent experiments, each dot is a separate biological replicate, * p<0.05, **p<0.01.
To test the importance of this system in an integrated model, we challenged naïve mice with Alternaria intranasally and measured CysLTs in acetone-precipitated nasal lavage. Alternaria-elicited CysLT generation was detected in the lavage from WT mice but was significantly reduced in that from BrC-deficient Pou2f3−/− mice (Fig. 4F). These results define BrCs as a major source of IgE-independent airway CysLT generation triggered by aeroallergens (Fig. S6). Taken together, we identify CysLT generation as a third distinct BrC effector function and P2Y2 as a novel BrC sensor. Furthermore, our findings demonstrate a sentinel role for BrCs in immune recognition of aeroallergens and suggest that they play a likely role in recognition of additional luminal danger signals that elicit tissue damage.
Finally, to assess the potential importance of BrCs in airway inflammation, we evaluated the recruitment of inflammatory cells in the bronchoalveolar lavage (BAL) in two previously established models of Alternaria-elicited lung inflammation that rely on either innate immune pathways at 24 hours after a single inhalation or a mix of innate and adaptive pathways triggered by repetitive inhalation (27, 65–67). As expected, WT mice developed BAL eosinophilia 24 hours after a single Alternaria challenge and eosinophilia increased after 4 challenges over 9 days (Fig. 4G). BrC-deficient Pou2f3−/− mice had reduced BAL eosinophilia at each timepoint, but no reduction in neutrophilia or mononuclear cell infiltrate, pointing to a highly relevant role of BrCs in airway immunity to this complex fungal aeroallergen.
Discussion
BrCs are a unique family of chemosensory cells thus far known to participate in respiratory host defense through their generation of IL-25 and acetylcholine (6, 9, 27, 29). Several groups have reported a common transcriptional profile of chemosensory EpCs that includes eicosanoid pathway enzymes (15, 25), but the capacity of any chemosensory EpC to generate eicosanoids and the mechanism(s) that might mediate their generation had not been clarified. Here we find that two populations of nasal EpCs, residing in the olfactory and respiratory epithelium, share the BrC transcriptional signature including the expression of CysLT biosynthetic enzymes. We demonstrate that BrCs generate CysLTs in response to ATP and to aeroallergens that elicit ATP release (63). In each case, CysLT generation was dependent on P2Y2. Taken together, these findings suggest that CysLT generation is a common chemosensory cell effector mechanism. Finally, BrC deficiency results in reduced aeroallergen-induced eosinophilic airway inflammation implicating BrCs in innate defense mechanisms beyond the recognition of bacterial products.
Our findings provide the first functional evidence for BrC production of CysLTs. Although previous transcriptional studies have suggested that airway BrCs express the requisite enzymes of the CysLT generation cascade (2, 3, 25, 27), the capacity of BrCs to generate CysLTs at levels higher than tissue-resident leukocytes is surprising. On a per cell basis, nasal BrCs generated 10-fold more CysLTs than naïve tissue-resident hematopoietic cells. Furthermore, our in vivo data confirm that BrCs are the dominant source of both ATP- and aeroallergen-induced CysLTs released into the nasal airway lumen immediately after inhalation. While we did not directly compare BrCs to sorted nasal mast cells, we did find that ATP-elicited CysLTs in BrCs and BMMCs are comparable on a per cell basis, suggesting that BrCs are a potent and relevant source of CysLTs in the airways. Furthermore, both the current study and previous transcriptional analyses of BrCs and tuft cells from the airway and intestine respectively have identified the CysLT transcriptional machinery as part of the core profile of chemosensory epithelial cells (2, 3, 25, 27, 34). Thus, across morphologically and transcriptionally distinct chemosensory subsets, a defining feature is their potential to generate CysLTs, positioning them as key epithelial effectors.
CysLTs are recognized as effector molecules in established airway inflammation for their potent induction of airway vascular permeability and cellular chemotaxis (68, 69). Classically regarded as the products of activated leukocytes, CysLTs are produced in tissue-resident macrophages and mast cells after IgE receptor crosslinking (40). CysLTs are also generated by macrophages and mast cells on exposure to microbial stimuli such as IgG-opsonized microbes (70, 71), bacterial peptidoglycan (72), dust mites (42), zymosan (73), and other fungi (42). In human airways, CysLTs are generated not only in the setting of IgE cross-linking but also in response to respiratory viruses in exacerbations of asthma (74) and in infants without pre-existing lung disease (75–78). Collectively, these studies suggest that CysLT generation is a common response of innate immunity to respiratory pathogens. Our findings demonstrate that extracellular nucleotides in the airway also elicit CysLT generation, further highlighting the conserved nature of this innate pathway in the response to diverse respiratory insults.
While chemosensory EpCs in the airways and intestine share expression of the CysLT biosynthetic pathway, their activating receptors vary between compartments. Canonical taste receptors are more highly expressed in chemosensory tracheal BrCs and thymic chemosensory cells than in intestinal chemosensory tuft cells (9, 25, 27). Similarly, intestinal chemosensory cells and tracheal BrCs, but not those from the nose, express the SUCNR1 receptor, a receptor for the tricyclic acid metabolite succinate that is required for the type 2 immune intestinal response to protozoa (25, 26, 53). This heterogeneity suggests that their sensing apparatus might be modulated to respond to relevant local stimuli.
We found that all airway BrCs express high levels of two purinergic receptors – P2Y2 and P2X4, allowing them to rapidly respond to both luminal signals and EpC damage. Furthermore, we found that inhibition of P2Y2 reduced BrC CysLT generation elicited by ATP, by UTP and by Alternaria. Previous pharmacologic studies have demonstrated that both MVCs from the olfactory mucosa and SCCs from mouse and human respiratory mucosa are activated by ATP with increases in calcium flux (17, 31, 79), but an effector function and relevant receptor was not defined. Thus, our findings extend this area significantly by identifying P2Y2 as a novel BrC sensor for CysLT generation and demonstrating the relevance of this pathway in aeroallergen recognition (Fig. S6) (63). Notably, as ATP, CysLTs, and the EpC generation of IL-25 are all highly upregulated in the setting of respiratory viral infections (75, 77, 80–83), P2Y2-dependent BrC activation may play a greater role in airway sensing and immunity than is uncovered here.
Our unbiased transcriptional analysis of nasal cholinergic EpC subsets detected differences in the expression level of key genes between the two nasal BrC subsets including a number of taste receptors, taste receptor signal transcduction machinery (Gnat3) and the BrC/tuft cell marker DCLK1. However, unsupervised cluster analysis and Euclidean distance measurements demonstrate that olfactory BrCs/MVCs are closer to tracheal BrCs than to non-cholinergic EpCs in the nasal mucosa. As previously reported, they express ChAT (22, 28, 29), the cation channel TRPM5 (14, 20, 22, 84) and the transcription factor Pou2f3 (18, 19, 24). Furthermore, we find that both subsets express core BrC effector programs including Il25 and CysLT biosynthetic enzymes and are enriched for 73% of the genes reported as a tuft/BrC signature. Thus, we find that the repertoire olfactory BrCs/MVCs share with chemosensory EpCs extends beyond the previously reported generation of acetyl choline (28)(22) and points to shared effector functions of these cells and suggests that these cells should be included in the brush/tuft cell family.
Finally, our aeroallergen challenge data confirm a broader importance of BrCs in the airway inflammatory response to aeroallergens. Moreover, since the BrC population expands in the setting of aeroallergen challenge (27, 85) or respiratory viral infection (86), our data suggest that BrC-derived CysLTs might also contribute to the enhanced CysLT generating capacity of the airway in these settings and in diseases such as chronic rhinosinusitis and asthma.
Our study raises several questions that remain unanswered. We previously reported that the innate, Alternaria-elicited generation of nasal CysLTs mediates important airway effector functions including: 1) the activation of epithelial goblet cells for mucin release, 2) the dramatic dilatation of submucosal vessels, and 3) the degranulation of mast cells in the nasal submucosa (44). Our study demonstrates that MVCs, best characterized for their ability to modulate olfactory responses through calcium depolarization and acetylcholine release (30, 31), can generate CysLTs, but how BrC-derived CysLTs contribute to the function and integrity of the olfactory epithelium remains to be elucidated. Additionally, our findings that olfactory BrCs also express high levels of the IL-25 transcript suggests a possible previously unappreciated immune function for these cells. Finally, although our in vivo aeroallergen challenge data suggest that BrCs play an important role in aeroallegen-triggered airway inflammation, the specific contribution of BrC effector molecules (IL-25, CysLTs, prostaglandins) and their potential synergy remains to be elucidated.
Taste receptor signaling in airway BrCs triggers mast cell degranulation and vasodilatation, which depend on BrC activation of trigeminal peptidergic sensory nerves (28). However, whether CysLTs-dependent nasal mucosal defense depends on sensory neural signaling remains to be determined. Interestingly, CysLTs have been shown to directly activate and modulate the excitability of sensory neurons (87, 88) and several recent studies detected high levels of transcript for the type 2 CysLT receptor (CysLT2R) in sensory neurons in the vagal (88), dorsal root (89) and trigeminal ganglia (90, 91). Thus, future studies will be needed to determine whether the protective functions elicited by BrC-derived CysLTs might be transmitted through the nervous system.
Materials and Methods
Study Design:
The aim of this study was to define the capacity of airway BrCs to generate CysLTs and to define activating receptors on BrCs responsible for CysLT generation. We characterized the transcriptional profile of two populations of murine nasal BrCs compared to tracheal BrCs and other EpCs using low-input RNA-seq on sorted cells from a fluorescent BrC reporter mouse. Using an ex vivo stimulation system, we defined the CysLT generating capacity of nasal BrCs. Using receptor specific ligands, inhibitors and mice with a genetic deletion of the receptor in the ex vivo assay, we defined P2Y2 as an activating GPCR required for CysLT generation. Finally, the contribution of BrCs to the total CysLT generating capacity of the nasal mucosa was evaluated in an in vivo model of ATP and aeroallergen inhalation using a mouse strain lacking nasal BrCs.
Experimental Model and Mice
ChATBAC-eGFP (B6.Cg-Tg(RP23-268L19-EGFP)2Mik/J) mice were purchased from Jackson Laboratories (Bar Harbor, ME). Ltc4s‒/‒ mice were generated on a 129Sv background and backcrossed for 15 generations onto the C57BL/6 background (92). Pou2f3‒/‒ mice were generated as described previously (93) and backcrossed for more than 10 generations onto the C57BL/6 background (8, 24). P2ry2‒/‒ mice were described previously (94). Age- and sex-matched C57BL/6 mice bred in house (originally from Charles River Laboratories, Wilmington, MA) were used as controls. Mice were bred and housed in a specific pathogen free facility at the Brigham and Women’s Hospital; pups were weaned between 19-28 days after birth. Purchased mice were maintained for at least 4 weeks in the same mouse facility before being used for the experiments. Male and female mice 3-8 months of age were used. All mice were housed in groups of 4-5 mice per cage with a standard light/dark cycle of 12 h and provided food and water ad libitum. All experiments were performed during the day. The use of mice for these studies was in accordance with review and approval by the Animal Care and Use Committee of Brigham and Women’s Hospital.
Mouse snout isolation for histology and single cell preparations
Mouse snouts were harvested from euthanized mice. The lower jaw was dissected with scissors and the head was separated at the base of the skull. The skin and brain remnant were removed together with the connective tissue surrounding the snout. For histology, the entire snout was fixed in 4% paraformaldehyde (PFA) for at least 18 hours, changed to PBS and either immediately used for imaging of the nasal septum or decalcified using 14% (w/v) EDTA in NH4w–7.4) for 7–14 d to prepare histological sections.
Confocal microscopy of nasal septum
For confocal microscopy of the nasal septum, the nasal cavity of ChATBAC-eGFP mice was opened and the nasal septum was separated. The septum was then permeabilized in a PBS-based blocking buffer containing 0.1% Triton X, 0.1% saponin, 3% bovine serum albumin and 3% normal donkey serum for 3 hours (7). Primary goat anti-GFP (Abcam), primary rabbit anti-DCLK1 antibody (Abcam) and APC anti-mouse CD326 (EpCAM)(Biolegend) antibody were added directly to the blocking buffer for incubation at 4°C for 48-72 hours. The specimens were rinsed with PBS containing 0.1% Triton X for 3-4 hours and immunoreactivity to GFP and DCLK1 was detected with secondary donkey anti-goat secondary antibody, Alexa Fluor 488 (Life Technologies) and donkey anti-rabbit secondary antibody, Alexa Fluor 594 (Life Technologies), respectively, applied for 72 hours at 4 °C. Nuclear staining was performed with Hoechst 33342 nuclear stain (Sigma). The nasal septum was embedded using a glycerol-based cover slipping solution. Images were acquired at the Brigham and Women’s Hospital Confocal Microscopy Core Facility with a Zeiss LSM 800 with Airyscan confocal system on a Zeiss Axio Observer Z1 inverted microcope with a 10 x Zeiss, 0.30NA, 20 x Zeiss, 0.8NA and a 63 x Zeiss oil, 1.4NA objectives.
Histology of cross-sections
For immunofluorescence of tissue sections, the snouts were sectioned behind the incisors and in between the first three palatal ridges, yielding four coronal sections through the nasal cavity (44). The tissue was embedded in paraffin and tissue sections (6 μm) and paraffin-embedded sections were deparaffinized in xylene and rehydrated in ethanol solutions. Heat-induced epitope retrieval was performed in Target Retrieval solution (Dako). Slides were blocked in a PBS-based blocking with 0.1% Triton X-100, 0.1% saponin, 3% bovine serum albumin and 3% normal donkey serum for 1 hour. Samples were stained with polyclonal rabbit anti-mouse antibodies against Gα-gustducin (Santa Cruz) and a polyclonal goat anti-GFP antibody (Abcam) and a rabbit and goat polyclonal IgG control overnight at 4 oC. Immunoreactivity to GFP, DCLK1 and gustducin was detected with secondary donkey anti-goat secondary antibody, Alexa Fluor 488 (Life Technologies) and donkey anti-rabbit secondary antibody, Alexa Fluor 594 (Life Technologies), respectively, applied for 2 hours at room temperature. Nuclear staining was performed with Hoechst 33342 nuclear stain (Sigma).
Single cell preparations for ex vivo cell stimulation and RNA sequencing
For single-cell preparations, the ethmoid bone overlying the nasal cavity was removed to expose the nasal mucosa and the whole snout was incubated in pre-warmed PBS solution containing 16 U/ml dispase (Gibco) and 20 μg/mL DNase I (Sigma) for 30-40 min at room temperature with gentle shaking. The dispase activity was quenched by dilution with cold DMEM + 5% FCS. The mucosa of the whole nasal cavity was carefully scraped off with scalpel under a dissecting microscope. The epithelium together with the remainders of the nasal septum were incubated in HEPES-Tyrode’s buffer without calcium (Boston Bioproducts, Boston, MA) containing 20 μL/ml papain (28 units/mg, Sigma), 10 μl/ml L-cysteine (25 mg/ml Sigma) and 20 μg/mL DNase I (Sigma) for 30 min at 37 °C with gentle agitation. Papain digestion was terminated with Tyrode’s solution (Boston Bioproducts) with 2 μL/ml leupeptin (5 mg/mL; Sigma). The digested tissue was vortexed for 30 seconds and thoroughly triturated using a 21-gauge needle. The digested cells were passed through a 100 μm filter, washed once and stained using monoclonal antibodies against CD45 and EpCAM, followed by dead cell exclusion with propidium iodide before sorting. BrCs were identified as CD45low/−EpCAM+eGFP+ cells. Nasal EpCs were identified as CD45low/−EpCAM+eGFP− cells and CD45+ cells were separately sorted. For RNA sequencing experiments, the BrCs were subdivided into two populations: olfactory BrCs were defined as CD45low/−EpCAM+eGFP− FSClow SSClow BrCs, while respiratory BrC were defined as CD45low/−EpCAM+eGFP− FSChiSSChi (Fig. 1A). Sorted CD45+ cells were further characterized based on their expression of CD11b, CD11c, CD4, CD8, MHCII, kit and Fcεr1 (Fig. S3). In a separate set of experiments, BrCs were isolated from WT C57BL/6, Ltc4s−/− Pou2f3−/− and P2ry2−/− mice based on their high expression of EpCAM and low expression of CD45 (Fig. S5). Cells were sorted at the Brigham and Women’s Human Immunology Flow Core using a BD FACSAria Fusion cell sorter. For RNA sequencing, at least 300 but no more than 500 cells from 1 naïve digested mouse nose were sorted into TCL buffer (Qiagen) supplemented with RNAprotect Cell Reagent (Qiagen). Tracheal BrCs and tracheal EpCs were sorted as previously described (27).
Ex vivo stimulation of nasal BrCs
Sorted EpC and BrC were plated in DMEM-based EpC proliferation media which includes: DMEM/F-12, 5% fetal bovine serum, 10 mM HEPES, 100 μg/mL Penicillin-Streptomycin, 10 μg/mL insulin, 5 μg/mL transferrin, 25 ng/mL epithermal growth factor, 0.1 μg/mL cholera toxin, 30 μg/mL bovine pituitary extract, 10 μg/mL BSA ,100 nM retinoic acid and 0.25 μg/mL amphotericin B (95). Sorted CD45+ cells were reconstituted in R10 media (RPMI medium 1640 containing 10% (vol/vol) FBS, 2 mM L-glutamine, 0.1 mM nonessential amino acids, penicillin (100 units/ mL), streptomycin (100 μg/mL) and 1% culture supernatant from Chinese hamster ovary cells expressing mouse interleukin-3(44). The cells were plated into 96-well culture plates at a concentration of 400,000 cells/ml in 50 μL of media per well and rested for 18 hours. The cells were stimulated with Alternaria alternata (lot# 151774) (200-800 μg/ml), Dermatophagoides pteronyssinus (lot# 45415) (200-800μg/ml) (both from Greer Laboratories, Lenoir, NC), A23187 (0.1-10 mM, Cayman), ATPγS (0.1-10mM, Abcam), or UTP (0.5-2 mM, Sigma) in 50 μl of HBSS with calcium and magnesium for 30 min. For inhibition experiments, the cells were pre-treated with HBSS with calcium and magnesium with or without the FLAP inhibitor MK-886 (1 μM, Cayman), P2Y2 receptor inhibitor AR-C 118925XX (10 μM, Tocris) or P2X4 receptor inhibitor 5-BDBD (10 μM, Tocris) for 15 mins before the stimulus was added. After stimulation, the reaction was stopped by centrifugation at 350 × g for 5 min at 4 °C, and the supernatants were retained for assays of CysLTs.
LDH release assay
Isolated BrCs were incubated in 96-well plates in 100 µL DMEM-based EpC proliferation media at a concentration of 14,000 to 15,000 cells per well for 1 or 18 hours. Both DMEM without additives and DMEM-based EpC proliferation media containing serum were used as controls to account for background due to FBS-related LDH activity (96). LDH release was measured using LDH Cytotoxicity Assay Kit (ThermoFisher) by adding either 10 μL of Lysis Buffer or 10 μL of sterile, ultrapure water to wells containing BrCs. The plate was further incubated at 37°C, 5% CO2 for 45 minutes. 50 µL of each sample, serum-free medium and proliferation medium were transferred to a new plate and mixed with 50 µL of Reaction Mixture. After 30 min of incubation at room temperature protected from light, the reaction was stopped by adding 50 µL of Stop Solution. The absorbance was measured at 490 nm and 680 nm, with 490 nm representing LDH-induced red formazan conversion and 680 nm standing for machine background. The results of the LDH assay are presented as OD differences between 490 nm and 680 nm measurements.
In vivo CysLT generation assay
Mice received a single intranasal (i.n.) application of Alternaria alternata culture filtrate, or ATPγS after anesthesia with an intraperitoneal injection of ketamine (10 mg/kg) and xylazine (20 mg/kg) for full sedation. The aeroallergen extract and ATPγS were delivered in a total volume of 20 μl of sterile phosphate-buffered saline (PBS). Mice were euthanized 30 min after the i.n. challenge with isoflurane overdose, and nasal lavage was performed with 200 μl of PBS through a tracheal opening. The nasal lavage fluid protein was precipitated with ice cold acetone and after high speed centrifugation, the lipid fraction was retrieved for CysLT measurements. Mice of a given genotype were randomized to treatment dose, and challenges were performed in groups organized by genotype and treatment dose.
Aeroallergen challenge for BAL fluid analysis
Mice received 30 μg of Alternaria or PBS i.n. once for an analysis of BAL fluid 24 hours later or four times over 9 days for BAL fluid analysis 48 hours after the last challenge. After euthanasia, the trachea was cannulated and BAL was obtained by three repeated lavages with 0.75 ml of PBS with 1 mM EDTA. The BAL fluid was centrifuged at 500x g for 5 min. Cells were resuspended in 0.2 ml of PBS, and the total cells were counted manually with a hemocytometer. For the differential cell counts of macrophages, neutrophils, and eosinophils, the cells were cytospun onto a glass slide and stained with Diff-Quik. Cell types in a total of 200 cells were identified by morphologic criteria (97).
BMMC culture and stimulation
BMMCs were generated as previously described (92). In brief, bone marrow was collected from femurs and tibiae of mice and cultured for 4-6 wk in R10 media described previously. The culture medium for the BMMC was changed every week, and the cell density was adjusted to 3×105/mL at every passage. After 4 weeks, more than 97% of the cells were BMMCs as assessed by staining with Wright–Giemsa and toluidine blue.
BMMCs were harvested from culture and plated at a concentration of 400,000 cells/ml in 50 μL of R10 media per well and rested for 2 hours. The cells were stimulated with ATPγS (0.1–10mM, Abcam) in 50 μl of HBSS with calcium and magnesium for 30 min, centrifuged at 350 × g for 5 min at 4 °C and the supernatants were retained for ELISA assays to measure CysLT generation.
CysLT detection
CysLT generation in the supernatants of BrCs, EpCs, CD45+ sorted cells, cultured BMMCs and acetone-precipitated nasal lavage fluid was measured by a commercially available ELISA according to the manufacturer’s protocol (Cayman) based on competition between CysLTs and CysLT-acetylcholinesterase conjugate for a limited amount of CysLT ELISA Monoclonal Antibody. The lower limit of detection was 60 pg/mL and the following reported reactivity: Leukotriene C4 (100%), Leukotriene D4 (100%), Leukotriene E4 (79%), 5,6-DiHETE (3.7%), Leukotriene B4 (1.3%), 5(S)-HETE (0.04%), Arachidonic Acid (<0.01%).
Low input RNA sequencing
Cells from nasal single cell suspensions were processed at the Broad Institute Technology Labs using low-input eukaryotic Smart-seq 2. Briefly, Smart-seq2 libraries were sequenced on an Illumina NextSeq500 using a High Output kit to generate 2 × 25 bp reads (plus dual index reads). Each sample’s sequences were aligned utilizing STAR v2.4.2a (98) and the following parameters: --twopassMode Basic, -- alignIntronMax 1000000, --alignMatesGapMax 1000000, --sjdbScore 2, --quantMode TranscriptomeSAM, --sjdbOverhang 24. Alignments were processed using Picard v1.1073 tools to add read groups and other sequencing information, reorder the BAM file to the reference, and mark duplicate sequences. The data was assessed for quality using RNA-SeQC (99). RNA sequencing results were analyzed using R Bioconductor. Raw counts were generated using the FeatureCounts function of Rsubread (100). Raw input was used for normalization and differential expression analysis by DeSeq2 software. STAR aggregated mappings on a per-gene basis were used as raw input for normalization by DESeq2 software (101). Any genes with a mean normalized read count < 10 in any BrC subset were removed from analysis. Genes were considered to be significantly differentially regulated based on a fold change in expression of > 2-fold and a false discovery rate of <0.05 based on a Benjamini-Hochberg adjusted p-value to correct for multiple comparisons. Principal component analysis was conducted using the top 100 differentially regulated genes by false discovery rate.
Statistics
Analysis was performed with GraphPad Prism software (version 8, GraphPad, La Jolla, CA). Nonparametric two-sided Mann–Whitney and unpaired T tests were used to determine significance in pairwise comparison of responses in the in vivo and ex vivo stimulation models. For dose responses, the overall significance was determined using a one-way ANOVA or Kruskall-Wallis tests and pairwise multiple comparison was performed with Dunn’s tests. A value of p <0.05 was considered significant. Sample sizes were not predetermined by statistical methods.
Supplementary Material
Table S2. Raw data in Excel file.
Fig. S1. Nasal ChAT-eGFP+ cells share the tuft cell signature.
Fig. S2. Nasal BrCs are enriched for phosphatidylinositol – phospholipase signaling and CysLT biosynthetic pathway transcripts.
Fig. S3. BrCs are the dominant source of CysLTs in the naïve nasal mucosa.
Fig. S4. Taste receptors are highly expressed in tracheal and respiratory nasal but not olfactory nasal BrCs.
Fig. S5. CD45 is expressed on nasal BrCs
Fig. S6. Aeroallergen-triggered pathways of BrC activation.
Table S1. Reagents.
Acknowledgements:
We thank Adam Chicoine (Brigham and Women’s Hospital Human Immunology Center Flow Core) for his help with FACS sorting.
Funding: This work was supported by NIH Grants R01 HL120952 and R01 AI134989 (to N.A.B.), U19 AI095219 (to N.A.B., Y.K., L.G.B), by R01 HD098363 (to W.J.), by R01 DC017503 (to I.M.), K08 AI132723 (to L.G.B), by the AAAAI/ALA Respiratory Disease Award (to N.A.B.), by the AAAAI Foundation Faculty Development Award (to L.G.B.), by the Steven and Judy Kaye Young Innovators Award (to N.A.B.), by the Department of Defense Investigator-Initiated Research Award W81XWH-17-1-0527 (to Y.K. and N.A.B.), by the Joycelyn C. Austen Fund for Career Development of Women Physician Scientists (to L.G.B.), and by a generous donation by the Vinik family (to L.G.B.).
Footnotes
Competing interests:
The authors declare that they have no competing interests.
Data and Materials Availability:
The RNA sequencing data for this study have been deposited to the NCBI Gene Expression Omnibus and are accessible under accession number GSE139014. All other data needed to evaluate the conclusions in the paper are present in the paper or the Supplementary Materials. Skn-1a/Pou2f3 knockout mice are available from Dr. Matsumoto through a Material Transfer Agreement with Monell Chemical Senses Center.
References:
- 1.Lambrecht BN, Hammad H, Allergens and the airway epithelium response: gateway to allergic sensitization. The Journal of allergy and clinical immunology 134, 499–507 (2014). [DOI] [PubMed] [Google Scholar]
- 2.Montoro DT et al. , A revised airway epithelial hierarchy includes CFTR-expressing ionocytes. Nature 560, 319–324 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Plasschaert LW et al. , A single-cell atlas of the airway epithelium reveals the CFTR-rich pulmonary ionocyte. Nature 560, 377–381 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ordovas-Montanes J et al. , Allergic inflammatory memory in human respiratory epithelial progenitor cells. Nature 560, 649–654 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Krasteva G, Kummer W, “Tasting” the airway lining fluid. Histochem Cell Biol 138, 365–383 (2012). [DOI] [PubMed] [Google Scholar]
- 6.von Moltke J, Ji M, Liang HE, Locksley RM, Tuft-cell-derived IL-25 regulates an intestinal ILC2-epithelial response circuit. Nature 529, 221–225 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Howitt MR et al. , Tuft cells, taste-chemosensory cells, orchestrate parasite type 2 immunity in the gut. Science 351, 1329–1333 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gerbe F et al. , Intestinal epithelial tuft cells initiate type 2 mucosal immunity to helminth parasites. Nature 529, 226–230 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Krasteva G et al. , Cholinergic chemosensory cells in the trachea regulate breathing. Proceedings of the National Academy of Sciences of the United States of America 108, 9478–9483 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Finger TE et al. , Solitary chemoreceptor cells in the nasal cavity serve as sentinels of respiration. Proceedings of the National Academy of Sciences of the United States of America 100, 8981–8986 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.DiMaio MF, Kattan M, Ciurea D, Gil J, Dische R, Brush cells in the human fetal trachea. Pediatric pulmonology 8, 40–44 (1990). [DOI] [PubMed] [Google Scholar]
- 12.Reid L et al. , The mysterious pulmonary brush cell: a cell in search of a function. American journal of respiratory and critical care medicine 172, 136–139 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ohmoto M, Matsumoto I, Yasuoka A, Yoshihara Y, Abe K, Genetic tracing of the gustatory and trigeminal neural pathways originating from T1R3-expressing taste receptor cells and solitary chemoreceptor cells. Mol Cell Neurosci 38, 505–517 (2008). [DOI] [PubMed] [Google Scholar]
- 14.Lin W, Ogura T, Margolskee RF, Finger TE, Restrepo D, TRPM5-expressing solitary chemosensory cells respond to odorous irritants. J Neurophysiol 99, 1451–1460 (2008). [DOI] [PubMed] [Google Scholar]
- 15.Bezencon C et al. , Murine intestinal cells expressing Trpm5 are mostly brush cells and express markers of neuronal and inflammatory cells. J Comp Neurol 509, 514–525 (2008). [DOI] [PubMed] [Google Scholar]
- 16.Kaske S et al. , TRPM5, a taste-signaling transient receptor potential ion-channel, is a ubiquitous signaling component in chemosensory cells. BMC Neurosci 8, 49 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gulbransen BD, Clapp TR, Finger TE, Kinnamon SC, Nasal solitary chemoreceptor cell responses to bitter and trigeminal stimulants in vitro. J Neurophysiol 99, 2929–2937 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ohmoto M et al. , Pou2f3/Skn-1a is necessary for the generation or differentiation of solitary chemosensory cells in the anterior nasal cavity. Biosci Biotechnol Biochem 77, 2154–2156 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yamashita J, Ohmoto M, Yamaguchi T, Matsumoto I, Hirota J, Skn-1a/Pou2f3 functions as a master regulator to generate Trpm5-expressing chemosensory cells in mice. PloS one 12, e0189340 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hansen A, Finger TE, Is TrpM5 a reliable marker for chemosensory cells? Multiple types of microvillous cells in the main olfactory epithelium of mice. BMC Neurosci 9, 115 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lin W, Ezekwe EA Jr., Zhao Z, Liman ER, Restrepo D, TRPM5-expressing microvillous cells in the main olfactory epithelium. BMC Neurosci 9, 114 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ogura T et al. , Cholinergic microvillous cells in the mouse main olfactory epithelium and effect of acetylcholine on olfactory sensory neurons and supporting cells. J Neurophysiol 106, 1274–1287 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Genovese F, Tizzano M, Microvillous cells in the olfactory epithelium express elements of the solitary chemosensory cell transduction signaling cascade. PloS one 13, e0202754 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Yamaguchi T et al. , Skn-1a/Pou2f3 is required for the generation of Trpm5-expressing microvillous cells in the mouse main olfactory epithelium. BMC Neurosci 15, 13 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nadjsombati MS et al. , Detection of Succinate by Intestinal Tuft Cells Triggers a Type 2 Innate Immune Circuit. Immunity 49, 33–41 e37 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Schneider C et al. , A Metabolite-Triggered Tuft Cell-ILC2 Circuit Drives Small Intestinal Remodeling. Cell 174, 271–284 e214 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bankova LG et al. , The cysteinyl leukotriene 3 receptor regulates expansion of IL-25-producing airway brush cells leading to type 2 inflammation. Science Immunology 3, (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Saunders CJ, Christensen M, Finger TE, Tizzano M, Cholinergic neurotransmission links solitary chemosensory cells to nasal inflammation. Proceedings of the National Academy of Sciences of the United States of America 111, 6075–6080 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Krasteva G, Canning BJ, Papadakis T, Kummer W, Cholinergic brush cells in the trachea mediate respiratory responses to quorum sensing molecules. Life Sci 91, 992–996 (2012). [DOI] [PubMed] [Google Scholar]
- 30.Lemons K et al. , Lack of TRPM5-Expressing Microvillous Cells in Mouse Main Olfactory Epithelium Leads to Impaired Odor-Evoked Responses and Olfactory-Guided Behavior in a Challenging Chemical Environment. eNeuro 4, (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Fu Z, Ogura T, Luo W, Lin W, ATP and Odor Mixture Activate TRPM5-Expressing Microvillous Cells and Potentially Induce Acetylcholine Release to Enhance Supporting Cell Endocytosis in Mouse Main Olfactory Epithelium. Front Cell Neurosci 12, 71 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Prawitt D et al. , TRPM5 is a transient Ca2+-activated cation channel responding to rapid changes in [Ca2+]i. Proceedings of the National Academy of Sciences of the United States of America 100, 15166–15171 (2003). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yau WM, Dorsett JA, Youther ML, Calcium-dependent stimulation of acetylcholine release by substance P and vasoactive intestinal polypeptide. Eur J Pharmacol 120, 241–243 (1986). [DOI] [PubMed] [Google Scholar]
- 34.Haber AL et al. , A single-cell survey of the small intestinal epithelium. Nature 551, 333–339 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Yoshimoto T, Soberman RJ, Spur B, Austen KF, Properties of highly purified leukotriene C4 synthase of guinea pig lung. The Journal of clinical investigation 81, 866–871 (1988). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lam BK, Owen WF Jr., Austen KF, Soberman RJ, The identification of a distinct export step following the biosynthesis of leukotriene C4 by human eosinophils. The Journal of biological chemistry 264, 12885–12889 (1989). [PubMed] [Google Scholar]
- 37.Rouzer CA, Samuelsson B, Reversible, calcium-dependent membrane association of human leukocyte 5-lipoxygenase. Proceedings of the National Academy of Sciences of the United States of America 84, 7393–7397 (1987). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Nalefski EA et al. , Delineation of two functionally distinct domains of cytosolic phospholipase A2, a regulatory Ca(2+)-dependent lipid-binding domain and a Ca(2+)-independent catalytic domain. The Journal of biological chemistry 269, 18239–18249 (1994). [PubMed] [Google Scholar]
- 39.Choi OH et al. , Antigen and carbachol mobilize calcium by similar mechanisms in a transfected mast cell line (RBL-2H3 cells) that expresses ml muscarinic receptors. Journal of immunology (Baltimore, Md. : 1950) 151, 5586–5595 (1993). [PubMed] [Google Scholar]
- 40.Razin E et al. , IgE-mediated release of leukotriene C4, chondroitin sulfate E proteoglycan, beta-hexosaminidase, and histamine from cultured bone marrow-derived mouse mast cells. The Journal of experimental medicine 157, 189–201 (1983). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Suram S et al. , Pathways regulating cytosolic phospholipase A2 activation and eicosanoid production in macrophages by Candida albicans. The Journal of biological chemistry 285, 30676–30685 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Barrett NA, Maekawa A, Rahman OM, Austen KF, Kanaoka Y, Dectin-2 recognition of house dust mite triggers cysteinyl leukotriene generation by dendritic cells. Journal of immunology (Baltimore, Md. : 1950) 182, 1119–1128 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Buczynski MW et al. , TLR-4 and sustained calcium agonists synergistically produce eicosanoids independent of protein synthesis in RAW264.7 cells. The Journal of biological chemistry 282, 22834–22847 (2007). [DOI] [PubMed] [Google Scholar]
- 44.Bankova LG et al. , Leukotriene E4 elicits respiratory epithelial cell mucin release through the G-protein-coupled receptor, GPR99. Proceedings of the National Academy of Sciences of the United States of America 113, 6242–6247 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Tallini YN et al. , BAC transgenic mice express enhanced green fluorescent protein in central and peripheral cholinergic neurons. Physiol Genomics 27, 391–397 (2006). [DOI] [PubMed] [Google Scholar]
- 46.Ogura T, Krosnowski K, Zhang L, Bekkerman M, Lin W, Chemoreception regulates chemical access to mouse vomeronasal organ: role of solitary chemosensory cells. PloS one 5, e11924 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zhang Y et al. , Coding of sweet, bitter, and umami tastes: different receptor cells sharing similar signaling pathways. Cell 112, 293–301 (2003). [DOI] [PubMed] [Google Scholar]
- 48.Topilko P et al. , Krox-20 controls myelination in the peripheral nervous system. Nature 371, 796–799 (1994). [DOI] [PubMed] [Google Scholar]
- 49.Du N et al. , EGR2 is critical for peripheral naive T-cell differentiation and the T-cell response to influenza. Proceedings of the National Academy of Sciences of the United States of America 111, 16484–16489 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lazarevic V et al. , The gene encoding early growth response 2, a target of the transcription factor NFAT, is required for the development and maturation of natural killer T cells. Nature immunology 10, 306–313 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lee JH et al. , Anatomically and Functionally Distinct Lung Mesenchymal Populations Marked by Lgr5 and Lgr6. Cell 170, 1149–1163 e1112 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.O’Leary CE, Schneider C, Locksley RM, Tuft Cells-Systemically Dispersed Sensory Epithelia Integrating Immune and Neural Circuitry. Annu Rev Immunol, (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lei W et al. , Activation of intestinal tuft cell-expressed Sucnr1 triggers type 2 immunity in the mouse small intestine. Proceedings of the National Academy of Sciences of the United States of America 115, 5552–5557 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Rafehi M, Burbiel JC, Attah IY, Abdelrahman A, Muller CE, Synthesis, characterization, and in vitro evaluation of the selective P2Y2 receptor antagonist AR-C118925. Purinergic Signal 13, 89–103 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Communi D, Janssens R, Suarez-Huerta N, Robaye B, Boeynaems JM, Advances in signalling by extracellular nucleotides. the role and transduction mechanisms of P2Y receptors. Cell Signal 12, 351–360 (2000). [DOI] [PubMed] [Google Scholar]
- 56.Junger WG, Immune cell regulation by autocrine purinergic signalling. Nature reviews. Immunology 11, 201–212 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jacobson KA, Balasubramanian R, Deflorian F, Gao ZG, G protein-coupled adenosine (P1) and P2Y receptors: ligand design and receptor interactions. Purinergic Signal 8, 419–436 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lustig KD, Shiau AK, Brake AJ, Julius D, Expression cloning of an ATP receptor from mouse neuroblastoma cells. Proceedings of the National Academy of Sciences of the United States of America 90, 5113–5117 (1993). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Coddou C, Yan Z, Obsil T, Huidobro-Toro JP, Stojilkovic SS, Activation and regulation of purinergic P2X receptor channels. Pharmacol Rev 63, 641–683 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Feng C, Mery AG, Beller EM, Favot C, Boyce JA, Adenine nucleotides inhibit cytokine generation by human mast cells through a Gs-coupled receptor. Journal of immunology (Baltimore, Md. : 1950) 173, 7539–7547 (2004). [DOI] [PubMed] [Google Scholar]
- 61.Mellor EA, Austen KF, Boyce JA, Cysteinyl leukotrienes and uridine diphosphate induce cytokine generation by human mast cells through an interleukin 4-regulated pathway that is inhibited by leukotriene receptor antagonists. The Journal of experimental medicine 195, 583–592 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ledderose C et al. , Purinergic P2X4 receptors and mitochondrial ATP production regulate T cell migration. The Journal of clinical investigation 128, 3583–3594 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.O’Grady SM et al. , ATP release and Ca2+ signalling by human bronchial epithelial cells following Alternaria aeroallergen exposure. The Journal of physiology 591, 4595–4609 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ramu S et al. , Allergens produce serine proteases-dependent distinct release of metabolite DAMPs in human bronchial epithelial cells. Clinical and experimental allergy : journal of the British Society for Allergy and Clinical Immunology 48, 156–166 (2018). [DOI] [PubMed] [Google Scholar]
- 65.Bartemes KR et al. , IL-33-responsive lineage- CD25+ CD44(hi) lymphoid cells mediate innate type 2 immunity and allergic inflammation in the lungs. Journal of immunology (Baltimore, Md. : 1950) 188, 1503–1513 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Doherty TA et al. , STAT6 regulates natural helper cell proliferation during lung inflammation initiated by Alternaria. American journal of physiology. Lung cellular and molecular physiology 303, L577–588 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Doherty TA et al. , Alternaria induces STAT6-dependent acute airway eosinophilia and epithelial FIZZ1 expression that promotes airway fibrosis and epithelial thickness. Journal of immunology (Baltimore, Md. : 1950) 188, 2622–2629 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Bankova LG, Boyce JA, A new spin on mast cells and cysteinyl leukotrienes: Leukotriene E4 activates mast cells in vivo. The Journal of allergy and clinical immunology 142, 1056–1057 (2018). [DOI] [PubMed] [Google Scholar]
- 69.Peters-Golden M, Henderson WR Jr., Leukotrienes. The New England journal of medicine 357, 1841–1854 (2007). [DOI] [PubMed] [Google Scholar]
- 70.Claesson HE, Lindgren JA, Gustafsson B, Opsonized bacteria stimulate leukotriene synthesis in human leukocytes. Biochim Biophys Acta 836, 361–367 (1985). [DOI] [PubMed] [Google Scholar]
- 71.Rouzer CA, Scott WA, Cohn ZA, Blackburn P, Manning JM, Mouse peritoneal macrophages release leukotriene C in response to a phagocytic stimulus. Proceedings of the National Academy of Sciences of the United States of America 77, 4928–4932 (1980). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.McCurdy JD, Olynych TJ, Maher LH, Marshall JS, Cutting edge: distinct Toll-like receptor 2 activators selectively induce different classes of mediator production from human mast cells. Journal of immunology (Baltimore, Md. : 1950) 170, 1625–1629 (2003). [DOI] [PubMed] [Google Scholar]
- 73.Olynych TJ, Jakeman DL, Marshall JS, Fungal zymosan induces leukotriene production by human mast cells through a dectin-1-dependent mechanism. The Journal of allergy and clinical immunology 118, 837–843 (2006). [DOI] [PubMed] [Google Scholar]
- 74.Drazen JM et al. , Recovery of leukotriene E4 from the urine of patients with airway obstruction. The American review of respiratory disease 146, 104–108 (1992). [DOI] [PubMed] [Google Scholar]
- 75.Da Dalt L et al. , Nasal lavage leukotrienes in infants with RSV bronchiolitis. Pediatric allergy and immunology : official publication of the European Society of Pediatric Allergy and Immunology 18, 100–104 (2007). [DOI] [PubMed] [Google Scholar]
- 76.Sznajer Y et al. , Airway eicosanoids in acute severe respiratory syncytial virus bronchiolitis. J Pediatr 145, 115–118 (2004). [DOI] [PubMed] [Google Scholar]
- 77.Piedimonte G et al. , Leukotriene synthesis during respiratory syncytial virus bronchiolitis: influence of age and atopy. Pediatric pulmonology 40, 285–291 (2005). [DOI] [PubMed] [Google Scholar]
- 78.van Schaik SM et al. , Increased production of IFN-gamma and cysteinyl leukotrienes in virus-induced wheezing. The Journal of allergy and clinical immunology 103, 630–636 (1999). [DOI] [PubMed] [Google Scholar]
- 79.Lee RJ et al. , Bitter and sweet taste receptors regulate human upper respiratory innate immunity. The Journal of clinical investigation 124, 1393–1405 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Oymar K, Halvorsen T, Aksnes L, Mast cell activation and leukotriene secretion in wheezing infants. Relation to respiratory syncytial virus and outcome. Pediatric allergy and immunology : official publication of the European Society of Pediatric Allergy and Immunology 17, 37–42 (2006). [DOI] [PubMed] [Google Scholar]
- 81.Idzko M, Ferrari D, Eltzschig HK, Nucleotide signalling during inflammation. Nature 509, 310–317 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Idzko M et al. , Extracellular ATP triggers and maintains asthmatic airway inflammation by activating dendritic cells. Nature medicine 13, 913–919 (2007). [DOI] [PubMed] [Google Scholar]
- 83.Beale J et al. , Rhinovirus-induced IL-25 in asthma exacerbation drives type 2 immunity and allergic pulmonary inflammation. Science translational medicine 6, 256ra134 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Tizzano M, Cristofoletti M, Sbarbati A, Finger TE, Expression of taste receptors in solitary chemosensory cells of rodent airways. BMC pulmonary medicine 11, 3 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Patel NN et al. , Fungal extracts stimulate solitary chemosensory cell expansion in noninvasive fungal rhinosinusitis. Int Forum Allergy Rhinol 9, 730–737 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Rane CK et al. , Development of solitary chemosensory cells in the distal lung after severe influenza injury. American journal of physiology. Lung cellular and molecular physiology 316, L1141–L1149 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Taylor-Clark TE, Nassenstein C, Undem BJ, Leukotriene D4 increases the excitability of capsaicin-sensitive nasal sensory nerves to electrical and chemical stimuli. British journal of pharmacology 154, 1359–1368 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Wang J et al. , Distinct and common expression of receptors for inflammatory mediators in vagal nodose versus jugular capsaicin-sensitive/TRPV1-positive neurons detected by low input RNA sequencing. PloS one 12, e0185985 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Chiu IM et al. , Transcriptional profiling at whole population and single cell levels reveals somatosensory neuron molecular diversity. Elife 3, (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Lopes DM, Denk F, McMahon SB, The Molecular Fingerprint of Dorsal Root and Trigeminal Ganglion Neurons. Front Mol Neurosci 10, 304 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Solinski HJ et al. , Nppb Neurons Are Sensors of Mast Cell-Induced Itch. Cell reports 26, 3561–3573 e3564 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Kanaoka Y, Maekawa A, Penrose JF, Austen KF, Lam BK, Attenuated zymosan-induced peritoneal vascular permeability and IgE-dependent passive cutaneous anaphylaxis in mice lacking leukotriene C4 synthase. The Journal of biological chemistry 276, 22608–22613 (2001). [DOI] [PubMed] [Google Scholar]
- 93.Matsumoto I, Ohmoto M, Narukawa M, Yoshihara Y, Abe K, Skn-1a (Pou2f3) specifies taste receptor cell lineage. Nat Neurosci 14, 685–687 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Homolya L, Watt WC, Lazarowski ER, Koller BH, Boucher RC, Nucleotide-regulated calcium signaling in lung fibroblasts and epithelial cells from normal and P2Y(2) receptor (−/−) mice. The Journal of biological chemistry 274, 26454–26460 (1999). [DOI] [PubMed] [Google Scholar]
- 95.You Y, Richer EJ, Huang T, Brody SL, Growth and differentiation of mouse tracheal epithelial cells: selection of a proliferative population. American journal of physiology. Lung cellular and molecular physiology 283, L1315–1321 (2002). [DOI] [PubMed] [Google Scholar]
- 96.Thomas MG, Marwood RM, Parsons AE, Parsons RB, The effect of foetal bovine serum supplementation upon the lactate dehydrogenase cytotoxicity assay: Important considerations for in vitro toxicity analysis. Toxicol In Vitro 30, 300–308 (2015). [DOI] [PubMed] [Google Scholar]
- 97.Barrett NA et al. , Dectin-2 mediates Th2 immunity through the generation of cysteinyl leukotrienes. The Journal of experimental medicine 208, 593–604 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Dobin A et al. , STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.DeLuca DS et al. , RNA-SeQC: RNA-seq metrics for quality control and process optimization. Bioinformatics 28, 1530–1532 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Liao Y, Smyth GK, Shi W, The Subread aligner: fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res 41, e108 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Love MI, Huber W, Anders S, Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 15, 550 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S2. Raw data in Excel file.
Fig. S1. Nasal ChAT-eGFP+ cells share the tuft cell signature.
Fig. S2. Nasal BrCs are enriched for phosphatidylinositol – phospholipase signaling and CysLT biosynthetic pathway transcripts.
Fig. S3. BrCs are the dominant source of CysLTs in the naïve nasal mucosa.
Fig. S4. Taste receptors are highly expressed in tracheal and respiratory nasal but not olfactory nasal BrCs.
Fig. S5. CD45 is expressed on nasal BrCs
Fig. S6. Aeroallergen-triggered pathways of BrC activation.
Table S1. Reagents.