

Abstract
Autophagy is a cellular process to clear pathogens. Salmonella enterica serovar Enteritidis (S.E) has emerged as one of the most important food-borne pathogens. However, major studies still focus on Salmonella enterica serovar Typhimurium. Here, we reported that AvrA, a S. Enteritidis effector, inhibited autophagy to promote bacterial survival in the host. We found that AvrA regulates the conversion of LC3 I into LC3 II and the enrichment of lysosomes. Beclin-1, a key molecular regulator of autophagy, was decreased after AvrA expressed strain colonization. In S.E-AvrA−-infected cells, we found the increases of protein levels of p-JNK and p-c-Jun and the transcription level of AP-1. AvrA-reduction of Beclin-1 protein expression is through the JNK pathway. The JNK inhibitor abolished the AvrA-reduced Beclin-1 protein expression. Moreover, we identified that the AvrA mutation C186A abolished its regulation of Beclin-1 expression. In addition AvrA protein was found interacted with Beclin-1. In organoids and infected mice, we explored the physiologically related effects and mechanism of AvrA in reducing Beclin-1 through the JNK pathway, thus attenuating autophagic responses. This finding not only indicates an important role of S. Enteritidis effector in reducing host protein as a strategy to suppress autophagy, but also suggests manipulating autophagy as a new strategy to treat infectious diseases.
Keywords: autophagy, effector, infection, organoids, paneth cells
Introduction
Salmonella is a Gram-negative, facultative anaerobe and an intracellular pathogen to both humans and animals. Salmonella enterica serovar Enteritidis has emerged as one of the most important food-borne pathogens for humans, and it is mainly associated with the consumption of contaminated poultry meat and egg (1, 2). Infection caused by Salmonella Enteritidis is the second most common cause of bacterial gastroenteritis in the developed world, and results in significant economic loss to the poultry industry and places a substantial burden on the healthcare system (2–4). Salmonella Enteritidis is an important pathogen with a public concern, thus demanding further studying. However, the majority of basic researches in Salmonela field still prefer to use Salmonella enterica serovar Typhimurium (Salmonella Typhimurium) as research model to study host-microbial interactions.
To survive in host cells, Salmonella use mechanisms to prevent clearance from host cells, such as escaping from the phagosome, inhibiting phagosome-lysosome fusion, and inhibiting apoptosis and autophagy in host cells (5–11). Among the numerous host defense systems against pathogens, anti-bacterial autophagy provides potent cell-autonomous immunity against bacterial attempts to colonize the cytosol of host cells (12, 13). During this process, the phagophore expands and engulfs pathogens, and closes to originate the autophagosome that fuses with the lysosome, at which the degradation of the pathogens takes place (14). There are more than 20 ATG proteins (many of which are evolutionarily conserved) that are essential for the execution of autophagy (15). Notably, the mammalian autophagy protein Beclin-1, an ortholog of the Atg6 in yeast, is a key molecule regulator of autophagy. Beclin-1 interacts with several cofactors (e.g., Atg14L, HMGB1, IP3R, PINK, and survivin) to regulate the lipid kinase Vps-34 protein and promote the formation of Beclin-1-Vps34-Vps15 core complexes, thereby inducing autophagy (16, 17).
Salmonella possesses a range of effector proteins that are translocated into the host cells via a type III secretion system (T3SS). These effector proteins are generally assumed to influence the host's cellular functions to facilitate Salmonella invasion and intracellular carriage (18–20). AvrA is one of the Salmonella effectors secreted by the Salmonella pathogenicity island 1 (SPI-1) T3SS. The AvrA protein in Salmonella Typhimurium is an anti-inflammatory effector that possesses acetyltransferase activity and inhibits the host c-Jun N-terminal kinase (JNK)/AP-1 and NF-κB signaling pathways. Through these methods, AvrA inhibits the host inflammatory response and stabilizes the intestinal tight junctions to the benefit of bacterial survival (8, 21–25). However, the role of AvrA in the interaction between Salmonella infection and host autophagic response is unexplored.
Here, we hypothesize that Salmonella Enteritidis effector AvrA inhibits the autophagic response by decreasing Beclin-1 expression. We used wild-type, Salmonella Enteritidis C50336 and established a deletion Salmonella Enteritidis mutant S.E-AvrA− and a plasmid-mediated complementary strain S.E-AvrA−/pAvrA+ (S.E-AvrA+) (25). In cell cultures, organoids and infected mice, we explored the physiologically related effects and molecular mechanism of AvrA regulation of autophagy in intestinal epithelial cells, whereas most studies on bacterial effectors and autophagy only use cell cultures. Our study provides new insights into the mechanisms of the bacterial effects in regulating host-microbial interactions.
Materials and Methods
Animals and Ethics Statement
C57BL/6 mice (female, 6–8 weeks) were obtained from the Jackson Laboratory (Jackson Laboratory, Bar Harbor, ME, USA). All the animal work was approved by the University of Illinois at Chicago Committee on Animal Resources. Euthanasia was accomplished via sodium pentobarbital administration (100 mg per kg body weight, i.p.) followed by cervical dislocation. All procedures were conducted in accordance with the approved guidelines of the Committees on Animal Resources.
Bacterial Strains and Growth Conditions
The wild-type Salmonella Enteritidis strain C50336 (S.E WT) was obtained from the National Institute for the Control of Pharmaceutical and Biological Products (NICPBP), China. The Salmonella Enteritidis AvrA mutant strain S.E-AvrA− was derived from C50336, and the AvrA complemented strain S.E-AvrA+ was constructed as in previous studies by our laboratory (25). The bacteria and plasmids used in this study are listed in Table 1. The bacterial growth conditions were as follows: the non-agitated microaerophilic bacterial cultures were prepared by inoculating 10 ml of Luria-Bertani (LB) broth with 0.01 ml of a stationary phase culture followed by an overnight incubation (>18 h) at 37°C (27).
Table 1.
Bacterial strains and plasmids used in this study.
| Strains/Plasmids | Characteristics | References |
|---|---|---|
| Strains | ||
| S.E-WT | Wild-type Salmonella Enteritidis CMCC(B)50336 | NICPBP, China |
| S.E-AvrA− | C50336 AvrA-deficient mutant | (25) |
| S.E-AvrA+ | C50336 AvrA-deficient mutant carrying pBR322-AvrA (Ampr) | (25) |
| Plasmids | ||
| pCMV-c-myc-AvrA | Plasmid of S.E full-length AvrA gene, AvrA WT plasmid, c-myc tag | (21) |
| pCMV-c-myc-AvrA(C186A) | Plasmid of S.E full-length AvrA gene with one-point mutant C186A, AvrA C186A mutant plasmid, c-myc tag | (21) |
| pcDNA4-HA-Beclin-1 | Plasmid of human BECN1 gene, Beclin-1 WT plasmid, HA tag | (26) |
| pGL3-AP1 | Plasmid of AP1 promoter gene, fluorescein tag | (21) |
Cell Culture
Human epithelial HCT116, Caco-2 BBE, and SKCO-15 cells were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), streptomycin-penicillin and L-glutamine. Bacterial colony forming units (CFU) were determined by plating diluted cell lysates onto LB agar culture plates and incubating the cultures at 37°C overnight (25).
Mouse Intestinal Organoid Isolation, Culture, and Passage
The mouse small intestines were removed immediately after cervical dislocation. The stool was then flushed out with ice-cold PBS (penicillin, 100 I.U./mL/streptomycin, 100 μg/mL), and the small intestines were dissected and opened longitudinally and cut into small (~1 cm) pieces. The tissues were rocked in PBS with 2 mmol/L ethylenediamine tetraacetic acid (EDTA) for 30 min at 4°C and were then switched to PBS with 54.9 mmol/L D-sorbitol and 43.4 mmol/L sucrose. The tissues were then vortexed for 1–2 min and were filtered through a 70 μm sterile cell strainer. The crypts were collected by centrifugation at 150 g for 10 min at 4°C. Approximately 500 crypts were suspended in 50 μL of growth factor-reduced phenol-free Matrigel (BD Biosciences, San Jose, CA). Next, a 50 μL droplet of the Matrigel/crypt mix was placed in the center well of a 12-well plate. After 30 min of polymerization, 650 μL of the mini gut medium was overlain (28, 29). The mini gut medium (advanced DMEM/F12 supplemented with HEPES, L-glutamine, N2, and B27) was added to the culture, along with R-Spondin, Noggin, and EGF. The medium was changed every 2–3 days. For passage, the organoids were removed from the Matrigel and broken up mechanically by passing them through a syringe and needle (27 G, BD Biosciences, San Jose, CA), and then, they were transferred to fresh Matrigel. The passage was performed every 7–10 days with a 1:4 split ratio.
Bacterial Colonization
Polarized human epithelial cells were grown in DMEM with 10% FBS. At 90–100% confluence, the cells were colonized with an equal number of the indicated Salmonella Enteritidis strain for 30 min, washed with Hank's balanced salt solution (HBSS), and incubated in DMEM containing gentamicin (100 μg/ml) for 30 min. The first 30 min of the incubation allowed the bacteria to contact the epithelial cell surface and inject the effectors into the host cells (30, 31). After extensive HBSS washing, the extracellular bacteria were washed away. The incubation with gentamicin inhibited the bacterial growth (32). At the indicated times, post-colonization, the cells samples were harvested for the analysis.
The organoids (6 days after passage) were colonized with the indicated Salmonella Enteritidis strain for 30 min, washed with HBSS, and incubated in mini gut media containing gentamicin (500 mg/mL) for the indicated times, as described in our previous studies (29). After extensive HBSS washing, the extracellular bacteria were washed away. The incubation with gentamicin inhibited the growth of the bacteria. Samples were collected for a Western blot analysis or immunofluorescence after the organoids were colonized with Salmonella for 30 min and were then incubated in medium with gentamicin for 1 h.
Cell Treatment With the JNK Inhibitor SP600125
The JNK inhibitor SP600125 (50 mM, EMD Biosciences, San Diego, CA, USA) was added directly to the culture medium 1 h before Salmonella treatment. The HCT116 cells were pretreated with SP600125 and were incubated with the indicated Salmonella for 30 min, washed 3 times in HBSS, incubated in DMEM containing 100 μg/ml gentamicin and 50 μM SP600125 for 30 min, and harvested. The protein levels were determined by Western blotting.
Lysotracker Staining
Lysotracker staining was performed following the manufacturer's protocol (Life technologies). The HCT116 cells were grown in the Lab-Tek Chambered Coverglass System (154526, Thermo Scientific, Rockford, IL, USA), and the cells were then incubated with 100 nM LysoTracker Deep Red Probe (L12492, Life technologies, Eugene, OR, USA) in cell growth medium at 37°C for 60 min. After washing with HBBS, the cells were incubated with the indicated Salmonella for 30 min, washed 3 times in HBSS, incubated in DMEM containing 100 μg/ml gentamicin for 30 min, and the cells were detected by fluorescence microscopy (ECLIPSE E600, Nikon Instruments, Inc., Melville, NY, USA.).
Streptomycin Pre-treated Salmonella Mouse Model
Water and food were withdrawn 4 h before an oral gavage with 7.5 mg/mouse streptomycin. Afterward, the animals were supplied with water and food. Twenty hour after the streptomycin treatment, water and food were once again withdrawn for 4 h before the mice were infected with 1.0 × 108 CFU of Salmonella (100 μl bacterial suspension in HBSS) or treated with sterile HBSS (control) by oral gavage, as previously described (33). At the indicated times post-infection, the mice were sacrificed, and the intestinal tissue samples were removed for the analysis.
Immunoblotting and Antibodies
Mouse epithelial cells were scraped and lysed in lysis buffer (1% Triton X-100, 150 mM NaCl, 10 mM Tris pH 7.4, 1 mM EDTA, 1 mM EGTA pH 8.0, 0.2 mM sodium orthovanadate, and protease inhibitor cocktail), and the protein concentration was measured using Protein Assay Kits (Bio-Rad, Hercules, CA, USA) (30). The cultured cells were rinsed twice in ice-cold HBSS and were lysed in protein loading buffer (50 mM Tris pH 6.8, 100 mM dithiothreitol, 2% SDS, 0.1% bromophenol blue, and 10% glycerol), and the remaining cells were scraped off the dish and sonicated to shear the DNA and reduce the sample viscosity. The organoid cells were rinsed three times in ice-cold HBSS and were then suspended in ice-cold HBSS. The organoid cells were then spun down at 900 rpm for 10 min at 4°C. Next, using a pipette to aspirate the PBS at the top, the organoid cells were lysed in lysis buffer and were then sonicated (29). The protein concentration was then measured. Equal amounts of protein were separated by SDS-polyacrylamide gel electrophoresis and were transferred to nitrocellulose membranes. The non-specific sites were blocked with 5% bovine serum albumin (34) in TBST (50 mM Tris, 150 mM NaCl, and 0.05% Tween 20 adjusted to pH 7.6 with HCl), and the membranes were then incubated with dilutions of the primary antibodies as recommended by the manufacturers. The primary antibodies included the following: anti-p62 (1:1,000, AP2183B, ABGENT, San Diego, CA,USA); anti-LC3B (1:1,000, 2775), anti-SAPK/JNK (1:1,000, 9258), anti-phospho-SAPK/JNK (Thr183/Tyr185, 1:1,000, 9251), anti-c-jun (60A8, 1:1,000, 9165), anti-phospho-c-jun (S63, 1:1,000, 9261) (Cell Signaling, Beverly, MA, USA); anti-BECN1/Beclin-1 (1:1,000, SC-10086), anti-c-myc (PE10, 1:1,000, SC-40), anti-HA-probe (F-7, 1:1,000, SC-7392) (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA); anti-β-actin (1:2,000, A5441, Sigma-Aldrich, Milwaukee, WI, USA); anti-ubiquitin (1:1,000, UG9511, ENZO, Farmingdale, NY, USA); and anti-AvrA [1:1,000, custom-made, as reported in our previous studies (35)]. The membranes were washed and incubated with an HRP-conjugated secondary antibody (anti-mouse, 1:5,000, 31430; anti-rabbit, 1:5,000, 31460; anti-goat, 1:5,000, R-21459, Invitrogen, Grand Island, NY, USA). The membranes were then washed again, treated with the ECL Western Blotting Substrate (Thermo Scientific, Rockford, IL, USA) and visualized on X-ray film. The membranes that were sequentially probed with more than one antibody were stripped in stripping buffer (Thermo Scientific, Rockford, IL, USA) before re-probing.
Immunoprecipitation
The cells were rinsed twice in ice-cold HBSS and were lysed in ice-cold immunoprecipitation buffer (1% Triton X-100, 150 mmol/L NaCl, 10 mmol/L Tris, pH 7.4, 1 mmol/L EDTA, 1 mmol/L ethylene glycol bis [β-aminoethyl ether)-N,N,N',N'-tetraacetic acid, pH 8.0, 0.2 mmol/L sodium orthovanadate, and protease inhibitor cocktail (Roche Diagnostics, Basel, Switzerland)]. The samples were prepared as previously described (36). The blots were probed with anti-HA-probe (F-7, 1:1,000, SC-7392, Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and anti-c-myc (PE10, SC-40, Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) antibodies.
Immunofluorescence
The ileal tissues from the distal portion of the ileum were freshly isolated and paraffin-embedded after fixation with 10% neutral buffered formalin. Immunofluorescence was performed on the paraffin-embedded sections (5 μm). After preparation of the slides, as described previously (31), the tissue samples were incubated with the indicated primary antibody, anti-lysozyme (1:100, sc27958, Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), at 4°C overnight. The samples were then incubated with the sheep anti-goat Alexa Fluor 594 (A11058, Life Technologies, Grand Island, NY, USA) and DAPI (D1306, Life Technologies, Grand Island, NY, USA) for 1 h at room temperature. The tissues were mounted with SlowFade (s2828, Life technologies, Grand Island, NY, USA), followed by a coverslip, and the edges were sealed to prevent drying. The specimens were examined with a Zeiss laser scanning microscope 710 (Carl Zeiss, Inc., Oberkochen, Germany).
Paneth Cell Staining and Counting
The Paneth cells in the mouse ileal tissue were counted after the anti-lysozyme immunofluorescence staining, according to our previous study (37). The patterns of the lysozyme expression in the Paneth cells were classified into four categories as follows: normal (D0); disordered (D1); depleted (D2) and diffuse (D3), according to previously published methods (38).
AP-1 Transcriptional Activity Assay
The cells were transiently transfected with 1 μg of pGL3-AP1 plasmid using the Lipofectamine 3,000 transfection kit, according to the manufacturer's instructions (Invitrogen, Carlsbad, CA). The pRL-TK vector was used as an internal control reporter. After 24 h post-transfection, the cells were colonized with equal numbers of bacteria for 30 min, washed, and incubated in DMEM containing gentamicin (100 μg/ml) for 30 min. The luciferase activity was monitored using the dual luciferase assay system (Promega).
Statistical Analysis
Data are expressed as the mean ± SE. All statistical tests were 2-sided. The p-values < 0.05 were considered statistically significant. The differences between two samples were analyzed using Student's t-test; the differences among three or more groups were analyzed using one-way ANOVA. The Tukey's method was used to adjust multiple comparisons to ensure results accurately. All statistical analyses were performed by GraphPad Prism 5(GraphPad Software, La Jolla, CA) or SAS version 9.4 (SAS Institute, Inc., Cary, NC, USA).
Results
Salmonella Enteritidis AvrA Decreases Autophagy Markers and Enhanced Bacterial Invasion in vitro
Autophagy is an important cell-autonomous defense mechanism required for pathogen clearance (12, 13). LC3 and p62 are well-recognized markers for autophagic activity (39). In this study, we found that in human intestinal epithelial HCT116 cells, the conversion of LC3 I into LC3 II was increased after S.E-AvrA− infection compared to that after wild-type S.E or S.E-AvrA+ infection. Notably, p62, which is a bona fide target of autophagosomal degradation, was decreased in the cells infected with the S.E-AvrA mutant strain compared with the expression in cells infected with the S.E-WT or S.E-AvrA+ strains (Figure 1A). A densitometry analysis showed a significant difference between the cells infected with the different S.E strains (Figures 1B,C). Meanwhile, HCT116 cells pre-treated with LysoTracker showed more lysosomes in the cells infected with the S.E-AvrA mutant strain than in the cells infected with the AvrA expressed strains (Figures 1D,E). The role of AvrA in Salmonella Enteritidis invasion is unknown. We further compared the invasion ability of Salmonella Enteritidis strains with or without AvrA expression. We found that the bacterial adhesion to human epithelial cells had no significant difference among S.E-WT, S.E-AvrA− and S.E-AvrA+(Figure 1F). However, S.E-AvrA− colonized human epithelial cells showed a decreased intracellular bacterial load compared to those colonized with wild type S.E or S.E-AvrA+ (Figure 1G). We observed similar trends in autophagic activity following S.E-AvrA−, S.E-AvrA+ and S.E-WT infection in human Caco-2 BBE and SKCO-15 cells (data not shown). Taken together, these data suggest that the Salmonella Enteritidis effector AvrA inhibits autophagy in vitro.
Figure 1.
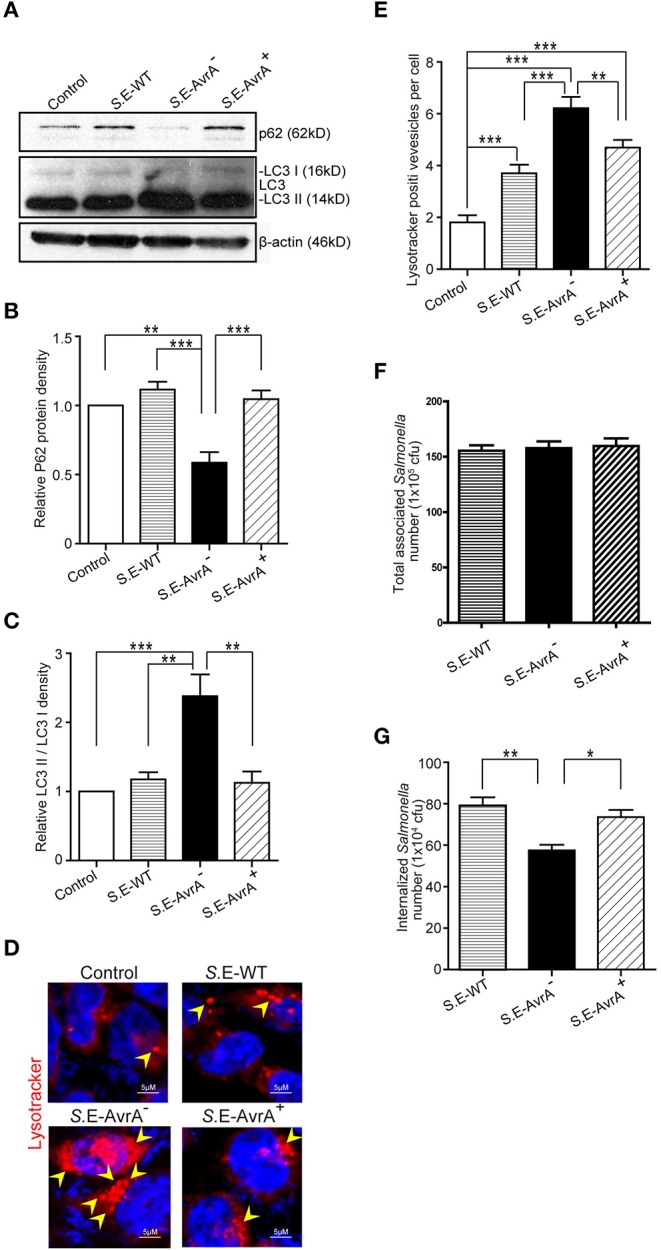
Salmonella Enteritidis AvrA inhibits autophagy in cell models. AvrA-regulated expression levels of autophagy related proteins. (A) The indicated HCT116 cell lines were infected with S.E WT, S.E-AvrA− and S.E-AvrA+ strains (MOI:1:1, 1 h incubation before harvested, n = 4) as shown and analyzed for protein expression by immunoblotting. The immunoblotting of P62/LC3 was used to track the expression of P62 and the conversion of LC3 I into LC3 II for autophagic activity in the HCT116 cells after infection with the different S.E. The relative density of P62 (B) and LC3 II/LC3 I (C) was determined using Quantity One 4.6.2 software (Bio-Rad, Hercules, CA, USA). N = 4, **adjusted P < 0.01, ***adjusted P < 0.001 by ANOVA test. (D) HCT116 cell lines were incubated with 100 nM LysoTracker Deep Red Probe, then infected with S.E WT, S.E-AvrA- and S.E-AvrA+ strains (MOI:1:1, 1 h incubation before microscopic examination) to check the lysosomes staining. The immunofluorescence indicated that the HCT116 cells, pre-treated with LysoTracker, showed more lysosomes in the cells infected with the S.E-AvrA− bacteria compared with the cells infected by the AvrA present strains. (E) Quantification of the number of lysotracker positive vesicles. The data are reported as the mean ± SE from three independent experiments, and a total of 100 cells per condition were analyzed. n = 100, **adjusted P < 0.01, ***adjusted P < 0.001 by ANOVA test. The numbers of associated Salmonella and internalized Salmonella in human epithelial cells colonized with wild-type Salmonella Enteritidis or AvrA mutant or AvrA-complemented strains. Human epithelial cells were grown on an insert, colonized with an equal number of the indicated bacteria for 30 min, after washed with HBSS, the number of associated Salmonella (Salmonella adhesion) was determined (F). Alternatively, cells were incubated in DMEM containing gentamicin (100 μg/ml) for 30 min, and the number of internalized intracellular Salmonella (Salmonella invasion) was then determined (G). Data are reported as the mean ± SE from six independent experiments, *adjusted P < 0.05, **adjusted P < 0.05 by ANOVA test.
AvrA Reduces Beclin-1 at the Protein Levels and Interacts With Beclin-1
Beclin-1, a key molecular regulator of autophagy, interacts with several cofactors to regulate the lipid kinase Vps-34 protein and promote the formation of Beclin-1-Vps34-Vps15 core complexes, thereby inducing autophagy (16, 17). Thus, we determined whether the protein level of Beclin-1 was changed by the infection with the different S.E strains. As shown in Figures 2A,B, Beclin-1 protein expression was significantly decreased after colonization of the AvrA present strains for 1 h. In contrast, the cells colonized with the S.E-AvrA− bacteria maintained Beclin-1 protein expression. To verify that AvrA affects the protein expression of Beclin-1, we transfected an AvrA WT plasmid or an AvrA C186A mutant [mutated at the key cysteine required for AvrA activity (21)] plasmid into HCT116 cells. As expected, the AvrA WT plasmid decreased endogenous Beclin-1 protein expression. However, the AvrA C186A mutant plasmid maintained the endogenous Beclin-1 protein expression (Figures 2C,D). We further determined the interaction of AvrA/Beclin-1 in the HCT116 cells by immunoprecipitation. Vps34 was used as a positive control. We found that exogenous AvrA (c-myc tag) co-immunoprecipitated with exogenous Beclin-1 (HA- tag), suggesting that AvrA interacted with Beclin-1 (Figure 2E). Therefore, the data suggest that the S.E effector AvrA changes Beclin-1 protein levels to inhibit autophagy, in addition AvrA can interact with Beclin-1 protein.
Figure 2.

AvrA changes Beclin-1 protein levels and interacts with Beclin-1 in HCT116 cells. AvrA changes Beclin-1 protein levels and interacts with Beclin-1 in HCT116 cells. (A) The Western blot shows the expression of Beclin-1 in the HCT116 cells after colonization with wild-type Salmonella Enteritidis or AvrA mutant or AvrA-complemented strains (MOI:1:1, 1 h incubation before harvested, n = 4). The relative density of Beclin-1 (B) was determined using Quantity One 4.6.2 software (Bio-Rad, Hercules, CA, USA). n = 4, *adjusted P < 0.05 by ANOVA test. (C) The Western blot shows the expression of Beclin-1 in the HCT116 cells after transfection with the AvrA wild-type and AvrA mutant plasmids (200 ng/μl, 24 h incubation, n = 5). The relative density of Beclin-1 (D) was determined using Quantity One 4.6.2 software (Bio-Rad, Hercules, CA, USA). n = 5, *adjusted P < 0.05, **adjusted P < 0.01 by ANOVA test. (E) The HCT116 cells were cotransfected with the AvrA WT plasmid (c-myc tag) and the Beclin-1 WT plasmid (HA-tag) (200 ng/μl, 24 h incubation, n = 4). At the indicated times, immunoprecipitation was performed with an anti-c-myc mouse monoclonal antibody. Pre-immune mouse IgG was used as a negative control. VPS34 was used as a positive control. The Western blot analyses of the pre-immunoprecipitation (Input) and immunoprecipitated samples (IP) were performed with an anti-c-myc mouse monoclonal antibody or with an anti-HA mouse monoclonal antibody. These results shown are representative of three independent experiments.
AvrA Inhibits the JNK Signaling Pathway to Decrease Beclin-1
Beclin-1 is regulated by the JNK signaling pathway (40). Previous studies have shown that Salmonella AvrA inhibits the activation of the JNK signaling pathways (8, 25). Using Western blotting, we found that the protein levels of p-JNK and p-c-Jun were higher in the S.E-AvrA−-infected cells than in the cells infected by the S.E-WT or S.E-AvrA+ strains (Figure 3A). Meanwhile, a luciferase reporter assay showed that AP-1 transcription was increased, as a consequence of the activation of JNK/c-JUN (Figure 3B). These data suggest that the JNK/c-jun pathway and AP-1 transcription are more highly activated in the S.E-AvrA−-infected cells than in the cells infected with the AvrA present strains. Interestingly, after treatment with the JNK inhibitor SP600125, the level of Beclin-1 and P62 protein expression was not different between the cells infected with AvrA or without AvrA (Figures 3A,B), indicating that the AvrA-related responses were abolished by the JNK inhibitor. Thus, our data indicate that the S.E effector AvrA inhibits the autophagic response by decreasing Beclin-1 at the protein level, and it occurs by inhibiting the JNK/c-Jun/AP-1 signaling pathways.
Figure 3.
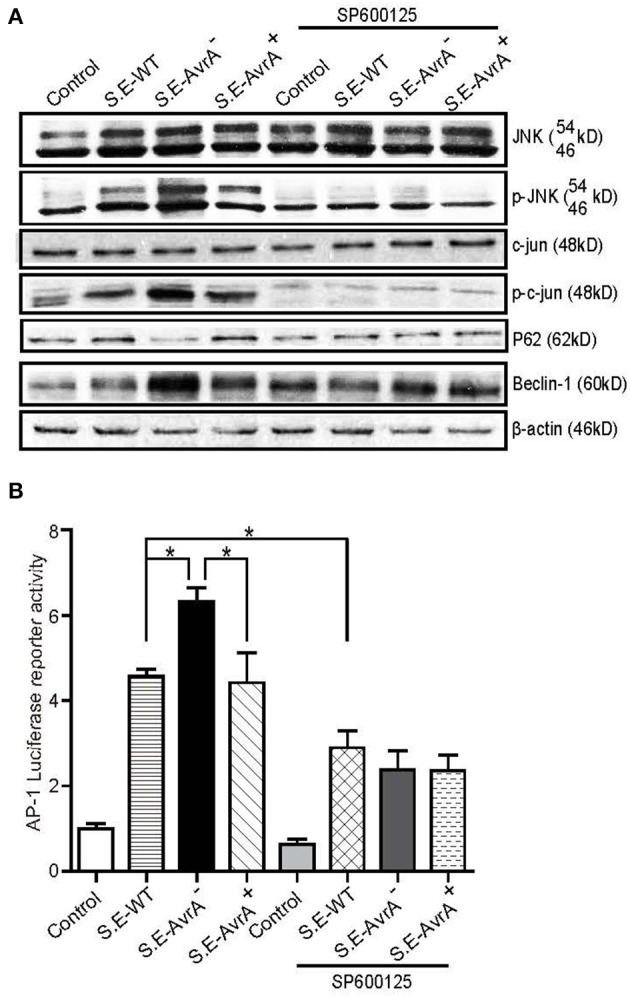
AvrA inhibits the JNK/c-jun/AP-1 signaling pathway to decrease Beclin-1 in HCT116 cells. AvrA inhibited the JNK pathway and decreased Beclin-1 in HCT116 cells, and the effect of AvrA on Beclin-1 expression was abolished by the JNK inhibitor SP600125. (A) The Western blot shows a change in the JNK pathway markers and Beclin-1 in the cells treated with the JNK inhibitor SP600125 (50 μM, 30 min) and those infected with the different S.E strains (MOI:1:1, 1 h incubation before harvested, n = 4), These results shown are representative of four independent experiments. (B) The luciferase reporter assay shows a change in AP-1 transcription in the cells treated with the JNK inhibitor SP600125 and in those infected with the different S.E strains (MOI:1:1, 1 h incubation before harvested, n = 6). *adjusted P < 0.05 by ANOVA test.
The AvrA Mutant C186A Plasmid Expression Abolishes the Regulation of Exogenous Beclin-1 Expression
To further study the function of the AvrA protein, we cotransfected an AvrA WT plasmid or an AvrA C186A mutant plasmid with a Beclin-1 WT plasmid. The AvrA C186A mutation is known to abolish the enzyme activity of AvrA (21). Our data showed that the AvrA WT plasmid decreased not only the endogenous Beclin-1 protein but also the exogenous Beclin-1 protein. In contrast, the AvrA C186A mutant plasmid abolished its regulation on Beclin-1 expression (Figures 4A,B). Moreover, we found that the protein levels of p-JNK, p-c-Jun and Beclin-1 were decreased in the AvrA WT plasmid transfected cells compared with the cells transfected with the AvrA C186A mutant plasmid. These data verified that AvrA decreased Beclin-1 by inhibiting the JNK signaling pathways, whereas the AvrA C186A mutation abolished its regulation (Figure 4C). We did not observe the change of the Beclin-1 at the mRNA level (data not shown).
Figure 4.
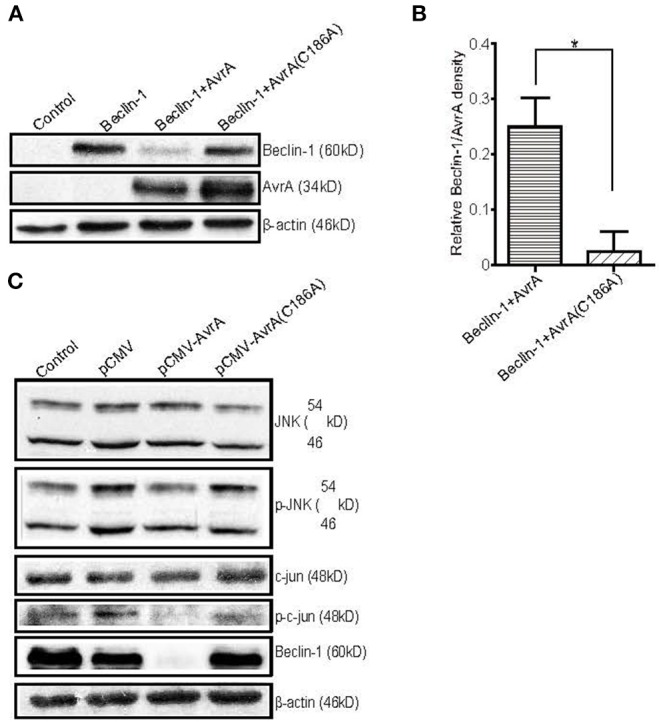
The AvrA C186A mutant plasmid abolishes the regulation of exogenous Beclin-1. The HCT116 cells were transfected with the indicated plasmids (200 ng/μl, 24 h incubation, n = 4). At the indicated times, the changes in the target proteins were measured. (A) The Western blot shows the expression of Beclin-1 in the HCT116 cells after transfection with the indicated plasmids. The relative density of Beclin-1 (B) was determined using Quantity One 4.6.2 software (Bio-Rad, Hercules, CA, USA). n=4, *P < 0.05 by student's t-test. (C) The Western blot shows the change in the JNK pathway markers and Beclin-1 in the HCT116 cells after transfection with the AvrA wild-type and AvrA mutant plasmids. These results shown are representative of four independent experiments.
AvrA Expressing Bacteria Reduce the Levels of Beclin-1 Protein in Mouse Organoids
Intestinal organoid culture is a newly developed 3D system to determine the bacterial–epithelial interactions post Salmonella infection (29). We found that the Beclin-1 protein expression was significantly decreased after the infection of the S.E-WT expressing AvrA. In contrast, the organoids colonized with the S.E-AvrA− mutant strain increased Beclin-1 protein expression, whereas S.E-AvrA+ could reduce the Beclin-1 expression (Figures 5A,B). Moreover, we found AvrA-associated changes of P62, LC3 II / LC3 I, and p-JNK in the Salmonella-infected organoids (Figures 5A,C,D). Decreased autophagy marker P62 was further confirmed in organoids colonized with the S.E-AvrA− strain in comparison with organoids infected by the S.E-WT and S.E-AvrA+ strains by Immunostaining (Figure 5E). These changes in the 3D organoids were consistent with AvrA-suppressed autophagy observed in the 2D cultured cell lines.
Figure 5.

Salmonella Enteritidis AvrA changes the levels of Beclin-1 in mouse intestinal organoids. Salmonella Enteritidis infection and invasion in the mouse organoids. (A) The Western blot shows the expression of Beclin-1, P62, LC3, and the JNK pathway markers in the organoids after infection with wild-type Salmonella Enteritidis or AvrA mutant or AvrA-complemented strains (MOI:1:1, 1.5 h incubation before harvested, n = 3). The relative density of Beclin-1 (B), P62 (C), and LC3 II/LC3 I (D) was determined using Quantity One 4.6.2 software (Bio-Rad, Hercules, CA, USA). n = 3, *adjusted P < 0.05, **adjusted P < 0.01, #unadjusted P < 0.05 by ANOVA test. (E) The representative images of the immunostaining of P62 in the organoids after infection with wild-type Salmonella Enteritidis or AvrA mutant or AvrA-complemented strains (MOI:1:1, 1.5 h incubation).
AvrA Changes the Levels of Beclin-1 and Affects the Function of Paneth Cell Granules of the Ileal Tissues in a Mouse Model
To study the role of the S.E effector AvrA in an in vivo model of natural intestinal infection, we used the streptomycin pretreatment mouse model of enteric salmonellosis (33, 41). C57BL/6 mice (female, 6–8 weeks) were pretreated with streptomycin for 24 h before infection with the S.E-WT, S.E-AvrA− or S.E-AvrA+ strains by oral gavage. In the Ileum samples from the S.E-WT-infected mice, Beclin-1 protein expression was significantly decreased compared to the expression in the samples from the S.E-AvrA−-infected mice (Figures 6A,B). As expected, decreased P62, increased conversion of LC3 I into LC3 II and activation of the JNK pathway were also found in mice infected with the S.E-AvrA− strain compared with those in the mice infected by the S.E-WT strain (Figures 6A,C,D). These data suggest that AvrA inhibits the JNK signaling pathway to decrease Beclin-1 expression and impair the autophagic response in vivo.
Figure 6.
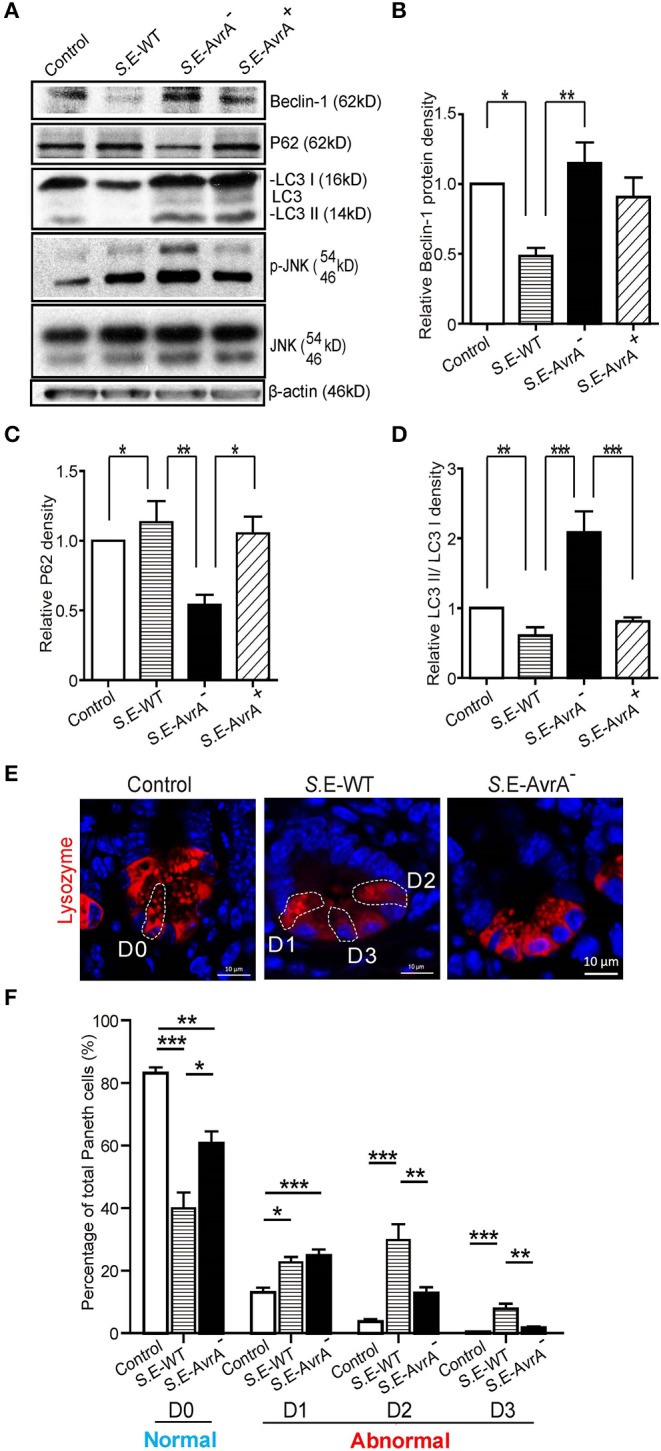
AvrA changes the levels of Beclin-1 and affects the function of Paneth cell granules of the ileal tissues in a mouse model. Wild-type Salmonella Enteritidis, AvrA mutant or AvrA-complemented strains were used to infect C57BL/6 mouse models (1.0 × 108 CFU per mouse, i.g., n = 6). At 8 h post-infection, the ileal tissue was harvested for immunoblotting and immunofluorescence staining. (A) Western blot shows the expression of Beclin-1, P62, LC3, and the JNK pathway markers in the ileal tissue after infection with the different S.E strains. The relative density of Beclin-1 (B), P62 (C), and LC3 II/LC3 I (D) was determined using Quantity One 4.6.2 software (Bio-Rad, Hercules, CA, USA). n = 5–6, *adjusted P < 0.05, **adjusted P < 0.01 by ANOVA test. (E) The representative images of the indirect immunofluorescence of the sections stained for lysozymes (42) in the ileal crypts of the C57BL/6 mice following infected with wild-type Salmonella Enteritidis or AvrA mutant strains. (F) The percentage of Paneth cells displaying a normal (D0) and abnormal (D1–D3) pattern of lysozyme expression. n = 5–6, *adjusted P < 0.05, **adjusted P < 0.01, ***adjusted P < 0.001 by ANOVA test.
Deficits in the autophagy pathway impair Paneth cell function in intestine (37, 43). Thus, we counted the number of Paneth cells using a previously reported method to stain lysozymes (37, 38). The abnormal Paneth cells were grouped as D1 (disordered), D2 (depleted), or D3 (diffuse) (Figure 6E). We found fewer normal Paneth cells (D0) in the mice infected with the S.E-WT strain than in the mice infected with the S.E-AvrA− strain (Figures 6E,F). Consequently, the number of abnormal Paneth cells (D1–D3) increased in the mice infected with the wild-type S.E strain (Figure 6F).
Discussion
In the current study, we report that the S. Enteritidis effector protein AvrA decreased Beclin-1 expression, thus impairing the autophagic response, for the benefit of Salmonella survival. AvrA-mediated regulation of host autophagy involves blocking the JNK signaling pathway, which was demonstrated in vitro and in vivo (Figure 7). The S. Enteritidis effector protein AvrA inhibited the JNK signaling pathways in epithelial cells, impaired autophagy by decreasing Beclin-1 expression at the protein level in vitro and in vivo, and affected the function of Paneth cell granules in a mouse model. Moreover, the JNK inhibitor SP600125 abolished the AvrA-reduced Beclin-1 protein expression. Together, these data suggest that the S.E effector AvrA inhibits the JNK/c-Jun/AP-1 signaling pathway to decrease Beclin-1 expression. In this way, Salmonella Enteritidis impairs the autophagic response to the benefit of the pathogen's survival. Moreover, AvrA affected the function of Paneth cell granules, potentially by inhibiting autophagy (Figure 7).
Figure 7.
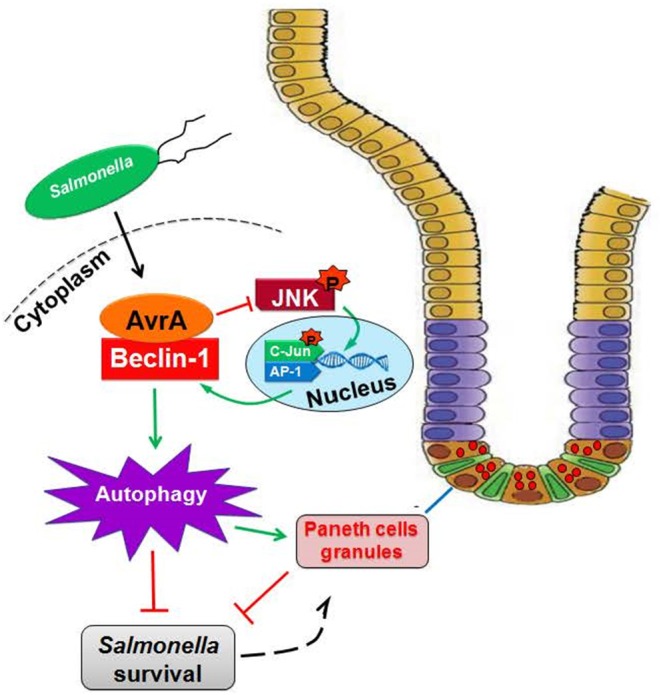
Working model of AvrA inhibition of Beclin-1-dependent autophagy. Salmonella Enteritidis AvrA inhibits the autophagic response by decreasing Beclin-1 protein. At the molecular level, AvrA interacts with Beclin-1. This process occurs through the inhibition of the JNK signaling pathway. At the cellular level, AvrA affects the function of Paneth cell granules by inhibiting autophagy.
The AvrA from S. Typhimurium is known as an anti-inflammatory effector that possesses acetyltransferase activity toward specific host MAPKKs and inhibits the host JNK/c-Jun/AP-1 and NF-κB pathways, but the role of AvrA from S. Enteritidis is less explored (8, 21–25). Here, we demonstrated that S. Enteritidis AvrA inhibited the autophagic response. Beclin-1 protein levels were reduced with AvrA present strain infection. In contrast, S.E-AvrA mutant strain infection maintained Beclin-1 protein expression. Earlier studies on the mechanism underlying the regulation of autophagy in cancer cells showed that the activation of the JNK pathway may be involved in the regulation of Beclin-1 expression, and the latter event could be responsible for the induction of the autophagic response (40). Blocking the JNK pathway might be the reason that AvrA decreases Beclin-1 expression. To verify our results, we used a specific JNK inhibitor SP600125 to block the JNK signaling pathway. After treatment with the JNK inhibitor SP600125, Beclin-1 protein expression did not differ between infection with and without AvrA. The results demonstrated that the S. Enteritidis effector AvrA inhibits the JNK/c-Jun/AP-1 signaling pathway to decrease Beclin-1 expression. Beclin-1 and JNK are also known to regulate apoptosis. Previous studies have demonstrated the effects of AvrA in inhibiting apoptosis and promoting proliferation in vitro and in vivo (21, 22). Taken together, our study has demonstrated a strategy that Salmonella used to impair the autophagic response to the benefit of the pathogen's survival.
Our data using AvrA WT and AvrA C186A mutant plasmids indicated that cysteine 186 is the key amino acid required for the AvrA regulation of Beclin-1 expression. We further demonstrated that the single mutation of Cys186 blocked JNK activity and abolished the AvrA-induced downregulation of Beclin-1.
In the current study, we observed that the S.E WT strain with AvrA expression and complementary strain induced weak autophagic activity, compared to the autophagic activity following S.E AvrA mtuant strain infection. Numerous lines of evidence indicate that Salmonella infection can activate robust host cell autophagy (44–46). This difference maybe due to the serotype difference. The existing research tends to use Salmonella Typhimurium to study the interaction between host autophagic activity and bacterial invasion. However, Salmonella Enteritidis, which belongs to another Salmonella serotype, could be much difference from Salmonella Typhimurium in host-adaptability, virulence, intracellular survival and so on (47–49).
Interestingly, our data showed that AvrA interacted with Beclin-1 and decreased the ubiquitination of Beclin-1 (Figure S1). These results suggested that AvrA decreased the ubiquitination of Beclin-1. A recent study identified S-phase kinase-associated protein 2 (SKP2) as E3 ligase that executes ubiquitination of Beclin-1, thus promoting its proteasomal degradation (50). Inhibition of SKP2 decreases Beclin-1 ubiquitination, decreases BECN1 degradation and enhances autophagic flux. A previous study also has shown that the ubiquitination of Beclin-1 enhances its association with Vps34 to promote Vps34 activity, which plays key role in activation of PI3KC3 complex at the initiation stage of autophagy process (51). AvrA may suppress Beclin-1 ubiquitination to inactivate Vps34 activity, leading to the suppression of autophagy. Certainly, this hypothesis requires further study.
Here, we highlighted the organoid system for investigating the host-bacterial interactions. Our previous studies have demonstrated that the intestinal organoid culture is a newly developed 3D system to study Salmonella infection (29, 52). Our data in the organoid system have shown that AvrA expressing bacteria reduce the levels of Beclin-1, thus suppressing autophagy in the host. In the future, human organoids could be used to further understand how bacterial effectors manipulate the host responses.
Paneth cells are specialized epithelial cells that are primarily located in the small intestine. The granules of Paneth cells contain AMPs–α-defensins, lysozyme, and secretory phospholipase A2 (53–55). Among these, lysozyme is a useful marker of the Paneth cell secretory granule (56). Our data showed that the normal expression pattern of the Paneth cells decreased in the mice infected with the AvrA present strain compared with that in the mice infected with the S.E-AvrA mutant strain. The ileum, after infection with the AvrA present strain, contained an increased proportion of Paneth cells with disorganized or diminished granules or exhibited diffuse cytoplasmic lysozyme staining. These findings suggest that AvrA impairs the autophagic response, and then, the autophagy affects the biosynthesis or quality control of the lysosomal pathway in the Paneth cell granules. Thus, there were fewer normal Paneth cells in the mice infected with the S.E-WT strain. Recently, a study showed that S. Typhimurium invaded Paneth cells, and the invasion was associated with an elevated number of LC3+ puncta in the Paneth cells (57). It is not clear whether S. Enteritidis interact with Paneth cells and affect autophagy through the AvrA effector. Alternatively, the Paneth cells with fewer granules might be a result of Paneth cell exhaustion (43), compensating for the changes elsewhere in the epithelium, due to the survival benefit of Salmonella caused by AvrA suppressing autophagy.
Taken together, our data reveal a new role for AvrA in S. Enteritidis in the reduction of Beclin-1 protein expression through the JNK pathway and the attenuation of the autophagic response in intestinal epithelial cells. Bacterial effector proteins paralyze or reprogram host cells to the benefit of the pathogens. Our findings indicate an important role of S. Enteritidis effector in reducing host protein as a strategy to suppress autophagy. Manipulating autophagic activity through the JNK pathway may be a novel therapeutic approach to treat infectious diseases.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by The University of Illinois at Chicago Committee on Animal Resources.
Author Contributions
YJ, ZL, YZ, CM, and RL: data acquisition, analysis and interpretation, and drafting of the manuscript. YJ, YZ, and JS: wrote the main manuscript text and prepared the figures. YX: statistical analysis and drafting of the manuscript. ZP, XJ, and XX: technical or material support and drafting of the manuscript. JS: study concept and design, critical revision of the manuscript for important intellectual content, and study supervision. All the authors reviewed the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This manuscript has been released as a preprint at Jiao et al. (58).
Glossary
Abbreviations
- AvrA C186A mutation
mutated at the key cysteine required for AvrA activity
- BSA
bovine serum albumin
- CFU
colony forming units
- DMEM
Dulbecco's modified Eagle's medium
- EDTA
ethylenediamine tetraacetic acid
- FBS
fetal bovine serum
- HBSS
Hank's balanced salt solution
- IP
immunoprecipitated
- JNK
c-Jun N-terminal kinase
- LB
Luria-Bertani
- Salmonella Enteritidis
S. Enteritidis, S.E, Salmonella enterica serovar Enteritidis
- Salmonella Typhimurium
Salmonella enterica serovar Typhimurium
- S.E-WT
Salmonella Enteritidis wild-type strain C50336
- S.E-AvrA−
Salmonella Enteritidis AvrA deletion mutant
- S. E-AvrA+
plasmid mediated complementary strain S.E-AvrA−/pAvrA+
- SPI-1
Salmonella pathogenicity island 1
- T3SS
type III secretion system
- ub-Beclin-1
ubiquitinated Beclin-1.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.00686/full#supplementary-material
AvrA decrease the ubiquitination of Beclin-1. The HCT116 cells were transfected with the indicated plasmids (200 ng/μl, 24 h incubation, n = 3) and were incubated for 2 h with the proteasome inhibitor MG262 (40 μmol/L). The total cell lysates were analyzed for ubiquitinated Beclin-1 by an immunoblot. The higher-molecular weight ubiquitinated Beclin-1 is indicated by bracket. The data shown are from a single experiment and are representative of three separate experiments.
References
- 1.Patrick ME, Adcock PM, Gomez TM, Altekruse SF, Holland BH, Tauxe RV, et al. Salmonella enteritidis infections, United States, 1985-1999. Emerg Infect Dis. (2004) 10:1–7. 10.3201/eid1001.020572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wright AP, Richardson L, Mahon BE, Rothenberg R, Cole DJ. The rise and decline in Salmonella enterica serovar Enteritidis outbreaks attributed to egg-containing foods in the United States, 1973-2009. Epidemiol Infect. (2016) 144:810–9. 10.1017/S0950268815001867 [DOI] [PubMed] [Google Scholar]
- 3.Scallan E, Hoekstra RM, Angulo FJ, Tauxe RV, Widdowson MA, Roy SL, et al. Foodborne illness acquired in the United States–major pathogens. Emerg Infect Dis. (2011) 17:7–15. 10.3201/eid1701.P11101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Majowicz SE, Musto J, Scallan E, Angulo FJ, Kirk M, O'Brien SJ, et al. The global burden of nontyphoidal Salmonella gastroenteritis. Clin Infect Dis. (2010) 50:882–9. 10.1086/650733 [DOI] [PubMed] [Google Scholar]
- 5.Steele-Mortimer O. The Salmonella-containing vacuole: moving with the times. Curr Opin Microbiol. (2008) 11:38–45. 10.1016/j.mib.2008.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Buchmeier NA, Heffron F. Inhibition of macrophage phagosome-lysosome fusion by Salmonella typhimurium. Infect Immun. (1991) 59:2232–8. 10.1128/IAI.59.7.2232-2238.1991 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Knodler LA, Finlay BB. Salmonella and apoptosis: to live or let die? Microbes Infect. (2001) 3:1321–6. 10.1016/S1286-4579(01)01493-9 [DOI] [PubMed] [Google Scholar]
- 8.Jones RM, Wu H, Wentworth C, Luo L, Collier-Hyams L, Neish AS. Salmonella AvrA coordinates suppression of host immune and apoptotic defenses via JNK pathway blockade. Cell Host Microbe. (2008) 3:233–44. 10.1016/j.chom.2008.02.016 [DOI] [PubMed] [Google Scholar]
- 9.Owen KA, Meyer CB, Bouton AH, Casanova JE. Activation of focal adhesion kinase by Salmonella suppresses autophagy via an Akt/mTOR signaling pathway and promotes bacterial survival in macrophages. PLoS Pathog. (2014) 10:e1004159. 10.1371/journal.ppat.1004159 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ganesan R, Hos NJ, Gutierrez S, Fischer J, Stepek JM, Daglidu E, Kronke M, et al. Salmonella Typhimurium disrupts Sirt1/AMPK checkpoint control of mTOR to impair autophagy. PLoS Pathog. (2017) 13:e1006227. 10.1371/journal.ppat.1006227 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jiao Y, Sun J. Bacterial manipulation of autophagic responses in infection and inflammation. Front Immunol. (2019) 10:2821. 10.3389/fimmu.2019.02821 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Randow F, Munz C. Autophagy in the regulation of pathogen replication and adaptive immunity. Trends Immunol. (2012) 33:475–87. 10.1016/j.it.2012.06.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Li S, Wandel MP, Li F, Liu Z, He C, Wu J, et al. Sterical hindrance promotes selectivity of the autophagy cargo receptor NDP52 for the danger receptor galectin-8 in antibacterial autophagy. Sci Signal. (2013) 6:ra9. 10.1126/scisignal.2003730 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gomes LC, Dikic I. Autophagy in antimicrobial immunity. Mol Cell. (2014) 54:224–33. 10.1016/j.molcel.2014.03.009 [DOI] [PubMed] [Google Scholar]
- 15.Mizushima N, Yoshimori T, Ohsumi Y. The role of Atg proteins in autophagosome formation. Ann Rev Cell Dev Biol. (2011) 27:107–32. 10.1146/annurev-cellbio-092910-154005 [DOI] [PubMed] [Google Scholar]
- 16.Kang R, Zeh HJ, Lotze MT, Tang D. The Beclin 1 network regulates autophagy and apoptosis. Cell Death Diff. (2011) 18:571–80. 10.1038/cdd.2010.191 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liang XH, Jackson S, Seaman M, Brown K, Kempkes B, Hibshoosh H, et al. Induction of autophagy and inhibition of tumorigenesis by beclin 1. Nature. (1999) 402:672–6. 10.1038/45257 [DOI] [PubMed] [Google Scholar]
- 18.Dalbey RE, Kuhn A. Protein traffic in Gram-negative bacteria–how exported and secreted proteins find their way. FEMS Microbiol Rev. (2012) 36:1023–45. 10.1111/j.1574-6976.2012.00327.x [DOI] [PubMed] [Google Scholar]
- 19.Ellermeier JR, Slauch JM. Adaptation to the host environment: regulation of the SPI1 type III secretion system in Salmonella enterica serovar Typhimurium. Curr Opin Microbiol. (2007) 10:24–9. 10.1016/j.mib.2006.12.002 [DOI] [PubMed] [Google Scholar]
- 20.Figueira R, Holden DW. Functions of the Salmonella pathogenicity island 2 (SPI-2) type III secretion system effectors. Microbiology. (2012) 158:1147–61. 10.1099/mic.0.058115-0 [DOI] [PubMed] [Google Scholar]
- 21.Ye Z, Petrof EO, Boone D, Claud EC, Sun J. Salmonella effector AvrA regulation of colonic epithelial cell inflammation by deubiquitination. Am J Pathol. (2007) 171:882–92. 10.2353/ajpath.2007.070220 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Collier-Hyams LS, Zeng H, Sun J, Tomlinson AD, Bao ZQ, Chen H, et al. Cutting edge: Salmonella AvrA effector inhibits the key proinflammatory, anti-apoptotic NF-kappa B pathway. J Immunol. (2002) 169:2846–50. 10.4049/jimmunol.169.6.2846 [DOI] [PubMed] [Google Scholar]
- 23.Du F, Galan JE. Selective inhibition of type III secretion activated signaling by the Salmonella effector AvrA. PLoS Pathog. (2009) 5:e1000595. 10.1371/journal.ppat.1000595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Naydenov NG, Hopkins AM, Ivanov AI. C-Jun N-terminal kinase mediates disassembly of apical junctions in model intestinal epithelia. Cell Cycle. (2009) 8:2110–21. 10.4161/cc.8.13.8928 [DOI] [PubMed] [Google Scholar]
- 25.Lin Z, Zhang YG, Xia Y, Xu X, Jiao X, Sun J. Salmonella enteritidis effector AvrA stabilizes intestinal tight junctions via the JNK pathway. J Biol Chem. (2016) 291:26837–49. 10.1074/jbc.M116.757393 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sun Q, Fan W, Chen K, Ding X, Chen S, Zhong Q. Identification of Barkor as a mammalian autophagy-specific factor for Beclin 1 and class III phosphatidylinositol 3-kinase. Proc Natl Acad Sci USA. (2008) 105:19211–6. 10.1073/pnas.0810452105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.McCormick BA, Colgan SP, Delp-Archer C, Miller SI, Madara JL. Salmonella typhimurium attachment to human intestinal epithelial monolayers: transcellular signalling to subepithelial neutrophils. J Cell Biol. (1993) 123:895–907. 10.1083/jcb.123.4.895 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wang N, Zhang H, Zhang BQ, Liu W, Zhang Z, Qiao M, et al. Adenovirus-mediated efficient gene transfer into cultured three-dimensional organoids. PLoS ONE. (2014) 9:e93608. 10.1371/journal.pone.0093608 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhang YG, Wu S, Xia Y, Sun J. Salmonella-infected crypt-derived intestinal organoid culture system for host-bacterial interactions. Physiol Rep. (2014) 2:e12147. 10.14814/phy2.12147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wu S, Liao AP, Xia Y, Li YC, Li JD, Sartor RB, et al. Vitamin D receptor negatively regulates bacterial-stimulated NF-kappaB activity in intestine. Am J Pathol. (2010) 177:686–97. 10.2353/ajpath.2010.090998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lu R, Wu S, Liu X, Xia Y, Zhang YG, Sun J. Chronic effects of a Salmonella type III secretion effector protein AvrA in vivo. PLoS ONE. (2010) 5:e10505. 10.1371/journal.pone.0010505 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sun J, Hobert ME, Rao AS, Neish AS, Madara JL. Bacterial activation of beta-catenin signaling in human epithelia. Am J Physiol. (2004) 287:G220–7. 10.1152/ajpgi.00498.2003 [DOI] [PubMed] [Google Scholar]
- 33.Barthel M, Hapfelmeier S, Quintanilla-Martinez L, Kremer M, Rohde M, Hogardt M, et al. Pretreatment of mice with streptomycin provides a Salmonella enterica serovar Typhimurium colitis model that allows analysis of both pathogen and host. Infect Immun. (2003) 71:2839–58. 10.1128/IAI.71.5.2839-2858.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.McCullough ML, Zoltick ES, Weinstein SJ, Fedirko V, Wang M, Cook NR, et al. Circulating vitamin D and colorectal cancer risk: an international pooling project of 17 cohorts. J Natl Cancer Inst. (2018) 111:158–69. 10.1093/jnci/djy087 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Boyle EC, Brown NF, Finlay BB. Salmonella enterica serovar Typhimurium effectors SopB, SopE, SopE2 and SipA disrupt tight junction structure and function. Cell Microbiol. (2006) 8:1946–57. 10.1111/j.1462-5822.2006.00762.x [DOI] [PubMed] [Google Scholar]
- 36.Sun J, Hobert ME, Duan Y, Rao AS, He TC, Chang EB, et al. Crosstalk between NF-kappaB and beta-catenin pathways in bacterial-colonized intestinal epithelial cells. Am J Physiol Gastrointest Liver Physiol. (2005) 289:G129–37. 10.1152/ajpgi.00515.2004 [DOI] [PubMed] [Google Scholar]
- 37.Wu S, Zhang YG, Lu R, Xia Y, Zhou D, Petrof EO, et al. Intestinal epithelial vitamin D receptor deletion leads to defective autophagy in colitis. Gut. (2015) 64:1082–94. 10.1136/gutjnl-2014-307436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Cadwell K, Liu JY, Brown SL, Miyoshi H, Loh J, Lennerz JK, et al. A key role for autophagy and the autophagy gene Atg16l1 in mouse and human intestinal Paneth cells. Nature. (2008) 456:259–63. 10.1038/nature07416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Jiang P, Mizushima N. LC3- and p62-based biochemical methods for the analysis of autophagy progression in mammalian cells. Methods. (2015) 75:13–8. 10.1016/j.ymeth.2014.11.021 [DOI] [PubMed] [Google Scholar]
- 40.Li DD, Wang LL, Deng R, Tang J, Shen Y, Guo JF, et al. The pivotal role of c-Jun NH2-terminal kinase-mediated Beclin 1 expression during anticancer agents-induced autophagy in cancer cells. Oncogene. (2009) 28:886–98. 10.1038/onc.2008.441 [DOI] [PubMed] [Google Scholar]
- 41.Vijay-Kumar M, Wu H, Jones R, Grant G, Babbin B, King TP, et al. Flagellin suppresses epithelial apoptosis and limits disease during enteric infection. Am J Pathol. (2006) 169:1686–700. 10.2353/ajpath.2006.060345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Duan Y, Liao AP, Kuppireddi S, Ye Z, Ciancio MJ, Sun J. beta-Catenin activity negatively regulates bacteria-induced inflammation. Lab Invest. (2007) 87:613–24. 10.1038/labinvest.3700545 [DOI] [PubMed] [Google Scholar]
- 43.Deretic V, Master S, Singh S. Autophagy gives a nod and a wink to the inflammasome and Paneth cells in Crohn's disease. Dev Cell. (2008) 15:641–2. 10.1016/j.devcel.2008.10.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Benjamin JL, Sumpter R, Jr, Levine B, Hooper LV. Intestinal epithelial autophagy is essential for host defense against invasive bacteria. Cell Host Microbe. (2013) 13:723–34. 10.1016/j.chom.2013.05.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wang L, Yan J, Niu H, Huang R, Wu S. Autophagy and Ubiquitination in Salmonella infection and the related inflammatory responses. Front Cell Infect Microbiol. (2018) 8:78. 10.3389/fcimb.2018.00078 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Schulthess J, Pandey S, Capitani M, Rue-Albrecht KC, Arnold I, Franchini F, et al. The Short Chain Fatty Acid Butyrate Imprints An Antimicrobial Program In Macrophages. Immunity. (2019) 50:432–45.e7. 10.1016/j.immuni.2018.12.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Saleh S, Van Puyvelde S, Staes A, Timmerman E, Barbe B, Jacobs J, et al. Salmonella Typhi, Paratyphi A, Enteritidis and Typhimurium core proteomes reveal differentially expressed proteins linked to the cell surface and pathogenicity. PLoS Negl Trop Dis. (2019) 13:e0007416. 10.1371/journal.pntd.0007416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Khan SB, Khan MA, Ahmad I, Ur Rehman T, Ullah S, Dad R, et al. Phentotypic, gentotypic antimicrobial resistance and pathogenicity of Salmonella enterica serovars Typimurium and Enteriditis in poultry and poultry products. Microb Pathog. (2019) 129:118–24. 10.1016/j.micpath.2019.01.046 [DOI] [PubMed] [Google Scholar]
- 49.Lamas A, Miranda JM, Regal P, Vazquez B, Franco CM, Cepeda A. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol Res. (2018) 206:60–73. 10.1016/j.micres.2017.09.010 [DOI] [PubMed] [Google Scholar]
- 50.Gassen NC, Niemeyer D, Muth D, Corman VM, Martinelli S, Gassen A, et al. SKP2 attenuates autophagy through Beclin1-ubiquitination and its inhibition reduces MERS-Coronavirus infection. Nat Commun. (2019) 10:5770. 10.1038/s41467-019-13659-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Xia P, Wang S, Du Y, Zhao Z, Shi L, Sun L, et al. WASH inhibits autophagy through suppression of Beclin 1 ubiquitination. EMBO J. (2013) 32:2685–96. 10.1038/emboj.2013.189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Zhang YG, Zhu X, Lu R, Messer JS, Xia Y, Chang EB, et al. Intestinal epithelial HMGB1 inhibits bacterial infection via STAT3 regulation of autophagy. Autophagy. (2019) 5:1935–53. 10.1080/15548627.2019.1596485 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Koslowski MJ, Beisner J, Stange EF, Wehkamp J. Innate antimicrobial host defense in small intestinal Crohn's disease. Int J Med Microbiol. (2010) 300:34–40. 10.1016/j.ijmm.2009.08.011 [DOI] [PubMed] [Google Scholar]
- 54.Wehkamp J, Wang G, Kubler I, Nuding S, Gregorieff A, Schnabel A, et al. The paneth cell alpha-defensin deficiency of ileal Crohn's disease is linked to Wnt/Tcf-4. J Immunol. (2007) 179:3109–18. 10.4049/jimmunol.179.5.3109 [DOI] [PubMed] [Google Scholar]
- 55.Menard S, Forster V, Lotz M, Gutle D, Duerr CU, Gallo RL, et al. Developmental switch of intestinal antimicrobial peptide expression. J Exp Med. (2008) 205:183–93. 10.1084/jem.20071022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wang D, Peregrina K, Dhima E, Lin EY, Mariadason JM, Augenlicht LH. Paneth cell marker expression in intestinal villi and colon crypts characterizes dietary induced risk for mouse sporadic intestinal cancer. Proc Natl Acad Sci USA. (2011) 108:10272–7. 10.1073/pnas.1017668108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Bel S, Pendse M, Wang Y, Li Y, Ruhn KA, Hassell B, et al. Paneth cells secrete lysozyme via secretory autophagy during bacterial infection of the intestine. Science. (2017) 357:1047–52. 10.1126/science.aal4677 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Jiao Y, Zhang Y, Lin Z, Lu R, Xia Y, Pan Z, et al. Salmonella enteritidis effector AvrA suppresses autophagy by reducing Beclin-1 protein. bioRxiv. (2020). 10.1101/2020.01.13.905596 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
AvrA decrease the ubiquitination of Beclin-1. The HCT116 cells were transfected with the indicated plasmids (200 ng/μl, 24 h incubation, n = 3) and were incubated for 2 h with the proteasome inhibitor MG262 (40 μmol/L). The total cell lysates were analyzed for ubiquitinated Beclin-1 by an immunoblot. The higher-molecular weight ubiquitinated Beclin-1 is indicated by bracket. The data shown are from a single experiment and are representative of three separate experiments.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.