

Abstract
Objective:
Anorexia nervosa (AN) has the highest mortality rate of any psychiatric condition, yet the pathophysiology of this disorder and its primary symptom—extreme dietary restriction—remains poorly understood. In states of hunger relative to satiety, the rewarding value of food stimuli normally increases to promote eating, yet individuals with AN avoid food despite emaciation. This study aimed to examine potential neural insensitivity to these effects of hunger in AN.
Method:
At two counterbalanced visits, one after a 16-hour fast and one after a standardized meal, 26 women remitted from AN (RAN; to avoid the confounding effects of malnutrition) and 22 matched control women (CW) received tastes of sucrose and water while functional MRI data were acquired. Within a network-of-interest responsible for food valuation and transforming taste signals into motivation to eat, we compared groups across conditions on blood-oxygen-level-dependent (BOLD) signal and task-based functional connectivity.
Results:
Subjects had similar BOLD responses to sucrose and water tastants. A Group x Condition interaction in the ventral caudal putamen indicated that hunger had opposite effects on tastant response in CW and RAN participants: CW showed an increase, and RAN showed a decrease, in BOLD response when hungry. Hunger had a similar opposite effect on insula-to-ventral caudal putamen functional connectivity in RAN compared with CW. Exploratory analyses indicated that lower caudate response to tastants when hungry was associated with higher harm avoidance among RAN participants.
Conclusions:
Reduced recruitment of neural circuitry that translates taste stimulation to motivated eating behavior when hungry may facilitate food avoidance and prolonged periods of extremely restricted intake in AN.
Keywords: anorexia nervosa, hunger, taste, fMRI, reward processing, motivation
INTRODUCTION
Individuals with anorexia nervosa (AN) are able to severely restrict food consumption and maintain an extremely low weight 1. Because there are no proven treatments that normalize core symptoms in adults with AN, this is often a chronic disorder resulting in high morbidity and mortality. In healthy humans, hunger is dysphoric and increases reward salience and subjective value of food to drive consumption 2. In contrast, individuals with AN often describe eating as anxiogenic, and food refusal may reduce dysphoric mood 3. Do individuals with AN have an altered response to the motivating signals of hunger? While individuals with AN have alterations in neural mechanisms coding motivation, salience, and valuation of food when ill and after weight restoration 4, few studies 5 have systematically examined whether there is altered functioning in this circuitry when hungry that might persist after recovery. Understanding the mechanisms contributing to the neurobiology of self-starvation in AN could identify new treatment targets.
One method of identifying food-related neural mechanisms coding motivation, salience, and valuation in humans is to measure brain response to palatable tastants, such as sucrose, using neuroimaging 6. This approach has identified a neural circuit in humans 7 that extends beyond pure gustatory chemosensory processing to include regions involved in motivation and reward processing, and that maps onto analogous circuits in rodents 8 and non-human primates 9. Specifically, tastes of sucrose signals are transmitted from sweet taste receptors in the tongue through the brainstem and thalamus to the primary gustatory cortex in the insula. In humans, the anterior insula receives chemosensory taste input from the mid-insula 10. Recent studies suggest that the insula taste cortex identifies sweetness, whereas the amygdala, which is involved in processing emotions, codes the valence, specifies its hedonic value, and elicits execution of selective behaviors 8. The hypothalamus, a homeostatic center, regulates metabolic processes, including hunger and food intake, with motivation-reward systems associated with the hedonic drive to eat 11. In primates, the orbitofrontal cortex processes the reward value of taste 7, while the ventromedial prefrontal cortex acts as a visceromotor area and governs the hypothalamus, amygdala, and insula. Ultimately, taste information is integrated with reward value and homeostatic drives 7, 11, via projections from the anterior insula and amygdala to the striatum 9, to guide motivated eating behavior (see Figure 4A and supplement). In healthy individuals, hunger, compared to satiety, increases brain response in these reward, attention, and motivation regions in response to palatable foods 6, 7, 12, 13.
Figure 4.
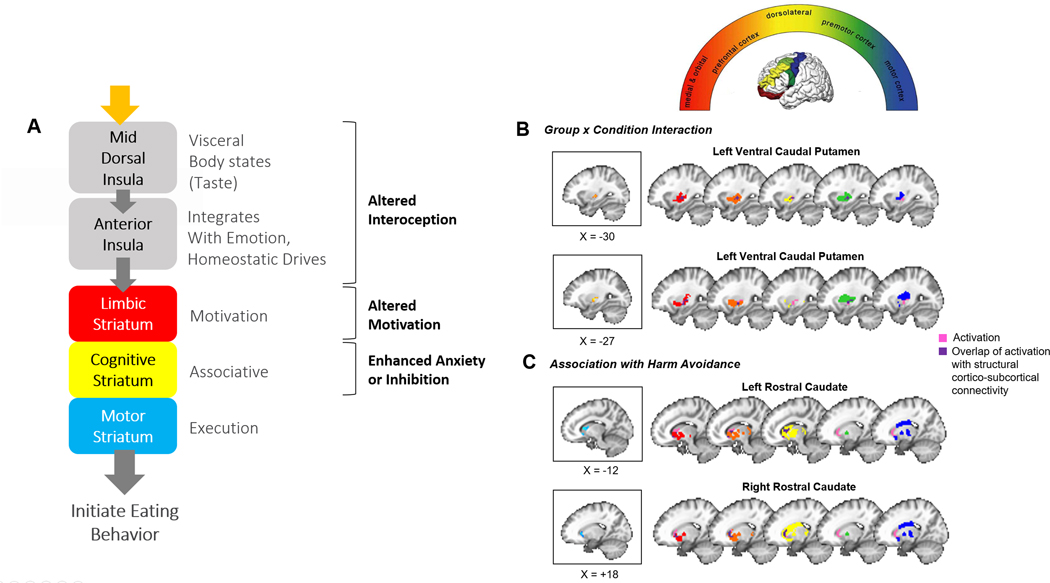
A) Schematic diagram of insula-striatal pathways. The mid insula relays somato- and viscero-sensory signals to the anterior insula and striatum. The anterior insula projects along the ventromedial axis of the striatum, including the rostral ventral striatum and ventral caudal putamen. Through feed-forward connections, DA-mediated information progresses from the limbic to cognitive to sensorimotor areas of the striatum 57 to generate action-selection related to the mediation and regulation of goal-directed behavior, such as the consumption of palatable food. B & C) The top panel within B reflects the rostro-caudal gradient of striatum-to-frontal-cortex structural connectivity as identified in healthy adults 27. Areas of the striatum are color-coded by their ultimate cortical projection targets, as indicated within the semicircle. The remaining panels depict BOLD activation from the current study in the context of the above identified structural circuits in healthy adults. Reference slices depicting significant clusters identified for each analysis in the current study (from Figure 2) are shown in boxes on the left. Colored areas shown on brain slices on the right reflect the gradients from the top semicircular panel. Overlap between clusters from our main analyses and these structural projection areas is shown in dark purple. Ventral caudal putamen clusters showing Group (AN, CW) x Condition (hungry, fed) interactions overlap primarily with areas of the putamen that have been shown previously in healthy adults to project to limbic (red, orange) and pre-motor (green) associative cortices. Rostral caudate clusters showing associations with harm avoidance in the RAN group overlap with areas of the caudate that have been shown previously to project to limbic (red, orange) and cognitive control-related dorsolateral prefrontal cortices (yellow). Structural connectivity maps adapted with permission from Draganski et al 2008 27.
This study used a palatable taste task with functional magnetic resonance imaging (fMRI) to investigate the response to hungry and fed states in the food reward-motivation circuit. We studied women remitted from AN (RAN) to avoid the confounding effects of malnutrition on this circuitry, in comparison to healthy control women (CW). Our previous studies 14, 15 supported our hypothesis that RAN relative to CW would show a diminished response to taste of sucrose in the insula and striatum when in a hungry compared to a fed state, suggesting that hunger may not generate signals to motivate eating7 in AN. We also considered whether AN might have an exaggerated response to being fed (enhanced satiety) and a normal response to hunger. Thus, we performed a Group x Condition X Tastant interaction analysis in the insula, striatum, amygdala, and medial OFC to test these hypotheses in regions that integrate sensory/hedonic aspects of taste and interoceptive awareness in the service of homeostasis.
METHODS
Subjects
Twenty-six RAN women with no history of binge-eating behavior (18 restricting only, 8 with purging behaviors) were compared to 22 age- and weight-matched healthy CW (see Table 1). RAN participants met DSM-IV-TR criteria for AN in the past 1, but for at least one year prior to participation did not endorse pathological eating behavior or cognitions and maintained above 85% of average body weight, weight stability (within 3 kg), and regular menstrual cycles 14. Individuals were excluded from study for the following: history of alcohol or drug abuse or dependence 3 months prior to study; meeting current diagnostic criteria for major depressive disorder, any anxiety disorder, or obsessive compulsive disorder (lifetime diagnosis was not exclusionary); medical or neurologic concerns; taking any psychoactive medication in the past 3 months; and MRI contraindications. The study was approved by the Institutional Review Board of the University of California, San Diego (UCSD), and all participants provided written informed consent and received compensation. See Supplement for details regarding participants and assessment tools.
Table 1.
Participant demographics and characteristics. Entries are of the form mean (standard deviation). CW: healthy control women; RAN: women remitted from anorexia nervosa; BMI: body mass index (kg/m2); WASI: Wechsler Abbreviated Scale of Intelligence; MDD: major depressive disorder; OCD: obsessive-compulsive disorder; STAI: State-Trait Anxiety Inventory; TCI: Temperament and Character Inventory; BDI: Beck Depression Inventory. One CW did not complete clinical assessments. (see Supplement)
| Characteristic | CW (N = 22) | RAN (N = 26) | p | |||
|---|---|---|---|---|---|---|
| Scanner | Signa Excite | 10 | 14 | n.s. | ||
| MR 750 | 12 | 12 | ||||
| Demographics | Age | 25.7 | (6.3) | 26.2 | (6.6) | n.s. |
| BMI | 22.0 | (2.1) | 21.9 | (1.7) | n.s. | |
| Lowest BMI | 20.4 | (1.5) | 14.7 | (1.5) | <0.01 | |
| Duration of illness (months) | -- | -- | 70.8 | (61.3) | ||
| Duration of recovery (months) | -- | -- | 67.0 | (60.5) | ||
| Education | 15.7 | (1.3) | 16.6 | (2.8) | n.s. | |
| WASI FSIQ estimate | 111.5 | (10.9) | 112.9 | (11.8) | n.s. | |
| Estradiol (pg/mL) | 12.8 | (7.7) | 12.5 | (5.9) | n.s. | |
| Lifetime Diagnosis (No.) | MDD | 0 | 15 | |||
| Any anxiety disorder | 1 | 10 | ||||
| OCD | 0 | 4 | ||||
| Past substance abuse/dependence (No.) | Alcohol | 0 | 3 | |||
| Cannabis | 0 | 1 | ||||
| Clinical Assessments | STAI State Anxiety | 24.6 | (5.9) | 29.7 | (9.2) | 0.03 |
| STAI Trait Anxiety | 23.9 | (1.0) | 28.7 | (1.4) | 0.01 | |
| TCI Harm Avoidance | 7.2 | (5.4) | 11.2 | (5.95) | 0.02 | |
| BDI | 1.0 | (3.4) | 2.0 | (2.4) | 0.25 | |
Experimental Design
Subjects were housed and meals were provided by the UCSD Clinical & Translational Research Institute for 72 hours to ensure dietary compliance. On day 1, subjects were evaluated and consumed the same standardized meals (calculated as 30 kcal/kg). On days 2 and 3, participants performed a taste task 14, 15 during fMRI on two visits scheduled 24 hours apart (see Supplement; Table S1). For the “hungry” condition, participants fasted for 16 hours prior to scanning, with ad libitum water permitted. During the “fed” condition, participants consumed standardized meals (calculated as 30 kcal/kg) on the day prior to study, as well as a weight-adjusted standardized breakfast (30% of overall daily caloric needs, approximating 450–500 kcal; 53% carbohydrates, 32% fat, and 15% protein) two hours before the 9:00 am scan. Study visit order on days 2 and 3 were randomized across participants and scheduled in the early follicular menstrual phase. Participants were scanned on one of two 3T scanners, and each participant was scanned on the same scanner for both visits 16. During the taste task, participants received pseudorandom delivery of 1.0 cc of 10% sucrose solution or ionic water over 80 trials (see Supplement). These solutions were chosen to enable comparison of current findings with those of our previous work that had limited control of food intake prior to scanning 14, 15. Participants also provided self-report ratings of hunger and thirst at both pre-scan and post-scan, and tastant pleasantness post-scan, for each imaging session (Figure 1).
Figure 1.
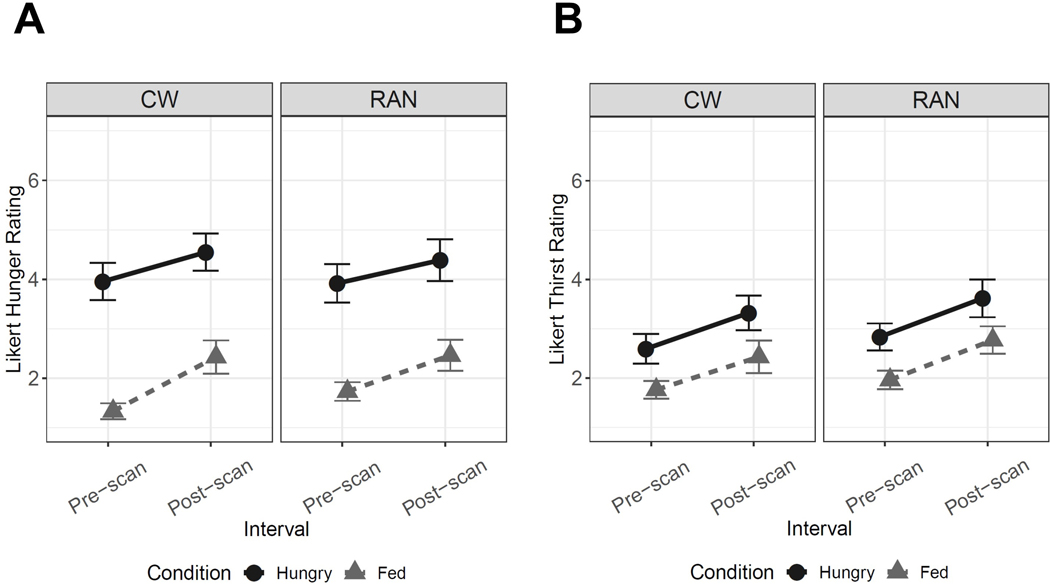
Line graphs reflecting self-report Likert visual analog scale values for a pre- and post-scan measure of hunger and thirst. A) We compared groups on hunger and thirst ratings across conditions and timepoints using a Group x Condition (hungry, fed) x Interval (pre-scan, post-scan) linear mixed effects model with subject as a random effect. For hunger, there was a main effect of Condition, F(1,134)=181.37, p<0.001, with post-hoc analyses suggesting all participants reported greater hunger during the hungry condition relative to fed condition, t(134)=13.47, p<0.001. There was a main effect of Interval, F(1,134)=17.59, p<0.001, with post-hoc analyses suggesting all participants reported greater hunger post-scan relative to pre-scan, t(134)=4.26, p<0.001. However, there was no main effect of Group, and interactions were not statistically significant. B) For thirst, similar effects were observed: There was a main effect of Condition, F(1,134)=40.55, p<0.001, with all participants reporting greater thirst in the hungry relative to fed condition, t(134)=6.27, p<0.001, and a main effect of Interval, F(1,134)=26.93, p<0.001, with all participants reporting greater thirst post-scan relative to pre-scan, t(134)=5.15, p<0.001. However, there was no main effect of Group, and interactions were not statistically significant.
Image Analysis
Following preprocessing (see Supplement), statistical analyses were performed using a generalized linear model (GLM), with individual events modeled using AFNI’s SPMG3 function. Six motion parameters (3 rotations and 3 translations) were used as nuisance regressors to account for motion artifact.
Task-Based Activation.
To examine whether RAN and CW differed in brain response to palatable tastants when hungry and fed, we employed a Group (RAN, CW) x Condition (Hungry, Fed) x Tastant (Sucrose, Water) linear mixed effects (LME) analysis. Group, Condition, and Tastant were treated as fixed effects. We designed this study to match our previous comparisons of sucrose and water 14 since we reasoned that, when hungry, RAN would have less response to a primary reward like sucrose. However, our initial analyses unexpectedly demonstrated no significant interactions for Tastant (i.e., no Group x Condition x Tastant, Group x Tastant, or Condition x Tastant). Therefore, we tested a simplified model that included the Group x Condition interaction (collapsed across Tastants) and the main effect of Tastant. Subject was treated as a random effect and nested within scanner, as recommended when multiple scanners are employed for data collection 17.
To improve power and reduce an inflated false discovery rate, primary analyses were restricted to a single mask (Figure S1) comprising a well-defined taste and motivation neurocircuit 7, 15: bilateral insula, amygdala, ventral striatum (nucleus accumbens and the most ventral parts of both the putamen and caudate 18), dorsal caudate, and putamen. The orbitofrontal cortex was excluded due to substantial susceptibility artifact. Intrinsic smoothness was estimated using the spatial autocorrelation function (acf) option in AFNI’s 3dFWHMx. Minimum cluster sizes were calculated with AFNI’s 3dClustSim to guard against false positives. For both ROI and whole-brain analyses, minimum cluster sizes corresponded to a voxel-wise probability of p<0.001 and a cluster wise probability of α=0.05 (two-sided) to correct for multiple comparisons. Exploratory voxel-wise analysis were also performed (see Supplement; Table S2).
Task-Based Functional Connectivity.
To follow-up on results from the analyses described above, we conducted a generalized psychophysiological interaction analysis (gPPI; 19). This analysis assessed the influence of hungry and fed states on group differences in functional connectivity within gustatory-reward circuitry during taste processing. Functionally-relevant seed regions were identified across all participants for the main effect of condition. This data-driven approach avoids circularity by accounting for the main effects of task condition but isolating effects distinct from the condition itself 19, 20 (see Supplement for details).
Relationship to Clinical Variables.
Within-group, within-visit exploratory voxel-wise Huber robust regressions 21 conducted in R examined associations of BOLD response for tastants (sucrose and water combined) with current BMI, age, harm avoidance, and trait anxiety. Additional analyses in RAN included lowest lifetime post-pubertal BMI, illness duration, and duration of remission. Significant clusters within our search region mask were identified using AFNI’s 3dClustSim for small volume correction, with a peak voxel of p<0.01. Results were Bonferroni-corrected for 4 experimental conditions and 4 clinical measures in CW (p<0.003) and for 6 clinical measures in RAN (p<0.002).
RESULTS
Demographics and Assessments.
CW and RAN had similar BMI, age, years of education, IQ, and depression scores (Table 1). RAN endorsed higher levels of anxiety and harm avoidance. Both CW and RAN reported significantly greater hunger during the hungry condition relative to the fed condition, and groups did not differ on ratings of hunger or thirst (Figure 1), or tastant pleasantness (Figure S2). Participants rated water as slightly more pleasant than the sucrose solution (see Supplement).
ROI Analysis.
At a voxel-wise p<0.001 (corrected α=0.05), a main effect of Condition (hungry>fed), seemingly driven by a greater response to hunger in CW, was found in the left ventral striatum (Figure S3, Table S3). No main effect of Tastant was observed. A Group x Condition interaction was detected in two clusters within the left ventral caudal putamen (Figure 2A, Table S3). Within-group post-hoc analyses revealed that in both clusters, CW were significantly more responsive to tastants when hungry versus fed. In contrast, RAN were significantly less responsive to tastants when hungry versus fed. Between-group post-hoc analyses indicated that when hungry, RAN showed lower response to tastants than CW. RAN and CW responses did not differ when fed. Using a less stringent threshold (voxel-wise p<0.01, corrected α=0.05), a similar Group x Condition interaction driven by reduced tastant response in RAN when hungry was detected in the left anterior insula (Table S3 Figure S4). A post-hoc analysis within the RAN group determined that AN subtype and lifetime history of depression or anxiety did not significantly contribute to our findings (see Supplement).
Figure 2.
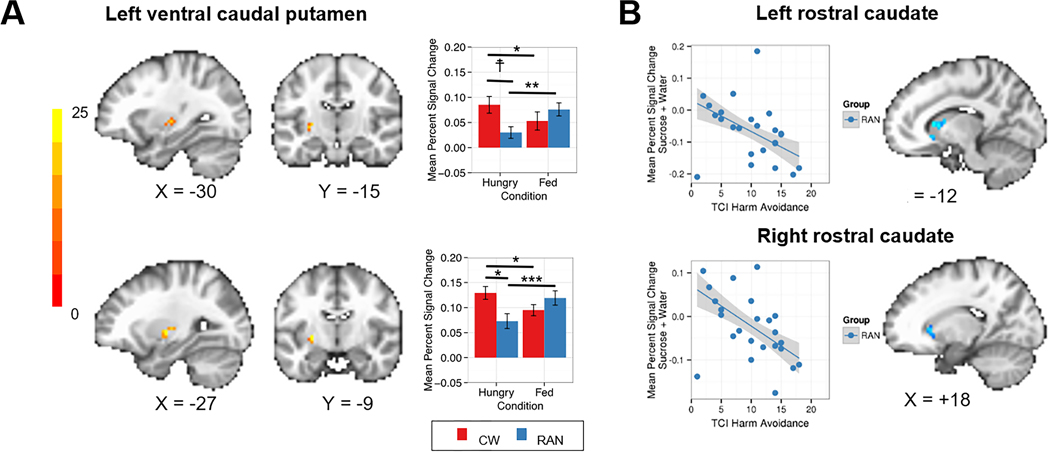
A) Linear mixed effect results showing two separate clusters demonstrating an interaction of Group (CW, RAN) by Condition (hungry, fed) in response to tastants (sucrose and water combined) within the left ventral caudal putamen (top peak coordinate: x=−30, y=−15, z=0; bottom peak coordinate: x=−27, y=−9, z=0). CW were significantly more responsive to tastants when hungry versus fed (ps<0.037). In contrast, RAN were significantly less responsive to tastants when hungry versus fed (ps<0.001). When hungry, RAN showed lower response to tastants than CW in both clusters, but this finding was statistically significant within only one cluster (shown in the bottom panel, p=0.035). Intrinsic smoothness was estimated using the spatial autocorrelation function (acf) option in AFNI’s 3dFWHMx. Minimum cluster sizes were calculated with AFNI’s 3dClustSim to guard against false positives (voxel-wise p<0.001, α=0.05). B) Plot demonstrating statistically significant relationships between the BOLD percent signal change response to tastants when hungry and harm avoidance within the bilateral rostral caudate (left peak coordinate: x=−12, y=+18, z=+6; right peak coordinate: x=+18, y=+24, z=+3) for RAN using Huber robust regression (left: t=−3.46, p=0.017; right: t=−3.94, p=0.009). CW: healthy control women; RAN: women remitted from anorexia nervosa; ☨p<0.1; *p<0.05; **p<0.01; ***p<0.001.
Task-Based Functional Connectivity Analysis.
Seed regions were derived from the main effect (across all subjects) of Condition and corresponded to the peak coordinates of two clusters: 1) right ventral caudal putamen (x=+33, y=−9, z=0), and 2) right dorsal mid-insula (x=+36, y=+6, z=0).
Group x Condition interactions indicated that metabolic state (i.e., hungry, fed) modulated group differences in functional connectivity between: 1) the right dorsal mid-insula seed and left ventral caudal putamen, right dorsal rostral putamen, and left anterior insula targets (Table S4, Figure 3A), and 2) the right ventral caudal putamen seed and left ventral caudal putamen, right dorsal rostral putamen, and right amygdala targets (Table S4 Figure 3B). Post-hoc analyses indicated that CW functional connectivity to tastants was greater when hungry than when fed, whereas RAN functional connectivity to tastants was lower when hungry than when fed (Table S4).
Figure 3.
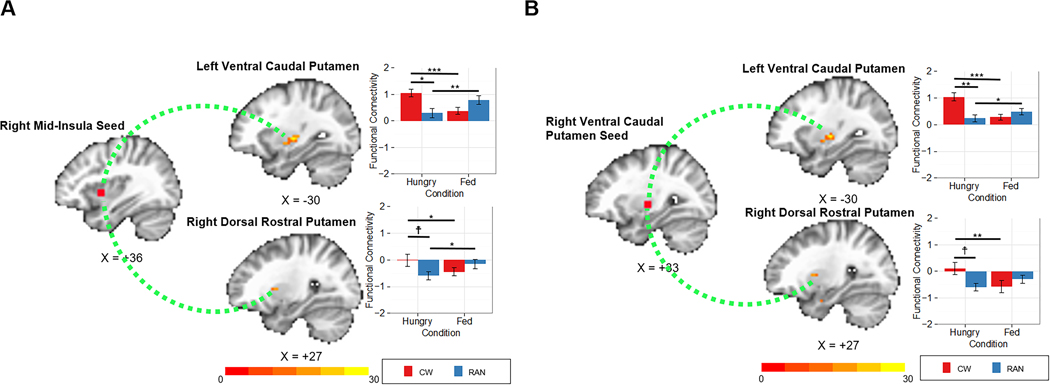
A)Functional connectivity analyses for the right caudal mid insula seed. Linear mixed effect results demonstrated an interaction of Group (CW, RAN) by Condition (hungry, fed) between the right insula seed and clusters within the left ventral caudal putamen (peak coordinate: x=−30, y=−15, z=0) and right dorsal rostral putamen (peak coordinate: x=+27, y=+9, z=+6) in response to tastants (sucrose and water combined). B) Functional connectivity analyses for the right ventral caudal putamen seed. Linear mixed effect results demonstrated an interaction of Group (CW, RAN) by Condition (hungry, fed) between the right putamen seed and clusters within the left ventral caudal putamen (peak coordinate: x=−30, y=−15, z=0) and right dorsal rostral putamen (peak coordinate: x=+27, y=+6, z=+6) in response to tastants (sucrose and water combined). CW: healthy control women; RAN: women remitted from anorexia nervosa; ☨p<0.1; *p<0.05; **p<0.01; ***p<0.001.
Relationship to Clinical Variables.
AN participants with the highest harm avoidance scores showed the lowest response to tastants when hungry in the left (t=−3.46, p<0.001) and right (t=−3.94, p<0.001) rostral caudate (Figure 2B). There were no relationships between response to tastants and other clinical variables tested in CW or RAN, such as duration of illness or remission.
DISCUSSION
The relentless ability to restrict eating and become severely emaciated has been one of the most puzzling symptoms of AN. In RAN, striatal and, at a lower statistical threshold, insular activation was abnormally unresponsive to taste stimulation in the hungry state. As expected, this circuitry, which guides motivated behavior, such as eating 7, was activated in CW when hungry compared to being fed 6, 12. Specifically, we identified a region of the ventral caudal putamen in which: 1) CW showed an increase, but RAN showed a decrease in BOLD response to tastants when hungry versus fed; and 2) RAN showed a lower BOLD response to tastants compared to CW only when hungry. Functionally, this region of the putamen is connected to the anterior and mid-insula 22, and, at a lower statistical threshold, RAN showed a similar response pattern that was opposite to controls in the left anterior insula. Moreover, CW showed increased mid-insula-to-ventral caudal putamen functional connectivity to tastants when hungry versus fed, whereas RAN showed decreased functional connectivity between these regions when hungry versus fed, and compared to CW when hungry. Overall, the interaction suggests that the RAN response to taste after eating may be “normal,” but the response to taste during hunger is abnormal. Future research in symptomatic groups using paradigms that measure food-specific reward value are needed; however, we speculate that our results are compatible with two possible main processes: the afferent metabolic signals that are translated into motivational behavior (food seeking) are attenuated and/or the afferent signals are excessively suppressed by top-down modulatory brain regions. Future investigation will need to disambiguate these possibilities.
We postulated that, when hungry, RAN would show less response to sucrose (a food and primary reward) compared to water. If there was a deficit specifically related to sucrose consumption, it might be possible to test this because water and sucrose have been shown to activate different groups of neurons in the primary taste cortex in non-human primates 23. Counter to our hypothesis, we did not detect differences between water and sucrose. However, fMRI studies in humans may not have the resolution to distinguish between response to water and sucrose in subregions of the primary taste cortex or related regions 12, 24. Furthermore, water and sucrose similarly activate taste circuitry and regions associated with emotion and motivation 7, 12. Because we did not detect differences between water and sucrose, we collapsed response across tastants to examine group and condition effects.
Reduced activation to tastants when hungry in the ventral caudal putamen and reduced functional connectivity between the right and left ventral caudal putamen are consistent with other studies showing disruption in limbic striatal circuitry in ill and remitted AN 16, 25, 26. Non-human primate studies show that the ventral caudal putamen receives sensory-limbic inputs from the insula, as well as the amygdala, orbitofrontal cortex, and temporal lobe, and is distinct from the sensorimotor dorsal putamen that is afferently regulated by the motor cortex 9. Human neuroimaging studies show it is functionally 22 and structurally 27 connected to prefrontal limbic and premotor cortex regions (Figure 4B). This circuitry is integral to reward processing and the preparation for and control of actions triggered by external stimuli 28. Reduced recruitment of the striatal aspects of this circuitry in response to tastants when hungry may impair the translation of taste reward value to motivated eating behavior in AN. However, the molecular mechanisms contributing to the selective sensitivity of this region to hunger signaling in AN require further study.
In addition, at a lower threshold, RAN showed reduced activation to tastants when hungry in the anterior insula (left anterior short gyrus and middle short gyrus) and reduced functional connectivity between the right anterior (middle short gyrus and posterior short gyrus) and mid-dorsal insula and ventral caudal putamen. The insula is a hub for interoception (the awareness and integration of internal body signals to regulate behavior) 10, and accumulating evidence suggests disturbances in anterior and mid-dorsal insula function in ill and remitted AN 14, 15, 29–31. The human gustatory cortex maps to the anterior and mid-insula 7. Moreover, the mid-insula is a chemosensory region involved in relaying various somato- and viscero-sensory signals to the anterior insula and striatum 7, 29. Specifically, the mid-dorsal insula integrates gustatory information with information about the body’s homeostatic needs (hunger and satiety signaling) to modulate feeding behavior. The anterior insula projects along the ventromedial axis of the striatum, including the rostral ventral striatum and ventral caudal putamen 9 to mediate and regulate goal-directed behavior, such as the consumption of palatable food 7. Together, these findings raise a question of whether there is a disconnect between the insula and striatum in AN, resulting in a failure to integrate taste information with motivational and homeostatic drives 10.
The current data are the first to suggest that RAN have an altered insula and striatal response to taste that is moderated by metabolic state. Our earlier studies of tastant responses in RAN similarly showed hypoactive response in the anterior insula, ventral putamen, and other striatal regions 14, 15. There was less supervision of pre-study eating in these early studies, raising the possibility of reduced food intake prior to scanning. Comparison of current findings to those of our earlier studies is also confounded by updates in neuroimaging statistical methods 32 and circuit identification (our earlier studies averaged activation across an anatomically defined ROI rather than performing voxel-wise analyses within an anatomical search region). Few studies have compared hungry and fed states in AN. Using a different design, Cowdrey 33 also found altered insula and striatal function during taste processing in RAN. Holsen 5 showed similar hungry-state hypoactivation in the anterior insula and limbic regions to food pictures versus objects in AN independent of illness state. Holsen 5 also noted that earlier studies show less consistent results, possibly due to methodological differences. Our study only assessed neural response to taste, but other studies suggest altered mid-insula and striatal responses to other interoceptive stimuli 30, 31, reward prediction error 25, 34, and food images 35. We previously showed reduced ventral striatal response to monetary rewards when hungry 16, suggesting that a dysfunction of homeostatic influences on neural processing of salient stimuli or reward is not restricted to food in AN but may generalize to secondary reinforcers (e.g., money) as well. Evidence that nutritional state strongly impacts reward and interoceptive processing in AN has significant implications for assessing these constructs moving forward and may explain the mixed findings in prior studies that did not manipulate or control for metabolic state.
More harm avoidant RAN participants showed less activation in response to taste when hungry in regions of the rostral caudate known to receive input from orbitofrontal and medial prefrontal cortices as well as the dorsolateral prefrontal cortex (Figures 2B and 4C). This raises the possibility that high harm avoidance is associated with reduced engagement of striatal limbic and associative control circuitry that translate food reward to motivated eating, and could be related to altered dopaminergic (DA) function. Harm avoidance, a construct of anxiety, inhibition, and inflexibility 36, and anxiety alone have been associated with caudate DA availability in healthy subjects 37 and rodents 38. Altered DA metabolism 39, 40 is found in RAN, and dorsal striatum function measured by BOLD response or DA metabolism has been linked to elevated anxiety, harm avoidance, and sensitivity to punishment in AN 25, 40, 41. Dorsal striatum DA signaling also plays a role in feeding 11. DA-depleted mice, which do not initiate feeding behavior, will resume normal eating after restoration of DA selectively in the dorsal striatum, but not the ventral striatum 42. Thus, dorsal striatum DA signaling may serve as a permissive, “action initiation” signal, promoting nutritive food retrieval and consumption in response to metabolic demand 42. In the context of these prior findings, we speculate that a lower dorsal striatum response to taste when hungry among RAN participants with the most pronounced behavioral inhibition may reflect a reduced eating action initiation signal. More research using tasks that include behavioral responses are needed to test this hypothesis. Other potential explanations of dorsal caudate dysfunction in AN including efficient cognitive control 43, development of habitual eating behavior 44, or anxiety associated with eating 41.
Taken together, these findings highlight circuitry that may play a key role in pathological eating in AN (Figure 4A). Of note, this circuitry has also been implicated in appetitive changes associated with major depression: increased appetite is associated with greater response to food stimuli in limbic reward circuitry, whereas appetite loss is associated with hypoactivation within the anterior and mid-insula 45. The pathophysiology driving these disturbances in AN remains to be determined. It is possible that top-down processes inhibit these signals or there is altered homeostatic system modulation 4. Interestingly, our RAN subjects reported hunger and fullness levels similar to those of CW (Figure 1) raising the possibility that bottom-up hunger signaling is intact, but not accurately translated within this insula-striatal network that motivates action. We propose (Figure 4A) that disturbance in the mid or anterior insula may result in a distorted signal about hunger or feedback about energy balance, the disturbance in the ventral caudal putamen may result in diminished motivation, and a lower caudate response may inhibit initiation among the most harm avoidant individuals, each serving to maintain food avoidance. Whether developing AN requires disturbances in all three of these processes (interoception, motivation, inhibition), or whether having any one disturbance is sufficient to impair the signal through this network remains uncertain. Factors such as a distorted body image, might initially drive restricted eating. If individuals are also vulnerable to failing to respond to homeostatic and reward signals that stimulate eating when starved, they thus may be able to maintain food avoidance.
Limitations and future directions
This study has several methodological strengths. We only included participants who were weight stable (<3 kg weight change) and physiologically and cognitively remitted from AN for at least one year, and we systematically monitored and manipulated pre-scan nutritional status. In terms of limitations, the passive task was designed specifically to characterize differences in hard-wired circuitry underlying neural sensitivity to gustatory processing, limiting our ability to assess top-down cognitive control, decision-making, or learning 46. Unexpectedly, all participants rated the water as more pleasant-tasting than the sucrose solution and both the water and sucrose solutions as moderately pleasant-tasting. This likely limited our ability to isolate activation associated with sweet taste reward. Additionally, these ratings were made only after the task was completed, and future studies should include pre- and post-scan pleasantness ratings. Signal drop-out precluded inclusion of the orbitofrontal cortex and hypothalamus in our gustatory-reward circuit mask, however, exploratory voxel-wise whole brain analyses (Supplement) were consistent with the ROI results with extended findings in thalamic, medial prefrontal, and parietal regions. Although we balanced diagnosis across scanners (χ2=.37, p=0.60) and participants were scanned on the same scanner for both scanning sessions, differences in the magnet hardware might have influenced findings between participants. Since ill AN have dysfunctional eating (unlike our RAN subjects), it is not clear from our cross-sectional study whether dysfunction within this insula and striatal network in RAN is a trait-level alteration or a scar of being underweight. Reward- and anxiety-related alterations associated with AN persist after symptom remission and may even be present in childhood 4, 47; we therefore hypothesized that our study would detect trait-like alterations in neural activation. Changes in other circuits not studied here (e.g., executive control circuits 48) may be required to compensate for these persistent reward- or anxiety-related alterations to promote normalized behavior. In our sample, BOLD response was not associated with duration of illness or remission, providing further support for trait-like alterations; however, longitudinal research among individuals at risk for developing AN is needed to understand alterations that may predispose individuals to AN and persist after remission. Moreover, future research using paradigms that directly assess food reward valuation and approach in currently symptomatic individuals with AN will be necessary to more rigorously test our hypothesized association between reduced hungry-state activation and reduced motivation to eat. Finally, our results indicate hungry-state differences in the neural response to predictable receipt of uncertain tastants (sucrose or water). These results cannot be generalized to all food stimuli. For example, other studies suggest different response to aversive taste processing 33, 49. It remains unknown why differences appear specific to hunger and not satiety, and future research is needed to understand the effects of hunger-state differences on unpredictable tastants.
Implications
Treatments for AN are only marginally effective because we lack an understanding of underlying neural mechanisms of the disorder. If reduced recruitment of neural circuitry implicated in translating reward signals to motivated behaviors when hungry interferes with food approach and eating initiation after restriction in AN, particularly among individuals with high anxiety, pharmacological and psychotherapeutic strategies that directly target these processes may be beneficial. For example, this circuitry involves dopaminergic projections. Recent findings 50–53 for olanzapine and aripiprazole suggest that investigation of medications targeting the dopamine system may improve eating behavior and reduce anxiety in AN. In addition, these findings support investigating behavioral strategies for enhancing initiation to eat or compensating for altered homeostatic drives 54, 55. Finally, a registry aimed at understanding how those with AN recover could be of much benefit in developing new treatment strategies. Such registries 56 have been of use for developing insights into successful weight loss in obesity.
Supplementary Material
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Acknowledgements:
Supported by NIH grants R01MH042984-17A1, R01MH042984-18S1, F32MH108311, 5T32MH018399-28, the Wismer Foundation, and the Price Foundation. The authors declare no conflict of interest.
Footnotes
Disclosures: none
References
- 1.American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-V (5th ed.). Washington, DC: American Psychiatric Association; 2013. [Google Scholar]
- 2.Goldstone A, Prechl de Hernandez C, Beaver J, et al. Fasting biases brain reward systems towards high-calorie foods. Eur J Neurosci. 2009;30(8):1625–35. [DOI] [PubMed] [Google Scholar]
- 3.Steinglass J, Sysko R, Mayer L, et al. Pre-meal anxiety and food intake in anorexia nervosa. Appetite. 2010;55(2):214–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Monteleone A, Castellini G, Volpe U, et al. Neuroendocrinology and brain imaging of reward in eating disorders: A possible key to the treatment of anorexia nervosa and bulimia nervosa. Prog Neuropsychopharmacol Biol Psychiatry. 2018;80(Pt B):132–42. [DOI] [PubMed] [Google Scholar]
- 5.Holsen L, Lawson E, Blum K, et al. Food motivation circuitry hypoactivation related to hedonic and nonhedonic aspects of hunger and satiety in women with active anorexia nervosa and weight-restored women with anorexia nervosa. J Psychiatry Neurosci. 2012;37(5):322–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Small D Central gustatory processing in humans. Adv Otorhinolaryngol. 2006;63:191–220. [DOI] [PubMed] [Google Scholar]
- 7.Rolls E Reward systems in the brain and nutrition. Ann Rev Nutrition. 2016;36:435–70. [DOI] [PubMed] [Google Scholar]
- 8.Wang L, Gillis-Smith S, Peng Y, et al. The coding of valence and identity in the mammalian taste system. Nature. 2018;558(7708):127–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Fudge J, Breibart M, Danish M, et al. Insular and gustatory inputs to the caudal ventral striatum in primates. J Comp Neurol. 2005;490(2):101–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Craig AD. How do you feel? Interoception: the sense of the physiological condition of the body. Nat Rev Neurosci. 2002;3(8):655–66. [DOI] [PubMed] [Google Scholar]
- 11.Ferrario C, Labouebe G, Liu S, et al. Homeostasis meets motivation in the battle to control food intake. J Neurosci. 2016;36(45):11469–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Haase L, Cerf-Ducastel B, Murphy C. Cortical activation in response to pure taste stimuli during the physiological states of hunger and satiety. Neuroimage. 2009;44(3):1008–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Stice E, Burger K, Yokum S. Caloric deprivation increases responsivity of attention and reward brain regions to intake, anticipated intake, and images of palatable foods. Neuroimage. 2013;67:322–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wagner A, Aizenstein H, Frank GK, et al. Altered insula response to a taste stimulus in individuals recovered from restricting-type anorexia nervosa. Neuropsychopharmacology. 2008;33(3):513–23. [DOI] [PubMed] [Google Scholar]
- 15.Oberndorfer T, Frank G, Fudge J, et al. Altered insula response to sweet taste processing after recovery from anorexia and bulimia nervosa. Am J Psychiatry. 2013;214(2):132–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wierenga C, Bischoff-Grethe A, Melrose A, et al. Hunger does not motivate reward in women remitted from anorexia nervosa. Biol Psychiatry. 2015;77(7):642–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Glover G, Mueller B, Turner J, et al. Function biomedical informatics research network recommendations for prospective multicenter functional MRI studies. J Magn Reson Imaging. 2012;36(1):39–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Alakurtti K, Johanson J, Tuokkola T, et al. Rostrocaudal gradients of dopamine D2/3 receptor binding in striatal subregions measured with [(11)C]raclopride and high-resolution positron emission tomography. Neuroimage. 2013;82:252–9. [DOI] [PubMed] [Google Scholar]
- 19.McLaren DRM, Xu G, Johnson S. A generalized form of context-dependent psychophysiological interactions (gPPI): a comparison to standard approaches. Neuroimage. 2012;61(14):1277–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Harrison T, McLaren D, Moody T, et al. Generalized psychophysiological interaction (PPI) analysis of memory related connectivity in individuals at genetic risk for Alzheimer’s Disease. J Vis Exp. 2017;14:129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Huber P Robust estimation of location parameter. Ann Mathematical Stats. 1964;35(1):73–101. [Google Scholar]
- 22.Choi E, Yeo B, Buckner R. The organization of the human striatum estimated by intrinsic functional connectivity. J Neurophysiol. 2012;108(8):2242–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yaxley S, Rolls E, Sienkiewicz Z. Gustatory responses of single neurons in the insula of the macaque monkey. J Neurophysiol. 1990;63(689–700). [DOI] [PubMed] [Google Scholar]
- 24.De Araujo I, Kringelbach M, Rolls ET, et al. Human cortical responses to water in the mouth, and the effects of thirst. J Neurophysiol. 2003;90:1865–76. [DOI] [PubMed] [Google Scholar]
- 25.DeGuzman M, Shott M, Yang T, et al. Association of elevated reward prediction error response with weight gain in adolescent anorexia nervosa. Am J Psychiatry. 2017;174(6):557–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Frank G, DeGuzman M, Shott M, et al. Association of brain reward learning response with harm avoidance, weight gain, and hypothalamic effective connectivity in adolescent anorexia nervosa. JAMA Psychiatry. 2018;75(10):1071–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Draganski B, Kherif F, Kloppel S, et al. Evidence for segregated and integrative connectivity patterns in the human basal ganglia. J Neurosci. 2008;28(28):7143–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Neuroscience Haggard P.. The sources of human volition Science. 2009;324(5928):731–3. [DOI] [PubMed] [Google Scholar]
- 29.Avery J, Gotts S, Kerr K, et al. Convergent gustatory and viscerosensory processing in the human dorsal mid-insula. Hum Brain Mapp. 2017;38(4):2150–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bischoff-Grethe A, Wierenga C, Berner L, et al. Neural hypersensitivity to pleasant touch in women remitted from anorexia nervosa. Transl Psychiatry. 2018;8(1):161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kerr K, Moseman S, Avery J, et al. Altered insula activity during visceral interoception in weight-restored patients with anorexia nervosa Neuropsychopharm. 2016;41(2):521–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Eklund A, Nichols T, Knutsson H. Cluster failure: Why fMRI inferences for spatial extent have inflated false-positive rates. Proc Natl Acad Sci USA. 2016;113(28):7900–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cowdrey F, Park R, Harmer C, et al. Increased neural processing of rewarding and aversive food stimuli in recovered anorexia nervosa. Biol Psych. 2011;70(8):736–43. [DOI] [PubMed] [Google Scholar]
- 34.Frank G, Collier S, Shott M, et al. Prediction error and somatosensory insula activation in women recovered from anorexia nervosa. J Psychiatry Neurosci. 2016;41(2):304–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lloyd E, Steinglass J. What can food-image tasks teach us about anorexia nervosa? A systematic review. J Eat Disord. 2018;1(6):31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cloninger CR, Bayon C, Svrakic DM. Measurement of temperament and character in mood disorders: a model of fundamental states as personality types. Journal of Affective Disorders. 1998;51:21–32. [DOI] [PubMed] [Google Scholar]
- 37.Kim J, Son Y, Kim H, et al. Association of harm avoidance with dopamine D2/3 receptor availability in striatal subdivisions: a high resolution PET study. Biol Psychology. 2011;87(1):164–7. [DOI] [PubMed] [Google Scholar]
- 38.Simon N, Montgomery K, Beas B, et al. Dopaminergic modulation of risky decision-making. J Neurosci. 2011;31(48):17460–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kaye WH, Frank GK, McConaha C. Altered dopamine activity after recovery from restricting-type anorexia nervosa. Neuropsychopharmacology. 1999;21(4):503–6. [DOI] [PubMed] [Google Scholar]
- 40.Frank G, Bailer UF, Henry S, et al. Increased dopamine D2/D3 receptor binding after recovery from anorexia nervosa measured by positron emission tomography and [11C]raclopride. Biological Psychiatry. 2005;58(11):908–12. [DOI] [PubMed] [Google Scholar]
- 41.Bailer U, Price J, Meltzer C, et al. Dopaminergic activity and altered reward modulation in anorexia nervosa-insight from multimodal imaging. Int J Eat Disord. 2017;50(5):593–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Palmiter R Dopamine signaling in the dorsal striatum is essential for motivated behaviors: lessons from dopamine-deficient mice. Ann NY Acad Sci. 2008;1129:35–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ehrlich S, Geisler D, Ritschel F, et al. Elevated cognitive control over reward processing in recovered female patients with anorexia nervosa. J Psychiatry Neurosci. 2015;40(5):307–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Foerde K, Steinglass J, Shohamy D, et al. Neural mechanisms supporting maladaptive food choices in anorexia nervosa. Nat Neurosci. 2015;18(11):1571–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Simmons W, Burrows K, Avery J, et al. Depression-related increases and decreases in appetite: dissociable patterns of aberrant activity in reward and interoceptive neurocircuitry. Am J Pschiatry. 2016;173(4):418–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Steinglass J, Foerde K. Reward system abnormalities in anorexia nervosa: Navigating a path forward. JAMA Psychiatry. 2018;75(10):993–4. [DOI] [PubMed] [Google Scholar]
- 47.Kaye W, Bulik C, Thornton L, et al. Comorbidity of anxiety disorders with anorexia and bulimia nervosa. Am J Psychiatry. 2004;161(12):2215–21. [DOI] [PubMed] [Google Scholar]
- 48.Uher R, Brammer M, Murphy T, et al. Recovery and chronicity in anorexia nervosa: brain activity associated with differential outcomes. Biol Psych. 2003;54:934–42. [DOI] [PubMed] [Google Scholar]
- 49.Monteleone A, Monteleone P, Esposito F, et al. Altered processing of rewarding and aversive basic taste stimuli in symptomatic women with anorexia nervosa and bulimia nervosa: An fMRI study. J Psychiatr Res. 2017;90:94–101. [DOI] [PubMed] [Google Scholar]
- 50.Attia E, Steinglass J, Walsh B, et al. Olanzapine versus placebo in adult outpatients with anorexia nervosa: A randomized clinical trial. Am J Psychiatry. 2019;176(6):449–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Frank G, Shott M, Hagman J, et al. The partial dopamine D2 receptor agonist aripiprazole is associated with weight gain in adolescent anorexia nervosa. Int J Eat Disord. 2017;50(4):447–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Marzola E, Desedime N, Giovannone C, et al. Atypical antipsychotics as augmentation therapy in anorexia nervosa. Plos One. 2015;10(4):e0125569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Trunko M, Schwartz T, Duvvuri V, et al. Aripiprazole in anorexia nervosa and low-weight bulimia nervosa: Case reports. Int J Eat Disord. 2011;44(3):269–75. [DOI] [PubMed] [Google Scholar]
- 54.Kaye W, Wierenga C, Knatz S, et al. Temperament-based treatment for anorexia nervosa. Eur Eat Disord Rev. 2015;23(1):12–8. [DOI] [PubMed] [Google Scholar]
- 55.Wierenga C, Hill L, Knatz Peck S, et al. The acceptability, feasibility, and possible benefits of a neurobiologically-informed 5-day multifamily treatment for adults with anorexia nervosa. Int J Eat Disord. 2018;51(8):863–9. [DOI] [PubMed] [Google Scholar]
- 56.Montesi L, El Ghoch M, Brodosi L, et al. Long-term weight loss maintenance for obesity: a multidisciplinary approach. Diabetes Metab Syndr Obes. 2016;9:37–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Martinez D, Slifstein M, Broft A, et al. Imaging human mesolimbic dopamine transmission with positron emission tomography. Part II: amphetamine-induced dopamine release in the functional subdivisions of the striatum. J Cereb Blood Flow Metab. 2003;23(3):285–300. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.