

Abstract
Type 2 diabetes mellitus (T2DM), characterized by insulin resistance and unclear pathogenesis, is a serious menace to human health. Bile acids are the end products of cholesterol catabolism and play an important role in maintaining cholesterol homeostasis. Furthermore, increasing studies suggest that bile acids may regulate glucose tolerance, insulin sensitivity, and energy metabolism, suggesting that bile acids may represent a potential therapeutic target for T2DM. This study summarizes the metabolism of bile acids and, more importantly, changes in their concentrations, constitution, and receptors in diabetes. Furthermore, we provide an overview of the mechanisms underlying the role of bile acids in glucose and lipid metabolism, as well as the occurrence and development of T2DM. Bile acid-targeted therapy may represent a valid approach for T2DM treatment.
1. Introduction
T2DM is generally considered a progressive, incurable, and increasingly prevalent illness characterized by insulin resistance and a deficiency in the absence of autoimmune beta-cell destruction, accounting for 90-95% of all diabetes cases [1]. Furthermore, it has become the most challenging endocrine disease and a leading cause of mortality worldwide, which by all predictions will only increase [2]. With rapid urbanization, economic growth, and changes in lifestyles, the prevalence of T2DM in China is increasing significantly, representing a serious problem that causes a significant burden on society [3]. The pathogenesis of T2DM is not yet entirely clear, and some evidence suggests it may be related to obesity, ethnicity, and environmental risk factors [4]. Recently, emerging evidence has suggested that insulin resistance may be the most important contributor [5]. Obese individuals tend to develop inflammation in their fat tissue, reducing the sensitivity of fat cells to insulin and inducing the development of T2DM and diminished effects of insulin [1]. Furthermore, T2DM frequently causes microvascular pathological changes, leading to stroke, heart failure, renal failure, and myocardial infarction in late stages, leading to poor prognosis and quality of life in patients [6]. Currently, treatment of T2DM comprises five primary methods: education for patients, self-monitoring of blood glucose, diet, exercise, and medication [7]. However, these methods are expensive and inefficient. Hence, diagnosis is often delayed until complications have arisen and financial costs for the treatment of T2DM have become significant, indicating the urgent need to develop new and efficient therapies and prevention methods for the control of type 2 diabetes.
Current studies have shown that gut hormones, such as ghrelin, play an important role in mediating feeding, which is significant to the development of T2DM [8]. As the endogenous ligand for the growth hormone secretagogue receptor (GHS-R) [9], ghrelin not only induces growth hormone release but also enhances food intake and stimulates adiposity [10, 11]. Primarily synthesized by X/A-like cells in the gastric oxyntic glands [9], acyl ghrelin releases NPY and AgRP by activating tGHS-Rs to stimulate food intake, body weight gain, and diabetic hyperphagia [12]. Additionally, gut hormones such as ghrelin have a vital influence on glucose metabolism [13] and T2DM remission; one study revealed that mice lacking acyl ghrelin demonstrated reduced fasting blood glucose levels and improved insulin sensitivity for controlling feeding blood glucose [14]. Moreover, other gut hormones, such as glucose-dependent insulinotropic polypeptide and glucagon-like peptide 1, hold the same promising potential in T2DM research [8].
Bile acids are the end products of cholesterol catabolism and play an important role in maintaining cholesterol homeostasis and preventing the buildup of toxic metabolites, as well as the accumulation of cholesterol [15]. Studies have demonstrated that bile acids are closely associated with the intestinal microbiota, which intimately affects gut hormones [16]. Regulation of feeding, metabolism, disease development, and homeostasis may be the result of their interactions and mutual influence [17]. On the one hand, bile acids not only facilitate transport of lipids and intestinal absorption but are also inflammatory agents and signaling molecules that effectively activate cell signaling pathways that regulate glucose, lipids, and energy metabolism [18]. On the other hand, accumulating studies have suggested that bile acids could activate certain receptors, such as the farnesoid X receptor (FXR) and the transmembrane G protein-coupled receptor 5 (TGR5), which improves glucose tolerance, insulin sensitivity, and energy metabolism [19]. The effects of these receptors suggest that bile acids may represent a potential therapeutic target for treating T2DM. This review is aimed at summarizing the effects of and changes in bile acids and their main receptors, such as FXR and TGR5, in T2DM development and their promise of representing potential treatment targets for T2DM.
2. Metabolism of Bile Acids
Bile acids (BAs) are significant bile components synthesized in the liver by cholesterol, secreted into the bile duct, and concentrated in the gallbladder, and they serve as amphipathic biological detergents for lipid metabolism [20]. Most bile acids are reabsorbed and recycled via enterohepatic circulation, and approximately 5% are lost in feces or serve as substrates for biotransformation and metabolism in the intestinal microbiota [20]. These bacteria are responsible for the dissociation of bile acids from glycine or taurine mediated by hydrolytic enzymes and hydroxyl oxidation [20].
The primary bile acids chenodeoxycholic acid (CDCA) and cholic acid (CA) are synthesized via two pathways utilizing approximately fifteen enzymes from cholesterol in the liver [20]. The rate-limiting enzyme, cytochrome P450 cholesterol 7α-hydroxylase (CYP7A1), triggers the classical pathway, converting cholesterol into 7α-hydroxycholesterol and producing most of the BA pool [21]. CA is formed through subsequent modification by a series of enzyme cascades (CYP8B1, AKR1D1, AKR1C4, and CYP27A1), while CDCA is synthesized by the same enzyme cascades except for CYP8B1 [21]. The rest of the BA pool is synthesized by cytochrome P450 27α-hydroxylase (CYP27A1) via an alternative pathway. First, cholesterol may be oxidized to 27-hydroxycholesterol with the help of CYP7B1. Then, 27-hydroxycholesterol is transformed into CDCA by CYP7B1 [21] (Figure 1). The dissociative BAs mentioned above transform into conjugated BAs after their conjugation to either glycine (primarily in humans) or taurine (predominantly in mice) by bile acid-amino acid transferase (BAT) and bile acid-CoA synthase (BACS) enzymes. Next, conjugated BAs are secreted into the bile canaliculi via the bile salt export pump and BA transporters MRP2 and MDR1A [22]. Subsequently, they accumulate and are stored and concentrated in the gallbladder. When cholecystokinin (CCK) is secreted by enteroendocrine I cells, the gallbladder is stimulated to contract and secrete bile into the duodenum to participate in the digestion and solubilization of ingested lipids. Approximately 95% of intestinal BAs are reabsorbed by enterocytes from the distal ileum through the apical sodium-dependent BA transporter (ASBT/SLC10A2) or the ileal bile acid transporter (IBAT) [23, 24]. These absorbed BAs are cleared by active transporters in the sinusoidal membrane of hepatocytes (NTCP, OAT, OATP, and mEH) when they return to the liver through the superior mesenteric and portal veins [25]. These redissociated BAs return to hepatocytes, along with newly formed bile acids, and are then secreted into the bile ducts, a process known as enterohepatic circulation. A portion of reabsorbed BAs will successfully escape hepatic recapture and reach the peripheral tissues via systemic circulation, performing signaling functions on several peripherally expressed BA receptors [21, 26]. The remaining 5% of BAs are excreted in the feces or serve as substrates for biotransformation, where they will be converted into secondary bile acids by the intestinal microbiota: deoxycholic acid (DCA) is formed from CA, lithocholic acid (LCA) is formed from CDCA, and ursodeoxycholic acid (UDCA) is formed in humans via 7α-dehydroxylation [24, 27].
Figure 1.

Bile acid synthesis and metabolism. Schematic representation of synthetic pathways of primary bile acids in hepatocytes and secondary bile acids in the intestine.
3. Bile Acid Metabolism Alterations in T2DM
Recently, accumulating studies have shown that bile acids are involved in systemic metabolism, pancreatic islet insulin resistance, hyperglycemia regulation, and energy expenditure [28, 29]. Abnormal BA metabolism is closely related to a variety of metabolic diseases, such as obesity, dyslipidemia, and nonalcoholic fatty liver disease. In addition, bile acids have been proven to be involved in glucose and lipid metabolism [22]. Therefore, we will focus on changes in BA metabolism in T2DM.
3.1. Changes in Total Bile Acids in T2DM
Some studies have shown that during the feeding state, total BA concentration in the setting of T2DM is significantly elevated compared to nondiabetic controls [30–32]. Furthermore, elevated total bile acid concentrations were positively correlated with increasing meal fat content [30]. For T2DM patients, total bile acid levels were positively correlated with triglycerides, insulin resistance index, blood pressure, and BMI, suggesting a relationship between total BA content and T2DM [33]. These findings suggest that total BA concentrations tend to increase in the setting of diabetes. Although the repeatability and authenticity of these experiments need to be further verified and the mechanism and causes of increased total BA concentration are not fully understood, we speculate that increased levels of total bile acids might be either a manifestation or cause of T2DM or may represent a link in the causal chain.
3.2. Changes in Bile Acid Composition in T2DM
Changes in bile acid composition have been verified in both clinical trials and animal models of type 2 diabetes mellitus. Increased concentrations of deoxycholic acid and decreased concentrations of chenodeoxycholic acid were observed in T2DM patients [34–38]. In T2DM patients, CDCA, DCA, and CA were also significantly increased, and CDCA, CA, and to a lesser extent DCA were positively associated with insulin resistance. Another study demonstrated that compared to nondiabetic controls, glyco-BAs are elevated in T2DM [39]. Moreover, glucose and insulin can boost histone acetylation of CYP7A1 chromatin, leading to stimulation of CYP7A1. CYP7A1 stimulation then activates the classic BA synthesis pathway and increases serum bile acids, leading to higher proportions of CA and DCA to CDCA and suggesting that abnormal glucose metabolism may affect bile acid metabolism in T2DM [40, 41]. In mouse models, inhibition of CA synthesis improves glucose homeostasis and prevents diet-induced obesity. However, increased levels of CA may contribute to dyslipidemia, diabetes, and obesity by stimulating cholesterol absorption [40–42].
Clearly, changes in the concentration and composition of the BA pool should not be ignored in T2DM. Although a few studies have been performed, evidence of these changes is still under investigation. Understanding changes in the bile acid pool is of great significance to the pathogenesis of T2DM.
4. Bile Acids in the Regulation of Glucose Homeostasis
Currently, with far more understanding of the regulation of bile acid metabolism, we now know that it is an important pathway in glucose metabolism [20] (Figure 2). Bile acid metabolism is regulated by precise feedback mechanisms from two receptors called FXR and TGR5, and the differential affinity of the bile acids towards FXR and TGR5 was summarized in Table 1.
Figure 2.

Bile acids in regulation of glucose homeostasis.
Table 1.
The differential affinity of the subtypes of bile acids towards FXR and TGR5.
| Subtypes of bile acids | |
|---|---|
| FXR agonists | 6-ECDCA43 (synthetic)>CDCA44,45,46>CA>DCA>LCA |
| FXR antagonists | TαMCA, TβMCA47 |
| TGR5 agonists | INT-77748 (a derivative of CDCA)>LCA49,50>DCA51>CDCA>CA>UDCA |
| TGR5 antagonists | SBI-11552 (synthetic) |
4.1. Regulation of Glucose Homeostasis and Bile Acid Metabolism by FXR
The nuclear receptor farnesoid X receptor (FXR), a key regulator of glucose metabolism, is significant not only to bile acid metabolism in the liver but also to biliary BA secretion and intestinal BA absorption [42–60]. FXR is easily activated by both free and conjugated bile acids due to its high expression levels in the liver. Activation of FXR in the liver subsequently increases the excretion of bile acids into the intestine and inhibits activity of CYP7A1, the rate-limiting enzyme of the classic pathway mentioned above, by increasing transcription of the inhibitory small heterodimer partner (SHP) [61]. FXR activation prevents the accumulation of bile acids in the liver via transcriptional induction of apical transporters, such as BSEP and MRP2 [62, 63]. To evaluate the function of FXR in regulating BA homeostasis and detoxification in response to bile duct ligation (BDL), many experiments have been performed in FXR knockout mice. These mice exhibited enhanced CYP7A1 mRNA expression and increased BA pools [64, 65]. Accompanied by a lack of CYP7A1 inhibition, FXR null mice presented with more serious hepatotoxicity when fed a CA diet. In addition, activation of FXR has many other mechanisms to downregulate bile acid synthesis and govern the composition of the bile acid pool [21, 66–73]. Additionally, activation of FXR in the liver may also regulate and inhibit CYP8B1 in some ways [61].
Numerous studies have shown that FXR is closely related to glucose metabolism. Diabetic rats induced by streptozotocin exhibited reduced FXR expression in the liver, which can be restored by insulin supplementation [74]. FXR knockout mice exhibited a distinct response to refeeding in hepatic expression of glucose metabolism genes [74]. Elevated glucose influx and activation of insulin signaling may lead to postprandial bile acid synthesis [75]. A previous study demonstrated that high concentrations of bile acids stimulate ligand-dependent FXR transactivating activity and increase cellular glucose flux [76]. During the postprandial phase, FXR may be activated to regulate glucose homeostasis [75, 77, 78]. Recent studies suggested that FXR regulates the sensitivity and secretion of peripheral insulin and promotes glycogen synthesis by inducing FGF15 (FGF19 in humans) in the intestine [70]. In addition, FGF19 directly activates SHP by combining with FGFR4 with the help of klotho β in hepatocytes [60] (Figure 3).
Figure 3.

The relationship between total bile acids, FXR, and FGF19.
Though the concrete role of FXR in the regulation of hepatic glucose metabolism remains debatable, the probability that FXR activation inhibits hepatic glucose synthesis to decrease fasting plasma glucose has been universally acknowledged [79]. Its natural association with T2DM may decrease the quantity, activation, or sensitivity of FXR in some ways, and one study by Staels and Fonseca suggested that insulin suppresses expression of the FXR gene, speculating that diabetes may be associated with the dysbiosis of FXR expression [71–73].
4.2. Regulation of Glucose Homeostasis and Bile Acid Metabolism by TGR5
Transmembrane G protein-coupled receptor 5 (TGR5) is expressed in nearly the entire body, particularly in a variety of liver cells [61]. Multiple studies have shown that there is a tight connection between TGR5 and bile acid metabolism [79–81]. TGR5 might also regulate bile acid metabolism [82]. Distinct from FXR, the affinity of bile acids for hydrophobicity to TGR5 is LCA>DCA>CDCA>CA>UDCA [61]. Combined with its agonists, TGR5, with the help of its cofactors α, β, and γ, activates protein kinase A (PKA) signaling pathways by activating adenyl cyclase, leading to the swift growth of intracellular cAMP production [77]. Then, PKA pathways lead to the phosphorylation and induce the expression of the target genes of the transcription factor cAMP-responsive element-binding protein (CREB) [83] (Figure 4).
Figure 4.
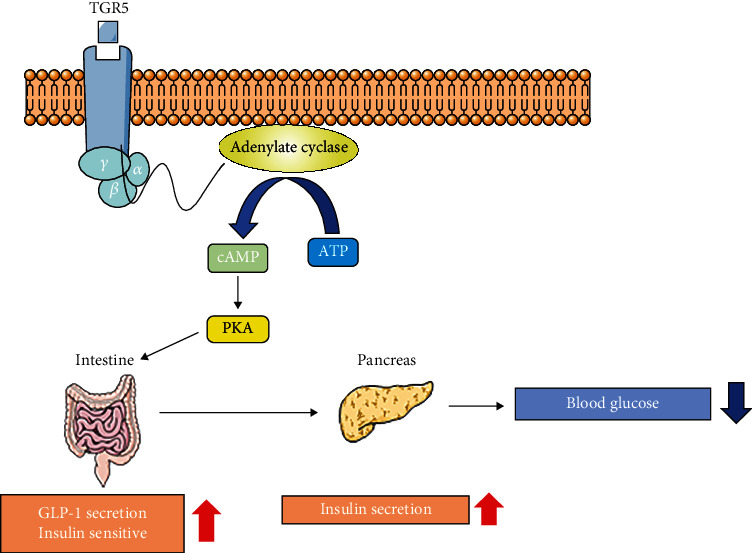
The relationship between TGR5, PKA, and GLP-1.
Glucose regulation improves when TGR5 signaling increases [81, 82]. Glucagon-like peptide 1 (GLP-1) promotes insulin secretion by islet beta cells in a glucose-dependent manner and reduces glucagon secretion by islet alpha cells, thereby lowering blood glucose. Secretion of GLP-1 is enhanced in response to TGR5 signaling activation in gastrointestinal enteroendocrine L cells [82]. In addition to its glucose-dependent insulinotropic effect, GLP-1 has similar properties to glucagon and induces satiety. Bile acids may regulate glucose homoeostasis, appetite, and even body weight via TGR5 [84–86]. Some studies have reported that TGR5 is a downstream target of FXR and it is required for promoting GLP-1 secretion through L cell FXR signaling [78]. The specific role of TGR5 in the regulation of glucose homeostasis requires further study, but activation of TGR5 does convey an apparent beneficial effect on glucose homeostasis.
4.3. Role of FXR, TGR5, and Total Bile Acids in Glucose Homeostasis and T2DM Remission after Bariatric Surgery
Although T2DM lacks a specific treatment, bariatric surgery, including Roux-en-Y gastric bypass (RYGB) and vertical sleeve gastrectomy (VSG), conveys long-term disease mitigation [87, 88]. Fibroblast growth factor 19 (FGF19), an intestinal feedback signal of bile acids, has been implicated in the glucometabolic changes that take place after Roux-en-Y gastric bypass (RYGB). Bariatric surgery improves glucose regulation by promoting bile acid signaling, which may then increase circulating bile acid concentrations or anatomical rearrangement of the gastrointestinal tract [89–91]. Studies have revealed that FXR and TGR5 contribute to the metabolic benefits of bariatric surgery. FXR is beneficial to weight loss and improves glucose regulation in response to VSG and TGR5 [89–91]. Improvements in signaling are associated with a TGR5-dependent pathway that decreases the hydrophobicity of the circulating bile acid pool [53]. This beneficial spectrum change in bile acids is associated with decreased expression of TGR 5-dependent liver CYP8B1 protein, with no effect on the expression of liver CYP7A1 [89].
A recent study investigating the relationship between total bile acids, FGF19, and T2DM in bariatric surgery identified important roles of total bile acids and FGF19 in T2DM remission after sleeve gastrectomy (SG) by comparing postprandial gut hormone patterns between patients undergoing laparoscopic gastric bypass (GB) and laparoscopic sleeve gastrectomy 2 years after surgery [92]. This research revealed that both laparoscopic GB and laparoscopic SG have significant effects after surgery but do have discrepancies not associated with insulin secretion, weight loss, or hindgut effect that are connected with reduced insulin resistance and duodenal exclusion in GB in T2DM remission. Moreover, differential regulation of different subtypes of ghrelin and total bile acids might be involved in the differing insulin resistance and T2DM remission responses between GB and SG procedures [92].
5. Bile Acid-Based Therapy for T2DM
5.1. Bile Acids as Therapeutic Drugs
Bile acids have been used for cholestatic liver diseases and metabolic diseases for years [93]. DCA, transformed by rectal taurocholate (TCA), increases GLP-1 secretion and insulin, leading to decreased serum glucose by activating intestinal bile acid receptors FXR and TGR5 [94, 95]. Although UDCA is not used for T2DM directly, it has been used to treat obese patients [96]. Short-term UDCA administration activates FXR to stimulate bile acid and cholesterol synthesis, while circulation of FGF19 decreases. Metformin is a noted drug prescribed for T2DM that improves insulin resistance and sensitivity. One of the drug's effects is increasing TUDCA and GUDCA by altering the gut microbiota. In addition, TUDCA and GUDCA act as antagonists of intestinal FXR to improve hyperglycemia in T2DM [72]. Therefore, another way to treat T2DM may be to increase concentrations of TUDCA and GUDCA directly.
5.2. Bile Acid Sequestrants
Bile acid sequestrants were observed to improve glycemic control in T2DM patients as early as the 1990s [31]. Since bile acids have been proven to play an important role in glucose metabolism, which is relevant to T2DM and insulin resistance, bile acid sequestrants were explored.
First used as a treatment for hypercholesterolemia, bile acid sequestrants, such as cholestyramine, colesevelam, colestimide, and colestipol, are nonabsorbable resins that combine negatively charged bile salts into a complex in the intestinal lumen, which are then excreted in the feces; therefore, bile acids are diverted from the enterohepatic cycle and excreted from the body [97]. Bile acid sequestrants decrease circulating concentrations of LDL cholesterol, the substrate for bile acid production, via increasingly delivering LDL cholesterol to the liver and enhancing cholesterol synthesis and upregulation of LDL receptors [31]. Cholestyramine functions as a combination of bile acids and is a therapeutic option for some metabolic syndromes, such as dyslipidemia [98]. It stimulates cholesterol transformation into bile acids by decreasing the concentration of bile acids returned to the liver via enterohepatic cycling.
Recently, bile acid sequestrants have been approved in the USA for the treatment of T2DM, despite the mechanisms of action still not being completely understood [99]. Bile acid sequestrants successfully treat T2DM due to their hypoglycemic effect. Bile acid sequestrants could alter the bile acid pool composition. However, this hypothesis was not supported by clinical research findings, which suggested that alterations of bile acid pool composition are not a significant pathway in the glucose-lowering action of bile acid sequestrants [61]. In 2008, colesevelam was approved by the FDA for the treatment of T2DM [28, 29]. Two weeks of colesevelam treatment in T2DM patients altered the synthesis of specific bile acids, which affected the concrete composition of the total pool size [42]. However, animal studies indicated that colesevelam improved oral glucose tolerance by activating TGR5 on L cells with subsequent GLP-1 secretion. Although the hypothesis that BAs may activate TGR5 to modulate human intestinal GLP-1 release and glucose homoeostasis remains to be further understood, the TGR5 signaling pathway may represent a target that may provide a highly promising strategy for the treatment of T2DM.
5.3. Farnesoid X Receptor Agonists
Currently, studies of FXR-targeting therapies for T2DM are extremely limited. Obeticholic acid (OCA), a semisynthetic bile acid, is 30 times more effective in activating FXR than CDCA. Although it was not used for diabetes treatment, its effects in inhibiting bile acid synthesis and improving liver function have been verified in the treatment of metabolic liver diseases [100, 101]. Since FXR activation inhibits hepatic glucose synthesis to decrease fasting plasma glucose, it is likely that similar to OCA, targeting FXR represents a novel strategy for improving hyperglycemia in T2DM.
5.4. G Protein-Coupled Bile Acid Receptor Agonists
Intestine-selective TGR5 agonists may represent a potential strategy for T2DM therapy, as they have been observed to improve glucose homeostasis [21]. Insulin resistance is a characteristic of T2DM, although the mechanism of its occurrence has yet to be elucidated, but there are two primary theories. First, lipid overload, in which fat cell enlargement leads to increased levels of circulating free fatty acids (FFAs) and its metabolites, as well as deposition in nonfat cells, inhibits insulin signaling. Second, the inflammation doctrine postulates that enlarged fat cells attract macrophages, which secrete inflammatory signaling molecules, such as TNF-α, resistin, and IL-6 [102]. These two theories intersect and complement each other. INT-777, a semisynthetic acid that activates TGR5, can decrease lipid loading and macrophage inflammation by inhibiting nuclear factor κB (NF-κB) and production of proinflammatory cytokines (Figure 5).
Figure 5.
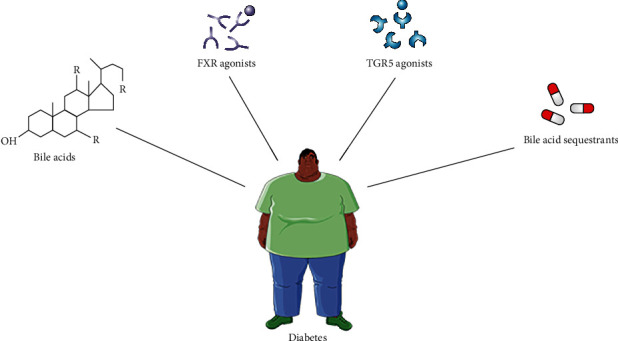
Bile acid-based therapy for T2DM.
6. Conclusions
In this review, we focused on the role of bile acids in glucose metabolism and the occurrence and development of T2DM, as well as the possibility of bile acids representing a new target for treating T2DM. Without a doubt, bile acids are exceedingly significant in glucose metabolism. Their important roles in the occurrence and development of T2DM have drawn plentiful attention. A mass of research findings have shown that there might be transformations of bile acids in their metabolism concomitant with the occurrence of T2DM, to some extent, leading to abnormal glucose metabolism and insulin resistance. However, additional studies are needed to verify these alterations and to understand the specific interactions that are occurring. Additionally, further animal experiments and clinical trials are required to support the safety and efficacy of bile acids and their sequestrants. Importantly, novel methods of targeted therapy for T2DM have recently been identified, and the future of T2DM treatment is becoming more promising.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Authors' Contributions
An Zhou, Li Tang, Yuanyuan Lei, Bo Tang, and Linjing Zhang reviewed the literature and discussed relevant articles. Yingjie Wu and Linjing Zhang prepared the manuscript.
References
- 1.American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care. 2011;35(Supplement 1):S64–S71. doi: 10.2337/dc12-s064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Greenhill C. Designing a new therapy for T2DM. Nature Reviews Endocrinology. 2019;15(12):p. 684. doi: 10.1038/s41574-019-0272-9. [DOI] [PubMed] [Google Scholar]
- 3.Balbus J. M., Barouki R., Birnbaum L. S., et al. Early-life prevention of non-communicable diseases. Lancet. 2013;381(9860):3–4. doi: 10.1016/S0140-6736(12)61609-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Defronzo R. A. Banting lecture-from the triumvirate to the ominous octet: a new paradigm for the treatment of type 2 diabetes mellitus. Diabetes. 2009;58(4):773–795. doi: 10.2337/db09-9028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cuschieri S. The genetic side of type 2 diabetes - a review. Diabetes and Metabolic Syndrome: Clinical Research and Reviews. 2019;13(4):2503–2506. doi: 10.1016/j.dsx.2019.07.012. [DOI] [PubMed] [Google Scholar]
- 6.Braunwald E. Diabetes, heart failure, and renal dysfunction: the vicious circles. Progress in Cardiovascular Diseases. 2019;62(4):298–302. doi: 10.1016/j.pcad.2019.07.003. [DOI] [PubMed] [Google Scholar]
- 7.Cattin L. Il diabete mellito: etiopatogenesi ed inquadramento clinico [Diabetes Mellitus: etiology, pathophysiology and clinical classification] Giornale Italiano di Nefrologia (Italian Journal of Nephrology) 2016;33(S68) gin/33.S68.6. [PubMed] [Google Scholar]
- 8.Chen C.-Y., Fujimiya M., Laviano A., Chang F.-Y., Lin H.-C., Lee S.-D. Modulation of ingestive behavior and gastrointestinal motility by ghrelin in diabetic animals and humans. Journal of the Chinese Medical Association. 2010;73(5):225–229. doi: 10.1016/S1726-4901(10)70048-4. [DOI] [PubMed] [Google Scholar]
- 9.Kojima M., Hosoda H., Date Y., Nakazato M., Matsuo H., Kangawa K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature. 1999;402(6762):656–660. doi: 10.1038/45230. [DOI] [PubMed] [Google Scholar]
- 10.Woods S. C. Gastrointestinal satiety signals I. An overview of gastrointestinal signals that influence food intake. American Journal of Physiology. Gastrointestinal and Liver Physiology. 2004;286(1):G7–13. doi: 10.1152/ajpgi.00448.2003. [DOI] [PubMed] [Google Scholar]
- 11.Tschöp M., Smiley D. L., Heiman M. L. Ghrelin induces adiposity in rodents. Nature. 2000;407(6806):908–913. doi: 10.1038/35038090. [DOI] [PubMed] [Google Scholar]
- 12.Huang H. H., Chen C. Y. Alpha-melanocyte stimulating hormone in ghrelin-elicited feeding and gut motility. Journal of the Chinese Medical Association. 2019;82(2):87–91. doi: 10.1097/JCMA.0000000000000007. [DOI] [PubMed] [Google Scholar]
- 13.Chen C.-Y., Asakawa A., Fujimiya M., Lee S.-D., Inui A. Ghrelin gene products and the regulation of food intake and gut motility. Pharmacological Reviews December. 2009;61(4):430–481. doi: 10.1124/pr.109.001958. [DOI] [PubMed] [Google Scholar]
- 14.Sun Y., Asnicar M., Saha P. K., Chan L., Smith R. G. Ablation of ghrelin improves the diabetic but not obese phenotype of ob/ob mice. Cell Metabolism. 2006;3(5):379–386. doi: 10.1016/j.cmet.2006.04.004. [DOI] [PubMed] [Google Scholar]
- 15.Kusumoto Y., Irie J., Iwabu K., et al. Bile acid binding resin prevents fat accumulation through intestinal microbiota in high-fat diet-induced obesity in mice. Metabolism. 2017;71:1–6. doi: 10.1016/j.metabol.2017.02.011. [DOI] [PubMed] [Google Scholar]
- 16.Ridlon J. M., Kang D. J., Hylemon P. B., Bajaj J. S. Bile acids and the gut microbiome. Current Opinion in Gastroenterology. 2014;30(3):332–338. doi: 10.1097/MOG.0000000000000057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Li Y., Tang R., Leung P. S. C., Gershwin M. E., Ma X. Bile acids and intestinal microbiota in autoimmune cholestatic liver diseases. Autoimmunity Reviews. 2017;16(9):885–896. doi: 10.1016/j.autrev.2017.07.002. [DOI] [PubMed] [Google Scholar]
- 18.Kiriyama Y., Nochi H. The biosynthesis, signaling, and neurological functions of bile acids. Biomolecules. 2019;9(6):p. 232. doi: 10.3390/biom9060232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nguyen A., Bouscarel B. Bile acids and signal transduction: role in glucose homeostasis. Cellular Signalling. 2008;20(12):2180–2197. doi: 10.1016/j.cellsig.2008.06.014. [DOI] [PubMed] [Google Scholar]
- 20.Brønden A., Albér A., Rohde U., et al. The bile acid-sequestering resin sevelamer eliminates the acuteGLP‐1 stimulatory effect of endogenously released bile acids in patients with type 2 diabetes. Diabetes, Obesity & Metabolism. 2017;20(2):362–369. doi: 10.1111/dom.13080. [DOI] [PubMed] [Google Scholar]
- 21.Morville T., Sahl R. E., Trammell S. A. J., et al. Divergent effects of resistance and endurance exercise on plasma bile acids, FGF19, and FGF21 in humans. JCI Insight. 2018;3(15, article e122737) doi: 10.1172/jci.insight.122737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chávez-Talavera O., Tailleux A., Lefebvre P., Staels B. Bile acid control of metabolism and inflammation in obesity, type 2 diabetes, dyslipidemia, and nonalcoholic fatty liver disease. Gastroenterology. 2017;152(7):1679–1694.e3. doi: 10.1053/j.gastro.2017.01.055. [DOI] [PubMed] [Google Scholar]
- 23.Staley C., Weingarden A. R., Khoruts A., Sadowsky M. J. Interaction of gut microbiota with bile acid metabolism and its influence on disease states. Applied Microbiology and Biotechnology. 2017;101(1):47–64. doi: 10.1007/s00253-016-8006-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Friedman E. S., Li Y., Shen T. C. D., et al. FXR-dependent modulation of the human small intestinal microbiome by the bile acid derivative obeticholic acid. Gastroenterology. 2018;155(6):1741–1752.e5. doi: 10.1053/j.gastro.2018.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Steiner C., Holleboom A. G., Karuna R., et al. Lipoprotein distribution and serum concentrations of 7α-hydroxy-4-cholesten-3-one and bile acids: effects of monogenic disturbances in high-density lipoprotein metabolism. Clinical Science (London, England) 2012;122(8):385–400. doi: 10.1042/CS20110482. [DOI] [PubMed] [Google Scholar]
- 26.Ridlon J. M., Harris S. C., Bhowmik S., Kang D. J., Hylemon P. B. Consequences of bile salt biotransformations by intestinal bacteria. Gut Microbes. 2016;7(1):22–39. doi: 10.1080/19490976.2015.1127483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Dayer-Berenson L., Finckenor M. Expanded colesevelam administration options with oral suspension formulation for patients with diabetes and hypercholesterolemia. Postgraduate Medicine. 2015;126(3):126–134. doi: 10.3810/pgm.2014.05.2762. [DOI] [PubMed] [Google Scholar]
- 28.Hansen M., Scheltema M. J., Sonne D. P., et al. Effect of chenodeoxycholic acid and the bile acid sequestrant colesevelam on glucagon-like peptide-1 secretion. Diabetes, Obesity & Metabolism. 2016;18(6):571–580. doi: 10.1111/dom.12648. [DOI] [PubMed] [Google Scholar]
- 29.Hansen M., Sonne D. P., Mikkelsen K. H., Gluud L. L., Vilsbøll T., Knop F. K. Bile acid sequestrants for glycemic control in patients with type 2 diabetes: a systematic review with meta-analysis of randomized controlled trials. Journal of Diabetes and its Complications. 2017;31(5):918–927. doi: 10.1016/j.jdiacomp.2017.01.011. [DOI] [PubMed] [Google Scholar]
- 30.Popescu I. R., Helleboid-Chapman A., Lucas A., et al. The nuclear receptor FXR is expressed in pancreatic beta-cells and protects human islets from lipotoxicity. FEBS Letters. 2010;584(13):2845–2851. doi: 10.1016/j.febslet.2010.04.068. [DOI] [PubMed] [Google Scholar]
- 31.Vincent R. P., Omar S., Ghozlan S., et al. Higher circulating bile acid concentrations in obese patients with type 2 diabetes. Annals of Clinical Biochemistry. 2013;50(4):360–364. doi: 10.1177/0004563212473450. [DOI] [PubMed] [Google Scholar]
- 32.Liang B., Hongwei L., Dongmei Z., Juan S., Changjiang Y., Wei L. Relationship between serum total bile acid and type 2 diabetes. Modern Medicine. 2019;47(3):250–254. [Google Scholar]
- 33.Besseling J., Hutten B. A. Is there a link between diabetes and cholesterol metabolism? Expert Review of Cardiovascular Therapy. 2016;14(3):259–261. doi: 10.1586/14779072.2016.1133292. [DOI] [PubMed] [Google Scholar]
- 34.Zhao P., Wendt D., Goodin S. Z., Ravichandran S., Chouinard T. E., Strader A. D. Adaptation of intestinal and bile acid physiology accompany the metabolic benefits following ileal interposition in the rat. Obesity Surgery. 2018;28(3):725–734. doi: 10.1007/s11695-017-2886-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Shapiro H., Kolodziejczyk A. A., Halstuch D., Elinav E. Bile acids in glucose metabolism in health and disease. The Journal of Experimental Medicine. 2018;215(2):383–396. doi: 10.1084/jem.20171965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zheng X., Huang F., Zhao A., et al. Bile acid is a significant host factor shaping the gut microbiome of diet-induced obese mice. BMC Biology. 2017;15(1):p. 120. doi: 10.1186/s12915-017-0462-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Li T., Francl J. M., Boehme S., et al. Glucose and insulin induction of bile acid synthesis: mechanisms and implication in diabetes and obesity. The Journal of Biological Chemistry. 2012;287(3):1861–1873. doi: 10.1074/jbc.M111.305789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Fiorucci S., Distrutti E. Bile acid-activated receptors, intestinal microbiota, and the treatment of metabolic disorders. Trends in Molecular Medicine. 2015;21(11):702–714. doi: 10.1016/j.molmed.2015.09.001. [DOI] [PubMed] [Google Scholar]
- 39.Haeusler R. A., Pratt-Hyatt M., Welch C. L., Klaassen C. D., Accili D. Impaired generation of 12-hydroxylated bile acids links hepatic insulin signaling with dyslipidemia. Cell Metabolism. 2012;15(1):65–74. doi: 10.1016/j.cmet.2011.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Slätis K., Gåfvels M., Kannisto K., et al. Abolished synthesis of cholic acid reduces atherosclerotic development in apolipoprotein E knockout mice. Journal of Lipid Research. 2010;51(11):3289–3298. doi: 10.1194/jlr.M009308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kaur A., Patankar J. V., de Haan W., et al. Loss of Cyp8b1 improves glucose homeostasis by increasing GLP-1. Diabetes. 2015;64(4):1168–1179. doi: 10.2337/db14-0716. [DOI] [PubMed] [Google Scholar]
- 42.Bertaggia E., Jensen K. K., Castro-Perez J., et al. Cyp8b1ablation prevents Western diet-induced weight gain and hepatic steatosis because of impaired fat absorption. American Journal of Physiology-Endocrinology and Metabolism. 2017;313(2):E121–E133. doi: 10.1152/ajpendo.00409.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Pellicciari R., Fiorucci S., Camaioni E., et al. 6α-Ethyl-chenodeoxycholic acid (6-ECDCA), a potent and selective FXR agonist endowed with anticholestatic activity. Journal of Medicinal Chemistry. 2002;45(17):3569–3572. doi: 10.1021/jm025529g. [DOI] [PubMed] [Google Scholar]
- 44.Parks D. J., Blanchard S. G., Bledsoe R. K., et al. Bile acids: natural ligands for an orphan nuclear receptor. Science. 1999;284(5418):1365–1368. doi: 10.1126/science.284.5418.1365. [DOI] [PubMed] [Google Scholar]
- 45.Makishima M., Okamoto A. Y., Repa J. J., et al. Identification of a nuclear receptor for bile acids. Science. 1999;284(5418):1362–1365. doi: 10.1126/science.284.5418.1362. [DOI] [PubMed] [Google Scholar]
- 46.Wang H., Chen J., Hollister K., Sowers L. C., Forman B. M. Endogenous bile acids are ligands for the nuclear receptor FXR/BAR. Molecular Cell. 1999;3(5):543–553. doi: 10.1016/S1097-2765(00)80348-2. [DOI] [PubMed] [Google Scholar]
- 47.Sayin S. I., Wahlström A., Felin J., et al. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metabolism. 2013;17(2):225–235. doi: 10.1016/j.cmet.2013.01.003. [DOI] [PubMed] [Google Scholar]
- 48.Kawamata Y., Fujii R., Hosoya M., et al. A G protein-coupled receptor responsive to bile acids. The Journal of Biological Chemistry. 2003;278(11):9435–9440. doi: 10.1074/jbc.M209706200. [DOI] [PubMed] [Google Scholar]
- 49.Chen X., Lou G., Meng Z., Huang W. TGR5: a novel target for weight maintenance and glucose metabolism. Experimental Diabetes Research. 2011;2011:5. doi: 10.1155/2011/853501.853501 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Maruyama T., Miyamoto Y., Nakamura T., et al. Identification of membrane-type receptor for bile acids (M-BAR) Biochemical and Biophysical Research Communications. 2002;298(5):714–719. doi: 10.1016/S0006-291X(02)02550-0. [DOI] [PubMed] [Google Scholar]
- 51.Watanabe M., Houten S. M., Mataki C., et al. Bile acids induce energy expenditure by promoting intracellular thyroid hormone activation. Nature. 2006;439(7075):484–489. doi: 10.1038/nature04330. [DOI] [PubMed] [Google Scholar]
- 52.Masyuk T. V., Masyuk A. I., Lorenzo Pisarello M., et al. TGR5 contributes to hepatic cystogenesis in rodents with polycystic liver diseases through cyclic adenosine monophosphate/Gαs signaling. Hepatology. 2017;66(4):1197–1218. doi: 10.1002/hep.29284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Hansen M., Sonne D. P., Knop F. K. Bile acid sequestrants: glucose-lowering mechanisms and efficacy in type 2 diabetes. Current Diabetes Reports. 2014;14(5):p. 482. doi: 10.1007/s11892-014-0482-4. [DOI] [PubMed] [Google Scholar]
- 54.Takebayashi K., Aso Y., Inukai T. Role of bile acid sequestrants in the treatment of type 2 diabetes. World Journal of Diabetes. 2010;1(5):146–152. doi: 10.4239/wjd.v1.i5.146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Shin D. J., Wang L. Bile acid-activated receptors: a review on FXR and other nuclear receptors. Handbook of Experimental Pharmacology. 2019;256:51–72. doi: 10.1007/164_2019_236. [DOI] [PubMed] [Google Scholar]
- 56.Cipriani S., Mencarelli A., Palladino G., Fiorucci S. FXR activation reverses insulin resistance and lipid abnormalities and protects against liver steatosis in Zucker (fa/fa) obese rats. Journal of Lipid Research. 2010;51(4):771–784. doi: 10.1194/jlr.M001602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Cariou B., van Harmelen K., Duran-Sandoval D., et al. The farnesoid X receptor modulates adiposity and peripheral insulin sensitivity in mice. Biological Chemistry. 2006;281(16):11039–11049. doi: 10.1074/jbc.M510258200. [DOI] [PubMed] [Google Scholar]
- 58.Kim K. H., Choi S., Zhou Y., et al. Hepatic FXR/SHP axis modulates systemic glucose and fatty acid homeostasis in aged mice. Hepatology. 2017;66(2):498–509. doi: 10.1002/hep.29199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Gonzalez F. J., Jiang C., Patterson A. D. An intestinal microbiota-farnesoid X receptor axis modulates metabolic disease. Gastroenterology. 2016;151(5):845–859. doi: 10.1053/j.gastro.2016.08.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mencarelli A., Renga B., D'Amore C., et al. Dissociation of intestinal and hepatic activities of FXR and LXR supports metabolic effects of terminal ileum interposition in rodents. Diabetes. 2013;62(10):3384–3393. doi: 10.2337/db13-0299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Swanson H. I., Wada T., Xie W., et al. Role of nuclear receptors in lipid dysfunction and obesity related diseases. Drug Metabolism and Disposition. 2012;41(1):1–11. doi: 10.1124/dmd.112.048694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Li T., Chiang J. Y. L. Bile acid signaling in metabolic disease and drug therapy. Pharmacological Reviews. 2014;66(4):948–983. doi: 10.1124/pr.113.008201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Allen R. M., Marquart T. J., Albert C. J., et al. miR-33 controls the expression of biliary transporters, and mediates statin- and diet-induced hepatotoxicity. EMBO Molecular Medicine. 2012;4(9):882–895. doi: 10.1002/emmm.201201228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Massafra V., van Mil S. W. C. Farnesoid X receptor: a “homeostat” for hepatic nutrient metabolism. Biochimica et Biophysica Acta - Molecular Basis of Disease. 2018;1864(1):45–59. doi: 10.1016/j.bbadis.2017.10.003. [DOI] [PubMed] [Google Scholar]
- 65.Kim K. H., Moore D. D. Regulation of liver energy balance by the nuclear receptors farnesoid X receptor and peroxisome proliferator activated receptor α. Digestive Diseases. 2017;35(3):203–209. doi: 10.1159/000450912. [DOI] [PubMed] [Google Scholar]
- 66.Drucker D. J. Mechanisms of action and therapeutic application of glucagon-like peptide-1. Cell Metabolism. 2018;27(4):740–756. doi: 10.1016/j.cmet.2018.03.001. [DOI] [PubMed] [Google Scholar]
- 67.Slijepcevic D., van de Graaf S. F. J. Bile acid uptake transporters as targets for therapy. Digestive Diseases. 2017;35(3):251–258. doi: 10.1159/000450983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Kliewer S. A., Mangelsdorf D. J. Bile acids as hormones: the FXR-FGF15/19 pathway. Digestive Diseases. 2015;33(3):327–331. doi: 10.1159/000371670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Zhang R., Ran H., Peng L., et al. Farnesoid X receptor regulates vasoreactivity via angiotensin II type 2 receptor and the kallikrein-kinin system in vascular endothelial cells. Clinical and Experimental Pharmacology & Physiology. 2016;43(3):327–334. doi: 10.1111/1440-1681.12535. [DOI] [PubMed] [Google Scholar]
- 70.Sonne D. P., van Nierop F. S., Kulik W., Soeters M. R., Vilsbøll T., Knop F. K. Postprandial plasma concentrations of individual bile acids and FGF-19 in patients with type 2 diabetes. Clinics in Endocrinology and Metabolism. 2016;101(8):3002–3009. doi: 10.1210/jc.2016-1607. [DOI] [PubMed] [Google Scholar]
- 71.Gao L., Wang X., Tang Y., Huang S., Hu C. A. A., Teng Y. FGF19/FGFR4 signaling contributes to the resistance of hepatocellular carcinoma to sorafenib. Journal of Experimental & Clinical Cancer Research. 2017;36(1):p. 8. doi: 10.1186/s13046-016-0478-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Pathak P., Xie C., Nichols R. G., et al. Intestine farnesoid X receptor agonist and the gut microbiota activate G-protein bile acid receptor-1 signaling to improve metabolism. Hepatology. 2018;68(4):1574–1588. doi: 10.1002/hep.29857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Brown A. E., Walker M. Genetics of insulin resistance and the metabolic syndrome. Current Cardiology Reports. 2016;18(8):p. 75. doi: 10.1007/s11886-016-0755-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Pathak P., Liu H., Boehme S., et al. Farnesoid X receptor induces Takeda G-protein receptor 5 crosstalk to regulate bile acid synthesis and hepatic metabolism. Biological Chemistry. 2017;292(26):11055–11069. doi: 10.1074/jbc.M117.784322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Sun L., Xie C., Wang G., et al. Gut microbiota and intestinal FXR mediate the clinical benefits of metformin. Nature Medicine. 2018;24(12):1919–1929. doi: 10.1038/s41591-018-0222-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Han S., Chiang J. Y. L. Mechanism of vitamin D receptor inhibition of cholesterol 7alpha-hydroxylase gene transcription in human hepatocytes. Drug Metabolism and Disposition. 2009;37(3):469–478. doi: 10.1124/dmd.108.025155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Péan N., Doignon I., Garcin I., et al. The receptor TGR5 protects the liver from bile acid overload during liver regeneration in mice. Hepatology. 2013;58(4):1451–1460. doi: 10.1002/hep.26463. [DOI] [PubMed] [Google Scholar]
- 78.Berrabah W., Aumercier P., Gheeraert C., et al. Glucose sensing O-GlcNAcylation pathway regulates the nuclear bile acid receptor farnesoid X receptor (FXR) Hepatology. 2014;59(5):2022–2033. doi: 10.1002/hep.26710. [DOI] [PubMed] [Google Scholar]
- 79.Wang X. X., Wang D., Luo Y., et al. FXR/TGR5 dual agonist prevents progression of nephropathy in diabetes and obesity. Journal of the American Society of Nephrology. 2017;29(1):118–137. doi: 10.1681/ASN.2017020222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wahlström A., Sayin S. I., Marschall H. U., Bäckhed F. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism. Cell Metabolism. 2016;24(1):41–50. doi: 10.1016/j.cmet.2016.05.005. [DOI] [PubMed] [Google Scholar]
- 81.Donepudi A. C., Boehme S., Li F., Chiang J. Y. L. G-protein-coupled bile acid receptor plays a key role in bile acid metabolism and fasting-induced hepatic steatosis in mice. Hepatology. 2017;65(3):813–827. doi: 10.1002/hep.28707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Guo C., Chen W. D., Wang Y. D. TGR5, not only a metabolic regulator. Frontiers in Physiology. 2016;7 doi: 10.3389/fphys.2016.00646. Published 2016 Dec 26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Martinot E., Sèdes L., Baptissart M., et al. Bile acids and their receptors. Molecular Aspects of Medicine. 2017;56:2–9. doi: 10.1016/j.mam.2017.01.006. [DOI] [PubMed] [Google Scholar]
- 84.Kim H., Fang S. Crosstalk between FXR and TGR5 controls glucagon-like peptide 1 secretion to maintain glycemic homeostasis. Laboratory Animal Research. 2018;34(4):140–146. doi: 10.5625/lar.2018.34.4.140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Biagioli M., Carino A., Cipriani S., et al. The bile acid receptor GPBAR1 regulates the M1/M2 phenotype of intestinal macrophages and activation of GPBAR1 rescues mice from murine colitis. Journal of Immunology. 2017;199(2):718–733. doi: 10.4049/jimmunol.1700183. [DOI] [PubMed] [Google Scholar]
- 86.Adrian T. E., Gariballa S., Parekh K. A., et al. Rectal taurocholate increases L cell and insulin secretion, and decreases blood glucose and food intake in obese type 2 diabetic volunteers. Diabetologia. 2012;55(9):2343–2347. doi: 10.1007/s00125-012-2593-2. [DOI] [PubMed] [Google Scholar]
- 87.Thomas C., Gioiello A., Noriega L., et al. TGR5-mediated bile acid sensing controls glucose homeostasis. Cell Metabolism. 2009;10(3):167–177. doi: 10.1016/j.cmet.2009.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Sonne D. P., Rehfeld J. F., Holst J. J., Vilsbøll T., Knop F. K. Postprandial gallbladder emptying in patients with type 2 diabetes: potential implications for bile-induced secretion of glucagon-like peptide 1. European Journal of Endocrinology. 2014;171(4):407–419. doi: 10.1530/EJE-14-0309. [DOI] [PubMed] [Google Scholar]
- 89.Adams T. D., Davidson L. E., Litwin S. E., et al. Health benefits of gastric bypass surgery after 6 years. Journal of the American Medical Association. 2012;308(11):1122–1131. doi: 10.1001/2012.jama.11164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Schauer P. R., Bhatt D. L., Kirwan J. P., et al. Bariatric surgery versus intensive medical therapy for diabetes–3-year outcomes. New England Journal of Medicine. 2014;370(21):2002–2013. doi: 10.1056/NEJMoa1401329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.McGavigan A. K., Garibay D., Henseler Z. M., et al. TGR5 contributes to glucoregulatory improvements after vertical sleeve gastrectomy in mice. Gut. 2017;66(2):226–234. doi: 10.1136/gutjnl-2015-309871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Lee W. J., Chen C. Y., Chong K., Lee Y. C., Chen S. C., Lee S. D. Changes in postprandial gut hormones after metabolic surgery: a comparison of gastric bypass and sleeve gastrectomy. Surgery for Obesity and Related Diseases. 2011;7(6):683–690. doi: 10.1016/j.soard.2011.07.009. [DOI] [PubMed] [Google Scholar]
- 93.Ding L., Sousa K. M., Jin L., et al. Vertical sleeve gastrectomy activates GPBAR-1/TGR5 to sustain weight loss, improve fatty liver, and remit insulin resistance in mice. Hepatology. 2016;64(3):760–773. doi: 10.1002/hep.28689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Ryan K. K., Tremaroli V., Clemmensen C., et al. FXR is a molecular target for the effects of vertical sleeve gastrectomy. Nature. 2014;509(7499):183–188. doi: 10.1038/nature13135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Kohli R., Bradley D., Setchell K. D., Eagon J. C., Abumrad N., Klein S. Weight loss induced by Roux-en-Y gastric bypass but not laparoscopic adjustable gastric banding increases circulating bile acids. The Journal of Clinical Endocrinology and Metabolism. 2013;98(4):E708–E712. doi: 10.1210/jc.2012-3736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Ferrell J. M., Chiang J. Y. L. Understanding bile acid signaling in diabetes: from pathophysiology to therapeutic targets. Diabetes and Metabolism Journal. 2019;43(3):257–272. doi: 10.4093/dmj.2019.0043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Mueller M., Thorell A., Claudel T., et al. Ursodeoxycholic acid exerts farnesoid X receptor-antagonistic effects on bile acid and lipid metabolism in morbid obesity. Journal of Hepatology. 2015;62(6):1398–1404. doi: 10.1016/j.jhep.2014.12.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Beysen C., Murphy E. J., Deines K., et al. Effect of bile acid sequestrants on glucose metabolism, hepatic de novo lipogenesis, and cholesterol and bile acid kinetics in type 2 diabetes: a randomised controlled study. Diabetologia. 2012;55(2):432–442. doi: 10.1007/s00125-011-2382-3. [DOI] [PubMed] [Google Scholar]
- 99.Henry R. R., Aroda V. R., Mudaliar S., Garvey W. T., Chou H. S., Jones M. R. Effects of colesevelam on glucose absorption and hepatic/peripheral insulin sensitivity in patients with type 2 diabetes mellitus. Diabetes, Obesity & Metabolism. 2012;14(1):40–46. doi: 10.1111/j.1463-1326.2011.01486.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Mudaliar S., Henry R. R., Sanyal A. J., et al. Efficacy and safety of the farnesoid X receptor agonist obeticholic acid in patients with type 2 diabetes and nonalcoholic fatty liver disease. Gastroenterology. 2013;145(3):574–582.e1. doi: 10.1053/j.gastro.2013.05.042. [DOI] [PubMed] [Google Scholar]
- 101.Jiang C., Xie C., Lv Y., et al. Intestine-selective farnesoid X receptor inhibition improves obesity-related metabolic dysfunction. Nature Communications. 2015;6(1) doi: 10.1038/ncomms10166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Burhans M. S., Hagman D. K., Kuzma J. N., Schmidt K. A., Kratz M. Contribution of adipose tissue inflammation to the development of type 2 diabetes mellitus. Comprehensive Physiology. 2018;9(1):1–58. doi: 10.1002/cphy.c170040. [DOI] [PMC free article] [PubMed] [Google Scholar]