

Abstract
Senolytics are a class of drugs that selectively clear senescent cells (SC). The first senolytic drugs Dasatinib, Quercetin, Fisetin and Navitoclax were discovered using a hypothesis‐driven approach. SC accumulate with ageing and at causal sites of multiple chronic disorders, including diseases accounting for the bulk of morbidity, mortality and health expenditures. The most deleterious SC are resistant to apoptosis and have up‐regulation of anti‐apoptotic pathways which defend SC against their own inflammatory senescence‐associated secretory phenotype (SASP), allowing them to survive, despite killing neighbouring cells. Senolytics transiently disable these SCAPs, causing apoptosis of those SC with a tissue‐destructive SASP. Because SC take weeks to reaccumulate, senolytics can be administered intermittently – a ‘hit‐and‐run’ approach. In preclinical models, senolytics delay, prevent or alleviate frailty, cancers and cardiovascular, neuropsychiatric, liver, kidney, musculoskeletal, lung, eye, haematological, metabolic and skin disorders as well as complications of organ transplantation, radiation and cancer treatment. As anticipated for agents targeting the fundamental ageing mechanisms that are ‘root cause’ contributors to multiple disorders, potential uses of senolytics are protean, potentially alleviating over 40 conditions in preclinical studies, opening a new route for treating age‐related dysfunction and diseases. Early pilot trials of senolytics suggest they decrease senescent cells, reduce inflammation and alleviate frailty in humans. Clinical trials for diabetes, idiopathic pulmonary fibrosis, Alzheimer’s disease, COVID‐19, osteoarthritis, osteoporosis, eye diseases and bone marrow transplant and childhood cancer survivors are underway or beginning. Until such studies are done, it is too early for senolytics to be used outside of clinical trials.
Keywords: senolytics, dasatinib, quercetin, fisetin, senescent cell anti‐apoptotic pathways, unitary theory of fundamental aging processes

Content List ‐ Read more articles from the symposium: “Cardiovascular program and cardiovascular retreat”.
Introduction
Ageing is associated with increasing risk for developing multiple chronic diseases, the geriatric syndromes, impaired physical resilience and mortality [1, 2, 3, 4]. These risks increase exponentially in the last 25% of the lifespan [5]. Amongst the diseases for which chronological age is a leading risk factor are congestive heart failure, myocardial infarction, dementias, strokes, most cancers, diabetes and metabolic diseases, renal dysfunction, chronic lung diseases, osteoporosis, arthritis, blindness and many others [6, 7, 8]. The geriatric syndromes include frailty, sarcopenia, falls, incontinence and mild cognitive impairment amongst others. Decreased resilience can be manifested as delayed recovery following myocardial infarction, strokes, injuries or surgery, increased severity of infections such as influenza and coronaviruses and impaired development of antibodies after immunization [3]. Combinations of several of these diseases, geriatric syndromes and resiliency impairments frequently occur in older people. Indeed, the incidence of multi‐morbidity, with 3 or more conditions being present simultaneously, increases exponentially beginning in adulthood [5]. Analogous increases in disease burden occur with increasing age across multiple species.
The Geroscience Hypothesis posits that fundamental ageing mechanisms are ‘root cause’ contributors to the increasing burden of disorders and diseases with advancing age that are responsible for the bulk of morbidity, mortality and health costs in the developed and developing worlds [9]. These fundamental ageing processes include: (1) chronic low grade ‘sterile’ (absence of bacteria, fungi, etc.) inflammation often accompanied by fibrosis, 2) macromolecular dysfunction (e.g. DNA damage, telomere uncapping, protein misfolding and aggregation, decreased autophagy, increased advanced glycation end‐products [AGEs], lipotoxicity and accumulation of bioactive lipids) and organelle dysfunction (altered nuclear membranes related to deficient lamin B, mitochondrial dysfunction leading to reduced fatty acid metabolism, higher glucose utilization, depletion of NAD+ and increased reactive oxygen species [ROS] generation, etc.), 3) stem, progenitor and immune cell dysfunction (including altered proliferative capacity and dysdifferentiation with failure to develop into functional mature cells, declines in ‘geroprotective’ factors [e.g. α‐Klotho], contributing to stem and progenitor cell dysfunction) and 4) cellular senescence. Our Unitary Theory of Fundamental Aging Processes hypothesizes that these fundamental ageing processes may be interlinked. If this Unitary Theory is correct, then accelerating or targeting any one fundamental ageing process (e.g. cellular senescence) genetically or with drugs should affect many or perhaps all of the rest. Indeed, consistent with the Unitary Theory, senescent cells contribute to inflammation, fibrosis, DNA damage, development of protein aggregates, failed autophagy, lipotoxicity, mitochondrial dysfunction, depletion of NAD+, ROS generation and stem, progenitor and immune cell dysfunction [10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24].
Cellular senescence
Cellular senescence, first reported in 1961 by Hayflick and Moorehead, is a cell fate that entails essentially irreversible replicative arrest, sustained viability with resistance to apoptosis, and frequently, increased metabolic activity [6, 8, 25, 26, 27] (Fig. 1 ). Intra and extracellular signals that can contribute to cells’ entering the senescent cell fate mainly include signals related to tissue or cellular damage and/or cancer development. These include DNA damage, telomeric uncapping or dysfunction, exposure to extracellular DNA, oncogene activation, replicative stress or inducers of proliferation (such as growth hormone/IGF‐1), protein aggregates, misfolded proteins, failed protein removal through decreased autophagy, presence of AGEs due to the reaction of reducing sugars with amino groups in proteins (e.g. Haemoglobin A1c is an AGE), saturated lipids and other bioactive lipids (bradykines, certain prostaglandins, etc.), reactive metabolites (e.g. ROS, hypoxia or hyperoxia), mechanical stress (e.g. bone‐on‐bone stress in osteoarthritis or shear stress such as occurs on the venous side of AV fistulae for haemodialysis or around atherosclerotic plaques), inflammatory cytokines (e.g. TNFα), damage‐associated molecular patterns (DAMPs, e.g. released intracellular contents signalling breakage of neighbouring cells), and pathogen‐associated molecular patterns (PAMPs, e.g. bacterial endotoxins). These inducers active one or more senescence‐promoting transcription factor cascades, in some cases involving p16INK4a‐retinoblastoma protein (Rb), in others, p53 and p21CIP1, both of these pathways, or other pathways. These transcription factor cascades enforce replicative arrest and cause altered expression of hundreds of genes as well as epigenetic changes in DNA. Cellular senescence takes longer to become established than other cell fates, such as replication, differentiation, apoptosis or necrosis. From initiation to the attainment of a completed state of cellular senescence takes from 10 days to 6 weeks, at least in cell culture, depending on the cell type and the inducers driving the cell into the senescent fate.
Figure 1.
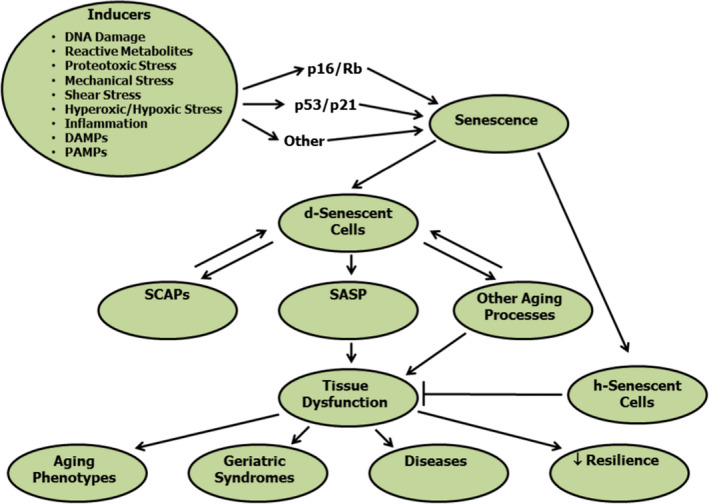
Cellular senescence: causes, mechanisms and consequences. A number of factors can combine to cause a cell to enter the senescent cell fate, as opposed to apoptosis or necrosis, through transcription factor cascades that can involve p16INK4a, Rb, p53, p21CIP1 or others. Note that not every senescent cell exhibits increased p16INK4a, Rb, p53 or p21CIP1. It can take up to 6 weeks for a cell to become a fully senescent, nondividing cell. This indicates that senescent cells can be removed intermittently, rather than having to treat with agents continuously to remove these cells. Some, but not all senescent cells can develop a SASP. This SASP is a ‘root cause contributor’ to the stem cell, progenitor, tissue and systemic dysfunction that predispose to multiple disorders, which account for the bulk of morbidity, mortality and health costs.
Some, but not all senescent cells can acquire a senescence‐associated secretory phenotype (SASP) [28, 29], which we term here as deleterious (D‐) senescent cells (Fig. 2 ). The SASP can include: 1) inflammatory, pro‐apoptotic, insulin resistance‐inducing cytokines, such as TNFα [30], interleukin‐ (IL‐) 6, IL‐8 and others, 2) chemokines that attract, activate and anchor immune cells, 3) matrix metalloproteinases (MMPs), such as MMP‐3, −9 and −12 that cause tissue destruction, 4) TGFβ family members that can contribute to fibrosis and stem cell and progenitor dysfunction, 5) activins and inhibins that also induce stem cell and progenitor dysfunction and dysdifferentiation [12], 6) factors such as the serpines (e.g. plasminogen activator inhibitor [PAI] −1 and −2) that can cause blood clotting and fibrosis [31, 32], 7) growth factors that can exacerbate tumour spread [33], 8) bioactive lipids that also contribute to inflammation and tissue dysfunction (e.g. bradykines, ceramides or prostaglandins) [34], 9) micro‐RNAs (miRNAs) that contribute to stem and progenitor cell dysfunction, inflammation and insulin resistance [35] and 10) exosomes that can carry cytotoxic and senescence‐inducing cargos locally and systemically [36]. D‐senescent cells can comprise 30 to 70 % of the senescent cell population. SASP components produced by D‐senescent cells depend on the cell type that became senescent and the cause of senescence (e.g. mechanical stress, repeated replication, radiation, chemotherapy, PAMPs, etc.).
Figure 2.
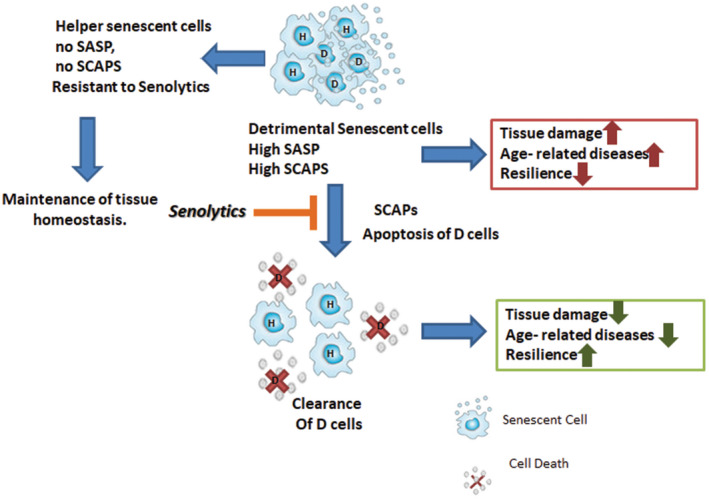
Helper (H) and (D) senescent cells. D‐senescent cells have a pro‐inflammatory, pro‐apoptotic, tissue‐damaging SASP and are susceptible to senolytics. H‐senescent cells do not have a SASP and do not up‐regulate SCAPS and are not cleared by senolytics. H‐ senescent cells appear to promote stem and progenitor cell determination into appropriate lineages and functions and have a beneficial impact on tissue homoeostasis.
The SASP can be modulated by hormones, drugs, pathogens and other factors. For example, endogenous or pharmacological glucocorticoids [37] can attenuate release of a subset of SASP factors, as can rapamycin and metformin [38, 39]. Bacterial toxins such as lipopolysaccharide can exacerbate the SASP, as can TNFα [40] (unpublished observations). The SASP can vary over time due to factors inside or outside of senescent cells. For example, after a few months, transpositional events can begin to occur in senescent cells involving LINE‐1 or other elements [41, 42]. This can lead to rearrangement of chromosomes and activation of cGAS/STING, cellular pathways that recognize DNA damage and exacerbate production of inflammatory SASP factors through interferon‐1. Thus, the SASP varies considerably depending on the tissue where D‐senescent cells are located, the originating cell type, senescence inducers, hormonal milieu, drugs, pathogens and passage of time.
Other senescent cells, termed here helper‐ (H‐) senescent cells, do not appear to release inflammatory and pro‐apoptotic factors substantially and may be involved in promoting stem and progenitor cell determination into appropriate lineages and functions [43, 44] (unpublished observations). H‐senescent cells might account for 30 to 70% of senescent cells. We speculate there could be mechanisms through which D‐ and H‐senescent cells can interconvert, but this remains to be fully explored.
D‐ and H‐senescent cells may both have beneficial functions, including remodelling of tissues during development, release of factors by placental senescent cells during childbirth that promote passage of the infant through the birth canal, removal of damaged tissue during initial phases of wound repair, and cancer defence [44, 45, 46, 47]. Cellular senescence can protect against cancer: senescence is induced by oncogenic insults, leading to replicative arrest of cancerous cells, as well as a pro‐apoptotic SASP that can eliminate surrounding tumour cells. Additionally, interfering with mechanisms involved in ability of cells to become senescent by decreasing capacity to express p16INK4a, Rb, p53 or p21CIP1 promotes cancer development [48, 49]. However, clearing already formed senescent cells, many of which harbour oncogenic mutations or cause cancer‐promoting inflammation due to their SASP, prevents or delays cancer development [46]. Indeed, senolytic drugs, agents that selectively eliminate senescent cells [50], delay development of multiple forms of cancer in old mice as well as cancer‐prone, DNA repair‐deficient Ercc1‐/Δ mice [51, 52]. Furthermore, although capacity for cells to become senescent is a cancer defence mechanism, already formed, chronically persisting senescent cells can accelerate cancer growth because of suppression of immune responses, growth factors in the SASP, production of proteases that can interfere with encapsulation of tumours and therefore can enhance spread of cancer, and promotion of mesenchymal transition of cells [10, 53]. Thus, targeting transcription factor cascades that allow cells to become senescent can exacerbate cancer development, whilst clearing already formed senescent cells with senolytics can prevent cancer development, growth and spread.
There are no completely sensitive or specific markers of senescent cells. Most senescent cells are larger than nonsenescent cells. They generally have multiple DNA damage foci identifiable by immunostaining for γ‐histone‐2AX (γH2AX), but cancer cells and other cell types, including normal cells, can also have γH2AX‐positive foci [54]. In senescent cells, frequently 2 or more γH2AX foci can be located within telomeric DNA (telomere‐associated foci [TAFs]), one of the more sensitive and specific indicators of senescent cells [55]. Senescent cells can develop peri‐centromeric distended satellite DNA (senescence‐associated distension of satellites [SADS]), which can be detected with antibodies to CENP‐B, another marker for detecting senescent cells [56]. Senescent cells can have increased lysosomal β‐galactosidase activity (senescence‐associated β‐galactosidase [SA‐β‐gal]), but so can macrophages and other cell types [57, 58]. Some, but not all senescent cells may have high expression of p16INK4a, but p16INK4a expression can be high in activated macrophages and many other cell types [59]. Like p16INK4a, p21CIP1 is not fully sensitive or specific for detecting senescent cells [60]. A subset of senescent cells can express SASP factors or damage signals, such as cytosolic HMGB1 [61], but again, these indicators are not fully sensitive or specific. Because of the less than perfect specificity and sensitivity of markers for cellular senescence, generally several need to be tested. This will especially be the case for the initial clinical trials of senolytics for different diseases that are already underway.
Some senescent cells have enlarged, fused mitochondria and changes in mitochondrial function, including a shift from utilizing fatty acids for energy generation towards utilizing glucose, much like the ‘Warburg shift’ in cancer cells [62, 63]. This shift to dependence on glycolysis can be associated with lipid accumulation, resulting in lipid droplet accumulation and lipotoxicity, contributing to an adipocyte‐like appearance of senescent mesenchymal cells or ‘MAD cells’ (mesenchymal adipocyte‐like default cells) that are insulin‐insensitive [15]. An example is the accumulation of pro‐inflammatory, perilipin‐expressing, lipid‐laden senescent ependymal MAD cells around the third ventricle in obesity [24]. These cells contribute to neuro‐inflammation, failed brain microcirculation, impaired neurogenesis and neuropsychiatric dysfunction in obese, insulin‐resistant mice.
Although senescent cells can appear at any point during life in vertebrates, even in 32 cell stage human blastocysts [45], senescent cells accumulate in multiple tissues during the latter part of the lifespan. For example, skin senescent cells increase sharply in otherwise healthy humans at a point usually between ages 60 and 80 (although there has been debate about this) [64, 65, 66, 67, 68]. The same is true for human adipose tissue biopsied from healthy subjects donating a kidney as well as adipose depots of ageing mice [22, 69]. Senescent cells can accumulate at aetiological sites of multiple diseases throughout the lifespan. For example, in humans, senescent cells accumulate in adipose tissue in diabetes and obesity, in the hippocampi and frontal cortex in Alzheimer’s disease (AD), the substantia nigra in Parkinson’s disease (PD), bone and marrow in age‐related osteoporosis, lungs in idiopathic pulmonary fibrosis, liver in cirrhosis, retinae in macular degeneration, plaques in psoriasis, kidneys in diabetic kidney disease, endothelium in pre‐eclampsia, and the heart and major arteries in cardiovascular disease, amongst many other conditions [6, 7, 13, 19, 23, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80].
Consistent with a causal contribution of senescent cells to chronic diseases, transplanting small numbers of autologous or syngeneic senescent cells near the knee joints of younger mice causes an osteoarthritis‐like state, with radiographic findings similar to those of human spontaneous osteoarthritis, as well as knee pain and impaired mobility, whilst transplanting nonsenescent cells does not [81]. Transplanting 106 syngeneic adipose cell progenitors or autologous ear fibroblasts made senescent by irradiation, doxorubicin exposure or repeated replication is sufficient to cause frailty and accelerate death from all causes, not just one or a few, in middle‐aged mice, whilst transplanting nonsenescent cells does not [51]. Transplanting 106 senescent human cells into immunodeficient mice also causes frailty. Additionally, transplanting senescent porcine renal scattered tubular‐like cells into the aorta of young mice causes kidney fibrosis and inflammation in the recipients, whilst transplanting nonsenescent cells does not [82].
There is a lag between senescent cell accumulation and development of functional effects. Frailty was evident a month after transplanting senescent cells into middle‐aged mice and increased mortality became apparent 3 months later [51]. This study demonstrated that if only 1/10,000 cells in recipient mice is a transplanted senescent cell, an accelerated ageing‐like state ensues. Transplanting 500 000 senescent cells did not do this, suggesting a senescent cell ‘threshold effect’. Consistent with this, transplanting 500 000 senescent cells into old mice, as opposed to middle‐aged recipients, did cause frailty. Furthermore, transplanting 500 000 senescent cells into middle‐aged but diet‐induced obese (DIO) mice caused frailty, whilst 106 senescent cells had to be transplanted into age‐matched, lean mice for frailty to occur. Thus, pre‐existing senescent cell burden, together with acute acquisition of yet more senescent cells, can surpass a threshold and cause clinical effects.
A potential reason for this ‘threshold effect’ is that D‐senescent cells can spread senescence to normal cells, even at a distance [51]. If senescent cells are labelled with luciferase, so transplanted senescent cells can be distinguished from the recipients’ own cells, and the labelled senescent cells are transplanted into the abdominal cavity where they remain, senescent cells originating from the recipient’s own cells appear throughout the recipient, including in the limbs. Since senescent cells resist cell death and are normally removed by the immune system [10], the threshold effect may be a result of the rate of spread of senescence exceeding the capacity of the immune system to clear them. This’Threshold Theory of Cellular Senescence’ may explain the rapid increase in senescent cell abundance in skin and adipose tissue of otherwise healthy people at a point after age 60 [22, 64, 65, 66, 67], as well as the lag between the first histological evidence for increased senescent cell abundance and subsequent development of symptoms.
Discovery of senolytic drugs
An article by the Sharpless group in 2004 led us to start searching for interventions that remove senescent cells [83]. They reported delayed accumulation of p16Ink4a‐, SA‐β‐gal‐positive cells in mice in which healthspan had been extended by caloric restriction or a mutation that decreases growth hormone signalling. This indicates there is an inverse association between senescent cell burden and healthspan. Beginning in 2005, we attempted to develop fusion proteins with a senescent cell recognition site and a toxin to kill senescent cells. We then attempted to find chemicals that would kill senescent but not nonsenescent cells using high‐throughput screens of human senescent vs. nonsenescent cells. Neither approach was successful. The high‐throughput screening approach suffered from difficulties in selecting the right controls. Parallel nondividing, nonsenescent cells had to be serum‐starved or confluent in order to prevent replication. However, serum starvation or confluence both cause artefactual effects. Alternatively, dividing nonsenescent cells could be used as controls. However, these dividing control cells have to be compared to senescent cells that, by definition, cannot divide, introducing a confounder that interferes with interpretation of the results.
Thus, rather than random high‐throughput drug library screening or other traditional drug development strategies, we turned to a hypothesis‐driven, mechanism‐based drug discovery paradigm to develop the first senolytic drugs. Our strategy was based on the 1995 observation by E. Wang that senescent cells resist apoptosis [84]. This apoptosis resistance occurs despite senescent cell production of pro‐apoptotic SASP factors. The existence of the SASP had been discussed within the field for a couple of years before it was published [28, 29, 85]. Our hypotheses upon which senolytic drug discovery was based were: 1) Senescent cells can resist apoptotic stimuli, implying increased pro‐survival ‒ anti‐apoptotic defences and 2) In some respects, senescent cells are like cancer cells that do not divide, including apoptosis resistance and metabolic shifts. We speculated that senescent cells might prevent their own removal through protective Senescent Cell Anti‐Apoptotic Pathway (SCAP) networks [86]. From proteomic and transcriptomic databases, we found that SCAP pathways were more highly expressed by senescent than nonsenescent cells [86]. We interrogated the SCAP network using RNA interference and demonstrated that senescent cells with a pro‐apoptotic SASP do indeed depend on this network to evade self‐destruction.
The small interfering RNA’s (siRNA’s) against anti‐apoptotic regulators that were effective in selectively decreasing senescent cell viability depended on the senescent cell type. For example, we found that siRNA’s that decrease BCL‐xL or other BCL‐2 family members were effective in selectively causing apoptosis of senescent vs. nonsenescent human umbilical vein endothelial primary cells (HUVEC’s) [86], as was confirmed by others [87]. However, the same siRNA’s were ineffective in selectively inducing apoptosis of senescent vs. nonsenescent human primary adipocyte progenitor cells. Furthermore, more than one SCAP pathway had to be targeted to kill some types of senescent cells, indicating that senescent cell defences against apoptosis are in some cases redundant.
Next, we used bioinformatics approaches to find compounds whose mechanisms of action involve targeting the SCAP network nodes that we had identified as being critical for protecting senescent cells from apoptosis. We identified 46 potentially senolytic compounds. To accelerate translation into clinical application, the compounds that we focused on initially were drugs and natural products already in use for other indications in humans, many as anti‐cancer agents. These included the tyrosine kinase inhibitor, Dasatinib (D), which has been approved for clinical use in the United States since 2006, and Quercetin (Q), the naturally occurring flavonoid that makes apple peels taste bitter. Unlike other tyrosine kinase inhibitors such as Imatinib, which are not senolytic, D promotes apoptosis caused by dependence receptors, such as the ephrins, in part by inhibiting Src kinase [88, 89]. Fisetin (F) is another naturally occurring flavonoid that is senolytic [52, 90]. The senolytic flavonoids act in part by inhibiting BCL‐2 family members such as BCL‐xL as well as HIF‐1α and other SCAP network components.
Based on their known targets, we predicted that D would target senescent human cultured primary adipocyte progenitor cells, whilst Q would target senescent cultured HUVEC’s. These predictions proved to be correct. The combination of D + Q caused apoptosis of both senescent human primary adipocyte progenitor cells and senescent HUVEC’s, but not nonsenescent adipocyte progenitor cells or HUVEC’s. Furthermore, likely because of redundancy of SCAP pathways, in some cell types, for example, mouse embryonic fibroblasts, neither D nor Q are senolytic on their own, whilst the combination of D + Q is senolytic [86].
The target of true senolytics is senescent cells, not a single receptor, enzyme or biochemical pathway. Efforts to discover senolytics by finding agents that act on multiple SCAP network nodes effectively increases specificity for senescent cells, reduces off‐target effects on nonsenescent cells compared with candidates that act on a single molecule, such as a receptor or an enzyme, and likely flatten side‐effect profiles. Consistent with this prediction, candidates that do act upon single or limited targets, for example, Navitoclax (N) [91, 92] or Nutlin3a [93], have substantial off‐target apoptotic effects on nonsenescent cell types, such as platelets and immune cells [94, 95], making them potentially ‘panolytic’. Also, nutlins can actually cause senescence [96] and drugs like N, which act on a single or few SCAP nodes, eliminate only a restricted range of senescent cells. For example, N and other BCL‐2 family member inhibitors such as A1331852 or A1155463, are not senolytic against human adipocyte progenitors, one of the most abundant senescent cell types in the elderly as well as patients with diabetes and obesity [90, 91]. Thus, candidates that act on only one or a limited range of SCAP‐related molecules are generally less specifically senolytic and are more ‘panolytic’, causing apoptosis or dysfunction of multiple cell types other than senescent cells, than agents or combinations of agents that act on multiple nodes across the SCAP network.
The first senolytics were discovered using bioinformatics approaches to identify agents that transiently disable the SCAP networks that allow D‐senescent cells to survive the hostile microenvironment they create that kills the nonsenescent cells around them (Table 1 ). In this respect, the development of senolytics has been distinct from the conventional approaches for developing drugs usually followed in other fields. Approaches for developing senolytics has held more in common with those used to develop antibiotics than the traditional one target‐one drug‐one disease approach. A second generation of senolytic agents is now being identified using more traditional drug discovery methods, including random high‐throughput drug library screens, such as one in which both senescent and nonsenescent cells are within the same wells, overcoming the control cell problem [97]. Several other strategies have recently been used to develop potentially senolytic interventions. These include the development of galacto‐oligosaccharide‐coated nanoparticles with toxic cargos that are engulfed by senescent cells and opened by SA‐β‐gal, vaccines and immunomodulators [10, 98, 99, 100]. Perhaps some of these will make it through preclinical studies and into clinical trials within the next few years.
Table 1.
Routes to discovering senolytics
| 1st generation senolytics: hypothesis‐driven, mechanism‐based | |
|---|---|
| Agent | References |
| Dasatinib | [87] |
| Quercetin | [87] |
| Fisetin | [52, 90] |
| Luteolin | [52] |
| Curcumin | [52] |
| Curcumin Analog EF24 | [102] |
| Navitoclax (ABT263) | [91, 92] |
| A1331852 | [91] |
| A1155463 | [91] |
| Geldanamycin | [103] |
| Tanespimycin | [103] |
| Alvespimycin | [103] |
| Piperlongumine | [104] |
| FOXO4‐related peptide | [105] |
| Nutlin3a (although Nutlin3a can also cause senescence | [93, 96] |
| Cardiac glycosides: Ouabain, Proscillaridin A, Digoxin, others | [105, 106] |
D + Q and F have been shown to reduce senescent cell abundance in vivo whilst not destroying nonsenescent cells [51, 52]. After intraperitoneal transplantation of 105 radiation‐induced syngeneic senescent adipocyte progenitors, which could be detected due to constitutive SFFV promoter‐driven luciferase expression, into middle‐aged mice, treatment with oral D + Q for 3 days substantially decreased luminescence 2 days after the last dose [51]. No decrease in luminescence was evident after treating senescent cell‐transplanted mice with vehicle, nor was decreased luminescence found after treating mice transplanted with nonsenescent luciferase‐expressing cells with either D + Q or vehicle. Oral treatment with F of progeroid Ercc‐/Δ mice expressing luciferase driven by a p16Ink4a promoter element also resulted in decreased luminescence, whilst treating these mice with vehicle had no effect. Thus, D + Q and F are senolytic in mice, each eliminating 30 to 70% of transplanted or endogenous senescent cells, respectively.
To test if D + Q is senolytic in human tissue, abdominal subcutaneous adipose tissue fragments that were freshly isolated during surgery from patients with diabetes and obesity, who have extensive adipose tissue senescent cell accumulation [108, 109, 110, 111, 112], were immediately placed in organ culture and exposed to D + Q or vehicle [51]. Within 2 days, abundance of senescent cells detected by TAFs had decreased by 70%. This was accompanied by increases in cleaved caspase‐3, confirming that the decrease was caused by apoptosis. Furthermore, the SASP factors, IL‐6, IL‐8, MCP‐1, PAI‐1 and GM‐CSF, decreased significantly in conditioned medium from the organ cultures treated with D + Q compared to vehicle. Also, F reduced senescent cell number in adipose tissue organ cultures [52]. Thus, D + Q and F cause decreased senescent cell abundance by apoptosis in human tissue within 2 days.
Senolytics alleviate multiple disorders in experimental animals
Senolytic drugs alleviate a number of disorders in mouse disease models [6, 7, 8, 108] (Table 2 ). Intermittent oral administration of D + Q enhances cardiac and vascular function in 24‐month‐old mice (equivalent to roughly 80 years in humans), with partial reversal of age‐related declines in cardiac ejection fraction, reduction in conduit vessel calcification and restoration of conduit vessel reactivity to nitroprusside and acetylcholine [113]. D + Q decreases cardiac senescent cell burden, as manifested by lowering of p16Ink4a, and partly reverses age‐related cardiomyocyte hypertrophy and left ventricular fibrosis [13]. Senescent cells can develop in the venous arm of arterial‐venous (AV) fistulae of the type surgically fashioned for haemodialysis, potentially contributing to clotting and failure of the AV fistulae [114]. In mice, D + Q decreases senescent cell abundance in AV fistulae. Furthermore, D + Q reduces senescent cell burden and calcification in the aortic media of high fat‐fed ApoE‐/‐ mice, a mouse model of atherosclerosis [113]. N is also senolytic in the hearts of old mice [18]. Following 1 month of N, there was decreased cardiac fibrosis and a 40% decrease in TAF+ cardiomyocytes (it appears nondividing cells such as cardiomyocytes can develop a senescent‐like state). Thus, senolytics alleviate age‐ and lipid‐related cardiovascular dysfunction in mice.
Table 2.
Conditions with emerging evidence for a causal contribution of cellular senescence or benefits of senolytics
| Condition | References |
|---|---|
| Diabetes/ Obesity | [108, 109, 111, 115, 116, 117] |
| Cardiac dysfunction | [13, 18, 113] |
| Vascular hyporeactivity/ calcification | [114] |
| AV fistulae | [115] |
| Frailty | [21, 51, 52, 86] |
| Age‐related muscle loss (Sarcopenia) | [119] |
| Chemotherapy complications | [21, 51, 92, 119, 120] |
| Radiation complications | [122] |
| Cancers | [51] |
| Bone marrow transplant complications | [120] |
| Organ transplantation complications | [122, 123] |
| Myeloma/ MGUS | [125] |
| Age‐related cognitive dysfunction | [126] |
| Alzheimer’s disease | [23, 126] |
| Parkinson’s disease | [76] |
| Amyotrophic lateral sclerosis | [128] |
| Ataxia | [87] |
| Obesity‐related neuropsychiatric dysfunction | [24] |
| Renal dysfunction | [110, 128] |
| Urinary incontinence | [87] |
| Osteoporosis | [14, 71, 129] |
| Osteoarthritis | [131] |
| Age‐related intervertebral disc disease | [86, 131] |
| Idiopathic pulmonary fibrosis | [21, 132] |
| Hyperoxic lung damage | [134] |
| Chronic obstructive pulmonary disease | [135] |
| Tobacco | [136] |
| Hepatic steatosis | [137] |
| Cirrhosis | [78] |
| Primary biliary cirrhosis | [138] |
| Progerias | [52, 86] |
| Pre‐eclampsia | [72] |
| Macular degeneration | [74, 138] |
| Glaucoma | [139, 140, 141] |
| Cataracts | [143] |
| Prostatic hypertrophy | [143, 144, 145] |
| Psoriasis | [81] |
| Healthspan | [51, 52, 86] |
| Lifespan | [51, 52, 86] |
Intermittent oral D + Q administration enhances insulin sensitivity in DIO as well as genetically obese mice, as manifested by improvements in glucose and insulin tolerance [111]. D + Q does so mainly by reducing peripheral insulin resistance. This is in part due to restoring adipose tissue AKT responses to insulin and in part to restoration of the capacity of adipocyte progenitors to differentiate into insulin‐responsive adipocytes. Furthermore, intermittent D + Q reduces the adipose tissue inflammation that is a causal contributor to the pathogenesis of type 2 diabetes. D + Q does so in part by reducing homing of monocytes to adipose tissue: there is decreased trafficking of luciferase‐expressing monocytes injected into the tail vain into adipose tissue of genetically obese mice treated with D + Q, but not in mice treated with vehicle. D + Q does not kill the activated macrophages that can accumulate in adipose tissue in diabetes [51]. These macrophages have increased p16 INK4a expression but are not senescent [59]. Consistent with this, senolytic drugs also do not target atherosclerotic foam cells, which are essentially activated macrophages, as opposed to the truly senescent cells that are deeper within plaques or the media of atherosclerotic blood vessels that are ablated by D + Q [113]. N, which is not effective in targeting senescent adipocyte progenitors [91], does decrease senescent pancreatic β cells, enhancing pancreatic function in DIO mice [115]. Thus, senolytics can alleviate diabetes by decreasing SASP‐expressing adipose tissue senescent cells, leading to increased peripheral insulin sensitivity, reducing chemo‐attraction and activation of pro‐inflammatory macrophages that further contribute to insulin resistance, and/or enhancing pancreatic insulin production.
Senolytics decrease several complications of diabetes in mouse models. Liver fat accumulation and consequent cirrhosis and liver failure are increasingly common complications of diabetes and obesity [108, 109]. D + Q reduces liver senescent cell accumulation related to hepatic steatosis in genetically obese mice [136]. Additionally, proteinuria and renal dysfunction are frequent complications of diabetes. DIO mice develop increased kidney senescent cell burden manifested by p16Ink4a and p53 expression and SASP factors in renal tubular cells along with glomerulomegaly, decreased renal function and impaired cortical oxygenation compared to lean mice [128]. Proteinuria is reduced in DIO mice treated with D + Q [110]. DIO mice with renal dysfunction that were treated with oral Q for 5 days every 2 weeks for 10 weeks had decreased kidney senescent cells, decreased SASP factors, decreased renal fibrosis, increased renal cortical oxygenation and decreased plasma creatinine levels compared to mice treated with vehicle [128].
Another complication of diabetes/obesity is neuropsychiatric dysfunction, particularly anxiety and its complications [24]. In genetically obese mice, senescent adipocyte‐like ependymal MAD cells can accumulate near the third ventricles near the limbic system, which regulates emotions. D + Q reduces the burden of these senescent adipocyte‐like periventricular ependymal cells. Administering D + Q also increases neuronal markers, indicating improved neurogenesis and reduces neuro‐inflammation in obese mice. Importantly, intermittent D + Q alleviates anxiety in these obese mice, as manifest by reduced fear in open‐field testing. Thus, senolytics not only restore peripheral insulin responsiveness in diabetic/obese mice, they alleviate complications of diabetes for which no effective mechanism‐based treatments currently exist.
In addition to diabetic/obesity‐related neuropsychiatric dysfunction, senolytics alleviate Alzheimer’s‐like changes in Tau+ as well as β‐amyloid+ (Aβ) mouse models of Alzheimer’s disease [23, 126]. Accumulation of Tau protein aggregates is a common pathology across multiple degenerative brain diseases, including AD, progressive supranuclear palsy, traumatic brain injury and over twenty others. Furthermore, Tau‐containing neurofibrillary tangles correlate with cognitive decline and brain cell loss. Neurofibrillary tangle‐containing neurons from postmortem human AD brains have expression profiles that are consistent with cellular senescence. Also, in both mice and humans, p16INK4a expression correlates directly with brain atrophy and neurofibrillary tangles. In 23‐month‐old tauNFT‐Mapt0/0 mice that overexpress Tau, D + Q every 2 weeks for 3 months reduced total neurofibrillary tangle density, neuronal loss, neuro‐inflammation, gliosis, small vessel hypoperfusion and ventricular enlargement and increased cortical volume and neurogenesis, despite the advanced age of these mice. Administering D + Q to Aβ+ mice was also effective, with selective removal of senescent cells from the AD‐associated plaque environment, reduced neuro‐inflammation, decreased Aβ load and alleviation of deficits in cognitive function, with improved memory and executive function [126]. Thus, senolytic drugs may potentially prove to be effective, mechanism‐based interventions for alleviating AD and other chronic brain diseases, depending on results of the clinical trials that are underway.
Senolytics alleviate dysfunction in mouse models of chronic lung diseases. Idiopathic pulmonary fibrosis (IPF) is a progressive, fatal, senescence‐driven lung disease for which there are no highly effective interventions apart from lung transplantation [21]. It affects older patients, leading to respiratory failure, pronounced frailty and weight loss. Targeting senescent cells with D + Q was effective in alleviating frailty as shown by improved treadmill endurance, reducing extent of weight loss, and enhancing lung compliance in a mouse bleomycin inhalation‐induced pulmonary fibrosis model of IPF. Another lung condition that might be alleviated by senolytics is hyperoxia‐induced damage to airway structure and function [133]. Supplemental O2 can be a necessary intervention in premature infants, but appears to contribute to development of neonatal and paediatric asthma. At least in cell culture, D + Q shows promise for alleviating complications of supplemental O2. Foetal airway smooth muscle (ASM) exposed for 7 days to 40% O2 developed markers of senescence, including SA‐βgal and increased p16INK4a, p21CIP1, p‐p53, γH2A.X and pro‐fibrotic and inflammatory SASP factors. Treating naïve ASM cells with media from hyperoxia‐exposed senescent ASM cells increased collagen, fibronectin and contractility. D + Q selectively reduced the number of hyperoxia‐induced senescent ASM cells. Hence, senolytic drugs may become interventions for multiple lung diseases. Indeed, in the first clinical trial of senolytics, D + Q alleviated physical dysfunction in a preliminary study in patients with IPF [70] (see below). Additionally, the senolytic, F, reduces mortality in mice infected with mouse β‐coronavirus and COVID‐19 viral antigens exacerbate the SASP in human senescent cells (unpublished observations). Based on these discoveries, a clinical trial of F for older hospitalized COVID‐19 patients to prevent progression to cytokine storm and acute respiratory distress syndrome has been approved by the US Food and Drug Administration (FDA) and is about to start.
Age‐related, as opposed to hormone‐deficiency‐related, osteoporosis appears to be associated with senescent cell accumulation, particularly senescent‐like osteocytes and bone marrow cells [71, 129]. Osteoclasts cause bone resorption. Conditioned medium from senescent cells increases early monocyte osteoclast progenitors due to enhanced survival of these cells [14]. The SASP factors, IL‐6, IL‐8 and PAI‐1, contribute to this increased survival of osteoclast progenitors. In old (20 months) mice with osteoporosis, D + Q treatment once monthly for 4 months led to significant reductions in bone p16Ink4a mRNA and SADS+ senescent osteocytes [14]. In D + Q‐treated mice, trabecular bone histomorphometry demonstrated reduced osteoclasts vs. in vehicle‐treated mice. D + Q treatment increased femoral cortical thickness and bone strength. This was associated with reduced endocortical surface osteoclast numbers and increases in endocortical numbers of osteoblasts, the cells that form new bone, mineral apposition rate and bone formation rate. These beneficial effects of targeting senescent cells resulted in suppression of bone resorption accompanied by either maintenance (trabecular) or increased (cortical) bone formation. Thus, unlike the coupled decreases in bone resorption and formation that results from currently available antiresorptive treatments for osteoporosis, D + Q causes decreased bone resorption whilst not causing a coupled decrease in bone formation. These beneficial effects result from a decrease in development of bone‐resorbing osteoclasts and an increase in development of bone‐forming osteoblasts from their respective stem cells/progenitors. Furthermore, intermittent D + Q was as effective as continuous D + Q, consistent with our contention that a ‘hit‐and‐run’ approach of senolytic administration is effective and could decrease adverse effects. A clinical trial of D + Q for age‐related osteoporosis is currently underway (see below). N also decreases bone senescent cells in old mice but, unlike D + Q, does not alleviate age‐related osteoporosis because, consistent with its ‘panolytic’ side‐effect profile, N is directly cytotoxic to bone‐forming osteoblasts [146].
In addition to age‐related osteoporosis [129] and osteoarthritis [130], senescent cells contribute to another skeletal disorder, age‐related intervertebral disc disease. Glycosaminoglycan content of the nucleus pulposus of intervertebral discs declines with ageing in progeroid mice [86]. D + Q increases spinal glycosaminoglycan levels in progeroid mice compared to animals receiving vehicle only. Since senolytics increase vertebral proteoglycans in preclinical models, they might prove to be an effective treatment for intervertebral disc disease.
D + Q or F alleviate physical dysfunction across a range of mouse models in which senescent cells are linked to frailty: naturally aged mice, progeroid Ercc‐/Δ mice, senescent cell‐transplanted younger mice and mice with bleomycin‐induced pulmonary fibrosis [21, 51, 52, 86]. In these mouse models, senolytics prevented frailty or restored physical function as assessed by daily activity (movement detected by laser measurement), distance run on a treadmill, treadmill speed, grip strength, time able to hang on a suspended wire, and/or ability to remain on a rotating spindle (RotaRod test). D + Q as well as F alone both extended remaining lifespan in old mice by up to 35% [52, 86]. Consistent with the Geroscience Hypothesis, D + Q delayed death in old mice not due to any single age‐related disease, but from multiple disorders that cause death in naturally aged, untreated mice [86].
As would be expected from their use as anticancer drugs, the senolytics D + Q and F delay death from cancers in mice [51, 52]. Over 50% of deaths in most mouse strains are caused by cancers, and D + Q and F delay deaths of old mice by up to 35 and 17%, respectively. Furthermore, even though F might cause aneuploidy in cultured nonsenescent cells [147], F delays cancers in cancer‐prone Ercc1‐/Δ mice that have a DNA repair enzyme mutation [52].
In these preclinical studies, it would be desirable to prove that a candidate agent alleviates a phenotype because it is actually a senolytic drug. An approach for proving senolytic effects is to use a modified set of Koch’s postulates. If an agent is truly senolytic, then: 1) Senescent cells should be present in association with the phenotype, 2) Individuals without senescent cells should not have the phenotype, 3) Inducing accumulation of senescent cells should cause the phenotype, 4) Clearing these induced senescent cells should alleviate the phenotype, 5) Clearing naturally occurring senescent cells should alleviate the phenotype, 6) The drug should induce few or no effects related to the phenotype in young individuals without senescent cells, 7) Administering the senolytic candidate intermittently should be effective and 8) The senolytic candidate should alleviate multiple age‐related conditions. In the case of D + Q, these modified Koch’s postulates have been met in mice for age‐ and cellular senescence‐related physical dysfunction, insulin resistance, cognitive dysfunction, osteoporosis, osteoarthritis and cancers in the published studies cited above and in unpublished studies. Evidence consistent for a true senolytic effect of F is mounting. To our knowledge, there is not yet comprehensive evidence for a true senolytic effect of N or Nutlin3a that meets this modified set of Koch’s postulates.
Clinical trials
Since clearing senescent cells with senolytic drugs is a completely new therapeutic paradigm, a novel strategy for translating senolytics into interventions for humans is needed (Fig. 3 ). In particular, ethical risk/benefit concerns have to be addressed: situations in which senolytic drugs enter the first clinical trials should ideally be serious and be those for which currently available treatments are ineffective (Fig. 3 ), such as IPF, complications of advanced diabetes, diastolic heart failure, advanced osteoporosis, dementias or COVID‐19 infections in hospitalized elderly patients. To accelerate progress, our approach has been to start multiple small, parallel trials for such conditions instead of conducting trials one after the other in series. If these trials in serious disorders show acceptable side‐effect profiles, if the agents are well tolerated, if target engagement (i.e. killing senescent cells) can be demonstrated, and if the candidate senolytic drugs are effective in treating serious diseases, then use of senolytics could go on to be tested for less serious conditions related to cellular senescence. Depending on results of such trials, the step after that would be to test effectiveness of the candidate senolytic drugs in delaying or preventing clinically manifest disease in subjects in whom presence of senescent cells can be demonstrated by blood, imaging, saliva or urine tests or biopsies. This could be feasible, since we discovered that there is a lag between accumulation of senescent cells and appearance of clinically apparent disease: after transplanting senescent cells into middle‐aged mice, it took time for frailty to develop [51]. As this progression from: 1) administration of senolytics in clinical trials for serious diseases, 2) to less serious conditions, 3) to prevention in at‐risk individuals proceeds, clinical trials will need to become progressively larger, longer and more expensive.
Figure 3.
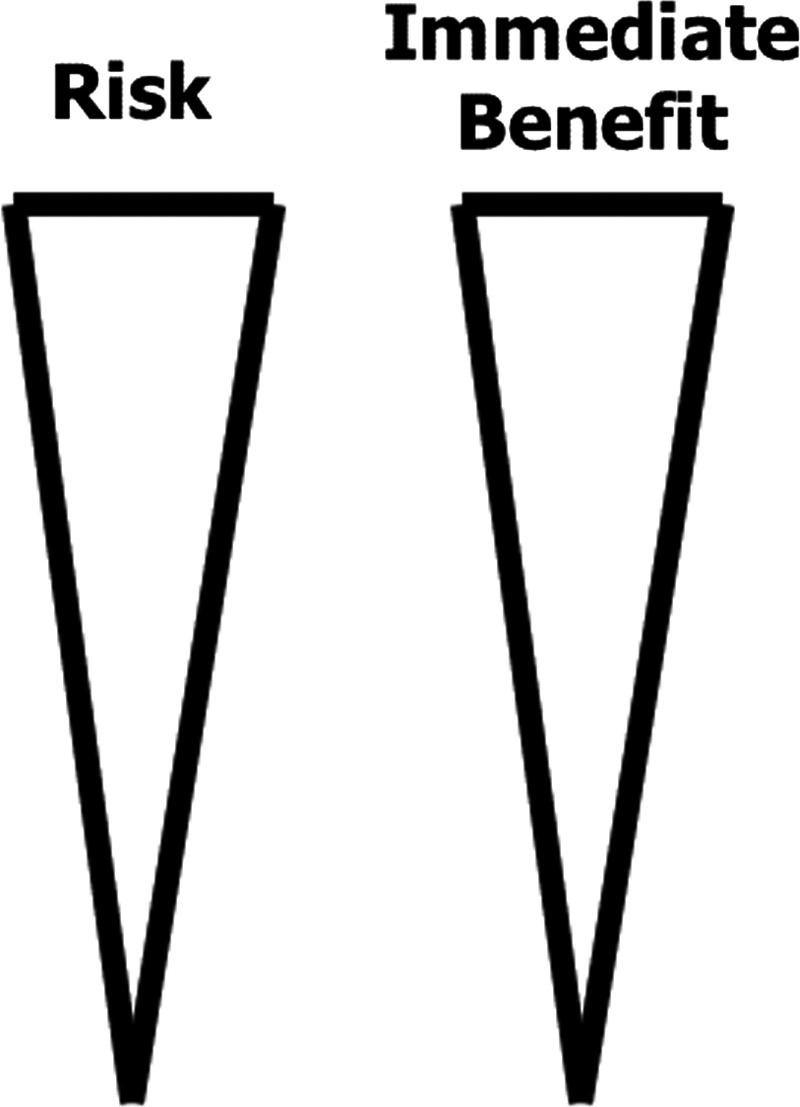
Early clinical studies of senolytics: Balancing benefit and risk. Since senolytic drugs are at the forefront of a completely new potential therapeutic paradigm – targeting fundamental ageing processes to delay, prevent or alleviate age‐related phenotypes, multiple diseases, geriatric syndromes and reductions in physical resilience – a careful risk‐benefit balance must be struck within first‐in‐human senolytic drug clinical trials, since potential short‐ and long‐term side‐effects from clearing senescent cells are not yet fully known.
Strategies for translating agents targeting fundamental ageing processes into treatments
Ideally, the first clinical trials should be with agents that have, insofar as possible, an established clinical safety profile, with strong preclinical data available about the drugs and indications to be tested. Intermittent administration, as opposed to continuous dosing, could reduce off‐target side‐effects. The clinical trials should be carefully monitored for safety, tolerability, target engagement (removal of senescent cells) and effectiveness. To speed translation, ideally several of these trials would be conducted in parallel across several diseases, rather than conducting one trial after the other in series. If safe and effective in initial trials for serious diseases, moving a senolytic drug towards less serious conditions and, eventually, to prevention in humans with an increased senescent cell burden and who are at risk for subsequently developing cell senescence‐associated diseases, could be a translational strategy. It is currently too early for practicing physicians to prescribe senolytic agents outside the context of carefully monitored clinical trials.
A consideration is whether systemic or local administration of candidate senolytic drugs is the best approach for initial clinical trials. For agents with an unknown or poor safety profile, local administration for senescence‐related conditions could be an option, but the following points run counter to this: 1) Senescence spreads from cell to cell [51], meaning that apparently local accumulations of senescent cells could be the ‘tip of the iceberg’, with an increased systemic senescent cell burden being a possible consequence. This could be the case even if effects of this systemic senescent cell burden are not yet apparent – as discussed above, there is a lag between senescent cell accrual and appearance of clinically apparent symptoms. 2) Also consistent with the concern that local injection of senolytics would be less beneficial than systemic administration is the Geroscience Hypothesis: multiple age‐ and senescence‐related diseases cluster within individuals, reducing the possible benefits from local injection. 3) Locally injected drugs generally do not stay local, particularly if the injection is into, for example, an osteoarthritic knee joint in which the synovial cavity is damaged. In the absence of an intact synovial membrane, local injection into the damaged knee joint could be effectively systemic, more like an intramuscular injection than a truly intra‐articular injection into a patient with an intact knee joint. 4) The previous points notwithstanding, local skin application, particularly of poorly absorbed senolytics, might be a viable strategy for treating senescence‐related skin disorders, such a melanotic naevi or psoriasis. Therefore, in general, systemic senolytic administration in initial clinical trials appears to be a reasonable approach, especially in the cases of candidates with an established safety profile, such as D, which has been approved for clinical prescription in the United States since 2006, or the natural product, Q. This would be less true for N or A1331852, which are not yet approved for general use as prescription drugs in the United States.
In a preliminary report from an early, open‐label clinical trial of 3 days of D + Q administration orally in patients with diabetes complicated by renal dysfunction, evidence for target engagement was found. Compared to biopsies before D + Q administration, 11 days after the last dose of D + Q (which has an 11 hour elimination half‐life), p16INK4a+ and SA‐βgal+ senescent cells had decreased in the diabetic subjects’ adipose tissue [116, 117]. Furthermore, D + Q decreased adipose tissue inflammation as evident by reduced macrophages and immune infiltration‐related crown‐like structures. A composite score of blood SASP factors was also decreased by D + Q in these patients. Thus, as in mice, D + Q can successfully decease senescent cell burden in humans.
The first‐in‐human trial of senolytics was of D + Q for patients with IPF [70] (N = 14 patients with stable IPF) in an open‐label study of intermittent D + Q (3 days/week over 3 weeks). Physical function was evaluated by testing 6‐minute walk distance, 4‐metre gait speed and chair‐stands time. These assessments were significantly and clinically meaningfully improved by D + Q (P < 0.05). Furthermore, correlations were found between change in function and change in SASP‐related pro‐inflammatory cytokines, matrix‐remodelling proteases and micro‐RNAs (23/48 markers; r ≥ 0.50). This pilot study supports the feasibility of further clinical trials using senolytics. It provided initial evidence that senolytics can alleviate physical dysfunction in IPF, supporting evaluation of D + Q in larger randomized, controlled trials for senescence‐associated diseases.
Several clinical trials of senolytics are currently underway or planned (Table 3 ). These include Phase I or II trials of D + Q for IPF, age‐related osteoporosis, age‐related frailty, frailty in bone marrow transplant survivors with an accelerated ageing‐like syndrome, frailty in childhood cancer survivors in their late 30’s and 40’s also with an accelerated ageing‐like syndrome, the ongoing trial in diabetic chronic kidney disease, Alzheimer’s disease (Tau+, symptomatic [clinical dementia rating 1]) and trials for improving outcomes after transplanting organs from old donors. Phase I or II trials of F are underway or planned for age‐related frailty, age‐related osteoporosis, osteoarthritis and to delay or prevent cytokine storm and acute respiratory distress syndrome in elderly patients infected by COVID‐19. Trials of injected Nutlin3a for osteoarthritis and of injected N for retinal degeneration are underway or planned. Preclinical studies are already underway and clinical trials are being planned in Sweden, including preclinical studies of senolytics for Critical Illness Myopathy at the Karolinska and a clinical trial for heritable type II diabetes at the University of Gothenburg.
Table 3.
RCT in process, completed and planned studies with senolytics
| Indication | Study ID (clinicaltrials.gov) | Senolytic regimen | Status |
|---|---|---|---|
| Idiopathic pulmonary fibrosis | NCT028749819 | Dasatinib + Quercetin |
Results of pilot study were published [132] A double‐blind, placebo‐controlled study is recruiting |
| Haematopoietic Stem Cell Transplant Survivors Study (HTSS) | NCT02652052 | Dasatinib + Quercetin | Recruiting |
| Alzheimer’s | NCT0463124 | Dasatinib + Quercetin | Recruiting |
| Diabetic chronic kidney disease | NCT02848131 | Dasatinib + Quercetin | Preliminary report published [117]; Recruiting |
| Childhood cancer survivors | Dasatinib + Quercetin | Funded; About to start | |
| First degree relatives of type II diabetics | Dasatinib + Quercetin | Planned | |
| Age‐related osteoporosis | Dasatinib + Quercetin; Fisetin | Funded; About to start | |
| Osteoarthritis |
UBX0101 |
Phase 1 completed, Phase 2 in process | |
| Osteoarthritis | NCT04210986 | Fisetin | Recruiting |
| Frailty | NCT03675724 | Fisetin | Recruiting |
| Complications due to COVID‐19 | Fisetin | Funded; FDA‐approved; About to start | |
| Macular degeneration |
UBX1967 UBX1325 |
Planned |
Multicentre alliances have been established to conduct clinical trials of senolytics and other interventions that target fundamental ageing processes, including the NIH‐funded Translational Geroscience Network (Mayo Clinic, Harvard, Johns Hopkins, and Wake Forest Universities, the Universities of Michigan, Texas [San Antonio], Minnesota, and Connecticut, and partners at other institutions) and the Alliance for Healthy Aging (Mayo Clinic, the Universities of Minnesota, Newcastle, and Copenhagen, and Groningen University).
Conclusions
It should be understood that the target of senolytics is senescent cells, not a single molecule or pathway, since targeting networks might yield more truly senolytic, less panolytic, drugs. Additionally, consistent with true senolytic activity, intermittent treatment with D + Q or F appears to be as effective as continuous administration, despite their short elimination half‐lives. D + Q already satisfies a modified set of Koch’s postulates for certain disorders and mounting evidence indicates that these postulates could also be satisfied for a number of other conditions in the cases of D + Q and F. True senolytic effects remain to be established for N or Nutlin3a. Consistent with the Unitary Theory, senolytics appear to restore progenitor dysfunction, attenuate tissue inflammation and alleviate age‐ and disease‐related metabolic dysfunction across cell types and tissues. Senolytics appear to delay, prevent or alleviate multiple age‐related conditions and chronic diseases and enhance healthspan and lifespan in experimental animals. Therefore, these agents could lead to interventions for humans that delay, prevent or treat senescence‐ and age‐related conditions – if clinical trials continue to demonstrate effectiveness and low toxicity. However, unless and until such clinical trials are completed and demonstrate safety, tolerability, target engagement and effectiveness, candidate senolytics should not be prescribed or used by general patient populations. They should only be administered in the course of carefully monitored clinical trials.
Conflicts of interest statement
The authors have a financial interest related to this research. Patents on senolytic drugs are held by Mayo Clinic. This research has been reviewed by the Mayo Clinic Conflict of Interest Review Board and was conducted in compliance with Mayo Clinic Conflict of Interest policies.
Author contribution
James Leslie Kirkland: Conceptualization (equal); Writing‐original draft (lead); Writing‐review & editing (equal). Tamar Tchkonia: Conceptualization (equal); Writing‐original draft (supporting); Writing‐review & editing (equal).
Acknowledgements
Dr. Yi Zhu worked with the authors in 2013 to discover the first senolytic drugs as reported in March, 2015 [87]. The authors acknowledge support from the Journal of Internal Medicine, US NIH grants R37 AG013925, P01 AG062413 and R33 AG061456 (Translational Geroscience Network), the Connor Fund, Robert J. and Theresa W. Ryan and the Noaber Foundation.
Kirkland JL, Tchkonia T (Mayo Clinic Robert and Arlene Kogod Center on Aging, Rochester, MN, USA). Senolytic drugs: from discovery to translation (Review). J Intern Med 2020;288:518–536. 10.1111/joim.13141
Content List ‐ Read more articles from the symposium: “Cardiovascular program and cardiovascular retreat”.
References
- 1. Kirkland JL. Translating the science of aging into therapeuticiInterventions. Cold Spring Harbor Perspect Med 2016; 6: a025908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Kirkland JL. Translating advances from the basic biology of aging into clinical application. Exp Gerontol 2013; 48: 1–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Kirkland JL, Stout MB, Sierra F. Resilience in aging mice. J Gerontol A Biol Sci Med Sci 2016; 71: 1407–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Olshansky SJ. Life expectancy and education: the author replies. Health Aff 2013; 32: 822. [DOI] [PubMed] [Google Scholar]
- 5. St Sauver JL, Boyd CM, Grossardt BR et al. Risk of developing multimorbidity across all ages in an historical cohort study: differences by sex and ethnicity. BMJ open 2015; 5: e006413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Kirkland JL, Tchkonia T. Cellular senescence: a translational perspective. EBioMedicine 2017; 21: 21–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Kirkland JL, Tchkonia T, Zhu Y, Niedernhofer LJ, Robbins PD. The Clinical potential of senolytic drugs. J Am Geriat Soc 2017; 65: 2297–301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Tchkonia T, Kirkland JL. Aging, cell senescence, and chronic disease: emerging therapeutic strategies. J Am Med Assoc 2018; 320: 1319–20. [DOI] [PubMed] [Google Scholar]
- 9. Kennedy BK, Berger SL, Brunet A et al. Geroscience: linking aging to chronic disease. Cell 2014; 159: 709–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Prata L, Ovsyannikova IG, Tchkonia T, Kirkland JL. Senescent cell clearance by the immune system: Emerging therapeutic opportunities. Semin Immunol 2018; 40: 101275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Chini C, Hogan KA, Warner GM et al. The NADase CD38 is induced by factors secreted from senescent cells providing a potential link between senescence and age‐related cellular NAD(+) decline. Biochem Biophys Res Comm 2019; 513: 486–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Xu M, Palmer AK, Ding H et al. Targeting senescent cells enhances adipogenesis and metabolic function in old age. eLife 2015; 4: e12997 10.7554/eLife.12997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Lewis‐McDougall FC, Ruchaya PJ, Domenjo‐Vila E et al. Aged‐senescent cells contribute to impaired heart regeneration. Aging Cell 2019; 18: e12931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Farr JN, Xu M, Weivoda MM et al. Targeting cellular senescence prevents age‐related bone loss in mice. Nat Med 2017; 23: 1072–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Kirkland JL, Tchkonia T, Pirtskhalava T, Han J, Karagiannides I. Adipogenesis and aging: Does aging make fat go MAD? Exp Gerontol 2002; 37: 757–67. [DOI] [PubMed] [Google Scholar]
- 16. Chapman J, Fielder E, Passos JF. Mitochondrial dysfunction and cell senescence: deciphering a complex relationship. FEBS Lett 2019; 593: 1566–79. [DOI] [PubMed] [Google Scholar]
- 17. Passos JF, Miwa S, von Zglinicki T. Measuring reactive oxygen species in senescent cells. Meth Mol Biol 2013; 965: 253–63. [DOI] [PubMed] [Google Scholar]
- 18. Anderson R, Lagnado A, Maggiorani D et al. Length‐independent telomere damage drives post‐mitotic cardiomyocyte senescence. EMBO J 2019; 38: e100492 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Kirkland JL. Inflammation and cellular senescence: potential contribution to chronic diseases and disabilities with aging. Public Policy Aging Rep 2013; 23: 12–5. [Google Scholar]
- 20. Nacarelli T, Lau L, Fukumoto T et al. NAD(+) metabolism governs the proinflammatory senescence‐associated secretome. Nat Cell Biol 2019; 21: 397–407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Schafer MJ, White TA, Iijima K et al. Cellular senescence mediates fibrotic pulmonary disease. Nat Comm 2017; 8: 14532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Xu M, Tchkonia T, Ding H et al. JAK inhibition alleviates the cellular senescence‐associated secretory phenotype and frailty in old age. Proc Natl Acad Sci (USA) 2015; 112: E6301–E10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Musi N, Valentine JM, Sickora KR et al. Tau protein aggregation is associated with cellular senescence in the brain. Aging Cell 2018; 17: e12840 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Ogrodnik M, Zhu Y, Langhi LGP et al. Obesity‐induced cellular senescence drives anxiety and impairs neurogenesis. Cell Metab 2019; 29: 1061–77. 10.1016/j.cmet.2018.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Hayflick L, Moorehead P. The serial cultivation of human diploid strains. Exp Cell Res 1961; 25: 585–621. [DOI] [PubMed] [Google Scholar]
- 26. Tchkonia T, Morbeck DE, von Zglinicki T et al. Fat tissue, aging, and cellular senescence. Aging Cell 2010; 9: 667–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Kirkland JL. The biochemistry of mammalian senescence. Clin Biochem 1992; 25: 61–75. [DOI] [PubMed] [Google Scholar]
- 28. Coppé JP, Patil C, Rodier F et al. Senescence‐associated secretory phenotypes reveal cell‐nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol 2008; 6: 2853–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Kuilman T, Peeper DS. Senescence‐messaging secretome: SMS‐ing cellular stress. Nat Rev Cancer 2009; 9: 81–94. [DOI] [PubMed] [Google Scholar]
- 30. Tchkonia T, Cartwright M, Wise B et al. Increased TNFa and CCAAT/enhancer binding protein homologous protein (CHOP) with aging predispose preadipocytes to resist adipogenesis. Am J Physiol 2007; 293: E1810–E9. [DOI] [PubMed] [Google Scholar]
- 31. Jiang C, Liu G, Luckhardt T et al. Serpine 1 induces alveolar type II cell senescence through activating p53–p21‐Rb pathway in fibrotic lung disease. Aging Cell 2017; 16: 1114–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Yamamoto K, Takeshita K, Saito H. Plasminogen activator inhibitor‐1 in aging. Semin Thrombosis Hemostasis 2014; 40: 652–9. [DOI] [PubMed] [Google Scholar]
- 33. Yang L, Fang J, Chen J. Tumor cell senescence response produces aggressive variants. Cell Death Discovery 2017; 3: 17049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Trayssac M, Hannun YA, Obeid LM. Role of sphingolipids in senescence: implication in aging and age‐related diseases. J Clin Invest 2018; 128: 2702–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Victoria B, Nunez Lopez YO, Masternak MM. MicroRNAs and the metabolic hallmarks of aging. Mol Cell Endocrinol 2017; 455: 131–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Xu D, Tahara H. The role of exosomes and microRNAs in senescence and aging. Advan Drug Delivery Rev 2013; 65: 368–75. [DOI] [PubMed] [Google Scholar]
- 37. Laberge RM, Zhou L, Sarantos MR et al. Glucocorticoids suppress selected components of the senescence‐associated secretory phenotype. Aging Cell 2012; 11: 569–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Christy B, Demaria M, Campisi J et al. p53 and rapamycin are additive. Oncotarget 2015; 6: 15802–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Moiseeva O, Deschenes‐Simard X, St‐Germain E et al. Metformin inhibits the senescence‐associated secretory phenotype by interfering with IKK/NF‐kappaB activation. Aging Cell 2013; 12: 489–98. [DOI] [PubMed] [Google Scholar]
- 40. Kandhaya‐Pillai R, Miro‐Mur F, Alijotas‐Reig J, Tchkonia T, Kirkland JL, Schwartz S. TNFalpha‐senescence initiates a STAT‐dependent positive feedback loop, leading to a sustained interferon signature, DNA damage, and cytokine secretion. Aging 2017; 9: 2411–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. De Cecco M, Ito T, Petrashen AP et al. Author Correction: L1 drives IFN in senescent cells and promotes age‐associated inflammation. Nature 2019; 572: E5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. De Cecco M, Ito T, Petrashen AP et al. L1 drives IFN in senescent cells and promotes age‐associated inflammation. Nature 2019; 566: 73–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Saito Y, Chikenji TS, Matsumura T, Nakano M, Fujimiya M. Exercise enhances skeletal muscle regeneration by promoting senescence in fibro‐adipogenic progenitors. Nat Commun 2020; 11: 889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Demaria M, Ohtani N, Youssef SA et al. An essential role for senescent cells in optimal wound healing through secretion of PDGF‐AA. Dev Cell 2014; 31: 722–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Meuter A, Rogmann LM, Winterhoff BJ, Tchkonia T, Kirkland JL, Morbeck DE. Markers of cellular senescence are elevated in murine blastocysts cultured in vitro: molecular consequences of culture in atmospheric oxygen. J Ass Reprod Gene 2014; 31: 1259–67. 10.1007/s10815-014-0299-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Campisi J. Aging, cellular senescence, and cancer. Ann Rev Physiol 2013; 75: 685–705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Menon R, Behnia F, Polettini J, Saade GR, Campisi J, Velarde M. Placental membrane aging and HMGB1 signaling associated with human parturition. Aging 2016; 8: 216–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Takeuchi S, Takahashi A, Motoi N et al. Intrinsic cooperation between p16INK4a and p21Waf1/Cip1 in the onset of cellular senescence and tumor suppression in vivo. Cancer Res 2010; 70: 9381–90. [DOI] [PubMed] [Google Scholar]
- 49. Larsen CJ. pRB, p53, p16INK4a, senescence and malignant transformation. Bull Cancer 2004; 91: 399–402. [PubMed] [Google Scholar]
- 50. Kirkland JL, Tchkonia T. Clinical strategies and animal models for developing senolytic agents. Exp Gerontol 2014; 68: 19–25. 10.1016/j.exger.2014.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Xu M, Pirtskhalava T, Farr JN et al. Senolytics improve physical function and increase lifespan in old age. Nat Med 2018; 24: 1246–56. 10.1038/s41591-018-0092-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Yousefzadeh MJ, Zhu Y, McGowan SJ et al. Fisetin is a senotherapeutic that extends health and lifespan. EBioMedicine 2018; 36: 18–28. 10.1016/j.ebiom.2018.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Santos F, Moreira C, Nobrega‐Pereira S, Bernardes de Jesus B. New insights into the role of epithelial–mesenchymal transition during aging. Int J Mol Sci 2019; 20: 891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Sedelnikova OA, Bonner WM. GammaH2AX in cancer cells: a potential biomarker for cancer diagnostics, prediction and recurrence. Cell Cycle 2006; 5: 2909–13. [DOI] [PubMed] [Google Scholar]
- 55. Hewitt G, Jurk D, Marques FD et al. Telomeres are favoured targets of a persistent DNA damage response in ageing and stress‐induced senescence. Nat Commun 2012; 3: 708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Giunta S, Funabiki H. Integrity of the human centromere DNA repeats is protected by CENP‐A, CENP‐C, and CENP‐T. Proc Natl Acad Sci USA 2017; 114: 1928–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Dimri GP, Lee X, Basile G et al. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc Natl Acad Sci USA 1995; 92: 9363–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Hall BM, Balan V, Gleiberman AS et al. p16(Ink4a) and senescence‐associated beta‐galactosidase can be induced in macrophages as part of a reversible response to physiological stimuli. Aging 2017; 98: 1867–84. 10.18632/aging.101268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Frescas D, Hall B, Strom E et al. Murine mesenchymal cells that express elevated levels of the CDK inhibitor p16(Ink4a) in vivo are not necessarily senescent. Cell Cycle 2017; 16: 1526–33. 10.1080/15384101.2017.1339850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Hsu CH, Altschuler SJ, Wu LF. Patterns of early p21 dynamics determine proliferation‐senescence cell fate after, Chemotherapy. Cell 2019; 178: 361–373.e12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Davalos AR, Kawahara M, Malhotra GK et al. p53‐dependent release of Alarmin HMGB1 is a central mediator of senescent phenotypes. J Cell Biol 2013; 201: 613–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Wiley CD, Campisi J. From ancient pathways to aging cells‐connecting metabolism and cellular senescence. Cell Metab 2016; 23: 1013–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Birch J, Barnes PJ, Passos JF. Mitochondria, telomeres and cell senescence: Implications for lung ageing and disease. Pharma Therap 2018; 183: 34–49. [DOI] [PubMed] [Google Scholar]
- 64. Schneider EL. Aging and cultured human skin fibroblasts. J Invest Dermatol 1979; 73: 15–8. [DOI] [PubMed] [Google Scholar]
- 65. Schneider EL. Cell replication and aging: in vitro and in vivo studies. Fed Proc 1979; 38: 1857–61. [PubMed] [Google Scholar]
- 66. Schneider EL, Mitsui Y. The relationship between in vitro cellular aging and in vivo human age. Proc Natl Acad Sci USA 1976; 73: 3584–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Schneider EL, Sternberg H, Tice RR et al. Cellular replication and aging. Mechan Ageing Dev 1979; 9: 313–24. [DOI] [PubMed] [Google Scholar]
- 68. Ressler S, Bartkova J, Niederegger H et al. p16INK4A is a robust in vivo biomarker of cellular aging in human skin. Aging Cell 2006; 5: 379–89. [DOI] [PubMed] [Google Scholar]
- 69. Kirkland JL, Hollenberg CH, Gillon WS. Age, anatomic site, and the replication and differentiation of adipocyte precursors. Am J Physiol 1990; 258: C206–C10. [DOI] [PubMed] [Google Scholar]
- 70. Justice JN, Nambiar AM, Tchkonia T et al. Senolytics in idiopathic pulmonary fibrosis: Results from a first‐in‐human, open‐label, pilot study. EBioMed 2019; 40: 554–63. 10.1016/j.ebiom.2018.12.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Farr JN, Rowsey JL, Eckhardt BA et al. Independent roles of estrogen deficiency and cellular senescence in the pathogenesis of osteoporosis: evidence in young adult mice and older humans. J Bone Mineral Res 2019; 34: 1407–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Suvakov S, Cubro H, White WM et al. Targeting senescence improves angiogenic potential of adipose‐derived mesenchymal stem cells in patients with preeclampsia. Biol Sex Diff 2019; 10: 49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Bian X, Griffin TP, Zhu X et al. Senescence marker activin A is increased in human diabetic kidney disease: association with kidney function and potential implications for therapy. BMJ Open Diabet Res Care 2019; 7: e000720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Sun S, Cai B, Li Y et al. HMGB1 and Caveolin‐1 related to RPE cell senescence in age‐related macular degeneration. Aging 2019; 11: 4323–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Chinta SJ, Woods G, Demaria M et al. Cellular senescence is induced by the environmental neurotoxin paraquat and contributes to neuropathology linked to parkinson's disease. Cell Rep 2018; 22: 930–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Liu RM, Liu G. Cell senescence and fibrotic lung diseases. Exp Gerontol 2020; 132: 110836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Moncsek A, Al‐Suraih MS, Trussoni CE et al. Targeting senescent cholangiocytes and activated fibroblasts with Bcl‐xL inhibitors ameliorates fibrosis in Mdr2‐/‐ mice. Hepatology 2017;67 :247–59. 10.1002/hep.29464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Blasiak J, Piechota M, Pawlowska E, Szatkowska M, Sikora E, Kaarniranta K. Cellular senescence in age‐related macular degeneration: can autophagy and dna damage response play a role? Oxidat Med Cell Long 2017; 2017: 5293258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Zhou B, Wan Y, Chen R et al. The emerging role of cellular senescence in renal diseases. J Cell Mol Med 2020; 24: 2087–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Covre LP, Martins RF, Devine OP et al. Circulating senescent T cells are linked to systemic inflammation and lesion size during human cutaneous leishmaniasis. Front Immunol 2018; 9: 3001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Xu M, Bradley EW, Weivoda MM et al. Transplanted senescent cells induce an osteoarthritis‐like condition in mice. J Gerontol A Biol Sci Med Sci 2017; 72: 780–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Kim SR, Jiang K, Ferguson CM et al. Transplanted senescent renal scattered tubular‐like cells induce injury in the mouse kidney. Am J Physiol Renal Physiol 2020; 318: F1167–F76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Krishnamurthy J, Torrice C, Ramsey MR et al. Ink4a/Arf expression is a biomarker of aging. J Clin Invest 2004; 114: 1299–307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Wang E. Senescent human fibroblasts resist programmed cell death, and failure to suppress bcl2 is involved. Cancer Res 1995; 55: 2284–92. [PubMed] [Google Scholar]
- 85. Kuilman T, Michaloglou C, Vredeveld LC et al. Oncogene‐induced senescence relayed by an interleukin‐dependent inflammatory network. Cell 2008; 133: 958–61. [DOI] [PubMed] [Google Scholar]
- 86. Zhu Y, Tchkonia T, Pirtskhalava T et al. The Achilles' heel of senescent cells: from transcriptome to senolytic drugs. Aging Cell 2015; 14: 644–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Yosef R, Pilpel N, Tokarsky‐Amiel R et al. Directed elimination of senescent cells by inhibition of BCL‐W and BCL‐XL. Nat Commun 2016; 7: 11190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Breccia M, Alimena G. Activity and safety of dasatinib as second‐line treatment or in newly diagnosed chronic phase chronic myeloid leukemia patients. BioDrugs 2011; 25: 147–57. [DOI] [PubMed] [Google Scholar]
- 89. Chan CM, Jing X, Pike LA et al. Targeted inhibition of Src kinase with dasatinib blocks thyroid cancer growth and metastasis. Clin Cancer Res 2012; 18: 3580–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Zhu Y, Doornebal EJ, Pirtskhalava T et al. New agents that target senescent cells: the flavone, fisetin, and the BCL‐XL inhibitors, A1331852 and A1155463. Aging 2017; 9: 995–63. 10.18632/aging.101202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Zhu Y, Tchkonia T, Fuhrmann‐Stroissnigg H et al. Identification of a novel senolytic agent, navitoclax, targeting the Bcl‐2 family of anti‐apoptotic factors. Aging Cell 2016; 15: 428–435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Chang J, Wang Y, Shao L et al. Clearance of senescent cells by ABT263 rejuvenates aged hematopoietic stem cells in mice. Nat Med 2016; 22: 78–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Vilgelm AE, Pawlikowski JS, Liu Y et al. Mdm2 and aurora kinase a inhibitors synergize to block melanoma growth by driving apoptosis and immune clearance of tumor cells. Cancer Res 2015; 75: 181–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Wilson WH, O'Connor OA, Czuczman MS et al. Navitoclax, a targeted high‐affinity inhibitor of BCL‐2, in lymphoid malignancies: a phase 1 dose‐escalation study of safety, pharmacokinetics, pharmacodynamics, and antitumour activity. Lancet Oncol 2010; 11: 1149–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Mahfoudhi E, Lordier L, Marty C et al. P53 activation inhibits all types of hematopoietic progenitors and all stages of megakaryopoiesis. Oncotarget 2016; 7: 31980–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Xu X, Liu Q, Zhang C et al. Inhibition of DYRK1A‐EGFR axis by p53‐MDM2 cascade mediates the induction of cellular senescence. Cell Death Dis 2019; 10: 282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Fuhrmann‐Stroissnigg H, Santiago FE, Grassi D, Ling Y, Niedernhofer LJ, Robbins PD. SA‐beta‐galactosidase‐based screening assay for the identification of senotherapeutic drugs. JoVE 2019;148 : 1–12. 10.3791/58133. [DOI] [PubMed] [Google Scholar]
- 98. Munoz‐Espin D, Rovira M, Galiana I et al. A versatile drug delivery system targeting senescent cells. EMBO Mol Med 2018; 10: e9355. 10.15252/emmm.201809355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Chen Z, Hu K, Feng L et al. Senescent cells re‐engineered to express soluble programmed death receptor‐1 for inhibiting programmed death receptor‐1/programmed death ligand‐1 as a vaccination approach against breast cancer. Cancer Sci 2018; 109: 1753–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Nakagami H. Cellular senescence and senescence‐associated T cells as a potential therapeutic target. Geriat Gerontol Int 2020; 20: 97–100. [DOI] [PubMed] [Google Scholar]
- 101. Li W, He Y, Zhang R, Zheng G, Zhou D. The curcumin analog EF24 is a novel senolytic agent. Aging 2019; 11: 771–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Fuhrmann‐Stroissnigg H, Ling YY, Zhao J et al. Identification of HSP90 inhibitors as a novel class of senolytics. Nat Commun 2017; 8: 422. 10.1038/s41467-017-00314-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Liu X, Wang Y, Zhang X et al. Senolytic activity of piperlongumine analogues: Synthesis and biological evaluation. Bioorganic Med Chem 2018; 26: 3925–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Baar MP, Brandt RM, Putavet DA et al. Targeted apoptosis of senescent cells restores tissue homeostasis in response to chemotoxicity and aging. Cell 2017; 169: 132–147.e16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Triana‐Martinez F, Picallos‐Rabina P, Da Silva‐Alvarez S et al. Identification and characterization of Cardiac Glycosides as senolytic compounds. Nat Commun 2019; 10: 4731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Guerrero A, Herranz N, Sun B et al. Cardiac glycosides are broad‐spectrum senolytics. Nature Metabol 2019; 1: 1074–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Weiland T, Lampe J, Essmann F et al. Enhanced killing of therapy‐induced senescent tumor cells by oncolytic measles vaccine viruses. Int J Cancer 2014; 134: 235–43. [DOI] [PubMed] [Google Scholar]
- 108. Palmer AK, Gustafson B, Kirkland JL, Smith U. Cellular senescence: at the nexus between ageing and diabetes. Diabetologia 2019; 62: 1835–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Palmer AK, Kirkland JL. Aging and adipose tissue: potential interventions for diabetes and regenerative medicine. Exp Gerontol 2016;86: 97–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110. Palmer AK, Tchkonia T, LeBrasseur NK, Chini EN, Xu M, Kirkland JL. Cellular senescence in type 2 diabetes: a therapeutic opportunity. Diabetes 2015; 64: 2289–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Palmer AK, Xu M, Zhu Y et al. Targeting senescent cells alleviates obesity‐induced metabolic dysfunction. Aging Cell 2019; 18: e12950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Minamino T, Orimo M, Shimizu I et al. A crucial role for adipose tissue p53 in the regulation of insulin resistance. Nat Med 2009; 15: 1082–7. [DOI] [PubMed] [Google Scholar]
- 113. Roos CM, Zhang B, Palmer AK et al. Chronic senolytic treatment alleviates established vasomotor dysfunction in aged or atherosclerotic mice. Aging Cell 2016. 10.1111/acel.12458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114. Nath KA, O'Brien DR, Croatt AJ et al. The murine dialysis fistula model exhibits a senescence phenotype: pathobiologic mechanisms and therapeutic potential. Am J Physiol Renal Physiol 2018; 315: F1493–99. 10.1152/ajprenal.00308.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Aguayo‐Mazzucato C, Andle J, Lee TB Jr et al. Acceleration of beta cell aging determines diabetes and senolysis improves disease outcomes. Cell Metabol 2019; 30: 129–42e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Hickson LJ, Langhi Prata LGP, Bobart SA et al. Senolytics decrease senescent cells in humans: Preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease. EBioMedicine. 2019; 47: 446–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Hickson LJ, Langhi Prata LGP, Bobart SA et al. Corrigendum to 'Senolytics decrease senescent cells in humans: Preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease' EBioMedicine 47 (2019) 446–456. EBioMedicine 2020; 52: 102595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. Larsson L, Degens H, Li M et al. Sarcopenia: aging‐related loss of muscle mass and function. Physiolog Rev 2019; 99: 427–511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Cupit‐Link MC, Kirkland JL, Ness KK et al. Biology of premature ageing in survivors of cancer. ESMO Open 2017; 2: e000250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Ness KK, Armstrong GT, Kundu M, Wilson CL, Tchkonia T, Kirkland JL. Frailty in childhood cancer survivors. Cancer 2014; 121: 1540 –47. 10.1002/cncr.29211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Pan J, Li D, Xu Y et al. Inhibition of Bcl‐2/xl with ABT‐263 selectively kills senescent type II pneumocytes and reverses persistent pulmonary fibrosis induced by ionizing radiation in mice. Int J Radiation Oncol Biol Phys 2017; 99: 353–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. van Willigenburg H, de Keizer PLJ, de Bruin RWF. Cellular senescence as a therapeutic target to improve renal transplantation outcome. Pharma Res 2018; 130: 322–30. [DOI] [PubMed] [Google Scholar]
- 123. Lau A, Kennedy BK, Kirkland JL, Tullius SG. Mixing old and young: enhancing rejuvenation and accelerating aging. J Clin Invest 2019; 129: 4–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Andre T, Meuleman N, Stamatopoulos B et al. Evidences of early senescence in multiple myeloma bone marrow mesenchymal stromal cells. PLoS One 2013; 8: e59756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Govoni S, Amadio M, Battaini F, Pascale A. Senescence of the brain: focus on cognitive kinases. Curr Pharm Design 2010; 16: 660–71. [DOI] [PubMed] [Google Scholar]
- 126. Zhang P, Kishimoto Y, Grammatikakis I et al. Senolytic therapy alleviates Abeta‐associated oligodendrocyte progenitor cell senescence and cognitive deficits in an Alzheimer's disease model. Nat Neurosci 2019; 22: 719 –28. 10.1038/s41593-019-0372-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Vazquez‐Villasenor I, Garwood CJ, Heath PR, Simpson JE, Ince PG, Wharton SB. Expression of p16 and p21 in the frontal association cortex of ALS/MND brains suggests neuronal cell cycle dysregulation and astrocyte senescence in early stages of the disease. Neuropathol App Neurobiol 2019; 46: 171–85. 10.1111/nan.12559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Kim SR, Jiang K, Ogrodnik M et al. Increased renal cellular senescence in murine high‐fat diet: effect of the senolytic drug quercetin. Trans Res 2019; 213: 112–23. 10.1016/j.trsl.2019.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Farr JN, Fraser DG, Wang H et al. Identification of senescent cells in the bone microenvironment. J Bone Min Res 2016; 31: 1920–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Xu M, Bradley EW, Weivoda MM et al. Transplanted senescent cells induce an osteoarthritis‐like condition in mice. J Gerontol A Biol Sci Med Sci 2016;72 6:780–5. 10.1093/gerona/glw154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Patil P, Dong Q, Wang D et al. Systemic clearance of p16(INK4a) ‐positive senescent cells mitigates age‐associated intervertebral disc degeneration. Aging Cell 2019; 18: e12927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132. Justice JN, Gregory H, Tchkonia T et al. Cellular senescence biomarker p16INK4a+ cell burden in thigh adipose is associated with poor physical function in older women. J Gerontol A 2018; 73: 939–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. Parikh P, Britt RD Jr, Manlove LJ et al. Hyperoxia‐induced cellular senescence in fetal airway smooth muscle cells. Am J Respirat Cell Mol Biol 2018; 61: 51–60. 10.1165/rcmb.2018-0176OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Barnes PJ, Baker J, Donnelly LE. Cellular senescence as a mechanism and target in chronic lung diseases. Am J Respirat Cell Mol Biol 2019; 200: 556–64. [DOI] [PubMed] [Google Scholar]
- 135. Nyunoya T, Monick MM, Klingelhutz A, Yarovinsky TO, Cagley JR, Hunninghake GW. Cigarette smoke induces cellular senescence. Am J Respirat Cell Mol Biol 2006; 35: 681–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Ogrodnik M, Miwa S, Tchkonia T et al. Cellular senescence drives age‐dependent hepatic steatosis. Nat Comm 2017; 8: 15691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137. Tabibian JH, O'Hara SP, Splinter PL, Trussoni CE, LaRusso NF. Cholangiocyte senescence by way of N‐ras activation is a characteristic of primary sclerosing cholangitis. Hepatology 2014; 59: 2263–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Blasiak J. Senescence in the pathogenesis of age‐related macular degeneration. Cell Mol Life Sci 2020; 77: 789–805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Liton PB, Challa P, Stinnett S, Luna C, Epstein DL, Gonzalez P. Cellular senescence in the glaucomatous outflow pathway. Exp Gerontol 2005; 40: 745–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Phatak NR, Stankowska DL, Krishnamoorthy RR. Bcl‐2, Bcl‐xL, and p‐AKT are involved in neuroprotective effects of transcription factor Brn3b in an ocular hypertension rat model of glaucoma. Mol Vis 2016; 22: 1048–61. [PMC free article] [PubMed] [Google Scholar]
- 141. Caprioli J. Glaucoma: a disease of early cellular senescence. Invest Opthalmol Vis Sci 2013; 54: ORSF60. [DOI] [PubMed] [Google Scholar]
- 142. Fu Q, Qin Z, Yu J et al. Effects of senescent lens epithelial cells on the severity of age‐related cortical cataract in humans: a case‐control study. Medicine 2016; 95: e3869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143. Castro P, Giri D, Lamb D, Ittmann M. Cellular senescence in the pathogenesis of benign prostatic hyperplasia. Prostate 2003; 55: 30–8. [DOI] [PubMed] [Google Scholar]
- 144. Choi J, Shendrik I, Peacocke M et al. Expression of senescence‐associated beta‐galactosidase in enlarged prostates from men with benign prostatic hyperplasia. Urology 2000; 56: 160–6. [DOI] [PubMed] [Google Scholar]
- 145. Vital P, Castro P, Tsang S, Ittmann M. The senescence‐associated secretory phenotype promotes benign prostatic hyperplasia. Am J Pathol 2014; 184: 721–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Sharma AK, Roberts RL, Benson RD et al. The senolytic drug navitoclax (ABT‐263) causes trabecular bone loss and impaired osteoprogenitor function in aged mice. Front Cell Dev Biol 2020;8: 354. 10.3389/fcell.2020.00354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Gollapudi P, Hasegawa LS, Eastmond DA. A comparative study of the aneugenic and polyploidy‐inducing effects of fisetin and two model Aurora kinase inhibitors. Mutat Res Genet Toxicol Environ Mutagen 2014; 767: 37–43. [DOI] [PMC free article] [PubMed] [Google Scholar]