

Abstract
The European Commission asked EFSA for a scientific opinion on the risks for animal and human health related to the presence of glycoalkaloids (GAs) in feed and food. This risk assessment covers edible parts of potato plants and other food plants containing GAs, in particular, tomato and aubergine. In humans, acute toxic effects of potato GAs (α‐solanine and α‐chaconine) include gastrointestinal symptoms such as nausea, vomiting and diarrhoea. For these effects, the CONTAM Panel identified a lowest‐observed‐adverse‐effect level of 1 mg total potato GAs/kg body weight (bw) per day as a reference point for the risk characterisation following acute exposure. In humans, no evidence of health problems associated with repeated or long‐term intake of GAs via potatoes has been identified. No reference point for chronic exposure could be identified from the experimental animal studies. Occurrence data were available only for α‐solanine and α‐chaconine, mostly for potatoes. The acute dietary exposure to potato GAs was estimated using a probabilistic approach and applying processing factors for food. Due to the limited data available, a margin of exposure (MOE) approach was applied. The MOEs for the younger age groups indicate a health concern for the food consumption surveys with the highest mean exposure, as well as for the P95 exposure in all surveys. For adult age groups, the MOEs indicate a health concern only for the food consumption surveys with the highest P95 exposures. For tomato and aubergine GAs, the risk to human health could not be characterised due to the lack of occurrence data and the limited toxicity data. For horses, farm and companion animals, no risk characterisation for potato GAs could be performed due to insufficient data on occurrence in feed and on potential adverse effects of GAs in these species.
Keywords: glycoalkaloids (GAs), solanine, chaconine, potato, margin of exposure (MOE), food, feed
Short abstract
This publication is linked to the following EFSA Supporting Publications article: http://onlinelibrary.wiley.com/doi/10.2903/sp.efsa.2020.EN-1905/full
Summary
The European Commission asked EFSA for a scientific opinion on the risks for animal and human health related to the presence of glycoalkaloids (GAs) in feed and food, in particular in potatoes and potato‐derived products. This risk assessment covers edible parts of potato plants and other food plants containing GAs, in particular tomato and aubergine. Non‐edible parts of GA containing plants have not been considered, with the exception of potato sprouts. The Panel developed the draft scientific opinion which underwent a public consultation from 27 February 2020 to 15 April 2020. The comments received and how they were taken into account when finalising the scientific opinion were published in an EFSA Technical Report (EFSA, 2020).
GAs are present in many plants of the family of Solanaceae, and contribute to plant resistance against pests and pathogens. GAs are composed of a steroidal aglycone and an oligosaccharide side chain. In commercial potato cultivars (S. tuberosum), the main GAs are α‐chaconine and α‐solanine, consisting of the aglycone solanidine and chacotriose and solatriose as oligosaccharide side chains, respectively. The aubergine fruit (S. melongena) contains primarily the GAs α‐solamargine and α‐solasonine, composed of the aglycone solasodine and chacotriose and solatriose, respectively. In tomato fruit (S. lycopersicum), α‐tomatine and α‐dehydrotomatine are the major GAs, with lycotetraose coupled to the aglycones tomatidine and tomatidenol, respectively.
Human risk assessment
In experimental animals, the potato GAs, α‐solanine and α‐chaconine, show a relatively low oral bioavailability, with differences between species. Hamsters exhibit higher absorption and slower excretion rates for both substances when compared to rats. Due to the limited information, the metabolic profiles of potato GAs in experimental animals could not be characterised.
In humans, α‐solanine and α‐chaconine are systemically absorbed following ingestion. For both substances, relatively long serum half‐lives were reported suggesting a possible accumulation. The blood clearance of the respective aglycone solanidine appears to be slow. Accordingly, levels of solanidine were regularly detected in the blood of human volunteers in several studies, suggesting hydrolysis of GAs. No further information is available on metabolism and excretion of potato GAs in humans.
There are no toxicokinetic data on tomato and aubergine GAs and their aglycones in experimental animals and humans.
In acute oral toxicity studies, no adverse effects of α‐solanine were observed at doses of 250 mg/kg body weight (bw) per day in rats and 1,000 mg/kg bw per day in mice. Reliable data on other potato GAs or tomato and aubergine GAs and their aglycones are missing.
In repeated oral dose studies on potato GAs, rodents showed non‐specific effects, such as reduced body weight and relative liver weight with indication of similar potencies of α‐solanine and α‐chaconine. Hamsters exhibited these symptoms after a 5‐day treatment with 100 mg of α‐solanine or α‐ chaconine/kg bw per day, while mice showed these effects after one week of daily treatments with 416.6 mg of α‐solanine or 409 mg of α‐chaconine/kg bw. Solanidine, however, increased the absolute and relative liver weight at 190.9 mg/kg bw per day in mice, suggesting a different effect of the aglycone compared to the GAs.
The tomato GA, α‐tomatine, and its aglycone, tomatidine, exerted no effects in rats, when applied at 20 mg/kg bw per day for a period of 200 day. At higher doses, α‐tomatine reduced the cholesterol uptake and increased fecal sterol and coprostanol excretion in hamsters and rats. In mice, a 1‐ to 2‐week treatment with the aubergine GA, α‐solasonine, increased the body weight gain at 424.4 mg/kg bw per day, while its aglycone, solasodine, decreased body weight gain and caused gastric gland degeneration and liver toxicity at 160 mg/kg bw per day.
Developmental studies have been performed mainly in hamsters treated with potato GAs and their aglycones for only one day or for a short, very restricted time period during gestation. Outcomes were mainly analysed in late gestational embryos and comprised effects in the central nervous system, predominantly exencephaly, encephalocele and anophthalmia. These malformations occurred at doses of 165 mg/kg bw per day and above for GAs, and of 115 mg/kg bw per day and above for the aglycones. No no‐observed‐adverse‐effect level (NOAEL) or lowest‐observed‐adverse‐effect level (LOAEL) could be identified from these studies. Reduced postnatal survival of pups due to insufficient milk production was reported when pregnant Holtzman rats had been exposed to 3.6 mg of α‐solanine/kg bw per day. Studies on the male fertility in dogs have been performed only with the aubergine aglycone, solasodine. Decreased epididymal weight and cauda epididymal epithelial height, and also an epididymal lumen depleted of sperm occurred in dogs after 80 mg/kg bw per day given for 1 month. Similar effects were observed in Rhesus monkeys exposed to 100 mg/kg bw per day for 5 months.
From the limited number of studies available, there was no evidence for genotoxicity of the potato GAs α‐solanine and α‐chaconine, and the aglycone, solanidine, as well as for the aubergine GA, α‐solamargine. However, there is not sufficient information to conclude on the genotoxic potential of these GAs.
No long‐term chronic toxicity/carcinogencity study for potato, tomato or aubergine GAs or for the respective aglycones could be identified.
In humans, acute toxic effects following ingestion of potato GAs include gastrointestinal symptoms of varying severity such as vomiting, diarrhoea and abdominal pain, which may occur from a total potato GAs (potato TGA) intake of 1 mg/kg bw or more. Further symptoms, including drowsiness, apathy, confusion, weakness, vision disturbances, rapid and weak pulse and low blood pressure, may be the consequence of dehydration following vomiting and diarrhoea.
In severe cases, paralysis, respiratory insufficiency, cardiac failure, coma and death have been reported. Doses in the range of 3–6 mg potato TGAs/kg bw are considered to be potentially lethal for humans. Results from limited volunteer studies suggest possible differences in the human population with respect to the individual susceptibility towards adverse effects associated with the intake of potato GAs.
Regarding the mode of action, adverse effects of GAs may be due to their ability to complex with membrane 3β‐hydroxy sterols, thereby causing disruption and loss of integrity of cell membranes. After oral exposure, these effects may affect the mucosa of the gastrointestinal tract and cause the symptoms observed in intoxicated humans, such as nausea, vomiting and diarrhoea.
GAs inhibit acetylcholinesterase (AChE) and serum butyrylcholinesterase (BuChE) by a reversible, competitive mode of action. The relative potency of inhibition of α‐solanine and α‐chaconine appears to be similar. The aglycones exert weak or no inhibitory effects. The excess of acetylcholine at the neuronal and neuromuscular junctions upon inhibition of the enzymes might also contribute to the symptoms described for intoxications with GAs.
At high doses, α‐tomatine may form a non‐absorbable complex with cholesterol and other sterols in the enteral lumen which may impair the absorption of cholesterol. As a consequence, blood cholesterol levels were lowered in rodents.
The CONTAM Panel considered that the use of rodent data on acute toxicity was not appropriate to establish a reference point for acute exposure to potato GAs in humans. The CONTAM Panel selected the LOAEL of 1 mg potato TGA/kg bw per day as the reference point for acute risk characterisation, based on human data from case reports, outbreaks and studies in volunteers. The available data on acute toxicity were considered insufficient to establish a health‐based guidance value. Instead, the Panel used the margin of exposure (MOE) approach to assess a possible health concern from acute exposure to potato TGAs via food.
Assuming the main symptoms to be mainly due to local irritation of the gastrointestinal mucosa rather than inhibition of AChE activity, the Panel considered that the possible inter‐individual variability in toxicodynamics is more relevant than the interindividual variability in toxicokinetics. Accordingly, an MOE higher than 10 indicates that there is no health concern. This MOE of 10 takes into account the extrapolation from a LOAEL to a NOAEL (a factor of 3) and the interindividual variability in toxicodynamics (a factor of 3.2).
The experimental data available for repeated dose toxicity are not sufficient to identify a reference point for chronic exposure to potato GAs. In humans, no evidence of health problems associated with repeated or long‐term intake of GAs via potatoes has been identified.
Regarding GAs or aglycones occurring in edible parts of food plants other than S. tuberosum, no suitable study for determining a reference point for tomato or aubergine GAs or aglycones was identified.
Occurrence data were only available for α‐solanine and α‐chaconine, and mostly for ‘Main‐crop potatoes’ and ‘New potatoes’. Few data were available for processed food. No data on the occurrence of tomato and aubergine GAs and their aglycones were submitted to EFSA.
Since the occurrence data on potato GAs did not cover all the food categories containing potatoes in the Consumption Database, it was decided that the best approach for the exposure assessment would be to use the occurrence data in the raw primary commodities (RPC) (main‐crop potatoes and new potatoes) and the RPC Consumption Database. The Panel decided to combine the occurrence of ‘New potatoes’ with that of ‘Main‐crop potatoes’ and the mean upper bound (UB) occurrence (sum of α‐solanine and α‐chaconine) for these two groups was 51.2 mg/kg and the P95 occurrence was 116.8 mg/kg. The minimum and maximum reported concentrations were 1.1 and 276.6 mg/kg, respectively.
The acute dietary exposure to potato TGAs was estimated using a probabilistic approach, including only days in which there was consumption of main‐crop potatoes. As no occurrence data were available for GAs in tomato and aubergine, these foods were not included in the exposure assessment.
Processing of potatoes has been reported to reduce the content of GAs in the final processed product. In general, and according to the literature, the peeling of potatoes reduced the GA content by 25–75%, boiling in water and blanching of peeled potatoes by 5–65% and frying in oil of peeled potatoes by 20–90%. Microwave and oven baking of unpeeled potatoes may cause a reduction in the GA content by 3–45% and by 20–50%, respectively. No information has been found about the chemical nature of the GA degradation products. For the exposure assessment, processing factors for the major food processing steps, comprising peeling and heat processing (boiling, frying, baking), were applied to the occurrence data as follows: processing factors between 0.25 and 0.75 were attributed to the peeling of potatoes, between 0.1 and 0.8 for frying and deep frying, and between 0.35 and 0.95 for all other cooking methods.
Information about the peeling of potatoes was not available in the consumption database, but it was assumed that 90% of the potatoes are consumed as peeled. Where information of the cooking method was not available, a cooking method was randomly attributed to the eating event based on the relative frequency of cooking methods reported.
The mean UB exposure to potato TGAs across surveys ranged from 23.3 μg/kg bw per day in adults to 174.0 μg/kg bw per day in toddlers. The 95th percentile exposure ranged from 78.3 μg/kg bw per day in adults to 535.1 μg/kg bw per day in toddlers (up to 822.9 μg/kg bw per day in the upper limit of the 95% confidence interval).
Comparing the LOAEL for potato TGAs of 1 mg/kg bw per day with the acute exposure estimates, the MOEs for the younger age groups indicate a health concern for the food consumption surveys with the highest mean exposure, as well as for the P95 exposure in all surveys. For adult age groups, the MOEs indicate a health concern only for the food consumption surveys with the highest P95 exposures.
The CONTAM Panel calculated the mean percentage of days with potato consumption across surveys per age group on which the potato TGA intake may be below the MOE of 10. The highest number of survey days with intake of potatoes below the MOE of 10 was estimated for toddlers (56%) followed by children (50%). For the other age groups, the estimated TGA intake was below the MOE of 10 in up to 22–40% of the survey days.
For tomato and aubergine GAs, the risk to human health could not be characterised due to the lack of occurrence data in food and the limited information on the adverse effects in experimental animals and humans.
The CONTAM Panel considered that the impact of the uncertainties on the risk assessment of acute exposure to potato GAs in food is moderate, and that overall, the identified uncertainties may either cause an over‐ or underestimation of the risk.
Farm animals, horses and companion animals risk assessment
Information on the toxicokinetics of GAs was limited to ruminants, for which the data suggest an extensive conversion of α‐solanine and α‐chaconine to aglycones in rumen, and a low potential of solanidine to transfer into cows’ milk.
No data on the potential adverse effects of potato GAs in horses, companion animals (cats and dogs) or fur animals were identified. Due to an insufficient database on the adverse effects of GAs in ruminants, pigs, poultry, rabbits and fish, an acute reference dose could not be derived.
Potatoes are not grown specifically as feed for livestock, but when supply exceeds market requirements for human consumption whole (raw) potatoes may be used as feed for ruminants and pigs. Some by‐products of potato processing and starch extraction are used as feeds for farmed livestock, principally non‐ruminants, and for companion animals.
Data on potato GAs in feed were insufficient to perform an exposure assessment.
Thus, no risk characterisation could be performed due to insufficient occurrence data of GAs for feed and the lack of, or limited, data on the adverse effects of GAs in farm animals, horses or companion animals.
Recommendations
The following needs have been identified to improve the risk assessment for humans and reduce the uncertainties:
Research on the occurrence of GAs and their aglycones and other potentially toxicologically relevant secondary plant metabolites in the potato cultivars available on the market and on new potato cultivars resulting from breeding experiments.
Occurrence data on GAs and their aglycones in potato processed products, including foods for infants.
Occurrence data on GAs and their aglycones in tomato and aubergine and products thereof.
Data on the toxicokinetics of potato, tomato and aubergine GAs and aglycones in experimental animals and humans.
Data on repeated dose toxicity, including reproductive and developmental toxicity of potato, tomato and aubergine GAs and aglycones in experimental animals.
Studies in humans linking dietary exposure, biomarkers of exposure and adverse effects.
The following needs have been identified to improve the risk assessment for farm animals, horses and companion animals and reduce the uncertainties:
Occurrence data on potato GAs and their aglycones in feed.
Studies on the kinetics and the potential adverse effects from feed material containing GAs of potato GAs in farm animals, horses and companion animals.
1. Introduction
1.1. Background and Terms of Reference as provided by the requestor
1.1.1.
Background
Many plants in the family Solanaceae contain glycoalkaloids, and they are considered to be natural toxins. The plant glycoalkaloids are toxic steroidal glycosides and the commonest types found in food plants are α‐solanine and α‐chaconine. Their natural function is probably to serve as stress metabolites or phytoalexins for the protection of the plant when attacked by insects, fungi, etc.
Amongst the most widely cultivated food crops, aubergines, tomatoes and potatoes are in the Solanaceae family, but the levels of glycoalkaloids in tomatoes and aubergines are generally quite low.
The glycoalkaloids of most relevance to food safety are those occurring in the potato. The predominant toxic steroidal glycosides in potato are α‐solanine and α‐chaconine. They occur in potato tubers, peel, sprouts, berries, leaves and blossoms and their concentration in tubers depends on a number of factors, such as cultivar, maturity and environmental factors. Concentrations of glycoalkaloids are 3–10 times greater in the peel than in the flesh. There is considerable variation in glycoalkaloid content among potato cultivars. Storage conditions, especially light and temperature, are mainly responsible for increases in solanine. Although the glycoalkaloid content can increase in the dark, the rate of formation is only about 20% the rate of formation in light. Increases of solanine in the potato peel are closely associated with greening (synthesis of chlorophyll) of the peel. These biochemical processes are independent of each other, but are both activated by light.
Bitter or burning sensation in the mouth are sensory impressions which may accompany glycoalkaloid poisoning symptoms from potatoes that include flu‐like symptoms such as nausea, vomiting, stomach and abdominal cramps, and diarrhoea. More severe cases of glycoalkaloid poisoning may be accompanied by a variety of neurological effects (i.e. drowsiness, apathy, restlessness, shaking, confusion, weakness, and disturbed vision). There are a few reports of deaths being attributed to glycoalkaloid exposure from the consumption of potatoes, potato leaves, and potato berries.
Potatoes and potato‐derived products are listed in the Catalogue of feed materials.1
Terms of Reference
In accordance with Art. 29 (1) of Regulation (EC) No 178/2002, the European Commission asks the European Food Safety Authority for a scientific opinion on the risks for animal and human health related to the presence of glycoalkaloids in feed and food, in particular in potatoes and potato‐derived products.
1.2. Interpretation of the Terms of Reference
The CONTAM Panel considered that the opinion should cover edible parts of potato plants and also of other food plants containing glycoalkaloids (GAs), e.g. tomato and aubergine. Non‐edible parts of GA containing plants have not been considered, with the exception of potato sprouts. In particular, the CONTAM Panel concluded this Opinion should comprise the:
-
a)
evaluation of the toxicity of GAs in feed and food, in particular in potatoes and potato‐derived products, for farm and companion animals and humans considering all relevant toxicological end points;
-
b)
evaluation of the alkaloid profile (i.e. composition of the alkaloids and their concentration) of the food and feed samples submitted to EFSA;
-
c)
estimation of the dietary exposure of the European population to GAs in food, in particular in potatoes and potato‐derived products, including the consumption patterns of specific groups of the population if appropriate;
-
d)
estimation of the dietary exposure of farm and companion animals to GAs in feed, in particular in potatoes and potato‐derived products;
-
e)
assessment of the human health risks for the European population, including specific groups of the population if appropriate, as the consequence of the estimated dietary exposure;
-
f)
assessment of the farm and companion animal health risks in Europe as the consequence of the estimated dietary exposure. Exposure to GAs from weeds containing GA is only addressed in this Opinion in the context of accidental intake by farm animals.
When referring to GAs in potatoes, the term total GAs (TGA) refers to a material comprising α‐solanine and α‐chaconine as major fraction, with no specification on the occurrence of minor GAs as well as β‐ and γ‐forms of solanine and chaconine. Similarly, when referring to tomato and aubergine, the term TGA refers to the GAs from the corresponding species and forms thereof.
1.3. Supporting information for the assessment
1.3.1. Chemistry
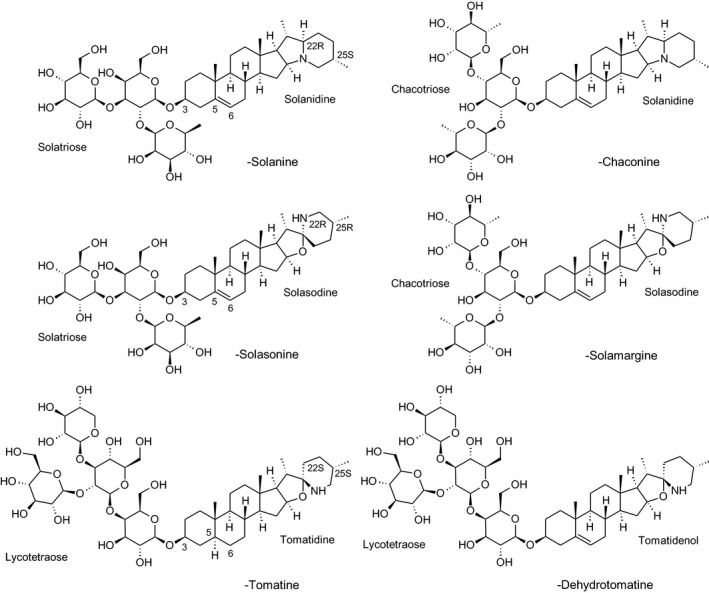
Solanine is one of the first alkaloids that has been isolated from nature, by Desfosses in 1820 (Friedman et al., 1997). In 1861, Zwenger and Kind reported that solanine contains a glycoside side chain (Zwenger and Kind, 1861). Only in 1954, it was shown that solanine extracted from potato is in fact a mixture of two glycoalkaloids (GAs), α‐solanine and α‐chaconine, that share the same solanidine aglycone (Kuhn and Löw, 1954). Since then, at least 90 different GAs have been isolated and fully structurally elucidated from over 300 species of the Solanaceae family (Sánchez‐Mata et al., 2010; Al Sinani and Eltayeb, 2017). The chemical structures and some physical properties of the most important ones are listed in Appendix A.
GAs are composed of a steroidal aglycone and an oligosaccharide side‐chain attached to the 3β‐hydroxy group of the aglycone (see Figure 1) (Friedman et al., 1997; Friedman, 2006; Milner et al., 2011). The GAs of relevance can be divided into the (i) solanidane group, with solanidine as the steroid backbone, and the (ii) spirosolane group, with either the solasodine or the tomatidenol/tomatidine backbone. GAs often contain a double bond between C5 and C6, but the corresponding 5a, 6‐hydrogenated forms are also common and, in some species (e.g. tomato), they constitute the major components. The stereochemistry at carbons C22 and C25 is well defined: in solanidine, the configuration is 22R, 25S; in solasodine, it is 22R, 25R; and in tomatidenol/tomatidine, the configuration is 22S, 25S (Friedman et al., 1997).
Figure 1.
Structure of the most important glycoalkaloids (GAs) found in potato (α‐solanine and α‐chaconine), aubergine (α‐solasonine and α‐solamargine) and tomato (α‐tomatine and α‐dehydrotomatine)
Further diversification is generated by the composition of the glycoside side‐chain. Most GAs contain either a trisaccharide (chacotriose or solatriose) or a tetrasaccharide (lycotetraose) as carbohydrate. In commercial potato cultivars (Solanum tuberosum), mostly α‐chaconine and α‐solanine (composed of the solanidine aglycone and chacotriose and solatriose, respectively) are present (Figure 1). Wild S. tuberosum varieties may contain a much wider range of GAs (Friedman et al., 1997; Distl and Wink, 2009). The aubergine fruit (derived from S. melongena) contains primarily α‐solamargine and α‐solasonine (composed of the solasodine aglycone and chacotriose and solatriose, respectively). In tomato fruit (derived from S. lycopersicum varieties), α‐tomatine and α‐dehydrotomatine are the major compounds (composed of the aglycones tomatidine and tomatidenol, respectively, coupled to lycotetraose) (Friedman, 2002).
The prefix alpha (α‐) refers to the intact glycoside while the prefixes beta (β‐), gamma (γ‐) and delta (δ‐) refer to the corresponding GAs with progressively truncated carbohydrate side‐chains due to the action of enzymatic or acidic hydrolysis (Friedman, 2006; Milner et al., 2011).
GAs as pure standards are stable to temperatures up to 150°C (Takagi et al., 1990) and to alkaline conditions. However, acidic hydrolysis using strong acid and elevated temperatures may cleave the glycosidic bonds and liberate the aglycone together with the corresponding reducing sugars (Friedman, 2006). The aglycones due to their steroid nature are not soluble in water, but the intact GAs are slightly soluble (e.g. solubility of α‐solanine in water: 25 mg/L). GAs as well as aglycons are readily soluble in organic solvents such as ethanol and methanol.
1.3.2. Analytical methods
Many different analytical techniques have been applied over the last century for the detection and quantification of GAs. With older analytical methods (gravimetric, colourimetric), it was often not possible to discriminate the individual GAs. These non‐specific methods are currently no longer used for quantitative purposes, but they are included because these methods were used in some of the older toxicological studies and case reports of human intoxications. Nowadays most confirmatory methods use liquid chromatography (LC) to separate the intact GAs and detection is performed either by ultraviolet‐visible (UV‐vis) detection or by mass spectrometry (MS). Assays based on thin layer chromatography (TLC) or immunoassays (i.e. enzyme‐linked immunosorbent assay (ELISA)) are also available and can be applied for screening.
For reliable quantification of GAs, reference standards of high purity are required, which are available for the most common ones (α‐solanine, α‐chaconine, α‐tomatine, α‐solasodine, α‐solamargine and the corresponding aglycones). For many of the less common GAs (Appendix A), no reference standards are currently available.
1.3.2.1. Extraction and clean‐up
When analysing the intact GAs, an important aspect is the stability of the compounds. Enzymatic degradation of the glycoside side‐chain due to the action of glycosidases present in fresh potato tubers may occur during extraction when not properly controlled (Friedman et al., 1997). This may result in the underestimation of the GA content, if only the intact GAs are monitored. Two approaches are generally followed to prevent from losses due to enzymatic activity. One approach is to cut or slice the potato tubers, add liquid nitrogen and to lyophilise by freeze‐drying. The freeze‐dried material is ground and homogenised to a fine powder (Friedman and Dao, 1992). Extraction of the lyophilised material is typically performed with an acidic aqueous solvent (e.g. acetic acid), mixed with a suitable organic solvent (e.g. methanol, acetonitrile). The other approach is to shred the potato tubers in a food processor, add liquid nitrogen and grind to a fine powder (Hellenäs, 1986; AOAC, 2000). Extraction of the frozen material is conducted with a mixture of 5% acidic acid to which 0.5% sodium bisulfite (NaHSO3) has been added. For both methods a further clean‐up is necessary before the extracts can be analysed by high‐performance LC (HPLC)‐based methods. Clean‐up is preferably conducted using solid‐phase extraction (SPE) cartridges (typically C18‐based, but cartridges with NH2, CN stationary phases have also been used) (Friedman et al., 1997).
1.3.2.2. Detection methods
Gravimetric methods
In gravimetric methods, the GAs are measured by alkaline precipitation from potato extracts. GAs are extracted from the tubers using a strong acid, such as sulfuric or hydrochloric acid, that results in hydrolysis of the GAs to the corresponding aglycone. Addition of a surplus of alkaline, e.g. calcium hydroxide, to the solution results in a precipitation of the alkaloids, that can be collected, dried and weighted (Bömer and Mattis, 1924). Gravimetric methods have a limit of quantification (LOQ) of approximately 10 mg total GAs (TGA)/kg. Gravimetric methods have been in general use until the development of colourimetric methods in the 1930s and 1940s. Gravimetric methods have a number of disadvantages, such as losses occurring during precipitation, lengthy procedures and the fact that other alkaloids may co‐precipitate (Lepper, 1949; Friedman et al., 1997).
Colourimetric methods
The first colourimetric method to quantify the content of solanine in potato tubers was developed by Alberti in 1932, using sulfuric acid and formaldehyde to generate a coloured derivative (Alberti, 1932). Many different methods using different reagents have been described in the literature in the following decades. Clement and Verbist (1980) compared nine different colourimetric methods for the analysis of solanidine in potato tubers. They noted that a considerable variation in the results was obtained, which was attributed to unspecific reactions with other plant components with the staining reagents used (resulting in overestimation of the content in some methods) or to losses during extraction (leading to an underestimation of the content in other methods). It was reported that sensitivity between methods differed up to 20‐fold, but limits of detection (LODs)/LOQs were not determined (Clement and Verbist, 1980).
Another important drawback of colourimetric methods is that they require hydrolysis of the glycoside side‐chain prior to derivatisation. As a result, only the TGA content can be determined and no information on the individual glycosides is obtained. Since the 1980s, colourimetric methods have been largely replaced by HPLC‐based methods.
TLC with UV‐Vis detection
TLC is often used as a quick, qualitative, screening tool to assess the presence of GAs in plant extracts. However, quantitative methods using high‐performance TLC (HPTLC)‐densitometry have also been described. Bodart et al. (2000) described a method for the determination of α‐solanine and α‐chaconine in potato extracts by means of HPTLC‐densitometry. The GAs extracted from potato were separated on the TLC plate and derivatised to generate chromophoric compounds that were measured at 507 nm with a densitometer. The LOQ for both GAs was 100 ng on plate, corresponding to 40 mg/kg in dehydrated peeled potato powder and to 10 mg/kg in fresh peeled potato tubers. A modified HPTLC method was described by Skarkova et al. (2008), with an LOQ of 5 mg/kg for α‐solanine and α‐chaconine for peeled, uncooked potatoes.
Enzyme‐linked immunosorbent assays (ELISA)
A few reports using immunoassays (ELISA) for the quantification of GAs have been published. Morgan et al. (1985) developed an ELISA assay to determine the TGA content in potato tubers and compared their assay with two existing colourimetric methods (Morgan et al., 1985). The sensitivity of the ELISA assay was excellent (0.02 mg/kg) and did not show cross reactivity with tomatidine derivatives. Comparison with a HPLC‐ultraviolet (UV) method showed a good linear relationship, when corrected for recovery losses (Hellenäs, 1986).
Stanker et al. (1994) developed an ELISA based on monoclonal antibodies that showed affinity for solanidine, tomatidine and solasodine derivatives. The commercialised version of the ELISA kit was found to work well for the quantification of TGAs in potato tubers, sprouts, potato fries and chips (Friedman et al., 1998). The ELISA displayed good cross reactivity for solanidine derivatives (α‐solanine, α‐, β‐, γ‐chaconine), lower sensitivity for α‐tomatine and very limited sensitivity for solasodine derivatives. For all investigated products, a good correlation for TGA content was observed with results obtained by HPLC‐UV analysis. The LOD for TGAs was estimated at 1 mg/kg.
Gas chromatography (GC) with FID, MS or NPD detection
Analysis of intact GAs by GC‐flame ionisation detector (FID) or GC‐MS is possible only after derivatisation of the glucoside hydroxyl groups (Herb et al., 1975). However, it is more common to analyse the aglycones after hydrolysis, because derivatisation in that case is not necessary (Van Gelder, 1985; Friedman et al., 1997). Bushway et al. (1984) have developed a GC‐nitrogen‐phosphorus detector (NPD) method for solanidine in milk, with an LOQ of 0.28 mg/L. Nowadays LC‐based methods have replaced GC‐based methods.
HPLC with UV‐VIS detection
Intact GAs as well as the aglycones can be measured by HPLC‐UV detector at wavelengths around 200 nm (Houben and Brunt, 1994; Friedman et al., 1997; AOAC Official Method 997.13, 2000). Separation of α‐solanine and α‐chaconine, as well as the separation of α‐tomanine and α‐dehydrotomatine, can be achieved on reversed phase C18 chromatographic columns using aqueous phosphate buffers in combination with an organic modifier, typically methanol or acetonitrile (Kozukue et al., 2004). It is possible to separate these GAs by a simple isocratic elution, but for the analysis of a wider range of GAs gradient elution is necessary (Eanes et al., 2008; Maurya et al., 2013). Due to the fact that GAs do not show significant UV absorption at wavelengths above 215 nm, the analysis needs to be conducted at low, unspecific, wavelengths. It is therefore necessary to include purification steps, e.g. by SPE clean‐up, prior to analysis by HPLC‐UV (Keukens et al., 1994; Friedman et al., 1997). The LOD is around 2.5 mg/kg for the individual compounds.
LC‐MS methods (LC‐MS/MS, LC‐HRMS)
LC‐MS‐based methods are becoming increasingly popular in the analysis of GAs. The most important advantage of MS detection above UV detection is the much higher selectivity of the MS. Combined with the high sensitivity of modern mass spectrometers, sample preparation and clean‐up can often be minimised and simple dilution of extracts may be sufficient. Methods based on LC coupled to tandem mass spectrometry (LC‐MS/MS) are often used for targeted analysis of GAs (Jandrić et al., 2011; Hossain et al., 2015; Nara et al., 2019), while methods using LC coupled to high resolution MS (LC‐HRMS) or to Orbitrap MS (LC‐Orbitrap‐MS) are more commonly used for metabolomic studies and exploratory research (Moco et al., 2007; Caprioli et al., 2014). Hossain et al. (2015) reported LOQs for α‐solanine and α‐chaconine of 2 μg/L in potato extract, corresponding to 40 μg/kg in dehydrated potato powder. Nara et al. (2019) reported an LOQ of 1 and 2 μg/L in whole blood for α‐solanine and α‐chaconine, respectively.
1.3.2.3. Analytical standards, certified reference materials and proficiency testing schemes
Most of the main GAs and their aglycons are commercially available as analytical reference standards. However, the less common GAs (e.g. α‐dehydrotomatine and α‐dehydrotomatidine) as well as the partially hydrolysed forms of GAs (e.g. β‐ and γ‐solanine and β‐chaconine) are not commercially available. Isotopically labelled analogues of GAs are not available. No certified materials are commercially available and no proficiency testing schemes for testing of GAs, potatoes or potato products have been identified.
1.3.3. Sources
Botanical sources of GAs with a steroidal aglycone are mainly found within the dicotyledoneous family Solanaceae (see Section 1.3.1), and to a lesser extent in the monocotyledonous family Liliaceae (Roddick, 1996). From the latter family only few intact GAs have been isolated, and their fully structure elucidated, while a large number of steroidal alkaloids have been found and fully identified in extracts, particularly of Veratrum and Fritillaria species (Liliaceae) following acidic hydroflysis step (Li et al., 2006). In general, the two monocotyledonous genera are not known to be of any use as sources of food, although two species of so‐called ‘riceroot’ (Fritillaria camschatcensis [L.] Ker‐Gawl. and F. lanceolata Pursh) are reported to have been important root foods for the Indians of the Northwest Coast of North America (Turner and Kuhnlein, 1983).
This section focuses on the species of importance of today's food production from the genus Solanum, together with a few other Solanum species known as possible sources of intoxications in man and farm animals.
The genus Solanum L. that contains the GAs as defined in Section 1.3.1, include around 1,500 species, and as such the largest genus in the family Solanaceae (Knapp, 2013; Tepe et al., 2016). The genus comprises the potato (S. tuberosum), the tomato (S. lycopersicum L. – synonyms Lycopersicon lycopersicum (L.) H. Karst, and Lycopersicon esculentum Mill.), and the aubergine (S. melongena L.) (Milner et al., 2011). Less well‐known plants are gboma (S. macrocarpon L.) and the scarlet aubergine (S. aethiopicum L.) (Sánchez‐Mata et al., 2010). Other consumed food plants belonging to the genus Solanum L. are the pepino (S. muricatum, also known as sweet pepino or sweet cucumber) from which the ripe fruit is eaten (Sánchez‐Mata et al., 2010), and the wild aubergine (S. torvum) from which the fruits, known as susumber or turkey berries, are eaten as vegetables in the traditional Jamaican cuisine (Smith et al., 2008; Karmakar et al., 2015).
Non‐food Solanum species that produce fruits that might be accidentally ingested are the soda apple (S. aculeastrum Dunal, also known as goat apple or poison apple) and the wolf apple (S. lycocarpum St. Hil) (Bhat and Pallyath, 2016). The latter is a common bush in parts of Brazil, and the fruits are used in folk medicine, although the unripe fruits are slightly toxic, especially to pregnant women (Schwarz et al., 2007). Bittersweet nightshade (S. dulcamara) and black nightshade (S. nigrum L.) are common toxic weed plants (Milner et al., 2011).
For the potato (S. tuberosum), it is the starchy stem tuber that is eaten. Both sexual and vegetative reproduction occurs in the ancestral wild species and the improved cultivars. Thus, potatoes flower and set true seeds in berries (the fruit) following natural pollination by insects, while the tubers propagate clonally. For the tomato (S. lycopersicum), both the mature and the immature (green) fruits are used as food (Martínez‐Valverde et al., 2002). The food use of the aubergine (S. melongena) concentrates around the mature fruit. For the production, the plant is grafted onto other species of Solanum, such as S. torvum (Miceli et al., 2014).
Different plant parts contain different concentrations of the individual GAs and TGAs. Furthermore, both the total TGA content and the relative proportions in a plant part (organ) will change as a result of the developmental stage of the organ.
The three main food crops (potatoes, tomatoes and aubergine) are discussed in more detail below.
1.3.3.1. Potatoes
S. tuberosum is the most cultivated tuber of the Solanum species. Potatoes are one of the main staple foods produced worldwide. In 2018, the global production volume was approximately 370 million tonnes (FAOSTAT, 2020a), which is about half the yearly volume produced of wheat and of rice. In the EU, Germany is the biggest producer of potatoes, with around 9 million tonnes in 2018, followed by Poland, France, The Netherlands and the UK with production volumes in the range of 5–8 million tonnes.
Potato breeding programmes have for quite a long time used many primitive forms of cultivated potatoes and their wild relatives as a valuable source of genetic variation (Distl and Wink, 2009; Bradshaw et al., 2006; van Gelder, 1989). Especially breeding programmes concerning disease resistance and environmental tolerance have been successful. Several methods to overcome hybridisation barriers have been developed (Machida‐Hirano, 2015).
Studies on the occurrence of GAs in commercial (common) potatoes (S. tuberosum ssp. Tuberosum) measured and reported only on α‐chaconine and α‐solanine, while the tubers of wild plants of the subspecies andigena have been reported to also contain, e.g. β‐chaconine (Distl and Wink, 2009). Both the common potato and other potato species, whether cultivated and consumed or not, may contain GAs other than α‐solanine and α‐chaconine. However, in general, the tuber concentrations of these other GAs are less than 5% of the total GA content (Friedman et al., 1997; Milner et al., 2011). In case these species are used in breeding, transfer (depending on the crossing methods used) of the biosynthetic pathway for the formation of these other GAs can occur.
According to Milner et al. (2011), the TGAs in different parts of the cultivated potato plant (S. tuberosum) will often be found within the concentration ranges given in Table 1 (see Figure 2 for the internal structure of a potato tuber). Using MALDI‐TOF‐MS imaging, the localisation and development of GAs in skin and sprouts were visualised by Ha et al. (2012). Berries of potatoes have about 10 times the levels of GAs compared to the tubers of commercial potato cultivars (Friedman, 1992).
Table 1.
Total glycoalkaloids (TGA) concentration (mg/kg fresh weight (fw)) in various cultivated potato (S. tuberosum) plant partsa
| Plant part | Concentration range (mg/kg fw) | Original Referencea |
|---|---|---|
| Flowers | 2,150–5,000 | (1), (2) |
| Leaves | 230–1,000 | (3) |
| Stems | 23–33 | (4) |
| Roots | 180–400 | (4) |
| Bitter‐tasting tubers | 250–800 | (5) |
| Whole tuber | 10–150 | (6) |
| Skin (2–3% of tuber) | 300–640 | (1), (2) |
| Peel (10–12% of tuber) | 150–168 | (1), (2) |
| Cortex | 125 | (6) |
| Flesh | 12–100 | (1), (2) |
| Pithb | Not detected–0.1 | (6) |
| Sprout | 2,000–7,300 | (1), (2) |
Figure 2.
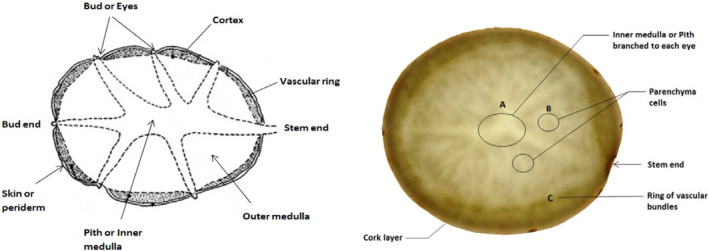
Internal structure of a potato tuber (from Faridnia et al. (2015)2)
In many regions of the world, the potato today is cultivated all year round by using different types of cropping and different cultivars adapted to these, i.e. to different temperature, day length and growing period among others (Mori et al., 2015). Besides the cultivar characteristics and the type of cropping, also the end use of the given potato matters. Potato use can be classified into four types, namely for ‘table use’, ‘industrial food processing use(s)’, ‘starch production’ and ‘other purposes’ (Mori et al., 2015). Due to the constant breeding for new cultivars, the GA content ranges given in Table 1 must be considered as approximate. Concerning potatoes with a food end use, the TGA content as well as the relative concentration of the different GAs has been reported in several studies. Details of the variation of GAs in different cultivars are discussed in Section 3.2.2.
The fact that these GAs are regarded as being important components of plant resistance against pests and pathogens means that breeders work to enhance the GA content in the non‐edible parts (Ginzberg et al., 2009). To do so, detailed knowledge of the biosynthetic pathway(s) and the regulation is essential. The major steps in the biosynthesis of α‐chaconine and α‐solanine have been summarised by Ginzberg et al. (2009) (Figure 3). GAs occur in most tissues of the S. tuberosum plant, except in the tuber pith (Table 1). Concerning the regulation of the biosynthesis and catabolism of the GAs, Sarquis et al. (2000) concluded that this most likely is happening at the tissue or organ level. This is because there is no evidence on their internal transport, and there is a poor correlation between the total content found in the tuber as compared to the foliage of the same cultivar (Sarquis et al., 2000; Ginzberg et al., 2009).
Figure 3.
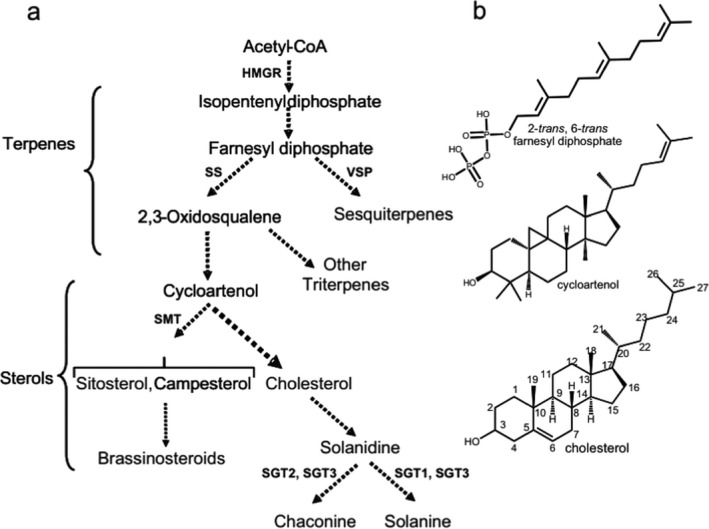
Simplified biosynthetic pathway for steroidal glycoalkaloids (GAs) (from Ginzberg et al., 2009)3
- (a) The biosynthetic pathway is presented from acetyl‐CoA to steroidal GAs with additional pathways included that use the same substrate. Dashed arrows indicate multiple enzymatic steps. Terpenes refer to low‐molecular weight isoprenoids. (b) Structural formulas for several intermediates of steroidal GA biosynthesis. HMGR: 3‐hydroxy‐3‐methylglutaryl coenzyme A reductase. SS: squalene synthase. VSP: vetispiradiene cyclase. SMT: sterol C24‐methyltransferase. SGT: solanidine glycosyltransferase.
The outer layers of the potato tuber (i.e. the periderm) contains the highest concentration of the GAs (Table 1), particularly in the 1.5 mm of tissue located under the surface of the tuber (Valkonen et al., 1996). It has been suggested that the biosynthesis of tuber GAs occurs in the phelloderm (Ginzberg et al., 2009), which is the inner part of the potato periderm.
Some reviews, especially in earlier times, have reported relatively small variations for the ratio between the concentrations of α‐chaconine and α‐solanine in potatoes (peel and flesh). However, later investigations have shown that this ratio may differ to a very great extent, depending on the cultivar as well as the part of the potato. Jin et al. (2018) analysed three different cultivars (‘Shepody’, ‘Russet Burbank’ and ‘Atlantic’) and found the ratio α‐chaconine:α‐solanine in the whole potato tuber to differ from 0.41 (‘Russet Burbank’), to 1.36 (‘Shepody’) and 3.61 (‘Atlantic’). Romanucci et al. (2016) investigated 15 cultivars and reported even higher ratio spans, e.g. in freshly harvested tubers, the ratio α‐chaconine:α‐solanine in the peel varied from < 0.06 to 5.7. After 20 days of storage, the ratio in the peel was still highly variable, but a general trend towards an increased content of α‐chaconine was noted.
Several factors have been studied whether they could influence the formation of GAs in potatoes. Preharvest factors such as the potato cultivar, the tuber size, climate, damage of the plants by insects, the soil composition, the use of fertilisers and growth conditions have been reported to potentially influence the content of GAs (e.g. Morris and Petterman, 1985; Gosselin et al., 1988; Mondy and Munshi, 1990; Hlywka et al., 1994; Love et al., 1994; Friedman et al., 1997; Bejarano et al., 2000; Zarzecka and Gugala, 2007; Najm et al., 2012). Some of these factors, such as the potato cultivar, location and farming system, are further discussed in Section 3.2.2.1.1.
Processing and other post‐harvest factors that affect the content of GAs in potatoes are described in Section 3.2.3.
1.3.3.2. Tomatoes
Besides being consumed fresh tomatoes are used in many processed foods such as canned and sun‐dried tomatoes, juices, ketchup, pastes, purees, salads, sauces and soups (Friedman, 2002). In 2018, worldwide production of tomatoes was 182 million tonnes (FAOSTAT, 2020b). Within the EU, Italy (5.8 million tonnes in 2018) and Spain (4.8 million tonnes) are the most important producers, followed by Portugal (1.3 million tonnes). In Europe, tomatoes are grown for the fresh market as well as for processing (tomato paste, juice, peeled tomatoes). Italy and Spain are major producers of processed tomato products.
The two most important GAs found in tomato (S. lycopersicum) are α‐tomatine and α‐dehydrotomatine, both of which are found in all parts of the tomato plant (Friedman, 2002). The first discovered GA, the fungitoxic and insect repellent α‐tomatine, has been shown to gradually disappear from the fruit with the appearance of lycopene (Sander, 1956, 1958). In addition to α‐tomatine and α‐dehydrotomatine, a variety of additional GAs have been identified in tomato fruits, including (dehydro)lycopersoside A to H, (dehydro)esculeoside A, B1, B2 and tomatoside A (Moco et al., 2007; Baldina et al., 2016). Some of these other GAs may be present, particularly in the ripe fruits, in higher concentrations than the marker compound α‐tomatine (Iijima et al., 2009; Baldina et al., 2016).
An example for the variations seen for the two most abundant GAs in the cultivated tomato plant parts is provided by Milner et al. (2011) (Table 2). From Table 2, it follows that the relative α‐dehydrotomatine content varies, depending on the plant part, from 6% to 25%. In the (unripe) fruits, α‐dehydrotomatine is present at approximately 10‐fold lower concentrations than α‐tomatine.
Table 2.
Glycoalkaloid (GAs) concentration (mg/kg fresh weight (fw)) and relative composition of parts of the cultivated tomato (S. lycopersicum) plant (Friedman, 2002)
| Tomato plant part | α‐tomatine (mg/kg fw) | α‐dehydrotomatine (mg/kg fw) | α‐tomatine (% of total) | α‐dehydrotomatine (% of total) |
|---|---|---|---|---|
| Large immature green fruit | 144 | 14 | 91 | 9 |
| Small immature green fruit | 465 | 54 | 90 | 10 |
| Root | 118 | 33 | 77 | 23 |
| Calyxes | 795 | 62 | 93 | 7 |
| Leaves | 975 | 71 | 93 | 7 |
| Small stems | 896 | 138 | 87 | 13 |
| Large stems | 465 | 142 | 75 | 25 |
| Flowers | 1,100 | 190 | 85 | 15 |
| Senescent leaves | 4,900 | 330 | 94 | 6 |
As for the potato plant (see above), the biosynthesis of the tomato GAs (α‐tomatine and α‐dehydrotomatine) is under separate genetic control in each plant part (Kozukue et al., 2004).
The TGA concentration in tomato depends, as for potato, on the cultivar and the fruit ripening stage. Eltayeb and Roddick (1984) studied the changes in the α‐tomatine content as a result of the fruit growth (e.g. given by the relative diameter of the berry) and/or the stage of ripening (e.g. defined by the development of the fruit colour: from dark green to full red or yellow colour depending on the cultivar), in commercially grown tomato cultivars and mutants with different ripening characteristics. For three normal‐ripening cultivars developing red‐, orange‐ and yellow‐ripe fruits, respectively, it was found that the highest concentrations occurred in very young green fruits of around 4 days of age (3,100–4,400 mg α‐tomatine/kg fw). In subsequent stages of fruit development, the concentration dropped to around 700 mg α‐tomatine/kg fw at day 10. In the same period, the fruit diameter increased from 7 mm at day 4–20 mm at day 20. By 30 days, when the tomato was approximately 30 mm in diameter, 20 g in weight and still green, α‐tomatine concentrations for the three cultivars had decreased to 3–6 mg/kg fw (Eltayeb and Roddick, 1984). Similar results were obtained by Kozukue and Friedman (2003), who reported an inverse relationship between fruit weight and α‐tomatine content in green ‘Momotaro’ tomatoes, with α‐tomatine concentrations of 900 mg/kg fw at 1.5–1.9 g of fruit weight, and of 150 mg/kg fw at 218.0–220.5 g of fruit weight. Choi et al. (2010) studied the α‐tomatine and α‐dehydrotomatine content of the tomato cultivar ‘Doturakworld’ for 60 days, discerning 11 stages of ripeness. The concentrations of α‐tomatine and α‐dehydrotomatine were highest during stage 1 at day 3, amounting to, respectively, 3,600 and 480 mg/kg fw. At growth stage 3 (14 days), α‐tomatine and α‐dehydrotomatine concentrations had dropped to 320 and 28 mg/kg fw, respectively. A steady but modest reduction was observed up to stage 7 (47 days), when the tomatoes were full grown, but still green (α‐tomatine and α‐dehydrotomatine concentrations, 140 and 15 mg/kg fw, respectively). When ripening set in at stage 8 (52 days), α‐tomatine and α‐dehydrotomatine concentrations had dropped below the LOQ (level not indicated) (Choi et al., 2010). It has been shown that during the later ripening stages, α‐tomatine is converted into esculeoside A and related glycosides (Fujiwara et al., 2004; Iijima et al., 2009; Nohara et al., 2010). The conversion of α‐tomatine into esculeoside A is correlated with enhanced levels of ethylene (Iijima et al., 2009). Moco et al. (2007) reported that concentrations of α‐tomatine, α‐dehydrotomatine and the lycoperosides A, B, C, F, G and H were highest in the epidermis, followed by the jelly parenchyma. During ripening the concentrations of lycoperosides F and G as well as that of esculeoside A strongly increased, while for the other GAs, a substantial decline was noted. A recent study of Tamasi et al. (2019) revealed that α‐tomatine and dehydrotomatidine are primarily located in the locular gel of the tomato fruits, with much lower concentrations in the skin and almost no TGAs present in the pulp.
For further details on the concentrations of α‐tomatine in tomato and tomato products reported in the literature, see Section 3.2.2.
1.3.3.3. Aubergine
The name aubergine refers to three crops from the genus Solanum subgenus Leptostemonum (Vorontsova et al., 2013). The three cultivated aubergine species are the Brinjal aubergine (S. melongena L.), the scarlet aubergine (S. aethiopicum L.) and the ghoma aubergine (S. macrocarpon). In this opinion, the term aubergine alone refers to S. melongena, unless stated otherwise.
The aubergine is generally cultivated in most tropical and subtropical regions of the world. The two other species (scarlet and ghoma) are less known and mostly grown in sub‐Saharan Africa (Taher et al., 2017), not only for their edible fruits but also for the young leaves (Caruso et al., 2017). The worldwide production of aubergines was 54 million tonnes in 2018 (FAOSTAT, 2020c). Production in the EU is relatively modest: around 0.85 million tonnes were produced in 2018, with Italy being the largest producer (300 ktonnes), followed by Spain (240 ktonnes) and Romania (140 ktonnes).
The plants are all berry‐producing vegetables. The aubergine complex shows a large morphological diversity, including small fruited spiny plants to large fruited non‐spiny plants. The long history of breeding has furthermore resulted in a wide range of cultivars with edible fruits of different sizes, shapes and colours (Caruso et al., 2017). Cultivars of aubergine are conventionally grouped into ‘Occidental’ and ‘Oriental’. The first are preferred in North Africa, Europe and the Americas, while the latter mostly are grown in East and South East Asia. They vary from one another with respect to their overall plant morphology and physiology, but also concerning their fruit size, colour and shape (Ceriola et al., 2013). Grafting on rootstocks of related Solanum species (e.g. S. torvum, S. aethiopicum, S. macrocarpon) or tomato (S. lycopersicum) hybrids has become an important area of research to improve the overall quality of the fruits and to increase resistance to soil‐borne diseases and abiotic stresses (Mennella et al., 2012; Sabatino et al., 2018).
For food purposes, aubergines are harvested at an immature stage and have a short storage period. Depending on the cultivar, the colour at the unripe stage may vary indeed very much, while the physiologically ripe fruits are brown or yellow (Caruso et al., 2017).
Together with α‐solamargine, α‐solasonine makes up the major content of GAs in this plant species. Both have the steroid solasodine as their aglycone, however, the sugar part in α‐solasonine is the trisaccharide solatriose where that of α‐solamargine is the trisaccharide chacotriose (see Figure 1). While the biosynthesis of α‐solasonine starts with the attachment of a D‐galactose to the aglycone, the biosynthesis of α‐solamargine is initiated with the attachment of a D‐glucose. Hence, the expression of solasodine galactosyltransferase 1 (SGT1) specifically reflects the biosynthesis of the GA α‐solasonine (Bagheri et al., 2017).
Mennella et al. (2012) studied the TGA content (α‐solasonine and α‐solamargine) in a total of 10 different genotypes from two different typologies (D3R and NAS), in commercially unripe, ripe and overripe (physiologically ripe) fruits. In the commercially unripe and ripe stages, the TGA content was relatively low (mean content: 125 and 230 mg/kg dry weight (dw), respectively), while in the commercially overripe stage, a strong increase of the TGA content (mean 1,937 mg/kg dw) was observed. The average ratio between α‐solasonine and α‐solamargine was approximately 1:2 in the commercially unripe and ripe stage, but changed to 1:1 in the overripe stage (Mennella et al., 2012).
Bagheri et al. (2017) compared two different aubergine genotypes, of which one (D1) had ‘sweet’ fruits and the other (J10) had ‘bitter’ fruits, with regard to the content of α‐solasonine (α‐solamargine was not analysed in this study) as well as concerning the expression of SGT1 in the leaves, flower buds, young fruits, mature fruits and physiologically ripe fruits. The levels of α‐solasonine closely followed the expression of the SGT1 gene in the same plant tissues. In all tissues, the concentration of α‐solasonine in the bitter variety was between two and three times higher than in the sweet variety. Concentrations of α‐solasonine reported for the D1 and J10 genotypes, respectively, were leaves: 113.3 and 249.3 mg/kg dw; flower buds: 135.6 and 478.8 mg/kg dw; young fruits: 61.3 and 160.4 mg/kg dw; mature fruits: 21.6 and 45.6 mg/kg dw and physiologically ripe fruits: 74.7 and 185.0 mg/kg dw (Bagheri et al., 2017).
1.3.4. Previous risk assessments
Previous human risk assessments
The general statement, that a level of 200 mg TGA/kg of unpeeled, uncooked potatoes could be considered as safe, is often found in the scientific literature and can be traced back for decades. Bömer and Mattis (1924) were probably the first to conclude that this TGA level provided sufficient protection to the consumer. They based their statement on the results of a study they conducted on batches of German potatoes from the 1922 harvest that was exceptionally high in solanine content. Batches of potatoes with a TGA content of 257–583 mg/kg potatoes reportedly caused intoxications or other health problems, while these effects were not reported for potatoes with levels < 200 mg/kg (concentrations determined with a gravimetric method).
The toxicity of α‐solanine and α‐chaconine was assessed by the Joint Food and Agricultural Organization/World Health Organisation (FAO/WHO) Expert Committee on Food Additives (JECFA) (JECFA, 1992, 1993). JECFA evaluated the kinetic aspects (absorption, distribution, biotransformation and excretion), toxicity and teratogenicity, primarily in rodents and humans, as available at the time of the evaluation. JECFA concluded that with the available experimental and epidemiological data, a safe level of intake could not be determined. The Committee reported that the occurrence of α‐solanine and α‐chaconine in potatoes is about 20–100 mg/kg, but the levels can increase due to mechanical damage, blight, sprouting, processing and storage. JECFA considered that daily consumption of potatoes containing normal GA levels was of no concern, provided that the potatoes were properly handled and grown.
In 1990, the Swedish National Food Administration carried out an ‘Assessment of Health‐risks Related to Glycoalkaloids (‘Solanine’) in Potatoes: A Nordic View’, and concluded that ‘Based on the present knowledge, a TGA concentration of 200 mg/kg potato for the potato varieties currently available on the Nordic Market appears to be the maximum level which can be accepted. However, because of the small safety margin, efforts should be made to reduce the levels of GA’. They recommended that the average TGA concentration in new potato varieties should not exceed 100 mg/kg (Nordic Working Group on Food Toxicology and Risk Assessment, 1990).
In 2018, and as a result of an intoxication of the members of a family in Baden–Württemberg (Germany) from potatoes that occurred in 2015, the German Federal Institute for Risk Assessment (BfR) published an assessment on the acute toxicity of potato GAs (BfR, 2018a,b). Jacket potatoes and potatoes boiled in the skin had been consumed before symptoms such as stomach ache and vomiting had developed. Analytical results showed that the potatoes contained 236 mg TGAs/kg (141 mg α‐solanine and 95 mg α‐chaconine per kg). Based on available human data in the literature, BfR identified a LOAEL of 1 mg GAs/kg body weight (bw) per day and extrapolated from this by applying an uncertainty factor of 2 to a NOAEL of 0.5 mg GAs/kg bw per day. This uncertainty factor of 2 was considered by BfR to also protect persons with higher susceptibility. In addition, BfR recommended that the intake of GAs should be below the NOAEL of 0.5 mg/kg bw per day to ensure a ‘margin of safety’ to the NOAEL being > 1. To avoid an exceedance of this NOAEL, BfR recommended that the TGA content of potatoes should not be higher than 100 mg/kg fresh weight. With this recommendation, particularly sensitive populations are covered too. In view of significant data gaps, BfR considered its conclusions as preliminary. Since case reports in humans indicate a lethal dose of 3–6 mg/kg bw, whereas the LD50 for mice and rats is at least 300‐fold higher, BfR pointed to the considerable higher sensitivity of humans compared to rodents.
Previous risk assessments on farm and companion animals
No risk assessments on farm and companion animals have been identified.
1.3.5. Legislation and other standards
In order to protect public health, Article 2 of the Council Regulation (EEC) No 315/934 stipulates that, where necessary, maximum tolerances for specific contaminants shall be established. Thus, a number of maximum tolerances for contaminants as well as natural plant toxicants are currently laid down in Commission Regulation (EC) No 1881/20065. However, no maximum levels for GAs in food or feed have been established at EU level under this or under another Regulation.
Some European countries have national legislation or recommendations on the maximum limits of TGAs, mainly in potato and potato products. Hungary has a national regulation of 100 mg/kg as the maximum limit of solanine equivalents of raw, unpeeled potatoes. In Finland, a maximum level for GAs (solanine glycosides, such as α‐solanine and α‐chaconine) for potatoes of 200 mg/kg6 exists, as well as in Sweden, where the National Food Administration's Regulations on Certain Foreign Substances in Food7 established in 2004 a maximum content of TGAs (total content of solanidine GAs, such as α‐chaconine and α‐solanine) in potatoes, raw and unpeeled, of 200 mg/kg. Also Denmark uses the Nordic guideline of 200 mg GAs/kg for known potatoes varieties, and 100 mg/kg for new potatoes varieties.
In the Netherlands, potato producers agreed to observe a limit of 100 mg/kg fresh weight for new potato cultivars (Bal, 1989, as cited in Essers et al., 1998). In Austria, no maximum level is laid down, but in the Regulation for marketing of food potatoes, there are minimum requirements for marketing of food potatoes to avoid a high content of GAs, such as the potatoes (i) must be free of noticeable green tubers, (ii) for class I potatoes: a slight green colour of not more than 1/8 of the surface is allowed, and (iii) for class II potatoes: a slight green colour, which can be removed by normal peeling is allowed.
In Germany, BfR recommended that the GA content of potatoes should not be higher than 100 mg/kg fresh weight (BfR, 2018a,b).
In Canada, a maximum level of 200 mg/kg for the sum of α‐solanine and α‐chaconine in fresh potato tubers was established, on the basis that most reported cases of adverse effects associated with potato GA exposure have occurred at concentrations above this value. This value was confirmed in 2014,8 indicating that it is achievable when good agricultural, manufacturing and storage practices are followed.9 In the USA, a maximum acceptable TGA content of potato tubers of 20–25 mg per 100 g of fresh potato (equivalent to 200–250 mg/kg) has been set according to the US‐Food and Drug Administration (FDA) poisonous Plant Database.10
The Organisation for Economic Co‐operation and Development (OECD) published in 2002 its ‘Consensus Document on Compositional Considerations for New Varieties of Potatoes: Key Food and Feed Nutrients, Anti‐Nutrients and Toxicants’.11 Under the considerations for the assessment of new potato varieties, it indicated that the comparison of the chemical composition of tubers from a modified variety with tubers from the non‐modified comparator should include, among others, the determination of GAs. It reported on the ‘widely accepted safety limit for the level of TGA in tubers’ of 200 mg/kg fresh weight (based on Bömer and Mattis, 1924 and Smith et al., 1996), and concluded that if the analyses of GAs (and other parameters) indicate that a novel variety is within the ranges given in the literature it can be considered equivalent with respect to its overall composition.
2. Data and methodologies
The CONTAM Panel applied the general principles for the assessment of chemicals in food as described by WHO/International Programme on Chemical Safety (IPCS) (WHO/IPCS, 2009). In addition, EFSA guidances pertaining to risk assessment were followed for the development of the risk assessment when necessary (EFSA, 2005, 2007, 2009, 2010a,b, 2011a,b; EFSA Scientific Committee, 2011, 2012a, b, 2017a, b, c, 2017d).
2.1. Methodology for data collection, selection of evidence and study appraisal
EFSA outsourced a call for an ‘Extensive literature search and selection for relevance of studies related to the chemistry and toxicity of GAs and quinolizidine alkaloids in food and feed’ aiming at identifying and evaluating literature related to the present assignment (and to another mandate of the CONTAM Panel on quinolizidine alkaloids which is not further considered here). The call was launched as a reopening competition for a specific contract under multiple framework contract CT/EFSA/AMU/2014/01 Lot 2. The University of Chemistry and Technology Prague was awarded with the contract and a final project report was delivered in November 2017 and was published (University of Chemistry and Technology Prague, 2019). The aim of the assignment was to identify and collect all relevant literature regarding GAs (and quinolizidine alkaloids) in food and feed covering the following areas: (i) chemistry and analysis, (ii) toxicokinetics in experimental animals, farm/companion animals, humans, and from in vitro studies and on biomarkers, (iii) in vitro and in vivo mode of action of toxicity, (iv) toxicity in vivo in laboratory animals, (v) toxicity in vitro, (vi) observations in humans, including epidemiological studies, case reports, outbreaks, intervention studies, (vii) adverse effects in farm and companion animals, (viii) occurrence in food, (ix) occurrence in feed and animal exposure and (x) influence of cultivars, maturity, environmental conditions, storage and processing in the concentrations of the alkaloids in the products. Details of the methodology and the results are reported in the University of Chemistry and Technology Prague (2019).
In addition to the literature searches outsourced by EFSA, additional complementary searches were performed. A literature search for studies regarding observations in humans was performed in October 2017 (for details see Appendix B, Section B.1). An extra search to identify studies on the toxicity of GAs was conducted in April 2018 (for details, see Appendix B, Section B.2). The literature searches were performed using Web of Science,12 PubMed13 and Scopus. The references resulting from the literature search were imported and saved using a software package (EndNote14), which allows effective management of references and citations.
Reviews, relevant scientific evaluations and toxicity studies by national or international bodies were also considered for the current risk assessment, i.e. JECFA (JECFA, 1992, 1993). When relevant papers were identified during the risk assessment process (e.g. from other studies or reviews), they were also considered.
In addition, a search for recent reviews was conducted to identify scientific publications dealing with methods of analysis, chemistry, formation in food, exposure and occurrence of GAs in food and feed.
A search for original studies reporting on the occurrence of GAs in food and feed samples was carried out, since it was expected that only limited number of occurrence data of GAs in food and feed were going to be submitted to EFSA (see Section 2.3.1).
An update of the literature available in the public domain was done regularly, on 17 December 2018, 3 May 2019 and 14 August 2019. Since that date, the literature was monitored to identify studies relevant for the risk assessment until the time of endorsement.
Any publications were used in the assessment if they were considered as relevant by applying expert judgement. In addition, for the studies on the toxicity of GAs in experimental animals and farm and companion animals, the selection criteria in Table 3 were used. For the sections on repeated dose toxicity and developmental and reproductive effects, animal studies based on i.p. or i.v. application of GAs or aglycones were excluded (see also Appendix G).
Table 3.
Selection criteria for the studies on the toxicity of glycoalkaloids (GAs) in experimental animals and in farm animals, horses and companion animals
| Inclusion criteria: | Exclusion criteria: |
|---|---|
|
|
The draft scientific opinion underwent a public consultation from 27 February 2020 to 15 April 2020. The comments received and how they were taken into account when finalising the scientific opinion were published in an EFSA Technical Report (EFSA, 2020).
2.2. Food and feed occurrence data submitted to EFSA
2.2.1. Data collection and validation
Following a mandate from the European Commission to EFSA, a call for annual collection of chemical contaminant occurrence data in food was issued by the former EFSA Dietary and Chemical Monitoring Unit (now DATA Unit)15 in December 2010 with a closing date of 1 October of each year.16 European national authorities and similar bodies, research institutions, academia, food business operators and other stakeholders were invited to submit analytical data on GAs in food through this call. The data submitted to EFSA on GAs were provided by national authorities from Germany, Sweden and the Netherlands.
The data submission to EFSA followed the requirements of the EFSA Guidance on Standard Sample Description for Food and Feed (EFSA, 2010a), and the occurrence data were managed following the EFSA standard operational procedures (SOPs) on ‘Data collection and validation’ and on ‘Data analysis of food consumption and occurrence data’.
By the 2 September 2019, a total of 1,300 analytical results on GAs in food (representing 651 food samples) were available in the EFSA database. Data received after that date were not included in the data set used for further evaluation for this opinion. All data submitted to EFSA concerned potato and potato products. No data were submitted concerning GAs in tomato, aubergine and food products containing tomato or aubergine.
In addition, in January–February 2019, a consultation was held with registered EFSA Stakeholders and other stakeholders to inform the work of the CONTAM Panel and its working group in developing this scientific opinion. The aim of the consultation was to collect information which were not readily available in the public domain regarding GAs occurrence data in food and feed, factors that influence the content of GAs and toxicity (see Section on Documentation provided to EFSA). Following this consultation, EFSA invited the stakeholders to submit occurrence data in food and feed within the annual collection of chemical contaminant occurrence data, following the requirements of the EFSA Guidance on Standard Sample Description for Food and Feed (EFSA, 2010a) for inclusion in the risk assessment. As a result, on 2 July 2019, the European Starch Industry Association submitted occurrence data to EFSA concerning 1,728 samples on dietary fibre and potato proteins, pulp, juice, starch and on potatoes used for starch production. See Section 3.2.1 for further details.
2.2.2. Data analysis
Following the EFSA SOP on ‘Data analysis of food consumption and occurrence data’ to guarantee an appropriate quality of the data used in the exposure assessment, the initial data set was carefully evaluated applying several data cleaning and validation steps. Special attention was paid to different parameters such as ‘Sampling strategy’, ‘Reporting unit’ and the codification of the different samples under FoodEx classification. The outcome of the data analysis is presented in Section 3.1.1.
The left‐censored data (results below LOD or below LOQ) were treated by the substitution method as recommended in the ‘Principles and Methods for the Risk Assessment of Chemicals in Food’ (WHO/IPCS, 2009). The same method is indicated in the EFSA Scientific Report ‘Management of left‐censored data in dietary exposure assessment of chemical substances’ (EFSA, 2010b) as an option in the treatment of left‐censored data. The guidance suggests that the lower bound (LB) and upper bound (UB) approach should be used for chemicals likely to be present in the food (e.g. naturally occurring contaminants, nutrients and mycotoxins). The LB is obtained by assigning a value of zero (minimum possible value) to all samples reported as lower than the LOD (< LOD) or LOQ (< LOQ). The UB is obtained by assigning the numerical value of LOD to values reported as < LOD and LOQ to values reported as < LOQ (maximum possible value), depending on whether LOD or LOQ is reported by the laboratory.
2.3. Food and feed consumption data
2.3.1. Food consumption data
The EFSA Comprehensive European Food Consumption Database (Comprehensive Database) provides a compilation of existing national information on food consumption at individual level. Details on how the Comprehensive Database is used are published in the Guidance of EFSA (EFSA, 2011a). Consumption data were collected using single or repeated 24‐ or 48‐h dietary recalls or dietary records covering from 3 to 7 days per subject. Subjects in the Comprehensive Database are classified according to the following subpopulations:
Infants: < 12 months old
Toddlers: ≥ 12 months to < 36 months old
Other children: ≥ 36 months to < 10 years old
Adolescents: ≥ 10 years to < 18 years old
Adults: ≥ 18 years to < 65 years old
Elderly: ≥ 65 years to < 75 years old
Very elderly: ≥ 75 years old.
Two additional surveys provided information on specific population groups: ‘Pregnant women’ (≥ 15 years to ≤ 45 years old; Latvia) and ‘Lactating women’ (≥ 28 years to ≤ 39 years old; Greece).
Owing to the differences in the methods used for data collection, direct country‐to‐country comparisons can be misleading. Overall, the food consumption data gathered by EFSA in the Comprehensive Database are the most complete and detailed data currently available in the EU.
Some of these data, however, cannot be used in exposure assessments if the relevant occurrence data are reported at the level of the raw primary commodities (RPCs). The RPC model developed by EFSA in 2019 aimed to bridge this gap by transforming food consumption data from the Comprehensive Database for composite foods (foods consisting of multiple components) and RPC derivatives (single‐component foods which have been physically changed by processing) into their corresponding quantities of RPCs. The RPC database also contains information on products derived from potato that have undergone a further refining, distillation and/or isolation process, i.e. potato starch and alcoholic beverages produced from potato by‐products by distillation. Details on how the RPC database was developed are published in a dedicated technical report (EFSA, 2019). The RPC Consumption Database, based on the Comprehensive Database version published on the 31 March 2018, contains 51 dietary surveys from 23 different countries. These surveys cover a total of 94,532 subjects and 26,573,088 RPC consumption records.
The use of the RPC Consumption Database was considered as the most appropriate approach for the current assessment for which occurrence data in the RPC main‐crop potato were available (see Section 2.5). As no occurrence data were available for GAs in tomato and aubergine, these foods were not included in the consumption and exposure assessment.
Detailed information on the different dietary surveys used in this report for the acute exposure assessment is shown in Annex A.1, including the number of subjects and days available for each age class.
2.3.2. Feed consumption data
Of the large number of species within the Solanaceae family, only potatoes and by‐products derived from them are used as feed for livestock. Due to the high cost of growing the crop, and the value of potatoes as food for human consumption, they are not usually grown as a feed for livestock, and whole (raw) potatoes are only used as feed when production is in surplus to requirements as food, or when they are discarded because they do not meet market requirements because of their size, appearance, diseases or physical damage. As a result, the use of raw tubers as livestock feed tends to be opportunistic, and the CONTAM Panel have not identified any data on quantities used in this way.
Feed grade potato tubers are mainly used as feeds for ruminants but may also be fed to horses. They are highly palatable and nutritious and dairy cows will typically consume up to 25–30 kg of potatoes (Crawshaw, 2001). For growing cattle, up to 30% of the diet (dry matter) may be fed (Fiems et al., 2013).
Raw potatoes are relatively unpalatable, and the starch indigestible for pigs and poultry, and therefore, it is recommended that they are cooked or steamed before being fed (Whittemore, 1977). They may be included at up to 15% of the diet dry matter for finishing pigs and sows (Ewing, 2002), and up to 50% of the dietary dry matter for growing and finishing pigs (Pond and Maner, 1984). Inclusion of cooked potatoes in poultry grower diets at up to 20% have been recommended, while for layers, a dietary inclusion rate of 15% potato meal did not reduce egg production (Whittemore, 1977).
In addition to raw potatoes, the EU Catalogue of Feed Materials17 lists 14 different by‐products from potato processing industries, including cuttings, scrapings, flakes and pulp. The suitability of these as feeds for different livestock depends on the degree of processing to which they have been exposed, and the nutritional requirements of the livestock.
Steamed potato peel is a highly nutritious and palatable feed for pigs, and its availability has increased in recent years in line with the increase in the production of French fries. Low‐GA potato protein concentrate is often added to low protein pig starter diets to reduce the incidence of diarrhoea in newly weaned pigs (Stein, 2002).
The use of potato protein concentrate in poultry diets is generally limited, with some research reporting an increase in necrotic enteritis in broilers.
Potato protein concentrate was used as an alternative to fishmeal in the diets of rainbow trout, with limited success (Tusche et al., 2011), and has been included in specialist feeds for dogs.18 However, information on amounts of this or other potato by‐products used as animal feeds within the EU is not publicly available. Since the highest concentrations of GAs are found in the skin (Milner et al., 2011), and these are removed at an early stage in the processing, levels of GAs in by‐products such as potato starch and potato protein concentrate are likely to be low.
2.4. Food classification
Consumption data were classified according to the FoodEx classification system (EFSA, 2011b). FoodEx is a food classification system developed by EFSA in 2009 with the objective of simplifying the linkage between occurrence and food consumption data when assessing the exposure to hazardous substances. It contains 20 main food categories (first level), which are further divided into subgroups having 140 items at the second level, 1,261 items at the third level and reaching about 1,800 end points (food names or generic food names) at the fourth level.
In 2011, a new version of FoodEx, named FoodEx2, has been developed and is described in the scientific document ‘Report on the development of a Food Classification and Description System for exposure assessment and guidance on its implementation and use’ (EFSA, 2011b). The last release of FoodEx2 complements the previous hierarchical classification system of basic codes with more detailed food levels and gives the possibility of reporting additional information through the use of facets and facet descriptors (EFSA, 2015).
For this opinion, the amount consumed for each reported FoodEx code was converted to the equivalent amount of RPC (main‐crop potatoes) by disaggregating complex foods into their components (the original RPC and RPC derivatives) and applying relevant reverse yield factors taking into account effects of processing, as described by the RPC model (EFSA, 2019).
2.5. Methodology for Exposure assessment
2.5.1.
Human exposure assessment
The CONTAM Panel estimated the acute exposure to GAs (see Section 3.1.6).
Since available occurrence data did not cover all the food categories containing potatoes in the Consumption Database (see Section 3.2.1), it was decided that the best approach for the exposure assessment would be to use the occurrence values in the RPC (main‐crop potatoes and new potatoes) and the RPC Consumption Database (see Section 2.3.1). A scenario including only days in which there was consumption of main‐crop potatoes (or new potatoes, see Section 3.2.1) was considered the one that best characterises the acute exposure.
The acute dietary exposure to GAs was estimated by using a probabilistic approach based on the method of random sampling with replacement of occurrence data, of processing steps where the information was not available, and of processing factors. The random sampling captures the variability of occurrence values and of processing factors. Random sampling of consumption events was not performed because potatoes are a widely consumed food and GAs are present in all samples. Thus, the CONTAM Panel concluded this would have a limited impact on the results. The exposure was calculated on a per day basis. Acute exposure was assessed for each reporting day by multiplying the total daily consumption amount of main‐crop potatoes by one occurrence level randomly drawn among the individual results available. For the calculations of exposure, the UB values for the occurrence were used as considered more adequate to represent acute exposure also considering that in this assessment, the LB and UB of the analysed samples do not differ substantially. In the final data set, only four analytical results for α‐solanine and 22 results for α‐chaconine were left censored, and in only two samples, both GAs were not quantified. The LOQ for samples with left censored results ranged from 0.75 to 5 mg/kg.
Since peeling of potatoes and heat processing decrease the content of GAs (see Section 3.2.3), the CONTAM Panel applied processing factors linked to peeling (where applicable) and to heat processing (to all eating events) to estimate the GA levels in the final food consumed based on available information as follows:
-
–
Information on peeling was not available in the RPC Consumption Database; thus, the CONTAM Panel assumed that in 90% of all eating events, potatoes were consumed peeled. Possible differences among countries or regions were not taken into consideration. A probabilistic approach was used to assign to each eating event the ‘peeled’ vs. ‘unpeeled’ status. Based on studies in the literature (see Section 3.2.3), a processing factor between 0.25 and 0.75 due to peeling was attributed to the peeled potatoes only.
-
–
Information about the heat processing methods was only available in a limited number of cases.
-
–
A probabilistic approach was also used to assign one of the heat processing methods to each eating event. The proportion of each of the heat processing methods within all eating events was based on available information in the RPC consumption database (frying: 23%, deep frying: 12%, cooking in water: 47%, drying, flaking and roasting: 5% each, and stewing: 2%). Possible differences among countries or regions were not taken into consideration.
-
–
Based on literature studies that reported on the percentage reduction for specific processing steps (see Section 3.2.3), processing factors between 0.1 and 0.8 were attributed to frying and deep frying, and between 0.35 and 0.95 for all other heat processing methods.
-
–
The processing factors for peeling and heat processing assigned to each eating event were extracted from a normal distribution, where the minimum and maximum allowed values were the ranges described above. Results were compared with an exposure assessment scenario where processing factors were extracted from an uniform distribution rather than a normal distribution and the exposure outcome was very similar (see Table A.7 in Annex A).
Exposures from the different eating events of one day (for the considered subject) were summed and divided by the individual's body weight to obtain the total exposure per kg bw per day.
Consumption of main‐crop potatoes linked to the consumption of alcoholic beverages (vodka and distilled spirits from potato) was excluded from the consumption and exposure assessment, as the CONTAM Panel considered the transfer of GAs from the potatoes during the distillation and refining process to be negligible. Exposure to potato GAs from potato starch was included in the exposure assessment. Due to insufficient occurrence data in potato starch and the lack of processing factors related to starch production, the occurrence data and processing factors for peeling and heat processing for main‐crop potatoes were used. The Panel noted that the overall concentration of GAs in potato starch may be low (Saito et al., 1990; Driedger and Sporns, 1999), and that in most of the consumption surveys, the contribution of potato starch to the overall potato consumption is below 5%. The Panel considered the impact of potato starch in the overall GA intake to be low.
The mean and P95 daily acute UB exposure were then calculated per survey and age class.
To model the uncertainty, the process was iterated 1,000 times. The mean of the 1,000 mean and P95 daily acute UB exposures per survey and per age class was then calculated. The 95% confidence interval (CI) defined as the interval between the 2.5th and 97.5th percentiles obtained from the 1,000 iterations was determined to indicate the uncertainty around the mean value.
In Table A.1 of Annex A, the number of available subjects for each age class used in the acute exposure assessment is provided.
Animal exposure assessment
Due to insufficient data on levels of GAs in either potatoes or potato by‐products used as animal feeds, and lack of data with which to calculate reference points for any of the farm animal species (see Sections 3.1.4 and 3.1.7), no estimates of exposure to GAs have been made.
2.6. Methodology for Risk characterisation
The CONTAM Panel applied the general principles of the risk characterisation process for chemicals in food as described by the WHO/IPCS (2009) and the relevant EFSA guidance documents.
3. Assessment
3.1. Hazard identification and characterisation
3.1.1. Toxicokinetics
3.1.1.1. Experimental animals
The studies identified on the toxicokinetics of GAs in experimental animals are described below, and details are given in Appendix C.
3.1.1.1.1. α‐Solanine
Nishie et al. (1971) studied the metabolic fate and distribution of α‐solanine in male Fisher rats. [3H]‐α‐solanine was administered as a single treatment either by gavage at 5 mg/kg bw or i.p. injection at 5–25 mg/kg bw to an unreported number of rats. Following oral exposure, fecal elimination of radioactivity reached 84% of the administered dose by day 4, with urinary excretion accounting for less than 10%. 24 h after the oral exposure, radioactivity distributed mainly in the liver (about 1.5% of the administered dose), followed by blood, kidney and lungs. Following i.p. injection of 5–15 mg/kg bw of labelled α‐solanine, radioactivity was eliminated at similar rates in urine and feces (approximately 15–20% of the administered doses by 24 h). Decreased urinary excretion and complete absence of fecal elimination were observed at the highest dose administered (25 mg/kg bw), with a corresponding increase in the radioactivity retention in the gastrointestinal tract, similarly to what reported by Norred et al. (1976) for α‐chaconine (see Section 3.1.1.1.2). Lesions of increasing severity were observed in the peritoneal cavities of animals exposed to ≥ 10 mg/kg bw. A metabolic study was performed using TLC. Following oral exposure, 65% of the radioactivity present in feces was identified as solanidine, whereas only 6% of the dose excreted in urine was identified as solanidine, with two other non‐identified metabolites accounting for 80% and 13% of the radioactivity excreted in urine.
Groen et al. (1993) studied the toxicokinetics of [3H]‐α‐solanine in male rats and hamsters. [3H]‐labelled α‐solanine was administered via i.v. injection or via gavage at a dose of 54 and 170 μg/kg bw, respectively. Following i.v. administration, a similar toxicokinetics of radioactivity in blood and plasma was observed in rats and hamsters. Plasma levels of total radioactivity and unchanged α‐solanine showed three‐phase profiles (see Table 4). Compared to i.v. treated animals, the oral bioavailability of α‐solanine and metabolites (based on total radioactivity) in animals treated by gavage was 29% and 57% in rats and hamsters, respectively. The oral bioavailability for the parent compound α‐solanine was 1.6% and 3.2% in rats and hamsters, respectively. Plasma clearance of unchanged α‐solanine following oral administration was significantly faster in rats (t1/2 = 7.79 ± 0.83 h) than in hamster (t1/2 = 19.7 ± 4.2 h); however, similar plasma clearance was observed for total radioactivity in the two species (82.2 ± 6.1 h in rats, and 94 ± 33 h in hamsters). Excretion of total radioactivity in urine and feces was significantly faster in rats than in hamsters treated by gavage. Seven days after the oral treatment, total excretion of the administered dose was 89% in rats, with approximately 3% in urine and 86% in feces; in hamsters, total excretion was 39%, with approximately 10% in urine and 29% in feces. Macroscopic observation performed at the necropsy of the animals sacrificed 7 days after the treatment revealed damages in the intestinal walls of some hamsters following oral treatment.
Table 4.
Plasma half‐lives (tri‐phase elimination) of total radioactivity and unchanged α‐solanine following i.v. injection of 54 mg/kg bw [3H]‐α‐solanine in rats and hamsters (Groen et al., 1993)
| T1/2α (h) | T1/2β (h) | T1/2τ (h) | ||
|---|---|---|---|---|
| Total radioactivity | Rat | 1.4 ± 1.6 | 8.9 ± 2.2 | 61 ± 13 |
| Hamster | 1.5 ± 0.4 | 7.7 ± 3.0 | 62 ± 22 | |
| Unchanged α‐solanine | Rat | 1.9 ± 1.4 | 9.9 ± 0.8 | 32 ± 6.5 |
| Hamster | 2.1 ± 1.4 | 8.9 ± 2.8 | 39 ± 13 |
3.1.1.1.2. α‐Chaconine
Norred et al. (1976) studied the toxicokinetics of [3H]‐α‐chaconine (tritiated in the aglycone moiety at carbon atoms adjacent to the double bond and to the nitrogen atom) in male Sprague‐Dawley rats (200–300 g bw). Rats were dosed either by gavage (5 mg/kg bw) or i.p. injection (5–25 mg/kg bw) and housed in metabolic cages up to at least 100 h after treatment. Following oral administration of 5 mg/kg bw, fecal elimination of 60% and 80% of the administered radioactivity dose was observed after 24 and 48 h, respectively. Urinary excretion reached a plateau of 10% of the administered radioactivity dose by 24 h. Maximum levels in tissues were measured after 6–12 h, with higher levels in liver (about 1.3% of the radioactivity dose) and blood (about 0.2%). The radioactivity recovery considering the sum of fecal elimination, urinary excretion and gastrointestinal content, ranged approximately from 85% to 100% across the selected time points. Following i.p. injection of 5–10 mg/kg bw, elimination took place via the urine (10–15% of the administered radioactivity dose excreted 24 h after treatment) and the feces (< 10%). However, reduced urinary excretion was observed at the highest dose administered (25 mg/kg bw); complete absence of fecal elimination was also observed at 15 and 25 mg/kg bw with a corresponding increase in the radioactivity detected in the gastrointestinal tract. A metabolic study was performed using TLC, which the CONTAM Panel considers to be of limited value. Following oral treatment with 5 mg/kg bw, about 63% and 16% of the radioactivity present in feces and urine was extracted with ether, respectively. TLC analysis of the feces and urine extracts revealed the presence of a main metabolite with the same retention factor of solanidine. About 25% of radioactivity detected in feces was attributable to the parent compound.
The distribution of radioactivity in subcellular fractions of hepatocytes in female Swiss‐Webster mice exposed to α‐chaconine was investigated by Sharma et al. (1983). Following oral administration of 10 mg/kg [3H]‐α‐chaconine, mice were sacrificed at 3, 6, 14, 72 and 120 h after exposure and liver tissue and blood were sampled. The distribution of radioactivity in subcellular fractions of hepatocytes was studied. Higher radioactivity levels in liver homogenate were measured from 3 until 14 h after the exposure, with the nuclear subcellular fraction containing the highest levels, followed by mitochondria and microsomes. In a second experiment, mice were orally exposed to different doses (1, 3 or 10 mg/kg) and sacrificed after 14 h. The radioactivity distribution in liver homogenate and in subcellular fractions was observed to increase consistently with the administered dose. Finally, mouse pre‐treatment with the cytochrome P450 inducer dibenzo[a]pyrene resulted in lower radioactivity levels measured in all the subcellular fractions, possibly suggesting an increased metabolism of α‐chaconine due to enzyme induction by dibenzo[a]pyrene.
Alozie et al. (1979a) studied the toxicokinetics of [3H]‐α‐chaconine in male Golden hamsters administered a dose of 10 mg/kg bw by gavage or i.p. injection. Hamsters were sacrificed at regular time intervals (3 animals per time point) up to 168 h after treatment and radioactivity was measured in different tissues. Urine and feces were collected and measured for radioactivity. Subcellular distribution of labelled α‐chaconine was also studied in various tissues. After the oral exposure, distribution of radioactivity in various tissues was observed at the different time points, in particular higher levels were observed in lungs, liver and skeletal muscles. Radioactivity levels remained high in the intestine throughout the oral study and also relatively high in the i.p. study, possibly suggesting high biliary excretion. Following oral exposure, the excretion of radioactivity occurred mainly via the urine reaching about 21% of the administered dose after 168 h, whereas only a minor fraction was detected in feces. The study of subcellular distribution showed α‐chaconine or its metabolites to be distributed mainly in the nuclear, microsomal and soluble cellular fractions in various tissues including brain, heart and liver. The residual radioactivity present in the cellular subfractions following a wash procedure with methanol suggested that a fraction of α‐chaconine (or α‐chaconine metabolites) could bind to macromolecules (type not specified) in these tissues.
In a follow‐up publication, Alozie et al. (1979b) studied the metabolic fate of α‐chaconine in Golden hamsters following oral exposure to 10 mg/kg bw [3H]‐ α‐chaconine. Urine and feces were collected at different time points up to 168 h after exposure and subsequently extracted with water or chloroform. These extracts were analysed by TLC. In urine, initially unaltered α‐chaconine appeared to be the only excreted substance (100% of detected radioactivity at the 12 h time point, corresponding to 0.25% of the administered dose), showing a declining trend towards the end of the study. Solanidine appeared in urine 24 h after the exposure and reached a maximum level after 72 h. Four other metabolites could be detected but not identified, two of them representing the major peaks at 168 h. The total excretion in feces represented only a minor amount of the administered dose (see Alozie et al., 1979a) and was mainly constituted by the parent compound.
3.1.1.2. Humans
The studies identified on the toxicokinetics of GAs in humans are described below, and details are given in Appendix C.
3.1.1.2.1. Mixtures of α‐solanine and α‐chaconine
The kinetics of α‐solanine and α‐chaconine in human volunteers was studied by Hellenäs et al. (1992). Seven healthy male adults were administered a peeled mashed potato meal containing 82 mg/kg α‐solanine and 118 mg/kg α‐chaconine. The study subjects were given a portion adjusted to reach a dose of 1 mg TGAs/kg bw. Blood was sampled at regular intervals prior the exposure and up to 25 h following the exposure and the content of α‐solanine, α‐chaconine and solanidine were determined via HPLC analysis. Average serum Cmax for α‐solanine and α‐chaconine were 7.7 and 14.4 ng/mL, with average Tmax of 5.1 and 6 h, respectively. Solanidine reached the maximum level after 8 h (ranging from 1.0 to 4.8 ng/mL across subjects) and remained apparently constant until the end of the study. Average apparent biological half‐lives were calculated to be approximately 11 and 19 h for α‐solanine and α‐chaconine, respectively, with low variability across subjects. The CONTAM Panel noted that the reliability of the calculated half‐lives was limited by the short observation period (25 h). Adverse/undesirable effects were observed in most of the study volunteers, as described in Section 3.1.3.
Mensinga et al. (2005) studied the pharmacokinetics and toxicity of α‐solanine and α‐chaconine in six male and eight female healthy volunteers treated with escalating doses of the two GAs either via solutions (0.30, 0.50 or 0.70 mg TGA/kg bw) or in a form of a mashed potato meal (0.95, 1.10 or 1.25 mg total TGA/kg bw). Both the solution and the potato meals contained approximately equal amounts of the two GAs. Blood was sampled from all subjects at various time points as from 1 h before treatment till 96 h after treatment and serum GA levels were measured by HPLC analysis. Both α‐solanine and α‐chaconine showed similar tmax in serum ranging from 4 to 8 h without correlation with the administered doses. Maximum serum concentrations (Cmax) showed strong positive correlations for α‐solanine and α‐chaconine both for administration via solution (correlation coefficient 0.98) and via potato meal (0.97) (see Table 5). A similar result was obtained for the 0–24 h areas under the curve (AUC0‐24), whereas a significant correlation was observed only in subjects treated with mashed potatoes for the AUC from point zero to infinity (AUC0‐∞). Serum half‐lives showed a high inter‐individual variability and were generally longer for α‐chaconine (27–84 h, average of 44 h) than for α‐solanine (5–42 h, average of 21 h). One volunteer exposed to 1.25 mg/kg bw experienced adverse effects as described in Section 3.1.3.1.2.
Table 5.
Pharmacokinetic parameters (ranges) of α‐solanine and α‐chaconine in human volunteers after oral exposure (adapted from Mensinga et al., 2005)
| GAs solution | GAs in mashed potatoes | |||||
|---|---|---|---|---|---|---|
| Dose (mg/kg bw) | 0.30 | 0.50 | 0.70 | 0.95 | 1.10 | 1.25 |
| Number of subjects | 2 | 2 | 2 | 3 | 3 | 2 |
| α‐Chaconine | ||||||
| C max (μg/L) | 2.8–3.2 | 3.2–4.5 | 3.3–6.3 | 5.2–7.4 | 2.9–7.8 | 6.3–9.7 |
| T max (h) | 5–6 | 5–6 | 4‐5 | 5–7 | 5‐8 | 7–12 |
| T 1/2 (h) | 32–37 | 49–84 | 29–39 | 27–49 | 49–60 | 37–45 |
| AUC 0‐∞ (μg h/L) | 117–130 | 148–242 | 128–227 | 237–345 | 168–316 | 299–419 |
| α‐Solanine | ||||||
| C max (μg/L) | 2.9–3.9 | 3.5–4.7 | 3.1–6.5 | 7.0–10.7 | 3.8–10.6 | 8.0–11.8 |
| T max (h) | 5–6 | 5–6 | 4–4 | 5–5 | 5–8 | 7–8 |
| T 1/2 (h) | 17–35 | 5–28 | 8–17 | 14–18 | 1–‐42 | 18–19 |
| AUC 0‐∞ (μg h/L) | 84–89 | 47–110 | 50–86 | 164–163 | 111–200 | 204–256 |
3.1.1.2.2. Solanidine
Claringbold (1982) administered an unreported dose of [3H]‐solanidine by a single i.v. injection in two male and one female volunteers. Blood was sampled at various times and urine and feces were collected. The decrease of radioactivity in plasma followed a three‐phase trend and levels were detected up to 150 h after the treatment. The authors estimated that in blood, about 70% of radioactivity detected was retained by erythrocytes, which could explain the slow clearance from plasma. After 24 h, only 3–7% of the administered radioactivity was excreted in urine and feces.
The same authors also studied the presence of solanidine in five post‐mortem liver samples. The extraction procedure from liver homogenates identified four solanidine containing fractions (free, glucuronide, glycoside and a residual fraction obtained following acid hydrolysis). In two out of five liver samples, no solanidine was detected in any of the four fractions. In another sample, no free solanidine was found and solanidine was observed mainly as the glucuronide (70%). In the two remaining samples, free solanidine accounted for 25 and 15%, respectively. In their acid hydrolysed fraction, solanidine accounted for 42 and 60%, respectively.
3.1.1.2.3. Biomarkers of exposure
Three studies were identified in which a radioimmunoassay method was developed to quantify solanidine in human plasma. In the first study, Matthew et al. (1983) described the method and reported the results on 34 human donors. One limitation of this method was that the rabbit immune serum developed for the radioimmunoassay had a 100% cross‐reactivity with α‐solanine, α‐chaconine and demissidine. However, a preliminary extraction step using chloroform allowed the specific determination of solanidine. The method was applied on plasma of 34 human donors (7 males and 27 females), randomly selected from a hospital clinic. All plasma samples showed quantifiable levels of solanidine, with mean levels of 1.56 ± 1.17 and 1.20 ± 0.93 ng/mL calculated for males and females, respectively.
The assay developed by Matthew et al. (1983) was subsequently applied by Harvey et al. (1985a) to study the levels of solanidine in 57 volunteers (30 males and 27 females). The consumption of potatoes and potato products of 33 subjects was recorded for one month, during winter. Two male volunteers maintained a potato‐free diet for several weeks while their serum was regularly analysed for solanidine levels. The mean levels of solanidine were 10.8 ± 5.4 ng/mL and 7.9 ± 4.3 ng/mL in males and females, respectively. A positive correlation between consumption of potatoes and potato products and serum solanidine levels was found both in male and female subjects in the subgroup that recorded their diets. Avoidance of potato consumption corresponded to a drop in the serum levels of solanidine, reaching minimum levels (lower than the assay sensitivity of 0.5 ng/mL) 3 weeks after the cessation of potato intake.
In a subsequent study, Harvey et al. (1985b) adapted the radioimmunoassay method to quantify both solanidine and the TGA levels in serum and saliva. Two groups of volunteers (18 males and 15 females from UK, and 5 males and 5 females from Sweden) were analysed for levels of GAs during summer. All the subjects maintained their standard diet during the study, with the exclusion of three Swedish volunteers, who had daily consumption of two potato cultivars known to be high in GAs for a week before serum and saliva sampling. Mean levels of solanidine and TGAs (all subjects) were, respectively, 14.5 (5.8 ng solanidine/mL) and 38.5 nmol/L in serum, and 3.2 (1.3 ng solanidine/mL) and 3.7 nmol/L in saliva. No marked differences were observed among the UK and Swedish subjects following standard diets. Significant correlations were shown between serum and salivary levels both for solanidine and TGAs, and between serum solanidine and serum TGA levels. The three subjects consuming potato varieties high in GA levels had substantially higher serum and salivary levels compared to the other Swedish subjects (e.g. in serum solanidine mean levels were 10.5 nmol/L (4.2 ng solanidine/mL) in the seven subjects following standard diets, and 80.8 nmol/L (32.3 ng solanidine/mL) in the three subjects with high‐alkaloid diets).
3.1.1.3. Farm animals, horses and companion animals
Only scant information is available on the toxicokinetics of GAs in ruminants.
King and McQueen (1981) studied the effect of rumen on α‐solanine and α‐chaconine in vitro. Rumen contents were collected from a rumen‐fistulated cow before the morning feeding and processed to remove feed particles. A mixture of α‐solanine and α‐chaconine was isolated from potato blossom and added to the rumen inoculum at a concentration of 1 mg/mL. At different time points up to 48 h, chloroform extracts were analysed by GC‐NPD. The analyses revealed the formation of solanidine, that reached a maximum concentration at 12 h, when a subsequent conversion to a 5,6‐dihydro derivative of solanidine, identified as 5β‐solanidan‐3β‐ol, was observed. By 24 h, the hydrolysis of the GA mixture reached a plateau and the sum of the two aglycones recovered from the rumen inoculum were on average 83% of the test concentration (approximately 30% constituted by solanidine and 50% by its 5,6‐dihydro derivative).
Bushway et al. (1984) studied the potential transfer of GAs into cow's milk. Three samples of potato meal were analysed for their GA content and incorporated at 10 or 20% (w/w) in bovine feed ration, corresponding to 2.25 and 4.5 kg potato meal per day, respectively. The mean contents of α‐chaconine and α‐solanine in the potato meal samples were 515 and 285 mg/kg dw, respectively. This resulted in estimated daily exposure of 3.6 and 2.0 mg/kg bw for α‐chaconine and α‐solanine, respectively, at the highest level of potato meal intake, and of 1.78 and 0.99 mg/kg bw, respectively, at the lower level of dietary inclusion.19 Considering the evidence from the King and McQueen (1981) study, that GAs are almost completely converted to aglycones in rumen, milk samples from dairy cows fed with spiked bovine feed ration were analysed by GC‐NPD for the presence of solanidine. No solanidine was detected in any milk sample (LOD: 0.14 mg/L).
3.1.1.4. Summary on toxicokinetics
Overall, limited conclusions on the toxicokinetics profiles of α‐solanine and α‐chaconine can be drawn based on the available evidence.
Limited information is available from toxicokinetic studies in experimental animals. Both α‐solanine and α‐chaconine showed a relatively low bioavailability in experimental animals. Different toxicokinetic profiles were observed in rats and hamsters. In hamsters, there is indication for higher absorption and slower excretion rates for both substances in comparison with rats. In particular in rats, following oral exposure to labelled α‐solanine or α‐chaconine, radioactivity was excreted mainly via the feces (about 60% in 24 h and more than 80% after 4 days) as solanidine. Urinary excretion accounted for about 3–10% of the doses for α‐solanine and 10% α‐chaconine. Conversely, in hamsters, a lower fecal excretion was observed, with urinary excretion reaching about 10% for α‐solanine and 21% for α‐chaconine 7 days after the exposure. Information on the metabolic profiles of α‐solanine and α‐chaconine were mainly obtained via TLC techniques and are of low reliability.
The blood kinetics profiles of α‐solanine and α‐chaconine in humans were relatively well studied in two volunteer studies, using HPLC methods and showing comparable results. In volunteers exposed to mixtures of α‐solanine and α‐chaconine in the range of 0.3–1.25 mg TGA/kg bw via consumption of mashed potato meals, peak levels in serum were reached approximately 6–8 h after exposure, independently from the doses. Both substances showed long serum half‐lives, suggesting a possible accumulation. α‐Chaconine showed longer serum half‐life than α‐solanine (44 and 21 h, respectively). In addition, in one study, the blood clearance of α‐solanine and α‐chaconine corresponded to the increase in solanidine levels, which reached a maximum level after 8 h and remained apparently constant up to 24 h following exposure. No further information on metabolism and no information on excretion was available in these studies. A volunteer study using tritiated solanidine support the evidence of slow clearance from plasma associated with a high retention of radioactivity in erythrocytes, with a corresponding slow excretion in urine and feces. In a separate experiment, solanidine was detected in three of five post‐mortem liver samples, either in the free form or in conjugated form (e.g. as glucuronide‐conjugate). Levels of solanidine were regularly detected in the blood of human volunteers in a series of studies using a radioimmuno assay for the detection of solanidine or TGAs. In particular, in one experiment, solanidine could be detected in serum of two volunteers up to 3 weeks after they started a potato‐free diet. Overall, these studies provide some limited information on the kinetics of potato GAs, mainly suggesting their hydrolysis to solanidine following absorption and the slow clearance of solanidine from blood.
Only scant information is available on the toxicokinetics profile of potato GAs in ruminants. In an in vitro study, α‐solanine and α‐chaconine were shown to be extensively converted to solanidine and subsequently to a 5,6‐dihydro derivative of solanidine, identified as 5β‐solanidan‐3β‐ol. Another study showed no detection of solanidine in cows’ milk upon consumption of feed spiked with a potato meal contaminated with α‐solanine or α‐chaconine, suggesting a low potential to transfer into milk for GA. No information was retrieved for other farm animals, horses or companion animals.
Toxicokinetic data on tomato and aubergine GAs and their aglycones could not be identified, neither for experimental animals nor for humans.
3.1.2. Toxicity in experimental animals
The selection criteria for studies relevant to inform the toxicity in experimental animals are described in Section 2.1.
In the descriptions below, the doses are reported as mg/kg bw per day. If the doses were reported on molar basis in the original study, these were converted using the molecular weights given in Appendix A. Due to the weight of the oligosaccharide in the GAs, the aglycones have a lower molecular weight by a factor of ~ 2. This should be considered when comparing the dose–response relationships of GAs and aglycones at doses given in mg/kg bw per day.
3.1.2.1. Acute toxicity studies
3.1.2.1.1. GAs from edible parts of S. tuberosum
Regarding acute oral toxicity of potato GAs (Table 6), a single oral application of 250 mg/kg bw of α‐solanine did not affect serum levels of liver transaminases or serum cholinesterase activity in rats (Dalvi, 1985). Mice showed no lethality at a dose of 1,000 mg/kg bw (Nishie et al., 1971).
Table 6.
Acute toxicity in experimental animals: oral administration of glycoalkaloids (GAs) (α‐solanine) or mixtures, from edible parts of S. tuberosum
| Test compounds | Species Dose route Doses | Observed effects | Highest dose with no effect (mg/kg bw) | Lowest dose with effect (mg/kg bw) | LD50 (mg/kg bw) | Reference |
|---|---|---|---|---|---|---|
| α‐Solaninea |
Sprague‐Dawley rats (M) No/sex/group: 5 Single gavage: 0, 20 or 250 mg/kg bw |
– | 250 | Dalvi (1985) | ||
| α‐Solaninea |
Albino mice (M) No/sex/group: n.r. Single oral application: 0 or 1,000 mg/kg bw |
– | 1,000 | Nishie et al. (1971) | ||
| Mixture of α‐solanine and unknown proportion of α‐chaconineb |
White rats (M/F) No/sex/group: 9 Single gavage: 300, 600 or 1,200 mg/kg bw |
Diarrhoea Lethality |
300 | 590 | Gull (1960) |
F: females. M: males. n.r.: not reported.
Chemical standard obtained from a commercial supplier, with high purity (95% pure or higher).
Poorly characterised chemical standard (e.g. purity or composition not defined).
An LD50 value of 590 mg/kg bw was identified for α‐solanine in male and female rats (Gull, 1960). However, it is uncertain whether pure α‐solanine was tested, as it appears likely that the standard (an extract of potato sprouts, purified by crystallisation) contained an unknown amount of α‐chaconine. Therefore, the study cannot be used for risk assessment. Diarrhoea was reported to be the most frequent symptom at lower doses of the extract.
When compared to oral toxicity, considerably lower doses of potato GA, aglycones or mixtures caused symptoms or were lethal when applied i.p. or i.v. (see Table 7). For example, LD50 values for α‐solanine ranged from 32.3 to 42 mg/kg bw in mice to 67 mg/kg bw in rats. Reported symptoms were diarrhoea and reduced spontaneous motor activity (Gull, 1960). The LD50 of α‐chaconine, applied i.p., appears to be similar, i.e. 19.2 or 32.3 mg/kg bw in mice and 84 mg/kg bw in rats (Nishie et al., 1975; Chaube and Swinyard, 1976; Sharma et al., 1979). Rabbits showed a similar sensitivity as rats and mice towards α‐solanine and α‐chaconine, i.e. lethality occurred when α‐solanine was applied at an i.v. dose of 10 mg/kg kg bw, and at i.p. doses of either 20 or 40 mg/kg bw or when animals received α‐chaconine at an i.p. dose of 50 mg/kg bw (Nishie et al., 1971, 1975). Swinyard and Chaube (1973) reported an LD50 value of < 40 mg of α‐solanine per kg bw for rhesus monkeys (administered i.p.).
Table 7.
Acute toxicity in experimental animals: non‐oral administration of glycoalkaloids (GAs) (α‐solanine, α‐chaconine), mixtures and the aglycone (solanidine), from edible parts of S. tuberosum
| Test compound | Species/dose route | Observed effects | Highest dose with no effect (mg/kg bw) | Lowest dose with effect (mg/kg bw) | LD50 (mg/kg bw) | Reference |
|---|---|---|---|---|---|---|
| α‐Solaninea | Rats (M/F)/i.p. | ↓ Hepatic glycogen content | 5 | Satoh (1967) | ||
| α‐Solaninea | Rats (M)/i.p. |
↑ Serum liver transaminases ↓ Serum cholinesterase activity |
20 | Dalvi (1985) | ||
| α‐Solanineb | Wistar rats (F)/i.p. | Lethality | 67 | Chaube and Swinyard (1976) | ||
| α‐Solaninea | Albino mice (M)/i.p. |
Lethality ↓ Spontaneous motor activity |
5 | 10 | 42 | Nishie et al. (1971) |
| α‐Solaninea | Balb/c mice (F)/i.p. | Lethality | 10 | 20 | 40 | Mohsenikia et al. (2013) |
| α‐Solaninea | Swiss‐Webster mice (M)/i.p. | Lethality | 32.3 | Patil et al. (1972) | ||
| α‐Solanineb | Swiss‐Webster mice (M)/i.p. | Lethality | 32.3 | Sharma et al. (1979) | ||
| α‐Solaninea | Swiss‐Webster mice (M)/i.p. | Lethality | 34.5 | Nishie et al. (1975) | ||
| α‐Solaninea | Balb‐c mice (M)/i.p. | 40 | Khodayari et al. (2013) | |||
| α‐Solaninea | Rabbits (sex n.r.)/i.p. or i.v. |
EEG alterations (i.p.) ↑ Respiratory rate, ↓ blood pressure (i.v.) Lethality (i.p.) Lethality (i.v.) |
< 15 |
> 15 2 20 10 |
Nishie et al. (1971) | |
| α‐Solaninea | White New Zealand rabbits (sex n.r.)/i.p. | Lethality | 40 | Nishie et al. (1975) | ||
| α‐Solaninea | Rhesus monkey (sex n.r.)/i.p. | Lethality (100%) |
40 2 × 20 |
< 40 | Swinyard and Chaube (1973) | |
| α‐Chaconineb | Swiss‐Webster mice (M)/i.p. | Lethality | 32.3 | Nishie et al. (1975) | ||
| α‐Chaconinea | Swiss‐Webster mice (M)/i.p. | Lethality | 19.2 | Sharma et al. (1979) | ||
| α‐Chaconineb | Wistar rats (F)/i.p. | Lethality | 84 | Chaube and Swinyard (1976) | ||
| α‐Chaconineb | Sprague‐Dawley rats (M)/i.p. |
Altered EEG of occipital cortex Bradycardia |
10 |
10 20 |
Aldous et al. (1980) | |
| α‐Chaconineb | White New Zealand rabbits (sex n.r.)/i.p. | Lethality | 50 | Nishie et al. (1975) | ||
| Mixture of α‐solanine and unknown proportion of α‐chaconinec | Rats (M/F)/i.p. | 75 | Gull (1960) | |||
|
Mixturea α‐solanine: α‐chaconine (1:1) |
Syrian hamster (M)/i.p. | Lethality | 10 | 25 | > 10 < 25 | Phillips et al. (1996) |
|
Mixtured α‐solanine, α‐chaconine and ß‐chaconine comprised ~ 20% of the extract at a ratio of 1:2:0.3, respectively |
Wistar rats (F)/i.p. | 60 | Chaube and Swinyard (1976) | |||
| Solanidinea | Albino mice (M)/i.p. | – | 500 | Nishie et al. (1971) |
EEG: electroencephalogram; F: female; M: male; n.r.: not reported.
Chemical standard obtained from a commercial supplier, with high purity (95% pure or higher).
Chemical standard obtained by in‐house isolation from plant material, and checked for purity (95% pure or higher).
Poorly characterised chemical standard (e. g. purity or composition not defined).
A non‐ or only partly purified extract or material for which the glycoalkaloid composition and concentration is given.
Regarding single i.p. application of defined mixtures of α‐solanine and α/ß‐chaconine, Syrian hamsters were relatively sensitive showing an LD50 value between 10 and 25 mg/kg bw, while in rats, the LD50 value was 60 or 75 mg/kg bw (Gull, 1960; Chaube and Swinyard, 1976; Phillips et al., 1996).
The aglycone solanidine was tested in mice only at a single i.p. dose of 500 mg/kg bw, which exerted no lethal effects (Nishie et al., 1971).
3.1.2.1.2. GAs from edible parts of food plants other than S. tuberosum
The studies identified are reported in Tables 8 and 9.
Table 8.
Acute toxicity in experimental animals: oral administration of glycoalkaloids (GAs) (α‐tomatine) from edible parts of S. lycopersicum and extracts from edible parts of S. melongena
| Substance (purity) | Species (sex) Dose route Doses | Observed effects | Highest dose with no effect (mg/kg bw) | Lowest dose with effect (mg/kg bw) | LD50 (mg/kg bw) | Reference |
|---|---|---|---|---|---|---|
| α‐Tomatinea |
Rats (M/F) No/sex/group: n.r. Oral dose of 900–1,000 mg/kg bw given within 24 h |
100% lethality | 900–1,000 | Wilson et al. (1961) | ||
| Water suspensions of ground lyophilised fruits of ripe S. melongena b |
Syrian hamster (sex n.r.) No/sex/group: 2 Single oral application: 6,316 mg suspension/kg bw TGA content of dried matter: 0.016% (corresponding to 1.0 mg TGA/kg bw) |
Haemorrhagic gastritis | 1 | Baker et al. (1989) |
F: females; M: males; n.r.: not reported.
Chemical standard obtained by in‐house isolation from plant material, may contain α‐dehydrotomatine as an impurity.
A non‐ or only partly purified extract or material for which the GA composition and concentration is given.
Table 9.
Acute toxicity in experimental animals: non‐oral administration of glycoalkaloids (GAs) (α‐tomatine, α‐solamargine) from edible parts of S. lycopersicum or S. melongena
| Substance (purity) | Species (sex) Dose route | Observed effects | Highest dose with no effect (mg/kg bw) | Lowest dose with effect (mg/kg bw) | LD50 (mg/kg bw) | Reference |
|---|---|---|---|---|---|---|
| α‐Tomatineb | Mice (sex n.r) i.v. | Lethality | 18 | Wilson et al. (1961) | ||
| α‐Tomatinea | Swiss‐Webster mice (M) i.p. | Lethality | 32.4 | Nishie et al. (1975) | ||
| α‐Tomatineb | Mice (sex n.r) i.p. | Lethality | 25 | Sackmann et al. (1959) | ||
| α‐Tomatineb | Mice (sex n.r) s.c. | Lethality | > 1,000 | Sackmann et al. (1959) | ||
| α‐Tomatinea | White New Zealand rabbits (sex n.r.) i.p. | – | 100 | Nishie et al. (1975) | ||
| α‐Solamarginec | Wistar rats (M) i.p. | Lethality | 42 | Al Chami et al. (2003) |
M: male. n.r.: not reported.
Chemical standard obtained from a commercial supplier, may contain α‐dehydrotomatine as an impurity.
Chemical standard obtained by in‐house isolation from plant material, may contain α‐dehydrotomatine as an impurity.
Poorly characterised chemical standard (e. g. purity or composition not defined).
Wilson et al. (1961) reported on 100% lethality in rats after oral application of 900–1,000 mg/kg bw of α‐tomatine to rats. Histologic analyses revealed an acute mucosal erosion of the stomach associated with a mild acute inflammation in the submucosa.
Regarding mixtures, Baker et al. (1989) treated hamsters by a single oral application of water suspensions of lyophilised seeds of aubergine containing 160 mg TGA/kg dry weight (dw). The authors reported that solasodine was the only aglycone detectable. A dose of 6,316 mg suspension/kg bw (approximately 1 mg TGA/kg bw) caused haemorrhagic gastritis in hamsters.
When compared to oral toxicity, considerably lower doses of tomato GAs were lethal when applied i.p. or i.v. The LD50 values for α‐tomatine in mice ranged from 18 mg/kg bw (i.v.) to 32.4 mg/kg bw (i.p.) (Sackmann et al., 1959; Wilson et al., 1961; Nishie et al., 1975). After s.c. administration, the LD50 was raised to > 1,000 mg/kg bw (Sackmann et al., 1959). Rabbits appear to be less sensitive than mice, since a single i.p. dose of α‐tomatine at 100 mg/kg bw did not cause death of the animals (Nishie et al., 1975).
The LD50 value of α‐solamargine was determined to be 42 mg/kg bw, when applied i.p. to rats (Al Chami et al., 2003).
3.1.2.1.3. Summary on acute toxicity studies
Regarding oral exposure to α‐solanine, an LD50 of 590 mg/kg bw was identified for rats, while mice showed no lethality at a dose of 1,000 mg/kg bw. When compared to gavage, considerably lower i.p. doses elicited effects, e.g. in mice, the LD50 was 40–42 mg/kg i.p., and in rats, increased liver transaminases in serum were reported at 20 mg/kg bw i.p.
Relative potency of toxicity of different GAs can be deduced after i.p. application and appear to be similar, i.e. in rats, the LD50 was 67 mg/kg bw for α‐solanine and 84 mg/kg bw for α‐chaconine, and in mice, it was 32.3–42 mg/kg bw for α‐solanine and 19–32.3 mg/kg bw for α‐chaconine. Lethality occurred in New Zealand rabbits starting at 40 mg/kg bw of α‐solanine and at 50 mg/kg bw for α‐chaconine. In mice, the aglycone solanidine exerted no effect when applied at 500 mg/kg bw i.p., contrasting to the pronounced toxicity of α‐solanine and α‐chaconine after the identical route of exposure.
Lethal doses of α‐tomatine were approximately 900–1,000 mg/kg bw p.o. in rats. Acute lesions comprised erosion, and also inflammation occurred in the stomach. When applied i.v. or i.p., LD50 values of 18–32.4 mg/kg bw were found in rodents. Again, the parenteral route was found to increase the toxicity of GAs from tomato.
3.1.2.2. Repeated dose toxicity studies
The studies on effects following repeated oral doses are summarised in Tables 10, 11, 12 and 13. The no‐observed‐effect level (NOEL) (or lowest‐observed‐effect level, LOEL) and NOAEL (LOAEL) values indicated, have been derived by the CONTAM Panel, unless stated otherwise.
Table 10.
Repeated dose oral toxicity studies with glycoalkaloids (GAs) (α‐solanine and α‐chaconine) and the aglycone (solanidine) from edible parts of S. tuberosum
| Test compound | Species/Experimental design and doses | Most sensitive endpoint | Highest dose with no effect (mg/kg bw per day) | Lowest dose with effect (mg/kg bw per day) | Reference |
|---|---|---|---|---|---|
| α‐Solanineb |
Swiss‐Webster mice (F) No/sex/group: 8 Diet: 0 or 2.4 mmol/kg diet (equivalent to 0 or 416.6 mg/kg bw per dayd) Duration: 7 days |
↓ bw gain ↓ absolute liver weight |
416.6 416.6 |
Friedman et al. (1996) | |
| α‐Solaninec |
White rats (M) No/sex/dose: 5–10 Diet: 0, 20, 50, 100, 250, 500, 1,000, 2,000 mg/kg diet (equivalent to 0, 1.8, 4.5, 9, 22.5, 45, 90, or 180 mg/kg bw per dayd) Duration: 5 weeks |
↓ growth rate | 90 | 180 | Gull (1960) |
| α‐Solaninea |
Golden Syrian hamster (F) No/sex/group: 4–8 Gavage: 0 or 100 mg/kg bw per day Duration: 5 days |
Lethality (50% of animals) ↓ food intake and bw gain ↓ relative liver weight ↑ RBC, HGB, HCT ↑ serum glucose, creatine ↓ serum alk phosphatase, albumin |
100 100 100 100 100 100 |
Langkilde et al. (2008) For mixtures of α‐solanine and α‐chaconine, see Table 12 |
|
| α‐Chaconineb |
Swiss‐Webster mice (F) No/sex/group: 8 Diet: 0 or 2.4 mmol/kg diet (equivalent to 0 or 409 mg/kg bw per dayd) Duration: 7 days |
↓ bw gain (day 7) ↓ absolute and relative liver weight (day 7) |
409 409 |
Friedman et al. (1996) | |
| α‐Chaconinea |
Golden Syrian hamster (F) No/sex/group: 4–8 Gavage: 0 or 100 mg/kg bw per day Duration: 5 days |
Lethality ↓ bw gain ↓ relative liver weight ↑ RBC, HGB, HCT ↑ serum glucose, creatine |
0 |
100 100 100 100 100 |
Langkilde et al. (2008) For mixtures of α‐solanine and α‐chaconine, see Table 12 |
| Solanidinea |
Swiss‐Webster mice (F) No/sex/group: 8–16 Diet: 0, 1.2, 2.4 or 4.8 mmol/kg diet (equivalent to 0, 95.5, 190.9 or 381.8 mg/kg bw per dayd) Duration: 7, 14 or 28 days |
↓ food intake (day 14) ↓ bw gain (day 7) ↓ bw gain (day 14) ↑ absolute and relative liver weight (day 7) ↑ absolute and relative liver weight (day 14, 28) |
190.9 (no 95.5 group) 190.9 (no 95.5 group) 95.5 |
381.8 190.9 381.8 190.9 190.9 |
Friedman et al. (1996) |
| Solanidinea |
Swiss‐Webster mice (F) No/sex/group: 10 Diet: 0 or 2.4 mmol/kg diet (equivalent to 0 or 190.9 mg/kg bw per dayd) Duration: 14 days |
↓ bw gain ↑ absolute and relative liver weight |
190.9 190.9 |
Friedman et al. (2003a) |
F: female; M: male; bw: body weight; RBC: red blood cell; HGB: haemoglobin; HTC: haematocrit.
Chemical standard obtained from a commercial supplier, with high purity (95% pure or higher).
Chemical standard obtained by in‐house isolation from plant material, and checked for purity (95% pure or higher).
Poorly characterised chemical standard (e.g. purity or composition not defined).
Applying EFSA default values.
Table 11.
Repeated dose oral toxicity studies with defined mixtures of glycoalkaloids (GAs) (α‐solanine and α‐chaconine) from edible parts of S. tuberosum
| Test compound | Species Dose route Doses | Observed effects | Highest dose with no effect GA (α‐solanine/α‐chaconine) (mg/kg bw per day) | Lowest dose with effect GA (α‐solanine/α‐chaconine) (mg/kg bw per day) | Reference |
|---|---|---|---|---|---|
| Potatoes containing 6.4 mg TGA (20 mg α‐solanine; 37 mg α‐chaconine) per kg fw, admixed as powder to dietb |
Swiss‐Webster mice (sex n.r) No/sex/group: 4–5 Oral application (diet): 0, 1, 5, 10, 20 or 40% potato powder in the diet with approximately 0, 2.9, 14.6, 29.1, 58.2 or 116.4 mg TGA/kg diete (equivalent to 0, 0.6, 5.8, 11.6 and 23.2 mg TGA/kg bw per dayd) Duration: 7 and 14 days |
↑ absolute and relative pancreas weight | ~ 11.6 (3.63/6.71) | ~ 23.2 (7.25/13.4) | Friedman (1992) |
| α‐Solanine:α‐chaconine (1:1)a |
Syrian hamster (M) No/sex/group: 2 Gavage: 0, 5, 10, 25 or 50 mg GA/kg bw per day Duration: 3 days |
No effect | 50 (25/25) | Phillips et al. (1996) | |
| α‐Solanine:α‐chaconine (1:2.5)a |
Golden Syrian hamster d No/sex/group: 4–8 Gavage: 0, 75 or 100 mg/kg bw per day Duration: 5 days |
Lethality (in 25%) ↓ food intake ↓ bw gain ↓ relative liver weight ↑ relative adrenal weight ↑ HGB, HCT ↑ RBC ↑ serum creatine |
75 (21.4/53.6) 75 (21.4/53.6) 75 (21.4/53.6) 75 (21.4/53.6) |
75 (21.4/53.6) 75 (21.4/53.6) 100 (28.6/71.4) 75 (21.4/53.6) 100 (28.6/71.4) 75 (21.4/53.6) 100 (28.6/71.4) 100 (28.6/71.4) |
Langkilde et al. (2008) For single compound controls, see Table 11 |
| α‐Solanine:α‐chaconine (1:3.7)a |
Golden Syrian hamster d No/sex/group: 4 Gavage: 0, 0.1, 0.33, 1, 3.3, 10 or 33.3 mg/kg bw per day Duration: 28 days |
↑WBC and RBC ↓albumin, urea and plasma proteins ↑serum AST Distended stomach and intestine |
10f (2.1/7.9) 10f (2.1/7.9) 10f (2.1/7.9) 10f (2.1/7.9) |
33 (7/26) 33 (7/26) 33 (7/26) 33 (7/26) |
Langkilde et al. (2009) |
| α‐Solanine:α‐chaconine (1:70)a |
Golden Syrian hamster d No/sex/group: 4 Gavage: 0, 0.1, 0.33, 1, 3.3, 10 or 33.3 mg/kg bw per day Duration: 28 days |
Mortality (25%) ↑WBC and RBC ↓albumin, urea and plasma proteins ↑serum AST Distended stomach and intestine |
10 (0.1/9.9) 10f (0.1/9.9) 10f (0.1/9.9) 10f (0.1/9.9) 10f (0.1/9.9) |
33.3 (0.5/32.8) 33.3 (0.5/32.8) 33.3 (0.5/32.8) 33.3 (0.5/32.8) 33.3 (0.5/32.8) |
Langkilde et al. (2009) |
|
α‐Solanine:α‐chaconine (1:3.1) S. tuberosum Desiree (added to chow as freeze‐dried powder)b, c |
Golden Syrian hamster d No/sex/group: 10 Diet: TGA at 0, 6.9, 13.8, 20.6 or 33d mg/kg bw per day Duration: 90 days |
No effects | 33 (8/25) | Langkilde et al. (2012) | |
|
α‐Solanine:α‐chaconine (1:25.5‐26) S. tuberosum SGT9‐2 (added to chow as freeze‐dried powder) and spiked with GA from commercial sourceb |
Golden Syrian hamster d No/sex/group: 10 Diet: TGA at 0, 6.9, 13.8 or 20.6 mg/kg bw per day Duration: 90 days |
No effects | 20.6 (1.0/19.6) | Langkilde et al. (2012) | |
| α‐Solanine:α‐chaconine (1:25.5–26) from S. tuberosum SGT 9‐2 (added to chow as freeze‐dried powder)b |
Golden Syrian hamster d No/sex/group: 10 Diet: TGA at 0 or 8.6 mg/kg bw per day Duration: 90 days |
No effects | 8.6 (0.3/8.5) | Langkilde et al. (2012) |
F: females; M: males; n.r.: not reported; TGA: total GA content; AST: aspartate aminotransferase; WBC: white blood cells; RBC: red blood cells; HCT: haematocrit; HBG: haemoglobin.
Chemical standard obtained from a commercial supplier, with high purity (95% pure or higher).
Chemical standard obtained by in‐house isolation from plant material, and checked for purity (95% pure or higher).
Poorly characterised chemical standard (e. g. purity or composition not defined) or with a non‐ or only partly purified extract or material for which the GA composition and concentration is given.
A non‐ or only partly purified extract or material for which the GA composition and concentration is given.
Spiked with GAs from commercial source.
Applying EFSA default values
Anticipating 78% water content of potatoes, the GA content would be 291 mg TGA, 91 mg α‐solanine and 168 mg α‐chaconine per kg potato powder.
Approximative values read from the figures of the study.
Table 12.
Repeated dose oral toxicity studies with edible plant parts of S. tuberosum and sprouts
| Test compound | Species Experimental design | Observed effects | Highest dose with no effect (mg/kg bw per day) | Lowest dose with effect (mg/kg bw per day) | Reference |
|---|---|---|---|---|---|
| Sprouts of S. tuberosum (dried) admixed to chow as powder; no trypsin inhibitor activitya |
Wistar rats (M) No/sex/group: 6 Diets: GA at 0, 60, 120, 180, 240 or 300 mg/kg (equivalent to: 0, 7.2, 14.4, 21.6, 28.8 or 36 mg/kg bw per dayb) Duration: 28 days |
No effects | 36 | Tuśnio et al. (2013a) | |
| Protein concentrate of S. tuberosum, admixed to chow as powder; with trypsin inhibitor activitya |
Wistar rats (M) No/sex/group: 6 Diet: GA at 0, 116, 125, 164, 215, 331 or 439 mg/kg diet (equivalent to 0, 13.9, 15, 19.7, 25.8, 39.7 or 52.7 mg/kg bw per dayb) Duration: 24 days |
↓relative weight of stomach ↑relative weight of intestine ↑serum ALT |
25.8 39.7 |
13.9 39.7 52.7 |
Tuśnio et al. (2013a) |
|
S. tuberosum; ripe or green as chowa 287.6 mg TGA/kg fw of green potato; 82.5 mg TGA/kg fw of normal potato |
Rabbits (sex n.r.) No/group: 10 Diet: Dose of TGA: 21.5 mg (ripe) or 64.2 mg (green)/kg diet; Equivalent to 21.6 and 82.3 mg/kg bw per daye Equivalent to 4.8 and 14 mg/kg bw per dayc NO CONTROL Duration: 30 days |
↓bw and relative heart weight ↑relative liver weight ↑plasma glucose and Ca2+ concentration ↓Na+, K+ and protein concentration in plasma |
82.3d 82.3d 82.3d 82.3d |
Azim et al. (1982) | |
|
S. tuberosum; ripe or green as chowa 203.8 mg TGA/kg of green potato; 75 mg TGA/kg TGA in normal potato |
Rabbits (sex n.r.) No/group: 4 Diet: Dose of GA: 14–25 or 48–59 mg/kg bw per day NO CONTROL Duration: 20 days |
↓bw ↑fecal protein excretion ↓protein digestibility |
48–59d 48–59d 48–59d |
Azim et al. (1983) | |
| S. tuberosum; ripe or green as chowa |
Rabbits (sex n.r.) No/group: 5 Diet: Dose of TGA: 16.1–17.9 mg (ripe) or 75–75.4 mg (green)/kg bw per day NO CONTROL Duration: 45 days |
↓RBC counts and HGB concentration | 75.2d | Azim et al. (1984) | |
| Strain B5141‐6, diced and admixed to the dieta |
Rhesus monkey, mated but not pregnant (F) No/sex/group: 4 Diet: 3.47 mg TGA/kg bw per day NO CONTROL Duration: 25 days |
No effects | 3.47 (0.87 α‐solanine) | Swinyard and Chaube (1973) |
F: female; M: male; bw: body weight; n.r.: not reported; RBC: red blood cells; HGB: haemoglobin.
A non‐ or only partly purified extract or material for which the GA composition and concentration is given.
Applying EFSA default values.
For the exposure estimates, a live weight of 2 kg and a daily feed intake of 75 g/kg bw were assumed (derived from Carabano and Piquer, 1998).
Significant when compared to lower dose group.
Table 13.
Repeated dose oral toxicity studies with glycoalkaloids (GAs) (α‐tomatine, α‐solasonine) and aglycones (tomatidine, solasodine) from edible parts of S. lycopersicum or S. melongena
| Test compound | Species Experimental design Doses | Observed effects | Highest dose with no effect (mg/kg bw per day) | Lowest dose with effect (mg/kg bw per day) | Reference |
|---|---|---|---|---|---|
| α‐Tomatine (given as hydrochloride)d |
Rats (M) No/sex/group: 5 Gavage of 250 mg/kg bw per day Duration: 5 days |
250 | Wilson et al. (1961) | ||
| α‐Tomatine (given as hydrochloride)d |
Albino rats d No/sex/group: 5 Diet: 0.0025–0.04% (equivalent to 1.25–20 mg/kg bw per daye) Duration: 200 days |
20 | Wilson et al. (1961) | ||
| α‐Tomatinec |
Albino rats (M) No/sex/group: 6 Diet: 0, 0.5 or 2% (equivalent to 0, 600 or 2,400 mg/kg bw per daye) Diets were supplied with 1% cholesterol Duration: 7 days |
↓ intrahepatic cholesterol | 600 | Cayen (1971) | |
| α‐Tomatinec |
Albino rats (M) No/sex/group: 6–9 Diet: 0 or 1% (equivalent to 0 or 1,200 mg/kg bw per daye) Duration: 14 days |
↓ uptake of dietary cholesterol by liver ↑ hepatic and intestinal cholesterol synthesis ↑ fecal sterol excretion without affecting bile acid excretion ↓ serum HDL, LDL and triglyceride levels |
1,200 1,200 1,200 1,200 |
Cayen (1971) | |
| α‐Tomatinec |
Swiss‐Webster mice f No/sex/group: 8 Diet: 0 or 2.4 mmol/kg diet (equivalent to 0 or 496 mg/kg bw per daye) Duration: 7 days |
No effect on bw or liver weight | 496 | Friedman et al. (1996) | |
| α‐Tomatinec |
Golden hamster (M) No/sex/group: 6–8 Animals kept on a high‐fat and high‐cholesterol diet Diet: 0, 0.05, 0.1 or 0.2% in diet (equivalent to 0, 30.5, 61 or 122 mg/kg bw per dayf) Duration: 21 days |
↓ LDL cholesterol ↓ sum of VLDL‐, LDL‐ and HDL‐cholesterol ↓ triglyceride ↑ fecal sterol and coprostanol excretion |
61 61 61 |
122 122 122 30.5 |
Friedman et al. (2000a) |
| α‐Solasoninea |
Swiss‐Webster mice f No/sex/group: 8 Diet: 0 or 2.4 mmol/kg diet (equivalent to 0 or 424.4 mg/kg bw per daye) Duration: 7 days |
↑ bw gain | 424.4 | Friedman et al. (1996) | |
| Tomatidineb |
Rats (M) No/sex/group: n.r. Diet: 0.0025–0.04% (equivalent to 1.25–20 mg/kg bw per daye Duration: 200 days |
20 | Wilson et al. (1961) | ||
| Tomatidinec |
Swiss‐Webster mice f No/sex/group: 8 Diet: 0, 1.2, 2.4 or 4.8 mmol/kg diet (equivalent to 0, 99.8, 199.5 or 399 mg/kg bw per daye) Duration: 7, 14 or 28 days |
↓ food intake (day 7) ↑ absolute and relative liver weight (day 7) ↑ absolute and relative liver weight (day 14, 28) |
199.5 (no value for 99.8) 199.5 |
399 199.5 399 |
Friedman et al. (1996) |
| Tomatidinea |
Swiss‐Webster mice f No/sex/group: 14 Diet: 0 or 2.4 mmol/kg diet (equivalent to 199.5 mg/kg bw per dayd) Duration: 14 days |
↓ bw gain ↑ relative liver weight |
199.5 199.5 |
Friedman et al. (2003a) | |
| Solasodinea |
Swiss‐Webster mice (n.r.) No/sex/group: 4–5 Diet: 0, 50, 100, 200, 400, 800 or 1,600 mg/kg diet (equivalent to 10, 20, 40, 80, 160 or 320 mg/kg bw per daye) Duration: 7 and 14 days |
↓ bw gain (day 7 and 14) Gastric gland degeneration Cholangiohepatitis ↑ relative liver weight ↑ serum GOT, GPT, bilirubin |
80 80 80 80 80 |
160 160 160 160 160 |
Friedman (1992) |
| Solasodinea |
Swiss‐Webster mice f No/sex/group: 8–16 Diet: 0, 1.2, 2.4 or 4.8 mmol/kg diet (equivalent to 0, 99.3, 198.6 or 397.2 mg/kg bw per daye) Duration: 7, 14 or 28 days |
↓ food intake (day 7) ↑ absolute and relative liver weight (day 7, 14, 28) |
198.6 99.3 |
397.2 198.6 |
Friedman et al. (1996) |
| Solasodinea |
Swiss‐Webster mice f No/sex/group: 24 Diet: 0 or 2.4 mmol/kg diet (equivalent to 0 or 198.6 mg/kg bw per daye) Duration: 14 days |
↓ bw gain ↑ absolute and relative liver weight |
198.6 198.6 |
Friedman et al. (2003a) |
F: female; M: male; n.r.: not reported; bw: body weight; HDL: high‐density lipoprotein; LDL: low‐density lipoprotein; VLDL: very low‐density lipoprotein; GOT: glutamic oxaloacetic transaminase; GPT: glutamic pyruvic transaminase.
Chemical standard obtained from a commercial supplier, with high purity (95% pure or higher).
Chemical standard obtained by in‐house isolation from plant material, and checked for purity (95% pure or higher).
Chemical standard obtained from a commercial supplier, may contain α‐dehydrotomatine as an impurity.
Chemical standard obtained by in‐house isolation from plant material, may contain α‐dehydrotomatine as an impurity.
Applying EFSA default values.
A mean body weight of 130 g and daily consumption of 8 g per animal were assumed; taken from Gattermann et al. (2002).
3.1.2.2.1. GAs and aglycones from edible parts of S. tuberosum
α‐Solanine
α‐Solanine reduced body weight gain and the absolute liver weight when given at 416.6 mg/kg bw per day (the only dose administered) for 1 week to Swiss Webster mice (Friedman et al., 1996). The food intake remained unaffected.
A lower dose (180 mg α‐solanine/kg bw per day) reduced the growth rate in rats treated over a period of 5 weeks which was not due to an altered food intake of the animals. No further compound‐related symptoms were reported (Gull, 1960). Based on the lower body weight at 180 mg/kg bw per day, a NOAEL of 90 mg α‐solanine/kg bw per day was derived.
Hamsters exhibited lethality in 50% of the animals and reduced growth and relative liver weights when exposed daily to 100 mg α‐solanine/kg bw (the only dose administered) for 5 days. The lowered body weight gain might have been caused by a decreased food intake by the animals. Serum glucose, serum creatine, red blood cell (RBC), haemoglobin and haematocrit were elevated (Langkilde et al., 2008).
α‐Chaconine
α‐Chaconine lowered body and liver weights at 409 mg/kg bw per day (the only dose administered) in mice over a period of 7 days (Friedman et al., 1996). The food intake by the animals was unaffected.
In hamsters, 100 mg/kg bw per day (the only dose administered) for 5 days induced lethality (in 25% of the animals) and reduced body weight gain and relative liver weights despite an unaltered food intake (Langkilde et al., 2008). Serum glucose, serum creatine, RBC, haemoglobin and haematocrit were elevated.
Solanidine
The aglycone solanidine was administered to mice for up to 28 days at doses ranging from 95.5 to 381.8 mg/kg bw per day (Friedman et al., 1996, 2003a). Adverse effects were seen at 190.9 mg/kg bw per day, such as decreased body weight gain and increased absolute and relative liver weights (Friedman et al., 1996). Feed intake was lowered at a daily dose of 381.8 mg/kg bw. Based on increases in absolute and relative liver weights at day 14 and 28 of treatment, a NOAEL of 95.5 mg solanidine/kg bw per day was derived.
Mixtures of GAs
Friedman (1992) fed mice potatoes containing 64 mg TGAs (20 mg α‐solanine and 37 mg α‐chaconine)/kg fresh weight. α‐Solanine and α‐chaconine were quantified by HPLC‐UV, while TGA were determined by a colourimetric method (possibly detecting other GAs as well, see Section 1.3.2). The potatoes were admixed as powder at high concentrations (up to 40%) to the diet presumably causing dietary imbalances of macro‐ and micronutrients. Increased absolute and relative weights of pancreas were observed at the highest TGA dose of 23.2 mg/kg bw per day. The CONTAM Panel agrees with the assumption of the authors that the effects on the pancreas may be due to presence of trypsin inhibitor activity in the potato powder and not caused by GAs. Consequently, no NOAEL was derived for this study.
For a period of 3 days, Phillips et al. (1996) treated male hamsters orally with doses up to 50 mg TGA/kg bw per day at a ratio of α‐solanine: α‐chaconine of approximately 1:1. No adverse effects could be observed. However, the study was conducted with a low number of animals per treatment group.
In a series of experiments, Langkilde et al. (2008, 2009, 2012) tested defined mixtures of α‐solanine and α‐chaconine in female hamsters. At a ratio α‐solanine:α‐chaconine of 1:2.5, gavage of 75 mg TGA/kg bw per day induced lethality (in 25% of the animals), lowered food intake, decreased the relative liver weight and increased haemoglobin and haematocrit within 5 days of treatment (Langkilde et al., 2008). At gavage of 100 mg TGA/kg bw per day, lethality was seen again in 25% of the animals, body weight gain was lowered and the relative weight of adrenals, serum creatine and RBC were elevated (Langkilde et al., 2008). The LOAEL in this study was 75 mg TGAs/kg bw per day based on the lethality and the decreased relative liver weight in female hamsters.
An α‐solanine:α‐chaconine ratio of 1:3.7 with a dose of 33 mg TGA/kg bw, applied by daily gavage, raised white blood cells (WBC), RBC and serum aspartate aminotransferase (AST), and decreased albumin, proteins and urea in blood within 4 weeks (Langkilde et al., 2009). When the ratio α‐solanine:α‐chaconine was 1:70 at a dose of 33.3 mg TGAs/kg bw per day, death occurred in one of the four treated animals 2 h after gavage. Weight of adrenal glands, AST, WBC and RBC were affected (Langkilde et al., 2009). Thus, the NOAEL was 10 mg TGAs/kg bw per day irrespective of whether α‐solanine and α‐chaconine were applied at a 1:3.7 or 1:70 ratio.
Langkilde et al. (2012) investigated the putative effects of a wild‐type (‘Desiree’) and a genetically modified potato line (SGT 9‐2). While the wild‐type potato contained α‐solanine:α‐chaconine at a ratio of 1:3.1, the SGT 9‐2 line exhibited a ratio of 1:25.5–26 and a considerably lower TGA content (determined by HPLC‐UV). All animals of the study received diets, which were identical in the concentrations of minerals, vitamins, sucrose/dextrin, soybean oil and cellulose but contained 0%, 20%, 40% or 60% freeze‐dried powder of control or SGT 9‐2 potatoes (trypsin inhibitor activity of the powders was not excluded). The variable proportions of potato powder in the diets were compensated to some extent by adjusted contents in caseinate and corn/potato starch. Since the TGA content of the SGT 9‐2 line was considerably lower than in the controls, commercially available α‐solanine and α‐chaconine were added to the diets with SGT 9‐2 in order to treat animals with TGA doses as occurring in control potatoes. However, the ratio between the two GAs was kept as being typical for the SGT 9‐2 line. In addition, one of the diets (40% powder of control potatoes) was spiked with commercially available α‐solanine and α‐chaconine to elevate the TGA dose to 33 mg/kg bw per day. When the relative proportion of α‐solanine:α‐chaconine was 1:3.1, a daily TGA dose of 33 mg/kg bw exerted no effect over a period of 90 days. At a ratio of 1:25.5–26 again no effects were seen at TGA doses of up to 20.6 mg/kg bw per day (Langkilde et al., 2012). In any case, the NOEL in this study was 33 mg TGA/kg bw per day at a ratio α‐solanine:α‐chaconine of 1:3.1 (the highest dose tested).
In contrast to data published in 2009, Langkilde et al. (2012) did not find adverse effects when hamsters were exposed to 33 mg TGAs/kg bw per day for 90 days with a ratio of α‐solanine:α‐chaconine of approximately 1:3.1. In this study, the NOEL for TGAs was 33 mg/kg bw per day which is considerably higher than in the study appearing in 2009. The different outcome may be based on the fact, that in the study of 2009 purified standards were applied to the animals as bolus by gavage, whereas in the study published in 2012, freeze‐dried potato powder, partly spiked with commercial standards, were administered via the diet.
Preparations based on edible parts of S. tuberosum and sprouts
In some experiments, the effects of plant parts of potatoes were studied. Tuśnio et al. (2013a) used potato sprout meal with 41.85 g TGAs/kg admixed to the diet to provide daily doses of up to 36 mg TGAs/kg bw to rats (no information on the ratio of α‐solanine:α‐chaconine provided). The potato sprout meal did not exhibit any trypsin inhibitor activity. This treatment did not elicit any effect when applied over a period of 28 days (Tuśnio et al., 2013a). Thus, the NOEL was 36 mg TGAs/kg bw per day (the highest dose tested).
In a second experiment, commercial potato protein concentrates were applied, which contained 677–3,185 mg TGAs/kg (Tuśnio et al., 2013a) and which did exhibit trypsin inhibitor activity. Again, the ratio of α‐solanine:α‐chaconine was not given and might have been variable. TGA doses of 13.9–52.7 mg/kg bw per day affected the gastrointestinal tract and caused decreased relative stomach weight, increased relative weight of the intestine and elevated serum alanine aminotransferase (ALT) levels. The study authors speculated that these alterations were rather due to trypsin inhibitor activity than the GA content in the diet (Tuśnio et al., 2013a). Consequently, no NOAEL or LOAEL could be derived.
Azim and colleagues performed three studies in rabbits and compared the effects of green or ripe potatoes, fed as chow (Azim et al., 1982, 1983, 1984). In all studies, the GA content in potatoes was determined leading to estimated intakes between 4.8 and 82.3 mg/kg bw per day, but no ratio of α‐solanine:α‐chaconine was given. Furthermore, the studies lack negative controls. Generally, decreased body weight and increased relative liver weights were reported. However, the data are difficult to interpret since untreated controls are missing. No NOEL or LOEL could be derived.
Swinyard and Chaube (1973) used potato strain B5141‐6, containing a mean TGA content of 260 mg/kg fw (range: 150–350 mg/kg fw). α‐Solanine accounted for 25% of the TGA content, while the remaining GAs were not specified, but probably contained α‐chaconine. The tubers were diced and admixed to the diet. By this procedure, Rhesus monkeys were treated with approximately 3.47 mg TGAs/kg bw per day for 25 days, which did not elicit any significant effect on food consumption and body weight. However, the study lacks an untreated control group. Therefore, no NOAEL could be derived.
Summary of repeated dose toxicity studies with GAs and aglycones from edible parts of S. tuberosum and sprouts
To conclude, α‐solanine and α‐chaconine reduced body and liver weights in mice after 1 week at 416.6 and 409 mg/kg bw per day, respectively. A NOAEL was set for α‐solanine at 90 mg/kg bw per day based on reduced growth of rats, treated for 5 weeks. In hamster, body and relative liver weights were lowered by 100 mg/kg bw of either α‐solanine or α‐chaconine, applied for 5 days. The aglycone solanidine caused adverse effects in mice treated with 190.9 mg/kg bw per day up to 4 weeks; the NOAEL was 95.5 mg/kg bw per day.
Mixtures of α‐solanine and α‐chaconine were applied to hamsters over 90 days (Langkilde et al., 2012). The NOEL was 20.6 mg/kg bw per day (α‐solanine:α‐chaconine ratio of 1:25.5–26) and 33 mg/kg bw per day (α‐solanine:α‐chaconine ratio of 1:3.1) (the highest doses tested). The NOEL of 33 mg/kg bw per day is identical to the LOAEL reported in another study, in which 33 mg/kg (α‐solanine:α‐chaconine ratio of 1:3.7), applied via gavage for 4 weeks, caused gastrointestinal effects in hamsters (Langkilde et al., 2009). The different outcome at the identical dose may be due to the bolus effect of gavage in the 2009 study and the matrix effects of the basal chow in the 2012 study.
Regarding edible parts of potato and sprouts, the NOEL was 36 mg TGA/kg bw per day, fed to rats for 28 days (the highest dose tested). No effects were seen in monkeys at a daily dose of 3.47 mg TGA/kg bw, applied for 25 days.
No data with regard to the repeated dose oral toxicity of ß‐ or y‐forms of potato GAs could be identified.
3.1.2.2.2. GAs and aglycones from edible parts of food plants other than S. tuberosum
The studies identified are reported in Table 13.
α‐Tomatine
α‐Tomatine was tested in rats at doses of 1.25–250 mg/kg bw per day over periods of 5–200 days. No effects were reported at 250 mg/kg bw per day when given for 5 days, and at 20 mg/kg bw applied over a period of 200 days (Wilson et al., 1961). A NOAEL of 20 mg/kg bw per day for rats was identified from this study (the highest dose applied in this study). A dose of 600 mg/kg bw caused decreased intrahepatic cholesterol after 7 days of administration (Cayen, 1971). Based on the lower hepatic cholesterol concentration, the LOEL was 600 mg α‐tomatine/kg bw per day in male rats. When the effects on the cholesterol metabolism were studied in rats in more detail, Cayen (1971) reported on reduced uptake of dietary cholesterol by the liver, an elevated hepatic and intestinal cholesterol synthesis and increased fecal sterol excretion without affecting bile acid excretion at 1,200 mg α‐tomatine/kg bw per day for 2 weeks (see also Section 3.1.2.9).
In mice, 496 mg α‐tomatine/kg bw per day (the only dose administered) for 1 week did not affect body or liver weights (Friedman et al., 1996).
When hamsters were treated with 30.5 mg/kg bw per day for 3 weeks, the LOEL was 30.5 mg α‐tomatine/kg bw per day, based on the elevated fecal sterol and coprostanol excretion (Friedman et al., 2000a) (see also Section 3.1.2.9).
α‐Solasonine
α‐Solasonine increased the body weight gain in mice at 424.4 mg/kg bw per day (the only dose administered) for 7 days despite an unaltered food intake by the animals. No further effects were reported (Friedman et al., 1996).
Tomatidine
Wilson et al. (1961) obtained the aglycone tomatidine by hydrolysis, as described by Fontaine et al. (1948). When given to rats daily at 1.25–20 mg/kg bw for 200 days, tomatidine exerted no significant effects (Wilson et al., 1961). The NOEL in this study was 20 mg tomatidine/kg bw per day for rats (the highest dose tested). Two studies in mice reported an elevated absolute and/or relative liver weight at 199.5 mg/kg bw per day, when applied for 7 or 14 days (Friedman et al., 1996, 2003a). Further observations were decreased feed intake (day 7 at 399 mg/kg bw per day) and reduced body weight gain (day 14 at 199.5 mg/kg bw per day) (Friedman et al., 1996, 2003a). The LOAEL was 199.5 mg/kg bw per day, based on the reduced body weight gain and elevated absolute and relative liver weight in female mice.
Solasodine
The aglycone solasodine was tested in Swiss‐Webster mice at doses of up to 397.2 mg/kg bw per day (Friedman, 1992; Friedman et al., 1996, 2003a). In the first study, Friedman (1992) reported on decreased body weight gain (despite unaltered food intake), gastric gland degeneration, cholangiohepatitis, elevated relative liver weight and increased serum glutamic oxaloacetic transaminase (GOT), glutamic pyruvic transaminase (GPT) and bilirubin, when animals received 160 mg/kg bw per day for 7 and/or 14 days. This indicates that the gastrointestinal tract and the liver were affected severely by this compound. The NOAEL was 80 mg/kg bw per day, based on the effects on body weight and on the gastrointestinal tract at 160 mg/kg bw per day in mice. In the two subsequent studies, the same study authors administered partly higher doses than in the study published in 1992 and focused on compound effects on body and liver weights (Friedman et al., 1996, 2003a). Decreased body weight gain (despite unaltered food intake) and increased relative and/or absolute liver weights were induced at 198.6 mg solasodine/kg bw per day (Friedman et al., 2003a). At 397.2 mg/kg bw per day, feed intake was reduced at day 7 of treatment (Friedman et al., 1996). Due to the high level and the limited dose ranges applied, these studies were not used to identify a NOAEL.
Friedman et al. (1996) compared the relative potencies of equimolar doses of the aglycones with regard to the increase in liver weight in mice and found that solanidine, solasodine and tomatidine were nearly equal (see also Table 10). This was confirmed by Friedman et al. (2003a). In contrast, equimolar doses of the GA α‐solasonine decreased insignificantly the liver weight (Friedman et al., 1996). Wilson et al. (1961) applied α‐tomatine and tomatidine at 20 mg/kg bw per day for a period of 200 days to rats. When considering the different molecular weights of the compounds, the α‐tomatine dose was 20 μmol/kg bw per day and the tomatidine dose 50 μmol/kg bw per day. Nevertheless, tomatidine did not elicit any effect, as was true also for α‐tomatine.
Summary of repeated dose toxicity studies with GAs and aglycones from edible parts of food plants other than S . tuberosum
To conclude, α‐tomatine exerted no effects in mice receiving 496 mg/kg bw per day for 1 week. A NOEL of 20 mg/kg bw per day (the highest dose tested) was identified for rats, treated for 200 days. In hamsters, the LOEL was 30.5 mg/kg bw per day, based on an increased fecal sterol and coprostanol excretion. α‐Solasonine increased the body weight gain in mice when treated daily with 424.4 mg/kg bw for 1 week (the only dose applied). For tomatidine, a LOAEL of 199.5 mg/kg bw per day was derived in mice, and a NOEL of 20 mg/kg bw per day (the highest dose applied) in rats. For solasodine, a NOAEL of 80 mg/kg bw per day was identified in mice, based on decreased body weight gain, gastric gland degeneration and liver toxicity at higher doses.
3.1.2.3. Developmental and reproductive toxicity studies
3.1.2.3.1. Developmental effects
In 1972, Renwick published a hypothesis that there might be a causal relationship between anencephaly and spina bifida and maternal consumption of blighted potatoes. He further proposed that this may be preventable by avoiding specific, at the time unidentified, compounds from potato tubers (Renwick, 1972).
3.1.2.3.1.1. GAs from edible parts of S. tuberosum
The studies identified are described below and details given in Table 14.
Table 14.
Developmental toxicity studies on glycoalkaloids (GAs) (α‐solanine, α‐chaconine) and aglycones (solanidine, demissidine) from edible parts of S. tuberosum
| Test compound | Species, experimental design and doses | Observed effects | Highest dose with no effect (mg/kg bw per day) | Lowest dose with effect (mg/kg bw per day) | Reference | ||
|---|---|---|---|---|---|---|---|
| Mother | Offspring | Mother | Offspring | ||||
| α‐Solanine | |||||||
| α‐Solaninea |
Wistar rats Gavage Exposure at different periods during gestation, sacrifice on day 22 (I) Exposure during GD6–15: 0, 0.3, 1, 3 mg/kg bw per day (9 or 10 litters) (II) Exposure during GD7–10: 0, 6 mg/kg bw per day (9 litters) (III) Exposure during GD8–11: 0, 2, 10, 25 mg/kg bw per day (2, 4, 3, 4 litters, respectively) |
all exposure groups: No significant effects | No effects | Ruddick et al. (1974) | |||
| α‐Solanineb (purified from sprouts of the British cultivar Arran Pilot) |
Syrian hamsters Number of dams not specified, 85 litters and 964 fetuses in control, 55 litters and 573 fetuses in α‐solanine group Gavage on GD8 200 mg/kg bw per day Sacrifice on GD15 |
maternal death (3 dams) 59% malformed littersf 23% CNS malformed fetuses (gavage GD8) |
200 |
200 200 |
Renwick et al. (1984) | ||
| α‐Solanineb |
Pregnant syrian hamsters No/group: 24 Gavage on GD8 0.28 mmol/kg bw (equivalent to 243 mg/kg bw per day) Sacrifice on GD15 |
21% abnormal littersf 5.5% abnormal fetuses (exencephaly, encephalocele, anophthalmia) ↑ maternal fatality |
243 243 243 |
Gaffield and Keeler (1996a) | |||
| α‐Chaconine | |||||||
| α‐Chaconined |
Wistar rats Gavage on GD6–15 Exposure GD6–15: 1.5 mg/kg bw per day (4 litters) Sacrifice on day 22 |
No statistically significant effect | Ruddick et al. (1974) | ||||
|
α‐Chaconineb Purified from sprouts of the British cultivar Arran Pilot |
Syrian hamsters Number of dams not specified, 85 litters and 964 fetuses in control, 60 litters and 645 fetuses in the α‐chaconine groups, no specified how many animals per dosage group Gavage on GD8 only 0, 165, 180 mg/kg bw per day |
maternal death (4 of 6 dams) 63% malformed littersf 23% CNS malformed fetuses |
165 |
165 165 |
Renwick et al. (1984) | ||
| α‐Chaconineb |
Pregnant Syrian hamsters No/Group: 15 and 1 litter Gavage on GD8 0.21, 0.29 mmol/kg bw (equivalent to 179 and 247 mg/kg bw) Sacrifice on GD15 |
maternal lethality (3/4) 20% abnormal littersf 2.7% abnormal fetuses (exencephaly and encephalocele) |
247 |
179 179 |
Gaffield and Keeler (1996a) | ||
| Solanidine | |||||||
| Solanidine N‐oxidec |
Pregnant Syrian hamsters No/group: 3–14 litters Gavage on GD8 0.14, 0.29, 0.43 mmol/kg bw (equivalent to 58, 120, 178 mg/kg) Sacrifice on GD15 |
50% abnormal littersf 16% abnormal fetuses (exencephaly and encephalocele) |
120 120 |
178 178 |
Gaffield and Keeler (1996a) | ||
| Solanidinec |
Pregnant Syrian hamsters No/group: 12 litters Gavage on GD8 0.44 mmol/kg bw (or 176 mg/kg bw)e Sacrifice on GD15 |
50% abnormal littersg 24% abnormal fetuses (exencephaly, encephalocele, anophthalmia, external viscera) |
176 176 |
Gaffield and Keeler (1993) | |||
| Solanidineb |
Pregnant Syrian hamsters No/group: 21 litters Gavage on GD8 0.29 mmol/kg bw (equivalent to 115 mg/kg bw) Sacrifice on GD15 |
19% abnormal littersf 2.3% abnormal fetuses (encephalocele and anophthalmia) |
115 115 |
Gaffield and Keeler (1996a) | |||
| Solanidinea |
Swiss Webster mice (time pregnant/time mated (not clear)) No/group: 20 pregnant females Diet. Exposure: Start GD4, Necroscopy GD17–18 2.4 mmol/kg diet (equivalent to 159 mg/kg bw per day) |
↓maternal feed intake ↑maternal relative liver weight ↓litter size ↓litter weight ↓litter weight/body weight ↓average fetus weight |
159 159 159 |
159 159 159 |
Friedman et al. (2003a) | ||
| Demissidine | |||||||
| Demissidinec |
Pregnant Syrian hamsters No/group: 14 litters Gavage on GD8 0.44 mmol/kg bw (or 176 mg/kg bw)e Sacrifice on GD15 |
14% abnormal littersg 3% abnormal fetuses (both values not statistically significant) |
176 176 |
Gaffield and Keeler (1993) | |||
| Demissidinec |
Pregnant Syrian hamsters No/group: 14 litters Gavage on GD8 0.44 mmol/kg bw (equivalent to 176 mg/kg bw) Sacrifice on GD15 |
14% abnormal littersf 3% abnormal fetuses (both values not statistically significant) |
176 176 |
Gaffield and Keeler (1996a) | |||
GD: gestational day; MW: molecular weight (in g/mol); CNS: central nervous system.
Chemical standard obtained from a commercial supplier, with high purity (95% pure or higher).
Chemical standard obtained by in‐house isolation from plant material, and checked for purity (95% pure or higher).
Chemical standard obtained by in‐house synthesis, and checked for purity (95% pure or higher).
Poorly characterised chemical standard (e.g. purity or composition not defined).
Doses as reported by the authors in the original publication.
Percentage of litters containing at least one malformed fetus.
No definition given on whether this refers to the litters or to individuals in the litter.
Blighted potatoes
After Renwick (1972) had raised the hypothesis above, a number of studies were published using material from blighted potatoes for exposure (Poswillo et al., 1972, 1973a,b; Chaube et al., 1973; Allen et al., 1977; Sharma et al., 1978). The common limitation of these studies is that no reference to the GA content of the test material is given. The studies were therefore not used for further evaluation.
α‐Solanine
Ruddick et al. (1974) performed a teratology study for α‐solanine and α‐chaconine in Wistar rats. They performed a comparative study with uncharacterised raw material and purified GAs. The three experimental arms investigating the teratogenic potential of α‐solanine included an experiment in which first nine litters each were examined from primipari mothers, which were exposed by gavage during gestational day (GD) 6–15 at doses of 0.3, 1 and 3 mg/kg bw per day. In the second experiment, nine litters from mothers, exposed with 6 mg/kg bw per day during GD7–10 were examined, and in the third experimental arm 3–4 primipari pregnant Wistar rats were treated by gavage with 2, 10 and 25 mg α‐solanine/kg bw per day. Three–four litters were examined in this experiment. In each experiment, animals only treated with the vehicle of GAs served as control. At GD22, females were sacrificed and the following parameters were evaluated: corpora lutea, resorption sites, litter size and weight, as well as potential gross visceral and skeletal anomalies. One out of 117 offspring animals of the 3 mg/kg bw per day group showed craniorachischisis and exopthalmos, and one out of 108 of the 0.3 mg/kg bw per day group showed twisted pelvic limbs and absent tail. The authors did not judge these effects as treatment related. Besides the effects detectable in these two animals, no evidence for teratogenicity was observed.
Renwick et al. (1984) purified α‐solanine and α‐chaconine from sprout material of the British cultivar ‘Arran Pilot’. They performed a comparative oral gavage study in pregnant Syrian hamsters with poorly characterised extract material and with pure compounds. For the latter, pregnant dams were exposed by gavage on GD8 to α‐solanine at a dose of 200 mg/kg bw per day. On day 15, the dams were sacrificed and necropsied. Maternal death was observed for three dams out of 38 dams (estimated from the three reported maternal deaths, the one resorbed litter and the 34 live litters), whereas no dam died in the control group. Examination of fetuses revealed 59% of litters with malformed fetuses. A total of 35 cases of cranial blebs and 39 cases of exencephaly were identified, which amounted to 23% of fetuses with malformation associated with the development of brain structures. Only one fetus in more than 300 fetuses investigated in the control group showed this phenotype.
Gaffield and Keeler (1996a) performed a comparative study investigating the potential teratogenic effects of GAs and some of their aglycones. Pregnant female Syrian hamsters were gavaged on GD8 with α‐solanine at a dose of 243 mg/kg bw per day. Dams were sacrificed and necropsied on GD15. The dose of 243 mg/kg bw per day of α‐solanine produced 21% of abnormal litters and 5.5% of abnormal fetuses. The rate of resorption was increased. Abnormalities were exencephaly, encephalocoele and anophthalmia.
α‐Chaconine
Ruddick et al. (1974) performed a teratology study for α‐solanine and α‐chaconine in Wistar rats. They performed a comparative study with uncharacterised raw material and purified GAs. In the experimental arms investigating the teratogenic potential of α‐chaconine (in‐house prepared standard) four litters from primiparity mothers were investigated following treatment of mothers by gavage during GD6–15 with 1.5 mg α‐chaconine/kg bw per day. Vehicle treated animals served as a control. At GD22, females were sacrificed and the following parameters were evaluated: corpora lutea, resorption sites, litter size and weight, as well as potential gross visceral and skeletal anomalies. No evidence for teratogenicity was observed in this experiment.
In the above‐mentioned study by Renwick et al. (1984), pregnant Syrian hamsters dams (number of animals not explicitly mentioned) were gavaged with α‐chaconine at a dose of 165 and 180 mg/kg bw per day on GD8. On day 15, the dams were sacrificed and necropsied. Maternal death was observed for 4 out of 24 dams in the 165 mg/kg bw per day group (estimated from the four reported maternal deaths, the one resorbed litter and the 19 live litters) and for 6 out of 15 dams in the 180 mg/kg bw per day group (estimated from the six reported maternal deaths, the one resorbed litter and the eight live litters), whereas no dam died in the control group (one resorbed and 37 live litters reported). Examination of fetuses revealed 27 cases of cranial blebs and 19 cases of exencephaly in the 165 mg/kg bw per day exposure group, and 10 cases of cranial blebs and four cases of exencephaly in the higher dose group, which amounts to 23% and 18% of fetuses, respectively, with malformation in the central nervous system. Overall, the percentage of malformed litters as of live litters was lower in the 165 mg/kg bw per day dose group (63%) than in the 180 mg/kg bw per day dose group. Only one fetus in more than 300 fetuses investigated in the control group showed this phenotype.
In the Gaffield and Keeler (1996a) study, pregnant female Syrian hamsters were gavaged with α‐chaconine at a dose of 179 or 247 mg/kg bw per day on GD8. Dams were sacrificed and necropsied on GD15. The dose of 247 mg/kg bw per day of α‐chaconine (n = 1 L) resulted in an abnormal litter. The dose of 179 mg/kg bw per day of α‐chaconine produced 20% of abnormal litters and 2.7% of abnormal fetuses. The rate of resorption was increased. Abnormalities were encephalocoele and anophthalmia.
Solanidine N‐oxide
In the Gaffield and Keeler (1996a) study, pregnant female Syrian hamsters were gavaged with solanidine N‐oxide at doses of 58, 120 or 178 mg/kg bw per day on GD8. Solanidine N‐oxide was reported by the authors as a putative metabolite of solanidine. Dams were sacrificed and necropsied on GD15. The dose of 178 mg/kg bw per day of solanidine N‐oxide produced 50% of abnormal litters and 16% of abnormal fetuses. The rate of resorption was increased. The two other doses were without effects. Abnormalities were exencephaly and encephalocoele.
Solanidine
Gaffield and Keeler (1993) performed a comparative study based on the hypothesis of C5, C6 unsaturation being a key structural factor in steroidal alkaloid‐induced mammalian teratogenesis. Pregnant female Syrian hamsters were gavaged with solanidine at a dose of 176 mg/kg bw per day on GD8. Dams were sacrificed and necropsied on GD15. The dose of 176 mg/kg bw per day produced 50% of abnormal litters and 24% of abnormal fetuses. Abnormalities were exencephaly, encephalocoele, anophthalmia and external viscera.
Gaffield and Keeler (1996a) studied solanidine again. Pregnant female Syrian hamsters were gavaged with solanidine at a dose of 115 mg/kg bw per day on GD8 and dams were sacrificed and necropsied on GD15. The dose of 115 mg/kg bw per day produced 19% of abnormal litters and 2.3% of abnormal fetuses. Abnormalities were encephalocoele and anophthalmia.
Potential teratogenic effects of solanidine were also investigated by Friedman et al. (2003a). For the experiment the authors purchased time pregnant Swiss Webster mice, many of which apparently were not pregnant. Following acclimatisation, exposure to experimental chow was started on GD4 and terminated either at GD17 or GD18. Exposure to 205 mg solanidine/kg bw per day resulted in reduced maternal body weight, probably caused by reduced maternal feed intake. In maternal animals, an increased maternal relative liver weight was observed in addition. For the offspring, the authors identified reduction of litter size, litter weight, litter weight/body weight and average fetus weight in response to treatment.
Demissidine
In an earlier study by Gaffield and Keeler (1993), pregnant female Syrian hamsters were gavaged with demissidine at a dose of 176 mg/kg bw per day on GD8, and this dose produced 14% of abnormal litters and 3% of abnormal fetuses. Both effects did not reach the level of statistical significance. Abnormalities were not specified.
In a later study by Gaffield and Keeler (1996a), pregnant female Syrian hamsters were gavaged with demissidine at a dose of 176 mg/kg bw per day on GD8. The dose of 176 mg/kg bw per day produced 14% of abnormal litters and 3% of abnormal fetuses. Abnormalities were exencephaly, encephalocoele, cebocephaly, cleft palate and anophthalmia.
3.1.2.3.1.2. GAs from edible parts of food plants other than S. tuberosum
The studies identified are described below and details given in Table 15.
Table 15.
Developmental toxicity studies on glycoalkaloids (GAs) (α‐tomatine) and aglycones (tomatidine, solasodine, dihydrosolasodine) from edible parts of food plants other than S. tuberosum
| Test compound | Species, experimental design and doses | Observed effects | Highest dose with no effect (mg/kg bw per day) | Lowest dose with effect (mg/kg bw per day) | Reference | ||
|---|---|---|---|---|---|---|---|
| Mother | Offspring | Mother | Offspring | ||||
| α‐Tomatine | |||||||
| α‐Tomatined |
Pregnant Simonsen hamsters No/groups: 34 dams Gavage on GD8 0, 530 mg/kg bw per day Sacrifice on GD15 |
Maternal death 6% deformed littersg from surviving dams |
530 | 530 | Keeler et al. (1990) | ||
| Tomatidine | |||||||
| Tomatidinec,e |
Pregnant Syrian hamsters No/groups: 15–24 pregnant dams Gavage on GD8 0, 1,184, 2,664 mg/kg bw per day Sacrifice on GD15 |
No effects | 2,664 | 2,664 | Keeler et al. (1976) | ||
| Tomatidinee |
Swiss Webster time pregnant/time mated (not clear) mice No/group: 23 pregnant mice Diet. Exposure GD4–18 2.4 mmol/kg diet (equivalent to 167 mg/kg bw per day) |
↓maternal bw ↑maternal relative liver weight |
167 | Friedman et al. (2003a) | |||
| Solasodine | |||||||
| Solasodinec |
Pregnant Syrian hamsters No/group: 11–50 dams per group 0, 1,184, 1,480 and 1,628 mg/kg bw per day gavage on GD7, sacrifice on GD15 0, 1,184 and 1,480 mg/kg bw on GD8, sacrifice on GD15 |
GD7 exposure: Maternal death 25% abnormal littersh 5% abnormal offspring GD8 exposure: Maternal death 36% abnormal littersh 6% abnormal offspring |
1,184 1,184 |
1,184 1,184 1,184 1,184 |
Keeler et al. (1976) | ||
| Solasodinec |
Pregnant Syrian hamsters No/group: 8 litters Gavage on GD8 3.4 mmol/kg bw (or 1,400 mg/kg bw)f Sacrifice on GD15 |
75% abnormal littersh 29% abnormal fetuses (encephalocele, exencephaly, anophthalmia) |
1,400 1,400 |
Gaffield and Keeler (1993) | |||
| Solasodinea |
Swiss Webster time pregnant/time mated (not clear) mice No/group: 45 pregnant females Diet. Exposure GD4–18 2.4 mmol/kg diet (equivalent to 156 mg/kg bw per day) |
↓ maternal bw ↓ maternal feed intake ↑ maternal relative liver weight ↓ litter weight ↓ litter weight/bw ↓ average fetus weight |
156 156 156 |
156 156 156 |
Friedman et al. (2003a) | ||
| Dihydrosolasodine | |||||||
| Dihydrosolasodineb (Soladulcidine) |
Pregnant Syrian hamsters No/group: 11 litters Gavage on GD8 3.4 mmol/kg bw (1,400 mg/kg bwf) Sacrifice on GD15 |
45% abnormal littersh 6% abnormal fetuses (encephalocele, cebocephaly, exencephaly, anophthalmia) |
1,400 1,400 |
Gaffield and Keeler (1993) | |||
GD: gestational day; MW: molecular weight (in g/mol); bw: body weight.
Chemical standard obtained from a commercial supplier of high purity (95% pure or higher).
Chemical standard obtained by in‐house synthesis, and checked for purity (95% pure or higher).
Chemical standard obtained from a commercial supplier, purity not stated.
Chemical standard obtained from a commercial supplier, may contain dehydrotomatine as an impurity.
Chemical standard obtained from a commercial supplier, may contain dehydrotomatidine as an impurity.
Doses as reported by the authors in the original publication.
Percentage of litters containing at least one malformed fetus.
No definition given on whether this refers to the litters or to individuals in the litter.
α‐Tomatine
In 1990, Keeler et al. (1990) published a review on existing data on congenital craniofacial malformation following exposure to spirosolane‐containing solanum species. To this review, some original data were added, among them, on α‐tomatine. Pregnant Simonsen hamsters were gavaged with 530 mg/kg bw per day of α‐tomatine on GD8. Nineteen dams died from overdose symptoms largely related gastrointestinal tract lesions. From surviving dams, a percentage of 6% deformed litters was recorded.
Tomatidine
Keeler et al. (1976) investigated potential teratogenic effects of tomatidine and exposed pregnant Syrian hamsters on GD8 with 1,184 or with 2,664 mg/kg bw per day of tomatidine by gavage. Neither abnormal litters nor abnormal offspring, which was used as readout, was reported.
In Swiss Webster mice, potential teratogenic effects of tomatidine were investigated by Friedman et al. (2003a) following the same scheme as that described for solanidine above. Exposure to 167 mg/kg bw per day of tomatidine resulted in reduced maternal body weight and increased maternal relative liver weight in response to treatment. No effects were detectable which affected offspring animals.
Solasodine
Keeler et al. (1976) comparatively studied the consequences of solasodine exposure at GD7 (1,184, 1,480 and 1,628 mg/kg bw per day) and at GD8 (1,184 and 1,480 mg/kg bw per day) in pregnant female Syrian hamsters (n = 115). The authors reported that a total of 10 dams distributed over all dose groups died from overdose symptoms, except for the 1,480 mg/kg bw per day dose group if exposed on GD7. For the remaining dams, the major effect was abnormal litters, which did not follow a clear dose dependency if exposed at GD7 (25–29%). Following exposure on GD8, the percentage of abnormal litters increased dose dependently in a statistically significant manner from 36% (1,184 mg/kg bw per day) to 44% (1,480 mg/kg bw per day). The most often detected phenotypes in abnormal offspring comprised spina bifida, exencephaly and cranial bleb. The percentage of abnormal offspring was 5–7% if exposure occurred on GD7, and 6–15% following exposure on GD8.
In the study by Gaffield and Keeler (1993), pregnant female Syrian hamsters were gavaged with solasodine at a dose of 1,400 mg/kg bw per day on GD8, that dose produced 75% of abnormal litters and 29% of abnormal fetuses. Abnormalities were encephalocoele, exencephaly and anophthalmia.
Using Swiss Webster mice, Friedman et al. (2003a) reported that exposure to 195 mg solasodine/kg bw per day resulted in reduced maternal body weight, probably caused by reduced maternal feed intake. In maternal animals, an increased maternal relative liver weight was observed in addition. For the offspring authors identified reduction of litter size, litter weight, litter weight/body weight and average fetus weight in response to treatment.
Dihydrosolasodine (soladulcidine)
Gaffield and Keeler (1993) also studied the effects of dihydrosolasodine gavaged to pregnant female Syrian hamsters on GD8 at a dose of 1,400 mg/kg bw per day. That dose produced 45% of abnormal litters and 6% of abnormal fetuses. Abnormalities were encephalocoele, exencephaly, cebocephaly and anophthalmia.
3.1.2.3.1.3. Comparative evaluation of studies
To make use of the studies in which single doses have been used, studies are evaluated comparatively regarding relative potency of compounds. This approach represents rather a qualitative than a quantitative assessment. Only hamsters, for which the widest sets of data are available, have been used. Further, in the comparative studies, authors used similar molar exposure levels. However, these exposure scenarios are mostly only identical within a study but not across studies. Further, in order to be able to compare potency, endpoints which have been investigated across studies have to be selected. Finally, a reference point for comparison would be helpful, but is not available.
Keeler et al. (1990) were the first to take the attempt to order GAs according to their teratogenic potency. They took the same database and some additional data to establish a relative potency list of GAs (Gaffield and Keeler, 1996b). This list contained data on GAs from Brown and Keeler (1978), Keeler et al. (1976), Renwick et al. (1984), Keeler et al. (1990), Gaffield and Keeler (1993) as well as Gaffield and Keeler (1996a). In this assessment, the authors aimed to compare relative teratogenic potencies of jervine, solanidine and spirosolane alkaloids. Relative potencies can only be compared by their induction of terata if a molar equivalent basis of exposure is applied (see Appendix D) or if effects are allocated to an ascending exposure level in mg/kg bw. The data which were consistently reported across studies were percentage of abnormal litters and percentage of abnormal fetuses. As can be seen from Table 1D. (Appendix D), a clear relative potency pattern cannot be delineated. It can also be seen that there are huge differences between values from different laboratories, e.g. for α‐chaconine and α‐solanine, if results from the studies of Renwick et al. (1984) and from Gaffield and Keeler (1996a) are compared (see Appendix D). Further, the relative order of potency reported by Gaffield and Keeler (1996b) may only be used for a qualitative comparison, according to the authors. However, α‐chaconine may be misplaced in this order, because at dosage 0.29 mmol/kg maternal lethality became apparent, not allowing to follow up the fate of the offspring.
3.1.2.3.1.4. Summary on developmental effects
The experimental developmental studies indicate that effects occur at oral doses of 165 mg/kg bw per day and above for GAs, and of 115 mg/kg bw per day and above for the aglycones. Among others, these effects to a large majority were observed in the central nervous system and most frequently comprised exencephaly, encephalocele and anophthalmia.
In comparative single dose assessment studies, the results suggest differences in the potency of GAs and their aglycones. However, due to limitations in the studies available (e.g. lack of dose–response study design), an order of potency cannot be established.
3.1.2.3.2. Reproductive effects
Kline et al. (1961) investigated the effects of α‐solanine on the survival of rat pups (see Table 16). They exposed pregnant female Holtzman rats (duration not given, starting exposure following detection of pregnancy by weighing until delivery) via the diet to either commercial α‐solanine at 3.6 or 4.8 mg/kg bw per day or to α‐solanine, isolated from the potato sprouts (purity unknown) at 3.6 mg/kg bw per day. Compared to controls, all treatment procedures produced a decrease in the percentage of weaned pups and a comparable increase in the number of zero litters (litters in which all pups died). No dose dependency was seen for the commercial α‐solanine. However, the effects were more pronounced when applying the isolated α‐solanine (23.6% weaned pups compared to controls) than the commercial α‐solanine (37.5% for both dosages, compared to controls). Starvation may have contributed to the early death of the pups as no milk was found in the stomach of these animals, pointing to a lactational deficit. Cross fostering studies were performed, but results were inconclusive. Adult males were investigated as controls and no noticeable effects were detectable (Kline et al., 1961).
Table 16.
Reproductive toxicity studies on glycoalkaloids (GAs) (α‐solanine) from edible parts of S. tuberosum
| Test compound | Species Experimental design Doses | Observed effects | Highest dose with no effect (mg/kg bw per day) | Lowest dose with effect (mg/kg bw per day) | Reference | ||
|---|---|---|---|---|---|---|---|
| Mother | Offspring | Mother | Offspring | ||||
| α‐Solanine | |||||||
|
α‐Solanine (commercial)a α‐Solanine (isolated from sprouts)a |
Female pregnant Holtzman rats N = 10/4 litters per group for testing of commercial/isolated α‐solanine Duration of exposure not clear (assumption that the study duration was 14 days) Diet: 0, 30, 40 mg α‐solanine (commercial) per kg chow (0, ≈ 3.6, ≈ 4.8 mg/kg bw per day) Diet: 0, 30 mg α‐solanine (isolated) per kg chow (≈ 3.6 mg/kg bw per day) |
↑ number of litters in which pups died within the first 3 days ↓ percentage of weaned pups (37.5% weaned pups, compared to controls) ↓ percentage of weaned pups (23.6% weaned pups, compared to controls) |
3.6 mg/kg bw per day (for commercial and isolated α‐solanine) 3.6 mg/kg bw per day (for commercial α‐solanine) 3.6 mg/kg bw per day (for isolated α‐solanine) |
Kline et al. (1961) | |||
Poorly characterised chemical standard (purity and composition not defined, may have contained α‐chaconine).
Effects of solasodine on fertility were also investigated in male Rhesus monkeys (Dixit et al., 1989). Ten Rhesus monkeys were divided into two groups of five animals each, which were either treated for 150 days by placebo or 100 mg/kg bw per day of solasodine encapsulated into a banana (see Table 17). On day 151, testes and epididymis were surgically removed. Sperm count and motility were assessed in the cauda epididymis. Pathological parameters were assessed from tissue section of paraffin‐embedded material. Biochemical parameters were measured from frozen tissue samples. Interstitial testicular cells, like Leydig cells and their precursors were counted. 100 mg/kg bw per day reduced testis weight, weight of the epididymis, the diameter of the seminiferous tubule and the Leydig cell nuclear diameter. In terms of fertility parameters, the authors detected a reduced number of spermatids, spermatozoa, immature and mature Leydig cells. The number of degenerating Leydig cells was found to be unaltered. Analysis of the cauda of the epididymis revealed a dramatic reduction of sperm density and sperm motility. On the biochemical level total testicular protein content, content of sialic acid and of glycogen were reduced, whereas testicular testosterone levels were elevated. The CONTAM Panel notes that treatment of Rhesus monkeys with 100 mg/kg bw per day of solasodine encapsulated into a banana affected testicular function and male reproduction.
Table 17.
Reproductive toxicity studies on aglycones (solasodine) from edible parts of S. melongena
| Test compound | Species Experimental design Doses | Observed effects | Lowest dose with effect (mg/kg bw per day) | Reference |
|---|---|---|---|---|
| Solasodine | ||||
| Solasodinea |
Captured Rhesus monkey (no further specifications) 0, 100 mg/kg bw per day for 150 days Oral administration by encapsulation into a banana On day 151 surgical removal of testes and epididymis |
↓ testes weight ↓ epididymal weight ↓ seminiferous tubule diameter ↓ Leydig cell nuclei diameter ↓ spermatid count ↓ spermatozoa ↓ immature and mature Leydig cells ↓ sperm density and motility in cauda epididymis ↓ total protein, sialic acid, glycogen in testes ↑ cholesterol in testes |
100 100 100 100 100 100 100 100 100 100 |
Dixit et al. (1989) |
|
Solasodinea Isolated from berries of S. xanthocarpum |
Dogs (M, 12–17 kg, 5 dogs per group) 0, 80 mg/kg bw per day Oral in a mutton meet pellet Duration: 30 days |
↓ epididymal weight ↓ cauda epididymal epithelial height ↓ total protein, sialic acid, glycogen in the cauda epididymis ↑ cholesterol in the cauda epididymis |
80 80 80 80 |
Gupta and Dixit (2002) |
M: males.
Chemical standard obtained by in‐house isolation from plant material, highly purified.
The impact of solasodine following oral application on male fertility was investigated in dogs (Gupta and Dixit, 2002). Five dogs were treated with 80 mg/kg bw per day of solasodine for 30 days with the compound embedded in a capsule of mutton meet (see Table 18). In this study, only effects on the epididymis were investigated. The treatment did not affect the body weight of the animals but decreased the epididymal weight and cauda epididymal epithelial height. The epididymal lumen was found to be empty of sperm. On a biochemical level, the total protein content, content of sialic acid and of glycogen were found to be reduced in the epididymis in response to 80 mg/kg bw per day of solasodine. Epidydimal levels of cholesterol were found to be elevated. In conclusion, oral treatment of dogs with 80 mg/kg bw per day of solasodine impacts on the integrity and the function of the epididymal gland in male dogs. The epididymal findings in this study using oral dosage are in line with a previous study which investigated effects of solasodine on male reproductive organs following i.p. application of 20 mg/kg bw per day for 30 days (Dixit and Gupta, 1982).
Table 18.
Tests for genotoxicity/mutagenicity of potato glycoalkaloids (GAs) (α‐solanine, α‐chaconine) and the aglycone (solanidine)
| Test compound (Source) | Test system | Cells/animals | Concentration/Treatment | Result | Comment | Reference |
|---|---|---|---|---|---|---|
| α‐Solaninea | Reverse mutation assay | S. Typhimurium TA98 and 100 +/−S9 | 10–50 μg/plate | Negative | No positive control | Ness et al. (1984) |
| α‐Solaninea | Reverse mutation assay |
S. Typhimurium TA98, TA100 +/−S9 |
0.07–2.3 μmol/platec |
Negative Positive controls: expected results |
Friedman and Henika (1992) | |
| α‐Solaninea | Micronucleus assay (fetal blood cells) | Swiss‐Webster mice (F) | Single i.p. application at GD15–16: 0.01, 0.02, 0.045 mmol/kg bwd (equivalent to 8.7, 17.4 or 39.1 mg/kg bw) |
Negative Positive controls: expected results |
Number of fetuses per dose group: 54–55 | Friedman and Henika (1992) |
| α‐Solaninea | Micronucleus assay (peripheral blood cells) |
Swiss‐Webster mice (M) No/sex/group: 8 |
Single i.p. application: 0.02, 0.045, 0.09 mmol/kg bwd (equivalent to 17.4, 39.1 or 78.1 mg/kg bw) |
Negative Positive controls: expected results |
Friedman and Henika (1992) | |
| α‐Chaconinea | Reverse mutation assay |
S. Typhimurium TA98, TA100 +/−S9 |
0.07–2.3 μmol/platec |
Negative Positive controls: expected results |
Friedman and Henika (1992) | |
| α‐Chaconinea | Micronucleus assay (peripheral blood cells) |
Swiss‐Webster mice (M) No/sex/group: 8 |
Single i.p. application: 0.01, 0.02, 0.045 mmol/kg bwd (equivalent to 8.5, 17.0 or 38.3 mg/kg bw) |
Negative Positive controls: expected results |
Friedman and Henika (1992) | |
| α‐Chaconina | Micronucleus assay (fetal blood cells) |
Swiss‐Webster mice (F) No of dams/sex/group: n.r. |
Single i.p. application at GD15–16: 0.005, 0.01, 0.02, 0.045 mmol/kg bwd (equivalent to 4.3, 8.5, 17.0 or 38.3 mg/kg bw) |
Negative Positive controls: expected results |
Number of fetuses per dose group: 63–65 (0.005 and 0.01 mol/kg) | Friedman and Henika (1992) |
| Solanidinea | Reverse mutation assay |
S. Typhimurium TA98, TA100 +/−S9 |
0.04–0.58 μmol/platec |
Negative Positive controls: expected results |
Friedman and Henika (1992) | |
| Solanidinea | Micronucleus assay (peripheral blood cells) |
Swiss‐Webster mice (M) No/sex/group: 8 |
Single i.p. application: 0.01, 0.02, 0.045 mmol/kg bwd (equivalent to 4.0, 8.0, 17.9 mg/kg bw) |
Negative Positive controls: expected results |
Friedman and Henika (1992) | |
| Solanidinea | Micronucleus assay (fetal blood cells) |
Swiss‐Webster mice (F) No of dams/sex/group: n.r. |
Single i.p. application at GD15–16: 0.01, 0.02, 0.045 mmol/kg bwd (equivalent to 4.0, 8.0, 17.9 mg/kg bw) |
Negative Positive controls: expected results |
Number of fetuses per does group: 46–50 | Friedman and Henika (1992) |
| Extract of S. tuberosum (plant part not specified)b | Reverse mutation assay | S. Typhimurium TA98 and 100 +/−S9 | 7.5 μg/plate | Negative | No positive control | Ness et al. (1984) |
F: female; M: male; N.r.: not reported; GD: gestational day.
Chemical standard obtained from a commercial supplier, with high purity (95% pure or higher).
A non‐ or only partly purified extract or material for which the GA composition and concentration is given.
The authors reported on the application of 0.07–2.3 mM/plate of α‐solanine (equivalent to 60.7–1996.6 mg per plate) of 0.07–2.3 mM/p of α‐chaconine (equivalent to 59.6 to 1,959.8 mg/plate) and of 0.04–0.58 mM/plate of solanidine (equivalent to 15.9–230.7 mg per plate). The CONTAM Panel assumed a typo and the application of the compounds in μmol/plate as realistic doses.
The authors reported on the application of 0.01, 0.02, 0.045, or 0.09 mol/kg bw of α‐solanine (equivalent to 8.7, 17.4, 39.1 or 78.1 g/kg bw), of 0.01, 0.02 or 0.045 mol/kg bw of α‐chaconine (equivalent to 8.5, 17.0 or 38.3 g/kg bw) and of 0.01, 0.02 or 0.045 mol/kg bw of solanidine (equivalent to 4.8, 8.0 or 17.9 g/kg bw). The CONTAM Panel assumed a typo and the application of the compounds in mmol/kg bw as realistic doses.
In summary, reduced postnatal survival of pups due to insufficient milk production was found following exposure of pregnant Holtzman rats to α‐solanine. Doses of approximately 3.6 mg/kg bw per day from different sources of α‐solanine resulted in a percentage of successfully weaned pups between 23.6% and 37.8% if compared to controls. The observed decrease most likely is due to lactational failure of dams and results in the death of the pups by starvation. The CONTAM Panel noted the limitations in the description of the study design.
In terms of fertility, effects were observed following exposure of male rhesus monkeys or male dogs to solasodine. In rhesus monkeys, 100 mg/kg bw per day reduced weight of testis and epididymis and the diameter of the seminiferous tubule. In addition, a reduced number of spermatids, spermatozoa, immature and mature Leydig cells was observed, as well as a reduction of sperm density and sperm motility in the cauda epididymis. Following oral exposure of male dogs to 80 mg/kg bw per day, decreased epididymal weight and cauda epididymal epithelial height as well as an epididymal lumen depleted of sperm were found.
3.1.2.4. Immunotoxicity studies
No standard assays for immunotoxicity of GA and aglycones could be identified.
Plant extracts of S. tuberosum as well as GA and aglycones exerted pronounced anti‐inflammatory effects in in vitro and in vivo experiments, as outlined below (see Section 3.1.5). Choi and Koo (2005) applied an ethanolic extract of the peeled tuber of S. tuberosum to mice at an oral dose of 100 or 200 mg/kg bw. One hour later, paw oedema was induced by carrageenan or formalin treatment. Pretreatment with the extract reduced significantly the oedema. The same group (Choi et al., 2005) induced arthritis by injecting Freund's adjuvant into an angle of a murine paw. Symptoms of the arthritis (oedema) were reduced when animals were pretreated orally with the ethanolic extract of S. tuberosum at 100 or 200 mg/kg bw per day for 21 days. Similar effects of the ethanolic extract of S. tuberosum were reported in an autoimmune disease model in mice (Choi, 2007).
Single GAs and aglycones, e.g. α‐solanine, α‐chaconine and solasodine, were also effective anti‐inflammatory compounds in vivo. For example, Shin et al. (2016) applied orally α‐solanine (0, 2.5, 5 or 10 mg/kg) to mice prior to an i.p. lipopolysaccharide (LPS) injection. The pretreatment with α‐solanine suppressed the expression of pro‐inflammatory genes, such as tumour necrosis factor α (TNF‐α) and interleukin 1 β (IL‐1β), in the liver. In another study, mice received orally α‐chaconine at 0.5 or 2 mg/kg bw before a single i.p. LPS injection (Lee et al., 2015). This pretreatment reduced the LPS‐induced lethality and the intrahepatic production levels of several pro‐inflammatory cytokines. Emmanuel et al. (2006, abstract only) applied solasodine, extracted from plant material as single gavage of 50 mg/kg bw to rats before carrageenan was injected to induce inflammation. Pretreatment with solasodine reduced the swelling of the paw.
To conclude, pronounced anti‐inflammatory effects were seen in rodents after single oral applications of α‐solanine, α‐chaconine, solasodine or extracts of S. tuberosum. The CONTAM Panel notes that there is not sufficient information to conclude on the immunotoxic potential of GAs.
3.1.2.5. Studies on cardiovascular effects
No studies were identified. Mechanistic studies have been identified as described in Section 3.1.5.2.3.
3.1.2.6. Neurotoxicity studies
No studies were identified. Mechanistic studies have been identified as described in Section 3.1.5.2.3 .
3.1.2.7. Genotoxicity
3.1.2.7.1. GAs from edible parts of S. tuberosum
Several assays were performed to test the genotoxic potential of potato GA, the aglycone solanidine, as well as extracts of S. tuberosum (see Table 18). The studies, applying the Ames test, reported on results obtained in only two strains of Salmonella Typhimurium, which is a major limitation regarding the quality of data.
α‐Solanine was subjected to the Ames test by two independent groups, both providing negative results. Ness et al. (1984) applied 10–50 μg per plate which did not increase the number of revertants in the strains TA98 and TA100 with or without metabolic activation. Friedman and Henika (1992) performed two independent Ames assays with α‐solanine. One of the assays was weakly positive in TA100 with S‐9 activation but only at the highest (cytotoxic) dose. The CONTAM Panel considered this study as negative. The same group applied several doses of α‐solanine via single i.p. administration to pregnant female mice at GD15‐16, which did not increase the frequency of micronuclei in the fetal blood cells (Friedman and Henika, 1992). Similarly, there was no increase in the frequency of micronuclei in peripheral blood cells of adult male mice after single i.p application of a large‐dose range of α‐solanine (Friedman and Henika, 1992).
α‐Chaconine was tested in the same assays as α‐solanine (Friedman and Henika, 1992). There was a small increase in the number of revertants in TA98 without S9 activation only at a cytotoxic dose. The CONTAM Panel considered this study as negative. The micronucleus tests performed on peripheral blood cells of male mice and fetal blood cells from exposed pregnant female mice were both negative.
The aglycone solanidine was tested by Friedman and Henika (1992) at concentrations/doses being equimolar to those of α‐solanine and α‐chaconine and was found to be uniformly negative in all the assays, i.e. in the Ames test and in the micronucleus assays on peripheral blood cells in both, the fetuses as well as in the male adult mice.
No increase in the number of revertants was observed when 7.5 μg extract/plate of S. tuberosum (plant part not specified) was assayed in strains TA98 and TA100 with or without metabolic activation (Ness et al., 1984). No conclusions can be drawn from this study since no information is provided on the method of extraction and the extent of its purification.
To conclude, from the limited number of studies available, there is no evidence of genotoxicity for the potato GAs α‐solanine and α‐chaconine, and for the aglycone solanidine. However, there is not sufficient information to conclude on the genotoxic potential of these GAs.
3.1.2.7.2. GAs from edible parts of food plants other than S. tuberosum
Several assays were performed to test the genotoxic potential of aubergine GAs, α‐solamargine and α‐solasonine (see Table 19). Almeida et al. (2010) used highly purified α‐solamargine, extracted from fruits of S. palinacanthum Dunal, in an Ames test with S. Typhimurium strains TA98, TA97a, TA100 and TA102 with and without metabolic activation. There was some weak mutagenicity in TA98 strain in the absence of metabolic activation.
Table 19.
Tests for genotoxicity/mutagenicity of aubergine glycoalkaloids (GAs) (α‐solasonine and α‐solamargine)
| Test compound | Test system | Cells/animals | Concentration/Treatment | Result | Comment | Reference |
|---|---|---|---|---|---|---|
| α‐Solamarginea | Reverse mutation assay | S. Typhimurium TA100, TA98, TA97a and TA102, +/− S9 (rat liver) |
Preincubation test 0, 1.25, 2.5, 3.5, 4.5, and 5.0 mg/plate Solvent: DMSO Appropriate positive controls |
Positive in TA98 without S9 from 4.5 mg/plate onwards (mutagenic index > 2) Statistical increase in the number of revertants in TA100 +S9 from 2.5 mg/plate onwards, but not biologically significant (mutagenic index < 2) Positive controls: expected results |
The CONTAM Panel noted the limited significance of this finding | Almeida et al. (2010) |
| α‐Solasonine and α‐solamargine (1:1)b | In vitro Comet assay |
Chinese hamster lung fibroblasts (V79) −S9 |
Preliminary cytotoxicity: 1–256 μg/mL Main test: 0, 4, 8, 16, and 32 μg/mL Positive control: MMS |
Cytotoxicity observed from 64 μg/mL onwards Negative MMS as positive control: expected result |
Munari et al. (2012) | |
| α‐Solasonine and α‐solamargine (1:1)b | In vitro chromosomal aberration assay |
Chinese hamster lung fibroblasts (V79) −S9 |
Preliminary cytotoxicity: 1– 256 μg/mL Main test: 0, 4, 8, 16, and 32 μg/mL Positive control: MMS Exposure: 18 h and harvesting at the end of treatment |
Cytotoxicity observed from 64 μg/mL onwards Negative MMS as positive control: expected result |
Munari et al. (2012) | |
| α‐Solasonine and α‐solamargine (1:1)b | In vivo micronucleus assay (peripheral blood cells) | Swiss mice (M) |
Preliminary toxicity test: 15–1,000 mg/kg bw Gavage: 0 (water),15, 30 or 60 mg/kg bw per day for 14 days Blood samples collected 48h, 7 days and 14 days after start of treatment |
Doses above 60 mg/kg bw were toxic No cytotoxicity observed Negative MMS as positive control: expected results |
Munari et al. (2014) | |
| α‐Solasonine and α‐solamargine (1:1)b | In vivo micronucleus assay (bone marrow cells) | Swiss mice (M) |
Gavage: 0 (water) or 60 mg/kg bw per day for 14 days Blood samples collected at end of treatment |
No cytotoxicity observed Negative MMS as positive control: expected result |
Munari et al. (2014) | |
| α‐Solasonine and α‐solamargine (1:1)b | In vivo Comet assay (liver cells) | Swiss mice (M) | Gavage: 0 (water), 60 mg/kg bw per day for 14 days |
No statistically significant decrease of cell viability Negative MMS as positive control: expected result |
Munari et al. (2014) |
MMS: Methyl methanesulfonate; DMSO: Dimethyl sulfoxide.
Extracted from S. palinacanthum Dunal, highly purified.
Extracted from S. lycocarpum St. Hill., 89% purity.
Munari et al. (2012) generated an extract of S. lycocarpum St. Hill, which was 89% pure and contained α‐solasonine and α‐solamargine in a ratio of 1:1. The clastogenic potential of the extract was tested in vitro in V97 Chinese hamster lung fibroblasts in the absence of metabolic activation with negative results both in the Comet and chromosomal aberrations assays. Using the same type of extract, Munari et al. (2014) performed in vivo assays in mice. In a first test, male Swiss mice were exposed by gavage to 0, 15, 30 or 60 mg/kg bw per day for 14 days and peripheral blood samples were analysed 48 h, 7 days and 14 days after treatment. No increase in the frequency of micronucleated polychromatic erythrocytes (PCE) was observed. In subsequent tests, male Swiss mice received the extract at 60 mg/kg bw per day by gavage for a duration of 14 days. Negative results were obtained both in the bone marrow micronucleus test and in the Comet assay.
In conclusion, from the limited number of studies available, there was no evidence for genotoxicity of the aubergine GA α‐solamargine. However, there is not sufficient information to conclude on the genotoxic potential of this GA.
No studies could be identified investigating the genotoxicity of tomato GAs.
3.1.2.8. Carcinogenicity studies
No long‐term carcinogenicity studies on any of the GAs and aglycones addressed in this opinion could be identified.
3.1.2.9. Studies on metabolic effects
3.1.2.9.1. GAs from edible parts of S. tuberosum
No studies on potato GA and aglycones were identified.
3.1.2.9.1.1. GAs from edible parts of food plants other than S. tuberosum
There are several studies focusing on the impact of α‐tomatine on the cholesterol metabolism. Cayen (1971) tested whether α‐tomatine forms complexes with cholesterol in the intestinal tract (see also Section 3.1.2.2). Rats received 600 mg of α‐tomatine/kg bw per day via chow, which lowered significantly the uptake of dietary cholesterol by the liver. As compensation, 14 days treatment with 1,200 mg/kg bw per day elevated the rate of acetate incorporation into neutral lipids and de novo cholesterol biosynthesis being more pronounced in hepatic than in the intestinal cells. Nevertheless, the cholesterol concentrations in the low‐density lipoproteins (LDL) and high‐density lipoproteins (HDL) fraction were reduced. Furthermore, α‐tomatine increased fecal excretion of sterol but not of bile acids, suggesting that dietary α‐tomatine forms a nonabsorbable complex with cholesterol in the intestinal tract and leaves the bile acids unaffected.
Friedman et al. (2000a) also tested whether α‐tomatine may lower dietary cholesterol absorption and plasma levels of cholesterol and triglycerides (see also Section 3.1.2.2). Hamsters were treated for 21 days with a high‐fat/high‐cholesterol diet containing from 500 to 2,000 mg/kg α‐tomatine. The highest dose (122 mg/kg bw per day) decreased the cholesterol in the serum LDL without changing the cholesterol concentration in HDL. When compared to the controls, the fecal excretion of cholesterol and coprostanol was elevated dose‐dependently up to four‐ to fivefold. This again confirmed the hypothesis of the generation of an insoluble complex between α‐tomatine and cholesterol complex in the intestinal lumen.
Friedman et al. (2000b) treated hamsters (8 per group, sex not specified) for 21 days with semipurified diets containing (i) lyophilised ripe S. lycopersicum, admixed as powder (with 0.7 mg α‐tomatine per kg dw) at a concentration of 610 g/kg chow: estimated uptake of α‐tomatine at roughly 0.05 mg/kg bw per day, or (ii) lyophilised unripe (green) S. lycopsersicum, admixed as powder (with 743 mg α‐tomatine per kg dw) at a concentration of 593.8 g/kg chow: estimated uptake of roughly 46.7 mg/kg bw per day. Compared to the control diet without tomatoes, there was a reduction in body weight and absolute liver weight, most probably due to dietary imbalances. The green and red tomatoes caused a reduction of LDL‐cholesterol by 59% and 44%, very low density lipoprotein (VLDL)‐cholesterol by 45% and 35% and of plasma triglyceride concentrations by 47% and 31%, respectively. However, the treatments did not impact on plasma levels of HDL but elevated fecal deoxycholic in hamsters fed green fruits, and fecal lithocholic acid concentrations in animals on both green and red tomato diets. However, feeding of both green and red tomato powder resulted in considerably reduced feed intake and weight gain, which impairs the interpretation of data.
To conclude, there is experimental evidence for the formation of undigestible complexes between α‐tomatine and cholesterol in the gastrointestinal tract of rodents, which may enhance fecal elimination of sterols and lower the cholesterol concentration in the plasma lipoproteins.
Khaserao and Somani (2017) described an anti‐obesity effect of the aubergine aglycone solasodine in obese rats kept on a high‐fat diet. Rising concentrations of solasodine (25, 50 and 100 mg/kg bw per day), given for 6 weeks, reduced dose‐dependently body weight, abdomen circumference, retroperitoneal adipose tissue weight and serum total cholesterol, triglyceride and glucose levels. The induction of a fatty liver was suppressed as well. It is presently unclear whether the blood lipid lowering effect of solasodine is based on a mode of action being similar to the formation of unresorbable complexes of α‐tomatine and cholesterol in the gastrointestinal tract.
3.1.3. Observations in humans
3.1.3.1. GAs from S. tuberosum
The CONTAM Panel noted that some of the authors of the reports described in this section used a terminology (e.g. ‘solanine poisoning’) not distinguishing between solanine, chaconine, α‐solanine, α‐chaconine, solanine:chaconine mixtures and total potato GAs (potato TGAs). This might also be the case for the chemical analysis reported by the authors. The authors’ terminology has been kept in this chapter.
Poisonings of humans by GAs, particularly from potato consumption, have been repeatedly reviewed, e.g. by Morris and Lee (1984), Van Gelder (1989), JECFA (1993), Tice (1998), Lee (2006), Barceloux (2009), Koleva et al. (2012) and Milner et al. (2011), who report on more than 10 separate outbreaks of potato GA poisonings affecting more than 2,000 subjects including fatalities in more than 1% of the cases. Besides showing membrane disruption activity, potato GAs inhibit the enzyme acetylcholinesterase (AChE), thereby affecting the nervous system directly (see Section 3.1.5). At doses > 1 mg/kg bw, potato GAs are considered to be toxic to humans (JECFA, 1993). The studies report that signs of intoxication may occur immediately after intake or with a latency period up to 2 days. Potato GAs induce symptoms such as sweating, vomiting, diarrhoea, severe abdominal pain, drowsiness, apathy, confusion, weakness, vision disturbances, fever, rapid and weak pulse, low blood pressure and bronchospasm. In severe cases, paralysis, respiratory insufficiency, cardiac failure, coma and death have been reported. Doses in the range of 3–6 mg potato TGAs/kg bw are described to be potentially lethal for humans (JECFA, 1993). However, estimated doses associated with toxicity are regarded as uncertain (McMillan and Thompson, 1979; Morris and Lee, 1984; Hellenäs et al., 1992; JECFA, 1993; Smith et al., 1996; Milner et al., 2011; Koleva et al., 2012).
Toxic GA concentrations are reported to be associated with taste alterations. Bitterness, e.g. when chewing a small piece of the raw peel, is indicative of a high GA concentration in potatoes (Wood and Young, 1974; Beier and Nigg, 1992). Normal levels of GAs in potatoes of less than 100 mg/kg are usually not detectable by taste. GA levels exceeding 100 mg/kg cause a slowly within 15–30 sec developing, burning sensation of the tongue and back of the mouth with a persistent irritation. Consumption of potatoes that contain more than 200 mg GAs/kg results in an immediate burning sensation (Wood and Young, 1974). Allergic reactions to raw and cooked potatoes, including cases of anaphylaxis, are only rarely reported and not associated with their content of GAs (e.g. Beausoleil et al., 2001; Majamaa et al., 2001).
In the following sections, only reports that are relevant for the risk assessment of the intake of potato GAs, e.g. giving information on dose–effect relationships, are presented in detail.
3.1.3.1.1. Reports on intoxications
Outbreaks
Several large outbreaks of potato poisonings in humans are known and include those in France (Cortical, 1888, as cited by Rothe, 1919), Germany (Schmiedeberg, 1892, as cited by Bömer and Matthis, 1924; Pfuhl, 1899), East Glasgow (Harris and Cockburn, 1918), Cyprus (Willimott, 1933), North Korea (Reelakh and Keem, 1958), South London (McMillan and Thompson, 1979) and Canada (Canada Diseases Weekly Report, 1984). Most of the victims of potato poisonings were only moderately affected. However, in some cases, serious poisonings including deaths occurred (Damon, 1928; McMillan and Thompson, 1979; Lee, 2006).
In an outbreak in 1898 in Germany, consumption of a large portion of potatoes resulted in 56 soldiers with typical symptoms of ‘solanine poisoning’, including irritation of the throat, rise of temperature, headache, abdominal pain, diarrhoea and partly vomiting or nausea, loss of consciousness and jaundice. In one case, convulsions were observed. Analysis revealed that the cooked peeled potatoes contained 240 mg/kg and the unpeeled, uncooked potatoes 380 mg ‘solanine’/kg (Pfuhl, 1899). The author estimated that for those who consumed the whole meal, the intake of ‘solanine’ had been 300 mg. In the JECFA (1993) assessment, the intake of ‘solanine’ was estimated to have been 3.4–5.1 mg/kg bw.
In 1917, 61 persons from 18 households in Glasgow in Scotland showed the typical picture of ‘solanine poisoning’ immediately or within 3 h after eating potatoes. Observed symptoms were vomiting, diarrhoea, headaches and debility. A 5‐year‐old boy died, and the post‐mortem examination showing that death was due to bowel strangulation, was interpreted as a consequence of extreme retching and vomiting. The analysis of the potatoes consumed in the boy's family contained 410 mg ‘solanine’/kg (Harris and Cockburn, 1918). JECFA (1993) concluded that, assuming a body weight of 18 kg and a consumption of 200 g, the lethal dose for the boy may be estimated at 4.5 mg/kg bw. Furthermore, for adults an intake of 3.4 mg ‘solanine’/kg bw was estimated under the assumption that they ate 500 g potatoes and had a bw of 60 kg.
Rothe (1919) reported on an outbreak in Leipzig (Germany) in 1919, which affected 14 persons after consumption of potato dishes with immediate onset of gastrointestinal pain, followed by vomiting and diarrhoea. The author conducted a self‐experiment from which he concluded that intake of 150 mg to 200 mg ‘solanine’ with a meal of cooked peeled potatoes by a male person could lead to gastric discomfort and nausea after 90 min, followed by accelerated pulse, symptoms fading within a day (Rothe, 1919).
McMillan and Thompson (1979) reported on an intoxication involving 78 British junior schoolboys (11–15 years of age), who became ill with diarrhoea and vomiting after eating two small boiled peeled potatoes each (weight of potatoes not indicated) as part of their midday meal. Symptoms developed mainly between 20.00 and 22.00 o'clock the same evening. Seventeen out of the 78 boys were hospitalised with symptoms of vomiting, severe diarrhoea, along with generalised abdominal pain, commencing together 7–19 h after the meal. Their feces were watery and in 13 cases showed markedly green colour by the second or third day as was the case for some of the vomitus. Fifteen of the 17 boys developed fever being maximal just before admission and subsiding within 1–3 days. Restlessness, sometimes extreme, was a common symptom often combined with other central nervous system (CNS) effects, such as stupor, delirium, hallucinations, confusion, drowsiness or disorientation. The three most seriously ill boys were comatose or stuporose on admission and had a peripheral circulatory collapse. All of the 17 boys were discharged home within 6–11 days following admission, and when seen 4–5 weeks later were without sequelae. From the 61 boys not admitted to hospital, only for 24 of them Public Health Department records were available. According to these records, all these boys suffered from abdominal pain, diarrhoea and/or vomiting. Twelve boys experienced pyrexia and 13 headaches. Other symptoms reported were delirium, dryness of the throat, trembling, dizziness and weakness, sometimes amounting to a feeling of paralysis. Bacteriological examinations of blood, serum, vomit, urine and feces as well as of samples of food recovered from the meal have all been proved to be negative. Testing of 22 samples of the school meal, 10 samples of the boys’ urine and 23 of feces for excess lead, copper, arsenic and zinc revealed negative results as well. Assays for the presence of organic compounds, such as nicotine, organophosphorus or organochlorine pesticides, metaldehyde or defoliant were also negative. Six days after eating the meal, pseudocholinesterase plasma levels in 10 out of 17 schoolboys were subnormal and returned to the normal range 4 or 5 weeks later, for all but one value. The source of poisoning was seen in a bag of potatoes left from the previous term that had been considered ‘unfit’ for consumption upon inspection, but that had inadvertently been cooked. Peeled boiled potatoes left‐over, uneaten after the meal, were extracted and analysed for their anticholinesterase activity. Based on the enzyme activity, the concentration was estimated to amount to 250–300 mg ‘solanine’/kg peeled, boiled potato. The authors claimed that this concentration considerably exceeded that of normal fresh peeled tuber (normal value for whole unpeeled tuber: less than 100 mg/kg, with most GAs in the peel). JECFA (1993) estimated the intake of ‘solanine’ by the schoolboys to have been approximately 1.4–1.6 mg/kg bw.
In 1983, a poisoning incident occurred in Alberta (Canada) when 61 out of 109 school children and staff members suffered from a sudden onset of symptoms after eating baked potatoes containing 494 mg solanine/kg (Canada Diseases Weekly Report, 1984). Predominant symptoms with frequency of occurrence indicated were the following: nausea (69%), abdominal cramps (43%), headache (33%), vomiting (25%), fever (11%) and diarrhoea (8%). The affected persons had recovered in about 3 h. From the affected individuals, 44% had noted a bitter or unusual taste and 18% had felt a burning sensation in the throat when eating the baked potatoes. According to the observations of several staff members the potatoes had a slight tinge of green. Testing of the potatoes for the presence of bacteria, viruses, moulds, pesticides or other chemicals yielded negative results (Canada Diseases Weekly Report, 1984). JECFA (1993) estimated the exposure in this poisoning outbreak at about 2.5 mg ‘solanine’/kg bw assuming that the children consumed 200 g potatoes and had a body weight of 40 kg (JECFA, 1993).
Case Reports
While early reports of potato poisoning (Muncke, 1845, as cited by Bömer and Matthis, 1924; Morris, 1859, as cited by Van Gelder, 1989) refer to severe or fatal cases associated with the consumption of green, unripe or ‘new’ tubers, some studies in the years 1923 and 1924 addressed health impairments related to the intake of potato tubers of normal appearance (Alfa and Heyl, 1923; Bömer and Mattis, 1923, 1924; Griebel, 1923). Bömer and Mattis (1923, 1924) reported that, although measured ‘solanine’ contents in samples of the consumed potatoes were relatively high and ranged from 257 to 583 mg/kg (raw, unpeeled tubers), observed symptoms were not severe and included burning sensation in the mouth, irritation of mucous membranes, gastrointestinal disturbance and vomiting. Similar symptoms were reported by Griebel (1923) and Alfa and Heyl (1923) after intake of potatoes with ‘solanine’ levels ranging from 196 to 790 mg/kg (raw, unpeeled tubers) or 109 to 468 mg/kg (raw, peeled tubers), respectively.
In 1924, seven members of a family living in Illinois (USA) were poisoned by consumption of greened potatoes. Two of them died, while the others recovered. Symptoms were epigastric pain, nausea and vomiting. Patients were apathetic and extremely exhausted. Respirations were accelerated. In the two fatal cases, consciousness was lost 3 to 4 h before death (Hansen, 1925).
A food control report from Switzerland reported on two samples of potatoes, which caused burning in the throat and nausea after consumption, ‘solanine’ levels of 230 and 350 mg/kg (Viollier, 1941). No further information was available.
In the UK, a poisoning incident occurred involving four members of a family on three consecutive Sunday evenings about 9 h after they had consumed baked potatoes eating the flesh and the skin of the tubers. Symptoms consisted of sickness, abdominal pain, diarrhoea and general malaise. Two family members who had only eaten one potato suffered only slightly, a third one, who had two potatoes, suffered more severely and the fourth one who ate three potatoes suffered very severely. A fifth person, who only ate the flesh of the potatoes, did not develop any symptoms. The patients recovered within 24 h. The potatoes contained 500 mg ‘solanine’/kg (Wilson, 1959). JECFA (1993) estimated the intake of ‘solanine’ at 1.25–3.2 mg/kg bw assuming a weight of 150 g per potato.
A 70‐year‐old woman developed symptoms of poisoning, including vomiting, diarrhoea and bloody feces, after consumption of a juice prepared from partly peeled potatoes without sprouts (Gonmori et al., 1993). She was admitted to hospital and discharged after 27 days (no information on recovery available, only abstract in English). Her blood was analysed by GC and GC‐MS on the 4th and 20th days without ‘solanine’ being detected. The potatoes from which the juice was prepared contained a 15‐fold higher ‘solanine’ concentration compared to normal potatoes (levels not given by the authors) (Gonmori et al., 1993).
In November 2015, members of a family in Baden‐Württemberg (Germany) experienced vomiting and stomach ache after the consumption of potato dishes (jacket potatoes and potatoes boiled in the skin). Due to the bitter taste, a salad prepared from the same potato charge was not consumed. Chemical analysis showed that the potatoes, which were without damage, green sites, sprouts or microbiological contamination, contained 236 mg potato TGAs/kg (141 mg α‐solanine and 95 mg α‐chaconine per kg). In the potato salad, a lower concentration of 124 mg potato TGA/kg was found. No information was available on the quantity of potatoes consumed, if all members of the family were affected by the intoxication and on recovery (BfR, 2018a,b).
3.1.3.1.2. Studies in human volunteers
In 1919, Rothe conducted a self‐experiment from which he concluded that intake of 150 mg to 200 mg ‘solanine’ with a meal of cooked peeled potatoes by a male person could lead to gastric discomfort and nausea after 90 min, followed by accelerated pulse, symptoms fading within a day (Rothe, 1919).
In three Swedish adult volunteers (two males and one female), who consumed daily for 1 week 200–300 g of cooked unpeeled potatoes of varieties high in GAs (180–300 mg/kg), no signs of acute toxicity were noted. However, the three subjects reported on a bitter taste (Harvey et al., 1985a,b). JECFA (1993) estimated the intake of TGA at 1 mg/kg bw.
In a kinetic study, seven healthy male adults have been given a potato meal, which consisted of mashed potatoes prepared from two parts peeled potatoes and one part low‐fat milk (Hellenäs et al., 1992, see Section 3.1.1). The dish contained 200 mg GAs (82 mg α‐solanine and 118 mg α‐chaconine) per kg. The amount consumed was adjusted in relation to each subject's body weight to standardise the TGA intake for 1 mg/kg bw. All except one of the volunteers experienced signs of toxicity (mainly light to severe nausea, in one case combined with diarrhoea) and an unpleasant off‐taste after ingestion of the meal. In most subjects, symptoms started within about 30 min after exposure and lasted for 3–4 h. The peak serum concentrations for α‐solanine and α‐chaconine were on average reached after 5.1 and 6.0 h, respectively, and amounted on average to 7.7 and 14.4 ng/mL (8.8 and 16.9 nmol/L), respectively. During the 25‐h period, the highest solanidine values varied among the subjects between 1.0 and 4.8 ng/ml (2.5 and 12 nmol/L). No correlation between the severity or duration of adverse effects and GA serum levels was seen and the highest maximum blood serum concentrations for α‐solanine and α‐chaconine of 11.3 and 21.4 ng/mL, respectively, were exhibited in the one subject not experiencing signs of toxicity (solanidine serum level not given). According to the authors, the rapid onset and short duration of the symptoms compared to the GA serum profiles suggest that the symptoms might be the consequence of local direct effects on the gastrointestinal tract rather than of systemic interactions. The authors noted that the study was not focusing on the toxic effects of the GAs and was not conducted double‐blinded. Therefore, a placebo effect could not be excluded.
Bushway (1987, personal communication, as cited by Van Gelder, 1989) reported on a study in six male adult volunteers who consumed potato tubers which contained 320 mg potato TGA/kg. The potatoes were baked and consumed unpeeled. The amounts of GAs ingested ranged from 1.75 to 2.58 mg/kg bw. Symptoms of intoxications occurred in all of the subjects, started about 2–4 h after ingestion and included nausea, vomiting and diarrhoea. Symptoms faded away after 8–10 h.
In a study in 14 volunteers (6 males, 8 females), aged 18–45 years, the toxicity and toxicokinetics of different oral single doses of the potato GAs, α‐chaconine and α‐solanine, were investigated (Mensinga et al., 2005, see Section 3.1.1). The subjects (2–3 per treatment) received solutions with potato TGA doses of 0.30, 0.50 or 0.70 mg/kg bw, or mashed potatoes adjusting TGA doses to 0.95, 1.10 or 1.25 mg/kg bw under standardised conditions. The solutions contained 50% α‐solanine and 50% α‐chaconine. The mashed potatoes had a TGA concentration of 199 mg/kg (51% α‐solanine, 49% α‐chaconine), which was recognised as upper limit of safety according to the authors. No adverse events did occur in any of the treatments, except for one of the two volunteers in the high‐dose group receiving 1.25 mg potato TGA/kg bw in mashed potatoes. About 4 h after administration, this subject developed nausea and started vomiting, which was associated by the authors with GA toxicity, even though the doses were lower than those generally reported in the literature to cause gastrointestinal disturbances. According to the authors, the serum GA concentrations of this subject were comparable to those of the subject receiving a similar dose and did not show any symptoms. For both subjects, maximum blood serum concentrations were ranging for α‐solanine between 8.0 and 11.8 ng/mL and for α‐chaconine between 6.3 and 9.7 ng/mL, no individual levels being indicated for each subject. The authors concluded from the results that some subjects may be more susceptible for adverse effects of potato GAs than others.
3.1.3.1.3. Epidemiological studies
Congenital anomalies
Intake of potato GAs, particularly with blighted or injured potatoes have been proposed as an aetiological factor in the pathogenesis of neural tube defects (NTD) (anencephaly and spina bifida) (Renwick, 1972; Harvey et al., 1986). This hypothesis has been addressed in two epidemiological studies (Nevin and Merrett, 1975; Harvey et al., 1986). Maternal consumption of sprouted potatoes as risk factor of NTD has also been investigated in two Chinese studies (Wang et al., 2008; Ni et al., 2018).
In the prospective study of Nevin and Merrett (1975) in women with a previous infant with either anencephaly and/or spina bifida and advised to avoid potato consumption, the incidence of NTD was 8.7% in the group of women adhering to the advice (n = 27), and 3.6% in the group of non‐compliers (n = 61) (p = 0.58). In the prospective study by Harvey et al. (1986) conducted in NTD screening centres, outcome of pregnancy in 380 patients resulted in 210 NTD cases and 170 normal offspring, whereby in most of the nine centres studied, serum total potato alkaloids levels and serum ‘solanidine’ levels were higher in the women with a normal fetus than in those with a fetus affected by NTD. For both groups serum total potato alkaloid concentrations varied widely between individuals as well as between each of the participating centres, whereby levels for individual subjects ranged from < 10 to < 150 nmol/L.
Two Chinese case–control studies investigated the risk factors of NTD in the Shanxi province in Northern China (Wang et al., 2008; Ni et al., 2018). The results indicate that, among other factors, maternal consumption of sprouted potatoes during the periconceptional period may increase the risk of NTDs or orofacial clefts (OFCs). Subjects recruited between 2002 and 2007 by Ni et al. (2018) included 622 cases with NTD, 135 cases with orofacial clefts (OFCs) and 858 controls. Data collection comprised information on maternal sprouted potato consumption, lifestyle behaviours and folic acid supplementation. Intake of sprouted potatoes was not associated with spina bifida or encephalocele, but with increased odds of total NTDs (OR = 2.20; 95% CI, 1.12–4.32) and anencephaly (OR = 2.48; 95% CI, 1.10–5.58) and with elevated risk of total OFCs (OR = 3.49; 95% CI, 1.29–9.49) and cleft lip with or without cleft palate (CL ± P) (OR = 4.03; 95% CI, 1.44–11.28). The authors also found that maternal consumption of sprouted potatoes was significantly associated with NTDs and OFCs among mothers with lower consumption of B vitamins, but not among mothers with a higher consumption of B vitamins. Another case–control study was conducted by Wang et al. (2008) in subjects genotyped for methylenetetrahydrofolate reductase (MTHFR) C667T polymorphism. Multivariate conditional logistic regression analysis indicated that four maternal factors were linked to NTDs: frequency of pregnancy (OR = 2.87, 95%CI: 1.28–6.44), contacting chemicals in early pregnancy (OR = 16.18, 95%CI: 1.18–221.59), frequent taking of germinated potatoes in early pregnancy (OR = 4.66, 95%CI: 1.78–12.17) and MTHFR C677T mutation (OR = 2.13, 95%CI: 1.08–4.21). The CONTAM Panel noted that no information on GA levels in the sprouted potatoes or GA intake was given in the two studies.
The CONTAM Panel noted that the hypothesis of Renwick (1972) has not been supported by the outcome of the epidemiological studies of Nevin and Merrett (1975) and Harvey et al. (1986). The design and results of the two Chinese studies (Wang et al., 2008; Ni et al., 2018) do not allow conclusions on causal relationships of GA exposure and developmental effects.
Cancer
The relationship between the intake of potatoes and risks of cancer in the gastrointestinal tract has been investigated. As reviewed by Hopkins (1995) and Tice (1998), results of epidemiological studies were contradictory and remained altogether inconclusive. A causal relationship between diets with the consumption of large quantities of potatoes and increased risks of cancers of the brain, breast, endometrium, lung and thyroid could not be proven (Hopkins, 1995; Tice, 1998).
The CONTAM Panel noted that the design and outcome of existing studies do not allow concluding on the causal relationship between intake of potato GAs and cancer risks.
3.1.3.1.4. Summary
Overall, the CONTAM Panel noted that the information from the available human data is limited. However, results from kinetic studies in volunteers (Harvey et al., 1985a,b; Hellenäs et al., 1992; Mensinga et al., 2005) and from reports on intoxications (e.g. Wilson, 1959; Bushway, 1987, personal communication, as cited by Van Gelder, 1989) indicate that acute toxic effects, such as nausea, vomiting and diarrhoea, may occur in adults from a potato TGA intake of 1 mg/kg bw or more (Table 20). Doses in the range of 3–6 mg potato TGAs/kg bw are considered to be potentially lethal for humans.
Table 20.
Summary of reports on outbreaks and cases of intoxications in humans and on volunteer studies relevant for the identification of the reference point for acute toxicity
| Affected subjects | Potato type/preparation | Concentration of solanine or TGAs in the potatoes or potato preparation, respectivelya | Intake per person | Estimated single dose of solanine or potato TGA/kg bwb | Taste, symptoms and outcome | Reference |
|---|---|---|---|---|---|---|
| 3 adult volunteers (two males and one female) | potato varieties high in GAs | 180–300 mg ‘TGA’ mg/kg potatoes | for 1 week daily 200–300 g of cooked unpeeled potatoes | 1 mg ‘TGA’/kg bwc | bitter taste, no signs of acute toxicity | Harvey et al. (1985) |
| 6 out of 7 male adult volunteers | preparation of mashed peeled potatoes with milk | 200 mg TGAs (82 mg α‐solanine and 118 mg α‐chaconine)/kg potato preparation | adjusted to bw resulting in a dose of 1 mg TGAs/kg bwd | 1 mg TGAs/kg bwd | unpleasant off‐taste;nausea, which in one case was combined with diarrhoea; all recovered within 4 h | Hellenäs et al. (1992) |
| 6 male adult volunteers | baked potatoes, consumed unpeeled | 320 mg TGAs/kg | n.r. | 1.75–2.58 mg TGAs/kg bwd | nausea, vomiting, diarrhoea, all recovered within 8–10 h | Bushway (1987, personal communication, as cited in Van Gelder, 1989) |
| 1 out of 2 adult volunteers | mashed potatoes | 199 mg TGA/kg fw (51% α‐solanine, 49% α‐chaconine) | adjusted to bw resulting in a dose of 1.25 mg TGAs/kg bwd | 1.25 mg TGAs/kg bwd | nausea, vomiting | Mensinga et al. (2005) |
| 4 adults | baked potatoes (flesh and skin eaten) | 500 mg ‘solanine’/kg potatoes | 1–3 potatoes | 1.25–3.25 mg ‘solanine’/kg bwc | sickness, abdominal pain, diarrhoea, general malaise; all recovered within 24 h | Wilson (1959) |
| 78 school‐boys (11–15 years old) | old potatoes, ‘unfit’ for consumption | 250–300 mg ‘solanine’/kg peeled boiled potatoes | two small potatoes | 1.4–1.6 mg ‘solanine’/kg bwc | diarrhoea, vomiting, abdominal pain, 17 boys hospitalised due to severe symptoms including 15 subjects with fever and 3 being comatose or stuporose; all recovered | McMillan and Thompson (1979) |
| 61 out of 109 school children and staff members | baked potatoes with a ‘slight green tinge’ | 494 mg ‘solanine’/kg potatoes | n.r. | 2.5 mg ‘solanine’/kg bwc | bitter or unusual taste (44%), burning sensation in the throat (18%);nausea (69%), abdominal cramps (43%), headache (33%), vomiting (25%), fever (11%), diarrhoea (8%); all recovered in about 3 h | Canada Diseases Weekly Report (1984) |
Concentrations are given as ‘potato TGAs’, ‘solanine’, ‘α‐solanine’ or ‘α‐chaconine’ depending on the subject of analysis as described in the referenced publication.
Doses are given as ‘TGAs’, or ‘solanine’, depending on the subject of analysis as described in the referenced publication.
As estimated in JECFA (1993).
As estimated by the authors.
Results of studies conducted under standardised conditions (Hellenäs et al., 1992; Mensinga et al., 2005) suggest that differences in the individual susceptibility towards adverse effects associated with the intake of potato GAs may be expected. The available human data do not allow to draw conclusions if children or other subgroups of the population, such as individuals with inflammatory bowel diseases, may be more susceptible to adverse effects associated with potato GAs than others.
No data showing convincing evidence of GA‐related health problems in humans associated with the long‐term intake of potatoes have been identified.
3.1.3.2. GAs from food plants other than S. tuberosum
The occurrence of GAs in foods other than potatoes has been reviewed by Tice (1998) and Dolan et al. (2010) (see Sections 1.3.1 and 1.3.3). For example, tomato fruits, particularly the green unripe fruits, are known to contain, i.a. α‐tomatine. However, according to Dolan et al. (2010), there is no evidence to consider α‐tomatine as a substance of concern and there are no reports of acute toxicity in humans due to ingestion of green tomatoes.
3.1.3.2.1. Case Reports
Smith et al. (2008) reported on intoxications occurring on two different occasions with susumber berries originating from the plant S. torvum. Poisonings with susumber berries personally imported from Jamaica occurred in three persons in 2003 in Toronto and in three persons in 2006 in York City. Symptoms included diarrhoea, weakness, facial paralysis, slurred speech and ataxia. The most seriously affected patients developed hypertension, confusion and respiratory failure requiring mechanical ventilation. The authors noted that the imported poisonous berries ingested appeared indistinguishable from non‐toxic varieties commercially available on the market. α‐Solasonine and α‐solamargine, which share the same aglycone, solasodine, were isolated from the toxic berries but not from their non‐toxic counterparts (Smith et al., 2008).
The French Agency for Food, Environmental and Occupational Health and Safety (Anses) reported in 2014 on 26 cases of poisoning in France associated with consumption of cherry tomatoes originating from Morocco (Anses, 2014). Gastrointestinal symptoms, including vomiting, abdominal cramps and irritation of the pharynx, were accompanied by an unpleasant bitter taste. The symptoms were observed in most cases 5–30 min after ingestion of the cherry tomatoes. Based on chemical analysis using LC‐HRMS, rubijervine was postulated as a metabolite present in the cherry tomatoes. However, it was not possible to confirm the identity or to perform a quantification. Neither α‐tomatine nor α‐solanine could be detected in the tomatoes. Rubijervine is the 12α‐hydroxy metabolite of solanidine which is known to occur also in other plants such as those of the genus Veratrum. Metabolome profiling studies conducted on wild and cultivated tomato species using similar techniques have not reported the presence of rubijervine in any of the investigated samples (Moco et al., 2007; Iijima et al., 2013; Schwahn et al., 2014), a fact also mentioned by the authors of the Anses report. The authors concluded that the intoxications could have been caused by GAs, but that further investigations would be necessary to identify the causative substances.
3.1.4. Adverse effects in farm animals, horses and companion animals
The selection criteria for studies relevant to inform the adverse effects in farm and companion animals are described in Section 2.1.
3.1.4.1. Ruminants
Three studies on ruminants could be identified (see Table 21), which lack significance due to the low number of animals investigated and due to lack of information on the exact composition and the degree of purity of the test compounds. These limited studies indicate that acute toxicity may be expected to occur at oral doses above 40 mg/kg bw in sheep and cows.
Table 21.
Acute oral toxicity of glycoalkaloids (GAs) (α‐solanine) and aglycones (solasodine) in ruminants
| Test compound | Species Dose route Doses | Observed effects | Highest dose with no effect (mg/kg bw) | Lowest dose with effects (mg/kg bw) | LD50 (mg/kg bw) | Reference |
|---|---|---|---|---|---|---|
| α‐Solanineb |
Sheep (F) No/sex/group: 1 Single oral dose: 100 or 225 mg/kg bw NO CONTROL |
Anisocytosis, poikilocytosis, leucocytosis | 225 | König and Staffe (1953) | ||
| α‐Solaninea |
Adolescent bull (M) No/sex/group: 1 Single gavage: 77 mg/kg bw NO CONTROL |
77 | Liebenow (1970) | |||
| Solasodinea |
Adolescent bull (M) No/sex/group: 1 Singe gavage: 43 mg/kg bw NO CONTROL |
43 | Liebenow (1970) |
F: female; M: male.
Chemical standard obtained by in‐house isolation from plant material, and checked for purity (95% pure or higher).
Poorly characterised chemical standard (purity or composition not defined, may contain α‐chaconine).
3.1.4.2. Pigs
Kerr et al. (1998) treated weanling pigs with control diet containing 3% spray‐dried animal plasma or diets with additional 2.6% or 5.1% potato proteins for 28 days (see Table 22). GA content of potato proteins was determined by HPLC‐UV. No information is provided on the concentration of α‐solanine and/or α‐chaconine. Increasing concentrations of potato proteins had no effect on the average daily body weight gain or food intake of the animals.
Table 22.
Repeated dose toxicity of glycoalkaloids (GA) containing potato protein in pigs
| Test compound | Species Dose route Doses | Observed effects | Highest dose with no effect (mg/kg bw) | Lowest dose with effects (mg/kg bw) | LD50 (mg/kg bw) | Reference |
|---|---|---|---|---|---|---|
| Potato proteina |
Pigs (M,F) No/sex/group: ~ 18 Diet: 0, 4.6 or 8.5 mg TGA/kg bw/day Duration: 28 days |
8.5 | Kerr et al. (1998) |
F: female; M: male.
A non‐purified material for which the TGA concentration is given.
3.1.4.3. Poultry
The studies identified are reported in Table 23.
Table 23.
Repeated dose toxicity of glycoalkaloids (GAs) (α‐solanine) and potato protein concentrates in poultry
| Test compound | Species Dose route Doses | Observed effects | Highest dose with no effect (mg/kg bw per day) | Lowest dose with effect (mg/kg bw per day) | Reference |
|---|---|---|---|---|---|
| α‐Solaninea |
Acres chicken (M) No/sex/group: 3 Diet: 0, 12.5, 25 or 50 mg/kg (equivalent to 0, 1.1–3, 2.2–6 or 4.4–12 mg/kg bw per day) Duration: 46 days |
↑bw gain | 2.2–6 | 4.4–12 | Vogt and Stute (1969) |
| Potato protein concentrate containing GAs and also trypsin inhibitors at various concentrationsb |
Chicken (F) No/sex/group: 14 Diet: 0, 86.2, 87.7, 154.2, 274.5 or 318.5 mg/kg bw per day Duration: 23 days |
↓ decreased height of villi in jejunum + ileum ↓depth of crypts in jejunum |
87.7 87.7 |
154.2 154.2 |
Tuśnio et al. (2013b) |
| Potato protein concentrate containing GAs and also trypsin inhibitors at various concentrationsb |
Chicken (F) No/sex/group: 24 Diet: 0, 33.8, 67.7 or 101.5 mg/kg bw per day Duration: 35 days |
↓feed intake ↓bw gain ↑relative weight of pancreas and liver ↑relative weight of gastrointestinal tract |
67.7 67.7 33.8 67.7 |
101.5 101.5 67.7 101.5 |
Tuśnio et al. (2013b) |
F: female. M: male. Bw: body weight,
Poorly characterised chemical standard (e. g. purity or composition not defined).
A non‐ or only partly purified extract or material for which the GA composition and concentration is given.
Vogt and Stute (1969) used an extract of S. tuberosum as a source of α‐solanine. However, information is lacking whether the extract contained also α‐chaconine or further GA or aglycones. The extract was admixed to a diet containing dried ground potatoes with a α‐solanine content of 100–150 mg/kg. This treatment increased the body weight gain in young male Acres chicken when treated daily with 4–12 mg α‐solanine per kg bw for 46 days.
Tuśnio et al. (2013b) used two different potato protein concentrates containing 677–3,185 mg/kg TGA. No information is given on the concentrations of α‐solanine and α‐chaconine, and the ratio between these two GA is likely to be different in the potato concentrates. A GA dose of 101.5 mg/kg bw per day reduced feed intake and body weight gain in female chicken when treated over a period of 35 days. Furthermore, an increased relative weight of liver and pancreas could be observed in this study at a dose of 67.7 mg/kg bw per day. When applying 154.2 mg/kg bw per day, the authors observed morphological changes in the intestinal tract of female chicken. The interpretation of these observations is hampered by the fact that the diet contained also trypsin inhibitors at various concentrations, which might have affected the digestibility of the feed components and the activity of the gastrointestinal tract.
3.1.4.4. Rabbits
No studies on the adverse effects in rabbits have been identified other than the ones described already in Sections 3.1.2.1 and 3.1.2.2. In the repeated‐dose toxicity studies, no NOAEL or LOAEL could be derived, since untreated controls were missing.
3.1.4.5. Fish
The studies identified in fish are reported in Table 24. Xie and Jokumse (1997) applied potato protein concentrate, containing solanine (no more details reported) at 1 g/kg dw. This concentrate was admixed to a fishmeal‐based diet and fed to rainbow trouts for a period of 4 weeks. At an uptake of solanine of daily 1.12 mg/kg bw, the specific growth rate of the trouts was reduced significantly.
Table 24.
Repeated dose toxicity of potato protein concentrates in fish
| Test compound | Species Dose route Doses | Observed effects | Highest dose with no effect (mg/kg bw per day) | Lowest dose with effect (mg/kg bw per day) | Reference |
|---|---|---|---|---|---|
| Potato protein concentrate containing solanine at 1,000 mg/kg dwa |
Rainbow trout Application oral Control diet (fish meal) admixed with potato protein concentrate at isontrogenous and isoenergetic levels containing solanine at 0, 22, 56, 89 and 111 mg/kg diet (equivalent to: 0, 0.44, 1.12, 1.78 or 2.22 mg/kg bw per dayb) Duration: 28 days |
Specific growth rate | 0.44 | 1.12 | Xie and Jokumsen (1997) |
| Potato protein concentrate containing GA at 7.41 mg/kg dwa |
Rainbow trout Application oral Diet containing α‐solanine/α‐chaconine at 0/0, 0.4/0.6, 0.8/1.2, 1.2/1.7 or 1.6/2.3 mg/kg dw (equivalent to: 0/0, 0.008/0.012, 0.016/0.024, 0.024/0.034 or 0.032/0.048 mg/kg bw per dayb) Duration: 84 days |
↓ bw ↓Serum glucose ↓Serum protein Altered liver histology |
0.016/0.024 |
0.008/0.012 0.024/0.034 0.008/0.012 0.008/0.012 |
Tusche et al. (2011) |
| Potato protein concentrate containing GA at 2,150 mg/kg dwa |
Rainbow trout Application oral Diet containing α‐solanine/α‐chaconine at 0/0, 166/149, 306/298, 459/446 or 612/595 mg/kg dw (equivalent to: 0/0, 3.32/2.98, 6.12/5.96, 9.18/8.92, 12.24/11.9 mg/kg bw per dayb) Duration: 84 days |
↓ bw ↓Relative liver weight ↓Serum glucose and protein ↓Serum triglyceride Altered liver, intestinal and stomach histology |
3.32/2.98 3.32/2.98 3.32/2.98 3.32/2.98 3.32/2.98 |
Tusche et al. (2011) |
dw: dry weight; bw: body weight.
A non‐ or only partly purified extract or material for which the GA composition and concentration is given.
Body weight of 2 kg and average feed consumption of 40 g per day were assumed (EFSA, 2011c).
One 84‐day study in rainbow trout indicated that the most sensitive end points are decreased body weight, lowered serum proteins and an altered hepatic histology occurring at the lowest doses of α‐solanine/α‐chaconine of 0.008/0.012 mg/kg bw per day (Tusche et al., 2011). No NOAEL or LOAEL could be identified.
3.1.4.6. Horses
No studies were identified.
3.1.4.7. Companion animals (cats and dogs)
No studies were identified.
3.1.4.8. Fur animals
No studies were identified.
3.1.4.9. Reports on intoxications
The risks of feeding green potatoes or sprouts to livestock have been well understood for many years, and therefore, there have been relatively few recent reports of intoxications following ingestion of these feeds. Outbreaks of poisoning of cattle in the UK fed green or sprouting potatoes have been reported (Milligan, 1941), while in Poland, outbreaks of ‘solanine’ poisoning in pigs were reported between 1949 and 1961. These involved 180 pigs, and resulted in 64% mortality, but levels of GAs were not given (Cooper and Johnson, 1984). Sheep are also susceptible to poisoning following consumption of green potatoes. In the USA, 14 (of 35) became weak, uncoordinated and subsequently died after consuming green potatoes which had been spread on the pasture (Bolin, 1962)
Poisoning of livestock following consumption of other GA‐containing plants, such as S. dulcamara and S. nigrum, are uncommon. Because of their strong odour and bitter taste, animals do not eat the growing plants as long as other feed is available. However, these plants may grow as weeds in hay and silage crops, where they can be harvested with the crop and then fed to livestock. Drying or ensiling does not destroy the toxin (USDA, 2011), and as a result cases of accidental poisoning (or suspected poisoning) have been reported in horses, cattle, sheep, goats and pigs where livestock have consumed hay or silage, which have been contaminated with these plants (Cooper and Johnson, 1984).
Norman et al. (2012) reported a case in which six horses were fed poor quality hay containing S. eleagnifolium (Silverleaf nightshade) and S. dimidiatum (Western horse nettle). S. eleagnifolium is known to contain solasodine‐type GAs (Álvarez et al., 1994). Symptoms included gastrointestinal upset, respiratory depression and central nervous system depression, obtundation, ataxia, pyrexia, muscle fasciculations and ileus. Therapy for these horses was poorly described, but all the affected horses recovered. Gunning (1950) also reported a case of poisoning and death in a horse fed greened potato sprouts.
Poultry appear to be less susceptible to ‘solanine’ poisoning; in feeding trials laying hens tolerated green potato sprouts and ground tubers when included at 10% of their diet for long periods (Temperton, 1943).
Kees et al. (2015) reported a case of GA toxicity in a 10‐week‐old male Labrador retriever dog as a result of ingesting dried stems and unripe berries of S. dulcamara. S. dulcamara may contain a variety of GAs such as α‐solasonine, α‐solamargine, α‐solamarine, β‐solamarine and soladulcine A (Calf et al., 2018). The level of exposure to GAs was not determined.
3.1.5. Mode of action
Information from the available human data on poisoning with potato GAs indicate that acute toxic effects of GAs comprise nausea, vomiting, abdominal cramps and diarrhoea (see Section 3.1.3). These symptoms may be caused by local irritating effects in the mucosa of the gastrointestinal tract referring to the ability of potato and tomato GAs to form complexes with membrane 3β‐hydroxy sterols, thereby disrupting the integrity of cell membranes. The acute toxic effects in humans might be related also to the cholinergic toxidrome or toxic syndrome, comprising a set of symptoms caused by a group of toxic chemicals leading to overstimulation of cholinergic receptors.20 The various effects of GAs are outlined in detail below.
3.1.5.1. Membrane effects with implications for the gastrointestinal tract
GAs, like α‐solanine, α‐chaconine and α‐tomatine, may exert saponine‐like effects which may injure membranes. This membrane toxicity has been documented in several tissues or cell lines, such as rat liver cells (specifically in mitochondria) (Keukens et al., 1996), mammalian erythrocytes (Keukens et al., 1996), rodent cell lines (Gee et al., 1996) as well as in human colorectal carcinoma (Glynn et al., 2017; Arena et al., 2018), embryonic intestinal (Gee et al., 1996; Keukens et al., 1996; Patel et al., 2002) and MCF7 breast cancer cell lines (Sucha et al., 2013).
The molecular mechanism of toxicity may be based on membrane destabilisation (Friedman, 2006). The presence of sterols in the membrane appears to be an absolute requirement for the membrane disruptive action of the GAs (Keukens et al., 1992). It has been suggested that the initial insertion of the aglycone part into the membranes is followed by the generation of a 1:1 complex with membrane sterols, which disrupts the structure of the lipid bilayers. GAs interact with the sterols in the following potency order: α‐tomatine > α‐chaconine > α‐solanine (Friedman, 2006). Deletion of one or more mono‐saccharides from the GAs resulted in an almost complete loss of activity. Accordingly, solanidine was weakly active (Keukens et al., 1992, 1995). Keukens et al. (1995) proposed that on the one side the length of the sugar chain of the GA and on the other side a planar ring structure, a 3β‐OH‐group as well as a side‐chain at position 24 of the sterol, are critical. As a consequence of this interaction, the permeability, stability, flexibility and fluidity of cell membranes may be considerably altered (Friedman, 2006).
Manrique‐Moreno et al. (2014) showed that the aubergine‐derived aglycone solasodine may cause structural perturbations in the membranes of human erythrocytes. In more detail, X‐ray diffraction studies demonstrated that solasodine interacted mainly with dimyristoylphosphatidylcholine, one of the major phospholipids in the outer and inner erythrocyte membrane affecting the polar head and acyl chain regions of the molecule. Interestingly, the strongest effect was seen with the aglycone solasodine and the lowest with the GA α‐solanine. It has not been investigated whether the potato aglycone soladinine exerts similar effects in phospholipids as solasodine.
The disintegrating effect of GA on membranes may be of significance in the gastrointestinal tract after enteral uptake of the compounds. For example, Gee et al. (1996) reported that GAs reduced the integrity of isolated rat mucosa, which impaired active nutrient transport and enabled high molecular weight compounds (such as allergens or GAs) to cross the epithelial barrier. The communication between intestinal cells by gap junctions may also be a target of GAs, as observed in Caco‐2 human epithelial colon carcinoma cells after treatment with α‐chaconine and α‐solanine (Keukens et al., 1996; Glynn et al., 2017). GAs also disturb the transport of Ca2+ and Na+ across cell membranes. For example, Blankemeyer et al. (1995, 1997) found that the transepithelial active transport of sodium decreased considerably when incubating frog skin with α‐tomatine, α‐chaconine or α‐solanine. In the rat duodenum, 5 mM α‐solanine applied via drinking water, inhibited the active calcium transport in a non‐competitive way (Michalska et al., 1985). The extent of these effects may depend on the pH within the intestinal lumen, e.g. GAs show a low solubility at neutral pH and a higher solubility around a pH of 6, as occurring in the proximal jejunum (Gee et al., 1996). However, it has to be considered that the intestinal mucosa in the intact organism of healthy individuals is protected by a layer of mucus, which may antagonise the effects of GAs.
Disruption of epithelial barrier integrity by GAs may be important in triggering inflammatory bowel disease in susceptible individuals. To test this hypothesis, Patel et al. (2002) treated homozygous IL‐10 gene‐deficient mice with overt enterocolitis with a 1:1 mixture of α‐chaconine:α‐solanine at 3 mg/kg bw per day via drinking water for 21 days. This treatment aggravated intestinal injury, as determined by pathohistological diagnosis, while in control animals no effect could be seen. The same group used Il‐10‐gene‐deficient male mice as well as male mice with dextran sodium sulfate‐induced colitis exposed to fried skin from potatoes with low/medium (154 mg GA/kg skin, i.e. 30.8 mg/kg bw per day) or high GA content (460 mg GA/kg skin, i.e. 92 mg/kg bw per day) for 20 days (no further feed was provided to the mice) (Iablokov et al., 2010). Exposure to the low/medium GA skins elevated interferon‐gamma (IFN‐γ) in the Il‐10‐gene‐deficient mice, and IFN‐γ, TNF‐α and IL‐17 in mice with the dextran sodium sulfate‐induced colitis. In the latter model also enhanced colonic permeability could be observed.
To conclude, the cytotoxicity in the gastrointestinal mucosa may contribute to symptoms observed after GA intoxication, such as necrosis in experimental animals or diarrhoea and vomiting in humans (see Sections 3.1.2 and 3.1.3). It remains to be elucidated in more detail whether susceptible individuals are at an elevated risk for the development of inflammatory bowel disease after ingestion of GAs.
3.1.5.2. Inhibition of cholinesterases (ChEs)
3.1.5.2.1. Comparative determination of inhibition of ChEs in vitro
ChEs have to be subdivided into two classes of enzymes: acetylcholinesterase (AChE, EC 3.1.1.721) and butyrylcholinesterase (BuChE, EC 3.1.1.821). This distinction is important because in older studies the term ChE has been used in a general sense, not discriminating between the two classes of enzymes. The BuChE class of enzymes was also called ‘pseudocholinesterase’ or in older studies, ‘unspecific ChE’, representing the serum ChE.
The primary function of ChEs is to inactivate acetylcholine (ACh) by cleavage, thereby avoiding overstimulation of cholinergic processes. These cholinolytic functions of ChEs require the catalytic (enzymatic) function of the protein. This function is required in the case of the classical synaptic function (Vogel‐Höpker et al., 2012). Within the enzymatic mode of action AChE most likely functions in cooperation with other components of the cholinergic system including the substrate ACh, both the nicotinic and muscarinic acetylcholine receptors, choline acetyltransferase, and potential ACh transporters.
Besides cleavage of ACh, BuChE is primarily known for its activity in metabolism of bioactive esters from food and medications, particularly for its role to inactivate the muscle relaxant succinylcholine, used to facilitate abdominal surgery (Brimijoin et al., 2016). Otherwise, it was not known to play a direct role in mammalian physiology and was regarded as an orphan enzyme. This changed when it was recognised that BuChE cleaves ghrelin, a hormone involved in appetite control.
Each class of enzymes can be inhibited by two typical modes of action, the competitive reversible and the non‐competitive irreversible mode of inhibition. Reversible competitive inhibition of ChEs is used for clinical purposes using substances with a positively charged nitrogen, competing for the ligand binding domain of the enzyme. Chemically, these inhibitors include compounds with different functional groups (carbamate, quaternary or tertiary ammonium group). They are/were applied for diagnostic and/or treatment purposes for diseases such as myasthenia gravis, Alzheimer disease or glaucoma.
Non‐competitive inhibitors represent largely organophosphates which are esters or thiols derived from phosphoric, phosphonic, phosphinic or phosphoramidic acid. They exert their main toxicological effects through non‐reversible phosphorylation of esterases in the central nervous system thereby irreversibly inactivate the catalytic centre of the enzyme (Čolović et al., 2013). GAs are inhibitors of ChEs and follow the reversible, competitive mode of action.
Regarding inhibition of ChEs by GAs, Pokrovskii (1956) was the first to describe inhibition of enzymatic activity by α‐solanine. The first description of inhibition of serum ChE in the English literature was by Orgell et al. (1958), who showed that a peel extract is up to 40‐fold more potent than in the inner part of the tuber. In the meantime, there is a broad range of evidence supporting the observation that GAs, particularly α‐chaconine and α‐solanine, are potent inhibitors of ChEs, and that the GAs are more potent than the aglycones.
Inhibition of ChEs by GAs was reported in two different ways, representing two different experimental scientific approaches. Some studies described percentage of inhibition obtained at tested concentration(s) of the GAs or their aglycones against control, while others determined inhibitory constants (Ki). Consequently, results are summarised in two different tables representing percentage of inhibition values (Appendix E, Table E.1) and determined Ki values (Appendix E, Table E.2). A direct comparison of the potency of inhibition of the two ChE enzyme classes is difficult, because enzymes were measured in different organs or body fluids and concentrations used to study percentage of inhibition by the investigated GA differed considerably. However, from direct comparative studies, some general findings can be delineated. The most potent GAs in term of inhibition of ChEs were α‐solanine and α‐chaconine, with α‐chaconine in tendency being slightly more potent. BuChE appears to be a slightly more sensitive target for inhibition by GAs than AChE. Human ChEs reacted equally if not more sensitive to inhibition by GAs than their counterparts from animals (see Appendix E).
Table E.1.
Inhibition of cholinesterases by GAs
| Compound | Species | AChE | BuChE | Citation | ||
|---|---|---|---|---|---|---|
| Dose | % inhibition | Dose | % inhibition | |||
| α‐Solanine | Bovine erythrocyte | 5 μM | 27.7 | Roddick (1989) | ||
| α‐Solanine | Bovine erythrocyte | 10 μM | 44.3 | |||
| α‐Chaconine | Bovine erythrocyte | 5 μM | 25.9 | |||
| α‐Chaconine | Bovine erythrocyte | 10 μM | 54.4 | |||
| α‐Solanine + α‐chaconine | Bovine erythrocyte | 5+5 μM | 37.3 | |||
| α‐Solasonine | Bovine erythrocyte | 50 μM | –12 | |||
| α‐Solasonine | Bovine erythrocyte | 100 μM | 16.7 | |||
| α‐Solamargine | Bovine erythrocyte | 50 μM | –3.6 | |||
| α‐Solamargine | Bovine erythrocyte | 100 μM | 13 | |||
| Solanindine (0 h) | Bovine erythrocyte | 100 μM | 11.1 | Roddick (1989) | ||
| Solanidine (1 h) | Bovine erythrocyte | 100 μM | 5.7 | |||
| Solanidine (2 h) | Bovine erythrocyte | 100 μM | 2.0 | |||
| Solanidine (0 h) | Human erythrocyte | 100 μM | –6.2 | |||
| Solanidine (1 h) | Human erythrocyte | 100 μM | 3.6 | |||
| Solanidine (2 h) | Human erythrocyte | 100 μM | 14.1 | |||
| Tomatidine (0 h) | Bovine erythrocyte | 100 μM | 15.6 | |||
| Tomatidine (1 h) | Bovine erythrocyte | 100 μM | –7.7 | |||
| Tomatidine (2 h) | Bovine erythrocyte | 100 μM | –4.5 | |||
| Tomatidine (0 h) | Human erythrocyte | 100 μM | –13.8 | |||
| Tomatidine (1 h) | Human erythrocyte | 100 μM | –13.8 | |||
| Tomatidine (2 h) | Human erythrocyte | 100 μM | –10.6 | |||
| Solasodine (0 h) | Bovine erythrocyte | 100 μM | 8.9 | |||
| Solasodine (1 h) | Bovine erythrocyte | 100 μM | –15 | |||
| Solasodine (2 h) | Bovine erythrocyte | 100 μM | –1.5 | |||
| Solasodine (0 h) | Human erythrocyte | 100 μM | –8.3 | |||
| Solasodine (1 h) | Human erythrocyte | 100 μM | –2.4 | |||
| Solasodine (2 h) | Human erythrocyte | 100 μM | 0.6 | |||
| α‐Chaconine | Eel | 33.8 ppm (39.6 μM) | 26.8 | Bushway et al. (1987) | ||
| β‐Chaconine | Eel | 33.8 ppm (40 μM) | 23.3 | |||
| α‐Solanine | Eel | 33.4 ppm (38.5 μM) | 26.3 | |||
| Dehydrocommersonine | Eel | 33.7 ppm (32.2 μM) | 18.5 | |||
| Commersonine | Eel | 34.5 ppm (32.9 μM) | 20.2 | |||
| Demissine | Eel | 34.3 ppm (33.6 μM) | 21.6 | |||
| α‐Tomatine | Eel | 34 ppm (32.8 μM) | 4.2 | |||
| Solanidine | Eel | 33.8 ppm (85.1 μM) | 15.4 | |||
| Demissidine | Eel | 40 ppm (100 μM) | 11.3 | |||
| Tomatidine | Eel | 41.3 ppm (99.5 μM) | 4.2 | |||
| Potato peel | Human serum (unusual phenotype) | (85 g potato peel extracted by 300 mL water) | 77–83 | Harris and Whittaker (1959) | ||
| Potato peel | Human serum (intermediate phenotype) | (85 g potato peel extracted by 300 mL water) | 53–67 | |||
| Potato peel | Human serum (atypical phenotype) | (85 g potato peel extracted by 300 mL water) | 16–24 | |||
| α‐Solanine | Human serum (unusual phenotype) | 2.88 μM | 86.2 | Harris and Whittaker (1962) | ||
| α‐Solanine | Human serum (intermediate phenotype) | 2.88 μM | 65.8 | |||
| α‐Solanine | Human serum (atypical phenotype) | 2.88 μM | 21 | |||
| Solanidine | Human serum (unusual phenotype) | 3.14 μM | 80 | |||
| Solanidine | Human serum (intermediate phenotype) | 3.14 μM | 53.2 | |||
| Solanidine | Human serum (atypical phenotype) | 3.14 μM | 0.8 | |||
| α‐Chaconine | Rat plasma isoenzyme 1 | 100 μM | 0 | Alozie et al. (1978) | ||
| α‐Chaconine | Rat plasma isoenzyme 2 | 100 μM | 0 | |||
| α‐Chaconine | Rat plasma isoenzyme 3 | 100 μM | 78.5 | |||
| α‐Chaconine | Rat plasma isoenzyme 4 | 100 μM | 81.7 | |||
| α‐Chaconine | Rat plasma isoenzyme 5 | 100 μM | 90 | |||
| α‐Chaconine | Rat plasma isoenzyme 6 | 100 μM | 100 | |||
| α‐Chaconine | Rat erythrocyte isoenzyme 1 | 100 μM | 0 | |||
| α‐Chaconine | Rat erythrocyte isoenzyme 2 | 100 μM | 42 | |||
| α‐Chaconine | Rat erythrocyte isoenzyme 3 | 100 μM | 42 | |||
| α‐Chaconine | Rat erythrocyte isoenzyme 4 | 100 μM | 100 | |||
| α‐Chaconine | Rat erythrocyte isoenzyme 5 | 100 μM | 100 | |||
| α‐Chaconine | Rat erythrocyte isoenzyme 6 | 100 μM | 100 | |||
| α‐Chaconine | Rat brain isoenzyme 1 | 100 μM | 23 | |||
| α‐Chaconine | Rat brain isoenzyme 2 | 100 μM | 27.1 | |||
| α‐Chaconine | Rat brain isoenzyme 3 | 100 μM | 15.6 | |||
| α‐Chaconine | Rat brain isoenzyme 4 | 100 μM | 19.5 | |||
| α‐Chaconine | Rat brain isoenzyme 5 | 100 μM | 67.3 | |||
| α‐Chaconine | Rat brain isoenzyme 6 | 100 μM | 100 | |||
| α‐Chaconine | Human purified | 100 μM | 67.3 | 100 μM | 92.8 | McGehee et al. (2000) |
| α‐Solanine | Human purified | 100 μM | 76.8 | 100 μM | 91.5 | |
| α‐Chaconine (0/10/30 min) | Human serum | 2.88 μM | 68.3/70.2/67.3 | Nigg et al. (1996) | ||
| α‐Solanine (0/10/30 min) | Human serum | 2.88 μM | 50/50/50 | |||
Note: In older papers, e.g. Orgell (1963), existence of BuChE was not known, data were reported as AChE activity with no discrimination made.
Table E.2.
Determined Ki values
| Compound | Species | AChE | BuChE | |
|---|---|---|---|---|
| Ki (μM) | Ki (μM) | Citation | ||
| α‐Solanine | Human plasma | 5 | Orgell (1963) | |
| α‐Tomatine | Human plasma | 100 | ||
| Chaconine (TLC preparation) | Purified bovine cholinesterase | 8,3 | Alozie et al. (1978) | |
| Horse serum cholinesterase | 4 | |||
| α‐Chaconine | equine BuChE | 1.39 | Benilova et al. (2006) | |
| α‐Solanine | equine BuChE | 3.33 | ||
| α‐Tomatine | equine BuChE | 1.66 | ||
| α‐Chaconine | human BuChE | 0.17 | ||
| α‐Solanine | human BuChE | 0.22 | ||
| α‐Tomatine | human BuChE | 1.3 | ||
| α‐Solanine | Human serum (D/D; wild type, homozygous) | 2.9 | Sternfeld et al. (1997) | |
| α‐Solanine | Human serum (D/G; heterozygous) | 3.6 | ||
| α‐Solanine | Human serum (G/G; homozygous) | 165 | ||
| α‐Solanine | Human recombinant (D/D; wild type, homozygous) | 2.0 | ||
| α‐Solanine | Human recombinant (D/G; heterozygous) | 3.5 | ||
| α‐Solanine | Human recombinant (G/G; homozygous) | 171 | ||
| α‐Solanine | Human, purified | 14 | 0.17 | McGehee et al. (2000) |
| α‐Chaconine | Human, purified | 17 | 0.066 | |
| α‐Solanine | Human wild type | 3.3 | Loewenstein‐Lichtenstein et al. (1996) | |
| α‐Solanine | Human atypical (D70G) | 78 | ||
| α‐Solanine | Various artificial mutations | 0.3–78 |
Harris and Whittaker (1959) extracted 85 g of potato peel with 300 mL water and tested these extracts on human serum ChE. By electrophoretic procedures, they could discriminate between three subtypes: the unusual phenotype, the intermediate phenotype and an atypical phenotype. The percentage of inhibition recorded was: 77–83% for the unusual phenotype, 53–67% for the intermediate phenotype and 16–24% for the atypical phenotype. Using the same discrimination of enzyme subtypes for human serum ChEs, the same authors made a comparative investigation of the inhibitory activity of α‐solanine at 2.88 μM and its aglycone solanidine at 3.14 μM (Harris and Whittaker, 1962). For α‐solanine, the authors found 86.2% inhibition for the unusual phenotype, 65.8% inhibition for the intermediate phenotype and 21% for the atypical phenotype. Inhibition by 3.14 μM solanidine was slightly lower, with 80% inhibition for the unusual phenotype, 53.2% for the intermediate phenotype and 0.8% for the atypical phenotype. Overall, the inhibitory activity of the aglycone seems to be lower than that of the glycoside. Nigg et al. (1996) did a comparative assessment of α‐chaconine and α‐solanine. They also used human serum ChE, specifically targeted BuChE using α‐naphthylacetate as substrate at a concentration of 2.88 μM and had three time points of measurement (0, 10, 30 min). Whereas the inhibitory activity for α‐solanine was 50% at all time points measured, α‐chaconine treatment lead to an inhibition of BuChE of 68.3% at time point 0, 70.2% at 10 min and 67.3% at 30 min. These results are indicative for a higher inhibitory of α‐chaconine if compared to α‐solanine.
The comparative approach of estimation of an inhibitory activity at a defined substrate concentration for both enzymes was used only once, meaning direct comparison of the inhibitory activity of α‐chaconine and α‐solanine (McGehee et al., 2000). Using human purified enzymes and substrate concentration of 100 μM, the authors determined the percentage of inhibition of AChE and BuChE. α‐Chaconine inhibited 67.3% of AChE activity and 92.8% of BuChE activity. α‐Solanine showed a similar activity, inhibiting AChE by 76.5% and BuChE by 91.5%, pointing to the fact that BuChE may be more sensitive to inhibition by GAs than AChE.
Alozie et al. (1978) performed an inhibitory assay with 100 μM of α‐chaconine on ChEs in serum, erythrocytes and brains of rats. By incubating electrophorese gels, they were able to discriminate six isoenzymes. The inhibitory activities for isoenzymes 1–6 from plasma were 0%, 0%, 78.5%, 81.7%, 90% and 100%, respectively, from erythrocytes were 0%, 42%, 42%, 100%, 100% and 100%, respectively, and from brain were 23%, 27.1%, 15.6%, 19.5%, 67.3% and 100%, respectively.
Using ChE from erythrocytes, Roddick (1989) investigated three questions: (a) to compare the relative potency of α‐chaconine and α‐solanine alone and the individual activities to those of mixtures of both compounds, (b) to compare the activity of the glycosides to those of aglycones and (c) to compare the relative potencies of the aglycones against bovine and human ChE from erythrocytes. α‐Chaconine and α‐solanine were tested at concentrations of 5 μM and 10 μM, as well as in a mixture containing 5 μM of each compound. α‐Chaconine inhibited bovine erythrocyte ChE by 25.9% (5 μM) and by 54.4% (10 μM), whereas α‐solanine inhibited by 27.7% (5 μM) and by 44.3% (10 μM). The 1:1 mixture of both compounds at 5 μM each leads to a 37.3% inhibition, indicative of an additivity of the two compounds. α‐Solasonine and solamargine were tested at 50 and 100 μM each. While 50 μM concentrations of both compounds slightly increased enzyme activity, an inhibition was detectable at 100 μM, with 16.7% inhibition for α‐solasonine and 13% inhibition for solamargine. In addition, Roddick (1989) comparatively assessed the inhibitory activity of the aglycones solanidine, tomatidine and solasodine. At 100 μM, no consistent inhibitory activity using bovine and human erythrocyte AChE was detected.
3.1.5.2.2. Determination of inhibitory constants (Ki) for GAs on inhibition of ChEs in vitro
Most data for the assessment of the Ki are available for α‐solanine. For human AChE, the range of determined Ki values stretches from a Ki of 5 μM (Orgell, 1963) to a Ki of 14 μM (McGehee et al., 2000). For bovine and equine AChE, the Ki values were determined to be 8.3 μM and 4 μM, respectively.
A lot more data are available for the Ki determination of α‐solanine for inhibition of human BuChE. The range of the determined Ki values is between a higher nM and a lower μM range. McGehee et al. (2000) described a Ki of 0.17 μM for the inhibition of human BuChE by α‐solanine. Benilova et al. (2006) determined an almost identical Ki of 0.22 μM for this enzyme using a biosensor‐based method. A similarly low Ki was also reported by Loewenstein‐Lichtenstein et al. (1996), although for an artificial mutant which was expressed recombinantly. The same authors determined a Ki of 3.3 μM for the wild‐type human BuChE. This observation is in line with the findings of Sternfeld et al. (1997) who determined a Ki of 2.9 μM for the human placental BuChE and a Ki of 2.0 μM for the recombinantly expressed human BuChE. Ki values for the inhibition of equine BuChE were found to be higher (Benilova et al., 2006).
The Ki values determined for the inhibition of BuChE by α‐chaconine were even lower than those for α‐solanine. McGehee et al. (2000) reported a Ki of 0.066 μM for the purified human enzyme, whereas Benilova et al. (2006) reported a Ki of 0.17 μM. Like for α‐solanine, human BuChE appears to react more sensitive than equine BuChE.
The data for α‐tomatine are inconclusive. Orgell (1963) reported a Ki of 100 μM for human plasma ChE, whereas Benilova et al. (2006) determined a Ki of 1.3 μM for human BuChE and 1.66 μM for equine BuChE using a biosensor method.
In summary, biochemical data on inhibition of ChEs by GAs can be summarised as follows below and as illustrated in Tables 25 and 26 (for details refer to Appendix E):
the sugar moiety is a crucial determinant of ChE inhibition, as GAs are more potent inhibitors than their aglycones (Table 25),
α‐chaconine seems to be a slightly more potent inhibitor of ChEs than α‐solanine, particularly of BuChE, and
GA exposure leads to a slightly higher inhibition of BuChE activity than of AChE activity.
Table 25.
Percent inhibition of bovine erythrocyte cholinesterase (ChE) by glycoalkaloids (GAs) and their aglycones (Roddick, 1989)
| Compound | Concentration (μM) | % Inhibitiona |
|---|---|---|
| α‐Solanine | 10 | 44.3 |
| α‐Chaconine | 10 | 54.4 |
| α‐Solanine + α‐chaconine | 5 + 5 | 37.3 |
| Solanidine, first subexperiment | 100 | 2–11 |
| Solanidine, second subexperiment | 100 | –14.1––6.2 |
% inhibition of the activity measured for the solvent control DMSO.
Table 26.
Inhibitory constants for glycoalkaloids (GAs) on human acetylcholinesterase (AChE) and human butyrylcholinesterase (BuChE)(a)
| Compound | Human AChE; Ki (μM) | Human BuChE; Ki (μM) |
|---|---|---|
| α‐Solanine | 5–14a | 0.22–3.3b |
| α‐Chaconine | 17c | 0.06–0.17d |
3.1.5.2.3. Inhibition of ChEs in vivo
Inhibition of ChEs in serum and erythrocytes
Inhibition of AChE by α‐solanine is effective not only in vitro but also in vivo, as shown by several experimental studies. When α‐solanine was applied i.p. to two rabbits of each sex, AChE activity declined over time both in serum and in erythrocytes and was lowest at time of death (range of lowest remaining activity: 55.2–73.2% in serum and 66.6–81.3% in erythrocytes). One male animal recovered. In this animal study, AChE activity remained constant between time points 140 min after treatment and 210 min after treatment in the case of serum AChE, and even recovered from 71.4% to 82.9% in the same time range in erythrocytes (Patil et al., 1972).
Inhibition of ChEs in the central nervous system
As mentioned on Section 3.1.3, in humans, acute effects of intoxication comprise sweating, vomiting, diarrhoea, severe abdominal pain, drowsiness, apathy, confusion, weakness, vision disturbances, fever, rapid and weak pulse, low blood pressure and bronchospasm in cases of milder intoxications, as well as paralysis, respiratory insufficiency, cardiac failure, coma and death in severe cases (see Section 3.1.3.1). As only cholinergic neurons mediate both pre‐ and postganglionic activities of the parasympathic nervous system, and in addition, part of the preganglionic activities in the sympathic nervous system, it appears likely that anticholinolitic activities of GAs in this part of the nervous system result in the effects observed following human intoxication. Similar symptoms have been described for non‐competitive organophosphorus inhibitors (Pereira et al., 2014). These effects may mechanistically explain the severe symptoms described for GA intoxication.
Since the 70s experimental evidence was gained that GAs may inhibit ChE also in the central nervous system. In rats, treated i.p. with 0, 10, 30, 60, 90 mg/kg bw per day of α‐chaconine, brain AChE activity dose dependently decreased at 10, 30 and 60 mg/kg bw to 79%, 55% and 18%, respectively (Alozie et al., 1978). These effects could not be related to the availability of ACh, as in Sprague Dawley rats receiving i.p. 3, 8 or 20 mg/kg bw of α‐chaconine either three or 12 h prior to sacrifice, brain levels of ACh remained unchanged (Aldous et al., 1980). In addition to neurotransmitter levels in the brain, the authors assessed neuro‐ and electrophysiological parameters related to both brain function. After i.p. injection of 10, 20 and 30 mg α‐chaconine/kg bw, both in the parietal and occipital EEGs, the slow wave activity was affected at 20 mg/kg bw per day. The disappearance of the 6 Hertz peak in parietal records and the dose‐dependent disappearance of 6 Hertz peak in occipital records occurred at/started at 10 mg/kg bw per day (Aldous et al., 1980).
Baker et al. (1988) reported on effects on brain AChE activity in Syrian hamsters gavaged with increasing amounts of lyophilised sprout material. At 300 mg/kg bw of the material, a statistically significant increase in brain AChE activity was observed, followed by significantly decreased activity at 400 and 500 mg/kg bw. This decrease was not as pronounced as the decrease seen for the organophosphate diethyl‐tri‐chloro‐phosphorothioate (Dursban 2E), which was investigated against control in a parallel experiment and acts through a non‐competitive mechanism.
Inhibition of cholinesterases in cardiac and neuromuscular transmission
The effect of α‐solanine on the electrocardiogram (ECG) and respiration was also investigated in rabbits (Nishie et al., 1971). Below a dose of 15 mg/kg i.p., no effect was detectable. Tachycardia was observed both at low (10 mg/kg bw per day) and at very high doses (40 mg/kg bw per day). Intermediate doses (20 and 30 mg/kg bw per day) lead to bradycardia. Symptoms which became apparent at low doses were sedation, respiratory impairment and constriction of the abdominal muscles which became apparent at both 8 and 10 mg/kg bw per day. In conclusion, GAs may influence heart function at high systemic levels.
McGehee et al. (2000) investigated whether GAs would impact on mivacurium‐induced neuromuscular blockage. Female and male New Zealand white rabbits received an i.v. administration of mivacurium (27 nmol/kg in 30 s), followed by an i.v. administration of 12 μmol/kg of α‐solanine or α‐chaconine for 20–60 min. A decrease in plasma BuChE, responsible for the detoxification, e.g. of cocaine, in response to infusion of the GAs resulted in elevated mivacurium levels and consequently for a prolonged time for recovery from mivacurium‐induced neuromuscular blockage. The effect of α‐chaconine was thereby more pronounced than the effect of α‐solanine.
3.1.5.2.4. Developmental and reproductive effects of GAs and their aglycones
Teratology occurred at relatively high doses of both GAs and their aglycones, and the teratogenic phenotypes observed included exencephaly, encephalocele, cebocephaly, cleft palate and anophthalmia (see Section 3.1.2.3). Experimental evidence suggests that these phenotypes in the central nervous system are presumably not, or not exclusively, caused by inhibition of the catalytic function of AChE. First, the AChE knock‐out mouse does not show a phenotype in the central nervous system. High sensitivity against BuChE inhibitors indicates that this enzyme may be able to compensate for the missing AChE (Xie et al., 2000). Comparative immunohistochemical examination between brain tissues of knock‐out and wild‐type animals not even revealed differences at the microscopic level in neuronal, dendritic, astrocytic, synaptic, microglial and endothelial tissue compartments (Rice et al., 2007). AChE knock‐out mice died around PND14 from their inability to obtain sufficient nutrients. This phenotype could be rescued by a special diet and enabled examination of older knock‐out animals. The phenotypes detected comprised body tremors, abnormal gait and posture, absent grip strength, inability to eat solid food, pinpoint pupils, decreased pain response, vocalisation and early death caused by seizures or gastrointestinal tract ileus (Duysen et al., 2002), with body tremors resembling the human situation. Second, the teratologic phenotypes were caused by both the GAs and their aglycones, the latter being much weaker inhibitors of ChE catalytic activity than the GAs or do not inhibit at all (see Appendix C). However, the papers dealing with the phenotype of the AChE −/− mice point to the hypothesis that BuChE may substitute for AChE protect from excess acetylcholine (Mesulam et al., 2002). As GAs inhibit both enzymes, still the possibility remains that inhibition of ChEs is involved in causing the teratology, neurodevelopmental phenotypes.
There is increasing notion that ChE shows also non‐cholinolytic enzymatic side activities like acylamidase activity (Vogel‐Höpker et al., 2012; Layer et al., 2013). Furthermore, the number of descriptions of non‐enzymatic roles of ChEs has increased over the years, including non‐catalytic activities of the enzymes involved in processes of neuronal and non‐neuronal developmental (Vogel‐Höpker et al., 2012). Developmental neuronal functions comprise among others, involvement of proliferative control of neuronal cells, their migration, neurite outgrowth and synaptogenesis, as well as cell adhesion. Non‐neuronal developmental functions concern association with the development of the immune system, haemopoietic stress responses, as well as of limb and bone development (Vogel‐Höpker et al., 2012; Lionetto et al., 2013). It remains to be studied whether GA and their aglycones interfere with any of these non‐catalytic functions of ChE and whether such effect could have impact on the neuronal development of the unborns.
Reduced postnatal survival of pups occurred at the relative low doses of 3.6 and 4.8 mg/kg bw per day, producing a decrease in the percentage of weaned pups, due to early death of the pups. This was presumably due to starvation as no milk was found in the stomach of these animals. Most clinically used AChE inhibitors represent competitive AChE inhibitors. They are believed to be compatible with human breast feeding, except for the use of neostigmine or combinations of neostigmine and atropine (Tsiaoussis et al., 2018). However, the use of AChE inhibitors is intended to increase the levels of Ach, and this increase in ACh may have an impact on lactation at three different levels. First, in animals, the systemic or central administration of ACh or AChE inhibitors leads a decrease in the prolactin release from the anterior pituitary gland (Grandison et al., 1974), which may be mediated through an increased oxytocin release from the neurohypophysis. This lowering of prolactin may affect the growth and lactation within the mammary gland. Second, the sustained reflex release of oxytocin during suckling leads to an increase in intramammary pressure through muscarinic and nicotinergic AChE receptor mechanisms (Clarke et al., 1978). And third, by the decreased prolactin release, the transcription of casein mRNA is decreased or eliminated, leading to a decreased synthesis of α‐lactalbumin.
Negative (or dysfunctional) effects of solasodine on fertility were accompanied by morphological alterations in the testes, the epididymis and by reduced sperm quantity and particularly quality in dogs and monkeys. The mechanisms causing these effects are largely unknown. Inhibition of ChEs as cause of this male reproductive phenotype can presumably be ruled out, because solasodine is a poor inhibitor of ChEs. Similar findings to those reported in the studies with Rhesus monkey and with orally treated dogs as described above (Dixit et al., 1989) were observed in dogs following i.p. injection of 20 mg/kg bw per day solasodine (Dixit and Gupta, 1982). In both dog studies, the authors interpreted their findings as anitiandrogenic based on the reduced sialic acid content, an androgenic response marker. This interpretation is plausible as in both studies the number of androgen‐producing Leydig cells is decreased. On the other hand, i.p. or oral treatment of animals with solasodine and testosterone propionate did not restore the testicular and epididymal phenotypes, pointing to the fact that hormonal effects may be involved in the disruption of male reproductive organ function by solasodine, but hormonal effects alone may not be sufficient to explain the observed effects entirely. Further, an effect of GAs on male reproductive organs may be compound specific. An i.p. treatment of mice with 2.5 mg/kg bw of α‐solanine every third day for 10 times did not result in a reduced testis weight, but decreased mRNA levels of cyclin D1 and of Inhibin A and B in the testis, which is in line with the hypothesis that hormonal effects contribute to the observed effects (Park et al., 2009). Finally, i.p. injection of 50 mg/kg bw per day of α‐solamargine showed no effect on the weight of the testes and epididymis or the number of spermatozoa in male Wistar rats (Al Chami et al., 2003).
In summary, α‐solanine in rats impacts on the availability of milk to the pups. Regarding the mechanisms underlying the effects of solasodine with the structure and function of male reproductive organs, little is known.
3.1.5.2.5. Inhibition of cholinesterases and effects in the immune system
Plant extracts of the peeled tuber of S. tuberosum as well as GA and aglycones exerted pronounced anti‐inflammatory effects in in vitro and in vivo experiments (as outlined in Section 3.1.2.4). With regard to the putative mode of action of these anti‐inflammatory effects, it should be noted that a tight cross‐talk between the immune system and the cholinergic neurotransmitter system has been demonstrated, the so‐called ‘anti‐inflammatory non‐neuronal cholinergic pathway’. In particular, stimulation of a specific acetylcholine‐receptor (α−7 nAChR) suppresses the release of pro‐inflammatory mediators from macrophages. The stimulation of α−7 nAChR may be due to non‐neuronal ACh, which is secreted by many cell types, such as macrophages, lymphocytes or dendritic cells. These cells also express AChE and BuChE to regulate the levels of ACh. There are reports that an increased BuChE activity may suppress the cholinergic anti‐inflammatory responses and amplify systemic inflammation, a pathogenetic mechanism discussed for multiple sclerosis and other diseases (Pohanska, 2014; Reale et al., 2018). It may be speculated that the GA‐induced lowered BuChE activity may result in an anti‐inflammatory outcome. The mode of action underlying the anti‐inflammatory effect of the aglycones, with weak impact on BuChE activity, remains to be clarified.
3.1.5.3. Interference with metabolism
Elkahoui et al. (2018) tested whether dietary supplementation with potato peel powder would impact on body weight gain in mice on high fat diet containing 25% of lard. The powder was prepared from four different potato cultivars with well‐defined amounts of α‐solanine and α‐chaconine. The isocaloric and isonitrogenous peel‐containing diets dose dependently reduced both weight gain and epididymal adipose tissue in male C57B1/6J mice, although the food consumption remained unaltered. Similar observations were reported for α‐solanine (Gull, 1960) or α‐chaconine (Friedman et al., 1996; Langkilde et al., 2008). The reasons for the lowered gain in body weights in GA‐treated rodents are presently unclear.
Recently, it was discovered that BuChE acts also as an esterase of ghrelin (Brimijoin et al., 2016, 2018). Ghrelin is an orexigenic hormone, produced by the pancreas and mucosa of the stomach and stimulating the appetite via inducing the secretion of neuropeptide Y in the hypothalamus. Immediately after food intake, ghrelin is degraded into smaller peptide fragments, involving the activity of BuChE. Ghrelin plays a role also in the regulation of glucose homeostasis through the modulation of insulin secretion and insulin sensitivity (Heppner et al., 2011). It is presently unclear whether BuChE inhibition by GAs may affect food intake, body weight and the metabolism via altered ghrelin levels and/or impact on insulin sensitivity.
There is experimental evidence that dietary α‐tomatine forms a non‐absorbable complex with cholesterol and other sterols in the gut (see Sections 3.1.2.2 and 3.1.2.9). It appears likely that the underlying mechanisms are similar to those described in Section 3.1.5.1. The insoluble complexes in the intestinal lumen are excreted via feces. Accordingly, an α‐tomatine‐containing diet elevated the fecal elimination of cholesterol and sterols in rats and hamsters (Friedman et al., 2000a). While α‐tomatine did not affect bile acid secretion in the rats, in hamsters there were significant increases in fecal deoxycholic and lithocholic acid content after treatment with green and/or red tomatoes (Friedman et al., 2000b). This supports that α‐tomatine interferes with enterohepatic recirculation of sterols.
In rats, the increased fecal efflux and the concomitant decreased uptake of dietary cholesterol by the liver was counteracted by an increased de novo synthesis of hepatic and intestinal cholesterol (Cayen, 1971). Nevertheless, plasma HDL‐cholesterol and triglycerides were reduced when rats received α‐tomatine via diet for a period of 14 days. α‐Tomatine also lowered plasma levels of LDL‐cholesterol and triglycerides in hamsters, while HDL cholesterol remained unchanged (Friedman et al., 2000a,b).
To conclude, α‐tomatine reduces enteral absorption of cholesterol and serum cholesterol levels in rodents. However, it remains to be studied whether similar effects of α‐tomatine or other GAs are to be expected in humans.
3.1.6. Considerations of critical effects and dose–response analysis for the human risk assessment
3.1.6.1. GAs from edible parts of S. tuberosum
3.1.6.1.1. Considerations of critical effects and dose‐response analysis
Acute toxicity
In humans, the limited human data available from kinetic studies in volunteers (2–3 per treatment) showed that potato TGA (50% α‐solanine; 50% α‐chaconine) administered as a solution at 0.30, 0.50 or 0.70 mg/kg bw p.o. did not elicit symptoms. Gastric discomfort and nausea may occur within ~ 30 min after ingestion of ~1 mg potato TGA/kg bw contained in a potato meal (see Section 3.1.3.1, in particular Table 20). Subsequently, the volunteers developed diarrhoea and started to vomit. A number of reports on outbreaks and cases of intoxications substantiate that acute toxic effects may occur in adults from a total potato GAs intake of 1 mg/kg bw or more. In severe cases of intoxications hallucinations, paralysis, respiratory insufficiency and cardiac failure have been reported. Doses in the range of 3–6 mg potato TGAs/kg bw are considered to be potentially lethal for humans. Results from limited volunteer studies suggest differences in the human population with respect to the individual susceptibility towards adverse effects associated with the intake of potato GAs.
In experimental animals, no lethality was observed in mice at a single dose of 1,000 mg α‐solanine/kg bw p.o., and no effects at 500 mg of solanidine/kg bw p.o. In rats, no adverse effects were reported for a single oral application of 250 mg/kg bw of α‐solanine. Diarrhoea occurred after application of α‐solanine/α‐chaconine mixtures at 300 mg/kg bw and above (see Section 3.1.2.1). This indicates acute toxicity of GAs at doses above 250 mg/kg bw in rats and mice with diarrhoea being the most frequent symptom.
The acute symptoms, described in humans and experimental animals, may be due to the ability of α‐solanine and α‐chaconine to complex with membrane sterols, thereby causing disruption and loss of integrity of cell membranes. Deletion of one or more mono‐saccharides from the GAs resulted in reduced activity. After oral exposure of potato GAs, these effects may lead to local irritation of the mucosa of the gastrointestinal tract leading to gastric discomfort, vomiting and diarrhoea. The adverse effects may also be due to the inhibitory effect of α‐solanine and α‐chaconine on the activity of AChE. The aglycone solanidine exerted almost no inhibitory effects, suggesting that the sugar side chain is obligatory for inhibiting the AChE activity. As a consequence of the impaired termination of the cholinergic transmission at the neuronal and neuromuscular junctions, intoxications with α‐solanine and α‐chaconine might lead to the development also of a cholinergic toxidrome (see Section 3.1.5), with diarrhoea and vomiting being predominant. Further symptoms described in case reports and outbreaks may have developed secondary to dehydration following vomiting and diarrhoea, such as low blood pressure, dizziness, cardio‐respiratory insufficiency or coma. However, some prototypical symptoms of a cholinergic toxidrome that may not develop secondary to dehydration, such as miosis and fasciculations of the skeletal musculature, have not been reported so far.
In mice, the aglycone solanidine exerted no effect at 500 mg (1.26 mM)/kg bw i.p., contrasting to the pronounced toxicity of GAs after the identical route of exposure, i.e. LD50 of α‐solanine was 42 mg (48.3 μM)/kg bw, and of α‐chaconine was 32.3 mg (37.9 μM)/kg bw. The relative potency of toxicity of α‐solanine and α‐chaconine appear to be similar also in rats and rabbits, e.g. in rats LD50 for α‐solanine and α‐chaconine were reported to be 67 mg (77.2 μM)/kg bw and 84 mg (98.6 μM)/kg bw, respectively. These findings are substantiated by (i) a similar potency of α‐solanine and α‐chaconine to interact with sterols and disintegrate cellular membranes and act as inhibitors of AChE activity and (ii) the weak effects of the aglycone with regard to membrane disruption and AchE inhibition. Based on these experimental findings, the CONTAM Panel considered that (i) the aglycone solanidine is of lower acute toxicity than the respective GAs and that (ii) there is the potential for additivity of α‐solanine and α‐chaconine, recognising that they share common modes of action and exert similar adverse effects. Therefore, the CONTAM Panel performed the risk assessment on the basis of the total content of GAs (TGA) in potatoes comprising α‐solanine and α‐chaconine, assuming equal potency.
When comparing experimental rat and human data on acute oral toxicity of potato TGAs, it may be deduced that:
in humans, gastric discomfort and nausea occurred rapidly after ingestion of ~ 1 mg/kg bw, followed by vomiting and diarrhoea. Studies in volunteers give some indication on differing individual susceptibilities towards adverse effects of GAs,
gastric discomfort and vomiting are symptoms not covered by bioassays in rats. In this species, the most frequent symptom is diarrhoea occurring at a single oral potato TGA application of 300 mg/kg bw,
based on the likelihood to develop diarrhoea after ingestion of potato TGA, humans show an approximately 300‐fold higher susceptibility than rats.
The reasons for these marked species differences are presently unclear. It may relate to anatomical, histological and/or functional features of the gastrointestinal tract differing considerably between humans and rodents, e.g. the unique murine non‐glandular forestomach, an intragastric pH around 3–4 in rodents and around 1 in humans, and a relatively long small and large intestine and a large caecum in rats and mice. These factors may affect the local concentration of the GA in the gastrointestinal tract and the vulnerability of the mucosa (Hugenholtz and de Vos, 2018). The differences in susceptibility may also be due to a relatively high oral bioavailability and low clearance rates of potato TGA in humans when compared to rodents. Furthermore, it is not well documented whether there are differences in the sensitivity of human and rat AChE towards the inhibitory effects of GAs.
Due to these uncertainties, the CONTAM Panel considered that the use of rodent data on acute toxicity was not appropriate to establish a reference point for acute exposure in humans.
Based on human data on case reports, outbreaks and studies in volunteers (see Section 3.1.3.1, in particular Table 20), the CONTAM Panel selected the LOAEL of 1 mg potato TGA/kg bw per day as the reference point for evaluating the risk following acute oral exposure.
Repeated dose toxicity
In humans, no evidence of health problems associated with repeated or long‐term intake of GAs via potatoes has been identified. Thus, the CONTAM Panel was not able to identify critical chronic effects in humans.
In experimental animals, toxicological studies have been carried out in rodents and monkeys using different experimental designs with administration of different doses and with durations of up to 90 days. However, no long‐term chronic toxicity or carcinogenicity study for GAs or their aglycones could be identified.
The purity of the GA preparations tested varied considerably. Furthermore, a number of studies were performed according to non‐standard experimental protocols and do not provide results that are suitable for risk assessment purposes.
In repeated oral dose toxicity studies, the main effects for α‐solanine and α‐chaconine were reduced body weight gain (accompanied by unaltered or, as reported occasionally, reduced feed intake) and decreased absolute and relative liver weight. Solanidine lowered also food intake and body weight gain but elevated relative and absolute liver weights.
In hamsters, mortality was reported after oral administration of 100 mg/kg bw per day of α‐solanine or α‐chaconine given for 5 days. In rats, an oral dose of 90 mg α‐solanine/kg bw per day applied over 5 weeks did not cause any effect. Mice exhibited reduced body and absolute liver weights at 416.6 mg α‐solanine/kg bw per day, and reduced body and absolute and relative liver weights at 409 mg α‐chaconine/kg bw per day, applied over 7 days. These findings indicate that hamsters may be more sensitive towards the adverse effects of α‐solanine and α‐chaconine than mice and rats. This is substantiated by a marked species difference in the toxicokinetics of the GAs, with hamsters showing higher oral bioavailability and lower clearance rates of GAs than rats (see Section 3.1.1.1).
Relative potency of oral toxicity of α‐solanine and α‐chaconine can be partly deduced from two independent studies with durations of up to 7 days. Identical doses of these two GAs elicited comparable effects (i.e. reducing the body weight) at a similar frequency in the two species tested (mouse and hamster). These findings are supported by a similar potency of α‐solanine and α‐chaconine with regard to acute toxicity. Oral administration of solanidine did not lower but increased the absolute and relative murine liver weight, which is some indication that the adverse effects in repeated dose studies may differ between the GA and the respective aglycones.
A NOEL for potato TGA of 33.3 mg/kg bw per day was identified by Langkilde et al. (2012). This study was performed in hamsters over a period of 90 days, the longest duration of all studies identified. The animals were treated via diet, thus accounting for dietary matrix effects and simulating GA uptake via food. However, the CONTAM Panel noted several limitations of the study: the animals were dosed with freeze‐dried powder of two different potato cultivars, partly spiked with commercial standards of α‐solanine and α‐chaconine to obtain different ratios between α‐solanine and α‐chaconine from 1:3.1 to 1:70. To obtain these highly variable ratios, different proportions of potato powder of the two cultivars were admixed to the chow, creating considerable nutritional imbalances between the treatment and the control groups (for details, see Section 3.1.2.2.1). In addition, the authors did not state whether the potato powder contained trypsin inhibitor activity or not. All other studies identified were either not on hamster, of short duration and/or applied GAs via bolus not mimicking intake of GAs via the diet.
For the aglycone solanidine, there are only two toxicity studies in female mice over a period of 28 days. It is considered that the sugar moieties of the ingested α‐solanine and α‐chaconine are cleaved off during metabolism and that any toxicity testing of the two GAs will cover the adverse effects of solanidine. Therefore, the CONTAM Panel considered it was not appropriate to identify an additional reference point for a human risk assessment of solanidine.
To conclude, the data available for repeated dose toxicity are not sufficient to identify a reference point for chronic exposure to potato GAs.
Genotoxicity
Several assays were performed to test for mutagenicity and clastogenicity of potato GAs, the aglycone solanidine, as well as extracts of S. tuberosum. No genotoxicity was observed in the available tests. However, some tests provided limited information (see Section 3.1.2.7, Table 18). Overall, the CONTAM Panel concluded that there is no evidence of genotoxicity for the potato GAs α‐solanine and α‐chaconine, and for the aglycone solanidine, and that there is not sufficient information to conclude on the genotoxic potential of these GAs.
Developmental toxicity
Developmental toxicity studies in experimental animals are limited in number and quality and show effects of potato GAs at high oral doses. In hamsters, severe malformations, affecting mainly the central nervous system, occurred in offspring when dams received 200 mg α‐solanine/kg bw or 165 mg α‐chaconine/kg bw per day on GD7.5/8. The teratogenic phenotypes comprised exencephaly, encephalocele, cebocephaly, cleft palate and anophthalmia. Interestingly, the aglycones solanidine and demissidine caused also malformations in the central nervous system in hamsters. Exencephaly, encephalocele and/or anophthalmia occurred in offspring at maternal doses of 176 mg demissidine/kg bw or 115 mg solanidine/kg bw applied orally on GD8 (see Section 3.1.2.3).
There are no dose‐response studies, no indications that species other than hamster may be affected, and no mechanistic explanations for the malformations described above.
Several epidemiological studies failed to provide evidence that the intake of potato GAs may be an aetiological factor in the pathogenesis of NTD. Two studies indicated a possible association between maternal consumption of sprouted potatoes during the periconceptional phase and an elevated risk for NTDs or orofacial clefts. However, incomplete reporting impairs any conclusion on causal relationship of GA exposure and developmental effects from these studies.
3.1.6.1.2. Derivation of a health‐based guidance value (HBGV) or margin of exposure (MOE) approach
Based on the available information, the CONTAM Panel considered the LOAEL of 1 mg potato TGA/kg bw per day based on human data from case reports, outbreaks and studies in volunteers (see Section 3.1.3.1, in particular Table 20), as the reference point for evaluating the risk following acute oral exposure.
The available data on acute toxicity were considered insufficient to establish an HBGV, and instead the Panel used the margin of exposure (MOE) approach to assess a possible health concern from acute exposure to potato TGAs via food.
An MOE higher than 10 indicates that there is no health concern. This MOE of 10 takes into account the interindividual variability in toxicodynamics (a factor of 3.2), and extrapolation from a LOAEL to a NOAEL (a factor of 3). As outlined in Section 3.1.6.1.1, the CONTAM Panel considered that the critical effects of acute exposure to potato TGAs may be mainly due to the local effects in the gastrointestinal tract, following probably local irritation rather than inhibition of AChE activity. Thus, the possible interindividual variability in toxicodynamics is more relevant for the evaluation of the MOE than the interindividual variability in toxicokinetics.
No reference point could be identified to characterise the risk following chronic exposure.
3.1.7. GAs from edible parts of food plants other than S. tuberosum
3.1.7.1. Considerations of critical effects and dose‐response analysis
With respect to GAs from food plants other than S. tuberosum, reports of intoxications or outbreaks, on the effects of acute or chronic ingestion of tomato‐ or aubergine‐derived GAs or aglycones or relevant epidemiological studies are missing.
In experimental animals, acute and repeated dose toxicity studies on relatively high doses of tomato GAs showed mainly reduced absorption of cholesterol and bile acids from the gastrointestinal tract and lowered blood lipid levels.
With regard to the mode of action, tomato and aubergine GAs interfere with sterols in cell membranes and/or the lumen of the gastrointestinal tract, which may impair the resorption and enterohepatic recirculation of cholesterol and bile acids. The inhibitory effect on ChE activity is much weaker for tomato and aubergine GAs than for potato GAs.
Experimental developmental toxicity studies indicate that in hamster severe malformation (mainly of the central nervous system) occurred in offspring, when dams received doses of 1,400 mg of solasodine or dihydrosolasodine per kg bw per day on GD8. However, dose‐response studies, studies in other experimental species, studies covering not only one or few days of the gestation period and mechanistic studies are missing.
Adverse effects on male fertility (decreased epididymal weight and cauda epididymal epithelial height, lack of sperm in epididymal lumen) occurred when dogs or rhesus monkeys received the aglycone solasodine orally at 80 or 100 mg/kg bw per day, respectively.
From the limited number of studies available, there was no evidence for genotoxicity of the aubergine GA, α‐solamargine. However, there is not sufficient information to conclude on the genotoxic potential of this GA.
No data for determining a reference point for GAs or aglycones occurring in edible parts of food plants other than S. tuberosum are available.
3.1.7.2. Consideration of critical effects and dose‐response analysis for the farm animal, horses and companion animals risk assessment
The CONTAM Panel reviewed the available studies in order to identify a NOAEL or LOAEL for farm animals, horses and companion animals.
No studies were identified in horses, companion animals (cats and dogs) or fur animals.
The few studies identified on the adverse effects of GAs in ruminants, pigs, poultry and fish could not be used for the identification of a reference point.
For rabbits, no studies have been located other than those described under acute toxicity (i.p or i.v.) and repeated dose toxicity (see Sections 3.1.2.1 and 3.1.2.2). No reference point could be identified.
3.2. Occurrence data
3.2.1. Occurrence data submitted to EFSA
Occurrence data on food
An initial number of 1,300 results for 651 food samples analysed on GA from three European countries were available for the assessment. The number of results and samples submitted for each food category are provided in Table A.2 of Annex A. The major contributor of data on GA in terms of number of results was Germany (73% of the results) while the Netherlands and Sweden contributed with 21% and 6%, respectively. Data were reported on samples collected between the years 2005 and 2017. Only data on GAs in potato or potato products were reported. No data on GAs present in tomato or aubergine or products were reported.
Free text information was analysed to better specify the food category to which the sample was referring. The number of samples and results available for each food category is shown in Table 27.
Table 27.
Number of samples and results analysed for the glycoalkaloids (GAs) content, submitted to EFSA through the continuous call for annual collection of chemical contaminant occurrence data in food
| Foodex category | Results | Samples |
|---|---|---|
| Potatoes and potatoes products | 14 | 7 |
| New potatoes | 171 | 86 |
| Main‐crop potatoes | 1,069 | 535 |
| Starch potato | 2 | 1 |
| Potato flakes | 2 | 1 |
| Mashed potato powder | 16 | 8 |
| Ready‐to‐eat meal for infants and young children | 6 | 3 |
| Ready‐to‐eat meal for children, vegetable‐based | 4 | 2 |
| Ready‐to‐eat meal for children, cereal‐based | 2 | 1 |
| Ready‐to‐eat meal for children, meat/fish‐based | 14 | 7 |
| Total | 1,300 | 651 |
All samples were analysed for both α‐solanine and α‐chaconine besides two samples that were analysed only for α‐solanine and were excluded from the final cleaned data set as the total GA content (sum of α‐solanine and α‐chaconine) was used for the exposure assessment.
A total of 86 samples were available for potatoes categorised as ‘New potatoes’, while 535 samples were categorised as ‘Main‐crop potatoes’, with occurrence data to be similar. However, the CONTAM Panel considered the distinction between ‘New potatoes’ and ‘Main‐crop potatoes’ to be imprecise and decided to combine the occurrence data from both categories. In the following text, samples from these two categories will be referred to as samples from main‐crop potatoes
A total of 29 samples referred to potato products including only few food categories plus a generic ‘Potatoes and potatoes product’, thus not covering the variety of composite food categories containing potatoes in the Consumption Database. Based on this, the CONTAM Panel decided that the best approach for the exposure assessment would be to use the occurrence values in the RPC, main‐crop potatoes (combined with the new potatoes as described above) and the RPC Consumption Database (see Section 2.3.1).
Two samples were removed because they were reported as sampled based on ‘Suspect sampling’.
In addition, seven main‐crop potato samples were excluded as they were reported as ‘processed’, thus not relevant for the calculation of the occurrence in the raw primary commodity.
Information about the potato cultivar was available for 210 samples and only few samples were available for each cultivar. Information on the cultivar is also not reported in the Consumption Database, thus the CONTAM Panel considered any analysis based on possible difference among cultivars not feasible. One sample referring to starch potato cultivar was excluded.
Information about the growing method (available for only 86 samples) was not taken into account in the analysis for the same reasons.
No information was available on the length of storage between harvest and the date of sampling to allow a comparison of the occurrence data based on the storage length.
Specific information about the peeling was available for 33 samples. Of these, only seven were reported as ‘peeled’ potatoes. However, most of the samples were reported explicitly as ‘unprocessed’. The CONTAM Panel assumed samples for which no information about the peeling nor the processing was available (131 samples), to be unpeeled and they were included in the final data set. The seven samples of peeled potatoes were excluded. An overview of the excluded samples by each exclusion criteria is given in Table A.3 of Annex A.
The occurrence data of total GAs (sum of α‐chaconine and α‐solanine) in unpeeled main‐crop potatoes were used for the exposure assessment. The final data set contained 604 samples referring to unpeeled potatoes and individual occurrence values are available in Annex A.4. A summary of the available samples by country and year is provided in Table 28. Summary statistics of the occurrence UB values for main‐crop potatoes (merged with new‐potatoes) in the final data set used in the assessment and for the other food categories for which data were available are provided in Table 29.
Table 28.
Number of samples by country and year of sampling in the final GAs occurrence data set
| Country/Year | 2005 | 2007 | 2015 | 2016 | 2017 | Total by country |
|---|---|---|---|---|---|---|
| Germany | 222 | 8 | 8 | 9 | 185 | 432 |
| Netherlands | – | – | 81 | 52 | – | 133 |
| Sweden | – | – | 35 | – | 4 | 39 |
| Total by year | 222 | 8 | 124 | 61 | 189 | 604 |
Table 29.
Summary statistics of the total potato glycoalkaloid (potato TGA) concentrations (mg/kg, upper bound) for ‘Main‐crop potatoes’ (merged with ‘New potatoes’) in the final data set used in the assessment and for the other food categories for which data were available
| Food category | N | Mean | Min | Max | P95a |
|---|---|---|---|---|---|
| Main‐crop potatoes (including new potatoes) | 604 | 51.2 | 1.1 | 276.6 | 116.8 |
| Potatoes and potato products | 7 | 214.7 | 85.0 | 381.3 | – |
| Potato flakes | 1 | 95.6 | – | – | – |
| Mashed potato powder | 8 | 73.9 | 14.0 | 205.0 | – |
| Ready‐to‐eat meal for infants and young children | 3 | 0.8 | 0.1 | 1.4 | – |
| Ready‐to‐eat meal for children, vegetable‐based | 2 | 3.3 | 2.0 | 4.6 | – |
| Ready‐to‐eat meal for children, cereal‐based | 1 | 0.1 | – | – | – |
| Ready‐to‐eat meal for children, meat/fish‐based | 7 | 1.1 | 0.1 | 2.7 | – |
The 95th percentiles obtained on occurrence data with fewer than 60 analytical results may not be statistically robust (EFSA, 2011b) and are therefore not reported in the table.
The mean UB occurrence in the RPC main‐crop potatoes was 51.2 mg/kg with a P95 of 116.8 mg/kg. The minimum and maximum reported concentrations were 1.1 mg/kg and 276.6 mg/kg, respectively.
As described in Section 2.5, processing factors for peeling and heat processing were applied to these occurrence data.
Distribution of α‐solanine and α‐chaconine in the food samples
The samples in the final data set were analysed for the distribution between α‐solanine and α‐chaconine, using the UB results. In Figure 4, the results are expressed as ratio α‐chaconine to α‐solanine, while in Figure 5, the results are expressed as the contribution of α‐chaconine to the total GA content (sum of α‐solanine and α‐chaconine) in the potato samples. From Figures 4 and 5, it is evident that there is a wide range reported for the ratio of α‐chaconine to α‐solanine. The ratio reported ranges from 0.02 to 7.4, equivalent to a relative contribution of α‐chaconine to the TGA ranging from 1.5% to 88.1%. The average ratio of α‐chaconine to α‐solanine is 1.39 and the median is 1.28. The average contribution of α‐chaconine to the total is 51.5% and the median contribution is 56.1%, showing a slight overall preference for α‐chaconine in the samples. In 52.6% of the samples, α‐chaconine contributes between 50% and 70% to the TGA. The average ratio between α‐solanine and α‐chaconine was found to differ between data submissions, which may in part be related to the different analytical methods used by the laboratories.
Figure 4.
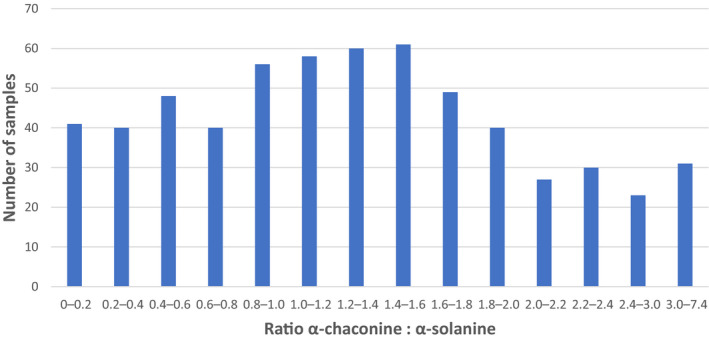
Ratio of α‐chaconine to α‐solanine in potato samples present in the EFSA occurrence database (N = 604)
Figure 5.
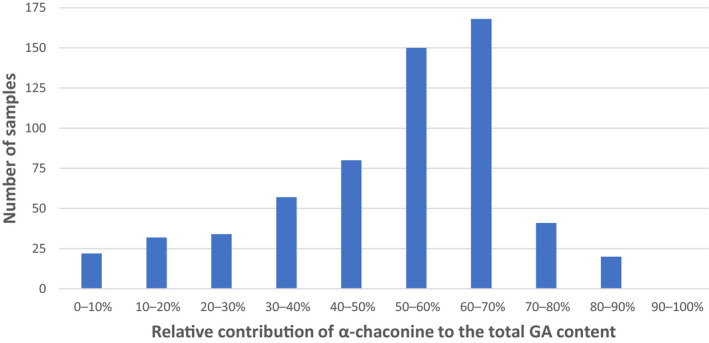
Relative contribution of α‐chaconine to the total glycoalkaloid (GA) content in potato samples present in the EFSA occurrence database (N = 604)
It was noted by the CONTAM Panel that in the database, there are 22 samples with a very low α‐chaconine content compared to the α‐solanine content (Figure 5) while there are no samples with a similarly low content of α‐solanine. This is related to the fact that in 22 samples, α‐chaconine was not detected above the LOQ of the method, while this was the case for α‐solanine in only four samples. In these cases, the UB value (LOQ) of the method is used. It should be noted that there is some evidence that α‐chaconine is more susceptible to enzymatic degradation (producing β2‐chaconine and solanidine) than α‐solanine (Swain et al., 1978; Friedman and McDonald, 1995). Partial degradation of α‐chaconine may occur during sample preparation and analysis unless specific conditions are applied (Friedman et al., 1997; see also Section 1.3.2.1). For the data sets submitted to EFSA, it could not be determined to what extent the results for individual samples could be affected by enzymatic hydrolysis. This would require detailed information on the sample preparation and analytical method used by the laboratory, and this is not part of the information requested in the SSD format for data submission.
Additional occurrence data received
The European Starch Industry Association submitted data to EFSA concerning 1,728 samples including samples on dietary fibre and potato proteins, pulp, juice and starch and on potatoes used for starch production (Table 30). Detailed results are available in Annex A.5.
Table 30.
Number of samples submitted by the European Starch Industry Association by product type and country of sampling for which total glycoalkaloid (GA) occurrence was available
| Product/Country | DK | FI | FR | DE | NL | SE | Total |
|---|---|---|---|---|---|---|---|
| Dietary fibre | 6 | – | – | 40 | – | 7 | 53 |
| Isolated proteins and other protein products | – | – | – | – | 505 | – | 505 |
| Potato juice, concentrated (feed) | – | – | – | – | 27 | – | 27 |
| Potato protein (feed) | 25 | 1 | 45 | 55 | 415 | 28 | 569 |
| Potato pulp (feed) | – | – | 9 | – | – | – | 9 |
| Potato starch | – | 1 | 10 | 49 | 41 | – | 101 |
| Potatoes (for starch production) | – | – | – | – | 450 | – | 450 |
| Total | 31 | 2 | 64 | 144 | 1,438 | 35 | 1,714 |
DK: Denmark; FI: Finland; FR: France: DE: Germany; NL: the Netherlands; SE: Sweden.
Note: Fourteen samples reporting on only one GA were excluded from the table.
Due to uncertainty on the occurrence data reported (e.g. it was not known if the occurrence data referred to dry or wet weight and it was not always known if the samples referred to feed or food for human consumption) and the difficulties expressed by the data provider in retrieving this information, these data were not included in the exposure assessment. Mean occurrence values for each category are provided in Annex A. Values should be taken with caution as individual values might have referred to different concentration expressions (information on the water content of the samples was not provided, data may reflect results based on dry vs. wet weight).
3.2.2. Previously reported occurrence data in the open literature
3.2.2.1. Literature on occurrence data on food
3.2.2.1.1. Occurrence data on GAs in potatoes
Occurrence data on GAs in potato samples obtained from field studies
In this section, studies are discussed that focussed on the effect of pre‐harvest factors on the GA content of potatoes, such as the genotype, year‐to‐year variability, location, altitude, environmental conditions and farming system under controlled field conditions. In Table 31, an overview of the studies that have investigated the TGA content of potatoes under field conditions is presented. The studies cover the effects on the TGA content of cultivars grown on different locations, in different years and with different production methods (e.g. organic, integrated or conventional). The studies which investigated the effects of location and environmental conditions are described, while those studies focussing mainly on the effect of the farming system are discussed under a separate heading. In addition, data from the registration and authorisation of seed potatoes bred in the Netherlands are presented, that includes the measurement of TGA levels under field conditions.
Table 31.
Summary results of field studies conducted on total glycoalkaloid (TGA) concentrations in potatoes
| Country | Year of survey | Cultivars/location/year/farming system | N | TGA average (mg/kg fw) | Range (mg/kg fw) | Ratio α‐chaconine: α‐solanine | Analytical technique | Reference |
|---|---|---|---|---|---|---|---|---|
| CZ | 1993–1994 | 11 cultivars, 3 locations, 2 years | 66 | 74 | 31–166 | n.r. | HPLC‐UV | Panovská et al. (1997) |
| 1996–1999 | 8 cultivars, 2 locations, 4 years, 2 systems | 82 | 70 | 15–245 | n.r. | HPLC‐UV | Hajšlová et al. (2005) | |
| 2006–2008 | 5 cultivars, 2 locations, 3 years, 2 systems | 120 | 87 | 49–134 | 1.26–2.52 | LC‐MS/MS | Bártová et al. (2013) | |
| 2009–2011 | 14 cultivars, 2 locations, 3 years | 82 | 54 | 19–102 | n.r. | LC‐MS/MS | Hamouz et al. (2014) | |
| 2012–2014 | 16 cultivars, 2 locations, 3 years | 96 | 81 | 34–168 | 1.18–3.78 | LC‐MS/MS | Urban et al. (2018) | |
| DE | 1970–1974 | 10 cultivars, 5 locations, 5 years | 184 | 29 | 3–92 | n.d. | Colourimetric | Ross et al. (1978) |
| HU | 2002–2004 | 10 cultivars, 1–3 years | 20 | 33 | 1–150 | n.r. | HPLC‐UV | Tömösközi‐Farkas et al. (2006) |
| 2007–2009 | 3 cultivars, 3 years, 2 systems | 18 | 43 | < 1–281 | n.r. | HPLC‐UV | Tömosközi‐Farkas et al. (2014) | |
| IT | < 2006 | 9 cultivars, 1 yeara | 9 | 32 | 10–51 | 2.91–5.93 | HPLC‐UV | Finotti et al. (2006) |
| < 2010 | 9 cultivars, 2 locations, 1 year | 18 | 95 | 50–209 | n.r. | LC‐MS | Ieri et al. (2011) | |
| LV | 2010–2011 | 20 cultivars, 1 location, 1 year, 2 systems | 40 | 21 | 3–104 | 0.30–4.60 | LC‐MS | Skrabule et al. (2013) |
| NL | 1985–1986 | 3 cultivars, 2 locations, 3 years | 60 | 81 | 40–146 | n.r. | HPLC‐UV | Van Gelder et al. (1988) |
| Ongoing | All cultivarsb | 464 | 48 | 3–152 | n.r. | HPLC‐UV | Website NAK | |
| PT | 2003 | 2 cultivars, 1 location, 1 year, 3 systems | 6 | 51 | 37–80 | 0.81–1.82 | HPLC‐UV | Abreu et al. (2007) |
| SE | 1986–1987 | 6 cultivars, 3 locations, 2 years | 12 | 120 | 51–210 | n.r. | HPLC‐UV | Hellenäs et al. (1995a) |
| 2009 | 21 cultivars, 1 location, 1 year | 39 | 127 | 46–293 | 1.09–2.67 | HPLC‐UV | Petterson et al. (2013) | |
| UK | 1980–1982 | 36 cultivars, 7 locations, 3 yearsc | 141 | 71 | 36–175 | n.d. | Colourimetric | Bintcliffe et al. (1982); Parnell et al. (1984) |
| 1994–1995 | 2 cultivars, 1 location, 2 years | 4 | 87 | 40–163 | 2.32‐3.08 | HPLC‐UV | Papathanasiou et al. (1999) | |
| USA | 1970–1971 | 5 cultivars, 39 locations, 2 yearsd | 255 | 79 | 14–387 | n.d. | Colourimetric | Sinden and Webb (1972, 1974) |
| < 2003 | 8 cultivars, 1 location, 1 year | 8 | 52 | 7–187 | 1.4–2.2 | HPLC‐UV | Friedman et al. (2003b) |
TGA: total glycoalkaloid; UV: ultraviolet. LC: liquid chromatography; MS: mass spectrometry; MS/MS: tandem MS; n.r.; not reported; n.d.; not determined.
From wholesale.
For details, see Figure 6.
The publication by Bintcliffe et al. (1982) reports the results for 1980–1981.
The variety B5146‐6 was excluded.
Effect of location, environmental conditions and year‐to-year variability on the GA content of potatoes
Sinden and Webb (1972) conducted a large 2‐year field trial including five commercial cultivars grown on 39 locations in the USA and Alaska. Overall, the average TGA content ranged from 43 mg/kg for cultivar ‘Red Pontiac’ in 1970–100 mg/kg for the cultivar ‘Kennebec’ in 1971. For three cultivars tested, relatively small variations between years for the average TGA content were found. A significant correlation between the location and the TGA content was noted, with potatoes from Alaska and North Dakota containing the highest average TGA content for the five cultivars (150 mg/kg and 143 mg/kg, respectively), while lowest TGA contents were found in Florida (31 mg/kg) (data for 1970). Important factors influencing the GA content were the (local) climatic conditions, length of daylight and soil type. A cold and wet, as well as a hot climate appears correlated with an elevated TGA content, as well as locations with extended day lengths of 18–20 h. Higher TGA contents can be associated with immaturity of the tubers, which is in turn related to the climate conditions and extended daytime length (Sinden and Webb, 1972).
The importance of location and the environmental conditions was also noted in a German study covering several locations over a span of 5 years (Ross et al., 1978). The variation in TGA content between locations amounted to 50% and the variation between years (strongly related to the weather conditions) to 78%. A 3‐year study conducted in the UK also showed that although the TGA content is primarily determined by the genotype (cultivar), location and environment are important co‐influencing factors (Bintcliffe et al., 1982; Parnell et al., 1984). A significant yearly variation was found in a 3‐year study conducted in Hungary with five different cultivars (Tömösközi‐Farkas et al., 2006). This was also concluded from a 2‐year study on six early potato cultivars in Sweden, as the TGA content for the six cultivars in 1986 was 30–130% higher compared to 1987 (Hellenäs et al., 1995a). Papathanasiou et al. (1999) noted a significant difference in the amount of yearly variation in TGA content for two cultivars (‘Pentland Dell’ and ‘Estima’) studied. The cultivar ‘Pentland Dell’, which has intrinsically a higher TGA content, was much more sensitive to differences in environmental conditions than the cultivar ‘Estima’, which has a low TGA content. The effect of location on 14 cultivars of different flesh‐colours (white/yellow, red or blue) was investigated during a 3‐year period by Hamouz et al. (2014). With respect to TGA content, genotype was the most important factor, followed by the year‐to‐year variation, while location was the least important. In a follow‐up study, the effect of altitude, climatological conditions and soil type using 16 cultivars on two locations (298–460 m) were studied over a 3‐year period (Urban et al., 2018). The authors concluded that differences found in TGA content were mainly correlated to the genotype, followed by differences in environmental conditions and the soil type. Ieri et al. (2011) studied the TGA content of nine purple or red skinned cultivars, grown on two locations in Italy. The average TGA content was slightly higher in the tubers grown in central Italy (100 mg/kg) compared to the tubers grown at higher altitude (1,000 m) in the south (91 mg/kg). It was noted that the difference was mainly related to the TGA content in the flesh of the tubers, which averaged to 8.6 mg/kg in south Italy and 28.3 mg/kg in central Italy. Panovská et al. (1997) studied the effect of altitude (110 m, 460 m, 860 m) on the TGA content of 11 cultivars, but no significant effects were noted.
Several studies reported besides the TGA content, also the concentration of α‐chaconine and α‐solanine. From these data, the ratio of α‐chaconine to α‐solanine was calculated. The reported ratio of α‐chaconine to α‐solanine ranges from 0.30 to 5.93. From the data it follows that, generally, α‐chaconine is present in higher concentrations than α‐solanine. There is, however, a substantial variation in the ratio between α‐chaconine and α‐solanine, not only for individual samples within a study but also between studies.
In the Netherlands, the Nederlandse Algemene Keuringsdienst (NAK) (Dutch General Inspection Service) is responsible for the registration and authorisation of seed potatoes. New cultivars are grown for at least 2 years at two locations under controlled conditions together with the reference potato cultivar Innovator, which has an average TGA content of 152 mg/kg. To be allowed to the market, the TGA content of the new cultivars must be lower than the TGA content of the Innovator cultivar. TGA content is determined according to the HPLC‐UV method as described by Houben and Brunt (1994).
The NAK list of registered potato cultivars in 201822 contained 552 entries, and for 464 cultivars, the TGA content is listed. Before 2000, it was not mandatory to establish the TGA content and in particularly for older cultivars, this information has not been recorded. The TGA distribution in the cultivars is shown in Figure 6, where it can be seen that the large majority of potatoes grown under the Dutch test conditions have a TGA content of < 100 mg/kg. The average content of the registered cultivars is 48.4 mg/kg.
Figure 6.
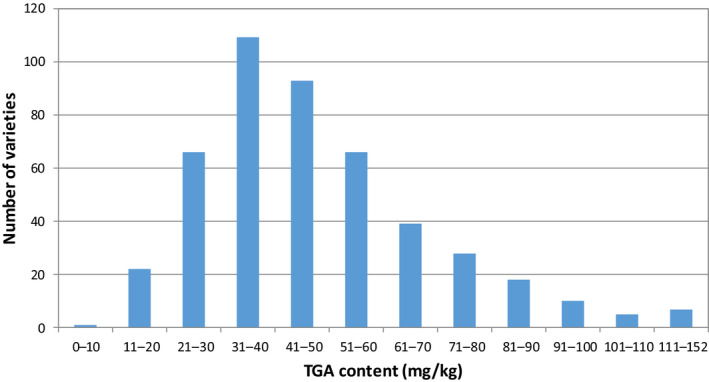
Distribution of total glycoalkaloid (TGA) content in 464 potato cultivars registered in the Netherlands22
The NAK list22 includes also 77 potato cultivars grown specifically for starch production. For 70 cultivars, the TGA content is listed. Starch potatoes23 contain higher amounts of GAs: the average TGA content is 107.7 mg/kg, while 34 cultivars exceed 100 mg/kg and 7 exceed 200 mg/kg. The cultivar ‘Kuras’ has the highest TGA content (338 mg/kg) and also the cultivars ‘Aventra’ and ‘Elles’ (both 260 mg/kg) are also high in TGA content.
Overall, it can be concluded from the field studies that genotype is the most important factor determining the TGA content. However, also environmental conditions, location and year‐to‐year variability contribute to the differences in TGA content observed during the preharvest stage.
Differences in GAs content between organic and conventionally grown potatoes
Several studies evaluated the effect of farming systems (organic versus conventional) on the GAs content in potato tubers as shown in Table 31 and some further details are given in Table 32.
Table 32.
Total glycoalkaloids (TGAs) content in potato tubers grown in conventional and organic farming systems
| Country | Potato cultivars | Year of harvest | TGA content conventional (mg/kg fw) | TGA content organic (mg/kg fw) | Analytical technique | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| α‐solanine | α‐chaconine | TGA | α‐solanine | α‐chaconine | TGA | |||||
| CZ | Karin | 1995–1997 | 53–113 | 59–157 | HPLC‐UV | Hamouz et al. (2005) | ||||
| CZ | Christa, Karin, Koruna, Krasa, Krystala, Monalisa, Rosara, Rosella | 1996–1999 | 15–245 | 16–157 | HPLC‐UV | Hajšlová et al. (2005) | ||||
| CZ | Bionta, Karin, Marabel, Rosara, Satina | 2006–2008 | 35.3 | 55.6 | 90.9 | 30.4 | 53.2 | 83.6 | LC‐MS/MS | Bártová et al. (2013) |
| HU | Hópehely, Rioja, White Lady | 2007–2009 | 0.0–15 | 0.0–28 | HLPC‐UV | Tömösközi‐Farkas et al. (2014) | ||||
| LV | 20 potato genotypes | 2010–2011 | 1.0–54.5 | 1.5–42.5 | 1.5–59.0 | 1.5–48.5 | LC‐MS | Skrabule et al. (2013) | ||
| PT | Raja, Santé | 2003 | 17.7–43.9 | 19.6–35.5 | 37.3–79.5 | 13.6–23.3 | 21.3–24.8 | 38.4–44.6 | HPLC‐UV | Abreu et al. (2007) |
TGA: total glycoalkaloid; UV: ultraviolet; LC: liquid chromatography; MS: mass spectrometry; MS/MS: tandem MS.
In the study of Abreu et al. (2007), higher TGA concentrations were found in the conventionally grown Raja tubers, compared to the organically grown ones, while for the Santé tubers, no clear difference in TGA content was observed between the farming systems. Skrabule et al. (2013) evaluated GAs in 20 potato genotypes grown under organic and conventional farming systems in the years 2010 and 2011. The genotype did significantly influence the GAs content, while the effect of farming system was not significant, except for the content of α‐solanine in the 2011 season. Bártová et al. (2013) showed the importance of the individual reaction of the genotypes to stress conditions existing in organic and conventional crop management. Crop management seemed to only influence the GA content in tubers in interaction with other evaluated agro‐ecological factors (cultivar, year and locality). Although a significantly lower concentration of α‐solanine was found in organically grown tubers, with respect to the TGA content the difference between the two farming systems was not significant. Hajšlová et al. (2005) studied the tuber quality of eight potato cultivars grown in organic and conventional farming systems. No statistically significant difference was found between the TGA contents in tubers produced by the two production systems. The observed difference in TGA content seems to be caused by a substantial variation in the content of GAs between years, geographical area of cultivation and cultivar. Additionally, Tömösközi‐Farkas et al. (2014) investigated the GA content in three Hungarian potato species grown in organic and conventional farming systems. The respective farming system was found to have no effect on the level of GAs, and was mostly influenced by season and genotype. Diehl and Wedler (1977) studied the effect based on the rate of fertilisers applied. No difference was found between α‐solanine content in potatoes in relation to organic and mineral fertilisation.
Distribution of GAs between potato peel and flesh
In a number of field studies, the distribution of GAs between the peel and the flesh was studied. The most comprehensive study is that of Valcárcel et al. (2014) in which the α‐solanine and α‐chaconine content of potato skin and flesh of 60 potato cultivars grown in two locations in Ireland over a 2‐year period was investigated. TGA levels varied from 4 to 957 mg/kg dw in the potato flesh and from 150 to 8,133 mg/kg dw in the potato peels. The ratio of α‐chaconine to α‐solanine was higher in the peels (average ratio: 1.44; range 0.7–4.5) than in the flesh (average ratio: 0.87; range 0.3–1.6). Deußer et al. (2012) conducted a similar study on 17 cultivars grown in Luxembourg. TGA contents were determined for the peel (1–2 mm thickness), the outer flesh (1 cm) and inner flesh. The peels contained between 545 and 5,342 mg TGA/kg dw, while the outer and inner flesh contained 19–466 and 7–187 mg TGA/kg dw, respectively. The α‐chaconine:α‐solanine ratio varied between 1.05 and 3.45 (average: 1.94) in the peel, between 0.83 and 2.36 (average: 1.46) in the outer flesh and between 1.00 and 2.38 (average: 1.69) in the inner flesh. Friedman et al. (2003a) reported for eight potato cultivars a range of 84–3,526 mg TGA/kg dw in potato peels, and of 6–592 mg TGA/kg dw in potato flesh. A study on 12 potato cultivars grown in Mexico reported for potato peels 176–5,497 mg TGA/kg dw and for flesh < 5–642 mg TGA/kg dw (Sotelo and Serrano, 2000).
Occurrence data on GAs in potato samples obtained from retail surveys
In Table 33, an overview is given of surveys conducted on GAs levels in potatoes collected from retail outlets. Information on such surveys is available only for three different countries in the EU: Denmark, Sweden and the UK.
Table 33.
Summary results of surveys conducted on total glycoalkaloids (TGAs) levels in potatoes for human consumption sampled in retail outlets
| Country | Year of survey | Potato cultivars | N | TGA average (mg/kg fw)b | Range (mg/kg fw) | Analytical technique | Reference |
|---|---|---|---|---|---|---|---|
| CZ | 1993–1994 | All cultivars (14) (tuber size 100–160 g)a | 14 | 63 | 23–95 | HPLC‐UV | Panovská et al. (1997) |
| All cultivars (14) (tuber size 20–50 g)a | 14 | 110 | 40–237 | ||||
| DK | 2001–2002 | All cultivars (6) | 28 | 102 | 45–167 | HPLC‐UV | Knuthsen et al. (2009) |
| 2002–2003 | All cultivars (14) | 50 | 64 | 26–159 | |||
| 2003–2004 | All cultivars (10) | 54 | 97 | 26–280 | |||
| 2004–2005 | All cultivars (28) | 99 | 67 | 15–223 | |||
| 2005–2006 | All cultivars (20) | 64 | 54 | 5–109 | |||
| 2006–2007 | All cultivars (22) | 91 | 49 | < 5–105 | |||
| All DK samples | 386 | 67 | < 5–280 | ||||
| SE | 1986 | ‘Magnum Bonum’ | 332 | 254 | 61–665 | HPLC‐UV | Hellenäs et al. (1995a,b) |
| Others (8 main‐crop cultivars) | 105 | 108 | 35–315 | ||||
| 1987 | ‘Magnum Bonum’ | 16 | 151 | 60–293 | HPLC‐UV | Hellenäs et al. (1995a,b) | |
| Others (8 early cultivars) | 126 | 120 | 31–344 | ||||
| 1997 | Main‐crop potatoes (8 cultivars) | 113 | 105 | 36‐244 | HPLC‐UV | Branzell and Hellenäs (1999) | |
| Early potatoes (7 cultivars) | 127 | 98 | 35‐267 | ||||
| 2015 | All cultivars (31) | 38 | 95 | 24–277 | HPLC‐UV | Swedish NFA (2016) | |
| All SE samples but excluding ‘Magnum Bonum’ | 509 | 107 | 24–344 | ||||
| UK | 1980–1981 | Foreign early potatoes (8 cultivars) | 42 | 123 | 56–208 | Colourimetric | Davies and Blincow (1984) |
| UK early potatoes (5 cultivars) | 28 | 115 | 70–212 | ||||
| UK main‐crop potatoes (3 cultivars) | 63 | 105 | 60–195 | ||||
| All UK samples | 133 | 113 | 56–212 |
LC: liquid chromatography; UV: ultraviolet.
From wholesale.
Average as reported by the authors or calculated based on the provided individual data.
Panovská et al. (1997) studied the TGA content in 14 Czech cultivars available on the Czech market. It was found that for all cultivars, larger sized tubers (100–160 g) consistently contained lower GA levels than smaller sized tubers (20–50 g). TGA levels in all large‐sized tubers were less than 100 mg/kg, while for the small‐sized tubers for two cultivars (‘Javor’ and ‘Oreb’) levels above 200 mg/kg were found.
The Danish survey spanned 7 years, from 2001 to 2007, and a total of 386 potato tuber samples were analysed (Knuthsen et al., 2009). The number of samples and the potato cultivars collected varied from year to year. There was a significant yearly variation noted in the average TGA content of the potatoes sampled; a relatively high average TGA content was obtained for the seasons 2001–2002 and 2004–2005. In total, 56 samples exceeded 100 mg TGA/kg fw, and three exceeded 200 mg/kg fw. Cultivars that exceeded 200 mg/kg included ‘Sava’ (1 sample out of 38 analysed, origin Denmark), an unknown cultivar (origin: Cyprus) and ‘Grenailles’ (origin: Italy).
When excessive levels of GAs were detected in Swedish potatoes, particularly in the cultivar ‘Magnum Bonum’, a broad survey was conducted in Sweden. The survey was conducted in 1986–1987 and a total of 617 potato samples were analysed, including 348 ‘Magnum Bonum’ samples (Hellenäs et al., 1995a,b). Besides the ‘Magnum Bonum’ cultivar, high TGA levels were also found in potatoes of other cultivars, but to a lower extent. The most prominent of the other cultivars that regularly exceeded 200 mg/kg were the early potato ‘Ulster Chieftain’ and the main‐crop potato ‘Snöboll (British Queen)’. In a follow‐up study in 1997, 240 samples of main‐crop and early potato cultivars were sampled and analysed (Branzell and Hellenäs, 1999). TGA levels were only slightly lower than in 1986–1987. The six samples collected from the early potato ‘Ulster Chieftain’ all exceeded 200 mg/kg and also about 30% of the early potato ‘Pentland Javelin’ samples also exceeded this limit. A further survey was conducted by the Swedish Food Safety Authority in 2015, in which 38 samples of potato tubers were collected and analysed for GA content (Swedish NFA, 2016). The 38 samples represented 31 different cultivars. Twelve samples had a TGA content in excess of 100 mg/kg, and five exceeded 200 mg/kg: the cultivars ‘Caro’, ‘Folva’, ‘Rosagold’, ‘Sava’ (for each cultivar, one sample analysed) and ‘Snöboll’ (one out of two samples analysed). Overall, the TGA levels were slightly lower than in 1986–1987 and similar to the 1997 surveys.
In the UK study of 1981, 514 potato samples were collected from supermarkets, greengrocers and farm shops (Davies and Blincow, 1984). Samples were pooled to produce 133 composite samples. Sampling of cultivars was made on basis of available consumption data. Samples were grouped in three categories: foreign early potatoes (42 samples), UK early potatoes (28 samples) and UK main‐crop potatoes (63 samples). The TGA content was determined using a less specific colourimetric method. The average TGA content of the samples was 113 mg/kg fw, with a range of 56–212 mg/kg. No statistically significant differences in TGA content between the different groups could be found. One sample (cultivar ‘Pentland Javelin’, six samples tested) exceeded 200 mg TGA/kg.
Processed potato food products
Several studies have determined the levels of GAs in laboratory processed and in commercial potato products such as potato crisps (or potato chips) and French fries (Sizer et al., 1980; Jones and Fenwick, 1981; Bushway and Ponnampalam, 1981; Saito et al., 1990; Friedman and Dao, 1992; Rytel, 2012a; Liu et al., 2014; Tajner‐Czopek et al., 2014). The results are summarised in Table 34. In potato crisps, TGA levels ranging from 16 to 720 mg/kg have been reported. In French fries, the reported levels are lower, ranging from 0.4 to 58 mg/kg. Dehydrated products contained TGA levels between 15 and 75 mg/kg.
Table 34.
Total glycoalkaloids (TGAs) in laboratory processed and in commercial potato products
| Country | Year of study | Producta | N | TGA average (mg/kg fw)f | Range (mg/kg fw) | Analytical technique | Reference |
|---|---|---|---|---|---|---|---|
| USA | 1980 | Potato chipsb | 3 | 304 | 95–720 | Colourimetric | Sizer et al. (1980) |
| Potato chipsc |
4 (different production methodsd) Potato TGA content: 42.3 mg/kg |
166 | 123–236 | ||||
| UK | 1981 | Potato crispsb | 2 | 65 | 59–70 | Colourimetric | Jones and Fenwick (1981) |
| Pre‐cooked frozen chipsb | 2 | 39 | 23–55 | ||||
| USA | 1981 | Potato chipsb | 14 | 71 | 27–162 | HPLC‐UV | Bushway and Ponnampalam (1981) |
| Frozen fries (steak, French, crinkle cut)b | 18 | 22 | 3.3–58 | ||||
| Frozen fried potato ballsb | 4 | 19 | 14–23 | ||||
| Frozen mashed potatoesb | 3 | 3.3 | 2.0–4.6 | ||||
| Frozen baked potatoesb | 2 | 102 | 80–123 | ||||
| Frozen fried potatoesb | 5 | 6.4 | 4.3–8.4 | ||||
| Dehydrated potato flourb | 2 | 70 | 65–75 | ||||
| Dehydrated potato flakesb | 2 | 19 | 15–23 | ||||
| Canned potatoesb | 4 | 1.5 | 0.9–2.5 | ||||
| Canned home friesb | 2 | 1.3 | 1.1–1.5 | ||||
| JP | 1990 | Potato chipsb | 6 | 29 | 11–54 | Saito et al. (1990) | |
| Fried potatoesb | 1 | 33 | – | ||||
| Frozen fried potatoesb | 1 | 31 | – | ||||
| Croquettesb | 1 | 24 | – | ||||
| Frozen croquettesb | 1 | 67 | – | ||||
| Mashed potatoesb | 1 | 20 | – | ||||
| USA | 1992 | Potato chipsb | 3 | 61 | 23.5–109 | HPLC‐UV | Friedman and Dao (1992) |
| French friesb | 3 | 18 | 0.4–44 | ||||
| Pancake Powderb | 2 | 45 | 44–45 | ||||
| PL | 2012 | Dehydrated cooked potatoesc | 4, potato TGA content: 200–296 mg/kg dw; 5 processing stages | 65 | 61–69 | HPLC‐UV | Rytel (2012a) |
| CN | 2014 | Potato crispsb | 20 | n.d. | 16–61 | LC‐MS/MS | Liu et al. (2014) |
| PL | 2014 | French friesc |
4 (colour‐fleshed potatoes) Potato TGA content: 41–61 mg/kg; 5 processing stepse |
8 | 6–10 | HPLC‐UV | Tajner‐Czopek et al. (2014) |
dw: dry weight; LC: liquid chromatography; UV: ultraviolet; MS: mass spectrometry; MS/MS: tandem MS.
Potato chips (often just chips), or crisps, are thin slices of potato that have been deep fried or baked until crunchy. Regarding naming, British crisps are US chips, while British chips are US fries/French fries (Oxford dictionary online). In the above table each product is named according to the naming in the article.
Commercial product.
Experimental product.
Different frying temperature, frying time and slice thickness.
For details see Section 3.2.3.
Average as reported by the authors or calculated based on the provided individual data.
Only two studies have reported on the content of GAs in potato starch. Saito et al. (1990) reported a TGA content of 1.1–3.5 mg/kg in three commercial samples of potato starch. Driedger and Sporns (1999) reported that they did not detect GAs in the starch extracted from potato tubers using a labscale model of an industrial extraction process. Limits of detection were not reported for the MALDI‐MS method used by the authors.
Potato peel products
Studies that investigated the GA content of potato peel products are summarised in Table 35. Bushway et al. (1983) analysed a range of different potato products available on the US market and reported a range from 52 to 242 mg TGA/kg. They also studied the TGA content of baked, fried and baked‐fried potato peels (see Section 3.2.3.1.2). An increase in TGA content was noted compared to unprocessed peels, what could be explained by a lower water content in the processed products.
Table 35.
Total glycoalkaloids (TGAs) in laboratory processed and in commercial potato peel products
| Country | Year of study | Potato product | N | TGA average (mg/kg fw)c | TGA range (mg/kg fw) | Analytical technique | Reference |
|---|---|---|---|---|---|---|---|
| USA | 1981 | Fried peelsa | 2 | 142 | 139–145 | HPLC‐UV | Bushway and Ponnampalam (1981) |
| USA | 1983 | Unprocessed potato peelsb | 12 cultivars | 276 | 18–1,068 | HPLC‐UV | Bushway et al. (1983) |
| Baked potato peelsb | 12 cultivars | 317 | 24–1,098 | ||||
| Fried potato peelsb | 12 cultivars | 414 | 33–1,649 | ||||
| Baked‐fried potato peelsb | 12 cultivars | 444 | 42–1,556 | ||||
| Frozen peel wedgesa | 6 | 98 | 76–121 | ||||
| Frozen peel slicesa | 2 | 68 | 66–71 | ||||
| Restaurant fried peelsa | 3 | 57 | 52–63 | ||||
| Restaurant baked‐fried peelsa | 3 | 181 | 120–242 | ||||
| USA | 2017 | Potato peel powderb | 6 (3 cultivars, organic and conventional) | 1,541 (dw) | 639–3,580 (dw) | HPLC‐UV | Friedman et al. (2017) |
| USA | 2018 | Potato peel powdera | 4 (3 cultivars, organic and conventional) | 1,527 (dw) | 861–2,180 (dw) | HPLC‐UV | Elkahoui et al. (2018) |
Dw: dry weight; LC: liquid chromatography; UV: ultraviolet.
Commercial product.
Experimental product.
Average as reported by the authors or calculated based on the provided individual data.
Friedman et al. (2017) measured the GAs content of potato peel powder prepared from three conventional and three organic store‐bought potatoes (‘Gold’, ‘Red’ and ‘Russet’) by HPLC and LC‐MS and detected a wide variation in GAs content between the samples. Highest TGA levels were found in potato peel powder from the organically grown ‘Gold’ cultivar (3,580 ± 390 mg/kg dw) and the lowest in conventionally grown ‘Russet’ (639 ± 52 mg/kg dw). It was noted that the corresponding cultivars did not always have a higher TGA level when comparing the organic vs. the conventional samples. For the ‘Red’ cultivar, the TGA level in potato peel powder of conventionally grown potatoes was higher than in organically grown potatoes (1,709 ± 61 vs. 850 ± 120 mg/kg dw). In a follow‐up study, Elkahoui et al. (2018) reported the GAs content in potato peel powder prepared from three potato cultivars (‘Gold’, ‘Red’, ‘Russet’) grown under organic and/or conventional conditions. A higher TGA content was reported for the conventionally grown ‘Russet’ cultivar (1,128 ± 1 mg/kg dw) vs. the organically grown ‘Russet’ cultivar (861 ± 10), but higher TGA levels were reported for the other two cultivars grown only under organic conditions (1,940–2,180 mg/kg dw).
3.2.2.1.2. Occurrence data on GAs in tomatoes
A number of publications were identified on the occurrence of tomato GAs in tomatoes. The results are summarised in Table 36. The levels of α‐tomatine reported in ripe tomatoes are generally low, ranging from not detectable to 23 mg/kg fw. In green, but mature, tomatoes somewhat higher amounts of α‐tomatine were found, ranging from not detectable to 90 mg/kg fw. Green, immature, tomatoes contain the highest TGA levels, ranging from 35 to 550 mg/kg fw. It should be noted that the results were obtained with methods that had a wide range of LOQs (ranging from 0.05 to 2.5 mg/kg).
Table 36.
Summary results of literature reports on total glycoalkaloids (TGA) concentrations in tomatoes
| Country | Year of survey | Tomato | N | TGA average (mg/kg fw)e | Range (mg/kg fw) | Analytical technique | Reference |
|---|---|---|---|---|---|---|---|
| UK | 1984 | Ripe (3 cultivars)a | 3 | 3.7 | 3–6 | Radioimmunoassay | Eltayeb and Roddick (1984) |
| NL | 1987 | Ripea | 1 | 5 | – | GC‐FID and GC‐NPD | Van Gelder and De Ponti (1987) |
| NL | 1994 | Redb | 14 | 0.6 | < 0.5–4.1 | HPLC‐UV | Keukens et al. (1994) |
| Orangeb | 14 | 6.2 | < 0.5–18.2 | ||||
| Greenb | 14 | 26 | 4.5–90 | ||||
| USA | 1994 | Red (5 cultivars)b | 99 | – | < 2.5–23 (94 are ND) | HPLC‐UV | Bushway et al. (1994) |
| Green (4 cultivars)b | 80 | 18.3 | < 2.5–88 | ||||
| USA | 1994–2004 | Red, conventional (3 cultivars)c | 54 | 1.16f | < 0.05–5.6f | LC‐MS/MS | Koh et al. (2013) |
| Red, organic (3 cultivars)c | 54 | 2.11f | < 0.05–4.9f | ||||
| USA | 1994 | Red, riped | 4 | 1.1 | 0.3–2.8 | HPLC‐UV/HPLC‐PAD | Friedman et al. (1994) |
| Reda | 2 | 2.7 | 1.5–3.9 | ||||
| Breakera | 1 | 16.5 | – | ||||
| Green, maturea | 1 | 45 | – | ||||
| Green, immaturea | 1 | 69 | – | ||||
| USA | 1995 | Red, yellow, ripe (8 cultivars)d | 8 | 3.6 | 0.3–11 | HPLC‐PAD | Friedman and Levin (1995) |
| Green, unriped | 1 | 16 | – | ||||
| Reda | 2 | 1.8 | 1.2–2.5 | ||||
| Breakera | 2 | 10.5 | 6.7–14 | ||||
| Green, maturea | 2 | 40 | 25–58 | ||||
| Green, immaturea | 2 | 125 | 40–210 | ||||
| JP | 1997 | Green, maturea | 7 | 165 | 140–180 | Colourimetric | Furui et al. (1997) |
| Turninga | 4 | 44 | 30–60 | ||||
| Red, maturea | 6 | 3 | 1–8 | ||||
| USA | 1998 | Reda | 15 | 1.4 | 0.4–4.3 | HPLC‐PAD | Friedman and Levin (1998) |
| Green, immaturea | 15 | 205 | 35–550 | ||||
| IT | 2000 | Green‐orange, reda | 5 | 15 | ND–43 | HPLC‐UV | Leonardi et al. (2000) |
| IL | 2006 | Ripeningc | 3 | 2.5f | 0–6.7f | HPLC‐UV | Mulatu et al. (2006) |
| Green, maturec | 3 | 0.65f | 0–1.4f | ||||
| Green, immaturec | 3 | 1.4f | 0.13–3.5f | ||||
| JP | |||||||
| IT | 2016 | Ripe (15 landraces and 3 hybrids)c | 18 |
1.5g 5.1h |
0.6–4.0g 1.8–14.2h |
LC‐HRMS | Baldina et al. (2016) |
| IT | 2019 | Various ripening stagesd | 3 | 1.1g | 0.49–2.0g | LC‐MS/MS | Tamasi et al. (2019) |
TGA: total glycoalkaloids; LC: liquid chromatography; UV: ultraviolet; MS: mass spectrometry; MS/MS: tandem MS; HR: high resolution; HP: high‐performance; PAD: Pulsed amperometric detector; FID: flame ionisation detector. NPD: nitrogen‐phosphorous detector.
Greenhouse.
Wholesale.
Field trial.
Retail.
Average as reported by the authors or calculated based on the provided individual data.
Data were presented as dry weight. It has been assumed that the water content was 95%.
Results for α‐tomatine + α‐dehydrotomatine.
Results for all 7 GAs analysed (α‐tomatine, dehydrotomatine, esculeoside A, lycoperosides A, B and H and tomatoside A).
Baldina et al. (2016) investigated 15 landraces and 3 hybrids with different morphological characteristics (flattened/ribbed, pear/oxheart, round/elongate) for the presence of seven tomato glycosides (α‐tomatine, α‐dehydrotomatine, esculeoside A, lycoperosides A, B and H and tomatoside A). On average, slightly higher concentrations were found for tomatoside A (mean 14.4 mg/kg fw, range 0.25–5.5 mg/kg fw) than for α‐tomatine (mean 12.2 mg/kg fw, range 0.36–3.4 mg/kg fw), esculeoside A (0.91 mg/kg fw, range 0.36–2.7 mg/kg fw) and lycoperoside H (0.88 mg/kg fw, range 0.28–2.8 mg/kg fw). The other GAs were present at lower levels.
Differences in GAs content between organic and conventionally grown tomatoes
Koh et al. (2013) studied the α‐tomatine content in dried tomato samples grown by organic and conventional crop management over a period of 10 years. Mean levels of α‐tomatine in organically grown tomatoes (42.3 mg/kg dw) were significantly higher than in the conventionally (23.2 mg/kg dw) grown crops. The differences were attributed to environmental factors, external to nitrogen fertilisation, affecting the α‐tomatine content in tomatoes.
Processed tomato products
Most of the studies on GA content in processed tomato products have been made by the same research group (Friedman et al., 1994; Friedman and Levin, 1995; Friedman, 2004). Table 37 shows the results of a survey in which different tomato products were obtained from a local market. Pickled green tomatoes and tomato red sauce showed the highest concentrations of α‐tomatine (Kyzlink et al., 1981; Kibler et al., 1985; Friedman and Levin, 1995).
Table 37.
Total glycoalkaloid (TGA) content of processed tomato products reported in the literature
| Country | Year of study | Product | N | TGA average (mg/kg fw)f | TGA range (mg/kg fw) | Analytical technique | Reference |
|---|---|---|---|---|---|---|---|
| CZ | 1981 | Tomato pulpa , b | 1 | 193 | HLPC/colourimetrici | Kyzlink et al. (1981) | |
| Sweetened tomato juicea , b | 1 | 165 | |||||
| Tomato marmaladea , b | 1 | 182 | |||||
| Fruit buttera , b | 1 | 143 | |||||
| DE | 1985 | Green tomato marmaladea | 2 | 35g | 25‐44g | Colourimetrici | Kibler et al. (1985) |
| Sweet sour pickled green tomatoesa | 2 | 31g | 20‐42g | ||||
| CZ | 1992 | Green tomato salad with onion, steriliseda , c | 2 | 24 | 20–28 | HPLC‐UV | Voldrich et al. (1992) |
| Green tomato salad with onion and paprikaa | 1 | 8 | |||||
| Fermented green tomatoesa | 1 | 14h | |||||
| Tomato pureea | 3 | 18 | 16–20 | ||||
| USA | 1995 | Condensed tomato soup | 3 | 2.2 | 1.4–3.7 | HPLC‐PAD | Friedman and Levin (1995) |
| Stewed red tomatoes | 1 | 1.1 | |||||
| Sun dried tomatoes | 1 | 21 | |||||
| Fried green tomatoesa , d | 1 | 11 | |||||
| Microwaved green tomatoesa , e | 1 | 12 | |||||
| Tomato ketchup | 1 | 25 | |||||
| Tomato juice | 1 | 28 | |||||
| Tomato red sauce | 1 | 57 | |||||
| Green salsa | 2 | 9.5 | 0–19 | ||||
| Pickled green tomato product | 2 | 50 | 28–72 | ||||
| USA | 2017 | Red and tangerine tomato juice | 2 | 0.65 | 0.5‐0.8 | LC‐HRMS | Cichon et al., 2017 |
TGA: total glycoalkaloids; LC: liquid chromatography; UV: ultraviolet; MS: mass spectrometry; HR: high resolution; HP: high performance; PAD: Pulsed amperometric detector.
Prepared in the laboratory.
Prepared from unripe tomatoes.
1 sample sterilised at 90°C and 1 sample sterilised at 120°C.
Frying was done as follows: tomatoes were sliced and breaded with equal amounts of flour and corn meal after which they were fried in ¼ inch of oil.
Microwaving was for 5 min at ‘high’.
Average as reported by the authors or calculated based on the provided individual data.
Reported as solanine, probably α‐dehydrotomatine has been measured.
The brine was analysed as well, containing 7 mg/kg α‐tomatine.
Colourimetric methods cannot discriminate between α‐solanine, α‐chaconine and α‐dehydrotomatine.
3.2.2.1.3. Occurrence data on GAs in aubergines
The literature on the occurrence of GAs in the aubergine (S. melongena) is very limited. The data available are summarised in Table 38. Bajaj et al. (1979), using a colourimetric method, reported on the TGA content in mature fruits of 21 different cultivars (including round type, long type and oblong type fruits). The fruits contained GAs, expressed as α‐solanine equivalents, between 65 and 205 mg/kg fw (Bajaj et al., 1979). Using a similar approach, Jones and Fenwick (1981) analysed two aubergine samples imported to the UK. The samples contained 76 and 82 mg GAs (determined as α‐solanine) per kg fw.
Table 38.
Summary results of literature reports on total glycoalkloid (TGA) concentrations in aubergine
| Country | Year of survey | Aubergine species | N | TGA average (mg/kg fw)c | Range (mg/kg fw) | Analytical technique | Reference |
|---|---|---|---|---|---|---|---|
| IN | 1979 | S. melongena a | 21 | 113 | 65–205 | Colourimetric | Bajaj et al. (1979) |
| UK | 1981 | S. melongena b | 2 | 79 | 76–82 | Colourimetric | Jones and Fenwick (1981) |
| TR | 2006 | S. melongena b | 1 | < LOQ (10) | – | HPLC‐UV | Eanes et al. (2006) |
| ES | 2010 | S. melongena a | 3 | 14.3 | 11–20 | LC‐MS/MS | Sánchez‐Mata et al. (2010) |
| S. macrocarpon a | 4 | 1639 | 1,402–2,210 | ||||
| S. aethiopicum a | 3 | 19.5 | 10–54 | ||||
| IT | 2012 | S. melongena, unripea | 10 | 10.2d | 0.6–19.6d | HPLC‐UV | Mennella et al. (2012) |
| S. melongena, commercial statea | 10 | 20.6d | < 0.02–85d | ||||
| S. melongena, physiologically ripea | 10 | 163d | 25.0–403d | ||||
| S. aethiopicum, unripea | 1 | 527 | – | ||||
| S. aethiopicum, commercial stagea | 1 | 193 | – | ||||
| S. aethiopicum, physiologically ripea | 1 | 349 | – | ||||
| IT | 2016 | S. melongena, commercial statea | 3 | 311d | 231–430d | HPLC‐UV | Lo Scalzo et al. (2016) |
| IT | 2019 | S. melongena, commercial statea | 3 | 435d | – | LC‐HRMS | Lelario et al. (2019) |
TGA: total glycoalkaloids; LC: liquid chromatography; UV: ultraviolet; MS: mass spectrometry; MS/MS: tandem MS; HR: high resolution; HP: high performance.
Field trial.
Retail.
Average as reported by the authors or calculated based on the provided individual data.
Concentrations were presented as dry weight; fresh weight concentrations have been calculated based on the water content of the samples as provided in the publication.
Using a HPLC‐UV method, Eanes et al. (2008) reported for S. linneaeanum (a wild‐type aubergine) a solmargine content of 25,500 ± 11,200 mg/kg dw, while no GAs could be detected in a commercial sample of aubergine (reported LOQ was 100–200 mg/kg dw). Assuming a water content of 90%, the LOQ in fresh material was 10–20 mg/kg.
Using an LC‐MS/MS method, Sánchez‐Mata et al. (2010) reported for the fruits of four different varieties of the gboma aubergine a TGA content of 1,402–2,210 mg/kg fw. The majority corresponded to α‐solamargine (1,241–1,975 mg/kg), while the concentration of α‐solasonine (161–235 mg/kg) was lower. In three varieties of the scarlet aubergine, α‐solamargine (5.8–48.6 mg/kg) was also the major GA compared to α‐solasonine (4.1–10.0 mg/kg). Also, for three varieties of aubergine, α‐solamargine (8.5–16.1 mg/kg) was present in higher concentrations than α‐solasonine (1.7–4.0 mg/kg) (Sánchez‐Mata et al., 2010).
Mennella et al. (2012) studied the effect of ripening on the GA concentration in 10 experimental lines of aubergine. Interestingly, they noticed a substantial increase of GA concentration during ripening of the fruits. The TGA concentration increased from approximately 10 mg/kg fw in unripe aubergines to 20 mg/kg fw in commercially ripe aubergines to 160 mg/kg fw in physiologically ripe aubergines. In the immature and commercially ripe stage, the flesh appears soft and greenish, the peel glossy (unripe) or a bit dull (commercial stage). The physiologically ripe stage is typified by a brownish peel colour, spongy, fibrous and yellowish flesh and visible and hard mature seeds. The ratio between α‐solamargine and α‐solasonine fluctuated substantially between the different samples. In approximately 50% of the samples, α‐solasonine could not be detected (LOQ was 0.02 mg/kg fw). Overall, α‐solamargine contributed to the total content, for the unripe, commercially ripe and physiologically ripe stage, 77%, 83% and 69%, respectively (Mennella et al., 2012).
3.2.2.1.4. Occurrence data on GAs in other food products
The Rapid Alert System for Food and Feed (RASFF) has received, up to the endorsement of this Opinion, no notifications on the presence of high levels of GAs in potato, aubergine, tomato or other food products. However, within the period 1982–2019, there have been seven notifications on the presence of S. nigrum (black nightshade) in food products (see Appendix D). Three recent notifications (2015–2018) concern the presence of S. nigrum in food supplements originating from India. The four other notifications (1982–2013) report the presence of S. nigrum berries in green beans (canned or frozen). None of the notifications report on concentrations of GAs in the products. The potential of S. nigrum and the related species S. ptycanthum Dun (eastern black nightshade) to contaminate food crops, such as beans, peas and soybeans, is a problem noted by several authors (Eldridge and Hockridge, 1983; Ogg et al., 1981; Ogg and Rogers, 1989). Cavlovic et al. (2003) reported unripe berries of S. nigrum to contain 1,100 mg/kg fw α‐solasonine and 900 mg/kg fw α‐solamargine. In a follow‐up of the RASFF notification 2013.0804, six samples of S. nigrum green berries were analysed by LC‐MS/MS (P. Mulder, 2019, personal communication). The berries contained 1,540 ± 170 mg/kg fw α‐solasonine and 1,580 ± 260 mg/kg fw α‐solamargine. Other GAs (α‐solanine and α‐chaconine) as well as the aglycones solasodine and solanidine could not be detected (LOD = 1 mg/kg fw).
3.2.2.2. Literature occurrence data in feed
No surveys on the levels of GAs in potatoes and potato by‐products used as feeds for livestock or companion animals have been identified in the open literature.
3.2.3. Influence of storage and processing on the content of GAs
Post‐harvest, both storage and processing may influence the content of different GAs in edible parts of plants such as in the potato tuber and in the tomato fruit.
3.2.3.1. GAs from S. tuberosum
3.2.3.1.1. Storage of potatoes
The main factors that may influence the GA content during storage of potato tubers include the duration, temperature, humidity and light exposure (wavelength and intensity), as well as tuber damage and the matrix on/in which the tubers are stored. Several studies on the influence of such factors on the content of GAs in potatoes have been performed. In these studies, the potato tubers were usually exposed to a combination of two or more of these factors, and different potato cultivars have been tested in the different experiments.
In general, studies have reported increases in the GA content with storage time (e.g. Abdel‐Gawad et al., 1993; Amer et al., 2014; Abbasi et al., 2016; Romanucci et al., 2016), although no variations in the levels have been observed for certain cultivars (e.g. Edwards and Cobb, 1999; Amer et al., 2014), while Wilson et al. (1983) reported a decrease. Storage at low temperatures (4°C) has been shown to increase the levels in some cultivars compared to storage at 10°C (Cieslik and Praznik, 1998), while others showed the opposite (Kumar and Ezekiel, 2009) or no change (Edwards and Cobb, 1997).
Exposure to light has generally been reported to induce formation of GAs. Both the wavelength of the light source and the cultivar influence the degree to which the overall content of GAs is enhanced (Nema et al., 2008; Paradiso et al., 2019). In general, exposure to fluorescent light has been shown to induce higher levels of GAs (e.g. Dao and Friedman, 1994; Percival, 1999; Machado et al., 2006; Abbasi et al., 2016). While some cultivars have been found to be nearly insensitive to light exposure (e.g. ‘Maris Piper’), others were reported to be very sensitive (e.g. ‘Kestrel’) (Percival, 1999).
Exposure of potato tubers to light can also induce the formation of chlorophyll, which results in the greening of the potato skin. The intensity, duration and quality of the light can impact on the rate of greening in potatoes: a greater light intensity will result in a faster progression of greening and chlorophyll accumulation in the tuber (Liljemark and Widoff, 1960; Olsen et al., 2018). The green pigmentation can render the potato undesirable for the consumer since skin quality is one of the most important factors influencing consumers choice (Olsen et al., 2018). The light‐induced chlorophyll and GA formation are processes concurrent during greening progression, with both implicated in a non‐mevalonate synthetic pathway but with independent genetic controls in the plant tissue (Edwards et al., 1998; Kozukue et al., 2001; Friedman, 2006). Early studies observed that UV light at a wavelength about 300 nm induced GA synthesis, while it did not induce the synthesis of chlorophyll (Conner, 1937). The formation of chlorophyll in the outer 3 mm of the tuber is stimulated by irradiation with red light at higher wavelengths (between 600 and 700 nm) (Liljemark and Widoff, 1960). While some studies have reported a correlation between greening and an increase in the GA levels in the specific potato cultivars studied (Spoladore et al., 1983, 1985; Dale et al., 1993; Zgorska et al., 2006), others have not found such a correlation (Mori and Kozukue, 1995; Edwards et al., 1998; Grunenfelder et al., 2006). Slightly greened tubers can be high in GAs, while highly greened tubers can have low GA levels. Thus, a direct relationship between greening of the potato tubers and an increase in the GAs levels has not been established. Various factors, e.g. light, temperature, as well as the potato variety influence the formation of both chlorophyll and GAs.
The storage temperature affected the levels of GAs in response to light exposure. Storage at 10°C prior to 6 weeks storage at 4°C has been reported to reduce the sensitivity of the tubers to subsequent light exposure (Griffiths et al., 1998).
Haase (2010) showed long‐term storage at 10°C without sprouting inhibitors led to an increase in GAs in two out of three potato cultivars (‘Cilena’ and ‘Lolita’), while storage at 4°C slightly increased the concentrations in these two cultivars. In a second experiment by the same authors, it was concluded that sprout control leads to a decrease in the levels of GAs. Potato tubers from other varieties were exposed to artificial light for 3 weeks at 12‐h intervals to simulate conditions at the point of sale, e.g. a supermarket. The authors reported that potatoes tended to a blue‐green colour, but no substantial increase in GA levels was noted.
An increase in the levels of GAs after mechanical damage or wounding of the tuber has been observed (e.g. Ahmed and Műller, 1978; Fitzpatrick et al., 1978; Mondy et al., 1987; Dale et al., 1998; Petersson et al., 2013; Dastmalchi et al., 2019). The combination of light and wounding was also reported to increase the levels of GAs, especially around the wound surface (Petersson et al., 2013; Chen et al., 2018; Nie et al., 2019).
The influence of the matrix in which the potatoes are stored has been studied under different storage conditions, such as temperature and light, and for different potato cultivars. Potatoes packaged in low light transmission packaging (e.g. craft paper coated with black polyethylene, high density polyethylene) have been reported to contain lower levels than those packed in high light transmission packages (e.g. polyethylene blue, transparent plastic film) (Bitsch et al., 1974; Rosenfeld et al., 1995; Abbasi et al., 2016). Other studies have reported that potatoes stored in polyethylene bags showed higher GA levels that those packaged in mesh or paper (Gosselin and Mondy, 1989).
Different methods to reduce the content of GAs in stored potatoes for later distribution and consumption have also been investigated. Submersion in different solutions such as 0.1 M of NaOH (pH 12) has been reported to reduce the α‐solanine and α‐chaconine content by 43% and 27%, respectively (Romanucci et al., 2018). Methods that inhibit sprouting, such as surface coverage with hydrophobic nanosilica (Zhang et al., 2018) or γ‐irradiation (Bergers, 1981; Dale et al., 1997; Afify et al., 2012) have also been proposed, as well as the addition of sulfur‐containing compounds, such as methionine, thiamine and garlic, which have been reported to reduce the GA content up to 60% and more (Surjawan et al., 2001; El‐Said, 2013; Karaway, 2014).
Several national bodies have provided a general advice to consumers on the home storage of potato tubers in relation to GAs. For example, the BfR,24 the Netherlands Nutrition Centre Foundation (Voedingscentrum)25 and the Swedish National Food Agency26 advise to store potatoes in a dark and cool place.
Other bodies have provided advice to food business operators on the storage of potatoes but with regard to the potential formation of acrylamide after further processing (UK‐FSA,27 US‐FDA,28 Commission Regulation (EU) 2017/215829). The latter stipulates that the temperature shall be appropriate to the potato cultivar stored and it shall be above 6°C, the level of humidity shall be such as to minimise senescent sweetening, and sprouting shall be suppressed in long‐term stored potatoes where permitted, using appropriate agents.
3.2.3.1.2. Processing of potatoes for food consumption
Potato tubers in general are only eaten after they have undergone some processing, which may take place in private households or in the food industry. Private household processing (domestic cooking) of potatoes may include the following methods:
peeling
boiling, blanching or steaming in water,
frying and deep frying in hot oil (‘French fries’ and crisps),
oven baking,
microwaving,
grating or cutting (e.g. followed by frying (potato pancakes) or boiling (potato dumplings))
Industrial processing of potatoes intended for food consumption includes the above‐mentioned methods. In addition, other processing methods may be applied for the production of other products such as dehydrated potato flakes and potato granules, flour and starch.
The scientific literature identified includes studies on the influence of single processing steps mentioned above on the resulting final content of GAs in the processed potato, and also studies on the effect of a typically industrial, multistep processing to produce, e.g. frozen French fries or potato chips. In this section, a summary of these studies is provided with the focus on the effectiveness of the most important processing steps with regard to reducing the GAs content in potato products.
Tuber peeling
A number of investigations provide values for the reduction of GAs in potatoes and potato products as a result of peeling. Since the major part of the total GA content in the tuber is situated just under the cork layer, peeling is expected to result in a reduction of the GA content. The reduction depends on the depth and peeling technique used (Rytel, 2012a).
In households, peeling of cooked potatoes is normally performed using a peeling knife. In the industry, the peeling of potatoes may take place using different types of mechanical peeling machines (abrasion or knife peelers), steam peeling machines or using caustic peeling (lye peeling) where the potato is surface treated with a caustic solution prior to the peeling (Muneta et al., 1977; Dornow Food Technology, 2019).
In Table 39, the reduction in GA content found in studies published in the open literature is shown. Overall, the reduction is between 25% and 75% and the average reduction is 48%.
Table 39.
Reduction in the total content of total glycoalkaloids (TGA) in S. tuberosum tubers as a result of peeling (as part of a commercial or experimental processing)
| Studya | Peeling method | Number of cultivars | Analytical method | TGA initial content in unpeeled tuber (mg/kg) | TGA content in peeled tuber (mg/kg) | Reduction in TGA content (%) |
|---|---|---|---|---|---|---|
| Sizer et al. (1980) | Hand peeling (1–2 mm)b | 1 | Non‐aqueous titration | 42.3 (fw) | 23.7 (fw) | 44% |
| Hellenäs et al. (1995) | Manual peelingb | 1 | HPLC‐UV | Cultivar ‘Magnum Bonum’: 336 (fw) | 243 (fw) | 30% |
| Panovská et al. (1997) | Manual peelingb | 7 | HPLC‐UV | 178 (fw) Mean of 7 cultivars | 82 (fw) Mean of 7 cultivars | 54% Mean of 7 cultivars |
| Ostrý et al. (2010) | Hand peeling (1 mm)b | 3d | HPTLC |
56–215 (fw) Across cultivars |
47–177 (fw) Across cultivars |
44% Across cultivars |
| Rytel et al. (2018) | Manual peelingb | 2 (coloured‐fleshed) | HPLC‐UV |
Cultivar ‘Valfi’: 269 (dw) ‘Blaue Elise’: 238 (dw) |
Cultivar ‘Valfi’: 97 (dw) ‘Blaue Elise’: 63 (dw) |
69% Mean of 2 cultivars |
| Rytel et al. (2005) | n.r. (taken from a French fries production line)c | 2 | HPLC‐UV |
Cultivar ‘Santana’: 209 (dw)e ‘Innowator’: 186 (dw)e |
Cultivar ‘Santana’: 127 (dw)e ‘Innowator’: 141 (dw)e |
32% Mean of 2 cultivars |
| Pęksa et al. (2006) | n.r. (taken from a chip processing line)c | 2 | HPLC‐UV |
Cultivar ‘Karlena’: 129 (dw) ‘Saturna’: 165 (dw) |
Cultivar ‘Karlena’: 101 (dw) ‘Saturna’: 118 (dw) |
25% Mean of 2 cultivars |
| Tajner‐Czopek et al. (2008) | n.r. | 3 | HPLC‐UV |
17.4 (fw) Mean of 3 cultivars |
7,4 (fw) Mean of 3 cultivars |
58% Mean of 3 cultivars |
| Mäder et al. (2009) | Steam‐peelingc | 1 | HPTLC | 294 (dw)f | 67 (dw)f | 77% |
| Tajner‐Czopek et al. (2012) | n.r. | 2 (coloured‐fleshed) | HPLC‐UV |
Cultivar ‘Rosalinde’: 248 (dw)e ‘Blue Congo’: 274 (dw)e |
Cultivar ‘Rosalinde’: 199 (dw)e ‘Blue Congo’: 219 (dw)e |
20% Mean of 2 cultivars |
| Rytel (2012a) | Laboratory carborundum peelerc | 1 | HPLC‐UV |
254 (dw) Mean of 4 batches |
175 (dw) Mean of 4 batches |
31% Mean of 4 batches |
| Rytel (2012b) | Steam‐peelingc | 1 | HPLC‐UV | 160 (dw)f | 81 (dw)f | 50% |
| Rytel (2012c) | Laboratory carborundum peelerc | 3 | HPLC‐UV |
225 (dw) Mean of 3 cultivars |
147 (dw) Mean of 3 cultivars |
35% Mean of 3 cultivars |
| Rytel (2013) | Laboratory carborundum peelerc | 1 | HPLC‐UV |
249 (dw)f Mean of 2 batches |
169 (dw)f Mean of 2 batches |
32% Mean of 2 batches |
| Rytel et al. (2013) | n.r. | 4 (coloured‐fleshed | HPLC‐UV |
250 (dw) Mean of 4 cultivars |
69 (dw) Mean of 4 cultivars |
72% Mean of 4 cultivars |
| Tajner‐Czopek et al. (2014) | Mechanical peelerc | 4 (coloured‐fleshed) | HPLC‐UV |
50 (fw) Mean of 4 cultivars |
25 (fw) Mean of 4 cultivars |
50% Mean of 4 cultivars |
| Rytel et al. (2015) | Carborundum peeler and manual selectionc | 3 | HPLC‐UV |
282 (dw) Mean of 3 cultivars |
196 (dw) Mean of 3 cultivars |
31% Mean of 3 cultivars |
TGA: total glycoalkaloids; dw: dry weight; fw: fresh weight; n.r.: not reported; LC: liquid chromatography; UV: ultraviolet; MS: mass spectrometry; MS/MS: tandem MS; HR: high resolution; HP: high performance; HPTLC: high‐performance thin layer chromatography.
Study year is in this case the year of publication.
Househod‐like peeling.
Industrial‐like peeling.
A total of 20 samples was tested.
Results were presented in figures, for α‐solanine and α‐chaconine.
Results were presented for α‐solanine and α‐chaconine.
Boiling, blanching or steaming in water
The effects of boiling, blanching and steaming in water on the content of GAs have been studied alone or as part of studies that included peeling and/or other methods of processing. This thermal process can further reduce the content of GAs, and the degree of reduction may differ depending on the applied temperatures and degree of disintegration of the raw materials (Rytel et al., 2018).
In Table 40, the results of some of these studies are shown. It should be noted that the reduction is given as the percent reduction in the GA content after peeling.
Table 40.
Reduction in the total content of total glycoalkaloids (TGA) in S. tuberosum tubers as a result of boiling, blanching and/or steaming (as part of a commercial or experimental processing)
| Studya | Processing step(s) | Number of cultivars | Analytical method | TGA initial content in peeled tuber (mg/kg) | TGA content in cooked tuber (mg/kg) | Reduction in TGA content (%) |
|---|---|---|---|---|---|---|
| Takagi et al. (1990) | Boiling in water | 3 | HPLC‐UV | 91–135e | 90–126e |
5% Mean of 3 cultivars |
| Tajner‐Czopek et al. (2008) | Boiling | 3 | HPLC‐UV |
7.4 (fw) Mean of 3 cultivars |
3,9 (fw) Mean of 3 cultivars |
48% Mean of 3 cultivars |
| Mulinacci et al. (2008) | Boiling in waterc | 3 (red‐ and blue‐fleshed) | HPLC‐UV |
Cultivar ‘Kennebec’: 46 (fw) ‘Vitelotte’: 79 (fw) ‘Highland Burgundy’: 180 (fw) |
Cultivar ‘Kennebec’: 29 (fw) ‘Vitelotte’: 66 (fw) ‘Highland Burgundy’: 153 (fw) |
23% Mean of 3 cultivars |
| Ostrý et al. (2010) | Boiling in salted water until edible stage | 3d | HPTLC |
117–217 (fw) Across cultivars |
102–180 (fw) Across cultivars |
19% Across cultivars |
| Tajner‐Czopek et al. (2012) | Boiling in water | 2 (coloured‐fleshed) | HPLC‐UV |
Cultivar ‘Rosalinde’: 199 (dw)f ‘Blue Congo’: 219 (dw)f |
Cultivar ‘Rosalinde’: 163 (dw)f ‘Blue Congo’: 156 (dw)f |
23% Mean of 2 cultivars |
| Tajner‐Czopek et al. (2012) | Boiling in waterc | 2 (coloured‐fleshed) | HPLC‐UV |
Cultivar ‘Rosalinde’: 248 (dw)f ‘Blue Congo’: 274 (dw)f |
Cultivar ‘Rosalinde’: 231 (dw)f ‘Blue Congo’: 247 (dw)f |
8% Mean of 2 cultivars |
| Lachman et al. (2013) | Boiling | 6 | LC‐MS/MS |
92.6 (dw) Mean of 6 cultivars |
79.7 (dw) Mean of 6 cultivars |
14% Mean of 6 cultivars |
| Rytel et al. (2005) | Blanching (2 stage) | 2 | HPLC‐UV |
Cultivar ‘Santana’: 127 (dw)e ‘Innowator’: 141 (dw)e |
Cultivar ‘Santana’: 55 (dw)e ‘Innowator’: 48 (dw)e |
62% Mean of 2 cultivars |
| Pęksa et al. (2006) | Blanching (including slicing and washing) | 2 | HPLC‐UV |
Cultivar ‘Karlena’: 101 (dw) ‘Saturna’: 118 (dw) |
Cultivar ‘Karlena’: 38 (dw) ‘Saturna’: 37 (dw) |
65% Mean of 2 cultivars |
| Mäder et al. (2009) | Blanching + steam cooking | 1 | HPTLC | 67 (dw)e | 34 (dw)e | 49% |
| Rytel (2012a) | Blanching | 1 | HPLC‐UV |
175 (dw) Mean of 4 batches |
126 (dw) Mean of 4 batches |
28% Mean of 4 batches |
| Rytel (2012a) | Steaming and cooling | 1 | HPLC‐UV |
175 (dw) Mean of 4 batches |
101 (dw) Mean of 4 batches |
42% Mean of 4 batches |
| Rytel (2012b) | Blanching | 1 | HPLC‐UV | 81 (dw)e | 60 (dw)e | 26% |
| Rytel (2012b) | Blanching + steaming | 1 | HPLC‐UV | 81 (dw)e | 38 (dw)e | 53% |
| Rytel (2012c) | Blanching of potato dices (including cutting and washing) | 3 | HPLC‐UV |
147 (dw) Mean of 3 cultivars |
110 (dw) Mean of 3 cultivars |
25% Mean of 3 cultivars |
| Rytel (2013) | Blanching of potato dices (5 min at 60°C) | 1 | HPLC‐UV |
169 (dw)f Mean of 2 batches |
123 (dw)f Mean of 2 batches |
27% Mean of 2 batches |
| Rytel (2013) | Blanching of potato dices (5 min at 75°C) | 1 | HPLC‐UV |
169 (dw)f Mean of 2 batches |
120 (dw)f Mean of 2 batches |
29% Mean of 2 batches |
| Rytel (2013) | Blanching of potato dices (5 min at 90°C) | 1 | HPLC‐UV |
169 (dw)f Mean of 2 batches |
100 (dw)f Mean of 2 batches |
41% Mean of 2 batches |
| Rytel et al. (2013) | Blanching of potato parts (5 min at 65°C) | 4 (coloured‐fleshed | HPLC‐UV |
69 (dw) Mean of 4 cultivars |
51 (dw) Mean of 4 cultivars |
26% Mean of 4 cultivars |
| Rytel et al. (2013) | Blanching of potato parts, (5 min at 75°C) | 4 (coloured‐fleshed | HPLC‐UV |
69 (dw) Mean of 4 cultivars |
45 (dw) Mean of 4 cultivars |
34% Mean of 4 cultivars |
| Rytel et al. (2013) | Blanching of potato parts (5 min at 85°C) | 4 (coloured‐fleshed | HPLC‐UV |
69 (dw) Mean of 4 cultivars |
35 (dw) Mean of 4 cultivars |
49% Mean of 4 cultivars |
| Tajner‐Czopek et al. (2014) | Blanching of potato strips (including cutting) | 4 (coloured‐fleshed) | HPLC‐UV |
111 (dw)f Mean of 4 cultivars |
93 (dw)f Mean of 4 cultivars |
16% Mean of 4 cultivars |
| Rytel et al. (2018)b | Blanching of potato strips (10 min at 75°C) | 2 (blue‐fleshed) | HPLC‐UV |
Cultivar ‘Valfi’: 97 (dw) ‘Blaue Elise’: 63 (dw) |
Cultivar ‘Valfi’: 60 (dw) ‘Blaue Elise’: 44 (dw) |
34% Mean of 2 cultivars |
| Rytel et al. (2018)b | Blanching of potato halves (20 min at 75°C) | 2 (blue‐fleshed) | HPLC‐UV |
Cultivar ‘Valfi’: 97 (dw) ‘Blaue Elise’: 63 (dw) |
Cultivar ‘Valfi’: 79 (dw) ‘Blaue Elise’: 51 (dw) |
19% Mean of 2 cultivars |
| Rytel et al. (2018)b | Blanching (20 min at 75°C) + steaming (20 min at 100°C) of potato halves | 2 (blue‐fleshed) | HPLC‐UV |
Cultivar ‘Valfi’: 97 (dw) ‘Blaue Elise’: 63 (dw) |
Cultivar ‘Valfi’: 64 (dw) ‘Blaue Elise’: 42 (dw) |
34% Mean of 2 cultivars |
dw: dry weight; fw: fresh weight; LC: liquid chromatography; UV: ultraviolet; MS: mass spectrometry; MS/MS: tandem MS; HR: high resolution; HP: high performance; HPTLC: high performance thin layer chromatography.
Study year is in this case the year of publication.
Study year is 2014–2015.
Experiments were conducted with unpeeled potatoes.
A total of 12 samples were tested.
Results were presented for α‐solanine and α‐chaconine.
Results provided in figures, for α‐solanine and α‐chaconine.
Note: the reduction is given as the percent reduction in the TGA content after peeling.
A considerable part of the GAs has been shown to be released into the boiling water in their intact form, both when the potatoes are boiled as no‐peeled and when boiled as peeled. Mondy and Gosselin (1988) reported higher TGA levels in potatoes boiled with peel than without peel (peel referred to in this study as the periderm and 1–2 mm of the cortex tissue). The authors also reported an increase in TGAs in the cortex tissue if the peel was not removed before boiling, concluding that GAs migrated from the peel into the cortex tissue during cooking.
Overall, boiling and certain blanching regimes result in reductions of between 5% and 65% and the average reduction is 33%.
Frying
A decrease in the content of TGA has been shown after frying in oil, e.g. for the production of French fries or potato crisps,30 and the results from several studies are summarised in Table 41.
Table 41.
Reduction in the total content of total glycoalkaloids (TGA) in S. tuberosum tubers as a result of frying for the production of French fries or potato crisps. Note: the reduction is given as the percent reduction in the TGA content after peeling
| Studya | Processing step(s) | Number of cultivars | Analytical method | TGA initial content in peeled tuber (mg/kg) | TGAs content in fried potato product (mg/kg) | Reduction in TGA content (%) |
|---|---|---|---|---|---|---|
| Takagi et al. (1990) | Frying (5 min at 150°C) | 3 | HPLC‐UV | 222–243c , d | 218–231d , e , f |
3% Mean of 3 cultivars |
| Takagi et al. (1990) | Frying (5 min at 170°C) | 4 | HPLC‐UV | 185–373c , d | 169–294d , e , f |
21% Mean of 4 cultivars |
| Takagi et al. (1990) | Frying (10 min at 210°C) | 3 | HPLC‐UV | 114–234c , d | 64–149d , e , f |
38% Mean of 3 cultivars |
| Rytel et al. (2005) | Frying (170–180°C, 2 stage) (French fries processing) | 2 | HPLC‐UV |
Cultivar ‘Santana’: 127 (dw)f ‘Innowator’: 141 (dw)f |
Cultivar ‘Santana’: 14 (dw)f ‘Innowator’: 6 (dw)f |
92% Mean of 2 cultivars |
| Pęksa et al. (2006) | Frying at 180°C (chip processingb) | 2 | HPLC‐UV |
Cultivar ‘Karlena’: 101 (dw) ‘Saturna’: 118 (dw) |
Cultivar ‘Karlena’: 24 (dw) ‘Saturna’: 23 (dw) |
79% Mean of 2 cultivars |
| Tajner‐Czopek et al. (2012) | Frying at 180°C (potato crisp processing)c | 2 (coloured‐fleshed) | HPLC‐UV |
Cultivar ‘Rosalinde’: 248 (dw)f ‘Blue Congo’: 274 (dw)f |
Cultivar ‘Rosalinde’: 41 (dw)f ‘Blue Congo’: 45 (dw)f |
83% Mean of 2 cultivars |
| Tajner‐Czopek et al. (2012) | Frying (175°C, 2 stage) (French fries processing) | 2 (coloured‐fleshed) | HPLC‐UV |
Cultivar ‘Rosalinde’: 199 (dw)f ‘Blue Congo’: 219 (dw)f |
Cultivar ‘Rosalinde’: 12 (dw)f ‘Blue Congo’: 16 (dw)f |
93% Mean of 2 cultivars |
| Tajner‐Czopek et al. (2014) | Frying (175°C, 2 stage) (French fries processing) | 4 (coloured‐fleshed) | HPLC‐UV |
111 (dw)f Mean of 4 cultivars |
7 (dw)f Mean of 4 cultivars |
94% Mean of 4 cultivars |
| Rytel et al. (2015) | Frying (2 min at 175°C) including slicing and washing (chips processing) | 3 | HPLC‐UV |
196 (dw) Mean of 3 cultivars |
29 (dw) Mean of 3 cultivars |
85% Mean of 3 cultivars |
| Rytel et al. (2018) | Frying (175°C, 2‐stage) (French fries processing) | 2 (blue‐fleshed) | HPLC‐UV |
Cultivar ‘Valfi’: 97 (dw) ‘Blaue Elise’: 63 (dw) |
Cultivar ‘Valfi’: 9.5 (dw) ‘Blaue Elise’: 7.0 (dw) |
89% Mean of 2 cultivars |
| Rytel et al. (2018) | Frying (2 min at 190°C)(potato crisps processing) | 2 (blue‐fleshed) | HPLC‐UV |
Cultivar ‘Valfi’: 97 (dw) ‘Blaue Elise’: 63 (dw) |
Cultivar ‘Valfi’: 53 (dw) ‘Blaue Elise’: 28 (dw) |
50% Mean of 2 cultivars |
TGA: total glycoalkaloids; dw: dry weight; fw: fresh weight; LC: liquid chromatography; UV: ultraviolet; HP: high performance.
Study year is in this case the year of publication.
It is not clear whether the final products refer to French fries or to potato crisps.
Experiment conducted with unpeeled potatoes.
Results expressed as raw weight, corrected for water loss during frying.
Results were presented for α‐solanine and α‐chaconine.
Results provided in figures, for α‐solanine and α‐chaconine.
Takagi et al. (1990) studied the effect of frying temperature on the TGA content in peeled potato cubes. At 150°C, there was only a minor loss (5%) of GAs; at 170°C, the loss increased to 21%; and at 210°C, TGA levels were reduced by 38%. It was remarked by the authors that 170°C is the standard temperature for frying, while frying at 150°C or at 210°C is outside the typical range.
Rytel et al. (2005) reported a reduction of 62–79% (from 48–55 mg/kg dw to 10–21 mg/kg dw) after first‐stage frying for 1 min of blanched potato strips, while a reduction of 75–87% (48–55 mg/kg dw to 6–14 mg/kg dw) was obtained after two‐stage frying for an additional 5 min. A reduction of approx. 92% was reported by Tajner‐Czopek et al. (2014) when blanched potato strips were fried at 175°C for 5–6 min. Rytel et al. (2018) reported a reduction of 84% for blanched potato strips that were fried for 5–6 min at 175°C (from 43.9–60.4 mg/kg dw to 7.0–9.5 mg/kg dw).
Pęksa et al. (2006) reported that during frying of blanched potato slices at 170–180°C to produce chips, the TGA levels were reduced by 39% from 82.1–85.3 mg/kg dw to 22.6–23.5 mg/kg dw). Chips produced from peeled raw potato slices that were fried for 2 min at 190°C had a 45–54% reduced TGA level (from 62.7–97.4 mg/kg dw to 28.3–53.0 mg/kg dw) (Rytel et al., 2018).
In some of these studies, the ratio α‐solanine:α‐chaconine was studied and it remained constant regardless of the different conditions (e.g. temperature and time) employed in the studies (Takagi et al., 1990; Pęksa et al., 2006; Rytel et al., 2018).
In conclusion, a reduction between 20% and 90% in the final fried potato product compared to the peeled potato is reported.
Other processing methods
The effects of baking and microwaving on the content GAs have also been studied. In Table 42, the results of some of these studies are shown. It should be noted that the reduction is given as the percent reduction compared to the original GA content in the unpeeled potato.
Table 42.
Reduction in the total content of total glycoalkaloids (TGA) in S. tuberosum tubers as a result of baking or microwaving
| Studya | Processing step(s) | Number of cultivars | Analytical method | TGA initial content in fresh tuber (mg/kg) | TGA content in processed tuber (mg/kg) | Reduction in TGA content (%) |
|---|---|---|---|---|---|---|
| Bushway and Ponnampalam (1981) | Bakingb | 2 | HPLC‐UV | 103–161 (fw) | 99–113 (fw) |
20% Mean of 2 cultivars |
| Lachman et al. (2013) | Bakingb | 6 | LC‐MS/MS |
267 (dw) Mean of 6 cultivars |
135 (dw) Mean of 6 cultivars |
49% Mean of 6 cultivars |
| Bushway and Ponnampalam (1981) | Microwavingb | 2 | HPLC‐UV | 103–161 (fw) | 124–133 (fw) |
3% Mean of 2 cultivars |
| Takagi et al. (1990) | Microwavingc | 3 | HPLC‐UV | 86–239d | 72–208d |
15% Mean of 3 cultivars |
| Lachman et al. (2013) | Microwavingb | 6 | LC‐MS/MS |
267 (dw) Mean of 6 cultivars |
146 (dw) Mean of 6 cultivars |
45% Mean of 6 cultivars |
| Mulinacci et al. (2008) | Microwavinga | 3 (red‐ and blue‐fleshed) | HPLC‐UV |
Cultivar ‘Kennebec’: 46 (fw) ‘Vitelotte’: 79 (fw) ‘Highland Burgundy’: 180 (fw) |
Cultivar ‘Kennebec’: 38 (fw) ‘Vitelotte’: 75 (fw) ‘Highland Burgundy’: 168 (fw) |
7% Mean of 3 cultivarsb |
TGA: total glycoalkaloids; LC: liquid chromatography; UV: ultraviolet; MS/MS: tandem mass spectrometry.; HP: high performance.
Study year is in this case the year of publication.
Experiments were conducted with unpeeled potatoes.
Experiments were conducted with peeled potatoes, cut into cubes of 5–10 mm.
Results were presented for α‐solanine and α‐chaconine.
Note: The reduction is given as the percent reduction compared to the original GA content in the unpeeled potato.
Treatment of non‐peeled tubers, cut into cubes, at 750 W for 10 min in a microwave oven resulted in an average reduction of the TGA content of 45% (from 267 to 146 mg/kg dw) (Lachman et al., 2013). Lower reductions (15%) were reported by Takagi et al. (1990), while Mulinacci et al. (2008) did not observe significant variations in the content of GAs in microwaved unpeeled potatoes compared to fresh potatoes. In a comparative study by Bushway and Ponnapalam et al. (1981), baking and microwaving of potatoes resulted in lower levels (99–113 and 124–133 mg/kg fw, respectively) compared to levels in raw potatoes (103–161 mg/kg fw). The study by Lachman et al. (2013) reported that baking non‐peeled tubers diced into small cubes resulted in nearly the same reduction, i.e. from 267 to 134 mg/kg dw as did microwaving (from 267 to 146 mg/kg).
When dried potato powder was heated to 150°C for 2 h, α‐solanine and α‐chaconine were found to be stable (Nie et al., 2018). Above 150°C both GAs started to degrade, α‐chaconine being somewhat less stable than α‐solanine.
Rytel et al. studied the effects of drying in a (semi‐)industrial setting for the production of diced potatoes, granulates and powder (Rytel, 2012a,b,c, 2013; Rytel et al., 2013, 2018). Drying of peeled and sliced potatoes at temperatures between 120°C and 160°C for 1–2 h, resulted in reduction of the TGA levels in the final product (dried dices, flakes, powder) of 29% to 67% (Table 43). A reduction in GA content of 57% was reported by Mäder et al. (2009) in the production of dried potato flakes from peeled potatoes.
Table 43.
Reduction in the total content of total glycoalkaloids (TGA) in S. tuberosum tubers as a result of drying (preceeded by boiling or blanching and cutting steps)
| Studya | Processing step(s) | Number of cultivars | Analytical method | TGA initial content in peeled tuber (mg/kg) | TGA content in processed tuber (mg/kg) | Reduction in TGA content (%) |
|---|---|---|---|---|---|---|
| Rytel et al. (2005) | Drying | 2 | HPLC‐UV |
Cultivar ‘Santana’: 127 (dw)b ‘Innowator’: 141 (dw)b |
Cultivar ‘Santana’: 43 (dw)b ‘Innowator’: 36 (dw)b |
30% Mean of 2 cultivars |
| Mäder et al. (2009) | Drum drying at 160°C | 1 | HPTLC | 67 (dw)b | 29 (dw)b | 57% |
| Rytel (2012a) | Drying (2 h at 160°C) | 1 | HPLC‐UV |
175 (dw) Mean of 4 batches |
65 (dw) Mean of 4 bathces |
63% Mean of 4 batches |
| Rytel (2012b) | Drying (pneumatic) | 1 | HPLC‐UV | 81 (dw)b | 34 (dw)b | 58% |
| Rytel (2012b) | Drying (fluidised bed) | 1 | HPLC‐UV | 81 (dw)b | 31 (dw)b | 62% |
| Rytel (2012b) | Drying (1 h at 120°C) | 3 | HPLC‐UV |
193 (dw) Mean of 3 cultivars |
81 (dw) Mean of 3 cultivars |
62% Mean of 3 cultivars |
| Rytel (2012c) | Drying (1 h at 120°C) (potato dices)a | 3 | HPLC‐UV |
147 (dw) Mean of 3 cultivars |
104 (dw) Mean of 3 cultivars |
29% Mean of 3 cultivars |
| Rytel (2013) | Drying (1 h at 120°C) (potato dices)a | 1 | HPLC‐UV |
169 (dw)b Mean of 2 batches |
113 (dw)b Mean of 2 batches |
33% Mean of 2 batches |
| Rytel (2013) | Drying (1 h at 140°C) (potato dices)a | 1 | HPLC‐UV |
169 (dw)b Mean of 2 batches |
108 (dw)b Mean of 2 batches |
36% Mean of 2 batches |
| Rytel (2013) | Drying (1 h at 160°C) (potato dices)a | 1 | HPLC‐UV |
169 (dw)b Mean of 2 batches |
100 (dw)b Mean of 2 batches |
41% Mean of 2 batches |
| Rytel et al. (2013) | Drying (1 h at 120°C) (potato dices)a | 4 (coloured‐fleshed) | HPLC‐UV |
69 (dw) Mean of 4 cultivars |
49 (dw) Mean of 4 cultivars |
29% Mean of 4 cultivars |
| Rytel et al. (2013) | Drying (1 h at 140°C) (potato dices)a | 4 (coloured‐fleshed) | HPLC‐UV |
69 (dw) Mean of 4 cultivars |
40 (dw) Mean of 4 cultivars |
43% Mean of 4 cultivars |
| Rytel et al. (2013) | Drying (1 h at 160°C) (potato dices)a | 4 (coloured‐fleshed) | HPLC‐UV |
69 (dw) Mean of 4 cultivars |
38 (dw) Mean of 4 cultivars |
46% Mean of 4 cultivars |
| Rytel et al. (2018) | Drying (2 h at 160°C) (potato puree) | 2 (blue‐fleshed) | HPLC‐UV |
Cultivar ‘Valfi’: 97 (dw) ‘Blaue Elise’: 63 (dw) |
Cultivar ‘Valfi’: 51 (dw) ‘Blaue Elise’: 35 (dw) |
46% Mean of 2 cultivars |
TGA: total glycoalkaloids; LC: liquid chromatography; UV: ultraviolet; MS/MS: tandem mass spectrometry; HP: high performance.
Study year is in this case the year of publication.
Experiments were conducted with peeled potatoes, cut into cubes of 10 mm.
Results were presented for α‐solanine and α‐chaconine.
Note: The reduction is given as the percent reduction in the TGA content after peeling.
Industrial multistep processing of potatoes
Studies presenting the complete process chain for the production of potato‐based products are available. Some of these studies provide data on the reduction of the GA content in the intermediate as well as the initial and final products. The methods of processing leading to the different final potato products may differ according to the end product as well as according to the producer. In general, the more recent methods will include the highest number of steps. Examples on production schemes for French fries and dehydrated potato flakes are shown in Figure 7.
Figure 7.
Examples of production schemes for (A) French fries (from Tajner‐Czopek et al., 2014, 31) and (B) dehydrated potato flakes from the cultivar ‘Karlena’ (from Mäder et al., 2009)
In the production of French fries, Tajner‐Czopek et al. (2014) reported reductions of up to 97.5% after exposing the potatoes to six steps of processing (Figure 7A). Similar results were obtained by Rytel et al. (2005) who reported that the levels in two potato cultivars processing (peeling, cutting, two‐stage blanching, drying and two‐stage frying) decreased from an initial content of 209 and 186 mg/kg dw (unpeeled potato tubers) to 14 and 6 mg/kg dw, i.e. reductions of around 93 and 98%, respectively, for the two cultivars studied. The same authors reported, in a later study, a reduction of 97% and of 84% in the production of French fries and potato crisps, respectively (Rytel et al., 2018).
The production of dehydrated potato flakes was studied using one cultivar with a starting content on TGA of 294 mg/kg dw, which after peeling, blanching, steam cooking, mashing and drying resulted in a TGA concentration of 29 mg/kg dw (corresponding to 5.8 mg/kg fw for reconstituted flakes), i.e. a reduction of around 90% (Mäder et al., 2009). Rytel et al. (2018) reported a reduction of 83% in the multi‐step production of dehydrated potato (puree).
Rytel (2012b) studied the effects of industrial potato processing to obtain potato granules (dehydrated) and the changes in GA content (α‐solanine and α‐chaconine) at the different stages. The processing included steam peeling, slicing, blanching, cooling in water, steaming and a two‐stage drying (pneumatic and fluidisation drying) to finally obtain a granulated product. After steam peeling, the TGA content decreased by 50% on average (from 160 to 81 mg/kg dw) (see Table 39). The authors observed a somewhat larger decrease for α‐solanine (55%) than for α‐chaconine (46%). After blanching, a further decrease of 26% compared to the peeled potatoes was observed (from 81 to 59.6 mg/kg dw). In this case, a larger decrease in α‐chaconine content (29%) compared to α‐solanine (21%) was observed. Cooling in water did not result in significant changes in the TGA content, but the subsequent steaming step decreased TGA by 35% on average (from 59.6 to 38.1 mg/kg dw). The last processing step of drying resulted in a further TGA decline from 38.1 to 31.1 mg/kg dw). The final granulated product contained 26.6 mg/kg dw, i.e. 16% of the original TGA content in unpeeled potatoes.
Potato peel
Bushway and Ponnampalam (1981) studied the effect of frying (4 min at 177°C) on the TGA content in raw potato peels. A reduction of 16–22% (678–763 mg/kg to 567–594 mg/kg, corrected for moisture loss) was observed.
Similar results were obtained by Bushway et al. (1983). Starting from raw potato peels, containing GAs in the range of 18.0–1,068 mg/kg across cultivars, the levels of GAs in baked potato peels were 24–1,098 mg/kg across cultivars, in fried peels 32.7–1,649 mg/kg and in baked‐fried 41.7–1,557 mg/kg (all concentrations uncorrected for moisture loss). The authors attributed this difference to the greater moisture loss during frying than during baking.
3.2.3.1.3. Processing of potatoes for feed
Whole potatoes intended for use as feeds for ruminants normally undergo no further processing,32 and therefore, no effects on GA concentrations would be expected. When the supply of potatoes exceeds demand, they may also be ensiled (either alone or with the addition of a green forage), but no reports have been identified on the effects of ensiling on the GA contents.
For pigs, it is generally recommended that potatoes are cooked or steamed before being fed (Whittemore, 1977), and the effects of these processes on GA concentrations when applied to the production of foods, as reported above, may also be expected to apply to feed.
A number of by‐products of potato processing are used as feeds for livestock and companion animals. These include potato protein and potato peelings. Where the peels have been removed by steaming, it should be noted that GAs are stable to decomposition at cooking temperatures below 150°C, and therefore, potato peelings obtained by steaming might represent a risk to livestock, depending on the extent of greening in the tubers (Bushway and Ponnampalam, 1981; Takagi et al., 1990).
Drying of industrial potato peels resulted in an increase of GA levels (Hossain et al., 2016). The levels of α‐solanine increased from ~ 100 mg/kg dw in the fresh peels to nearly 400 mg/kg dw after drying either by air‐, vacuum oven‐ or freeze‐drying. Similar behaviour was observed for α‐chaconine, increasing from ~ 50 to ~ 600 mg/kg dw. Air drying showed the highest increase in GA levels. The levels of solanidine and demissidine appeared not to be affected by the drying process.
Potato peelings produced as a by‐product from industrial processing were stored for 4 days at room temperature to investigate the effect on the GA content. A slow degradation of the GAs was observed with calculated half‐lifes in the range of 2.9–9.5 days, depending on the size of the potato peel wastes (Sepelevs et al., 2019).
3.2.3.2. GAs from food plants other than S. tuberosum
The α‐tomatine concentration decreases during fruit ripening (see Section 1.3.3.2). The concentration in the ripe fruit furthermore differs depending on the cultivar. While a number of published investigations on the effect of different processing steps on the GA concentration in potatoes and potato products exist, those regarding tomato and tomato products are scarce. Examples of the content of α‐tomatine in different tomato products after processing are shown in Table 37.
Friedman and Levin (1995) reported that microwaving of green tomatoes only caused a 7% loss of α‐tomatine, while preparation of fried green tomatoes caused a 22% loss, when adjusting for added ingredients and moisture changes. No significant reduction was noted for the content of α‐dehydrotomatine (measured as ‘solanine’33) upon preparation of green tomato marmalade, while preparation of pickled green tomatoes resulted in a loss of 11% (Kibler et al., 1985). Kyzlink et al. (1981) found that cooking for 5 min of unripe green tomatoes resulted in a loss of 0–15% α‐tomatine. Freeze‐drying of the produced tomato homogenate and subsequent storage for 4 weeks at room temperature, resulted in a reduction (corrected for water loss) of the α‐tomatine content by 82–85% (Kyzlink et al., 1981). Voldrich et al. (1992) also noted that the α‐tomatine content of tomatoes and tomato products (fresh or fermented) declines upon storage. Two batches of unripe green tomatoes did not contain measurable amounts of α‐tomatine after been stored at 4°C for 10 weeks (starting content 52 and 83 mg/kg fw, respectively). Similar results were found for products of green tomato salad, fermented green tomato and tomato puree that had been stored for 2 months (Voldrich et al., 1992). Stoyanova et al. (1988) reported for pickled immature tomatoes a reduction in α‐tomatine content from an initial 360–400 mg/kg fw to 240–340 mg/kg fw after 10 days and to 120–160 mg/kg fw after 180 days. After 6 months of storage, the tomatoes had lost their initial bitter taste.
A single study has been published on the effect of cooking and grilling on the content of GAs in three cultivars of aubergine. Boiling for 10 min reduced the TGA content by 6–35% due to leaching, while grilling for 5 min resulted in 8% to 107% increase in TGA content, what the authors contributed to a water loss of the grilled product (Lo Scalzo et al., 2016).
3.2.3.3. Summary on the influence of storage and processing on the levels of GAs
For potatoes, factors such as the duration of storage, temperature, light exposure, as well as tuber damage can affect the GA content during storage of potato tubers. Varying results have been obtained in the different studies depending on the conditions and the potato cultivar studied. In general, longer storage times, exposure to light and tuber damage have been reported to induce the formation of GAs. The influence of temperature is dependent on the potato cultivar and other conditions tested in the studies.
Processing of potatoes has been reported to reduce the content of GAs in the intermediate and/or final processed product. A substantial variation in processing factors has been reported in the literature. In general, the peeling of potatoes reduced the GA content by 25% up to 75%, depending on the peeling method used. Boiling in water and blanching/steaming reduced the GA content in peeled potatoes in the range of 5–65%. Frying in oil lowered the content from 20% up to 90% compared to the peeled potato tuber. Microwave and oven baking of unpeeled potatoes have also been reported to cause a reduction in the GA content by 3–45% and by 20–50%, respectively. In general, drying of peeled potato and final processing to flakes, granules or powder led to a reduction of the GA content by 29–67%. No specific studies on the effects of roasting were indentified. The reductions in TGA content observed for the various processing steps were used to define the processing factors that were applied in the exposure assessment as described in Section 2.5. Based on the available information, the CONTAM Panel decided to use for the peeling of potatoes a processing factor ranging from 0.25 to 0.75, for frying and deep frying a processing factor ranging from 0.1 to 0.8, and for all other heat treatment methods a processing factor ranging from 0.35 to 0.95.
No information has been found about the chemical nature of the GAs degradation products.
For tomatoes, the ripening process leads to a decrease of the content of α‐tomatine and α‐dehydrotomatine. Factors that may lead to an increase of α‐tomatine and α‐dehydrotomatine after harvesting have not been identified. Processing of tomatoes may result in modest losses of α‐tomatine of up to 15%. Storage at low temperatures may slow down the ripening of tomatoes and the concomitant decrease of α‐tomatine, but does not fully stop it. Storage of tomato products also results in a loss of α‐tomatine over time.
For aubergine, the ripening process has been reported to result in an increase of the GA content. Other factors that may influence the formation of GAs have not been reported. Boiling in water may reduce the TGA content by 6–35%, while grilling may lead to an increased TGA content in the grilled product, due to a lower water content.
3.3. Exposure assessment
3.3.1. Current acute dietary exposure assessment for humans
The CONTAM Panel assessed the acute dietary exposure to potato TGAs (sum of α‐solanine and α‐chaconine) in food following the methodology described in Section 2.5.
A scenario including only days in which there was consumption of main‐crop potatoes was considered the most relevant for assessing the risk following acute exposure. The summary statistics of the probabilistic acute dietary exposure assessment to potato TGAs across European dietary surveys by age group are shown in Table 44. Detailed results for each survey are shown in Annex A.6.
Table 44.
Summary statistics of the probabilistic acute dietary exposure assessment to potato TGAs across European dietary surveys (μg/kg bw per day) by age group including only days with consumption. The corresponding 95% confidence intervals are presented in the brackets
| Age class | Range across surveys of acute dietary exposure (μg/kg bw per day) | |||
|---|---|---|---|---|
| Mean | P95a | |||
| Minimum | Maximum | Minimum | Maximum | |
| Infants | 32.3 (20.5–49.9) | 119.9 (114.3–125.9) | 309.0 (256.1–374.4) | 375.5 (343.5–410.6) |
| Toddlers | 59.1 (57.2–61.2) | 174.0 (139.5–221.2) | 231.4 (218.3–245.5) | 535.1 (349.6–822.9) |
| Children | 45.6 (42.5–49.4) | 149.2 (135.3–165.0) | 149.2 (130.6–171.2) | 444.9 (374.4–539.4) |
| Adolescents | 28.8 (27.2–30.5) | 109.8 (101.3–118.2) | 94.1 (84.8–104.3) | 331.7 (287.9–381.5) |
| Adults | 23.3 (21.4–25.6) | 82.5 (79.2 – 86.2) | 78.3 (67.0–93.4) | 244.8 (227.7–264.6) |
| Elderly | 27.4 (22.7–33.5) | 72.0 (64.7–80.4) | 88.7 (83.2–94.6) | 210.7 (172.8–255.5) |
| Very elderly | 30.3 (27.1–34.1) | 70.7 (58.2–83.9) | 85.7 (76.1–96.5) | 214.0 (154.9–297.3) |
bw: body weight; P95: 95th percentile.
One dietary survey had less than 60 days in the Infants and Toddlers age groups; therefore, these were not included in calculation of the 95th percentile exposure.
The mean exposure across surveys ranged from 23.3 μg/kg bw per day in adults to 174.0 μg/kg bw per day in toddlers. The 95th percentile exposure ranged from 78.3 μg/kg bw per day in adults to 535.1 μg/kg bw per day in ‘Toddlers’ (up to 822.9 μg/kg bw per day in the upper limit of the 95% confidence interval).
Figure 8 shows the mean amounts (in grams per day) of main‐crop potatoes consumed by adults or toddlers via different food categories. Method details are given in Section 2.5. Data were available for 22 European countries. In the age group ‘Adults’ the largest part of main‐crop potatoes is consumed as ‘Starchy roots and tubers’, followed by the food categories ‘Composite food’ and ‘Grains and grain‐based products’. Food surveys for toddlers show that the food category ‘Starchy roots and tubers’ is predominant with no significant differences among the 11 European countries, for which data are available. These findings indicate that the main dietary GA source via potatoes and potato‐derived products for adults and toddlers in Europe are ‘Starchy roots and tubers’ while other food categories appear to be of considerably less importance.
Figure 8.
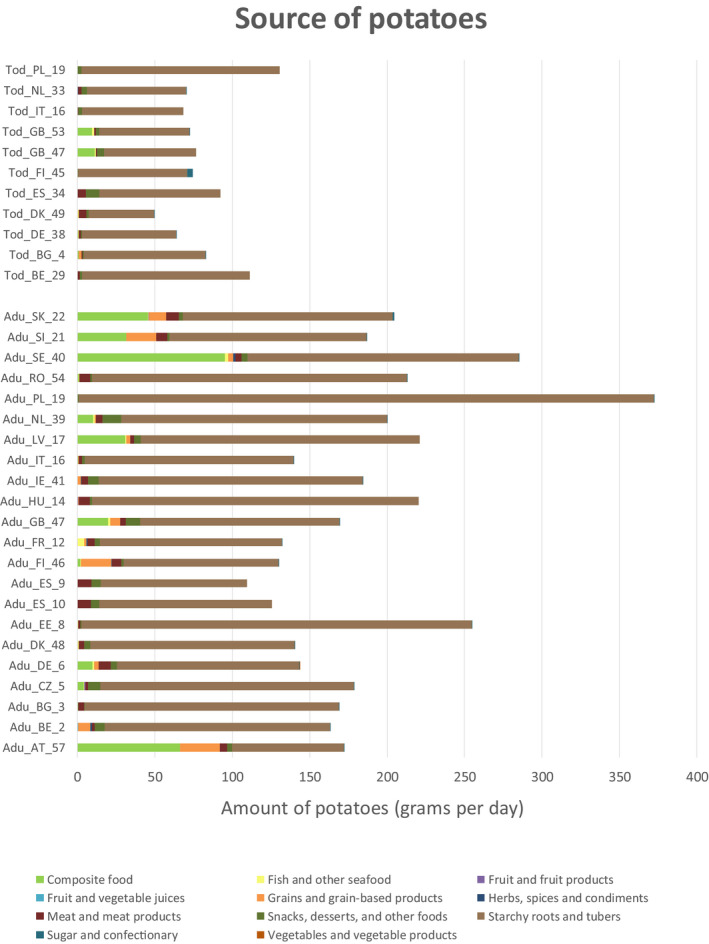
Mean daily contribution (in grams per day) of different food categories to the consumption of main‐crop potatoes calculated on survey days by Adults and Toddlers across different consumption surveys
3.3.2. Previously reported dietary exposure assessments
Ruprich et al. (2009) estimated the acute and chronic exposure to GAs using probabilistic modelling for Sweden, Czech Republic and the Netherlands. The authors combined the data on occurrence of α‐solanine and α‐chaconine in potato tubers available from the Czech Republic (n = 200 analytical results, peeled raw potatoes, year 2004–2005) and from Sweden (n = 239 analytical results, raw unpeeled potatoes, year 1997) to perform the overall exposure assessment for the three countries. Since the data from the Czech Republic referred to peeled potatoes, a factor of 1.25 was applied to correct for GAs losses during peeling. The final data set contained 439 results for raw unpeeled potatoes. In addition, for the acute exposure estimation, four other scenarios were considered to simulate a situation in which raw unpeeled potatoes contained limited quantities of GAs, by deleting concentrations over 200, 150, 100 or 50 mg/kg. This resulted in four occurrence data sets with the following median (average, range) concentrations:
Complete dataset (n = 439): 64.0 (70.4, 6.3–302.9) mg/kg
Cut‐off 200 mg/kg (n = 421): 62.5 (63.5, 6.3–197.8) mg/kg
Cut‐off 150 mg/kg (n = 400): 60.2 (58.0, 6.3–149.9) mg/kg
Cut‐off 100 mg/kg (n = 333): 48.1 (45.5, 6.3–99.8 mg/kg
Cut‐off 50 mg/kg (n = 173): 15.1 (18.2, 6.3–49.4) mg/kg
In addition, processing factors were included in the assessment to correct for losses of GAs during processing of potatoes. Factors of 0.8, 0.64 and 0.17, respectively, were considered for peeling, for peeling and cooking and for frying or baking in fat. Food consumption data were retrieved from the national databases and for the Czech Republic comprised subjects from 4 to 90 years old, for Sweden subjects from 17 to 74 years old, and for the Netherlands subjects from 1 to 97 years old. The consumption data on potato products were recalculated at raw agricultural commodity level. The probabilistic estimation was done using the Monte Carlo Risk Assessment software.
Mean (and upper 97.5% confidence limit) acute exposure estimates, when all occurrence data were considered were 80 (85), 80 (87) and 78 (84) μg/kg bw per day for the Czech Republic, Sweden and the Netherlands, respectively. The calculated P95 (and upper 97.5% confidence limit) acute dietary exposures were 358 (383), 339 (366), 319 (344) μg/kg bw per day for the Czech Republic, Sweden and the Netherlands, respectively. The estimates were lower when the different cut‐off scenarios were used (200, 150, 100 and 50 mg/kg cut off). For the Czech Republic, the mean acute exposure estimates for these scenarios were 72, 66, 52 and 21 μg/kg bw per day, respectively; for Sweden, they were 73, 67, 53 and 21 μg/kg bw per day, respectively; and for the Netherlands, they were 71, 64, 51 and 20 μg/kg bw per day, respectively. The P95 acute exposures using the different cut‐off scenarios were for the Czech Republic: 318, 288, 228 and 106 μg/kg bw per day, respectively; for Sweden: 302, 273, 217 and 100 μg/kg bw per day, respectively; and for the Netherlands, 283, 258, 205 and 94 μg/kg bw per day, respectively. Median (and upper 97.5% confidence limit) estimates of chronic intake were 74 (79), 73 (79) and 154 (166) μg/kg bw per day, respectively, for Czech Republic, Sweden and the Netherlands. The authors noted that the differences in intake estimates between countries were due not only to different consumption habits but also due to the structure and complexity of the food consumption database used in each case. Considering 1 mg/kg bw per day as the critical exposure dose for adverse effects, the authors concluded that 0.01% of the population in all three countries exceeded this value. No chronic estimates were above that level.
Pariera Dinkins and Peterson (2008) estimated the acute oral exposure to GAs (sum of α‐solanine and α‐chaconine) considering the occurrence in the inner tissue of tubers from plants defoliated by Colorado potato beetles and with no defoliation (control) for different age groups within the US population. The authors used a data set for peeled potatoes with mean (standard deviation) GAs concentrations of 174.5 ± 8.59 mg/kg dw for the control and 258.8 ± 23.3 mg/kg dw for Colorado potato beetle defoliation samples. Estimates of acute exposure were higher in the scenario with Colorado potato beetle defoliation samples compared to the control scenario. The median exposure estimates for the US population were 239 and 354 μg/kg bw per day for the control scenario and for the Colorado potato beetle defoliation scenario, respectively. Acute oral exposures were also estimated for seven population subgroups including infants, children 7–12 years old, youth 13–19 years old, women > 13 years old, women > 20 years old and males > 20 years old. The median of exposure in the control scenario ranged from 190 to 515 μg/kg bw per day, while in the potato beetle defoliation scenario, it ranged from 282 to 760 μg/kg bw per day. The estimated P95 acute exposure ranged from 406 to 1,131 μg/kg bw per day in the control scenario and from 603 to 1,676 μg/kg bw per day in the potato beetle defoliation scenario. Highest exposures were calculated for the age group children 1–6 years. It should be noted that it is not indicated whether processing factors for heat processing steps were applied to calculate the exposure.
A detailed comparison between these studies and the current exposure assessment is not possible, since the three studies used different data sets, age groups and exposure models. Nevertheless, all studies indicate that there is a likelihood that the LOAEL of 1 mg/kg bw per day is exceeded in a number of cases, with the largest probability for the younger age groups.
3.3.3. Current dietary exposure assessment for farm animals, horses and companion animals
While potatoes are not widely used as feeds for livestock, in some situations they may represent an important component of the ration, for example, when production exceeds market requirements for human consumption. Potato by‐products, particularly potato protein concentrate (PPC) and potato starch, are used to a limited extent in compound feeds for certain non‐ruminant livestock and companion animals. However, insufficient data were available on levels of GAs in either potatoes or potato by‐products used as animal feeds to allow estimates of exposure to be made. A number of studies have examined the potential use of PPC in diets for farmed fish as a replacement for fish meal. Dietary inclusion rates as low as 5% have resulted in reductions in feed intake, feed utilisation efficiency, growth rates and body composition for rainbow trout (Xie and Jokumsen, 1998) and salmon (Norwegian Scientific Committee for Food Safety, 2009). This has been attributed to the presence of solanidine GAs in the PPC. However, if the PPC is processed to remove these, inclusion rates of up to 20% are possible in diets of farmed salmon (Refstie and Tiekstra, 2003; Norwegian Scientific Committee for Food Safety, 2009).
3.4. Risk characterisation
3.4.1. Human health risk characterisation
3.4.1.1. GA from edible parts of S. tuberosum
The available data on acute toxicity were considered insufficient to establish a HBGV, and instead the CONTAM Panel used an MOE approach to assess a possible health concern from acute exposure to potato TGAs via food. An MOE higher than 10 indicates that there is no health concern. This MOE of 10 takes into account the interindividual variability in toxicodynamics (a factor of 3.2), and extrapolation from a LOAEL to a NOAEL (a factor of 3).
The LOAEL of 1 mg potato TGA/kg bw in humans was used as reference point to characterise the risk following acute exposure.
The CONTAM Panel calculated the acute dietary exposure using a probabilistic approach and applying processing factors to estimate the GA levels in the final food consumed as described in Sections 2.5, 3.2.1 and 3.3.1. Comparison of the acute exposure estimates (see Table 44) to the reference point of 1 mg potato TGAs/kg bw per day resulted in the MOE values that are presented in Table 45 for acute mean and P95 exposure values across surveys and age groups.
Table 45.
Margin of exposure (MOE) values for the range of acute mean and P95 exposure estimates across different surveys
| MOE | Mean exposure | P95 exposure | ||
|---|---|---|---|---|
| Minimum | Maximum | Minimum | Maximum | |
| Infants | 31 | 8 | 3 | 3 |
| Toddlers | 17 | 6 | 4 | 2 |
| Children | 22 | 7 | 7 | 2 |
| Adolescents | 35 | 9 | 11 | 3 |
| Adults | 43 | 12 | 13 | 4 |
| Elderly | 36 | 14 | 11 | 5 |
| Very elderly | 33 | 14 | 12 | 5 |
The MOEs for the younger age groups indicate a health concern for the food consumption surveys with the highest mean exposure, as well as for the P95 exposure in all surveys. For adult age groups, the MOEs indicate a health concern only for the food consumption surveys with the highest P95 exposures.
To estimate the consequences of these findings, the Panel calculated the mean percentage of days across surveys per age group on which the potato TGA intake may be below the MOE of 10 (Table 46). The highest number of survey days with intake of potatoes below the MOE of 10 was estimated for toddlers (56%) followed by children (50%). For the other age groups, the estimated TGA intake was below the MOE of 10 in up to 22–40% of the survey days.
Table 46.
Summary statistics of the % of survey days with an intake of potato TGAs leading to a margin of exposure (MOE) of below 10 calculated only for the days of potato consumption
| Age class | Range of mean % of days below the MOE of 10a | |
|---|---|---|
| Minimum | Maximum | |
| Infants | 4 (0–14) | 40 (38–42) |
| Toddlers | 18 (17–19) | 56 (45–65) |
| Children | 11 (9–13) | 50 (45–54) |
| Adolescents | 4 (3–5) | 37 (34–40) |
| Adults | 3 (2–4) | 27 (25–28) |
| Elderly | 3 (0–8) | 22 (18–26) |
| Very elderly | 3 (2–5) | 22 (15–29) |
The mean percentage of days with intakes leading to MOE of below 10 per age group for each survey was calculated by averaging the percentage of days below the reference for each subject and then averaged over all subjects within the age group for each survey. The process was iterated 1,000 times as for the exposure estimates, and a mean of the mean percentage of days below the reference per age group was calculated together with a 95% confidence interval.
Note: The corresponding 95% confidence intervals (95% CI) are given in the brackets.
The Panel also calculated the mean percentage of days across surveys per age group on which the potato TGA intake may be above the LOAEL of 1 mg/kg bw. Exposure was estimated to be above the LOAEL of 1 mg/kg bw for toddlers and children on maximally 1% and 0.6% of the survey days, respectively, and on less than 0.4% of the survey days for the other age groups (see Appendix H).
3.4.1.2. GAs from edible parts of food plants other than S. tuberosum
No occurrence data on tomato or aubergine‐derived GAs or aglycones were submitted to EFSA. The data from the literature indicate that the concentration of tomato GAs is lower than of potato GAs in the various Solanum species (see Section 3.2.2.1.2). There are few data on the occurrence of GAs in aubergines (see Section 3.2.2.1.3). Overall, the CONTAM Panel concluded that the data from the literature are too limited to attempt an exposure assessment for tomato or aubergine GAs.
In addition, the lack of any human study and insufficient experimental toxicity data for tomato and aubergine GAs (no long‐term studies, no dose‐response in repeated dose studies) impairs the identification of a reference point.
To conclude, there are insufficient data to perform a risk characterisation for tomato and aubergine GAs.
3.4.2. Farm animals, horses and companion animal risk characterisation
Since no studies on the adverse effects of GAs in farm animals, horses or companion animals were identified, which could be used for the identification of a reference points, no risk characterisation could be performed.
3.5. Uncertainty analysis
The evaluation of the inherent uncertainties in the assessment of exposure to GAs has been performed following the guidance of the Opinion of the Scientific Committee related to Uncertainties in the Dietary Exposure Assessment (EFSA, 2007). In addition, the report on ‘Characterizing and Communicating Uncertainty in Exposure Assessment’ has been considered (WHO/IPCS, 2008). The CONTAM Panel took note of the new guidance on uncertainties of the Scientific Committee (EFSA Scientific Committee, 2018), but it was not implemented in this opinion.
The CONTAM Panel assessed the risk for human health following acute exposure to potato GAs. The data available for repeated dose toxicity and on reproduction in experimental animals did not allow to draw conclusions or to establish a reference point for a chronic exposure risk assessment. In this respect, the Panel noted that: (i) the repeated dose studies on the aglycone solanidine in rodents showed adverse effects being different from those induced by the respective potato GAs, (ii) the developmental effects of potato GAs and aglycones have not been studied over a dose‐range, for the entire gestational period until weaning, and in species other than hamster, (iii) from the limited data available in humans relatively long serum half‐lives were reported for potato GAs suggesting a potential for bioaccumulation, (iv) no conclusive toxicity data on repeated exposure of humans with GAs and aglycones are available.
Due to limited information no risk assessment was possible for effects of potato GAs and their aglycones for farm animals, horses and companion animals.
Similarly, a risk assessment for the effects of tomato and aubergine GAs and their aglycones for humans could not be performed.
Therefore, the evaluation of uncertainties in this section focuses only on the assessment of the risk following acute exposure to potato GAs and aglycones in humans.
3.5.1. Assessment objectives
The objectives of the assessment are clarified in Section 1.2 on Interpretation of the Terms of Reference.
3.5.2. Exposure scenario/exposure model
Occurrence data in food
In most studies, only α‐solanine and α‐chaconine have been looked for and reported, while other, minor GAs as well as β‐ and γ‐forms of solanine and chaconine, have not been investigated. However, these would represent around 5% of the total potato GAs content, and thus represent a minor uncertainty.
In response to EFSA's request to submit occurrence data on GAs, only three countries provided data, thus country‐based differences in the levels of GA in main‐crop potatoes is not evenly represented in the available data, introducing an uncertainty on the representativeness of the overall statistics. Possible differences in GA levels in potatoes, due to the use of different cultivars, location, different growing conditions and year to year variability, across European countries may result in over‐ or underestimation of exposure for certain food consumption surveys.
For 95% of the samples, the information concerning specifically the peeling of the analysed potatoes was not reported. However, most of the samples were reported as unprocessed, implying that they were not peeled. For 131 samples, no information was available. For these samples, the CONTAM Panel considered it unlikely they were peeled, and thus, they were assumed to have been analysed unpeeled. In case (part of) these 131 samples were peeled, this might have caused an underestimation of the GA occurrence.
The analytical results used for the exposure assessment were obtained using different analytical methods, different detection techniques and with varying LODs/LOQs. The average ratio between α‐solanine and α‐chaconine was found to differ between data submissions, indicating differences between the analytical methodologies. This could result in an over‐ or underestimation of the exposure.
There is some evidence that partial degradation of α‐chaconine (to β‐ and γ‐forms) may occur during sample preparation and analysis, unless specific conditions are applied. For the data sets submitted to EFSA, it could not be determined to which extent the results for individual samples could have been affected by enzymatic hydrolysis. This could result in an underestimation of the exposure.
Food processing/processing factors
Important sources of uncertainty in the exposure assessment are related to the different assumptions made, such as that the occurrence values obtained from sampled potatoes are representative of consumed potatoes, and to the assignment of processing factors associated with peeling and heat processing. Literature studies, however, report a wide range of processing factors, introducing uncertainty on the actual reduction during the various processing steps. To take into account the reduction by peeling and heat processing, several assumptions were made (see Section 2.5). These assumptions contribute to the uncertainty of the exposure assessment and they could have led to an over‐ or underestimation of the exposure.
It is not known to which extent these food processing steps result in only a partial degradation of GAs (to β‐ and γ‐forms of solanine and chaconine) or degradation to the aglycone (solanidine). This may result in a slight underestimation of the TGA content present in food products as consumed.
Food Consumption data
Uncertainties and limitations related to the use of the EFSA Comprehensive Food Consumption Database were described by EFSA (EFSA, 2011a) and are not further detailed in this Scientific Opinion. These relate to the use of different dietary survey methodologies, standard portion sizes, representativeness of samples included in surveys or to the inclusion of consumption surveys covering only few days to estimate high percentiles of chronic exposure.
Uncertainties and limitations related to the use of the RPC model related to the process of converting the consumed amount of composite foods to the amount of raw primary commodity were also described by EFSA (EFSA, 2019). The main uncertainty is linked to the conversion of the amount consumed for a specific FoodEx code into the amount consumed of each raw primary commodity from which the food originated or was assumed to originate.
3.5.3. Hazard identification and characterisation
Toxicokinetics
The available studies did not allow for a full characterisation of the absorption, distribution, metabolism and excretion profile for α‐solanine and α‐chaconine. Differences in absorption and excretion between rats and hamsters were observed for these two GAs, with indications of higher absorption and slower excretion rates in the latter species. There is uncertainty on the most representative species for the human risk assessment.
Interactions
The possibility that components different from GAs in potatoes, such as spirostane saponins, may enhance the toxicity of GA has also to be considered. As discussed by McMillan and Thompson (1979) in their report on an outbreak of poisoning in schoolboys associated with the consumption of toxic potatoes, saponins might promote gastrointestinal absorption of GAs or contribute to irritation of the gastrointestinal tract. This might contribute to an overestimation of the risk from the potato GAs alone.
Observations in Humans
A NOAEL for local effects in the gastrointestinal tract of potato GAs could not be identified.
Data on adverse effects of potato GAs were only available for adults and school children, but are lacking for infants, toddlers and young children.
Available data suggest interindividual differences in the susceptibility to the adverse effects of potato GAs in the gastrointestinal tract.
Intake figures of potato GAs for case reports were in general estimates which causes uncertainties on the shape of the dose–response relationship and which may lead to over or under estimation.
Non‐specific symptoms like nausea and gastrointestinal discomfort may not always have been associated with intake of potato GAs. This might lead to under‐reporting and contribute to the uncertainty in the NOAEL/LOAEL.
3.5.4. Summary of uncertainties
In Table 47, a summary of the uncertainty evaluation is presented, highlighting the main sources of uncertainty and indicating an estimate of whether the respective source of uncertainty might have led to an over‐ or underestimation of the exposure or the resulting risk.
Table 47.
Summary of qualitative evaluation of the impact of uncertainties on the risk assessment of acute exposure to potato GAs in food
| Sources of uncertainty | Directiona |
|---|---|
| Occurrence | |
| Insufficient information on GA levels in potatoes of specific cultivars grown at different locations and conditions; variability between countries and years; data from only three countries; no data for potato cultivars used in industrial processing | +/– |
| In case of no information, it was assumed that potatoes were analysed as unpeeled | – |
| Estimated reduction of potato GAs concentration to account for loss during food processing (peeling and heat treatment) | +/– |
| Underreporting of GA levels in potatoes due to enzymatic degradation during sample preparation and analysis | – |
| Consumption data | |
| Different methodologies/representativeness/underreporting/misreporting/no portion size standard | +/– |
| Conversion of food consumed to raw primary commodity | +/– |
| Hazard identification and characterisation | |
| Uncertainty in the exposure levels of GAs estimated from case reports | +/– |
| Uncertainties on the shape of the dose–response relationship due to limited data available in humans | +/– |
| Putative interactions on the local irritating effect of potato GA with saponins | + |
| Possible interindividual differences in humans with regard to gastrointestinal symptoms | +/– |
| Non‐specific symptoms of intoxications in humans not associated with potato GAs intake, resulting in under‐reporting and uncertainties for derivation of LOAEL/NOAEL | +/– |
+ = uncertainty with potential to cause overestimation of exposure/risk; – = uncertainty with potential to cause underestimation of exposure/risk.
The CONTAM Panel considered that the impact of the uncertainties on the risk assessment of acute exposure to potato GAs in food is moderate. Overall, the identified uncertainties may either cause an over‐ or underestimation of the risk.
4. Conclusions
4.1. Hazard identification and characterisation
4.1.1. Toxicokinetics
The potato glycoalkaloids (GAs), α‐solanine and α‐chaconine, show a relatively low oral bioavailability in experimental animals, with differences between species. Hamsters exhibit higher absorption and slower excretion rates for both substances when compared to rats. Due to the limited information, the metabolic profiles of potato GAs in experimental animals could not be characterised.
In humans, α‐solanine and α‐chaconine are systemically absorbed following ingestion with relatively long serum half‐lives reported suggesting a possible accumulation.
Levels of solanidine were regularly detected in the blood of human volunteers in several studies suggesting hydrolysis of GAs. The blood clearance of solanidine appears to be slow.
No further information is available on metabolism and excretion of potato GAs in humans.
Toxicokinetic data on tomato and aubergine GAs and their aglycones could not be identified, neither for experimental animals nor for humans.
Information on the toxicokinetics of GAs in farm and companion animals was limited to ruminants, for which the data suggest an extensive conversion of α‐solanine and α‐chaconine to aglycones in rumen, and a low potential of solanidine to transfer into cows’ milk.
4.1.2. Toxicity in experimental animals
In acute, subacute and subchronic studies similar/identical doses of α‐solanine and α‐chaconine elicited comparable symptoms in several experimental animal species. This indicates a similar potency of α‐solanine and α‐chaconine with regard to toxicity.
In repeated oral dose studies in rodents, reduced body and liver weight were the most common effects for potato GAs, while the aglycone solanidine increased absolute and relative liver weight.
The tomato GA, α‐tomatine, and the aglycone, tomatidine, showed no adverse effects in rodents.
In mice, the aubergine GA, α‐solasonine, increased the body weight gain. However, its aglycone, solasodine, lowered the body weight and caused gastric gland degeneration and liver toxicity, indicating different effects of the aglycone when compared to the GA.
Malformations in the central nervous system occurred in hamsters at relatively high doses of potato GAs and the aglycone, solanidine, when applied for only one day or for a short, very restricted time period during gestation. No no‐observed‐adverse‐effect level (NOAEL) or lowest‐observed‐adverse‐effect level (LOAEL) could be identified from these studies.
Reduced postnatal survival of pups due to insufficient milk production was reported when pregnant Holtzman rats had been exposed to α‐solanine.
Exposure of male dogs to the aubergine aglycone solasodine decreased epididymal weight and cauda epididymal epithelial height. Also an epididymal lumen depleted of sperm was reported. Similar effects of solasodine were observed in Rhesus monkeys.
From the limited number of studies available, there was no evidence for genotoxicity of the potato GAs, α‐solanine and α‐chaconine, and the aglycone, solanidine, as well as the aubergine GA α‐solamargine. However, there is not sufficient information to conclude on the genotoxic potential of these GAs.
No long‐term chronic toxicity/carcinogenicity study for potato, tomato or aubergine GAs or for the respective aglycones could be identified.
4.1.3. Observations in humans
Acute toxic effects following ingestion of potato GAs include gastrointestinal symptoms of varying severity such as vomiting, diarrhoea and abdominal pain, which may occur from a total potato GAs (potato TGA) intake of 1 mg/kg bw or more.
In severe cases, paralysis, respiratory insufficiency, cardiac failure, coma and death have been reported. Doses in the range of 3–6 mg potato TGAs/kg bw are considered to be potentially lethal for humans.
4.1.4. Adverse effects in farm animals, horses and companion animals
No data on the potential adverse effects of potato GAs in horses, companion animals (cats and dogs) or fur animals were identified.
Due to an insufficient database on the adverse effects of GAs in ruminants, pigs, poultry, rabbits and fish, an acute reference dose could not be derived.
4.1.5. Mode of action
Adverse effects of GAs may be due to their ability to complex with membrane 3β‐hydroxy sterols, thereby causing disruption and loss of integrity of cell membranes. After oral exposure, these effects may affect the mucosa of the gastrointestinal tract and cause the symptoms observed in intoxicated humans, such as nausea, vomiting and diarrhoea.
GAs inhibit acetylcholinesterase (AChE) and serum butyrylcholinesterase (BuChE) by a reversible, competitive mode of action. The relative potency of inhibition of α‐solanine and α‐chaconine appear to be similar. The aglycones exert weak or no inhibitory effects. The excess of acetylcholine at the neuronal and neuromuscular junctions upon inhibition of the enzymes might also contribute to the symptoms described for intoxications with GAs.
4.1.6. Margin of exposure (MOE) approach
The CONTAM Panel considered that the use of rodent data on acute toxicity was not appropriate to establish a reference point for acute exposure to potato GAs in humans. The CONTAM Panel selected the LOAEL of 1 mg potato TGA/kg bw per day as the reference point for assessing the risk following acute exposure, based on human data from case reports, outbreaks and studies in volunteers.
The available data on acute toxicity were considered insufficient to establish a health‐based guidance value (HBGV). Instead, the MOE approach was used to assess a possible health concern from acute exposure to potato TGAs via food.
The main symptoms following acute exposure to potato GAs may be mainly due to local irritation of the gastrointestinal mucosa rather than inhibition of AChE activity. Therefore, the CONTAM Panel considered that the possible interindividual variability in toxicodynamics is more relevant than the interindividual variability in toxicokinetics.
An MOE higher than 10 indicates that there is no health concern. This MOE of 10 takes into account the extrapolation from a LOAEL to a NOAEL (a factor of 3) and the interindividual variability in toxicodynamics (a factor of 3.2).
The experimental data for repeated dose toxicity are not sufficient to identify a reference point for chronic exposure to potato GAs. In humans, no evidence of health problems associated with repeated or long‐term intake of GAs via potatoes has been identified.
No data for determining a reference point for tomato or aubergine GAs or aglycones are available.
4.2. Occurrence and exposure
4.2.1. Food
Occurrence data were only available for α‐solanine and α‐chaconine, and mostly for ‘Main‐crop potatoes’ and ‘New potatoes’. Few data were available for processed food categories. No occurrence data were available for GAs and their aglycones in tomato and aubergine.
The mean UB occurrence for ‘Main‐crop potatoes’ and ‘New potatoes’ was 51.2 mg/kg and the P95 occurrence was 116.8 mg/kg. The minimum and maximum reported concentrations were 1.1 mg/kg and 276.6 mg/kg, respectively.
The CONTAM Panel decided to use the occurrence data in the raw primary commodities due to lack of available data for processed foods.
The CONTAM Panel assessed the acute dietary exposure to potato TGAs (sum of α‐solanine and α‐chaconine) using a probabilistic approach.
Processing factors were estimated for the major food processing steps, comprising peeling (factor between 0.25 and 0.75) and heat processing of potatoes (factors between 0.1 and 0.8 for frying and deep frying, and between 0.35 and 0.95 for all other processing methods), and these were applied to the occurrence data. It was assumed that 90% of the potatoes are consumed as peeled.
The mean upper bound exposure to potato TGAs across surveys ranged from 23.3 μg/kg bw per day in adults to 174.0 μg/kg bw per day in toddlers. The 95th percentile exposure ranged from 78.3 μg/kg bw per day in adults to 535.1 μg/kg bw per day in toddlers (up to 822.9 μg/kg bw per day in the upper limit of the 95% confidence interval).
4.2.2. Feed
Potatoes and by‐products thereof can be used as feed but data on potato GAs in feed were insufficient to perform an exposure assessment.
4.3. Risk characterisation
4.3.1. Human health risk characterisation
For potato GAs, the MOEs for the younger age groups indicate a health concern for the food consumption surveys with the highest mean exposure, as well as for the P95 exposure in all surveys.
For adult age groups, the MOEs indicate a health concern only for the food consumption surveys with the highest P95 exposures.
The Panel calculated the mean percentage of days with potato consumption across surveys per age group on which the potato TGA intake may be below the MOE of 10. The highest number of survey days was estimated for toddlers (56%) followed by children (50%). For the other age groups, the estimated TGA intake was below the MOE of 10 in up to 22–40% of the survey days.
For tomato and aubergine GAs, the risk to human health could not be characterised due to the lack of occurrence data in food and the limited information on the adverse effects in experimental animals and humans.
4.3.2. Farm animals, horses and companion animal health risk characterisation
No risk characterisation could be performed due to insufficient occurrence data of GAs for feed and the lack of, or limited, data on the potential adverse effects of GAs in farm animals, horses and companion animals.
5. Recommendations
The following needs have been identified to improve the risk assessment for humans and reduce the uncertainties:
Research on the occurrence of GAs and their aglycones and other potentially toxicologically relevant secondary plant metabolites in the potato cultivars available on the market and on new potato cultivars resulting from breeding experiments.
Occurrence data on GAs and their aglycones in potato processed products, including foods for infants.
Occurrence data on GAs and their aglycones in tomato and aubergine and products thereof.
Data on the toxicokinetics of potato, tomato and aubergine GAs and aglycones in experimental animals and humans.
Data on repeated dose toxicity, including reproductive and developmental toxicity of potato, tomato and aubergine GAs and aglycones in experimental animals.
Studies in humans linking dietary exposure, biomarkers of exposure and adverse effects.
The following needs have been identified to improve the risk assessment for farm animals, horses and companion animals and reduce the uncertainties:
Occurrence data on potato GAs and their aglycones in feed.
Studies on the kinetics and the potential adverse effects from feed material containing GAs of potato GAs in farm animals, horses and companion animals.
Documentation provided to EFSA
Sicherheitsbewertung gentechnisch veränderter Lebensmittel: Glykoalkaloide in transgenen Kartoffeln. PhD thesis by Gessner Gerhard. Technisched Universität Műnchen. 2000. Submitted to EFSA by the German Federation for Food Law and Food Science (BLL) following the Consultation with stakeholders (see Section 2.2.1).
Occurrence data of GAs in ‘fresh potato’, ‘fresh potato with skin for industrial use’ and ‘potato crisps’. Data submitted by the European Snacks Association (ESA) following the Consultation with stakeholders (see Section 2.2.1). The occurrence data was not further submitted following the requirements of the EFSA Guidance on Standard Sample Description for Food and Feed (EFSA, 2010a), and they could not be used for the current dietary exposure assessment.
Comments on GAs in potato and potato products and occurrence data in potatoes and potato products such as ‘croquettes’, ‘French fries’, ‘potato flakes’, ‘potato pancakes’ and ‘potato wedges’. Data submitted by the European Potato Processors’ Association (EUPPA) following the Consultation with stakeholders (see Section 2.2.1). The occurrence data was not further submitted following the requirements of the EFSA Guidance on Standard Sample Description for Food and Feed (EFSA, 2010a), and they could not be used for the current dietary exposure assessment.
Comments on GAs in potato and potato products. Data submitted by the European Potato Trade Associations (Europatat) following the Consultation with stakeholders (see Section 2.2.1).
Occurrence data of GAs in potato and potato products such as ‘potato dietary fibre’, ‘potato proteins’, ‘potato pulp’, ‘potato juice’, ‘potato starch’ and ‘potatoes used for starch production’. Data submitted by the European Starch Industry Association following the Consultation with stakeholders (see Sections 2.2.1 and 3.2.1).
Abbreviations
- ACh
Acetylcholine
- AChE
Acetylcholinesterase
- ALT
Alanine aminotransferase
- Anses
French Agency for Food, Environmental and Occupational Health and Safety
- AOAC
Association of Official Analytical Chemists
- AST
Aspartate aminotransferase
- AUC0‐24
Areas Under the Curve (0–24 h)
- AUC0‐∞
Areas Under the Curve (from point zero to infinity)
- BfR
The German Federal Institute for Risk Assessment
- BuChE
Butyrylcholinesterase
- bw
Body weight
- ChEs
Cholinesterases
- CI
Confidence interval
- Cmax
Maximum serum concentrations
- CNS
Central nervous system
- CONTAM Panel
Panel on Contaminants in the Food Chain
- dw
Dry weight
- ECG
Electrocardiogram
- EEG
electroencephalogram
- ELISA
Enzyme‐linked immunosorbent assay
- F
female
- FAO
Food and Agricultural Organization
- FDA
Food and Drug Administration
- FID
Flame ionisation detector
- fw
Fresh weight
- GAs
Glycoalkaloids
- GC
Gas Chromatography
- GD
Gestational day
- GOT
Glutamic oxaloacetic transaminase
- GPT
Glutamic pyruvic transaminase
- h
hour
- HDL
High‐density lipoprotein
- HGB
Haemoglobin
- HMGR
3‐hydroxy‐3-methylglutaryl coenzyme A reductase
- HPLC
High‐performance liquid chromatography
- HPTLC
High performance thin layer chromatography
- HRMS
High resolution mass spectrometry
- HTC
Haematocrit
- IPCS
International Programme on Chemical Safety
- i.p.
Intraperitoneal injection
- i.v.
Intravenous injection
- IFN‐γ
Interferon‐gamma
- IL‐1β
Interleukin 1 beta
- JECFA
Joint FAO/WHO Expert Committee on Food Additives
- Ki
Inhibitory constants
- LB
Lower bound
- LC
Liquid chromatography
- LD50
Lethal dose, median
- LDL
Low‐density lipoproteins
- LOEL
Lowest‐observed‐effect level
- LOAEL
Lowest‐observed‐adverse‐effect level
- LOD
Limit of detection
- LOQ
Limit of quantification
- LPS
Lipopolysaccharide
- M
male
- MALDI
Matrix‐assisted laser desorption/ionisation
- MOE
Margin of exposure
- MS
Mass spectrometry
- MS/MS
Tandem mass spectrometry
- MTHFR
Methylenetetrahydrofolate reductase
- nAChR
Specific acetylcholine‐receptor
- NaHSO3
Sodium bisulfite
- NAK
Nederlandse Algemene Keuringsdienst (Dutch General Inspection Service)
- NOEL
No‐observed‐effect level
- NOAEL
No‐observed‐adverse‐effect level
- NPD
Nitrogen Phosphorous Detector
- NTD
Neural tube defects
- OECD
The Organisation for Economic Co‐operation and Development
- OFCs
Orofacial clefts
- PAD
Pulsed amperometric detector
- PCE
Polychromatic erythrocytes
- p.o.
per oral
- RASFF
Rapid Alert System for Food and Feed
- RBC
Red blood cell
- RPC
Raw primary commodity
- s.c.
subcutaneous
- SGT
Solanidine glycosyltransferase
- SMT
Sterol C24‐methyltransferase
- SOPs
Standard operational procedures
- SPE
Solid‐phase extraction
- SS
Squalene synthase
- TGA
Total glycoalkaloids
- TLC
Thin layer chromatography
- TNF‐α
Tumour necrosis factor‐alpha
- TOF
Time‐of-flight
- UB
Upper bound
- UK
United Kingdom
- USA
United States of America
- UV
Ultraviolet
- UV‐vis
Ultraviolet‐visible
- VLDL
Very low‐density lipoproteins
- Voedingscentrum
The Netherlands Nutrition Centre Foundation
- VSP
Vetispiradiene cyclase
- WBC
White blood cells
- WHO
World Health Organization
Appendix A – Major glycoalkaloids and their aglycones present in Solanum species
1.
-
A
The main aglycones found in Solanum species considered in this Scientific Opinion
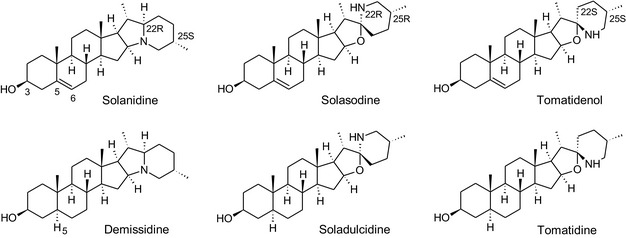
-
B
The main sugar moieties found in Solanum species considered in this Scientific Opinion
-
C
Glycoalkaloids considered in this Scientific Opinion: structures and physico‐chemical properties
|
Solanidine Synonym: solatubine CAS No: 80‐78‐4 Sugar moiety: none, aglycone C27H43NO MW: 397.647 LogP (est): 6.1 Solubility (est): 0.0004 g/L |
|
|
Solanidine N‐oxide Sugar moiety: none, aglycone C27H43NO2 MW: 413.636 LogP (est) : 5.5 |
|
|
α‐Solanine Synonym: solatunine CAS No: 20562‐02‐1 Sugar moiety: solatriose C45H73NO15 MW: 868.071 LogP (est): 1.8 Solubility (est): 0.62 g/L |
|
|
β 1 ‐Solanine CAS No: 142287‐76‐1 Sugar moiety: galactose‐rhamnose C39H63NO10 MW: 705.930 LogP (est): 3.4 |
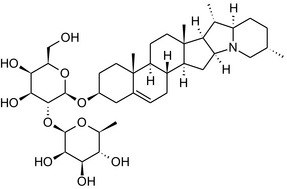
|
|
β 2 ‐Solanine CAS No: 61877‐94‐9 Sugar moiety: galactose‐glucose C39H63NO11 MW: 721.929 LogP (est): 2.9 Solubility (est): 0.25 g/L |
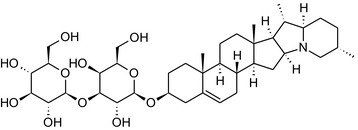
|
|
γ‐Solanine CAS No: 511‐37‐5 Sugar moiety: galactose C33H53NO6 MW: 559.788 LogP (est): 4.5 Solubility (est): 0.021 g/L |
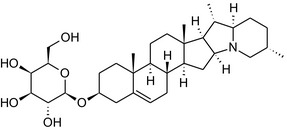
|
|
α‐Chaconine CAS No: 20562‐03‐2 Sugar moiety: chacotriose C45H73NO14 MW: 852.072 LogP (est): 1.8 Solubility (est): 0.29 g/L |
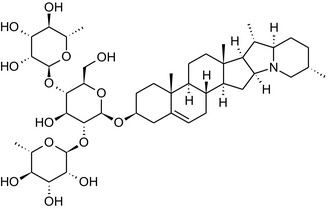
|
|
β 1 ‐Chaconine CAS No: 472‐51‐5 Sugar moiety: rhamnose‐glucose C39H63NO10 MW: 705.930 LogP (est): 3.4 Solubility (est): 0.092 g/L |
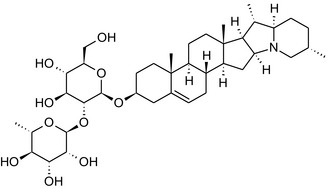
|
|
β 2 ‐Chaconine CAS No: 469‐14‐7 Sugar moiety: rhamnose‐glucose C39H63NO10 MW: 705.930 LogP (est): 2.8 Solubility (est): 0.088 g/L |
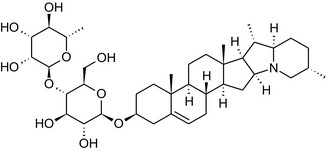
|
|
γ‐Chaconine CAS No: 511‐36‐4 Sugar moiety: glucose C33H53NO6 MW: 559.788 LogP (est): 4.5 Solubility (est): 0.021 g/L |
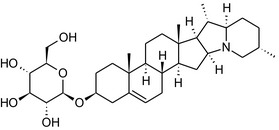
|
|
Rubijervine Synonym: 12α‐hydroxysolanidine CAS No: 79‐58‐3 Sugar moiety: none, aglycone C27H43NO2 MW: 413.646 LogP (est): 4.7 |
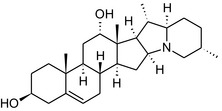
|
|
Leptinidine Synonym: 23β‐hydroxysolanidine CAS No: 24884‐17‐1 Sugar moiety: aglycone C27H43NO2 MW: 413.646 LogP (est): 5.1 |
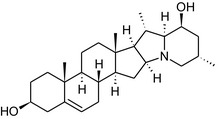
|
|
Leptinine I CAS No: 101009‐59‐0 Sugar moiety: chacotriose C45H73NO15 MW: 868.071 LogP (est): 0.8 |
|
|
Leptinine II CAS No: 100994‐57‐8 Sugar moiety: solatriose C45H73NO16 MW: 884.070 LogP (est): 0.8 |
|
|
Leptine I CAS No: 101030‐83‐5 Sugar moiety: chacotriose C47H75NO16 MW: 910.108 LogP (est): 1.4 |
|
|
Leptine II CAS No: 101054‐39‐1 Sugar moiety: solatriose C47H75NO17 MW: 926.107 LogP (est): 1.4 |
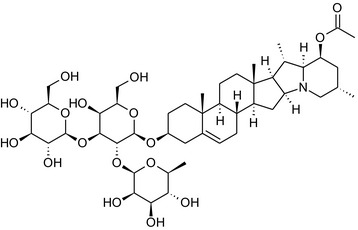
|
|
Dehydrodemissine CAS No: 195433‐57‐9 Sugar moiety: lycotetraose C50H81NO20 MW: 1,016.185 LogP (est): ‐0.8 Solubility (est): 1.69 g/L |
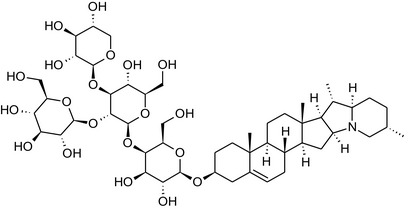
|
|
Dehydrocommersonine CAS No: 65428‐74‐2 Sugar moiety: commertetraose C51H83NO21 MW: 1,046.211 LogP (est): ‐0.8 Solubility (est): 1.69 g/L |
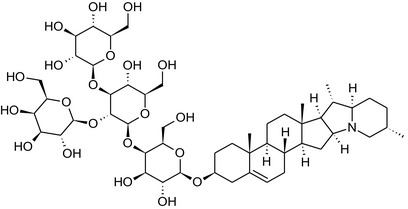
|
|
Demissidine Synonym: 5α,6‐dihydrosolanidine, 5α‐Solanidan‐3β‐ol CAS No: 474‐08‐8 Sugar moiety: none, aglycone C27H45NO MW: 399.663 LogP (est): 6.9 Solubility (est): 0.00022 g/L |
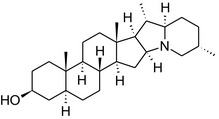
|
|
Demissine CAS No: 6077‐69‐6 Sugar moiety: lycotetraose C50H83NO20 MW: 1,018.201 LogP (est): 0.0 Solubility (est): 1.63 g/L |
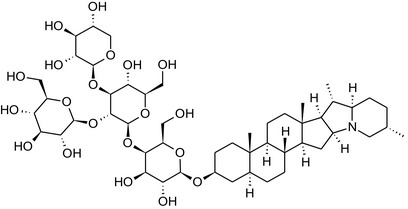
|
|
Commersonine CAS No: 60776‐42‐3 Sugar moiety: commertetraose C51H85NO21 MW: 1,048.227 LogP (est): 0.0 Solubility (est): 1.63 g/L |
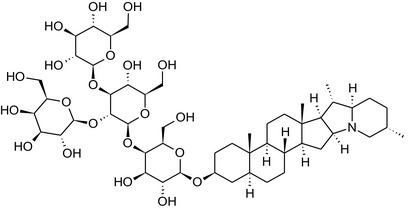
|
|
5β‐Solanidan‐3β‐ol Sugar moiety: none, aglycone C27H45NO MW: 399.663 |
|
|
Solasodine Synonym: purapuridine CAS No: 126‐17‐0 Sugar moiety none: aglycone C27H43NO2 MW: 413.646 LogP (est): 5.4 Solubility (est): 0.00084 g/L |
|
|
α‐Solasonine Synonym: purapurine, γ‐solanigrine CAS No: 19121‐58‐5 Sugar moiety: solatriose C45H73NO16 MW: 884.070 LogP (est): 1.1 Solubility (est): 0.41 g/L |
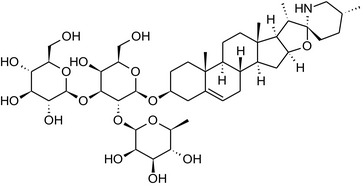
|
|
α‐Solamargine Synonym: δ‐solanigrine CAS No: 20311‐51‐7 Sugar moiety: chacotriose C45H73NO15 MW: 868.071 LogP (est): 1.1 Solubility (est): 0.2 g/L |
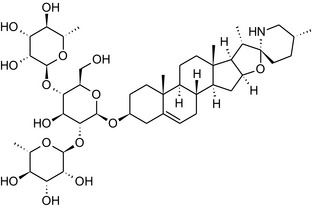
|
|
Soladulcidine Synonym: solasodan‐3β‐ol, dihydrosolasodine CAS No: 511‐98‐8 Sugar moiety: none aglycone C27H45NO2 MW: 415.662 LogP (est): 6.2 Solubility (est): 0.00019 g/L |
|
|
Soladulcine A CAS No: 156555‐52‐1 Sugar moiety: chacotriose C45H75NO15 MW: 870.087 LogP (est): |
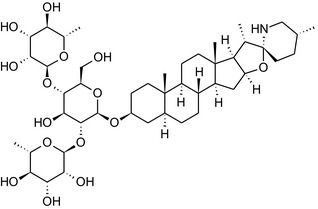
|
|
Tomatidenol Synonym: dehydrotomatidine CAS No: 546‐40‐7 Sugar moiety: none, aglycone C27H43NO2 MW: 413.646 LogP (est): 5.4 Solubility (est): 0.00084 g/L |
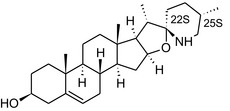
|
|
α‐Solamarine CAS No: 20318‐30‐3 Sugar moiety: solatriose C45H73NO16 MW: 884.07 LogP (est): 1.1 Solubility (est): 0.38 g/L |
|
|
β‐Solamarine CAS No: 3671‐38‐3 Sugar moiety: chacotriose C45H73NO15 MW: 868.071 LogP (est): 1.1 Solubility (est): 0.16 g/L |
|
|
α‐Dehydrotomatine CAS No: 157604‐98‐3 Sugar moiety: lycotetraose C50H81NO21 MW: 1,032.184 LogP (est): −1.5 Solubility (est): 1.82 g/L |
|
|
Tomatidine CAS No: 77‐59‐8 Sugar moiety: none, aglycone C27H45NO2 MW: 415.662 LogP (est): 6.2 Solubility (est): 0.00019 g/L |
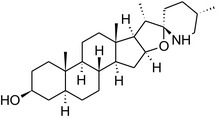
|
|
α‐Tomatine CAS No: 17406‐45‐0 Sugar moiety: lycotetraose C50H83NO20 MW: 1,034.188 LogP (est): −0.7 Solubility (est): 1.71 g/L |
|
|
Lycoperoside A Syn: 23R‐Acetoxytomatine CAS No: 176181‐33‐2 Sugar moiety: lycotetraose C52H85NO23 MW: 1,092.224 LogP (est): −1.1 |
|
|
Lycoperoside B Syn: 23S‐Acetoxysoladulcidine CAS No: 176300‐86‐0 Sugar moiety: lycotetraose C52H85NO23 MW: 1,092.224 LogP (est): −1.1 |
|
|
Lycoperoside C (23S,25S)‐23‐Acetoxy‐5α,22αN‐Spirosolan‐3β‐ol CAS No: 176300‐87‐1 Sugar moiety: lycotetraose C52H85NO23 MW: 1,092.224 LogP (est): −1.1 |
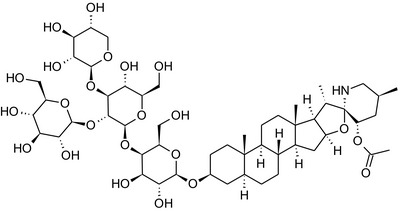
|
|
Lycoperoside F CAS No: 675828‐26‐9 Sugar moiety: Lycotetraose and glucose C58H95NO29 MW: 1,270.606 LogP (est): −3.6 |
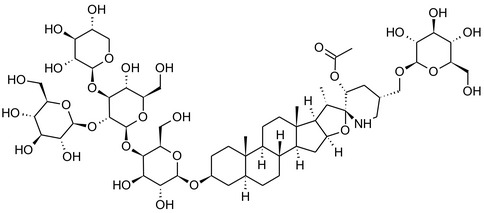
|
|
Lycoperoside G CAS No: 675828‐27‐0 Sugar moiety: Lycotetraose and glucose C58H95NO29 MW: 1,270.606 LogP (est): −3.6 |
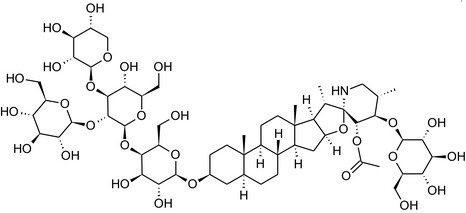
|
|
Lycoperoside H CAS No: 675828‐28‐1 Sugar moiety: lycotetraose C50H83NO22 MW: 1,050.188 LogP (est): 1.2 |
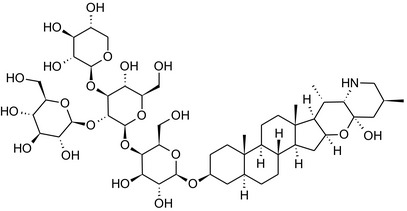
|
|
Esculeoside A CAS No: 532387‐86‐3 Sugar moiety: lycotetraose and glucose C58H95NO29 MW: 1,270.376 LogP (est): −3.9 |
|
|
Tomatoside A CAS No: 75557‐22‐1 Sugar moiety: triose and glucose C51H86O24 MW: 1,084.210 LogP (est): −1.4 |
|
Appendix B – Identification and selection of evidence relevant for the risk assessment of glycoalkaloids in feed and food
1.
In addition to the literature search outsourced by EFSA (see University of Chemistry and Technology Prague, 2019 for further details), some specific searches were performed to identify the scientific literature.
B.1. Literature search for hazard identification and characterisation
Adverse effects of GAs in humans
A literature search was performed to retrieve data on the adverse effects of GAs in humans. Three databases were used: Web of Science, Pubmed and Scopus. The search was performed without limits regarding the year/type of publication, and language. All the references were uploaded in Endnote. After removal of duplicates, the final number of hits is 330.
Solanine
Web of science
TOPIC: (solanin* OR 20562‐02‐1 OR 243‐879‐8) AND TOPIC: (adverse effect* OR epidemiolog* OR “case control” OR case report* OR case stud* OR clinical trial* OR clinical stud* OR cohort OR “cross sectional” OR human stud* OR human volunteer*)
Results: 276
Pubmed
(((solanin[All Fields] OR solanina[All Fields] OR solaninase[All Fields] OR solaninaza[All Fields] OR solaninbestimmung[All Fields] OR solanine[All Fields] OR solaninlosung[All Fields] OR solaninneubildung[All Fields] OR solaninom[All Fields] OR solaninum[All Fields] OR solaninvergiftung[All Fields]) OR “20562‐02‐1”[All Fields])) AND (“Epidemiologic Studies”[Mesh] OR “Drug‐Related Side Effects and Adverse Reactions”[Mesh] OR “Volunteers”[Mesh] OR “Human Experimentation”[Mesh] OR “Clinical Study” [Publication Type] OR “Clinical Studies as Topic”[Mesh] OR “Case Reports” [Publication Type] OR epidemiolog*[tiab] OR “case control”[tiab] OR case stud*[tiab] OR cohort[tiab] OR “cross sectional”[tiab] OR adverse effect*[tiab] OR “human volunteers”[tiab] or human stud*[tiab])
Results: 11
Scopus
(TITLE‐ABS‐KEY ((solanin* OR “20562‐02‐1” OR “243‐879‐8”)) AND TITLE‐ABS‐KEY ((human AND adverse AND effect*) OR (epidemiolog*) OR (case AND control) OR (case AND report*) OR (case AND stud*) OR (clinical AND trial*) OR (clinical AND stud*) OR (cohort) OR (cross AND sectional) OR (human AND stud*) OR (human AND volunteer*)))
Results: 144
All the retrieved references were uploaded in Endnote. Final number of hits after removal of duplicates: 356
Chaconine
Web of science
TOPIC: ((chaconin* OR 20562‐03‐2 OR 472‐51‐5)) AND TOPIC: (adverse effect* OR epidemiolog* OR case control OR case report* OR case stud* OR clinical trial* OR clinical stud* OR cohort OR cross sectional OR human stud* OR human volunteer*)
Results: 172
Pubmed
(((chaconin[All Fields] OR chaconine[All Fields] OR chaconinverhaltnisses[All Fields]) OR “20562‐03‐2”[All Fields] OR “472‐51‐5”[All Fields])) AND (((“Epidemiologic Studies”[Mesh] OR “Drug‐Related Side Effects and Adverse Reactions”[Mesh] OR “Volunteers”[Mesh] OR “Human Experimentation”[Mesh] OR “Clinical Study” [Publication Type] OR “Clinical Studies as Topic”[Mesh] OR “Case Reports” [Publication Type] OR epidemiolog*[tiab] OR “case control”[tiab] OR case stud*[tiab] OR cohort[tiab] OR “cross sectional”[tiab] OR adverse effect*[tiab] OR “human volunteers”[tiab] or human stud*[tiab])))
Results: 7
Scopus
(TITLE‐ABS‐KEY (chaconin* OR 20562‐03‐2 OR 472‐51‐5) AND TITLE‐ABS‐KEY ((human AND adverse AND effect*) OR (epidemiolog*) OR (case AND control) OR (case AND report*) OR (case AND stud*) OR (clinical AND trial*) OR (clinical AND stud*) OR (cohort) OR (cross AND sectional) OR (human AND stud*) OR (human AND volunteer*)))
Results: 79
All the retrieved references were uploaded in Endnote. Final number of hits after removal of duplicates: 206
Glycoalkaloids
Web of science
TOPIC: (glycoalkaloid*) AND TOPIC: (adverse effect* OR epidemiolog* OR case control OR case report* OR case stud* OR clinical trial* OR clinical stud* OR cohort OR cross sectional OR human stud* OR human volunteer*)
Results: 497
Pubmed
((glycoalkaloid[All Fields] OR glycoalkaloidal[All Fields] OR glycoalkaloidic[All Fields] OR glycoalkaloids[All Fields] OR glycoalkaloidsalpha[All Fields])) AND (((“Epidemiologic Studies”[Mesh] OR “Drug‐Related Side Effects and Adverse Reactions”[Mesh] OR “Volunteers”[Mesh] OR “Human Experimentation”[Mesh] OR “Clinical Study” [Publication Type] OR “Clinical Studies as Topic”[Mesh] OR “Case Reports” [Publication Type] OR epidemiolog*[tiab] OR “case control”[tiab] OR case stud*[tiab] OR cohort[tiab] OR “cross sectional”[tiab] OR adverse effect*[tiab] OR “human volunteers”[tiab] or human stud*[tiab])))
Results: 18
Scopus
(TITLE‐ABS‐KEY (glycoalkaloid*) AND TITLE‐ABS‐KEY ((human AND adverse AND effect*) OR (epidemiolog*) OR (case AND control) OR (case AND report*) OR (case AND stud*) OR (clinical AND trial*) OR (clinical AND stud*) OR (cohort) OR (cross AND sectional) OR (human AND stud*) OR (human AND volunteer*)))
Results: 167
All the retrieved references were uploaded in Endnote. Final number of hits after removal of duplicates: 541
B.2. Literature search for hazard identification and characterisation
Toxicity of glycoalkaloids
An extra search was conducted including all the relevant GAs from the mandate. The search included the search strings listed in the tables below, three different search databases were used. The GAs were divided into three groups for the search:
Tomato alkaloids: rubijervin, tomatidine, tomatine, tomatidenol, dehydrotomatine.
Solanine alkaloids: solasodine, solasonine, solamargine, soladulcine, soladulcidine, solamarine, solanidine.
Leptin alkaloids: leptinin, leptidine, leptinine I and II, leptine I and II
(a) Tomato alkaloids
Web of science
TOPIC: (rubijervin*) OR TOPIC: (tomatidin*) OR TOPIC: (tomatidenol*) OR TOPIC: (tomatin*) OR TOPIC: (dehydrotomatin)) AND (TOPIC: (toxic*) OR TOPIC: (adverse) OR TOPIC: (terato*))
Results: 78
Pubmed
((((((rubijervin*) OR tomatidin*) OR tomatin*) OR tomatidenol) OR dehydrotomatin*)) AND (((toxic*) OR adverse) OR terato*)
Results: 101
Scopus
((TITLE‐ABS‐KEY (rubijervin*) OR TITLE‐ABS‐KEY (tomatidin*) OR TITLE‐ABS‐KEY (tomatin*) OR TITLE‐ABS‐KEY (tomatidenol*) OR TITLE‐ABS‐KEY (dehydrotomatin*))) AND ((TITLE‐ABS‐KEY (toxic*) OR TITLE‐ABS‐KEY (adverse) OR TITLE‐ABS‐KEY (terato*)))
Results: 109
(b) Solanine alkaloids
Web of science
(TOPIC: (solasodin*) OR TOPIC: (solasonin*) OR TOPIC: (solamargin*) OR TOPIC: (soladulcin*) OR TOPIC: (solamarin*) OR TOPIC: (solanidin*)) AND (TOPIC: (toxic*) OR TOPIC: (adverse) OR TOPIC: (terato*))
Results: 103
Pubmed
(((((((((solasodin*) OR solasonin*) OR solamargin*) OR soladulcidin*) OR soladulcin*) OR solamarin*) OR soladinin*)) AND (((toxic*) OR adverse) OR terato*)
Results: 87
Scopus
((TITLE‐ABS‐KEY(solasodin*) OR TITLE‐ABS‐KEY (solasonin*) OR TITLE‐ABS‐KEY (solamargin*) OR TITLE‐ABS‐KEY (soladulcidin*) OR TITLE‐ABS‐KEY (soladulcin*) OR TITLE‐ABS‐KEY (solamarin*) OR TITLE‐ABS‐KEY (soladinin*))) AND ((TITLE‐ABS‐KEY (toxic*) OR TITLE‐ABS‐KEY (adverse) OR TITLE‐ABS‐KEY (terato*)))
Results: 107
(c) Leptin alkaloids
Web of science
TOPIC: (leptinin*) OR TOPIC: (leptinidin*) OR TOPIC: (leptin*) AND TOPIC: (toxic*) OR TOPIC: (adverse) OR TOPIC: (terato*) NOT TS= (fat OR adipos* OR adipocyt* OR diabetes OR obese OR obesity OR insulin* OR hormon* OR disease*)
Results:335
Pubmed
(((((leptinin*) OR leptinidin*) OR leptin*)) AND (((toxic*) OR adverse) OR terato*)) NOT (((((((((fat) OR adipos*) OR adipocyt*) OR diabetes) OR obese) OR obesity) OR insulin*) OR hormon*) OR disease*)
Results:294
Scopus
((TITLE‐ABS‐KEY (toxic*) OR TITLE‐ABS‐KEY (adverse) OR TITLE‐ABS‐KEY (terato*))) AND ((TITLE‐ABS‐KEY (leptinin*) OR TITLE‐ABS‐KEY (leptinidin*) OR TITLE‐ABS‐KEY (leptin*))) AND NOT ((TITLE‐ABS‐KEY (fat) OR TITLE‐ABS‐KEY (adipos*) OR TITLE‐ABS‐KEY (adipocyt*) OR TITLE‐ABS‐KEY (diabetes) OR TITLE‐ABS‐KEY (obese) OR TITLE‐ABS‐KEY (obesity) OR TITLE‐ABS‐KEY (insulin*) OR TITLE‐ABS‐KEY (hormon*) OR TITLE‐ABS‐KEY (disease*)))
Results: 321
All the retrieved references were uploaded in Endnote. Final number of hits after removal of duplicates were
Tomato alkaloids: 183 articles
Solanine alkaloids: 187 articles
Leptin alkaloids: 655 articles
The articles were screened for their relevance, exclusion criteria:
Tomato alkaloids: analytics/detection of compounds, synthesis of compounds, pest resistance, positive/pharmaceutical effects, plant defence mechanisms
Solanine alkaloids: analytics/detection of compounds, pest resistance, positive/pharmaceutical effects/clinical trials, toxicity on snails
Leptin alkaloids: synthesis of compounds, insects (e.g. Leptinotarsa), plant defence mechanism, pest resistance
Appendix C – Details of the study design of the toxicokinetic studies
1.
Table C.1.
Studies on the toxicokinetics of glycoalkaloids (GAs) in experimental animals
| Test compound | Study design (species, doses, administration routes, study duration) | Parameters | Reference |
|---|---|---|---|
| [3H]‐α‐chaconine (tritiated in the aglycone moiety at carbon atoms adjacent to the double bond and to the nitrogen atom), activity |
Male Sprague‐Dawley rats (200–300 g bw) Dosage by gavage (5 mg/kg bw) or i.p. injection (5, 10, 15 or 25 mg/kg bw) Rats housed in metabolic cages. 2 rats/treatment group euthanised at each time point Time points: 3, 6, 12, 24, 72 or 100 h following treatment |
Radioactivity distribution and elimination Metabolism studied by means of thin layer chromatography following ether extraction |
Norred et al. (1976) |
| [3H]‐α‐chaconine (no information on labelling position) | Female Swiss‐Webster mice (20 g bw) Experiments:
|
Subcellular distribution Accumulation in subcellular fractions Metabolism (enzyme induction) |
Sharma et al. (1983) |
| [3H]‐α‐chaconine (randomly labelled) |
Male Golden hamsters (130–150 g bw) Gavage or i.p. injection at a dose of 10 mg/kg bw 3 animals/time point euthanised at regular time intervals (3, 12, 24, 72 and 168 h following treatment) |
Tissue distribution Excretion Subcellular distribution |
Alozie et al. (1979a) |
| [3H]‐α‐chaconine (randomly labelled) |
Study design described in Alozie et al. (1979a) Collected urine and feces from animals orally exposed were repeatedly extracted with chloroform. The water soluble and chloroform soluble extracts were subject to TLC analysis for the identification of metabolites |
Metabolite identification | Alozie et al. (1979b) |
| [3H]‐α‐solanine (no information on the labelling position) |
Male rats (strain SPF Riv:TOX rats; 240–260 g bw; 12–13 weeks of age, implanted with a Colonisation Resistant Factor mouse flora) Male hamsters (SPF Charles River/Wiga Syrian golden hamsters; 130–160 g bw, 12–15 weeks of age) Dosage either by gavage (in 4 rats and 5 hamsters) at a dose of 170 μg/kg bw, or via i.v. injection (in 5 rats and 5 hamsters) at a dose of 54 μg/kg bw, respectively Animals were housed in metabolic cages for 7 days following the treatment. Urine, feces and exhaled air. Blood was regularly collected over the period after the treatment and analysed for the radioactivity level of unchanged α‐solanine |
Absolute bioavailability Kinetics in blood and plasma Excretion |
Groen et al. (1993) |
| [3H]‐α‐solanine (tritiated in the aglycone moiety at carbon atoms adjacent to the double bond and to the nitrogen atom) |
Male Fischer rats (180–250 g bw for rats subject to oral treatment, 95–170 g bw for rats treated by i.p. injection) Dosage by gavage (5 mg/kg bw) or i.p. injection (5, 10, 15 or 25 mg/kg bw). Number of rats/treatment group was not reported Rats were housed in metabolic cages and sacrificed after 3, 6, 12, 24, 48, 72 or 96 h after treatment Metabolites in urine and feces were studies by TLC analysis |
Tissue distribution Metabolic fate Excretion |
Nishie et al. (1971) |
bw: body weight; i.p.: intraperitoneal; i.v.: intravenous; TLC: thin‐layer chromatography.
Table C.2.
Details of studies reporting on the toxicokinetics of glycoalkaloids (GAs) in humans
| Test compound | Study design (subjects, doses, administration routes, study duration) | Parameters | Reference |
|---|---|---|---|
| α‐solanine, α‐chaconine and solanidine |
7 healthy male adults Treatment with a portion of peeled mashed potatoes containing 200 mg TGAs/kg (82 and 118 mg/kg for α‐solanine and α‐chaconine, respectively), adjusted to reach a dose of 1 mg TGAs/kg bw The study subjects were instructed to refrain from eating potatoes and potato products as from 48 h before and up to 25 h after the exposure Blood was sampled prior to the exposure and 1, 2, 3, 4, 5, 8 and 25 h following the exposure Levels of α‐solanine, α‐chaconine and solanidine in serum were measured via HPLC analysis |
Kinetic in blood serum (Cmax, Tmax, t1/2) | Hellenäs et al. (1992) |
| α‐solanine, α‐chaconine and solanidine |
6 male and 8 female healthy volunteers (age range: 18–45 years old) Dietary rules to refrain from other products containing GAs were followed as from 72 h prior to the treatment Treatments:
Blood was withdrawn 1 h and 30 min before treatment and 1, 2, 3, 4, 5, 6, 7, 8, 10, 12, 16, 20, 24, 32, 48, 56, 72 and 96 h after treatment Serum α‐solanine, α‐chaconine and solanidine levels were measured by HPLC analysis |
Kinetic in blood serum (Cmax, Tmax, t1/2) | Mensinga et al. (2005) |
| [3H]‐solanidine (tritiated in the aglycone moiety at carbon atoms adjacent to the nitrogen atom) |
Two male and one female volunteers Treatment via a single i.v. injection (doses not reported) Blood was sampled at various times up to 150 h Urine was collected at 2, 4, 8 and 12 h Feces were collected as individual voidings The distribution of solanidine in liver was studied in five post‐mortem liver samples. Four solanidine fractions were extracted:
|
Kinetic in plasma, urine and feces Distribution of solanidine in liver |
Claringbold (1982) |
| Solanidine |
34 human donors (7 males and 27 females), randomly selected from a hospital clinic Development of a radioimmunoassay method to quantify solanidine in human plasma. Serum from rabbits immunised using a solanine‐BSA conjugated was used. 100% cross reactivity was found for solanidine, α‐solanine, α‐chaconine and desmissidine. A chloroform extraction procedure showed allowed the specific quantification of solanidine |
Determination of solanidine in plasma | Matthew et al. (1983) |
| Solanidine |
57 volunteers (30 males, age range: 18–44 years old; 27 females, age range: 16–62 years old) Solanidine levels in plasma were analysed using the radioimmune assay developed by Matthew et al. (1983) The study subjects were asked to maintain their standard diet during winter (November–December), with the exclusion of two male volunteers who maintained a potato‐free diet for several weeks while their serum was regularly analysed for solanidine levels The consumption of potatoes and potato products of 33/57 subjects was recorded for one month |
Determination of solanidine in plasma | Harvey et al. (1985a) |
| Solanidine, TGA |
Two groups of volunteers:
GAs levels were analysed using a radioimmuno method adapted from Matthew et al. (1983) Exposure to GAs through normal diet (during summer), with the exclusion of three Swedish volunteers (2 m and 1 f), who had daily consumption of two potato cultivars known to be high in GAs (‘Ulster Chieftain’ and ‘SV72118’) for a week before serum and saliva sampling Blood and saliva samples were collected from all volunteers and analysed for the presence of solanidine (upon chloroform extraction) or total GA levels |
Solanidine and total GA levels in blood and saliva | Harvey et al. (1985b) |
HPLC: high‐performance liquid chromatography.
Appendix D – Comparison of developmental toxicity of single dose studies
1.
Table D.1.
Comparison of developmental toxicity of glycoalkaloids (GAs) and their aglycones from single dose studies
| Compound | Dose (mmol/kg bw) | Dose (mg/kg bw) | Abnormal litters (%) | Abnormal fetuses (%) | Reference |
|---|---|---|---|---|---|
| Solanidine N‐oxide | 0.14 | 58 | 0 | 0 | Gaffield and Keeler (1996a,b) |
| Solanidine | 0.29 | 115 | 19 | 2.3 | Gaffield and Keeler (1996a,b) |
| Solanidine N‐oxide | 0.29 | 120 | 0 | 0 | Gaffield and Keeler (1996a,b) |
| Solanidine | 0.43 | 175 | 50 | 16 | Gaffield and Keeler (1996a,b); other study |
| Solanidine | 0.44 | 176 | 50 | 24 | Gaffield and Keeler (1993) |
| Demissidine | 0.44 | 176 | 14 | 3 | Gaffield and Keeler (1996a,b) |
| Demissidine | 0.44 | 176 | 14 | 3 | Gaffield and Keeler (1993) |
| Solanidine N‐oxide | 0.43 | 178 | 50 | 16 | Gaffield and Keeler (1996a,b) |
| α‐Chaconine | 0.19 | 165 | 63 | 23 | Renwick et al. (1984) |
| α‐Chaconine | 0.21 | 179 | 20 | 2.7 | Gaffield and Keeler (1996a,b) |
| α‐Chaconine | 0.21 | 180 | 88 | 18 | Renwick et al. (1984) |
| α‐Solanine | 0.23 | 200 | 59 | 23 | Renwick et al. (1984) |
| α‐Solanine | 0.28 | 243 | 21 | 5.5 | Gaffield and Keeler (1996a,b) |
| α‐Chaconine | 0.29 | 247 | Not measurable, maternal lethality | Not measurable, maternal lethality | Gaffield and Keeler (1996a,b) |
| Solasodine | 3.4 | 1,400 | 75 | 29 | Gaffield and Keeler (1993) |
| Dihydrosolasodine | 3.4 | 1,400 | 45 | 6 | Gaffield and Keeler (1993) |
Appendix E – Inhibition of cholinesterases by GAs
1.
Appendix F – Rapid Alert System for Food and Feed (RASFF) reports on the presence of Solanum nigrum in food products
1.
Table F.1.
Rapid Alert System for Food and Feed (RASFF) reports on the presence of Solanum nigrum in food products
| Product category | Date | Reference | Product type | Notification type | Notification basis | Notified by | Countries concerned | Subject | Action taken | Distribution status | Risk decision |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Dietetic foods, food supplements, fortified foods | 9‐5‐2018 | 2018.1276 | Food | Information for follow‐up | Official control on the market | Poland | Cyprus (D), Germany (D), Greece (D), India (O), Italy (D), Latvia, Poland (D), Romania (D), Slovenia (D), Spain | Solanum nigrum and unauthorised novel food ingredients Cassia occidentalis, Achillea millefolium and Tamarix gallica in food supplement from India, via Latvia and via Spain | Distribution to other member countries | Undecided | |
| Dietetic foods, food supplements, fortified foods | 9‐5‐2018 | 2018.1278 | Food | Information for follow‐up | Official control on the market | Poland | Austria (D), Bulgaria (D), Cyprus (D), France (D), Germany (D), Greece (D), India (O), Italy (D), Latvia, Lithuania (D), Poland (D), Portugal (D), Romania (D), Slovakia (D), Slovenia (D), Spain (D), United Kingdom (D) | Solanum nigrum and unauthorised novel food ingredients Cassia occidentalis, Achillea millefolium and Tamarix gallica in food supplement from India, via Latvia and via Spain | Distribution to other member countries | Undecided | |
| Dietetic foods, food supplements, fortified foods | 6‐3‐2015 | 2015.AKE | Food | Border rejection | Border control – consignment detained | Portugal | India (O), Portugal | Unauthorised placing on the market (Solanum nigrum, Sida cordifolia Operculina turpethum and Nepeta hindostana) of food supplements from India | Import not authorised | Product not (yet) placed on the market | Undecided |
| Fruits and vegetables | 7‐6‐2013 | 2013.0804 | Food | Alert | Official control on the market | Germany | Germany (D), Netherlands (O) | Solanum nigrum (black nightshade) in young green beans from the Netherlands | Recall from consumers | Distribution restricted to notifying country | Serious |
| Fruits and vegetables | 18‐2‐2004 | 2004.086 | Food | Alert | Consumer complaint | France | Belgium (O), France (D) | Foreign body (50 green solanum nigrum in 2 samples) in frozen green beans | Product recall or withdrawal | Undecided | |
| Fruits and vegetables | 6‐11‐1985 | 1985.15 | Food | Alert | Germany | Germany, Netherlands (O) | Solanum nigrum (Berries of solanum nigrum) in beans – canned | Undecided | |||
| Fruits and vegetables | 13‐10‐1982 | 1982.07 | Food | Alert | Italy | Belgium (O), Italy | Solanum nigrum in Green beans – quick‐frozen | Undecided |
Appendix G – Studies on the toxicity of Glycoalkaloids not considered in the risk assessment
1.
| Excluded studies | Reason for exclusion |
|---|---|
| Baker DC, Keeler RF and Gaffield WP, 1988. Mechanism of death in Syrian hamsters gavaged potato sprout material. Toxicologic Pathology, 16, 333–339. | In the study, no GA concentrations were determined |
| Bell DP, Gibson JG, McCarroll AM and McClean GA, 1976. Embryotoxicity of solanine and aspirin in mice. Journal of Reproduction and Fertility, 46, 257–259. | i.p. study. Although the authors call it solanine, in fact a‐solanine was used (standard from Sigma) |
| Caldwell KA, Grosjean OK, Henika PR and Friedman M, 1991. Hepatic ornithine decarboxylase induction by potato glycoalkaloids in rats. Food Chemistry and Toxicology, 29, 531–535. | Excluded because additional application of dexamethasone in all treatment groups, no proper control |
| Candeias MF, Abreu P, Pereira A and Cruz‐Morais J, 2009. Effects of strictosamide on mouse brain and kidney Na+, K+‐ATPase and Mg2+‐ATPase activities. Journal of Ethnopharmacology, 12, 117–122. | Strictosamide is not a potato GA |
| Chaube S, Swinyard CA and Daines RH, 1973. Failure to induce malformations in fetal rats by feeding blighted potatoes to their mothers. Lancet, 1, 329–330. | In the study, no GA concentrations were determined |
| Crawford L and Myrh B, 1995. A preliminary assessment of the toxic and mutagenic potential of steroidal alkaloids in transgenic mice. Food Chemistry and Toxicology, 33, 191–194. | A preliminary study without proper controls and without value for the mutagenicitiy assessment |
| Friedman M, Rayburn JR and Bantle JA, 1992. Structural relationships and developmental toxicity of Solanum alkaloids in the frog embryo teratogenesis assay‐Xenopus. Journal of Agricultural and Food Chemistry, 40, 1617–1624. | Studies in Xenopus are not informative for the risk assessment |
| Gupta AK, Ganguly P, Majumder UK and Ghosal S, 2009. Antidiabetic and antihyperlipidaemic effects of Solanum Xanthocarpum total extract in alloxan induced diabetic rats. Pharmacologyonline, 1, 484–497. | In the study, no GA composition and concentration were determined |
| Hellenäs KE, Cekan E, Slanina P and Bergman K, 1992. Studies of embryotoxicity and the incidence of external malformations after continuous intravenous infusion of α‐chaconine in pregnant rats. Pharmacology and Toxicology, 70, 381–383. | i.v. excluded from the consideration in the experimental animal section, except in the acute toxicity for comparison with the oral route |
| Hornfeldt CS and Collins JE, 1990. Toxicity of nightshade berries (Solanum dulcamara) in mice. Clinical Toxicology, 28, 185–192. | In the study, no GA composition and concentration were determined |
| Ji Y, Sun J and Lang L, 2011. Analysis of testicular toxicity of solanine in mice. Advanced Materials Research, 282, 195–200. | i.p. administration |
| Patel VB, Rathod IS, Patel JM and Brahmbhatt MR 2010. Anti‐urolithiatic and natriuretic activity of steroidal constituents of Solanum xanthocarpum. Der Pharma Chemica, 2, 173–176. | No proper methodical description of the experiments and no proper description of isolation and purification procedure. Purity of the tested substances is highly questionable |
| Peičev P, Nikiforov N and Rusev R, 1967. Pharmacological properties of glycoalkaloids, obtained from Solanum tuberosum L. Experimental Medicine and Morphology. VI, 1. | Extract used of Herba Solani (leaves of S. tuberosum), purity of isolated GA is not described, documentation is incomplete |
| Poswillo DE, Sopher D and Mitchell S, 1972. Experimental induction of foetal malformation with ‘‘blighted’’ potato: a preliminary report. Nature, 239, 462–464. | In the study, no GA composition and concentration were determined |
| Poswillo DE, Sopher D, Mitchell SJ, Coxon DT, Curtis RF and Price KR, 1973. Further investigations into the teratogenic potential of imperfect potatoes. Symposium on Potatoes and Birth defects, 8, 339–347. | Only TGA content reported. Reported TGA content is much lower than generally found in (healthy) potato tubers, indicating an error in the analytical methodology used |
| Schwarz A, Pinto E, Haraguchi M, de Oliveira CA, Bernardi MM and de Souza Spinosa H, 2007. Phytochemical study of Solanum lycocarpum (St. Hil) unripe fruit and its effects on rat gestation. Phytotherapy Research, 21, 1025–1028. | Extracts of S. lycocarpum were tested, which is not a food plant |
| Sharma RP, Willhite CC, WU MT and Salunkhe DK, 1978. Teratogenic potential of blighted potato concentrate in rabbits, hamsters and miniature swine. Teratology, 18, 55–62. | In the study, no GA composition and concentration were determined |
| Wang XG, 1993. Teratogenic effect of potato glycoalkaloids. Zhonghua Fu Chan Ke Za Zhi, 28, 121–122. | Abstract only. Intra‐abdominal administration. Article in Chinese |
| Yoon DJ and Kirkowski A, 1979. Effects of dietary Solanaceous potatoes on the serum, calcium, phosphorus, magnesium and hydroxyproline levels in rats. Korean Journal of Animal Science, 21, 401–406. | In the study, no GA composition and concentration were determined |
| Allen et al., 1977. Teratogenicity studies on late blighted potatoes in nonhuman primates (Macaca mulatta and Saguinus labiatus). Teratology, 15, 17–23. | Only TGA content reported. Reported TGA content is much lower than generally found in (healthy) potato tubers, indicating an error in the analytical methodology used |
| Rao MV, 1988. Effects of alcoholic extract of Solanum xanthocarpum seeds in adult male rats. Indian Journal of Experimental Biology, 26, 95–98. | Extract of Solanum xanthocarpum seeds. In the study, no GA composition and concentration were determined |
| Mali PC, Chaturvedi M and Dixit VP, 1996. Antispermatogenic activity of Solanum xanthocarpum S and W (50 percent EtOH‐extract) in rats. Journal of Phytological Research, 9, 13–17. | Extract of Solanum xanthocarpum root. In the study, no GA composition and concentration were determined |
Appendix H – Additional scenario for the human risk characterisation
1.
Table H.1.
Summary statistics of the % of survey days with an intake of potato TGAs above the LOAEL of 1 mg/kg bw calculated only for days of potato consumption
| Age class | Range of mean % of days with an intake of potato TGAs above the LOAEL of 1 mg/kg bw per daya | |
|---|---|---|
| Minimum | Maximum | |
| Infants | 0.0 (0.0–0.0) | 0.4 (0.0–0.9) |
| Toddlers | < 0.1 (0.0–0.2) | 1.0 (0.0–3.3) |
| Children | < 0.1 (0.0–0.2) | 0.6 (0.0–1.4) |
| Adolescents | 0.0 (0.0–0.0) | 0.2 (0.0–0.7) |
| Adults | 0.0 (0.0–0.0) | < 0.1 (0.0–0.1) |
| Elderly | 0.0 (0.0–0.0) | < 0.1 (0.0–0.4) |
| Very elderly | 0.0 (0.0–0.0) | < 0.1 (0.0–0.0) |
The mean percentage of days with an intake of potato TGAs above the LOAEL of 1 mg/kg bw per age group for each survey was calculated by averaging the percentage of days above the reference for each subject and then averaged over all subjects within the age group for each survey. The process was iterated 1,000 times as for the exposure estimates, and a mean of the mean percentage of days above the reference per age group was calculated together with a 95% confidence interval.
Note: The corresponding 95% confidence intervals (95% CI) are given in the brackets.
Annex A – Occurrence data in food and feed submitted to EFSA and dietary exposure assessment for humans
1.
Annex A can be found as a separate document available online on the EFSA Knowledge Junction community at: http://doi.org/10.5281/zenodo.3939104
Description: This Annex is an Excel file which presents tables on GAs on occurrence data in food and feed, and dietary exposure assessment for humans.
Suggested citation: EFSA CONTAM Panel (EFSA Panel on Contaminants in the Food Chain) , Schrenk D, Bignami M, Bodin L, Chipman JK, del Mazo J, Hogstrand C, Hoogenboom LR, Leblanc J‐C, Nebbia CS, Nielsen E, Ntzani E, Petersen A, Sand S, Schwerdtle T, Vleminckx C, Wallace H, Brimer L, Cottrill B, Dusemund B, Mulder P, Vollmer G, Binaglia M, Ramos Bordajandi L, Riolo F, Roldán‐Torres R and Grasl‐Kraupp B, 2020. Scientific Opinion – Risk assessment of glycoalkaloids in feed and food, in particular in potatoes and potato‐derived products. EFSA Journal 2020;18(8):6222, 190 pp. 10.2903/j.efsa.2020.6222
Requestor: European Commission
Question number: EFSA‐Q‐2016‐00811
Panel members: Margherita Bignami, Laurent Bodin, James Kevin Chipman, Jesús del Mazo, Bettina Grasl‐Kraupp, Christer Hogstrand, Laurentius (Ron) Hoogenboom, Jean‐Charles Leblanc, Carlo Stefano Nebbia, Elsa Nielsen, Evangelia Ntzani, Annette Petersen, Salomon Sand, Dieter Schrenk, Tanja Schwerdtle, Christiane Vleminckx and Heather Wallace.
Acknowledgements: The Panel wishes to thank the following for the support provided to this scientific output: Kelly Niermans. The Panel wishes to acknowledge all European competent institutions, Member State bodies and other organisations that provided consumption and occurrence data for this scientific output.
Adopted: 7 July 2020
Reproduction of the images listed below is prohibited and permission must be sought directly from the copyright holder:
Figure 2: © Elsevier. Figure 3: © Springer. Figure 7: © American Chemical Society, Springer.
This publication is linked to the following EFSA Supporting Publications article: http://onlinelibrary.wiley.com/doi/10.2903/sp.efsa.2020.EN-1905/full
Notes
Commission Regulation (EU) No 6812013 of 16 January 2013 on the Catalogue of feed materials. OJL 29, 30.1.2013, p. 1.
Reprinted from Food research, 77, Faridnia F, Burritt DJ, Bremer PJ and Oey I, Innovative approach to determine the effect of pulsed electric fields on the microstructure of whole potato tubers: Use of cell viability, microscopic images and ionic leakage measurements, 556–564, 2015, with permission from Elsevier.
Reprinted by permission from Springer: Potato Research, Potato Steroidal Glycoalkaloids: Biosynthesis and Genetic Manipulation, Ginzberg I, Tokuhisa JG and Veilleux RE, 2009.
Council Regulation (EEC) No 315/93 of 8 February 1993 laying down Community procedures for contaminants in food. OJ L 37, 13.2.1993, p. 1–3.
Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. OJ L 364, 20.12.2006, p. 5–24.
Föreskrifter om ändring i Livsmedelsverkets föreskrifter (SLVFS 1993:36) om vissa främmande ämnen i livsmedel, LIVSFS 2002:16. Available online: https://www.livsmedelsverket.se/globalassets/om-oss/lagstiftning/nummerordning---upphord-lagstiftning/2004/livsfs-2004-7-frammande-amnen.pdf
In 2017 Health Canada proposed to transfer the existing maximum level for TGA in potato tubers from the List of Maximum Levels for Various Chemical Contaminants in Food to the regulatory List of Contaminants and Other Adultering Substances in Food, thereby removing this maximum level from the first list.
Web of Science (WoS), formerly ISI Web of Knowledge, Thomson Reuters. Available at: http://thomsonreuters.com/thomson-reuters-web-of-science/
PubMed, Entrez Global Query Cross‐Database Search System, National Center for Biotechnology Information (NCBI), National Library of Medicine (NLM), Department of the National Institutes of Health (NIH), United States Department of Health and Human Services. Available at: http://www.ncbi.nlm.nih.gov/pubmed/
EndNote X5, Thomson Reuters. Available at: http://endnote.com/
From 1 January 2014 onwards, Evidence Management Unit (DATA).
Current call: https://www.efsa.europa.eu/en/consultations/call/190410
Commission Regulation (EU) No 68/2013 of 16 January 2013 on the Catalogue of feed materials. OJ L 29, 30.1.2013, p. 1–64.
Source: http://www.4petsusa.com
Assuming 650 kg bw.
Report on the Toxic Chemical Syndrome. Definitions and Nomenclature Workshop. May 8‐9 2012. Available at: https://chemm.nlm.nih.gov/Report_from_Toxic_Syndrome_Workshop_final_with_ACMT_edits_cover.pdf
International Union of Biochemistry and Molecular Biology (IUBMB) enzyme nomenclature.
Starch potatoes are specifically grown for the production of potato starch. They are not used for human consumption.
https://www.food.gov.uk/print/pdf/node/281
https://www.fda.gov/food/chemicals/acrylamide‐and‐diet‐food‐storage‐and‐food‐preparation
Commission Regulation (EU) 2017/2158 of 20 November 2017 establishing mitigation measures and benchmark levels for the reduction of the presence of acrylamide in food. OJ L304/24.
In this opinion, the terminology ‘potato crisps’ refers to crunchy thin slices of deep‐fried/baked potato usually eaten as snacks, whereas ‘French fries’ refers to batons of deep‐fried potato usually served as an accompaniment during a meal. When citing literature data, the original food name is used. If it is different from the standard terminology used in the opinion, then the standard terminology is also indicated in brackets, when possible.
Reprinted with permission from Tajner‐Czopek A, Rytel E, Aniolowska M and Hamouz K, 2014. The influence of French fries processing on the glycoalkaloid content in coloured‐fleshed potatoes. European Food Research and Technology, 238, 895‐904. Copyright 2009, American Chemical Society.
They may be chopped or sliced to reduce the risk of choking.
The authors report the presence of ‘solanine’ in tomato and tomato products. It should be noted that the analytical method (colourimetry) used in this publication cannot distinguish between α‐solanine, α‐chaconine and α‐dehydrotomatine. It is therefore likely that the authors have measured α‐dehydrotomatine.
References
- Abbasi KS, Masud T, Qayyum A, Ahmad A, Bibi Y, Mehmood A and Sher A, 2016. Photo‐induced changes in quality attributes of potato tubers during storage. Journal of Applied Botany and Food Quality, 89, 315–321. [Google Scholar]
- Abdel‐Gawad A, Omer EA and Ahmed SS, 1993. Effect of storage on the glycoalkaloids and carbohydrate contents in different potato varieties. Egyptian Journal of Horticulture, 19, 81–92. [Google Scholar]
- Abreu P, Relva A, Matthew S, Gomes Z and Morais Z, 2007. High‐performance liquid chromatographic determination of glycoalkaloids in potatoes from conventional, integrated, and organic crop systems. Food Control, 18, 40–44. [Google Scholar]
- Afify AEMR, El‐Beltagi HS, Aly AA and El‐Ansary AE, 2012. The impact of γ‐irradiation, essential oils and iodine on biochemical components and metabolism of potato tubers during storage. Notulae Botanicae Horti Agrobotanici, 40, 129–139. [Google Scholar]
- Ahmed SS and Műller K, 1978. Effect of wond‐damages on the glycol‐alkaloid content in potato tubers and chips. Lebenmittel Wiss und Technology, 11, 144–146. [Google Scholar]
- Al Chami L, Mendez R, Chataing B, O'Callaghan J, Usubillaga A and LaCruz L, 2003. Toxicological effects of alpha‐solamargine in experimental animals. Phytotherapy Research, 17, 254–258. 10.1002/ptr.1122 [DOI] [PubMed] [Google Scholar]
- Al Sinani SSS and Eltayeb EA, 2017. The steroidal glycoalkaloids solamargine and solasonine in Solanum plants. South African Journal of Botany, 112, 253–269. [Google Scholar]
- Alberti B, 1932. Zum Nachweis von Solanin. Zeitschrift fuer Untersuchung der Lebensmittel, 64, 260–262. [Google Scholar]
- Aldous CN, Sharma RP and Salunkhe DK, 1980. Effects of α‐chaconine on brain biogenic amines, electroencephalogram, cardiac rates and respiratory response in rats. Journal of Food Safety, 2, 20–33. [Google Scholar]
- Alfa J and Heyl E, 1923. Kartoffeln 1922‐er ernte mit ausserordentlich hohem solaningehalt. Zeitschrift für Untersuchung der Nahrungs‐ und Genußmittel, sowie der Gebrauchsgegenstände, 46, 306–309. [Google Scholar]
- Allen JR, Marlar Dr RJ, Chesney CF, Helgeson JP, Kelman A, Weckel KG, Traisman E and White JW Jr, 1977. Teratogenicity studies on late blighted potatoes in nonhuman primates (Macaca rnulatta and Saguinus labiatus). Experimental Teratology, 15, 17–23. [DOI] [PubMed] [Google Scholar]
- Almeida AE, Cardoso CRP, Almeida DV, Moreira RRD, Silva M and Varanda EA, 2010. Mutagenic activity of glycoalkaloids from Solanum palinacanthum Dunal (Solanacea) found in the Brazilian cerrado. Latin American Journal of Pharmacy, 29, 122–126. [Google Scholar]
- Alozie SO, Sharma RP and Salunkhe DK, 1978. Inhibition of rat cholinesterase isoenzymes in vitro and in vivo by the potato alkaloid, α‐chaconine. Journal of Food Biochemistry, 2, 259–276. [Google Scholar]
- Alozie SO, Sharma RP and Salunkhe DK, 1979a. Physiological disposition, subcellular distribution and tissue binding of [3H]‐α‐chaconine. Journal of Food Safety, 1, 257–273. [Google Scholar]
- Alozie SO, Sharma RP and Salunkhe DK, 1979b. Excretion of alpha‐chaconine‐3H, a steroidal glycoalkaloid from Solanum tuberosum L. and its metabolites in hamsters. Pharmacological Research Communications, 11, 483–490. [DOI] [PubMed] [Google Scholar]
- Álvarez MA, Rodríguez Talou J, Paniego NB and Giulietti AM, 1994. Solasodine production in transformed organ cultures (roots and shoots) of Solanum eleagnifolium Cav. Biotechnology Letters, 16, 393–396. [Google Scholar]
- Amer FS, Reddivari L, Madiwale GP, Stone M, Holm DG and Vanamala J, 2014. Effect of genotype and storage on glycoalkaloid and acrylamide content and sensory attributes of potato chips. American Journal of Potato Research, 91, 632–641. [Google Scholar]
- Anses (Agence Nationale de Sécurité Sanitaire de l'Alimentation, de l'Environnment et du Travail), 2014. Appui scientifique et technique de l'Anses, Saisine n°2014‐SA-0115 Note d'appui scientifique et technique de l'Agence nationale de sécurité sanitaire de l'alimentation, de l'environnement et du travail relatif aux intoxications liées à la consommation de tomates cerise en provenance du Maroc. Maisons‐Alfort. 13 pp. Available online: https://www.anses.fr/fr/system/files/ERCA2014sa0115.pdf
- AOAC (Association of Official Agricultural Chemists), 2000. AOAC official method 997.13: Glycoalkaloids (a‐Solanine and a‐Chaconine) in potato tubers. 3.
- Arena MP, Govers C, Lotti C, Ricciardi L, Wichers HJ and Mes JJ, 2018. The effect of tomatine on gene expression and cell monolayer integrity in Caco‐2. Molecules, 23, 644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azim A, Shaikh HA and Ahmad R, 1982. Effect of feeding greened potatoes on different visceral organs and blood plasma of rabbits. Journal of the Science of Food and Agriculture, 33, 1275–1279. [DOI] [PubMed] [Google Scholar]
- Azim A, Shaikh HA and Ahmad R, 1983. Toxic effects of high glycoalkaloid feeding on the protein digestibility and growth of rabbits. Journal of Pharmacy, 2, 15–24. [Google Scholar]
- Azim A, Shaikh HA and Ahmad R, 1984. Toxic effects of high glycoalkaloid feeding on the RBC counts and hemoglobin concentration of rabbit blood. Journal of Pharmacy, 3, 43–49. [Google Scholar]
- Bagheri M, Bushehri AAS, Hassandokht MR and Naghavi MR, 2017. Evaluation of solasonine content and expression patterns of SGT1 gene in different tissues of two Iranian Eggplant (Solanum melongena L.) genotypes. Food Technology and Biotechnology, 55, 236–242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bajaj KL, Kaur G and Chadha ML, 1979. Glycoalkaloid content and other chemical constituents of the fruits of some egg plant (Solanum melongena, L.) varieties. Journal of Plant Foods, 3, 163–168. [Google Scholar]
- Baker DC, Keeler RF and Gaffield W, 1989. Pathology in hamsters administered Solanum plant species that contain steroidal alkaloids. Toxicon, 27, 1331–1337. [DOI] [PubMed] [Google Scholar]
- Baker DC, Keeler RF and Gaffield WP, 1988. Mechanism of death in Syrian hamstyers gavaged potato sprout material. Toxicologic Pathology, 16, 333–339. [DOI] [PubMed] [Google Scholar]
- Bal A, 1989. Visie van het Ministerie van Landbouw en Visserij op natuurlijke toxinen. [Vision of the Netherlands Ministry of Agriculture and Fisheries on natural toxins] De Ware(n)‐Chemicus 19 (November), 149–151. [Google Scholar]
- Baldina S, Picarella ME, Troise AD, Pucci A, Ruggieri V, Ferracane R, Barone A, Fogliano V and Mazzucato A, 2016. Metabolite profiling of Italian tomato landrces with different fruit types. Frontiers in Plant Science, 7, 664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barceloux DG, 2009. Potatoes, tomatoes, and solanine toxicity (Solanum tuberosum L., Solanum [lycopersicum L.).]. Disease‐a‐month: DM, 55, 391–402. 10.1016/j.disamonth.2009.03.009 [DOI] [PubMed] [Google Scholar]
- Bártová V, Diviš J, Bárta J, Brabcová A and Švajnerova M, 2013. Variation of nitrogenous components in potato (Solanum tuberosum L.) tubers produced under organic and conventional crop management. Euroepan Journal of Agronomy, 49, 20–31. [Google Scholar]
- Beausoleil JL, Spergel JM and Pawlowski NA, 2001. Anaphylaxis to raw potato. Annals of Allergy, Asthma and Immunology, 86, 68–70. 10.1016/s1081-1206(10)62359-9 [DOI] [PubMed] [Google Scholar]
- Beier RC and Nigg HN, 1992. Natural toxicants in foods. Phytochemical Resources for Medicine and Agriculture, 247–367. [Google Scholar]
- Bejarano L, mingolet E, Devaux A, Espinola N, Carrasaco E and Larondelle Y, 2000. Glycoalkaloids in potato tubers: the effect of variety and drought stress on the a‐solanine and a‐chaconine contents of potatoes. Journal of the Science of Food and Agriculture, 80, 2096–2100. [Google Scholar]
- Benilova IV, Soldatkin AP, Martelet C and Jaffrezic‐Renault N, 2006. Nonfaradaic impedance probing of potato glycoalkaloids interaction with butyrylcholinesterase immobilized onto gold electrode. Electroanalysis, 18, 1950–1956. 10.1002/elan.200603601 [DOI] [Google Scholar]
- Bergers WWA, 1981. Investigation of the contents of phenolic and alkaloidal compounds of gamma irradiated potatoes during storage. Food Chemistry, 7, 47–61. [Google Scholar]
- BfR (Bundesinstitut fűr Risikobewertung), 2018a. BfR Opinion No 010/2018 of 23 April 2018. Table potatoes should contain low levels of glycoalkaloids (solanine) (Abstract in English). Available online: http://www.bfr.bund.de/cm/349/table-potatoes-should-contain-low-levels-of-glycoalkaloids-solanine.pdf
- BfR (Bundesinstitut fűr Risikobewertung), 2018b. Stellungnahme Nr. 010/2018 des BfR vom 23. April 2018. Speisekartoffeln sollten niedrige Gehalte an Glykoalkaloiden (Solanin) enthalten (in German). Available online: http://www.bfr.bund.de/cm/343/speisekartoffeln-sollten-niedrige-gehalte-an-glykoalkaloiden-solanin-enthalten.pdf
- Bhat R and Pallyath G, 2016. In: Caballero B, Finglas PM. and Toldra F. (eds.). Fruits of tropical climates: dietary importance and health benefits, in ‘Encyclopedia of food and health’. Elsevier, Amsterdam: 144 pp. [Google Scholar]
- Bintcliffe EJB, Clydesdale A and Draper SR, 1982. Effect of genotype, site and season on the glycoalkaloid content of potato tubers. Journal of the National Institute of Agricultural Botanic, 16, 86–91. [Google Scholar]
- Bitsch R, Sultemeier J and Hotzel D, 1974. Quality retention of potatoes during transport and storage as influenced by various packaging materials. II. Effects of various packaging materials on solanine formation. Nahrung, 18, 395–408. [DOI] [PubMed] [Google Scholar]
- Blankemeyer JT, Atherton R and Friedman M, 1995. Effect of potato glycoalkaloids α‐chaconine and α‐solanine on sodium active transport in frog skin. Journal of Agricultural and Food Chemistry, 43, 636–639. [Google Scholar]
- Blankemeyer JT, White JB, Stringer BK and Friedman M, 1997. Effect of α‐tomatine and tomatidine on membrane potential of frog embryos and active transport of ions in frog skin. Food and Chemical Toxicology, 35, 639–646. [DOI] [PubMed] [Google Scholar]
- Bodart P, Kabengera C, Noirfalise A, Hubert P and Angenot L, 2000. Determination of α‐solanine and α‐chaconine in potatoes by high‐performance thin‐layer chromatography/densitometry. Journal of AOAC International, 83, 1468–1473. [PubMed] [Google Scholar]
- Bolin FM, 1962. Green potatoes can kill sheep. North Dakota Farm Research, 22, 15. [Google Scholar]
- Bömer A and Mattis H, 1923. Über hohe Solaningehalte bei Kartoffeln. Zeitschrift fűr Untersuchung der Nahrungs‐ und Genussmittel sowie der Gebrauchsgegenstaende, 45, 288–291. [Google Scholar]
- Bömer A and Mattis H, 1924. Der Solaningehalt in der Kartoffel (The solanine content of potatoes). Zeitschrift fűr Untersuchung der Nahrungs‐ und Genussmittel sowie der Gebrauchsgegenstaende, 47, 97–127. [Google Scholar]
- Bradshaw JE, Bryan GJ and Ramsay G, 2006. Genetic resources (including wild and cultivated Solanum species) and progress in their utilisation in potato breeding. Potato Research, 49, 49–65. [Google Scholar]
- Branzell C and Hellenäs K‐E, 1999. Undvik höga halter glykoalkaloider: Välj potatis av rätt sort!. Vår Föda, 3, 4–8. [Google Scholar]
- Brimijoin S, Chen VP, Pang YP, Geng L and Gao Y, 2016. Physiological roles for butyrylcholinesterase: a BChE‐ghrelin axis. Chemico‐Biological Interactions, 259, 271–275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brimijoin S, Gao Y, Geng L and Chen VP, 2018. Treating cocaine addiction, obesity, and emotional disorders by viral gene transfer of butyrylcholinesterase. Frontiers in Pharmacology, 9, 112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown D and Keeler RF, 1978. Structure‐activity relation of steroid teratogens. 3. Solanidan epimers. Journal of Agricultural and Food Chemistry, 26, 566–569. [DOI] [PubMed] [Google Scholar]
- Bushway RJ and Ponnampalam R, 1981. α‐Chaconine and α‐solanine content of potato products and their stability during several modes of cooking. Journal of Agricultural and Food Chemistry, 29, 814–817. [Google Scholar]
- Bushway RJ, Bureau JL and McGann DF, 1983. Alpha‐chaconine and alpha‐solanine content of potato peels and potato peel products. Journal of Food Science, 8, 84–86. [Google Scholar]
- Bushway RJ, McGann DF and Bushway AA, 1984. Gas chromatographic method for the determination of solanidine and its application to a study of feed‐milk transfer in the cow. Journal of Agricultural and Food Chemistry, 32, 548–551. [Google Scholar]
- Bushway RJ, Savage SA and Ferguson BS, 1987. Inhibition of acetyl cholinesterase by solanaceous glycoalkaloids and alkaloids. American Potato Journal, 64, 409–413. [Google Scholar]
- Bushway RJ, Perkins LB, Paradis LR and Vanderpan S, 1994. High‐performance liquid chromatographic determination of the tomato glycoalkaloid, tomatine, in green and red tomatoes. Journal of Agricultural and Food Chemistry, 42, 2824–2829. [Google Scholar]
- Calf OW, Huber H, Peters JL, Weinhold A and van Dam NM, 2018. Glycoalkaloid composition explains variation in slug resistance in Solanum dulcamara. Oecologia, 187, 495–506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canada Diseases Weekly Report , 1984. Solanine food poisoning associated with a school lunch program ‐ Alberta. May 1984. Canada Diseases Weekly Report, Health and Welfare Canada, 71, 10–18 Available online: http://publications.gc.ca/collections/collection_2016/aspc-phac/H12-21-1-10-18.pdf [Google Scholar]
- Caprioli G, Logrippo S, Cahill MG and James KJ, 2014. High‐performance liquid chromatography LTQ‐Orbitrap mass spectrometry method for tomatidine and non‐target metabolites quantification in organic and normal tomatoes. International Journal of Food Sciences and Nutrition, 65, 942–947. [DOI] [PubMed] [Google Scholar]
- Caruso G, Pokluda R, Sekara A, Kalisz A, Jezdinsky A, Kopta T and Grabowska A, 2017. Agricultural practices, biology and quality of eggplant cultivated in Central Europe. A review. Horticultural Science, 44, 201–212. [Google Scholar]
- Cavlovic P, Mankotia M, Pantazopoulos P and Scott PM, 2003. Liquid chromatographic determination of alpha‐solasonine in frozen green peas as an indicator of the presence of nightshade berries. Journal of AOAC International, 86, 759–763. [PubMed] [Google Scholar]
- Cayen MN, 1971. Effect of dietary tomatine on cholesterol metabolism in the rat. Journal of Lipid Research, 12, 482–490. [PubMed] [Google Scholar]
- Ceriola F, Portis E, Toppino L, Barchi L, Acciarri N, Ciriaci T, Sala T, Rotino GL and Lankri S, 2013. The population structure and diversity of eggplant from Asia and the Mediterranean Basin. PLoS ONE, 8, e73702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaube S and Swinyard CA, 1976. Teratological and toxicological studies of alkaloidal and phenolic compounds from Solanum tuberosum L. Toxicology and Applied Pharmacology, 36, 227–237. [DOI] [PubMed] [Google Scholar]
- Chaube S, Swinyard CA and Daines RH, 1973. Failure to induce malformations in fetal rats by feeding blighted potatoes to their mothers. The Lancet, 1, 329–330. [DOI] [PubMed] [Google Scholar]
- Chen X, Ding Y and Kan J, 2018. Changes in the content and influence factors of α‐solanine in potato during storage. Emirates Journal of Food and Agriculture, 30, 10–16. [Google Scholar]
- Choi E‐M, 2007. Suppressive effects of potato (Solanum tuberlosum) on type II [conagen‐induced arthritis in DBA/1J mice]. Food Science and Biotechnology, 16, 43–48. [Google Scholar]
- Choi E and Koo S, 2005. Anti‐nociceptive and anti‐inflammatory effects of the ethanolic extract [of potato (Solanum tuberosum). Food and Agricultural Immunology, 16, 29–39. 10.1080/09540100500064320 [DOI] [Google Scholar]
- Choi EM, Lee KH and Koo SJ, 2005. Effects of ethanol extract of potato (Solanum tuberlosum) on Freund's complete adjuvant‐induced model of chronic arthritis in mice. Food Science and Biotechnology, 14, 228–232. [Google Scholar]
- Choi S‐H, Lee SH, Kim HJ, Lee IS, Kozukue N, Levin CE and Friedman M, 2010. Changes in free amino acid, phenolic, chlorophyll, carotenoid, and glycoalkaloid contents in tomatoes during 11 stages of growth and inhibition of cervical and lung human cancer cells by green tomato extracts. Journal of gricultural and Food Chemistry, 58, 7547–7556. [DOI] [PubMed] [Google Scholar]
- Cichon MJ, Riedl KM and Schwartz SJ, 2017. A metabolomic evaluation of the phytochemical composition of tomato juices being used in human clinical trials. Food Chemistry, 28, 270–278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cieslik E and Praznik W, 1998. Changes of glycoalkaloid content in potato tubers of selected varieties during vegetation and storage. Polish Journal of Food and Nutrition Sciences, 7, 417–422. [Google Scholar]
- Claringbold , 1982. Kinetics and retention of solanidine in man. Xenobiotica, 12, 293–302. [DOI] [PubMed] [Google Scholar]
- Clarke G, Fall CHD, Lincoln DW and Merrick LP, 1978. Effects of cholinoceptor antagonists on suckling‐induced and experimentally evoked release of oxytocin. British Journal of Pharmacology, 63, 519–527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clement E and Verbist JF, 1980. The determination of solanine in Solanum tuberosum L. tubers: a comparative study of nine colorimetric methods. Lebensmittel‐Wissenschaft und ‐Technologie, 13, 202–206. [Google Scholar]
- Čolović MB, Krstić DZ, Lazarević‐Pašti TD, Bondžić AM and Vasić VM, 2013. Acetylcholinesterase Inhibitors: Pharmacology and Toxicology. Current Neuropharmacology, 11, 315–335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conner HW, 1937. Effect of light on solanine synthesis in the potato tuber. Plant Physiology, 12, 79–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper MR and Johnson AW, 1984. Poisonous plants in Britain and their effects on Animals and Man. Reference Book 161, Ministry of Agriculture, Fisheries and Food. HMSO, UK.
- Cortical , 1888. Accidents d'intoxication survenus au 139c d'infanterie à Lyon, le 11 et 12 Juillet 1888, et imputés à la consommation de pommes de terre de mauvaise qualité. Archives de Medicine et de Pharmacie militaire, 4, 3. As cited by Rothe (1919).
- Crawshaw R, 2001. Co‐product feeds: animal feeds from the food and drinks industries. Wiley‐Blackwell, Nottingham University.
- Dale MFB, Griffiths DW, Bain H and Goodman BA, 1997. The effect of gamma irradiation on glycoalkaloid and chlorophyll synthesis in seven potato cultivars. Journal of the Science of Food and Agriculture, 75, 141–147. [Google Scholar]
- Dale MFB, Griffiths DW and Bain HF, 1998. Effect of bruising on the total glycoalkaloid and chlorogenic acid content of potato (Solanum tuberosum) tubers of five cultivars. Journal of the Science of Food and Agriculture, 77, 499–505. [Google Scholar]
- Dale MFB, Griffiths DW, Bain H and Todd D, 1993. Glycoalkaloid increase in Solanum tuberosum on exposure to light. Annals of Applied Biology, 123, 411–418. [Google Scholar]
- Dalvi RR, 1985. Comparative assessment of the effect of solanine administered orally and intraperitoneally on hepatic dysfunction in male rats. Nihon Juigaku Zasshi. The Japanese Journal of Veterinary Science, 47, 657–659. [DOI] [PubMed] [Google Scholar]
- Damon SR, 1928. Food Infections and Food Intoxications. Chapter XI Potato poisoning. Williams & Wilkins, Baltimore, Maryland: pp. 153–157. [Google Scholar]
- Dao L and Friedman M, 1994. Chlorophyll, chlorogenic acid, glycoalkaloid, and protease inhibitor content of fresh and green potatoes. Journal of Agricultural and Food Chemistry, 42, 633–639. [Google Scholar]
- Dastmalchi K, Perez Rodriguez M, Lin J, Yoo B and Stark RE, 2019. Temporal resistance of potato tubers: Antibacterial assays and metabolite profiling of wound‐healing tissue extracts from contrasting cultivars. Phytochemistry, 159, 75–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies AMC and Blincow PJ, 1984. Glycoalkaloid content of potatoes and potato products sold in the UK. Journal of the Science of Food and Agriculture, 35, 553–557. [Google Scholar]
- Dean BB, 1994. Managing the potato production system. Food Products Press, New York. ISBN 1560220252. 56 pp. [Google Scholar]
- Deußer H, Guignard C, Hoffmann L and Evers D, 2012. Polyphenol and glycoalkaloid contents in potato cultivars grown in Luxembourg. Food Chemistry, 135, 2814–2824. [DOI] [PubMed] [Google Scholar]
- Diehl JF and Wedler A, 1977. Traditional versus “organic” farming. Comparative studies concerning the quality of the harvested products. Berichte der Bundesforschungsanstalt fuer Ernaehrung, 1, 144 pp. [Google Scholar]
- Distl M and Wink M, 2009. Identification and quantification of steroidal alkaloids from wild tuber‐bearing Solanum species by HPLC and LC‐ESI‐Ms. Potato Research, 52, 79–104. [Google Scholar]
- Dixit VP and Gupta RS, 1982. Antispermatogenic, antiandrogenic properties of solasodine (C27H43O2N) obtained from Solanum xanthocarpum berries on the male genital tract of dog (Cannis familiars): A histophysiological approach. International Journal of Andrology, 5, 295–307. [DOI] [PubMed] [Google Scholar]
- Dixit VP, Gupta RS and Gupta S, 1989. Antifertility plant products: testicular cell population dynamics following solasodine (C27H43O2N) administration in Rresus monkey (Macaca mulatta). Andrologia, 21, 542–546. [PubMed] [Google Scholar]
- Dolan LC, Matulka RA and Burdock GA, 2010. Naturally occurring food toxins. Toxins, 2, 2289–2332. 10.3390/toxins2092289 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dornow Food Technology , 2019. Applications ‐ steam peeling mechanical peeling. What peeling method is used where?. Available online: http://www.dornow.de/de/englisch/Q-Prospekte/Q104e5.pdf [Accessed: 4 February 2019].
- Driedger DR and Sporns P, 1999. Glycoalkaloid concentration in by‐products of potato starch extraction as measured by matrix‐assisted laser desorption/ionization mass spectrometry. Journal of Food Processing Preservation, 23, 377–390. [Google Scholar]
- Duysen EG, Stribley JA, Fry DL, Hinrichs SH and Lockridge O, 2002. Rescue of the acetylcholinesterase knockout mouse by feeding a liquid diet; phenotype of the adult acetylcholinesterase deficient mouse. Developmental Brain Research, 137, 43–54. [DOI] [PubMed] [Google Scholar]
- Eanes RC, Tek N, Kirsoy O, Frary A, Doganlar S and Almeida AE, 2008. Development of practical HPLC methods for the separation and determination of eggplant steroidal glycoalkaloids and their aglycones. Journal of Liquid Chromatography and Related Technologies, 31, 984–1000. [Google Scholar]
- Edwards EJ and Cobb AH, 1997. Effect of temperature on glycoalkaloid and chlorophyll accumulation in potatoes (Solanum tuberosum L cv King Edward) stored at low photon flux density, including preliminary modeling using an artificial neural network. Journal of Agricultural and Food Chemistry, 45, 1032–1038. [Google Scholar]
- Edwards EJ and Cobb AH, 1999. The effect of prior storage on the potential of potato tubers (Solanum tuberosum L) to accumulate glycoalkaloids and chlorophylls during light exposure, including artificial neural network modelling. Journal of the Science of Food and Agriculture, 79, 1289–1297. [Google Scholar]
- Edwards EJ, Saint RE and Cobb AH, 1998. Is there a link between greening and light‐enhanced glycoalkaloid accumulation in potato (Solanum tuberosum L) tubers? Journal of the Science of Food and Agriculture, 76, 327–333. [Google Scholar]
- EFSA (European Food Safety Authority), 2005. Opinion of the Scientific Committee on a request from EFSA related to a harmonised approach for risk assessment of substances which are both genotoxic and carcinogenic. EFSA Journal 2005;3(10):282, 33 pp. 10.2903/j.efsa.2005.282 [DOI] [Google Scholar]
- EFSA (European Food Safety Authority), 2007. Guidance of the Scientific Committee on a request from EFSA related to Uncertainties in Dietary Exposure Assessment. EFSA Journal 2007;4(12):438, 54 pp. 10.2903/j.efsa.2007.438 [DOI] [Google Scholar]
- EFSA (European Food Safety Authority), 2009. Guidance of the Scientific Committee on transparency in the scientific aspects of risk assessments carried out by EFSA. Part 2: general principles. EFSA Journal 2009;7(5):1051, 22 pp. 10.2903/j.efsa.2009.1051 [DOI] [Google Scholar]
- EFSA (European Food Safety Authority), 2010a. Standard sample description for food and feed. EFSA Journal 2010;8(1):1457, 54 pp. 10.2903/j.efsa.2010.1457 [DOI] [Google Scholar]
- EFSA (European Food Safety Authority), 2010b. Management of left‐censored data in dietary exposure assessment of chemical substances. EFSA Journal 2010;8(3):1557, 96 pp. 10.2903/j.efsa.2010.1557 [DOI] [Google Scholar]
- EFSA (European Food Safety Authority), 2011a. Use of the EFSA Comprehensive European Food Consumption Database in Intakes Assessment. EFSA Journal 2011;9(3):2097, 34 pp. 10.2903/j.efsa.2011.2097 [DOI] [Google Scholar]
- EFSA (European Food Safety Authority), 2011b. Report on the development of a food classification and description system for exposure assessment and guidance on its implementation and use. EFSA Journal 2011;9(12):2489. 84 pp. 10.2903/j.efsa.2011.2489 [DOI] [Google Scholar]
- EFSA (European Food Safety Authority), 2011c. EFSA Panel on Contaminants in the Food Chain (CONTAM); Scientific Opinion on the risks for animal and public health related to the presence of T‐2 and HT‐2 toxin in food and feed. EFSA Journal 2011;9(12):2481, 187 pp. https://doi:10.2903/j.efsa.2011.2481 [Google Scholar]
- EFSA (European Food Safety Authority), 2015. The food classification and description system FoodEx2 (revision 2). EFSA supporting publication 2015;EN‐804: 90 pp. 10.2903/sp.efsa.2015.EN-804 [DOI]
- EFSA (European Food Safety Authority), Dujardin B and Kirwan L, 2019. Technical report on the raw primary commodity (RPC) model: strengthening EFSA's capacity to assess dietary exposure at different levels of the food chain, from raw primary commodities to foods as consumed. EFSA supporting publication 2019;EN‐1532, 30 pp. 10.2903/sp.efsa.2019.EN-1532 [DOI]
- EFSA (European Food Safety Authority), 2020. Outcome of a public consultation on the draft risk assessment of glycoalkaloids in feed and food, in particular in potatoes and potato‐derived products. EFSA supporting publication 2020:EN‐1905, 63 pp. 10.2903/sp.efsa.2020.EN-1905 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA Scientific Committee , 2011. Scientific Opinion on genotoxicity testing strategies applicable to food and feed safety assessment. EFSA Journal 2011;9(9):2379, 69 pp. 10.2903/j.efsa.2011.2379 [DOI] [Google Scholar]
- EFSA Scientific Committee , 2012a. Guidance on selected default values to be used by the EFSA Scientific Committee, Scientific Panels and Units in the absence of actual measured data. EFSA Journal 2012;10(3):2579, 32 pp. 10.2903/j.efsa.2012.2579 [DOI] [Google Scholar]
- EFSA Scientific Committee , 2012b. Scientific Opinion on Risk Assessment Terminology. EFSA Journal 2012;10(5):2664, 43 pp. 10.2903/j.efsa.2012.2664 [DOI] [Google Scholar]
- EFSA Scientific Committee , Hardy A, Benford D, Halldorsson T, Jeger MJ, Knutsen KH, More S, Mortensen A, Naegeli H, Noteborn H, Ockleford C, Ricci A, Rychen G, Silano V, Solecki R, Turck D, Aerts M, Bodin L, Davis A, Edler L, Gundert‐Remy U, Sand S, Slob W, Bottex B, Abrahantes JC, Marques DC, Kass G and Schlatter JR, 2017a. Update: use of the benchmark dose approach in risk assessment. EFSA Journal 2017;15(1):4658, 41 pp. 10.2903/j.efsa.2017.4658 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA Scientific Committee , Hardy A, Benford D, Halldorsson T, Jeger MJ, Knutsen HK, More S, Naegeli H, Noteborn H, Ockleford C, Ricci A, Rychen G, Schlatter JR, Silano V,Solecki R, Turck D, Benfenati E, Chaudhry QM, Craig P, Frampton G, Greiner M, Hart A, Hogstrand C,Lambre C, Luttik R, Makowski D, Siani A, Wahlstroem H, Aguilera J, Dorne J‐L, Fernandez Dumont A,Hempen M, Valtueña Martínez S, Martino L, Smeraldi C, Terron A, Georgiadis N and Younes M, 2017b. Scientific Opinion on the guidance on the use of the weight of evidence approach in scientific assessments. EFSA Journal 2017;15(8):4971, 69 pp. 10.2903/j.efsa.2017.4971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA Scientific Committee , Hardy A, Benford D, Halldorsson T, Jeger M, Knutsen HK, More S, Naegeli H, Noteborn H, Ockleford C, Ricci A, Rychen G, Silano V, Solecki R, Turck D, Younes M, Aquilina G, Crebelli R, Gurtler R, Hirsch‐Ernst KI, Mosesso P, Nielsen E, van Benthem J, Carfi M, Georgiadis N, Maurici D, Parra Morte J and Schlatter J, 2017c. Scientific Opinion on the clarification of some aspects related to genotoxicity assessment. EFSA Journal 2017;15(12):5113, 25 pp. 10.2903/j.efsa.2017.5113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA Scientific Committee , Hardy A, Benford D, Halldorsson T, Jeger MJ, Knutsen HK, More S, Naegeli H, Noteborn H, Ockleford C, Ricci A, Rychen G, Schlatter JR, Silano V,Solecki R, Turck D, Younes M, Bresson J‐L, Griffin J, Hougaard Benekou S, van Loveren H, Luttik R,Messean A, Penninks A, Ru G, Stegeman JA, van der Werf W, Westendorf J, Woutersen RA, Barizzone F, Bottex B, Lanzoni A, Georgiadis N and Alexander J, 2017d. Guidance on the assessment of the biological relevance of data in scientific assessments. EFSA Journal 2017;15(8):4970, 73 pp. 10.2903/j.efsa.2017.4970 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA Scientific Committee , Benford D,Halldorsson T, Jeger MJ, Knutsen HK, More S, Naegeli H, Noteborn H, Ockleford C, Ricci A, Rychen G,Schlatter JR, Silano V, Solecki R, Turck D, Younes M, Craig P, Hart A, Von Goetz N, Koutsoumanis K,Mortensen A, Ossendorp B, Martino L, Merten C, Mosbach‐Schulz O and Hardy A, 2018. Guidance on Uncertainty Analysis in Scientific Assessments. EFSA Journal 2018;16(1):5123, 39 pp. 10.2903/j.efsa.2018.5123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eldridge AC and Hockridge ME, 1983. High‐performance liquid chromatographic separation of Eastern black nightshade (Solanum ptycanthum) glycoalkaloids. Journal of Agricultural and Food Chemistry, 31, 1218–1220. [Google Scholar]
- Elkahoui S, Bartley GE, Yokoyama WH and Friedman M, 2018. Dietary Supplementation of Potato Peel Powders Prepared from Conventional and Organic Russet and Non‐organic Gold and Red Potatoes Reduces Weight Gain in Mice on a High‐Fat Diet. Journal of Agricultural and Food Chemistry, 66, 6064–6072. [DOI] [PubMed] [Google Scholar]
- El‐Said SM, 2013. Removal of pharmacological undesirable compounds from potato tuber. Research and Reviews in BioSciences, 7, 129–135. [Google Scholar]
- Eltayeb EA and Roddick JG, 1984. Changes in the Alkaloid Content of Developing Fruits of Tomato (Lycopersicon esculentum Mill.): I. Analyses of cultivars and mutants with different ripening characteristics. Journal of Experimental Botany, 35, 252–260. [Google Scholar]
- Emmanuel S, Ignacimuthu S, Perumalsamy R and Amalraj T, 2006. Antiinflammatory activity of Solanum trilobatum . Fitoterapia, 77, 611–612. [DOI] [PubMed] [Google Scholar]
- Essers AJ, Alink GM, Speijers GJ, Alexander J, Bouwmeister PJ, van den Brandt PA, Ciere S, Gry J, Herrman J, Kuiper HA, Mortby E, Renwick AG, Shrimpton DH, Vainio H, Vittozzi L and Koeman JH, 1998. Food plant toxicants and safety Risk assessment and regulation of inherent toxicants in plant foods. Environmental Toxicology and Pharmacology, 5, 155–172. [DOI] [PubMed] [Google Scholar]
- Ewing WN, 2002. The Feeds Directory Branded Products Guide. Context, Ashby de la Zouch, UK. [Google Scholar]
- FAOSTAT (Food and Agriculture Organization of the United Nations), 2020a. FAOSTAT database on tomato production. Rome, Italy: FAO. Retrieved in 2020 from http://www.fao.org/faostat/en/#data
- FAOSTAT (Food and Agriculture Organization of the United Nations), 2020b. FAOSTAT database on potato production. Rome, Italy: FAO. Retrieved in 2020 from http://www.fao.org/faostat/en/#data
- FAOSTAT (Food and Agriculture Organization of the United Nations), 2020c. FAOSTAT database on aubergine production. Rome, Italy: FAO. Retrieved in 2020 from http://www.fao.org/faostat/en/#data
- Faridnia F, Burritt DJ, Bremer PJ and Oey I, 2015. Innovative approach to determine the effect of pulsed electric fields on the microstructure of whole potato tubers: Use of cell viability, microscopic images and ionic leakage measurements. Food Research International, 77, 556–564. [Google Scholar]
- Fiems LO, De Boever JL, Vanacker JM and De Brabander DL, 2013. Effect of cull potatoes in the diet for finishing Belgian Blue double‐muscled cows. Animal, 7, 93–100. [DOI] [PubMed] [Google Scholar]
- Finotti E, Bertone A and Vivanti V, 2006. Balance between nutrients and anti‐nutrients in nine Italian potato cultivars. Food Chemistry, 99, 698–701. [Google Scholar]
- Fitzpatrick TJ, McDermott JA and Osman SF, 1978. Evaluation of injured commercial potato samples for total glycoalkaloid content. Journal of Food Science, 43, 1617–1618. [Google Scholar]
- Fontaine TD, Irving GW, Poole RM and Dooluttle SP, 1948. Isolation and partial characterization of crystalline tomatine, an antibiotic agent from the tomato plant. Archives of Biochemistry, 18, 467–475. [PubMed] [Google Scholar]
- Friedman M, 1992. Composition and safety evaluation of potato berries, potato and tomato seeds, potatoes, and potato alkaloids. Food Safety Assessment, Chapter, 35, 429–462. [Google Scholar]
- Friedman M, 2002. Tomato glycoalkaloids: role in the plant and in the diet. Journal of Agricultural and Food Chemistry, 50, 5751–5780. [DOI] [PubMed] [Google Scholar]
- Friedman M, 2004. Analysis of biologically active compounds in potatoes (Solanum tuberosum), tomatoes (Lycopersicon esculentum), and jimson weed (Datura stramonium) seeds. Review. Journal of Chromatography A, 1054, 143–155. [DOI] [PubMed] [Google Scholar]
- Friedman M, 2006. Potato glycoalkaloids and metabolites: roles in the plant and in the diet. Journal of Agricultural and Food Chemistry, 54, 8655–8681. [DOI] [PubMed] [Google Scholar]
- Friedman M and Dao L, 1992. Distribution of glycoalkaloids in potato plants and commercial potato products. Journal of Agricultural and Food Chemistry, 40, 419–423. [Google Scholar]
- Friedman M and Henika PR, 1992. Absence of genotoxicity of potato alkaloids alpha‐chaconine, alpha‐solanine and solanidine in the Ames Salmonella and adult and foetal erythrocyte micronucleus assays. Food and Chemical Toxicology, 30, 689–694. [DOI] [PubMed] [Google Scholar]
- Friedman M and Levin CE, 1995. Tomatine Content in Tomato and Tomato Products Determined by HPLC with Pulsed Amperometric Detection. Journal of Agricultural and Food Chemistry, 1995(43), 1507–1511. [Google Scholar]
- Friedman M and Levin CE, 1998. Dehydrotomatine content in tomatoes. Journal of Agricultural and Food Chemistry, 46, 4571–4576. [Google Scholar]
- Friedman M and McDonald GM, 1995. Acid‐catalyzed partial hydrolysis of carbohydrate groups of the potato glycoalkaloid a‐chaconine in alcoholic solutions. Journal of Agricultural and Food Chemistry, 43, 1501–1506. [Google Scholar]
- Friedman M, Levin CE and McDonald GM, 1994. a‐Tomatine determination in tomatoes by HPLC using pulsed amperometric detection. Journal of Agricultural and Food Chemistry, 42, 1959–1964. [Google Scholar]
- Friedman M, Henika PR and Mackey BE, 1996. Feeding of potato, tomato and eggplant alkaloids affects food consumption and body and liver weights in mice. Journal of Nutrition, 126, 989–999. [DOI] [PubMed] [Google Scholar]
- Friedman M, McDonald GM and Filadelfi‐Kesyi M, 1997. Potato glycoalkaloids: chemistry, analysis, safety, and plant physiology. Critical Reviews in Plant Sciences, 16, 55–132. [Google Scholar]
- Friedman M, Bautista FF, Stanker LH and Larkin KA, 1998. Analysis of potato glycoalkaloids by a new ELISA Kit. Journal of Agricultural and Food Chemistry, 46, 5097–5102. [Google Scholar]
- Friedman M, Fitch TE and Yokoyama WE, 2000a. Lowering of plasma LDL cholesterol in hamsters by the tomato glycoalkaloid tomatine. Food and Chemical Toxicology, 38, 549–553. [DOI] [PubMed] [Google Scholar]
- Friedman M, Fitch TE, Levin CE and Yokoyama WH, 2000b. Feeding Tomatoes to Hamsters Reduces their Plasma Low‐density Lipoprotein Cholesterol and Triglycerides. Journal of Food Science, 65, 897–900. [Google Scholar]
- Friedman M, Henika PR and Mackey BE, 2003a. Effect of feeding solanidine, solasodine and tomatidine to non‐pregnant and pregnant mice. Food and Chemical Toxicology, 41, 61–71. [DOI] [PubMed] [Google Scholar]
- Friedman M, Roitman JN and Kozukue N, 2003b. Glycoalkaloid and calystegine contents of eight potato cultivars. Journal of Agricultural and Food Chemistry, 51, 2964–2973. [DOI] [PubMed] [Google Scholar]
- Friedman M, Kozukue N, Kim H‐J, Choi S‐H and Mizuno M, 2017. Glycoalkaloid, phenolic, and flavonoid content and antioxidative activities of conventional nonorganic and organic potato peel powders from commercial gold, red, and Russet potatoes. Journal of Food Composition and Analysis, 62, 69–75. [Google Scholar]
- Fujiwara Y, Takaki A, Uehara Y, Ikeda T, Okawa M, Yamauchi K, Ono M, Yoshimitsu H and Nohara T, 2004. Tomato steroidal alkaloid glycosides, esculeosides A and B, from ripe fruits. Tetrahedron, 60, 4915–4920. [Google Scholar]
- Furui H, Inakuma T, Ishiguro Y and Kiso M, 1997. Absorptiometric measurement of tomatine in tomatoes. Nippon Nogei Kagaku Kaishi, 71, 777–782. [Google Scholar]
- Gaffield W and Keeler RF, 1993. Implication of C‐5, C‐6 unsaturation as a key structural factor in steroidal alkaloid‐induced mammalian teratogenesis. Experientia, 49, 922–924. [DOI] [PubMed] [Google Scholar]
- Gaffield W and Keeler RF, 1996a. Induction of terata in hamsters by solanidane alkaloids derived from Solanum tuberosum . Chemical Research in Toxicology, 9, 426–433. [DOI] [PubMed] [Google Scholar]
- Gaffield W and Keeler RF, 1996b. Craniofacial malformations induced in hamsters by steroidal alkaloids. Journal of Natural Toxins, 5, 25–38. [Google Scholar]
- Gattermann R, Fritzsche P, Weinandy R and Neumann K, 2002. Comparative studies of body mass, body measurements and organ weights of wild derived and laboratory golden hamsters (Mesocricetus auratus). Laboratory Animals, 36, 445–454. [DOI] [PubMed] [Google Scholar]
- Gee JM, Wortley GM, Johnson IT, Price KR, Rutten AA, Houben GF and Penninks AH, 1996. Effects of saponins and glycoalkaloids on the permeability and viability of mammalian intestinal cells and on the integrity of tissue preparations in vitro. Toxicology In Vitro, 10, 117–128. [DOI] [PubMed] [Google Scholar]
- Ginzberg I, Tokuhisa JG and Veilleux RE, 2009. Potato Steroidal Glycoalkaloids: Biosynthesis and Genetic Manipulation. Potato Research, 52, 1–15. [Google Scholar]
- Glynn A, Igra AM, Sand S, Ilbäck NG, Hellenäs KE, Rósen J and Aspenström‐Fagerlund B, 2017. Are additive effects of dietary surfactants on intestinal tight junction integrity an overlooked human health risk? ‐ A mixture study on Caco‐2 monolayers. Food and Chemical Toxicology, 106, 314–323. [DOI] [PubMed] [Google Scholar]
- Gonmori K, Meguro H, Lu YQi, Yoshioka N, Hori K and Kikkawa S, 1993. The risk of solanine poisoning in a folk remedy and solanine production in potato. Research and Practice in Forensic Medicine, 36, 5. Only abstract in English. [Google Scholar]
- Gosselin B and Mondy NI, 1989. Effect of packaging materials on the chemical composition of potatoes. Journal of Food Science, 54, 629–631. [Google Scholar]
- Gosselin B, Mondy NI and Evans WD, 1988. Effect of method of irrigation on the total glycoalkaloid and nitrate‐nitrogen content of Rosa potatoes. American Potato Journal, 65, 99–103. [Google Scholar]
- Grandison L, Gelato M and Meites J, 1974. Inhibition of prolactin secretion by cholinergic drugs. Proceedings of the Society for Experimental Biology and Medicine, 145, 1236–1239. [DOI] [PubMed] [Google Scholar]
- Griebel C, 1923. Solaninreiche gesundheitsschädliche Kartoffeln. Zeitschrift fuer Untersuchung der Nahrungs‐ und Genussmittel sowie der Gebrauchsgegenstaende, 45, 175–183. [Google Scholar]
- Griffiths DW, Bain H and Dale FB, 1998. Effect of Storage Temperature on Potato (Solanum tuberosum L.) Tuber Glycoalkaloid Content and the Subsequent Accumulation of Glycoalkaloids and Chlorophyll in Response to Light Exposure. Journal of Agricultural and Food chemistry, 46, 5262–5268. [Google Scholar]
- Groen K, Pereboom‐de Fauw DP, Besamusca P, Beekhof PK, Speijers GJ and Derks HJ, 1993. Bioavailability and disposition of 3H‐solanine in rat and hamster. Xenobiotica, 23, 995–1005. [DOI] [PubMed] [Google Scholar]
- Grunenfelder LA, Knowles LO, Hiller LK and Knowles NR, 2006. Glycoalkaloid development during greening of fresh market potatoes (Solanum tuberosum L.). Journal of Agricultural and Food Chemistry, 54, 5847–5854. [DOI] [PubMed] [Google Scholar]
- Gull DD, 1960. Chlorophyll and solanine changes in tubers of Solanum Tuberosum induced by fluorescent light, and a study of solanine toxicology by bioassay technique. PhD, Cornell University, Michigan, 149, 11–139. [Google Scholar]
- Gunning OV, 1950. Suspected potato poisoning in a mare. British Veterinary Journal, 106, 32–33. [Google Scholar]
- Gupta RS and Dixit VP, 2002. Effects of short term treatment of solasodine on cauda epididymis in dogs. Indian Journal of Experimental Biology, 40, 169–173. [PubMed] [Google Scholar]
- Ha M, Kwak JH, Kim Y and Zee OP, 2012. Direct analysis for the distribution of toxic glycoalkaloids in potato tuber tissue using matrix‐assisted laser desorption/ionization mass spectrometric imaging. Food Chemistry, 133, 1155–1162. [Google Scholar]
- Haase NU, 2010. Glycoalkaloid concentration in potato tubers related to storage and consumer offering. Potato Research, 53, 297–307. [Google Scholar]
- Hajšlová J, Schulzová V, Slanina P, Janné K, Hellenäs KE and Andersson CE, 2005. Quality of organically and conventionally grown potatoes: Four‐year study of micronutrients, metals, secondary metabolites, enzymic browning and organoleptic properties. Food Additives and Contaminants, 22, 514–534. [DOI] [PubMed] [Google Scholar]
- Hamouz K, Lachman J, Dvořák P and Pivec V, 2005. The effect of ecological growing on the potatoes yield and quality. Plant, Soil and Environment, 51, 397–402. [Google Scholar]
- Hamouz K, Pazderu K, Lachman J, Orsak M, Pivec V, Hejtmankova K, Tomasek J and Cizek M, 2014. Effect of cultivar, flesh colour, location and year of cultivation on the glycoalkaloid content in potato tubers. Plant Soil and Environment, 60, 512–517. [Google Scholar]
- Hansen AA, 1925. Two fatal cases of potato poisoning. Science, 61, 340–341. [DOI] [PubMed] [Google Scholar]
- Harris FW and Cockburn T, 1918. Alleged poisoning by potatoes. Analyst, 43, 133–137. [Google Scholar]
- Harris H and Whittaker M, 1959. Differential response of human serum cholinesterase types to an inhibitor in potato. Nature, 183, 1808–1809. [DOI] [PubMed] [Google Scholar]
- Harris H and Whittaker M, 1962. Differential inhibition of the serum cholinesterase phenotypes by solanine and solanidine. Annals of Human Genetics, 26, 73–76. [DOI] [PubMed] [Google Scholar]
- Harvey MH, McMillan M, Morgan MR and Chan HW, 1985a. Solanidine is present in sera of healthy individuals in amounts dependent on their dietary potato consumption. Human Toxicology, 4, 187–194. [DOI] [PubMed] [Google Scholar]
- Harvey MH, Morris BA, McMillan M and Marks V, 1985b. Measurement of potato steroidal alkaloids in human serum and saliva by radioimmunoassay. Human Toxicology, 4, 503–512. [DOI] [PubMed] [Google Scholar]
- Harvey MH, Morris BA, McMillan M and Marks V, 1986. Potato steroidal alkaloids and neural tube defects: serum concentrations fail to demonstrate a causal relation. Human Toxicology, 5, 249–253. [DOI] [PubMed] [Google Scholar]
- Hellenäs KE, 1986. A simplified procedure for quantification of potato glycoalkaloids in tuber extracts by HPLC; comparison with ELISA and a colorimetric method. Journal of the Science of Food and Agriculture, 37, 776–782. [Google Scholar]
- Hellenäs KE, Nyman A, Slanina P, Lööf L and Gabrielsson J, 1992. Determination of potato glycoalkaloids and their aglycone in blood serum by high‐performance liquid chromatography. Application to pharmacokinetic studies in humans. Journal of Chromatography, 573, 69–78. [DOI] [PubMed] [Google Scholar]
- Hellenäs KE, Branzell C, Johnsson H and Slanina P, 1995a. Glycoalkaloid content of early potato varieties. Journal of the Science of Food and Agriculture, 67, 125–128. [Google Scholar]
- Hellenäs K‐E, Branzell C, Johnsson H and Slanina P, 1995b. High levels of glycoalkaloids in the established Swedish potato variety Magnum Bonum. Journal of the Science of Food and Agriculture, 68, 249–255. [Google Scholar]
- Heppner KM, Tong J, Kirchner H, Nass R and Tschöp MH, 2011. The ghrelin O‐acyltransferase‐ghrelin system: a novel regulator of glucose metabolism. Current Opinion in Endocrinology, Diabetes and Obesity, 18, 50–55. [DOI] [PubMed] [Google Scholar]
- Herb SF, Fitzpatrick TJ and Osman SF, 1975. Separation of potato glycoalkaloids by gas chromatography. Journal of Agricultural and Food Chemistry, 23, 520–523. [DOI] [PubMed] [Google Scholar]
- Hlywka JJ, Stephenson GR, Sears MK and Yada RY, 1994. Effects of insect damage on glycoalkaloid content in potatoes (Solanum tuberosum). Journal of Agricultural and Food Chemistry, 42, 2545–2550. [Google Scholar]
- Hopkins J, 1995. The glycoalkaloids: naturally of interest (but a hot potato?). Food and Chemical Toxicology, 33, 323–328. [DOI] [PubMed] [Google Scholar]
- Hossain MB, Rai DK and Brunton NP, 2015. Optimisation and validation of ultra‐high performance liquid chromatographic‐tandem mass spectrometry method for qualitative and quantitative analysis of potato steroidal alkaloids. Journal of Chromatography B, 997, 110–115. [DOI] [PubMed] [Google Scholar]
- Hossain MB, Brunton NP and Rai DK, 2016. Effect of drying methods on the steroidal alkaloid content of potato peels, shoots and berries. Molecules, 21, 403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Houben RJ and Brunt K, 1994. Determination of glycoalkaloids in potato tubers by reversed‐phase high‐performance liquid chromatography. Journal of Chromatography A, 661, 169–174. [Google Scholar]
- Hugenholtz F and de Vos WM, 2018. Mouse models for human intestinal microbiota research: a critical evaluation. Cellular and Molecular Life Sciences, 75, 149–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iablokov V, Sydora BC, Foshaug R, Meddings J, Driedger D, Churchill T and Fedorak RN, 2010. Naturally occurring glycoalkaloids in potatoes aggravate intestinal inflammation in two mouse models of inflammatory bowel disease. Digestive Diseases and Sciences, 55, 3078–3085. [DOI] [PubMed] [Google Scholar]
- Ieri F, Innocenti M, Andrenelli L, Vecchio V and Mulinacci N, 2011. Rapid HPLC/DAD/MS method to determine phenolic acids, glycoalkaloids and anthocyanins in pigmented potatoes (Solanum tuberosum L.) and correlations with variety and geographical origin. Food Chemistry, 125, 750–759. [Google Scholar]
- Iijima Y, Fujiwara Y, Tokita T, Ikeda T, Nohara T, Aoki K and Shibata D, 2009. Involvement of ethylene in the accumulation of esculeoside a during fruit ripening of tomato (Solanum lycopersicum). Journal of Agricultural and Food Chemistry, 57, 3247–3252. [DOI] [PubMed] [Google Scholar]
- Iijima Y, Watanabe B, Sasaki R, Takenaka M, Ono H, Sakurai N, Umemoto N, Suzuki H, Shibata D and Aoki K, 2013. Steroidal glycoalkaloid profiling and structures of glycoalkaloids in wild tomato fruit. Phytochemistry, 95, 145–157. [DOI] [PubMed] [Google Scholar]
- Jandrić Z, Rathor MN, Švarc‐Gajić J, Maestroni BM, Sasanya JJ, Djurica R and Cannavan A, 2011. Development of a liquid chromatography‐tandem mass spectrometric method for the simultaneous determination of tropane alkaloids and glycoalkaloids in crops. Food Additives and Contaminants, 28, 1205–1219. [DOI] [PubMed] [Google Scholar]
- JECFA (Joint FAO/WHO Expert Committee on Food Additives), 1992. Evaluation of certain food additives and naturally occurring toxicants. Thirty‐ninth report of the Joint FAO/WHO Expert Committee on Food Additives. World Health Organisation Technical Report Series, 828, 1‐49. [PubMed]
- JECFA (Joint FAO/WHO Expert Committee on Food Additives), 1993. Toxicological evaluation of certain food additives and naturally occurring toxicants. WHO Food Additives Series, Vol. 30. World Health Organization, Geneva.
- Jin CY, Liu H, Xu D, Zeng FK, Zhao YC, Zhang H and Liu G, 2018. Glycoalkaloids and phenolic compounds in three commercial potato cultivars grown in Hebei, China. Food Science and Human Wellness, 7, 156–162. [Google Scholar]
- Jones PG and Fenwick GR, 1981. The glycoalkaloid content of some edible solanaceous fruits and potato products. Journal of the Science of Food and Agriculture, 32, 419–421. [DOI] [PubMed] [Google Scholar]
- Karaway MA, 2014. Removal of pharmacological undesirable compounds from potato tuber. Research and Reviews in BioSciences, 8, 73–77. [Google Scholar]
- Karmakar K, Islam A, Chhanda SA, Tuhin TI, Muslim T and Rahman A, 2015. Secondary metabolites from the fruits of Solanum torvum SW. Journal of Pharmacognosy and Phytochemistry, 4, 160–163. [Google Scholar]
- Keeler RF, Young S and Brown D, 1976. Spina bifida, exencephaly, and cranial bleb produced in hamsters by the solanum alkaloid solasodine. Research Communications in Chemical Pathology and Pharmacology, 13, 723–730. [PubMed] [Google Scholar]
- Keeler RF, Baker DC and Gaffield W, 1990. Spirosolane‐containing Solanum species and induction of congenital craniofacial malformations. Toxicon, 28, 873–884. [DOI] [PubMed] [Google Scholar]
- Kees M, Beckel N and Sharp C, 2015. Successful treatment of Solanum dulcamara intoxication in a Labrador retriever puppy. Canadian Veterinary Journal, 56, 1283–1286. [PMC free article] [PubMed] [Google Scholar]
- Kerr CA, Goodband RD, Smith II JW, Musser RE, Bergstro¨m JR, Nessmith WG Jr, Tokach MD and Nelssen JL, 1998. Evaluation of Potato Proteins on the Growth Performance of Early‐Weaned Pigs. Journal of Animal Science, 76, 3024–3033. [DOI] [PubMed] [Google Scholar]
- Keukens EAJ, Vrije Td, Fabrie CHJP, Demel t’ RA, Jongen WMF and Kruijff Bd, 1992. Dual specificity of sterol‐mediated glycoalkaloid induced membrane disruption. Biochunica et Biophysica Acta, 1110, 127–136. [DOI] [PubMed] [Google Scholar]
- Keukens EAJ, Hop MECM and Jongen WMF, 1994. Rapid high‐performance liquid chromatographic method for the quantification of a‐tomatine in tomato. Journal of Agricultural and Food Chemistry, 42, 2475–2477. [Google Scholar]
- Keukens EAJ, deVrije T, vandenBoom C, deWaard P, Plasman HH, Thiel F, Chupin V, Jongen WMF and deKruijff B, 1995. Molecular basis of glycoalkaloid induced membrane disruption. Biochimica et Biophysica Acta, 1240, 216–228. [DOI] [PubMed] [Google Scholar]
- Keukens EAJ, deVrije T, Jansen LAM, deBoer H, Janssen M, deKroon AIPM, Jongen WMF and deKruijff B, 1996. Glycoalkaloids selectively permeabilize cholesterol containing biomembranes. Biochimica et Biophysica Acta, 1279, 243–250. [DOI] [PubMed] [Google Scholar]
- Khaserao S and Somani R, 2017. Evaluation of anti‐obesity activity of solasodine in high fat diet‐induced obesity in rat. International Journal of Pharmacy and Pharmaceutical Sciences, 9, 23–29. [Google Scholar]
- Khodayari S, Alizadeh AM, Kouhpayeh SA, Mohsenikia M, Karimi AA, Khodayari H, Abdolahi AR, Sarbiyaii A, Erfani S, Kazemi S and Zakeri H, 2013. The acute and chronic toxicity effects of alpha‐solanine in mice. Journal of Babol University of Medical Sciences, 15, 24–31. [Google Scholar]
- Kibler R, Lang H and Ziegler W, 1985. Einfluß küchentechnischer Maßnahmen auf den Solaningehalt grüner Tomatenfrüchte. Deutsche Lebensmittel‐Rundschau, 81, 111–113. [Google Scholar]
- King RR and McQueen RE, 1981. Transformations of potato glycoalkaloids by rumen microorganisms. Journal of Agricultural and Food Chemistry, 29, 1101–1103. [DOI] [PubMed] [Google Scholar]
- Kline BE, von Elbe H, Dahle NA and Morris Kupchan S, 1961. Toxic effects of potao sprouts and of solanine fed to pregnant rats 107 pp. [DOI] [PubMed] [Google Scholar]
- Knapp S, 2013. Typification of Solanum species (Solanaceae) described by Casamiro Gómez Ortega. Anales del Jardín Botánico de Madrid, 70, 56–61. [Google Scholar]
- Knuthsen P, Jensen U, Schmidt B and Larsen IK, 2009. Glycoalkaloids in potatoes: content of glycoalkaloids in potatoes for consumption. Journal of Food Composition and Analysis, 22, 577–581. [Google Scholar]
- Koh E, Kaffka S and Mitchell AE, 2013. A long‐term comparison of the influence of organic and conventional crop management practices on the content of the glycoalkaloid alpha‐tomatine in tomatoes. Journal of the Science of Food Agriculture, 93, 1537–1542. [DOI] [PubMed] [Google Scholar]
- Koleva II, van Beek TA, Soffers AEMF, Dusemund B and Rietjens IMCM, 2012. Alkaloids in the human food chain ‐ Natural occurrence and possible adverse effects. Molecular Nutrition and Food Research, 56, 30–52. [DOI] [PubMed] [Google Scholar]
- König H and Staffe A, 1953. Beitrag zur Wirkung des Solanins auf Blutbild und ‐Katalase beim Schaf (Action of solanine upon blood picture and blood catalase of sheep). Deutsche Tierärztliche Wochenschrift, 60, 150–153. [Google Scholar]
- Kozukue N and Friedman M, 2003. Tomatine, chlorophyll, β‐carotene and lycopene content in tomatoes during growth and maturation. Journal of the Science of Food and Agriculture, 83, 195–200. [Google Scholar]
- Kozukue N and Mizuno S, 1990. Effects of light exposure and storage temperature on greening and glycoalkaloid content in potato tubers. Journal of the Japanese Society for Horticultural Science, 59, 673–677. [Google Scholar]
- Kozukue N, Kozukue E and Mizuno S, 1987. Glycoalkaloids in potato plants and tubers. HortScience, 22, 294–296. [Google Scholar]
- Kozukue N, Tsuchida H and Friedman M, 2001. Tracer Studies on the Incorporation of [2‐14C]‐DL‐Mevalonate into Chlorophylls a and b, a‐chaconine, and a‐Solanine of Potato Sprouts. Journal of Agricultural and Food Chemistry, 49, 92–97. [DOI] [PubMed] [Google Scholar]
- Kozukue N, Han JS, Lee KR and Friedman M, 2004. Dehydrotomatine and alpha‐tomatine content in tomato fruits and vegetative plant tissues. Journal of Agricultural and Food Chemistry, 52, 2079–2083. [DOI] [PubMed] [Google Scholar]
- Kuhn R and Löw I, 1954. Die Konstitution des Solanins. Angewandte Chemie, 66, 639–640. [Google Scholar]
- Kumar D and Ezekiel R, 2009. Changes in glycoalkaloids and phenolic contents in potato tubers stored under different conditions. Journal of Food Science and Technology, 46, 480–483. [Google Scholar]
- Kyzlink V, Míková K and Jelínek R, 1981. Tomatine, solanine and embryotoxicity of unripe tomatoes. Sbornik Vysoké Školy Chemicko‐Technologické v Praze, Potraviny, E51, 69–83. [Google Scholar]
- Lachman J, Hamouz K, Musilová J, Hejtmánková K, Kotíková Z, Pazderů K, Domkářová J, Pivec V and Cimr J, 2013. Effect of peeling and three cooking methods on the content of selected phytochemicals in potato tubers with various colour of flesh. Food Chemistry, 138, 1189–1197. [DOI] [PubMed] [Google Scholar]
- Lampitt LH, Bushill JH, Rooke HS and Jackson EM, 1943. Solanine, glycoside of the potato. II. Distribution in the potato plant. Journal of the Society of Chemical Industry, London, 62, 48–51. [Google Scholar]
- Langkilde S, Schroder M, Stewart D, Meyer O, Conner S, Davies H and Poulsen M, 2008. Acute toxicity of high doses of the glycoalkaloids, alpha‐solanine and alpha‐chaconine, in the Syrian Golden hamster. Journal of Agricultural and Food Chemistry, 56, 8753–8760. [DOI] [PubMed] [Google Scholar]
- Langkilde S, Mandimika T, Schroder M, Meyer O, Slob W, Peijnenburg A and Poulsen M, 2009. A 28‐day repeat dose toxicity study of steroidal glycoalkaloids, alpha‐solanine and alpha‐chaconine in the Syrian Golden hamster. Food and Chemical Toxicology, 47, 1099–1108. [DOI] [PubMed] [Google Scholar]
- Langkilde S, Schroder M, Frank T, Shepherd LVT, Conner S, Davies HV, Meyer O, Danier J, Rychlik M, Belknap WR, McCue KF, Engel K‐H, Stewart D, Knudsen I and Poulsen M, 2012. Compositional and toxicological analysis of a GM potato line with reduced alpha‐solanine content ‐ a 90‐day feeding study in the Syrian Golden hamster. Regulatory Toxicology and Pharmacology, 64, 177–185. [DOI] [PubMed] [Google Scholar]
- Layer PG, Klaczinski J, Salfelder A, Sperling LE, Thangaraj G, Tuschl C and Vogel‐Höpker A, 2013. Cholinesterases in development: AChE as a firewall to inhibit cell proliferation and support differentiation. Chemico‐Biological Interactions, 203, 269–276. [DOI] [PubMed] [Google Scholar]
- Lee MR, 2006. The Solanaceae: foods and poisons. The journal of the Royal College of Physicians of Edinburgh, 36, 162–169. [PubMed] [Google Scholar]
- Lee C and De Luca C, 2019. Potato propagation North Dacota State University. Plant Propagation PLSC 368. Available online: https://www.ndsu.edu/pubweb/chiwonlee/plsc368/student/papers06/carina%20de%20luca/carinadeluca.htm [Accessed: January 2019].
- Lee KG, Lee SG, Lee HH, Lee HJ, Shin JS, Kim NJ, An HJ, Nam JH, Jang DS and Lee KT, 2015. α‐Chaconine isolated from a Solanum tuberosum L. cv Jayoung suppresses lipopolysaccharide‐induced pro‐inflammatory mediators via AP‐1 inactivation in RAW 264.7 macrophages and protects mice from endotoxin shock. Chemico‐Biological Interactions, 235, 85–94. [DOI] [PubMed] [Google Scholar]
- Lelario F, De Maria S, Rivelli AR, Russo D, Milella L, Bufo SA and Scrano L, 2019. A Complete Survey of Glycoalkaloids Using LC‐FTICR‐MS and IRMPD in a Commercial Variety and a Local Landrace of Eggplant (Solanum melongena L.) and their Anticholinesterase and Antioxidant Activities. Toxins, 11, 230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leonardi C, Ambrosino P, Esposito F and Fogliano V, 2000. Antioxidative activity and carotenoid and tomatine contents in different [typologies of fresh consumption tomatoes.]. Journal of Agricultural and Food Chemistry, 48, 4723–4727. [DOI] [PubMed] [Google Scholar]
- Lepper W, 1949. Solanine content of 58 varieties of potatoes. Zeitschrift für Lebensmittel‐Untersuchung und ‐Forschung, 89, 264–273. [Google Scholar]
- Li H‐J, Jiang Y and Li P, 2006. Chemistry, bioactivity and geographical diversity of steroidal alkaloids from the Liliaceae family. Natural Products Reports, 23, 735–752. [DOI] [PubMed] [Google Scholar]
- Liebenow H, 1970. Solanum nigrum as a nitrate‐containing plant and the determination of its alkaloid content. Wissenschaftliche Zeitschrift der Humboldt‐Universität zu Berlin, Mathematisch‐Naturwissenschaftliche Reihe, 19, 59–71. [Google Scholar]
- Liljemark A and Widoff E, 1960. Greening and solanine development of white potato in fluorescent light. American Potato Journal, 37, 379–389. [Google Scholar]
- Lionetto MG, Caricato R, Calisi A, Giordano ME and Schettino T, 2013. Acetylcholinesterase as a Biomarker in Environmental and Occupational Medicine: New Insights and Future Perspectives. Biomedical Research. International, 321213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu W, Zhang N, Li B, Fan S, Zhao R, Li Li‐P WuG‐H and Zhao Y, 2014. Determination of α‐chaconine and α‐solanine in commercial potato crisps by QuEChERS extraction and UPLC‐MS/MS. Chemical Papers, 68, 1498–1504. [Google Scholar]
- Lo Scalzo R, Fibinani M, Francese G, D′Alessandro A, Rotino GL, Conte P and Mennella G, 2016. Cooking influence on physicochemical fruit characteristics of eggplant (Solanum melongena L.). Food Chemistry, 194, 835–842. [DOI] [PubMed] [Google Scholar]
- Loewenstein‐Lichtenstein Y, Glick D, Gluzman N, Sternfeld M, Zakut H and Soreq H, 1996. Overlapping drug interaction sites of human butyrylcholinesterase dissected by site‐directed mutagenesis. Molecular Pharmacology, 50, 1423–1431. [PubMed] [Google Scholar]
- Love SL, Herrman TJ, Thompsonjohns A and Baker TP, 1994. Effect and interaction of crop management factors on the glycoalkaloid concentration of potato tubers. Potato Research, 37, 77–85. [Google Scholar]
- Machado RMD, Toledo MCF and García LC, 2006. Effect of light and temperature on the formation of glycoalkaloids in potato tubers. Food Control, 18, 503–508. [Google Scholar]
- Machida‐Hirano R, 2015. Diversity of potato genetic resources. Breeding Science, 65, 26–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mäder J, Rawel H and Kroh LW, 2009. Composition of phenolic compounds and glycoalkaloids α‐ solanine and α‐chaconine during commercial potato processing. Journal of Agricultural and Food Chemistry, 57, 6292–6297. [DOI] [PubMed] [Google Scholar]
- Majamaa H, Seppala U, Palosuo T, Turjanmaa K, Kalkkinen N and Reunala T, 2001. Positive skin and oral challenge responses to potato and occurrence of immunoglobulin E antibodies to patatin (Sol t 1) in infants with atopic dermatitis. Pediatric Allergy and Immunology, 12, 283–288. [DOI] [PubMed] [Google Scholar]
- Manrique‐Moreno M, Londoño‐Londoño J, Jemioła‐Rzemińska M, Strzałka K, Villena F, Avello M and Suwalsky M, 2014. Structural effects of the Solanum steroids solasodine, diosgenin and solanine on human erythrocytes and molecular models of eukaryotic membranes. Biochimica et Biophysica Acta, 1838, 266–277. [DOI] [PubMed] [Google Scholar]
- Martínez‐Valverde I, Periago MJ, Provan G and Chesson A, 2002. Phenolic compounds, lycopene and antioxidant activity in commercial varieties of tomato (Lycopersicum esculentum). Journal of the Science of Food and Agriculture, 82, 323–330. [Google Scholar]
- Matthew JA, Morgan MRA, McNerney R, Chan HW‐S and Coxon DT, 1983. Determination of solanidine in human plasma by radioimmunoassay. Food and Chemical Toxicology, 21, 637–640. [DOI] [PubMed] [Google Scholar]
- Maurya A, Manika N, Verma RK, Singh SC and Srivastava SK, 2013. Simple and Reliable Methods for the Determination of Three Steroidal Glycosides in the Eight Species of Solanum by Reversed‐phase HPLC Coupled with Diode Array Detection. Phytochemical Analysis, 24, 87–92. [DOI] [PubMed] [Google Scholar]
- McGehee DS, Krasowski MD, Fung DL, Wilson B, Gronert GA and Moss J, 2000. Cholinesterase inhibition by potato glycoalkaloids slows mivacurium metabolism. Anesthesiology, 93, 510–519. [DOI] [PubMed] [Google Scholar]
- McMillan M and Thompson JC, 1979. An outbreak of suspected solanine poisoning in schoolboys: Examinations of criteria of solanine poisoning. The Quarterly Journal of Medicine, 48, 227–243. [PubMed] [Google Scholar]
- Mennella G, Lo Scalzo R, Fibiani M, D'Alessandro A, Francese G, Toppino L, Acciarri N, de Almeida AE and Rotino GL, 2012. Chemical and bioactive quality traits during fruit ripening in eggplant (S. melongena L.) and allied species. Journal of Agricultural and Food Chemistry, 60, 11821–11831. [DOI] [PubMed] [Google Scholar]
- Mensinga TT, Sips AJAM, Rompelberg CJM, van Twillert K, Meulenbelt J, van den Top HJ and van Egmond HP, 2005. Potato glycoalkaloids and adverse effects in humans: an ascending dose study. Regulatory Toxicology and Pharmacology, 41, 66–72. [DOI] [PubMed] [Google Scholar]
- Mesulam MM, Guillozet A, Shaw P, Levey A, Duysen EG and Lockridge O, 2002. Acetylcholinesterase knockouts establish central cholinergic pathways and can use butyrylcholinesterase to hydrolyze acetylcholine. Neuroscience, 110, 627–639. [DOI] [PubMed] [Google Scholar]
- Miceli A, Sabatino L, Moncada A, Vetrano F and D'Anna F, 2014. Nursery and field evaluation of eggplant grafted onto unrooted cuttings of Solanum torvum Sw. Scientia Horticulturae, 178, 203–210. [Google Scholar]
- Michalska L, Nagel G, Swiniarski E and Zydowo MM, 1985. The effect of alpha‐solanine on the active calcium transport in rat intestine. General Pharmacology, 16, 69–70. [DOI] [PubMed] [Google Scholar]
- Milligan JB, 1941. Suspected potato poisoning in cattle. Veterinary Record, 53, 512. [Google Scholar]
- Milner SE, Brunton NP, Jones PW, O'Brien NM, Collins SG and Maguire AR, 2011. Bioactivities of glycoalkaloids and their aglycones from Solanum species. Journal of Agricultural and Food Chemistry, 59, 3454–3484. [DOI] [PubMed] [Google Scholar]
- Moco S, Capanoglu E, Tikunov Y, Bino RJ, Boyacioglu D, Hall RD, Vervoort J and de Vos RCH, 2007. Tissue specialization at the metabolite level is perceived during the development of tomato fruit. Journal of Experimental Botany, 58, 4131–4146. [DOI] [PubMed] [Google Scholar]
- Mohsenikia M, Alizadeh AM, Khodayari S, Khodayari H, kouhpayeh SA, Karimi A, Zamani M, Azizian S and Mohagheghi MA, 2013. The protective and therapeutic effects of alpha‐solanine on mice breast cancer. European Journal of Pharmacology, 718, 1–9. [DOI] [PubMed] [Google Scholar]
- Mondy NI and Gosselin B, 1988. Effect of peeling on total phenols, total glycoalkaloids, discoloration and flavor of cooked potatoes. Journal of Food Science, 53, 756–759. [Google Scholar]
- Mondy NI and Munshi CB, 1990. Effect of nitrogen fertilization on glycoalkaloid and nitrate content of potatoes. Journal of Agricultural and Food Chemistry, 38, 565–567. [Google Scholar]
- Mondy NI, Leja M and Gosselin B, 1987. Changes in total phenolic, total glycoalkaloid, and ascorbic acid content of potatoes as a result of bruising. Journal of Food Science, 52, 631–633. [Google Scholar]
- Morgan MRA, Coxon DT, Bramham S, Chan HWS, Van Gelder WMJ and Allison MJ, 1985. Determination of the glycoalkaloid content of potato tubers by three methods including enzyme‐linked immunosorbent assay. Journal of the Science of Food and Agriculture, 36, 282–288. [Google Scholar]
- Mori M and Kozukue N, 1995. The glycoalkaloid contents of potato tubers: differences in tuber greening in various varieties and lines. Report of the Kyushu Branch of the Crop Science Society of Japan, 61, 77–79. [Google Scholar]
- Mori K, Asano K, Tamiya S, Nakao T and Mori M, 2015. Challenges of breeding potato cultivars to grow in various environments and to meet different demands. Breed Science, 65, 3–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris T, 1859. Case of poisoning by potatoes‐berries. British Medical Journal, 719–720. As cited by Van Gelder, 1989. [Google Scholar]
- Morris SC and Lee TH, 1984. The toxicity and teratogenicity of Solanaceae glycoalkaloids, particularly those of the potato (Solanum tuberosum): a review. Food Technology in Australia, 36, 118–124. [Google Scholar]
- Morris SC and Petterman JB, 1985. Genetic and environmental effects on levels of glycoalkaloids in cultivars of potato (Solanum tuberosum L.). Food Chemistry, 18, 271–282. [Google Scholar]
- Mulatu B, Applebaum SW, Zerem Z and Coll M, 2006. Tomato fruit size, maturity and a‐tomatine content influence the performance of larvae of potato tuber moth Phthorimaea operculella (Lepidoptera: Gelechiidae). Bulletin of Entomological research, 96, 173–178. [DOI] [PubMed] [Google Scholar]
- Mulder P, 2019. Personal communication provided via email.
- Mulinacci N, Ieri F, Giaccherini C, Innocenti M, Andrenelli L, Canova G, Saracchi M and Casiraghi MC, 2008. Effect of cooking on the anthocyanins, phenolic acids, glycoalkaloids, and resistant starch content in two pigmented cultivars of Solanum tuberosum L. Journal of Agricultural Food Chemistry, 56, 11830–11837. [DOI] [PubMed] [Google Scholar]
- Munari CC, de Oliveira PF, de Souza Lima IM, de Paula Lima Martins S, de Carvalho da Costa J, Bastos JK and Tavares DC, 2012. Evaluation of cytotoxic, genotoxic and antigenotoxic potential of Solanum lycocarpum fruits glicoalkaloid extract in V79 cells. Food Chemistry and Toxicology, 50, 3696–3701. [DOI] [PubMed] [Google Scholar]
- Munari CC, de Oliveira PF, Leandro LF, Pimenta LM, Ferreira NH, da Costa J de C, Bastos JK and Tavares DC, 2014. In vivo assessment of genotoxic, antigenotoxic and anticarcinogenic activities of Solanum lycocarpum fruits glycoalkaloidic extract. PLoS ONE, 9, 1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muncke , 1845. Med. Ann. Heidelberg XI:298. As cited by Bömer and Matthis, 1924.
- Muneta P, Shen W‐W, Jennings S, Everson D and Edwarsa L, 1977. Potassium hydroxide for peeling potatoes. American Potato Journal, 54, 83–90. [Google Scholar]
- Najm AA, Haidi MRHS, Fazeli F, Darzi MT and Rahi A, 2012. Effect of Integrated Management of Nitrogen Fertilizer and Cattle Manure on the Leaf Chlorophyll, Yield, and Tuber Glycoalkaloids of Agria Potato. Communications in Soil Science and Plant Analysis, 43, 912–923. [Google Scholar]
- Nara A, Saka K, Yamada C, Kodama T and Takagi T, 2019. Forensic analysis using ultra‐high‐performance liquid chromatography–tandem mass spectrometry with solid‐phase extraction of α‐solanine and α‐chaconine in whole blood. Forensic Toxicology, 37, 197–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nema PK, Ramayya N, Duncan E and Niranjan K, 2008. Potato glycoalkaloids: formation and strategies for mitigation. Journal of the Science of Food and Agriculture, 88, 1869–1881. [Google Scholar]
- Ness E, Joner PE and Dahle HK, 1984. Alpha‐solanine tested for mutagenicity with Ames test. Acta veterinaria Scandinavica, 25, 145–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nevin NC and Merrett JD, 1975. Potato avoidance during pregnancy in women with a previous infant with either anencephaly and/or spina bifida. British Journal of Preventive and Social Medicine, 29, 111–115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ni W, Tian T, Zhang L, Wang L and Ren A, 2018. Maternal periconceptional consumption of sprouted potato and risks of neural tube defects and orofacial clefts. Nutrition Journal, 28, 17–112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nie X, Zhang G, Lv S and Guo H, 2018. Steroidal glycoalkaloids in potato foods as affected by cooking methods. International Journal of Food Properties, 21, 1. [Google Scholar]
- Nie X, Li C, Zhang G, Shao Z, Wang X, Shi H and Guo H, 2019. Light exposure and wounding: synergistic effects on steroidal glycoalkaloid accumulation in potato tubers during storage. International Journal of Food Science and Technology, 54, 2939–2948. [Google Scholar]
- Nigg HN, Ramos LE, Graham EM, Sterling J, Brown S and Cornell JA, 1996. Inhibition of human plasma and serum butyrylcholinesterase (EC 3.1.1.8) by alpha‐chaconine and alpha‐solanine. Fundamental and Applied Toxicology, 33, 272–281. [DOI] [PubMed] [Google Scholar]
- Nishie K, Gumbmann MR and Keyl AC, 1971. Pharmacology of solanine. Toxicology and Applied Pharmacology, 19, 81–92. [DOI] [PubMed] [Google Scholar]
- Nishie K, Norred WP and Swain AP, 1975. Pharmacology and toxicology of chaconine and tomatine. Research Communications in Chemical Pathology and Pharmacology, 12, 657–668. [PubMed] [Google Scholar]
- Nohara T, Ono M, Ikeda T, Fujiwara Y and El‐Aasr , 2010. The tomato saponin, Esculeoside A. Journal of Natural Products, 73, 1734–1741. [DOI] [PubMed] [Google Scholar]
- Nordic Working Group on Food Toxicology and Risk Assessment , 1990. Assessment of health‐risks related to glycoalkaloids (‘Solanine’) in potatoes: a nordic view. Compiled by Slanina P. Vår Föda, 43, Supplement 1, 14 pp.
- Norman TE, Chaffin MK, Norton PL, Coleman MC, Stoughton WB and Mays T, 2012. Concurrent ivermectin and Solanum spp. toxicosis in a herd of horses. Journal of Veterinary Internal Medicine, 26, 1439–1442. [DOI] [PubMed] [Google Scholar]
- Norred WP, Nishie K and Osman SF, 1976. Excretion, distribution and metabolic fate of 3H‐alpha‐chaconine. Research Communications in Chemical Pathology and Pharmacology, 13, 161–171. [PubMed] [Google Scholar]
- Norwegian Scientific Committee for Food Safety , 2009. Opinion of the Panel on Animal Feed of the Norwegian Scientific Committee for Food Safety. Criteria for safe use of plant ingredients in diets for aquacultured fish. Available online: https://vkm.no/download/18.2994e95b15cc5450716356b2/1498419374011/1232b28f3b.pdf
- Ogg AG and Rogers BS, 1989. Taxonomy, distribution, biology, and control and black nightshade (Solanum nigrum) and related species in the United States and Canada. Reviews of Weed Science, 4, 25–58. [Google Scholar]
- Ogg AG Jr, Rogers BS and Schilling EE, 1981. Characterization of black nightshade (Solanum‐nigrum) and related species in the United States. Weed Science, 29, 27–32. [Google Scholar]
- Olsen NL, Brandt T and Price WJ, 2018. The Impact of Retail Light Source on Greening of Russet Burbank Potato Tubers. American Journal of Potato Research, 95, 123–129. [Google Scholar]
- Orgell WH, 1963. Inhibition of human plasma cholinesterase in vitro by alkaloids, glycosides, and other natural substances. Lloydia, 26, 36–43. [Google Scholar]
- Orgell WH, Vaidya KA and Dahm PA, 1958. Inhibition of human plasma cholinesterase in vitro by extracts of solanaceous plants. Science, 128, 1136–1137. [DOI] [PubMed] [Google Scholar]
- Ostrý V, Ruprich J and Skarkova J, 2010. Glycoalkaloids in potato tubers: the effect of peeling and cooking in salted water. Acta Alimentaria, 39, 130–135. [Google Scholar]
- Panovská Z, Hajšlová J, Kosinková P and Čepl J, 1997. Glycoalkaloid content of potatoes sold in Czechia. Nahrung, 41, 146–149. [DOI] [PubMed] [Google Scholar]
- Papathanasiou F, Mitchell SH and Harvey BMR, 1999. Variation in glycoalkaloid concentration of potato tubers harvested from mature plants. Journal of the Science of Food and Agriculture, 79, 32–36. [Google Scholar]
- Paradiso R, Arena C, Rouphael Y, d'Acquino L, Makris K, Vitaglione P and De Pascale S, 2019. Growth, photosynthetic activity and tuber quality of two potato cultivars in controlled environment as affected by light source. Plant Biosystems, 153, 725–735. [Google Scholar]
- Pariera Dinkins CL and Peterson RKD, 2008. A human dietary risk assessment associated with glycoalkaloid responses of potato to Colorado potato beetle defoliation. Food and Chemical Toxicology, 46, 2837–2840. [DOI] [PubMed] [Google Scholar]
- Park S, Park MY, Song G and Lim W, 2009. Alpha‐solanine inhibits cell proliferation via mitochondrial dysfunction and inhibin synthesis in mouse testis In vitro and In vivo. Chemosphere, 235, 271–279. [DOI] [PubMed] [Google Scholar]
- Parnell A, Bhuva VS and Bintcliffe EJB, 1984. The glycoalkaloid content of potato varieties. Journal of the National Institute of Agricultural Botanic, 16, 535–541. [Google Scholar]
- Patel B, Schutte R, Sporns P, Doyle J, Jewel L and Fedorak RN, 2002. Potato glycoalkaloids adversely affect intestinal permeability and aggravate inflammatory bowel disease. Inflammatory Bowel Diseases, 8, 340–346. [DOI] [PubMed] [Google Scholar]
- Patil BC, Sharma RP, Salunkhe DK and Salunkhe K, 1972. Evaluation of solanine toxicity. Food and Cosmetics Toxicology, 10, 395–398. [DOI] [PubMed] [Google Scholar]
- Pęksa A, Golubowska G, Aniolowski K, Lisinska G and Rytel , 2006. Changes of glycoalkaloids and nitrate content in potatoes during chip processing. Food Chemistry, 97, 151–156. [Google Scholar]
- Percival G, 1999. Light‐induced glycoalkaloid accumulation of potato tubers (Solanum tuberosum L). Journal of the Science of Food and Agriculture, 79, 1305–1310. [Google Scholar]
- Pereira EFR, Aracava Y, DeTolla LJ Jr, Beecham EJ, Basinger GW Jr, Wakayama EJ and Albuquerque EX, 2014. Animal Models That Best Reproduce the Clinical Manifestations of Human Intoxication with Organophosphorus Compounds. Journal of Pharmacology and Experimental Therapy, 350, 313–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petersson EV, Arif U, Schulzova V, Krtková V, Hajšlová J, Meijer J, Andersson HC, Jonsson L and Sitbon F, 2013. Glycoalkaloid and Calystegine Levels in Table Potato Cultivars Subjected to Wounding, Light, and Heat Treatments. Journal of Agricultural and Food Chemistry, 61, 5893–5902. [DOI] [PubMed] [Google Scholar]
- Pfuhl E, 1899. Regarding an outbreak of illness due to poisoning by solanine in potatoes. Deutsche Medizinische Wochenschrift, 25, 753–754. [Google Scholar]
- Phillips BJ, Hughes JA, Phillips JC, Walters DG, Anderson D and Tahourdin CSM, 1996. A study of the toxic hazard that might be associated with the consumption of green potato tops. Food and Chemical Toxicology, 34, 439–448. [DOI] [PubMed] [Google Scholar]
- Pohanska M, 2014. Inhibitors of Acetylcholinesterase and Butyrylcholinesterase Meet Immunity. International Journal of Molecular Sciences, 15, 9809–9825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pokrovskii AA, 1956. Effect of alkaloids of growing potatoes on cholinesterase. Biokhimiia, 21, 683–688. [PubMed] [Google Scholar]
- Pond WG and Maner JH, 1984. Swine production and nutrition. AVI Publishing Co., Westport, Connecticut: AVI Pub. Co, 1984. ISBN: 0870554506. Available online: https://trove.nla.gov.au/version/26200281 [Google Scholar]
- Poswillo DE, Sopher D and Mitchell S, 1972. Experimental induction of foetal malformation with “blighted” potato: A preliminary report. Nature, 239, 462–464. [DOI] [PubMed] [Google Scholar]
- Poswillo DE, Sopher D, Mitchell SJ, Coxon DT, Curtis RF and Price KR, 1973a. Investigations into the Teratogenic Potential of Imperfect Potatoes. Teratology, 8, 339–348. [DOI] [PubMed] [Google Scholar]
- Poswillo DE, Sopher D and Mitchell S, 1973b. Further investigations into the Teratogenic Potential of Imperfect potatoes. Nature, 224, 367–368. [DOI] [PubMed] [Google Scholar]
- Reale M, Costantini E, Di Nicola M, D'Angelo C, Franchi S, D'Aurora M, Di Bari M, Orlando V, Galizia S, Ruggieri S, Stuppia L, Gasperini C, Tata AM and Gatta V, 2018. Butyrylcholinesterase and Acetylcholinesterase polymorphisms in Multiple Sclerosis patients: implication in peripheral inflammation. Scientific Reports, 8, 1319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reelakh LA and Keem A, 1958. Sov. Med. 22, 129. As cited by McMillan and Thompson (1979).
- Refstie S and Tiekstra HAJ, 2003. Potato protein concentrate with low content of solanidine glycoalkaloids in diets for Atlantic salmon (Salmo salar). Aquaculture, 216, 283–298. [Google Scholar]
- Renwick JH, 1972. Hypothesis: anencephaly and spina bifida are usually preventable by avoidance of a specific but unidentified substance present in certain potato tubers. British Journal of Preventive and Social Medicine, 26, 67–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Renwick JH, Calringbold WDB, Earthy ME, Few JD and McLean ACS, 1984. Neural‐Tube Defects Produced in Syrian Hamsters by Potato Glycoalkaloids. Terotology, 30, 371–381. [DOI] [PubMed] [Google Scholar]
- Rice SG, Nowak L, Duysen EG, Lockridge O, Lahiri DK and Reyes PF, 2007. Neuropathological and immunochemical studies of brain parenchyma in acetylcholinesterase knockout mice: Implications in Alzheimer's disease. Journal of Alzheimers Disease, 11, 481–489. [DOI] [PubMed] [Google Scholar]
- Roddick JG, 1989. The acetylcholinesterase‐inhibitory activity of steroidal glycoalkaloids and their aglycones. Phytochemistry, 28, 2631–2634. [Google Scholar]
- Roddick JG, 1996. Steroidal Glycoalkaloids: Nature and Consequences of Bioactivity. Advances in Experimental Medicine and Biology, 404, 277–295. [DOI] [PubMed] [Google Scholar]
- Romanucci V, Pisanti A, Di Fabio G, Davinelli S, Scapagnini G, Guaragna A and Zarrelli A, 2016. Toxin levels in different variety of potatoes: Alarming contents of α‐chaconine. Phytochemistry Letters, 16, 103–107. [Google Scholar]
- Romanucci V, Di Fabio G, Di Marino C, Davinelli S, Scapagnini G and Zarrelli A, 2018. Evaluation of new strategies to reduce the total content of α‐solanine and α‐chaconine in potatoes. Phytochemistry Letters, 23, 116–119. [Google Scholar]
- Rosenfeld HJ, Sundell HA and Ringstad PLM, 1995. Influence of packaging materials and temperature on the glycoalkaloid content of potato tubers. Food Research International, 28, 481–484. [Google Scholar]
- Ross H, Pasemann P and Nitzsche W, 1978. Der Glykoalkaloidgehalt von Kartoff elsorten in seiner Abhangigkeit von Anbauort und ‐jahr und seiner Beziehung zum Geschmack. Z. Pflanzenziichtg., 80, 64–79. [Google Scholar]
- Rothe JC, 1919. Über Erkrankungen nach Genuss von solaninhaltigen Kartoffeln. Zeitsch Hyg, 88, 1–12. [Google Scholar]
- Ruddick JA, Harwig J and Scott PM, 1974. Nonteratogenicity in rats of blighted potatoes and compounds contained in them. Teratology, 9, 165–168. [DOI] [PubMed] [Google Scholar]
- Ruprich J, Rehurkova I, Boon PE, Svensson K, Moussavian S, Van der Voet H, Bosgra S, Van Klaveren JD and Busk L, 2009. Probabilistic modelling of exposure doses and implications for health risk characterization: Glycoalkaloids from potatoes. Food and Chemical Toxicology, 47, 2899–2905. [DOI] [PubMed] [Google Scholar]
- Rytel E, 2012a. Changes in the levels of glycoalkaloids and nitrates after the dehydration of cooked potatoes. American Journal of Potato Research, 89, 501–507. [Google Scholar]
- Rytel E, 2012b. The effect of industrial potato processing on the concentrations of glycoalkaloids and nitrates in potato granules. Food Control, 28, 380–384. [Google Scholar]
- Rytel E, 2012c. Changes in glycoalkaloid and nitrate content in potatoes during dehydrated dice processing. Food Control, 25, 349–354. [Google Scholar]
- Rytel E, 2013. Effect of technological factors on glycoalkaloids and nitrates content in dehydrated potato. Italian J. Food Sci, 25, 142–148. [Google Scholar]
- Rytel E, Gołubowska G, Linińska G, Pęksa A and Aniołowski K, 2005. Changes in glycoalkaloid and nitrate contents in potatoes during French fries processing. Journal of the Science of Food and Agriculture, 85, 879–882. [Google Scholar]
- Rytel E, Tajner‐Czopek A, Aniołowska M and Hamouz K, 2013. The influence of dehydrated potatoes processing on the glycoalkaloids content in coloured‐fleshed potato. Food Chemistry, 141, 2495–2500. [DOI] [PubMed] [Google Scholar]
- Rytel E, Tajner‐Czopek A, Kita A, Miedzianka J and Bronkowska M, 2015. The influence of washing and selection processes on the contents of glycoalkaloid and other toxic compounds during industrial chip production. International Journal of Food Science & Technology, 50, 1737–1742. [Google Scholar]
- Rytel E, Tajner‐Czopek A, Kita A, Kucharska AZ, Sokół‐Łeztowska A and Hamouz K, 2018. Content of anthocyanins and glycoalkaloids in blue‐fleshed potatoes and changes in the content of a‐solanine and a‐chaconine during manufacture of fried and dried products. International Journal of Food Science and Technology, 53, 719–727. [Google Scholar]
- Sabatino L, Iapichino G, D'Anna F, Palazzolo E, Mennella G and Rotino GL, 2018. Hybrids and allied species as potential rootstocks for eggplant: Effect of grafting on vigour, yield and overall fruit quality traits. Scientia Horticulturae, 228, 81–90. [Google Scholar]
- Sackmann W, Kern H and Wiesmann E, 1959. Untersuchungen über die biologischen Wirkungen von Tomatin and Solanin (Studies on the biological effects of tomatin and solanine). Schweizerische Zeitschrift fur Pathologie und Bakteriologie. Revue Suisse de Pathologie et de Bacteriologie, 22, 557–563. [PubMed] [Google Scholar]
- Saito K, Horie M, Hoshino Y, Nose N and Nakazawa H, 1990. High‐performance liquid chromatographic determination of glycoalkaloids in potato products. Journal of Chromatography, 508, 141–147. [Google Scholar]
- Sánchez‐Mata MC, Yokoyama WE, Hong Y‐J and Prohens J, 2010. Alpha‐solasonine and alpha‐solamargine contents of gboma (Solanum macrocarpon L.) and scarlet (Solanum aethiopicum L.) eggplants. Journal of Agricultural and Food Chemistry, 58, 5502–5508. [DOI] [PubMed] [Google Scholar]
- Sander H, 1956. Studien über Bildung und Abbau von Tomatin in der Tomatenpflanze (The formation and degradation of tomatin in the tomato plant). Planta, 47, 374–400. [Google Scholar]
- Sander H, 1958. Tomatin ein möglicher Ausgangsstoff für die Lycopinbildung. Naturwissenschaften, 45, 59. [Google Scholar]
- Sarquis JI, Coria NA, Aguilar I and Rivera A, 2000. Glycoalkaloid content in Solanum species and hybrids from a breeding program for resistance to late blight (Phytophthora infestans). American Journal of Potato Research, 77, 295–302. [Google Scholar]
- Satoh T, 1967. Glycemic effects of solanine in rats. Japanese Journal of Pharmacology, 17, 652–658. [PubMed] [Google Scholar]
- Schmiedeberg O, 1892. Űber die toxikologische Bedeutung des Solaningehaltes der Kartoffeln. Archiv für exp. Pathologie und Pharmakologie, Bd. XXXVI, S. 373. As cited by Bömer and Matthis (1924).
- Schwahn K, de Souza LP, Fernie AR and Tohge T, 2014. Metabolomics‐assisted refinement of the pathways of steroidal glycoalkaloid biosynthesis in the tomato clade. Journal of Integrative Plant Biology, 56, 864–875. 10.1111/jipb.12274 [DOI] [PubMed] [Google Scholar]
- Schwarz A, Pinto E, Haraguchi M, de Oliveira CA, Bernardi MM and de Souza Spinosa H, 2007. Phytochemical study of Solanum lycocarpum (St. Hil) unripe fruit and its effect on rat gestation. Phytotherapy Research, 21, 1025–1028. [DOI] [PubMed] [Google Scholar]
- Sepelevs I, Nakurte I and Galoburda R, 2019. Biologically active compound stability in the industrial potato processing by‐products. Foodbalt, 131–136, ref. 40. [Google Scholar]
- Sharma RP, Willhite CC, Wu MT and Salunkhe DK, 1978. Teratogenic potential of blighted potato concentrate in rabbits, hamster and miniature swine. Teratology, 18, 55–62. [DOI] [PubMed] [Google Scholar]
- Sharma RP, Willhite CC, Shupe JL and Salunkhe DK, 1979. Acute toxicity and histopathological effects of certain glycoalkaloids and extracts of Alternaria solani or Phytophthora infestans in mice. Toxicology Letters, 3, 349–355. [Google Scholar]
- Sharma RP, Taylor MJ and Bourcier DR, 1983. Subcellular distribution of alpha‐chaconine in mouse hepatocytes. Drug and Chemical Toxicology, 6, 219–234. [DOI] [PubMed] [Google Scholar]
- Shin JS, Lee KG, Lee HH, Lee HJ, An HJ, Nam JH, Jang DS and Lee KT, 2016. alpha‐Solanine Isolated From Solanum Tuberosum L. cv Jayoung Abrogates LPS‐Induced Inflammatory Responses Via NF‐kappaB Inactivation in RAW 264.7 Macrophages and Endotoxin‐Induced Shock Model in Mice. Journal of Cellular Biochemistry, 117, 2327–2339. [DOI] [PubMed] [Google Scholar]
- Sinden SL and Webb RE, 1972. Effect of variety and location on the glycoalkaloid content of potatoes. American Potato Journal, 49, 334–338. [Google Scholar]
- Sinden SL and Webb RE, 1974. Effect of environment ion glycoalkaloid content of six potato varieties at 39 locations. Technical Bulleting No 1472. Agricultural Research Service. United States Department of Agriculture.
- Sinden SL, Sanford LL and Webb RE, 1984. Genetic and environmental control of potato glycoalkaloids. American Potato Journal, 61, 141–155. [Google Scholar]
- Sizer CE, Maga JA and Craven CJ, 1980. Total glycoalkaloids in potatoes and potato chips. Journal of Agricultural and Food Chemistry, 28, 578–579. [DOI] [PubMed] [Google Scholar]
- Skarkova J, Ostry V and Ruprich J, 2008. Determination of Glycoalkaloids (α‐solanine and α‐chaconine) in Peeled Potato Tubers Using High‐Performance Thin‐Layer Chromatography. Journal of Planar Chromatography ‐ Modern TLC, 21, 113–117. [Google Scholar]
- Skrabule I, Muceniece R and Kirhnere I, 2013. Evaluation of Vitamins and Glycoalkaloids in Potato Genotypes Grown Under Organic and Conventional Farming Systems. Potato Research, 56, 259–276. [Google Scholar]
- Smith DB, Roddick JG and Jones JL, 1996. Potato glycoalkaloids: some unanswered questions. Trends in Food Science and Technology, 7, 126–131. [Google Scholar]
- Smith SW, Giesbrecht E, Thompson M, Nelson LS and Hoffman RS, 2008. Solanaceous steroidal glycoalkaloids and poisoning by Solanum torvum, the normally edible susumber berry. Toxicon, 52, 667–676. [DOI] [PubMed] [Google Scholar]
- Sotelo A and Serrano B, 2000. High‐Performance Liquid Chromatographic Determination of the Glycoalkaloids r‐Solanine and r‐Chaconine in 12 Commercial Varieties of Mexican Potato. Journal of Agricultural and Food Chemistry, 48, 2472–2475. [DOI] [PubMed] [Google Scholar]
- Stanker LH, Kamps‐Holtzapple C and Friedman M, 1994. Development and characterization of monoclonal antibodies that differentiate between potato and tomato glycoalkaloids and aglycons. Journal of Agricultural and Food Chemistry, 42, 2360–2366. [Google Scholar]
- Stein HH, 2002. Experience of feeding pigs without antibiotics: A European Perspective. Animal Biotechnology, 13, 85–95. [DOI] [PubMed] [Google Scholar]
- Sternfeld M, Rachmilewitz J, Loewenstein‐Lichtenstein Y, Andres C, Timberg R, BenAri S, Glick D, Soreq H and Zakut H, 1997. Normal and atypical butyrylcholinesterases in placental development, function, and malfunction. Cellular and Molecular Neurobiology, 17, 315–332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoyanova LA, Codreanu EP, Bobeiko VA and Balashova NN, 1988. Changes in the content of glycoalkaloids during processing of tomatoes. Piŝevaâ promyšlennostʹ (Moskva), 10, 55–56. [Google Scholar]
- Spoladore DS, Teixeira JPF, Zullo MAT, Teixeira PRM, Coelho SMBM and Filho FSM, 1983. Occurrence of glycoalkaloids and greening in newly harvested and stored potato tubers. Bragantia, 42, 221–231. [Google Scholar]
- Spoladore DS, Zullo MAT and Teixeira JPF, Coelho SMBM and Filho FSM, 1985. Synthesis of chlorophylls and glycoalkaloids in mature potato tubers stored under natural light. Bragantia, 44, 197–208. [Google Scholar]
- Sucha L, Hroch M, Rezacova M, Rudolf E, Havelek R, Sispera L, Cmielova J, Kohlerova R, Bezrouk A and Tomsik P, 2013. The cytotoxic effect of α‐tomatine in MCF‐7 human adenocarcinoma breast cancer cells depends on its interaction with cholesterol in incubation media and does not involve apoptosis induction. Oncology Reports, 30, 2593–2602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Surjawan I, Dougherty MP, Bushway RJ, Bushway AA, Briggs JL and Camire ME, 2001. Sulfur Compounds Reduce Potato Toxins during Extrusion Cooking. Journal of Agricultural and Food Chemistry, 49, 2835–2838. [DOI] [PubMed] [Google Scholar]
- Swain AP, Fitzpatrick TJ, Talley EA, Herb SF and Osman SF, 1978. Anzymatic hydrolysis of a‐chaconine and cc‐solanine. Phytochemistry, 17, 800–801. [Google Scholar]
- Swedish NFA (Swedish National Food Agency), 2016. Samordnade kontrollprojekt 2015 − Glykoalkaloider, kadmium och bly i potatis. Livsmedelsverkets rapportserie nr 23/2015.
- Swinyard CA and Chaube S, 1973. Are potatoes teratogenic for experimental animals. Teratology, 8, 349–357. [DOI] [PubMed] [Google Scholar]
- Taher D, Solberg SØ, Prohens J, Chou Y‐Y, Rakha M and Wu T‐H, 2017. World vegetable center eggplant collection: origin, composition, seed dissemination and utilization in breeding. Frontiers. Plant Science, 8, Article 1484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tajner‐Czopek A, Jarych‐Szyszka M and Lisinska G, 2008. Changes in glycoalkaloids content of potatoes destined for consumption. Food Chemistry, 106, 706–711. [Google Scholar]
- Tajner‐Czopek A, Rytel E, Kita A, Pe˛ksa A and Hamouz K, 2012. The influence of thermal process of coloured potatoes on the content of glycoalkaloids in the potato products. Food Chemistry, 133, 1117–1122. [Google Scholar]
- Tajner‐Czopek A, Rytel E, Aniolowska M and Hamouz K, 2014. The influence of French fries processing on the glycoalkaloid content in coloured‐fleshed potatoes. European Food Research and Technology, 238, 895–904. [Google Scholar]
- Takagi K, ToyodaM Fujiyama Y and Saito Y, 1990. Effect of cooking on the contents of α‐chaconine and α‐solanine in potatoes. Shokuhin Eiseigaku Zasshi, 31, 67–73. [Google Scholar]
- Tamasi G, Pardini A, Bonechi C, Donati A, Pessina F and Marcolongo P, 2019. Characterization of nutraceutical components in tomato pulp, skin and locular gel. European Food Research and Technology, 245, 907–918. [Google Scholar]
- Temperton H, 1943. The effects of feeding green and sprouted potatoes to laying pullets. Veterinary Record, 55, 359. [Google Scholar]
- Tepe EJ, Anderson GJ, Spooner DM and Bohs L, 2016. Relationships among wild relatives of the tomato, potato and pepino. Taxon, 65, 262–276. [Google Scholar]
- Tice RR, 1998. α‐Chaconine [20562‐03-2] and α‐Solanine [20562‐02-1]. Review of toxicological literature, Integrated Laboratory Systems: p. 103. [Google Scholar]
- Tömösközi‐Farkas R, Daood HG, Polgár ZS and Hajós GY, 2006. Determination of Glycoalcaloids in Hungarian Potatoes by HPLC. Chromatographia Supplement, 63, 115–118. [Google Scholar]
- Tömösközi‐Farkas R, Polgár ZS, Nagy‐Gasztonyi M, Horváth V, Renkecz T, Simon K, Boross F, Fabulya Z and Daood H, 2014. Changes of potentially antinutritive components in 8 Hungarian potatoes under organic and conventional farming. Acta Alimentaria, 43, 676–683. [Google Scholar]
- Tsiaoussis J, Hatzidaki E, Docea AO, Nikolouzakis TK, Petrakis D, Burykina T, Mamoulakis C, Makrigiannakis A and Tsatsakis A, 2018. Molecular and clinical aspects of embryotoxicity induced by acetylcholinesterase inhibitors. Toxicology, 409, 137–143. [DOI] [PubMed] [Google Scholar]
- Turner NJ and Kuhnlein HV, 1983. Camas (Camasia spp.) and Riceroot (Fritillaria spp.): two Liliaceous ‘root’ foods of the Northwest Coast Indians. Ecology of Food and Nutrition, 13, 199–219. [Google Scholar]
- Tusche T, Wuertz S, Susenbeth A and Schulz C, 2011. Feeding fish according to organic aquaculture guidelines EC710/2009: Influence of potato protein concentrates containing various glycoalkaloid levels on health status and growth performance of rainbow trout (Oncorhynchus mykiss). Aquaculture, 319, 122–131. [Google Scholar]
- Tuśnio A, Pastuszewska B, Taciak M, Barszcz M and Skomiał J, 2013a. The response of rats to solanidine glycoalkaloids and trypsin inhibitor present in potato protein concentrates, and to glycoalkaloids provided by potato sprouts. Journal of Animal and Feed Sciences, 22, 130–136. [Google Scholar]
- Tuśnio A, Pastuszewska B, Taciak M, Mieczkowska A and Smulikowska S, 2013b. Response of growing chicken to potato protein concentrates providing different amounts of solanidine glycoalkaloids and trypsin inhibitor. Archiv fűr Geflűgelkunde, 77, 51–58. [Google Scholar]
- University of Chemistry and Technology Prague , 2019. Extensive literature search and selection for relevance of studies related to the chemistry and toxicity of glycoalkaloids and quinolizidine alkaloids in food and feed. 10.2903/sp.efsa.2019.en-1348 [DOI]
- Urban J, Hamouz K, Lachman J, Pulkrábek J and Pazderů K, 2018. Effect of genotype, flesh colour and environment on the glyco‐alkaloid content in potato tubers from integrated agriculture. Plant, Soil and Environment, 64, 186–191. [Google Scholar]
- USDA (United States Department of Agriculture), 2011. Plants poisonous to livestock in the Western States. Agricultural Research Service Information Bulletin 415.
- Valcárcel J, Reilly K, Gaffney M and O'Brien N, 2014. Effect of Genotype and Environment on the Glycoalkaloid Content of Rare, Heritage, and Commercial Potato Varieties. Journal of Food Science, 79, 1039–1048. [DOI] [PubMed] [Google Scholar]
- Valkonen JPT, Keskitalo M, Vasara T and Pietila L, 1996. Potato glycoalkaloids: a burden or a blessing? Critical Reviews in Plant Sciences, 15, 1–20. [Google Scholar]
- Van Denburgh RW, Hiller L‐K- and Koller DC, 1986. Ultrastructural Changes in Potato Tuber Pith Cells during Brown Center Development. Plant Physiology, 81, 167–170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Gelder WMJ, 1985. Determination of the total C27‐steroidal alkaloid composition of Solanum species by high‐resolution gas chromatography. Journal of Chromatography A, 331, 285–293. [Google Scholar]
- Van Gelder WMJ, 1989. Steroidal glycoalkaloids in Solanum species: consequences for potato breeding and food safety. Van Gelder, Dissertation. Centrale landbouwcatalogus 000 0352 3277, ISN 506943.
- Van Gelder WMJ, 1991. Chemistry, toxicology, and occurrence of steroidal glycoalkaloids: potential contaminants of the potato In Rizk AFM, ed. Poisonous Plant Contamination of Edible Plants. CRC Press, Boca Raton, FL, pp. 121–143. [Google Scholar]
- Van Gelder WMJ and De Ponti OMB, 1987. α‐Tomatine and other steroidal glycoalkaloids in fruits of tomato lines resistant to the glasshouse whitefly (Trialeurodes vaporariorum Westw.). Euphytica, 36, 555–561. [Google Scholar]
- Van Gelder WMJ, Vinket JH and Scheffer JJC, 1988. Steroidal glycoalkaloids in tubers and leaves of Solanum species used in potato breeding. Euphytica, 39, 147–158. [Google Scholar]
- Viollier R, 1941. Lebensmittelkontrolle und Berichte von Untersuchungsämtern. Zeitschrift fuer Untersuchung der Lebensmittel, 192.
- Vogel‐Höpker A, Sperling LE and Layer PG, 2012. Co‐opting Functions of Cholinesterases in Neural, Limb and Stem Cell Development. Protein and Peptide Letters, 19, 155–164. [DOI] [PubMed] [Google Scholar]
- Vogt H and Stute K, 1969. Dried ground potatoes in poultry rations. II. Solanine determination and effects of solanine. Archiv fűr Geflűgelkunde, 33, 316–320. [Google Scholar]
- Voldrich M, Ondroušek S and Dobiáš J, 1992. Steroid glycoalkaloids in fresh and processed tomatoes. Potrav. Vêdy, 10, 23–30. [Google Scholar]
- Vorontsova MS, Stern S, Bohs L and Knapp S, 2013. African spiny Solanum subgenus Leptostemonum, Solanaceae): a thorny phylogenetic tangle. Botanical Journal of the Linnean Society, 173, 176–193. [Google Scholar]
- Wang F, Yang YF and Li PZ, 2008. A case‐control study on the risk factors of neural tube defects in Shanxi province. Zhonghua Liu Xing Bing Xue Za Zhi, 29, 771–774. In Chinese, abstract in English. [PubMed] [Google Scholar]
- Whittemore CT, 1977. The potato (Solanum tuberosum) as a source of nutrients for pigs, calves and fowl — A review. Animal Feed Science and Technology, 2, 171–190. [Google Scholar]
- WHO/IPCS (World Health Organisation/International Programme on Chemical Safety), 2008. Uncertainty and data quality in exposure assessment. Harmonisation project document No. 6. ISBN 978924156376. Available online: https://apps.who.int/iris/handle/10665/44017
- WHO/IPCS (World Health Organisation/International Programme on Chemical Safety), 2009. Principles and methods for the risk assessment of chemicals in food. Environmental Health Criteria 240. International Programme on Chemical Safe. ISBN: 9789241572408. Available online: https://www.who.int/foodsafety/publications/chemical-food/en/
- Willimott SG, 1933. An investigation of solanine poisoning. Analyst, 58, 431–439. [Google Scholar]
- Wilson GS, 1959. A small outbreak of solanine poisoning. Monthly bulletin of the Ministry of Health and the Public Health Laboratory Service, 18, 207–210. [PubMed] [Google Scholar]
- Wilson RH, Poley GW and De Eds F, 1961. Some pharmacologic and toxicologic properties of tomatine and its derivatives. Toxicology and Applied Pharmacology, 3, 39–48. [DOI] [PubMed] [Google Scholar]
- Wilson AM, McGann DF and Bushway RJ, 1983. Effect of growth‐location and length of storage on glycoalkaloid content of roadside‐stand potatoes as stored by consumers. Journal of Food Protection, 46, 119–121. [DOI] [PubMed] [Google Scholar]
- Wood F and Young D, 1974. TGA in potatoes. Agriculture Canada, 1533, 3. [Google Scholar]
- Xie S and Jokumsen A, 1997. Incorporation of potato protein concentrate in diets for rainbow trout: effects on feed intake, growth and feed utilization. Aquacult. Nutr, 3, 223–226. [Google Scholar]
- Xie S and Jokumsen A, 1998. Effects of dietary incorporation of potato protein concentrate and supplementation of methionine on growth and feed utilization of rainbow trout. Aquacult. Nutr., 4, 183–186. [Google Scholar]
- Xie W, Stribley JA, Chatonnet A, Wilder PJ, Rizzino A, McComb RD, Taylor P, Hinrichs SH and Lockridge O, 2000. Postnatal developmental delay and supersensitivity to organophosphate in gene‐targeted mice lacking acetylcholinesterase. Journal of Pharmacology and Experimental Therapeutics, 293, 896–902. [PubMed] [Google Scholar]
- Zarzecka K and Gugala M, 2007. Changes in the content of glycoalkaloids in potato tubers according to soil tillage and weed control methods. Plant, Soil and Environment, 53, 247–251. [Google Scholar]
- Zgorska K, Czerko Z and Grudzinska M, 2006. The effect of light exposure on greening and accumulation of chlorophyll and glycoalkaloids in potato tubers. Zywnosc, 13, 222–228. [Google Scholar]
- Zhang L, Li M, Zhang G, Wu L, Cai D and Wu Z, 2018. Inhibiting Sprouting and Decreasing α‐Solanine Amount of Stored Potatoes Using Hydrophobic Nanosilica. ACS Sustainable Chemistry and Engineering, 6, 10517–10525. [Google Scholar]
- Zitnak A, 1961. The occurrence and distribution of free alkaloid solanidine in netted gem potatoes. Canadian Journal of Biochemistry and Physiology, 39, 1257–1265. [DOI] [PubMed] [Google Scholar]
- Zwenger C and Kind A, 1861. Ueber das Solanin und dessen Spaltungs‐producte. Ann. Chem. Pharm., 118, 129–151. [Google Scholar]