

Abstract
Introduction:
In patients with OSA, substantial increases in genioglossus (GG) activity during hypopneas/apneas usually fail to restore normal airflow. The present study was undertaken to evaluate if this phenomenon can be explained by reduced activation of other peri-pharyngeal muscles.
Methods:
We recorded EMGs of the GG and four other peri-pharyngeal muscles (accessory dilators, AD), in 8 patients with OSA and 12 healthy subjects, during wakefulness and sleep. Repetitive events of flow limitation were induced during sleep. The events with the highest increases in AD activity were evaluated, to assess if combined activation of both the GG and AD to levels higher than while awake ameliorate airflow reduction during sleep.
Results:
Flow limitation triggered large increases in GG-EMG, but only modest augmentation in AD activity. Nevertheless, phasic EMG activity was present in 40 % of the ADs during sleep. In multiple events, increases of both GG and AD activity to levels substantially higher than while awake were not associated with improvement in airflow.
Conclusions:
We conclude that sleep-induced reduction in AD response to airway obstruction cannot completely explain the failure of upper airway dilators to maintain pharyngeal patency. We speculate that reduction in dilator muscle efficacy may be due to the alterations in motor units recruitment patterns during sleep.
Keywords: Obstructive sleep apnea, Upper airway, Genioglossus, Upper airway dilator muscle
1. Introduction
Obstructive sleep apnea (OSA) is a common disorder characterized by repetitive pharyngeal obstructions during sleep. The human pharyngeal airway is collapsible and tends to obstruct above (OSA patients) or below (healthy subjects) atmospheric pressure in the absence of active dilator muscle force (Pham and Schwartz, 2015; Younes, 2003). Obesity, soft tissue and bony abnormalities and other structural alterations play key roles in the pathogenesis of OSA. However, all OSA patients manage easily to maintain adequate airflow during wakefulness, thereby compensating for the many anatomic alterations implicated in the pathogenesis of OSA. Therefore, it is obvious that sleep-induced changes in pharyngeal neuromuscular control is most relevant and combines with anatomic characteristics to determine pharyngeal airflow mechanics during sleep (Younes, 2003; Patil et al., 2007).
While most anatomic alterations contributing to sleep apnea are relatively easy to comprehend, the neuromuscular factors are more complex and difficult to explain. The human pharynx is characterized by instability and varying levels of collapsibility that depend on the level of consciousness: during wakefulness, reflex activation of upper airway dilator muscles prevents pharyngeal collapse and maintains adequate upper airway patency (Malhotra et al., 2000; Doherty et al., 2008). During sleep, these protective mechanisms are deranged (Eckert et al., 2007), and pharyngeal stability of patients with OSA is waxing and waning, moving between “active”, relatively stable periods, with little or no disturbance to airflow, and periods of “passive” state with flow limitation up to complete obstruction (McGinley et al., 2008). Anesthesia increases pharyngeal collapsibility relative to sleep and causes a dose-dependent increase in pharyngeal critical pressure (Pcrit) (Eastwood et al., 2005; Maddison et al., 2019), resulting in upper airway obstruction in almost every person (Oliven et al., 2008), as described also during the state of coma (Safar et al., 1959). Pharyngeal collapsibility depends also on sleep stage (Carberry et al., 2016). These observations suggest that neuromuscular protective mechanisms are present even in the “passive” state of sleep in healthy subjects, and contribute to their negative Pcrit. However, this activity is insufficient in OSA patients, due to their increased structural load and possibly also secondary to poorly understood neuromuscular control mechanisms (McGinley et al., 2008).
Since the pioneering studies of Remmers et al. it is believed that the forces preventing pharyngeal obstruction are produced primarily by the upper airway dilator muscles (Remmers et al., 1978). Numerous studies have shown that these muscles exhibit respiratory activity that rises with increasing respiratory drive, when higher tone is needed to prevent pharyngeal obstruction (Van Lunteren and Strohl, 1986). The diminution of the reflex response of the genioglossus (GG) to negative pressure (Wheatley et al., 1993), chemical drive (Pillar et al., 2000) and resistive load (Pillar et al., 1997) during sleep, and the reduction in GG EMG activity with the onset of sleep (Sauerland and Harper, 1976; Mezzanotte et al., 1996; Fogel et al., 2005) suggested that the increase in pharyngeal collapsibility during sleep is due primarily to sleep-associated decrease in dilator muscle activity.
However, it is well documented that the decline in dilator EMG activity at sleep onset does not persist throughout sleep, and shortly after sleep onset GG activity begins to rise again and recovers to waking (or slightly greater) levels within a few breaths (Remmers et al., 1978; Onal et al., 1980; Tangel et al., 1992; Okabe et al., 1993; Berry et al., 1997; Worsnop et al., 1998; Younes et al., 2012). Pharyngeal obstruction is known to trigger an increasing drive to the respiratory muscles, and a gradually increasing GG EMG activity could be documented even in the first few breaths of flow limitation (Schwartz et al., 1998). Nevertheless, this enhanced activity during hypopneas and apneas usually fails to restore pharyngeal patency. The cause for this phenomenon remained unclear. Remmers et al. suggested that the dilator muscles fail to counteract the large negative intraluminal pressures that develop at and downstream to the obstruction (Remmers et al., 1978). It was also suggested that the GG was unable to maintain pharyngeal patency due to a decrease in activity of other upper airway muscles that do not exhibit phasic activity (“tonic muscles”) (Tangel et al., 1992; Fogel et al., 2004; Jordan and White, 2008). More recently, Younes et al. suggested that GG activation during apneas and hypopneas fails to reach a threshold required to prevent collapse (Younes et al., 2012).
We have recently conducted several studies that challenged these explanations (Dotan et al., 2013, 2015; Oliven et al., 2018). In spontaneously breathing OSA patients under mild propofol anesthesia we observed that even dramatic increases in GG tonic and phasic activity, along with lower phasic activity of other peri-pharyngeal muscles, elicited by prolonged flow limitation, did not increase pharyngeal luminal size (pharyngoscopy) or inspiratory airflow. In contrast, immediately upon awakening from anesthesia, the pharynx enlarged and airflow normalized, despite concomitant decreases in dilator muscle activity to minimal levels (Dotan et al., 2013). In two later studies we found the same phenomenon also during sleep, both in OSA patients (Dotan et al., 2015; Oliven et al., 2018) and healthy subjects (Oliven et al., 2018): while awake, breathing through external loads was associated with increases in the activity of all dilator muscles. During sleep, upper airway obstruction was associated with progressive increases in GG activity, which exceeded substantially the level of activity observed at equal levels of negative esophageal pressure during wakefulness. In contrast, the activity of other peri-pharyngeal muscles did not respond at all to sleep-induced flow limitation, or increased much less than the GG. We concluded that the coordinated activation of lingual protrusor, retractors and other peri-pharyngeal muscles was no longer preserved during sleep. As coordinated recruitment and activation of several peri-pharyngeal muscles is required to maintain pharyngeal patency to airflow, our findings suggested that the deficient recruitment of ADs impedes muscle synchronisation during sleep and might play a role in the failure of the activated GG to stabilize the pharynx and prevent flow limitation.
In our previous articles (Dotan et al., 2015; Oliven et al., 2018) we focussed our analysis on GG EMG and limited our analysis to the events with the highest excitation of the GG. This approach was undertaken because the primary goal of these studies was to question the abovementioned assumptions that failure of the GG to alleviate flow limitation during sleep is due to insufficient rise in activity required to overcome large negative intra-pharyngeal pressures (Remmers et al., 1978) or to reach wakefulness levels (Younes et al., 2012). The present work was undertaken to assess the role of muscles other than the GG in the pathogenesis of pharyngeal collapse during sleep. In contrast to the GG, the main pharyngeal dilator (Odeh et al., 1995), we labelled other peri-pharyngeal muscles that may dilate and/or stabilize the pharynx as accessory dilators (AD). We hypothesised that if pharyngeal collapse in the presence of increased GG activity is due to deficient recruitment of AD, sufficient co-activation of AD during sleep, even occasionally, should prevent or ameliorates flow limitation. We therefore re-analyzed the experimental recordings used for our previous study (Oliven et al., 2018), with special emphasis placed on activation patterns of AD. Reviewing all sleep studies in OSA patients and healthy subjects, we now searched for events of flow limitation during which AD were recruited and became more active in the course of sleep than during wakefulness, and examined the flow-mechanical effect of such co-activation of the GG and AD.
2. Methods
The methods used in this study were previously described in detail (Oliven et al., 2018). Briefly, subjects, all males, were 8 patients with severe OSA and 12 healthy subjects (7 age-matched to the OSA patients and 5 young subjects), all previously diagnosed in a conventional fullnight sleep study in the Technion Sleep Laboratory. Anthropometric and sleep data of the study subjects are given in Table 1. All studies were performed in the respiratory research laboratory of Bnai-Zion Medical Centre. The aims and potential risks of the study were explained, and informed consent was obtained from all subjects. The study was approved by the institutional Human Investigations Review Board.
Table 1.
Anthropometric and sleep data of the study subjects. AHI and SO2 data were obtained from conventional sleep studies performed before the current study.
| OSA (n = 8) | healthy | ||
|---|---|---|---|
| older (n = 7) | young (n = 5) | ||
| AHI | 58.4 ± 9.4 | 7.6 ± 2.0*** | 3.8 ± 1.2*** |
| BMI | 33.5 ± 4.8 | 27.2 ± 3.5* | 23.6 ± 1.5** |
| age | 53.6 ± 8.7 | 52.6 ± 10.0 | 21.2 ± 0.4*** |
| lowest SO2 | 75.4 ± 8.5 | 90.4 ± 2.1*** | 93.8 ± 1.1** |
| %time SO2 < 90 % | 18.9 ± 19.1 | 0.2 ± 0.5** | 0* |
| Pcrit | 1.98 ± 1.4 | −6.2 ± 3.1*** | −5.7 ± 0.9*** |
AHI – apnea/hypopnea index. SO2 – oxygen saturation.
p < 0.05;
p < 0.01;
p < 0.001 for comparison with OSA.
EMGs were recorded via pairs of Teflon coated hook-wire electrodes as previously described (Oliven et al., 2018). In addition to the GG, four additional AD were evaluated: styloglossus (SG), geniohyoid (GH), sterno-cleido-mastoid (SCM) and sternohyoid (SH). Electrodes were positioned under direct vision and palpation during volitional muscle contraction and (for GH and SH) based also on ultrasound guidance. Adequate position of all electrodes was verified by the presence of muscle contraction during short bursts of 40 Hz electrical stimulation (Dynex III, Medtronic Inc, Minneapolis, MN). The GG and 1–2 additional muscles were studied in each subject. EMG signals were amplified using P122 amplifiers and band-pass filtered at 30–1000 Hz (Grass Technologies, Warwick, RI), and a 50 Hz notch filter was applied. Raw EMG signals were rectified and processed with leaky integrators with a time constant of 100 ms to yield an averaged EMG envelop. All EMG signals were expressed as % of the maximal value obtained for each muscle, as previously described (Oliven et al., 2018). C4-A1 and C3-A2 EEG were employed to monitor sleep. Breathing was monitored using a pneumotachometer mounted on a mouthpiece or nose mask during wakefulness and sleep, respectively. Flow signals were recorded without filters to document the vibrations characteristic to flow limitation and snoring (Beck et al., 1995), and filtered later on (5 Hz low pass) to enable measurement of flow. Intrathoracic pressure (Pes) was measured with a Millar catheter, and nasal pressure (Pn) was monitored with a catheter connected to a side port of the mask. Analogue-to-digital acquisition of all parameters was performed for monitoring and data storage on a digital polygraphic data acquisition system (LabVIEW, National Instruments, Austin TX).
The study protocol involved recording of respiratory and EMG activity during wakefulness and sleep. During sleep, variable levels of positive (CPAP) and negative (CNAP) continuous airway pressure to patients with OSA and healthy subjects, respectively, were applied to induce events of flow limitation. Flow levels at the 4th breath after Pn drop were used to calculate Pcrit. Patients fell asleep on CPAP levels sufficient to prevent flow limitation, and multiple pressure drops were performed to levels at which flow limitation triggered high increases in EMG activity without arousal.
The integrated EMG signal during expiration (average activity between the post-inspiratory inspiratory activity and the rise of activity before the next inspiration) was considered as tonic activity, and peak inspiratory activity represented the sum of tonic and phasic activity. Wakefulness data was obtained by averaging 6 breaths during quiet, undisturbed breathing. To characterize the maximal activation of AD during sleep, we averaged for each subjects the 3 highest EMG levels of these muscles observed during the events of flow limitation, as well as the concomitant activity of the GG. All data are presented as mean ± SD. Shapiro-Wilk Normality Test was used to test normal distribution of data. For normally distributed data, paired and unpaired t-tests were used to compare data between sleep and wakefulness and between subject groups and different muscles, respectively. For data not-normally distributed, Wilcoxon signed-rank and Mann-Whitney tests were used for paired and two-sample comparisons, respectively. Chi square test was used to compare binary variables. P < 0.05 was considered as statistically significant.
3. Results
The averaged EMG activity of the AD is presented in Table 2. During wakefulness, phasic activity was seldom observed, and low-level phasic activity was recorded in only 2 AD muscles of OSA and healthy subjects, respectively. During sleep-associated flow limitation EMG activity increased in 13 (41 %) of the AD studied. The increase in peak activity was statistically significant in each of the two neck muscles (p < 0.05). Phasic activity could be triggered by flow limitation in 12 AD muscles (38 %, p < 0.05 compared to wakefulness). This increase in phasic activity occurred similarly in OSA and healthy subjects. It was more commonly seen in the neck muscles (SH and SCM) (50 % vs. 31 % in the GH and SG, p = 0.11). Nevertheless, average tonic and peak activities during both wakefulness and sleep were not significantly different in the four AD. We therefore grouped the AD muscles together to enable comparison of the subject groups.
Table 2.
Averaged EMG levels observed in both OSA patients and healthy subjects, awake and before arousal during induced flow limitation, in the four accessory dilators studied. Differences between muscles in both tonic and peak activity, awake and asleep, were not statistically significant. Average peak activity triggered by flow limitation was slightly higher than during wakefulness, but this increase reached significance only for the SH.
| GH (n = 9) | SG (n = 7) | SH (n = 7) | SCM (n = 9) | ||
|---|---|---|---|---|---|
| awake | tonic | 4.1 ± 2.5 | 6.0 ± 2.6 | 3.1 ± 1.4 | 3.3 ± 1.2 |
| Peak inspiratory | 4.2 ± 2.5 | 6.7 ± 2.9 | 3.5 ± 1.1 | 3.4 ± 1.0 | |
| sleep | tonic | 3.8 ± 1.7 | 7.7 ± 8.9 | 4.8 ± 2.9 | 3.3 ± 1.2 |
| Peak inspiratory | 4.6 ± 2.1 | 8.9 ± 5.6 | 7.5 ± 4.11 | 4.4 ± 1.32 |
Data are presented as %max, mean ± SD.
GH – geniohyoid; SG – styloglossus; SH – stylohyoid; SCM – sterno-cleido mastoid.
p < 0.05 for the comparison of peak SH, awake vs. sleep.
p = 0.052 for the comparison of peak SCM awake vs. sleep.
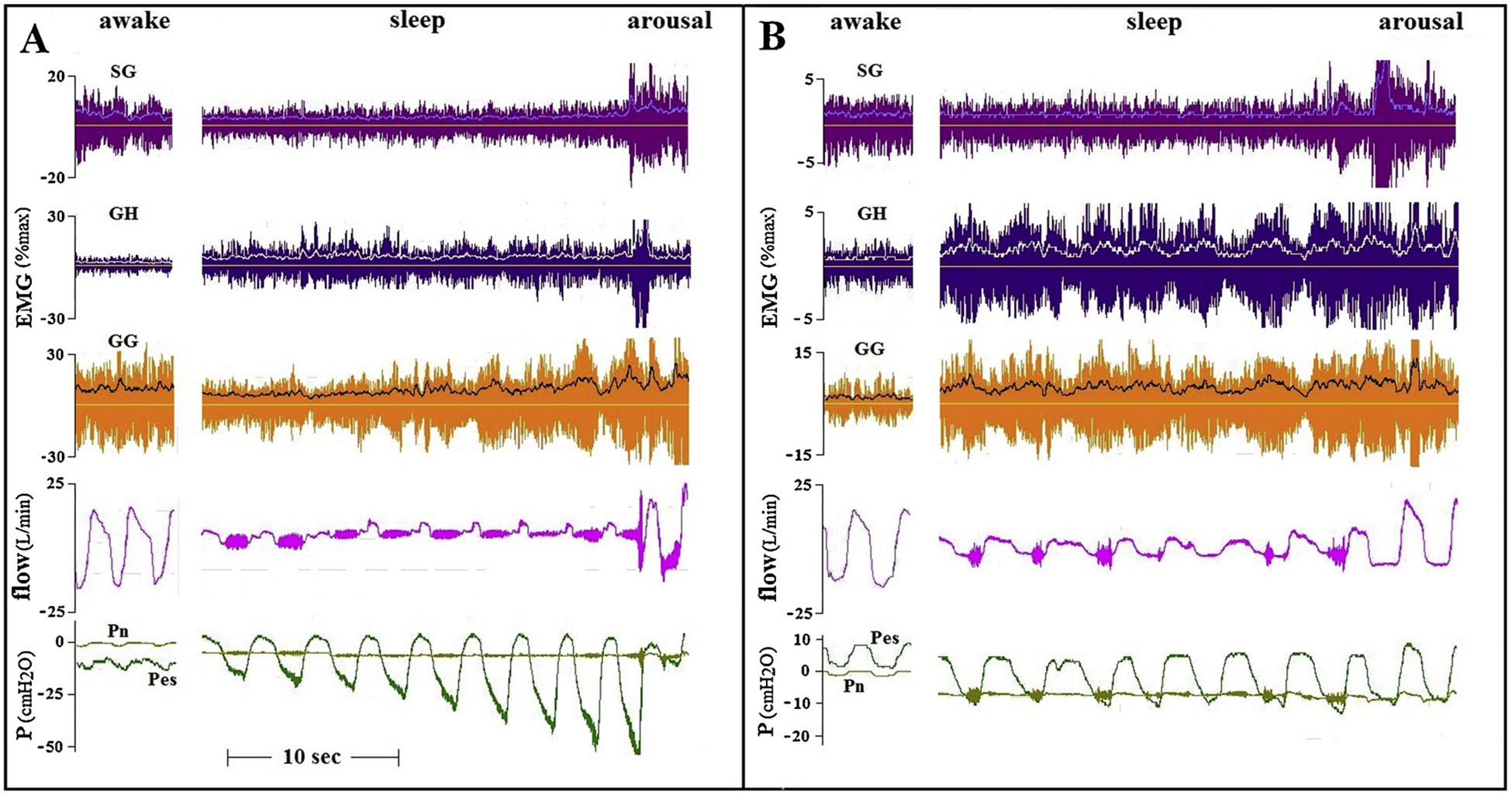
The most common finding during induced flow limitation was a marked increase in GG activity, with small or no increase in AD activity. Two examples are presented in Fig. 1. One OSA patient had phasic GG activity while awake, the other maintained upper airway patency with minimal tonic activity. Despite marked increases in GG activity and unchanged tonic activity of the ADs, substantial flow limitation occurred and persisted at relatively high CPAP.
Fig. 1.
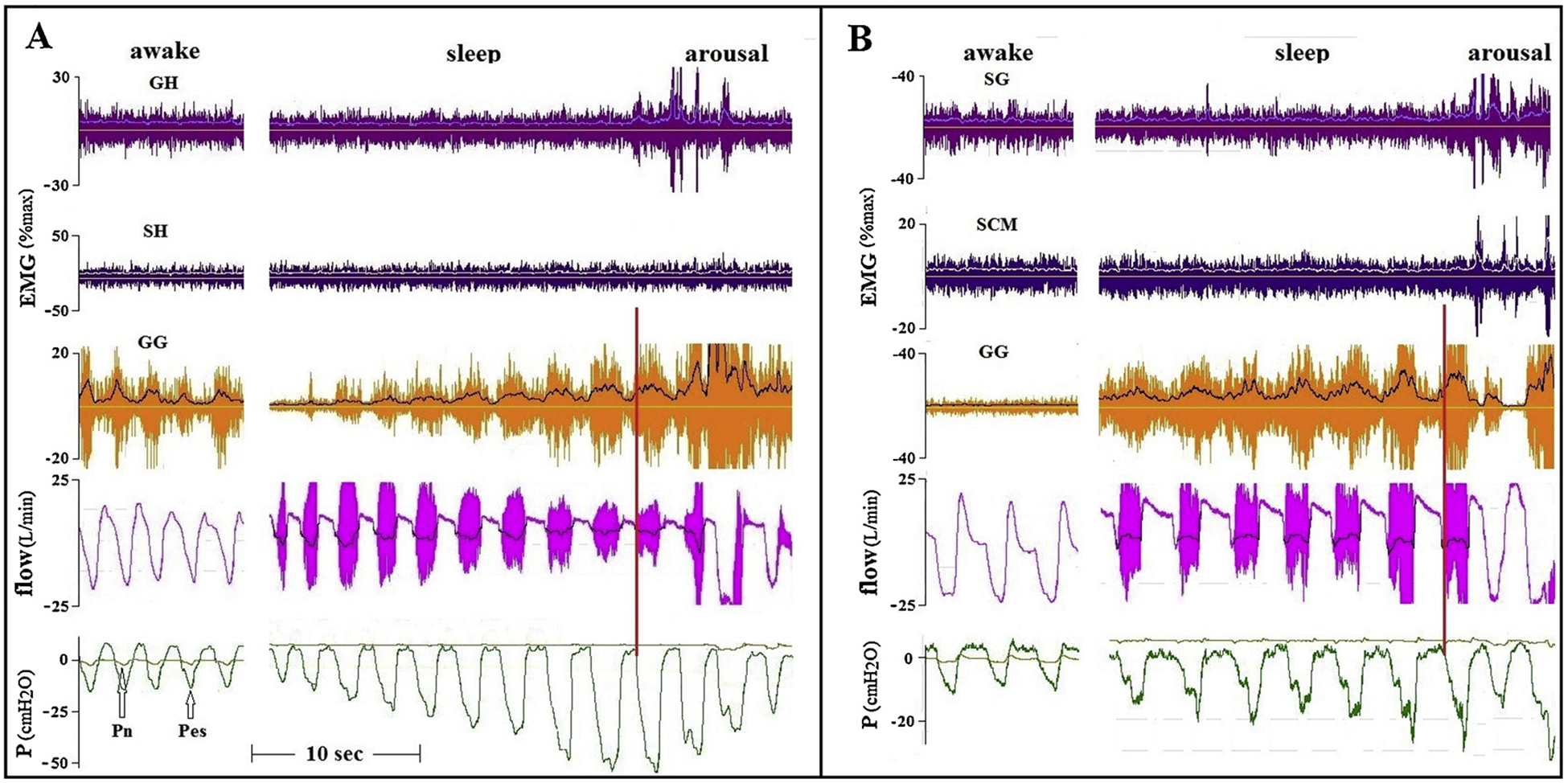
Tracings of two OSA patients, comparing GG and AD activity during wakefulness and sleep. EMG tracings present raw with superimposed integrated EMG tracings. Flow during sleep presents raw tracings showing marked vibrations (“snoring”), with superimposed filtered tracings. The CPAP of patient A (AHI 70, BMI39) was lowered from holding pressure to 7 cmH2O. In patient B (AHI 59, BMI 37) CPAP was lowered to 5 cmH2O. Note that in patient A, flow limitation was getting worse despite an increase in GG activity. In patient B flow increased immediately upon arousal, despite a marked reduction in GG activity. As indicated by the red vertical lines, flow limitation started after the initial increase in phasic GG activity, but well before the development of substantial negative intra-luminal (downstream) pressures. GH – geniohyoid; SH – sternohyoid; SG – styloglossus; SCM – sternocleidomastoid; GG – genioglossus; Pn – nasal (mask) pressure; Pes – esophageal pressure.
Average levels of EMG activity of the GG and AD and the presence of phasic activity are presented in Tables 3 and 4, respectively. Peak inspiratory flow during wakefulness was measured while subjects breathed at ambient pressure. The levels during sleep were measured before arousal from apneas/hypopneas, induced by low-level CPAP in the OSA patients, and continuous negative airway pressure (CNAP) in the healthy subjects. In contrast to the AD, peak GG activity during relaxed breathing in awake OSA patients was significantly higher than in healthy subjects (p < 0.05), and 4 of the patients exhibited phasic activity, as compared to one healthy subject (p < 0.05). During sleep-associated flow limitation, however, GG activity increased significantly and phasic activity was present in all OSA patients and healthy subjects. In contrast, peak AD activity increased significantly during sleep-associated flow limitation, compared to wakefulness, only in the OSA patients. The increase in the number of AD exhibiting phasic activity during flow limitation did not reach statistical significance. Consequently, peak inspiratory GG activity was significantly higher than that reached by the AD muscles. Of note, the marked flow-limitation-induced increase in both tonic and phasic GG activity, and some increases in the activity of the AD during sleep, as compared to wakefulness, was not associated with relief of severe flow limitation. Also, there was no significant difference in the level of activity of either the GG nor the AD during sleep when comparing OSA patients to healthy subjects.
Table 3.
Tonic and peak-inspiratory EMG levels of the genioglossus (GG) and the evaluated accessory dilators (AD), during wakefulness and sleep. Increases in GG activity during sleep-associated flow limitation, compared to wakefulness, were significant in each of the three groups, both during expiration (tonic activity, p < 0.05) and during inspiration (peak activity, P < 0.005). AD peak activity increased significantly only in the OSA patients. Also, in each of the three groups, GG peak activity reached during sleep was higher than that of the AD (p < 0.001)
| OSA (n = 8) | HO (n = 7) | HY (n = 5) | Flow (L/min) | |||||
|---|---|---|---|---|---|---|---|---|
| GG (n = 8) | AD (n=14) | GG (7) | AD (11) | GG (5) | AD (10) | |||
| awake | tonic | 5.0 ± 2.7 | 3.6 ± 1.2 | 3.3 ± 1.0 | 3.3 ± 1.6 | 5.2 ± 1.9 | 6.5 ± 3.6 | 24.5 ± 2.9 |
| peak | 7.4 ± 4.1 | 3.7 ± 1.1 | 3.4 ± 0.91 | 3.5 ± 1.4 | 5.2 ± 1.9 | 7.1 ± 3.5 | ||
| sleep | tonic | 9.0 ± 5.0 | 4.3 ± 1.52 | 6.4 ± 2.6 | 3.8 ± 2.8 | 10.4 ± 2.7 | 7.2 ± 5.4 | 8.4 ± 2.8 |
| peak | 30.6 ± 13.7 | 5.9 ± 3.03 | 19.0 ± 8.3 | 5.3 ± 3.6 | 24.2 ± 4.7 | 8.3 ± 4.8 | ||
Data are presented as %max, mean ± SD. OSA – patients with obstructive sleep apnea. HO – healthy older subjects, age matched to the OSA patients. YO – healthy young subjects.
p < 0.05 for the comparison of peak GG activity during wakefulness, OSA vs. OH.
p < 0.005 for the comparison of tonic activity during sleep, GG vs. AD.
p < 0.05 for the comparison of peak AD activity in OSA patients, wakefulness vs. flow limitation during sleep.
Table 4.
Presence of phasic activity in OSA patients and healthy subjects, during wakefulness and induced flow-limitation during sleep.
| group | OSA | healthy | ||
|---|---|---|---|---|
| muscles | GG (n = 8) | AD (n = 14) | GG (n = 12) | AD (n = 21) |
| awake | 4 (50 %)1 | 1 (7 %) | 1 (8 %) | 3 (14 %) |
| sleep | 8 (100 %)2 | 6 (43 %)3 | 12 (100 %)4 | 8 (38 %) |
GG – genioglossus; AD – accessory dilators.
p < 0.05, for the comparison of GG activity during wakefulness, OSA vs. healthy subjects.
p < 0.05, for the comparison of GG activity of OSA patients, awake vs. sleep.
p < 0.02, for the comparison of AD activity of OSA patients, awake vs. sleep.
p < 0.01, for the comparison of GG activity of healthy subjects, awake vs. sleep.
Our main emphasis in the current work was to identify subjects with events that were associated with substantial increases in AD activity during flow limitation (in addition to the increase in GG activity that could be triggered in all subjects), and assess the flow response to this co-activation. Figs. 2 and 3 present tracings of an older and a young healthy subjects in whom flow limitation was induced by the application of continuous negative airway pressure (CNAP). In these subjects, a relationship between increases in GG and AD activity and airflow seems to be present, abolishing pharyngeal vibrations associated with flow limitation, although this increased activity was not always associated with the restoration of normal airflow.
Fig. 2.
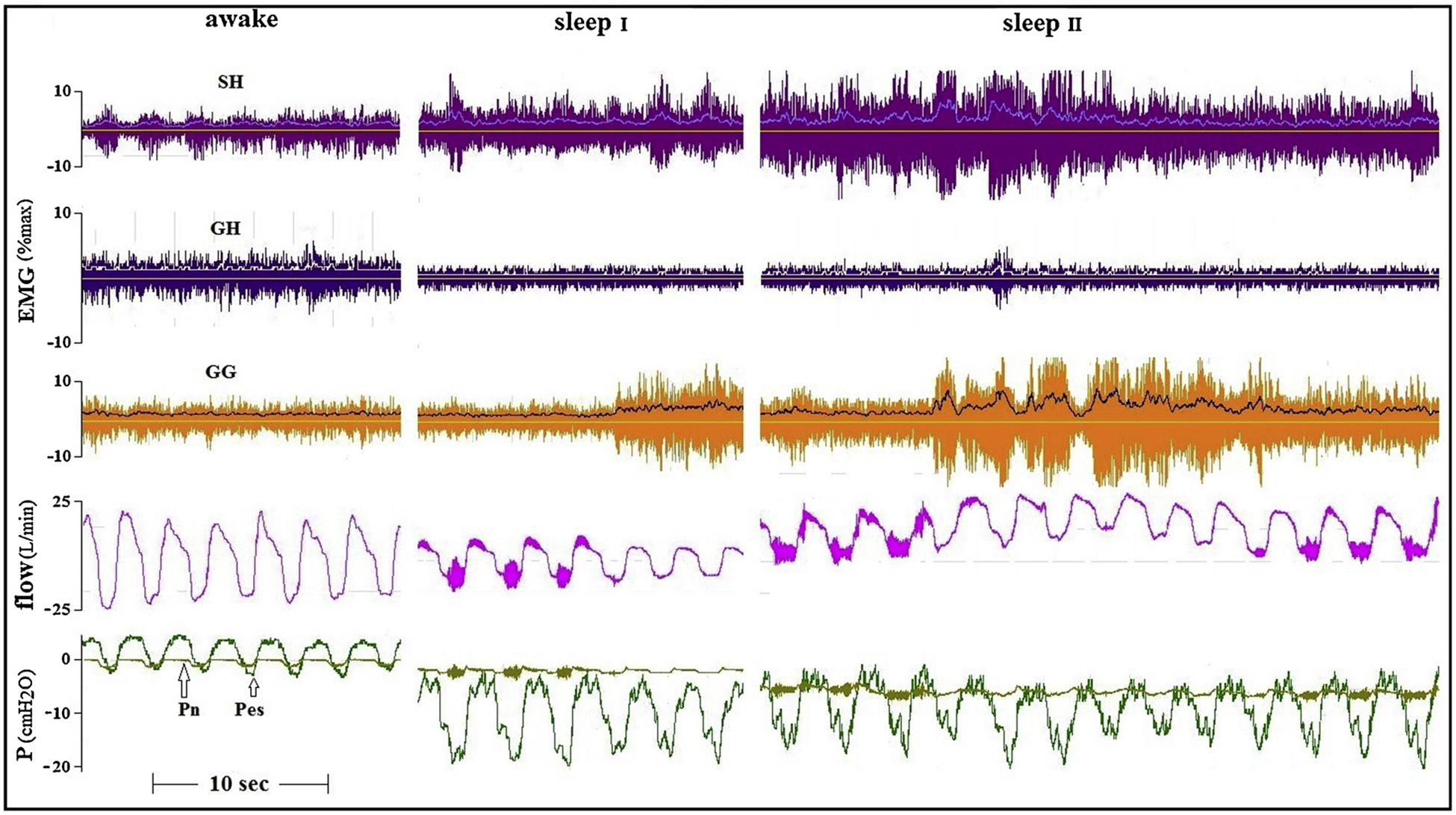
Awake and two sleep tracings of an older (40 yrs) healthy subject. Flow limitation was produced by applying CNAP. Flow vibrations disappear about when GG and SH activity increase, but flow limitation persists.
Fig. 3.
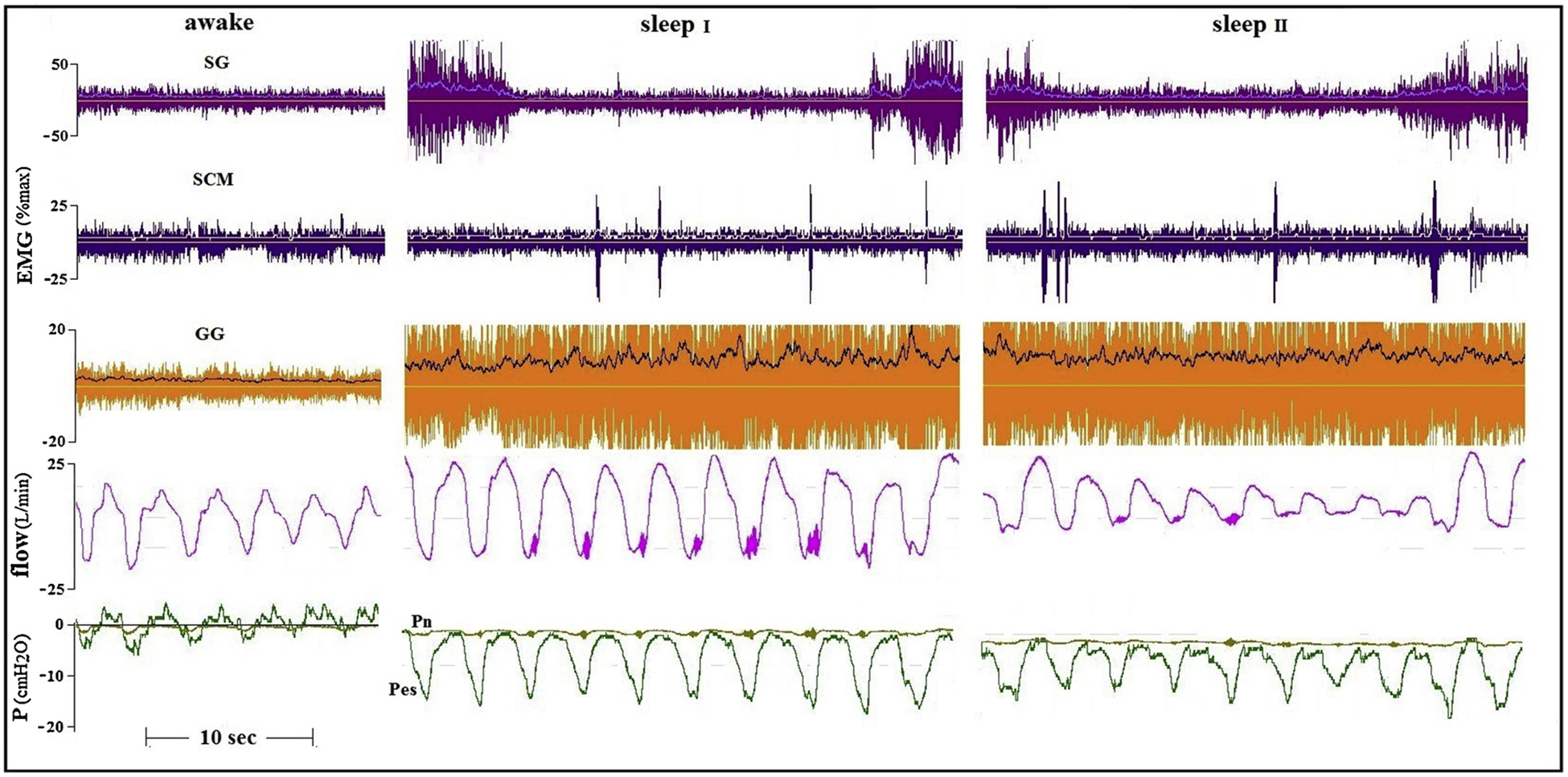
Awake and two sleep tracings of a younger (21 yrs) healthy subject. In this subject CNAP mainly increased tonic activity of the GG. However, only the intermittent co-activation of the SG was associated with disappearance of flow vibrations (sleep I, Pn= −2cmH2O) and also abolished flow limitation (sleep II, Pn= −4cmH2O) without arousal.
However, a possible association between increases in peri-pharyngeal muscle EMG activity and airflow during sleep was found only in these two subjects, and in none of the OSA patients. In contrast, in all other subjects in whom increases in AD activity could be triggered during sleep (Table 4), no such relationship was found in multiple trials. Examples are presented in Figs. 4 and 5.
Fig. 4.
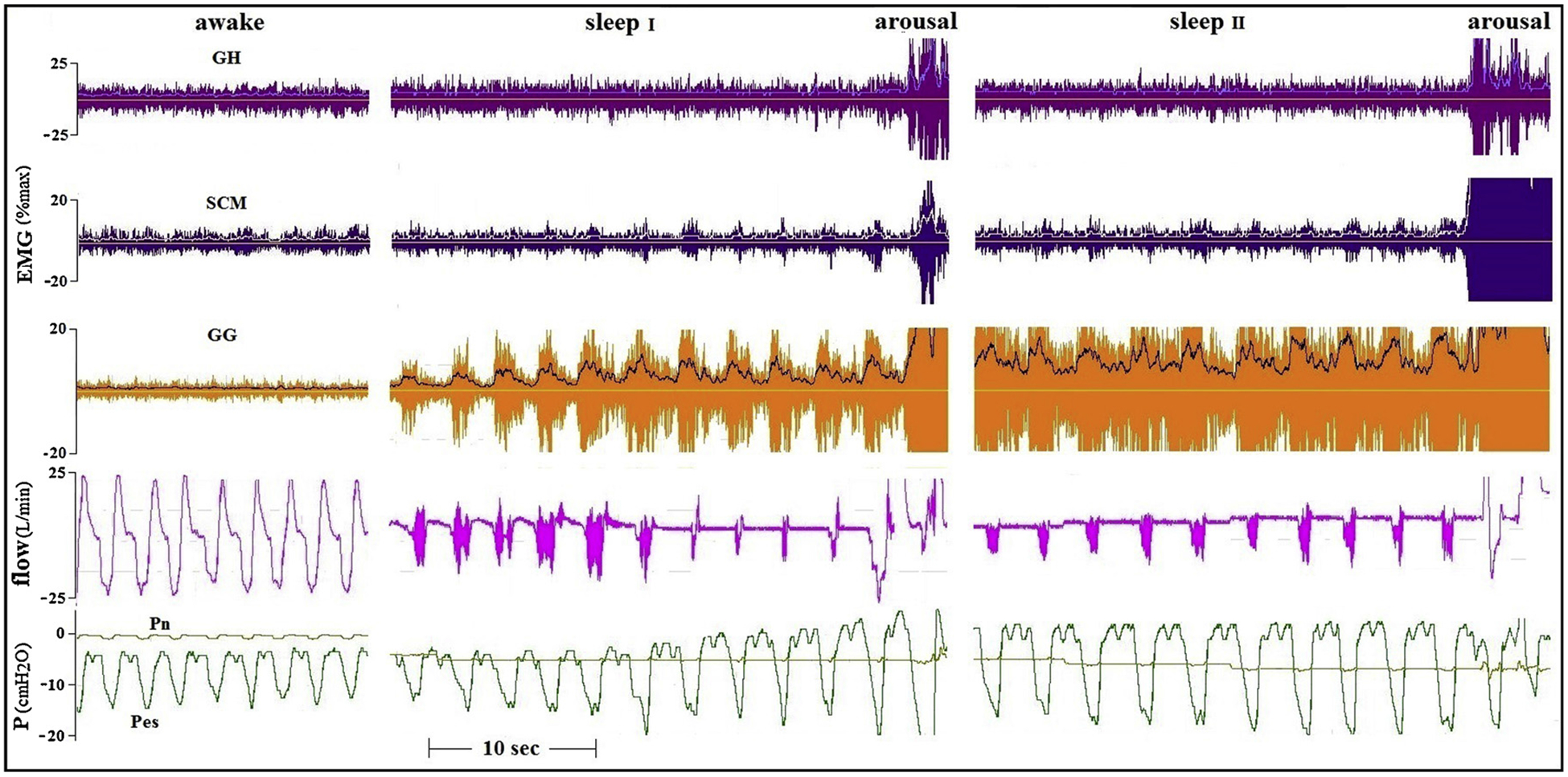
Tracings of a 22 years old healthy man, in whom the application of CNAP produced severe flow limitation. The presence of phasic SCM activity did not help the large rise in GG activity to ameliorate airflow.
Fig. 5.
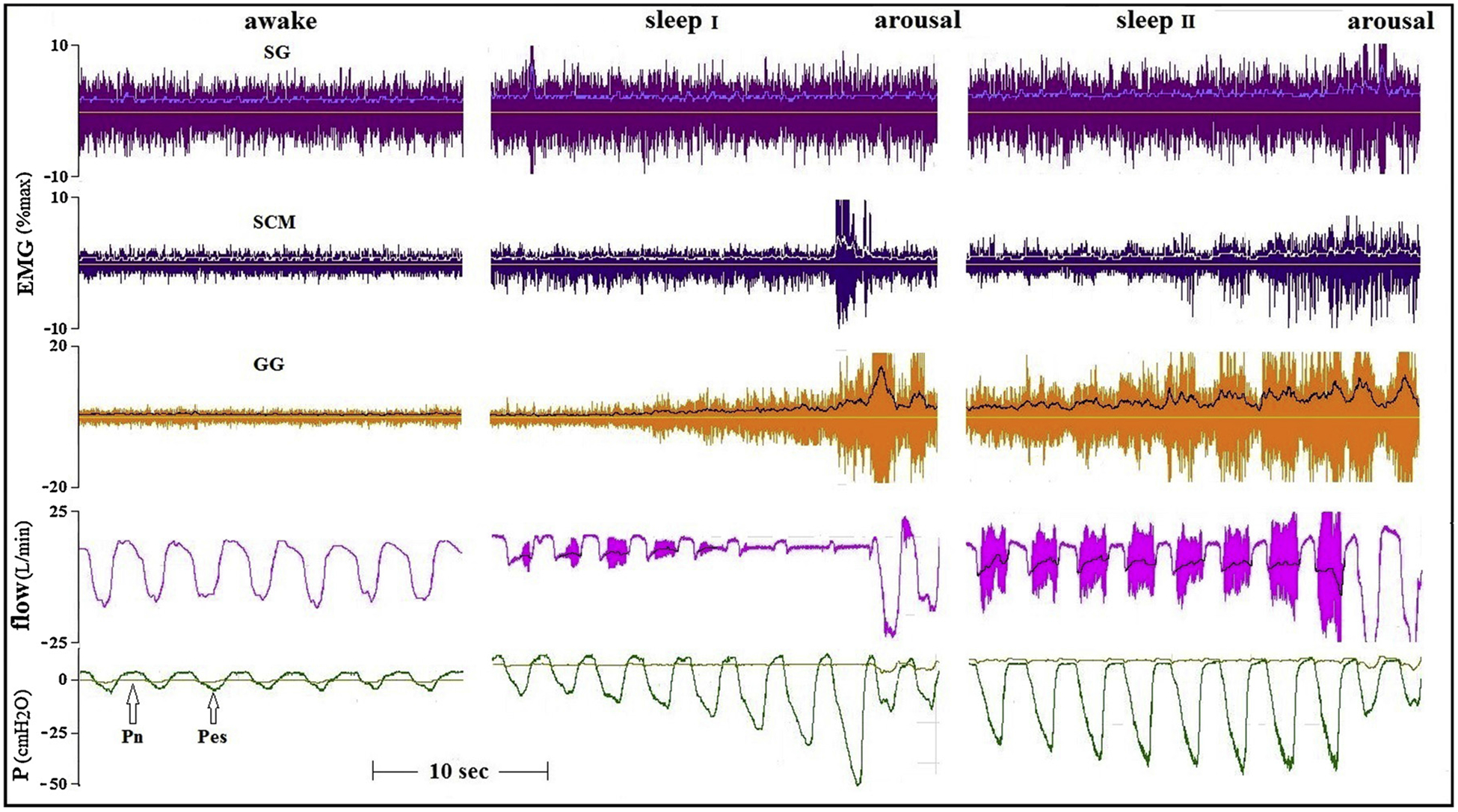
Tracings of a patient with sever OSA (AHI 56, BMI 39.7) recorded while awake and with CPAP of 6 (sleep I) and 8 (sleep II) cmH2O. Increases in tonic activity of the SG and phasic activity of SCM, in addition to the marked increase in GG activity, were not associated with any improvement in flow.
Both in healthy subjects and in OSA patients, the occurrence or increase in phasic activity of AD during induced hypopnea, in addition to high GG activity, did not increase airflow. In fact, even large phasic co-activation of two AD did not contribute to the large augmentation in GG activity and did not affect airflow (Figs. 6 and 7).
Fig. 6.
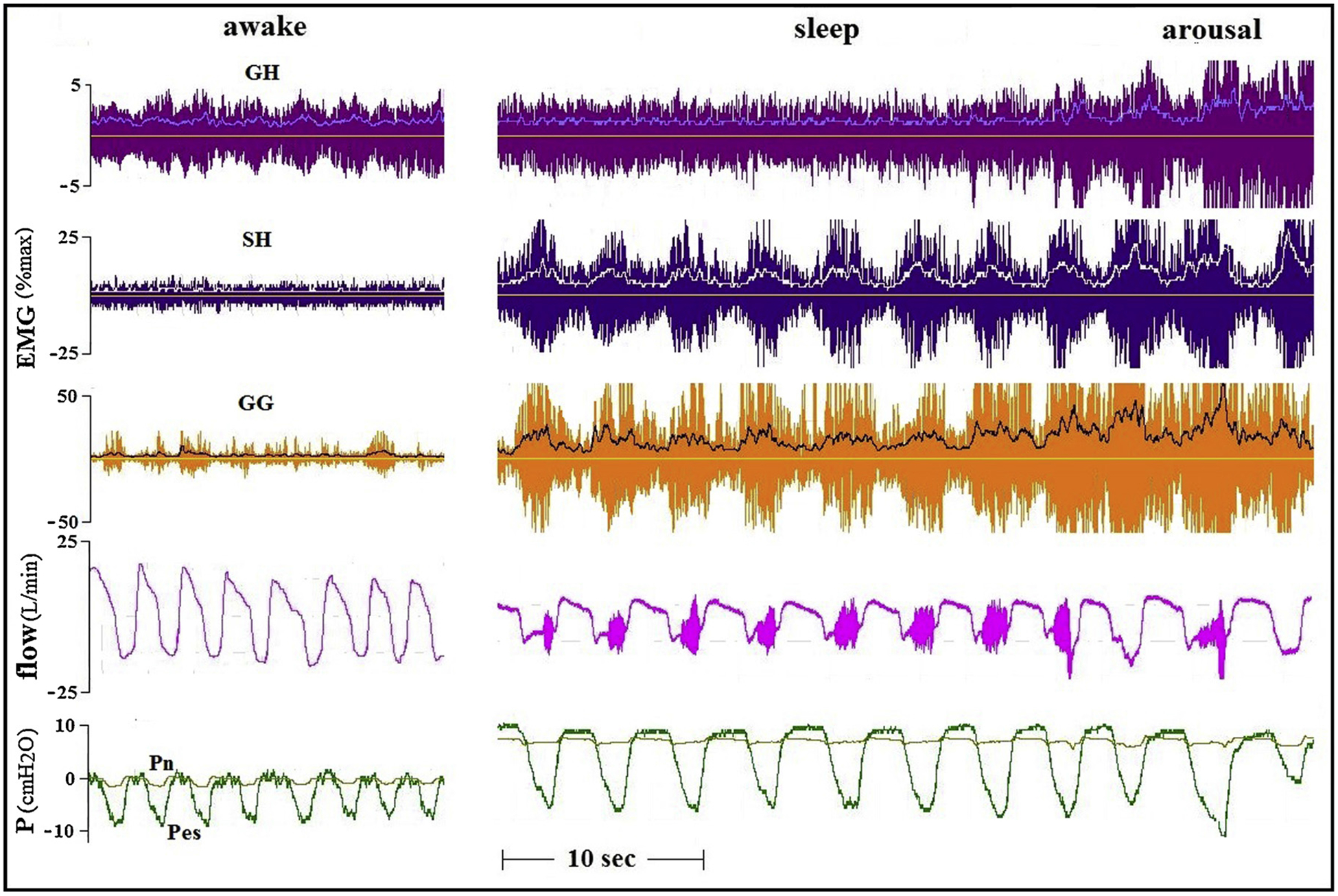
Tracings of an OSA patient (AHI 46, BMI 27.2) recorded while awake and with CPAP of 7 cmH2O. Large GG and SH activity was not associated with any change in flow limitation. It is possible that the appearance of phasic GH activity did have some flow-mechanical effect in this event.
Fig. 7.
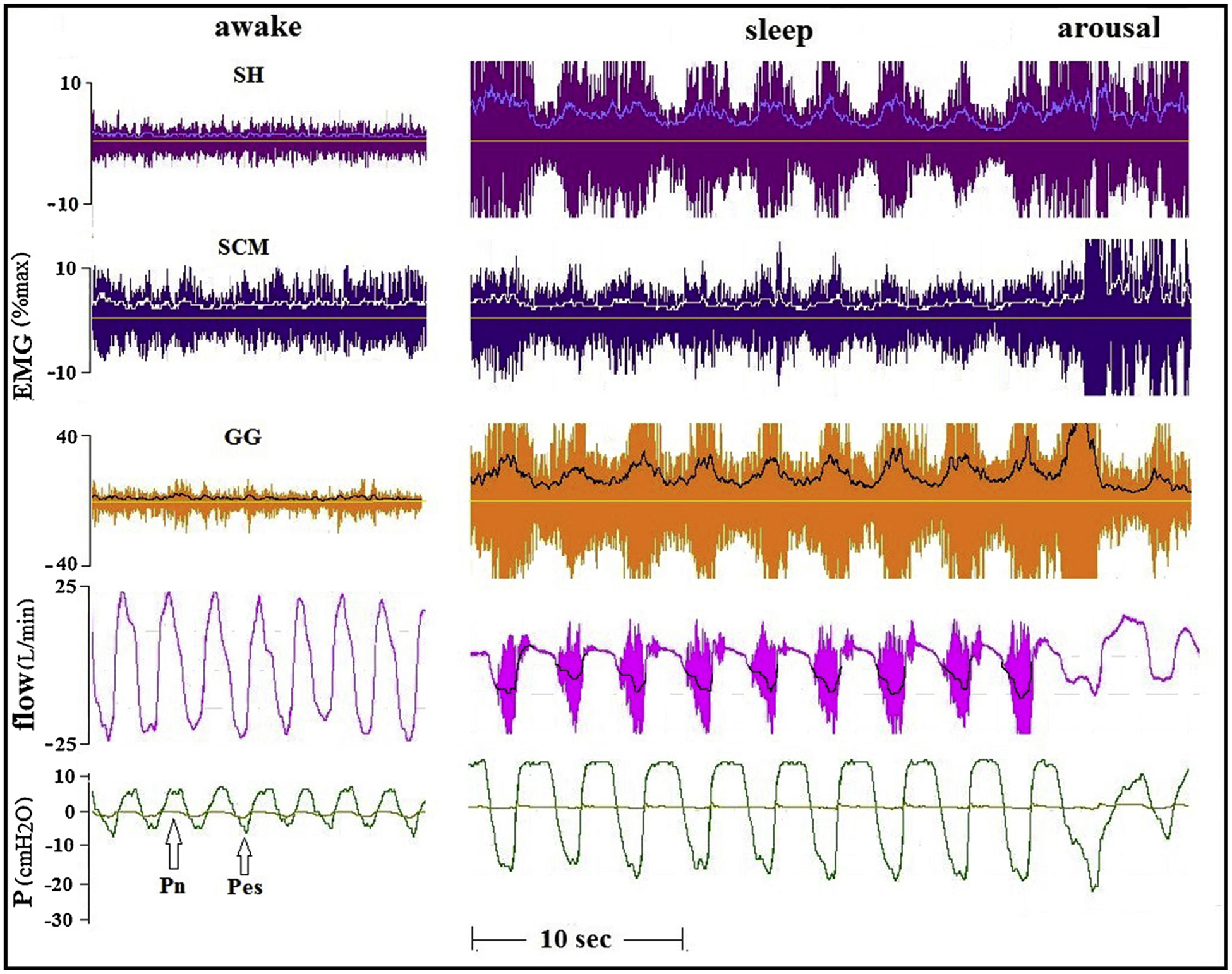
Tracings of another OSA patient (AHI 49, BMI 28.4) recorded while awake and during sleep with CPAP of 1 cmH2O. Large phasic activity of all muscles studied in this patient were not associated with any lessening in the severity of inspiratory flow limitation.
4. Discussion
Fig. 8 depicts a schematic presentation of the study design and main findings. This article focuses on the activity of upper airway dilators other than the GG (AD) and their recruitment and flow-mechanical effect when the pharynx collapses during sleep. The main findings are:1. The AD are recruited less than the GG during sleep-associated flow limitation; 2. Even when AD are activated to levels higher than while awake and exhibit phasic activity, this combined co-activation of AD and GG is not usually associated with improvement in airflow obstruction during sleep.
Fig. 8.
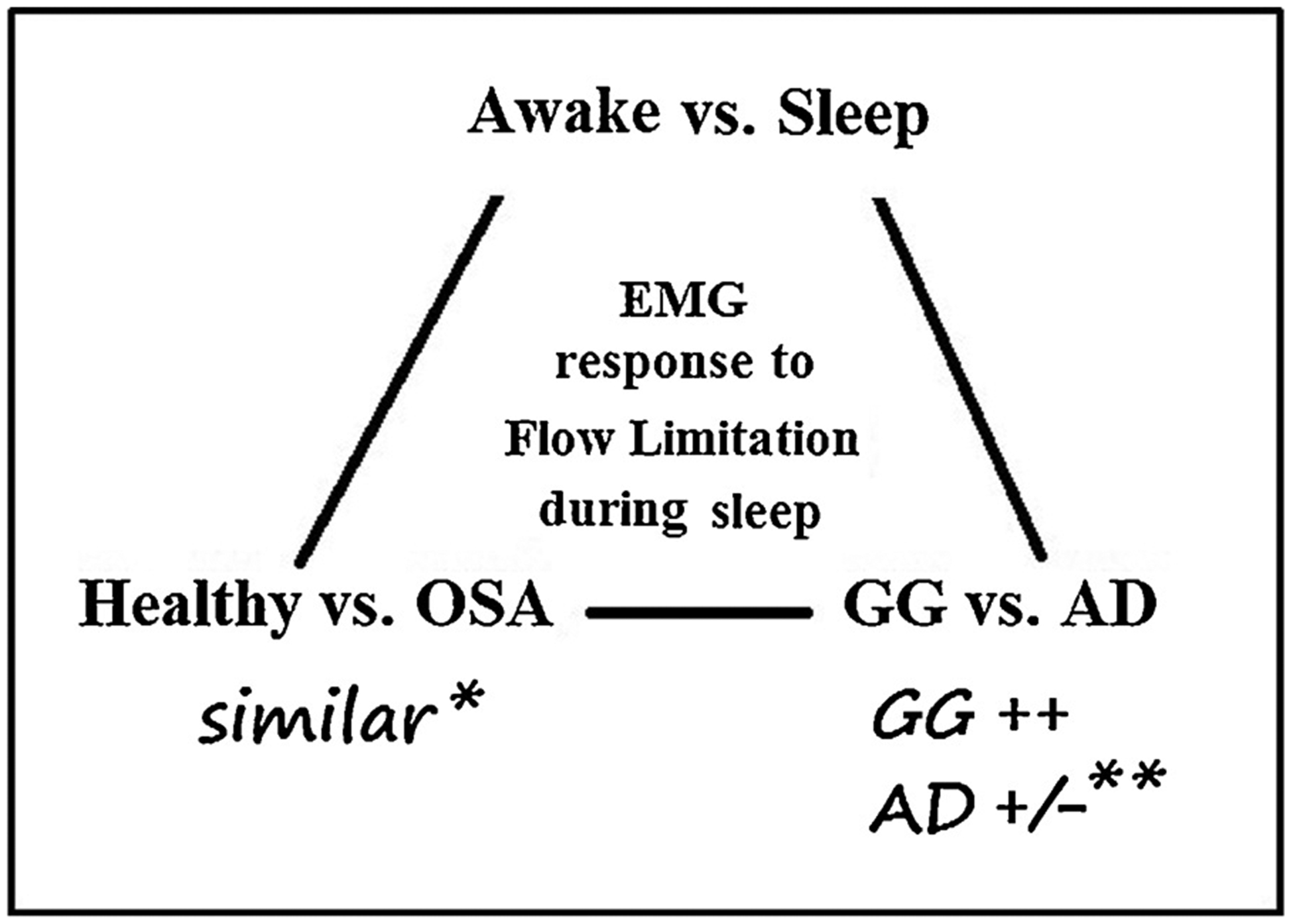
Schematic presentation of the study design and main findings. * - after adjustment of Pn to compensate for the difference in collapsibility (Pcrit). ** - main finding of this study: even when both GG and AD EMG activity was higher than while awake, it usually did not prevent or ameliorate flow limitation.
It has been known for many years that many peri-pharyngeal muscles may exhibit phasic activity during respiratory stimulation (Worsnop et al., 1998). Therefore, it is well accepted that all these muscles are involved in the maintenance of pharyngeal patency in the face of the sub-atmospheric intraluminal pressures produced during inspiration (Pham and Schwartz, 2015; Younes, 2003). Nevertheless, most of the experimental work carried out to explore control of pharyngeal patency in human subjects has centered on evaluating respiratory activity of the main tongue protrusor, the GG, and most other peri-pharyngeal muscles remained underexplored. We recorded EMGs from two muscles innervated, like the GG, by the hypoglossus nerve (GH and SG), and two neck muscles (SH and SCM), and found that AD muscles are activated during sleep much less than during wakefulness in the presence of increased airway resistance (Oliven et al., 2018). As seen in Table 3, although we selected only events with the highest activation of AD, their activity is considerably and significantly lower than that of the GG (expressed as %max) during the same flowlimited events. In fact, the increase in AD activity during hypopneas reached significance only in the OSA patients. Another interesting finding was that it was not rare to find only tonic activity most of the time in relaxed awake patients with severe OSA. As shown in Table 4, half of the OSA patients had no phasic activity while awake. Nevertheless, we found that peak inspiratory activity in OSA patients was, on the average, higher than in the healthy subjects, as previously reported, and phasic activity was more frequent. A partial explanation for the low awake EMG activity could be that the patients were breathing through a mouthpiece, and that in the present study our goal was to demonstrate that during wakefulness pharyngeal patency could be maintained with low EMG activity of the peri-pharyngeal muscles, while substantially higher activity failed to prevent flow limitation during sleep.
The finding that, on the average, the four AD studied reached similar peak activity during flow limitation does not imply that all peri-pharyngeal muscles are controlled similarly. It has been previously described that the activity of neurons and upper airway muscles that exhibit phasic respiratory activity while awake tends to be preserved during sleep, while the activity of tonic muscles tends to decline (Tangel et al., 1992; Orem et al., 1985). In the present study, the division of upper airway muscles into phasic and tonic was less clear, as the GG often exhibited only tonic activity during wakefulness, and phasic activity could be triggered by flow limitation in all AD in some of the patients. In addition, when stimulated by flow limitation, all ADs could be activated, in some of the patients, to levels higher than during wakefulness (Figs. 2,4,5,6). We found that whenever phasic activity appeared in the GH and SG in the course of induced flow limitation (n = 6), it started always after phasic GG activity was already present. In contrast, whenever the neck muscles exhibited phasic activity (n = 8), it started with or even before GG phasic activity. This finding suggests variable thresholds for recruitment of different AD.
The lack of improvement in flow in the face of increasing GG activity can also be seen in tracings of several recently published studies that evaluated the response of GG EMG to induced flow limitation (Carberry et al., 2016; Eckert et al., 2013; Osman et al., 2018; Jordan et al., 2011). As previously reported, this finding is not uniform (Jordan et al., 2011). We found repeatedly some increase in flow that paralleled the increase in GG activity in two subjects. For example, we found a substantial improvement in flow in a healthy subject in whom we recorded GG EMG also in a posterior site (in addition to the near-mandibular location studied in all subjects), when GG activity increased in both sites (Fig. 9). This observation is in accordance with the finding that high GG activity may occasionally alleviate flow limitation without EEG or other signs of arousal (Younes et al., 2012). However, as seen in Fig. 9, despite large increases in GG activity compared to wakefulness, flow limitation was not abolished, as indicated by the persistence of snoring vibrations. Also, the magnitude of increase in airflow seen in Fig. 9 was not seen in any other subject, including two other subjects in whom GG-EMG was recorded similarly in two sites: in both the two GG-EMG signals increased without any effect on flow.
Fig. 9.
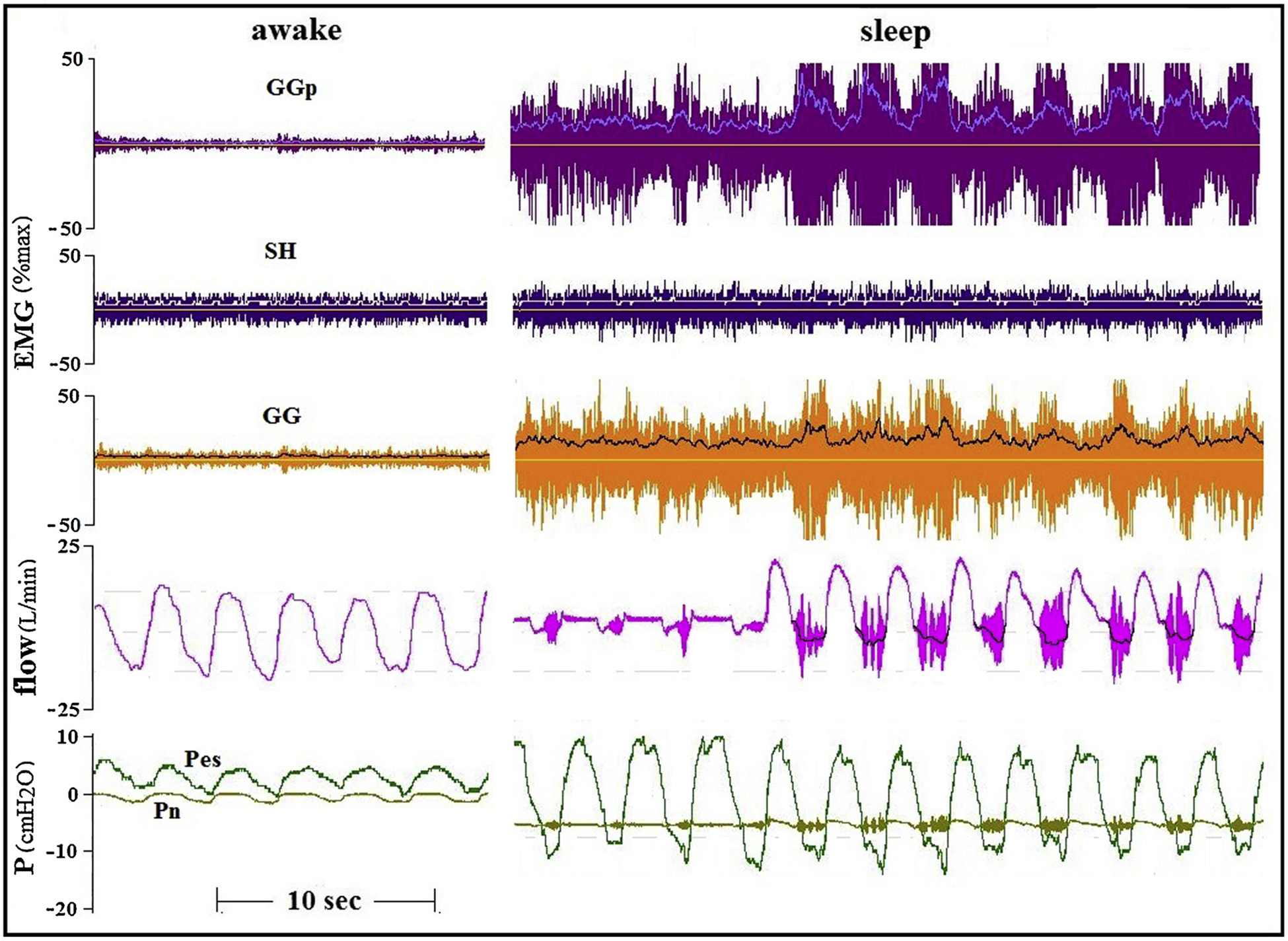
Marked improvement in flow during increased activity of the GG in a healthy subject during sleep. In this subject GG EMG was recorded concomitantly in the anterior site as in all other subjects (GG), and also in a more posterior site of the transverse part of the GG (GGp).
While one may argue that the increase in GG (and AD) EMG may be insufficient to overcome the negative pressure (CNAP) required to produce flow limitation in healthy subjects, this cannot explain the same phenomenon in OSA subjects, in whom flow limitation occurred despite the application of CPAP (see also Fig. 1). Another important consideration is that the application of CPAP could affect the results in our patients, as CPAP is known to reduce GG EMG activity, and reduction of CPAP may render the pharynx more collapsible than in the awake state. However, once CPAP is reduced, EMG activity of the peri-pharyngeal muscles increases gradually, breath by breath, well above the levels recorded during wakefulness, yet remains largely ineffective in restoring pharyngeal patency. Moreover, sleep was associated with flow limitation even when CPAP levels up to 8 cmH2O were applied (Figs. 5,6), as commonly seen in patients undergoing clinical CPAP titration. In contrast, similar CPAP levels would not be expected to produce flow limitation or to increase EMG activity during wakefulness. Hence, low EMG activity during wakefulness was sufficient to maintain airway patency yet was unable to stabilize the airway even with the support of CPAP during sleep. Accordingly, we believe that while the use of CPAP could potentially attenuate our results, it cannot explain the dissociation between EMG and airflow-mechanics during wakefulness and sleep.
As GG activity could always be augmented far above levels observed during wakefulness by induced flow limitation during sleep, one may speculate that the failure of the activated GG to prevent pharyngeal collapse can be due to reduced activity of AD muscles during sleep. If so, it would be expected that whenever activated during sleep, the AD should have at least some impact on airflow during pharyngeal collapse. However, as seen both in the tracings presented and in the cumulative, averaged results, amelioration of flow in conjunction with increases in AD activity was an uncommon event. The failure of the coactivated GG and AD to prevent pharyngeal collapse was independent of the severity of pharyngeal obstruction: it occurred both when mild flow limitation resulted in a stable obstructed ventilation, and when more severe flow limitation induced a pattern of increasing respiratory efforts terminated by arousal (Fig. 10).
Fig. 10.
Tracings of induced flow limitation in two healthy subjects in whom EMGs of the same muscles were recorded. In one subject (A, 21yrs old, BMI 26.5, AHI 5) Pn was lowered to −5 cmH2O, resulting in severe flow limitation and a gradual increase in the negative fluctuations of Pes, similar to those seen in OSA patients. In the other subject (B, 65yrs old, BMI 20, AHI 6) Pn was lowered to −7 cmH2O, resulting in moderate flow limitation with intermittent vibrations of the flow tracing. Ventilation is mostly stable, as are the EMGs. Changes in EMG during sleep, compared to wakefulness level, were not related to the severity of flow limitation and changes in downstream pressure.
While documenting the inefficiency of both GG and the AD to prevent or ameliorate flow limitation during sleep even when the activity of both exceeds wakefulness levels, our data do not provide a direct explanation for this phenomenon. Physiologically, there is no reason why the upper airway muscles should be mechanically less efficient during sleep (or anesthesia, as long as excitation-contraction relationship is not affected), when activated equally to wakefulness, or even more. Upper airway dilators do not differ from other skeletal muscles: with adequate methodology it was simple to demonstrate that increases in EMG activity of the GH is associated with increased contractile force production also during anesthesia (Brodsky et al., 2016). There is no doubt that central drive to the GG is modified considerably during sleep: the GG responds less to mechanical and chemical drive, reflex responses are altered (Wheatley et al., 1993; Pillar et al., 2000, 1997), and a poorly understood “wakefulness stimulus” (Orem, 1990) disappears. All these processes result in a reduction in GG activity during the transition from wakefulness to sleep, and were used to explain the propensity of the pharynx to collapse at the onset of sleep. However, once GG activity increases back and reaches during sleep the same or even higher levels than those observed during wakefulness, the contraction force and, hence, the flow-mechanical effect of the GG and other upper airway dilators is not expected to be reduced. Therefore, other mechanisms need to be involved to explain the inability of the active dilator muscles to maintain pharyngeal patency during sleep.
Obviously, the relationship between EMG and the mechanical effect of a striated muscle during movements is complex, depends on a large number of variables and is therefore seldom linear (Kuriki et al., 2012). While force production by a stimulated muscle depends primarily on intrinsic muscle characteristics, the actual mechanical product of this force depends also on concomitant contraction of neighbouring muscles and external loads. Accordingly, GG contraction does not necessarily imply shortening of the GG and tongue protrusion, as contraction may be isometric or even eccentric. However, flow limitation occurs well before high negative intraluminal pressures (i.e., external load) could prevent GG shortening (Fig. 1), and is not prevented by GG activity higher than the activity that prevents pharyngeal obstruction at equal negative intra-luminal pressures during wakefulness (Dotan et al., 2015; Oliven et al., 2018). Therefore, the fast decline in the ability of the dilator muscles to prevent pharyngeal collapse following transition from wakefulness to sleep seems to result from a state dependent alteration in the pattern of co-activation or coordination of muscles involved in the maintenance of pharyngeal patency. Because the pharynx is collapsible at multiple tangents, numerous muscle groups need to be coordinated and act in concert to prevent pharyngeal collapse. Even robust GG contraction elicited by electrical stimulation of the hypoglossal nerve is only partially effective, and is insufficient to restore pharyngeal stability in many patients with severe OSA (Strollo et al., 2014). Thus, explanation of the apparent inefficiency of pharyngeal dilators during sleep require to understand the central control of multiple AD groups, and how these muscles work in unison.
The low response of the AD to flow limitation may be due, in part, to arousal that ends apneas and hypopneas and interrupts further AD recruitment. However, in a previous study, we induced prolonged hypopneas in propofol-anesthetized OSA patient, thereby preventing arousal. This experimental setup resulted in activation of all AD studied. Nevertheless, as during sleep, this recruitment of AD failed to enlarge the collapsible segment of the pharynx and to improve airflow, despite large concomitant increase in tonic and phasic GG activity (Dotan et al., 2013). A potential limitation is that we have evaluated only four out of a large number of AD. However, these muscles are located in different areas around the pharynx and have different mechanical functions. Moreover, in the study performed under anesthesia (Dotan et al., 2013), we evaluated also the masseter and internal pterygoid muscle, that participate in determining the position of the mandible. Phasic activity was triggered by flow limitation in these AD in all subjects. Another consideration is that muscles that may obstruct the pharynx may be activated during sleep. However, we found that tongue retractors are less active than the GG during sleep (Dotan et al., 2015; Oliven et al., 2018), and pharyngeal constrictors were previously found to be inactive during sleep (Kuna and Smickley, 1997). Undoubtedly, as seen in Table 3, the average activity of the AD during sleep, even when only the maximal AD EMG values observed during flow limitation are considered, is rather low. However, the activity of both GG and AD was raised during sleep to levels significantly higher than while awake, but failed to improve airflow in OSA patients even with partial CPAP support.
Referring to a specific muscle function (e.g. improving pharyngeal patency), based on EMG recorded in a single location, particularly in a muscle able to perform multiple complex tasks like the GG, is a crude simplification. The GG fibers are arranged like a fan in the sagittal plane of the tongue. Its horizontal fibers advance the posterior belly of the tongue and enlarge the pharynx, while preferential contraction of vertical fibers depress the tongue and may narrow the pharynx (Dotan et al., 2011). Not only does the site of recording affects the magnitude of GG EMG (Eastwood et al., 2003; Vranish and Bailey, 2015), EMG recorded near the mandibular insertion of both the vertical and horizontal fibers of the GG does not reveal the movement of the tongue, as both protrusion and retraction may be associated with a similar EMG. Similarly, the EMG may appear unchanged although the balance of activation (and movement) of the two parts of the tongue may change during sleep.
More importantly, the conventional multiunit EMG is the summation of all motor units (MUs) activated in the vicinity of the recording electrodes. Its integrated activity increases when additional MUs become active, and/or when spike frequency increases (recruitment and rate coding). The multiple tasks performed by the tongue require precise central control of the many MUs of its muscles, but alterations in this delicate fine-tuning may not be recognizable from the multiunit EMG. In fact, similar multiunit GG EMGs were recorded when our subjects were requested to perform various isotonic and isometric tongue movements, although it is reasonable to assume that these different movements were not performed by similar activation of the same MU groups. In other words, substantial differences in the “content” of the multiunit EMG, in term of active MUs, may occur without being recognized from the traditional multiunit EMG. We believe that the considerable reduction in mechanical efficacy of apparently highly active GG and AD multiunit EMG indicates that sleep is associated with an alteration in upper airway dilator coordination, particularly during increased respiratory drive, not only in the “macro-level” (muscles and muscle parts), but also in the “micro-level” (MUs and MU pools).
Mechanically, the tongue functions as a muscular hydrostat, requiring high level of internal synchronization to perform its many tasks (Kier and Smith, 1985). Similarly, peri-pharyngeal muscles need to act in concert to maintain pharyngeal patency to airflow (Van Lunteren and Strohl, 1986). Recent work suggests that the lingual motor control is not based on muscles or groups of muscles (Bailey, 2011). Neither can specific functions be ascribed to muscles based on their morphology (i.e., intrinsic vs extrinsic) or function (i.e., protrusor vs retractor). For example, tongue retractors participate in, and are important for, protrusive movements (Fuller et al., 1998), and can be viewed, therefore, as an integral part of pharyngeal dilators. Data arising from MU recordings suggest that pharyngeal dilatation and/or stiffness are not modulated by activation of whole muscles, but results from activation of MU groups, based on their compartmental location and mechanical effect (Bailey, 2011). GG MUs of awake healthy subjects have been found to maintain a typical discharge pattern, i.e., some MUs are active only during inspiration, others also during expiration etc. (Saboisky et al., 2006). However, this pattern of activation is not an inherent characteristic of MUs, but rather a flexible and modifiable feature (Bailey, 2011; Bailey et al., 2007). A large proportion (~50 %) of GG MUs exhibit significant state-related changes in respiratory modulation of discharge pattern, i.e. MUs active during inspiration in wakefulness may switch to a tonic pattern (i.e. including expiration) during sleep. These changes appear to balance out and may occur without recognizable effect on the summed output (i.e. the multiunit GG EMG), including its phasic and tonic components (Bailey, 2011; Bailey et al., 2007). This finding is in agreement with our finding that falling asleep is associated with a change in the power spectrum of both GG and AD EMGs, that remained nearly constant throughout sleep, and returned to baseline upon arousal (Oliven et al., 2019). The significance of flexibility in MU firing behavior remains unclear. Recent work suggests that inspiratory motoneurons output is geared to the greatest mechanical advantage of the muscles that they innervate (Butler et al., 2014). Accordingly, it seems reasonable to assume that the activation of peri-pharyngeal MUs is coordinated most efficiently during wakefulness. If we expand on this idea further, we can speculate that the described state-related change from the awake pattern of coordination is less effective and may reduce the mechanical efficacy of the MUs ensemble activated during sleep. Incoordination of motoneural output is known to cause distinctive defects in muscle performance in pathological conditions like ataxia. In contrast, the mechanical consequence of physiological, state-related alteration in the pattern of peri-pharyngeal MUs coordination (Bailey, 2011; Bailey et al., 2007) is unknown. Neither do we know about the effect of sleep on the activation of skeletal muscle MUs, as respiratory muscles are the only ones required to perform the same coordinated non-uniform action during wakefulness and sleep.
Inputs to hypoglossal MUs arise from multiple sources, including the primary motor cortex and several brainstem central pattern generators. Maintaining the upper airway open requires orchestrating many thousands of MUs around the pharynx: the hypoglossal motoneuron pool alone comprises more than 8000 motoneurons per side (O’Kusky and Norman, 1995). Pharyngeal patency needs to be maintained not just during quiet breathing, but also during respiratory stimulation and concomitant critical functions such as chewing, swallowing and speaking. Obviously, this task requires a complex neural network, spanning over the spinal cord, medulla and supra-medullary structures up to the cortex. Very little is known about neural circuity governing this kind of complex movements, largely because we have lacked the ability to elucidate and then probe the underlying neuronal “wiring diagrams”. However, there is sufficient evidence that these control mechanisms are state dependent. Functional MRI studies have documented that human sleep is an active state during which brain activity is temporally re-organized in a regionally specific manner (Drummond et al., 2004). Widespread sleep-associated changes in brain activity are recognizable across multiple cortical and subcortical regions (Picchioni et al., 2013). These sleep-associated changes in organization may affect also the activation and coordination of the upper airway dilator muscles. As in states of stupor (e.g. encephalopathy, intoxication etc.), when large muscle force but no coordinated tasks can be performed, sleep may be associated with both intra and inter-muscular alterations in MU coordination, that impedes their mechanical efficacy to dilateand/or stiffen the pharynx. We suggest that MU coordination is state dependent and constitutes a vital “wakefulness process”, that is partially lost during sleep, explaining why high levels of GG and AD activity fail to alleviate pharyngeal obstruction during sleep and sedation, while much lower activity is sufficient to maintain adequate upper airway patency while awake.
The “sleep-induced alteration in coordination” theory does not preclude the use of multiunit GG EMG parameters as tools to assess central control of the GG during sleep, even if increases in GG activity are not necessarily associated with improvement in flow mechanics. Moreover, it also does not negate a (reduced) mechanical effect of dilator muscles during sleep. For example, a more stable breathing, often encountered in slow wave sleep, is associated with increased GG EMG activity (Hicks et al., 2017). The level of activity of the dilator muscles, even when augmented by flow limitation, is relatively low during sleep. In anesthetized animals exposed to extreme chemical and mechanical stimulation, we found that intense activation of dilator muscles resulted in improvement in airflow similar to that achieved by electrical stimulation of the hypoglossus (Brodsky et al., 2016). In addition, it should be kept in mind that all patients evaluated in this study had severe OSA. Although similar findings were observed also in healthy subjects, it is possible that less severe patients may well have more instances of coordinated neuromuscular recovery.
In conclusion, most studies that evaluated neural control of upper airway muscles in human selected for study the GG, hoping that the activity of this muscle yields insights into pharyngeal airway muscle activities more generally. Our findings indicate that other peri-pharyngeal muscles respond much less to flow limitation during sleep than the GG, both in healthy subjects and OSA patients. More importantly, even when AD are activated substantially more than during wakefulness, this co-activation along with the augmented GG activity seldom leads to improvement in airflow. We believe that this finding can be explained only by the assumption that alteration in the pattern of recruitment of MUs during sleep affects their coordination in a way that disrupts their flow-mechanical efficacy.
Funding
This work was supported by United States-Israel Binational Science Foundation (BSF) grant 2011491.
References
- Bailey EF, 2011. Activity of human genioglossus motor units. Respir. Physiol. Neurobiol 179, 14–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey EF, Fridel KW, Rice AD, 2007. Sleep/wake firing patterns of human genioglossus motor units. J. Neurophysiol 98, 3284–3291. [DOI] [PubMed] [Google Scholar]
- Beck R, Odeh M, Oliven A, Gavriely N, 1995. The acoustic properties of snores. Eur. Respir. J 8, 2120–2128. [DOI] [PubMed] [Google Scholar]
- Berry RB, McNellis MI, Kouchi K, Light RW, 1997. Upper airway anesthesia reduces phasic genioglossus activity during sleep apnea. Am. J. Respir. Crit. Care Med 156, 127–132. [DOI] [PubMed] [Google Scholar]
- Brodsky A, Dotan Y, Samri M, Schwartz AR, Oliven A, 2016. Differential effects of respiratory and electrical stimulation-induced dilator muscle contraction on mechanical properties of the pharynx in the pig. J. Appl. Physiol 121, 606–614. [DOI] [PubMed] [Google Scholar]
- Butler JE, Hudson AL, Gandevia SC, 2014. The neural control of human inspiratory muscles. Prog. Brain Res 209, 295–308. [DOI] [PubMed] [Google Scholar]
- Carberry JC, Jordan AS, White DP, Wellman A, Eckert DJ, 2016. Upper airway collapsibility (Pcrit) and pharyngeal dilator muscle activity are sleep stage dependent. Sleep 39, 511–521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doherty LS, Cullen JP, Nolan P, McNicholas WT, 2008. The human genioglossus response to negative airway pressure: stimulus timing and route of delivery. Exp. Physiol 93, 288–295. [DOI] [PubMed] [Google Scholar]
- Dotan Y, Golibroda T, Oliven R, Netzer A, Gaitini L, Toubi A, Oliven A, 2011. Parameters affecting pharyngeal response to genioglossus stimulation in sleep apnoea. Eur. Respir. J 38, 338–347. [DOI] [PubMed] [Google Scholar]
- Dotan Y, Pillar G, Tov N, Oliven R, Steinfeld U, Gaitini L, Odeh M, Schwartz AR, Oliven A, 2013. Dissociation of EMG and mechanical response in sleep apnoea during propofol anaesthesia. Eur. Respir. J 41, 74–84. [DOI] [PubMed] [Google Scholar]
- Dotan Y, Pillar G, Schwartz AR, Oliven A, 2015. Asynchrony of lingual muscle recruitment during sleep in obstructive sleep apnea. J. Appl. Physiol 118, 1516–1524. [DOI] [PubMed] [Google Scholar]
- Drummond SP, Smith MT, Orff HJ, Chengazi V, Perlis ML, 2004. Functional imaging of the sleeping brain: review of findings and implications for the study of insomnia. Sleep Med. Rev 8, 227–242. [DOI] [PubMed] [Google Scholar]
- Eastwood PR, Allison GT, Shepherd KL, Szollosi I, Hillman DR, 2003. Heterogeneous activity of the human genioglossus muscle assessed by multiple bipolar fine-wire electrodes. J. Appl. Physiol 94, 1849–1858. [DOI] [PubMed] [Google Scholar]
- Eastwood PR, Platt PR, Shepherd K, Maddison K, Hillman DR, 2005. Collapsibility of the upper airway at different concentrations of propofol anesthesia. Anesthesiology 103, 470–477. [DOI] [PubMed] [Google Scholar]
- Eckert DJ, McEvoy RD, George KE, Thomson KJ, Catcheside PG, 2007. Genioglossus reflex inhibition to upper-airway negative-pressure stimuli during wakefulness and sleep in healthy males. J. Physiol 581 (Pt 3), 1193–1205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eckert DJ, White DP, Jordan AS, Malhotra A, Wellman A, 2013. Defining phenotypic causes of obstructive sleep apnea. Am. J. Respir. Crit. Care Med 188, 996–1004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fogel RB, Malhotra A, White DP, 2004. Pathophysiology of obstructive sleep apnoea/hypopnoea syndrome. Thorax 59, 159–163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fogel RB, Trinder J, White DP, Malhotra A, Raneri J, Schory K, Kleverlaan D, Pierce RJ, 2005. The effect of sleep onset on upper airway muscle activity in patients with sleep apnoea versus controls. J. Physiol 564 (Pt 2), 549–562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuller D, Mateika JH, Fregosi RF, 1998. Co-activation of tongue protrudor and retractor muscles during chemoreceptor stimulation in the rat. J. Physiol 507, 265–276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hicks A, Cori JM, Jordan AS, Nicholas CL, Kubin L, Semmler JG, Malhotra A, McSharry DGP, Trinder JA, 2017. Mechanisms of the deep, slow-wave, sleeprelated increase of upper airway muscle tone in healthy humans. J. Appl. Physiol 122, 1304–1312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordan AS, White DP, 2008. Pharyngeal motor control and the pathogenesis of obstructive sleep apnea. Respir. Physiol. Neurobiol 160, 1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordan AS, Eckert DJ, Wellman A, Trinder JA, Malhotra A, White DP, 2011. Termination of respiratory events with and without cortical arousal in obstructive sleep apnea. Am. J. Respir. Crit. Care Med 184, 1183–1191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kier WM, Smith KK, 1985. Tongues, tentacles and trunks: the biomechanics and movement of muscular hydrostats. Zool. J. Linn. Soc 83, 207–324. [Google Scholar]
- Kuna ST, Smickley JS, 1997. Superior pharyngeal constrictor activation in obstructive sleep apnea. Am. J. Respir. Crit. Care Med 156, 874–880. [DOI] [PubMed] [Google Scholar]
- Kuriki HU, de Azevedo FM, Ota Takahashi LS, Mello EM, de Faria Negrão Filho R, Alves N, 2012. In: Schwartz Mark (Ed.), The Relationship between Electromyography and Muscle Force EMG Methods for Evaluating Muscle and Ntronerve Function. InTech Available from: http://www.intechopen.com/books/emg-methods-for-evaluating-muscleand-nerve-function/the-relationship-betweenelectromyography-and-muscle-force. [Google Scholar]
- Maddison KJ, Walsh JH, Shepherd KL, Bharat C, Lawther BK, Platt PR,Eastwood PR, Hillman DR, 2019. Comparison of collapsibility of the human upper airway during anesthesia and during sleep. Anesth. Analg 10.1213/ANE.0000000000004070. [DOI] [PubMed] [Google Scholar]
- Malhotra A, Fogel RB, Edwards JK, Shea SA, White DP, 2000. Local mechanisms drive genioglossus activation in obstructive sleep apnea. Am. J. Respir. Crit. Care Med 161, 1746–1749. [DOI] [PubMed] [Google Scholar]
- McGinley BM, Schwartz AR, Schneider H, Kirkness JP, Smith PL, Patil SP, 2008. Upper airway neuromuscular compensation during sleep is defective in obstructive sleep apnea. J. Appl. Physiol 105, 197–205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mezzanotte WS, Tangel DJ, White DP, 1996. Influence of sleep onset on upper-airway muscle activity in apnea patients versus normal controls. Am. J. Respir. Crit. Care Med 153, 1880–1887. [DOI] [PubMed] [Google Scholar]
- O’Kusky JR, Norman MG, 1995. Sudden infant death syndrome: increased number of synapses in the hypoglossal nucleus. J. Neuropathol. Exp. Neurol 54, 627–634. [DOI] [PubMed] [Google Scholar]
- Odeh M, Schnall R, Gavriely N, Oliven A, 1995. Dependency of upper airway patency on head position: the effect of muscle contraction. Respir. Physiol 100, 239–244. [DOI] [PubMed] [Google Scholar]
- Okabe S, Chonan T, Hida W, Satoh M, Kikuchi Y, Takishima T, 1993. Role of chemical drive in recruiting upper airway and inspiratory intercostal muscles in patients with obstructive sleep apnea. Am. Rev. Respir. Dis 147, 190–195. [DOI] [PubMed] [Google Scholar]
- Oliven A, Aspandiarov E, Gankin I, Gaitini L, Tov N, 2008. Collapsibility of the relaxed pharynx and risk of sleep apnoea. Eur. Respir. J 32, 1309–1315. [DOI] [PubMed] [Google Scholar]
- Oliven R, Cohen G, Dotan Y, Somri M, Schwartz AR, Oliven A, 2018. Alteration in upper airway dilator muscle co-activation during sleep: comparison of patients with OSA and healthy subjects. J. Appl. Physiol 124, 421–429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oliven R, Cohen G, Somri M, Schwartz AR, Oliven A, 2019. Spectral analysis of peri-pharyngeal muscles’ EMG in patients with OSA and healthy subjects. Respir. Physiol. Neurobiol 260, 53–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Onal E, Lopata M, O’Connor TD, 1980. Diaphragmatic and genioglossal electromyogram responses to CO2 rebreathing in humans. J. Appl. Physiol 50, 1052–1055. [DOI] [PubMed] [Google Scholar]
- Orem J, 1990. The nature of the wakefulness stimulus for breathing. Prog. Clin. Biol. Res 345, 23–30. [PubMed] [Google Scholar]
- Orem J, Osorio I, Brooks E, Dick T, 1985. Activity of respiratory neurons duringNREM sleep. J. Neurophysiol 54, 1144–1156. [DOI] [PubMed] [Google Scholar]
- Osman AM, Carter SG, Carberry JC, Eckert DJ, 2018. Obstructive sleep apnea: current perspectives. Nat. Sci. Sleep 10, 21–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patil SP, Schneider H, Marx JJ, Gladmon E, Schwartz AR, Smith PL, 2007. Neuromechanical control of upper airway patency during sleep. J. Appl. Physiol 102, 547–556. [DOI] [PubMed] [Google Scholar]
- Pham LV, Schwartz AR, 2015. The pathogenesis of obstructive sleep apnea. J. Thorac.Dis 15 (7), 1358–1372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Picchioni D, Duyn JH, Horovitz SG, 2013. Sleep and the functional connectome. Neuroimage 80, 387–396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pillar G, Schnall RP, Peled N, Oliven A, Lavie P, 1997. Impaired respiratory response to resistive loading during sleep in healthy offspring of patients with obstructive sleep apnea. Am. J. Respir. Crit. Care Med 155, 1602–1608. [DOI] [PubMed] [Google Scholar]
- Pillar G, Malhotra A, Fogel RB, Beauregard J, Slamowitz DI, Shea SA, White DP, 2000. Upper airway muscle responsiveness to rising PCO2 during NREM sleep. J. Appl. Physiol 89, 1275–1282. [DOI] [PubMed] [Google Scholar]
- Remmers JE, deGroot WJ, Sauerland EK, Anch AM, 1978. Pathogenesis of upper airway occlusion during sleep. J. Appl. Physiol 44, 931–938. [DOI] [PubMed] [Google Scholar]
- Saboisky JP, Butler JE, Fogel RB, Taylor JL, Trinder JA, White DP, Gandevia SC, 2006. Tonic and phasic respiratory drives to human genioglossus motoneurons during breathing. J. Neurophysiol 95, 2213–2221. [DOI] [PubMed] [Google Scholar]
- Safar P, Lourdes AE, Francis C, 1959. Upper airway obstruction in the unconscious patient. J. Appl. Physiol 14, 760–764. [DOI] [PubMed] [Google Scholar]
- Sauerland EK, Harper RM, 1976. The human tongue during sleep. Electromyographic activity of the genioglossus muscle. Exp. Neurol 51, 160–170. [DOI] [PubMed] [Google Scholar]
- Schwartz AR, O’Donnell CP, Baron J, Schubert N, Alan D, Samadi SD, Smith PL, 1998. The hypotonic upper airway in obstructive sleep apnea. Role of structures and neuromuscular activity. Am. J. Respir. Crit. Care Med 157, 1051–1057. [DOI] [PubMed] [Google Scholar]
- Strollo PJ Jr, Soose RJ, Maurer JT, de Vries N, Cornelius J, Froymovich O,Hanson RD, Padhya TA, Steward DL, Gillespie MB, Woodson BT, Van de Heyning PH, Goetting MG, Vanderveken OM, Feldman N, Knaack L, Strohl KP, STAR Trial Group, 2014. Upper-airway stimulation for obstructive sleep apnea. N. Engl. J. Med 370, 139–149. [DOI] [PubMed] [Google Scholar]
- Tangel DJ, Mezzanote WS, Sandberg EJ, White DP, 1992. Influences of NREM sleep on the activity of tonic vs. inspiratory phasic muscles in normal men. J. Appl. Physiol 73, 1058–1066. [DOI] [PubMed] [Google Scholar]
- Van Lunteren E, Strohl KP, 1986. The muscles of the upper airways. Clin. Chest Med 7,171–188. [PubMed] [Google Scholar]
- Vranish JR, Bailey EF, 2015. A comprehensive assessment of genioglossus electromyographic activity in healthy adults. J. Neurophysiol 113, 2692–2699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheatley JR, Mezzanotte WS, Tangel DJ, White DP, 1993. Influence of sleep on genioglossus muscle activation by negative pressure in normal men. Am. Rev. Respir. Dis 148, 597–605. [DOI] [PubMed] [Google Scholar]
- Worsnop C, Kay A, Pierce R, Kim Y, Trinder J, 1998. Activity of respiratory pump and upper airway muscles during sleep onset. J. Appl. Physiol 85, 908–920. [DOI] [PubMed] [Google Scholar]
- Younes M, 2003. Contributions of upper airway mechanics and control mechanisms to severity of obstructive apnea. Am. J. Respir. Crit. Care Med 168, 645–658. [DOI] [PubMed] [Google Scholar]
- Younes M, Andrea HS, Loewen ASH, Ostrowski M, Laprairie J, Maturino F, Hanly PJ, 2012. Genioglossus activity available via non-arousal mechanisms vs. That required for opening the airway in obstructive apnea patients. J. Appl. Physiol 112, 249–258. [DOI] [PubMed] [Google Scholar]