

Abstract
Background
During fertility treatment, diminished ovarian reserve (DOR) is a challenge that can seriously affect a patient’s reproductive potential. However, the pathogenesis of DOR is still unclear and its treatment options are limited. This study aimed to explore DOR’s molecular mechanisms.
Methods
We used R software to analyze the mRNA microarray dataset E-MTAB-391 downloaded from ArrayExpress, screen for differentially expressed genes (DEGs), and perform functional enrichment analyses. We also constructed the protein-protein interaction (PPI) and miRNA-mRNA networks. Ovarian granulosa cells (GCs) from women with DOR and the control group were collected to perform untargeted metabolomics analyses. Additionally, small molecule drugs were identified using the Connectivity Map database.
Results
We ultimately identified 138 DEGs. Our gene ontology (GO) analysis indicated that DEGs were mainly enriched in cytokine and steroid biosynthetic processes. According to the Kyoto Encyclopedia of Genes and Genomes (KEGG), the DEGs were mainly enriched in the AGE (advanced glycation end-product)-RAGE (receptor for AGE) signaling pathway in diabetic complications and steroid biosynthesis. In the PPI network, we determined that JUN, EGR1, HMGCR, ATF3, and SQLE were hub genes that may be involved in steroid biosynthesis and inflammation. miRNAs also played a role in DOR development by regulating target genes. We validated the differences in steroid metabolism across GCs using liquid chromatography-tandem mass spectrometry (LC-MS/MS). We selected 31 small molecules with potentially positive or negative influences on DOR development.
Conclusion
We found that steroidogenesis and inflammation played critical roles in DOR development, and our results provide promising insights for predicting and treating DOR.
Keywords: Diminished ovarian reserve, Bioinformatics analysis, Steroid, Inflammation, Metabolomics
Introduction
Diminished ovarian reserve (DOR), defined as a decline in the number or quality of follicles and oocytes, reduces a female patient’s reproductive potential (Sharara, Scott Jr & Seifer, 1998). The incidence of DOR ranges from 9% to 24% in women undergoing in vitro fertilization (IVF) (Kyrou et al., 2009). The most common effective treatment for DOR is the use of assisted reproductive techniques. Ovarian reserve naturally declines with age, but some women experience DOR much earlier than average. Young women with DOR thus have an accelerated physiological decline in ovarian reserve. DOR is one of the greatest challenges for reproductive endocrinologists because it is characterized by poor ovarian response to gonadotrophin stimulation, low pregnancy rates, and high rates of pregnancy loss (Levi et al., 2001; Navot, Rosenwaks & Margalioth, 1987).
An ovarian follicle is a complex structure comprised of oocytes, cumulus cells (CCs), and granulosa cells (GCs). The bidirectional communication between the oocyte and its companion somatic cells is essential for follicular development and oocyte growth (Anderson & Albertini, 1976; Matzuk et al., 2002). In mice, the oocyte’s transcriptional activity was modulated when in vitro cultured with GCs, but not without GCs (De La Fuente & Eppig, 2001). Another study showed that DOR patients had an increase in GC apoptosis, which is associated with a poor ovarian response and oocyte yield (Fan et al., 2019). Considering this coadjutant relationship between oocyte and GC, exploring the alteration of GCs from women with DOR may provide a deeper understanding for DOR pathogenesis. Previous studies have investigated the mRNA expression profiles of ovarian CCs (Greenseid et al., 2011) and GCs (Chin et al., 2002; Skiadas et al., 2012) in addition to miRNA expression patterns (Chen et al., 2017; Woo et al., 2018) in DOR patients. These studies typically had different inclusion criteria for DOR patients. miRNAs are small noncoding RNAs that play critical roles in many diseases and biological processes, including reproduction, and regulate mRNA translation and stability (Fabian, Sonenberg & Filipowicz, 2010). Exploring how miRNA affects female reproduction (Sabry et al., 2019) could reveal miRNA therapeutics as a possible DOR treatment option.
Steroid hormones are a type of steroid involved in many biological and physiological functions. Cholesterol is the precursor for steroid hormone synthesis (Greaves et al., 2014). Steroid hormone levels affect the follicular growth and development processes (Chou & Chen, 2018). Inflammation triggers a wide range of physiological and pathological processes (Medzhitov, 2008). Aberrant inflammation has a negative effect on folliculogenesis and ovulation, and polycystic ovary syndrome (PCOS) is associated with the chronic endogenous production of low-grade pro-inflammatory cytokines (Boots & Jungheim, 2015). Inflammation and steroidogenesis abnormalities may also be involved in the development of DOR.
In this study, we performed bioinformatic analyses on the mRNA expression profiles of DOR GCs from the publicly available dataset E-MTAB-391 in order to identify differentially expressed genes (DEGs). We constructed protein-protein interaction (PPI) networks based on the DEGs, and a miRNA-mRNA network using the differentially expressed miRNAs (DEMs) extracted from a previous study (Woo et al., 2018). Small molecule drugs with potential synergistic or antagonistic effects on DOR were also screened using the Connectivity Map (CMap) database (Lamb et al., 2006). Moreover, we applied liquid chromatography-tandem mass spectrometry (LC-MS/MS) on our samples to explore the metabolic alteration of GCs. This study may shed light on the future of DOR prognosis and treatment.
Materials & Methods
Data collection
We screened GEO and ArrayExpress (https://www.ebi.ac.uk/arrayexpress/) for expressed profiles of GCs from patients with and without DOR. We excluded datasets that: (1) were without detailed sample information, (2) used samples based on cell lines or animal models, (3) had sample sizes <10, and (4) had quite different definitions of DOR or used different participant age ranges than our study. Only one mRNA microarray dataset, E-MTAB-391 (which included 13 DOR samples and 13 normal ovarian reserve (NOR) samples (Skiadas et al., 2012), met our criteria and was downloaded from ArrayExpress. E-MTAB-391′s platform was the A-MEXP-1564-IIIumina HumanRef-8 WG-DASL v3 Expression BeadChip. We directly extracted the data from the DOR miRNAs used in a previous study (Woo et al., 2018).
Identifying DEGs
We used the limma package in R software (Ritchie et al., 2015) to identify DEGs across the DOR and NOR samples. The adjusted P-value (false discovery rate) was obtained using the Benjamini–Hochberg algorithm when screening the DEGs. We set P < 0.05 and —log 2 (fold change; FC) —>0.58 as the DEG cut-off criteria. The DEGs were divided into upregulated and downregulated DEGs and saved for subsequent analyses.
Functional and enrichment analyses of DEGs
Gene ontology (GO) annotation is widely used to study the biological functions of multiple genes, and is comprised of three independent ontologies: biological process (BP), molecular function (MF), and cellular component (CC) (Ashburner et al., 2000). The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis is a valuable tool used to assess the interaction networks of genes and their products (Kanehisa et al., 2017). In this study, we used the clusterProfiler package in R software to obtain the GO and KEGG pathway enrichment for the DEGs (Yu et al., 2012). We set the value of P < 0.05 as the threshold for significance.
PPI network construction and module analysis
The STRING database offers PPI assessment and integration (Szklarczyk et al., 2017). To achieve a system-wide understanding across DEGs, we constructed the PPI network using STRING (version 10.5; https://string-db.org/cgi/input.pl) with a combined cutoff score ≥ 4. The network was visualized using Cytoscape (version 3.6.1; https://cytoscape.org/), which can integrate biomolecular interaction networks into a unified conceptual framework (Shannon et al., 2003). The nodes and edges of the network represent proteins and protein-protein associations, respectively. We performed a module analysis of the PPI network based on the Molecular Complex Detection (MCODE) feature of the Cytoscape software using the following parameters: degree cut-off = 2, node score cut-off = 0.2, max depth = 100, and k-score = 2. We performed subsequent GO and KEGG pathway analyses of the selected modules using the clusterProfiler package.
Exploring DEM target genes
We extracted 105 DEMs from the GCs of women diagnosed with DOR in a previous study (Woo et al., 2018). The multiMiR package integrated 11 miRNA-target databases (three validated and eight predicted miRNA-target databases) and three disease-/drug-related miRNA databases (Ru et al., 2014) to retrieve interactions between the DEMs and screened DEGs. The DEM target genes were only screened from the 11 miRNA-target databases. Finally, we visualized the regulatory miRNA-mRNA network using Cytoscape software.
Identifying small molecules
The CMap database (https://portals.broadinstitute.org/cmap) contains 7,000 gene-expression profiles from cultured human cells treated with bioactive small molecules (Lamb et al., 2006), and is a valuable resource when looking for connections between diseases, genetic perturbation, and drug action. We mapped the upregulated and downregulated genes to the CMap database to identify potential small molecule drugs, which either have antagonistic or synergistic effects on DOR. We regarded n (the number of instances) ≥ 4, enrichment >0.7, and P-value <0.01 as statistically significant.
Collecting GCs
This study was approved by the ethics committee of the First Hospital of Lanzhou University (LDYYLL2019-44), and we obtained written informed consent from all participants. Ovarian GCs were collected from women with DOR (n = 3) and women with NOR (n = 3). All participants were ≤ 35 years old to eliminate age as a potential confounding variable. DOR was identified using FSH levels (12 ≤ FSH <25) and ovarian response (the number of follicles on the day of the ovulatory human chorion gonadotropin (hCG) trigger injection ≤ 7). We selected infertile women undergoing IVF due to male or tubal factor infertility for the NOR group. Controlled ovarian stimulation was performed and follicular development was monitored using a transvaginal ultrasound. Oocyte retrieval was performed 36 h after hCG administration and GCs were isolated from fluid aspirates using previously-described methods (Vanacker et al., 2011).
LC-MS/MS experiments
We performed metabolite extraction and LC-MS/MS analysis at Beijing Genomics Institute (BGI). Each frozen GCs sample was thawed and weighed into 1.5 mL Eppendorf tubes. We added the internal standard solution and 800 µL of methanol/acetonitrile/water solvent (2:2:1, v/v/v) to homogenize. Mixtures were centrifuged at 25,000 rcf for 15 min, and the supernatant was transferred out and vacuum dried. We then re-extracted the metabolite extract in 200 µL of a methanol/water mixture (1:9, v/v). After vortexing, the samples were centrifuged again. We collected the supernatant and inspected each sample using 20 µL of supernatant, the Waters 2D UPLC (Waters, Milford, MA, USA), and a Q Exactive high-resolution mass spectrometer (Thermo Fisher Scientific, Waltham, MA, USA). The analytical column we used was an ACQUITY UPLC BEH C18 (1.7 µm, 2. 1 ×100 mm, Waters). In positive ion mode, the mobile phase was MS-grade water with 0.1% formic acid (A) and 100% methanol with 0.1% formic acid (B). In negative ion mode, the mobile phase was MS-grade water with 10 mM of ammonium formate (A) and 95% methanol with 10 mM of ammonium formate (B). The extracts were gradient-eluted with a flow rate of 0.35 ml/min. The full scan and fragment acquisition resolutions were 70,000 and 17,500, respectively. The ESI parameters were set as follows: sheath gas flow rate of 40 L min-1, auxiliary gas flow rate of 10 L min-1, spray voltage of 3800 V (positive mode) and 3200 V (negative mode), capillary temperature of 320 °C, and auxiliary gas heater temperature of 350 °C. The LC-MS/MS data were processed using Compound Discoverer 3.0 software (Thermo Fisher Scientific). We identified the differential metabolites using a combination of principal component analysis (PCA) and univariate analysis.
Results
Identifying DEGs related to DOR
In this study, we downloaded the normalized expression data from the E-MTAB-391 dataset (Fig. S1). A total of 18,128 genes were available for further DEG identification. Using our criteria, we selected 138 DEGs from the DOR and NOR samples, including 55 upregulated and 83 downregulated genes. The volcano plot in Fig. 1 shows the distribution of all screened genes. The heat map of all DEGs based on unsupervised hierarchical clustering is shown in Fig. S2.
Figure 1. Volcano plot of all DEGs.

The orangered and blue dots represent significantly upregulated and downregulated DEGs, respectively. DEGs, differentially expressed genes; FC, fold change.
GO and KEGG pathway enrichment analyses of DEGs
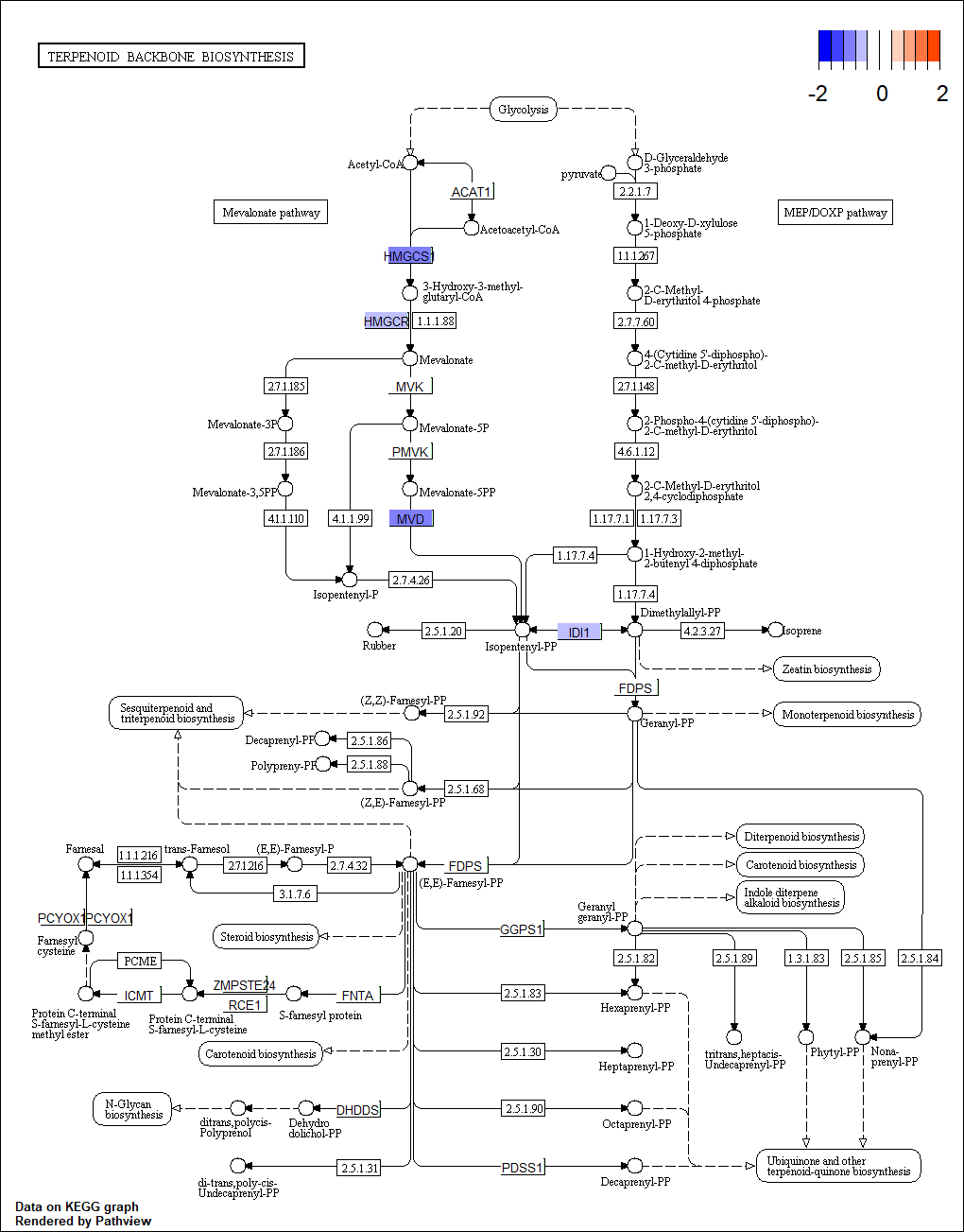
According to the GO BP analysis, the upregulated DEGs were mainly enriched in skeletal muscle cell differentiation and regulation of transcription from RNA polymerase II promoter in response to stress. Downregulated DEGs were mainly enriched in the steroid biosynthetic and cholesterol biosynthetic processes. Figure 2 shows the top 20 GO BP up- and down-regulated DEG terms in detail. Additionally, we performed KEGG pathway enrichment analysis and the results can be found in Table 1. The upregulated DEGs were significantly enriched in the AGE (advanced glycation end-product)-RAGE (receptor for AGE) signaling pathway in diabetic complications and human T-cell leukemia virus 1 infection, and the downregulated DEGs were mainly enriched in steroid biosynthesis (Fig. 3) and terpenoid backbone biosynthesis (Fig. S3).
Figure 2. The GO enrichment analysis of the DEGs.

(A) Top 20 enriched BP terms of downregulated genes. (B) Top 20 enriched BP terms of upregulated genes. The length of bars represents the number of genes, the color of bars represents corresponding adjusted P-value. GO, Gene Ontology; DEGs, differentially expressed genes; BP, biological process.
Table 1. Top five KEGG pathways of the DEGs.
| Pathway ID | Description | P-value | Count | Genes | State |
|---|---|---|---|---|---|
| hsa04933 | AGE-RAGE signaling pathway in diabetic complications | 0.000389 | 4 | SERPINE1, EGR1, CXCL8, JUN | upregulated |
| hsa05166 | Human T-cell leukemia virus 1 infection | 0.000905 | 5 | EGR2, ZFP36, SERPINE1, SRF, JUN | upregulated |
| hsa05161 | Hepatitis B | 0.002359 | 4 | EGR3, EGR2, CXCL8, JUN | upregulated |
| hsa04657 | IL-17 signaling pathway | 0.004284 | 3 | FOSB, CXCL8, JUN | upregulated |
| hsa05203 | Viral carcinogenesis | 0.005122 | 4 | EGR3, EGR2, SRF, JUN | upregulated |
| hsa00100 | Steroid biosynthesis | 2.49E−11 | 7 | CYP51A1, FDFT1, MSMO1, EBP, NSDHL, SC5D, SQLE | downregulated |
| hsa00900 | Terpenoid backbone biosynthesis | 1.39E−05 | 4 | HMGCS1, HMGCR, IDI, MVD | downregulated |
| hsa04913 | Ovarian steroidogenesis | 0.00035 | 4 | LHCGR, STAR, HSD3B2,CYP19A1 | downregulated |
| hsa04066 | HIF-1 signaling pathway | 0.000908 | 5 | HK2, PGK1, ALDOC, ENO2, SLC2A1 | downregulated |
| hsa00010 | Glycolysis / Gluconeogenesis | 0.001222 | 4 | HK2, PGK1, ALDOC, ENO2, | downregulated |
Notes.
- KEGG
- Kyoto Encyclopedia of Genes and Genomes
- DEGs
- differentially expressed genes
Figure 3. Pathway of steroid biosynthesis from KEGG.
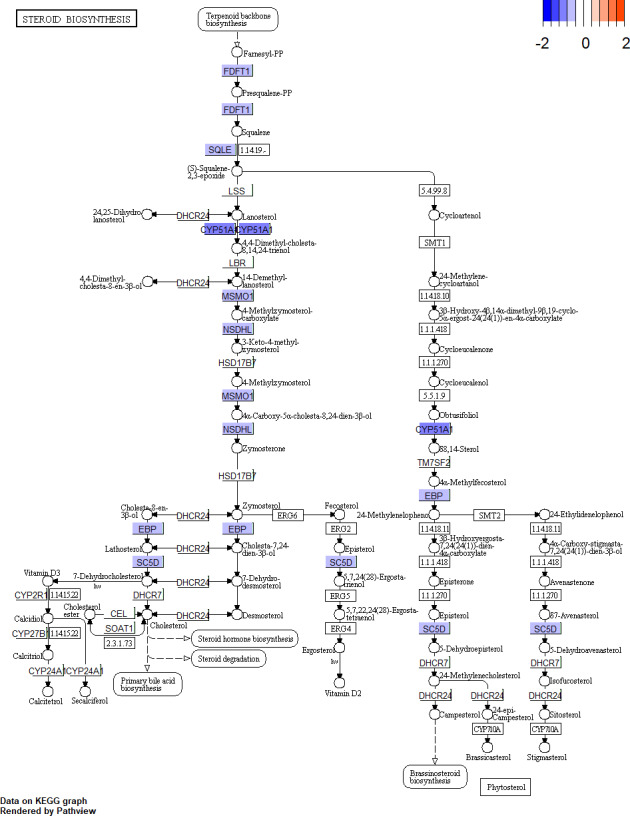
The genes with blue are downregulated DEGs. KEGG, Kyoto Encyclopedia of Genes and Genomes; DEGs, differentially expressed genes.
Constructing PPI networks and module analysis
We imported all DEGs related to DOR into the STRING database when constructing the PPI network, which included 99 nodes and 294 edges when the cut-off combined score was set at ≥ 0.7. The PPI network was visualized using Cytoscape software (Fig. 4A). The genes with higher node degrees were Jun proto-oncogene, AP-1 transcription factor subunit (JUN, degree=24); early growth response 1 (EGR1, degree=18); 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR, degree=17); activating transcription factor 3 (ATF3, degree=15); and squalene epoxidase (SQLE, degree=15). Additionally, we filtered out the top two modules from the PPI network in order to implement further GO and KEGG pathway analyses. Module 1 (Fig. 4B) contained 12 downregulated genes enriched in the cholesterol biosynthetic process of the GO BP term. Module 2 (Fig. 4C) contained 10 upregulated genes that were mainly enriched in skeletal muscle cell differentiation of the GO BP term. The detailed results of the modules’ GO and KEGG pathway analyses are shown in Table 2.
Figure 4. The PPI network and modular analysis.

(A) The PPI network of DEGs. (B) Module 1. (C) Module 2.Node represents gene, node size represents node degree. The orangered and blue nodes represent upregulated and downregulated genes, respectively. The depth of color represents the level of differential expression and the width of lines represents the combined score between two proteins. PPI, protein-protein interaction; DEGs, differentially expressed genes.
Table 2. Enriched GO BP terms (top five) and significantly enriched KEGG pathways of genes in the top two modules.
| Modules | Description | P.adjust | Count | |
|---|---|---|---|---|
| module 1 | BP terms | cholesterol biosynthetic process | 3.53E−24 | 11 |
| secondary alcohol biosynthetic process | 3.53E−24 | 11 | ||
| sterol biosynthetic process | 4.99E−24 | 11 | ||
| cholesterol metabolic process | 2.88E−21 | 11 | ||
| secondary alcohol metabolic process | 3.39E−21 | 11 | ||
| KEGG pathway | Steroid biosynthesis | 1.02E−15 | 7 | |
| Terpenoid backbone biosynthesis | 1.09E−07 | 4 | ||
| module 2 | BP terms | skeletal muscle cell differentiation | 2.30E−05 | 4 |
| muscle organ development | 0.00019411 | 5 | ||
| skeletal muscle tissue development | 0.00019411 | 4 | ||
| skeletal muscle organ development | 0.00019411 | 4 | ||
| regulation of nuclear-transcribed mRNA poly(A) tail shortening | 0.001616513 | 2 | ||
| KEGG pathway | Human T-cell leukemia virus 1 infection | 0.019978459 | 3 | |
| C-type lectin receptor signaling pathway | 0.043059356 | 2 | ||
| Osteoclast differentiation | 0.043059356 | 2 |
Notes.
- GO
- Gene Ontology
- BP
- biological process
- KEGG
- Kyoto Encyclopedia of Genes and Genomes
Predicting DEM target genes and constructing the DEM-DEG regulatory network
We analyzed the DEMs associated with DOR, including 85 upregulated and 20 downregulated genes, using the multiMiR package to predict their target genes. We then used the identified DEM-DEG pairs (comprised of 91 DEMs and 109 DEGs) to construct the regulatory network. Among the pairs, miR-155-5p, miR-16-5p, let-7b-5p, miR-107, and miR-103a-3p had the most target genes. The detailed interactions between the DEMs and DEGs are shown in Fig. 5.
Figure 5. The DEMs-DEGs regulatory network.

(A) The upregulated DEMs and targeted DEGs. (B) The downregulated DEMs and targeted DEGs. Circle nodes represent DEGs and triangle nodes represent DEMs. Orangered represent upregulation and blue represent downregulation. DEGs, differentially expressed genes; DEMs, differentially expressed miRNAs.
Screening small molecule drugs
In order to screen out small molecule drugs, we compared all DEGs to the gene expression profiles in CMap. We identified 31 small molecules, seven of which had negative scores with the potential to reverse DOR. The detailed results are shown in Fig. 6.
Figure 6. Result of CMap analysis.

The orangered dots represent the synergistic small molecule drugs. The blue dots represent antagonistic small molecule drugs. CMap, Connectivity Map.
Metabolic differences between DOR and NOR GCs
Metabolites are essential for cellular function and untargeted metabolomics analyses can provide information on their associations with diseases. We analyzed the GC samples from the DOR and NOR groups using LC-MS/MS in both positive and negative ion modes. After data processing and metabolite identification, we screened the differential metabolites using a threshold p-value <0.05 and FC ≥ 1.2 or ≤ 0.83. We did detect metabolic differences between the GCs of the DOR and NOR samples. The detailed differences in the steroids and metabolites observed in the GCs of the two groups are listed in Table 3.
Table 3. LC-MS/MS detected steroids that varied in GCs of DOR with significant difference.
| Components | Formula | m/z | FC | p-value | Class | Sub class | label |
|---|---|---|---|---|---|---|---|
| Hydroxyprogesterone | C21H30O3 | 330.2190 | 0.0404 | 0.0381 | Steroids and steroid derivatives | Pregnane steroids | down |
| Progesterone | C21H30O2 | 314.2244 | 0.0785 | 0.0025 | Steroids and steroid derivatives | Pregnane steroids | down |
| 3 alpha-hydroxydesogestrel | C22H30O2 | 326.2241 | 0.0657 | 0.0027 | Steroids and steroid derivatives | Estrane steroids | down |
| (6beta,8xi,11beta,14xi,16alpha)-9-fluoro-6,11,17,21-tetrahydroxy-16-methylpregna-1,4-diene-3,20-dione | C22H29FO6 | 408.1960 | 0.0290 | 0.0093 | Steroids and steroid derivatives | Hydroxysteroids | down |
| (6beta,8xi,11beta,14xi,16alpha)-9-fluoro-6,11,17,21-tetrahydroxy-16-methylpregna-1,4-diene-3,20-dione | C22H29FO6 | 408.1959 | 0.0340 | 0.0179 | Steroids and steroid derivatives | Hydroxysteroids | down |
| 4,6-cholestadien-3-one | C27H42O | 382.3229 | 0.0343 | 0.0352 | Steroids and steroid derivatives | Cholestane steroids | down |
| Cholest-4-en-3-one | C27H44O | 384.3385 | 0.0796 | 0.0273 | Steroids and steroid derivatives | Cholestane steroids | down |
Notes.
- LC-MS/MS
- liquid chromatography-tandem mass spectrometry
- GCs
- granulosa cells
- DOR
- diminished ovarian reserve
- FC
- fold change
Discussion
A patient with DOR has a reduced number of retrieved oocytes compared to other women of a similar age. In some women, DOR can progress to a diagnosis of primary ovarian insufficiency (POI), which is an extreme form of ovarian dysfunction (Cooper et al., 2011). Because of this serious threat to a patient’s reproductive health, there is an urgent need to further examine DOR etiology. Previous studies have used the GC mRNA/miRNA expression profiles from DOR patients to explore the molecular mechanisms of DOR. In this study, we focused on altered GCs from young women with DOR, and performed LC-MS/MS experiments and bioinformatic analyses to explore the differences between DOR and NOR. We obtained raw mRNA expression patterns and DEM data from previous publications (Skiadas et al., 2012; Woo et al., 2018) with similar inclusion criteria to ours. The inclusion criteria are presented in Table S1.
Sex steroid hormones (progestogens, androgens, and estrogens) have a steroid nucleus structure and are typically synthesized from cholesterol in the gonads and adrenal glands (Greaves et al., 2014). These hormones play important roles in female reproduction. The synthesis and secretion of estrogen are promoted by the elevated FSH levels found in patients with DOR (Practice Committee of the American Society for Reproductive Medicine, 2015). However, whether there is a difference in the estrogen levels of patients with DOR and NOR remains controversial. In our study, we found that downregulated DEGs were mainly enriched in the steroid biosynthetic process of the GO BP term (Fig. 2A). KEGG pathway analysis (Table 1) showed that downregulated DEGs were mainly enriched in steroid biosynthesis and terpenoid backbone biosynthesis. Therefore, a range of steroidogenesis substances may play a major role in DOR development. Consistent with our PPI network results (Fig. 4), we found several key genes in the top 20 (including HMGCR, SQLE, CYP51A, HMGCS1, FDFTI, SC5D, NSDHL, IDI1, EBP, and MSMO1) in steroid biosynthesis and terpenoid backbone biosynthesis pathways (Fig. 3 and Fig. S3). All these genes were downregulated in patients with DOR from our dataset. Previous studies have found that HMGCR catalyzes the first rate-limiting step in cholesterol synthesis (Howe et al., 2017); HMGCS1 condenses acetyl-CoA to form 3-hydroxy-3-methylglutaryl CoA, which is the substrate for HMGCR (Mathews et al., 2014); and CYP51A also participates in cholesterol synthesis, which can catalyze the removal of the 14α-methyl group from lanosterol (Sharpe & Brown, 2013). Upstream biological disruptions lead to a series of metabolomic changes. According to our untargeted metabolomics analysis, several steroids (Table 3) were significantly lower in GCs from patients with DOR compared to the control group. Among these steroids, progesterone plays an essential role in female reproductive events (ovulation, implantation, and pregnancy maintenance) and serves as an intermediate during estrogen biosynthesis (Gellersen, Fernandes & Brosens, 2009). Prior evidence suggests that GCs can directly produce progesterone before entering theca cells to convert into androgens (Oktem et al., 2017). Hydroxyprogesterone acts as an intermediate during the conversion of progesterone to androgens that are transported into GCs and converted into estrogen. The relationships between steroids and ovarian function (Table 3) have been rarely reported on and require more study. It has been demonstrated that several steroidogenic gene disturbances induced by bisphenol A can cause developmental impairments of ovary tissue (Liu et al., 2019). Overall, patients with DOR may have impaired hormone synthesis, which could be compensated for by elevated FSH levels. The perturbation of steroidogenic genes may be responsible for DOR development.
Aberrant inflammation has been suggested to influence follicular growth and development (Boots & Jungheim, 2015). Our results showed that upregulated genes were enriched in the AGE-RAGE signaling pathway (Table 1). The AGE-RAGE signaling pathway has been shown to induce reactive oxygen species (ROS) burst and inflammation, eventually leading to POI (Huang et al., 2019). EGR1 plays a proinflammatory role in numerous pathological processes and human diseases (Schmidt et al., 2008). A recent study found that in mice, EGR1 was increased in aged ovaries compared to young ovaries (Yuan et al., 2016). Our results showed that EGR1 as a key gene in the PPI network was upregulated in women with DOR. During cholestasis, EGR1 regulates the production of inflammatory mediators, including cytokines and adhesion molecules, that promote the accumulation and activation of inflammatory cells, causing liver injuries (Bonetti et al., 2010). According to the results of our GO analysis of upregulated DEGs (Fig. 2B), cytokines may be associated with DOR development. Cytokines play a key role in inflammation, and can be found in the immune cells of the ovary (Tabibzadeh, 1994; Vinatier et al., 1995). Accumulated evidence suggests that inflammation is closely related to ovarian dysfunction. Women diagnosed with PCOS often present with chronic low-grade inflammation due to overactive interleukin-1 (IL-1), a proinflammatory cytokine (Popovic, Sartorius & Christ-Crain, 2019). Additionally, multiple autoimmune diseases have adverse effects on female fertility via premature DOR (Sen et al., 2014). Therefore, anti-inflammatory treatment may be able to alleviate the progression of DOR. In a POI rat model, resveratrol counteracted inflammatory signaling induced by ionizing radiation, and preserved the entire ovarian follicle pool (Said et al., 2016). More studies are needed to confirm the role of inflammation in DOR development and whether controlling inflammation is an option for DOR treatment.
miRNAs, a class of endogenous non-coding small molecule RNA, play an important role in gene expression modulation at the post-transcriptional level. Previous studies have shown that miRNAs help regulate reproductive functions, particularly follicular development, oocyte maturation, corpus function, pregnancy establishment, and early embryonic development (Eisenberg et al., 2015; Tesfaye et al., 2016). The role of miRNAs in ovarian function has been demonstrated primarily by the conditional knockout of Dicer (Luense, Carletti & Christenson, 2009), a cytoplasmic RNase; required for miRNA production in mammals. In a mouse model, the conditional knockout of Dicer in ovarian GCs led to decreased ovulation rates (Nagaraja et al., 2008) and compromised folliculogenesis and POI in oocytes (Yuan et al., 2014). In our study, we found that steroidogenic genes were regulated by differentially expressed miRNAs. HMGCS1 was regulated by 25 DEMs, including miR-155-5p, miR-16-5p, let-7b-5p, miR-107, and miR-103a-3p. Furthermore, a single miRNA can target multiple genes. MiR-107 targets 21 DEGs, including five steroidogenic genes: HMGCS1, FDFT1, CYP51A1, SQLE, and EBP. miRNA-107 expression in murine ovarian GCs exposed to cadmium was significantly different from expression in the control group, and miRNA-107 can regulate kit ligand (kitl) expression (Wang et al., 2018). Kitl plays an important role in the recruitment of primitive follicles (Parrott & Skinner, 1999), the proliferation and differentiation of GCs, the recruitment of theca cells, and early steroid hormone synthesis (Flanagan, Chan & Leder, 1991). Therefore, miRNAs may contribute to DOR development by regulating target genes. microRNA therapies for several diseases have reached clinical testing stages with promising results (Rupaimoole & Slack, 2017). miRNAs should be researched for potential DOR treatments.
We also conducted CMap analysis to quickly identify molecule drugs with antagonistic or synergistic effects on DOR based on their gene expression profiles. We found seven agents with negative scores that had potential for DOR treatment (Fig. 6). Among these, H-7 (1-(5-isoquinolinesulfonyl)-2-methylpiperazine), an inhibitor of protein kinase C, has been found to reduce the release of oocytes from rat ovaries (Shimamoto, Yamoto & Nakano, 1993). Estriol is a form of estrogen, and a meta-analysis concluded that luteal estradiol stimulation in assisted reproductive technology could decrease cycle cancellation rates and increase clinical pregnancy rates in poor responders exposed to controlled ovarian hyperstimulation (Reynolds et al., 2013). Therefore, we hypothesized that these molecule drugs, identified by bioinformatic analysis, may provide novel DOR treatment. Further validation of their effects is still needed.
Despite our study’s promising findings, there were still limitations. We identified possible DEGs using —log2FC—>0.58 (the approximate fold change was >1.5), which is a relatively lower criterion than those used by other studies. DOR is an early stage ovarian reserve impairment that may take several years to develop into POI, and subtle alterations may have broader significance during its development. Small gene expression changes are also worth noting. In addition, we derived the E-MTAB-391 data from a large sample size. Nevertheless, we validated the steroid metabolism differences between DOR and NOR samples using LC-MS/MS. Therefore, our findings are reliable and provide valuable insight into DOR development.
Conclusion
We used bioinformatics approaches to investigate the perturbed steroidogenic and inflammation-related genes that may be regulated by miRNAs in women with DOR. Using metabolomics, we found that steroid metabolites were reduced in the GCs from DOR samples. Additionally, several small molecule drugs (e.g., the steroid hormone estriol) with potential antagonistic or synergistic effects on DOR were screened out. Our results suggest that steroidogenesis and inflammation play critical roles in DOR development, and should be pursued in future studies on DOR prediction and treatment.
Supplemental Information
The red represents DOR samples and the black represents NOR samples. The lateral axis is the name of samples and longitudinal axis is the level of genes expression. DOR, diminished ovarian reserve; NOR, normal ovarian reserve.
{kind=link}
The color change from red to blue represents the values change of gene expression from high to low. DEGs, differentially expressed genes.
{kind=link}
The genes with blue are downregulated DEGs. KEGG, Kyoto Encyclopedia of Genes and Genomes; DEGs, differentially expressed genes.
{kind=link}
*In publication of Woo et al., women who met two or more of the four criteria in the table except for age were defined as diminished ovarian reserve, and the parameter that was set to 7 represented for antral follicle count (AFC).
Funding Statement
This study was supported by the National Natural Science Foundation of China (no. 81960278 and 81601351), and the Natural Science Foundation of the First Hospital of Lanzhou University (no. ldyyyn2018-52). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
The authors declare there are no competing interests.
Author Contributions
Ruifen He conceived and designed the experiments, performed the experiments, authored or reviewed drafts of the paper, and approved the final draft.
Zhongying Zhao performed the experiments, analyzed the data, prepared figures and/or tables, and approved the final draft.
Yongxiu Yang analyzed the data, prepared figures and/or tables, and approved the final draft.
Xiaolei Liang conceived and designed the experiments, authored or reviewed drafts of the paper, and approved the final draft.
Human Ethics
The following information was supplied relating to ethical approvals (i.e., approving body and any reference numbers):
This study was approved by the Ethics Committee of The First Hospital of Lanzhou University (LDYYLL2019-44).
Data Availability
The following information was supplied regarding data availability:
The raw data are available in the Supplemental File.
References
- Anderson & Albertini (1976).Anderson E, Albertini DF. Gap junctions between the oocyte and companion follicle cells in the mammalian ovary. Journal of Cell Biology. 1976;71:680–686. doi: 10.1083/jcb.71.2.680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashburner et al. (2000).Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, Kasarskis A, Lewis S, Matese JC, Richardson JE, Ringwald M, Rubin GM, Sherlock G. Gene ontology: tool for the unification of biology. The gene ontology consortium. Nature Genetics. 2000;25:25–29. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonetti et al. (2010).Bonetti TC, Salomao R, Brunialti M, Braga DP, Borges Jr E, Silva ID. Cytokine and hormonal profile in serum samples of patients undergoing controlled ovarian stimulation: interleukin-1beta predicts ongoing pregnancy. Human Reproduction. 2010;25:2101–2106. doi: 10.1093/humrep/deq171. [DOI] [PubMed] [Google Scholar]
- Boots & Jungheim (2015).Boots CE, Jungheim ES. Inflammation and human ovarian follicular dynamics. Seminars in Reproductive Medicine. 2015;33:270–275. doi: 10.1055/s-0035-1554928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen et al. (2017).Chen D, Zhang Z, Chen B, Ji D, Hao Y, Zhou P, Wei Z, Cao Y. Altered microRNA and Piwi-interacting RNA profiles in cumulus cells from patients with diminished ovarian reserve. Biology of Reproduction. 2017;97:91–103. doi: 10.1093/biolre/iox062. [DOI] [PubMed] [Google Scholar]
- Chin et al. (2002).Chin KV, Seifer DB, Feng B, Lin Y, Shih WC. DNA microarray analysis of the expression profiles of luteinized granulosa cells as a function of ovarian reserve. Fertility and Sterility. 2002;77:1214–1218. doi: 10.1016/s0015-0282(02)03114-x. [DOI] [PubMed] [Google Scholar]
- Chou & Chen (2018).Chou CH, Chen MJ. The effect of steroid hormones on ovarian follicle development. Vitamins and Hormones. 2018;107:155–175. doi: 10.1016/bs.vh.2018.01.013. [DOI] [PubMed] [Google Scholar]
- Cooper et al. (2011).Cooper AR, Baker VL, Sterling EW, Ryan ME, Woodruff TK, Nelson LM. The time is now for a new approach to primary ovarian insufficiency. Fertility and Sterility. 2011;95:1890–1897. doi: 10.1016/j.fertnstert.2010.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De La Fuente & Eppig (2001).De La Fuente R, Eppig JJ. Transcriptional activity of the mouse oocyte genome: companion granulosa cells modulate transcription and chromatin remodeling. Developmental Biology. 2001;229:224–236. doi: 10.1006/dbio.2000.9947. [DOI] [PubMed] [Google Scholar]
- Eisenberg et al. (2015).Eisenberg I, Kotaja N, Goldman-Wohl D, Imbar T. microRNA in human reproduction. Advances in Experimental Medicine and Biology. 2015;888:353–387. doi: 10.1007/978-3-319-22671-2_18. [DOI] [PubMed] [Google Scholar]
- Fabian, Sonenberg & Filipowicz (2010).Fabian MR, Sonenberg N, Filipowicz W. Regulation of mRNA translation and stability by microRNAs. Annual Review of Biochemistry. 2010;79:351–379. doi: 10.1146/annurev-biochem-060308-103103. [DOI] [PubMed] [Google Scholar]
- Fan et al. (2019).Fan Y, Chang Y, Wei L, Chen J, Li J, Goldsmith S, Silber S. Apoptosis of mural granulosa cells is increased in women with diminished ovarian reserve. 2019. 36:1225–1235. [DOI] [PMC free article] [PubMed]
- Flanagan, Chan & Leder (1991).Flanagan JG, Chan DC, Leder Transmembrane form of the kit ligand growth factor is determined by alternative splicing and is missing in the Sld mutant. Cell. 1991;64:1025–1035. doi: 10.1016/0092-8674(91)90326-t. [DOI] [PubMed] [Google Scholar]
- Gellersen, Fernandes & Brosens (2009).Gellersen B, Fernandes MS, Brosens JJ. Non-genomic progesterone actions in female reproduction. Human Reproduction Update. 2009;15:119–138. doi: 10.1093/humupd/dmn044. [DOI] [PubMed] [Google Scholar]
- Greaves et al. (2014).Greaves RF, Jevalikar G, Hewitt JK, Zacharin MR. A guide to understanding the steroid pathway: new insights and diagnostic implications. Clinical Biochemistry. 2014;47:5–15. doi: 10.1016/j.clinbiochem.2014.07.017. [DOI] [PubMed] [Google Scholar]
- Greenseid et al. (2011).Greenseid K, Jindal S, Hurwitz J, Santoro N, Pal L. Differential granulosa cell gene expression in young women with diminished ovarian reserve. Reproductive Sciences. 2011;18:892–899. doi: 10.1177/1933719111398502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howe et al. (2017).Howe V, Sharpe LJ, Prabhu AV, Brown AJ. New insights into cellular cholesterol acquisition: promoter analysis of human HMGCR and SQLE, two key control enzymes in cholesterol synthesis. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 2017;1862:647–657. doi: 10.1016/j.bbalip.2017.03.009. [DOI] [PubMed] [Google Scholar]
- Huang et al. (2019).Huang Y, Hu C, Ye H, Luo R, Fu X, Li X, Huang J, Chen W, Zheng Y. Inflamm-aging: a new mechanism affecting premature ovarian insufficiency. Journal of Immunology Research. 2019;2019:8069898. doi: 10.1155/2019/8069898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharara, Scott Jr & Seifer (1998).Sharara FI, Scott Jr RT, Seifer DB. The detection of diminished ovarian reserve in infertile women. American Journal of Obstetrics and Gynecology. 1998;179:804–812. doi: 10.1016/s0002-9378(98)70087-0. [DOI] [PubMed] [Google Scholar]
- Kanehisa et al. (2017).Kanehisa M, Furumichi M, Tanabe M, Sato Y, Morishima K. KEGG: new perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Research. 2017;45:D353–d361. doi: 10.1093/nar/gkw1092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyrou et al. (2009).Kyrou D, Kolibianakis EM, Venetis CA, Papanikolaou EG, Bontis J, Tarlatzis BC. How to improve the probability of pregnancy in poor responders undergoing in vitro fertilization: a systematic review and meta-analysis. Fertility and Sterility. 2009;91:749–766. doi: 10.1016/j.fertnstert.2007.12.077. [DOI] [PubMed] [Google Scholar]
- Lamb et al. (2006).Lamb J, Crawford ED, Peck D, Modell JW, Blat IC, Wrobel MJ, Lerner J, Brunet JP, Subramanian A, Ross KN, Reich M, Hieronymus H, Wei G, Armstrong SA, Haggarty SJ, Clemons PA, Wei R, Carr SA, Lander ES, Golub TR. The connectivity map: using gene-expression signatures to connect small molecules, genes, and disease. Science. 2006;313:1929–1935. doi: 10.1126/science.1132939. [DOI] [PubMed] [Google Scholar]
- Levi et al. (2001).Levi AJ, Raynault MF, Bergh PA, Drews MR, Miller BT, Scott Jr RT. Reproductive outcome in patients with diminished ovarian reserve. Fertility and Sterility. 2001;76:666–669. doi: 10.1016/s0015-0282(01)02017-9. [DOI] [PubMed] [Google Scholar]
- Liu et al. (2019).Liu Y, Wang L, Zhu L, Ran B, Wang Z. Bisphenol A disturbs transcription of steroidogenic genes in ovary of rare minnow Gobiocypris rarus via the abnormal DNA and histone methylation. Chemosphere. 2019;240:124935. doi: 10.1016/j.chemosphere.2019.124935. [DOI] [PubMed] [Google Scholar]
- Luense, Carletti & Christenson (2009).Luense LJ, Carletti MZ, Christenson LK. Role of dicer in female fertility. Trends in Endocrinology & Metabolism. 2009;20:265–272. doi: 10.1016/j.tem.2009.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mathews et al. (2014).Mathews ES, Mawdsley DJ, Walker M, Hines JH, Pozzoli M, Appel B. Mutation of 3-hydroxy-3-methylglutaryl CoA synthase I reveals requirements for isoprenoid and cholesterol synthesis in oligodendrocyte migration arrest, axon wrapping, and myelin gene expression. Journal of Neuroscience. 2014;34:3402–3412. doi: 10.1523/jneurosci.4587-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matzuk et al. (2002).Matzuk MM, Burns KH, Viveiros MM, Eppig JJ. Intercellular communication in the mammalian ovary: oocytes carry the conversation. Science. 2002;296:2178–2180. doi: 10.1126/science.1071965. [DOI] [PubMed] [Google Scholar]
- Medzhitov (2008).Medzhitov R. Origin and physiological roles of inflammation. Nature. 2008;454:428–435. doi: 10.1038/nature07201. [DOI] [PubMed] [Google Scholar]
- Nagaraja et al. (2008).Nagaraja AK, Andreu-Vieyra C, Franco HL, Ma L, Chen R, Han DY, Zhu H, Agno JE, Gunaratne PH, DeMayo FJ, Matzuk MM. Deletion of dicer in somatic cells of the female reproductive tract causes sterility. Molecular Endocrinology. 2008;22:2336–2352. doi: 10.1210/me.2008-0142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navot, Rosenwaks & Margalioth (1987).Navot D, Rosenwaks Z, Margalioth EJ. Prognostic assessment of female fecundity. Lancet. 1987;2:645–647. doi: 10.1016/s0140-6736(87)92439-1. [DOI] [PubMed] [Google Scholar]
- Oktem et al. (2017).Oktem O, Akin N, Bildik G, Yakin K, Alper E, Balaban B, Urman B. FSH stimulation promotes progesterone synthesis and output from human granulosa cells without luteinization. Human Reproduction. 2017;32:643–652. doi: 10.1093/humrep/dex010. [DOI] [PubMed] [Google Scholar]
- Parrott & Skinner (1999).Parrott JA, Skinner MK. Kit-ligand/stem cell factor induces primordial follicle development and initiates folliculogenesis. Endocrinology. 1999;140:4262–4271. doi: 10.1210/endo.140.9.6994. [DOI] [PubMed] [Google Scholar]
- Popovic, Sartorius & Christ-Crain (2019).Popovic M, Sartorius G, Christ-Crain M. Chronic low-grade inflammation in polycystic ovary syndrome: is there a (patho)-physiological role for interleukin-1? Seminars in Immunopathology. 2019;41:447–459. doi: 10.1007/s00281-019-00737-4. [DOI] [PubMed] [Google Scholar]
- Practice Committee of the American Society for Reproductive Medicine (2015).Practice Committee of the American Society for Reproductive Medicine Testing and interpreting measures of ovarian reserve: a committee opinion. Fertility and Sterility. 2015;103:e9–e17. doi: 10.1016/j.fertnstert.2014.12.093. [DOI] [PubMed] [Google Scholar]
- Reynolds et al. (2013).Reynolds KA, Omurtag KR, Jimenez PT, Rhee JS, Tuuli MG, Jungheim ES. Cycle cancellation and pregnancy after luteal estradiol priming in women defined as poor responders: a systematic review and meta-analysis. Human Reproduction. 2013;28:2981–2989. doi: 10.1093/humrep/det306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ritchie et al. (2015).Ritchie ME, Phipson B, Wu D, Hu Y, Law CW, Shi W, Smyth GK. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Research. 2015;43:e47. doi: 10.1093/nar/gkv007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ru et al. (2014).Ru Y, Kechris KJ, Tabakoff B, Hoffman P, Radcliffe RA, Bowler R, Mahaffey S, Rossi S, Calin GA, Bemis L, Theodorescu D. The multiMiR R package and database: integration of microRNA-target interactions along with their disease and drug associations. Nucleic Acids Research. 2014;42:e133. doi: 10.1093/nar/gku631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rupaimoole & Slack (2017).Rupaimoole R, Slack FJ. MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nature Reviews Drug Discovery. 2017;16:203–222. doi: 10.1038/nrd.2016.246. [DOI] [PubMed] [Google Scholar]
- Sabry et al. (2019).Sabry R, Yamate J, Favetta L, LaMarre J. MicroRNAs: potential targets and agents of endocrine disruption in female reproduction. Journal of Toxicologic Pathology. 2019;32:213–221. doi: 10.1293/tox.2019-0054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Said et al. (2016).Said RS, El-Demerdash E, Nada AS, Kamal MM. Resveratrol inhibits inflammatory signaling implicated in ionizing radiation-induced premature ovarian failure through antagonistic crosstalk between silencing information regulator 1 (SIRT1) and poly(ADP-ribose) polymerase 1 (PARP-1) Biochemical Pharmacology. 2016;103:140–150. doi: 10.1016/j.bcp.2016.01.019. [DOI] [PubMed] [Google Scholar]
- Schmidt et al. (2008).Schmidt J, Stoffels B, Moore BA, Chanthaphavong RS, Mazie AR, Buchholz BM, Bauer AJ. Proinflammatory role of leukocyte-derived Egr-1 in the development of murine postoperative ileus. Gastroenterology. 2008;135:926–936. doi: 10.1053/j.gastro.2008.05.079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sen et al. (2014).Sen A, Kushnir VA, Barad DH, Gleicher N. Endocrine autoimmune diseases and female infertility. Nature Reviews Endocrinology. 2014;10:37–50. doi: 10.1038/nrendo.2013.212. [DOI] [PubMed] [Google Scholar]
- Shannon et al. (2003).Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Research. 2003;13:2498–2504. doi: 10.1101/gr.1239303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharpe & Brown (2013).Sharpe LJ, Brown AJ. Controlling cholesterol synthesis beyond 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR) Journal of Biological Chemistry. 2013;288:18707–18715. doi: 10.1074/jbc.R113.479808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimamoto, Yamoto & Nakano (1993).Shimamoto T, Yamoto M, Nakano R. Possible involvement of protein kinase C in gonadotropin-induced ovulation in the rat ovary. Endocrinology. 1993;133:2127–2132. doi: 10.1210/endo.133.5.8404662. [DOI] [PubMed] [Google Scholar]
- Skiadas et al. (2012).Skiadas CC, Duan S, Correll M, Rubio R, Karaca N, Ginsburg ES, Quackenbush J, Racowsky C. Ovarian reserve status in young women is associated with altered gene expression in membrana granulosa cells. Molecular Human Reproduction. 2012;18:362–371. doi: 10.1093/molehr/gas008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szklarczyk et al. (2017).Szklarczyk D, Morris JH, Cook H, Kuhn M, Wyder S, Simonovic M, Santos A, Doncheva NT, Roth A, Bork P, Jensen LJ, Mering Cvon. The STRING database in 2017: quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Research. 2017;45:D362–d368. doi: 10.1093/nar/gkw937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabibzadeh (1994).Tabibzadeh S. Cytokines and the hypothalamic-pituitary-ovarian-endometrial axis. Human Reproduction. 1994;9:947–967. doi: 10.1093/oxfordjournals.humrep.a138621. [DOI] [PubMed] [Google Scholar]
- Tesfaye et al. (2016).Tesfaye D, Salilew-Wondim D, Gebremedhn S, Sohel MM, Pandey HO, Hoelker M, Schellander K. Potential role of microRNAs in mammalian female fertility. Reproduction, Fertility, and Development. 2016;29:8–23. doi: 10.1071/rd16266. [DOI] [PubMed] [Google Scholar]
- Vanacker et al. (2011).Vanacker J, Camboni A, Dath C, Van Langendonckt A, Dolmans MM, Donnez J, Amorim CA. Enzymatic isolation of human primordial and primary ovarian follicles with Liberase DH: protocol for application in a clinical setting. Fertility and Sterility. 2011;96:379–383. doi: 10.1016/j.fertnstert.2011.05.075. [DOI] [PubMed] [Google Scholar]
- Vinatier et al. (1995).Vinatier D, Dufour P, Tordjeman-Rizzi N, Prolongeau JF, Depret-Moser S, Monnier JC. Immunological aspects of ovarian function: role of the cytokines. European Journal of Obstetrics & Gynecology and Reproductive Biology. 1995;63:155–168. doi: 10.1016/0301-2115(95)02227-9. [DOI] [PubMed] [Google Scholar]
- Wang et al. (2018).Wang W, Chen J, Luo L, Li Y, Liu J, Zhang W. Effect of cadmium on kitl pre-mRNA alternative splicing in murine ovarian granulosa cells and its associated regulation by miRNAs. Journal of Applied Toxicology. 2018;38:227–239. doi: 10.1002/jat.3516. [DOI] [PubMed] [Google Scholar]
- Woo et al. (2018).Woo I, Christenson LK, Gunewardena S, Ingles SA, Thomas S, Ahmady A, Chung K, Bendikson K, Paulson R, McGinnis LK. Micro-RNAs involved in cellular proliferation have altered expression profiles in granulosa of young women with diminished ovarian reserve. Journal of Assisted Reproduction and Genetics. 2018;35:1777–1786. doi: 10.1007/s10815-018-1239-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu et al. (2012).Yu G, Wang LG, Han Y, He QY. clusterProfiler: an R package for comparing biological themes among gene clusters. Omics. 2012;16:284–287. doi: 10.1089/omi.2011.0118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan et al. (2014).Yuan S, Ortogero N, Wu Q, Zheng H, Yan W. Murine follicular development requires oocyte DICER, but not DROSHA. Biology of Reproduction. 2014;91:39. doi: 10.1095/biolreprod.114.119370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan et al. (2016).Yuan S, Wen J, Cheng J, Shen W, Zhou S, Yan W, Shen L, Luo A, Wang S. Age-associated up-regulation of EGR1 promotes granulosa cell apoptosis during follicle atresia in mice through the NF-kappaB pathway. Cell Cycle. 2016;15:2895–2905. doi: 10.1080/15384101.2016.1208873. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The red represents DOR samples and the black represents NOR samples. The lateral axis is the name of samples and longitudinal axis is the level of genes expression. DOR, diminished ovarian reserve; NOR, normal ovarian reserve.
The color change from red to blue represents the values change of gene expression from high to low. DEGs, differentially expressed genes.
The genes with blue are downregulated DEGs. KEGG, Kyoto Encyclopedia of Genes and Genomes; DEGs, differentially expressed genes.
*In publication of Woo et al., women who met two or more of the four criteria in the table except for age were defined as diminished ovarian reserve, and the parameter that was set to 7 represented for antral follicle count (AFC).
Data Availability Statement
The following information was supplied regarding data availability:
The raw data are available in the Supplemental File.