

Abstract
More than 50 years of efforts to identify the major cytokine responsible for red blood cell (RBC) production (erythropoiesis) led to the identification of erythropoietin (EPO) in 1977 and its receptor (EPOR) in 1989, followed by three decades of rich scientific discovery. We now know that an elaborate oxygen-sensing mechanism regulates the production of EPO, which in turn promotes the maturation and survival of erythroid progenitors. Engagement of the EPOR by EPO activates three interconnected signaling pathways that drive RBC production via diverse downstream effectors and simultaneously trigger negative feedback loops to suppress signaling activity. Together, the finely tuned mechanisms that drive endogenous EPO production and facilitate its downstream activities have evolved to maintain RBC levels in a narrow physiological range and to respond rapidly to erythropoietic stresses such as hypoxia or blood loss. Examination of these pathways has elucidated the genetics of numerous inherited and acquired disorders associated with deficient or excessive RBC production and generated valuable drugs to treat anemia, including recombinant human EPO and more recently the prolyl hydroxylase inhibitors, which act partly by stimulating endogenous EPO synthesis. Ongoing structure–function studies of the EPOR and its essential partner, tyrosine kinase JAK2, suggest that it may be possible to generate new “designer” drugs that control selected subsets of cytokine receptor activities for therapeutic manipulation of hematopoiesis and treatment of blood cancers.
Keywords: Erythropoiesis, hypoxia-induced transcription factor, prolyl hydroxylase inhibitor, erythropoietin receptor signaling, JAK2, Janus Kinase 2
Introduction
Healthy human adults produce about 200 billion red blood cells (RBCs) daily to replace those lost by senescence. This process, termed erythropoiesis, is exquisitely regulated by an oxygen-sensing mechanism that has evolved to maintain RBC numbers within a narrow physiological range 1– 3. Central to this mechanism is erythropoietin (EPO), a cytokine secreted by the kidney in response to low blood oxygen tension. Circulating EPO binds its cognate receptor (EPOR) on bone marrow erythroid progenitors, triggering multiple signaling pathways that support differentiation into mature RBCs. Inherited and acquired abnormalities in EPO production, its downstream activities, or its regulation cause numerous human diseases associated with too many or too few RBCs. The same pathways have been shaped by evolution for adaptation to life under chronic hypoxia at high altitudes. Although much is known about the production of EPO and its biological activities after 40 years of research, the topic remains a rich source for biomedical discovery and therapeutics. This review focuses on recent insights into the oxygen-regulated production of EPO and its actions on post-natal bone marrow erythropoiesis. It is important to note that EPO–EPOR signaling also drives RBC production during embryogenesis through similar but distinct mechanisms 4– 6.
History of EPO
In 1875, Denis Jourdanet and Paul Bert described anemia-like symptoms in patients living at high altitude and identified low blood oxygen level to be the primary mechanism 7. Building on this finding about 30 years later, Carnot and Deflandre discovered that infusion of serum from anemic rabbits into normal ones caused a rise in RBC count, predicting the existence of a circulating factor that stimulates erythropoiesis 3. In the early 1950s, studies using parabiotic rats validated the concept of a humoral erythropoiesis-stimulating agent 8, 9 that was shown by Erslev 9 to originate from the kidney 10. In 1977, Goldwasser’s group reported the purification of EPO from 2550 liters of urine collected from patients with aplastic anemia 11. Molecular cloning of the EPO gene in 1985 facilitated the manufacture of recombinant human EPO (rhEPO) protein for treating various forms of anemia 12, 13. This work led to discoveries of the EPOR by Lodish’s group in 1989 14 and subsequently multiple downstream signaling pathways were characterized by many laboratories. An elaborate oxygen-sensing mechanism that regulates EPO production was discovered in the early 1990s by William Kaelin Jr., Sir Peter Ratcliffe, and Gregg Semenza, who received the 2019 Nobel Prize in Physiology or Medicine for this work 15– 20.
Erythropoietic activities of EPO and EPOR
Multi-potent hematopoietic stem cells undergo a series of differentiation steps that successively restrict developmental potential, giving rise to lineage-committed progenitors ( Figure 1) 5. The first identifiable erythroid progenitor, termed “burst-forming unit-erythroid” (BFU-E), is defined by its ability to generate large colonies with scattered clusters of erythroblasts in semi-solid medium. Differentiation of BFU-E produces “colony-forming units-erythroid” (CFU-E) that generate smaller colonies containing about 50 cells. Proerythroblasts, the first recognizable erythroid precursor, undergo further maturation steps, which include specialized cell divisions, reduced cell size, elimination of most organelles, development of a specialized cell membrane to facilitate microcirculatory transit, and accumulation of hemoglobin for oxygen transport 1, 21, 22. Terminal erythroid maturation occurs in bone marrow erythroblastic islands composed of erythroid precursors surrounding a central macrophage 23. The morphological and functional definitions of committed erythroid progenitors have been augmented by the identification of stage-specific cell surface markers 24– 31 and, more recently, the discovery of their transcriptional states using single-cell RNA sequencing (scRNAseq) 32, 33.
Figure 1. Erythropoietin (EPO) activity during erythropoiesis.

Classic hierarchy of hematopoiesis with stages of red blood cell (RBC) development shown in greater detail. The major site of EPO action is indicated. Genetic and cell culture studies have shown that EPO is required for the development of CFU-E into late-stage erythroblasts. NK, natural killer. Multi-potent hematopoietic progenitors include the following: CLP, common lymphoid progenitor; CMP, common myeloid progenitor; LT-HSC, long-term engrafting hematopoietic stem cell; MEP, megakaryocytic-erythroid progenitor; ST-HSC, short-term hematopoietic stem cell. Committed erythroid progenitors include the following: BFU-E, burst-forming unit-erythroid; CFU-E, colony-forming unit-erythroid. Erythroid precursors include the following: BasoE, basophilic erythroblast; OrthoE, orthochromatic erythroblast; PolyE, polychromatic erythroblast; ProE, proerythroblast; Retic, reticulocyte.
Although multiple cytokines support erythropoiesis 34, EPO is the key physiological regulator. Loss of EPO or derangements in EPO signaling in mice or humans cause anemia 4, 35 while excessive EPO production or EPOR signaling or both cause pathologically increased RBC numbers 36– 38. EPO acts mainly on CFU-E progenitors and proerythroblasts to maintain their survival and facilitate terminal maturation ( Figure 1) 25, 39– 41. Additionally, EPO can stimulate cell proliferation and drive multi-potent hematopoietic progenitors toward an erythroid fate 40, 42 but is not required for erythroid lineage commitment 4. In vivo administration of EPO leads to rapid skewing of multi-potential progenitors away from myeloid and toward the erythroid lineage and to altered gene expression in BFU-E and CFU-E progenitors 32.
An oxygen-sensitive feedback loop regulates EPO production
Post-natal EPO production occurs mainly in peritubular fibroblast-like interstitial cells of the kidney 43– 50 but also in liver, spleen, bone marrow, lungs, and brain 51– 53 and is regulated by blood oxygen levels through a transcriptional feedback loop ( Figure 2) 15– 19. The hypoxia-inducible transcription factor (HIF) complex binds hypoxia response elements in the EPO gene promoter to stimulate its transcription. Functional HIF is a heterodimer composed of an α subunit (HIFα) and a β subunit (HIFβ, also known as aryl hydrocarbon receptor nuclear translocator or ARNT). The stability of HIF is regulated by prolyl hydroxylase domain (PHD) enzymes, which use oxygen and 2-oxoglutarate to catalyze the hydroxylation of specific proline residues in HIFα, thereby stimulating binding of the HIF heterodimer to the von Hippel–Lindau protein (pVHL) component of an E3 ubiquitin ligase complex 3, 54, 55. Subsequent polyubiquitination of HIF leads to its proteasomal degradation. At low cellular oxygen concentrations, the PHD proteins are inactive and HIF is stabilized for target gene activation. Another 2-oxoglutarate–dependent oxygenase, factor inhibiting HIF (FIH), stimulates the oxygen-dependent hydroxylation of a specific asparagine residue in HIFα, which inhibits its activity by blocking HIFα binding to the transcriptional co-activator p300 55, 56. In these ways, the PHD and FIH enzymes act as oxygen sensors that inhibit the production of EPO and other HIF targets under oxygen-replete conditions. Remarkably, HIF also activates hundreds of genes besides EPO. Other HIF target genes encode glycolytic enzymes, angiogenic factors, and iron uptake proteins, representing a concerted hypoxia response to increase RBC production, manufacture hemoglobin, enhance tissue perfusion, and promote oxygen-independent metabolism through glycolysis 54, 57– 59.
Figure 2. Regulation of endogenous erythropoietin (EPO) gene transcription by the oxygen-sensitive hypoxia-inducible factor (HIF) pathway.
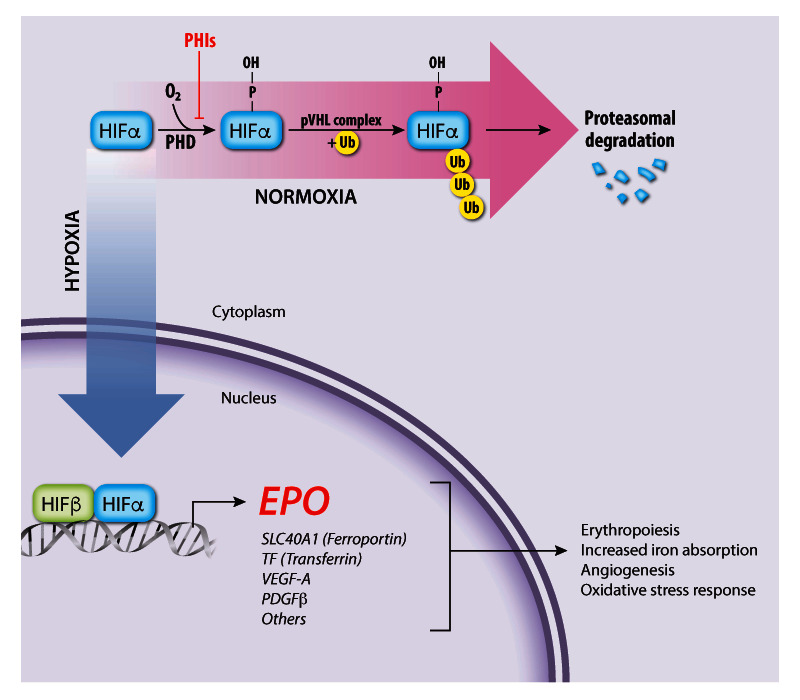
The HIF transcription factor heterodimer (HIFα–HIFβ) activates the EPO gene and numerous other genes that promote tissue oxygen delivery. At high oxygen concentrations, prolyl hydroxylase (PHD) enzymes hydroxylate the HIFα subunit, targeting it for ubiquitination by the von Hippel–Lindau protein (pVHL) ubiquitin ligase complex followed by proteasomal degradation. Under hypoxia, PHD enzymes are inactive, thereby stabilizing HIF, which activates transcription of EPO and other target genes involved in tissue oxygen delivery. PHD inhibitors (PHIs) such as roxadustat and vadadustat stabilize HIFα and are under investigation for treating anemia associated with chronic renal failure. PDGFβ, platelet-derived growth factor beta; SLC40A1, solute carrier family 40 member 1; TF, Transferrin; VEGF-A, vascular endothelial growth factor A.
Mammals express three HIFα isoforms (HIF-1α, -2α, and -3α) and three PHD isoforms (PHD1, 2, and 3), each encoded by separate genes with overlapping but distinct tissue distributions and functions 60. The production of EPO in adult life is regulated mainly by HIF-2α and PHD2 61. Perhaps not surprisingly, germline and somatic mutations affecting the PHD–HIF–EPO regulatory pathway are associated with erythrocytosis, anemia, abnormal angiogenesis, and cancer 62, 63. In mice and humans, loss-of-function mutations in PHD2 and VHL or gain-of-function missense mutations that stabilize HIF-2α by inhibiting its binding to PHD2 or VHL cause erythrocytosis 64. An interesting gain-of-function mutation in the EPO gene (c.32delG) was recently identified to cause autosomal dominant erythrocytosis in a multi-generational pedigree 65. The single-nucleotide deletion introduces a frameshift into the main EPO mRNA but initiates excess production of EPO from what is normally a non-coding EPO mRNA transcribed from an alternative promoter in intron 1. Variants in the PHD–HIF–EPO pathway have also been selected for in evolution as an adaptive mechanism to living at high altitude. Some of these variants attenuate hypoxia-induced erythrocytosis that can cause deleterious hyperviscosity syndromes 64, 66– 69. These clinical observations highlight the exquisite and complex genetic regulation of EPO production and erythropoiesis. Of note, only one isoform of FIH has been identified. Ablation of the corresponding gene in mice causes metabolic alterations but does not appear to alter the canonical HIF functions in erythropoiesis or angiogenesis 70.
EPO activities are mediated through the EPOR
EPO drives erythropoiesis by stimulating the EPOR on the surface of erythroid progenitors. The EPOR is a member of the type I cytokine receptor family distinguished by a conserved extracellular WSXWS amino acid motif, a single-transmembrane domain, and a cytoplasmic tail that lacks intrinsic tyrosine kinase activity 71. The proximal cytoplasmic domain of EPOR is bound by the JAK2 tyrosine kinase. Binding of a single EPO molecule to two EPOR molecules triggers a conformational change that stimulates JAK2 to initiate a multi-tiered signaling cascade ( Figure 3) 72, 73. Activated JAK2 phosphorylates itself and several tyrosine residues on the EPOR cytoplasmic tail, which serve as docking sites to engage SH2-containing signaling molecules such as the STAT5 (signal transducer and activator of transcription 5) transcription factor. Following phosphorylation and activation by JAK2, STAT5 enters the nucleus to activate numerous target genes 74. Biologically important erythroid STAT5 target genes include the following: BCL2L1, which prevents apoptosis of late-stage erythroblasts 75– 77; ID1, which promotes erythroblast expansion and survival 78; TRIB3, which regulates erythroid maturation 79; SPI2A, which encodes a serpin protease with antioxidant activities 80; and TFRC (transferrin receptor protein 1), which mediates iron uptake 81, 82. The recently discovered STAT5 target gene erythroferrone ( ERFE) encodes a hormone that acts on hepatocytes to inhibit their production of hepcidin, a different hormone that blocks intestinal iron absorption and release of iron stores from macrophage 83. By stimulating the production of ERFE in erythroblasts, EPO increases bioavailable iron for hemoglobin synthesis 83, 84. In addition to STAT5, EPOR activates the canonical Ras/mitogen-activated protein kinase (MAPK) and phosphoinositide-3 kinase (PI3K)/Akt pathways to enhance erythroid progenitor survival, proliferation, and differentiation 3, 85– 89. The Akt kinase also activates FOXO3, a transcription factor that induces genes that control antioxidant pathways 90, 91, cell polarity, and enucleation 92. Other signaling molecules activated by EPOR include Lyn kinase and PLCγ, although their contributions to erythropoiesis are less clear 93, 94.
Figure 3. Activation of erythropoietin receptor (EPOR) by EPO.
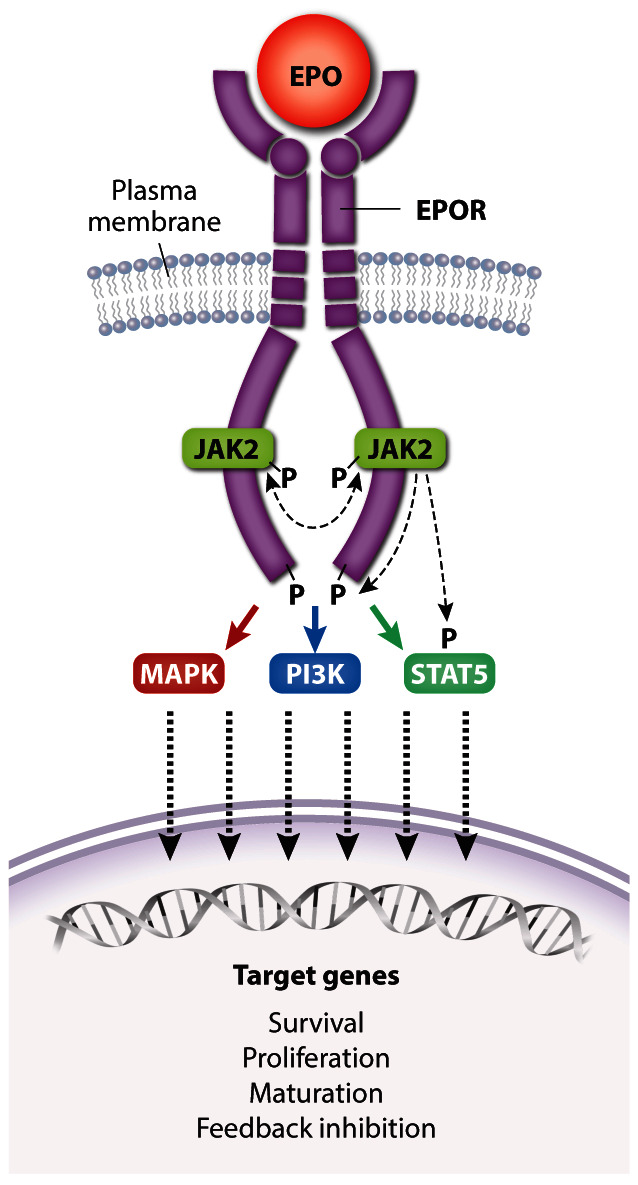
A single EPO molecule binds and stabilizes EPOR–JAK2 complex dimers, inducing a conformational change that initiates JAK2 trans-phosphorylation and activation. Active JAK2 phosphorylates multiple tyrosine residues on STAT5 and the cytoplasmic domain of EPOR, triggering a signaling cascade that activates numerous effector pathways contributing to biological activity. Dashed lines represent kinase activity. For simplicity, the kinase activity of only one JAK2 protein is indicated. Major signaling pathways activated by EPOR include Ras/MAPK, STAT5, and PI3K/Akt, which drive the expression of genes that promote erythroid progenitor survival, proliferation, and differentiation as well as feedback inhibition of EPOR signaling. MAPK, mitogen-activated protein kinase; PI3K, phosphoinositide-3 kinase; STAT5, signal transducer and activator of transcription 5.
Although some EPO–EPOR effectors can be linked directly to activation of a single linear signaling pathway, overgeneralizing this concept may be biologically inaccurate. As postulated for cytokine receptor signaling in general 95, the biological functions of EPOR are likely to be regulated by cross-communications between its numerous downstream signaling pathways and signaling by other cytokine receptors. In regard to the latter, cooperative signaling between the EPOR and stem cell factor receptor (KIT) is believed to promote erythropoiesis 96– 99. The EPOR also binds the type 2 transferrin receptor (TFR2), which is expressed in hepatocytes and erythroid progenitors. In hepatocytes, the TFR2 stimulates hepcidin production and germline TFR2 mutations cause iron overload (hemochromatosis type 3) 100. In erythroid progenitors, TFR2 binds EPOR in the endoplasmic reticulum and facilitates its transport to the cell surface 101. The effects of TFR2 in erythroid progenitors appear to be context-dependent and are not fully resolved. In cultured erythroblasts, suppression of TFR2 inhibits erythropoiesis 101. In contrast, hematopoietic-specific ablation of the Tfr2 gene in mice enhances erythropoiesis, likely by modulating EPO sensitivity 102. Expression of TFR2 in the kidney may inhibit EPO production 103. Interaction with iron-bound transferrin stabilizes TFR2 at the cell surface 104, representing a potential mechanism by which TFR2 coordinates erythropoietic rate and enteral iron uptake with circulating iron level.
Signaling through EPO–EPOR promotes both basal erythropoiesis, which maintains homeostasis by replacing erythrocytes lost by normal senescence, and “stress erythropoiesis” associated with increased synthetic demands caused by bleeding, excessive RBC destruction, or hypoxia. Relatively low concentrations of EPO during basal erythropoiesis are thought to act mainly by inhibiting apoptosis of erythroid progenitors, while stress erythropoiesis induces higher EPO concentrations that can drive hematopoietic differentiation toward the erythroid fate 25, 32, 40, 41. In line with this notion, EPO is able to act like a dimmer switch in activating STAT5 over a wide concentration range 105, and functional genomic approaches are beginning to identify direct targets of EPO-activated STAT5 in erythropoiesis 74. Different EPO concentrations during basal and stress erythropoiesis are likely to engage distinct signaling modalities, as revealed by a “knock-in” mouse strain in which EPOR is replaced with a truncated version (EPOR-HM) that binds and activates JAK2 but lacks the cytoplasmic portion containing all JAK2 tyrosine substrates 106. EPOR-HM mice are viable with a mild defect in steady-state erythropoiesis but are unable to support stress erythropoiesis 106. Thus, phosphotyrosine signaling from the EPOR is selectively required for stress erythropoiesis. The signaling pathways that are activated in response to high EPO concentrations differ depending on whether stress is chronic or acute. For example, STAT5-mediated activation of BCL2L1 occurs rapidly after acute bleeding or hypoxia, and then decays, even if high levels of EPO persist. In contrast, persistent or chronic stress conditions such as β-thalassemia elicit a distinct set of EPOR signaling pathways that include the EPOR-mediated suppression of pro-apoptosis genes FAS and BCL2L11 (formerly BIM) 32.
EPO signal termination
Activation of EPOR by EPO is balanced by complex negative feedback mechanisms that fine-tune and inhibit signaling to prevent excessive RBC production. Initial evidence for this came from studies of a Finnish family ascertained through an Olympic cross-country skier 107. This family and others discovered subsequently were found to have erythrocytosis caused by EPOR-truncating mutations that eliminate portions of the cytoplasmic domain, which later was found to negatively regulate EPOR signaling by recruiting various inhibitory proteins, including the tyrosine phosphatase PTPN6, members of the suppressor of cytokine signaling (SOCS) protein family, SH2B adapter protein 3 (SH2B3, LNK), and the p85 regulatory subunit of PI3K 38. Mechanistically, PTPN6 attenuates EPOR signaling by dephosphorylating JAK2 108, 109. CISH and SOCS3 block access of STAT5 to the EPOR, whereas SOCS1 binds to the JAK2 kinase domain and reduces its tyrosine kinase activity 110. Transcription of SOCS1, SOCS3, and CISH are induced by STAT5, forming a negative feedback loop 111. Mutations in JAK2 at the SOCS3 binding site and mutations in SOCS3 occur in patients with erythrocytosis 112, 113. The SH2B3 protein (LNK) is upregulated and phosphorylated in response to EPO and inhibits EPOR signaling by binding phosphotyrosine residues in JAK2 and the cytoplasmic tail of EPOR 114, 115. Sh2b3 −/− mice exhibit features of myeloproliferative neoplasms (MPNs) such as splenomegaly and extramedullary hematopoiesis, and inactivating SH2B3 mutations are associated with myeloproliferative disease in humans 115. Genome-wide association studies have identified a hypomorphic SH2B3 variant associated with elevated hemoglobin and RBC counts 116, 117, and suppression of SH2B3 production by RNA interference improved the production of RBCs by in vitro differentiation of human CD34 + cells and embryonic stem cells 118.
The EPOR is also negatively regulated at the protein level by several mechanisms 119, 120. First, the p85 protein, which facilitates EPOR signaling as a regulatory subunit for PI3K 121, also promotes EPOR endocytosis and degradation 122, 123. Upon EPO stimulation, the casitas B-lineage lymphoma (CBL) protein ubiquitinates p85 bound to the cytoplasmic domain of EPOR, facilitating interaction with the adaptor protein Epsin-1 to promote endocytosis. Second, prolyl hydroxylase D3 (PHD3)-mediated proline hydroxylation of EPOR stimulates its proteasomal degradation 124. Third, iron deficiency reduces the expression of EPOR through interactions with TFR2 and Scribble, a scaffold protein that facilitates EPOR recycling 125. This mechanism may explain EPO resistance associated with iron deficiency. Dipeptidylpeptidase (DPP4, CD26) expressed on hematopoietic and stromal cells truncates EPO into inactive fragments, reducing its plasma activity 126. These examples illustrate how EPOR signaling is terminated by many proteins acting through multiple mechanisms, most of which are components of a negative feedback loop triggered by EPOR activation.
Recent insights into EPO–EPOR signaling
The discovery of activating JAK2 mutations in MPNs has fueled the development of ruxolitinib and other JAK2 inhibitors. Ruxolitinib induces clinical responses and improves survival in some patients with MPN but its overall effects and therapeutic index are relatively modest 127. New structure–function studies of EPOR and JAK2 may inform the rational design of novel drugs for MPNs. Binding of JAK2 to nascent EPOR in the endoplasmic reticulum facilitates its trafficking to the plasma membrane 128. The importance of this protein interaction is freshly reinforced by findings that MPN-associated JAK2 mutants use EPOR as a scaffold for recruiting downstream substrates in order to drive EPO-independent erythrocytosis 129. Moreover, changes in the JAK2 pseudokinase domain, which does not interact with EPOR directly, can affect EPOR–JAK2 association ( Figure 4) 129. Single-molecule fluorescence microscopy showed that EPO stimulates self-association of EPOR-bound JAK2 through its pseudokinase domain and that MPN-associated JAK2 mutations strengthen this interaction in the absence of EPO 130. Thus, mutant JAK2 proteins drive EPO-independent EPOR signaling by enhancing dimerization of EPOR–JAK2 complexes. Mutant JAK2 also drives MPN by stimulating ligand-independent activation of the thrombopoietin receptor, which is structurally similar to the EPOR. The authors note that MPNs might be treated by drugs which inhibit self-interaction of the JAK2 pseudokinase domain. Although JAK2 binds EPOR through its cytoplasmic box 1, subsequent activation requires another EPOR conserved region, termed the “hydrophobic switch” 131. Crystallographic data suggest that this region positions EPOR–JAK2 molecules into a specific conformation that facilitates JAK2 activation 132.
Figure 4. JAK2 regulates dimerization of the EPOR–JAK2 complex.
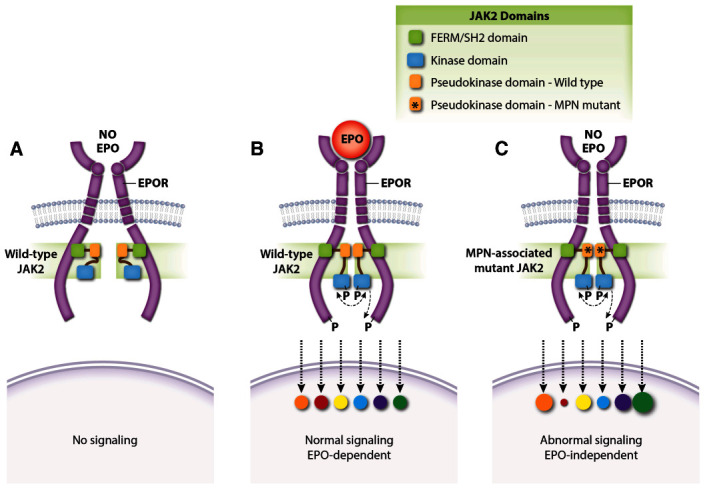
( A) Normal EPOR–JAK2 complexes are inert without EPO. ( B) EPO binding to EPOR stabilizes the EPOR–JAK2 complex and triggers downstream signaling by activating JAK2. Colored circles represent normalized activation levels of EPOR signaling targets. ( C) Myeloproliferative neoplasm (MPN)-associated mutations in the JAK2 pseudokinase domain, which does not interact directly with EPOR, stabilize dimerization of EPOR–JAK2 complexes and activate JAK2 in the absence of EPO. Mutations in the linker region separating the FERM-SH2 and pseudokinase domains (exon 12) act similarly (not shown). Constitutive activation of the EPOR by MPN-associated mutations causes abnormal downstream signaling relative to that induced by EPO 142– 146. EPO, erythropoietin; EPOR, erythropoietin receptor.
Activated EPOR triggers multiple signaling pathways that interact to specify the activation of different effectors and biological output. Medically relevant insights into this problem were gained by the discovery of a patient with pure red cell aplasia (Diamond–Blackfan anemia) caused by a homozygous EPO gene missense mutation (R150Q) 133. Although the mutant EPO protein exhibited only threefold reduced steady-state affinity for EPOR, kinetic studies revealed faster on-rate (k on) and off-rate (k off) ( Figure 5). Abnormally rapid release of the mutant EPO from EPOR was associated with impaired EPOR dimerization and reduced JAK2 activation. Remarkably, alterations in downstream phospho-signaling elicited by the mutant EPO were highly selective. Thus, erythropoietic failure was not caused by complete loss of EPO activity but rather by altered function. This study shows how variability in ligand-induced conformational changes of a cytokine receptor (in this case, EPO on- and off-rates) can selectively alter downstream signaling and biology.
Figure 5. A pathological erythropoietin (EPO) mutant with altered binding kinetics to EPO receptor (EPOR) causes qualitative changes in downstream signaling.
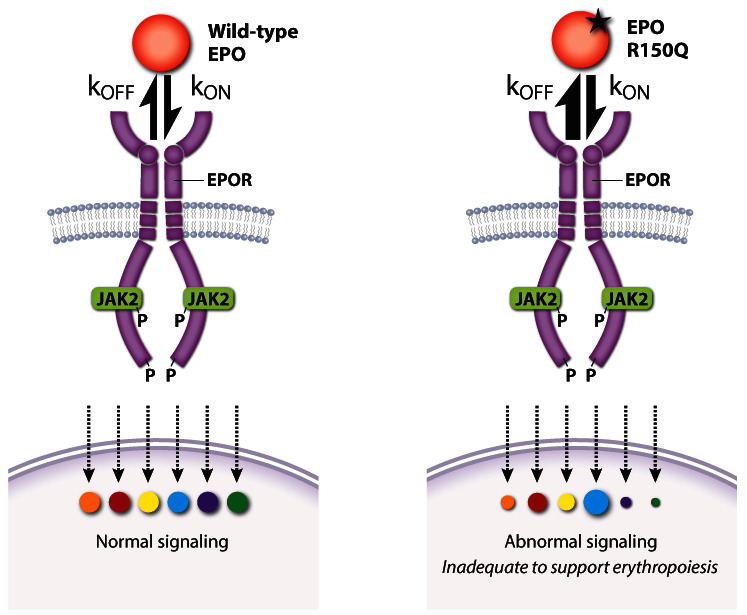
A homozygous p.R150Q EPO mutation was discovered in a patient with severe anemia caused by pure red cell aplasia (Diamond–Blackfan anemia). High levels of the mutant EPO failed to restore erythropoiesis despite an only threefold reduction in its overall affinity for EPOR. Compared with wild-type EPO, the mutant EPO interaction with EPOR was kinetically biased with higher on- and off-rates that altered the activation of specific EPOR effector pathways. k on, rate of association; k off, rate of dissociation. This figure was created using data from Kim et al. 2017 133.
Two other studies examined EPO–EPOR structure–function relationships more systematically by designing a series of EPOR ligands that generate different homodimer topologies, resulting in qualitative variation in signaling output 73, 134. These findings have potential medical implications. For example, one study showed that different artificial ligands that resulted in different angles and distance between EPOR homodimer subunits generated unique signaling patterns with stage-selective effects on hematopoiesis ( Figure 6) 134. The other study 73 identified artificial EPOR ligands that can block EPO-independent signaling by the MPN-associated mutation JAK2V617F, which may inform new therapies for MPNs 134. Overall, understanding and controlling the signaling output of different EPOR–JAK2 homo-dimer conformations may be used to precisely manipulate hematopoiesis or suppress pathologically active signaling. In this regard, many such studies performed on EPOR are generalizable to other cytokine receptors 135– 137.
Figure 6. Altered topology of the erythropoietin receptor (EPOR) extracellular domains (ECDs) induced by engineered ligands produces qualitative changes in downstream signaling.

( A) Wild-type erythropoietin (EPO) causes normal activation of EPOR signaling targets. Engineered EPOR ligands that modify the angle ( B) or distance ( C) between EPOR ECDs produce selective alterations in the activation of downstream signaling targets. This figure was created using data from Mohan et al., 2019 134.
Pharmacologic stimulation of erythropoiesis
rhEPO is used to treat anemia associated with a variety of diseases 13, 138. The most common indication is anemia of chronic renal failure where rhEPO increases blood hemoglobin levels and improves quality of life 139. In general, patients with renal failure have high levels of hepcidin, which limits iron absorption and availability for erythropoiesis 140, 141. Thus, rhEPO therapy usually requires administration of intravenous iron 139. Although the benefits of rhEPO in renal failure are clear, its overaggressive use is associated with increased rates of arteriovenous fistula thrombosis, venous thromboembolism, congestive heart failure, myocardial infarction, and death 138, 147– 150. Similarly, rhEPO use for cancer-related anemia has been associated with reduced survival 151. Elevated blood viscosity caused by increased RBC mass probably contributes to these adverse events. Additionally, some adverse effects of rhEPO may result from stimulation of EPOR signaling in non-erythroid tissues or tumor cells or both 151– 157, although this point is complicated by technical difficulties in establishing the presence of EPOR in non-erythroid tissues because of non-specific antibody interactions 158. Regardless, current guidelines recommend careful titration of rhEPO dosing in patients with renal failure 139. A recent study showed that, compared with low-dose intravenous iron sucrose, high-dose intravenous iron sucrose therapy in chronic renal failure resulted in reduced dosage requirements for rhEPO, fewer major adverse cardiovascular events, and lower death rates 159.
The routine use of rhEPO in most cancer patients who are undergoing curative chemotherapy should be avoided 160. rhEPO remains an important drug for treating anemia associated with myelodysplastic syndrome, although responses are often transient 161– 163. From a historical perspective, the first rhEPO (epoetin alfa) was approved for clinical use in 1989 and the longer-acting darbepoetin was approved in 2001. Combined sales reached about $5 billion per year in 2005 and then declined by 40% over the next 6 years as the price of the drugs dropped and the potential adverse effects became recognized 164.
The prolyl hydroxylase inhibitors (PHIs), which act by stabilizing HIFα to stimulate endogenous EPO production ( Figure 2), are promising new agents for treating anemia of chronic kidney disease and perhaps other etiologies 165, 166. Numerous clinical studies have shown that PHIs are effective for raising hemoglobin levels in subjects with chronic renal failure 166– 170. Compared with rhEPO, PHIs offer several potential advantages, including oral administration, improved iron utilization possibly due to suppression of hepcidin, lowering of plasma lipids and cholesterol, and efficacy at relatively low plasma concentrations of endogenous EPO, which may reduce cardiovascular toxicities. Three PHIs are in advanced phase III clinical development, and one was recently approved for clinical use in China 171. Although the drugs have been shown to be relatively safe in clinical trials, there are numerous theoretical concerns related to on-target effects given the extensive number of genes and biological pathways that are regulated by the HIF transcription factors. Potential adverse effects include alterations in metabolism, immune response, vascular tone, and angiogenesis. Monitoring for these problems is required in more extended clinical trials pre- and post-marketing.
Conclusions
EPO and its receptor are essential for the differentiation of CFU-E progenitors into mature RBCs. The complex, multi-layered biochemical pathways that regulate EPO production, signal through EPO engagement of EPOR, and extinguish EPOR signaling are all geared to maintain circulating RBC numbers in a narrow physiological range at steady state and during erythropoietic stress. Examination of these processes over more than 40 years has elucidated fundamental concepts of general biology, defined the mechanisms of human diseases associated with over- or under-production of RBCs, and produced a remarkably useful biological drug to treat some forms of anemia. The development of rhEPO, and more recently PHD inhibitors, arose from basic biological research and represents excellent paradigms for “bench to bedside and back” therapeutic development. Despite the tremendous knowledge gained through extensive studies of EPO and EPOR over many years, the field remains a fruitful area of research, as illustrated by ongoing efforts to better understand the complexities of the PHD–HIF–EPO pathway, structure–function regulation of EPO–EPOR–JAK2 signaling, and mechanisms of human disease caused by germline and somatic alterations in genes tied to EPO biology. In fact, interesting and medically relevant research problems related to EPO are too numerous to cover in a single review. Topics not covered here include the biology of EPOR signaling in non-erythroid tissues and its role in metabolic pathways 149, 151, 172– 179. Thus, it is likely that laboratory scientists and clinical researchers who study EPO-related biology and medicine will continue to generate exciting and clinically useful findings for many years to come.
Acknowledgments
We thank Anita Impagliazzo, CMI (Anita Impagliazzo Medical Illustration) for assistance with the figures and Merav Socolovsky (Department of Molecular, Cell and Cancer Biology and Department of Pediatric, Division of Hematology/Oncology at the University of Massachusetts Medical School) for critical review of the manuscript.
Editorial Note on the Review Process
F1000 Faculty Reviews are commissioned from members of the prestigious F1000 Faculty and are edited as a service to readers. In order to make these reviews as comprehensive and accessible as possible, the referees provide input before publication and only the final, revised version is published. The referees who approved the final version are listed with their names and affiliations but without their reports on earlier versions (any comments will already have been addressed in the published version).
The referees who approved this article are:
Vijay G Sankaran, Division of Hematology/Oncology, Boston Children's Hospital, Harvard Medical School, Boston, MA, USA; Department of Pediatric Oncology, Dana-Farber Cancer Institute, Harvard Medical School, Boston, MA, USA; Broad Institute of Harvard and Massachusetts Institute of Technology (MIT), Cambridge, MA, USA; Harvard Stem Cell Institute, Cambridge, MA, USA
Mohandas Narla, New York Blood Center, New York, NY, USA
Constance Tom Noguchi, Molecular Medicine Branch, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD, USA
Wolfgang Jelkmann, Institute of Physiology, University of Lübeck (retired), Lübeck, Germany
Funding Statement
LJH is funded by National Institutes of Health (NIH) grant R01 HL089966. MJW is funded by NIH grants R01 DK61692 and P01 HL053749, the Doris Duke Charitable Foundation, and the American Lebanese Syrian Associated Charities (ALSAC).
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
[version 1; peer review: 4 approved]
References
- 1. Moras M, Lefevre SD, Ostuni MA: From Erythroblasts to Mature Red Blood Cells: Organelle Clearance in Mammals. Front Physiol. 2017;8:1076. 10.3389/fphys.2017.01076 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Koury MJ: Abnormal erythropoiesis and the pathophysiology of chronic anemia. Blood Rev. 2014;28(2):49–66. 10.1016/j.blre.2014.01.002 [DOI] [PubMed] [Google Scholar]
- 3. Bunn HF: Erythropoietin. Cold Spring Harb Perspect Med. 2013;3(3):a011619. 10.1101/cshperspect.a011619 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Wu H, Liu X, Jaenisch R, et al. : Generation of committed erythroid BFU-E and CFU-E progenitors does not require erythropoietin or the erythropoietin receptor. Cell. 1995;83(1):59–67. 10.1016/0092-8674(95)90234-1 [DOI] [PubMed] [Google Scholar]
- 5. Palis J: Primitive and definitive erythropoiesis in mammals. Front Physiol. 2014;5:3. 10.3389/fphys.2014.00003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Malik J, Kim AR, Tyre KA, et al. : Erythropoietin critically regulates the terminal maturation of murine and human primitive erythroblasts. Haematologica. 2013;98(11):1778–87. 10.3324/haematol.2013.087361 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. West JB, Richalet JP: Denis Jourdanet (1815-1892) and the early recognition of the role of hypoxia at high altitude. Am J Physiol Lung Cell Mol Physiol. 2013;305(5):L333–40. 10.1152/ajplung.00128.2013 [DOI] [PubMed] [Google Scholar]
- 8. Reissmann KR: Studies on the mechanism of erythropoietic stimulation in parabiotic rats during hypoxia. Blood. 1950;5(4):372–80. [PubMed] [Google Scholar]
- 9. Erslev A: Humoral Regulation of Red Cell Production. Blood. 1953;8(4):349–57. [PubMed] [Google Scholar]
- 10. Jacobson LO, Goldwasser E, Fried W, et al. : Role of the kidney in erythropoiesis. Nature. 1957;179:633–4. 10.1038/179633a0 [DOI] [PubMed] [Google Scholar]
- 11. Miyake T, Kung CK, Goldwasser E: Purification of human erythropoietin. J Biol Chem. 1977;252(15):5558–64. [PubMed] [Google Scholar]
- 12. Lin FK, Suggs S, Lin CH, et al. : Cloning and expression of the human erythropoietin gene. Proc Natl Acad Sci U S A. 1985;82(22):7580–4. 10.1073/pnas.82.22.7580 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Kalantar-Zadeh K: History of Erythropoiesis-Stimulating Agents, the Development of Biosimilars, and the Future of Anemia Treatment in Nephrology. Am J Nephrol. 2017;45(3):235–47. 10.1159/000455387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. D’Andrea AD, Lodish HF, Wong GG: Expression cloning of the murine erythropoietin receptor. Cell. 1989;57(2):277–85. 10.1016/0092-8674(89)90965-3 [DOI] [PubMed] [Google Scholar]
- 15. Semenza GL, Nejfelt MK, Chi SM, et al. : Hypoxia-inducible nuclear factors bind to an enhancer element located 3' to the human erythropoietin gene. Proc Natl Acad Sci U S A. 1991;88(13):5680–4. 10.1073/pnas.88.13.5680 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Maxwell PH, Wiesener MS, Chang GW, et al. : The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature. 1999;399(6733):271–5. 10.1038/20459 [DOI] [PubMed] [Google Scholar]
- 17. Wang GL, Jiang BH, Rue EA, et al. : Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci U S A. 1995;92(12):5510–4. 10.1073/pnas.92.12.5510 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Ivan M, Kondo K, Yang H, et al. : HIFalpha targeted for VHL-mediated destruction by proline hydroxylation: Implications for O2 sensing. Science. 2001;292(5516):464–8. 10.1126/science.1059817 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 19. Jaakkola P, Mole DR, Tian YM, et al. : Targeting of HIF-alpha to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science. 2001;292(5516):468–72. 10.1126/science.1059796 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 20. Johnson R: How cells sense and adapt to oxygen availability.Stockholm: Nobel Foundation;2019[cited 2020 June]. Reference Source [Google Scholar]
- 21. Dzierzak E, Philipsen S: Erythropoiesis: Development and differentiation. Cold Spring Harb Perspect Med. 2013;3(4):a011601. 10.1101/cshperspect.a011601 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Jelkmann W: Functional significance of erythrocytes.In: Lang F, Foller M, editors. Erythrocytes. London: Imperial College Press;20121–56. 10.1142/9781848166219_0001 [DOI] [Google Scholar]
- 23. Lee SH, Crocker PR, Westaby S, et al. : Isolation and immunocytochemical characterization of human bone marrow stromal macrophages in hemopoietic clusters. J Exp Med. 1988;168(3):1193–8. 10.1084/jem.168.3.1193 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Chen K, Liu J, Heck S, et al. : Resolving the distinct stages in erythroid differentiation based on dynamic changes in membrane protein expression during erythropoiesis. Proc Natl Acad Sci U S A. 2009;106(41):17413–8. 10.1073/pnas.0909296106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Liu Y, Pop R, Sadegh C, et al. : Suppression of Fas-FasL coexpression by erythropoietin mediates erythroblast expansion during the erythropoietic stress response in vivo. Blood. 2006;108(1):123–33. 10.1182/blood-2005-11-4458 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Pronk CJH, Rossi DJ, Månsson R, et al. : Elucidation of the phenotypic, functional, and molecular topography of a myeloerythroid progenitor cell hierarchy. Cell Stem Cell. 2007;1(4):428–42. 10.1016/j.stem.2007.07.005 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 27. Hu J, Liu J, Xue F, et al. : Isolation and functional characterization of human erythroblasts at distinct stages: Implications for understanding of normal and disordered erythropoiesis in vivo. Blood. 2013;121(16):3246–53. 10.1182/blood-2013-01-476390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Ludwig LS, Lareau CA, Bao EL, et al. : Transcriptional States and Chromatin Accessibility Underlying Human Erythropoiesis. Cell Rep. 2019;27(11):3228–3240.e7. 10.1016/j.celrep.2019.05.046 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 29. Li J, Hale J, Bhagia P, et al. : Isolation and transcriptome analyses of human erythroid progenitors: BFU-E and CFU-E. Blood. 2014;124(24):3636–45. 10.1182/blood-2014-07-588806 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. An X, Schulz VP, Li J, et al. : Global transcriptome analyses of human and murine terminal erythroid differentiation. Blood. 2014;123(22):3466–77. 10.1182/blood-2014-01-548305 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 31. Schulz VP, Yan H, Lezon-Geyda K, et al. : A Unique Epigenomic Landscape Defines Human Erythropoiesis. Cell Rep. 2019;28(11):2996–3009.e7. 10.1016/j.celrep.2019.08.020 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 32. Tusi BK, Wolock SL, Weinreb C, et al. : Population snapshots predict early haematopoietic and erythroid hierarchies. Nature. 2018;555(7694):54–60. 10.1038/nature25741 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 33. Pellin D, Loperfido M, Baricordi C, et al. : A comprehensive single cell transcriptional landscape of human hematopoietic progenitors. Nat Commun. 2019;10(1):2395. 10.1038/s41467-019-10291-0 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 34. Lodish H, Flygare J, Chou S: From stem cell to erythroblast: Regulation of red cell production at multiple levels by multiple hormones. IUBMB Life. 2010;62(7):492–6. 10.1002/iub.322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Parganas E, Wang D, Stravopodis D, et al. : Jak2 Is Essential for Signaling through a Variety of Cytokine Receptors. Cell. 1998;93(3):385–95. 10.1016/s0092-8674(00)81167-8 [DOI] [PubMed] [Google Scholar]
- 36. Hammond D, Winnick S: Paraneoplastic erythrocytosis and ectopic erythropoietins. Ann N Y Acad Sci. 1974;230:219–27. 10.1111/j.1749-6632.1974.tb14452.x [DOI] [PubMed] [Google Scholar]
- 37. Gross M, Ben-Califa N, McMullin MF, et al. : Polycythaemia-inducing mutations in the erythropoietin receptor (EPOR): Mechanism and function as elucidated by epidermal growth factor receptor-EPOR chimeras. Br J Haematol. 2014;165(4):519–28. 10.1111/bjh.12782 [DOI] [PubMed] [Google Scholar]
- 38. Huang LJ, Shen YM, Bulut GB: Advances in understanding the pathogenesis of primary familial and congenital polycythaemia. Br J Haematol. 2010;148(6):844–52. 10.1111/j.1365-2141.2009.08069.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Koury M, Bondurant M: Erythropoietin retards DNA breakdown and prevents programmed death in erythroid progenitor cells. Science. 1990;248(4953):378–81. 10.1126/science.2326648 [DOI] [PubMed] [Google Scholar]
- 40. Spivak JL, Pham T, Isaacs M, et al. : Erythropoietin is both a mitogen and a survival factor. Blood. 1991;77(6):1228–33. [PubMed] [Google Scholar]
- 41. Koury MJ, Bondurant MC, Atkinson JB: Erythropoietin control of terminal erythroid differentiation: maintenance of cell viability, production of hemoglobin, and development of the erythrocyte membrane. Blood Cells. 1987;13(1–2):217–26. [PubMed] [Google Scholar]
- 42. Grover A, Mancini E, Moore S, et al. : Erythropoietin guides multipotent hematopoietic progenitor cells toward an erythroid fate. J Exp Med. 2014;211(2):181–8. 10.1084/jem.20131189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Bachmann S, Le Hir M, Eckardt KU: Co-localization of erythropoietin mRNA and ecto-5'-nucleotidase immunoreactivity in peritubular cells of rat renal cortex indicates that fibroblasts produce erythropoietin. J Histochem Cytochem. 1993;41(3):335–41. 10.1177/41.3.8429197 [DOI] [PubMed] [Google Scholar]
- 44. Lacombe C, Da Silva JL, Bruneval P, et al. : Peritubular cells are the site of erythropoietin synthesis in the murine hypoxic kidney. J Clin Invest. 1988;81(2):620–3. 10.1172/JCI113363 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Koury ST, Bondurant MC, Koury MJ: Localization of erythropoietin synthesizing cells in murine kidneys by in situ hybridization. Blood. 1988;71(2):524–7. [PubMed] [Google Scholar]
- 46. Zeisberg M, Kalluri R: Physiology of the Renal Interstitium. Clin J Am Soc Nephrol. 2015;10(10):1831–40. 10.2215/CJN.00640114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Fisher JW, Koury S, Ducey T, et al. : Erythropoietin production by interstitial cells of hypoxic monkey kidneys. Br J Haematol. 1996;95(1):27–32. 10.1046/j.1365-2141.1996.d01-1864.x [DOI] [PubMed] [Google Scholar]
- 48. Shanks JH, Hill CM, Lappin TR, et al. : Localization of erythropoietin gene expression in proximal renal tubular cells detected by digoxigenin-labelled oligonucleotide probes. J Pathol. 1996;179(3):283–7. [DOI] [PubMed] [Google Scholar]
- 49. Pan X, Suzuki N, Hirano I, et al. : Isolation and characterization of renal erythropoietin-producing cells from genetically produced anemia mice. PLoS One. 2011;6(10):e25839. 10.1371/journal.pone.0025839 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Kurtz A: Endocrine functions of the renal interstitium. Pflugers Arch. 2017;469(7–8):869–76. 10.1007/s00424-017-2008-9 [DOI] [PubMed] [Google Scholar]
- 51. Lacombe C, Da Silva JL, Bruneval P, et al. : Erythropoietin: Sites of synthesis and regulation of secretion. Am J Kidney Dis. 1991;18(4 Suppl 1):14–9. [PubMed] [Google Scholar]
- 52. Jelkmann W: Regulation of erythropoietin production. J Physiol. 2011;589(Pt 6):1251–8. 10.1113/jphysiol.2010.195057 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 53. Maxwell PH, Ferguson DJ, Nicholls LG, et al. : Sites of erythropoietin production. Kidney Int. 1997;51(2):393–401. 10.1038/ki.1997.52 [DOI] [PubMed] [Google Scholar]
- 54. Haase VH: Regulation of erythropoiesis by hypoxia-inducible factors. Blood Rev. 2013;27(1):41–53. 10.1016/j.blre.2012.12.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Schofield CJ, Ratcliffe PJ: Oxygen sensing by HIF hydroxylases. Nat Rev Mol Cell Biol. 2004;5(5):343–54. 10.1038/nrm1366 [DOI] [PubMed] [Google Scholar]
- 56. Mahon PC, Hirota K, Semenza GL: FIH-1: A novel protein that interacts with HIF-1alpha and VHL to mediate repression of HIF-1 transcriptional activity. Genes Dev. 2001;15(20):2675–86. 10.1101/gad.924501 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 57. Peyssonnaux C, Nizet V, Johnson RS: Role of the hypoxia inducible factors HIF in iron metabolism. Cell Cycle. 2008;7(1):28–32. 10.4161/cc.7.1.5145 [DOI] [PubMed] [Google Scholar]
- 58. Masoud GN, Li W: HIF-1α pathway: Role, regulation and intervention for cancer therapy. Acta Pharm Sin B. 2015;5(5):378–89. 10.1016/j.apsb.2015.05.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Ratcliffe PJ: HIF-1 and HIF-2: Working alone or together in hypoxia? J Clin Invest. 2007;117(4):862–5. 10.1172/JCI31750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Kaplan JM, Sharma N, Dikdan S: Hypoxia-Inducible Factor and Its Role in the Management of Anemia in Chronic Kidney Disease. Int J Mol Sci. 2018;19(2):389. 10.3390/ijms19020389 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 61. Appelhoff RJ, Tian YM, Raval RR, et al. : Differential function of the prolyl hydroxylases PHD1, PHD2, and PHD3 in the regulation of hypoxia-inducible factor. J Biol Chem. 2004;279(37):38458–65. 10.1074/jbc.M406026200 [DOI] [PubMed] [Google Scholar]
- 62. Simonson TS, Yang Y, Huff CD, et al. : Genetic Evidence for High-Altitude Adaptation in Tibet. Science. 2010;329(5987):72–5. 10.1126/science.1189406 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 63. Chappell JC, Payne LB, Rathmell WK: Hypoxia, angiogenesis, and metabolism in the hereditary kidney cancers. J Clin Invest. 2019;129(2):442–51. 10.1172/JCI120855 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 64. Lappin TR, Lee FS: Update on mutations in the HIF: EPO pathway and their role in erythrocytosis. Blood Rev. 2019;37:100590. 10.1016/j.blre.2019.100590 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 65. Zmajkovic J, Lundberg P, Nienhold R, et al. : A Gain-of-Function Mutation in EPO in Familial Erythrocytosis. N Engl J Med. 2018;378(10):924–30. 10.1056/NEJMoa1709064 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 66. Bigham AW: Genetics of human origin and evolution: High-altitude adaptations. Curr Opin Genet Dev. 2016;41:8–13. 10.1016/j.gde.2016.06.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Julian CG, Moore LG: Human Genetic Adaptation to High Altitude: Evidence from the Andes. Genes (Basel). 2019;10(2):150. 10.3390/genes10020150 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 68. van Tissot Patot MC, Gassmann M: Hypoxia: Adapting to High Altitude by Mutating EPAS-1, the Gene Encoding HIF-2α. High Alt Med Biol. 2011;12(2):157–67. 10.1089/ham.2010.1099 [DOI] [PubMed] [Google Scholar]
- 69. Yi X, Liang Y, Huerta-Sanchez E, et al. : Sequencing of 50 human exomes reveals adaptation to high altitude. Science. 2010;329(5987):75–8. 10.1126/science.1190371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Zhang N, Fu Z, Linke S, et al. : The asparaginyl hydroxylase factor inhibiting HIF-1alpha is an essential regulator of metabolism. Cell Metab. 2010;11(5):364–78. 10.1016/j.cmet.2010.03.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Constantinescu SN: Mechanism of erythropoietin receptor activation.In: Elliott S, Foote M, Molineux G editors. Erythropoietins, Erythropoietic Factors, and Erythropoiesis.Basel: Birkhäuser Basel;2009;175–98. Reference Source [Google Scholar]
- 72. Constantinescu SN, Keren T, Socolovsky M, et al. : Ligand-independent oligomerization of cell-surface erythropoietin receptor is mediated by the transmembrane domain. Proc Natl Acad Sci U S A. 2001;98(8):4379–84. 10.1073/pnas.081069198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Moraga I, Wernig G, Wilmes S, et al. : Tuning cytokine receptor signaling by re-orienting dimer geometry with surrogate ligands. Cell. 2015;160(6):1196–208. 10.1016/j.cell.2015.02.011 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 74. Gillinder KR, Tuckey H, Bell CC, et al. : Direct targets of pSTAT5 signalling in erythropoiesis. PLoS One. 2017;12(7):e0180922. 10.1371/journal.pone.0180922 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Rhodes MM, Kopsombut P, Bondurant MC, et al. : Bcl-xL prevents apoptosis of late-stage erythroblasts but does not mediate the antiapoptotic effect of erythropoietin. Blood. 2005;106(5):1857–63. 10.1182/blood-2004-11-4344 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Socolovsky M, Fallon AEJ, Wang S, et al. : Fetal anemia and apoptosis of red cell progenitors in Stat5a-/-5b-/- mice: a direct role for Stat5 in Bcl-X(L) induction. Cell. 1999;98(2):181–91. 10.1016/s0092-8674(00)81013-2 [DOI] [PubMed] [Google Scholar]
- 77. Cui Y, Riedlinger G, Miyoshi K, et al. : Inactivation of Stat5 in mouse mammary epithelium during pregnancy reveals distinct functions in cell proliferation, survival, and differentiation. Mol Cell Biol. 2004;24(18):8037–47. 10.1128/MCB.24.18.8037-8047.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Wood AD, Chen E, Donaldson IJ, et al. : ID1 promotes expansion and survival of primary erythroid cells and is a target of JAK2V617F-STAT5 signaling. Blood. 2009;114(9):1820–30. 10.1182/blood-2009-02-206573 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Dev A, Asch R, Jachimowicz E, et al. : Governing roles for Trib3 pseudokinase during stress erythropoiesis. Exp Hematol. 2017;49:48–55.e5. 10.1016/j.exphem.2016.12.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Dev A, Byrne SM, Verma R, et al. : Erythropoietin-directed erythropoiesis depends on serpin inhibition of erythroblast lysosomal cathepsins. J Exp Med. 2013;210(2):225–32. 10.1084/jem.20121762 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Zhu BM, McLaughlin SK, Na R, et al. : Hematopoietic-specific Stat5-null mice display microcytic hypochromic anemia associated with reduced transferrin receptor gene expression. Blood. 2008;112(5):2071–80. 10.1182/blood-2007-12-127480 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Kerenyi MA, Grebien F, Gehart H, et al. : Stat5 regulates cellular iron uptake of erythroid cells via IRP-2 and TfR-1. Blood. 2008;112(9):3878–88. 10.1182/blood-2008-02-138339 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Kautz L, Jung G, Valore EV, et al. : Identification of erythroferrone as an erythroid regulator of iron metabolism. Nat Genet. 2014;46(7):678–84. 10.1038/ng.2996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Coffey R, Ganz T: Erythroferrone: an erythroid regulator of hepcidin and iron metabolism. Hemasphere. 2018;2(2):e35. 10.1097/HS9.0000000000000035 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 85. Rainville N, Jachimowicz E, Wojchowski DM: Targeting EPO and EPO receptor pathways in anemia and dysregulated erythropoiesis. Expert Opin Ther Targets. 2016;20(3):287–301. 10.1517/14728222.2016.1090975 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Wojchowski DM, Sathyanarayana P, Dev A: Erythropoietin receptor response circuits. Curr Opin Hematol. 2010:17(3):169–76. 10.1097/MOH.0b013e328338008b [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Lodish HF, Ghaffari S, Socolovsky M, et al. : Intracellular signaling by the erythropoietin receptor.In: Elliott S, Foote M, Molineux G, editors. Erythropoietins, Erythropoietic Factors, and Erythropoiesis.Basel: Birkhäuser Basel;2009;155–74. 10.1007/978-3-7643-8698-6_10 [DOI] [Google Scholar]
- 88. Watowich SS: The erythropoietin receptor: Molecular structure and hematopoietic signaling pathways. J Investig Med. 2011;59(7):1067–72. 10.2310/JIM.0b013e31820fb28c [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Kumkhaek C, Aerbajinai W, Liu W, et al. : MASL1 induces erythroid differentiation in human erythropoietin-dependent CD34+ cells through the Raf/MEK/ERK pathway. Blood. 2013;121(16):3216–27. 10.1182/blood-2011-10-385252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Marinkovic D, Zhang X, Yalcin S, et al. : Foxo3 is required for the regulation of oxidative stress in erythropoiesis. J Clin Invest. 2007;117(8):2133–44. 10.1172/JCI31807 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 91. Kashii Y, Uchida M, Kirito K, et al. : A member of Forkhead family transcription factor, FKHRL1, is one of the downstream molecules of phosphatidylinositol 3-kinase-Akt activation pathway in erythropoietin signal transduction. Blood. 2000;96(3):941–9. 10.1182/blood.V96.3.941 [DOI] [PubMed] [Google Scholar]
- 92. Liang R, Campreciós G, Kou Y, et al. : A Systems Approach Identifies Essential FOXO3 Functions at Key Steps of Terminal Erythropoiesis. PLoS Genet. 2015;11(10):e1005526. 10.1371/journal.pgen.1005526 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93. Karur VG, Lowell CA, Besmer P, et al. : Lyn kinase promotes erythroblast expansion and late-stage development. Blood. 2006;108(5):1524–32. 10.1182/blood-2005-09-008243 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Schnöder TM, Arreba-Tutusaus P, Griehl I, et al. : Epo-induced erythroid maturation is dependent on Plcγ 1 signaling. Cell Death Differ. 2015;22(6):974–85. 10.1038/cdd.2014.186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Liongue C, Sertori R, Ward AC: Evolution of Cytokine Receptor Signaling. J Immunol. 2016;197(1):11–8. 10.4049/jimmunol.1600372 [DOI] [PubMed] [Google Scholar]
- 96. Li K, Miller C, Hegde S, et al. : Roles for an Epo Receptor Tyr-343 Stat5 Pathway in Proliferative Co-signaling with Kit. J Biol Chem. 2003;278(42):40702–9. 10.1074/jbc.M307182200 [DOI] [PubMed] [Google Scholar]
- 97. Wu H, Klingmüller U, Besmer P, et al. : Interaction of the erythropoietin and stem-cell-factor receptors. Nature. 1995;377(6546):242–6. 10.1038/377242a0 [DOI] [PubMed] [Google Scholar]
- 98. Wessely O, Bauer A, Quang CT, et al. : A Novel Way to Induce Erythroid Progenitor Self Renewal: Cooperation of c-Kit with the Erythropoietin Receptor. Biol Chem. 1999;380(2):187–202. 10.1515/BC.1999.027 [DOI] [PubMed] [Google Scholar]
- 99. Wu H, Klingmüller U, Acurio A, et al. : Functional interaction of erythropoietin and stem cell factor receptors is essential for erythroid colony formation. Proc Natl Acad Sci U S A. 1997;94(5):1806–10. 10.1073/pnas.94.5.1806 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Roetto A, Mezzanotte M, Pellegrino RM: The Functional Versatility of Transferrin Receptor 2 and Its Therapeutic Value. Pharmaceuticals (Basel). 2018;11(4):115. 10.3390/ph11040115 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 101. Forejtnikovà H, Vieillevoye M, Zermati Y, et al. : Transferrin receptor 2 is a component of the erythropoietin receptor complex and is required for efficient erythropoiesis. Blood. 2010;116(24):5357–67. 10.1182/blood-2010-04-281360 [DOI] [PubMed] [Google Scholar]
- 102. Nai A, Lidonnici MR, Rausa M, et al. : The second transferrin receptor regulates red blood cell production in mice. Blood. 2015;125(7):1170–9. 10.1182/blood-2014-08-596254 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Wortham AM, Goldman DC, Chen J, et al. : Extrahepatic deficiency of transferrin receptor 2 is associated with increased erythropoiesis independent of iron overload. J Biol Chem. 2020;295(12):3906–17. 10.1074/jbc.RA119.010535 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 104. Johnson MB, Enns CA: Diferric transferrin regulates transferrin receptor 2 protein stability. Blood. 2004;104(13):4287–93. 10.1182/blood-2004-06-2477 [DOI] [PubMed] [Google Scholar]
- 105. Porpiglia E, Hidalgo D, Koulnis M, et al. : Stat5 signaling specifies basal versus stress erythropoietic responses through distinct binary and graded dynamic modalities. PLoS Biol. 2012;10(8):e1001383. 10.1371/journal.pbio.1001383 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Zang H, Sato K, Nakajima H, et al. : The distal region and receptor tyrosines of the Epo receptor are non-essential for in vivo erythropoiesis. EMBO J. 2001;20(12):3156–66. 10.1093/emboj/20.12.3156 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Juvonen E, Ikkala E, Fyhrquist F, et al. : Autosomal dominant erythrocytosis caused by increased sensitivity to erythropoietin. Blood. 1991;78(11):3066–9. [PubMed] [Google Scholar]
- 108. Jiao H, Berrada K, Yang W, et al. : Direct association with and dephosphorylation of Jak2 kinase by the SH2-domain-containing protein tyrosine phosphatase SHP-1. Mol Cell Biol. 1996;16(12):6985–92. 10.1128/mcb.16.12.6985 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Baker SJ, Rane SG, Reddy EP: Hematopoietic cytokine receptor signaling. Oncogene. 2007;26(47):6724–37. 10.1038/sj.onc.1210757 [DOI] [PubMed] [Google Scholar]
- 110. Wormald S, Hilton DJ: Inhibitors of cytokine signal transduction. J Biol Chem. 2004;279(2):821–4. 10.1074/jbc.R300030200 [DOI] [PubMed] [Google Scholar]
- 111. Yoshimura A, Ito M, Chikuma S, et al. : Negative Regulation of Cytokine Signaling in Immunity. Cold Spring Harb Perspect Biol. 2018;10(7):a028571. 10.1101/cshperspect.a028571 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 112. Suessmuth Y, Elliott J, Percy MJ, et al. : A new polycythaemia vera-associated SOCS3 SH2 mutant (SOCS3F136L) cannot regulate erythropoietin responses. Br J Haematol. 2009;147(4):450–8. 10.1111/j.1365-2141.2009.07860.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113. Hookham MB, Elliott J, Suessmuth Y, et al. : The myeloproliferative disorder-associated JAK2 V617F mutant escapes negative regulation by suppressor of cytokine signaling 3. Blood. 2007;109(11):4924–9. 10.1182/blood-2006-08-039735 [DOI] [PubMed] [Google Scholar]
- 114. Tong W, Zhang J, Lodish HF: Lnk inhibits erythropoiesis and Epo-dependent JAK2 activation and downstream signaling pathways. Blood. 2005;105(12):4604–12. 10.1182/blood-2004-10-4093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. McMullin MF, Cario H: LNK mutations and myeloproliferative disorders. Am J Hematol. 2016;91(2):248–51. 10.1002/ajh.24259 [DOI] [PubMed] [Google Scholar]
- 116. van der Harst P, Zhang W, Mateo Leach I, et al. : Seventy-five genetic loci influencing the human red blood cell. Nature. 2012;492(7429):369–75. 10.1038/nature11677 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. McMullin MF, Wu C, Percy MJ, et al. : A nonsynonymous LNK polymorphism associated with idiopathic erythrocytosis. Am J Hematol. 2011;86(11):962–4. 10.1002/ajh.22154 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. Giani FC, Fiorini C, Wakabayashi A, et al. : Targeted Application of Human Genetic Variation Can Improve Red Blood Cell Production from Stem Cells. Cell Stem Cell. 2016;18(1):73–8. 10.1016/j.stem.2015.09.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Neumann D, Wikström L, Watowich SS, et al. : Intermediates in degradation of the erythropoietin receptor accumulate and are degraded in lysosomes. J Biol Chem. 1993;268(18):13639–49. [PubMed] [Google Scholar]
- 120. Bulut GB, Sulahian R, Ma Y, et al. : Ubiquitination Regulates the Internalization, Endolysosomal Sorting, and Signaling of the Erythropoietin Receptor. J Biol Chem. 2011;286(8):6449–57. 10.1074/jbc.M110.186890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Jimenez C, Hernandez C, Pimentel B, et al. : The p85 regulatory subunit controls sequential activation of phosphoinositide 3-kinase by Tyr kinases and Ras. J Biol Chem. 2002;277(44):41556–62. 10.1074/jbc.M205893200 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 122. Sulahian R, Cleaver O, Huang LJS: Ligand-induced EpoR internalization is mediated by JAK2 and p85 and is impaired by mutations responsible for primary familial and congenital polycythemia. Blood. 2009;113(21):5287–97. 10.1182/blood-2008-09-179572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Bulut GB, Sulahian R, Yao H, et al. : Cbl ubiquitination of p85 is essential for Epo-induced EpoR endocytosis. Blood. 2013;122(24):3964–72. 10.1182/blood-2013-05-506212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Heir P, Srikumar T, Bikopoulos G, et al. : Oxygen-dependent Regulation of Erythropoietin Receptor Turnover and Signaling. J Biol Chem. 2016;291(14):7357–72. 10.1074/jbc.M115.694562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Khalil S, Delehanty L, Grado S, et al. : Iron modulation of erythropoiesis is associated with Scribble-mediated control of the erythropoietin receptor. J Exp Med. 2018;215(2):661–79. 10.1084/jem.20170396 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 126. Broxmeyer HE, Hoggatt J, O'Leary HA, et al. : Dipeptidylpeptidase 4 negatively regulates colony-stimulating factor activity and stress hematopoiesis. Nat Med. 2012;18(12):1786–96. 10.1038/nm.2991 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Vainchenker W, Leroy E, Gilles L, et al. : JAK inhibitors for the treatment of myeloproliferative neoplasms and other disorders [version 1; peer review: 2 approved]. F1000Res. 2018;7:82. 10.12688/f1000research.13167.1 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 128. Huang LJ, Constantinescu SN, Lodish HF: The N-Terminal Domain of Janus Kinase 2 Is Required for Golgi Processing and Cell Surface Expression of Erythropoietin Receptor. Mol Cell. 2001;8(6):1327–38. 10.1016/s1097-2765(01)00401-4 [DOI] [PubMed] [Google Scholar]
- 129. Yao H, Ma Y, Hong Z, et al. : Activating JAK2 mutants reveal cytokine receptor coupling differences that impact outcomes in myeloproliferative neoplasm. Leukemia. 2017;31(10):2122–31. 10.1038/leu.2017.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Wilmes S, Hafer M, Vuorio J, et al. : Mechanism of homodimeric cytokine receptor activation and dysregulation by oncogenic mutations. Science. 2020;367(6478):643–52. 10.1126/science.aaw3242 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 131. Constantinescu SN, Huang LJ, Nam H, et al. : The Erythropoietin Receptor Cytosolic Juxtamembrane Domain Contains an Essential, Precisely Oriented, Hydrophobic Motif. Mol Cell. 2001;7(2):377–85. 10.1016/s1097-2765(01)00185-x [DOI] [PubMed] [Google Scholar]
- 132. Ferrao RD, Wallweber HJ, Lupardus PJ: Receptor-mediated dimerization of JAK2 FERM domains is required for JAK2 activation. eLife. 2018;7:e38089. 10.7554/eLife.38089 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 133. Kim AR, Ulirsch JC, Wilmes S, et al. : Functional Selectivity in Cytokine Signaling Revealed Through a Pathogenic EPO Mutation. Cell. 2017;168(6):1053–1064.e15. 10.1016/j.cell.2017.02.026 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 134. Mohan K, Ueda G, Kim AR, et al. : Topological control of cytokine receptor signaling induces differential effects in hematopoiesis. Science. 2019;364(6442): eaav7532. 10.1126/science.aav7532 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 135. Wootten D, Christopoulos A, Marti-Solano M, et al. : Mechanisms of signalling and biased agonism in G protein-coupled receptors. Nat Rev Mol Cell Biol. 2018;19(10):638–53. 10.1038/s41580-018-0049-3 [DOI] [PubMed] [Google Scholar]
- 136. Thomas C, Moraga I, Levin D, et al. : Structural linkage between ligand discrimination and receptor activation by type I interferons. Cell. 2011;146(4):621–32. 10.1016/j.cell.2011.06.048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137. McKeithan TW: Kinetic proofreading in T-cell receptor signal transduction. Proc Natl Acad Sci U S A. 1995;92(11):5042–6. 10.1073/pnas.92.11.5042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Ng T, Marx G, Littlewood T, et al. : Recombinant erythropoietin in clinical practice. Postgrad Med J. 2003;79(933):367–76. 10.1136/pmj.79.933.367 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Mikhail A, Brown C, Williams JA, et al. : Renal association clinical practice guideline on Anaemia of Chronic Kidney Disease. BMC Nephrol. 2017;18(1):345. 10.1186/s12882-017-0688-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. van Swelm RPL, Wetzels JFM, Swinkels DW: The multifaceted role of iron in renal health and disease. Nat Rev Nephrol. 2020;16(2):77–98. 10.1038/s41581-019-0197-5 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 141. Ganz T, Nemeth E: Iron Balance and the Role of Hepcidin in Chronic Kidney Disease. Semin Nephrol. 2016;36(2):87–93. 10.1016/j.semnephrol.2016.02.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142. Won HH, Park I, Lee E, et al. : Comparative analysis of the JAK/STAT signaling through erythropoietin receptor and thrombopoietin receptor using a systems approach. BMC Bioinformatics. 2009;10(Suppl 1):S53. 10.1186/1471-2105-10-S1-S53 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143. Shi J, Yuan B, Hu W, et al. : JAK2 V617F stimulates proliferation of erythropoietin-dependent erythroid progenitors and delays their differentiation by activating Stat1 and other nonerythroid signaling pathways. Exp Hematol. 2016;44(11):1044–1058.e5. 10.1016/j.exphem.2016.07.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144. Funakoshi-Tago M, Sumi K, Kasahara T, et al. : Critical Roles of Myc-ODC Axis in the Cellular Transformation Induced by Myeloproliferative Neoplasm-Associated JAK2 V617F Mutant. PLoS One. 2013;8(1):e52844. 10.1371/journal.pone.0052844 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145. Cai X, Jia H, Liu Z, et al. : Polyhydroxylated fullerene derivative C 60(OH) 24 prevents mitochondrial dysfunction and oxidative damage in an MPP + -induced cellular model of Parkinson's disease. J Neurosci Res. 2008;86(16):3622–34. 10.1002/jnr.21805 [DOI] [PubMed] [Google Scholar]
- 146. Funakoshi-Tago M, Nagata T, Tago K, et al. : Fullerene derivative prevents cellular transformation induced by JAK2 V617F mutant through inhibiting c-Jun N-terminal kinase pathway. Cell Signal. 2012;24(11):2024–34. 10.1016/j.cellsig.2012.06.014 [DOI] [PubMed] [Google Scholar]
- 147. Eschbach JW, Aquiling T, Haley NR, et al. : The long-term effects of recombinant human erythropoietin on the cardiovascular system. Clin Nephrol. 1992;38 Suppl 1:S98–103. [PubMed] [Google Scholar]
- 148. Guglin ME, Koul D: Cardiovascular effects of erythropoietin: Anemia and beyond. Cardiol Rev. 2006;14(4):200–4. 10.1097/01.crd.0000195223.85556.8e [DOI] [PubMed] [Google Scholar]
- 149. Gupta N, Wish JB: Erythropoietin and its cardiovascular effects. In: Rangaswami J, Lerma EV, Ronco C, editors. Cardio-Nephrology.Cham: Springer;2017;119–28. 10.1007/978-3-319-56042-7_11 [DOI] [Google Scholar]
- 150. Casati S, Passerini P, Campise MR, et al. : Benefits and risks of protracted treatment with human recombinant erythropoietin in patients having haemodialysis. Br Med J (Clin Res Ed). 1987;295(6605):1017–20. 10.1136/bmj.295.6605.1017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Debeljak N, Solár P, Sytkowski AJ: Erythropoietin and cancer: The unintended consequences of anemia correction. Front Immunol. 2014;5:563. 10.3389/fimmu.2014.00563 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152. Jelkmann W: Physiology and Pharmacology of Erythropoietin. Transfus Med Hemother. 2013;40(5):302–9. 10.1159/000356193 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153. Pradeep S, Huang J, Mora EM, et al. : Erythropoietin Stimulates Tumor Growth via EphB4. Cancer Cell. 2015;28(5):610–22. 10.1016/j.ccell.2015.09.008 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 154. Remuzzi G, Ingelfinger JR: Correction of anemia--payoffs and problems. N Engl J Med. 2006;355(20):2144–6. 10.1056/NEJMe068233 [DOI] [PubMed] [Google Scholar]
- 155. Singh AK, Szczech L, Tang KL, et al. : Correction of anemia with epoetin alfa in chronic kidney disease. N Engl J Med. 2006;355(20):2085–98. 10.1056/NEJMoa065485 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 156. Pfeffer MA, Burdmann EA, Chen CY, et al. : A Trial of Darbepoetin Alfa in Type 2 Diabetes and Chronic Kidney Disease. N Engl J Med. 2009;361(21):2019–32. 10.1056/NEJMoa0907845 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 157. Miller CP, Lowe KA, Valliant-Saunders K, et al. : Evaluating erythropoietin-associated tumor progression using archival tissues from a phase III clinical trial. Stem Cells. 2009;27(9):2353–61. 10.1002/stem.156 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Elliott S, Sinclair AM: The effect of erythropoietin on normal and neoplastic cells. Biologics. 2012;6:163–89. 10.2147/BTT.S32281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159. Macdougall IC, White C, Anker SD, et al. : Intravenous Iron in Patients Undergoing Maintenance Hemodialysis. N Engl J Med. 2019;380(5):447–58. 10.1056/NEJMoa1810742 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 160. Bohlius J, Bohlke K, Castelli R, et al. : Management of cancer-associated anemia with erythropoiesis-stimulating agents: ASCO/ASH clinical practice guideline update. Blood Adv. 2019;3(8):1197–210. 10.1182/bloodadvances.2018030387 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 161. Platzbecker U, Symeonidis A, Oliva EN, et al. : A phase 3 randomized placebo-controlled trial of darbepoetin alfa in patients with anemia and lower-risk myelodysplastic syndromes. Leukemia. 2017;31(9):1944–50. 10.1038/leu.2017.192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162. Park S, Greenberg P, Yucel A, et al. : Clinical effectiveness and safety of erythropoietin-stimulating agents for the treatment of low- and intermediate-1-risk myelodysplastic syndrome: A systematic literature review. Br J Haematol. 2019;184(2):134–60. 10.1111/bjh.15707 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 163. Steensma DP: Hematopoietic Growth Factors in Myelodysplastic Syndromes. Semin Oncol. 2011;38(5):635–47. 10.1053/j.seminoncol.2011.04.014 [DOI] [PubMed] [Google Scholar]
- 164. Whoriskey P: The rise and fall of a billion-dollar drug. Washington Post2012. Reference Source [Google Scholar]
- 165. Kaplan J: Roxadustat and Anemia of Chronic Kidney Disease. N Engl J Med. 2019;381(11):1070–2. 10.1056/NEJMe1908978 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 166. Sanghani NS, Haase VH: Hypoxia-Inducible Factor Activators in Renal Anemia: Current Clinical Experience. Adv Chronic Kidney Dis. 2019;26(4):253–66. 10.1053/j.ackd.2019.04.004 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 167. Chen N, Hao C, Liu BC, et al. : Roxadustat Treatment for Anemia in Patients Undergoing Long-Term Dialysis. N Engl J Med. 2019;381(11):1011–22. 10.1056/NEJMoa1901713 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 168. Brigandi RA, Johnson B, Oei C, et al. : A Novel Hypoxia-Inducible Factor−Prolyl Hydroxylase Inhibitor (GSK1278863) for Anemia in CKD: A 28-Day, Phase 2A Randomized Trial. Am J Kidney Dis. 2016;67(6):861–71. 10.1053/j.ajkd.2015.11.021 [DOI] [PubMed] [Google Scholar]
- 169. Besarab A, Provenzano R, Hertel J, et al. : Randomized placebo-controlled dose-ranging and pharmacodynamics study of roxadustat (FG-4592) to treat anemia in nondialysis-dependent chronic kidney disease (NDD-CKD) patients. Nephrol Dial Transplant. 2015;30(10):1665–73. 10.1093/ndt/gfv302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170. Provenzano R, Besarab A, Sun CH, et al. : Oral Hypoxia–Inducible Factor Prolyl Hydroxylase Inhibitor Roxadustat (FG-4592) for the Treatment of Anemia in Patients with CKD. Clin J Am Soc Nephrol. 2016;11(6):982–91. 10.2215/CJN.06890615 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171. Dhillon S: Roxadustat: First Global Approval. Drugs. 2019;79(5):563–72. 10.1007/s40265-019-01077-1 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 172. Ma S, Chen J, Chen C, et al. : Erythropoietin Rescues Memory Impairment in a Rat Model of Chronic Cerebral Hypoperfusion via the EPO-R/JAK2/STAT5/PI3K/Akt/GSK-3β Pathway. Mol Neurobiol. 2018;55(4):3290–9. 10.1007/s12035-017-0568-5 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 173. Kimáková P, Solár P, Solárová Z, et al. : Erythropoietin and Its Angiogenic Activity. Int J Mol Sci. 2017;18(7):1519. 10.3390/ijms18071519 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174. Broxmeyer HE: Erythropoietin: Multiple targets, actions, and modifying influences for biological and clinical consideration. J Exp Med. 2013;210(2):205–8. 10.1084/jem.20122760 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175. Brines M, Grasso G, Fiordaliso F, et al. : Erythropoietin mediates tissue protection through an erythropoietin and common beta-subunit heteroreceptor. Proc Natl Acad Sci U S A. 2004;101(41):14907–12. 10.1073/pnas.0406491101 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 176. Razak A, Hussain A: Erythropoietin in perinatal hypoxic-ischemic encephalopathy: A systematic review and meta-analysis. J Perinat Med. 2019;47(4):478–89. 10.1515/jpm-2018-0360 [DOI] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 177. Tsai TH, Lu CH, Wallace CG, et al. : Erythropoietin improves long-term neurological outcome in acute ischemic stroke patients: A randomized, prospective, placebo-controlled clinical trial. Crit Care. 2015;19(1):49. 10.1186/s13054-015-0761-8 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation
- 178. Nekoui A, Blaise G: Erythropoietin and Nonhematopoietic Effects. Am J Med Sci. 2017;353(1):76–81. 10.1016/j.amjms.2016.10.009 [DOI] [PubMed] [Google Scholar]
- 179. Suresh S, Rajvanshi PK, Noguchi CT: The Many Facets of Erythropoietin Physiologic and Metabolic Response. Front Physiol. 2020;10:1534. 10.3389/fphys.2019.01534 [DOI] [PMC free article] [PubMed] [Google Scholar]; Faculty Opinions Recommendation