

Abstract
Neurological diseases are a major threat to global public health and prosperity. The number of patients with neurological diseases is increasing due to the population aging and increasing life expectancy. Autophagy is one of the crucial mechanisms to maintain nerve cellular homeostasis. Numerous studies have demonstrated that autophagy plays a dual role in neurological diseases. Long noncoding RNAs (lncRNAs) are a vital class of noncoding RNAs with a length of more than 200 nucleotides and cannot encode proteins themselves but are expressed in most neurological diseases. An early phase, emerging knowledge has revealed that long noncoding RNAs (lncRNAs) are crucial in autophagy regulation. Furthermore, autophagy-associated lncRNAs can promote the development of neurological diseases or slow their progression. In this review, we introduce a general overview of lncRNA functional mechanisms and summarizes the recent progress of lncRNAs on autophagy regulation in neurological diseases to reveal possible novel therapeutic targets or useful biomarkers.
1. Introduction
Neurological diseases are important causes of human disability and death worldwide. According to the pathology, they can be divided into several groups, including cerebrovasculardiseases, neurodegenerative diseases, demyelinating diseases, infectious diseases, brain tumors, epilepsy, and headache. Most neurological diseases occur after the developmental maturation of CNS [1]. Autophagy is an important catabolic process during which the unwanted cytoplasmic components such as damaged organelles and protein aggregates are sequestered and engulfed by double-membrane vesicles called autophagosomes, and then autophagosomes fuse with lysosomes to form autolysosomes where the cargos are degraded and recycled [2–4]. Autophagy can be divided into basal autophagy and induced autophagy [5]. Several studies have demonstrated that basal autophagy plays a housekeeping role in eukaryotes through degrading useless and dysfunctional proteins and organelles to maintain cellular homeostasis and promote cell growth and development [3, 6]. Compared with basal autophagy, the degree of induced autophagy is significantly increased, which is a defensive response of the body to external stimuli and may cause autophagic cell death [5]. Neuronal autophagy plays an essential role in synaptic plasticity, oligodendrocyte development, anti-inflammatory function in glial cells, and myelination process [7, 8]. As postmitotic cells, nerve cells are unable to dispose toxic or misfolded proteins through cell division. Therefore, proper autophagy is crucial for nerve cells to remove harmful cellular components; however, insufficient activation of autophagy or pathological stress induced-autophagy will lead to the accumulation of those harmful constituents and eventually causes neuronal dysfunction, which is associated with neurological diseases such as Alzheimer's disease (AD), Parkinson's disease (PD), ischemic stroke (IS), and glioma [9].
Recently, noncoding RNA, such as long noncoding RNA (lncRNA), have been verified to regulate cell autophagy by several mechanisms and further contribute to many of the characteristics of disease phenotypes [10]: (1) lncRNA can sponge some miRNAs which directly target autophagy-related proteins to regulate autophagy. For example, lncRNA APF regulates miR-188-3p, thereby affecting the expression of ATG7 which is an autophagy factor [11]. FLJ11812 can regulate the level of the miR-4459 target ATG13 (autophagy-related 13) and then promote autophagy [12]. Similarly, lncRNA TGFB2-OT1 has been found to regulate the expression of autophagy-related proteins CERS1, NAT8L, and LARP1 by binding to miR-3960, miR-4488, and miR-4459 [13]. (2) lncRNA can also target autophagy-related signaling pathways, such as lncRNA H19 and lncRNA HOTAIR. In this review, we will discuss not only the current knowledge about the relationship between autophagy and neurological diseases but also the possible biological functions of lncRNA in regulating autophagy and summarize some specific studies that have provided novel insights into the underlying mechanism of the lncRNA-autophagy axis for neurological disease pathogenesis and therapeutic intervention.
2. lncRNA Characteristics, Classification, and Functional Mechanisms
Long noncoding RNAs (lncRNAs) are a vital class of noncoding RNAs with a length of more than 200 nucleotides. Surprisingly, although lncRNAs cannot encode proteins themselves, they are known to be 5′ capped, 3′ polyadenylated, and spliced similar to mRNAs. According to their location in the genome relative to nearby protein-coding genes, lncRNAs are generally divided into major groups: sense lncRNAs, antisense lncRNAs, bidirectional lncRNAs, enhancer lncRNAs, intronic lncRNAs, and intergenic lncRNAs [14–16]. They are widespread in most eukaryotic transcriptome and constitute a significant fraction of mammalian genomes, which are involved in cell growth, proliferation, differentiation, apoptosis, metabolism, and other biological processes [17, 18]. With the rapid development of high-throughput sequencing and gene chip technology, a large number of studies have indicated that lncRNA participates in the pathophysiology of various diseases, such as cancer, aging, cardiovascular and cerebrovascular diseases, and neurodegenerative disorders; however, its underlying molecular mechanism is still not well clarified [19–21]. The latest study showed that lncRNA functions generally by five modes of action: signals, decoys, sponges, guides, and scaffold [22, 23] (Figure 1). (1) In response to cell signaling or other stimuli from extracellular environments, lncRNAs that serve as signals will be transcribed and directly regulate the transcription of the downstream gene (Figure 1(a)). This process is directly regulated by lncRNA and does not involve protein translation, so it can respond to external stimuli quickly [24]. (2) lncRNAs can act as a decoy to bind to transcription factors or transcriptional regulating factors and then block its molecular action or other signaling components that regulate the transcription of downstream genes [23] (Figure 1(b)). (3) Interestingly, lncRNAs often function as competitive endogenous RNA (ceRNAs), namely, miRNA sponges or antagonists [25]. More specifically, lncRNAs can bind to miRNA with base pair sequence complementarity, which will repress the binding of miRNA to the 3′ untranslated regions (UTRs) of its target mRNA, protecting mRNA from degrading and consequently regulating protein translation [26, 27] (Figure 1(c)). (4) Besides, lncRNAs are also involved in posttranscriptional modification and alternative splicing of mRNA [28] (Figure 1(d)). (5) lncRNAs which serve as guide RNAs are classified into two categories cis-acting and trans-acting. These lncRNAs can combine with proteins such as transcription factors and transcriptional regulating factors and direct these RNA-protein complexes to the specific DNA sites, contributing to regulating transcription precisely [23] (Figure 1(e)).
Figure 1.
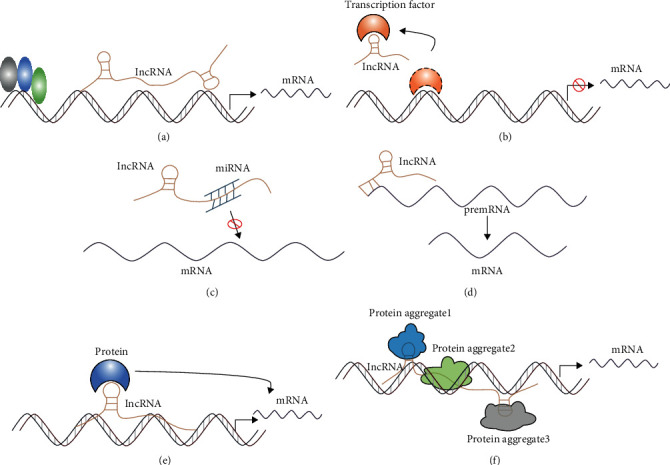
The modes of action of lncRNA. (a) lncRNAs function as signals to directly regulate the transcription of the downstream gene. (b) lncRNAs function as decoys to regulate the binding of proteins to DNA or other proteins. (c) lncRNAs interact with miRNA as miRNA sponges. (d) lncRNAs regulate alternative splicing. (e) lncRNAs serve as guide RNAs to impart specificity at genomic positions through either RNA-DNA or RNA-protein-DNA interactions. (f) lncRNAs serve as scaffolds to allow the formation of larger RNA-protein complexes.
(6) lncRNAs that act as a scaffold are like a central platform, and multiple relevant transcription factors can be bound to these lncRNAs to regulate the activity of transcription factors. Besides, multiple signaling pathways are activated or inhibited, and their downstream effector molecules can be bound to the platform to achieve information intersection and integration between different signaling pathways [29–31] (Figure 1(f)). It is worth noting that the above-mentioned modes of action of lncRNA do not exist independently, but are interrelated and interact with each other. Numerous studies have revealed that lncRNAs are highly expressed in a mammalian brain tissue and play a significant role in regulating protein-coding gene expression through the epigenetic, transcriptional, and posttranscriptional levels and are involved in multiple signaling pathways of various neurological diseases, which has a very broad clinical application prospect [32–34].
3. lncRNA as Regulators of Autophagy in Neurological Diseases
Autophagy is one of the crucial mechanisms to maintain nerve cellular homeostasis; however, numerous researches have shown that autophagy is a double-edged sword that can either protect cells against apoptosis or promote autophagic cell death. The activation of autophagy has been documented in various diseases such as cancer, AD, PD, and vascular diseases [35, 36]. Similar to other signaling pathways, autophagy is regulated by several factors, including transcription factors and noncoding RNAs such as lncRNAs and miRNAs. There is growing evidence that noncoding RNAs regulate diverse pathophysiological processes in vivo and in vitro, from cell proliferation to aging, are both closely associated with autophagy. Besides this, the deregulation of lncRNAs will contribute to numerous neurological diseases [37]. In this section, we will introduce some recent findings that shed light on the role of lncRNA-mediated autophagy in some common neurological diseases (Table 1).
Table 1.
lncRNAs associated with autophagy regulation in neurological diseases.
| Neurological diseases | lncRNA | Main target gene | Expression | Mechanism | Function | Models | Ref. |
|---|---|---|---|---|---|---|---|
| Alzheimer's disease | 17A | GABABR2 | Up | 17A downregulation promotes autophagy by elevating GABABR2 expression | Inducing Aβ secretion and increment of the Aβ42/40 ratio | SH-SY5Y cells treated with amyloid β peptide1-42 | [41, 42] |
| NEAT1 | NEDD4L and PINK1 | Up | Inhibiting PINK1-independent mitophagy by promoting NEDD4L-mediated PINK1 ubiquitination and degradation | Increasing Aβ accumulation | APP/PS1 mice | [46] | |
| Parkinson's disease | NEAT1 | PINK1 | Up | Promoting autophagy through stabilizing PINK1 protein | Alleviating dopaminergic neuronal injury | MPTP mouse model and MPP+-induced SH-SY5Y | [51] |
| SNHG1 | miR-221/222 | Up | SNHG1 downregulation increased LC3-II levels through the miR-221/222/p27/mTOR pathway | Enhancing α-synuclein aggregation thus leads to dopaminergic toxicity | MPTP mouse model and MPP+-induced MN9D cells | [56] | |
| HOTAIR | LRRK2 | Up | Promoting autophagy by improving LRRK2 mRNA stability and activating ERK/MAPK pathways | Increasing the loss of striatal dopamine and promoting PD progression | MPTP mouse model and MPP+-induced SH-SY5Y | [57] | |
| miR-126-5p | Suppressing autophagy trough HOTAIR/miR-126-5p/RAB3IP axis | Elevating the loss of striatal dopamine and the accumulation of α-synuclein | MPTP rat model and MPP+-induced SH-SY5Y | [59] | |||
| HAGLROS | miR-100 | Up | Inhibiting autophagy via regulating miR-100/ATG10 axis and PI3K/AKT/mTOR pathway activation | Contributing to the development of PD | MPTP mouse model and MPP+-induced SH-SY5Y | [61] | |
| Ischemic stroke | H19 | DUSP5 | Up | Promoting autophagy through regulating DUSP5-ERK1/2 axis | Aggravating cerebral ischemia-reperfusion injury | SH-SY5Y cell culture | [71] |
| SNHG12 | Unknown | Up | Activating beclin1-dependent autophagy | Alleviating cerebral ischemia-reperfusion injury | SH-SY5Y cell culture | [76] | |
| MALAT1 | miR-26b | Up | Activating autophagy through upregulation of ULK2 | Attenuating ischemia-reperfusion injury | BMEC culture | [74] | |
| miR-200c-3p | Promoting autophagy by regulating miR-200c-3p/Sirt1 axis | [75] | |||||
| miR-30a | Promoting autophagy via MALAT1/miR-30a/beclin1 axis | Aggravating ischemic injury | Mouse brain and N2a cells | [73] | |||
| KCNQ1OT1 | miR-200a | Up | Promoting the formation of autophagosomes through the mir-200a/FOXO3/ATG7 axis | Aggravating neurological impairments | Mouse brain and SH-SY5Y cell culture | [78] | |
| Epilepsy | MALAT1 | PI3K/AKT | Up | Activating autophagy by inhibiting PI3K/AKT signaling pathway | Promoting the degeneration and necrosis of hippocampal neurons | Rats with EP | [81] |
| Glioma | MEG3 | PI3K/AKT/mTOR | Down | Activating autophagy by downregulating of the PI3K/AKT/mTOR pathways and the upregulating of Sirt7 | Repressing cell proliferation and migration | U251 cell lines | [92] |
| Unknown | Suppressing cisplatin-induced excessive autophagy | Increasing glioma cell sensitive to cisplatin | U87 cell lines | [93] | |||
| PVT1 | miR-186 | Up | Activating autophagy through upregulation of ATG7 and Beclin1 | Promoting proliferation, migration and angiogenesis | hCMEC/D3 | [94] | |
| MALAT1 | miR-101 | Up | Activating autophagy through upregulation | Promoting cell proliferation | Glioma tissue from patients, U87, U118, | [96] | |
| STMN1, RAB5A, and ATG4D | U251, U373, and D247 cell lines | ||||||
| miR-384 | Activating autophagy through upregulation GOLM1 | Enhancing glioma migration and invasion | Glioma tissue from patients, SHG-44, and LN229 cells | [100] | |||
| GAS5 | mTOR | Up | Suppressing cisplatin-induced excessive autophagy in an mTOR-independent manner | Increasing glioma cell sensitive to cisplatin | U87 and u138 cell lines | [103] |
3.1. lncRNA-Mediated Autophagy in Alzheimer's Disease
Alzheimer's disease (AD) is one of the most common age-related neurodegenerative diseases and causes progressive memory impairment and cognitive dysfunction, accounting for 70% of cases of dementia [38]. However, the etiology and pathogenesis of AD remain unclear. At present, most scholars believe that the extracellular disposition of β-amyloid (Aβ) and the intraneuronal accumulation of tau protein are two major neuropathological features of AD. Studies have shown that autophagy inducers seem to prevent the accumulation of β-amyloid plaques and neurofibrillary tangles by degrading these aggregates in the early stages of AD, while the promotion of autophagy could aggravate the impaired autophagosome-lysosomal fusion and lysosomal dysfunction the late stages of AD [39]. Despite great advances in diagnostic and therapeutic drugs of AD in recent years, there are no effective treatments to prevent or reverse AD progression. Hence, understanding the regulatory mechanisms underlying autophagy will be the focus of treatment for AD. Recent studies found that lncRNAs were differentially expressed in the blood and CSF from patients with AD compared to the healthy elderly and played a key role in AD pathogenesis [40]. Nevertheless, there are few reports about the role and molecular mechanism of lncRNAs on autophagy in AD; here, we introduce lncRNAs involved in autophagy regulation in the AD, which contributes to AD pathogenesis and provides new diagnostic biomarkers or therapeutic targets for AD.
3.1.1. lncRNA 17A
lncRNA 17A is transcribed from the antisense strand of the GABABR2 gene and expressed in the human brain. Massone et al. suggested that the expression of lncRNA 17A was upregulated in the brain tissue of patients with AD and regulated Aβ secretion [41]. Wang et al. found that compared to lncRNA 17A overexpressed cells, lncRNA 17A knockdown increased the expression levels of LC3-II which is a hallmark of autophagosome formation. At the same time, the level of Aβ42 was diminished in shRNA-17A-transfected SH-SY5Y cells. Aβ42 is considered the main component of senile plaques and the primary mechanistic factor in AD pathology. Moreover, GABABR2 was found to be upregulated when lncRNA 17A was overexpressed and downregulated when lncRNA was knocked down. Therefore, the researchers speculated that depletion of lncRNA 17A promoted autophagy by regulating alternative splicing of GABABR2 and thus alleviated the accumulation of protein aggregates, which effectively suppress the progression of AD [42]. However, the effects of lncRNA 17A are needed to be examined in vivo.
3.1.2. lncRNA NEAT1
lncRNA NEAT1 has been reported to be a vital transcriptional regulator in cancer cell growth [43]. NEAT1 was well documented to be upregulated in the brain tissue from patients with AD, and its role in the pathophysiology of AD has received much attention in recent years [44, 45]. lncRNA NEAT1 is significantly upregulated in old APP/PS1 mice (over 6 months old) in a time-dependent manner but not in younger littermates, and the levels of Aβ showed a similar expression pattern. A further study found that overexpression of lncRNA NEAT1 could improve the interaction of PINK1 and NEDD4L and facilitate PINK1 ubiquitination. Besides, protein levels of the autophagy adaptors such as P62, OPTN, and LC3 were decreased at the same time Aβ was increased. Furthermore, lncRNA NEAT1 knockdown can reverse the above-mentioned changes and ameliorates cognitive impairments in AD mice. These studies suggested that lncRNA NEAT1 can promote NEDD4L-mediated PINK1 ubiquitination and degradation and thus inhibit PINK1-dependent mitophagy, which finally escalates Aβ accumulation and cognitive decline [46]. It is well known that PINK1 together with Parkin can promote the recruitment of autophagy receptor OPTN and activate ubiquitin and autophagic proteins, contributing to the information of autophagosome [47]. From what has been discussed above, lncRNA NEAT1 aggravated Aβ-induced neuronal damage via promoting the ubiquitination and degradation of PINK1. Hence, lncRNA NEAT1 may be a useful biomarker for AD.
3.2. lncRNA-Mediated Autophagy in Parkinson's Disease
PD is the second most common neurodegenerative disorder after AD in aging individuals, the main characteristics of which are the progressive loss of dopaminergic neurons in the substantia nigra pars compacta and the formation of Lewy bodies within the cytoplasm, leading to motor dysregulation [48, 49]. Although the pathogenesis remains not fully understood, PD is widely assumed to be associated with both genetic and environmental factors [35]. Most PD cases are sporadic with an unclear etiology, but about 5% are familiar caused by genic mutations, including SNCA, LRRK2, PRKN, PINK1, MAPT, GBA, and PARK2 [50]. Mutations in these genes can cause the formation of cytotoxic aggregates, impaired actin remodeling, dysregulation of autophagy, and enhancement of proapoptotic signaling pathways and eventually lead to neuron degeneration. Recently, growing evidences indicated that lncRNA takes part in the pathogenesis of PD. In the following paragraphs, we focus on the role of lncRNA-mediated autophagy in PD.
3.2.1. lncRNA NEAT1
Similar to its role in AD, lncRNA NEAT1 has also been proved to play key roles in PD pathophysiology. MPTP can significantly promote the expression level of lncRNA NEAT1, PINK1, and LC3-II in vitro and in vivo models of PD, which also seems to increase with dose and time within a certain range [51, 52]. Upregulated NEAT1 expression can further increase PINK1 expression level by inhibiting CHX-induced PINK1 protein degradation, which occurs both in impaired and in intact mitochondria. Moreover, the downregulation of NEAT1 largely reversed the effect of MPP+ on SH-SY5Y cells, including decreased LC3-II and PINK1 protein levels. The accumulated PINK1 directly interacts with LC3-II and increases the accumulation of LC3-II in mitochondria, leading to abnormal mitochondrial autophagy [51]. Interestingly, the NEAT1/PINK1/ LC3-II axis is involved not only in the degradation of damaged mitochondria in PD models but also in the abnormal elimination of the healthy mitochondria leading to reduced ATP generation thereby causing neurodegeneration [53]. The above results indicated that lncRNA NEAT1 could induce abnormal autophagy by stabilizing PINK1 which was an LC3-II upstream regulatory factor and played a role in the pathogenesis of PD.
3.2.2. lncRNA SNHG1
It has been reported that the expression level of lncRNA SNHG1 was significantly upregulated in postmortem midbrain samples from PD patients compared to healthy people [54]. Besides, lncRNA SNHG1 also has been demonstrated to promote α-synuclein aggregation and enhance neurotoxicity resulting in dopaminergic neuronal loss [55]. Recent studies have found lncRNA SNHG1 gradually upregulated in cell and animal models in PD. Furthermore, silencing lncRNA SNHC1 could remarkably increase the expression of miR-221/222 and LC3-II and prevent MPP+-induced cell death. Further investigations indicated that lncRNA SNHG1 acted as a ceRNA and prevented miR-221/222 from interacting with target P27 mRNA which is a key regulatory factor for the phosphorylation of mTOR and cell death. The decreased expression of P27 can inhibit the mTOR pathway, thus promoting neural autophagy and alleviating MPP+-mediated cell injury [56]. Experiment result shows that downregulated lncRNA SNHG1 inhibits the mTOR pathway and initiates autophagy through sponging miR-221/222, which can reduce the death of dopaminergic neurons in PD patients.
3.2.3. lncRNA HOTAIR
Several studies have indicated that lncRNA HOTAIR obviously increased in vivo and vitro models of PD [1, 57]. As previously mentioned, mutations of LRRK2 are associated with familial PD and they can enhance autophagy by activating the ERK/MAPK pathway [58]. Overexpression of lncRNA HOTAIR could specifically improve LRRK2 mRNA stability and upregulate its expression, which promoted autophagy in PD models. On the contrary, silencing HOTAIR would rescue these alterations and increase cell viability [57]. These results strongly indicated that lncRNA HOTAIR can activate the ERK/MAPK pathway through improving the stability of LRRK2, which can inhibit autophagy and ultimately lead to PD. Recently, Lin et al. found that lncRNA HOTAIR was upregulated in MPP+-treated SH-SY5Y cell lines and contribute to PD while downregulating HOTAIR would increase the number of TH-positive cells and decrease the number of α-synuclein-positive cells. Similar results were also observed in in vivo experiments. A further study showed that miR-126-5p negatively regulated the expressions of RAB3IP which have been demonstrated to inhibit autophagy in mammalian cells. lncRNA HOTAIR could prevent miR-126-5P from interacting with RAB3IP by sponging miR-126-5P, which would inhibit autophagy and eventually caused the accumulation of α-synuclein in dopaminergic neurons [59]. Overall, the lncRNA HOTAIR/miR-126-5p/RAB3IP axis has been proved to be related to autophagy in PD, and its dysregulation was considered a therapeutic target for PD.
3.2.4. lncRNA HAGLROS
A recent study showed that lncRNA HAGLROS was upregulated and miR-100 was downregulated in MPTP-induced mice and MPP+-treated SH-SY5Y cells. Knockdown of lncRNA HAGLROS could increase the expression level of miR-100 and alleviated MPTP-induced autophagy. A further study found that ATG 10 which participated in the formation of autophagosomes and initiated autophagy was a direct target of miR-100 [60]. lncRNA HAGROS could prevent miR-100 from interacting with ATG10 by serving as a sponge of miR-100 and thus promote autophagy, leading to aggravating MPP+-induced cell injury. In addition, the PI3K/AKT/mTOR pathway was found to be inactivated in MPP+-intoxicated SH-SY5Y cell, which led to the decrease in the number of dopamine-positive cells and aggravated cell damage; meanwhile, lncRNA HAGLROS silencing could dramatically increase the phosphorylation levels of the PI3K/AKT/mTOR pathway and thus promote autophagy, contributing to protecting neurons from MPTP-induced damage [61]. Together, lncRNA HAGLROS regulates autophagy through regulating miR-100/ATG10 axis and PI3K/AKT/mTOR, which provide a potential therapeutic strategy for PD and need further evidence.
3.3. lncRNA-Mediated Autophagy in Ischemic Stroke
IS is a generic term for blood flow interruption and cerebral tissue necrosis caused by a thrombotic or embolic blockage of a cerebral artery, accounting for approximately 85% of all strokes [62, 63]. After brain ischemia, nerve cell membrane potential and cellular ion homeostasis are disrupted leading to a series of deleterious events in the brain such as excitotoxicity, oxidative stress, autophagy, and inflammation. These events affect each other, forming a positive feedback loop and causing ischemic cascading effects, which causes irreversible neuronal injury characterized by neuronal apoptosis or death in the core area, brain edema, and blood-brain barrier dysfunction [64, 65]. The current research indicated that autophagy was activated after cerebral ischemia; at this time, inhibition of autophagy plays a protective role whereas excessive activation of autophagy aggravates the injury. However, the inhibition of basal autophagy before ischemic stroke can aggravate subsequent cerebral ischemic injury [66, 67]. Therefore, autophagy induction may be a potential therapeutic target for IS [68]. We will review some regulatory lncRNAs on autophagy in IS in this section.
3.3.1. lncRNA H19
lncRNA H19, a maternally imprinted gene, generally declines after birth, whereas it increases in pathological situations such as cancer, oxidative stress, or hypoxia, which is important for early embryonic development [69]. The expression levels of lncRNA H19 are dramatically increased in the peripheral blood of ischemic patients and brain tissue, plasma, and white blood cells of mice with transient cerebral ischemia. Wang et al. found that compared to the control group, the human neuroblastoma cell line SH-SY5Y subjected to oxygen-glucose deprivation/reoxygenation (OGD/R) significantly induced the expression of lncRNA H19, and at the same time, the ratio of LC3II/I and beclin1 was increased, while P62 was decreased. Knocking out lncRNA h19 has been reported to reverse these changes. Further studies have shown that the overexpression of lncRNA H19 can activate autophagy by inhibiting DUSP5, a mitogen-activated protein kinase phosphatase, and thus activating ERK1/2 which is related to autophagy initiation [70, 71]. Taken together, lncRNA H19 can promote autophagy through regulating the DUSP5-ERK1/2 axis, contributing to cerebral ischemia-reperfusion injury. However, these results still need to be further verified by in vivo experiments.
3.3.2. lncRNA MALAT1
lncRNA MALAT1 was considered to be one of the most significantly upregulated lncRNAs in both in vivo and in vitro models of IS, accompanied by the upregulation of LC3-II and Beclin1 [72, 73]. Further studies by Guo et al. showed that lncRNA MALAT1 decreases Beclin1 expression by acting as a sponge for miR-30a and thus promotes Beclin1-dependent autophagy, leading to neuronal cell death after IS. Moreover, lncRNA MALAT1 silencing can alleviate ischemic brain injury by inhibiting autophagy. This suggests that the MALAT1/miR-30a/beclin1 (lncRNA/miRNA/mRNA) regulatory network may exist in ischemic stroke [73]. Interestingly, lncRNA MALAT1 and autophagy have a protective effect on BMECs during IS. MALAT1 functions as a ceRNA for miR-26b, which can promote ULK2 expression. ULK2 is a downstream target in the mTOR signaling pathway and is associated with autophagosome formation, suggesting that MALAT1 protected BMECs from ischemia-reperfusion injury by promoting autophagy [74]. Another study by Wang et al. reported that the in vitro BMEC model, lncRNA MALAT1, also serves as ceRNA and prevents miR-200c-3p from binding to Sirt1 which has been reported to stimulate the expression and deacetylation of autophagy-related genes, suggesting that lncRNA MALAT1 can activate autophagy and promote the survival of OGD/R-treated BMECs by regulating miR-200c-3p/Sirt1 [75]. Collectively, lncRNA MALAT1 can regulate autophagy in IS through sponging miRNA and abolishing their effects on autophagy-related factors.
3.3.3. lncRNA SNHG12
Recently, lncRNA SNHG12 was found to be significantly elevated in mouse MCAO models and OGD/R models in SH-SY5Y cells [72, 76]. An in vitro study has confirmed that overexpression of lncRNA SNHG12 could promote LC3-II and Beclin1 expression levels and the survival of SH-SY5Y cell lines after OGD/R, while downregulation of SNHG12 rescued these effects. Besides, autophagy inhibitor 3-MA can weaken the protective effect of lncRNA SNHG12 overexpression on I/R injury, suggesting that lncRNA SNHG12, as an autophagy inducer, can attenuate brain I/R injury and may be a new therapeutic target for ischemic stroke [76]. Does lncRNA SNHG12 regulate mTOR, a classical autophagy signaling pathway, or autophagy-related proteins? The mechanisms of lncRNA SNHG12 on autophagy following IS remain to be elucidated.
3.3.4. lncRNA KCNQ10T1
lncRNA KCNQ1OT1 is significantly upregulated in the peripheral blood of patients with ischemic stroke. Previous studies have shown that lncRNA KCNQ1OT1 is associated with risk factors for ischemic stroke, such as diabetes and myocardial infarction [77]. A recent study substantiated that lncRNA KCNQ1OT1 silencing reduced cerebral infarction volume and alleviated neurological deficits in mouse MCAO models as well as improved cell viability of OGD/R-treated SH-SY5Y cells. Besides, rapamycin, an autophagy inducer, reversed these effects, suggesting that the downregulation of lncRNA KCNQ1OT1 protects neurons from ischemia injury by inhibiting autophagy. Further studies found that lncRNA KCNQ1OT1 may be the ceRNA of miR-200a and prevented targeting of FOXO3 and thus promoted the expression of ATG7 which participates in vesicle elongation. These experiments indicated that the knockdown of KCNQ1OT1 may inhibit the formation of autophagosomes through the miR-200a/FOXO3/ATG7 axis and increase cell viability. This finding provides a potential novel strategy for the treatment of ischemic stroke [78].
3.4. lncRNA-Mediated Autophagy in Epilepsy
Epilepsy (EP), as one of the most common neurological disorders, is a kind of chronic syndrome mainly caused by abnormal discharge of brain neurons [79]. It is characterized by spontaneous seizures with a high recurrence rate of 60%. Despite the availability of antiepileptic drugs (AEDs), many patients still suffer refractory seizures and unacceptable side effects [80]. Currently, the pathogenesis of epilepsy has not been well defined and the reports about the role of autophagy in epilepsy remain rare. Wu et al. showed that lncRNA MALAT1 was obviously upregulated in the hippocampus of rats with EP and the expression of LC3II/LC3I and Beclin1 also upregulated compared to the control group, indicating that EP may lead to excessive autophagy in hippocampal neurons of rats [81]. In addition, further mechanism analysis showed that lncRNA MALAT1 activated autophagy by inhibiting PI3K/AKT signaling pathway while silencing MALAT1 could prohibit autophagy in hippocampal neurons of epileptic models. These results bring up a hint that downregulated lncRNA MALAT1 can protect the hippocampal neurons from excessive autophagy through activating PI3K/AKT signaling pathway, contributing to attenuating neuron injury after EP. These findings may help to elucidate the pathophysiology of epilepsy and provide a potential therapeutic target.
3.5. lncRNA-Mediated Autophagy in Glioma
Glioma has been considered to be the most common type of primary brain tumor with high mortality and poor prognosis, accounting for 30% of central nervous system tumors and 80% of all malignant brain tumors [82]. The growth and metastasis of glioma rely on angiogenesis, which is one of the main reasons for treatment failure [83]. Cisplatin, a chemotherapeutic agent, binds to DNA and causes DNA damage-induced tumor cell death, which is extensively used for the treatment of glioma currently [84]. The role of autophagy in cancer varies from person to person. On the one hand, autophagy contributes to maintaining cellular homeostasis and can suppress tumor growth; on the other hand, autophagy may also facilitate proliferation and survival of tumor cells thus promoting tumor growth, invasion, and metastasis [85, 86]. In recent years, more and more studies have shown that autophagy played a key role in the development of tumors, including cell proliferation, metastasis, and chemotherapy resistance [87].
Interestingly, the latest evidence suggested that lncRNAs are involved in the development and cisplatin sensitivity of glioma through regulating autophagy. In the following paragraphs, we will discuss the regulatory lncRNAs in autophagy during glioma development and their specific mechanisms to explore a more suitable therapeutic target for treating glioma.
3.5.1. lncRNA MEG3
lncRNA MEG3 is widely recognized as a tumor suppressor gene in several types of human cancers. lncRNA MEG3 was found to be markedly downregulated in glioma tissues and cell lines, which is an independent biomarker of poor prognosis in glioma [88]. It usually functioned as a ceRNA; for example, it could prevent miR-19a and miR-93 from interacting with their target mRNAs and elevate the expression levels of PTEN and PHLPP2 respectively, which inhibited PI3K/AKT/mTOR pathway and eventually suppressed the proliferation of glioma [89, 90]. Xu et al. revealed that overexpression of lncRNA MEG3 repressed cell proliferation and migration but promoted autophagy in U251 cells. Beclin1 and LC3-II/LC3-I were upregulated whereas P62 was downregulated; interestingly, these autophagy-related proteins' expressions were still unchanged after lncRNA MEG3 silencing, suggesting that autophagy in U251 cells was induced by inhibiting autophagosome degradation after overexpression of lncRNA MEG3, and inhibition of autophagy can improve cell viability. Furthermore, lncRNA MEG3 overexpression decreased the phosphorylation levels of key kinases in PI3K/AKT/mTOR pathways, indicating the inactivation of the PI3K/AKT/mTOR pathways and the upregulation of Sirt7 that is related to the deacetylation of autophagy- related genes and participated in various types of cancers [91]. However, lncRNA MEG3 or Sirt7 silencing exhibited the utter opposite effects. Collectively, lncRNA MEG3 decreased the phosphorylation levels of PI3K/AKT/mTOR pathways by enhancing Sirt7 and finally activates autophagy and improves the prognosis of glioma [92]. These results may provide novel strategies of glioma treatment, but the specific molecular mechanisms between MEG3 and Sirt7 require further investigation, and the functional role of MEG3 needs to be verified in in vivo experiments for future clinical application.
Another study showed that the expression levels of lncRNA MEG3 in U87 cells were induced by cisplatin in a time- and dose-dependent manner. Overexpression of lncRNA MEG3 enhanced the chemosensitivity of U87 cell lines to cisplatin through inhibiting cisplatin-induced autophagy, whereas knockdown of lncRNA MEG3 increased resistance of U87 cell lines to cisplatin by promoting cisplatin-induced autophagy [93]. These studies suggest that lncRNA MEG3 may be a potential target for the treatment of cisplatin-resistance glioma.
3.5.2. lncRNA PVTI
A recent study found that lncRNA PVT1 was upregulated in glioma vascular endothelial cells and miR-186 was downregulated. Moreover, lncRNA PVT1 overexpression or miR-186 knockdown increased the expression levels of ATG7, Beclin-1, and LC3-II/LC3-I whereas it decreased the level of P62, contributing to cell proliferation, migration, and angiogenesis. Meanwhile, autophagy inhibitors could reverse these effects. Further mechanism analysis suggested that lncRNA PVT1 was bound to miR-186 directly and abolished its negative effects of ATG7 and Beclin-1 which is essential for autophagy initiation and the formation of a double-membrane structure. These studies are a hint that lncRNA PVT1 induces autophagy and thus promotes proliferation, migration, and angiogenesis of glioma-conditioned vascular endothelial cells through regulating miR-186-ATG7/Beclin-1 expression [94]. lncRNA PVT1 and miR-186 would provide an antiangiogenic target for gliomas.
3.5.3. lncRNA MALAT1
It has been reported that lncRNA MALAT1 was highly expressed in glioma tissue and served as an indicator for poor prognosis in glioma patients; however, the regulatory mechanism of lncRNA MALAT1 in human glioma was rarely studied [95]. Recently, lncRNA MALAT1 was found to be highly expressed in glioma tissues compared with adjacent normal tissues, and its elevated expression was positively associated with the LC3-II level. In vitro experiments also showed that lncRNA Malat1 significantly promoted autophagy and proliferation of glioma cells. More importantly, the inhibition of autophagy by 3-MA alleviated MALAT1-induced glioma proliferation, suggesting that lncRNA MALAT1 could activate autophagy and eventually promote glioma proliferation. Further molecular mechanism analysis revealed that lncRNA MALAT1 could directly bind to miR-101 and prevent it from interacting with the 3′-UTR of STMN1, RAB5A, and ATG4D mRNA [96]. STMN1, RAB5A, and ATG4D were shown to be important autophagic regulators. STMN1 and RAB5A affected the fusion of autophagosomes with lysosomes, whereas ATG4D participated in autophagosome maturation [97–99]. These experimental results demonstrated that lncRNA MALAT1 promoted autophagy and proliferation of glioma cells by regulating the Malat1-miR-101-STMN1/RAB5A/ATG4D network [96]. In the latest study, lncRNA MALAT1 also acted as a miRNA sponge to regulate autophagy in glioma cells. Knockdown of lncRNA MALAT1 could depress glioma cell autophagy, migration, and invasion, whereas inhibiting miR-384 could eliminate these effects [100]. Besides, GOLM1, a downstream target of miR-384, was also identified to promote autophagy by activating protein kinase AKT [101], suggesting that lncRNA MALAT1, as a miR-384 sponge, promoted vesicle nucleation and thus enhanced glioma migration and invasion by upregulating GOLM1 [100]. This newly discovered lncRNA MALAT1/miR-384/GOLM1 axis may provide new insights into the mechanisms of glioma metastasis, and lncRNA MALAT1 may be a promising target for future glioma therapy.
3.5.4. lncRNA GAS5
It has been reported that lncRNA GAS5 suppressed glioma stem cell proliferation and high expression level of lncRNA GAS5 was associated with the 2-year overall survival rate of patients with glioma [102]. Recently, a study showed that lncRNA GAS5 downregulation reduced the sensitivity of U87 cells that had high GAS5 levels to cisplatin. In contrast, lncRNA GAS5 overexpression reduced U138 cells that had a relatively low GAS5 levels resistance to cisplatin. These results suggested that lncRNA GAS5 may increase the sensitivity of glioma cells to cisplatin and play an important role in glioma chemoresistance. A Further mechanism study revealed that exposure to cisplatin could increase the expression levels of LC3II and decrease P62 levels, thus leading to excessive autophagy. In addition, upregulated lncRNA GAS5 activated mTOR signaling that was restrained by cisplatin and eventually inhibited cisplatin-induced excessive autophagy. Interestingly, blocking the mTOR pathway could also overturn the positive effect of lncRNA GAS5 upregulation on chemosensitivity to cisplatin. However, how lncRNA GAS5 regulates the mTOR signaling pathway needs to be further studied. In a word, lncRNA GAS5 suppressed cisplatin-induced excessive autophagy and thus increased cisplatin sensitivity in an mTOR-independent manner, suggesting that lncRNA GAS5 was a potential and promising target for overcoming glioma chemoresistance [103].
4. Conclusion and Perspectives
The above-mentioned studies have corroborated the involvement of lncRNAs in autophagy and have opened the way for further investigations into the function and mechanism of lncRNAs in neurological diseases. At present, RNA-targeted or RNA-based therapeutic approaches include antisense oligonucleotides (ASOs), RNA interference (RNAi), and ribozymes with intrinsic catalytic activities and RNA aptamers [104]. It was shown that ASOs regulated noncoding RNAs via the following two main methods. One is to synthesize antagonists to inhibit noncoding RNAs binding to their mRNA targets. The other use of ASOs is to prepare miRNA mimetics to restore levels of miRNAs that have been reduced in pathogenic conditions [105]. In 2016, two ASOs, eteplirsen and nusinersen, were approved by the FDA for the treatment of Duchenne muscular dystrophy and spinal muscular atrophy respectively, which opened a new era of ASO therapies for neurodegenerative diseases [106]. Currently, a phase 1 clinical trial of a MAPT-targeting ASO and LRRK2 ASO BIIB094 has been initiated in mild AD patients (NCT03186989) and PD patients (NCT03976349), respectively. Unlike ASO therapeutics which can be ended by terminating treatment, RNAi-based therapeutics when delivered as an shRNA can develop nonreversible efficacy, which may be beneficial but also comes with risk since it may cause a decrease in untargeted proteins [107]. A growing body of evidence points towards the promise of nanoparticles as carriers for siRNA and shRNA therapeutics for neurodegenerative disorders, including shRNAs targeting a α-synuclein in a mouse model of PD, siRNAs targeting BACE1 and APP in mouse CNS toward treating AD [104]. Nevertheless, these promising RNA-based therapies also face substantial challenges. Firstly, most RNA-targeted drugs cannot cross the blood brain barrier and has to be delivered to the central nervous system through intrathecal injection, an invasive delivery method that limits its use. Secondly, it is well known that lncRNA has a variety of biological functions and complex potential mechanisms; however, at present, studies on the mechanism of lncRNA are generally limited to finding relevant miRNA or binding proteins, not to mention poor conservation of lncRNA among species. Lastly, neurons possess limited ability to regenerate so that considerable undetected and potentially irreversible damage is likely to have occurred before a patient reports symptom to a physician, which requires us to find reliable biomarkers for early diagnosis and treatment. For example, in vivo and in vitro, certain lncRNA dynamically changes over ischemia time, and thus, the level of lncRNA in blood samples may reflect the pathophysiological state of the brain, which may be used as a biomarker in clinical application like myocardial markers in the future. In conclusion, this review provides an overview of lncRNAs in autophagy regulation and new insights into the underlying mechanisms and may be able to provide new ideas for studying the possible role of lncRNAs on regulating the press of neurological diseases.
Acknowledgments
This work was supported by the National Nature Science Foundation of China (81671181), the Natural Science Foundation of Guangdong Province (2017A030310658), Guangdong Province Colleges and Universities Young Innovative Talents Project (2018KQNCX095), and Guangdong Province Universities and Colleges Pearl River Scholar Funding Scheme (2017).
Contributor Information
Wangtao Zhong, Email: 158980116@qq.com.
Yujie Cai, Email: musejie163@163.com.
Data Availability
No data were used to support this study.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- 1.Oe S., Kimura T., Yamada H. Regulatory non-coding RNAs in nervous system development and disease. Frontiers in Bioscience (Landmark edition) 2019;24(7):1203–1240. doi: 10.2741/4776. [DOI] [PubMed] [Google Scholar]
- 2.Wang J.-F., Mei Z.-G., Fu Y., et al. Puerarin protects rat brain against ischemia/reperfusion injury by suppressing autophagy via the AMPK-mTOR-ULK1 signaling pathway. Neural Regeneration Research. 2018;13(6):989–998. doi: 10.4103/1673-5374.233441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Dikic I., Elazar Z. Mechanism and medical implications of mammalian autophagy. Nature Reviews Molecular Cell Biology. 2018;19(6):349–364. doi: 10.1038/s41580-018-0003-4. [DOI] [PubMed] [Google Scholar]
- 4.Jiang Z., Watts L. T., Huang S., et al. The effects of methylene blue on autophagy and apoptosis in MRI-defined normal tissue, ischemic penumbra and ischemic core. PLoS One. 2015;10(6, article e0131929) doi: 10.1371/journal.pone.0131929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hou K., Xu D., Li F., Chen S., Li Y. The progress of neuronal autophagy in cerebral ischemia stroke: mechanisms, roles and research methods. Journal of the Neurological Sciences. 2019;400:72–82. doi: 10.1016/j.jns.2019.03.015. [DOI] [PubMed] [Google Scholar]
- 6.Xilouri M., Stefanis L. Autophagy in the central nervous system: implications for neurodegenerative disorders. CNS & Neurological Disorders Drug Targets. 2010;9(6):701–719. doi: 10.2174/187152710793237421. [DOI] [PubMed] [Google Scholar]
- 7.Conway O., Akpinar H. A., Rogov V. V., Kirkin V. Selective autophagy receptors in neuronal health and disease. Journal of Molecular Biology. 2020;432(8):2483–2509. doi: 10.1016/j.jmb.2019.10.013. [DOI] [PubMed] [Google Scholar]
- 8.Kesidou E., Lagoudaki R., Touloumi O., Poulatsidou K. N., Simeonidou C. Autophagy and neurodegenerative disorders. Neural Regeneration Research. 2013;8(24):2275–2283. doi: 10.3969/j.issn.1673-5374.2013.24.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Scrivo A., Bourdenx M., Pampliega O., Cuervo A. M. Selective autophagy as a potential therapeutic target for neurodegenerative disorders. The Lancet Neurology. 2018;17(9):802–815. doi: 10.1016/S1474-4422(18)30238-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Choudhry H., Harris A. L., McIntyre A. The tumour hypoxia induced non-coding transcriptome. Molecular Aspects of Medicine. 2016;47-48:35–53. doi: 10.1016/j.mam.2016.01.003. [DOI] [PubMed] [Google Scholar]
- 11.Wang K., Liu C.-Y., Zhou L.-Y., et al. APF lncRNA regulates autophagy and myocardial infarction by targeting miR-188-3p. Nature Communications. 2015;6(1, article 6779) doi: 10.1038/ncomms7779. [DOI] [PubMed] [Google Scholar]
- 12.Di Ge L. H., Huang S. Y., Peng N., et al. Identification of a novel MTOR activator and discovery of a competing endogenous RNA regulating autophagy in vascular endothelial cells. Autophagy. 2014;10(6):957–971. doi: 10.4161/auto.28363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Huang S. Y., Lu W., Di Ge N. M., et al. A new microRNA signal pathway regulated by long noncoding RNA TGFB2-OT1 in autophagy and inflammation of vascular endothelial cells. Autophagy. 2015;11(12):2172–2183. doi: 10.1080/15548627.2015.1106663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Derrien T., Johnson R., Bussotti G., et al. The GENCODE v7 catalog of human long noncoding RNAs: analysis of their gene structure, evolution, and expression. Genome Research. 2012;22(9):1775–1789. doi: 10.1101/gr.132159.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Akella A., Bhattarai S., Dharap A. Long noncoding RNAs in the pathophysiology of ischemic stroke. Neuromolecular Medicine. 2019;21(4):474–483. doi: 10.1007/s12017-019-08542-w. [DOI] [PubMed] [Google Scholar]
- 16.Guttman M., Garber M., Levin J. Z., et al. Ab initio reconstruction of cell type-specific transcriptomes in mouse reveals the conserved multi-exonic structure of lincRNAs. Nature Biotechnology. 2010;28(5):503–510. doi: 10.1038/nbt.1633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Carpenter S., Aiello D., Atianand M. K., et al. A long noncoding RNA mediates both activation and repression of immune response genes. Science (New York, N.Y.) 2013;341(6147):789–792. doi: 10.1126/science.1240925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Schaukowitch K., Kim T.-K. Emerging epigenetic mechanisms of long non-coding RNAs. Neuroscience. 2014;264:25–38. doi: 10.1016/j.neuroscience.2013.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhang X., Wu N., Wang J., Li Z. LncRNA MEG3 inhibits cell proliferation and induces apoptosis in laryngeal cancer via miR-23a/APAF-1 axis. Journal of Cellular and Molecular Medicine. 2019;23(10):6708–6719. doi: 10.1111/jcmm.14549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Guo J., Liu Z., Gong R. Long noncoding RNA: an emerging player in diabetes and diabetic kidney disease. Clinical science (London, England: 1979) 2019;133(12):1321–1339. doi: 10.1042/CS20190372. [DOI] [PubMed] [Google Scholar]
- 21.Monteiro J. P., Bennett M., Rodor J., Caudrillier A., Ulitsky I., Baker A. H. Endothelial function and dysfunction in the cardiovascular system: the long non-coding road. Cardiovascular Research. 2019;115(12):1692–1704. doi: 10.1093/cvr/cvz154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Barangi S., Hayes A. W., Reiter R., Karimi G. The therapeutic role of long non-coding RNAs in human diseases: a focus on the recent insights into autophagy. Pharmacological Research. 2019;142:22–29. doi: 10.1016/j.phrs.2019.02.010. [DOI] [PubMed] [Google Scholar]
- 23.Wang K. C., Chang H. Y. Molecular mechanisms of long noncoding RNAs. Molecular Cell. 2011;43(6):904–914. doi: 10.1016/j.molcel.2011.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bhat S. A., Ahmad S. M., Mumtaz P. T., et al. Long non-coding RNAs: mechanism of action and functional utility. Non-coding RNA Research. 2016;1(1):43–50. doi: 10.1016/j.ncrna.2016.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Han X., Yang F., Cao H., Liang Z. Malat1regulates serum response factor through miR-133 as a competing endogenous RNA in myogenesis. FASEB Journal. 2015;29(7):3054–3064. doi: 10.1096/fj.14-259952. [DOI] [PubMed] [Google Scholar]
- 26.An Y., Furber K. L., Ji S. Pseudogenes regulate parental gene expression via ceRNA network. Journal of Cellular and Molecular Medicine. 2017;21(1):185–192. doi: 10.1111/jcmm.12952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Guennewig B., Cooper A. A. The central role of noncoding RNA in the brain. International Review of Neurobiology. 2014;116:153–194. doi: 10.1016/B978-0-12-801105-8.00007-2. [DOI] [PubMed] [Google Scholar]
- 28.Li X., Wu Z., Fu X., Han W. lncRNAs: insights into their function and mechanics in underlying disorders. Mutation Research/Reviews in Mutation Research. 2014;762:1–21. doi: 10.1016/j.mrrev.2014.04.002. [DOI] [PubMed] [Google Scholar]
- 29.Saxena A., Carninci P. Long non-coding RNA modifies chromatin: epigenetic silencing by long non-coding RNAs. BioEssays: News and Reviews in Molecular, Cellular and Developmental Biology. 2011;33(11):830–839. doi: 10.1002/bies.201100084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tsai M.-C., Manor O., Wan Y., et al. Long noncoding RNA as modular scaffold of histone modification complexes. Science (New York, N.Y.) 2010;329(5992):689–693. doi: 10.1126/science.1192002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Guttman M., Donaghey J., Carey B. W., et al. lincRNAs act in the circuitry controlling pluripotency and differentiation. Nature. 2011;477(7364):295–300. doi: 10.1038/nature10398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Qureshi I. A., Mehler M. F. Long non-coding RNAs: novel targets for nervous system disease diagnosis and therapy. Neurotherapeutics. 2013;10(4):632–646. doi: 10.1007/s13311-013-0199-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bao M.-H., Szeto V., Yang B. B., Zhu S.-z., Sun H.-S., Feng Z.-P. Long non-coding RNAs in ischemic stroke. Cell Death & Disease. 2018;9(3):p. 281. doi: 10.1038/s41419-018-0282-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kopp F., Mendell J. T. Functional classification and experimental dissection of long noncoding RNAs. Cell. 2018;172(3):393–407. doi: 10.1016/j.cell.2018.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hashemzaei M., Heravi R. E., Rezaee R., Roohbakhsh A., Karimi G. Regulation of autophagy by some natural products as a potential therapeutic strategy for cardiovascular disorders. European Journal of Pharmacology. 2017;802:44–51. doi: 10.1016/j.ejphar.2017.02.038. [DOI] [PubMed] [Google Scholar]
- 36.Roos W. P., Thomas A. D., Kaina B. DNA damage and the balance between survival and death in cancer biology. Nature Reviews Cancer. 2016;16(1):20–33. doi: 10.1038/nrc.2015.2. [DOI] [PubMed] [Google Scholar]
- 37.Zhang J., Wang P., Wan L., Xu S., Pang D. The emergence of noncoding RNAs as Heracles in autophagy. Autophagy. 2017;13(6):1004–1024. doi: 10.1080/15548627.2017.1312041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ng R., Chan K.-H. Potential neuroprotective effects of adiponectin in Alzheimer's disease. International Journal of Molecular Sciences. 2017;18(3):p. 592. doi: 10.3390/ijms18030592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Nah J., Yuan J., Jung Y.-K. Autophagy in neurodegenerative diseases: from mechanism to therapeutic approach. Molecules and Cells. 2015;38(5):381–389. doi: 10.14348/molcells.2015.0034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhao Y., Zhang Y., Zhang L., Dong Y., Ji H., Shen L. The potential markers of circulating microRNAs and long non-coding RNAs in Alzheimer's disease. Aging and Disease. 2019;10(6):1293–1301. doi: 10.14336/AD.2018.1105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Massone S., Vassallo I., Fiorino G., et al. 17A, a novel non-coding RNA, regulates GABA B alternative splicing and signaling in response to inflammatory stimuli and in Alzheimer disease. Neurobiology of Disease. 2011;41(2):308–317. doi: 10.1016/j.nbd.2010.09.019. [DOI] [PubMed] [Google Scholar]
- 42.Wang X., Zhang M., Liu H. LncRNA17A regulates autophagy and apoptosis of SH-SY5Y cell line as anin vitromodel for Alzheimer's disease. Bioscience, Biotechnology, and Biochemistry. 2019;83(4):609–621. doi: 10.1080/09168451.2018.1562874. [DOI] [PubMed] [Google Scholar]
- 43.Nong W. Long non-coding RNA NEAT1/miR-193a-3p regulates LPS-induced apoptosis and inflammatory injury in WI-38 cells through TLR4/NF-κB signaling. American Journal of Translational Research. 2019;11(9):5944–5955. [PMC free article] [PubMed] [Google Scholar]
- 44.Ke S., Yang Z., Yang F., Wang X., Tan J., Liao B. Long noncoding RNA NEAT1 aggravates Aβ-induced neuronal damage by targeting miR-107 in Alzheimer's disease. Yonsei Medical Journal. 2019;60(7):640–650. doi: 10.3349/ymj.2019.60.7.640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhao M.-Y., Wang G.-Q., Wang N.-N., Yu Q.-Y., Liu R.-L., Shi W.-Q. The long-non-coding RNA NEAT1 is a novel target for Alzheimer's disease progression via miR-124/BACE1 axis. Neurological Research. 2019;41(6):489–497. doi: 10.1080/01616412.2018.1548747. [DOI] [PubMed] [Google Scholar]
- 46.Huang Z., Zhao J., Wang W., Zhou J., Zhang J. Depletion of LncRNA NEAT1 rescues mitochondrial dysfunction through NEDD4L-dependent PINK1 degradation in animal models of Alzheimer's disease. Frontiers in Cellular Neuroscience. 2020;14:p. 28. doi: 10.3389/fncel.2020.00028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.McWilliams T. G., Muqit M. M. K. PINK1 and Parkin: emerging themes in mitochondrial homeostasis. Current Opinion in Cell Biology. 2017;45:83–91. doi: 10.1016/j.ceb.2017.03.013. [DOI] [PubMed] [Google Scholar]
- 48.Lyu Y., Bai L., Qin C. Long noncoding RNAs in neurodevelopment and Parkinson's disease. Animal Models and Experimental Medicine. 2019;2(4):239–251. doi: 10.1002/ame2.12093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Martinez-Vicente M. Autophagy in neurodegenerative diseases: from pathogenic dysfunction to therapeutic modulation. Seminars in Cell & Developmental Biology. 2015;40:115–126. doi: 10.1016/j.semcdb.2015.03.005. [DOI] [PubMed] [Google Scholar]
- 50.Klein C., Westenberger A. Genetics of Parkinson's disease. Cold Spring Harbor Perspectives in Medicine. 2012;2(1, article a008888) doi: 10.1101/cshperspect.a008888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yan W., Chen Z.-Y., Chen J.-Q., Chen H.-M. LncRNA NEAT1 promotes autophagy in MPTP-induced Parkinson's disease through stabilizing PINK1 protein. Biochemical and Biophysical Research Communications. 2018;496(4):1019–1024. doi: 10.1016/j.bbrc.2017.12.149. [DOI] [PubMed] [Google Scholar]
- 52.Xie S.-P., Zhou F., Li J., Duan S.-j. NEAT1 regulates MPP+-induced neuronal injury by targeting miR-124 in neuroblastoma cells. Neuroscience Letters. 2019;708, article 134340 doi: 10.1016/j.neulet.2019.134340. [DOI] [PubMed] [Google Scholar]
- 53.Kawajiri S., Saiki S., Sato S., et al. PINK1 is recruited to mitochondria with parkin and associates with LC3 in mitophagy. FEBS Letters. 2010;584(6):1073–1079. doi: 10.1016/j.febslet.2010.02.016. [DOI] [PubMed] [Google Scholar]
- 54.Kraus T. F. J., Haider M., Spanner J., Steinmaurer M., Dietinger V., Kretzschmar H. A. Altered long noncoding RNA expression precedes the course of Parkinson's disease-a preliminary report. Molecular Neurobiology. 2017;54(4):2869–2877. doi: 10.1007/s12035-016-9854-x. [DOI] [PubMed] [Google Scholar]
- 55.Chen Y., Lian Y.-j., Ma Y.-q., Wu C.-j., Zheng Y.-k., Xie N.-c. LncRNA SNHG1 promotes α-synuclein aggregation and toxicity by targeting miR-15b-5p to activate SIAH1 in human neuroblastoma SH-SY5Y cells. Neurotoxicology. 2018;68:212–221. doi: 10.1016/j.neuro.2017.12.001. [DOI] [PubMed] [Google Scholar]
- 56.Qian C., Ye Y., Mao H., et al. Downregulated lncRNA-SNHG1 enhances autophagy and prevents cell death through the miR-221/222 /p27/mTOR pathway in Parkinson's disease. Experimental Cell Research. 2019;384(1):p. 111614. doi: 10.1016/j.yexcr.2019.111614. [DOI] [PubMed] [Google Scholar]
- 57.Wang S., Zhang X., Guo Y., Rong H., Liu T. The long noncoding RNA HOTAIR promotes Parkinson's disease by upregulating LRRK2 expression. Oncotarget. 2017;8(15):24449–24456. doi: 10.18632/oncotarget.15511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Plowey E. D., Cherra S. J., Liu Y.-J., Chu C. T. Role of autophagy in G2019S-LRRK2-associated neurite shortening in differentiated SH-SY5Y cells. Journal of Neurochemistry. 2008;105(3):1048–1056. doi: 10.1111/j.1471-4159.2008.05217.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Lin Q., Hou S., Dai Y., Jiang N., Lin Y. LncRNA HOTAIR targets miR-126-5p to promote the progression of Parkinson's disease through RAB3IP. Biological Chemistry. 2019;400(9):1217–1228. doi: 10.1515/hsz-2018-0431. [DOI] [PubMed] [Google Scholar]
- 60.Deng Z., Sheehan P., Chen S., Yue Z. Is amyotrophic lateral sclerosis/frontotemporal dementia an autophagy disease? Molecular Neurodegeneration. 2017;12(1):p. 90. doi: 10.1186/s13024-017-0232-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Expression of Concern:Long noncoding RNA HAGLROS regulates apoptosis and autophagy in Parkinson's disease via regulating miR-100/ATG10 axis and PI3K/Akt/mTOR pathway activation. Artificial Cells, Nano Medicine, and Biotechnology. 2020;48(1):p. 708. doi: 10.1080/21691401.2020.1741841. [DOI] [PubMed] [Google Scholar]
- 62.Wang W., Jiang B., Sun H., et al. Prevalence, incidence, and mortality of stroke in China. Circulation. 2017;135(8):759–771. doi: 10.1161/CIRCULATIONAHA.116.025250. [DOI] [PubMed] [Google Scholar]
- 63.Wu S., Wu B., Liu M., et al. Stroke in China: advances and challenges in epidemiology, prevention, and management. The Lancet Neurology. 2019;18(4):394–405. doi: 10.1016/S1474-4422(18)30500-3. [DOI] [PubMed] [Google Scholar]
- 64.Yemisci M., Caban S., Gursoy-Ozdemir Y., et al. Systemically administered brain-targeted nanoparticles transport peptides across the blood-brain barrier and provide neuroprotection. Journal of Cerebral Blood Flow and Metabolism: Official Journal of the International Society of Cerebral Blood Flow and Metabolism. 2015;35(3):469–475. doi: 10.1038/jcbfm.2014.220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Khoshnam S. E., Winlow W., Farzaneh M., Farbood Y., Moghaddam H. F. Pathogenic mechanisms following ischemic stroke. Neurological Sciences. 2017;38(7):1167–1186. doi: 10.1007/s10072-017-2938-1. [DOI] [PubMed] [Google Scholar]
- 66.Yang Z., Zhong L., Zhong S., Xian R., Yuan B. Hypoxia induces microglia autophagy and neural inflammation injury in focal cerebral ischemia model. Experimental and Molecular Pathology. 2015;98(2):219–224. doi: 10.1016/j.yexmp.2015.02.003. [DOI] [PubMed] [Google Scholar]
- 67.Carloni S., Girelli S., Scopa C., Buonocore G., Longini M., Balduini W. Activation of autophagy and Akt/CREB signaling play an equivalent role in the neuroprotective effect of rapamycin in neonatal hypoxia-ischemia. Autophagy. 2014;6(3):366–377. doi: 10.4161/auto.6.3.11261. [DOI] [PubMed] [Google Scholar]
- 68.Wu C., Yan X., Liao Y., et al. Increased perihematomal neuron autophagy and plasma thrombin-antithrombin levels in patients with intracerebral hemorrhage: an observational study. Medicine. 2019;98(39, article e17130) doi: 10.1097/MD.0000000000017130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Mehta S. L., Kim T., Vemuganti R. Long noncoding RNA FosDT promotes ischemic brain injury by interacting with REST-associated chromatin-modifying proteins. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 2015;35(50):16443–16449. doi: 10.1523/JNEUROSCI.2943-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Chi H.-C., Tsai C.-Y., Tsai M.-M., Yeh C.-T., Lin K.-H. Molecular functions and clinical impact of thyroid hormone-triggered autophagy in liver-related diseases. Journal of Biomedical Science. 2019;26(1):p. 24. doi: 10.1186/s12929-019-0517-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Wang J., Cao B., Han D., Sun M., Feng J. Long non-coding RNA H19 induces cerebral ischemia reperfusion injury via activation of autophagy. Aging and Disease. 2017;8(1):71–84. doi: 10.14336/AD.2016.0530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Zhang J., Yuan L., Zhang X., et al. Altered long non-coding RNA transcriptomic profiles in brain microvascular endothelium after cerebral ischemia. Experimental Neurology. 2016;277:162–170. doi: 10.1016/j.expneurol.2015.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Guo D., Ma J., Yan L., et al. Down-regulation of Lncrna MALAT1 attenuates neuronal cell death through suppressing Beclin1-dependent autophagy by regulating Mir-30a in cerebral ischemic stroke. Cellular Physiology and Biochemistry. 2017;43(1):182–194. doi: 10.1159/000480337. [DOI] [PubMed] [Google Scholar]
- 74.Li Z., Li J., Tang N. Long noncoding RNA Malat1 is a potent autophagy inducer protecting brain microvascular endothelial cells against oxygen-glucose deprivation/reoxygenation-induced injury by sponging miR-26b and upregulating ULK2 expression. Neuroscience. 2017;354:1–10. doi: 10.1016/j.neuroscience.2017.04.017. [DOI] [PubMed] [Google Scholar]
- 75.Wang S., Han X., Mao Z., Xin Y., Maharjan S., Zhang B. MALAT1 lncRNA induces autophagy and protects brain microvascular endothelial cells against oxygen-glucose deprivation by binding to miR-200c-3p and upregulating SIRT1 expression. Neuroscience. 2019;397:116–126. doi: 10.1016/j.neuroscience.2018.11.024. [DOI] [PubMed] [Google Scholar]
- 76.Yao X., Yao R., Huang F., Yi J. LncRNA SNHG12 as a potent autophagy inducer exerts neuroprotective effects against cerebral ischemia/reperfusion injury. Biochemical and Biophysical Research Communications. 2019;514(2):490–496. doi: 10.1016/j.bbrc.2019.04.158. [DOI] [PubMed] [Google Scholar]
- 77.Vausort M., Wagner D. R., Devaux Y. Long noncoding RNAs in patients with acute myocardial infarction. Circulation Research. 2014;115(7):668–677. doi: 10.1161/CIRCRESAHA.115.303836. [DOI] [PubMed] [Google Scholar]
- 78.Yu S., Yu M., He X., Wen L., Bu Z., Feng J. KCNQ1OT1 promotes autophagy by regulating miR-200a/FOXO3/ATG7 pathway in cerebral ischemic stroke. Aging Cell. 2019;18(3, article e12940) doi: 10.1111/acel.12940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Meng F., You Y., Liu Z., Liu J., Ding H., Xu R. Neuronal calcium signaling pathways are associated with the development of epilepsy. Molecular Medicine Reports. 2015;11(1):196–202. doi: 10.3892/mmr.2014.2756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Huang Y., Liu X., Liao Y., et al. MiR-181a influences the cognitive function of epileptic rats induced by pentylenetetrazol. International Journal of Clinical and Experimental Pathology. 2015;8(10):12861–12868. [PMC free article] [PubMed] [Google Scholar]
- 81.Wu Q., Yi X. Down-regulation of long noncoding RNA MALAT1 protects hippocampal neurons against excessive autophagy and apoptosis via the PI3K/Akt signaling pathway in rats with epilepsy. Journal of Molecular Neuroscience: MN. 2018;65(2):234–245. doi: 10.1007/s12031-018-1093-3. [DOI] [PubMed] [Google Scholar]
- 82.Song X., Zhang N., Han P., et al. Circular RNA profile in gliomas revealed by identification tool UROBORUS. Nucleic Acids Research. 2016;44(9):p. e87. doi: 10.1093/nar/gkw075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Jain R. K., di Tomaso E., Duda D. G., Loeffler J. S., Sorensen A. G., Batchelor T. T. Angiogenesis in brain tumours. Nature Reviews Neuroscience. 2007;8(8):610–622. doi: 10.1038/nrn2175. [DOI] [PubMed] [Google Scholar]
- 84.Kelland L. The resurgence of platinum-based cancer chemotherapy. Nature Reviews Cancer. 2007;7(8):573–584. doi: 10.1038/nrc2167. [DOI] [PubMed] [Google Scholar]
- 85.Li P., He J., Yang Z., et al. ZNNT1long noncoding RNA induces autophagy to inhibit tumorigenesis of uveal melanoma by regulating key autophagy gene expression. Autophagy. 2020;16(7):1186–1199. doi: 10.1080/15548627.2019.1659614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Zhang N., Li Z., Bai F., Zhang S. PAX5-induced upregulation of IDH1-AS1 promotes tumor growth in prostate cancer by regulating ATG5-mediated autophagy. Cell Death & Disease. 2019;10(10):p. 734. doi: 10.1038/s41419-019-1932-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Zhang F., Yan T., Guo W., et al. Novel oncogene COPS3 interacts with Beclin1 and Raf-1 to regulate metastasis of osteosarcoma through autophagy. Journal of Experimental & Clinical Cancer Research. 2018;37(1):p. 135. doi: 10.1186/s13046-018-0791-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Zhao H., Wang X., Feng X., et al. Long non-coding RNA MEG3 regulates proliferation, apoptosis, and autophagy and is associated with prognosis in glioma. Journal of Neuro-Oncology. 2018;140(2):281–288. doi: 10.1007/s11060-018-2874-9. [DOI] [PubMed] [Google Scholar]
- 89.Qin N., Tong G.-F., Sun L.-W., Xu X.-L. Long noncoding RNA MEG3 suppresses glioma cell proliferation, migration, and invasion by acting as a competing endogenous RNA of miR-19a. Oncology Research. 2017;25(9):1471–1478. doi: 10.3727/096504017X14886689179993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Jiang L., Wang C., Lei F., et al. miR-93 promotes cell proliferation in gliomas through activation of PI3K/Akt signaling pathway. Oncotarget. 2015;6(10):8286–8299. doi: 10.18632/oncotarget.3221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Shi H., Ji Y., Zhang D., Liu Y., Fang P. MicroRNA-3666-induced suppression of SIRT7 inhibits the growth of non-small cell lung cancer cells. Oncology Reports. 2016;36(5):3051–3057. doi: 10.3892/or.2016.5063. [DOI] [PubMed] [Google Scholar]
- 92.Xu D.‐. H., Chi G.‐. N., Zhao C.‐. H., Li D.‐. Y. Long noncoding RNA MEG3 inhibits proliferation and migration but induces autophagy by regulation of Sirt7 and PI3K/AKT/mTOR pathway in glioma cells. Journal of Cellular Biochemistry. 2018;120(5):7516–7526. doi: 10.1002/jcb.28026. [DOI] [PubMed] [Google Scholar]
- 93.Ma B., Gao Z., Lou J., et al. Long non-coding RNA MEG3 contributes to cisplatin-induced apoptosis via inhibition of autophagy in human glioma cells. Molecular Medicine Reports. 2017;16(3):2946–2952. doi: 10.3892/mmr.2017.6897. [DOI] [PubMed] [Google Scholar]
- 94.Ma Y., Wang P., Xue Y., et al. PVT1 affects growth of glioma microvascular endothelial cells by negatively regulating miR-186. Tumour Biology. 2017;39(3) doi: 10.1177/1010428317694326. [DOI] [PubMed] [Google Scholar]
- 95.Ma K.-x., Wang H.-j., Li X.-r., et al. Long noncoding RNA MALAT1 associates with the malignant status and poor prognosis in glioma. Tumour Biology. 2015;36(5):3355–3359. doi: 10.1007/s13277-014-2969-7. [DOI] [PubMed] [Google Scholar]
- 96.Fu Z., Luo W., Wang J., et al. Malat1 activates autophagy and promotes cell proliferation by sponging miR-101 and upregulating STMN1, RAB5A and ATG4D expression in glioma. Biochemical and Biophysical Research Communications. 2017;492(3):480–486. doi: 10.1016/j.bbrc.2017.08.070. [DOI] [PubMed] [Google Scholar]
- 97.Lu Y., Dong S., Hao B., et al. Vacuolin-1 potently and reversibly inhibits autophagosome-lysosome fusion by activating RAB5A. Autophagy. 2014;10(11):1895–1905. doi: 10.4161/auto.32200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Abreu S., Kriegenburg F., Gómez‐Sánchez R., et al. Conserved Atg8 recognition sites mediate Atg4 association with autophagosomal membranes and Atg8 deconjugation. EMBO Reports. 2017;18(5):765–780. doi: 10.15252/embr.201643146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Mo H., He J., Yuan Z., et al. PLK1 contributes to autophagy by regulating MYC stabilization in osteosarcoma cells. OncoTargets and Therapy. 2019;12:7527–7536. doi: 10.2147/OTT.S210575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Ma R., Zhang B.-W., Zhang Z.-B., Deng Q.-J. LncRNA MALAT1 knockdown inhibits cell migration and invasion by suppressing autophagy through miR-384/GOLM1 axis in glioma. European Review for Medical and Pharmacological Sciences. 2020;24(5):2601–2615. doi: 10.26355/eurrev_202003_20529. [DOI] [PubMed] [Google Scholar]
- 101.Xu R., Ji J., Zhang X., et al. PDGFA/PDGFRα-regulated GOLM1 promotes human glioma progression through activation of AKT. Journal of Experimental & Clinical Cancer Research. 2017;36(1):p. 193. doi: 10.1186/s13046-017-0665-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Shen J., Hodges T. R., Song R., et al. Serum HOTAIR and GAS5 levels as predictors of survival in patients with glioblastoma. Molecular Carcinogenesis. 2018;57(1):137–141. doi: 10.1002/mc.22739. [DOI] [PubMed] [Google Scholar]
- 103.Huo J.‐. F., Chen X.‐. B. Long noncoding RNA growth arrest-specific 5 facilitates glioma cell sensitivity to cisplatin by suppressing excessive autophagy in an mTOR-dependent manner. Journal of Cellular Biochemistry. 2019;120(4):6127–6136. doi: 10.1002/jcb.27900. [DOI] [PubMed] [Google Scholar]
- 104.Scoles D., Pulst S. Oligonucleotide therapeutics in neurodegenerative diseases. RNA Biology. 2018;15(6):707–714. doi: 10.1080/15476286.2018.1454812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Doxakis E. Therapeutic Antisense Oligonucleotides for Movement Disorders. Medicinal Research Reviews. 2020 doi: 10.1002/med.21706. [DOI] [PubMed] [Google Scholar]
- 106.de Bruin D., Bossert N., Aartsma-Rus A., Bouwmeester D. Measuring DNA hybridization using fluorescent DNA-stabilized silver clusters to investigate mismatch effects on therapeutic oligonucleotides. Journal of Nanobiotechnology. 2018;16(1):p. 37. doi: 10.1186/s12951-018-0361-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Dash D., Mestre T. A. Therapeutic update on Huntington's disease: symptomatic treatments and emerging disease-modifying therapies. Neurotherapeutics. 2020:1–15. doi: 10.1007/s13311-020-00891-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
No data were used to support this study.