

ABSTRACT
Ras proteins must localize to the plasma membrane (PM) for biological function. The membrane anchor of the K-Ras4B isoform comprises a farnesylated and methylated C-terminal cysteine together with an adjacent hexa-lysine polybasic domain (PBD). Traditionally, polybasic sequences have been thought to interact electrostatically with negatively charged membranes showing no specificity for anionic lipid head groups. By contrast we recently showed that the K-Ras membrane anchor actually exhibits a very high degree of specificity for phosphatidylserine (PtdSer). The selectivity for PtdSer is determined by a combinatorial code comprising the PBD sequence plus the prenyl anchor. Lipid binding specificity is therefore altered by PBD point mutations that in turn modulate signaling output. For example, mutating Lys177 or Lys178 to glutamine switches K-Ras4B lipid affinity from PtdSer to phosphoinositol 4,5-bisphosphate (PIP2). Changing the lipid anchor from farnesyl to geranylgeranyl or the PBD lysines to arginines also changes lipid binding specificity. All-atom molecular dynamics simulations reveal the structural basis for these K-Ras anchor lipid-binding preferences. Here we examine the PM interactions of a series of geranylgeranylated PBD mutants and provide further evidence that the precise PBD sequence and prenyl lipid determines lipid sorting specificity of the K-Ras anchor and hence biological function.
KEYWORDS: electron microscopy, K-Ras, lipid-anchored proteins, phosphatidylserine, plasma membrane
The Ras isoforms, H-Ras, N-Ras and K-Ras splice variants K-Ras4A and K-Ras4B regulate cell growth, survival, proliferation and migration and are frequently mutated in human tumors [1,2]. These Ras proteins have highly conserved G-domains (amino acids 1–165) that bind guanine nucleotides and interact with a common set of exchange factors and downstream effectors, but exhibit substantial sequence variation in their C-terminal hypervariable regions (HVRs, amino acids 166–188/189) [3,4]. The HVR region includes a C-terminal bi-partite membrane anchor. The membrane anchor comprises a C-terminal farnesyl-methyl ester, common to all Ras isoforms, that operates with distinct “second signals”. The second signal is dual palmitoyl chains on cysteines (Cys) 181 and Cys184 for H-Ras, a single additional palmitoyl chain on Cys181 for N-Ras, a contiguous hexa-lysine polybasic domain (PBD, Lys175-180) in K-Ras4B, or a single palmitoyl chain on Cys180 along with two short polybasic sequences RLKK (residues 167–171) and KIKK (residues 182–185) in K-Ras4A [5–8]. Ras anchors undergo complex interactions with plasma membrane (PM) lipids to drive the formation of nanometer-scaled domains, termed nanoclusters [9–12]. These nanoclusters possess distinct lipid compositions and are main sites for downstream effector binding and signal transmission [13–15]. Thus, lipid selectivity contributes significantly to Ras signal transduction and isoform specificity.
The PBD of K-Ras4B (hereafter referred to simply as K-Ras) has been traditionally thought to only sense electrical charge nonspecifically. Indeed, in in vitro assays, polybasic sequences without a lipid anchor associate with multivalent phosphoinositol 4,5-bisphosphate (PIP2) and phosphoinositol 3,4,5-trisphosphate (PIP3) more efficiently than monovalent anionic lipids [16,17]. However, farnesylated K-Ras in intact cells sorts monovalent phosphatidylserine (PtdSer) but not more highly charged PIP2 into nanoclusters [13–15,18–20]. This suggests that, aside from charges, other structural factors must contribute to K-Ras lipid sorting on the PM. This hypothesis was recently examined in some detail [15]. First, in cells acutely depleted of endogenous PtdSer and supplemented with synthetic PtdSer with different acyl chains it was found that K-Ras PM localization can be restored by any species of PtdSer with at least one desaturated acyl chain, but not by fully saturated di18:0 PtdSer. Strikingly, the lateral spatial distribution of K-Ras is only sensitive to asymmetric PtdSer (16:0/18:1 PtdSer or 18:0/18:1 PtdSer). Co-localization of K-Ras with its downstream effector CRAF is also only sensitive to asymmetric PtdSer. Taken together, these data suggest that K-Ras PBD senses distinct lipid acyl chain structures in addition to charged headgroups. To further evaluate K-Ras PBD, we took the constitutively active oncogenic mutant K-Ras.G12V and generated 6 PBD single-point mutants, each with a single lysine mutated to glutamine: K-RasG12V-K175Q, K-RasG12V-K176Q, K-RasG12V-K177Q, K-RasG12V-K178Q, K-RasG12V-K179Q and K-RasG12V-K180Q. Each PBD mutant still possesses same amount of charge (+5) and should therefore be expected to have similar electrostatic interactions with the PM. However, bivariate lipid mapping analysis demonstrates that each K-Ras PBD mutant has distinct lipid composition within its nanoclusters, suggesting that each lysine potentially has different roles in sorting PM lipids (Figure. 1) [15]. Further, a geranylgeranyl K-RasG12V shows enhanced association with almost all lipids tested (PtdSer, PIP2, PIP3 and cholesterol, Figure. 1) [15], suggesting that switching the prenyl anchor to a longer C20 chain enhances the global association of K-Ras with cell PM but diminishes its ability to differentiate among various PM lipid species.
Figure 1.
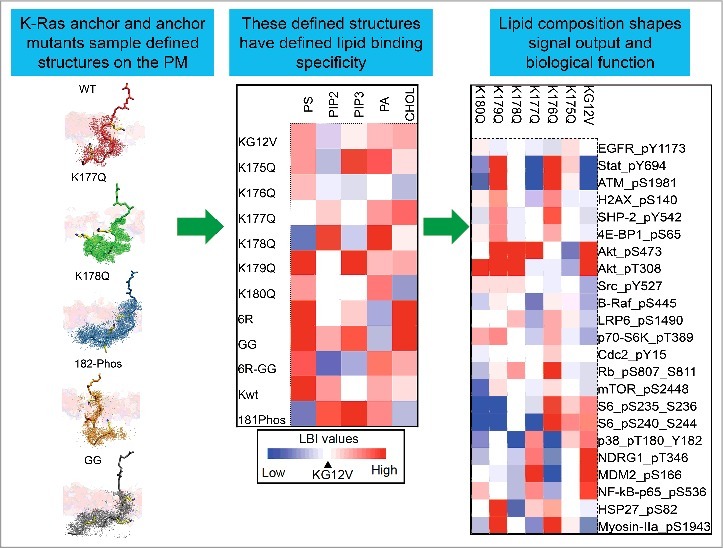
K-Ras membrane anchoring domain encodes a highly selective capacity for lipid sorting. K-Ras membrane anchor is composed of a hexa-lysine polybasic domain and an adjacent farnesyl methyl ester at the C-terminal end. The prenylated K-Ras PBD is more than just a charge sensor. The precise PBD amino acid sequence and the structure of the prenyl group combine to allow K-Ras anchor to sample well-defined conformational states, which results in distinct interactions with different lipid headgroups and acyl chains. Aggregation of a set of like anchors leads to assembly of nanoclusters with defined lipid compositions (shown as a heat map). The lipid composition of the resulting nanocluster shapes signal output (shown as a heat map of signaling pathway activity) [15].
Of particular interest is K177Q and K178Q PBD mutants, which switch their lipid preference from PtdSer to PIP2 [15]. GFP-K-RasG12V-K177Q and GFP-K-RasG12V-K178Q mutants mislocalize away from the PM whereas other PBD muatnts maintain similar PM affinity as K-RasG12V with the original hexa-lysine PBD, which is further validated in our confocal imaging [15]. Our univariate EM-spatial mapping and FLIM-FRET analyses further show that nanclustering of GFP-K-RasG12V-K177Q or GFP-K-RasG12V-K178Q is markedly disrupted on the PM, while other PBD mutants display similar extent of clustering as K-RasG12V with the original hexa-lysine PBD (Figure. 2). This is most likely because the PIP2 level in cell PM is much lower than that of PtdSer. Indeed, supplementation of exogenous PIP2 effectively rescues PM localization and nanoclustering of K177Q and K178Q mutants, but has no effect on K-RasG12V with the original hexa-lysine PBD. The switch of lipid specificity from PtdSer to PIP2 in K177Q and K178Q mutants is further validated in the same set of exogenous lipid supplementation experiments, where exogenous PtdSer markedly enhances PM localization and nanoclustering of GFP-K-RasG12V (with original hexa-lysine PBD), but has no effect on the K177Q or K178Q mutants [15]. Reverse phospho-protein arrays showed that BHK cells expressing individual PBD mutants display distinct kinase activities (Figure. 1), consistent with lipid sorting being biologically important [15].
Figure 2.
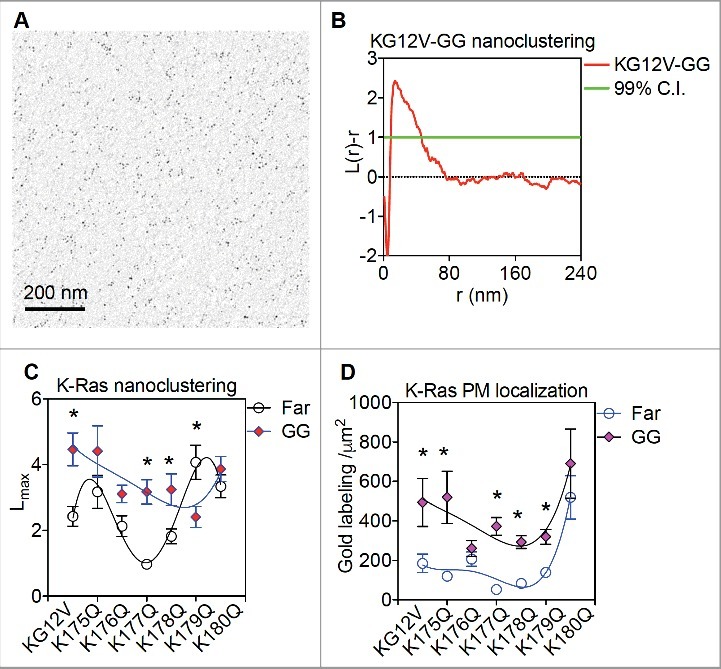
Geranylgeranyl anchor modifies PBD interactions of the K-Ras anchor (A) Intact PM sheets of BHK cells expressing GFP-tagged farnesylated (Far) or geranylgeranylated (GG) K-RasG12V PBD mutants were attached to copper EM grids, fixed using 4% paraformaldehyde (PFA) and 0.1% glutaraldehyde and immunolabeled with 4.5nm gold nanoparticles coupled to anti-GFP antibody. Gold labels were then imaged using transmission EM at 100,000X magnification and digitized using ImageJ. An intact 1μm2 PM sheet of a BHK cell expressing GFP-K-RasG12V-GG immunolabeled with 4.5nm gold conjugated to anti-GFP antibody is shown. (B) Univariate K-function was used to quantify the spatial distribution of gold particles on intact BHK PM sheets. Extent of nanoclustering, L(r)-r, was plotted against length scale, r. L(r)-r values above the 99% confidence interval (99%C.I.) of 1 (green line) indicate statistically meaningful clustering. (C) Peak values of the K-function curves shown in (B) were termed as Lmax and used to summarize the spatial data. (D) Number of the gold particles within a PM area of 1μm2 was counted and used as an estimate of K-Ras PM localization. At least 15 images were analyzed in each condition with mean ± SEM shown. Bootstrap tests were used to evaluate the clustering statistical significance between GFP-K-RasG12V-Far and GFP-K-RasG12V-GG PBD mutants (* indicates p <0.05). One-way ANOVA was used to evaluate the gold labeling data between GFP-K-RasG12V-Far and GFP-K-RasG12V-GG PBD mutants (* indicates p < 0.05) Spatial data of farnesylated K-Ras PBD mutants were previously shown [15].
Atomistic molecular dynamic (aMD) and metaMD simulations using the minimal anchoring domain of K-Ras (tK) revealed the molecular mechanism driving the PBD specificity for lipid sorting. Unlike previously thought, the prenyl-anchored hexa-lysine tK polybasic backbone adopts distinct conformational states with clearly distinguishable energy barriers: disordered (D), intermediate (I) or ordered (O) (Figure. 1) [15]. Wild-type tK with an intact hexa-lysine PBD and a C-terminal farnesyl anchor prefers the disordered conformation, which possesses higher ability to form H-bonds with PtdSer. Conversely, tK-K177Q and tK-K178Q adopt the ordered conformation with fewer tendencies to H-bond with PtdSer. This potentially drives the dissociation of K177Q and K178Q mutants from PtdSer. When the farnesyl anchor is switched to the longer geranylgeranyl (GG) anchor, tK-GG peptide backbone prefers the ordered conformational state and interacts with lipid bilayer core and acyl chains extensively (Figure. 1). This is indeed consistent with our lipid mapping analysis, showing that geranylgeranylated K-RasG12V enhances its association with almost all tested lipids (Figure. 1) [15]. These data suggest that geranylgeranylated K-Ras potentially loses its ability to specifically sort lipid headgroups.
Here, we show new data supporting our view that the prenyl anchor contributes to K-Ras PBD specific lipid sorting. We mutated the CAAX motif of the 6 K-RasG12V PBD mutants used in our recent study from CVIM to CCIL and so generated 6 geranylgeranylated K-RasG12V PBD mutants: GFP-K-RasG12V-K175Q-GG, GFP-K-RasG12V-176Q-GG, GFP-K-RasG12V-K177Q-GG, GFP-K-RasG12V-K178Q-GG, GFP-K-RasG12V-K179Q-GG and GFP-K-RasG12V-K180Q-GG. We then conducted univariate spatial analysis of these geranylgeranylated K-Ras PBD mutants on the inner PM leaflet of BHK cells (Figure. 2A and B). All geranylgeranylated PBD mutants displayed elevated PM localization when compared with their farnesylated cognates (Figure. 2D). This is consistent with the aMD / metaMD simulations and lipid mapping analysis, where geranylgeranylation enhances K-Ras PBD interactions with the membrane hydrophobic core. More interestingly, all geranylgeranylated PBD mutants showed a similar extent of nanoclustering (Figure. 2C). This is very different from their farnesylated counterparts, where K177Q and K178Q cluster less efficiently on the PM (Figure. 2C). These data suggest that geranylgeranylation markedly changes how the K-Ras PBD associates with PM lipids. These new data further support the view that the prenyl group plays a key role in modulating K-Ras PBD conformational orientations on the PM and in turn determines how the complete anchor sorts specific PM lipids.
K-Ras lipid binding selectivity determines the specificity with which downstream effectors are recruited and hence the activation profile of downstream signaling cascades [15]. Extrapolating from these findings we speculate that geranylgeranylated K-Ras by virtue of a different lipid binding selectivity will generate a different signaling output compared to farnesylated K-Ras. Thus, although geranylgeranylated K-Ras is capable of driving cell transformation [6], the molecular signaling mechanisms may differ from that of the farnesylated K-Ras. The geranylgeranyl modified hexa-lysine anchor showed strong binding to all anionic phospholipids indicating a loss of selectivity for lipid head groups; this was confirmed in the MD structures which showed reduced interactions of the lysine side chains with PtdSer and PC headgroups, but enhanced van der Waals interactions of the geranylgeranyl with lipid side chains in the hydrophobic core of the membrane. Lipid binding specificity was restored when the six lysines were replaced with six ariginines. It is tempting to speculate that the guanidinium groups of arginine allow for more extensive anionic lipid head group interactions than the protonated α-amino group of lysine in the context of a geranygeranyl lipid chain [15]. It is interesting to note in this context that other small GTPases that are known to be geranylgeranylated all contain arginines in their PBD; Rac1 for example has a mixed PBD composed of KKRKRK (amino acids 183–188) with a geranylgeranyl chain attached to an adjacent Cys189. Thus, although traditionally basic residues have been assumed to be interchangeable, these findings show that the exact sequence of the basic amino acids in a PBD in combination with the structure of the prenyl anchor actual encodes complex membrane binding, lipid sorting and signal transmission information.
Acknowledgment
This work is partially supported by the Cancer Research and Prevention Institute of Texas (CPRIT: RP170233) to JFH and the National Institutes of Health (NIH: P30 DK056338) to YZ.
Funding
National institute of health ID: DK056338 Cancer Prevention and Research Institute of Texas (CPRIT) ID: RP170233
References
- [1].Prior, IA., Hancock, JF.. Ras trafficking, localization and compartmentalized signalling. Semin Cell Dev Biol. 2012;23(2):p. 145–53. https://doi.org/ 10.1016/j.semcdb.2011.09.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Downward, J. Targeting RAS signalling pathways in cancer therapy. Nat Rev Cancer. 2003;3(1):p. 11–22. https://doi.org/ 10.1038/nrc969 [DOI] [PubMed] [Google Scholar]
- [3].Prior, IA., Lewis, PD., Mattos, C.. A comprehensive survey of ras mutations in cancer. Cancer Res. 2012;72(10):p. 2457–67. https://doi.org/ 10.1158/0008-5472.CAN-11-2612 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Prior, IA., Hancock, JF.. Compartmentalization of Ras proteins. J Cell Sci. 2001;114(Pt 9):p. 1603–8. [DOI] [PubMed] [Google Scholar]
- [5].Hancock, JF., et al., All ras proteins are polyisoprenylated but only some are palmitoylated. Cell. 1989;57(7):p. 1167–77. https://doi.org/ 10.1016/0092-8674(89)90054-8 [DOI] [PubMed] [Google Scholar]
- [6].Hancock, JF., et al., A CAAX or a CAAL motif and a second signal are sufficient for plasma membrane targeting of ras proteins. EMBO J. 1991;10(13):p. 4033–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Hancock, JF., Paterson, H., Marshall, CJ.. A polybasic domain or palmitoylation is required in addition to the CAAX motif to localize p21ras to the plasma membrane. Cell. 1990;63(1):p. 133–9. https://doi.org/ 10.1016/0092-8674(90)90294-O [DOI] [PubMed] [Google Scholar]
- [8].Tsai, FD., et al., K-Ras4A splice variant is widely expressed in cancer and uses a hybrid membrane-targeting motif. Proc Natl Acad Sci U S A. 2015;112(3):p. 779–84. https://doi.org/ 10.1073/pnas.1412811112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Prior, IA., et al., Direct visualization of Ras proteins in spatially distinct cell surface microdomains. J Cell Biol, 2003. 160: p. 165–170. https://doi.org/ 10.1083/jcb.200209091 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Hancock, JF., Ras proteins: different signals from different locations. Nat Rev Mol Cell Biol. 2003;4(5):p. 373–84. https://doi.org/ 10.1038/nrm1105 [DOI] [PubMed] [Google Scholar]
- [11].Plowman, SJ., et al., H-ras, K-ras, and inner plasma membrane raft proteins operate in nanoclusters with differential dependence on the actin cytoskeleton. Proc Natl Acad Sci U S A. 2005;102(43):p. 15500–5. https://doi.org/ 10.1073/pnas.0504114102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Zhou, Y., Hancock, JF., Ras nanoclusters: Versatile lipid-based signaling platforms. Biochim Biophys Acta. 2015;1853(4):p. 841–849. https://doi.org/ 10.1016/j.bbamcr.2014.09.008 [DOI] [PubMed] [Google Scholar]
- [13].Zhou, Y., et al., Signal Integration by Lipid-Mediated Spatial Cross Talk between Ras Nanoclusters. Mol Cell Biol. 2014;34(5):p. 862–76. https://doi.org/ 10.1128/MCB.01227-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Zhou, Y., et al., SIGNAL TRANSDUCTION. Membrane potential modulates plasma membrane phospholipid dynamics and K-Ras signaling. Science. 2015;349(6250):p. 873–6. https://doi.org/ 10.1126/science.aaa5619 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Zhou, Y., et al., Lipid-Sorting Specificity Encoded in K-Ras Membrane Anchor Regulates Signal Output. Cell. 2017;168(1-2):p. 239–251. e16. https://doi.org/ 10.1016/j.cell.2016.11.059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Gambhir, A., et al., Electrostatic sequestration of PIP2 on phospholipid membranes by basic/aromatic regions of proteins. Biophys J. 2004;86(4):p. 2188–207. https://doi.org/ 10.1016/S0006-3495(04)74278-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].McLaughlin, S., et al., PIP(2) and proteins: interactions, organization, and information flow. Annu Rev Biophys Biomol Struct. 2002;31:p. 151–75. https://doi.org/ 10.1146/annurev.biophys.31.082901.134259 [DOI] [PubMed] [Google Scholar]
- [18].Cho, KJ., et al., Staurosporines disrupt phosphatidylserine trafficking and mislocalize Ras proteins. J Biol Chem, 2012;287(52):p. 43573–84. https://doi.org/ 10.1074/jbc.M112.424457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Cho, KJ., et al., Inhibition of Acid Sphingomyelinase Depletes Cellular Phosphatidylserine and Mislocalizes K-Ras from the Plasma Membrane. Mol Cell Biol, 2015. 36(2):p. 363–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].van der Hoeven, D., et al., Fendiline inhibits K-Ras plasma membrane localization and blocks K-Ras signal transmission. Mol Cell Biol, 2013. 33(2): p. 237–51. https://doi.org/ 10.1128/MCB.00884-12 [DOI] [PMC free article] [PubMed] [Google Scholar]