

Abstract
Alcoholic and non-alcoholic liver diseases begin from an imbalance in lipid metabolism in hepatocytes as the earliest response. Both liver diseases share common disease features and stages (i.e., steatosis, hepatitis, cirrhosis, and hepatocellular carcinoma). However, the two diseases have differential pathogenesis and clinical symptoms. Studies have elucidated the molecular basis underlying similarities and differences in the pathogenesis of the diseases; the factors contributing to the progression of liver diseases include depletion of sulfhydryl pools, enhanced levels of reactive oxygen and nitrogen intermediates, increased sensitivity of hepatocytes to toxic cytokines, mitochondrial dysfunction, and insulin resistance. Endoplasmic reticulum (ER) stress, which is caused by the accumulation of misfolded proteins and calcium depletion, contributes to the pathogenesis, often causing catastrophic cell death. Several studies have demonstrated a mechanism by which ER stress triggers liver disease progression. Autophagy is an evolutionarily conserved process that regulates organelle turnover and cellular energy balance through decomposing damaged organelles including mitochondria, misfolded proteins, and lipid droplets. Autophagy dysregulation also exacerbates liver diseases. Thus, autophagy-related molecules can be potential therapeutic targets for liver diseases. Since ER stress and autophagy are closely linked to each other, an understanding of the molecules, gene clusters, and networks engaged in these processes would be of help to find new remedies for alcoholic and non-alcoholic liver diseases. In this review, we summarize the recent findings and perspectives in the context of the molecular pathogenesis of the liver diseases.
Keywords: Non-alcoholic steatohepatitis, Non-alcoholic fatty liver disease, Mitochondria, Endoplasmic reticulum stress, Autophagy
INTRODUCTION
Excessive and chronic alcohol consumption may lead to the progression of alcoholic liver disease (ALD) [1]. ALD is caused by chronic alcohol consumption (>20 g/day for women and >30 g/day for men) [2,3]. Moreover, repeated alcohol drinking may promote progression of simple steatosis to steatohepatitis and/or cirrhosis [4]. Another most common liver disease is non-alcoholic fatty liver disease (NAFLD), which is considered as the hepatic manifestation of the metabolic syndrome including hypertension, type 2 diabetes, insulin-resistance, obesity, dyslipidemia, and distinct hepatic histological features [5]. Both ALD and NAFLD share general histopathological spectrum from simple steatosis to steatohepatitis and fibrosis, which may progress to more severe diseases (i.e., cirrhosis and hepatocellular carcinoma) (Fig. 1) [4]. However, the two diseases differ from each other in a variety of properties, ranging from the molecular mechanisms of disease exacerbation to differences in clinical features. In particular, infiltration of inflammatory cells occurs to a greater degree in ALD than in NAFLD. In contrast, fat degeneration in hepatocytes is more pronounced in NAFLD than in ALD [6]. Despite the ongoing study of the pathology, causes, and risk factors for the diseases, we do not yet have an appropriate treatment regimen. Here, we aim to motivate intensive research on the diseases by reviewing the current understanding of the causes of ALD and NAFLD, and the trends in potential therapeutic approaches.
Figure 1.
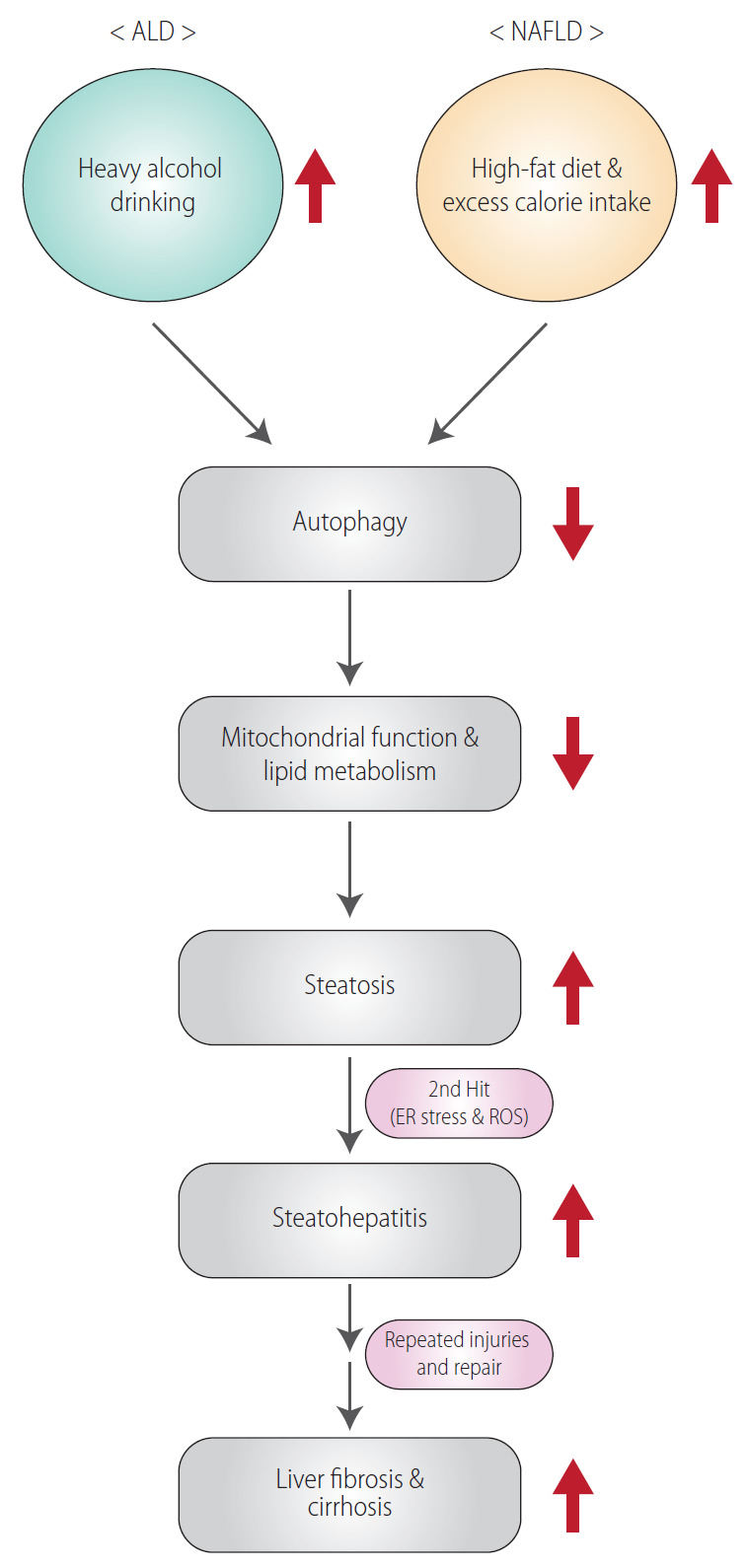
General pathological processes for ALD and NAFLD. ALD, alcoholic liver disease; NAFLD, non-alcoholic fatty liver disease; ER, endoplasmic reticulum; ROS, reactive oxygen species.
GENERAL MOLECULAR PATHOGENESIS
Definition of endoplasmic reticulum (ER) stress
ER is a structure of a membrane component that extends from the nuclear membrane and has two types: a ribosome attached rough ER and a ribosome-free smooth ER. Approximately one-third of the protein in the cell is translated from the messenger RNA to protein, which becomes an active protein structure through processes such as folding, assembly, glycation, and disulfide bond [7]. Smooth ER is the site for the synthesis of lipids and sterols, and functions as a calcium reservoir to regulate the cellular calcium concentrations [8,9]. However, if either immature protein flows into the ER above the capacity, or calcium is depleted in the ER, the function of ER is impaired. This condition is defined as ER stress [9-11]. When ER stress occurs, cells have a defense mechanism to survive, which is called the ER stress response [8]. Thus, ER stress is a cellular stress, in which the levels of synthetic proteins exceed the capacity of unfolded protein responses. Finally, when the ER stress becomes so severe that it cannot be overcome and the ER cannot recover its function, then cell death pathway is activated with inflammatory responses [9,12].
Cross-links between ER and mitochondria
In several cell and animal models, we found ER stress as a key stimulus of fatty liver transition to non-alcoholic steatohepatitis (NASH). So, ER stress is claimed as a secondary hit to provoke hepatocyte injury and inflammation, leading to liver fibrosis in a chronic situation (Fig. 1). Dysregulation of unfolded protein responses enhances ER stress and consequently may promote catastrophic cell death [13]. Additionally, ER stress, in association with reactive oxygen species (ROS) production, triggers the subsequent activation of cell death pathway due to imbalance of redox homeostasis [14,15]. Cytochrome P450 2E1, a high affinity enzyme for ethanol metabolism, is located in the membrane of ER, and thus the induction of cytochrome P450 2E1 would be also associated with ER stress in hepatocytes inflicted by sustained ethanol consumption [16]. This idea would be also supported by impairment of free fatty acids (FFAs) oxidation in mitochondria because mitochondria and ER physiologically work together through mitochondria-associated ER membrane [17].
The lipids are mainly metabolized in the liver, and lipid metabolism in the liver maintains homeostasis by balancing or storing fat for energy [18]. In particular, lipid absorption, esterification, oxidation and secretion of fatty acids are performed in liver cells [19]. The supply of FFAs to the liver contributes to the pathogenesis of hepatic steatosis; triglycerides and cholesterol are stored in the form of lipid droplets to protect cells from exposure to excessive amounts of FFAs that can damage signal pathways and metabolic homeostasis [20]. Impairment of oxidative metabolism of FFAs is also coupled to fat accumulation because FFAs are used as fuel for mitochondrial oxidation.
Compared to normal individuals, the incidence of ALD increases in patients with fatty liver by 2–3 times [21]. Thus, obesity is considered as an independent risk factor for alcoholic steatohepatitis since diet pattern and dietary fat content contribute to the progression of ALD. Mitochondrial dysfunction occurs not only by peroxidation of unsaturated fats of mitochondrial phospholipids, but by the attack of oxygen free radicals generated through ethanol metabolism. Hence, excessive alcohol consumption causes impairment of mitochondrial function [22]. In NAFLD patients, dysregulation of fatty acid metabolism causes hepatic steatosis and hyperlipidemia in a large patient population. In addition, NAFLD as a result of obesity and diabetes facilitates inflammatory cytokines production [23]. So, metabolic profile in hepatocytes greatly alters because of fuel source and consumption rates change. Together, metabolic disturbances caused by fat along with alcohol consumption stimulate worsening of alcoholic steatohepatitis, which may lead to more severe conditions.
MITOCHONDRIAL TARGETS
Peroxisome proliferator-activated receptor alpha regulates the expression of lipid metabolism-related genes (e.g., Cyp4a1, Acbp, and Acsl) [24]. In our preliminary study, we found that mitochondrial activity and oxidative energy metabolism are controlled by core molecules including sirtuin 1, peroxisome proliferator-activated receptor-γ coactivator 1-alpha and peroxisome proliferator-activated receptor alpha (i.e., a transcription complex required for mitochondrial biogenesis) [25,26]. In this study, we have shown that an activated form of G protein subunit alpha 12 (Gα12) stabilizes sirtuin 1 via ubiquitin specific peptidase 22 induction, which depends on hypoxia-inducible factor 1-alpha. The research results showed that Gα12 regulates sirtuin 1-dependent lipid oxidation in mitochondria through hypoxia-inducible factor 1-alpha-mediated ubiquitin specific peptidase 22 induction, identifying Gα12 as a linker between cell surface signaling and mitochondrial lipid oxidation (Fig. 2).
Figure 2.

Functional molecular biomarkers for the processes of ALD and NAFLD. ALD, alcoholic liver disease; AMPKα, AMP-activated protein kinase alpha; miR, microRNA; TXNIP, thioredoxin interacting protein; LXRα, liver X receptor α; ATG4B, autophagy related 4B cysteine peptidase; Gα12, G protein subunit alpha 12; HIF-1α, hypoxia-inducible factor 1-alpha; USP22, ubiquitin specific peptidase 22; SIRT1, sirtuin 1; ER, endoplasmic reticulum; ROS, reactive oxygen species; HSCs, hepatic stellate cells; JNK, c-Jun N-terminal kinase; ATG12-5, autophagy related 12-5 conjugate; NAFLD, non-alcoholic fatty liver disease.
Liver fibrosis is manifested by repeated wound healing processes, such as an increase in matrix protein and a decrease in matrix remodeling: liver fibrosis is characterized by decreased matrix remodeling and increased production of matrix proteins, and may lead to end-stage cirrhosis [27]. Liver fibrosis is defined by concerted actions of non-parenchymal cells of the liver, particularly macrophages (including Kupffer cells), hepatic stellate cells (HSCs), and endothelial cells [28]. Accumulation of aberrant extracellular matrix is induced by a diverse population of myofibroblasts, among which HSCs play a major role [29].
AUTOPHAGY TARGETS
Definition of autophagy
Studies have reported the correlation between hepatic diseases and autophagy. Autophagy may inhibit the progression of steatosis and fatty hepatitis by preventing hepatocyte injury [30,31].
The liver is the major detoxifying and metabolic organ. Autophagy, a “recycling mechanism” in hepatocytes, is evolved to salvage key metabolites and to provide energy for sustaining anabolism [32]. Thus, autophagy is a crucial regulator of cellular homeostasis [32-34]; autophagy targets damaged cellular constituents, such as denatured proteins, accumulated fat, and mitochondria that have lost their function to lysosomes for degradation [35,36]. Amino acids, insulin, and mammalian target of rapamycin signaling pathways inhibit the autophagic pathway [36]. Additionally, fasting regulates metabolic pathways, such as gluconeogenesis, ketone body formation, and β-oxidation. Fatty acids are mainly generated by selective autophagy of lipid droplets (lipophagy), while amino acids are generated by proteolysis through autophagy and are used for gluconeogenesis [33,37]. Therefore, autophagy plays a decisive role in regulating liver physiology and balancing liver metabolism [38].
Autophagic molecules associated with liver steatosis
Chronic alcohol intake or excessive energy intake causes damage to the regulation of autophagy. When it persists, the homeostatic balance breaks and metabolic diseases develop [39,40]. Although several studies have demonstrated the correlation between autophagy and metabolic liver disease, the role of autophagy in metabolic liver disease has not been completely elucidated. Moreover, the understanding of the regulatory molecules by which the disease progression is determined is still needed.
The role of autophagy in acute and chronic ALD is still complex and controversial. It is generally known that binge drinking enhances autophagy of hepatocytes, which limits hepatic cell damage, death and fat accumulation by selectively removing excess lipid droplets and damaged mitochondria [41]. In contrast, chronic alcohol intake has been reported to decrease lysosome function and increase ubiquitinated aggregates accumulation [42]. Thus, the duration of alcohol exposure affects autophagy [43]. 5’ AMP-activated protein kinase, an enzyme that plays a role in the metabolism of fatty acids (e.g., acetyl-CoA carboxylase), is a key player of autophagy. Ethanol consumption causes inhibition of 5’ AMP-activated protein kinase, suggestive of the role of autophagy in ALD. One of the studies claims a mechanism for suppressing autophagy during chronic alcohol intake in association with decrease of 5’ AMP-activated protein kinase activity, which may increase fat production through decrease of β-oxidation [44]. This information can be employed to understand the pathogenesis of ALD.
Although the evaluation of autophagy in animal and human samples is somewhat controversial [45], it is generally agreed that autophagy is reduced in NAFLD. Hepatocellular steatosis is aggravated in mice deficient in autophagy-related genes [46,47]. Inhibition of hepatocyte-specific run domain beclin-1-interacting and cysteine-rich domain-containing protein (beclin-1 interacting negative regulator for autophagosome fusion), overexpression of adenoviral-induced autophagy-related 7 [47,48], or treatment of an autophagy activator such as rapamycin can relieve ER stress and hepatic steatosis. The opposite effect was seen with the inhibitor [49]. Moreover, autophagy participates in the basic conversion process of lipids by decomposing lipid droplets under physiological conditions. Recently, we demonstrated that liver X receptor α (LXRα) functions as a transcriptional regulator of microRNAs (miRs) [50]. In this study, gene set enrichment analysis was used to discover the phenomenon that autophagy can be inhibited by LXRα, and focusing on this, we proved the regulatory mechanism of autophagy using animal model. As a mechanistic basis, it was noted that LXRα inhibits autophagy-related 4B cysteine peptidase and rasrelated protein Rab-8B, and this effect is due to the transcriptional induction of let-7a and miR-34a by LXRα. In addition, the same mechanism works in human samples and genetic obesity-derived animal (ob/ob, db/db) samples. Furthermore, LXRα and the above identified molecules all inhibit lipid degradation and mitochondrial function, supportive of a link between autophagy and NAFLD when LXRα is activated. The outcomes may provide key information for target discovery and potential strategy to treat NASH (Fig. 2).
Autophagic molecules associated with liver fibrosis
Autophagy is reported to be involved in the progression of liver fibrosis [51]. Activation of HSCs is a major phenomenon in liver fibrosis because these cells differentiate into myofibroblasts and produce a major extracellular matrix in the liver [52]. HSCs of the quiescent phenotype store vitamin A in lipids and are activated when vitamin A is decomposed by various stimuli. Thus, HSCs can be activated by promoting autophagy. Previously, we have shown that Gα12 activation increases autophagy in HSCs, accompanying c-Jun N-terminal kinase-dependent autophagy-related 12-5 conjugation [53]. In the study, miR-16 directly inhibits de novo synthesis of Gα12 and thereby modulations of miR-16 alter autophagy (Fig. 2). Consistently, patients with severe fibrosis exhibited downregulated levels of miR-16 [53]. The activation of HSCs by apoptotic hepatocytes or by transforming growth factor-β produced from activated Kupffer cells also contributes to fibrosis [54]. In this process, sustained ER stress, cytokines, saturated FFAs, and adipokines could be also involved [54].
Potential targeting of autophagy
Currently, there are no specific treatments for non-alcohol/alcohol-related liver disease, and the main treatment is dietary control or alcohol withdrawal. Recent studies have demonstrated the importance of autophagy in metabolic diseases and suggested new treatment strategies. Preclinical studies have reported that enhancing autophagy using carbamazepine or rapamycin decreases fat accumulation and liver damage in obese and chronic ethanolfed mice [49]. Although there is a Janus face of autophagy in chronic liver disease, autophagy is primarily a hepatoprotective mechanism; autophagy is protective against the early stages of hepatocellular carcinoma and promotes liver regeneration [55]. Conversely, autophagy may be deleterious during the late phase of hepatocellular carcinoma in addition to activation of HSCs [51,56]. Thus, the cell-specific and pathology-specific regulation of autophagy should be considered for devising strategies for liver disease treatment.
CHANGES IN LIPID PROFILES
Alcohol-induced hepatic dysfunction or injury is closely related to abnormal lipid profiling [57,58]. Cumulated evidence shows that liver steatosis is linked to insulin resistance and the consequent hyperinsulinemia. In addition to steatosis, chronic alcohol consumption induces insulin resistance in the liver as indicated by the suppression of insulin-responsive genes [59,60]. Moreover, heavy alcohol consumption may lead to impairment of mitochondrial membrane depolarization; dysregulation of cellular redox state results at least in part from changes in the nicotinamide adenine dinucleotide (NAD+)/nicotinamide adenine dinucleotide hydride (NADH) ratio [61,62]. Alcohol disrupts the sirtuin 1 signaling pathway, which is involved in the regulation of mitochondrial biogenesis and oxygen consumption, and consequently promotes hepatic steatosis, injury, and inflammation [63]. In particular, mitochondrial dysfunction contributes to this change to a large extent, in which replacement of old non-functional mitochondria is disturbed due to autophagy inhibition as well as impaired proteasome function [64,65]. Electron microscopy has revealed the presence of megamitochondria in the liver of patients with ALD [66,67].
Lipotoxicity not only causes inflammation, ER stress, and ROS, but also affects the biological function of organelles, the most important of which is the mitochondria [68]. Increasing autophagy removes damaged mitochondria in a non-alcoholic fatty liver animal model, improving fatty liver [69]. Conversely, when autophagy is inhibited, severe mitochondrial damage and hepatic steatosis occur [70]. Additionally, suppression of autophagy through drug or gene regulation can reduce the number of triglycerides and lipid particles in hepatocytes [71]. As a result, a decrease in FFAs supply from lipid particles reduces mitochondrial β-oxidation, and the consequential fat accumulation in turn increases mitochondrial dysfunction [72].
Therefore, the interactions among mitochondrial activity, biogenesis, and autophagy contribute to metabolic dysregulation of hepatocytes in alcoholic and non-alcoholic liver diseases.
ALD AND DRUG CANDIDATES
Alcoholic liver steatosis is considered as a benign condition and is thus reversible to normal state under the condition of abstinence [73,74]. One of the early hallmarks of alcohol-induced liver damage caused by alcohol is the accumulation of lipid droplets in hepatocytes [75]. Clinically, most patients experience no symptoms for alcohol-induced fatty liver. However, patients with alcoholic steatohepatitis or fibrosis may present symptoms such as nausea and vomiting, which are caused by dysregulated cytokines production [76-79]. In a small proportion of chronic and heavy alcohol drinkers, ALD may lead to the development of liver cirrhosis (Fig. 1) [80-84].
Electrons generated during alcohol metabolism promote production of reactive intermediates, including ROS and reactive nitrogen species, which are catalyzed by enzymes such as alcohol dehydrogenase and cytochrome P450 2E1 [85,86]. ROS and reactive nitrogen species oxidize lipids to generate lipid peroxidation end products such as isoprostane and malondialdehyde [87-89]. Mitochondria is another organelle, from which ROS is greatly generated through oxidative fuel consumption [90]. Since an excess amount of ROS production causes depletion of reduced sulfhydryl pools (e.g., glutathione) in hepatocytes, hepatocytes may be sensitized by the stresses of toxic cytokines (e.g., tumor necrosis factor-β) and thus are more susceptible to injurious challenges. Previously, we had demonstrated that a spectrum of liver injury, including pyroptotic hepatocyte death, occurs in an alcohol abuse animal model; alcohol induces pyroptosis in hepatocytes by promoting ‘thioredoxin interacting protein and NOD-, LRR- and pyrin domain-containing protein 3’ inflammation through decreases of miR-148a and forkhead box protein O1 [91]. Attention was also paid to methionine and S-adenosylmethionine in the field of metabolic mechanism studies, the levels of which may be changed by alcohol consumption as part of the evidence of changes in cellular glutathione pool [92].
As mentioned above, various mechanisms are involved in ALD, and based on these findings, clinical treatment with new and emerging molecular targets for ALD treatment is ongoing (Table 1).
Table 1.
Ongoing clinical trials or candidate targets of pharmacotherapies for the treatment of ALD or NAFLD
| Pathology | Target | Agent | Stage | Reference |
|---|---|---|---|---|
| ALD | Hepatic regeneration | IL-22 (F-652) | Phase 1 | [100] |
| IgG antibody to LPS | Bovine colostrum (IMM-124E) | Phase 2 | [101] | |
| Probiotic, restores gut microbiome | Lactobacillus rhamnosus GG | Phase 2 | [102] | |
| Antagonist to IL-1 receptor | Anakinra | Phase 2 | [103] | |
| FXR agonism | Obeticholic acid (INT-747) | Phase 2 | [104] | |
| Antibiotic amoxicillin plus clavulanic acid | Augmentin | Phase 3 | [105] | |
| Antioxidant and promotes abstinence | Metadoxine | Phase 4 | [106] | |
| Increase neutrophils, hepatic regeneration | G-CSF (filgrastim) | Phase 4 | [107] | |
| Glutamate | - | Target discovery stage | [108] | |
| HMGB1 | - | Target discovery stage | [109] | |
| ALDH2 | - | Target discovery stage | [110] | |
| Toll-like receptor 3 | - | Target discovery stage | [111] | |
| C3aR, C5aR1 | - | Target discovery stage | [112] | |
| Fructose, cytochrome P450-2E1 | - | Target discovery stage | [113] | |
| HIF-1α | - | Target discovery stage | [114] | |
| Lactobacillus GG | - | Target discovery stage | [115] | |
| Bile acid,FXR,FGF15 | - | Target discovery stage | [116] | |
| FGF19 | - | Target discovery stage | [117] | |
| FoxO1, miR-148a, TXNIP | - | Target discovery stage | [91] | |
| REG3 lectins | - | Target discovery stage | [118] | |
| NAFLD | Thyroid hormone receptor beta agonist | VK2809 | Phase 2 | [119] |
| PanPPAR agonist | Lanafibranor | Phase 2 | [120] | |
| FXR agonist and SGLT1/2 inhibitor | Tropifexor and licoglifozin | Phase 2 | [121] | |
| Engineered version of human hormone FGF19 | Aldafermin | Phase 2 | [122] | |
| FXR agonist | EDP-305 | Phase 2 | [123] | |
| ASBT inhibitor | Volixibat | Phase 2 | [124] | |
| Recombinant FGF21 | BMS-986036 | Phase 2 | [125] | |
| Pan-caspase inhibitor | Emricasan | Phase 2 | [126] | |
| Galectin 3 inhibitor | GR-MD-02 | Phase 2 | [127] | |
| CCR2/CCR5 receptor inhibitor | Cenicriviroc (AURORA) | Phase 3 | [128] | |
| SCD1 modulator | Aramchol | Phase 3 | [129] | |
| Thyroid hormone receptor beta agonist | Resmetirom | Phase 3 | [130] | |
| PPARα/δ ligand | Elafibranor (RESOLVE-IT) | Phase 3 | [131] | |
| FXR ligand | Obeticholic acid (REGENERATE) | Phase 3 | [132] | |
| FXR agonist | Obeticholic acid (REVERSE) | Phase 3 | [132] | |
| ASK1 inhibitor | Selonsertib | Phase 3 | [133] | |
| SGLT2 inhibitor | Gliflozin | Pilot | [134] | |
| LXRα, let-7a, miR-34a, ATG4B, Rab-8B | – | Target discovery stage | [50] | |
| STAT-1, STAT-3 | – | Target discovery stage | [135] | |
| Fructokinase | – | Target discovery stage | [136] | |
| Gα13, ITIH1 | – | Target discovery stage | [137] | |
| TAZ | – | Target discovery stage | [138] | |
| TNFAIP3 | – | Target discovery stage | [139] | |
| Glutaminase 1 | – | Target discovery stage | [140] | |
| USP22, Gα12, SIRT1 | – | Target discovery stage | [26] | |
| OTULIN | – | Target discovery stage | [141] |
ALD, alcoholic liver disease; NAFLD, non-alcoholic fatty liver disease; IL, interleukin; IgG, immunoglobulin G; LPS, lipopolysaccharide; FXR, farnesoid X receptor; G-CSF, granulocyte-colony stimulating factor; HMGB1, high mobility group box-1; ALDH2, aldehyde dehydrogenase 2; HIF, hypoxia-inducible factor; FGF, fibroblast growth factor; miR, microRNA; TXNIP, thioredoxin interacting protein; REG3, regenerating islet-derived protein 3; PanPPAR, pan-peroxisome proliferator-activated receptor; SGLT, sodium-glucose co-transporter; ASBT, apical sodium-dependent bile acid transporter; CCR, C-C chemokine receptor; SCD, stearoyl-CoA desaturase; PPAR, peroxisome proliferator-activated receptor; ASK1, apoptosis signal-regulating kinase 1; LXRα, liver X receptor α; ATG4B, autophagy related 4B cysteine peptidase; STAT, signal transducer and activator of transcription; Gα13, G protein subunit alpha 13; ITIH, inter-alpha-trypsin inhibitor heavy chain H1; TAZ, tafazzin; TNFAIP3, tumor necrosis factor alpha induced protein 3; USP, ubiquitin specific peptidase 22; SIRT, sirtuin 1; OTULIN, OTU domain-containing deubiquitinase with linear linkage specificity.
NAFLD AND DRUG CANDIDATES
Globally, NAFLD is the most common liver disease that encompasses diseases from inflamed steatosis to NASH, cirrhosis, and hepatocellular carcinoma [93]. In Asian and American populations, the prevalence of NAFLD is approximately 30% and the disease is often accompanied by type 2 diabetes and obesity [94]. Accurate diagnosis of NASH is essential because inflammation and/or fibrosis may determine the prognosis of NASH. Additionally, advanced conditions such as cirrhosis may require liver transplantation accordingly [95]. Currently, there are increased efforts to develop therapeutic strategies for NASH, which have no established treatment.
Previous studies have reported that multiple hits triggered NAFLD, which was recognized as a potential therapeutic goal. Several modulators of these targets and related pathways have been analyzed in clinical trials (phase II) or are currently in ongoing clinical trials (Table 1) [93]. The candidate drugs inhibit de novo lipogenesis or increase lipid export from the liver [94]. Additionally, liver or systemic insulin resistance is improved by diverse drugs or gastrointestinal hormones such as peroxisome proliferator-activated receptor γ agonists. Accumulation of excess lipids may facilitate simple steatosis progression to inflamed steatosis. This step usually provides effective targeting because inflammation generally precedes fibrosis. Currently, the inhibitors of apoptosis signal-regulating kinase 1, C-C chemokine receptor types 2/5, and chemokine receptors of C-C motif chemokine 5 and C-C motif chemokine 2 are under phase III clinical trials [96]. Several farnesoid X receptor agonists reduce gluconeogenesis and liver fat production, inhibit bile acid synthesis, improve peripheral insulin sensitivity, and thereby have a profound effect on various pathways for development of NAFLD [97,98]. Several farnesoid X receptor agonists have been tested in clinical trials (Table 1) [99]. Combination therapy may be an effective therapeutic strategy for NAFLD as it has a complex pathophysiology.
CONCLUSION
In this review, we attempted to summarize the molecular mechanisms underlying ALD and NAFLD. Both diseases are associated with unhealthy lifestyle habits, including increased consumption of alcohol and unhealthy diet. Extraordinary progress has been made in the understanding of the pathology of fatty liver disease so far, suggesting that several parallel hits are required to conquer the disease. Understanding the molecular mechanisms underlying ALD and NAFLD is important for medical professionals, especially physicians, general practitioners, hepatologists, and diabetologists. Because fatty liver disease has many clinical features, doctors should consider the patients in a multifaceted way. Identifying the various molecular mechanisms and finding therapeutic functions are key clinical prerequisites. In the future, the awareness of the disease progression and key targets would enable proper diagnosis, effective management, and treatment of comorbid diseases.
Acknowledgments
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (NRF-2018R1A2A1A05078694) and in part by Dongguk University fund.
Abbreviations
- ALD
alcoholic liver disease
- ER
endoplasmic reticulum
- FFAs
free fatty acids
- Gα12
G protein subunit alpha 12
- HSCs
hepatic stellate cells
- LXRα
liver X receptor α
- miR
microRNA
- NAD
nicotinamide adenine dinucleotide
- NADH
nicotinamide adenine dinucleotide hydride
- NAFLD
non-alcoholic fatty liver disease
- NASH
non-alcoholic steatohepatitis
- ROS
reactive oxygen species
Footnotes
Authors’ contribution
YSK and SGK contributed to the literature review, and manuscript preparation.
Conflicts of Interest: The authors have no conflicts to disclose.
REFERENCES
- 1.Ramaiah S, Rivera C, Arteel G. Early-phase alcoholic liver disease: an update on animal models, pathology, and pathogenesis. Int J Toxicol. 2004;23:217–231. doi: 10.1080/10915810490502069. [DOI] [PubMed] [Google Scholar]
- 2.Mathurin P, Bataller R. Trends in the management and burden of alcoholic liver disease. J Hepatol. 2015;62(1 Suppl):S38–S46. doi: 10.1016/j.jhep.2015.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zakhari S, Li TK. Determinants of alcohol use and abuse: impact of quantity and frequency patterns on liver disease. Hepatology. 2007;46:2032–2039. doi: 10.1002/hep.22010. [DOI] [PubMed] [Google Scholar]
- 4.Gual P, Gilgenkrantz H, Lotersztajn S. Autophagy in chronic liver diseases: the two faces of Janus. Am J Physiol Cell Physiol. 2017;312:C263–C273. doi: 10.1152/ajpcell.00295.2016. [DOI] [PubMed] [Google Scholar]
- 5.Younossi Z, Tacke F, Arrese M, Chander Sharma B, Mostafa I, Bugianesi E, et al. Global perspectives on nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Hepatology. 2019;69:2672–2682. doi: 10.1002/hep.30251. [DOI] [PubMed] [Google Scholar]
- 6.Tannapfel A, Denk H, Dienes HP, Langner C, Schirmacher P, Trauner M, et al. Histopathological diagnosis of non-alcoholic and alcoholic fatty liver disease. Virchows Arch. 2011;458:511–523. doi: 10.1007/s00428-011-1066-1. [DOI] [PubMed] [Google Scholar]
- 7.Kaufman RJ. Stress signaling from the lumen of the endoplasmic reticulum: coordination of gene transcriptional and translational controls. Genes Dev. 1999;13:1211–1233. doi: 10.1101/gad.13.10.1211. [DOI] [PubMed] [Google Scholar]
- 8.Schröder M, Kaufman RJ. The mammalian unfolded protein response. Annu Rev Biochem. 2005;74:739–789. doi: 10.1146/annurev.biochem.73.011303.074134. [DOI] [PubMed] [Google Scholar]
- 9.Oyadomari S, Araki E, Mori M. Endoplasmic reticulum stress-mediated apoptosis in pancreatic beta-cells. Apoptosis. 2002;7:335–345. doi: 10.1023/a:1016175429877. [DOI] [PubMed] [Google Scholar]
- 10.Mori K. Tripartite management of unfolded proteins in the endoplasmic reticulum. Cell. 2000;101:451–454. doi: 10.1016/s0092-8674(00)80855-7. [DOI] [PubMed] [Google Scholar]
- 11.Kaufman RJ, Scheuner D, Schröder M, Shen X, Lee K, Liu CY, et al. The unfolded protein response in nutrient sensing and differentiation. Nat Rev Mol Cell Biol. 2002;3:411–421. doi: 10.1038/nrm829. [DOI] [PubMed] [Google Scholar]
- 12.Araki E, Oyadomari S, Mori M. Impact of endoplasmic reticulum stress pathway on pancreatic beta-cells and diabetes mellitus. Exp Biol Med (Maywood) 2003;228:1213–1217. doi: 10.1177/153537020322801018. [DOI] [PubMed] [Google Scholar]
- 13.Xu C, Bailly-Maitre B, Reed JC. Endoplasmic reticulum stress: cell life and death decisions. J Clin Invest. 2005;115:2656–2664. doi: 10.1172/JCI26373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lieber CS. Hepatic, metabolic and toxic effects of ethanol: 1991 update. Alcohol Clin Exp Res. 1991;15:573–592. doi: 10.1111/j.1530-0277.1991.tb00563.x. [DOI] [PubMed] [Google Scholar]
- 15.Grunnet N, Kondrup J. The effect of ethanol on the beta-oxidation of fatty acids. Alcohol Clin Exp Res. 1986;10(6 Suppl):64S–68S. doi: 10.1111/j.1530-0277.1986.tb05182.x. [DOI] [PubMed] [Google Scholar]
- 16.Cho YE, Mezey E, Hardwick JP, Salem N, Jr, Clemens DL, Song BJ. Increased ethanol-inducible cytochrome P450-2E1 and cytochrome P450 isoforms in exosomes of alcohol-exposed rodents and patients with alcoholism through oxidative and endoplasmic reticulum stress. Hepatol Commun. 2017;1:675–690. doi: 10.1002/hep4.1066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Theurey P, Rieusset J. Mitochondria-associated membranes response to nutrient availability and role in metabolic diseases. Trends Endocrinol Metab. 2017;28:32–45. doi: 10.1016/j.tem.2016.09.002. [DOI] [PubMed] [Google Scholar]
- 18.Wang L, Liu X, Nie J, Zhang J, Kimball SR, Zhang H, et al. ALCAT1 controls mitochondrial etiology of fatty liver diseases, linking defective mitophagy to steatosis. Hepatology. 2015;61:486–496. doi: 10.1002/hep.27420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kawano Y, Cohen DE. Mechanisms of hepatic triglyceride accumulation in non-alcoholic fatty liver disease. J Gastroenterol. 2013;48:434–441. doi: 10.1007/s00535-013-0758-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Obrowsky S, Chandak PG, Patankar JV, Povoden S, Schlager S, Kershaw EE, et al. Adipose triglyceride lipase is a TG hydrolase of the small intestine and regulates intestinal PPARα signaling. J Lipid Res. 2013;54:425–435. doi: 10.1194/jlr.M031716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Parker R, Kim SJ, Im GY, Nahas J, Dhesi B, Vergis N, et al. Obesity in acute alcoholic hepatitis increases morbidity and mortality. EBioMedicine. 2019;45:511–518. doi: 10.1016/j.ebiom.2019.03.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fernández-Checa JC, Hirano T, Tsukamoto H, Kaplowitz N. Mitochondrial glutathione depletion in alcoholic liver disease. Alcohol. 1993;10:469–475. doi: 10.1016/0741-8329(93)90067-x. [DOI] [PubMed] [Google Scholar]
- 23.Fabbrini E, Sullivan S, Klein S. Obesity and nonalcoholic fatty liver disease: biochemical, metabolic, and clinical implications. Hepatology. 2010;51:679–689. doi: 10.1002/hep.23280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mandard S, Müller M, Kersten S. Peroxisome proliferator-activated receptor alpha target genes. Cell Mol Life Sci. 2004;61:393–416. doi: 10.1007/s00018-003-3216-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Shin SY, Kim TH, Wu H, Choi YH, Kim SG. SIRT1 activation by methylene blue, a repurposed drug, leads to AMPK-mediated inhibition of steatosis and steatohepatitis. Eur J Pharmacol. 2014;727:115–124. doi: 10.1016/j.ejphar.2014.01.035. [DOI] [PubMed] [Google Scholar]
- 26.Kim TH, Yang YM, Han CY, Koo JH, Oh H, Kim SS, et al. Gα12 ablation exacerbates liver steatosis and obesity by suppressing USP22/SIRT1-regulated mitochondrial respiration. J Clin Invest. 2018;128:5587–5602. doi: 10.1172/JCI97831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Weiskirchen R, Weiskirchen S, Tacke F. Organ and tissue fibrosis: molecular signals, cellular mechanisms and translational implications. Mol Aspects Med. 2019;65:2–15. doi: 10.1016/j.mam.2018.06.003. [DOI] [PubMed] [Google Scholar]
- 28.Weiskirchen R, Weiskirchen S, Tacke F. Recent advances in understanding liver fibrosis: bridging basic science and individualized treatment concepts. F1000Res. 2018;7:F1000 Faculty Rev-921. doi: 10.12688/f1000research.14841.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lotersztajn S, Julien B, Teixeira-Clerc F, Grenard P, Mallat A. Hepatic fibrosis: molecular mechanisms and drug targets. Annu Rev Pharmacol Toxicol. 2005;45:605–628. doi: 10.1146/annurev.pharmtox.45.120403.095906. [DOI] [PubMed] [Google Scholar]
- 30.Gao B, Bataller R. Alcoholic liver disease: pathogenesis and new therapeutic targets. Gastroenterology. 2011;141:1572–1585. doi: 10.1053/j.gastro.2011.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Marra F, Lotersztajn S. Pathophysiology of NASH: perspectives for a targeted treatment. Curr Pharm Des. 2013;19:5250–5269. doi: 10.2174/13816128113199990344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kaur J, Debnath J. Autophagy at the crossroads of catabolism and anabolism. Nat Rev Mol Cell Biol. 2015;16:461–472. doi: 10.1038/nrm4024. [DOI] [PubMed] [Google Scholar]
- 33.Madrigal-Matute J, Cuervo AM. Regulation of liver metabolism by autophagy. Gastroenterology. 2016;150:328–339. doi: 10.1053/j.gastro.2015.09.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lamming DW, Bar-Peled L. Lysosome: the metabolic signaling hub. Traffic. 2019;20:27–38. doi: 10.1111/tra.12617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Boya P, Reggiori F, Codogno P. Emerging regulation and functions of autophagy. Nat Cell Biol. 2013;15:713–720. doi: 10.1038/ncb2788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mizushima N, Levine B, Cuervo AM, Klionsky DJ. Autophagy fights disease through cellular self-digestion. Nature. 2008;451:1069–1075. doi: 10.1038/nature06639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schulze RJ, Drižytė K, Casey CA, McNiven MA. Hepatic lipophagy: new insights into autophagic catabolism of lipid droplets in the liver. Hepatol Commun. 2017;1:359–369. doi: 10.1002/hep4.1056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Singh R, Cuervo AM. Autophagy in the cellular energetic balance. Cell Metab. 2011;13:495–504. doi: 10.1016/j.cmet.2011.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Deretic V, Saitoh T, Akira S. Autophagy in infection, inflammation and immunity. Nat Rev Immunol. 2013;13:722–737. doi: 10.1038/nri3532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rubinsztein DC, Codogno P, Levine B. Autophagy modulation as a potential therapeutic target for diverse diseases. Nat Rev Drug Discov. 2012;11:709–730. doi: 10.1038/nrd3802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ding WX, Li M, Chen X, Ni HM, Lin CW, Gao W, et al. Autophagy reduces acute ethanol-induced hepatotoxicity and steatosis in mice. Gastroenterology. 2010;139:1740–1752. doi: 10.1053/j.gastro.2010.07.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Donohue TM, Jr, Osna NA, Kharbanda KK, Thomes PG. Lysosome and proteasome dysfunction in alcohol-induced liver injury. Liver Research. 2019;3:191–205. [Google Scholar]
- 43.Li Y, Wang S, Ni HM, Huang H, Ding WX. Autophagy in alcohol-induced multiorgan injury: mechanisms and potential therapeutic targets. Biomed Res Int. 2014;2014:498491. doi: 10.1155/2014/498491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Thomes PG, Trambly CS, Fox HS, Tuma DJ, Donohue TM., Jr Acute and chronic ethanol administration differentially modulate hepatic autophagy and transcription factor EB. Alcohol Clin Exp Res. 2015;39:2354–2363. doi: 10.1111/acer.12904. [DOI] [PubMed] [Google Scholar]
- 45.Lavallard VJ, Gual P. Autophagy and non-alcoholic fatty liver disease. BioMed Res Int. 2014;2014:120179. doi: 10.1155/2014/120179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Shibata M, Yoshimura K, Furuya N, Koike M, Ueno T, Komatsu M, et al. The MAP1-LC3 conjugation system is involved in lipid droplet formation. Biochem Biophys Res Commun. 2009;382:419–423. doi: 10.1016/j.bbrc.2009.03.039. [DOI] [PubMed] [Google Scholar]
- 47.Yang L, Li P, Fu S, Calay ES, Hotamisligil GS. Defective hepatic autophagy in obesity promotes ER stress and causes insulin resistance. Cell Metab. 2010;11:467–478. doi: 10.1016/j.cmet.2010.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Tanaka S, Hikita H, Tatsumi T, Sakamori R, Nozaki Y, Sakane S, et al. Rubicon inhibits autophagy and accelerates hepatocyte apoptosis and lipid accumulation in nonalcoholic fatty liver disease in mice. Hepatology. 2016;64:1994–2014. doi: 10.1002/hep.28820. [DOI] [PubMed] [Google Scholar]
- 49.Lin CW, Zhang H, Li M, Xiong X, Chen X, Chen X, et al. Pharmacological promotion of autophagy alleviates steatosis and injury in alcoholic and non-alcoholic fatty liver conditions in mice. J Hepatol. 2013;58:993–999. doi: 10.1016/j.jhep.2013.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kim YS, Nam HJ, Han CY, Joo MS, Jang K, Jun DW, et al. LXRα activation inhibits autophagy and lipophagy in hepatocytes by dysregulating ATG4B and Rab-8B, reducing mitochondrial fuel oxidation. Hepatology. 2020 Jun 17; doi: 10.1002/hep.31423. doi: 10.1002/hep.31423. [DOI] [PubMed] [Google Scholar]
- 51.Gracia-Sancho J, Guixé-Muntet S. The many-faced role of autophagy in liver diseases. J Hepatol. 2018;68:593–594. doi: 10.1016/j.jhep.2017.09.015. [DOI] [PubMed] [Google Scholar]
- 52.Tsuchida T, Friedman SL. Mechanisms of hepatic stellate cell activation. Nat Rev Gastroenterol Hepatol. 2017;14:397–411. doi: 10.1038/nrgastro.2017.38. [DOI] [PubMed] [Google Scholar]
- 53.Kim KM, Han CY, Kim JY, Cho SS, Kim YS, Koo JH, et al. Gα12 overexpression induced by miR-16 dysregulation contributes to liver fibrosis by promoting autophagy in hepatic stellate cells. J Hepatol. 2018;68:493–504. doi: 10.1016/j.jhep.2017.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Tran A, Gual P. Non-alcoholic steatohepatitis in morbidly obese patients. Clin Res Hepatol Gastroenterol. 2013;37:17–29. doi: 10.1016/j.clinre.2012.07.005. [DOI] [PubMed] [Google Scholar]
- 55.Wang S, Zhu M, Wang Q, Hou Y, Li L, Weng H, et al. Alpha-fetoprotein inhibits autophagy to promote malignant behaviour in hepatocellular carcinoma cells by activating PI3K/AKT/mTOR signalling. Cell Death Dis. 2018;9:1027. doi: 10.1038/s41419-018-1036-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Thoen LF, Guimarães EL, Dollé L, Mannaerts I, Najimi M, Sokal E, et al. A role for autophagy during hepatic stellate cell activation. J Hepatol. 2011;55:1353–1360. doi: 10.1016/j.jhep.2011.07.010. [DOI] [PubMed] [Google Scholar]
- 57.Wannamethee G, Shaper AG. Blood lipids: the relationship with alcohol intake, smoking, and body weight. J Epidemiol Community Health. 1992;46:197–202. doi: 10.1136/jech.46.3.197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Zilkens RR, Rich L, Burke V, Beilin LJ, Watts GF, Puddey IB. Effects of alcohol intake on endothelial function in men: a randomized controlled trial. J Hypertens. 2003;21:97–103. doi: 10.1097/00004872-200301000-00019. [DOI] [PubMed] [Google Scholar]
- 59.Volkow ND, Wang GJ, Baler RD. Reward, dopamine and the control of food intake: implications for obesity. Trends Cogn Sci. 2011;15:37–46. doi: 10.1016/j.tics.2010.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Seiler AE, Henderson A, Rubin R. Ethanol inhibits insulin receptor tyrosine kinase. Alcohol Clin Exp Res. 2000;24:1869–1872. [PubMed] [Google Scholar]
- 61.Lieber CS. New pathway of ethanol metabolism in the liver. Gastroenterology. 1970;59:930–937. [PubMed] [Google Scholar]
- 62.Lieber CS, DeCarli LM. Hepatotoxicity of ethanol. J Hepatol. 1991;12:394–401. doi: 10.1016/0168-8278(91)90846-4. [DOI] [PubMed] [Google Scholar]
- 63.Yin H, Hu M, Zhang R, Shen Z, Flatow L, You M. MicroRNA-217 promotes ethanol-induced fat accumulation in hepatocytes by down-regulating SIRT1. J Biol Chem. 2012;287:9817–9826. doi: 10.1074/jbc.M111.333534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Leung TM, Lu Y, Yan W, Morón-Concepción JA, Ward SC, Ge X, et al. Argininosuccinate synthase conditions the response to acute and chronic ethanol-induced liver injury in mice. Hepatology. 2012;55:1596–1609. doi: 10.1002/hep.25543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lehwald N, Tao GZ, Jang KY, Papandreou I, Liu B, Liu B, et al. β-catenin regulates hepatic mitochondrial function and energy balance in mice. Gastroenterology. 2012;143:754–764. doi: 10.1053/j.gastro.2012.05.048. [DOI] [PubMed] [Google Scholar]
- 66.Junge J, Horn T, Christoffersen P. Megamitochondria as a diagnostic marker for alcohol induced centrilobular and periportal fibrosis in the liver. Virchows Arch A Pathol Anat Histopathol. 1987;410:553–558. doi: 10.1007/BF00781692. [DOI] [PubMed] [Google Scholar]
- 67.Palma E, Ma X, Riva A, Iansante V, Dhawan A, Wang S, et al. Dynamin-1-like protein inhibition drives megamitochondria formation as an adaptive response in alcohol-induced hepatotoxicity. Am J Pathol. 2019;189:580–589. doi: 10.1016/j.ajpath.2018.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ertunc ME, Hotamisligil GS. Lipid signaling and lipotoxicity in metaflammation: indications for metabolic disease pathogenesis and treatment. J Lipid Res. 2016;57:2099–2114. doi: 10.1194/jlr.R066514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Wang Y, Viscarra J, Kim SJ, Sul HS. Transcriptional regulation of hepatic lipogenesis. Nat Rev Mol Cell Biol. 2015;16:678–689. doi: 10.1038/nrm4074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Williams JA, Ni HM, Ding Y, Ding WX. Parkin regulates mitophagy and mitochondrial function to protect against alcohol-induced liver injury and steatosis in mice. Am J Physiol Gastrointest Liver Physiol. 2015;309:G324–G340. doi: 10.1152/ajpgi.00108.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Singh R, Kaushik S, Wang Y, Xiang Y, Novak I, Komatsu M, et al. Autophagy regulates lipid metabolism. Nature. 2009;458:1131–1135. doi: 10.1038/nature07976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Ipsen DH, Lykkesfeldt J, Tveden-Nyborg P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell Mol Life Sci. 2018;75:3313–3327. doi: 10.1007/s00018-018-2860-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Teli MR, Day CP, Burt AD, Bennett MK, James OF. Determinants of progression to cirrhosis or fibrosis in pure alcoholic fatty liver. Lancet. 1995;346:987–990. doi: 10.1016/s0140-6736(95)91685-7. [DOI] [PubMed] [Google Scholar]
- 74.Zhou Z, Wang L, Song Z, Lambert JC, McClain CJ, Kang YJ. A critical involvement of oxidative stress in acute alcohol-induced hepatic TNF-alpha production. Am J Pathol. 2003;163:1137–1146. doi: 10.1016/s0002-9440(10)63473-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.McVicker BL, Rasineni K, Tuma DJ, McNiven MA, Casey CA. Lipid droplet accumulation and impaired fat efflux in polarized hepatic cells: consequences of ethanol metabolism. Int J Hepatol. 2012;2012:978136. doi: 10.1155/2012/978136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Crabb DW, Galli A, Fischer M, You M. Molecular mechanisms of alcoholic fatty liver: role of peroxisome proliferator-activated receptor alpha. Alcohol. 2004;34:35–38. doi: 10.1016/j.alcohol.2004.07.005. [DOI] [PubMed] [Google Scholar]
- 77.Bataller R, Rombouts K, Altamirano J, Marra F. Fibrosis in alcoholic and nonalcoholic steatohepatitis. Best Pract Res Clin Gastroenterol. 2011;25:231–244. doi: 10.1016/j.bpg.2011.02.010. [DOI] [PubMed] [Google Scholar]
- 78.McAlaney J, McMahon J. Establishing rates of binge drinking in the UK: anomalies in the data. Alcohol Alcohol. 2006;41:355–357. doi: 10.1093/alcalc/agl025. [DOI] [PubMed] [Google Scholar]
- 79.Mathurin P, Deltenre P. Effect of binge drinking on the liver: an alarming public health issue? Gut. 2009;58:613–617. doi: 10.1136/gut.2007.145573. [DOI] [PubMed] [Google Scholar]
- 80.Sozio M, Crabb DW. Alcohol and lipid metabolism. Am J Physiol Endocrinol Metab. 2008;295:E10–E16. doi: 10.1152/ajpendo.00011.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.French SW. Ethanol and hepatocellular injury. Clin Lab Med. 1996;16:289–306. [PubMed] [Google Scholar]
- 82.Savolainen VT, Liesto K, Männikkö A, Penttilä A, Karhunen PJ. Alcohol consumption and alcoholic liver disease: evidence of a threshold level of effects of ethanol. Alcohol Clin Exp Res. 1993;17:1112–1117. doi: 10.1111/j.1530-0277.1993.tb05673.x. [DOI] [PubMed] [Google Scholar]
- 83.Grant BF, Dufour MC, Harford TC. Epidemiology of alcoholic liver disease. Semin Liver Dis. 1988;8:12–25. doi: 10.1055/s-2008-1040525. [DOI] [PubMed] [Google Scholar]
- 84.MacSween RN, Burt AD. Histologic spectrum of alcoholic liver disease. Semin Liver Dis. 1986;6:221–232. doi: 10.1055/s-2008-1040605. [DOI] [PubMed] [Google Scholar]
- 85.Zimatkin SM, Pronko SP, Vasiliou V, Gonzalez FJ, Deitrich RA. Enzymatic mechanisms of ethanol oxidation in the brain. Alcohol Clin Exp Res. 2006;30:1500–1505. doi: 10.1111/j.1530-0277.2006.00181.x. [DOI] [PubMed] [Google Scholar]
- 86.Osna NA, Donohue TM, Jr, Kharbanda KK. Alcoholic liver disease: pathogenesis and current management. Alcohol Res. 2017;38:147–161. [PMC free article] [PubMed] [Google Scholar]
- 87.Milne GL, Sanchez SC, Musiek ES, Morrow JD. Quantification of F2-isoprostanes as a biomarker of oxidative stress. Nat Protoc. 2007;2:221–226. doi: 10.1038/nprot.2006.375. [DOI] [PubMed] [Google Scholar]
- 88.Morrow JD, Hill KE, Burk RF, Nammour TM, Badr KF, Roberts LJ., 2nd A series of prostaglandin F2-like compounds are produced in vivo in humans by a non-cyclooxygenase, free radical-catalyzed mechanism. Proc Natl Acad Sci U S A. 1990;87:9383–9387. doi: 10.1073/pnas.87.23.9383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Iuliano L, Praticò D, Greco C, Mangieri E, Scibilia G, FitzGerald GA, et al. Angioplasty increases coronary sinus F2-isoprostane formation: evidence for in vivo oxidative stress during PTCA. J Am Coll Cardiol. 2001;37:76–80. doi: 10.1016/s0735-1097(00)01040-8. [DOI] [PubMed] [Google Scholar]
- 90.Murphy MP. How mitochondria produce reactive oxygen species. Biochem J. 2009;417:1–13. doi: 10.1042/BJ20081386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Heo MJ, Kim TH, You JS, Blaya D, Sancho-Bru P, Kim SG. Alcohol dysregulates miR-148a in hepatocytes through FoxO1, facilitating pyroptosis via TXNIP overexpression. Gut. 2019;68:708–720. doi: 10.1136/gutjnl-2017-315123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Chen Y, Singh S, Matsumoto A, Manna SK, Abdelmegeed MA, Golla S, et al. Chronic glutathione depletion confers protection against alcohol-induced steatosis: implication for redox activation of AMP-activated protein kinase pathway. Sci Rep. 2016;6:29743. doi: 10.1038/srep29743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Rinella ME. Nonalcoholic fatty liver disease: a systematic review. JAMA. 2015;313:2263–2273. doi: 10.1001/jama.2015.5370. [DOI] [PubMed] [Google Scholar]
- 94.Targher G. Non-alcoholic fatty liver disease as driving force in coronary heart disease? Gut. 2017;66:213–214. doi: 10.1136/gutjnl-2016-312891. [DOI] [PubMed] [Google Scholar]
- 95.Lassailly G, Caiazzo R, Pattou F, Mathurin P. Perspectives on treatment for nonalcoholic steatohepatitis. Gastroenterology. 2016;150:1835–1848. doi: 10.1053/j.gastro.2016.03.004. [DOI] [PubMed] [Google Scholar]
- 96.European Association for the Study of the Liver (EASL) European Association for the Study of Diabetes (EASD) European Association for the Study of Obesity (EASO) EASL-EASD-EASO clinical practice Guidelines for the management of non-alcoholic fatty liver disease. J Hepatol. 2016;64:1388–1402. doi: 10.1016/j.jhep.2015.11.004. [DOI] [PubMed] [Google Scholar]
- 97.Ma K, Saha PK, Chan L, Moore DD. Farnesoid X receptor is essential for normal glucose homeostasis. J Clin Invest. 2006;116:1102–1109. doi: 10.1172/JCI25604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Ploton M, Mazuy C, Gheeraert C, Dubois V, Berthier A, Dubois-Chevalier J, et al. The nuclear bile acid receptor FXR is a PKA- and FOXA2-sensitive activator of fasting hepatic gluconeogenesis. J Hepatol. 2018;69:1099–1109. doi: 10.1016/j.jhep.2018.06.022. [DOI] [PubMed] [Google Scholar]
- 99.Brunt EM, Kleiner DE. Challenges in the hepatic histopathology in non-alcoholic fatty liver disease. Gut. 2017;66:1539–1540. doi: 10.1136/gutjnl-2016-313379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Arab JP, Sehrawat TS, Simonetto DA, Verma VK, Feng D, Tang T, et al. An open-label, dose-escalation study to assess the safety and efficacy of IL-22 agonist F-652 in patients with alcohol-associated hepatitis. Hepatology. 2020;72:441–453. doi: 10.1002/hep.31046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Spalinger MR, Atrott K, Baebler K, Schwarzfischer M, Melhem H, Peres DR, et al. Administration of the hyper-immune bovine colostrum extract IMM-124E ameliorates experimental murine colitis. J Crohns Colitis. 2019;13:785–797. doi: 10.1093/ecco-jcc/jjy213. [DOI] [PubMed] [Google Scholar]
- 102.Mantegazza C, Molinari P, D’Auria E, Sonnino M, Morelli L, Zuccotti GV. Probiotics and antibiotic-associated diarrhea in children: a review and new evidence on Lactobacillus rhamnosus GG during and after antibiotic treatment. Pharmacol Res. 2018;128:63–72. doi: 10.1016/j.phrs.2017.08.001. [DOI] [PubMed] [Google Scholar]
- 103.Petrasek J, Bala S, Csak T, Lippai D, Kodys K, Menashy V, et al. IL-1 receptor antagonist ameliorates inflammasome-dependent alcoholic steatohepatitis in mice. J Clin Invest. 2012;122:3476–3489. doi: 10.1172/JCI60777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Iracheta-Vellve A, Calenda CD, Petrasek J, Ambade A, Kodys K, Adorini L, et al. FXR and TGR5 agonists ameliorate liver injury, steatosis, and inflammation after binge or prolonged alcohol feeding in mice. Hepatol Commun. 2018;2:1379–1391. doi: 10.1002/hep4.1256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Hakami AY, Alshehri FS, Althobaiti YS, Sari Y. Effects of orally administered augmentin on glutamate transporter 1, cystine-glutamate exchanger expression and ethanol intake in alcohol-preferring rats. Behav Brain Res. 2017;320:316–322. doi: 10.1016/j.bbr.2016.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Di Miceli M, Gronier B. Pharmacology, systematic review and recent clinical trials of metadoxine. Rev Recent Clin Trials. 2018;13:114–125. doi: 10.2174/1574887113666180227100217. [DOI] [PubMed] [Google Scholar]
- 107.Nikou T, Ioannidis A, Zoga M, Tzavellas E, Paparrigopoulos T, Magana M, et al. Alteration in the concentrations of interleukin-7 (IL-7), interleukin-10 (IL-10) and granulocyte colony stimulating factor (G-CSF) in alcohol-dependent individuals without liver disease, during detoxification therapy. Drug Alcohol Depend. 2016;163:77–83. doi: 10.1016/j.drugalcdep.2016.03.022. [DOI] [PubMed] [Google Scholar]
- 108.Choi WM, Kim HH, Kim MH, Cinar R, Yi HS, Eun HS, et al. Glutamate signaling in hepatic stellate cells drives alcoholic steatosis. Cell Metab. 2019;30:877–889. doi: 10.1016/j.cmet.2019.08.001. e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Ge X, Antoine DJ, Lu Y, Arriazu E, Leung TM, Klepper AL, et al. High mobility group box-1 (HMGB1) participates in the pathogenesis of alcoholic liver disease (ALD) J Biol Chem. 2014;289:22672–22691. doi: 10.1074/jbc.M114.552141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Seo W, Gao Y, He Y, Sun J, Xu H, Feng D, et al. ALDH2 deficiency promotes alcohol-associated liver cancer by activating oncogenic pathways via oxidized DNA-enriched extracellular vesicles. J Hepatol. 2019;71:1000–1011. doi: 10.1016/j.jhep.2019.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Lee JH, Shim YR, Seo W, Kim MH, Choi WM, Kim HH, et al. Mitochondrial double-stranded RNA in exosome promotes interleukin-17 production through toll-like receptor 3 in alcohol-associated liver injury. Hepatology. 2020;72:609–625. doi: 10.1002/hep.31041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.McCullough RL, McMullen MR, Poulsen KL, Kim A, Medof ME, Nagy LE. Anaphylatoxin receptors C3aR and C5aR1 are important factors that influence the impact of ethanol on the adipose secretome. Front Immunol. 2018;9:2133. doi: 10.3389/fimmu.2018.02133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Cho YE, Kim DK, Seo W, Gao B, Yoo SH, Song BJ. Fructose promotes leaky gut, endotoxemia, and liver fibrosis through ethanolinducible cytochrome P450-2E1-mediated oxidative and nitrative stress. Hepatology. 2019 Apr 8; doi: 10.1002/hep.30652. doi: 10.1002/hep.30652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Shao T, Zhao C, Li F, Gu Z, Liu L, Zhang L, et al. Intestinal HIF-1α deletion exacerbates alcoholic liver disease by inducing intestinal dysbiosis and barrier dysfunction. J Hepatol. 2018;69:886–895. doi: 10.1016/j.jhep.2018.05.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Forsyth CB, Farhadi A, Jakate SM, Tang Y, Shaikh M, Keshavarzian A. Lactobacillus GG treatment ameliorates alcohol-induced intestinal oxidative stress, gut leakiness, and liver injury in a rat model of alcoholic steatohepatitis. Alcohol. 2009;43:163–172. doi: 10.1016/j.alcohol.2008.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Hartmann P, Hochrath K, Horvath A, Chen P, Seebauer CT, Llorente C, et al. Modulation of the intestinal bile acid/farnesoid X receptor/fibroblast growth factor 15 axis improves alcoholic liver disease in mice. Hepatology. 2018;67:2150–2166. doi: 10.1002/hep.29676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Brandl K, Hartmann P, Jih LJ, Pizzo DP, Argemi J, Ventura-Cots M, et al. Dysregulation of serum bile acids and FGF19 in alcoholic hepatitis. J Hepatol. 2018;69:396–405. doi: 10.1016/j.jhep.2018.03.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Wang L, Fouts DE, Stärkel P, Hartmann P, Chen P, Llorente C, et al. Intestinal REG3 lectins protect against alcoholic steatohepatitis by reducing mucosa-associated microbiota and preventing bacterial translocation. Cell Host Microbe. 2016;19:227–239. doi: 10.1016/j.chom.2016.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Zhou J, Waskowicz LR, Lim A, Liao XH, Lian B, Masamune H, et al. A liver-specific thyromimetic, VK2809, decreases hepatosteatosis in glycogen storage disease type Ia. Thyroid. 2019;29:1158–1167. doi: 10.1089/thy.2019.0007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Boubia B, Poupardin O, Barth M, Binet J, Peralba P, Mounier L, et al. Design, synthesis, and evaluation of a novel series of indole sulfonamide peroxisome proliferator activated receptor (PPAR) α/γ/δ triple activators: discovery of lanifibranor, a new antifibrotic clinical candidate. J Med Chem. 2018;61:2246–2265. doi: 10.1021/acs.jmedchem.7b01285. [DOI] [PubMed] [Google Scholar]
- 121.Tully DC, Rucker PV, Chianelli D, Williams J, Vidal A, Alper PB, et al. Discovery of tropifexor (LJN452), a highly potent non-bile acid FXR agonist for the treatment of cholestatic liver diseases and nonalcoholic steatohepatitis (NASH) J Med Chem. 2017;60:9960–9973. doi: 10.1021/acs.jmedchem.7b00907. [DOI] [PubMed] [Google Scholar]
- 122.He YL, Haynes W, Meyers CD, Amer A, Zhang Y, Mahling P, et al. The effects of licogliflozin, a dual SGLT1/2 inhibitor, on body weight in obese patients with or without diabetes. Diabetes Obes Metab. 2019;21:1311–1321. doi: 10.1111/dom.13654. [DOI] [PubMed] [Google Scholar]
- 123.Li S, Ghoshal S, Sojoodi M, Arora G, Masia R, Erstad DJ, et al. The farnesoid X receptor agonist EDP-305 reduces interstitial renal fibrosis in a mouse model of unilateral ureteral obstruction. FASEB J. 2019;33:7103–7112. doi: 10.1096/fj.201801699R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Newsome PN, Palmer M, Freilich B, Sheikh MY, Sheikh A, Sarles H, et al. Volixibat in adults with non-alcoholic steatohepatitis: 24-week interim analysis from a randomized, phase II study. J Hepatol. 2020;73:231–240. doi: 10.1016/j.jhep.2020.03.024. [DOI] [PubMed] [Google Scholar]
- 125.Sanyal A, Charles ED, Neuschwander-Tetri BA, Loomba R, Harrison SA, Abdelmalek MF, et al. Pegbelfermin (BMS-986036), a PEGylated fibroblast growth factor 21 analogue, in patients with non-alcoholic steatohepatitis: a randomised, double-blind, placebo-controlled, phase 2a trial. Lancet. 2019;392:2705–2717. doi: 10.1016/S0140-6736(18)31785-9. [DOI] [PubMed] [Google Scholar]
- 126.Barreyro FJ, Holod S, Finocchietto PV, Camino AM, Aquino JB, Avagnina A, et al. The pan-caspase inhibitor emricasan (IDN-6556) decreases liver injury and fibrosis in a murine model of non-alcoholic steatohepatitis. Liver Int. 2015;35:953–966. doi: 10.1111/liv.12570. [DOI] [PubMed] [Google Scholar]
- 127.Harrison SA, Marri SR, Chalasani N, Kohli R, Aronstein W, Thompson GA, et al. Randomised clinical study: GR-MD-02, a galectin-3 inhibitor, vs. placebo in patients having non-alcoholic steatohepatitis with advanced fibrosis. Aliment Pharmacol Ther. 2016;44:1183–1198. doi: 10.1111/apt.13816. [DOI] [PubMed] [Google Scholar]
- 128.Tacke F. Cenicriviroc for the treatment of non-alcoholic steatohepatitis and liver fibrosis. Expert Opin Investig Drugs. 2018;27:301–311. doi: 10.1080/13543784.2018.1442436. [DOI] [PubMed] [Google Scholar]
- 129.Iruarrizaga-Lejarreta M, Varela-Rey M, Fernández-Ramos D, Martínez-Arranz I, Delgado TC, Simon J, et al. Role of Aramchol in steatohepatitis and fibrosis in mice. Hepatol Commun. 2017;1:911–927. doi: 10.1002/hep4.1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Harrison SA, Bashir MR, Guy CD, Zhou R, Moylan CA, Frias JP, et al. Resmetirom (MGL-3196) for the treatment of non-alcoholic steatohepatitis: a multicentre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet. 2019;394:2012–2024. doi: 10.1016/S0140-6736(19)32517-6. [DOI] [PubMed] [Google Scholar]
- 131.Ratziu V, Harrison SA, Francque S, Bedossa P, Lehert P, Serfaty L, et al. Elafibranor, an agonist of the peroxisome proliferator-activated receptor-α and -δ, induces resolution of nonalcoholic steatohepatitis without fibrosis worsening. Gastroenterology. 2016;150:1147–1159. doi: 10.1053/j.gastro.2016.01.038. e5. [DOI] [PubMed] [Google Scholar]
- 132.Younossi ZM, Ratziu V, Loomba R, Rinella M, Anstee QM, Goodman Z, et al. Obeticholic acid for the treatment of non-alcoholic steatohepatitis: interim analysis from a multicentre, randomised, placebo-controlled phase 3 trial. Lancet. 2019;394:2184–2196. doi: 10.1016/S0140-6736(19)33041-7. [DOI] [PubMed] [Google Scholar]
- 133.Schuster S, Feldstein AE. NASH: novel therapeutic strategies targeting ASK1 in NASH. Nat Rev Gastroenterol Hepatol. 2017;14:329–330. doi: 10.1038/nrgastro.2017.42. [DOI] [PubMed] [Google Scholar]
- 134.Akuta N, Watanabe C, Kawamura Y, Arase Y, Saitoh S, Fujiyama S, et al. Effects of a sodium-glucose cotransporter 2 inhibitor in nonalcoholic fatty liver disease complicated by diabetes mellitus: preliminary prospective study based on serial liver biopsies. Hepatol Commun. 2017;1:46–52. doi: 10.1002/hep4.1019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Grohmann M, Wiede F, Dodd GT, Gurzov EN, Ooi GJ, Butt T, et al. Obesity drives STAT-1-dependent NASH and STAT-3-dependent HCC. Cell. 2018;175:1289–1306. doi: 10.1016/j.cell.2018.09.053. e20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Andres-Hernando A, Orlicky DJ, Kuwabara M, Ishimoto T, Nakagawa T, Johnson RJ, et al. Deletion of fructokinase in the liver or in the intestine reveals differential effects on sugar-induced metabolic dysfunction. Cell Metab. 2020;32:117–127. doi: 10.1016/j.cmet.2020.05.012. e3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Kim TH, Koo JH, Heo MJ, Han CY, Kim YI, Park SY, et al. Overproduction of inter-α-trypsin inhibitor heavy chain 1 after loss of Gα13 in liver exacerbates systemic insulin resistance in mice. Sci Transl Med. 2019;11:eaan4735. doi: 10.1126/scitranslmed.aan4735. [DOI] [PubMed] [Google Scholar]
- 138.Wang X, Cai B, Yang X, Sonubi OO, Zheng Z, Ramakrishnan R, et al. Cholesterol stabilizes TAZ in hepatocytes to promote experimental non-alcoholic steatohepatitis. Cell Metab. 2020;31:969–986. doi: 10.1016/j.cmet.2020.03.010. e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Liu D, Zhang P, Zhou J, Liao R, Che Y, Gao MM, et al. TNFAIP3 interacting protein 3 overexpression suppresses nonalcoholic steatohepatitis by blocking TAK1 activation. Cell Metab. 2020;31:726–740. doi: 10.1016/j.cmet.2020.03.007. e8. [DOI] [PubMed] [Google Scholar]
- 140.Simon J, Nuñez-García M, Fernández-Tussy P, Barbier-Torres L, Fernández-Ramos D, Gómez-Santos B, et al. Targeting hepatic glutaminase 1 ameliorates non-alcoholic steatohepatitis by restoring very-low-density lipoprotein triglyceride assembly. Cell Metab. 2020;31:605–622. doi: 10.1016/j.cmet.2020.01.013. e10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Verboom L, Martens A, Priem D, Hoste E, Sze M, Vikkula H, et al. OTULIN prevents liver inflammation and hepatocellular carcinoma by inhibiting FADD- and RIPK1 kinase-mediated hepatocyte apoptosis. Cell Rep. 2020;30:2237–2247. doi: 10.1016/j.celrep.2020.01.028. e6. [DOI] [PubMed] [Google Scholar]