

Abstract
Aerobic exercise induces oxidative stress and DNA damage, nevertheless, lowers cancer incidence. It remains unclear how genetic stability is maintained under this condition. Here, we examined the dynamic change of the tumor suppressor p16INK4a in cells of skeletal muscle among young men following 60-min of aerobic cycling at 70% maximal oxygen consumption (V̇O2max). Rg1 (5 mg, an immunostimulant ginsenoside) and placebo (PLA) were supplemented 1 h before exercise. Data from serial muscle biopsies shows unchanged p16INK4a+ cells after exercise followed by a considerable increase (+21-fold) in vastus lateralis muscle 3 h later. This increase was due to the accumulation of endothelial progenitor cells (p16INK4a+/CD34+) surrounding myofibers and other infiltrated nucleated cells (p16INK4a+/CD34-) in necrotic myofibers. During the Rg1 trial, acute increases of p16INK4a+ cells in the muscle occurred immediately after exercise (+3-fold) and reversed near baseline 3 h later. Rg1 also lowered IL-10 mRNA relative to PLA 3 h after exercise. Post-exercise increases in VEGF mRNA and CD163+ macrophages were similar for PLA and Rg1 trials. Conclusion: The marked increases in p16INK4a protein expression of endothelial progenitor cells in skeletal muscle implicates a protective mechanism for maintaining genetic stability against aerobic exercise. Rg1 accelerates resolution of the exercise-induced stress response.
Keywords: cancer, tumor, cell cycle arrest, skeletal muscle
INTRODUCTION
Most of the cells within the human body are short-lived and age rapidly [1], where birth, aging, and death of cells are continuously occurring to sustain a stable cell population. Capillary endothelial cells have an average lifespan ranging from 2 to 15 days in mammalian tissues [1, 2]. Endothelial progenitor cells (CD34+) from bone marrow are responsible for the rapid replacement of unhealthy endothelial cells [3], as well as the donation of their nuclei to injured myofibers for regeneration [4]. We have recently reported various amounts of p16INK4a+ endothelial progenitor cells in more than 40% of capillaries surrounding myofibers in human skeletal muscle of young men between 20-25 years of age [5].
Given a huge amount of daily cell turnover in human adults, a random genetic mutation is unavoidable following numerous cell division. The protein p16INK4a, known as a tumor suppressor, is a protein inhibitor of the DNA synthesis phase of the cell cycle expressed in replicable cells [6]. Both cellular senescence and stress elevate p16INK4a protein in replicable cells, which induces cell cycle arrest [6, 7], and illustrates the protective role of p16INK4a for the fidelity during cell division in large multicellular systems. Downregulation of p16INK4a has been observed in a large number of tumors in humans.
Aerobic exercise transiently increases oxidative stress and DNA damage [8]. However, participating aerobic exercise is associated with lower cancer incidence in both animals and humans [9]. The underlying mechanism remains unclear. In this study, distribution of p16INK4a+ endothelial progenitor cells in vastus lateralis muscle were examined in young adults. Induction of cellular senescence facilitates immune clearance of unhealthy replicable cells by activating immune responses [10]. To further investigate the role of the immune system on dynamical changes of p16INK4a+ senescent cells, Rg1 was orally ingested 1 h before exercise [11, 12]. Rg1 is an immunostimulant during inflammation, which activates macrophage function [11, 13].
RESULTS
Endurance cycling at 70% V̇O2max for 1 h does not produce significant increases in circulating LDH and myoglobin (Figure 1). The lipid peroxidation marker TBARS levels tends to increase by 35% during the PLA trial (Figure 1C). No detectable change in p16INK4a+ senescent cells was observed immediately after exercise (PLA trial). However, ~21-fold increases (P < 0.01) in p16INK4a+ senescent cells of skeletal muscle occurred 3 h after exercise. During the Rg1 trial, ~3-fold increases (P < 0.05) in p16INK4a+ senescent cells of skeletal muscle were observed immediately exercise followed by ~40% decline 3 h after exercise (P < 0.05) (Figure 2A, 2C). CD34+ cell-to-fiber ratio of skeletal muscle was not altered after exercise (Figure 2B, 2D). A great portion of senescent cell accumulation in exercised muscle is contributed by increased senescent endothelial progenitor cells (p16INK4a+/CD34+) (Figure 2E, 2G). Approximately 40% of the increases is associated with other types of nucleated cells (p16INK4a+/CD34-) localized mostly in necrotic myofibers (Figure 2F, 2H). Dramatic increases in p16INK4a+ cells 3 h after exercise is potently lowered by Rg1 supplementation (P < 0.05). A moderate correlation (r = 0.29, p = 0.08) was found between p16INK4a positive cells and ß-galactosidase positive cells of 36 biopsied muscle samples in men aged 20-26 y (Figure 3).
Figure 1.
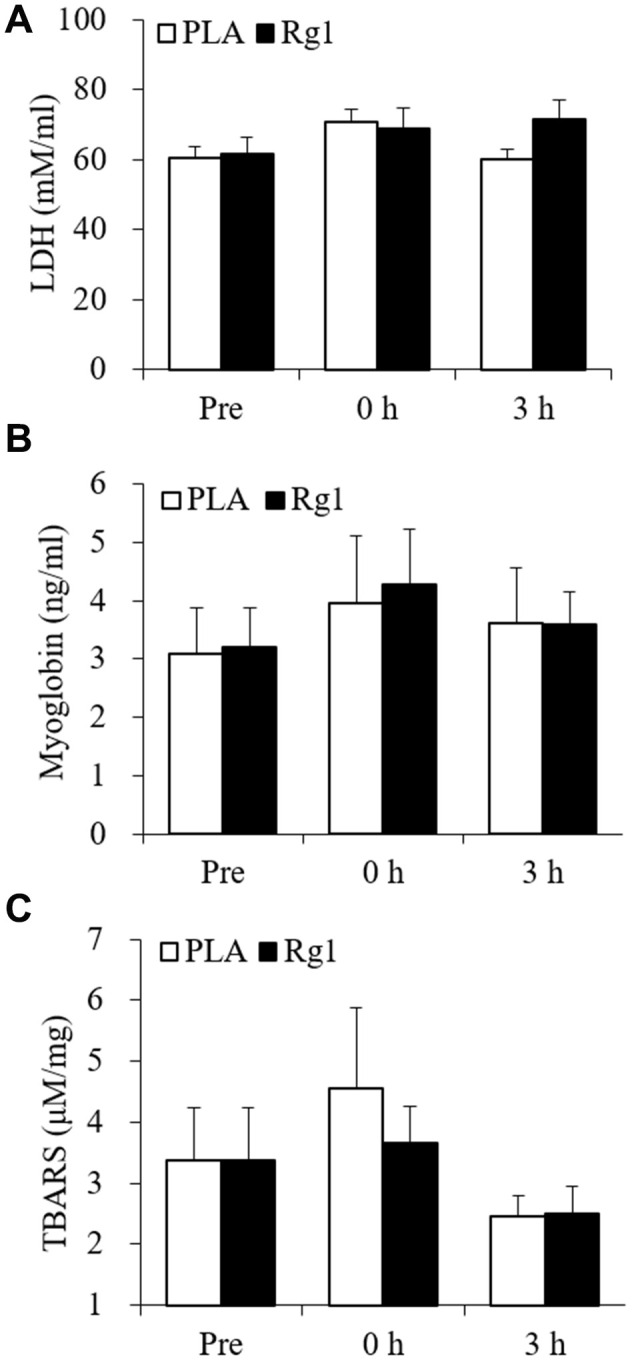
Plasma muscle damage markers unaltered following aerobic exercise. Lactate dehydrogenase (A), myoglobin (B), and TBARS (C) were not significantly increased after 1-h cycling exercise (70% V̇O2max). Data are expressed as mean and SEM. Abbreviation: TBARS, Thiobarbituric acid reactive substances.
Figure 2.

Massive increases of senescent endothelial progenitor cells in human skeletal muscle 3 h after aerobic exercise (70% V̇O2max). Approximately 21-fold increases in senescent endothelial progenitor cells occurred 3 h after 1-h cycling exercise (70% V̇O2max), while Rg1 supplementation advances the increase immediately after exercise (~3-fold) and decline to baseline 3 h after exercise (A, C). Total endothelial progenitor cells (CD34+) surrounding myofibers were unaltered after exercise for both trials (B, D). Approximately 60% of elevated senescent endothelial progenitor cells was contributed by endothelial progenitor cells (p16INK4a+ / CD34+) (E, G), while the rest of 40% was contributed by infiltrated nucleated cells (p16INK4a+ / CD34-) (F, H). Scale bar = 30 μm. Data are expressed as mean and SEM. * Significant difference against Pre (Baseline), P < 0.01; † Significant difference against Placebo, P < 0.01. Abbreviation: PLA, Placebo.
Figure 3.

Moderate correlation (r = 0.29, p = 0.08) between p16INK4a positive cells and ß-galactosidase positive cells of 36 biopsied vastus lateralis muscle in men aged 20-26 y. Abbreviation: ß-gal, ß-galactosidase.
Aerobic cycling (70% V̇O2max) increased CD163+ macrophage infiltration into human skeletal muscle (Figure 4). Placebo (PLA) and Rg1 trials show a similar magnitude of cell infiltration (PLA: 0 h, +63%, P < 0.05; 3 h, +56%, P < 0.05; Rg1: 0 h, +92%, P < 0.01; 3 h, +70%, P < 0.01).
Figure 4.

Aerobic exercise increases regenerative macrophage infiltration into human skeletal muscle. Arrows in the representative images indicate CD163+ cells (bright green) and nuclei (blue) surrounding myofibers in a muscle cross-section (A). This increase after 1-h cycling exercise (70% V̇O2max) was similar for both Placebo (PLA) and Rg1 trials (B). Scale bar = 50 μm. Data are expressed as mean and SEM. * Significant difference against Pre (Baseline), P < 0.01. Abbreviation: PLA, Placebo.
Data for exercise response in IL-10 mRNA, VEGF mRNA and PGC-1α mRNA of skeletal muscle are shown in Figure 5. IL-10 mRNA did not change after an acute bout aerobic exercise, while Rg1 significantly decreased IL-10 mRNA expression 3 h after exercise (-60%, P < 0.05) (Figure 5A). Both VEGF mRNA (Figure 5B) and PGC-1α mRNA (Figure 5C) increased significantly in challenged skeletal muscle. Similar increases were observed in VEGF mRNA of skeletal muscle for both PLA (0 h, +2-fold, P < 0.01; 3 h, +7-fold, P < 0.01) and Rg1 (0 h, +1-fold, P < 0.05; 3 h, +7-fold, P < 0.01) trials following exercise. PGC-1α mRNA also shows similar increases for PLA (0 h, +1-fold, P < 0.05; 3 h, +13-fold, P < 0.01) and Rg1 (0 h, +1-fold, P < 0.01; 3 h, +14-fold, P < 0.01) trials.
Figure 5.
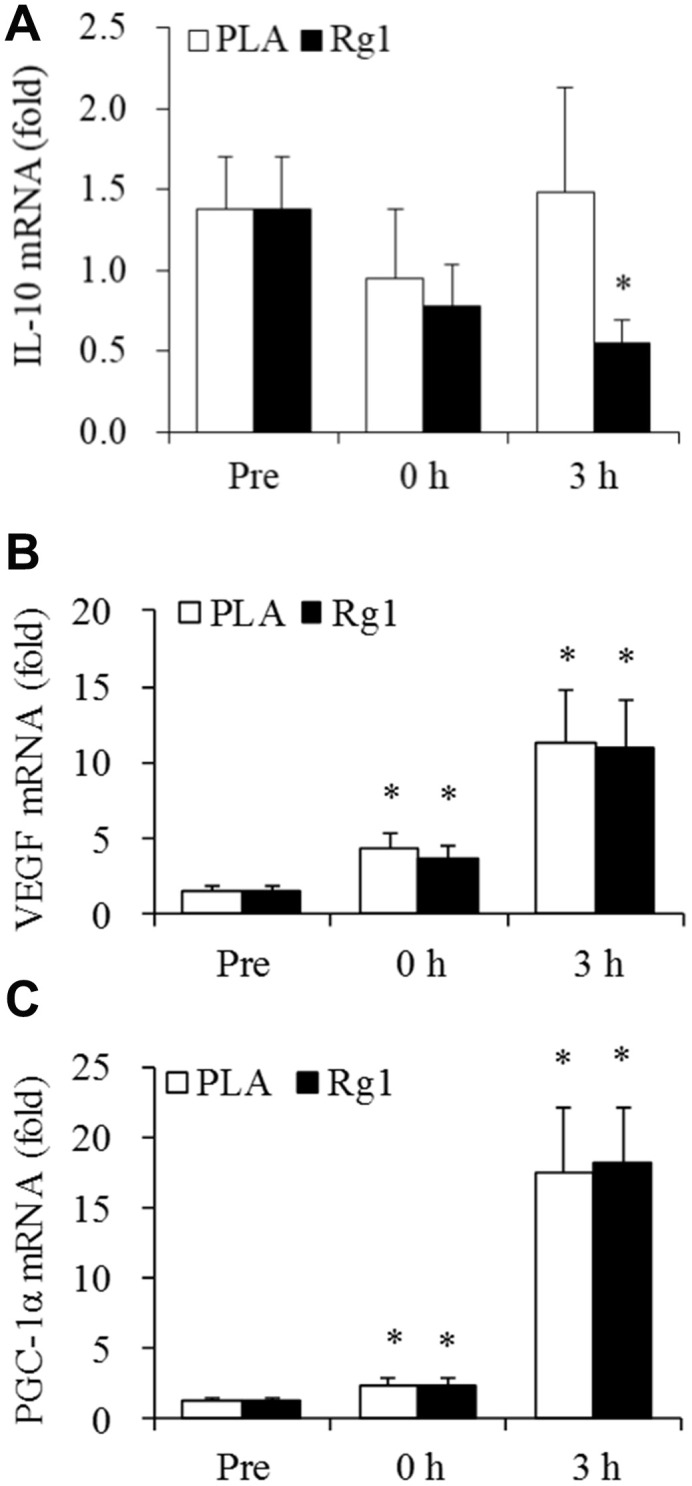
IL-10, VEGF, and PGC1-alpha expression after aerobic exercise. (A) Rg1 supplementation lowers IL-10 mRNA 3 h after exercise (-60%, P < 0.05). Dramatic elevations for VEGF mRNA (B) and PGC1-alpha mRNA (C) occurred 3 h after 1-h cycling exercise (70% V̇O2max). Data are expressed as mean and SEM. * Significant difference against Pre (Baseline), P < 0.05. Abbreviation: PLA, Placebo.
DISCUSSION
Aerobic exercise induces VEGF expression and stimulates angiogenesis [14]. During this process, endothelial progenitor cells are capable of self-replicating to increase cell number to renew damaged endothelial cells in the capillaries [15]. However, oxidative DNA damage is increased after an acute bout of aerobic exercise [8, 16]. In this study, we found a considerable rise in the tumor suppressor p16INK4a protein expression in endothelial progenitor cells of human skeletal muscle after an acute bout of aerobic exercise (70% V̇O2max). Furthermore, the immune-stimulant Rg1 accelerated the resolution of this stress response, evidenced by an earlier rise and fall of p16INK4a+ cell number in exercised muscle. Stress that causes DNA damage results in the increased expression of the p16INK4a protein, which is responsible for inhibition of cell division [6]. Thus, the transient increases in p16INK4a+ protein expression may mirror the magnitude of stress-related DNA damage [16] and suggests a protective mechanism for maintaining genetic stability of replicable cells against aerobic exercise.
Increasing p16INK4a protein expression is also known to stimulate tissue repair during inflammation [7, 17]. Inflammation is essential for recognition and phagocytic clearance of unhealthy cells that develops a senescence phenotype [18–20] followed by a protracted period of cell regeneration [21]. A recent study has reported an enhanced regenerative process after increasing p16INK4a+ mesenchymal stem cells during muscle inflammation [22]. Taken together, these recent findings suggest a new role of stress-induced p16INK4a protein expression in inflammation-mediated muscle regeneration. Rg1 is an immunostimulant that activates macrophage [11] and advances the progression of inflammation from M1 to M2 phenotype in exercised human skeletal muscle [23]. Therefore, pre-exercise Rg1 intake is likely to potentiate the inflammation response (preconditioning) and result in an early resolution of the exercise stress response.
M2 macrophage (CD163+) polarization occurs during the later period of inflammation, which is responsible for cell regeneration of muscle tissue after physical challenge [21, 24]. However, massive increases in p16INK4a+ endothelial progenitor cells observed in the study was not directly associated with an exercise-induced increment of M2 macrophage in skeletal muscle. The magnitude of increase in M2 macrophage was similar for PLA and Rg1 trials, while the exercise-induced response of p16INK4a+ cells in human skeletal muscle was lower than that in the PLA trial. However, our results do not preclude the possibility that accumulation of p16INK4a+ endothelial progenitor cells directly increase M2 macrophage activity in human skeletal muscle.
A limitation of the study is the difficulty in determining whether increased p16INK4a+ cells 3 h post-exercise was completely attributed cell senescence or simply due to a reversible induction of p16INK4a protein expression of well-functioned endothelial progenitor cells. Cellular senescence is featured by an irreversible form of cell-cycle arrest after prolonged stress. It is not possible to determine whether p16INK4a+ cell number is representing of irreversible cell-cycle arrest in human muscle tissue, since both p16INK4a+ cell accumulation and p16INK4a+ cell removal (immune clearance) can occur in the same muscle tissue following exercise. In this study, a moderate correlation between p16INK4a positive cells and ß-galactosidase positive cells in 36 muscle samples suggests that p16INK4a is not a perfect tissue senescence marker. This is in agreement with a previous study [25]. Whether p16INK4a is a reliable cell senescence marker in human tissues cannot be settled in this study.
Another limitation is inadequate time points for muscle biopsies, which prevented us from delineating the timings of the rise-and-fall pattern for p16INK4a+ cell accumulation in exercising muscle. The acute response of p16INK4a protein expression in endothelial cells of skeletal muscle after aerobic exercise is in sharp contrast to what has been observed after resistance exercise. We and others have previously shown decreased p16INK4a+ cells in muscle tissue of untrained active women and men after resistance training [5, 26]. Time required for resolution of the stress response and inflammation during and after exercise is associated with the magnitude of tissue damage, and varying by mode, intensity and duration of exercise. Furthermore, the local tissue response is influenced by distribution of the whole-body immune resource (white blood cells and stem cells from bone marrow). For example, aerobic exercise exerts a major challenge to the cardiopulmonary system in addition to skeletal muscle. The majority (> 60%) of white blood cells and stem cells are harbored in the lungs for constant local repair and regeneration [27, 28]. Aerobic exercise creates a competition between the lungs and muscle for immune resources for cell turnover [27]. In contrast, resistance exercise generates little challenge to the lungs, yet eccentric muscle contractions induce a substantial amount of muscle damage, which attracts more immune cells compared with aerobic exercise. Therefore, infiltration of immune cells into skeletal muscle after resistance exercise would be less likely to be compromised by competition with the lungs as occurs with aerobic exercise. Such differences can produce distinct rise-and-fall patterns for p16INK4a+ cells in human skeletal muscle [19, 29], and possibly explain the delayed response of p16INK4a+ cells in human skeletal muscle after aerobic exercise.
CONCLUSIONS
We observed a considerable increase in p16INK4a protein expression of endothelial progenitor cells in human skeletal muscle 3 h after aerobic exercise at 70% V̇O2max. The result of the study suggests that increased p16INK4a expression is a protective mechanism to maintain genetic stability of replicable cells during regenerative phase after aerobic stress. Early resolution of this stress response occurs when the immunostimulant Rg1 is orally taken 1 h before exercise.
MATERIALS AND METHODS
Participants
Twelve healthy young men 20-23 years of age (weight: 51-95 kg; height: 161-190 cm) with V̇O2max ranging between 43-55 ml−1 kg−1 min−1 volunteered to participate in this study. Participants were untrained and recreationally active non-smokers. They were fully informed of the risks and discomfort associated with the study, and all provided written consent before participation. This study was conducted in accordance with the guidelines contained in the Declaration of Helsinki and was approved by the Institutional Review Board of University of Taipei, Taipei, Taiwan.
Experimental design
A placebo-controlled, counter-balanced, crossover study was conducted. Participants were randomized into one of two groups: PLA (5 mg of cellulose) and Rg1 (5 mg). Rg1 was obtained from NuLiv Science, Inc. (Brea, CA, USA). Participants randomly assigned to the PLA group received cellulose before trial one, and Rg1 before trial two. Accordingly, participants randomly assigned to the Rg1 group received Rg1 before trial one and cellulose before trial two. Trials one and two were separated by ten days. The Rg1 and cellulose were provided in capsules 1 h before exercise. Capsules were coded by number for later identification. Exercise consisted of 1 h of continuous cycling at 70% V̇O2max on a cycle ergometer (Monark 839E, Stockholm, Sweden).
Experimental protocol
All participants were familiarized with the experimental procedures and equipment before testing started. Participants completed a V̇O2max test using a graded exercise protocol on a cycle ergometer one week before starting the experimental trials. All participants consumed a standard isocaloric diet 12 h prior to each experimental trial to limit any potential dietary effect that could influence the outcome of the study. Participants orally ingested 5 mg of Rg1 or PLA 1 h before cycling. After exercise, participants consumed a high carbohydrate meal (1.5 g carbohydrates per kg body weight: carbohydrate 80%, fat 8% and protein 12%; glycemic index: 80) within 10 min at the start of a 3-h recovery period. Water was provided ad libitum during and after the meal.
Muscle biopsy
Muscle samples were taken from vastus lateralis muscle before (Pre), immediately (0 h) and 3 h after the 1-h cycling exercise protocol. Biopsies were performed under local anesthesia (2% lidocaine) by a certified physician using an 18G Temno disposable cutting needle (Cardinal Health, Waukegan, IL, USA). Biopsies were taken from the vastus lateralis 3 cm in depth and 20 cm proximal to the knee. The baseline muscle biopsy (Pre) was conducted 3-4 weeks before the start of the first 1 h cycling exercise test. Two consecutive muscle biopsies were performed immediately (0 h) after and 3 h after each cycling test. The 0 h and 3 h biopsies were taken from opposite legs, but at the same position on the vastus lateralis. A portion of each muscle sample was immediately frozen in liquid nitrogen and stored at -70 °C until analyzed. The rest of the sample was immediately placed in a conical vial containing 10% formalin and used for immunohistochemical analysis. Paraffin-embedded tissue was sectioned no later than 3 h following muscle sample collection.
Immunohistochemistry
Serial sections of paraffin-embedded tissue were cut and analyzed for distribution of p16INK4a and CD34 in the vastus lateralis muscle. Paraffin sections (8 μm thick) were labeled using immunohistochemistry for binding of human monoclonal antibody p16INK4a (1:200, ab108349; Abcam, Cambridge, MA, USA) and CD34 (1:200, ab81289; Abcam, Cambridge, MA, USA). Immunofluorescence was used to detect regenerative macrophage CD163 (1:400, ab87099; Abcam, Cambridge, MA, USA) infiltration in the vastus lateralis after exercise. Optical images were analyzed using ImageJ (NIH, Bethesda, MD). Positive markers within cells were quantified and expressed as positive signal number/total skeletal muscle fiber number (%). An average of 600 muscle fibers per slide were included for analysis. All analyses were conducted by a specialist at the University of Taipei and a certified pathologist from Taipei Institute of Pathology with similar results. An additional 36 muscle biopsied samples were used to assess correlation between p16INK4a positive cells and ß-galactosidase positive cells (1: 150, NBP2-45731, Novus Biologicals, CO, USA).
Quantitative PCR
RNA was extracted from ~15 mg of skeletal muscle using TRI Reagent (T9424-200) (Sigma, St. Louis, MO, USA) for homogenization, followed by isopropanol precipitation, two ethanol washes, drying, and suspension in 20 μl nuclease-free water. One microgram of RNA in a total volume of 20 μl was reverse transcribed to cDNA using iScript cDNA Synthesis Kit (#170-8890) (Bio-Rad, Hercules, CA, USA). Real-time PCR was performed using MyiQ Single Color Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA), TaqMan Probe (Sigma-Aldrich, Singapore) and iQ Supermix kit (#170-8860) (Bio-Rad, Hercules, CA, USA). The PCR conditions for all genes consisted of one denaturing cycle at 90°C for 30 s, annealing at 60°C for 60 s and elongation at 72°C for 60 s. At the end of the PCR the samples were subjected to a melting curve analysis. To control for any variations due to efficiencies of the reverse transcription and PCR, 18S ribosomal RNA was used as an internal standard to determine relative expression levels of the target mRNAs. The primers and probes used to amplify target mRNA are 18S ribosomal (18S): Forward (5’-3’): ACAGGATTGACAGATTGATAGCTC, Reverse (5’-3’): TCGCTCCACCAACTAAGAACG, Probe (5’-3’): TGCACCACCACCCACGGAATCGAG; IL-10: Forward (5’-3’): CTTCCCTGTGAAAACAAG, Reverse (5’-3’): AGACCTCTAATTTATGTCCTA, Probe (5’-3’): AGTCGCCACCCTGATGTCTC; VEGF: Forward (5’-3’): TGAGATCGAGTACATCTTCAAGCC, Reverse (5’-3’): GGCCTTGGTGAGGTTTGATCC, Probe (5’-3’): CCTGTGTGCCCCTGATGCGATGCG; PGC-1α: Forward (5’-3’): CGAGGAATATCAGCACGAGAGG, Reverse (5’-3’): CATAAATCACACGGCGCTCTTC, Probe (5’-3’): TGCCTTCTGCCTCTGCCTCTCCCTC.
Serum LDH, Myoglobin and TBARS
A colorimetric assay kit was used to detect serum LDH activity according to the manufacturer’s instructions (Bio-Vision, # k726-500, CA, USA). Myoglobin was measured by ELISA using a commercially available kit (Immunology Consultants Laboratory, E-80MY, OR, USA). Serum samples were also used after further dilution for measurement of TBARS using a commercially available kit (Cayman Chemical, No. 10009055, MI, USA).
Statistical analysis
All data are expressed as means ± standard error. The data were analyzed using a two-factor repeated-measures ANOVA (SPSS 20.0). Post hoc paired comparison analysis was performed with the Fisher LSD method. Type I error of P ≤ 0.05 was considered significant. P ≤ 0.1 was considered moderately significant.
Footnotes
AUTHOR CONTRIBUTIONS: JW and CHK designed the experiments. JW, WHJ, CYH, TXYL, YYL, SS, and ISC performed the experiments. JW conducted the statistical analyses. JW and CHK wrote the manuscript. JLI help with the discussing, writing and editing of the manuscript. All authors read and approved the final manuscript.
CONFLICTS OF INTEREST: Chia-Hua Kuo and Jinfu Wu have submitted patents (Pub. No US2019/0374592 A1) for formulating senolytic ergogenic supplements with Nuliv Science related to the material studied. All other authors declare that they have no conflict of interest. The results of the study are presented clearly, honestly, and without fabrication, falsification, or inappropriate data manipulation. Publication fee is covered by funding agency listed above.
FUNDING: This study was sponsored by industrial research funds for product design from Joint fund of Nuliv Science Taiwan and Ministry of Science and Technology Taiwan (grant number: 106-2410-H-845), and the University of Taipei, Taiwan.
REFERENCES
- 1.Spalding KL, Bhardwaj RD, Buchholz BA, Druid H, Frisén J. Retrospective birth dating of cells in humans. Cell. 2005; 122:133–43. 10.1016/j.cell.2005.04.028 [DOI] [PubMed] [Google Scholar]
- 2.Erben RG, Odörfer KI, Siebenhütter M, Weber K, Rohleder S. Histological assessment of cellular half-life in tissues in vivo. Histochem Cell Biol. 2008; 130:1041–46. 10.1007/s00418-008-0470-3 [DOI] [PubMed] [Google Scholar]
- 3.Zampetaki A, Kirton JP, Xu Q. Vascular repair by endothelial progenitor cells. Cardiovasc Res. 2008; 78:413–21. 10.1093/cvr/cvn081 [DOI] [PubMed] [Google Scholar]
- 4.Zheng B, Cao B, Crisan M, Sun B, Li G, Logar A, Yap S, Pollett JB, Drowley L, Cassino T, Gharaibeh B, Deasy BM, Huard J, Péault B. Prospective identification of myogenic endothelial cells in human skeletal muscle. Nat Biotechnol. 2007; 25:1025–34. 10.1038/nbt1334 [DOI] [PubMed] [Google Scholar]
- 5.Yang C, Jiao Y, Wei B, Yang Z, Wu JF, Jensen J, Jean WH, Huang CY, Kuo CH. Aged cells in human skeletal muscle after resistance exercise. Aging (Albany NY). 2018; 10:1356–65. 10.18632/aging.101472 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sharpless NE. Ink4a/arf links senescence and aging. Exp Gerontol. 2004; 39:1751–59. 10.1016/j.exger.2004.06.025 [DOI] [PubMed] [Google Scholar]
- 7.Sarkar-Agrawal P, Vergilis I, Sharpless NE, DePinho RA, Rünger TM. Impaired processing of DNA photoproducts and ultraviolet hypermutability with loss of p16INK4a or p19ARF. J Natl Cancer Inst. 2004; 96:1790–93. 10.1093/jnci/djh307 [DOI] [PubMed] [Google Scholar]
- 8.Tryfidou DV, McClean C, Nikolaidis MG, Davison GW. DNA damage following acute aerobic exercise: a systematic review and meta-analysis. Sports Med. 2020; 50:103–27. 10.1007/s40279-019-01181-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ashcraft KA, Peace RM, Betof AS, Dewhirst MW, Jones LW. Efficacy and mechanisms of aerobic exercise on cancer initiation, progression, and metastasis: a critical systematic review of in vivo preclinical data. Cancer Res. 2016; 76:4032–50. 10.1158/0008-5472.CAN-16-0887 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Burton DG, Stolzing A. Cellular senescence: immunosurveillance and future immunotherapy. Ageing Res Rev. 2018; 43:17–25. 10.1016/j.arr.2018.02.001 [DOI] [PubMed] [Google Scholar]
- 11.Fan ZH, Isobe K, Kiuchi K, Nakashima I. Enhancement of nitric oxide production from activated macrophages by a purified form of ginsenoside (Rg1). Am J Chin Med. 1995; 23:279–87. 10.1142/S0192415X9500033X [DOI] [PubMed] [Google Scholar]
- 12.Joo SS, Won TJ, Lee DI. Reciprocal activity of ginsenosides in the production of proinflammatory repertoire, and their potential roles in neuroprotection in vivo. Planta Med. 2005; 71:476–81. 10.1055/s-2005-864145 [DOI] [PubMed] [Google Scholar]
- 13.Guo X, Zhang J, Liu M, Zhao GC. Protective effect of ginsenoside Rg1 on attenuating anti-GBM glomerular nephritis by activating NRF2 signalling. Artif Cells Nanomed Biotechnol. 2019; 47:2972–79. 10.1080/21691401.2019.1640712 [DOI] [PubMed] [Google Scholar]
- 14.Tsai HH, Lin CP, Lin YH, Hsu CC, Wang JS. High-intensity interval training enhances mobilization/functionality of endothelial progenitor cells and depressed shedding of vascular endothelial cells undergoing hypoxia. Eur J Appl Physiol. 2016; 116:2375–88. 10.1007/s00421-016-3490-z [DOI] [PubMed] [Google Scholar]
- 15.Urbich C, Dimmeler S. Endothelial progenitor cells: characterization and role in vascular biology. Circ Res. 2004; 95:343–53. 10.1161/01.RES.0000137877.89448.78 [DOI] [PubMed] [Google Scholar]
- 16.Wu J, Saovieng S, Cheng IS, Liu T, Hong S, Lin CY, Su IC, Huang CY, Kuo CH. Ginsenoside Rg1 supplementation clears senescence-associated β-galactosidase in exercising human skeletal muscle. J Ginseng Res. 2019; 43:580–88. 10.1016/j.jgr.2018.06.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Serrano M. Senescence helps regeneration. Dev Cell. 2014; 31:671–72. 10.1016/j.devcel.2014.12.007 [DOI] [PubMed] [Google Scholar]
- 18.Kay MM. Mechanism of removal of senescent cells by human macrophages in situ. Proc Natl Acad Sci USA. 1975; 72:3521–25. 10.1073/pnas.72.9.3521 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Prata LG, Ovsyannikova IG, Tchkonia T, Kirkland JL. Senescent cell clearance by the immune system: emerging therapeutic opportunities. Semin Immunol. 2018; 40:101275. 10.1016/j.smim.2019.04.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sagiv A, Biran A, Yon M, Simon J, Lowe SW, Krizhanovsky V. Granule exocytosis mediates immune surveillance of senescent cells. Oncogene. 2013; 32:1971–77. 10.1038/onc.2012.206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tidball JG. Regulation of muscle growth and regeneration by the immune system. Nat Rev Immunol. 2017; 17:165–78. 10.1038/nri.2016.150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chikenji TS, Saito Y, Konari N, Nakano M, Mizue Y, Otani M, Fujimiya M. p16INK4A-expressing mesenchymal stromal cells restore the senescence-clearance-regeneration sequence that is impaired in chronic muscle inflammation. EBioMedicine. 2019; 44:86–97. 10.1016/j.ebiom.2019.05.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hou CW, Lee SD, Kao CL, Cheng IS, Lin YN, Chuang SJ, Chen CY, Ivy JL, Huang CY, Kuo CH. Improved inflammatory balance of human skeletal muscle during exercise after supplementations of the ginseng-based steroid Rg1. PLoS One. 2015; 10:e0116387. 10.1371/journal.pone.0116387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wang Y, Wehling-Henricks M, Samengo G, Tidball JG. Increases of M2a macrophages and fibrosis in aging muscle are influenced by bone marrow aging and negatively regulated by muscle-derived nitric oxide. Aging Cell. 2015; 14:678–88. 10.1111/acel.12350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dungan CM, Peck BD, Walton RG, Huang Z, Bamman MM, Kern PA, Peterson CA. In vivo analysis of γH2AX+ cells in skeletal muscle from aged and obese humans. FASEB J. 2020; 34:7018–35. 10.1096/fj.202000111RR [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Justice JN, Gregory H, Tchkonia T, LeBrasseur NK, Kirkland JL, Kritchevsky SB, Nicklas BJ. Cellular senescence biomarker p16INK4a+ cell burden in thigh adipose is associated with poor physical function in older women. J Gerontol A Biol Sci Med Sci. 2018; 73:939–45. 10.1093/gerona/glx134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Adams GR, Zaldivar FP, Nance DM, Kodesh E, Radom-Aizik S, Cooper DM. Exercise and leukocyte interchange among central circulation, lung, spleen, and muscle. Brain Behav Immun. 2011; 25:658–66. 10.1016/j.bbi.2011.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Rochefort GY, Vaudin P, Bonnet N, Pages JC, Domenech J, Charbord P, Eder V. Influence of hypoxia on the domiciliation of mesenchymal stem cells after infusion into rats: possibilities of targeting pulmonary artery remodeling via cells therapies? Respir Res. 2005; 6:125. 10.1186/1465-9921-6-125 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Chang J, Wang Y, Shao L, Laberge RM, Demaria M, Campisi J, Janakiraman K, Sharpless NE, Ding S, Feng W, Luo Y, Wang X, Aykin-Burns N, et al. Clearance of senescent cells by ABT263 rejuvenates aged hematopoietic stem cells in mice. Nat Med. 2016; 22:78–83. 10.1038/nm.4010 [DOI] [PMC free article] [PubMed] [Google Scholar]