

Abstract
Sugar transporter (SUT) genes are associated with multiple physiological and biochemical processes in filamentous fungi, such as the response to various stresses. However, limited systematic analysis and functional information of SUT gene family have been available on Aspergillus oryzae (A. oryzae). To investigate the potential roles of SUTs in A. oryzae, we performed an integrative analysis of the SUT gene family in this study. Based on the conserved protein domain search, 127 putative SUT genes were identified in A. oryzae and further categorized into eight distinct subfamilies. The result of gene structure and conserved motif analysis illustrated functional similarities among the AoSUT proteins within the same subfamily. Additionally, expression profiles of the AoSUT genes at different growth stages elucidated that most of AoSUT genes have high expression levels at the stationary phase while low in the adaptive phase. Furthermore, expression profiles of AoSUT genes under salt stress showed that AoSUT genes may be closely linked to salt tolerance and involved in sophisticated transcriptional process. The protein-protein interaction network of AoSUT propounded some potentially interacting proteins. A comprehensive overview of the AoSUT gene family will offer new insights into the structural and functional features as well as facilitate further research on the roles of AoSUT genes in response to abiotic stresses.
1. Introduction
Aspergillus oryzae has been routinely used in the production of traditional fermented food such as soy sauce, miso, vinegar, and fermented soybean paste in Asian countries for centuries [1]. A. oryzae has been generally listed as “Generally Recognized as Safe (GRAS)” by the Food and Drug Administration (FDA) in the USA, and its safety was recognized by the World Health Organization (WHO) [2]. A. oryzae also has a prominent ability of secreting various hydrolases, liking amylase, protease, and lipase, to degrade large molecules such as polysaccharides and proteins into relatively low-molecular-weight metabolites, including smaller monosaccharides and amino acids [3]. Furthermore, it serves as a source of secondary volatile metabolites that contribute to the quality of fermented products; thus, it is also frequently expected as hosts for heterologous production of useful secondary metabolites [4–7].
Sugars are not only primary nutrients and structural precursors but also signaling molecules that trigger changes in gene regulation for signal transduction and stress responses [8–10]. In fungi, the function of sugar as signaling molecules was mainly achieved through sugar transport and sensing. Considerable researches focused on the transportation and utilization of sugar, such as the absorption and utilization of xylose and glucose [11, 12]. Meanwhile, accumulating researches paid much attention to the ability of stress response of SUT. For example, different SUT families in Aspergillus niger (A. niger) have a dynamic expression pattern during growth on distinct carbon sources and hence enable the organism to adapt to an extremely broad range of environments [13]. Contrary to A. niger, in response to different carbon sources, Cordyceps militaris (C. militaris) SUT genes within each subfamily harbor an analogous expression pattern with the exception of those involved in the pentose subfamily, which were highly expressed in the xylose culture [14]. Vankuyk et al. reported that regulators CreA regulated the expression of SUT genes in A. niger in response to extracellular pH changes [15]. In Arabidopsis, overexpression of rice monosaccharide transporter gene (OsMST6) could enhance the tolerance to the salinity [16]. In insects, sugar transporters might also act as receivers for virus entry. It was constitutively expressed in susceptible silkworm, thus leading to the host yield to viral infection [17]. What is more, in response to different stocking densities, intestinal section-dependent expression patterns of the three transporter genes (glut2, sglt1, and sglt4) in grass carp (Ctenopharyngodon idellus) did not alter. Nevertheless, high stocking density led to significant differences in segmental mRNA expression levels [18].
Whole transcriptome sequencing gives a valuable alternative to genome-wide sequencing for gene mining and functional identification [19]. It has enabled us to easily gain long-read or full-length transcripts with complete coding sequences (CDS) and characterization of gene families [20]. These approaches have become a primary strategy in characterizing gene expression profiles and elucidating the genetic networks in all fields, such as animals [17, 18], plants [21–23], and fungi [13, 24–26]. However, just a few of the SUTs have been reported in fungi, such as trehalose transporter in Beauveria bassiana (BbAGT1), putative pentose transporter in C. militaris (CCM_06358), and raffinose transporter in Metarhizium robertsii (Mrt) [14, 27, 28]. Nonetheless, there is no systematic analysis and characterization of SUT family genes in A. oryzae. Thus, in the present study, we performed the genome-wide identification of evolutionary, gene structures and basic physicochemical properties of AoSUT genes. Furthermore, expression profiles of AoSUT genes at different stages of development stage and in response to salt stress treatment were analyzed using the RNA-Seq data, in an attempt to understand their possible roles in salt stress. The findings of the current investigations will serve as a reference of sugar regulation mechanism for other related Aspergillus.
2. Materials and Methods
2.1. Identification of Sugar Transporter Genes and Physicochemical Properties
The genomic, CDS, and protein sequences of A. oryzae RIB40 were downloaded from the Aspergillus Genome Database (AspGD, http://www.aspergillusgenome.org/). The sugar transporter protein sequences of Saccharomyces cerevisiae (ScSUT) were retrieved from the Saccharomyces Genome Database (SGD, https://www.yeastgenome.org/). ScSUT proteins were employed as query sequences in a Base Local Alignment Search Tool (BLAST) [29]. To ensure the selected sequences were nonredundant sequences and the presence of the Sugar_tr domain (PF00083) in each candidate AoSUT protein, we downloaded the Sugar_tr domain from the Pfam database [30] (http://pfam.sanger.ac.uk/search). It was used to search the A. oryzae RIB40 protein database by HMMER software with a standard E value < 1 × 10−5 [31]. The Sequence Manipulation Suite online tool (http://www.detaibio.com/sms2/protein_iep.html) was used to estimate the basic physicochemical properties of the protein, such as the isoelectric point and molecular weight of the gene product for each member of A. oryzae sugar transporter gene family. Thereafter, the subcellular localization of sugar transporter genes was predicted by two online analysis tools, such as WoLF PSORT Prediction (https://wolfpsort.hgc.jp) and LocTree3 Prediction system (https://rostlab.org/services/loctree3/). The results were confirmed by more than one method.
2.2. Multiple Sequence Alignment and Phylogenetic Analysis
Multiple sequence alignments and phylogenetic analysis of sugar transporter proteins in A. oryzae were subjected to MEGA X [32]. Alignments were performed using the Muscle program with default parameters, and the results after that were subjected to construct unrooted phylogenetic tree with maximum likelihood (ML) using pairwise deletion option and Poisson correction model. To further verify the reliability phylogenetic tree, the Neighbor-Joining (NJ) tree was constructed with the Jones–Taylor–Thornton (JTT) model. Both of the phylogenetic trees were bootstrap value set as 1000 replications in order to choose the best result. The unrooted tree was visualized by the online tool Interactive Tree Of Life (https://itol.embl.de/).
2.3. Analysis of Conserved Motifs and Gene Structures
To obtain insights into the diversity of motif compositions in AoSUT proteins, SUT protein sequences were subjected to the MEME (version 5.1.1, http://meme-suite.org/tools/meme) [33]. It is based on the maximum expectation (EM) algorithm, which alternates the execution of steps E (expectation) and M (maximum). Meanwhile, motif sequences and annotations were further predicted by Pfam. MEME using the default parameters with the exception that maximum number of motifs set to 6 and optimum width was from 6 to 50. Following that, the results are visualized by the TBtools software.
To apprehend better the structure of each SUT gene, CDSs and corresponding genomic sequences of AoSUT were downloaded from the Aspergillus Genome Database. Gene Structure Display Server 2.0 (GSDS 2.0, http://gsds.cbi.pku.edu.cn/) was used to generate a schematic diagram of exon-intron organization by comparison of the genomic and CDS sequences of each AoSUT gene.
2.4. Analysis of Chromosomal Locations
To determine the chromosomal locations of all SUT genes in A. oryzae, the information of locus coordinates was obtained from the genomic sequences. The MapChart software was used for the mapping of AoSUT genes' chromosomal positions and relative distances on the basis of their ascending order of physical position (bp).
2.5. Expression Analysis of AoSUT Genes in Different Growth Stages and Salt Stress Treatments
The genome-wide transcriptome data of A. oryzae in different growth stages and salt stress treatments were obtained from NCBI SRA databases under BioProject Accession PRJNA407002 and PRJNA383095. The raw reads that contained adapter sequence or comprised of more than 10% unknown nucleotide base calls (N) were removed. Gene expression levels were normalized using the FPKM (Fragments Per Kilobase of transcript per Million mapped reads) method. According to the GeneID of AoSUT genes in A. oryzae expressed transcripts, the expression data of these genes in three different stages (24, 48, and 72 h, corresponding to the adaptive phase, logarithmic phase, and stationary phase) and four salt stress treatments (cultivated in potato dextrose agar medium supplied with 0, 5, 10, and 15% NaCl) were obtained. The expression profiles were displayed in a heat map generated with the heat map Illustrator software (v 1.0.3.7) by the default data normalization parameter and clustering method [34].
To further ascertain the expression profiles of SUT genes in A. oryzae, quantitative real-time RT-PCR (qRT-PCR) experiments of six selected genes (including AoSUT1, 5, 12, 23, 35, and 69) were performed. The genome-wide transcriptome data of A. oryzae at three growth stages and four salt stress treatments were obtained from databases. Specifically, three growth stages represent the adaptive, logarithmic, and stationary phases, separately. Four salt stress treatments are equivalent to control, slight stress, moderate stress, and severe stress, respectively. All the transcriptome data are the average of three biological replicates. Total RNA of all collected samples was extracted using the TRIzol Reagent (Takara, Beijing, China) following the instructions in our previous studies [35]. CFX96 Real-Time PCR Detection System (Bio-Rad, CA, USA) was used to perform the qRT-PCR analysis. The specific primers of sugar transporter genes in A. oryzae used for qRT-PCR were listed in Table S1. The reaction conditions were 30 s at 94°C, 45 cycles of 20 s at 94°C 20 s at 55°C, and 30 s at 72°C. The melting curves were analyzed from 60°C to 95°C to observe the specificity of the PCR products. The comparative 2−ΔΔCT method was employed to calculate the relative expression between samples [36].
2.6. Analysis of the Protein-Protein Interaction Network
Protein-protein interaction (PPI) data were gained from the online database of STRING (https://string-db.org/), which is an open source software interface for the exploration, analysis, and visualization of protein-protein associations. These interactions are obtained from text mining of literature of experimental validation including physical interactions and enzymatic reactions found in signal transduction pathways, high-throughput experimental data, and computational predictions containing those based on genomic context analysis and also derived from analyses of coexpressed genes [37]. The PPI data were preprocessed including removing redundancy and self-loops. Targets with a high confidence score > 0.7 were selected to construct the PPI networks. The PPI networks were visualized in Gephic software with the nodes representing proteins/genes and the edges representing interactions between any two proteins/genes.
2.7. Statistical Analysis
All values obtained from the studies were presented as mean ± SE. Data from the same period were evaluated by one-way nested analysis of variance (ANOVA), followed by least significant difference test (LSD) for mean comparison. All statistical analysis was performed with SAS 9.20 software (SAS Institute Inc., Cary, North Carolina, USA). Values with p < 0.05 were considered significant.
3. Results and Discussion
3.1. Genome-Wide Identification of SUT Gene Family in A. oryzae RIB40
Through a BLAST search and HMMER analysis, eventually, a total of 127 putative SUT genes in the A. oryzae RIB40 genome were identified and named based on their chromosomal locations (Table 1). The length of 127 AoSUT proteins varied from 115 (AoSUT102) to 767 (AoSUT69) amino acid residues, and the number of transmembrane domains ranged from 2 (AoSUT102, AoSUT117) to 12 (AoSUT3, AoSUT14, AoSUT15, etc.).
Table 1.
The member of SUT gene family in A. oryzae.
| Nomenclature | AspGeneID | CDS length (bp) | PL (aa) | PI | MW (kDa) | TMD | SCLpred |
|---|---|---|---|---|---|---|---|
| AoSUT1 | ASPL0000299362 | 1626 | 541 | 6.12 | 60.46 | 10 | Plas |
| AoSUT2 | ASPL0000348493 | 1596 | 531 | 8.24 | 58.64 | 10 | Plas |
| AoSUT3 | ASPL0000312952 | 1575 | 524 | 8.96 | 58.05 | 12 | Plas |
| AoSUT4 | ASPL0000330204 | 1590 | 529 | 8.64 | 58.34 | 12 | Plas |
| AoSUT5 | ASPL0000321372 | 1512 | 503 | 9.01 | 54.12 | 12 | Plas |
| AoSUT6 | ASPL0000298924 | 1635 | 544 | 7.06 | 60.5 | 11 | Vacu |
| AoSUT7 | ASPL0000350553 | 1608 | 535 | 8.15 | 59.69 | 12 | Plas |
| AoSUT8 | ASPL0000291308 | 1359 | 452 | 7.9 | 50.65 | 8 | Plas |
| AoSUT9 | ASPL0000301060 | 1533 | 510 | 9.28 | 55.97 | 10 | Plas |
| AoSUT10 | ASPL0000309712 | 1653 | 550 | 5.56 | 60.88 | 10 | Plas |
| AoSUT11 | ASPL0000344279 | 1899 | 632 | 7.22 | 71.65 | 10 | Plas |
| AoSUT12 | ASPL0000349681 | 1527 | 508 | 6.84 | 55.73 | 11 | Plas |
| AoSUT13 | ASPL0000301678 | 1563 | 520 | 7.29 | 57.38 | 12 | Plas |
| AoSUT14 | ASPL0000315710 | 1611 | 536 | 6.71 | 59.68 | 12 | Plas |
| AoSUT15 | ASPL0000287472 | 1755 | 584 | 8.59 | 62.91 | 12 | Plas |
| AoSUT16 | ASPL0000281504 | 1593 | 530 | 8.28 | 57.28 | 12 | Plas |
| AoSUT17 | ASPL0000337017 | 1629 | 542 | 8.18 | 59.11 | 12 | Plas |
| AoSUT18 | ASPL0000349981 | 1623 | 540 | 7.89 | 59.38 | 12 | Plas |
| AoSUT19 | ASPL0000323998 | 1623 | 540 | 7.49 | 60.5 | 8 | Plas |
| AoSUT20 | ASPL0000284520 | 1734 | 577 | 7.46 | 62.29 | 12 | Plas |
| AoSUT21 | ASPL0000322602 | 1602 | 533 | 6.66 | 59.46 | 9 | Plas |
| AoSUT22 | ASPL0000325484 | 1590 | 529 | 7.68 | 58.88 | 12 | Plas |
| AoSUT23 | ASPL0000294580 | 1710 | 569 | 7.99 | 63.22 | 11 | Vacu |
| AoSUT24 | ASPL0000288152 | 1626 | 541 | 9.28 | 59.47 | 12 | Mito |
| AoSUT25 | ASPL0000309860 | 1605 | 534 | 7.63 | 58.9 | 12 | Plas |
| AoSUT26 | ASPL0000279704 | 1464 | 487 | 7.08 | 53.42 | 11 | ER |
| AoSUT27 | ASPL0000335285 | 1485 | 494 | 8.91 | 54.07 | 11 | Plas |
| AoSUT28 | ASPL0000313172 | 1671 | 556 | 4.98 | 61.52 | 12 | Plas |
| AoSUT29 | ASPL0000283168 | 1299 | 432 | 8.89 | 46.44 | 12 | Plas |
| AoSUT30 | ASPL0000281742 | 1797 | 598 | 8.44 | 66.54 | 12 | Plas |
| AoSUT31 | ASPL0000324124 | 1515 | 504 | 7.88 | 54.98 | 11 | Plas |
| AoSUT32 | ASPL0000327738 | 1446 | 481 | 9.04 | 53.16 | 12 | Plas |
| AoSUT33 | ASPL0000334740 | 1506 | 501 | 6.51 | 54.28 | 11 | Plas |
| AoSUT34 | ASPL0000280136 | 1503 | 500 | 6.25 | 54.06 | 12 | Plas |
| AoSUT35 | ASPL0000300822 | 1575 | 524 | 6.78 | 58.61 | 12 | Plas |
| AoSUT36 | ASPL0000349699 | 1527 | 508 | 6.58 | 55.65 | 10 | Plas |
| AoSUT37 | ASPL0000338681 | 1671 | 556 | 8.11 | 61.07 | 12 | Plas |
| AoSUT38 | ASPL0000283036 | 1545 | 514 | 6.99 | 56.61 | 12 | Plas |
| AoSUT39 | ASPL0000289632 | 1545 | 514 | 6.36 | 56.07 | 10 | Plas |
| AoSUT40 | ASPL0000304806 | 1650 | 549 | 6.97 | 60.71 | 10 | Plas |
| AoSUT41 | ASPL0000340951 | 1578 | 525 | 6.24 | 57.49 | 10 | Plas |
| AoSUT42 | ASPL0000295608 | 1602 | 533 | 6.5 | 58.35 | 10 | Plas |
| AoSUT43 | ASPL0000337719 | 1680 | 559 | 6.08 | 61.71 | 10 | Mito |
| AoSUT44 | ASPL0000282628 | 1824 | 607 | 8.58 | 68.4 | 9 | Plas |
| AoSUT45 | ASPL0000346097 | 1593 | 530 | 9.61 | 58.02 | 12 | Plas |
| AoSUT46 | ASPL0000299188 | 1542 | 513 | 7.78 | 56.59 | 10 | Plas |
| AoSUT47 | ASPL0000294920 | 1671 | 556 | 8.85 | 60.69 | 12 | Plas |
| AoSUT48 | ASPL0000341497 | 1644 | 547 | 6.11 | 59.38 | 12 | Plas |
| AoSUT49 | ASPL0000284342 | 1809 | 602 | 9.08 | 67.21 | 12 | Plas |
| AoSUT50 | ASPL0000322158 | 1629 | 542 | 4.87 | 58.75 | 12 | Plas |
| AoSUT51 | ASPL0000327028 | 1617 | 538 | 7.03 | 59.01 | 12 | Plas |
| AoSUT52 | ASPL0000345761 | 1680 | 559 | 6.58 | 61.86 | 12 | Mito |
| AoSUT53 | ASPL0000288766 | 1842 | 613 | 7.26 | 67.86 | 12 | Plas |
| AoSUT54 | ASPL0000348009 | 1536 | 511 | 7.46 | 55.9 | 10 | Plas |
| AoSUT55 | ASPL0000308756 | 1578 | 525 | 7.78 | 56.95 | 9 | Plas |
| AoSUT56 | ASPL0000288748 | 1614 | 537 | 8.16 | 59.3 | 12 | Plas |
| AoSUT57 | ASPL0000342947 | 1521 | 506 | 6.74 | 56.62 | 12 | Plas |
| AoSUT58 | ASPL0000289192 | 546 | 181 | 8.33 | 19.9 | 4 | Plas |
| AoSUT59 | ASPL0000302542 | 1674 | 557 | 6.82 | 60.12 | 11 | Pero |
| AoSUT60 | ASPL0000297440 | 1623 | 540 | 6.31 | 57.65 | 12 | Plas |
| AoSUT61 | ASPL0000301470 | 1710 | 569 | 5.37 | 63.49 | 11 | Vacu |
| AoSUT62 | ASPL0000317896 | 1830 | 609 | 7.54 | 70.1 | 12 | Plas |
| AoSUT63 | ASPL0000305684 | 1587 | 528 | 8.63 | 58.55 | 12 | Plas |
| AoSUT64 | ASPL0000290346 | 1548 | 515 | 7.03 | 56.35 | 12 | Plas |
| AoSUT65 | ASPL0000290728 | 1548 | 515 | 6.29 | 56.95 | 10 | Plas |
| AoSUT66 | ASPL0000314400 | 1476 | 491 | 7.77 | 54.85 | 12 | Plas |
| AoSUT67 | ASPL0000307186 | 1467 | 488 | 9.14 | 51.94 | 12 | Vacu |
| AoSUT68 | ASPL0000322566 | 1488 | 495 | 8.03 | 55.42 | 12 | Plas |
| AoSUT69 | ASPL0000284218 | 2304 | 767 | 7.26 | 84.61 | 9 | Plas |
| AoSUT70 | ASPL0000291632 | 1617 | 538 | 6.76 | 59.05 | 12 | Vacu |
| AoSUT71 | ASPL0000316294 | 1572 | 523 | 5.71 | 57.42 | 12 | Plas |
| AoSUT72 | ASPL0000340407 | 1590 | 529 | 8.1 | 59.65 | 10 | Vacu |
| AoSUT73 | ASPL0000321912 | 1542 | 513 | 6.18 | 56.75 | 10 | Plas |
| AoSUT74 | ASPL0000327626 | 1620 | 539 | 6.6 | 58.86 | 12 | Plas |
| AoSUT75 | ASPL0000291430 | 1653 | 550 | 6.28 | 60.89 | 11 | Plas |
| AoSUT76 | ASPL0000330168 | 1641 | 546 | 8.88 | 59.22 | 12 | Plas |
| AoSUT77 | ASPL0000299472 | 1638 | 545 | 8.61 | 58.72 | 11 | Plas |
| AoSUT78 | ASPL0000348593 | 1566 | 521 | 6.5 | 56.46 | 12 | Plas |
| AoSUT79 | ASPL0000304244 | 1413 | 470 | 6.5 | 50.61 | 9 | Plas |
| AoSUT80 | ASPL0000322394 | 1482 | 493 | 8.62 | 53.88 | 10 | Mito |
| AoSUT81 | ASPL0000308688 | 1581 | 526 | 9.36 | 58.33 | 9 | Plas |
| AoSUT82 | ASPL0000348347 | 1521 | 506 | 7.37 | 55.82 | 12 | Plas |
| AoSUT83 | ASPL0000296900 | 1629 | 542 | 8.81 | 59.7 | 10 | Plas |
| AoSUT84 | ASPL0000327800 | 1593 | 530 | 6.7 | 57.98 | 12 | Plas |
| AoSUT85 | ASPL0000308836 | 1629 | 542 | 5.5 | 59.45 | 12 | Plas |
| AoSUT86 | ASPL0000284922 | 1614 | 537 | 7.22 | 59.13 | 12 | Plas |
| AoSUT87 | ASPL0000306294 | 1641 | 546 | 6.01 | 60.5 | 9 | Plas |
| AoSUT88 | ASPL0000310270 | 1611 | 536 | 6.98 | 59.53 | 10 | Plas |
| AoSUT89 | ASPL0000299794 | 1698 | 565 | 6.75 | 61.76 | 12 | Plas |
| AoSUT90 | ASPL0000343545 | 1788 | 595 | 9.47 | 66.32 | 12 | Mito |
| AoSUT91 | ASPL0000300438 | 1485 | 494 | 8.37 | 54.34 | 10 | Plas |
| AoSUT92 | ASPL0000336593 | 1542 | 513 | 6.39 | 56.75 | 12 | Plas |
| AoSUT93 | ASPL0000344697 | 1626 | 541 | 6.27 | 60.41 | 11 | Vacu |
| AoSUT94 | ASPL0000326072 | 1644 | 547 | 7.41 | 59.46 | 12 | Plas |
| AoSUT95 | ASPL0000304476 | 1614 | 537 | 8.81 | 59.73 | 12 | Plas |
| AoSUT96 | ASPL0000298998 | 1506 | 501 | 6.63 | 56.33 | 9 | Plas |
| AoSUT97 | ASPL0000307814 | 1614 | 537 | 7.49 | 60.39 | 11 | Plas |
| AoSUT98 | ASPL0000309592 | 1698 | 565 | 7.75 | 62.88 | 12 | Plas |
| AoSUT99 | ASPL0000281934 | 1629 | 542 | 5.44 | 59.99 | 10 | Plas |
| AoSUT100 | ASPL0000287394 | 1938 | 645 | 6.63 | 70.99 | 12 | Plas |
| AoSUT101 | ASPL0000289388 | 1689 | 562 | 6.08 | 62.31 | 12 | Plas |
| AoSUT102 | ASPL0000342469 | 348 | 115 | 9.15 | 13.21 | 2 | Mito |
| AoSUT103 | ASPL0000331538 | 1887 | 628 | 7.91 | 70.76 | 11 | Vacu |
| AoSUT104 | ASPL0000342747 | 1614 | 537 | 8.23 | 58.96 | 12 | Plas |
| AoSUT105 | ASPL0000300304 | 1662 | 553 | 7.77 | 62.09 | 12 | Mito |
| AoSUT106 | ASPL0000294524 | 1587 | 528 | 8.3 | 58.1 | 12 | Plas |
| AoSUT107 | ASPL0000295892 | 1665 | 554 | 6.87 | 61.72 | 12 | Plas |
| AoSUT108 | ASPL0000335375 | 1713 | 570 | 6.65 | 63.51 | 10 | Plas |
| AoSUT109 | ASPL0000333426 | 2004 | 667 | 7.33 | 75.14 | 9 | ER |
| AoSUT110 | ASPL0000313766 | 1554 | 517 | 6.74 | 57.24 | 11 | Plas |
| AoSUT111 | ASPL0000328104 | 1539 | 512 | 9.42 | 55.9 | 10 | Vacu |
| AoSUT112 | ASPL0000334670 | 1461 | 486 | 6.52 | 53.39 | 12 | Plas |
| AoSUT113 | ASPL0000340285 | 1584 | 527 | 6.31 | 57.33 | 12 | Plas |
| AoSUT114 | ASPL0000324582 | 1452 | 483 | 6.15 | 53.48 | 12 | Plas |
| AoSUT115 | ASPL0000284234 | 1872 | 623 | 7.23 | 70.73 | 8 | Plas |
| AoSUT116 | ASPL0000322906 | 1659 | 552 | 6.87 | 60.77 | 10 | Plas |
| AoSUT117 | ASPL0000323794 | 759 | 252 | 8.69 | 29.14 | 2 | Plas |
| AoSUT118 | ASPL0000332556 | 1596 | 531 | 8.49 | 59.42 | 12 | Plas |
| AoSUT119 | ASPL0000315516 | 1599 | 532 | 7.97 | 58.21 | 11 | Plas |
| AoSUT120 | ASPL0000348251 | 1596 | 531 | 8.45 | 58.98 | 9 | Plas |
| AoSUT121 | ASPL0000282924 | 1527 | 508 | 6.79 | 55.12 | 12 | Plas |
| AoSUT122 | ASPL0000342577 | 1614 | 537 | 5.94 | 59.11 | 10 | Plas |
| AoSUT123 | ASPL0000324822 | 1620 | 539 | 5.54 | 60.26 | 11 | Plas |
| AoSUT124 | ASPL0000301868 | 1701 | 566 | 9.31 | 63.52 | 12 | Plas |
| AoSUT125 | ASPL0000316738 | 1641 | 546 | 6.51 | 60.49 | 12 | Mito |
| AoSUT126 | ASPL0000349221 | 1536 | 511 | 7.85 | 56.35 | 12 | Mito |
| AoSUT127 | ASPL0000331760 | 1749 | 582 | 8.56 | 64.2 | 12 | Plas |
PL: protein length; MW: molecular weight; pI: isoelectric point; TMD: transmembrane domain number; SCLpred: predicted subcellular localization; Plas: plasma membrane; Vacu: vacuolar membrane; Mito: mitochondrion; ER: endoplasmic reticulum; Pero: peroxisome membrane.
The basic physicochemical properties of 127 AoSUT were estimated by the Sequence Manipulation Suite online tool (Table 1). From the results obtained, the molecular weight of these AoSUT proteins ranged from 13.21 (AoSUT102) to 84.61 (AoSUT69) kDa. What is more, half of the AoSUT proteins exhibited alkaline isoelectric points greater than 7.41, with the highest being 9.61 (AoSUT45), while 25 proteins had acidic isoelectric points of less than 6.5, of which AoSUT50 was the lowest at 4.87. Also, a few proteins, such as AoSUT26, AoSUT38, and AoSUT64, had relatively neutral isoelectric points that fell between 6.5 and 7.41.
WoLF PSORT Prediction and LocTree3 Prediction system were performed to precisely predict the subcellular localization for the products of the AoSUT gene family. Based on the predicted results, overwhelming majority of AoSUT proteins were located in the plasma membrane, demonstrating that these proteins as transcription factors play a transcriptional regulatory role directly in the plasma membrane.
3.2. Classification and Phylogenetic Analysis of AoSUT Gene Families
In order to better understand the genetic relationship within members of the SUT gene family in A. oryzae, the amino acid sequences of 127 AoSUT were used to construct a maximum likelihood phylogenetic tree. As shown in Figure 1, the AoSUT genes (except AoSUT2, AoSUT5, AoSUT67, AoSUT23, and AoSUT69) were classified into eight different clades according to the research of de Vries et al. [10], which include clades A-H. Additionally, the Neighbor-Joining (NJ) tree of SUT genes with 1000 repeat values displayed a topology which was analogous to ML topology (Figure S1). There were 32 members in the clade H subfamily, which was the largest subfamily, while only 8 members grouped in clades B and C.
Figure 1.
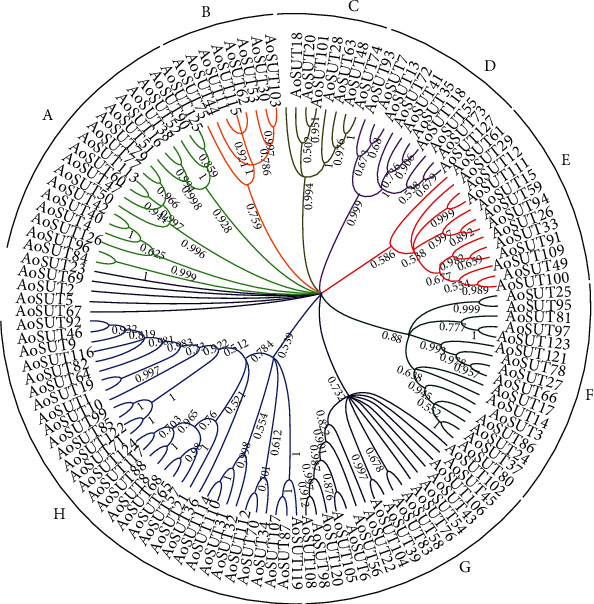
The phylogenetic relationship in A. oryzae SUT proteins. The maximum likelihood tree was constructed by the MEGA X with 1000 bootstrap replications. Eight clades were distinguished in different colors, and several unclassified genes were colored in black.
These AoSUT genes were grouped together suggesting that these homologous genes may have derived from multiple duplications after the speciation of A. oryzae during the evolution.
3.3. Conserved Motifs and Structures of AoSUT Gene Families
A total of six conserved motifs were identified in the SUT gene family of A. oryzae, and the location of these motifs within the protein is represented in Figure 2. As the figure shows, motif numbers varied from zero to eight. Motif 5 was included in all of AoSUT (except AoSUT102), and the majority of AoSUT genes contained two motif 5, such as AoSUT27 and AoSUT66. These results elucidated that motif 5 may play a vital role in AoSUT genes. Moreover, only a few protein sequences do not contain motif 2, demonstrating its significance for the AoSUT proteins in A. oryzae. And the motif number of several genes is under 3, AoSUT 26, AoSUT 29 and AoSUT 117 (3 motif), AoSUT111 (double motif 5). It is noteworthy that the AoSUT102 have no motif. The consensus amino acid sequence width of the conserved motifs ranged from 15 to 24 amino acids (Table 2). Furthermore, the amino acid frequency in the six conserved motifs of AoSUT proteins was investigated. It showed that the amino acid frequency of the conserved motifs is not consistent in different AoSUT proteins (Figure S2).
Figure 2.

Conserved motifs of the SUT gene family in A. oryzae. All conserved motifs of the AoSUT proteins were identified by the MEME program. Protein sequences are indicated by thick gray lines, and the conserved motifs are represented by different colored boxes. The length (the number of amino acids) of the protein and motif can be estimated using the scale bar at the bottom.
Table 2.
The motifs identified in AoSUT proteins.
| Motif | Conserved amino acid sequences | E values | Sites | Width |
|---|---|---|---|---|
| 1 | LPESPRWLVRKGRHEEARQVL | 7.3e − 980 | 111 | 21 |
| 2 | RFIAGJGVGILSATVPVYQSEIAP | 1.0e − 886 | 113 | 24 |
| 3 | PETKGRSLEEJDELF | 6.0e − 755 | 113 | 15 |
| 4 | QFFQQLSGINAISYYAPTJFZ | 2.4e − 661 | 101 | 21 |
| 5 | FLIDRFGRRKLLLIG | 1.8e − 498 | 126 | 15 |
| 6 | VPWLYPAEIFPLRLRA | 3.0e − 442 | 117 | 16 |
To obtain a further insight into the evolutionary trajectory and structural diversity of SUT genes, exon-intron boundaries were analyzed by the GSDS software. As the result showed in Figure S3, a considerable proportion of AoSUT genes contained short and abundant introns, and an analogous phenomenon has been found in the genomes of many lower eukaryotes [38]. The number of exons in AoSUT gene is diversified, ranging from one to 13, whereas the number of introns ranged from zero to 12. It is known that genes lacking introns (e.g., AoSUT99 and AoSUT117) would evolve faster than the rate of intron gain after gene duplication. Simultaneously, it should be noted that some members shared similar intron numbers but with different intron lengths, such as AoSUT19 and AoSUT30 and AoSUT65 and AoSUT69. The existence of plentiful exons and introns indicates that abundant energy is spent to synthesize introns and to maintain the nucleoprotein complex serving the intron-containing genes and that several mechanisms act to sustain vast genomic information [39, 40]. Variation in architectural features of AoSUT genes indicated that gains or losses of exons have occurred during the evolutionary process of the SUT gene family, which may give rise to functional diversity of closely associated SUT genes.
3.4. Chromosomal Distribution of AoSUT Genes
The distribution of 127 SUT genes in A. oryzae on 8 chromosomes was visualized by the MapChart program. It can be seen that all of the 127 AoSUT genes are nonrepetitive and randomly distributed. The great amounts of AoSUT genes were located on the proximate or the distal ends of the chromosomes (Figure 3). There were 11 and 10 AoSUT genes mapped onto chromosomes 5 and 7 whereas chromosomes 2 and 6 harbored the most AoSUT genes (20 and 21, respectively). In addition, 17 AoSUT genes were found on chromosomes 3 and 4.
Figure 3.
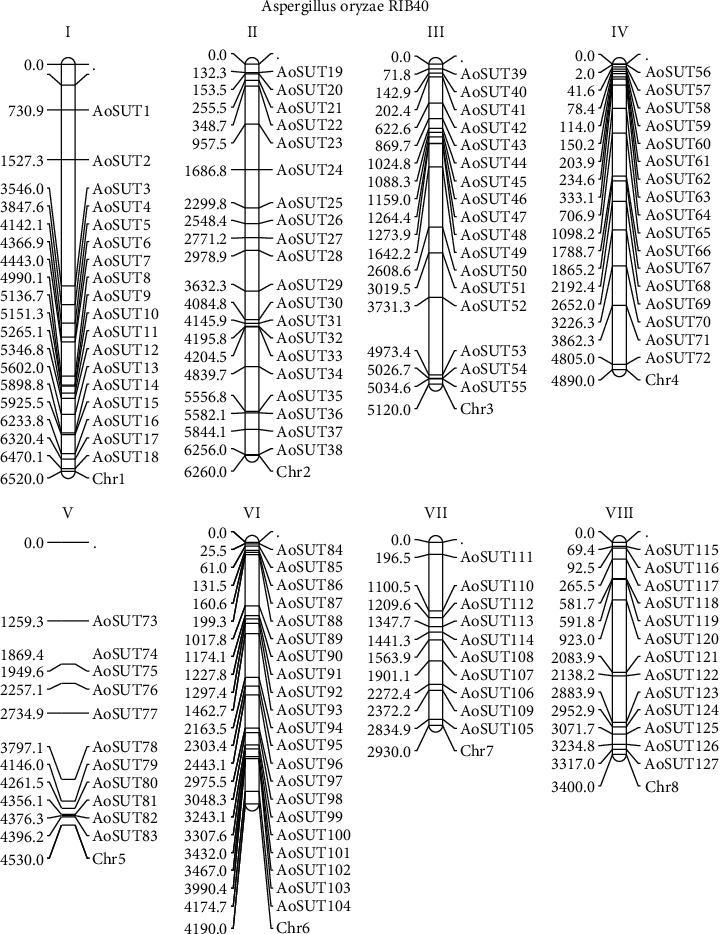
The chromosomal locations of AoSUT genes in the A. oryzae genome. Chromosomes I-VIII are depicted as bars. The AoSUT genes are indicated by black lines.
3.5. Expression Analysis of AoSUT Genes during Growth Stages
To investigate the transcript pattern of SUT family genes during growth stages, the expression patterns at three developmental stages of A. oryzae (from adaptive, logarithmic, to stationary phase) were analyzed by mining publicly accessible A. oryzae transcriptome datasets, which were released by our previous studies [36]. The results revealed that a large proportion of AoSUT genes are highly expressed at the logarithmic and stationary phases (Figure 4). Moreover, a portion of AoSUT genes showed apparent upregulation in the logarithmic phase, whereas it had low expression level in the stationary phase (e.g., AoSUT7, AoSUT44, and AoSUT96). Similarly, some AoSUT genes have high expression in the stationary phase while low in the adaptive phase (e.g., AoSUT8, AoSUT12, and AoSUT88), revealing that these AoSUT genes may be peculiarly involved in the growing processes during adaptive and logarithmic phases. The diversity of expression profiles of AoSUT genes indicated a broad array of biological functions during the growth and development of A. oryzae under normal conditions.
Figure 4.
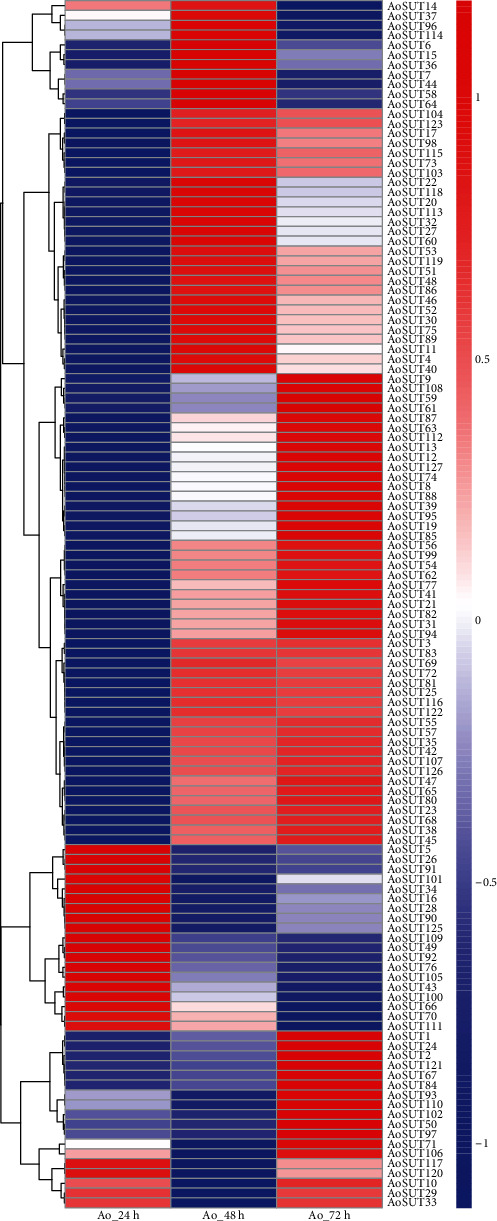
Expression profiles of AoSUT genes at different stages of development. Ao_24_48_72h indicated the expression level of AoSUT at 24 h, 48 h, and 72 h (corresponding to lag phase, logarithmic phase, and stationary phase, respectively). The color of the scale bar, ranging from blue to red, represents low and high expressions, respectively.
To test the accuracy of the gene expression determined from transcriptome data, six AoSUT genes were randomly selected to investigate the expression profiles at three developmental stages of A. oryzae (24 h, 48 h, and 72 h, corresponding to adaptive, logarithmic, and stationary phases) using qRT-PCR; among them the relative expression level of each gene at 24 h is used as a control and its value set to 1 for comparison. As shown in Figure 5, the expression level of AoSUT1, AoSUT12, AoSUT23, and AoSUT35 showed a gradual increase upon transition from the adaptive phase to the logarithmic and stationary phases. However, the expression profiles of AoSUT5 were dramatically decreased from the adaptive phase to the logarithmic and stationary phases. Additionally, the transcript levels of AoSUT69 reached the highest level at the logarithmic phase. The results of qRT-PCR were coordinated with those of transcriptome analysis.
Figure 5.
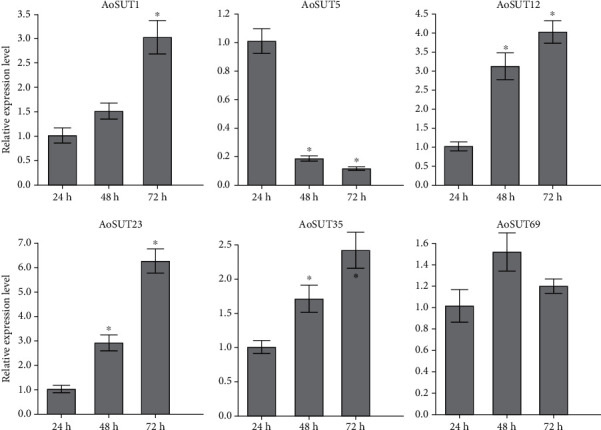
Expression profiles of six selected AoSUT genes at different stages of development determined by qRT-PCR. The bars represent the average (±SE) of biological repeats. Asterisks indicate statistical difference between groups (Student's t test): ∗p < 0.05.
3.6. Expression Analysis of AoSUT Genes under Salt Stress
The abiotic stress response ability of SUT genes has been reported in many fungi but not in A. oryzae, such as salt stress and ethanol stress. Therefore, we investigated the expression patterns of the AoSUT gene family under different concentrations of salt stress (cultivated in PDA medium supplied with 0, 5, 10, and 15% NaCl) in the present study. The heat map representation of expression patterns of these AoSUT genes, represented by FPKM values, exhibited differential expression patterns (Figure 6). Among them, AoSUT genes presented altered expression patterns, either induction or suppression. A majority of AoSUT genes exhibited relatively high expression in the control, whereas in response to slight and moderate salt stress, a large array of AoSUT genes are not expressed or only expressed at very low levels. There are also a small number of genes (approximately 1/5) that are highly expressed at high concentrations of salt stress. It elucidated that these genes may correlate with responding to salt stress or probably collaborating with other genes associated with the response of salinity. These results obtained indicated that AoSUT are components of a complex transcriptional network regarding the different concentrations of salt stress. The dynamic expression pattern under salt stress illustrated that the mechanism of AoSUT genes in response to osmotic regulation might be sophisticated and varied.
Figure 6.
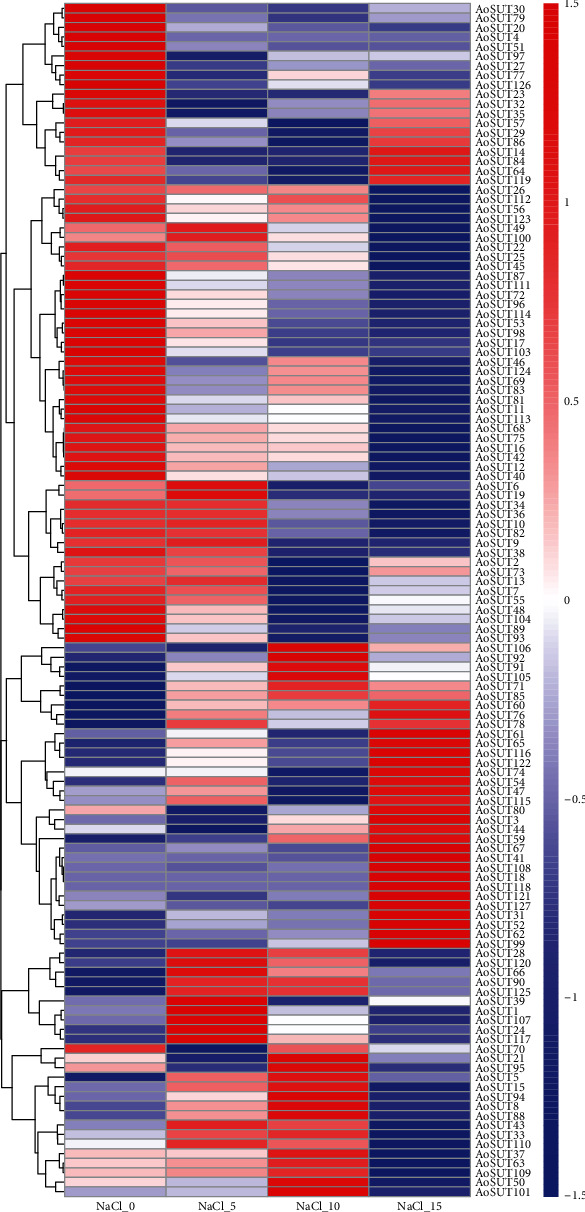
The expression levels of AoSUT genes under salt stress. NaCl_0_5_10_15 indicated samples cultivated in PDA medium supplied with 0, 5, 10, and 15% NaCl, respectively. The color scale bar, ranging from blue to red, represents low and high expressions, separately.
We also selected six AoSUT genes (including AoSUT1, AoSUT5, AoSUT12, AoSUT23, AoSUT35, and AoSUT69) to examine their expression levels under salt stress using qRT-PCR; of them the relative expression level of each gene at under 0% salt concentration was used as a control and its value set to 1 for comparison. Under salt stress, AoSUT1 was notably upregulated at 5% salinity treatment, around nine times the expression level of control (0% salinity treatment), while AoSUT5 had significantly increased expressions when treated in 5 and 10% salinity (Figure 7). In addition, the other four AoSUT genes (AoSUT12, AoSUT23, AoSUT35, and AoSUT69) were downregulated under salt stress, indicating their negative roles in response to salt stress.
Figure 7.
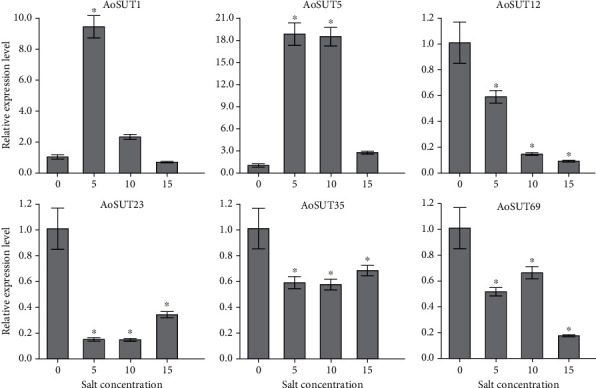
Expression profiles of six selected AoSUT genes in response to salt stress determined by qRT-PCR. The bars represent the average (±SE) of biological repeats. Asterisks indicate statistical difference between groups (Student's t test): ∗p < 0.05.
3.7. Analysis of AoSUT Protein Function Link Network
To elucidate the potential molecular mechanisms of A. oryzae sugar transporter protein, here, we constructed the PPI network using the STRING database. Twenty-five AoSUT proteins were included in this network (Figure 8). Besides AoSUT proteins, there were five proteins, including vacuolar transport chaperone, shikimate dehydrogenase, serine, neutral trehalase, and an uncharacterized protein, located in the important positions of the network. Moreover, various types of protein interactions exist in this figure, for instance, AoSUT22 and Serine associated with three proteins, respectively, and AoSUT80 only interacted with AoSUT6. Interestingly, we also found that shikimate dehydrogenase, AoSUT97, and uncharacterized protein interacted with each other. As the whole network shows, AoSUT44 and vacuolar transport chaperone featured prominently in the protein-protein network, illustrating that both of them play essential roles in maintaining the whole protein interactions in this network. The results obtained in the present study will further accelerate the identification of more vital proteins and biological modules which interacted with AoSUT proteins. The detailed information of the proteins in this PPI network is listed in Table S2.
Figure 8.
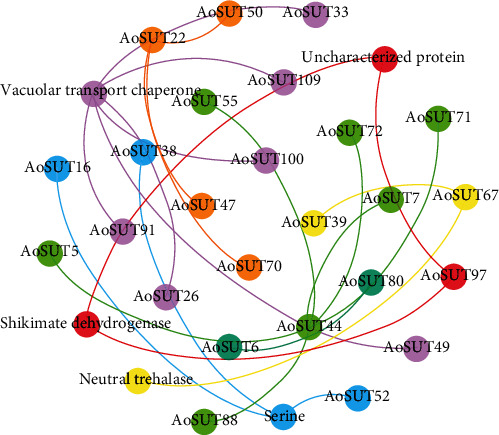
A protein-protein interaction (PPI) network of A. oryzae sugar transporter. The nodes represent proteins, and the edges represent the corresponding PPI. The confidence score was required to be greater than 0.7.
4. Discussion
According to literature mining, with considerable diversity in the numbers and types of SUT gene among fungi, insects, and plants, it can be seen that SUT genes are widespread in nature. The accessibility of the A. oryzae genome sequence has supplied a great opportunity to identify gene family members. In the current study, we identified a total of 127 AoSUT genes in eight clades (Figure 1). By searching the conserved SUT domain, researcher ascertained 86 putative SUT genes in A. niger, and 83 of these genes were classified into nine different clades supported by bootstrap values (over 60%). The remaining SUTs were outside the main clades [13]. What is more, previous studies reported 23 SUT genes in the black truffle Tuber melanosporum, which fell into two subgroups of genes: 16 “true” and 7 “potential” sugar transporter genes [41]. We also performed phylogenetic analysis between A. oryzae, A. niger, and Tuber melanosporum SUT proteins (Figure S4). From the result obtained, these SUTs were divided into nine clades and all were in the A. niger phylogenetic tree. Moreover, the topology of this phylogenetic tree was analogous to the A. oryzae unrooted tree. Among them, the abovementioned several unclassified AoSUT genes were grouped with A. niger and Tuber melanosporum SUT genes, which indicated the homology with each other. It is worth noting that Tuber melanosporum sugar transporter genes primary distributed in clades D-G and clade I. Above, it is not hard to conclude the variations in the distribution and number of SUT genes among filamentous fungi; this difference may be caused by the evolution of different strains in adapting to distinct external environments. Coincidentally, beyond comparative genomics, researchers identified 85 SUTs in C. militaris, which is closed to A. niger, clustering into nine subfamilies [14]. Moreover, 100 sugar transporter genes in the silkworm Bombyx mori genome (categorized into 11 distinct groups in the phylogenetic tree) and 20 SUT genes were identified from the cassava (Manihot esculenta) genome, which were further divided into four clades [17, 23]. To sum up, by comparing SUT number and type with other species, our results of the SUT gene family identification together with the previous studies strongly suggest the multiplicity of SUT genes in the biological world. It might be related to biochemical functions of SUT genes in various species. Our effort will be beneficial to explore SUT genes with similar functional characteristics within distinct species in the near future.
Additionally, we noticed that exon and intron numbers of AoSUT gene are various (exons ranging from one to 13, while introns from zero to 12), which showed that the majority of AoSUT genes possess short and abundant introns (Figure S3). The results might also reflect the mechanism of the expansion in genome size of A. oryzae to some extent. Comparatively, the exon-intron structure of SUT genes is different from other species. In aphids, for example, a great number of SUT genes have a multiexon structure (with 7-9 exons being typical). Among them, Ap_ST3 harbors 7 exons with alternative splicing giving two different exon 1 sequences in the 5′ untranslated region [42]. In plants, the exon-intron structure of SUT genes is also diversified. For example, the exon-intron structure of SUT genes in strawberry is different in various subfamilies (ranging from one to 40) and has substantial short introns [21], while the exon-intron structure of SUT genes in cabbages is relatively conserved, 72.7% of which were found to have four exons and three introns [22]. The previous studies revealed that the existence of exons and substantial introns played roles in maintaining vast genomic information content [43].
Although numerous studies have reported the crucial roles of SUT genes in sugar transport and responses to biotic and abiotic stresses in many species, such as A. niger [13], aphids [42], and wheat [44], the genome-wide expression profile of the SUT gene family was absent in A. oryzae. Here, we explored the expression profiles of the AoSUT genes during growth stages and under salt stress. As shown in Figure 4, AoSUT genes are highly expressed at the logarithmic and stationary phases. In these phases, the number of cells increases exponentially, and cells have been proved of vigorous energy and strong adaptability [45, 46]. The expression patterns of AoSUT genes indicate their primary roles in the permeability of cell membrane and stress responses. These results may lead to more directed understanding on the function of AoSUT genes in developmental biology and stress responses. Under salt stress, AoSUT genes presented altered expression patterns, either induction or suppression, indicating that AoSUT are components of a complex transcriptional network regarding the different concentrations of salt stress. The dynamic expression pattern under salt stress illustrated that the mechanism of AoSUT genes in response to osmotic regulation might be sophisticated and diversified. Similar results have been found in wheat. The expression pattern of two sugar transporter genes (SuT4 and SuT5) were differentially regulated during salt stress, which implies that the product of these SUT genes is critical in signal transduction pathway under osmotic stress due to the distinct salt environment [44]. Furthermore, Truernit et al. reported that the expression level of sugar transporter gene (AtSTP4) in Arabidopsis was significantly upregulated under pathogen attack and wounding [47]. These results indicate that the SUT genes may play vital roles in response to biotic and abiotic stresses. Despite the appreciable progress in the analysis of sugar transporters in A. oryzae, it should be interesting to perform functional genomics experiments, for further clarifying and experimentally demonstrating the important questions concerning the physiological functions of individual AoSUT genes in the near future, for instance, deletion of AoSUT genes in A. oryzae.
5. Conclusions
In summary, we performed a systematic and comprehensive analysis of the SUT gene family in A. oryzae, including phylogenetic relationships, basic classification, conserved motif, gene structure, chromosomal location, and the expression level during growth stages and under salt stress. In addition, qRT-PCR was employed to examine the expression profiles of the AoSUT genes during growth stages and in responses to salt treatments. The results revealed that the AoSUT gene family may mainly participate in the logarithmic and stationary phases of development and be involved in salt stress responses. These results supply an accurate reference of AoSUT genes, which lay a solid foundation for further characterization of the physiological and biochemical functions of individual AoSUT genes.
Acknowledgments
The authors thank Yunyong Wen for critical reading of this manuscript. This research was funded by the Natural Science Foundation of Jiangxi Province (Grant Nos. 20202BABL203043 and 20202BAB215003), the Science and Technology Research Project of Jiangxi Provincial Department of Education (GJJ170692), the National College Students' Entrepreneurship Training Program (202011318039X), and the Innovation Fund Designated for Graduate Students of Jiangxi Science and Technology Normal University (YC2020-X20).
Contributor Information
Bin Zeng, Email: zengtx001@aliyun.com.
Bin He, Email: hebin.li@foxmail.com.
Data Availability
Raw transcriptome data are available through the NCBI Sequence Read Archive databases (BioProject Accession PRJNA407002 and PRJNA383095). All samples were sequenced as 100 bp paired-end reads on an Illumina HiSeq 2000 sequencer.
Conflicts of Interest
The authors declare that there is no conflict of interest regarding the publication of this paper.
Supplementary Materials
Table S1: primer details for genes selected for qRT-PCR analysis in this study.
Table S2: the detailed information of the AoSUT proteins in the PPI network.
Figure S1: phylogenetic analysis of A. oryzae SUT proteins. An unrooted NJ tree of A. oryzae SUT amino acid sequences using 1000 bootstrap replicates by MEGA X. The eight clades are marked by different colors.
Figure S2: sequence logos for 6 motifs of sugar transporter (SUT) domains using the MEME program. MEME motifs are displayed by stacks of letters at each site. The total height of the stack is the “information content” of that site in the motif in bits. The height of each letter in a stack is the probability of the letter at that site multiplied by the total information content of the stack. Abscissa represents the width of the motif, and ordinate represents the bits of each letter, separately.
Figure S3: exon-intron structure of 127 sugar transporter genes identified in A. oryzae. Exons are shown as red boxes, and introns are shown as black lines.
Figure S4: the phylogenetic relationship between A. oryzae, A. niger, and Tuber melanosporum SUT proteins. The maximum likelihood tree was constructed by the MEGA X with 1000 bootstrap replications. Nine clades were distinguished in different colors.
References
- 1.Zhong Y., Lu X., Xing L., Ho S. W. A., Kwan H. S. Genomic and transcriptomic comparison of Aspergillus oryzae strains: a case study in soy sauce koji fermentation. Journal of Industrial Microbiology & Biotechnology. 2018;45(9):839–853. doi: 10.1007/s10295-018-2059-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.He B., Hu Z., Ma L., et al. Transcriptome analysis of different growth stages of Aspergillus oryzae reveals dynamic changes of distinct classes of genes during growth. BMC Microbiology. 2018;18(1):p. 12. doi: 10.1186/s12866-018-1158-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Smit G., Smit B. A., Engels W. J. Flavour formation by lactic acid bacteria and biochemical flavour profiling of cheese products. FEMS Microbiology Reviews. 2005;29(3):591–610. doi: 10.1016/j.fmrre.2005.04.002. [DOI] [PubMed] [Google Scholar]
- 4.Feng Y., Cui C., Zhao H., Gao X., Zhao M., Sun W. Effect of koji fermentation on generation of volatile compounds in soy sauce production. International Journal of Food Science & Technology. 2013;48(3):609–619. doi: 10.1111/ijfs.12006. [DOI] [Google Scholar]
- 5.Kum S.-J., Yang S.-O., Lee S. M., et al. Effects of Aspergillus species inoculation and their enzymatic activities on the formation of volatile components in fermented soybean paste (doenjang) Journal of Agricultural and Food Chemistry. 2015;63(5):1401–1418. doi: 10.1021/jf5056002. [DOI] [PubMed] [Google Scholar]
- 6.Munawar A., Marshall J. W., Cox R. J., Bailey A. M., Lazarus C. M. Isolation and characterisation of a ferrirhodin synthetase gene from the sugarcane pathogen Fusarium sacchari. Chembiochem. 2013;14(3):388–394. doi: 10.1002/cbic.201200587. [DOI] [PubMed] [Google Scholar]
- 7.Fujii R., Minami A., Tsukagoshi T., et al. Total biosynthesis of diterpene aphidicolin, a specific inhibitor of DNA polymerase alpha: heterologous expression of four biosynthetic genes in Aspergillus oryzae. Bioscience, Biotechnology, and Biochemistry. 2014;75(9):1813–1817. doi: 10.1271/bbb.110366. [DOI] [PubMed] [Google Scholar]
- 8.Walmsley A. R., Barrett M. P., Bringaud F., Gould G. W. Sugar transporters from bacteria, parasites and mammals: structure–activity relationships. Trends in Biochemical Sciences. 1998;23(12):476–481. doi: 10.1016/S0968-0004(98)01326-7. [DOI] [PubMed] [Google Scholar]
- 9.Gupta A. K., Kaur N. Sugar signalling and gene expression in relation to carbohydrate metabolism under abiotic stresses in plants. Journal of Biosciences. 2005;30(5):761–776. doi: 10.1007/BF02703574. [DOI] [PubMed] [Google Scholar]
- 10.de Vries R. P., Riley R., Wiebenga A., et al. Comparative genomics reveals high biological diversity and specific adaptations in the industrially and medically important fungal genus Aspergillus. Genome Biology. 2017;18(1):p. 28. doi: 10.1186/s13059-017-1151-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Colabardini A., Ries L. N., Brown N., et al. Functional characterization of a xylose transporter in Aspergillus nidulans. Biotechnology for Biofuels. 2014;7(1):p. 46. doi: 10.1186/1754-6834-7-46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.dos Reis T. F., Menino J. F., Bom V. L. P., et al. Identification of glucose transporters in Aspergillus nidulans. PLoS One. 2013;8(11, article e81412) doi: 10.1371/journal.pone.0081412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Peng M., Aguilar-Pontes M. V., de Vries R. P., Makela M. R. In silico analysis of putative sugar transporter genes in Aspergillus niger using phylogeny and comparative transcriptomics. Frontiers in Microbiology. 2018;9:p. 1045. doi: 10.3389/fmicb.2018.01045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sirithep K., Xiao F., Raethong N., et al. Probing carbon utilization of Cordyceps militaris by sugar transportome and protein structural analysis. Cells. 2020;9(2):p. 401. doi: 10.3390/cells9020401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.van Kuyk P. A., Diderich J. A., Mac Cabe A. P., Hererro O., Ruijter G. J. G., Visser J. Aspergillus niger mstA encodes a high-affinity sugar/H+ is regulated in response to extracellular pH. Biochemical Society. 2004;379:375–383. doi: 10.1042/BJ20030624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Monfared H. H., Chew J. K., Azizi P., et al. Overexpression of a rice monosaccharide transporter gene (OsMST6) confers enhanced tolerance to drought and salinity stress in Arabidopsis thaliana. Plant Molecular Biology Reporter. 2020;38(1):151–164. doi: 10.1007/s11105-019-01186-x. [DOI] [Google Scholar]
- 17.Govindaraj L., Gupta T., Esvaran V. G., Awasthi A. K., Ponnuvel K. M. Genome-wide identification, characterization of sugar transporter genes in the silkworm Bombyx mori and role in Bombyx mori nucleopolyhedrovirus (BmNPV) infection. Gene. 2016;579(2):162–171. doi: 10.1016/j.gene.2015.12.057. [DOI] [PubMed] [Google Scholar]
- 18.Liang X., Yan F., Gao Y., et al. Sugar transporter genes in grass carp (Ctenopharyngodon idellus): molecular cloning, characterization, and expression in response to different stocking densities. Fish Physiology and Biochemistry. 2020;46(3):1039–1052. doi: 10.1007/s10695-020-00770-3. [DOI] [PubMed] [Google Scholar]
- 19.Wang Z., Gerstein M., Snyder M. RNA-Seq: a revolutionary tool for transcriptomics. Nature Reviews. Genetics. 2009;10(1):57–63. doi: 10.1038/nrg2484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Koren S., Schatz M. C., Walenz B. P., et al. Hybrid error correction and de novo assembly of single-molecule sequencing reads. Nature Biotechnology. 2012;30(7):693–700. doi: 10.1038/nbt.2280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jiu S., Haider M. S., Kurjogi M. M., Zhang K., Zhu X., Fang J. Genome-wide characterization and expression analysis of sugar transporter family genes in woodland strawberry. Plant Genome. 2018;11(3):1–16. doi: 10.3835/plantgenome2017.11.0103. [DOI] [PubMed] [Google Scholar]
- 22.Zhang W., Wang S., Yu F., et al. Genome-wide identification and expression profiling of sugar transporter protein (STP) family genes in cabbage (Brassica oleracea var. capitata L.) reveals their involvement in clubroot disease responses. Genes. 2019;10(1) doi: 10.3390/genes10010071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Liu Q., Dang H., Chen Z., et al. Genome-wide identification, expression, and functional analysis of the sugar transporter gene family in cassava (Manihot esculenta) International Journal of Molecular Sciences. 2018;19(4) doi: 10.3390/ijms19040987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Nogueira K. M. V., de Paula R. G., Antoniêto A. C. C., et al. Characterization of a novel sugar transporter involved in sugarcane bagasse degradation in Trichoderma reesei. Biotechnology for Biofuels. 2018;11(1):p. 84. doi: 10.1186/s13068-018-1084-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Milner D. S., Attah V., Cook E., et al. Environment-dependent fitness gains can be driven by horizontal gene transfer of transporter-encoding genes. Proceedings of the National Academy of Sciences. 2019;116(12):5613–5622. doi: 10.1073/pnas.1815994116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sloothaak J., Tamayo-Ramos J. A., Odoni D. I., et al. Identification and functional characterization of novel xylose transporters from the cell factories Aspergillus niger and Trichoderma reesei. Biotechnology for Biofuels. 2016;9(1):p. 148. doi: 10.1186/s13068-016-0564-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Fang W., Leger R. J. S. Mrt, a gene unique to fungi, encodes an oligosaccharide transporter and facilitates rhizosphere competency in Metarhizium robertsii. Plant Physiology. 2010;154(3):1549–1557. doi: 10.1104/pp.110.163014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wang X. X., Ji X. P., Li J. X., Keyhani N. O., Feng M. G., Ying S. H. A putative α-glucoside transporter gene BbAGT1 contributes to carbohydrate utilization, growth, conidiation and virulence of filamentous entomopathogenic fungus Beauveria bassiana. Research in Microbiology. 2013;164(5):480–489. doi: 10.1016/j.resmic.2013.02.008. [DOI] [PubMed] [Google Scholar]
- 29.Stephen T. L. M., Altschul F., Schäffer A. A., et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research. 1997;25:3389–3402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.El-Gebali S., Mistry J., Bateman A., et al. The Pfam protein families database in 2019. Nucleic Acids Research. 2019;47(D1):D427–D432. doi: 10.1093/nar/gky995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Finn R. D., Clements J., Eddy S. R. HMMER web server: interactive sequence similarity searching. Nucleic Acids Research. 2011;39(suppl):W29–W37. doi: 10.1093/nar/gkr367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kumar S., Stecher G., Li M., Knyaz C., Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Molecular Biology and Evolution. 2018;35(6):1547–1549. doi: 10.1093/molbev/msy096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bailey T. L., Boden M., Buske F. A., et al. MEME SUITE: tools for motif discovery and searching. Nucleic Acids Research. 2009;37(Web Server):W202–W208. doi: 10.1093/nar/gkp335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Tang W., Tu Y., Cheng X., et al. Genome-wide identification and expression profile of the MADS-box gene family in Erigeron breviscapus. PLoS One. 2019;14(12, article e0226599) doi: 10.1371/journal.pone.0226599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tang W., Ouyang C., Liu L., et al. Genome-wide identification of the fatty acid desaturases gene family in four Aspergillus species and their expression profile in Aspergillus oryzae. AMB Express. 2018;8(1, article 169) doi: 10.1186/s13568-018-0697-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.He B., Ma L., Hu Z., et al. Deep sequencing analysis of transcriptomes in Aspergillus oryzae in response to salinity stress. Applied Microbiology and Biotechnology. 2018;102(2):897–906. doi: 10.1007/s00253-017-8603-z. [DOI] [PubMed] [Google Scholar]
- 37.Raman K. Construction and analysis of protein-protein interaction networks. Automated Experimentation. 2010;2(1):p. 2. doi: 10.1186/1759-4499-2-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Loftus B. J., Fung E., Roncaglia P., et al. The genome of the basidiomycetous yeast and human pathogen Cryptococcus neoformans. Science. 2005;307(5713):1321–1324. doi: 10.1126/science.1103773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ivashchenko A. T., Tauasarova M. I., Atambayeva S. A. Exon-intron structure of genes in complete fungal genomes. Molecular Biology. 2009;43(1):24–31. doi: 10.1134/S002689330901004X. [DOI] [PubMed] [Google Scholar]
- 40.Bondarenko V. S., Gelfand M. S. Evolution of the exon-intron structure in ciliate genomes. PLoS One. 2016;11(9, article e0161476) doi: 10.1371/journal.pone.0161476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ceccaroli P., Saltarelli R., Polidori E., et al. Sugar transporters in the black truffle Tuber melanosporum: from gene prediction to functional characterization. Fungal Genetics and Biology. 2015;81:52–61. doi: 10.1016/j.fgb.2015.05.006. [DOI] [PubMed] [Google Scholar]
- 42.Price D. R. G., Tibbles K., Shigenobu S., et al. Sugar transporters of the major facilitator superfamily in aphids; from gene prediction to functional characterization. Insect Molecular Biology. 2010;19(Suppl 2):97–112. doi: 10.1111/j.1365-2583.2009.00918.x. [DOI] [PubMed] [Google Scholar]
- 43.Kupfer D. M., Drabenstot S. D., Buchanan K. L., et al. Introns and splicing elements of five diverse fungi. Eukaryotic Cell. 2004;3(5):1088–1100. doi: 10.1128/EC.3.5.1088-1100.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Fahimeh Charkazi S. S. R., Soltanloo H. Expression pattern of two sugar transporter genes (SuT4 and SuT5) under salt stress in wheat. Plant Omics. 2010. pp. 194–198.
- 45.He B., Tu Y., Hu Z., et al. Genome-wide identification and expression profile analysis of the HOG gene family in Aspergillus oryzae. World Journal of Microbiology and Biotechnology. 2018;34(2) doi: 10.1007/s11274-018-2419-6. [DOI] [PubMed] [Google Scholar]
- 46.Han F., Pei H., Hu W., Zhang S., Han L., Ma G. The feasibility of ultrasonic stimulation on microalgae for efficient lipid accumulation at the end of the logarithmic phase. Algal Research. 2016;16:189–194. doi: 10.1016/j.algal.2016.03.014. [DOI] [Google Scholar]
- 47.Elisabeth Truernit J. S., Epple P., Illig J., Sauer N. The sink-specific and stress-regulated Arabidopsis SPP4 gene: enhanced expression of a gene encoding a monosaccharide transporter by wounding, elicitors, and pathogen challenge. The Plant Cell. 1996;8:2169–2182. doi: 10.1105/tpc.8.12.2169. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1: primer details for genes selected for qRT-PCR analysis in this study.
Table S2: the detailed information of the AoSUT proteins in the PPI network.
Figure S1: phylogenetic analysis of A. oryzae SUT proteins. An unrooted NJ tree of A. oryzae SUT amino acid sequences using 1000 bootstrap replicates by MEGA X. The eight clades are marked by different colors.
Figure S2: sequence logos for 6 motifs of sugar transporter (SUT) domains using the MEME program. MEME motifs are displayed by stacks of letters at each site. The total height of the stack is the “information content” of that site in the motif in bits. The height of each letter in a stack is the probability of the letter at that site multiplied by the total information content of the stack. Abscissa represents the width of the motif, and ordinate represents the bits of each letter, separately.
Figure S3: exon-intron structure of 127 sugar transporter genes identified in A. oryzae. Exons are shown as red boxes, and introns are shown as black lines.
Figure S4: the phylogenetic relationship between A. oryzae, A. niger, and Tuber melanosporum SUT proteins. The maximum likelihood tree was constructed by the MEGA X with 1000 bootstrap replications. Nine clades were distinguished in different colors.
Data Availability Statement
Raw transcriptome data are available through the NCBI Sequence Read Archive databases (BioProject Accession PRJNA407002 and PRJNA383095). All samples were sequenced as 100 bp paired-end reads on an Illumina HiSeq 2000 sequencer.