

Abstract
Gestational Diabetes Mellitus (GDM) is characterized by abnormal maternal D-glucose metabolism and altered insulin signaling. Dysregulation of thyroid hormones (TH) tri-iodethyronine (T3) and L-thyroxine (T4) Hormones had been associated with GDM, but the physiopathological meaning of these alterations is still unclear. Maternal TH cross the placenta through TH Transporters and their Deiodinases metabolize them to regulate fetal TH levels. Currently, the metabolism of TH in placentas with GDM is unknown, and there are no other studies that evaluate the fetal TH from pregnancies with GDM. Therefore, we evaluated the levels of maternal TH during pregnancy, and fetal TH at delivery, and the expression and activity of placental deiodinases from GDM pregnancies. Pregnant women were followed through pregnancy until delivery. We collected blood samples during 10–14, 24–28, and 36–40 weeks of gestation for measure Thyroid-stimulating hormone (TSH), Free T4 (FT4), Total T4 (TT4), and Total T3 (TT3) concentrations from Normal Glucose Tolerance (NGT) and GDM mothers. Moreover, we measure fetal TSH, FT4, TT4, and TT3 in total blood cord at the delivery. Also, we measured the placental expression of Deiodinases by RT-PCR, western-blotting, and immunohistochemistry. The activity of Deiodinases was estimated quantified rT3 and T3 using T4 as a substrate. Mothers with GDM showed higher levels of TT3 during all pregnancy, and an increased in TSH during second and third trimester, while lower concentrations of neonatal TT4, FT4, and TT3; and an increased TSH level in umbilical cord blood from GDM. Placentae from GDM mothers have a higher expression and activity of Deiodinase 3, but lower Deiodinase 2, than NGT mothers. In conclusion, GDM favors high levels of TT3 during all gestation in the mother, low levels in TT4, FT4 and TT3 at the delivery in neonates, and increases deiodinase 3, but reduce deiodinase 2 expression and activity in the placenta.
Introduction
Gestational Diabetes Mellitus (GDM) is defined as any degree of glucose intolerance with onset or first manifestation during pregnancy [1]. GDM is characterized by abnormal maternal D-glucose metabolism and altered insulin signaling in feto-placental circulation [2]. It has been described that a decrease in maternal Free L-Thyroxine (FT4) levels during the first trimester of pregnancy could also increase GDM risk [3–5]. Several studies relate alterations in Thyroid Hormones (TH) levels with GDM, but their results are controversial. While some studies associate a decrease in maternal FT4 and increase in Thyroid Stimulating Hormone (TSH) with GDM, others have related an increase in maternal Total Tri-iodothyronine (TT3, Free T3 plus protein bound T3) with this pathology [6–8].
Throughout the gestational stage, the mother provides these hormones to the fetus through the placenta [9]. Fetal THs are fully contributed by the mother until week 16 of gestation after that, the fetus begins to produce its own THs, due to the maturation of the hypothalamus/pituitary/thyroid axis [10]. Few studies have evaluated the behavior of fetal THs in GDM. A decrease in the neonatal TH from mothers with type 2 diabetes mellitus was evidenced [11], while on the other hand, the suppression of these fetal hormones was also associated with maternal glucose intolerance [12]. However, there are no other studies that evaluate the neonatal thyroid profile from pregnancies with GDM. During pregnancy, T4 and T3 cross the placenta through Thyroid Hormone Transporters (THT), entering the placenta where they are regulated by deiodinases (DIO) [13, 14]. DIO are seleno-enzymes responsible for the catabolism of specific iodine atoms of the iodothyronine molecule [15–18]. These have three subtypes: I, II, and III (DIO1, DIO2, and DIO3, respectively), which have fundamental functions in thyroid hormone regulation [18, 19]. DIO1 is characterized by generating T3 through the deiodination of T4; however, its role is less relevant since its activity is lower than the other deiodinases [18]. DIO2 produces T3 from a 5′-deiodination of T4, increasing the availability of T3, which in turn is essential for the development of the fetus during pregnancy [20]. Indeed, it has been reported that a physiological concentration of thyroid hormones stimulates trophoblast endocrine function [21], while alterations in TH availability are linked with alterations in fetal growth and weight [22]. Finally, DIO3 is highly localized in placenta, and is the enzyme responsible for converting T4 into reverse T3 (rT3, inactive form), and catalyzes the conversion of diiodothyronine (T2) from T3, decreasing the levels of T3 [23, 24].
Currently, the metabolism of TH in placentas from pregnancies with GDM is unknown, then, we evaluated the levels of maternal and neonatal TH, and the expression and activity of placental DIO in the context of GDM.
Materials and methods
Patients, biological samples, and study groups
181 pregnant women were recruited between the years 2017 and 2018 from 3 primary health centers of Concepción (Víctor Manuel Fernández, Tucapel, and Santa Sabina) with 12–14 weeks of pregnancy. Pregnant women were followed through pregnancy until delivery. We collected blood samples during 10–14, 24–28, and 36–40 weeks of pregnancy, and total umbilical blood during post-delivery. Blood samples were transported to laboratory at 4°C. Sera were aliquoted and stored at -80°C. The placentae were obtained post-delivery, and transported to laboratory at 4°C. For the current study, a subset of 71 pregnant women who complied all diagnosis and inclusion criteria were analyzed.
Pregnant women between the 24–28 weeks of pregnancy with basal glycemia >90 mg/dL, (i.e., overnight starvation) or with >140 mg/dL at 2 h after an oral glucose load (75 g) were diagnosed as GDM, and subjected to dietary treatment with 1500 kcal/day and a maximum of 200 g per day carbohydrates. Our study obtained 23 pregnant with GDM and 48 pregnant without alteration in basal glycemia and oral glucose load, named Normal Glucose Tolerant (NGT). The exclusion criteria included underage women, previous diagnosis of diabetes mellitus and or thyroid pathologies, and other pregnancy pathologies (i.e. preeclampsia, fetal growth restriction, ectopic pregnancy).
The following characteristics were collected from the mothers: age (years), height (m), weight during first and third pregnancy trimesters (kg), and Body Mass Index (BMI) was calculated. For the newborn, data of gestational age (weeks), weight, height and ponderal index were collected.
Ethic aspects
The investigation conforms to the principles outlined in the Declaration of Helsinki. Ethics Committee approval from the Comité Ético Científico de Servicio de Salud Concepción (CEC:23-2017-20) and informed consent of each pregnant women from the three Primary Health Centers of Concepción were obtained.
Clinical biochemistry analysis
Glucose was measured by a colorimetric assay (VITROS Immunodiagnostic Products, NY, USA, GLU Slides). TSH concentrations were measured by immunometric assay (VITROS Immunodiagnostic Products, NY, USA, TSH), while FT4, Total L-Thyroxine (TT4), and T T3 were measured by competitive immunoassay (VITROS Immunodiagnostic Products, NY, USA).
Expression analysis
Reverse transcription and quantitative RT-PCR
RNA was extracted using Trizol reagent (Life Technologies Corporation, Carlsbad, CA, USA) following the manufacturer's instructions. Reverse transcriptions were realized using ImProm-II™ Reverse Transcription System (Promega, Madison, WI, USA). 1 μg of RNA in presence of 0.5 μg of random primers mix is thermally denatured (70°C, 5 min) and cooled on ice. A reverse transcription reaction mix was assembled on ice to contain nuclease-free water (to a final volume of 15μl), reaction buffer 5x, 160UI of reverse transcriptase, 6mM of magnesium chloride, 0.5 mM of dNTPs, 20UI of ribonuclease inhibitor and 1UI of Recombinant RNasin® Ribonuclease Inhibitor (Promega). As a final step, the template-primer combination is added to the reaction mix. Following an initial annealing (25°C, 5 min), and the reaction is incubated at 42°C for up to one hour. This procedure outlines the method proposed to amplify the entire 20μl reaction.
cDNA amplifications were performed using a Step One real time PCR system (Applied Biosystem, CA, USA) in a reaction mix containing 0.2 μM primers and master mix provided in the brilliant SYBR green qPCR Master Mix (Applied Biosystem, CA, USA) as described [25]. Hot Start Taq DNA polymerase was activated (10 min, 94°C), and assays included a 94°C denaturation (30 s), annealing (30 s) at 54.0°C (DIO3 and 28S) or 57.2°C (DIO1 and DIO2), and extension at 72°C (DIO1, DIO2, DIO3 and 28S, 45 s). Fluorescent product was detected after 3-s step to 5°C below the product melting temperature (Tm). Product specificity was confirmed by agarose gel electrophoresis (2% w/v) and melting curve analysis. The product Tm values were 60°C for DIO1, 60.2°C for DIO2, 56.8°C for DIO3 and 86.7°C for 28S [25]. Oligonucleotide primers are as follows: DIO1 (sense) 5’ -AGCGACTAGAGGACACGACT- 3’, DIO1 (anti-sense) 5’ -ACCAGTGGCCTATTACCTTGC- 3’, DIO2 (sense) 5’ -TCGATGCCTACAAACAGGTGAA- 3’, DIO2 (anti-sense) 5’ -TTGCCACTGTTGTCACCTCC- 3’, DIO3 (sense) 5’ -TCGAGCGTCTCTATGTCATC- 3’, DIO3 (anti-sense) 5’ -TCATCATAGCGTTCCAACCA- 3’, 28S (sense) 5’ -TTGAAAATCCGGGGGAGAG- 3’, 28S (anti-sense) 5’ -ACATTGTTCCAACATGCCAG- 3’. Expected size products for DIO1 (115 bp), DIO2 (94 bp), DIO3 (109 bp) and 28S (100 bp) were confirmed in PCR experiments. The 28S rRNA ct was unaltered (P > 0.05, n = 16) in all experimental conditions (not shown). The Ct value was defined as the PCR cycle number at which the probes’ fluorescent signal exceeded the background signal and was used to calculate the gene-expression data with the 2–ΔΔCt method.
Western blotting
Proteins were extracted taking 1 mg of placenta in presence of lysis buffer (Tris 50mM pH 7.5; Triton X-100 0.2%; EDTA 1mM), and proteases inhibitor (HaltTM Protease cocktail, Thermo Fisher Scientific, Waltham, MA, USA)
Proteins (70 μg) separated by polyacrylamide gel (12%) electrophoresis were probed with a primary polyclonal rabbit anti-DIO2 (1:1000, v/v) (Thermo Fisher Scientific), rabbit anti-DIO3 (1:1000, v/v) (Thermo Fisher Scientific), or monoclonal mouse anti-β-actin (1:3000, v/v) (Santa Cruz Biotechnology, Santa Cruz, CA, USA) antibodies. Proteins were detected by enhanced chemiluminescence in a LI-COR C-DiGit™ Blot Scanner 3600 (Lincoln, NE, USA) and quantified by densitometry using software ImageJ (NIH, Bethesda, MD, USA).
Placental tissue analysis
Immunohistochemistry
A standard protocol for immunohistochemistry was applied to two sections of the same placenta sample (100mm3) to study each DIO. The tissue sections were mounted on a slide previously treated with 2% silane in acetone. The placenta sections were dewaxed with three xylol baths of 10 minutes each and then were rehydrated in an ethanol battery (absolute ethanol, 95%, 70%, respectively) of 1 minute each. Afterwards, an antigenic recovery was performed with 0.01M Sodium Citrate buffer solution pH 6.0 at a temperature of approximately 90°C for 30 minutes, using a steamer.
The samples were then cooled in the same solution with changes of distilled water. A wet chamber was needed for the following steps. The endogenous peroxidase was blocked using 3% hydrogen peroxide in absolute methanol for 10 minutes at room temperature, followed by a wash with 1X PBS Buffer at pH 7, 0.01M + Tween 20 0.05% (PBST). The samples were then incubated with the respective primary antibody at DIO1 1:50 (v/v), DIO2 1:100 (v/v), and DIO3 1:500 (v/v) concentrations with TBS antibody diluent, for 45 minutes at 37°C, followed by washing with PBST.
A detection polymer (Mouse / Rabbit PolyDetector Plus DAB HRP Brown Detection System Bio SB) was used for to detect the primary antibody, incubating each sample for 15 minutes at 37°C and subsequently washed with PBST. Finally, a nuclear contrast was carried out with Harris's Hematoxylin for 20 seconds, followed by a bluish in running water for 5 minutes, then by a dehydration battery of increasing ethanol (70%, 95%, absolute ethanol) of 1 minute each, to end with a xylol clearance for 3 minutes and the assembly of the slides with synthetic resin in hydrophobic medium.
Deiodinase activity
300 μg of placenta protein from cotyledon in presence of 1,4-Dithiothreitol (DTT, 20mM) were exposed to T4 (MP Biomedicals, LLC, Solon, OH, USA) (0–500 nM, 37°C, 15 min). The reaction was stopped with absolute ethanol (4°C). Then, the sample was centrifuged at 10.500g (4°C, 8 min), and the supernatant was used to measure T3 and rT3 [26, 27].
T3 and rT3 measure
T3 (Competitive ELISA, Invitrogen) and rT3 (Competitive ELISA, Invitrogen) were quantified from supernatants.
Determination of kinetic parameters
Saturable T4 conversion at initial rates was adjusted to the Michaelis-Menten asymptotic hyperbola. T4-conversion kinetic parameters maximal velocity (Vmax) and apparent Michaelis-Menten constant (Km) of transport were calculated as follows:
| (1) |
Where Vo correspond to initial velocity and [T4] the T4 concentration. Each assay was run in duplicate and activity was expressed in ng/mL/μg of protein/min (5’-deiodinase activity) and pg/L/μg of protein/min (5-deiodinase activity). To compare the GDM effect on deiodinase activity, we calculated the enzyme catalytic efficiency using the Vmax/Km ratio.
Statistical analysis
Sample size was determined by average comparison approach considering free T4 level reported [28] in Chilean pregnant women with the equation:
| (2) |
Replacing free T4 level during first trimester of pregnancy (S), Zα value of 0.05, Zβ value of 0.8 and d value of 0.5; the estimated sample size (n) per group was 70 pregnant women. Thus, this study required an estimated population of 140 pregnant women. Values are mean ± standard deviation (S.D.) (or range or SEM), with n = 71 different parameters from NGT or GDM pregnant women and their newborns. The normality of the data was calculated with Kolmogorov-Smirnov test. Comparisons between two and more groups were performed by means of Student’s unpaired t test and analysis of variance (ANOVA), respectively. If the ANOVA demonstrated a significant interaction between variables, post hoc analyses were performed by the multiple-comparison Bonferroni correction test. P < 0.05 was considered statistically significant. The statistical software GraphPad Prism 7.0a.65 (GraphPad Software Inc., San Diego, CA, USA) was used for data analysis.
Results and discussion
Maternal thyroid hormone profile is altered in GDM during pregnancy
Our study considered the following along pregnancy of 71 women with either NGT or GDM pregnancies. No significant changes were found in height, overall weight during first trimester and third trimester, and BMI between NGT and GDM. However, we observed an increased in weight gain in GDM with respect to NGT. According to diagnosis criteria, increases in basal glycaemia, glycaemia 2 hours post 75 grams of glucose were found during second trimester of GDM pregnancies compared with NGT pregnancies (Table 1).
Table 1. Maternal clinical and biochemical parameters in NGT and GDM pregnancies.
| Parameters | Units | NGT | GDM |
|---|---|---|---|
| n | 48 | 23 | |
| age | years | 29.9 ± 5.6 (27.9–31.9) | 29.5 ± 6.9 (26.0–33.1) |
| Height | M | 1.60 ± 0.1 (1.57–1.61) | 1.61 ± 0.1 (1.55–1.66) |
| Weight 1T | kg | 77.8 ± 5.6 (75.3–80.1) | 78.3 ± 5.2 (75.2–80.3) |
| Weight 3T | kg | 88.8 ± 9.3 (85.5–91.1) | 91.5 ± 5.5 (88.2–93.7) |
| Weight Gain (3T-1T) | kg | 11.0 ± 1.0 (9.9–11.8) | 13.2 ± 1.9* (12.3–14.1) |
| BMI 1T | kg/m2 | 30.3 ± 3.9 (28.5–32.0) | 30.2 ± 3.8 (28.1–32.3) |
| BMI 3T | kg/m2 | 32.3 ± 5.6 (29.1–35.5) | 31.5 ± 4.0 (29.5–33.4) |
| Basal Glycaemia 1T | mg/dl | 81.1 ± 5.5 (78.7–83.7) | 81.6 ± 2.1 (80.7–82.9) |
| Basal Glycaemia 2T | mg/dl | 82.4 ± 0.9 (81.9–82.8) | 87.7 ± 4.8* (85.9–90.1) |
| Glycaemia 2h post 75 g (2T) | mg/dl | 109.0 ± 9.6 (101.2–114.4) | 161.3 ± 9.7* (155.8–172.4) |
| Maternal thyroid hormone profile at the first trimester | |||
| TSH | mIU/L | 2.0 ± 1.3 (1.7–2.6) | 2.3 ± 1.9 (1.2–3.4) |
| FT4 | ng/dl | 1.0 ± 0.2 (0.9–1.1) | 0.9 ± 0.2 (0.8–1.0) |
| TT4 | μg/ml | 11.4 ± 1.9 (10.5–12.4) | 11.5 ± 1.7 (10.7–12.5) |
| TT3 | ng/ml | 1.7 ± 0.2 (1.6–1.8) | 1.9 ± 0.1* (1.9–2.0) |
| Maternal thyroid hormone profile at the second trimester | |||
| TSH | mIU/L | 2.0 ± 0.8 (1.6–2.4) | 2.9 ± 1.2* (2.3–3.5) |
| FT4 | ng/dl | 0.8 ± 0.1 (0.7–0.9) | 0.8 ± 0.3 (0.6–0.9) |
| TT4 | μg/ml | 12.7 ± 1.8 (11.6–13.4) | 13.3 ± 3.0 (11.8–14.9) |
| TT3 | ng/ml | 1.9 ± 0.3 (1.7–2.0) | 2.2 ± 0.2* (2.1–2.3) |
| Maternal thyroid hormone profile at the third trimester | |||
| TSH | mIU/L | 2.1 ± 0.4 (1.9–2.3) | 3.4 ± 1.0* (2.9–3.9) |
| FT4 | ng/dl | 0.7 ± 0.1 (0.6–0.8) | 0.7 ± 0.1 (0.6–0.7) |
| TT4 | μg/ml | 11.1 ± 1.7 (10.1–12.3) | 12.5 ± 1.4* (11.7–13.3) |
| TT3 | ng/ml | 1.6 ± 0.2 (1.5–1.7) | 2.3 ± 0.1*† (2.2–2.3) |
NGT: Normal tolerance glucose. GDM: gestational diabetes mellitus.1T: first trimester; 2T: second trimester; 3T: Third trimester; BMI: Body Mass Index. TSH: Thyroid Stimulant Hormone. FT4: Free Thyroxine. TT4: Total Thyroxine. TT3: Total triiodothyronine. Mean ± S.D. (range).
*P<0.05 versus NGT.
†P<0.05 versus TT3 at the first trimester.
We analyzed the thyroid hormone profile (i.e., TSH, FT4, TT4 and TT3) during pregnancy, in which FT4 levels were similar in both groups along pregnancy. However, women with GDM showed an increase in the TT3 levels in all three trimesters of pregnancy compared to NGT. In addition, TT4 levels was higher in third trimester, and TSH levels were higher in GDM at second and third trimester than respective value in NGT. TT3 levels at the third trimester were higher than levels measured at first trimester only in the GDM group. Similar results have been described by other authors. In a HAPO study with 600 participant women, TT3 levels were increased in GDM pregnant during second trimester, including a positive correlation with glycaemia at 2 hours post-glucose 75 grams [29]; however, no changes were observed for TSH levels. Another study measured Free T3 (FT3), observing an increase of this analyte during first and second trimesters; although FT3 levels during the third trimester were not evaluated in this study [8]. In a retrospective cohort study in Israel with singleton pregnant women who had a first trimester and at least one additional (either first, second, third trimesters, or after delivery) TSH measurement for the same pregnancy, low TSH levels during first trimester were correlated with lower rates of GDM [30]. Moreover, a relationship between low FT4 levels and high maternal weight during second trimester was associated with an increased GDM risk; however, T3 was not measured [2]. In the other hand, another study in Pakistan observed that GDM women had high TSH blood levels, which were correlated with poor glycemic control and BMI [22]. Interestingly, a similar condition known as Gestational Impaired Glucose Tolerant (GIGT), which is associated with glucose at 2 hours post-75 grams > 100mg/dl, but <140 mg/dl, shown an increase in FT4 and TT4 compared to controls, without changes for TT3 [12], which could indicate that a TT3 increase could be associated with GDM, but not with other diseases.
Thyroid hormone profile is altered in newborn from GDM pregnancies
Newborn clinical parameters did not show any change associated with height, ponderal index and sex proportion (Table 2). However, gestational age was reduced; and weight was increased in GDM pregnancies. In addition, the thyroid hormone profile obtained from umbilical cord blood from GDM pregnancies showed a reduction in TT4 (~13.2%) and FT4 (~25%) and TT3 (~47%) levels, and an increase in TSH levels compared with newborn from NGT pregnancies.
Table 2. Newborn clinical and biochemical parameters in NGT and GDM pregnancies.
| Parameters | Units | NGT | GDM |
|---|---|---|---|
| n | 48 | 23 | |
| Newborn characteristic | |||
| Gestational age | weeks | 38.1 ± 1.1 (38.4–38.7) | 37.3 ± 1.4* (36.5–38.0) |
| Weight | g | 3600 ± 350 (3450–3740) | 3900 ± 320* (3760–4050) |
| Height | cm | 50 ± 3 (48.5–51.5) | 51 ± 3 (49.5–52.5) |
| Ponderal index | g/cm3 | 2.88 ± 0.43 (2.66–3.02) | 2.94 ± 0.26 (2.81–3.07) |
| Sex | Female/Male | 23/25 | 11/12 |
| Thyroid hormone profile (umbilical cord blood) | |||
| TSH | mIU/L | 3.2 ± 0.5 (2.9–3.5) | 3.8 ± 0.8* (3.4–4.2) |
| FT4 | ng/dl | 1.2 ± 0.1 (1.1–1.2) | 0.9 ± 0.1* (0.8–0.9) |
| TT4 | ug/ml | 12.1 ± 1.4 (11.4–12.8) | 10.5 ± 1.8* (9.4–11.3) |
| TT3 | ng/ml | 1.9 ± 0.2 (1.8–2.0) | 1.0 ± 0.1* (0.9–1.0) |
NGT: Normal glucose tolerance; GDM: gestational diabetes mellitus; TSH: Thyroid Stimulant Hormone; FT4: Free Thyroxine: TT4: Total Thyroxine; TT3: Total triiodothyronine. Data correspond to mean ± S.D. (range).
*P<0.05 with respect to NGT.
There are few studies where TT4, FT4 or TT3 have been measured from GDM umbilical cord blood. A retrospective study with diet-treated GDM mothers, TSH levels in umbilical venous blood obtained at delivery was progressively increased with the severity of glucose intolerance in the mother, while FT4 levels in the fetus were normal, proposing the high TSH levels as a response to fetal hypoxic stress, rather than as a response to GDM [31]. Another study later observed a reduction in fetal TT4 and TT3 levels, and an increase in fetal TSH levels in conditions of GIGT with respect to control pregnancies [12]. These alterations were associated with a thyroid unbalance in the fetus. With this information is possible to suppose that placenta could play a role in the reduction of T3 and T4 levels in fetal blood via DIOs.
Deiodinase expression is altered in placenta from GDM pregnancies
We found a reduction (~47%) in the mRNA levels of DIO2; but an increase (~2.3 fold) in the mRNA levels of DIO3 in GDM compared to NGT pregnancies. mRNA levels of DIO1 were similar in both conditions (Fig 1). Accordingly, protein levels of DIO2 were reduced (~54%), but DIO3 was increased (~2.9 fold) in placenta from GDM pregnancies (Fig 2). Furthermore, it was not possible to determine DIO1 protein levels with this technique (not shown), as it has been evidenced in literature that DIO1 protein is not found in placenta [24, 32].
Fig 1. GDM effect on mRNA levels for deiodinases in human placenta.

Deiodinases (DIO1, DIO2, and DIO3) mRNA levels were measured using Q-RT-PCR (2-ΔΔCT) in placentas from gestational diabetes mellitus (GDM) and normal glucose tolerance (NGT) pregnancies. Values are mean ± SE. *P<0.05 compared to NGT pregnancies. n = 20 per group.
Fig 2. GDM effect on protein levels of deiodinase in placenta.

Deiodinase (DIO) protein levels were measured using western blot in placentae from normal glucose tolerant (NGT) and gestational diabetes mellitus (GDM) pregnancies. (A) DIO2 and (B) DIO3 protein levels in placentae from GDM and NGT pregnancies. β-actin was used as a load control. Results of densitometric analysis were expressed as DIO/β-actin. Values are mean ± SE. *P<0.05 compared to NGT. n = 8–9 per group.
To determine the expression and location of DIO2 and DIO3 proteins in the placenta, we used IHC. As mentioned above, we did not find any changes for DIO1 in placental tissue from NTG (Fig 3A–3D) and GDM (Fig 3G and 3H), which could be unspecified marker. Was determined that DIO2 and DIO3 are expressed mainly in the syncytiotrophoblast (Fig 3). Confirming previous results using Q-PCR and western blot, DIO2 had less immunoreactivity in GDM placentae (Fig 3H–3K) compared to NGT (Fig 3B–3E), whereas DIO3 had a major immunoreactivity in GDM (Fig 3I–3L) compared to NGT (Fig 3C–3F) placentae.
Fig 3. GDM effect on DIO2 and DIO 3 expression in placental tissue.

DIO2 and DIO3 immunodetection were realized by immunohistochemistry (IHC) in normal glucose tolerant (NTG, A-F) and gestational diabetes mellitus (GDM, G-L) pregnancies. In A-C and G-I 400x magnification, and in D-F and J-L 1000x magnification. White asterisk (*) in A-C and G-I indicates a magnification point in D-F and J-L, respectively. White arrow in D-F and J-L indicates syncytiotrophoblast in the placental tissue. Black bar: 50μm. N = 5 per group.
In this regard, other authors have described the presence of deiodinases in the placenta. Thus, mRNA expression levels of DIO2 were significantly higher in normal placenta during the first 20 weeks of pregnancy compared with term placenta, while in the other hand, DIO3 was significantly increased throughout gestation compared with term samples [33]. During the first trimester, the syncytiotrophoblast from normal placenta showed strong DIO3 and weak DIO2 immunoreactivities, while stronger DIO2 immunoreactivity compared to DIO3 during the third trimester was found [23, 33]. These results suggest that the higher expression of DIO3 may regulate T3 levels crossing from the mother to the fetus.
Currently, some studies have evidenced opposite changes in DIO2 and DIO3 in other pathologies. An increase in DIO2 and a decrease in DIO3 mRNA expression in the hippocampus, amygdala, and prefrontal cortex have been evidenced in mice with epilepsy [34]. On the other hand, in papillary thyroid tumors, it has been shown that the expression of DIO2 was reduced and DIO3 was increased, by the ERK signaling pathway [35]. Likewise, it has been observed that in an oxygen-glucose deprivation/reperfusion model (OGDR) of a human cardiomyocyte cell line (AC16, HCM), there is a decrease in the mRNA and protein levels of DIO2 and increase of DIO3 [36], which are similar results to those found in preterm infants coursing with intraventricular hemorrhage, where the authors suggest a potential mechanism mediated by hypoxia-ischemia and oxidative stress [37]. These findings are associated with studies in cerebral ischemia in rat [38], and oxidative stress in culture rat astrocytes [39]. All these changes suggest that these enzymes possibly are mediated by a competitive mechanism dependent of hypoxia and oxidative stress, which are conditions present in GDM as well [40, 41].
While no changes in deiodinases have been observed in immunoreactivity for other pregnancy pathologies such as preeclampsia [42], this is the first evidence showing significant difference in immunoreactivity between normal and GDM placenta for both DIO2 and DIO3.
Deiodinase activity is altered in placenta from GDM pregnancies
DIO activity is presented as kinetic enzymatic assays (Fig 4) and parameters (Table 3). We observed that T3 and rT3 formation from T4 were saturable in placentae from NGT and GDM pregnancies (Fig 4A–4C). Eadie-Hofstee plots were lineal in both groups of placentae. 5’deiodinase activity showed a reduction in Vmax (~35%), without changes in apparent Km in placentae from GDM compared to NGT (Table 3). However, the 5-deiodinase activity showed an increase in Vmax (~2.9 fold), without changes in apparent Km in GDM compared with NGT (Table 3).
Fig 4. GDM effect on 5’- and 5- deiodinase activity in human placenta.
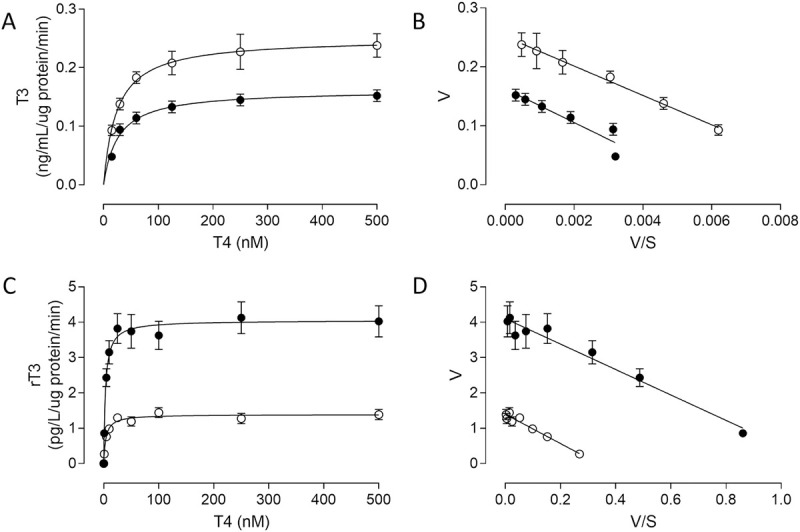
Kinetic of deiodinase activity was measured as T3 or rT3 formation from T4 (0–500 nM, 37°C, 15 min) in placental tissue (300μg placental protein) from Normal Glucose Tolerant (NGT, ○) and Gestational Diabetes Mellitus (GDM, ●) pregnancies. In A and C, T3 and rT3 formation from T4 adjusted to Michaelis-Menten equation. In B and D, Eadie-Hofstee plot from data in A and C, respectively. Mean ± SEM. N = 9 per group.
Table 3. Kinetic parameters of 5- and 5’- deiodinase activity.
| Kinetic parameters | NGT | GDM |
|---|---|---|
| 5-deiodinase activity (T4 → rT3) | ||
| Vmax (pg/L/μg of protein/min) | 1.39 ± 0.04 | 4.10 ± 0.09 * |
| Km (nM) | 4.14 ± 0.36 | 3.59 ± 0.27 |
| Vmax/Km (pg/L/μg of protein/min/nM) | 0.33 ± 0.03 | 1.14 ± 0.10* |
| 5’-deiodinase activity (T4 → T3) | ||
| Vmax (ng/mL/μg of protein/min) | 0.251 ± 0.003 | 0.163 ± 0.012* |
| Km (nM) | 24.92 ± 0.91 | 28.61 ± 5.64 |
| Vmax/Km (ng/mL/μg of protein/min/nM) | 0.010 ± 0.001 | 0.006 ± 0.002* |
Vmax: maximal velocity; Km: Michaelis-Menten constant; Vmax/Km: enzyme catalytic efficiency.
*P<0.05 with respect to NGT. N = 5 per group.
Deiodinase activity is classified in two types: (1) 5-deiodinase, favoring T3 formation from T4 mediated by DIO1 and DIO3, and (2) 5’-deiodinase, favoring rT3 formation from T4, or T2 from T3, mediated by DIO1 and DIO2.
An inverse relationship between FT4 and GDM has been confirmed, identifying a prevalence of GDM of 17.25% in the lowest FT4 quintile, decreasing to 11.62% in the highest FT4 quintile [6], which suggests changes in deiodinase activity in this type of patients. Another study confirmed the previous speculation that the T3/FT4 ratio (a measure of deiodinase activity) is related to maternal weight and glucose if weight-driven deiodinase activity may contribute to glycemic activity through T3 stimulation [29, 43].
The Km values that we obtained from 5-deiodinase activity are similar to those from previous experiments with that have measured DIO3 activity in placental tissue [44, 45], however, our Km values for 5’-deiodinase activity show less affinity for T4 than those that have been previously described for DIO2 activity in placental tissue [26, 46]. The reason for this could be that the methodology used (immunoassay versus radioactivity assay) the affinity for DIO2 in an unknown way, or that there is another, currently unknown enzyme or factor that affects it.
Studies in normal placenta have shown higher activity of DIO3 than DIO in first trimester and term placenta [33], while fetuses showed significantly lower TT3, but higher rT3 levels than newborns [47]. These results might be related with the DIO2 and DIO3 activity observed in the placenta. These changes in deiodinase activity have been associated with oxidative stress, specifically an increase in DIO3 activity and a decrease in DIO2 activity [39]. Our findings showing a change of deiodinases expression and activity in GDM placenta highlight the hypothesis that thyroid dysregulation occurs in the GDM fetus and that the required T3 for the fetus would not be optimal.
There were certain limitations in our study. Initially, the sample size was lower than total number of pregnant women recruited, due to miscarriage, suffering another, non-notified pregnancy pathology, or some of them no longer wanted to participate. Moreover, as we used term placentae, all the parameters analyzed are a reflect of the end of pregnancy and might not be accurate as a portrayal of the pregnancy course, specially of the first trimester.
Conclusions
An adequate concentration of THs is essential for the normal development of the fetus during pregnancy. At early stages, the mother provides THs, which are transported and regulated through the placenta before being released to the fetal blood. Deiodinases are responsible for the fine regulation of T3 and T4 that will be finally transported to the fetus, by inactivating T4 to rT3 (DIO2) or by converting T3 to T2 (DIO3), both rT3 and T2 having no apparent genomic biological functions. GDM mothers have higher levels of TT3, while expression and activity of DIOs in placenta are altered as well, as summarized in Fig 5. This leads to thyroid dysfunctions in the fetus even at early stages of the pregnancy. The higher levels of TT3 present in GDM mothers, regardless of the pregnancy trimester, are not reflected in the fetal blood, where both FT4 and TT3 are reduced in comparison to the mother. This could be explained as an altered compensatory mechanism to prevent an excess of T3 in the fetus, as any alteration could lead to an impaired development of the fetus [48, 49]. In conclusion, TH levels in GDM mothers are higher than those of normal mothers, peaking at the third trimester for both TT3 and TSH. The opposite was observed in the newborn, having decreased levels of FT4 and TT3 in the umbilical cord blood. This is associated with alteration of expression and activity of placental DIOs, observing a reduction of DIO2 and an increase of DIO3 in GDM pregnancies. The mechanism behind these alterations was not studied in this work, but it would be likely mediated by the hypoxia and oxidative stress present in GDM.
Fig 5. Proposal model for placental deiodinases in gestational diabetes mellitus (GDM) mothers.
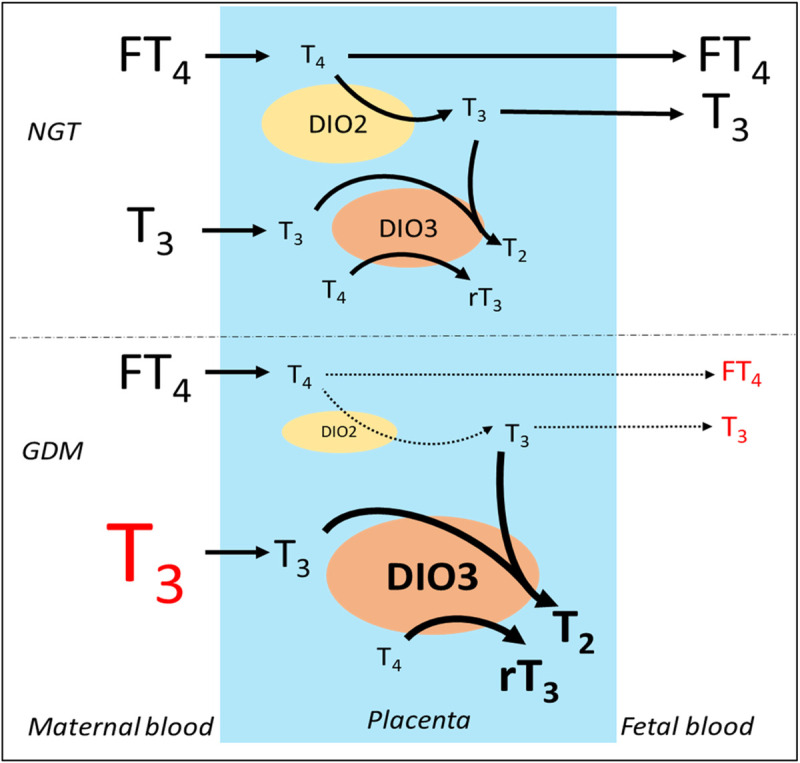
In normal glucose tolerant pregnancies (NGT), the thyroid hormones Free T4 (FT4) and T3 are transported from the maternal blood through the placenta, where their concentrations are regulated by deiodinase 2 (DIO2), which converts T4 to T3; and deiodinase 3 (DIO3), which converts T3 to T2, and T4 to rT3; in order to keep a steady and healthy concentration of thyroid hormones from the mother to the fetus. In GDM however, in order to prevent an influx of an excess of T3 from the mother to the fetus, the expression and activity of DIO3 increases while DIO2 decreases, causing less T3 to be transported to fetal blood by a currently unknown mechanism.
Therefore, this study constitutes the first approach regarding dysregulation of THs and placental DIO in the placenta from GDM. Another TH regulator involved in placenta, the Thyroid Hormone Transporters (THT), were not observed in this study, and will require further analysis. Therefore, the next goal is to understand how and why these changes happen are to observe placental THT in GDM mothers, to have the full picture behind this alteration.
Supporting information
(PPTX)
(XLSX)
Acknowledgments
We want to thank all the pregnant women and health personnel from the Family Health Centers (CESFAM) of Concepción (Chile): Santa Sabina, Tucapel and Dr. Victor Manuel Fernandez; and also Obstetrics and Gynecology Service staff of Hospital Guillermo Grant Benavente, who collaborated in the success of this work. AC, CE, MG, & EG-G are members of Group of Research and Innovation in Vascular Health (GRIVAS-Health, lead author: cescudero@ubiobio.cl).
Data Availability
All relevant data are within the manuscript and its Supporting Information files.
Funding Statement
This work was supported by the National Grant for Scientific and Technological Development of ANID, FONDECYT 11170710 and International Networks for Research REDI170287 (ANID) for EG-G, FONDECYT 1200250 for CE, and FONDECYT 1190250 for AL. DM is a PhD student of ANID (21190736). BO is a MSc student of ANID (22201750). Webpage: https://www.anid.cl/ The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.American Diabetes Association. 14. Management of Diabetes in Pregnancy: Standards of Medical Care in Diabetes-2019. Diabetes Care. 2019;42(Suppl 1):S165–S172. 10.2337/dc19-S014 [DOI] [PubMed] [Google Scholar]
- 2.Metzger BE, Buchanan TA, Coustan DR, de Leiva A, Dunger DB, Hadden DR, et al. Summary and recommendations of the Fifth International Workshop-Conference on Gestational Diabetes Mellitus. Diabetes Care. 2007;30 Suppl 2:S251–60. 10.2337/dc07-s225 [DOI] [PubMed] [Google Scholar]
- 3.Olivieri A, Valensise H, Magnani F, Medda E, De Angelis S, D'Archivio M, et al. High Frequency of Antithyroid Autoantibodies in Pregnant Women at Increased Risk of Gestational Diabetes Mellitus. Eur J Endocrinol. 2000;143(6):741–7. 10.1530/eje.0.1430741 [DOI] [PubMed] [Google Scholar]
- 4.Haddow JE, Craig WY, Neveux LM, Palomaki GE, Lambert-Messerlian G, Malone FD, et al. Free Thyroxine During Early Pregnancy and Risk for Gestational Diabetes. PLoS One. 2016;11(2):e0149065 10.1371/journal.pone.0149065 eCollection 2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zhang Y, Dai X, Yang S, Zhang C, Han M, Huang HF, et al. Maternal Low Thyroxin Levels Are Associated With Adverse Pregnancy Outcomes in a Chinese Population. PLoS One. 2017;12(5):e0178100 10.1371/journal.pone.0178100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Yang S, Shi FT, Leung PC, Huang HF, Fan J. Low Thyroid Hormone in Early Pregnancy Is Associated With an Increased Risk of Gestational Diabetes Mellitus. J Clin Endocrinol Metab. 2016;101(11):4237–4243. 10.1210/jc.2016-1506 [DOI] [PubMed] [Google Scholar]
- 7.Gong LL, Liu H, Liu LH. Relationship between hypothyroidism and the incidence of gestational diabetes: A meta-analysis. Taiwan J Obstet Gynecol. 2016;55(2):171–5. 10.1016/j.tjog.2016.02.004 [DOI] [PubMed] [Google Scholar]
- 8.Rawal S, Tsai MY, Hinkle SN, Zhu Y, Bao W, Lin Y, et al. A Longitudinal Study of Thyroid Markers Across Pregnancy and the Risk of Gestational Diabetes. J Clin Endocrinol Metab. 2018;103(7):2447–2456. 10.1210/jc.2017-02442 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Parkes IL, Schenker JG, Shufaro Y. Thyroid disorders during pregnancy. Gynecol Endocrinol. 2012;28(12):993–8. 10.3109/09513590.2012.692001 [DOI] [PubMed] [Google Scholar]
- 10.Pérez-López FR. Iodine and thyroid hormones during pregnancy and postpartum. Gynecol Endocrinol. 2007;23(7):414–28. 10.1080/09513590701464092 [DOI] [PubMed] [Google Scholar]
- 11.Wilker RE, Fleischman AR, Saenger P, Pan C, Surks MI. Thyroid hormone levels in diabetic mothers and their neonates. Am J Perinatol. 1984;1(3):259–62. 10.1055/s-2007-1000015 [DOI] [PubMed] [Google Scholar]
- 12.Shanmugam S, Dhiman P, Rajendiran S, Nimesh A, Maurya DK. Gestational impaired glucose tolerance (GIGT)-induced suppression of fetal thyroid secretion: effect on fetal outcome. J Matern Fetal Neonatal Med. 2019;32(12):1992–1996. 10.1080/14767058.2017.1422716 [DOI] [PubMed] [Google Scholar]
- 13.Guzmán-Gutiérrez E, Veas C, Leiva A, Escudero C, Sobrevia L. Is a low level of free thyroxine in the maternal circulation associated with altered endothelial function in gestational diabetes? Front Pharmacol. 2014;5:136 10.3389/fphar.2014.00136 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bianco AC, Dumitrescu A, Gereben B, Ribeiro MO, Fonseca TL, Fernandes GW, et al. Paradigms of Dynamic Control of Thyroid Hormone Signaling. Endocr Rev. 2019; 40(4):1000–1047. 10.1210/er.2018-00275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sugenoya A, Yamada Y, Kaneko G, Kobayashi M, Miyakawa M, Iida F. In vitro study on release of thyroid hormone in solitary autonomously functioning thyroid nodules using cell culture method. Endocrinol Jpn. 1984;31(6):749–53. 10.1507/endocrj1954.31.749 [DOI] [PubMed] [Google Scholar]
- 16.Rousset BA. Intracellular traffic and proteolytic cleavage of thyroglobulin, the thyroid prohormone. Ann Endocrinol (Paris). 1991;52(5):355–60. [PubMed] [Google Scholar]
- 17.Schussler GC. The thyroxine-binding proteins. Thyroid. 2000;10(2):141–9. 10.1089/thy.2000.10.141 [DOI] [PubMed] [Google Scholar]
- 18.Bianco AC, Kim BW. Deiodinases: implications of the local control of thyroid hormone action. J Clin Invest. 2006;116(10):2571–9. 10.1172/JCI29812 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Darras VM, Van Herck SL. Iodothyronine Deiodinase Structure and Function: From Ascidians to Humans. J Endocrinol. 2012;215(2):189–206. 10.1530/JOE-12-0204 [DOI] [PubMed] [Google Scholar]
- 20.Kilby MD, Barber K, Hobbs E, Franklyn JA. Thyroid hormone action in the placenta. Placenta. 2005;26(2–3):105–13. 10.1016/j.placenta.2004.08.004 [DOI] [PubMed] [Google Scholar]
- 21.Maruo T, Matsuo H, Mochizuki M. Thyroid hormone as a biological amplifier of differentiated trophoblast function in early pregnancy. Acta Endocrinol (Copenh). 1991;125(1):58–66. 10.1530/acta.0.1250058 [DOI] [PubMed] [Google Scholar]
- 22.Fatima SS, Rehman R, Butt Z, Asif Tauni M, Fatima Munim T, Chaudhry B, et al. Screening of subclinical hypothyroidism during gestational diabetes in Pakistani population. J Matern Fetal Neonatal Med. 2016;29(13):2166–70. 10.3109/14767058.2015.1077513 [DOI] [PubMed] [Google Scholar]
- 23.Huang SA, Dorfman DM, Genest DR, Salvatore D, Larsen PR. Type 3 iodothyronine deiodinase is highly expressed in the human uteroplacental unit and in fetal epithelium. J Clin Endocrinol Metab. 2003;88(3):1384–8. 10.1210/jc.2002-021291 [DOI] [PubMed] [Google Scholar]
- 24.Larsen PR, Zavacki AM. The role of the iodothyronine deiodinases in the physiology and pathophysiology of thyroid hormone action. Version 2. Eur Thyroid J. 2012;1(4):232–242. 10.1159/000343922 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Guzmán-Gutiérrez E, Westermeier F, Salomón C, González M, Pardo F, Leiva A, et al. Insulin-increased L-arginine transport requires A(2A) adenosine receptors activation in human umbilical vein endothelium. PLoS One. 2012;7(7):e41705 10.1371/journal.pone.0041705 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dora JM, Wajner SM, Costa JD, Pinto Ribeiro RV, Leiria LB, Lopes MG, et al. Type 2 deiodinase Thr92Ala polymorphism is associated with disrupted placental activity but not with dysglycemia or adverse gestational outcomes: a genetic association study. Fertil Steril. 2014;101(3):833–9. 10.1016/j.fertnstert.2013.11.018 [DOI] [PubMed] [Google Scholar]
- 27.Kundu S, Pramanik M, Roy S, De J, Biswas A, Ray AK. Maintenance of brain thyroid hormone level during peripheral hypothyroid condition in adult rat. Life Sci. 2006; 79(15):1450–5. 10.1016/j.lfs.2006.04.006 [DOI] [PubMed] [Google Scholar]
- 28.Mosso L, Martínez A, Rojas MP, Latorre G, Margozzini P, Lyng T, et al. Early pregnancy thyroid hormone reference ranges in Chilean women: the influence of body mass index. Clin Endocrinol (Oxf). 2016;85(6):942–948. 10.1111/cen.13127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Haddow JE, Metzger BE, Lambert-Messerlian G, Eklund E, Coustan D, Catalano P, et al. Maternal BMI, Peripheral Deiodinase Activity, and Plasma Glucose: Relationships Between White Women in the HAPO Study. J Clin Endocrinol Metab. 2019;104(7):2593–2600. 10.1210/jc.2018-02328 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Fraenkel M, Shafat T, Cahn A, Erez O, Novack V, Tsur A. Low thyroid-stimulating hormone and its persistence beyond the first trimester of pregnancy. Int J Gynaecol Obstet. 2018;142(3):270–276. 10.1002/ijgo.12540 [DOI] [PubMed] [Google Scholar]
- 31.Lao TT, Lee CP. Gestational diabetes mellitus and neonatal hyperthyrotropinemia. Gynecol Obstet Invest. 2002;53(3):135–9. 10.1159/000058363 [DOI] [PubMed] [Google Scholar]
- 32.Bianco AC, da Conceição RR. The Deiodinase Trio and Thyroid Hormone Signaling. Methods Mol Biol. 2018; 1801: 67–83. 10.1007/978-1-4939-7902-8_8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chan S, Kachilele S, Hobbs E, Bulmer JN, Boelaert K, McCabe CJ, et al. Placental iodothyronine deiodinase expression in normal and growth-restricted human pregnancies. J Clin Endocrinol Metab. 2003;88(9):4488–95. 10.1210/jc.2003-030228 [DOI] [PubMed] [Google Scholar]
- 34.Nascimento BPP, Bocco BMLC, Fernandes GW, Fonseca TL, McAninch EA, Cardoso CV, et al. Induction of Type 2 Iodothyronine Deiodinase After Status Epilepticus Modifies Hippocampal Gene Expression in Male Mice. Endocrinology. 2018;159(8):3090–3104. 10.1210/en.2018-00146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Romitti M, Wajner SM, Zennig N, Goemann IM, Bueno AL, Meyer EL, et al. Increased type 3 deiodinase expression in papillary thyroid carcinoma. Thyroid. 2012; 22(9):897–904. 10.1089/thy.2012.0031 [DOI] [PubMed] [Google Scholar]
- 36.Li Q, Qi X, Jia W. 3,3',5-triiodothyroxine inhibits apoptosis and oxidative stress by the PKM2/PKM1 ratio during oxygen-glucose deprivation/reperfusion AC16 and HCM-a cells: T3 inhibits apoptosis and oxidative stress by PKM2/PKM1 ratio. Biochem Biophys Res Commun. 2016;475(1):51–6. 10.1016/j.bbrc.2016.05.030 [DOI] [PubMed] [Google Scholar]
- 37.Vose LR, Vinukonda G, Jo S, Miry O, Diamond D, Korumilli R, et al. Treatment with thyroxine restores myelination and clinical recovery after intraventricular hemorrhage. J Neurosci. 2013;33(44):17232–46. 10.1523/JNEUROSCI.2713-13.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Margaill I, Royer J, Lerouet D, Ramaugé M, Le Goascogne C, Li WW, et al. Induction of type 2 iodothyronine deiodinase in astrocytes after transient focal cerebral ischemia in the rat. J Cereb Blood Flow Metab. 2005;25(4):468–76. 10.1038/sj.jcbfm.9600041 [DOI] [PubMed] [Google Scholar]
- 39.Lamirand A, Pallud-Mothré S, Ramaugé M, Pierre M, Courtin F. Oxidative stress regulates type 3 deiodinase and type 2 deiodinase in cultured rat astrocytes. Endocrinology. 2008;149(7):3713–21. 10.1210/en.2007-1462 [DOI] [PubMed] [Google Scholar]
- 40.Lappas M, Hiden U, Desoye G, Froehlich J, Hauguel-de Mouzon S, Jawerbaum A. The role of oxidative stress in the pathophysiology of gestational diabetes mellitus. Antioxid Redox Signal. 2011;15(12):3061–100. 10.1089/ars.2010.3765 [DOI] [PubMed] [Google Scholar]
- 41.Cvitic S, Desoye G, Hiden U. Glucose, insulin, and oxygen interplay in placental hypervascularisation in diabetes mellitus. Biomed Res Int. 2014;2014:145846 10.1155/2014/145846 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kurlak LO, Mistry HD, Kaptein E, Visser TJ, Broughton Pipkin F. Thyroid hormones and their placental deiodination in normal and pre-eclamptic pregnancy. Placenta. 2013;34(5):395–400. 10.1016/j.placenta.2013.02.009 [DOI] [PubMed] [Google Scholar]
- 43.Haddow JE, Lambert-Messerlian G, Eklund E, Neveux LM, Palomaki GE. Peripheral deiodinase activity: A potential explanation for the association between maternal weight and gestational hyperglycemia. Obstet Med. 2018;11(2):73–78. 10.1177/1753495X17733223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kuiper GG, Klootwijk W, Visser TJ. Substitution of cysteine for selenocysteine in the catalytic center of type III iodothyronine deiodinase reduces catalytic efficiency and alters substrate preference. Endocrinology. 2003;144(6):2505–13. 10.1210/en.2003-0084 [DOI] [PubMed] [Google Scholar]
- 45.Bessho K, Etani Y, Ichimori H, Miyoshi Y, Namba N, Yoneda A, et al. Increased type 3 iodothyronine deiodinase activity in a regrown hepatic hemangioma with consumptive hypothyroidism. Eur J Pediatr. 2010;169(2):215–21. 10.1007/s00431-009-1009-x [DOI] [PubMed] [Google Scholar]
- 46.Koopdonk-Kool JM, de Vijlder JJ, Veenboer GJ, Ris-Stalpers C, Kok JH, Vulsma T, et al. Type II and type III deiodinase activity in human placenta as a function of gestational age. J Clin Endocrinol Metab. 1996;81(6):2154–8. 10.1210/jcem.81.6.8964844 [DOI] [PubMed] [Google Scholar]
- 47.Santini F, Chiovato L, Ghirri P, Lapi P, Mammoli C, Montanelli L, et al. Serum iodothyronines in the human fetus and the newborn: evidence for an important role of placenta in fetal thyroid hormone homeostasis. J Clin Endocrinol Metab. 1999; 84(2):493–8. 10.1210/jcem.84.2.5439 [DOI] [PubMed] [Google Scholar]
- 48.Zimmerman D. Fetal and neonatal hyperthyroidism. Thyroid. 1999;9(7):727–33. 10.1089/thy.1999.9.727 [DOI] [PubMed] [Google Scholar]
- 49.Haddow JE, Palomaki GE, Allan WC, Williams JR, Knight GJ, Gagnon J, et al. Maternal thyroid deficiency during pregnancy and subsequent neuropsychological development of the child. N Engl J Med. 1999;341(8):549–55. 10.1056/NEJM199908193410801 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(PPTX)
(XLSX)
Data Availability Statement
All relevant data are within the manuscript and its Supporting Information files.