

Abstract
Nitric oxide (NO) is a gasotransmitter of great significance to developing the innate immune response to many bacterial and viral infections, while also modulating vascular physiology. The generation of NO from the upregulation of endogenous nitric oxide synthases serves as an efficacious method for inhibiting viral replication in host defense and warrants investigation for the development of antiviral therapeutics. With increased incidence of global pandemics concerning several respiratory-based viral infections, it is necessary to develop broad therapeutic platforms for inhibiting viral replication and enabling more efficient host clearance, as well as to fabricate new materials for deterring viral transmission from medical devices. Recent developments in creating stabilized NO donor compounds and their incorporation into macromolecular scaffolds and polymeric substrates has created a new paradigm for developing NO-based therapeutics for long-term NO release in applications for bactericidal and blood-contacting surfaces. Despite this abundance of research, there has been little consideration of NO-releasing scaffolds and substrates for reducing passive transmission of viral infections or for treating several respiratory viral infections. The aim of this review is to highlight the recent advances in developing gaseous NO, NO prodrugs, and NO donor compounds for antiviral therapies; discuss the limitations of NO as an antiviral agent; and outline future prospects for guiding materials design of a next generation of NO-releasing antiviral platforms.
Keywords: Nitric oxide, Viral infection, Severe acute respiratory distress, COVID-19, Inhalation therapy
Abbreviations: HI, Host Immunology; ACE, angiotensin converting enzyme; AP1, activator protein 1; IFN, interferon; IFNγ, interferon gamma; IKK, inhibitor of nuclear factor kappa B kinase; IRF-1, interferon regulatory factor 1; NF-κB, nuclear factor kappa-light-chain enhancer of activated B cells; P38-MAPK, P38 mitogen-activated protein kinases; PAMP, pathogen-associated molecular pattern; PKR, protein kinase R; STAT-1, signal transducer and activator of transcription 1; TAK1, transforming growth factor β-activated kinases-1; TLR, toll-like receptor; Medical Terminology: ARDS, acute respiratory distress syndrome; COVID-19, coronavirus disease 2019; ECMO, extracorporeal membrane oxygenation, FDA, United States Food and Drug Administration; I/R, pulmonary ischemia-reperfusion; PHT, pulmonary hypertension; SARS, severe acute respiratory syndrome; VAP, ventilator associated pneumonia; Nitric Oxide and Related Compounds: eNOS/NOS 3, endothelial nitric oxide synthase; gNO, gaseous nitric oxide; GNSO, S-nitrosoglutathione; iNOS/NOS 2, inducible nitric oxide synthase; mtALDH, mitochondrial aldehyde dehydrogenase; NO, nitric oxide; NOS, nitric oxide synthase; nNOS/NOS 1, neuronal nitric oxide synthase; RNI, reactive nitrogen intermediate; RSNO, S-nitrosothiol; SNAP, S-nitroso-N-acetyl-penicillamine; Viruses: CVB3, coxsackievirus; H1N1, influenza A virus subtype H1N1; HIV, human immunodeficiency virus; HPV, human papillomavirus; HSV, herpes simplex virus; PCV2, porcine circovirus type 2; SARS-CoV-2, severe acute respiratory syndrome coronavirus 2; Other: DNA, deoxyribonucleic acid; dsRNA, double stranded (viral) ribonucleic acid; IC50, inhibitory concentration 50; RNA, ribonucleic acid
Graphical Abstract

1. Introduction
The current and projected global health and economic ramifications of a viral pandemic emphasize the need to develop robust antiviral treatments and corresponding mitigative efforts to enable rapid response. Concerns with the inflexibility of current therapies for novel viruses have prompted two needs: 1) comprehensive antiviral treatments, and 2) enhancements to conventional medical devices (e.g. nasal cannulas, mechanical ventilators, and extracorporeal membrane oxygenation) to improve patient outcomes. Even at an accelerated rate, vaccine development for emergent viruses is optimistically expected to take at least 12–18 months [1]. Rapid mutations associated with many viruses including human immunodeficiency virus (HIV) and influenza present a significant challenge in vaccine development, emphasizing the need for nonspecific antiviral treatments. A broad-spectrum agent exhibiting multi-mechanistic antiviral activity could be used as a tool to combat infections caused by an array of viruses, reduce instances of drug resistance, and alleviate additional costs in developing virus-specific drugs [2]. Moreover, much like other viral pandemics, early reports on the 2019 severe acute respiratory syndrome coronavirus (SARS-CoV-2) outbreak have agreed that secondary infections are a significant predictor of patient outcome, affecting up to 50% of non-survivors [3], [4], [5], [6]. Secondary infections are not new – cumulatively, 86% of pneumonia infections in critically ill patients are related to mechanical ventilation [7]. Viral outbreaks coupled with frequent secondary bacterial infections have devastating effects on patient care – increasing disease severity, requiring supplementary treatment, and significantly impacting patient morbidity and mortality [8]. Therefore, in addition to the pressing need for broad-spectrum antiviral agents capable of treating a range of DNA and RNA viruses, there is significant interest in developing multifaceted therapeutic strategies that not only inhibit viral invasion but also prevent secondary bacterial infections.
Prior to its discovery as the endothelium derived relaxing factor in the 1980s, nitric oxide (NO) had been categorized as an atmospheric pollutant, a precursor of acid rain, and believed carcinogen [9,10]. After recognizing the role of NO as a cardiovascular signaling molecule, co-recipients Louis Ignarro, Robert Furchgott, and Ferid Murad were awarded the 1998 Nobel Prize in Physiology or Medicine, thus encouraging researchers to further examine other physiological roles of NO. Within the next decade, NO was found to play an important role in digestion, blood pressure regulation, neural function, platelet regulation, and immune response [10,11]. As a result, researchers at the turn of the century began to use antiviral NO-based therapeutics through three different strategies: 1) compounds that modulate endogenous production of NO, 2) gaseous NO (gNO) inhalation, and 3) direct NO donor compounds. More recently, NO-releasing and catalytic NO-generating surfaces have been developed for blood-contacting and other indwelling medical devices to combat bacterial infection and prevent blood thrombus formation that are often associated with urinary/vascular catheters, mechanical ventilators, and ex vivo blood circulation in dialysis and extracorporeal membrane oxygenation (ECMO) [12], [13], [14].
While the antibacterial activity of NO is well-documented, researchers have also shown NO-mediated antiviral mechanisms against several DNA and RNA viruses. During the 2002-2004 SARS outbreak, a rescue trial found that gNO treatments both improved oxygenation and reduced lung infiltrates associated with the viral infection, therefore warranting further investigation of NO strategies [15]. Preliminary findings of NO's role in viral clearance has merited further examination of its comprehensive antiviral and antibacterial capacities and has earmarked its potential through NO-related treatment strategies and modifications to existing medical devices to combat current and future infections. In this review, we provide an in-depth discussion of NO's role in the pathogenesis of viral infections, survey current NO-based treatment strategies, and identify key gaps for overcoming the limited clinical translation of antiviral NO-based technologies to date.
2. Biology and in vivo Metabolism of NO
NO contributes to and/or regulates many physiological responses, commonly through cyclic guanosine monophosphate (cGMP) pathways. Within the vasculature, NO released from the endothelium temporarily inhibits platelet activation – preventing thrombosis [16]. NO also causes vasodilation by signaling for smooth muscle cells to relax and promotes angiogenesis [17,18]. In the gastrointestinal tract, NO contributes to vascular tone regulation in addition to promoting gastric mucosal integrity [19]. Within the nervous systems, NO acts as a neurotransmitter by stimulating cGMP pathways, which modulate excitatory neurotransmission and inhibit synaptic transmission [20]. Finally, NO is an instrumental component of the immune system, with activated macrophages producing NO for antimicrobial, antitumor, and antiviral effects [21].
NO is produced endogenously from L-arginine via three NO synthase (NOS) isoforms: neuronal (nNOS or NOS1) found in neuronal and skeletal muscle tissues; inducible (iNOS or NOS2) found in macrophages; and endothelial (eNOS or NOS3) found in vascular endothelial cells [22]. Activation of these isozymes is mediated by binding to calmodulin, with nNOS and eNOS binding being calcium dependent. Whereas nNOS and eNOS are constitutive – producing low levels of NO, iNOS activity is insensitive to physiological changes of calcium which enables prolonged, higher concentrations of NO to be developed – making it a crucial mediator of the immune response [23]. While endogenous NO plays many beneficial roles in the human body, it is often considered a “double-edged sword” [20,24]. Different NO concentrations are related to different physiological responses, with low concentrations (nM range) facilitating cellular proliferation and higher concentrations (µM range) inducing cellular apoptosis and cell cycle arrest [25]. In infected M1 macrophages, NO concentrations are known to exceed several hundred µM [26]. Elevated and/or decreased levels of NO have been implicated in contributions to neuronal destruction, cardiovascular diseases, cancer, obesity, insulin resistance, and inflammatory conditions [24,[27], [28], [29]. Therefore, it is important to consider NO in a tissue and concentration-dependent context when developing NO-based therapeutics.
The balance between beneficial and detrimental effects of NO depends on its steady-state concentration, considering both its rates of formation and decomposition. As a free radical, NO has several modes of reaction in the human body: oxidation to form nitrosonium cations (NO+); reduction to form nitroxyl anions (NO−); autooxidation to form dinitrogen trioxide (N2O3), which rapidly converts to nitrite (NO2 −); and reaction with superoxide radical anions to form reactive nitrogen species such as peroxynitrite (ONOO−) [30,31]. Still further, the autooxidation process of NO forming N2O3 can lead to the nitrosation of protein Cys residues and the formation of S-nitrosothiols (RSNOs), which are heavily involved in cell signaling and regulatory processes [32]. In the vasculature, NO is rapidly converted to nitrate (NO3 −) via oxyhemoglobin, which contributes to its approximate half-life of 0.1–2 s as a local modulator of vasodilatory response [33]. Whether exogenously inhaled, endogenously synthesized, or otherwise formed from intravascular drug administration, NO and its metabolic derivatives undergo a variety of localized catabolic processes as summarized in Fig. 1 . Overall, several key factors have been shown to contribute to rates of NO's in vitro catabolism, including: (1) its reactivity with catalytic metal ions; (2) localized oxygen concentration and redox environment; (3) cell type; and (4) tissue diffusivity [34], [35], [36]. Readers are directed to several recent reviews on NO metabolism for further elaboration on the subject matter [31,36,37]. Any prospective antiviral therapy stemming from NO or its many derivatives must therefore make careful consideration of its concentration and localization specific effects in design of both the active agent and delivery mechanism.
Fig. 1.

Pathways for the production and consumption of nitric oxide (NO) and its related metabolic products. Exogenous nitrogen oxide species (NOx) and endogenously produced NO are shuffled to the vasculature. Basal levels of NO are stored in the blood via low molecular weight S-nitrosothiols (RSNOs). Otherwise, NO is rapidly converted to nitrite (NO2−) and nitrate (NO3−). Nitrites and nitrates are ultimately discharged directly or converted to other species such as ammonia (NH3) and nitrogen (N2) for removal.
3. Antiviral mechanisms of NO
Endogenous NO has long been recognized for its capacity to modulate vascular physiology and defend against microbial infection, though its role in antiviral mechanisms is less understood. The endogenous antiviral mechanism of NO begins with stimulation of iNOS through one of several mechanisms (see Fig. 2 A). NO production from iNOS is critical to the inflammatory response and is supported by type I interferon (IFN) host defense against viral infection in key cell types. Several pathways are activated when viruses invade cells and mediate iNOS upregulation to lead to NO production and virus elimination: The first of these pathways is based on toll-like receptors (TLRs) in immune and non-immune cells (macrophages, T and B lymphocytes, and epithelial cells). TLRs respond to both DNA and RNA viruses, detect pathogen-associated molecular patterns (PAMPs), and activate both nuclear factor kappa-light-chain enhancer of activated B cells (NF-κB) and activator protein 1 (AP1) via transforming growth factor β-activated kinases-1 (TAK1). AP1 enters cellular nuclei to upregulate iNOS and subsequent NO production [38]. A second pathway for iNOS regulation develops from the activation of signal transducer and activator of transcription 1 (STAT-1). Approximately 24–36 h after viral infection, interferon gamma (IFNγ) produced by lymphocytes regulates STAT-1 and leads to the upregulation of iNOS. The third and final major pathway targets double stranded viral RNAs (dsRNAs) involved in viral replication within the host cell. Viral dsRNAs are products of both DNA and RNA-based viral infections and are potent inducers of IFNs. Viral dsRNAs bind to and activate protein kinase-R (PKR), leading to signal transduction for iNOS upregulation [39]. Following iNOS upregulation by any one of the above three pathways, L-arginine is readily converted to L-citrulline to produce NO that freely diffuses across cellular membranes to inhibit viruses.
Fig. 2.

Overview of endogenous nitric oxide (NO) production in response to host viral infection. a) Host recognition of viral DNA initiates one of many signal transduction pathways that converge with iNOS upregulation, leading to increased endogenous NO. NO has several antiviral effects in host defense, including: b) nitrosylation of Cys residues leading to viral enzyme deactivation, c) formation of reactive nitrogen species such as peroxynitrite that induce DNA strand breaks, and d) suppression of viral transcription factors leading to inhibition of viral replication and disease state propagation.
NO has several direct modes of action as an antiviral agent, including: 1) inhibition of viral enzymes through nitrosylation of viral proteins; 2) damage to viral DNA via oxidative and nitrosative stress during pathogenesis; 3) modulation of viral-encoded transcription factors; and 4) further activation of host signaling pathways for adaptive response (see Fig. 2B-D) [38,[40], [41], [42]. NO appears to be effective in the clearance of both DNA and RNA viruses based on various in vitro and in vivo studies [38,43]. NO is a highly unstable free radical that reacts with oxygen, superoxide, and hydrogen peroxide to generate reactive nitrogen intermediates (RNIs), such as peroxynitrite (ONOO−), that in turn react with thiol moieties on cysteine residues of both viral and host proteins [41]. These nitrosylation reactions, especially on viral proteases, reductases, and reverse transcriptases interrupt the viral replication cycle, prevent further infectivity of virions, and enable more efficient host clearance [41]. Other antiviral effects of NO center on its activity towards viral transcription and replication. Specifically, NO has the observed effect of modulating early transcript levels of some viruses such as human papillomavirus (HPV), while preventing further viral DNA replication [44]. RNIs developed from NO such as peroxynitrite have shown only modest inhibition of viral replication in host cells, though viability of free virions was significantly impaired. These results corroborated against tests with direct NO donors have suggested that the reactive nitrogen intermediate formed can significantly affect viral clearance [45]. RNIs tend to induce strand breakages, deamination of adenine and guanine bases, and other oxidative damage near indiscriminately against both viral and host DNAs/RNAs, though host genomes are more robust in encoding repair nucleases and polymerases. Because of the inherent limitations of viral genome sizes, most viruses do not encode repair mechanisms for oxidative damage and instead rely on integration into the host genome to mediate repair by host machinery – in analogy to the pathogenesis of many RNA viruses [46]. Readers are directed to a recent review by Uehara et al. [47] that further covers the antiviral mechanisms of NO, it's potentially deleterious effects in certain disease states, and its use as a potential prognostic marker.
In many circumstances, NO's role in host viral infection is time and concentration dependent as well as contingent on viral pathogenesis. In some circumstances, endogenous NO production is inhibited, and exogenous NO may be necessary to provide an antiviral effect. For example, the overproduction of NO by iNOS can downregulate many inflammatory cytokines and chemokines based on the inhibition of both NF-κB and interferon regulatory factor 1 (IRF-1) dependent transcription in epithelial cells [48,49]. In a human rhinovirus infection study, endogenous NO production was limited by feedback inhibition through NF-κB and IRF-1 pathways, with exogeneous NO otherwise eliciting a desired antiviral effect [49]. Additionally, some antiviral components such as acetylsalicylic acid [50], chebulagic acid, and punicalagin [51] have been shown to suppress iNOS activity via the IKK-NF-κB and P38-MAPK signaling pathways. In other circumstances, viruses can actively suppress NO production through downregulation of iNOS expression via inhibition of NF-κB, as has been demonstrated with HIV-1 [52,53]. SARS-CoV is also reported to have its membrane protein (SARS-CoV M) suppress NF-κB activity through interaction with inhibitor of nuclear factor kappa-B kinase subunit beta, which further indicates the need for exogenous NO therapeutics [54].
Endogenous NO production must closely balance inflammation response with antiviral effects. Otherwise, excessive NO is produced and can lead to several complications such as: viral pneumonia from influenza [55,56]; various neurological diseases tied to HIV-1 [57] and Dengue viral infections [58]; and late stage upregulation of iNOS that leads to enhanced viral replication in the pathogenesis of HIV-1 infection [57]. In short, NO is by no means a “magic bullet” against host viral infections, but the development of NO-based therapies holds value for many host infections and the eradication of free virions [59].
4. Survey of NO-based antiviral therapies
NO-based antiviral therapies are broadly categorized into three strategies: 1) drugs that affect NOS regulation or endogenous production of NO; 2) gNO inhalation therapies; and 3) direct NO donor compounds. The bulk of existing antiviral NO therapeutic strategies target strains specific to respiratory tract infections, including: adenovirus, coronavirus, human respiratory syncytial virus, influenza A and B viruses, human parainfluenza viruses, and rhinovirus, among others [40,41,60]. Upper respiratory tract viral infections are often associated with increased lower airway exhaled NO, which as a host defense strategy has shown clearance ability against rhinoviruses [61,62], coronaviruses [63], herpes simplex virus (HSV) [64], and human influenza [65] in addition to many other bacterial, fungi, and viral strains [66]. Other chronic inflammatory diseases such as asthma and bronchiectasis are associated with increased endogenous NO [67]. Therefore, the primary considerations of any antiviral NO therapeutic strategy are: 1) the effect of NO release/scavenging on the underlying pathology, 2) the effect of the mode of delivery on therapeutic action, and 3) parallel effects of NO on pulmonary circulation and vascular physiology.
Reiss et al. provide an excellent summary of the roughly first decade of in vitro and in vivo studies with NO for viral infection [59]. A summary of more contemporary clinical trial work with gNO and dissolved NO donor therapies as well as other promising in vivo studies are summarized in Table 1 . Many in vitro and ex vivo studies performed to date demonstrate the potency of both gNO and direct NO donor compounds for reducing titers or otherwise inhibiting RNA replication of a number of viral strains, including: porcine circovirus [68], coronavirus [69], human influenzas A and B (including H1N1) [65,70], human rhinovirus [48,61], Hantaan hantavirus [45], Crimean Congo hemorrhagic fever virus [71], HPV [72,73], and HSV and vaccinia [74]. A summary of these in vitro studies, their dosages, and infection models is provided in Table 2 .
Table 1.
Summaries of clinical trial work and in vivo studies with gNO and NO donor compounds as active antiviral agents.
 |
Table 2.
Summaries of in vitro and ex vivo studies with gNO and NO donors as antiviral agents.
 |
4.1. Endogenous NO Regulating Drugs
Several antiviral compounds have been developed to attenuate the physiological formation of NO via their metabolic degradation into NO or modulation of iNOS expression. One such example is hydroxyurea, which has shown antiviral effects through the inhibition of viral replication as a ribonucleotide reductase inhibitor and modulator of intracellular deoxynucleotide availability [75]. Hydroxyurea is metabolically oxidized into NO and has shown promise in several therapies against HIV-1, parvovirus, and hepatitises B and C [76], [77], [78], [79]. Other strategies to create NO donor prodrugs include the glycosylation or vinylation of diazeniumdiolates, though no reports of their application as antiviral agents were found in literature to date [80,81]. Similarly, other NO prodrug classes such as furoxans, nitrates, and sydnonimines have shown efficacy in preclinical drug development for anti-cancer and Alzheimer's disease therapies, though to date little consideration has been made for their potential antiviral application [82], [83], [84], [85], [86].
Beyond NO donor prodrugs, other compounds have been designed to modulate iNOS activity for intended antiviral effect. Bilirubin has been shown to upregulate NO release in achieving viricidal activity against HSV-1 [87]. In contrast, many antiviral drug therapies selectively inhibit iNOS for antiviral effects, such as various dihydropyrazole derivatives against vaccinia virus [88] as well as ribavirin and acetylsalicylic acid against hepatitis C virus, wherein increased NO production may contribute towards the pathophysiology and clinical manifestation of the viral disease state [50,89]. Finally, several NO scavenging compounds such as PTIO (CAS 183900-00-6), hemoglobin derivatives, low molecular weight iron compounds, and cerium oxide have been investigated for treating NO mediated diseases, though not as antiviral compounds [90]. Because of the many possible deleterious effects from aiding viral immunopathogenesis by upregulating iNOS, many therapies have instead opted to explore direct application of gNO, prodrugs, and direct donor compounds for viral strains attenuated by increased NO production [47].
4.2. gNO Therapies
Direct gNO therapies have been investigated since the 1980’s for treating a variety of medical conditions, including: acute respiratory distress syndrome (ARDS) [91], persistent pulmonary hypertension (PHT) in neonates [92], reversible pulmonary vasoconstriction [93], bronchoconstriction [94], cystic fibrosis [95,96], and various pneumonias among other medical conditions [97], [98], [99]. Although therapies such as INOmaxTM have received FDA approval for treating PHT in neonates [100], gNO has received mixed reception in literature because of its dose sensitivity thresholds for antiviral and antimicrobial action in parallel to those for vasodilation and platelet quiescence [66,101]. One prominent example of this is seen in gNO treatments for ARDS – a condition prompted by diverse etiologies such as SARS-related coronavirus (SARS-CoV) infection, pulmonary ischemia-reperfusion (I/R) injury, and bacterial/viral pneumonias, among others [102]. Treatment with gNO for ARDS manifested from SARS-CoV infection has shown to improve arterial oxygenation and reduce pneumonia infiltrates in rescue trials [15]. On the other hand, some reports have shown that gNO can intensify I/R-induced lung injury and be a causative agent of endotoxin-induced ARDS in various in vivo models of acute lung injury in mice [103,104]. Recent retrospective analyses of gNO therapeutic clinical trials for refractory hypoxemia from ARDS have found insufficient evidence to support any overall benefit for its treatment with gNO [105,106]. These results are supported by similar studies of gNO inhalation therapies for severe influenza within in vivo mouse models, which found that intermittent treatment at even 160 ppm of NO failed to reduce lung viral load [107]. Regardless, a rescue trial [15] for SARS-CoV infections (during the 2002–2004 outbreak) evidenced against promising in vitro work with NO donors [69] has prompted several ongoing clinical trials that aim to develop gNO treatments against coronavirus disease 2019 (COVID-19) that address: 1) early stage infection to prevent further propagation (NCT04305457); 2) mechanically ventilated patients to improve oxygenation in hypoxic SARS-CoV-2 infection (NCT04306393); and 3) hospital-associated transmission among healthcare workers (NCT04312243) [108], [109], [110], [111]. Another ongoing trial (NCT03331445) is adapting gNO therapies originally conceived against non-tuberculosis mycobacteria for application as an antiviral therapy to reduce intervention with mechanical ventilators and the need for oxygen therapies [112].
To date, gNO has debatably had the most success as an FDA-approved drug under the tradename INOmaxTM for treating oxygen desaturation at recommended dosages of 20 ppm for up to two weeks in neonates with hypoxic respiratory failure and to help avoid further intervention with ECMO. gNO therapies are often used because of minimized systemic effects owed to the short half-life of NO in physiological conditions [113,114], though exact dosages needed for antiviral action may be well above those for attenuating pulmonary vasculature. For example, while less than 40 ppm is cited as eliciting pulmonary vasoconstriction without dilation [93], 80 ppm failed to elicit any antimicrobial effect in clinical trials [101]. Furthermore, most in vitro studies point to excesses of 160 ppm being needed for full antimicrobial effect, while still being largely dependent on the inhalation period and physiological environment [66,98,115]. Although a rescue trial of SARS patients during the 2002–2004 outbreak demonstrated reduced lung infiltrates with a stepped-down treatment regimen from 30 ppm over three days, the authors advised caution towards interpretation as an efficacious antiviral treatment strategy because of the small treatment group. Regardless, gNO was found effective in its primary role for improving oxygen saturation [15].
Currently, a large gap in knowledge remains in understanding differences in therapeutic outcomes observed in vitro with gNO treatments compared to those with dissolved NO donor compounds. Recent work testing the antimicrobial efficacy of gNO versus a NO-releasing soluble chitosan oligosaccharide demonstrated that excesses of 500 ppm of gNO were necessary to elicit the same 3-log reduction of Pseudomonas aeruginosa observed with the soluble donor, despite the gNO treatment reportedly leading to almost a 20x excess in dissolved NO compared to the donor compound [115]. Other studies have shown corroborative results and emphasize that the success of gNO therapies is contingent on continuous replenishment of gNO because of limitations in phase transfer to the physiological medium coupled with physiological buffering and the short half-life of gNO [96]. In contrast, dissolved NO donors were shown to achieve the same antimicrobial thresholds with a one-off treatment because of the enhanced stability and controlled degradation of NO-donating compounds in physiological conditions [96,115]. For these reasons as well as ease of application, storage stability, and ready clinical translation, NO-donating compounds as antiviral agents have been shown increased interest and will be the focus of the remainder of this section.
4.3. Direct NO donor therapies
NO donor compounds used as antiviral agents fall under three general classifications: nitrites, N-diazeniumdiolates, and S-nitrosothiols (RSNOs). The earliest antiviral clinical trial work with NO donors tested acidified nitrites in topical ointments against molluscum contagiosum infection but faced several issues with skin discoloration, irritation, and other side effects [116]. While there has been limited academic literature with NO donors in antiviral clinical trial work, significant effort has been made to commercialize an investigational product named SB206 (NOVAN Inc, Morrisville, NC, USA) for application against molluscum contagiosum and HPV infection. The active component of SB206 is a propriety NO-releasing compound NVN1000 (berdazimer sodium, CAS 1846565-00-1, NOVAN Inc.) that has previously shown promising antiviral capacity in vitro with HPV raft cultures [72]. Combined work on this product has been successful through a phase II clinical trial against HPV with significant improvements in wart clearance (NI-MC201) and is undergoing continued testing in phase III trials against molluscum contagiosum (NI-MC301, NI-MC302) [73,117,118].
Most other contemporary antiviral work with NO donor compounds has involved small molecule, synthetic RSNOs such as S-nitroso-N-acetylpenicillamine (SNAP) due to their favorable stability in physiological conditions (see Fig. 3 a). Most recently, Akaberi et. al [119] demonstrated reduction in SARS-CoV-2 protease activity with up to 99.42% replication inhibition using low doses of SNAP with a SARS-CoV-2 infection model in Vero E6 cells (see Fig. 3b). Studies such as these have been mostly relegated to in vitro models of viral infection [59,64,70,[119], [120], [121], [122]; however, SNAP and various other RSNO derivatives have also shown promising antibacterial, anti-thrombotic, and non-cytotoxic results in other in vivo models of blood circulation and urinary tract infection [12,14,123]. These applications incorporated NO donors such as SNAP or S-nitrosoglutathione (GSNO) into medical grade polymers, polysaccharides, and various macromolecular scaffolds via covalent modifications (see Fig. 3c), solvent swelling impregnation, or blending into bulk substrates [124], [125], [126]. Other degradable scaffolds are designed to expose microenvironments to NO from the controlled release of embedded NO donor compounds (see Fig. 3d) as well as from covalently attached NO donors through various biodegradation mechanisms [125,127]. Additionally, NO-releasing scaffolds and nanoparticles have been designed to target organs and tissues to deliver payloads following stimuli-response to differences in oxidation environment or the presence of catalytic metal ions [127], [128], [129], [130], [131]. Through rational design approaches, NO-releasing formulations could be developed to target specific viral disease states. Readers are directed to detailed reviews on NO-releasing macromolecular scaffolds [124] as well as others on antimicrobial and antithrombotic applications of NO donor compounds for further elaboration on this material chemistry design [132,133].
Fig. 3.

S-nitroso-N-acetyl-penicillamine (SNAP) is a synthetic NO donor used in a variety of biomedical applications. a) SNAP undergoes homolytic cleavage of its S-N bond in the presence of heat, light, or catalytic metal ions to afford NO. b) Treatment of SARS-CoV-2 infected Vero E6 cells with low doses of SNAP can lead to over 99% inhibition of viral replication. Reproduced with permission from Akaberi et al. [119]. c) Direct conjugation of SNAP onto bulk polymer networks enhances its degradation kinetics for enhanced NO released rates over several months. Reproduced with permission from Hopkins et al. [124]. d) Incorporation of SNAP into biodegradable poly lactic-co-glycolic acid (PLGA) microspheres allows for tunable NO release rates in the presence of catalytic copper ions (PBSACu). Modified with permission from Lautner et al. [127].
5. Pharmacological considerations for NO-based antiviral therapies
Designing therapeutic agents that discharge NO or produce it in a targeted tissue environment has been met with considerable challenges. The “double-edged” nature of NO in homeostasis coupled with the poor physiochemical properties of many NO prodrugs and direct donor compounds has greatly limited the translational application of the technology. Nonetheless, the rising success of many of these NO-based therapies for antiviral application motivates further consideration of their underlying pharmacological properties in comparison to existing alternatives. This section therefore surveys the pharmacokinetics and pharmacodynamics of gNO, NO prodrugs, and NO donor compounds previously developed in antiviral therapies and critically evaluates their therapeutic potential with respect to existing antiviral agents.
5.1. Developing pharmacokinetic and pharmacodynamic models
During the development of NO-based therapies for preclinical animal studies, NO becomes a difficult species to localize and quantify directly in physiological models. In these studies, NO generation and consumption from gaseous NO treatments, prodrugs, and other donor compounds are frequently measured indirectly, through the formation of nitrites and nitrates in blood, tissue dialysate, or urine samples. Detection then depends on combinations of colorimetric methods such as Griess assay, fluorometric assays with aromatic diamino compounds, dedicated high throughput high performance liquid chromatography methods, or combinations thereof – each with its inherent limitations in detection thresholds [134]. Because many of these assays are limited by sample preparation and impurity, alternatives such as chemiluminescence or electrochemical based methods are often considered, where direct NO detection is achieved via its light-emitting reaction with ozone or reduction to diazeniumdiolate (N2O2 2−) and nitrate (NO3 −), respectively [25]. NO-based therapy studies have therefore become critically dependent on the accuracy of their detection system for determining dose-response relationships.
Because NO is a key gasotransmitter in homeostasis, the administration route and pharmacokinetics are critical parameters for optimizing therapeutic systems to minimize off-target effects. Gaseous NO therapies are often associated with mechanical ventilation via endotracheal intubation with proprietary gas formulations, such as INOmaxTM, paired with injection systems that have been validated in lung models with external chemiluminescence-based readouts [135]. Since the past thirty years of preclinical and clinical research have focused on using gNO for improving pulmonary vasodilation and oxygenation in ARDS and neonatal hyperplasia, direct dose-response correlations with gNO for preclinical antiviral therapies have yet to be shown [91,105,136].
A similar lack of knowledge regarding dose-response correlations exists with NO prodrugs and donor compounds, for which antiviral work has been mostly limited to in vitro studies. Zell et al. [137] have pioneered work in this respect with organic nitrates for the treatment of coxsackievirus B3 (CVB3) linked to cardiac arrhythmias in a mouse model (see Table 3 ). In these studies, two organic nitrates – glyceryl trinitrate and isosorbide dinitrate, were studied for their pharmacodynamics with respect to iNOS expression, fibrinolysis, and viral titer in cardiac tissue. Vascular bioactivation of glyceryl trinitrate via soluble guanylyl cyclase occurs with excesses of ascorbate and cysteine in the presence of mitochondrial aldehyde dehydrogenase (mtALDH) [138]. Isosorbide dinitrate is alternatively activated through a mtALDH-independent pathway, though both prodrugs have been shown to generate similar dose-response curves in vasorelaxation [139]. Both nitrates demonstrated reductions in viral titer and associated cardiac fibrinolysis and showcase the potential of future work with other established NO prodrugs for antiviral applications [137]. Horton et al. [140] recently summarized the pharmacokinetics and pharmacodynamics of many NO prodrugs developed and tested in preclinical animal models within the past decade, including various nitrates, diazeniumdiolates, sydnonimines, and furoxans.
Table 3.
Summary of the pharmacological considerations in studies with NO prodrugs/donor compounds in preclinical animal models.
| Prodrug and Main Metabolite | Administration Route and Dose | NO Detection Method | Efficacy | Ref |
|---|---|---|---|---|
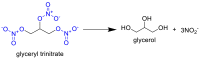 |
Intraoral (drinking water), 10 µg/mL | Griess assay on serum from treated mice | Elevated iNOS mRNA in heart tissue, decreased CVB3 viral titer, reduced heart fibrosis by day 14 | [137] |
 |
Direct intraoral administration, 1.5 mg/day | Griess assay on serum from treated mice | Elevated iNOS mRNA in heart tissue, decreased CVB3 viral titer, reduced heart fibrosis by day 28 | [137] |
| Nitric Oxide Donor Compound | Administration Route and Dose | NO Detection Method | Efficacy | Ref |
 |
Intraperitoneally, 0.336 mg/day | Griess assay on serum from treated mice | Improved spleen and thymus indices, decrease in PCV2 DNA copies in tissue autopsies | [68] |
Direct NO donor compounds have also seen limited study in preclinical animal models for antiviral application, with the only work coming from Liu et al. [68] with GSNO on porcine circovirus type 2 (PCV2) in a mouse model (see Table 3). GSNO has previously been studied through in vivo models for neuroprotection, gastrointestinal mucosal barrier integrity, inflammation, and platelet activation [141]. While Liu et al [68] demonstrated decreased PCV2 tissue copies after intraperitoneal GSNO administration, dose-response relationships for GSNO and other RSNOs in animal models of viral infection remain underdeveloped. Taken altogether, the effects of administration route, dose, and other factors such as metabolic activation or degradation are mostly unexplored for NO-based antiviral therapies and though reflective of the infancy of the field are critical to its clinical translation.
5.2. Comparison of NO-based therapies to other existing therapies
NO-based therapies are considered a multi-faceted tool for immune defense due to broad-spectrum antiviral and antimicrobial properties paired with their tunable anti-inflammatory effect. Comparable broad-spectrum antiviral agents generally focus on modulating host defense and targeting viral infectivity via one or more avenues including: viral attachment, membrane fusion and endocytosis, viral replication, viral assembly and budding, or viral envelope integrity [142]. Common broad-spectrum antivirals include alpha interferon, ribavirin, various peptide fusion inhibitors, antiviral peptides, nucleoside enantiomers, and other synthetic constructs. Direct comparisons in therapeutic efficacy of broad-spectrum antivirals to NO-based prodrugs and donors is complicated by NO's case-specific role in viral pathogenesis. For many respiratory specific strains such as syncytial virus, increased NO localization decreases replication, while for other strains such as hepatitis C it mediates viral persistence and pathogenic activity [143,144].
Few studies have considered the antiviral potency and corresponding cytocompatibility of NO prodrugs or donor compounds with respect to existing broad-spectrum alternatives. NO prodrug catalogs based on nucleoside analogues (see Table 4 ) have been studied extensively against other nucleosides and antivirals such as acyclovir which treat herpes and vesicular stomatitis viral infections [74,[145], [146], [147], [148]. Adding NO prodrug components to nucleosides generally lowered the IC50 – the concentration of agent required for 50% inhibition of the virus culture in vitro, and led to variable increases in cytocompatibility. Similarly, NO donors such as SNAP have seen extensive evaluation in vitro at treatment ranges from 100–400 µM with reports of negligible cytotoxicity against coxsackievirus, HSV 1, SARS, influenza virus, and vesicular stomatitis virus [64,70,120,122,149]. However, the current lack of IC50 data for many NO donors presents significant challenges in the clinical translation of NO-based therapies in addition to other overarching issues such as drug solubility, stability, and trafficking in vivo [150,151].
Table 4.
Comparison studies of some NO prodrugs to existing antiviral therapeutics.
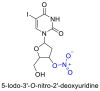
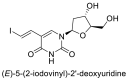
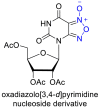

6. Future prospects
Based on NO's non-specific antiviral and antimicrobial properties, highly tunable NO-based therapeutics and materials provide promising potential in both directly combatting viruses and mitigating secondary infections associated with viral infections (see Fig. 4 ). Although gNO treatment has shown some antiviral efficacy in rescue trials and other preclinical in vivo models, its widespread use may be constrained by logistical and financial restrictions, as it has been considered one of the most expensive therapeutics in neonatal medicine. Additionally, gNO applications are mostly limited to respiratory viral infections. The use of NO-donor compounds, alone or in scaffolds, can address these shortcomings with prolonged and site-specific NO release for broader applications. Furthermore, incorporation of prolonged NO release outside of direct therapies could inhibit viral transmission and address complications in treatments. In this section, we discuss the potential of these compounds as direct antiviral agents and in preventative and indirect antiviral strategies.
Fig. 4.

Summary of current and prospective NO-based strategies for active elimination of viruses and further prevention of viral infection ranging from gaseous NO (gNO) to small molecule NO donor compounds and material strategies. Active NO release from or onto surfaces may reduce viral contamination, as illustrated by active (orange) virions becoming inert (gray) after exposure to NO.
6.1. Direct antiviral therapies
Sustained NO release from NO donor compounds or scaffolds can improve the logistics of gNO therapies for respiratory infections. In addition to NO releasing strategies, NO generating scaffolds that use catalytic metal ions or metal organic frameworks to generate NO from endogenous or supplemented RSNOs have shown promising kinetics in physiological conditions [152]. In clinical evaluations, numerous scaffolds containing RSNO moieties for enhanced NO delivery have already shown their effectiveness against microbial respiratory infections. For example, NO-releasing biodegradable alginates developed by Vast Therapeutics Inc. have proven effective in combatting lung bacterial infections in a cystic fibrosis-like environment, showing NO's potential in mucosal environments [153]. However, the antiviral effects of designs such as these remain unexplored to date. Given the substantial influence NO can have in ARDS and pneumonias, such constructs alone or in combination with other antiviral agents could provide enhanced pulmonary oxygenation and reduce infiltrates.
In addition to respiratory viral infections, the incorporation of NO drugs into established or new nasal/inhaled formulations currently under investigation has great potential to add or enhance targeted antiviral activity in decongestants and other commonly used therapeutics. Rapid mucociliary clearance is a common issue with nasal drug delivery systems, but significant research has shown the promise of mucoadherant polymers and nanoparticles as improved templates [154], [155], [156]. The inclusion of NO donors and/or catalytically generating compounds into these systems or other biodegradable systems is an emerging field and not well explored to date, but given that NO drugs have successfully been attached to or contained in many scaffolds and nanoparticles for other applications, the field appears promising [132].
6.2. Passive antiviral and secondary bacterial infection prevention
6.2.1. Mechanical ventilation
In addition to direct antiviral therapies, the incorporation of NO into medical devices for the prevention of secondary infections related to viral infection is an equally important endeavor. Numerous NO-releasing and generating materials have been explored for antimicrobial and hemocompatible applications (e.g. vascular catheters and ECMO) [157]. Many of these materials exhibit encouraging properties, good biocompatibility, and are undergoing commercialization [158], [159], [160]. While developing a better understanding of the antiviral properties of these materials should be explored, translating their use for improving viral infection treatment options is just as critical. As previously mentioned, treating complications from respiratory viral infections such as SARS-CoV and influenza notably requires mechanical ventilation and ECMO [161,162]. However, these therapies are often associated with further issues. Endotracheal intubation encumbers normal defense mechanisms of mucociliary clearance and introduces a foreign surface vulnerable to bacterial infiltration and biofilm formation on the endotracheal tube surface, ultimately resulting in high rates of ventilator associated pneumonia (VAP) [163]. NO-releasing endotracheal tubes have recently been shown to reduce bacterial infections in vitro and show promise for VAP reduction as a complication of infections [164]. Additionally, they may serve as a strategy to help manage concurrent viral infections as well as microbial infections. Other strategies explored to prevent VAP involve subglottic secretion suction, prophylactic antibiotic regimens, and surface modifications, including silver, gentamicin, and hydrogel-based coatings [158,159]. The combination of these surface designs with NO may result in a synergistic increase in antimicrobial efficacy with enhanced antiviral effects.
6.2.2. ECMO
In severe respiratory infections where mechanical ventilation is insufficient, ECMO is often used as a last resort. In addition to the potential for microbial and viral surface contamination, ECMO has broader fouling concerns with blood clot formation in the ex vivo circuit. Therefore, ECMO has simultaneous risks for both embolism and bacterial/viral sepsis [165]. The fouling process is mediated by the adsorption of blood plasma and bacterial proteins to the surfaces of the blood-contacting circuit, which sets off simultaneous processes of platelet/bacteria surface adhesion and eventual clot/biofilm formation. Any analogous process of viral protein adsorption onto the blood-contacting polymer substrates within ECMO circuits remains unexplored to date. However, the greater issue at hand with ECMO is its use to treat SARS in COVID-19 patients – when mechanical ventilation is often inadequate, in light of additional concerns about viral sepsis and advanced states of hypercoagulation [166], [167], [168], [169]. Traditionally, ECMO patients are systemically heparinized to prevent inline clotting. However, this strategy is problematic for patients with histories of cardiovascular disease, diabetes mellitus, and hypertension that are treated with angiotensin-converting enzyme (ACE) inhibitors because: 1) heparin is contraindicated due to the increased risks for hemorrhaging, thrombocytopenia, and hyperkalemia; 2) ACE2 is a functional receptor for SARS-CoV [170,171]; and 3) high molecular weight heparins have been shown to act as non-specific binding sites for SARS-CoV [172]. Even with emerging investigational coatings with immobilized heparin there exists the concurrent risks of enhanced viral fouling, further propagation of the viral disease state, and cardiovascular complications for this class of SARS patients. Other commercially available or otherwise investigational antifouling coatings include phosphorylcholine, amphiphilic polymers, immobilized albumin, zwitterions, and other “slippery” non-adhesive coatings [173]. However, none of these strategies contain active broad-spectrum antimicrobial potential, disperse mature biofilms, inhibit the coagulation cascade, or otherwise act as a direct viricidal agent [165].
With their notable antimicrobial/antiviral properties and anti-thrombotic effects, NO strategies alone or in combination with these other approaches have the best potential to augment antifouling and antiviral properties when incorporated into ECMO circuits (see Fig. 5 ). In particular, combinations of NO release and surface grafting of zwitterionic polymer brushes have shown excellent antimicrobial (>3-log reduction of viable bacteria in vitro) and antithrombotic efficacy (significant reduction in clot formation in a 7-day in vivo rabbit model of extracorporeal circulation) [174,175]. Similarly, catalytic NO-generating materials have also been tested in an in vivo extracorporeal circulation rabbit model and have shown reduction in platelet adhesion, but their applications may be limited by bioavailability of endogenous donors, catalyst stability, and toxicity of leachates [165,176]. Materials using combinations of antifouling strategies that incorporate NO release and/or generation appear to be the most promising way forward to prevent complications and simultaneously have antiviral effects in ECMO treatments (see Fig. 5). However, until such materials are commercially available NO sweep gas streams may prove useful as an alternative sterilization strategy for eradicating microbes and viruses [66,98].
Fig. 5.

Synergistic strategies combining nitric oxide release with a passive surface modification can enhance the antifouling surface properties and biocompatibility of medical device surfaces for a variety of antimicrobial and hemocompatible applications. These strategies combine tunable biocidal release rates of NO from a surface embedded donor compound with antifouling surface conjugates to reduce rates of surface protein adsorption. These approaches have not been investigated against viral surface contamination and could become prospects for antiviral surface development in mechanical ventilation and ECMO.
6.3. Clinical environments and beyond
NO-releasing surfaces and materials also have implicit application potential in passively preventing hospital-associated viral transmission. The shelf-stability and extended NO surface flux from these materials enables versatile application both in and outside of clinical environments to control infection rates, especially in outbreaks as rampant as COVID-19 [177]. Self-sterilizing surfaces with NO release would have a wide range of effects against most bacteria, fungi, and select viruses. Applying NO-releasing technology to facemasks could enable self-sterilization, increasing safety and decreasing disposal concerns. NO-releasing coatings could also be beneficial on many surfaces otherwise susceptible to high contamination rates, such as frequently touched objects in medical settings (e.g. doorknobs, elevator buttons, etc.), as NO can concurrently inhibit viral replication and kill microbial pathogens to thereby reduce the spread of infections.
7. Conclusions
Thirty years of research have led to NO emerging as an important therapeutic option for treating virus-based illnesses and has led to successful or otherwise ongoing commercialization of several gNO and NO-releasing therapies for respiratory viral infections, skin infections, and other medical conditions concurrent to viral infection. Significant progress has been made in understanding the limits of the antiviral capacity of NO both in host cells and as a passive mechanism for reducing free viral loads. This work is of critical importance to developing gNO and other NO donor therapies to aid in systemic clearance of viral infections while also providing insight into the basic biology of NO in the immune response. While gNO has begun to demonstrate its excellent potential as a broad antiviral agent, NO-releasing materials can have the additional benefits of prolonged release and vast delivery/application options both directly and indirectly addressing associated complications of viral infections. Future research should therefore focus on further developing gNO and NO-releasing delivery platforms for medical procedures related to hospitalization for viral infection, such as mechanical ventilation, therapeutic pulmonary delivery, ECMO, endotracheal intubation, and other blood-contacting interventions. Furthermore, NO releasing composite materials functionalized with additional antifouling strategies remain to be investigated in a clinical context for viral contamination and could develop into a new generation of materials for actively inhibiting viral replication and preventing further contamination of medical devices and surfaces. Ongoing and future development of NO chemistries and materials will expedite the clinical translation process and bring new avenues of therapeutic antiviral strategies to patient care.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
Acknowledgements
We thank Ms. Yujie Chen (University of Georgia) for assistance with graphical design. The authors declare no competing financial interests.
Funding
This work was supported by the National Institutes of Health [grant number R01HL134899]; and the National Science Foundation [grant number NSF 1842396].
References
- 1.Adalja A.A., Toner E., Inglesby T.V. Priorities for the US health community responding to COVID-19. JAMA. 2020;323:1343. doi: 10.1001/jama.2020.3413. 10.1001/jama.2020.3413 [DOI] [PubMed] [Google Scholar]
- 2.Rossignol J.-F. Nitazoxanide: a first-in-class broad-spectrum antiviral agent. Antivir Res. 2014;110:94. doi: 10.1016/j.antiviral.2014.07.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Huang C., Wang Y., Li X., Ren L., Zhao J., Hu Y., Zhang L., Fan G., Xu J., Gu X., Cheng Z., Yu T., Xia J., Wei Y., Wu W., Xie X., Yin W., Li H., Liu M., Xiao Y., Gao H., Guo L., Xie J., Wang G., Jiang R., Gao Z., Jin Q., Wang J., Cao B. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet North Am Ed. 2020;395:497. doi: 10.1016/S0140-6736(20)30183-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Liu K., Chen Y., Lin R., Han K. Clinical features of COVID-19 in elderly patients:a comparison with young and middle-aged patients. J Infect. 2020;80:e14. doi: 10.1016/j.jinf.2020.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ruan Q., Yang K., Wang W., Jiang L., Song J. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intens Care Med. 2020;46:846. doi: 10.1007/s00134-020-05991-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zhou F., Yu T., Du R., Fan G., Liu Y., Liu Z., Xiang J., Wang Y., Song B., Gu X., Guan L., Wei Y., Li H., Wu X., Xu J., Tu S., Zhang Y., Chen H., Cao B. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. Lancet North Am Ed. 2020;395:1054. doi: 10.1016/S0140-6736(20)30566-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Darouiche R.O., infections Device-associated. A macroproblem that starts with microadherence. Clin Infect Dis. 2001;33:1567. doi: 10.1086/323130. [DOI] [PubMed] [Google Scholar]
- 8.Morris D.E., Cleary D.W., Clarke S.C. Secondary bacterial infections associated with influenza pandemics. Front Microbiol. 2017;8:1041. doi: 10.3389/fmicb.2017.01041. https://dx.doi.org/10.3389%2Ffmicb.2017.01041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rong F., Tang Y., Wang T., Feng T., Song J., Li P., Huang W. Nitric oxide-releasing polymeric materials for antimicrobial applications: a review. Antioxidants. 2019;8:556. doi: 10.3390/antiox8110556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Culotta E., Koshland D.E., Jr. NO news is good news. Science. 1992;258:1862. doi: 10.1126/science.1361684. [DOI] [PubMed] [Google Scholar]
- 11.Snyder S.H., Bredt D.S. Biological roles of nitric oxide. Sci Am. 1992;266:68. doi: 10.1038/scientificamerican0592-68. [DOI] [PubMed] [Google Scholar]
- 12.Douglass M.E., Goudie M.J., Pant J., Singha P., Hopkins S., Devine R., Schmiedt C.W., Handa H. Catalyzed nitric oxide release via cu nanoparticles leads to an increase in antimicrobial effects and hemocompatibility for short-term extracorporeal circulation. ACS Appl Bio Mater. 2019;2:2539. doi: 10.1021/acsabm.9b00237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Homeyer K.H., Goudie M.J., Singha P., Handa H. Liquid-Infused nitric-oxide-releasing silicone foley urinary catheters for prevention of catheter-associated urinary tract infections. ACS Biomater Sci Eng. 2019;5:2021. doi: 10.1021/acsbiomaterials.8b01320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pant J., Goudie M.J., Chaji S.M., Johnson B.W., Handa H. Nitric oxide releasing vascular catheters for eradicating bacterial infection. J Biomed Mater Res B Appl Biomater. 2018;106:2849. doi: 10.1002/jbm.b.34065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chen L., Liu P., Gao H., Sun B., Chao D., Wang F., Zhu Y., Hedenstierna G., Wang C.G. Inhalation of nitric oxide in the treatment of severe acute respiratory syndrome: a rescue trial in Beijing. Clin Infect Dis. 2004;39:1531. doi: 10.1086/425357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Du X. A new mechanism for nitric oxide - and cGMP-mediated platelet inhibition. Blood. 2007;109:392. doi: 10.1182/blood-2006-10-053827. [DOI] [Google Scholar]
- 17.Chen K., Pittman R.N., Popel A.S. Nitric oxide in the vasculature: where does it come from and where does it go? A quantitative perspective. Antioxid Redox Signal. 2008;10:1185. doi: 10.1089/ars.2007.1959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Moncada S. Nitric oxide in the vasculature: physiology and pathophysiology. Ann N Y Acad Sci. 1997;811:60. doi: 10.1111/j.1749-6632.1997.tb51989.x. [DOI] [PubMed] [Google Scholar]
- 19.Lanas A. Role of nitric oxide in the gastrointestinal tract. Arthritis Res Ther. 2008;10(Suppl 2):S4. doi: 10.1186/ar2465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dzoljic E., Grbatinic I., Kostic V. Why is nitric oxide important for our brain? Funct Neurol. 2015;30:159. doi: 10.11138/fneur/2015.30.3.159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wink D.A., Hines H.B., Cheng R.Y., Switzer C.H., Flores-Santana W., Vitek M.P., Ridnour L.A., Colton C.A. Nitric oxide and redox mechanisms in the immune response. J Leukoc Biol. 2011;89:873. doi: 10.1189/jlb.1010550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mayer B., Hemmens B. Biosynthesis and action of nitric oxide in mammalian cells. Trends Biochem Sci. 1997;22:477. doi: 10.1016/s0968-0004(97)01147-x. [DOI] [PubMed] [Google Scholar]
- 23.Villanueva C., Giulivi C. Subcellular and cellular locations of nitric oxide synthase isoforms as determinants of health and disease. Free Radic Biol Med. 2010;49:307. doi: 10.1016/j.freeradbiomed.2010.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Snyder S.H. Janus faces of nitric oxide. Nature. 1993;364:577. doi: 10.1038/364577a0. [DOI] [PubMed] [Google Scholar]
- 25.Coneski P.N., Schoenfisch M.H. Nitric oxide release: part III. Measurement and reporting. Chem Soc Rev. 2012;41:3753. doi: 10.1039/c2cs15271a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jimenez W., Ros J., Morales-Ruiz M., Navasa M., Sole M., Colmenero J., Sort P., Rivera F., Arroyo V., Rodes J. Nitric oxide production and inducible nitric oxide synthase expression in peritoneal macrophages of cirrhotic patients. Hepatology. 1999;30:670. doi: 10.1002/hep.510300310. [DOI] [PubMed] [Google Scholar]
- 27.Ghimire K., Altmann H.M., Straub A.C., Isenberg J.S. Nitric oxide: what's new to NO? Am J Physiol Cell Physiol. 2017;312:C254. doi: 10.1152/ajpcell.00315.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sansbury B.E., Hill B.G. Regulation of obesity and insulin resistance by nitric oxide. Free Radic Biol Med. 2014;73:383. doi: 10.1016/j.freeradbiomed.2014.05.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Laroux F.S., Pavlick K.P., Hines I.N., Kawachi S., Harada H., Bharwani S., Hoffman J.M., Grisham M.B. Role of nitric oxide in inflammation. Acta Physiol Scand. 2001;173:113. doi: 10.1046/j.1365-201x.2001.00891.x. [DOI] [PubMed] [Google Scholar]
- 30.Pisoschi A.M., Pop A. The role of antioxidants in the chemistry of oxidative stress: a review. Eur J Med Chem. 2015;97:55. doi: 10.1016/j.ejmech.2015.04.040. [DOI] [PubMed] [Google Scholar]
- 31.Hughes M.N. Relationships between nitric oxide, nitroxyl ion, nitrosonium cation and peroxynitrite. Biochim et Biophys Acta (BBA) - Bioenerg. 1999;1411:263. doi: 10.1016/s0005-2728(99)00019-5. [DOI] [PubMed] [Google Scholar]
- 32.Smith B.C., Marletta M.A. Mechanisms of S-nitrosothiol formation and selectivity in nitric oxide signaling. Curr Opin Chem Biol. 2012;16:498. doi: 10.1016/j.cbpa.2012.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Thomas D.D., Liu X., Kantrow S.P., Lancaster J.R. The biological lifetime of nitric oxide: implications for the perivascular dynamics of NO and O2. Proc Natl Acad Sci. 2001;98:355. doi: 10.1073/pnas.98.1.355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ford P.C., Lorkovic I.M. Mechanistic aspects of the reactions of nitric oxide with transition-metal complexes. Chem Rev. 2002;102:993. doi: 10.1021/cr0000271. [DOI] [PubMed] [Google Scholar]
- 35.Hickok J.R., Vasudevan D., Jablonski K., Thomas D.D. Oxygen dependence of nitric oxide-mediated signaling. Redox Biol. 2013;1:203. doi: 10.1016/j.redox.2012.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Thomas D.D. Breathing new life into nitric oxide signaling: a brief overview of the interplay between oxygen and nitric oxide. Redox Biol. 2015;5:225. doi: 10.1016/j.redox.2015.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kelm M. Nitric oxide metabolism and breakdown. Biochim et Biophys Acta (BBA) - Bioenerg. 1999;1411:273. doi: 10.1016/s0005-2728(99)00020-1. [DOI] [PubMed] [Google Scholar]
- 38.Abdul-Cader M.S., Amarasinghe A., Abdul-Careem M.F. Activation of toll-like receptor signaling pathways leading to nitric oxide-mediated antiviral responses. Arch Virol. 2016;161:2075. doi: 10.1007/s00705-016-2904-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Xu W., Zheng S., Dweik R.A., Erzurum S.C. Role of epithelial nitric oxide in airway viral infection. Free Radic Biol Med. 2006;41:19. doi: 10.1016/j.freeradbiomed.2006.01.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Akaike T., Maeda H. Nitric oxide and virus infection. Immunology. 2000;101:300. doi: 10.1046/j.1365-2567.2000.00142.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Colasanti M., Persichini T., Venturini G., Ascenzi P. S-nitrosylation of viral proteins: molecular bases for antiviral effect of nitric oxide. IUBMB Life. 1999;48:25. doi: 10.1080/713803459. [DOI] [PubMed] [Google Scholar]
- 42.Saura M., Zaragoza C., McMillan A., Quick R.A., Hohenadl C., Lowenstein J.M., Lowenstein C.J. An antiviral mechanism of nitric oxide: inhibition of a viral protease. Immunity. 1999;10:21. doi: 10.1016/s1074-7613(00)80003-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mehta D.R., Ashkar A.A., Mossman K.L. The nitric oxide pathway provides innate antiviral protection in conjunction with the type I interferon pathway in fibroblasts. PLoS One. 2012;7:e31688. doi: 10.1371/journal.pone.0031688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wei L., Gravitt P.E., Song H., Maldonado A.M., Ozbun M.A. Nitric oxide induces early viral transcription coincident with increased DNA damage and mutation rates in human papillomavirus-infected cells. Cancer Res. 2009;69:4878. doi: 10.1158/0008-5472.CAN-08-4695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Klingstrom J., Akerstrom S., Hardestam J., Stoltz M., Simon M., Falk K.I., Mirazimi A., Rottenberg M., Lundkvist A. Nitric oxide and peroxynitrite have different antiviral effects against hantavirus replication and free mature virions. Eur J Immunol. 2006;36:2649. doi: 10.1002/eji.200535587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.E. Ryan, R. Hollingworth, R. Grand, Activation of the DNA damage response by RNA viruses, 6 (2016) 2, https://dx.doi.org/ 10.3390/biom6010002. [DOI] [PMC free article] [PubMed]
- 47.Uehara E.U., Shida Bde S., de Brito C.A. Role of nitric oxide in immune responses against viruses: beyond microbicidal activity. Inflamm Res. 2015;64:845. doi: 10.1007/s00011-015-0857-2. [DOI] [PubMed] [Google Scholar]
- 48.Koetzler R., Zaheer R.S., Newton R., Proud D. Nitric oxide inhibits IFN regulatory factor 1 and nuclear factor-kappaB pathways in rhinovirus-infected epithelial cells. J Allergy Clin Immunol. 2009;124:551. doi: 10.1016/j.jaci.2009.04.041. [DOI] [PubMed] [Google Scholar]
- 49.Koetzler R., Zaheer R.S., Wiehler S., Holden N.S., Giembycz M.A., Proud D. Nitric oxide inhibits human rhinovirus-induced transcriptional activation of CXCL10 in airway epithelial cells. J Allergy Clin Immunol. 2009;123:201. doi: 10.1016/j.jaci.2008.09.041. [DOI] [PubMed] [Google Scholar]
- 50.Rios-Ibarra C.P., Lozano-Sepulveda S., Munoz-Espinosa L., Rincon-Sanchez A.R., Cordova-Fletes C., Rivas-Estilla A.M. Downregulation of inducible nitric oxide synthase (iNOS) expression is implicated in the antiviral activity of acetylsalicylic acid in HCV-expressing cells. Arch Virol. 2014;159:3321. doi: 10.1007/s00705-014-2201-5. [DOI] [PubMed] [Google Scholar]
- 51.Xie F. Broad-spectrum antiviral effect of chebulagic acid and punicalagin on respiratory syncytial virus infection in a BALB/c model. Int. J. Clin. Exp. Pathol. 2016;9:611. doi: 10.1186/1471-2180-13-187. [DOI] [Google Scholar]
- 52.Pingle S.C., Jajoo S., Mukherjea D., Sniderhan L.F., Jhaveri K.A., Marcuzzi A., Rybak L.P., Maggirwar S.B., Ramkumar V. Activation of the adenosine A1 receptor inhibits HIV-1 tat-induced apoptosis by reducing nuclear factor-kappaB activation and inducible nitric-oxide synthase. Mol Pharmacol. 2007;72:856. doi: 10.1124/mol.106.031427. [DOI] [PubMed] [Google Scholar]
- 53.Loveless M.O., Phillips C.R., Giraud G.D., Holden W.E. Decreased exhaled nitric oxide in subjects with HIV infection. Thorax. 1997;52:185. doi: 10.1136/thx.52.2.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fang X., Gao J., Zheng H., Li B., Kong L., Zhang Y., Wang W., Zeng Y., Ye L. The membrane protein of SARS-CoV suppresses NF-kappaB activation. J Med Virol. 2007;79:1431. doi: 10.1002/jmv.20953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.L. A. Perrone, J. A. Belser, D. A. Wadford, J. M. Katz, T. M. Tumpey, Inducible nitric oxide contributes to viral pathogenesis following highly pathogenic influenza virus infection in mice, 207 (2013) 1576, https://dx.doi.org/ 10.1093/infdis/jit062. [DOI] [PubMed]
- 56.Akaike T., Noguchi Y., Ijiri S., Setoguchi K., Suga M., Zheng Y.M., Dietzschold B., Maeda H. Pathogenesis of influenza virus-induced pneumonia: involvement of both nitric oxide and oxygen radicals. Proc Natl Acad Sci U S A. 1996;93:2448. doi: 10.1073/pnas.93.6.2448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Torre D., Pugliese A., Speranza F. Role of nitric oxide in HIV-1 infection: friend or foe? Lancet Infect Dis. 2002;2:273. doi: 10.1016/s1473-3099(02)00262-1. [DOI] [PubMed] [Google Scholar]
- 58.De Souza K.P.R., Silva E., De Oliveira Rocha E., Figueiredo L., De Almeida-Leite C., Arantes R.M.E., De Assis Silva Gomes J., Ferreira G., De Oliveira J., Kroon E., Campos M. Nitric oxide synthase expression correlates with death in an experimental mouse model of dengue with CNS involvement. Virol J. 2013;10:267. doi: 10.1186/1743-422X-10-267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Reiss C.S., Komatsu T. Does nitric oxide play a critical role in viral infections? J Virol. 1998;72:4547. doi: 10.1128/jvi.72.6.4547-4551.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mannick J.B. The antiviral role of nitric oxide. Res Immunol. 1995;146:693. doi: 10.1016/0923-2494(96)84920-0. [DOI] [PubMed] [Google Scholar]
- 61.Sanders S.P., Siekierski E.S., Porter J.D., Richards S.M., Proud D. Nitric oxide inhibits rhinovirus-induced cytokine production and viral replication in a human respiratory epithelial cell line. J Virol. 1998;72:934. doi: 10.1128/jvi.72.2.934-942.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Sanders S.P., Proud D., Permutt S., Siekierski E.S., Yachechko R., Liu M.C. Role of nasal nitric oxide in the resolution of experimental rhinovirus infection. J Allergy Clin Immunol. 2004;113:697. doi: 10.1016/j.jaci.2004.01.755. [DOI] [PubMed] [Google Scholar]
- 63.Jung K., Gurnani A., Renukaradhya G.J., Saif L.J. Nitric oxide is elicited and inhibits viral replication in pigs infected with porcine respiratory coronavirus but not porcine reproductive and respiratory syndrome virus. Vet Immunol Immunopathol. 2010;136:335. doi: 10.1016/j.vetimm.2010.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Croen K.D. Evidence for antiviral effect of nitric oxide. inhibition of herpes simplex virus type 1 replication. J Clin Invest. 1993;91:2446. doi: 10.1172/jci116479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Regev-Shoshani G., Vimalanathan S., McMullin B., Road J., Av-Gay Y., Miller C. Gaseous nitric oxide reduces influenza infectivity in vitro. Nitric Oxide. 2013;31:48. doi: 10.1016/j.niox.2013.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.McMullin B.B., Chittock D.R., Roscoe D.L., Garcha H., Wang L., Miller C.C. The antimicrobial effect of nitric oxide on the bacteria that cause nosocomial pneumonia in mechanically ventilated patients in the intensive care unit. Respir Care. 2005;50:1451. http://rc.rcjournal.com/content/50/11/1451.abstract. [PubMed] [Google Scholar]
- 67.Sanders S.P. Asthma, viruses, and nitric oxide. Proc Soc Exp Biol Med. 1999;220:123. doi: 10.1046/j.1525-1373.1999.d01-19.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Liu C., Wen L., Xiao Q., He K. Nitric oxide-generating compound GSNO suppresses porcine circovirus type 2 infection in vitro and in vivo. BMC Vet Res. 2017;13:59. doi: 10.1186/s12917-017-0976-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Keyaerts E., Vijgen L., Chen L., Maes P., Hedenstierna G., Van Ranst M. Inhibition of SARS-coronavirus infection in vitro by S-nitroso-N-acetylpenicillamine, a nitric oxide donor compound. Int J Infect Dis. 2004;8:223. doi: 10.1016/j.ijid.2004.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Rimmelzwaan G.F., Baars M.M., de Lijster P., Fouchier R.A., Osterhaus A.D. Inhibition of influenza virus replication by nitric oxide. J Virol. 1999;73:8880. doi: 10.1128/jvi.73.10.8880-8883.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Simon M., Falk K.I., Lundkvist A., Mirazimi A. Exogenous nitric oxide inhibits Crimean Congo hemorrhagic fever virus. Virus Res. 2006;120:184. doi: 10.1016/j.virusres.2006.03.005. [DOI] [PubMed] [Google Scholar]
- 72.Banerjee N.S., Moore D.W., Wang H.K., Broker T.R., Chow L.T. NVN1000, a novel nitric oxide-releasing compound, inhibits HPV-18 virus production by interfering with E6 and E7 oncoprotein functions. Antivir Res. 2019;170 doi: 10.1016/j.antiviral.2019.104559. [DOI] [PubMed] [Google Scholar]
- 73.Tyring S.K., Rosen T., Berman B., Stasko N., Durham T., Maeda-Chubachi T. A phase 2 controlled study of SB206, a topical nitric oxide-releasing drug for extragenital wart treatment. J Drugs Dermatol. 2018;17:1100. https://www.ncbi.nlm.nih.gov/pubmed/30365591 [PubMed] [Google Scholar]
- 74.Naimi E., Zhou A., Khalili P., Wiebe L.I., Balzarini J., De Clercq E., Knaus E.E. Synthesis of 3′- and 5′-nitrooxy pyrimidine nucleoside nitrate esters: "nitric oxide donor" agents for evaluation as anticancer and antiviral agents. J Med Chem. 2003;46:995. doi: 10.1021/jm020299r. [DOI] [PubMed] [Google Scholar]
- 75.Spivak J.L., Hasselbalch H. Hydroxycarbamide: a user's guide for chronic myeloproliferative disorders. Expert Rev Anticancer Ther. 2011;11:403. doi: 10.1586/era.11.10. [DOI] [PubMed] [Google Scholar]
- 76.Nozaki A., Numata K., Morimoto M., Kondo M., Sugimori K., Morita S., Miyajima E., Ikeda M., Kato N., Maeda S., Tanaka K. Hydroxyurea suppresses HCV replication in humans: a Phase I trial of oral hydroxyurea in chronic hepatitis C patients. Antivir Ther. 2010;15:1179. doi: 10.3851/imp1668. [DOI] [PubMed] [Google Scholar]
- 77.Kashani H.H., Vossoughi A., Adibi P., Alavian S.M. Amazing results with hydroxyurea therapy in chronic hepatitis B: a preliminary report. Hepat Mon. 2010;10:215. https://www.ncbi.nlm.nih.gov/pubmed/22308141 [PMC free article] [PubMed] [Google Scholar]
- 78.Bonvicini F., Bua G., Conti I., Manaresi E., Gallinella G. Hydroxyurea inhibits parvovirus B19 replication in erythroid progenitor cells. Biochem Pharmacol. 2017;136:32. doi: 10.1016/j.bcp.2017.03.022. [DOI] [PubMed] [Google Scholar]
- 79.Lori F., Malykh A., Cara A., Sun D., Weinstein J.N., Lisziewicz J., Gallo R.C. Hydroxyurea as an inhibitor of human immunodeficiency virus-type 1 replication. Science. 1994;266:801. doi: 10.1126/science.7973634. [DOI] [PubMed] [Google Scholar]
- 80.Wu X., Tang X., Xian M., Wang P.G. Glycosylated diazeniumdiolates: a novel class of enzyme-activated nitric oxide donors. Tetrahedron Lett. 2001;42:3779. doi: 10.1016/S0040-4039(01)00614-1. [DOI] [Google Scholar]
- 81.Nandurdikar R.S., Keefer L.K., Saavedra J.E. Novel protection–deprotection strategies in diazeniumdiolate chemistry: synthesis of V-IPA/NO. Chem Commun. 2011;47:6710. doi: 10.1039/C1CC12130H. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Thatcher G.R. An introduction to NO-related therapeutic agents. Curr Top Med Chem. 2005;5:597. doi: 10.2174/1568026054679281. [DOI] [PubMed] [Google Scholar]
- 83.Thatcher G.R., Bennett B.M., Reynolds J.N. NO chimeras as therapeutic agents in Alzheimer's disease. Curr Alzheimer Res. 2006;3:237. doi: 10.2174/156720506777632925. [DOI] [PubMed] [Google Scholar]
- 84.Singh S., Gupta A.K. Nitric oxide: role in tumour biology and iNOS/NO-based anticancer therapies. Cancer Chemother Pharmacol. 2011;67:1211. doi: 10.1007/s00280-011-1654-4. [DOI] [PubMed] [Google Scholar]
- 85.Sorba G., Medana C., Fruttero R., Cena C., Di Stilo A., Galli U., Gasco A. Water Soluble Furoxan Derivatives as NO Prodrugs. J Med Chem. 1997;40:463. doi: 10.1021/jm960379t. [DOI] [PubMed] [Google Scholar]
- 86.Lundberg J.O., Weitzberg E., Gladwin M.T. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat Rev Drug Discov. 2008;7:156. doi: 10.1038/nrd2466. [DOI] [PubMed] [Google Scholar]
- 87.Santangelo R., Mancuso C., Marchetti S., Di Stasio E., Pani G., Fadda G. Bilirubin: an Endogenous Molecule with Antiviral Activity in vitro. Front Pharmacol. 2012;3:36. doi: 10.3389/fphar.2012.00036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Liu X.H., Ruan B.F., Li J., Chen F.H., Song B.A., Zhu H.L., Bhadury P.S., Zhao J. Synthesis and biological activity of chiral dihydropyrazole: potential lead for drug design. Mini Rev Med Chem. 2011;11:771. doi: 10.2174/138955711796355285. [DOI] [PubMed] [Google Scholar]
- 89.Wohl B.M., Smith A.A., Kryger M.B., Zelikin A.N. Narrow therapeutic window of ribavirin as an inhibitor of nitric oxide synthesis is broadened by macromolecular prodrugs. Biomacromolecules. 2013;14:3916. doi: 10.1021/bm401048s. [DOI] [PubMed] [Google Scholar]
- 90.Fricker S.P. Nitric oxide scavengers as a therapeutic approach to nitric oxide mediated disease. Expert Opin Investig Drugs. 1999;8:1209. doi: 10.1517/13543784.8.8.1209. [DOI] [PubMed] [Google Scholar]
- 91.Rossaint R., Gerlach H., Schmidt-Ruhnke H., Pappert D., Lewandowski K., Steudel W., Falke K. Efficacy of inhaled nitric oxide in patients with severe ARDS. Chest. 1995;107:1107. doi: 10.1378/chest.107.4.1107. [DOI] [PubMed] [Google Scholar]
- 92.Day R.W., Lynch J.M., White K.S., Ward R.M. Acute response to inhaled nitric oxide in newborns with respiratory failure and pulmonary hypertension. Pediatrics. 1996;98:698. https://pubmed.ncbi.nlm.nih.gov/8885949/ [PubMed] [Google Scholar]
- 93.Frostell C.G., Blomqvist H., Hedenstierna G., Lundberg J., Zapol W.M. Inhaled nitric oxide selectively reverses human hypoxic pulmonary vasoconstriction without causing systemic vasodilation. Anesthesiology. 1993;78:427. doi: 10.1097/00000542-199303000-00005. [DOI] [PubMed] [Google Scholar]
- 94.Hogman M., Frostell C., Arnberg H., Hedenstierna G. Inhalation of nitric oxide modulates methacholine-induced bronchoconstriction in the rabbit. Eur Respir J. 1993;6:177. https://pubmed.ncbi.nlm.nih.gov/8444288/ [PubMed] [Google Scholar]
- 95.Deppisch C., Herrmann G., Graepler-Mainka U., Wirtz H., Heyder S., Engel C., Marschal M., Miller C.C., Riethmuller J. Gaseous nitric oxide to treat antibiotic resistant bacterial and fungal lung infections in patients with cystic fibrosis: a phase I clinical study. Infection. 2016;44:513. doi: 10.1007/s15010-016-0879-x. [DOI] [PubMed] [Google Scholar]
- 96.Rouillard K.R., Hill D.B., Schoenfisch M.H. Antibiofilm and mucolytic action of nitric oxide delivered via gas or macromolecular donor using in vitro and ex vivo models. J Cyst Fibros. 2020 doi: 10.1016/j.jcf.2020.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.C. C. Miller, D. R. Hole, R. E. Lee, Use of high dose concentrations of gaseous nitric oxide, US 20080193566A1, (2008), https://patents.google.com/patent/US20080193566A1.
- 98.Miller C., McMullin B., Ghaffari A., Stenzler A., Pick N., Roscoe D., Ghahary A., Road J., Av-Gay Y. Gaseous nitric oxide bactericidal activity retained during intermittent high-dose short duration exposure. Nitric Oxide. 2009;20:16. doi: 10.1016/j.niox.2008.08.002. [DOI] [PubMed] [Google Scholar]
- 99.Ichinose F., Roberts J.D., Jr., Zapol W.M. Inhaled nitric oxide: a selective pulmonary vasodilator: current uses and therapeutic potential. Circulation. 2004;109:3106. doi: 10.1161/01.cir.0000134595.80170.62. [DOI] [PubMed] [Google Scholar]
- 100.FDACDER, approval package for: application number: NDA 20845/S17. https://www.accessdata.fda.gov/drugsatfda_docs/nda/2015/020845Orig1s017.pdf, 2015 (Accessed 1 July 2020).
- 101.Long R., Jones R., Talbot J., Mayers I., Barrie J., Hoskinson M., Light B. Inhaled nitric oxide treatment of patients with pulmonary tuberculosis evidenced by positive sputum smears. Antimicrob Agents Chemother. 2005;49:1209. doi: 10.1128/aac.49.3.1209-1212.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Chen H.I., Kao S.J., Wang D., Lee R.P., Su C.F. Acute respiratory distress syndrome. J Biomed Sci. 2003;10:588. doi: 10.1007/bf02256308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Lee R.P., Wang D., Kao S.J., Chen H.I. The lung is the major site that produces nitric oxide to induce acute pulmonary oedema in endotoxin shock. Clin Exp Pharmacol Physiol. 2001;28:315. doi: 10.1046/j.1440-1681.2001.03446.x. [DOI] [PubMed] [Google Scholar]
- 104.Kao S.J., Peng T.C., Lee R.P., Hsu K., Chen C.F., Hung Y.K., Wang D., Chen H.I. Nitric oxide mediates lung injury induced by ischemia-reperfusion in rats. J Biomed Sci. 2003;10:58. doi: 10.1007/bf02255998. [DOI] [PubMed] [Google Scholar]
- 105.Gebistorf F., Karam O., Wetterslev J., Afshari A. Inhaled nitric oxide for acute respiratory distress syndrome (ARDS) in children and adults. Cochrane Database Syst Rev. 2016 doi: 10.1002/14651858.cd002787.pub3. CD002787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Karam O., Gebistorf F., Wetterslev J., Afshari A. The effect of inhaled nitric oxide in acute respiratory distress syndrome in children and adults: a cochrane systematic review with trial sequential analysis. Anaesthesia. 2017;72:106. doi: 10.1111/anae.13628. [DOI] [PubMed] [Google Scholar]
- 107.Darwish I., Miller C., Kain K.C., Liles W.C. Inhaled nitric oxide therapy fails to improve outcome in experimental severe influenza. Int J Med Sci. 2012;9:157. doi: 10.7150/ijms.3880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Martel J., Ko Y.F., Young J.D., Ojcius D.M. Could nasal nitric oxide help to mitigate the severity of COVID-19? Microbes Infect. 2020 doi: 10.1016/j.micinf.2020.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Berra L., Lei C., Su B., Dong H., Safaee Fakhr B., Grassi L.G., Di Fenza R., Gianni S., Pinciroli R., Vassena E., Araujo Morais C.C., Bellavia A., Spina S., Kacmarek R., Carroll R. Protocol for a randomized controlled trial testing inhaled nitric oxide therapy in spontaneously breathing patients with COVID-19. medRxiv. 2020 doi: 10.1101/2020.03.10.20033522. 2020.03.10.20033522. [DOI] [Google Scholar]
- 110.Lei C., Su B., Dong H., Bellavia A., Di Fenza R., Safaee Fakhr B., Gianni S., Grassi L.G., Kacmarek R., Araujo Morais C.C., Pinciroli R., Vassena E., Berra L. Protocol of a randomized controlled trial testing inhaled Nitric Oxide in mechanically ventilated patients with severe acute respiratory syndrome in COVID-19 (SARS-CoV-2) medRxiv. 2020 doi: 10.1101/2020.03.09.20033530. 2020.03.09.20033530. [DOI] [Google Scholar]
- 111.Gianni S., Safaee Fakhr B., Morais C.C.A., Di Fenza R., Larson G., Pinciroli R., Houle T., Mueller A.L., Bellavia A., Kacmarek R., Carroll R., Berra L. Nitric oxide gas inhalation to prevent COVID-2019 in healthcare providers. medRxiv. 2020 doi: 10.1101/2020.04.05.20054544. 2020.04.05.20054544. [DOI] [Google Scholar]
- 112.J. Road, Inhaled Gaseous Nitric Oxide (gNO) Antimicrobial Treatment of Difficult Bacterial and Viral Lung (COVID-19) Infections (NCT03331445), (2020)
- 113.Hakim T.S., Sugimori K., Camporesi E.M., Anderson G. Half-life of nitric oxide in aqueous solutions with and without haemoglobin. Physiol Meas. 1996;17:267. doi: 10.1088/0967-3334/17/4/004. [DOI] [PubMed] [Google Scholar]
- 114.Vaughn M.W., Kuo L., Liao J.C. Estimation of nitric oxide production and reaction rates in tissue by use of a mathematical model. Am J Physiol. 1998;274:H2163. doi: 10.1152/ajpheart.1998.274.6.h2163. [DOI] [PubMed] [Google Scholar]
- 115.Hall J.R., Rouillard K.R., Suchyta D.J., Brown M.D., Ahonen M.J.R., Schoenfisch M.H. Mode of Nitric Oxide Delivery Affects Antibacterial Action. ACS Biomater Sci Eng. 2019;6:433. doi: 10.1021/acsbiomaterials.9b01384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Ormerod A.D., White M.I., Shah S.A., Benjamin N. Molluscum contagiosum effectively treated with a topical acidified nitrite, nitric oxide liberating cream. Br J Dermatol. 1999;141:1051. doi: 10.1046/j.1365-2133.1999.03204.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Hebert A.A., Siegfried E.C., Durham T., De León E.N., Reams T., Messersmith E., Maeda-Chubachi T. Efficacy and tolerability of an investigational nitric oxide–releasing topical gel in patients with molluscum contagiosum: a randomized clinical trial. J Am Acad Dermatol. 2020;82:887. doi: 10.1016/j.jaad.2019.09.064. [DOI] [PubMed] [Google Scholar]
- 118.NOVAN, novan reports top-line efficacy results from phase 3 trials of SB206 for Molluscum Contagiosum. http://investors.novan.com/news-releases/news-release-details/novan-reports-top-line-efficacy-results-phase-3-trials-sb206, 2020 (Accessed 1 July 2020).
- 119.Akaberi D., Krambrich J., Ling J., Luni C., Hedenstierna G., Järhult J.D., Lennerstrand J., Lundkvist Å. Mitigation of the replication of SARS-CoV-2 by nitric oxide in vitro. Redox Biol. 2020;37 doi: 10.1016/j.redox.2020.101734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Zaragoza C., Ocampo C.J., Saura M., McMillan A., Lowenstein C.J. Nitric oxide inhibition of coxsackievirus replication in vitro. J Clin Invest. 1997;100:1760. doi: 10.1172/jci119702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Lin Y.L., Huang Y.L., Ma S.H., Yeh C.T., Chiou S.Y., Chen L.K., Liao C.L. Inhibition of Japanese encephalitis virus infection by nitric oxide: antiviral effect of nitric oxide on RNA virus replication. J Virol. 1997;71:5227. doi: 10.1128/jvi.71.7.5227-5235.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Akerstrom S., Mousavi-Jazi M., Klingstrom J., Leijon M., Lundkvist A., Mirazimi A. Nitric oxide inhibits the replication cycle of severe acute respiratory syndrome coronavirus. J Virol. 2005;79:1966. doi: 10.1128/jvi.79.3.1966-1969.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Brisbois E.J., Davis R.P., Jones A.M., Major T.C., Bartlett R.H., Meyerhoff M.E., Handa H. Reduction in thrombosis and bacterial adhesion with 7 Day implantation of S-Nitroso-N-acetylpenicillamine (SNAP)-Doped Elast-eon E2As Catheters in Sheep. J Mater Chem B. 2015;3:1639. doi: 10.1039/c4tb01839g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Riccio D.A., Schoenfisch M.H. Nitric oxide release: part I. macromolecular scaffolds. Chem Soc Rev. 2012;41:3731. doi: 10.1039/c2cs15272j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Carpenter A.W., Schoenfisch M.H. Nitric oxide release: part II. therapeutic applications. Chem Soc Rev. 2012;41:3742. doi: 10.1039/c2cs15273h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Hopkins S.P., Pant J., Goudie M.J., Schmiedt C., Handa H. Achieving Long-Term Biocompatible Silicone via Covalently Immobilized S-Nitroso- N-acetylpenicillamine (SNAP) that exhibits 4 months of sustained nitric oxide release. ACS Appl Mater Interfaces. 2018;10:27316. doi: 10.1021/acsami.8b08647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Lautner G., Meyerhoff M.E., Schwendeman S.P. Biodegradable poly(lactic-co-glycolic acid) microspheres loaded with S-nitroso-N-acetyl-D-penicillamine for controlled nitric oxide delivery. J Control Release. 2016;225:133. doi: 10.1016/j.jconrel.2015.12.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Estes L.M., Singha P., Singh S., Sakthivel T.S., Garren M., Devine R., Brisbois E., Seal S., Handa H. Characterization of a Nitric Oxide (NO) donor molecule and Cerium Oxide Nanoparticle (CNP) interactions and their synergistic antimicrobial potential for biomedical applications. J Colloid Interface Sci. 2020 doi: 10.1016/j.jcis.2020.10.081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Gao S., Zhang W., Wang R., Hopkins S.P., Spagnoli J.C., Racin M., Bai L., Li L., Jiang W., Yang X., Lee C., Nagata K., Howerth E.W., Handa H., Xie J., Ma Q., Kumar A. Nanoparticles encapsulating nitrosylated maytansine to enhance radiation therapy. ACS Nano. 2020;14:1468. doi: 10.1021/acsnano.9b05976. [DOI] [PubMed] [Google Scholar]
- 130.Singha P., Workman C.D., Pant J., Hopkins S.P., Handa H. Zinc‐oxide nanoparticles act catalytically and synergistically with nitric oxide donors to enhance antimicrobial efficacy. J Biomed Mater Res Part A. 2019;107:1425. doi: 10.1002/jbm.a.36657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Quinn J.F., Whittaker M.R., Davis T.P. Delivering nitric oxide with nanoparticles. J Control Release. 2015;205:190. doi: 10.1016/j.jconrel.2015.02.007. [DOI] [PubMed] [Google Scholar]
- 132.Wo Y., Brisbois E.J., Bartlett R.H., Meyerhoff M.E. Recent advances in thromboresistant and antimicrobial polymers for biomedical applications: just say yes to nitric oxide (NO) Biomater Sci. 2016;4:1161. doi: 10.1039/c6bm00271d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Singha P., Locklin J., Handa H. A review of the recent advances in antimicrobial coatings for urinary catheters. Acta Biomater. 2017;50:20. doi: 10.1016/j.actbio.2016.11.070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Bryan N.S., Grisham M.B. Methods to detect nitric oxide and its metabolites in biological samples. Free Radic Biol Med. 2007;43:645. doi: 10.1016/j.freeradbiomed.2007.04.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Kirmse M., Hess D., Fujino Y., Kacmarek R.M., Hurford W.E. Delivery of inhaled nitric oxide using the Ohmeda INOvent Delivery System. Chest. 1998;113:1650. doi: 10.1378/chest.113.6.1650. [DOI] [PubMed] [Google Scholar]
- 136.Heidersbach R.S., Johengen M.J., Bekker J.M., Fineman J.R. Inhaled nitric oxide, oxygen, and alkalosis: dose-response interactions in a lamb model of pulmonary hypertension. Pediatr Pulmonol. 1999;28:3. doi: 10.1002/(sici)1099-0496(199907)28:1<3::aid-ppul2>3.0.co;2-s. 10.1002/(sici)1099-0496(199907)28:1%3C3::aid-ppul2%3E3.0.co;2-s. [DOI] [PubMed] [Google Scholar]
- 137.Zell R., Markgraf R., Schmidtke M., Gorlach M., Stelzner A., Henke A., Sigusch H.H., Gluck B. Nitric oxide donors inhibit the coxsackievirus B3 proteinases 2A and 3C in vitro, virus production in cells, and signs of myocarditis in virus-infected mice. Med Microbiol Immunol. 2004;193:91. doi: 10.1007/s00430-003-0198-6. [DOI] [PubMed] [Google Scholar]
- 138.Beretta M., Gruber K., Kollau A., Russwurm M., Koesling D., Goessler W., Keung W.M., Schmidt K., Mayer B. Bioactivation of nitroglycerin by purified mitochondrial and cytosolic aldehyde dehydrogenases. J Biol Chem. 2008;283:17873. doi: 10.1074/jbc.m801182200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Chen Z., Foster M.W., Zhang J., Mao L., Rockman H.A., Kawamoto T., Kitagawa K., Nakayama K.I., Hess D.T., Stamler J.S. An essential role for mitochondrial aldehyde dehydrogenase in nitroglycerin bioactivation. Proc Natl Acad Sci U S A. 2005;102:12159. doi: 10.1073/pnas.0503723102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Horton A., Schiefer I.T. Pharmacokinetics and pharmacodynamics of nitric oxide mimetic agents. Nitric Oxide. 2019;84:69. doi: 10.1016/j.niox.2019.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Broniowska K.A., Diers A.R., Hogg N. S-nitrosoglutathione. Biochim Biophys Acta. 2013;1830:3173. doi: 10.1016/j.bbagen.2013.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Zhu J.D., Meng W., Wang X.J., Wang H.C. Broad-spectrum antiviral agents. Front Microbiol. 2015;6:517. doi: 10.3389/fmicb.2015.00517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Ali-Ahmad D., Bonville C.A., Rosenberg H.F., Domachowske J.B. Replication of respiratory syncytial virus is inhibited in target cells generating nitric oxide in situ. Front Biosci. 2003;8:a48. doi: 10.2741/986. [DOI] [PubMed] [Google Scholar]
- 144.Majano P.L., Garcia-Monzon C. Does nitric oxide play a pathogenic role in hepatitis C virus infection? Cell Death Differ. 2003;10(Suppl 1):S13. doi: 10.1038/sj.cdd.4401115. [DOI] [PubMed] [Google Scholar]
- 145.Shi J.B., Gao J., Wang Y.P., Yao Q.Z. Synthesis and antiviral activity of 4H-[1,2,5]oxadiazolo[3,4-d]pyrimidine-5,7-dione 1-oxide nucleosides. Chin Chem Lett. 2009;20:404. doi: 10.1016/j.cclet.2008.12.009. [DOI] [Google Scholar]
- 146.Xu X., Wang J., Yao Q. Synthesis and quantitative structure-activity relationship (QSAR) analysis of some novel oxadiazolo[3,4-d]pyrimidine nucleosides derivatives as antiviral agents. Bioorg Med Chem Lett. 2015;25:241. doi: 10.1016/j.bmcl.2014.11.065. [DOI] [PubMed] [Google Scholar]
- 147.Mathe C., Gosselin G. L-nucleoside enantiomers as antivirals drugs: a mini-review. Antivir Res. 2006;71:276. doi: 10.1016/j.antiviral.2006.04.017. [DOI] [PubMed] [Google Scholar]
- 148.Shi J.B., Xu S., Wang Y.P., Li J.J., Yao Q.Z. Synthesis of new pyrimidine nucleoside derivatives with nitric oxide donors for antiviral activity. Chin Chem Lett. 2011;22:899. doi: 10.1016/j.cclet.2011.01.010. [DOI] [Google Scholar]
- 149.Bi Z., Reiss C.S. Inhibition of vesicular stomatitis virus infection by nitric oxide. J Virol. 1995;69:2208. doi: 10.1128/jvi.69.4.2208-2213.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Cheng J., He K., Shen Z., Zhang G., Yu Y., Hu J. Nitric Oxide (NO)-releasing macromolecules: rational design and biomedical applications. Front Chem. 2019;7:530. doi: 10.3389/fchem.2019.00530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Kumari S., Sammut I.A., Giles G.I. The design of nitric oxide donor drugs: s-nitrosothiol tDodSNO is a superior photoactivated donor in comparison to GSNO and SNAP. Eur J Pharmacol. 2014;737:168. doi: 10.1016/j.ejphar.2014.05.012. [DOI] [PubMed] [Google Scholar]
- 152.Harding J.L., Reynolds M.M. Metal organic frameworks as nitric oxide catalysts. J Am Chem Soc. 2012;134:3330. doi: 10.1021/ja210771m. [DOI] [PubMed] [Google Scholar]
- 153.Ahonen M.J.R., Dorrier J.M., Schoenfisch M.H. Antibiofilm efficacy of nitric oxide-releasing alginates against cystic fibrosis bacterial pathogens. ACS Infect Dis. 2019;5:1327. doi: 10.1021/acsinfecdis.9b00016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Chonkar A., Nayak U., Udupa N. Smart polymers in nasal drug delivery. Indian J Pharm Sci. 2015;77:367. doi: 10.4103/0250-474x.164770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Chaturvedi M., Kumar M., Pathak K. A review on mucoadhesive polymer used in nasal drug delivery system. J Adv Pharm Technol Res. 2011;2:215. doi: 10.4103/2231-4040.90876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Jiao J., Zhang L. Influence of intranasal drugs on human nasal mucociliary clearance and ciliary beat frequency. Allergy Asthma Immunol Res. 2019;11:306. doi: 10.4168/aair.2019.11.3.306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Yang T., Zelikin A.N., Chandrawati R. Progress and promise of nitric oxide-releasing platforms. Adv Sci (Weinh) 2018;5 doi: 10.1002/advs.201701043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.M. H. Schoenfisch, E. M. Hetrick, N. A. Stasko, C. B. Johnson, Use of nitric oxide to enhance the efficacy of silver and other topical wound care agents, US 8,399,005 B2, (2013), https://patents.google.com/patent/US8399005B2/en.
- 159.M. H. Schoenfisch, J. H. Shin, N. Stasko, Nitric oxide-releasing particles for nitric oxide therapeutics and biomedical applications, US 9.403,852 B2, (2016), https://patents.google.com/patent/US9403852B2/en.
- 160.Seabra A.B., Durán N. Nitric oxide-releasing vehicles for biomedical applications. J. Mater. Chem. 2010;20:1624. doi: 10.1039/B912493B. [DOI] [Google Scholar]
- 161.Sukhal S., Sethi J., Ganesh M., Villablanca P.A., Malhotra A.K., Ramakrishna H. Extracorporeal membrane oxygenation in severe influenza infection with respiratory failure: a systematic review and meta-analysis. Ann Card Anaesth. 2017;20:14. doi: 10.4103/0971-9784.197820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Yang Y., Peng F., Wang R., Guan K., Jiang T., Xu G., Sun J., Chang C. The deadly coronaviruses: The 2003 SARS pandemic and the 2020 novel coronavirus epidemic in China. J Autoimmun. 2020;109 doi: 10.1016/j.jaut.2020.102434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Xie X., Lyu J., Hussain T., Li M. Drug prevention and control of ventilator-associated Pneumonia. Front Pharmacol. 2019;10:298. doi: 10.3389/fphar.2019.00298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Homeyer K.H., Singha P., Goudie M.J., Handa H. S-Nitroso-N-acetylpenicillamine impregnated endotracheal tubes for prevention of ventilator-associated pneumonia. Biotechnol Bioeng. 2020 doi: 10.1002/bit.27341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Roberts T.R., Garren M., Handa H., Batchinsky A.I. Toward an artificial endothelium: development of blood-compatible surfaces for extracorporeal life support. J Trauma Acute Care Surg. 2020:1. doi: 10.1097/ta.0000000000002700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Bartlett R.H., Ogino M.T., Brodie D., McMullan D.M., Lorusso R., MacLaren G., Stead C.M., Rycus P., Fraser J.F., Belohlavek J., Salazar L., Mehta Y., Raman L., Paden M.L. Initial ELSO guidance document: ECMO for COVID-19 patients with severe cardiopulmonary failure. ASAIO J. 2020;66:472. doi: 10.1097/mat.0000000000001173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Kowalewski M., Fina D., Slomka A., Raffa G.M., Martucci G., Lo Coco V., De Piero M.E., Ranucci M., Suwalski P., Lorusso R. COVID-19 and ECMO: the interplay between coagulation and inflammation-a narrative review. Crit Care. 2020;24:205. doi: 10.1186/s13054-020-02925-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Li H., Liu L., Zhang D., Xu J., Dai H., Tang N., Su X., Cao B. SARS-CoV-2 and viral sepsis: observations and hypotheses. Lancet. 2020;395:1517. doi: 10.1016/s0140-6736(20)30920-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Beyls C., Huette P., Abou-Arab O., Berna P., Mahjoub Y. Extracorporeal membrane oxygenation for COVID-19-associated severe acute respiratory distress syndrome and risk of thrombosis. Br J Anaesth. 2020 doi: 10.1016/j.bja.2020.04.079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Jia H.P., Look D.C., Shi L., Hickey M., Pewe L., Netland J., Farzan M., Wohlford-Lenane C., Perlman S., McCray P.B., Jr. ACE2 receptor expression and severe acute respiratory syndrome coronavirus infection depend on differentiation of human airway epithelia. J Virol. 2005;79:14614. doi: 10.1128/jvi.79.23.14614-14621.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Xu H., Zhong L., Deng J., Peng J., Dan H., Zeng X., Li T., Chen Q. High expression of ACE2 receptor of 2019-nCoV on the epithelial cells of oral mucosa. Int J Oral Sci. 2020;12:8. doi: 10.1038/s41368-020-0074-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Lang J., Yang N., Deng J., Liu K., Yang P., Zhang G., Jiang C. Inhibition of SARS pseudovirus cell entry by lactoferrin binding to heparan sulfate proteoglycans. PLoS One. 2011;6:e23710. doi: 10.1371/journal.pone.0023710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Ontaneda A., Annich G.M. Novel Surfaces in Extracorporeal Membrane Oxygenation Circuits. Front Med (Lausanne) 2018;5:321. doi: 10.3389/fmed.2018.00321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Liu Q., Singha P., Handa H., Locklin J. Covalent grafting of antifouling phosphorylcholine-based copolymers with antimicrobial nitric oxide releasing polymers to enhance infection-resistant properties of medical device coatings. Langmuir. 2017;33:13105. doi: 10.1021/acs.langmuir.7b02970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Singha P., Goudie M.J., Liu Q., Hopkins S., Brown N., Schmiedt C.W., Locklin J., Handa H. Multipronged approach to combat catheter-associated infections and thrombosis by combining nitric oxide and a Polyzwitterion: a 7 day in vivo study in a Rabbit Model. ACS Appl Mater Interfaces. 2020;12:9070. doi: 10.1021/acsami.9b22442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Major T.C., Brant D.O., Burney C.P., Amoako K.A., Annich G.M., Meyerhoff M.E., Handa H., Bartlett R.H. The hemocompatibility of a nitric oxide generating polymer that catalyzes S-nitrosothiol decomposition in an extracorporeal circulation model. Biomaterials. 2011;32:5957. doi: 10.1016/j.biomaterials.2011.03.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Goudie M.J., Brisbois E.J., Pant J., Thompson A., Potkay J.A., Handa H. Characterization of an S-nitroso-N-acetylpenicillamine-based nitric oxide releasing polymer from a translational perspective. Int J Polym Mater. 2016;65:769. doi: 10.1080/00914037.2016.1163570. [DOI] [PMC free article] [PubMed] [Google Scholar]