

Abstract
Oleanane and ursane pentacyclic triterpenoids are secondary metabolites of plants found in various climatic zones and regions. This group of compounds is highly attractive due to their diverse biological properties and possible use as intermediates in the synthesis of new pharmacologically promising substances. By now, their antiviral, anti-inflammatory, antimicrobial, antitumor, and other activities have been confirmed. In the last decade, methods of microbial synthesis of these compounds and their further biotransformation using microorganisms are gaining much popularity. The present review provides clear evidence that industrial microbiology can be a promising way to obtain valuable pharmacologically active compounds in environmentally friendly conditions without processing huge amounts of plant biomass and using hazardous and expensive chemicals. This review summarizes data on distribution, microbial synthesis, and biological activities of native oleanane and ursane triterpenoids. Much emphasis is put on the processes of microbial transformation of selected oleanane and ursane pentacyclic triterpenoids and on the bioactivity assessment of the obtained derivatives.
Keywords: biological activity, biotransformation, glycyrrhetinic acid, oleanolic acid, ursolic acid
1. Introduction
Drugs derived from secondary plant metabolites make up about 25% of the global pharmaceutical market [1]. Secondary metabolites of plants are several groups of compounds; the most numerous (about 25,000 representatives) and diverse group is terpenic hydrocarbons and their oxygen-containing derivatives (terpenoids). Depending on the number of isoprene units (C5H8) in their structure, they contain a certain number of carbon atoms and are classified into mono-(C10), sesqui-(C15), di-(C20), triterpenoids (C30), etc.
Naturally occurring triterpenoids are represented by more than 100 various types of skeletons [2]. Native triterpenoids, in particular, oleanane and ursane representatives, are of interest for researchers due to their availability and multiple biological activities, including antimicrobial, anti-inflammatory, antitumor, cytotoxic, hepatoprotective, and other activities [3,4,5,6,7]. Triterpenic molecules, however, are highly hydrophobic which significantly limits their use as effective pharmacological agents. At present, one of the most common ways to increase the effectiveness and bioavailability of triterpenoids is by chemical modification. This usually requires high temperature and pH, use of expensive reagents, and introduction of protective groups of molecule reactive centers [8,9,10,11]. An alternative way to obtain valuable derivatives is by biotransformation under normal and environmentally friendly conditions employing the catalytic activity of microorganisms with high regio- and stereoselectivity in one technological stage. Furthermore, microbial conversion ensures specific modifications of triterpenic molecule sites that are either not modified or poorly modified by synthetic transformations [12]. Note that, among the known microbial biocatalysts, members of mycelial fungi are the most studied [13,14,15] whereas bacterial catalysts are only represented by a few gram-positive species [16,17,18,19,20]. The first papers on microbial transformation of triterpenoids were published in the 1960s [21]. The earliest information related to bioconversion processes of oleanane derivatives catalyzed by fungi, such as Curvularia lunata ATCC 13432, Trichotecium roseum ATCC 8685, Cunninghamella sp. ATCC 3229, Mucor griseo-cyanus ATCC 1207-a, Helicostylum piriforme ATCC 8992, Fusarium lini, and Cunninghamella blakesleana dates back to about the same time [22,23,24,25]. The data on bacterial transformation of oleanane triterpenoids by Streptomyces sp. G-20 and Chainia antibiotica IFO 12,246 were reported in the second half of the 1980s [26,27]. As for microbial transformations of ursane pentacyclic triterpenoids by both fungal (Mucor plumbeus ATCC 4740 [28]) and bacterial (Nocardia sp. NRRL 5646 [29]) strains, those studies were initiated only in the 2000s. Henceforth, the interest in the topic discussed has been increasing and the Active Triterpenoid Biocatalysts List has been expanded every year, as is the number of various bioactive triterpenic derivatives formed via biotransformations [30,31,32].
Now, preparation of biologically active compounds based on pentacyclic triterpenoids is an actual research discussed in plenty of experimental and review publications [8,12,32,33,34]. However, the reviews are overwhelmingly focused on chemical transformations or describe specific types of biological activities of triterpenoids. Less frequently, they deal with biological transformations. The latest of the few reviews on microbial transformations of pentacyclic triterpenoids include literature data up to 2016 [12,32,33,35]. Our review summarizes the data from 2013 to the present on distribution, microbial biosynthesis, biological activity, and mainly biotransformation of oleanane and ursane pentacyclic triterpenoids to obtain promising biologically active compounds or intermediates of their synthesis.
2. Distribution in Nature
Plant pentacyclic triterpenoids are represented by more than two dozen structural types; the most common are oleanane and ursane ones (Figure 1). In free forms, triterpenoids are nonvolatile lipophilic substances that are soluble in organic solvents and insoluble in water. The most available oleanane triterpenoids are oleanolic acid 1 (OA, 3β-hydroxy-olean-12-en-28-oic acid, C30H48O3, CAS 508-02-1) and glycyrrhetinic acid 2 (GA, 3β-hydroxy-11-oxo-18β-olean-12-en-30-oic acid, C30H46O4, CAS 471-53-4), and ursolic acid 3 (UA, 3β-hydroxy-urs-12-en-28-oic acid, C30H48O3, CAS 77-52-1) is the most available ursane triterpenoid. Biosynthesis of oleanane and ursane pentacyclic triterpenoids in plants occurs by conversion of the acyclic triterpene squalene (4) to 2,3-oxidosqualene (5) and its further cyclization by specific enzyme complexes (oxidosqualene cyclase) via β-amyrin (6) or α-amyrin (7), respectively [36,37]. The carbon skeletons of these triterpenoids consist of five condensed cyclohexane rings. What differentiates them is one methyl group (CH3-29) located in oleanane and ursane derivatives at C20 and C19 of the E ring, respectively.
Figure 1.

Representative skeletons of oleanane (A) and ursane (B) pentacyclic triterpenoids.
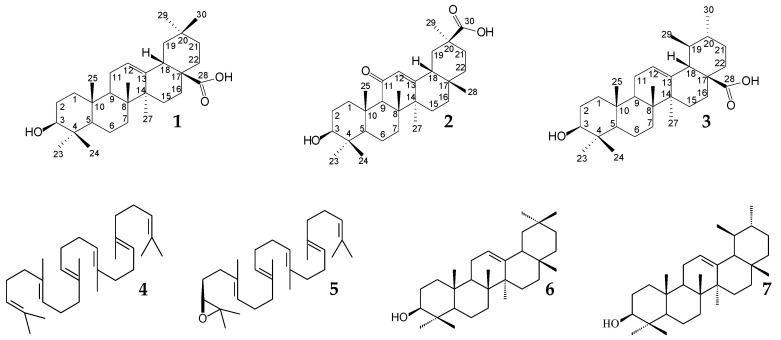
Representatives of various higher plant families are active producers of both oleanane and ursane triterpenoids (Table 1). Frequently, OA and UA are simultaneously detected in the same plant sources. OA and UA contents in Meconopsis henrici, Dracocephalum tanguticum, Comastoma pulmonaria, Corydalis impatiens, and Swertia racemosa—traditionally used in Chinese medicine—can reach 0.96 ± 0.01 mg/g and 0.64 ± 0.01 mg/g dry weight, respectively [38]. Flowers and leaves of the shrubs Ocimum tenuiflorum and Syzygium aromaticum and the herbs Origanum vulgare, Rosmarus officinalis, and Salvia officinalis used as condiments contain up to 15.3 mg/g OA and up to 26.2 mg/g UA (wet weight) [39]. The main source of OA is considered to be the fruits and leaves of Olea europaea. The acid content in olive leaves can reach 27.16 mg/g wet weight and 25.09 ± 0.72 mg/g dry weight [39,40]. GA is commonly extracted from herbaceous plants of the genus Glycyrrhiza [41,42,43]; the acid content in their roots can reach 10.2 ± 1.7 mg/g wet weight [44].
Table 1.
Some plant sources of oleanolic acid (OA), ursolic acid (UA), and glycyrrhetinic acid (GA).
| Compound | Plant Source | Part of Plant | Reference |
|---|---|---|---|
| Apocynaceae Juss., nom. cons. | |||
| OA, UA | Alstonia scholaris (L.) R.Br. | Leaves | [49] |
| OA, UA | Plumeria obtusa L. var. sericifolia | Leaves | [50] |
| Aquifoliaceae DC. ex. A.Rich. | |||
| OA, UA | Ilex guayusa Loes. | Leaves | [39] |
| Araliaceae Juss., nom. cons. | |||
| OA | Panax stipuleanatus H.T.Tsai & K.M.Feng | Roots | [51] |
| Asteraceae Bercht. & J.Presl, nom. cons. | |||
| OA | Baccharis uncinella DC. | Leaves | [52] |
| Betulaceae Gray | |||
| OA | Betula pendula Roth. | Bark | [3,53] |
| Cornaceae Bercht. & J.Presl, nom. cons. | |||
| UA | Cornus officinalis Torr. ex Dur. | Seeds | [54] |
| Ebenaceae Gürke, nom. cons. | |||
| OA, UA | Diospyros L. | Fruits | [55] |
| Ericaceae Juss., nom. cons. | |||
| OA, UA | Arctostaphylos uva-ursi (L.) Spreng. | Fruits | [39] |
| OA, UA | Vaccinium vitis-idaea L. | Fruits | [53] |
| Fabaceae Lindl. | |||
| GA | Glycyrrhiza glabra L. | Roots | [41] |
| Glycyrrhiza uralensis Fisch. | Roots | [42] | |
| Glycyrrhiza inflate Batalin | Roots | [43] | |
| Gentianaceae Juss., nom. cons. | |||
| OA, UA | Comastoma pulmonaria Turcz. | Sprouts, roots | [38] |
| Swertia racemosa Wall. ex Griseb. | Sprouts, roots | ||
| Lamiaceae Martinov | |||
| OA, UA | Ocimum tenuiflorum L. | Leaves | [39] |
| Lavandula angustifolia Mill. | Flowers | ||
| Origanum vulgare L. | Leaves | ||
| Rosmarinus officinalis L. | Leaves | ||
| Salvia officinalis L. | Leaves | ||
| Dracocephalum tanguticum Maxim | Sprouts, roots | [38] | |
| Lythraceae J.St.-Hil. | |||
| OA, UA | Punica granatum L. | Fruits | [38] |
| Punica granatum L.cv. Daqingpi | Flowers | [56] | |
| Malvaceae Juss., nom. cons. | |||
| OA | Durio zibethinus Murr. | Fruits | [7] |
| Myrtaceae Juss., nom. cons. | |||
| OA, UA | Syzygium aromaticum (L.) Merr. & L.M.Perry | Flowers | [39] |
| Eucalyptus globules Labill. | Leaves | ||
| Oleaceae Hoffmanns. & Link, nom. cons. | |||
| OA | Olea europaea L.cv. Picual | Fruits, leaves | [40] |
| Olea europaea L. cv. Cornezuelo | |||
| OA, UA | Olea europaea L. | Leaves | [39] |
| Papaveraceae Juss. | |||
| OA, UA | Meconopsis henrici Bureau & Franch. | Sprouts, roots | [38] |
| Corydalis impatiens (Pall.) Fisch. ex DC. | Sprouts, roots | ||
| Pinaceae Lindl. | |||
| UA | Picea abies (L.) H.Karst. | Resin, cons, sprouts | [57] |
| Rosaceae Juss. | |||
| OA, UA | Malus P. Mill. | Fruits | [55] |
| Pyrus L. | Fruits | ||
| UA | Fragaria ananassa (Duchesne ex Weston) Duchesne ex Rozier | Perianth | [4] |
| UA | Potentilla fulgens Wall. ex Hook. | Roots | [58] |
| OA, UA | Eriobotrya japonica (Thunb.) Lindl | Leaves | [59] |
| Rubiaceae Juss., nom. cons. | |||
| OA | Uncaria laevigata Wall. ex G.Don | Stem bark | [60] |
| UA | Emmenopterys henryi Oliv. | Leaves, sprouts | [61] |
| Vitaceae Juss., nom. cons. | |||
| OA | Vitis vinifera L. | Fruits | [55] |
Note: GA, glycyrrhetinic acid; OA, oleanolic acid; UA, ursolic acid.
The amount of pentacyclic triterpenoids in plants is not constant and can significantly vary depending on the activity of enzyme systems and external factors [45]. Thus, the fruits and leaves of olive (Olea europaea) of various varieties contained OA from 0.4 ± 0.1 mg/g to 0.81 ± 0.16 mg/g dry weight and from 29.2 ± 1.8 mg/g to 34.5 ± 3.1 mg/g dry weight, respectively [46,47]. The OA content decreased by 70‒80% during olive fruit ripening [40]. The same tendency was observed when grapes (Vitis vinifera) ripen [48]. Changes in pentacyclic triterpenoid concentrations in plant sources may be related to specific climate, season, landscape, and cultivation strategies [47].
3. Biosynthesis of Pentacyclic Triterpenic Acids Using Microorganisms
Today, pentacyclic triterpenoids and their natural derivatives are mainly obtained by extraction from plant sources. However, the extraction and separation of these compounds (often with organic solvents) are extremely labor-intensive, and energy- and time-consuming. Besides, most of pentacyclic triterpenoids are found in relatively low concentrations in plants, entailing the use of huge amounts of plant raw materials and the formation of waste biomass in large volumes [62]. An alternative source of pentacyclic triterpenoids seems to be highly efficient cell factories, increasingly popular in the last decade. They allow to obtain valuable biologically active compounds of plant origin in environmentally friendly conditions using available compounds as the sole source of carbon (glucose, galactose, and ethanol) [63]. Cell factories are usually yeast cells—natural catalysts of the mevalonate (MVA) pathway—with plant genes responsible for pentacyclic triterpenoid synthesis introduced into their genome. The MVA pathway includes formation of mevalonate involving 3-hydroxy-3-methylglutaryl-CoA reductases (HMG1). The mevalonate formed is further transformed into isopentenyl diphosphate and dimethylallyl diphosphate, being converted to farnesyl diphosphate by a farnesyl phosphate synthase (ERG20). This pathway provides natural synthesis of squalene (4)—a common precursor of triterpenoids—by the squalene synthase (ERG9) based on two molecules of farnesyl diphosphate and its further transformation into 2,3-oxidosqualene (5) by squalene epoxidase (ERG1) [64]. Subsequent synthesis of pentacyclic triterpenoids involves plant genes encoding amyrin synthase, CYP450, and CYP450 reductase (Scheme 1).
Scheme 1.

Biosynthesis of oleanane and ursane triterpenoids in engineered yeasts. Enzymes (in frames): HMG1, 3-hydroxy-3-methylglutaryl-CoA reductase; ERG20, farnesyl phosphate synthase; ERG9, squalene synthase; ERG1, squalene epoxidase; βAS, β-amyrin synthase; mix-AS, mixed amyrin synthase; CYP450, cytochrome P450. Metabolites: IPP, isopentenyl diphosphate; DMAPP, dimethylallyl diphosphate; and FPP, farnesyl diphosphate. Modified from Lu et al., 2018 and Zhao et al., 2018 [64,65].
Genes encoding β-amyrin synthase (βAS) that catalyze the formation of β-amyrin (6)—a precursor of oleanane pentacyclic triterpenoids—from 2,3-oxidosqualene (5) were isolated from the genomes of Glycyrrhiza glabra, Panax ginseng, Catharanthus roseus, Lotus japonicus, Artemisia annua, Chenopodium quinoa, and others [65,66,67,68,69]. Because no enzymes were found to synthesize exclusively α-amyrin (7), a precursor of ursane pentacyclic triterpenoids, this reaction involves mixed amyrin synthases (mix-AS) that catalyze the formation of both α- and β-amyrin from Eriobotrya japonica and C. roseus [64,70]. Medicago truncatula is most often used to search for genes encoding CYP450 enzymes that catalyze subsequent conversion of α- and β-amyrin [64,65,66,67,69,71]. Less frequently, Phaseolus vulgaris [72], Bupleurum falcatum [71], G. uralensis [62], C. roseus [64], Crataegus pinnatifida [70], Solanum lycoperum, P. ginseng [68], and others are used to search for these genes. Native microbial cytochrome P450 reductases are often unable to transfer electrons to foreign CYP450s, as required for catalysis, and the source of additional CYP450 reductases (CPR and ATR) is usually Arabidopsis thaliana [62,64,65,66,69,71]. In a few studies, CYP450 reductases were obtained from M. truncatula [65], L. japonicus [67], G. uralensis [62], and V. vinifera [70]. Various approaches, including modification, overexpression, or inactivation of microorganisms’ own genes; insertion of plant genes in the yeast genome by various techniques; as well as combinatorial biosynthesis, are used to enhance microbial biosynthesis and to achieve an increased yield of pentacyclic triterpenoids (Table 2).
Table 2.
Biosynthesis of pentacyclic triterpenoids using engineered yeasts.
| Parent Strain | Modification | Product (Yield) | Reference |
|---|---|---|---|
| Saccharomyces cerevisiae INVSc1 | Insertion of βAS, CPR (Lotus japonicus), CYP93E2, and CYP72A61 (Medicago truncatula) | β-Amyrin (6 **, 1.07 mg/L), 24-hydroxy-β-amyrin (13, 0.27 mg/L), and soyasapogenol B (21, 1.35 mg/L) | [67] |
| Insertion of βAS, CPR (L. japonicus), CYP716A12, and CYP72A68 (M. truncatula) | β-Amyrin (6, 0.55 mg/L), erythrodiol (8, 0.09 mg/L), and gypsogenic acid (22, 0.96 mg/L) | ||
| Insertion of βAS, CPR (L. japonicus), CYP716A12, and CYP93E2 (M. truncatula) | Erythrodiol (8), OA, 4-epi-hederagenin (24), and others (trace amount) | ||
| Insertion of βAS, CPR (L. japonicus), CYP72A63, and CYP716A12 (M. truncatula) | Erythrodiol (8), OA, queretaroic acid (25), and others (trace amount) | ||
| Insertion of βAS, CPR (L. japonicus), CYP93E2, and CYP72A63 (M. truncatula) | Probable 11-deoxo-GA (23) and others (trace amount) | ||
| S. cerevisiae BY4742 | Insertion of βAS (Glycyrrhiza glabra) | β-Amyrin (6, 36.2 ± 3.9 mg/L) | [71] |
| Insertion of βAS (M. truncatula) | β-Amyrin (6, 19.0 ± 1.0 mg/L) | ||
| S. cerevisiae TM3 | Insertion of AtATR1 (Arabidopsis thaliana), CYP716Y1 (Bupleurum falcatum), and CYP716A12 (M. truncatula) | β-Amyrin (6), erythrodiol (8), OA (1), oleanolic aldehyde (9), and 16α-hydroxy-oleanolic aldehyde (10) | |
| Insertion of CYP716Y1 (B. falcatum), CYP716A12 (M. truncatula), AtATR1 (A. thaliana), and UGT73C11 (Barbarea vulgaris) | 3-O-Glc-Echinocystic acid (11), 3-O-Glc-OA (12) | ||
| Insertion of AtATR1 (A. thaliana) and CYP93E2 (M. truncatula) | 24-Hydroxy-β-amyrin (13, 1.3%) | [72] | |
| Insertion of AtATR1 (A. thaliana) and CYP93E7 (Lens culinaris) | 24-Hydroxy-β-amyrin (13, 16.2%) | ||
| Insertion of AtATR1 (A. thaliana) and CYP93E8 (Pisum sativum) | 24-Hydroxy-β-amyrin (13, 37.6%), probable 3β-hydroxy-olean-12-en-24-oic acid (14, 3.2%) | ||
| Insertion of AtATR1 (A. thaliana) and CYP93E5 (Cicer arietinum) | 24-Hydroxy-β-amyrin (13, 51.3%), probable 3β-hydroxy-olean-12-en-24-oic acid (14, 7.6%) | ||
| Insertion of AtATR1 (A. thaliana) and CYP93E6 (G. glabra) | 24-Hydroxy-β-amyrin (13, 50.2%), probable 3β-hydroxy-olean-12-en-24-oic acid (14, 6.3%) | ||
| Insertion of AtATR1 (A. thaliana) and CYP93E4 (Arachis hypogaea) | 24-Hydroxy-β-amyrin (13, 47.0%), probable 3β-hydroxy-olean-12-en-24-oic acid (14, 5.5%) | ||
| Insertion of AtATR1 (A. thaliana) and CYP93E1 (Glycine max) | 24-Hydroxy-β-amyrin (13, 58.8%), probable 3β-hydroxy-olean-12-en-24-oic acid (14, 13.5%) | ||
| Insertion of AtATR1 (A. thaliana) and CYP93E9 (Phaseolus vulgaris) | 24-Hydroxy-β-amyrin (13, 67.6%), probable 3β-hydroxy-olean-12-en-24-oic acid (14, 11.8%) | ||
| S. cerevisiae BY4742 | Deletion of TRP1 | Squalene (4, 9.6 mg/L) | [66] |
| S. cerevisiae BY4742-TRP | Overexpression of tHMG1 and LYS2 | Squalene (4, 150.9 mg/L) | |
| S. cerevisiae BY-T1 | Insertion of βAS (G. glabra) Overexpression of ERG1 and ERG9 |
β-Amyrin (6, 107.0 mg/L), squalene (4, 183.4 mg/L) | |
| Insertion of βAS (Panax ginseng) Overexpression of ERG1 and ERG9 |
β-Amyrin (6, 1.9 mg/L) | ||
| S. cerevisiae BY-βA-G | Insertion of βAS (G. glabra), OAS (M. truncatula), and AtCPR1 (A. thaliana) | OA (1, 71.0 mg/L), β-amyrin (6, 88.6 mg/L), and squalene (4, 141.2 mg/L) | |
| S. cerevisiae INVSc1 | Insertion of βAS (G. glabra) | β-Amyrin (6, 4.16 mg/L) | [73] |
| Insertion of βAS (G. glabra) and ERG1 (Candida albicans) | β-Amyrin (6, 24.50 mg/L) | ||
| Insertion of βAS (G. glabra), ERG1 (C. albicans), IDI (Escherichia coli) Overexpression of ERG9 and ERG20 |
β-Amyrin (6, 36.50 mg/L, 75.50 mg/L *) | ||
| Insertion of βAS (G. glabra), ERG1 (C. albicans), IDI (Escherichia coli) Overexpression of ERG9 and ERG20 Reconstruction of SRE into promoter |
β-Amyrin (6, 85.78 mg/L, 108.60 mg/L *) | ||
| S. cerevisiae W303-1a | Overexpression of tHMG1 and ERG20 | Squalene (4, 165.28 mg/L) | [64] |
| S. cerevisiae WTE | Insertion of mix-AS (Catharanthus roseus) Overexpression of tHMG1 and ERG20 |
β-Amyrin (6, 1.64 mg/L), α-amyrin (7, 5.64 mg/L) | |
| Insertion of mix-AS (Catharanthus roseus) and ERG1 (C. albicans) Overexpression of tHMG1 |
β-Amyrin (6, 24.95 mg/L, 44.92 mg/L *), α-amyrin (7, 97.31 mg/L, 175.15 mg/L *) | ||
| S. cerevisiae ScLCZ08 | Insertion of CrOAS (C. roseus) and AtCPR1 (A. thaliana) | OA (1, 29.49 mg/L), UA (3, 24.58 mg/L) | |
| Insertion of MtOAS (M. truncatula) and LjCPR1 (L. japonicus) | OA (1, 24.34 mg/L), UA (3, 23.37 mg/L) | ||
| Insertion of CrOAS (C. roseus) and LjCPR1 (L. japonicus) | OA (1, 23.91 mg/L), UA (3, 22.96 mg/L) | ||
| Insertion of MtOAS (M. truncatula) and AtCPR1 (A. thaliana) | OA (1, 31.41 mg/L, 155.58 mg/L *), UA (3, 25.85 mg/L, 123.37 mg/L *) | ||
| S. cerevisiae JDY52 | Insertion of GgβAS (G. glabra), MtCYP716A12 (M. truncatula), and AtCPR1 (A. thaliana) | OA (1, 2.5 ± 0.2 mg/L) | [65] |
| Insertion of GgβAS (G. glabra), MtCYP716A12 (M. truncatula), and LjCPR (L. japonicus) | OA (1, 6.3 ± 0.3 mg/L) | ||
| Insertion of GgβAS (G. glabra), MtCYP716A12 (M. truncatula), and GuCPR (G. uralensis) | OA (1, 7.1 ± 0.5 mg/L) | ||
| Insertion of GgβAS (G. glabra), MtCYP716A12, and MtCPR (M. truncatula) | OA (1, 9.0 ± 0.7 mg/L) | ||
| Insertion of GgβAS (G. glabra), MtCYP716A12, and MtCPR (M. truncatula) Knocking out of GAL1 |
OA (1, 41.3 ± 3.4 mg/L) | ||
| Insertion of GgβAS (G. glabra), MtCYP716A12, and MtCPR (M. truncatula) Knocking out of GAL1 and GAL80 |
OA (1, 70.3 ± 7.0 mg/L) | ||
| Insertion of GgβAS (G. glabra), MtCYP716A12, and MtCPR (M. truncatula) Knocking out of GAL1 and GAL80 Overexpression of tHMG1, ERG1, and ERG9 |
OA (1, 186.1 ± 12.4 mg/L, 606.9 ± 9.1 mg/L *) | ||
| S. cerevisiae INVSc1 | Insertion of CYP88D6, CYP72A154 (G. uralensis), and ATR1 (A. thaliana) | GA (2, 20.4 ± 7.7 µg/L), 11-oxo-β-amyrin (15, 0.5 ± 0.1 mg/L), β-amyrin (6, 21.23 ± 1.64 mg/L), 11α-hydroxy-β-amyrin (16), 30-hydroxy-11-oxo-β-amyrin (17), and glycyrrhetaldehyde (18) (trace amount) | [62] |
| S. cerevisiae INVSc1 (diploid) | Insertion of CYP88D6, CYP72A154 (G. uralensis), and ATR1 (A. thaliana) | GA (2, 31.8 ± 6.6 µg/L), 11-oxo-β-amyrin (15, 0.5 ± 0.06 mg/L) | |
| S. cerevisiae SGib | Insertion of CYP88D6, CYP72A154 (G. uralensis), and ATR1 (A. thaliana) | GA (2, 33.7 ± 7.4 µg/L), 11-oxo-β-amyrin (15, 7.5 ± 0.5 mg/L) | |
| Insertion of CYP88D6 and ATR1 (A. thaliana) | 11-oxo-β-amyrin (15, 3.1 ± 0.05 mg/L) | ||
| Insertion of Unigene25647 and ATR1 (A. thaliana) | 11-oxo-β-amyrin (15, 6.0 ± 0.1 mg/L) | ||
| Insertion of CYP72A63, Unigene25647 (G. uralensis), and ATR1 (A. thaliana) | GA (2, 40.9 ± 6.4 µg/L), 11-oxo-β-amyrin (15, 9.16 ± 1.20 mg/L) | ||
| Insertion of CYP72A154, Unigene25647 (G. uralensis), and ATR1 (A. thaliana) | GA (2, 42.3 ± 5.8 µg/L), 11-oxo-β-amyrin (15, 7.22 ± 1.58 mg/L) | ||
| Insertion of CYP72A154, Unigene25647, and GuCPR1 (G. uralensis) | GA (2, 517.4 ± 35.5 µg/L), 11-oxo-β-amyrin (15, 15.3 ± 1.6 mg/L) | ||
| Insertion of CYP72A63, Unigene25647, and GuCPR1 (G. uralensis) | GA (2, 7.4 ± 1.0 mg/L, 18.9 ± 2.0 mg/L *), 11-oxo-β-amyrin (15, 22.6 ± 0.9 mg/L, ~80 mg/L *) | ||
| S. cerevisiae BY-OA | Insertion of CYP716C49 (Crataegus pinnatifida) | Maslinic acid (19, 0.06 mg/L/OD600) | [70] |
| Insertion of CaCYP716C49 (Centella asiatica) | Maslinic acid (19, 0.2 mg/L/OD600) | ||
| Insertion of codon-optimized CaCYP716C49 (C. asiatica) | Maslinic acid (19, 0.45 mg/L/OD600, 384.3 mg/L *) | ||
| Insertion of mix-AS (Eriobotrya japonica), VvCYP716A15, and CPR (Vitis vinifera) | UA (3, 1.76 mg/L/OD600), OA (2, 0.61 mg/L/OD600) | ||
| Insertion of mix-AS (E. japonica), codon-optimized CaCYP716C49 (C. asiatica), VvCYP716A15, and CPR (V. vinifera) | Corosolic acid (20, 0.39 mg/L/OD600, 141.0 mg/L*) | ||
| S. cerevisiae BY4742 | Insertion of AaβAS (Artemisia annua) | β-Amyrin (6, 10.8 ± 1.0 mg/L) | [68] |
| Insertion of CqβAS (Chenopodium quinoa) | β-Amyrin (6, 10.8 ± 1.0 mg/L) | ||
| Insertion of PtβAS (Polygala tenuifolia) | β-Amyrin (6, 9.0 ± 0.7 mg/L) | ||
| Insertion of LjβAS (L. japonicus) | β-Amyrin (6, 8.2 ± 1.0 mg/L) | ||
| Insertion of EtβAS (Euphorbium tirucalli) | β-Amyrin (6, 8.0 ± 0.2 mg/L) | ||
| Insertion of SlβAS (Solanum lycopersicum) | β-Amyrin (6, 2.9 ± 0.3 mg/L) | ||
| Insertion of CYP716AL1 (C. roseus) and ATR2 (A. thaliana) | OA (1, 14.3 ± 1.6 mg/L), erythrodiol (8), and oleanolic aldehyde (9) (trace amount) | ||
| Insertion of CYP716A52v2 (P. ginseng) and ATR2 (A. thaliana) | OA (1, 3.0 ± 0.0 mg/L), erythrodiol (8), and oleanolic aldehyde (9) (trace amount) | ||
| Insertion of SlβAS (S. lycopersicum), CYP716AL1 (C. roseus), and ATR2 (A. thaliana) | OA (1, 3.9 ± 0.2 mg/L) | ||
| Insertion of SlβAS (S. lycopersicum), CYP716A52v2 (P. ginseng), and ATR2 (A. thaliana) | OA (1, 2.8 ± 0.0 mg/L) | ||
| Insertion of AaβAS (A. annua), CYP716AL1 (C. roseus), and ATR2 (A. thaliana) | OA (1, 8.5 ± 0.2 mg/L) | ||
| Yarrowia lipolytica ATCC 201249 | Insertion of βAS (C. roseus) | β-Amyrin (6, 16.0 mg/L) | [69] |
| Insertion of βAS (C. roseus), CYP716A12 (M. truncatula), and ATR1 (A. thaliana) | OA (1, 16.3 mg/L) | ||
| Insertion of βAS (C. roseus), CYP716A12 (M. truncatula), and ATR1(A. thaliana) Overexpression of ERG1, ERG9, ERG20, tHMG1 |
OA (1, 92.1 mg/L) | ||
| Insertion of βAS (C. roseus) and CYP716A2-linker (GSTSSG)-t46ATR1 Overexpression of ERG9, ERG20, and tHMG1 |
OA (1, 129.9 mg/L, 540.7 mg/L*) |
Note: * Yield of the compound in a 5-L fermenter. ** The number in bold is the number of the compound structural formula. GA, glycyrrhetinic acid; OA, oleanolic acid; UA, ursolic acid.
Overexpression of genes ERG1, ERG9, ERG20, and, more frequently, tHMG1 (HMG1 with truncated N-terminal 511 amino acids) involved in the natural microbial synthesis of 2,3-oxidosqualene (Scheme 1) can enhance the biosynthesis of triterpenoids [64,65,66,69]. Along with overexpression, the enhancement can also be facilitated by inactivation of TRP1 (phosphoribosylanthranilate isomerase), GAL1 (galactokinase), and GAL80 (galactose/lactose metabolism regulatory protein) involved in metabolic processes that “distract” cells from biosynthesis of triterpenoids [65,66]. Thus, overexpression of tHMG1, ERG1, and ERG9 and inactivation of GAL1 and GAL80 in the chromosome of Saccharomyces cerevisiae JDY52 and its use in a 5-L fermenter with 40 g/L glucose resulted in 606.9 ± 9.1 mg/L OA after 144 h. To date, it is the highest yield reported [65]. In another study, the use of Yarrowia lipolytica ATCC 201249 with overexpressed ERG1, ERG9, ERG20, and tHMG1 and the inserted expression modules βAS and CYP716A2-linker(GSTSSG)-t46ATR1 (ATR1 with truncated N-terminal 46 amino acids) provided the yield of 540.7 mg/L OA after 82 h in a 5-L fermenter with 100 g/L glucose [69]. Despite the fact that S. cerevisiae JDY52 produced relatively higher OA amounts (606.9 ± 9.1 mg/L) [65], the productivity of Y. lipolytica was 6.59 mg/L/h and exceeded that of S. cerevisiae (4.214 mg/L/h).
Various techniques of yeast genome modification were also applied to intensify the biosynthesis of triterpenoids. It was shown that the expression of inserted plant genes from low-copy and single-copy plasmids was more effective than that from integrated, high-copy, and multicopy plasmids [71,73]. In Reference [71], the activities of S. cerevisiae TM30 and S. cerevisiae TM44 obtained from the same parent strain by including different plasmid variations were evaluated. In the first case, the strain expressing CYP716Y1 and CYP716A12 to obtain a self-processing polyprotein with two enzymes bound via oligopeptide 2A catalyzed the formation of β-amyrin (6), erythrodiol (8), OA, oleanolic aldehyde (9), and 16α-hydroxy-oleanolic aldehyde (10), while the second strain produced two self-processing polyproteins, one consisting of CYP716Y1 and CYP716A12 and the other consisting of AtATR1 and UDP-dependent glycosyl transferase UGT73C11, and catalyzed the formation of 3-O-Glc-echinocystic acid (11) and 3-O-Glc-OA (12).
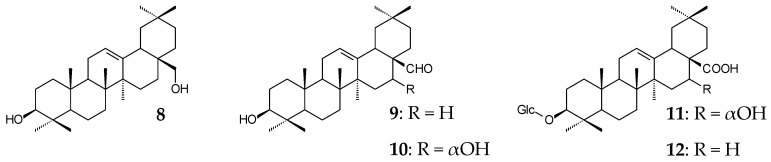
The most promising research area in this field is combinatorial biosynthesis. Change in the natural biosynthesis pathway or combination of synthesis genes from different sources leads to enhanced efficiency of microbial biosynthesis or to the formation of new compounds. The Moses group (2014) studied the C24-oxidizing activity of CYP93E2 orthologs from different species of the family Fabaceae. The obtained strains catalyzed the formation of 24-hydroxy-β-amyrin (13). Almost all strains (except one containing CYP93E7) simultaneously catalyzed the formation of 3β-hydroxy-olean-12-en-24-oic acid (14). The highest conversion (79.4%) of β-amyrin (6) to 24-hydroxy-β-amyrin was observed using saccharomycetes containing CYP93E9 (P. vulgaris) [72]. Dale et al. (2020) conducted a comparative study on catalytic activities of 12 βASs and 16 enzymes of the CYP716A subfamily. Of all the βASs studied, synthases derived from Artemisia annua and Chenopodium quinoa exhibited the highest catalytic activity (10.8 ± 1.0 mg/L β-amyrin). Comparatively, CYP716AL1 (C. roseus) showed the highest activity (14.3 ± 1.6 mg/L OA) of the 16 CYP716As studied. When a combination of genes encoding these enzymes was used, the OA yield made up 8.5 ± 0.2 mg/L. At the same time, saccharomycetes containing CYP716A75 (Maesa lanceolata), CYP716A79 (C. quinoa), CYP716A110 (Aquilegia coerulea), and CYP716A1 (A. thaliana) did not catalyze the formation of OA; erythrodiol (8) but not OA was detected in all cases [68]. Substitutions of CYP88D6 (C11 oxidase) for Unigene25647 (97% similarity) and ATR1 (A. thaliana) for GuCPR1 (G. uralensis) were performed, and a combination of GuCPR1 and two Unigene25647 cassettes was inserted into the genome of S. cerevisiae SGib. These manipulations resulted in the formation of 18.9 ± 2.0 mg/L GA and about 80.0 mg/L 11-oxo-β-amyrin (15) after 144 h of the fed-batch fermentation with ethanol (30 mL every 24 h) compared with the previously obtained 20.4 ± 7.7 µg/L GA and 0.5 ± 0.1 mg/L 11-oxo-β-amyrin (15) and β-amyrin (6) and trace amounts of 11α-hydroxy-β-amyrin (16), 30-hydroxy-11-oxo-β-amyrin (17), and glycyrrhetaldehyde (18) [62]. In strain S. cerevisiae BY-OA, substitution of CYP716C49 (C. pinnatifida) for the homologue (47.9% similarity) CaCYP716C49 (C. asiatica) allowed for obtaining 384.3 mg/L maslinic acid (19, 2α-hydroxy-OA) after 96-h incubation in a 5-L fermenter with glucose (5 g/L). When S. cerevisiae BY-T3 containing mix-AS, VvCYP716A15, CPR, and CaCYP716C49 from different plant sources were used in the same conditions; corosolic acid (20, 2α-hydroxy-UA) was formed after 144 h [70]. The activity of various combinations of CYP450 from M. truncatula was studied in Reference [67]. The main products of synthesis in different cases were soyasapogenol B (21), gypsogenic acid (22), or 11-deoxo-GA (23). Importantly, saccharomycetes expressing CYP716A12 and CYP93E2 catalyzed the formation of 4-epi-hederagenin (24), and the yeasts expressing CYP716A12 and CYP72A63 catalyzed the formation of queretaroic acid (25). These compounds were not previously detected in M. truncatula tissues, indicating the great potential of combinatorial biosynthesis using microorganisms [67].
4. Biological Activities of Triterpenic Acids and Their Native Derivatives
Extracts obtained from plant sources using various solvents and containing the pentacyclic triterpenoids reviewed in this paper usually exhibit a wide range of biological properties. Methanolic OA-containing extracts from various parts of Betula pendula exhibited antibacterial activity against test cultures Staphylococcus aureus and Bacillus subtilis [3]. Methanolic extracts from the aerial part of the tropical plant Baccharis uncinella containing OA and UA showed antiparasitic activity by limiting the growth of promastigote and amastigote forms of Leishmania amazonensis and enhanced the immune response in infected mice [52]. The ethyl acetate fraction of Glycyrrhiza uralensis root extract inhibited TNF-α-induced activation of NF-κB in HepG2 cells [42]. The ethyl acetate fraction of Potentilla fulgens root extract containing ursane triterpenoids showed antioxidant activity by inhibiting the production of free radicals [58]. An alcohol extract and ethyl acetate fraction of Fragaria ananassa perianth containing various triterpenoids had a pronounced cytotoxic effect on B16-F10 melanoma cells and inhibited their melanogenesis by 79.1% and 80.2%, respectively [4]. The allelopathic effect (inhibition of seed maturation and root growth of nearby plants) of Alstonia scholaris was also due to high UA content (2.5 ± 0.6 mg/g dry weight of leaves) [49].
Isolation of every triterpenic acid individually allows studying their bioactive properties in detail and explaining the pharmacological properties of some plants. The antibacterial property of birch bark was determined by a high content of pentacyclic triterpenoids, in particular, OA, which exhibits a pronounced antibacterial activity against S. aureus (minimal inhibitory concentration (MIC) 1.25%) and B. subtilis (MIC 0.625%) [3]. Antimicrobial activity of GA was manifested as the ability to reduce the motility of Pseudomonas aeruginosa cells and the level of biofilm formation. This can make a significant contribution to the development of effective antibiotic-free therapy for Pseudomonas infections [74]. UA was able to inhibit both the growth of Mycobacterium tuberculosis in vitro [75] and the replication of rotavirus in a dose-dependent manner [76]. In addition, OA and UA were shown to inhibit the COVID-19 (SARS-CoV-2) main protease, a key enzyme of the virus replication, through in silico studies [77,78].
OA, as an antitumor agent, increased the sensitivity of sarcoma cells to chemotherapeutic drugs in human soft tissues [79]. UA had similar properties, significantly increased the effectiveness of colorectal cancer chemotherapy, and reduced its side effects in vitro and in vivo [80]. The cytotoxic effect of the extract of Fragaria ananassa perianth on B16-F10 melanoma cells was determined partly by the presence of cytotoxic UA that suppressed the melanin production by 40.2% [4]. Additionally, UA reduced the spread of human myeloma cells by inhibiting the deubiquitinating protease USP7 [81] and caused apoptosis of gastric cancer cells by activating the caspases poly (ADP-ribose) polymerase and by inducing the release of reactive oxygen species [6].
The OA hepatoprotective activity was shown to be related to its inhibitory effect against carboxylesterase (therapeutic target for hypertriglyceridemia) and the hepatitis C virus (HCV) [5,82]. GA exhibited the hepatoprotective effect by inhibiting NO formation in rat hepatocytes, iNOS suppression, and COX-2 expression and by decreasing the activity of NF-κB transcription factor in HepG2 cells [42]. The ability of GA to stimulate a neuroprotective property of microglia and to suppress the MAPK signaling pathway of the central nervous system caused a decrease in the severity of experimental autoimmune encephalomyelitis in mice [83]. UA could be an effective antidiabetic agent due to its ability to inhibit α-glucosidase activity [60]. OA and UA exhibited their inhibitory effects against lypopolysaccharide (LPS)-induced NO production in RAW 264.7 cells that determined their anti-inflammatory activity [7].
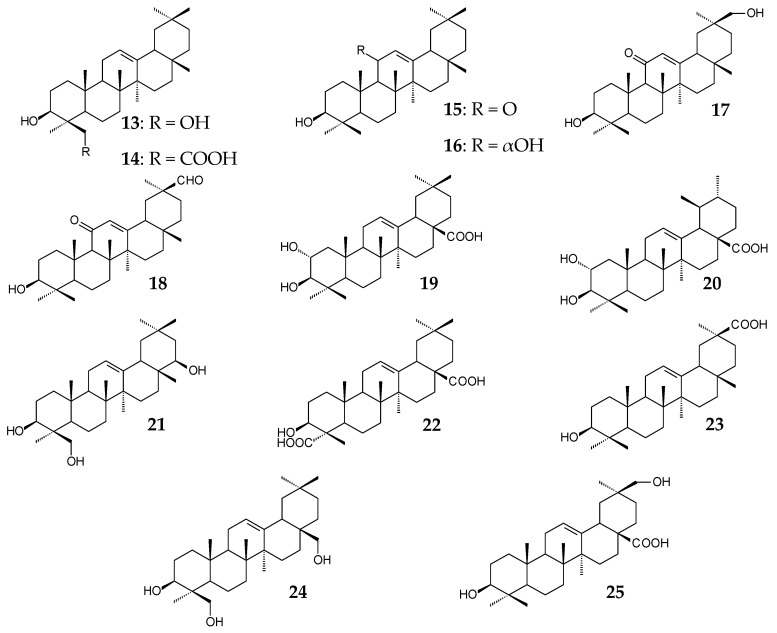
Along with their parent acids, the native derivatives also exhibit pronounced biological activities. Thus, the OA derivatives 22β-acetoxy-3,25-epoxy-3α-hydroxyolean-12-en-28-oic acid (26) and methyl 3,25-epoxy-3α-hydroxy-11-oxo-22β-senecioyloxyolean-12-en-28-oate (27) isolated from Lantana camara herb extract demonstrated antibacterial activity against a number of gram-positive and gram-negative bacteria [84].
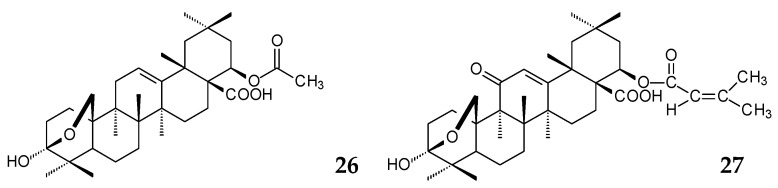
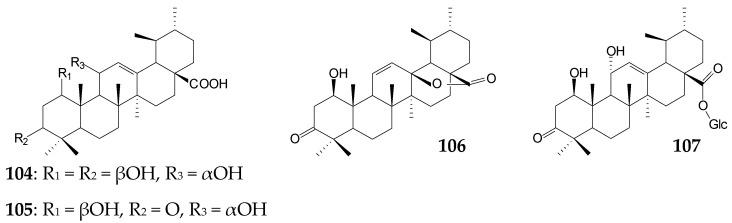
Oleanane 27-carboxy derivatives (28‒30) isolated from Chrysosplenium carnosum exhibited pronounced inhibitory activities against the mouse melanoma cell lines B16F10 and SP2/0 [85]. Natural 2α-hydroxylated OA derivative (20, corosolic acid) had cytotoxic effects on CaSki cells (human cervical cancer) by inducing apoptosis, by arresting the cell cycle in the G2/M phase, and by inhibiting the PI3K/Akt signaling pathway [86]. 2,3-seco-Derivative of the ursane type (31) from Siphonodon celastrineus showed pronounced cytotoxic activity against MOLT-3 cancer cells [87].
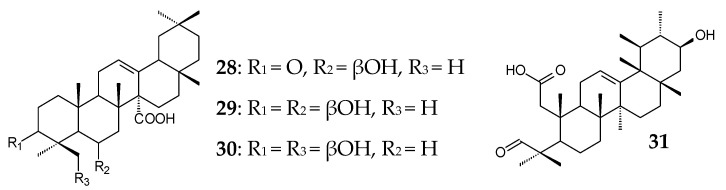
Oleanane derivatives from Panax stipuleanatus (C3, C28-diglycosides 32–34) and Astilbe rivularis (3β-trans-p-coumaroyloxy-olean-12-en-27-oic acid (35) and 6β-hydroxy-3-oxoolean-12-en-27-oic acid (36)) as well as oleanane and ursane polyhydroxylated derivatives (37–40) from Rosa laevigata exhibited anti-inflammatory activities [88,89,90] due to the suppression of TGFBIp-mediated hyperpermeability in vitro and in vivo as well [90]. Ursane derivatives from Durio zibethinus were shown to exhibit a more pronounced anti-inflammatory effect in LPS-induced NO production in RAW 264.7 cell inhibition tests compared to oleanane triterpenoids, for which the activity was reduced by C2 hydroxylation [7].
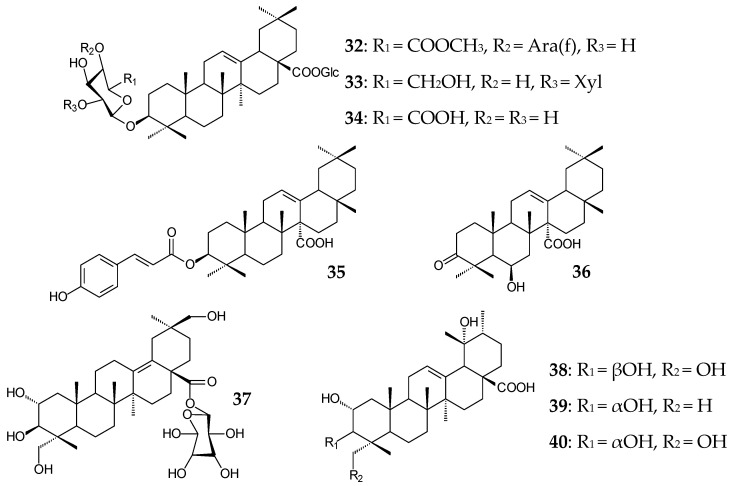
The pronounced inhibitory activity of Cecropia telentida root extracts against 11β-hydroxysteroid dehydrogenase could be determined by the presence of a new, probably 2α,20β-dihydroxylated UA derivative (41, isoyarumic acid) [91]. Ursane type 2,3-dihydroxy derivatives fulgic acids A (42) and B (43) isolated from Potentilla fulgens exhibited antioxidant effect by inhibiting the formation of free radicals [58]. Antiviral, antibacterial, hepatoprotective, anti-inflammatory, and antitumor activities of root extracts of Bupleurum chinense and B. scorzonerifolium species that are widespread in China were partially provided by the presence of bioactive oleanane saikosaponins (44–46) [92].
Due to the wide distribution and availability of the pentacyclic triterpenoids discussed in the review, they are often used to synthesize new semi-synthetic derivatives with antibacterial [93], antiviral [10,82,94], anti-inflammatory [95,96], hepatoprotective [5], antitumor [8,97,98,99], and other activities. Biological activities of oleanane and ursane semi-synthetic derivatives were studied in a number of reviews summarizing the data on derivative types [100,101,102] and biological activities [103,104,105].
5. Biological Transformation
Taking into account the relative availability of the discussed triterpenic acids in natural sources and their high bioactivity, it is interesting to assess the possibility of directed transformations of these compounds to expand the range of biologically active compounds and to increase their bioavailability. Chemical methods are currently the most tested and used to transform acids 1–3. However, chemical methods often require extreme acidity and temperature values, expensive catalysts, or protective groups of molecule reactive centers [8,9,10,106]. In contrast, biological transformation processes do not use aggressive reagents and can occur under normal eco-friendly conditions. Moreover, microorganisms are able to catalyze a wide range of regio- and stereoselective reactions that are difficult to perform chemically [12].
One of the most promising ways to highlight the pharmacological potential of native pentacyclic triterpenoids is the functionalization of their molecules by polyhydroxylation. Such functionalized derivatives hydroxylated by plant P450-dependent monooxygenases [107] are widespread in nature but are usually found in trace amounts or as part of a difficult-to-separate mixture. Enzymatic activity of microorganisms used for transformation of pentacyclic triterpenoids allows for obtaining hydroxylated derivatives with high yield and regioselectivity. Moreover, microbial hydroxylation occurs not only in the A ring but also at hard-to-reach positions on the B, D, and E rings. In addition to hydroxylation, microbial functionalization of pentacyclic triterpenoids can occur by less frequent reactions of carboxylation, glycosylation, lactone formation, and others.
5.1. Fungal Transformation
The described biotransformation processes of the compounds discussed in this review often occur using mycelial fungi of various species from the phyla Ascomycota (orders Glomerellales, Hypocreales) and Mucoromycota (order Mucorales). Fungal conversions of these compounds are accompanied by the formation of derivatives with hydroxyl groups at C1, C7, C15, C21, C24, or C30; oxo groups at C3, C7, or C21; glucopyranoside groups at C3, C28, or C30; lactone groups at C28/C13 or C3/C4, etc. as well as by the A ring fragmentation. The acid concentration used in biotransformation experiments usually ranges from 0.02 g/L to 1.0 g/L. The yield of transformation products (1.0% to 77.5%) and the duration of the processes (2 to 20 days) vary depending on the fungal catalyst characteristics (Table 3).
Table 3.
Biological activity of pentacyclic triterpenoid products derived by biotransformation.
| Microorganism | Initial Compound, Concentration | Derivative, Yield | Biological Activity | Reference |
|---|---|---|---|---|
| Fungi | ||||
| Rhizomucor miehei CECT 2749 | OA, 0.5 g/L | 1β,30-dihydroxy-OA (47), 5.0% | - * | [108] |
| 7β,30-dihydroxy-OA (48), 6.0% | - | |||
| 30-hydroxy-OA (25, queretaroic acid), 5.0% | Antitumor | |||
| OA methyl ester, 0.3 g/L | Methyl 3β,7β,30-trihydroxy-oleane-9(11),12-dien-28-oate (49), 15.0% | - | [112] | |
| Trichothecium roseum (M 95.56) | OA, 0.08 g/L | 7β,15α-dihydroxy-3-oxo-olean-12-en-28-oic acid (51), 7.5% | Anti-inflammatory | [114] |
| 15α-hydroxy-3-oxo-olean-12-en-28-oic acid (52), 6.25% | - | |||
| Circinella muscae AS 3.2695 | OA, 0.02 g/L | 7β,15α-dihydroxy-3-oxo-olean-12-en-28-oic acid (51), 6.1% | Anti-inflammatory | [15] |
| 7β-hydroxy-OA (53), 5.8% | Anti-inflammatory | |||
| 7β,21β-dihydroxy-OA (54), 4.2% | - | |||
| 7α,21β-dihydroxy-OA (55), 3.1% | - | |||
| 7β,15α-dihydroxy-OA (56), 4.9% | Anti-inflammatory | |||
| 7β-hydroxy-3-oxo-olean-12-en-28-oic acid (57), 3.8% | - | |||
| 7β,15α-dihydroxy-OA 28-O-β-d-glucopyranosyl ester (58), 4.7% | Anti-inflammatory | |||
| 21β-hydroxy-OA 28-O-β-D-glucopyranosyl ester (59), 5.1% | Anti-inflammatory | |||
| OA 28-O-β-D-glucopyranosyl ester (60), 5.5% | Anti-inflammatory | |||
| GA, 0.06 g/L | 7β-hydroxy-GA (61), 1.5% | Anti-inflammatory Antimicrobial |
[115] | |
| 15α-hydroxy-GA (62), 1.9% | Anti-inflammatory | |||
| 7β,15α-dihydroxy-GA (63), 1.8% | Anti-inflammatory Antioxidant Hepatoprotective |
|||
| 3,11-dioxo-7β-hydroxy-18β-olean-12-en-30-oic acid (64), 1.7% | Anti-inflammatory Antimicrobial |
|||
| 7β,15α-dihydroxy-3,11-dioxo-18β-olean-12-en-30-oic acid (65), 2.2% | Anti-inflammatory | |||
| 7β-hydroxy-11-oxo-18β-olean-12-en-30-oic acid 3-O-β-d-glucopyranoside (66), 1.8% | Anti-inflammatory | |||
| 7β-hydroxy-11-oxo-18β-olean-12-en-30-oic acid 3-O-β-d-6′-O-acetyl-glucopyranoside (67), 1.1% | Anti-inflammatory | |||
| 15α-hydroxy-11-oxo-18β-olean-12-en-30-oic acid 3-O-β-d-glucopyranoside (68), 2.5% | Anti-inflammatory | |||
| 15α-hydroxy-11-oxo-18β-olean-12-en-30-oic acid 3-O-β-d-6′-O-acetyl-glucopyranoside (69), 1.8% | Anti-inflammatory | |||
| 7β-hydroxy-GA 30-O-β-d-glucopyranoside (70), 2.2% | Anti-inflammatory | |||
| Rhizopus arrhizus AS 3.2893 | GA, 0.06 g/L | 7β-hydroxy-GA (61), 2.8% | Anti-inflammatory Antimicrobial |
[115] |
| 15α-hydroxy-GA (62), 2.2% | Anti-inflammatory | |||
| 7β,15α-dihydroxy-GA (63), 1.7% | Anti-inflammatory Antioxidant Hepatoprotective |
|||
| 3β-acetoxy-7β-hydroxy-11-oxo-18β-olean-12-en-30-oic acid (71), 1.7% | Anti-inflammatory | |||
| 7-oxo-GA (72), 1.3% | Anti-inflammatory | |||
| 7α-hydroxy-GA (73), 2.6% | Anti-inflammatory | |||
| 15α-hydroxy-7-oxo-GA (74), 1.3% | Anti-inflammatory | |||
| Colletotrichum lini AS 3.4486 | GA, 0.25 g/L | 7β,15α-dihydroxy-GA (63), NR ** | Anti-inflammatory Antioxidant Hepatoprotective |
[118] |
| Absidia pseudocylinderospora ATCC 24169 | GA, 1.0 g/L | 7β,15α-dihydroxy-GA (63), 18.0% | Anti-inflammatory Antioxidant Hepatoprotective |
[117] |
| Cunninghamella blakesleana CGMCC 3.970 | GA, 0.1 g/L | 15α,24-dihydroxy-GA (75), 0.11% | Anti-inflammatory | [14] |
| 15α,24-dihydroxy-3,11-dioxo-18β-olean-12-en-30-oic acid (76), 0.75% | - | |||
| 7β,24-dihydroxy-3,11-dioxo-18β-olean-12-en-30-oic acid (77), 0.16% | - | |||
| 3,11-dioxo-7β,15α,24-trihydroxy-18β-olean-12-en-30-oic acid (78), 0.29% | Anti-inflammatory | |||
| 7α,24-dihydroxy-3,11-dioxo-18β-olean-12-en-30-oic acid (79), 1.27% | Anti-inflammatory | |||
| C. blakesleana CGMCC 3.970 | GA, 0.3 g/L | 7β-hydroxy-GA (61), 30.0% | Anti-inflammatory Antimicrobial |
[116] |
| 3,11-dioxo-7β-hydroxy-18β-olean-12-en-30-oic acid (64), 25.0% | Anti-inflammatory Antimicrobial |
|||
| C. elegans TSY-0865 | GA, 0.4 g/L | 7β-hydroxy-GA (61), 2.5% | Anti-inflammatory Antimicrobial |
[119] |
| Syncephalastrum racemosum GCMCC 3.2500 | UA, 0.1 g/L | 7β,21β-dihydroxy-UA (80), 12.9% | - | [13] |
| 1β,21β-dihydroxy-UA (81), 3.9% | - | |||
| 1β-hydroxy-21-oxo-UA (82), 12.1% | - | |||
| 3β,21β-dihydroxy-urs-11-en-28,13-olide (83), 3.4% | Antihepatitis | |||
| 3β,7β,21β-trihydroxy-urs-11-en-28,13-olide (84), 2.9% | Antihepatitis | |||
| Gliocladium roseum CGMCC 3.3657 | UA, 0.1 g/L | 21β-hydroxy-3-oxo-urs-12-en-3,4-olide-28-oic acid (85), 8.0%) | - | [121] |
| 3,21-dioxo-urs-12-en-3,4-olide-28-oic acid (86), 6.25% | Antihepatitis | |||
| 21β-hydroxy-3,4-seco-ursane-4(23),12-diene-3,28-dioic acid (87), 1.5% | - | |||
| 21-oxo-3,4-seco-ursane-4(23),12-diene-3,28-dioic acid (88), 1.0% | Antihepatitis | |||
| Mucor spinosus AS 3.3450 | UA, 0.3 g/L | 7β-hydroxy-UA 28-ethanone (89), 5.04% | Antitumor | [122] |
| 7β,21β-dihydroxy-UA (81), 1.64% | - | |||
| 21β-hydroxy-urs-12-en-28-oic acid 3-O-β-d-glucopyranoside (90), 2.13% | - | |||
| Rhizopus chinensis CICC 40335 | OA, 0.2 g/L | 7β,21β-dihydroxy-OA (54), 53.75% | - | [34] |
| GA, 0.2 g/L | 7β-hydroxy-GA (61), 77.5% | Anti-inflammatory Antimicrobial |
||
| Bacteria | ||||
| Nocardia iowensis (DSM 45197, NRRL 5646) | OA, 0.3 g/L | Methyl 3β-hydroxy-olean-12-en-28-oat (91), 63.0% | Antitumor | [16] |
| Methyl 3-oxo-olean-12-en-28-oat (92, oleanonic acid methyl ester), 5.0% | Antitumor Antiprotozoal |
|||
| Metabolite 93 with hydroxyl and methyl groups, 2.0% | - | |||
| Streptomyces griseus ATCC 13273 | OA, 0.04 g/L | 3β-hydroxy-olean-12-ene-28,29-dioic acid (94), 21.9% | - | [128] |
| 3β,24-dihydroxy-olean-12-ene-28,29-dioic acid (95), 32.7% | - | |||
| 3β,21β,24-trihydroxy-olean-12-ene-28,29-dioic acid (96), 5.9% | Anti-inflammatory | |||
| S. griseus ATCC 13273 | UA, 0.04 g/L | 3β-hydroxy-urs-12-ene-28,30-dioic acid (97), 30.4% | - | [17] |
| 3β,24-dihydroxy-urs-12-ene-28,30-dioic acid (98), 31.6% | - | |||
| 3-oxo-UA, 0.04 g/L | 3-oxo-urs-12-ene-28,30-dioic acid (100), 24,1% | - | ||
| 24-hydroxy-3-oxo-urs-12-ene-28,30-dioic acid (101), 45.9% | - | |||
| 2α-hydroxy-UA, 0.04 g/L | 2α,3β-dihydroxy-urs-12-ene-28,30-dioic acid (102), 29.0% | - | ||
| 2α,3β,24-trihydroxy-urs-12-ene-28,30-dioic acid (103), 15.9% | - | |||
| Bacillus megaterium CGMCC 1.1741 | UA, 0.2 g/L | 1β,11α-dihydroxy-UA (104), 26.87% | - | [18] |
| 3-oxo-urs-12-en-28-oic acid (99), 6.23% | Anti-leishmania | |||
| 1β,11α-dihydroxy-3-oxo-urs-12-en-28-oic acid (105), 13.5% | Anti-inflammatory | |||
| 1β-hydroxy-3-oxo-urs-12-en-28,13-olide (106), 5.03% | Anti-inflammatory | |||
| 1β,11α-dihydroxy-3-oxo-urs-12-en-28-O-β-d-glucopyranoside (107), 8.57% | - | |||
| B. subtilis ATCC 6633 | 7β,21β-dihydroxy-OA, 0.2 g/L | 7β,21β-dihydroxy-olean-12-en-28-oic acid 3-O-β-d-glucopyranoside (108), 46.5% | - | [34] |
| GA, 0.2 g/L | GA 30-O-β-d-glucopyranoside (111), 27.5% | - | ||
| 7β-hydroxy-GA, 0.2 g/L | 7β-hydroxy-GA 30-O-β-d-glucopyranoside (68), 44.0% | - | ||
| S. griseus ATCC 13273 | 7β,21β-dihydroxy-OA, 0.2 g/L | 7β,21β,29-trihydroxy-OA (109), 26.0% | - | |
| 3β,7β,21β-trihydroxy-olean-12-ene-28,29-dioic acid (110), 15.0% | Neuroprotective | |||
Note: * Biological activity of the compound has not yet been detected. ** The value is not reported. GA, glycyrrhetinic acid; OA, oleanolic acid; UA, ursolic acid.
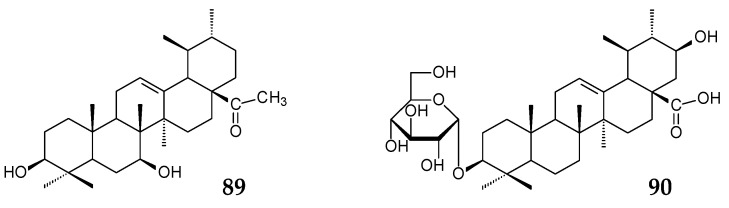
Abundantly found in nature, Rhizomucor miehei CECT 2749 partially metabolized OA (approximately 0.5 g/L) for 13 days to form of 1β,30-dihydroxy-OA (47), 7β,30-dihydroxy-OA (48), and 30-hydroxy-OA (25) in equivalent amounts (5.0‒6.0%) [108]. Compound 25, known as queretaroic acid, was first isolated from Lemaireocereus queretaroensis and L. beneckei endemic to Mexico [109]. This compound was shown to exhibit a moderate antitumor activity against HeLa cells [110]. Queretaroic acid 25 (3.3%) was obtained by the 24-h transformation of OA (0.2 g/L) by Escherichia coli cells expressing Nonomuraea recticatena CYP450 moxA and Pseudomonas redox partner camAB. At the same time, the use of cell-free reaction systems allowed to increase the yield of compound 25 to 17.0% [111]. Transformation of OA methyl ester (approximately 0.3 g/L) by R. miehei CECT 2749 during 13 days was accompanied by 7,30-dihydroxylation and, in addition, a 9(11),12-diene moiety formation in the C ring, producing 15.0% methyl 3β,7β,30-trihydroxy-oleane-9(11),12-dien-28-oate (49) [112].
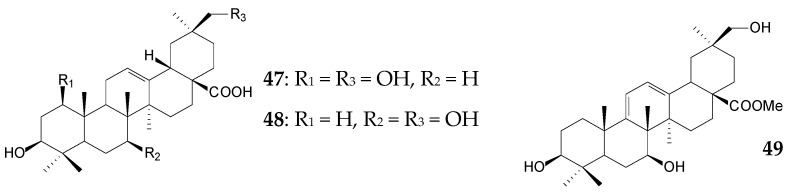
Colletotrichum lini AS 3.4486 was shown to catalyze the C15 hydroxylation of OA to form 15α-hydroxy-OA (50) [113]. In turn, ascomycete Trichothecium roseum (M 95.56) [114] and mucoromycete Circinella muscae AS 3.2695 [15] catalyzed the oxidation of OA (0.08 g/L and 0.02 g/L, respectively) to form 7β,15α-dihydroxy-3-oxo-olean-12-en-28-oic acid (51) on day 6 (7.5%) and day 7 (6.1%), respectively. At the same time, an intermediate (6.25%) of the dihydroxylation process—15α-hydroxy-3-oxo-olean-12-en-28-oic acid (52)—was also isolated from the culture medium of T. roseum (M 95.56) [114]. C. muscae AS 3.2695 simultaneously catalyzed a wide variety of hydroxylation and glycosylation reactions with the formation of 7β-hydroxy-OA (53), 7β,21β-dihydroxy-OA (54), 7α,21β-dihydroxy-OA (55), 7β,15α-dihydroxy-OA (56), 7β-hydroxy-3-oxo-olean-12-en-28-oic acid (57), 7β,15α-dihydroxy-OA 28-O-β-d-glucopyranosyl ester (58), 21β-hydroxy-OA 28-O-β-d-glucopyranosyl ester (59), and OA 28-O-β-d-glucopyranosyl ester (60) ranging from 3.1% to 5.8% [15]. C7 hydroxylation and C28 glycosylation presumably contributed to an increase in the anti-inflammatory activity of derivatives, while C21 hydroxylation led to a decreased ability of compounds to inhibit the release of LPS-induced nitric oxide by RAW 264.7 cells [15].
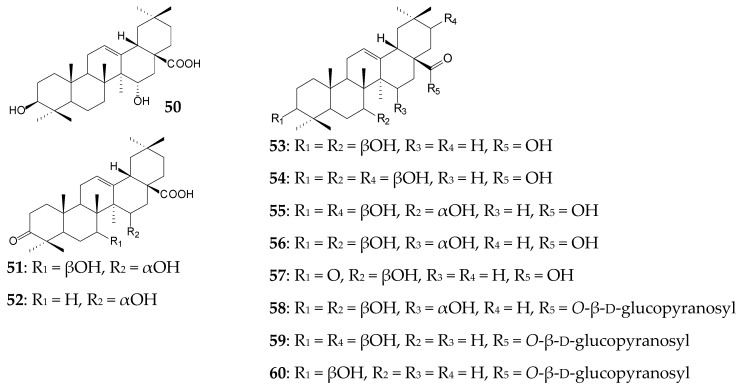
C. muscae AS 3.2695 was also active against GA (approximately 0.06 g/L) and initiated oxidation, acetylation, and glycosylation reactions with the formation of metabolites (yield did not exceed 2.4%) on day 7. The metabolites included 7β-hydroxy-GA (61), 15α-hydroxy-GA (62), 7β,15α-dihydroxy-GA (63), 3,11-dioxo-7β-hydroxy-18β-olean-12-en-30-oic acid (64), 7β,15α-dihydroxy-3,11-dioxo-18β-olean-12-en-30-oic acid (65), 7β-hydroxy-11-oxo-18β -olean-12-en-30-oic acid 3-O-β-d-glucopyranoside (66), 7β-hydroxy-11-oxo-18β-olean-12-en-30-oic acid 3-O-β-d-6′-O-acetyl-glucopyranoside (67), 15α-hydroxy-11-oxo-18β-olean-12-en-30-oic acid 3-O-β-d-glucopyranoside (68), 15α-hydroxy-11-oxo-18β-olean-12-en-30-oic acid 3-O-β-d-6′-O-acetyl-glucopyranoside (69), and 7β-hydroxy-GA 30-O-β-D-glucopyranoside (70) [115]. The above GA derivatives inhibited LPS-induced NO release by RAW 264.7 cells to different extents. Moreover, compounds 61 and 64 were shown to exhibit antimicrobial activity against the antibiotic-resistant strain Enterococcus faecalis [116], while compound 63 exhibited antioxidant and hepatoprotective properties [117].

The same authors reported Rhizopus arrhizus AS 3.2893 to perform oxidative transformation of GA (approximately 0.06 g/L) at C3, C7, and C15 for 7 days with the formation of 7β-hydroxy-GA (61), 15α-hydroxy-GA (62), 7β,15α-dihydroxy-GA (63), 3β-acetoxy-7β-hydroxy-11-oxo-18β -olean-12-en-30-oic acid (71), 7-oxo-GA (72), 7α-hydroxy-GA (73), and 15α-hydroxy-7-oxo-GA (74) (the yield of each compound did not exceed 2.8%). They also exhibited anti-inflammatory effects in the LPS-induced NO production inhibition test in RAW 264.7 cells [115].
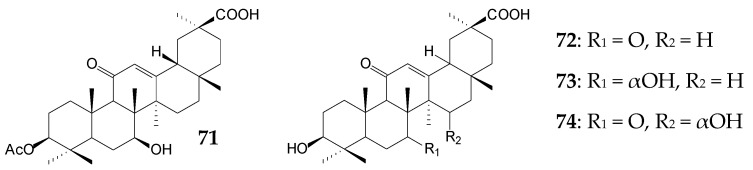
Biological hydroxylation of GA at concentrations of 0.25 g/L (48 h) and 1.0 g/L (14 days) at C7 and C15 producing the main metabolite 7β,15α-dihydroxy-GA (63) was also catalyzed by C. lini AS 3.4486 [118] and Absidia pseudocylinderospora ATCC 24169 [117], respectively. Moreover, A. pseudocylinderospora ATCC 24169, during long-term cultivation (14 days), catalyzed the formation of more than 18.0% of this compound (63) possessing antioxidant, hepatoprotective [117], and anti-inflammatory [115] activities.
Preparative biotransformation of 0.1 g/L GA by Cunninghamella blakesleana CGMCC 3.970 for 7 days resulted in a mixture of 15α,24-dihydroxy-GA (75), 15α,24-dihydroxy-3,11-dioxo-18β -olean-12-en-30-oic acid (76), 7β,24-dihydroxy-3,11-dioxo-18β-olean-12-en-30-oic acid (77), 3,11-dioxo-7β,15α,24-trihydroxy-18β-olean-12-en-30-oic acid (78), and 7α,24-dihydroxy-3,11 -dioxo-18β-olean-12-en-30-oic acid (79). The yield of each acid did not exceed 1.3% [14]. Compounds 75, 78, and 79 were found to effectively inhibit LPS-induced NO production in mouse microglia cells with IC50 values of 0.76 mmol/L, 0.94 mmol/L, and 0.16 mmol/L, respectively [14]. Interestingly, when C. blakesleana AS 3.970 was used, an increase in GA concentration to 0.3 g/L led to accumulation of two main products 7β-hydroxy-GA (61, 30.0%) and 3,11-dioxo-7β-hydroxy-18β-olean-12-en-30-oic acid (64, 25.0%) with pronounced antibacterial activity after 5 days [116]. In the case of C. elegans TSY-0865, only 2.5% of 7β-hydroxy-GA (61) was formed during 8 days [119].
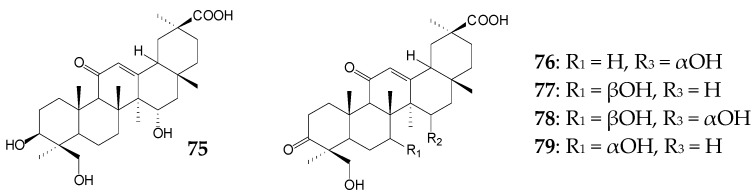
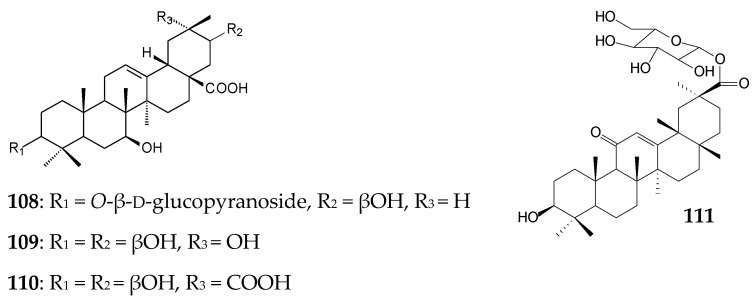
UA biotransformation processes include oxidation and lactone formation reactions. Thus, Syncephalastrum racemosum CGMCC 3.2500 transformed approximately 0.1 g/L UA to 7β,21β-dihydroxy-UA (80, 12.9%), 1β,21β-dihydroxy-UA (81, 3.9%), 1β-hydroxy-21-oxo-UA (82, 12.1%), 3β,21β-dihydroxy-urs-11-en-28,13-olide (83, 3.4%), and 3β,7β,21β-trihydroxy-urs-11-en-28,13-olide (84, 2.9%) within 10 days [13]. Similar bioconversion processes have been previously demonstrated using another strain S. racemosum AS 3.264 [120]. UA derivatives with a rare lactone moiety were shown to exhibit moderate inhibitory activity against HCV [13].
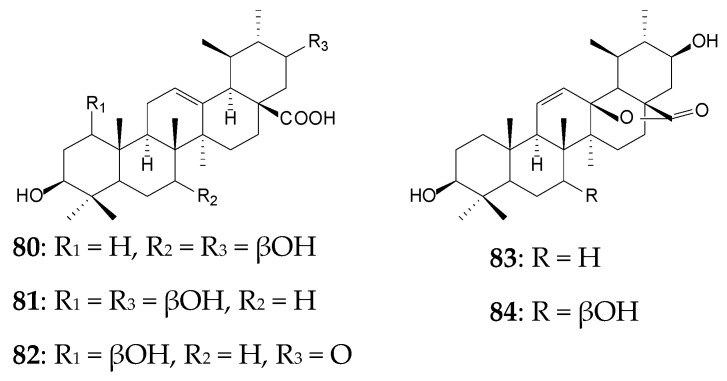
For 20 days, Gliocladium roseum CGMCC 3.3657 catalyzed oxidative transformation of the A ring of UA (0.1 g/L) by Baeyer–Villiger-type reaction and C21 oxidation to form 21β-hydroxy-3-oxo-urs-12-en-3,4-olide-28-oic acid (85, 8.0%), 3,21-dioxo-urs-12-en-3,4-olide-28-oic acid (86, 6.25%), 21β-hydroxy-3,4-seco-ursane-4(23),12-dien-3,28-dioic acid (87, 1.5%), and 21-oxo-3,4-seco-ursane-4(23),12-dien-3,28-dioic acid (88, 1.0%). Derivatives 86 and 88 containing a 21-oxo group showed the most pronounced anti-HCV activity [121].
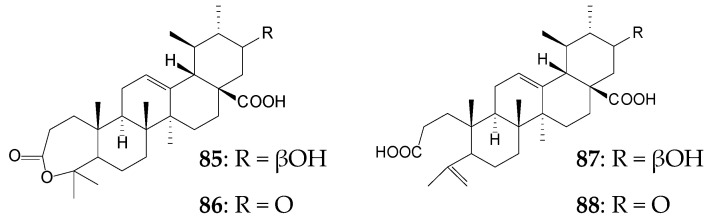
When Mucor spinosus AS 3.3450 was used to bioconvert UA, three metabolites, 7β-hydroxy-UA 28-ethanone (89, 5.04%), 7β,21β-dihydroxy-UA (81, 1.64%), and 21β-hydroxy-urs-12-en-28-oic acid 3-O-β-D-glucopyranoside (90, 2.13%), were formed within 96 h. Compound 89 was shown to exhibit pronounced (higher than that of UA) cytotoxic activity against HeLa, K562, and KB tumor cell lines [122].
5.2. Bacterial Transformation
The literature describes a few cases of pentacyclic triterpenoid bioconversion using gram-positive bacteria of the genera Bacillus, Nocardia, and Streptomyces and accompanied by the formation of C1, C2, C7, C11, C21, C24, or C29 hydroxylated derivatives, derivatives with a methyl ester group at C28; oxogroup at C3; additional carboxyl groups at C29 or C30; glucopyranoside groups at C3, C28, or C30; lactone group at C28/C13; and derivatives with a fragmented A ring. In biotransformation experiments, the compounds are usually used in concentrations ranging from 0.04 g/L to 0.3 g/L, and the yield of derivatives ranges from 5.0% to 60.0%. The duration of bioconversion is 3 to 5 days; only in the case of using Nocardia, it reached 13 days (Table 3).
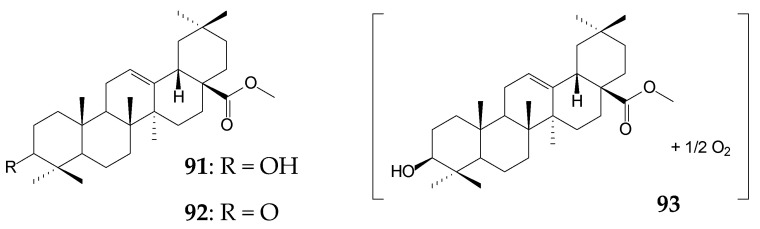
Actinobaceria of the genus Nocardia were capable of selective methylation of the C28-carboxylic group of pentacyclic triterpenoids [123]. The use of resting or immobilized cells of N. iowensis DSM 45197 as biocatalysts of the OA (approximately 0.3 g/L) transformation process for 13 days resulted in the formation of methyl OA (91) as the main bioconversion product (more than 60.0%), small amounts (≤5.0%) of methyl 3-oxo-olean-12-en-28-oat (92), and metabolite 93 unidentified by the authors [16]. 3-oxo-OA (92) was shown to have pronounced antimelanoma [124], antileishmanial, and antitrypanosomal effects [125]. Despite numerous successful examples to increase the efficiency of the biotransformation process by immobilizing microbial cells [126], the use of fixed Nocardia cells in alginate carriers led to a decrease in their catalytic activity, as confirmed by a 10-fold decrease in the formation of compound 91 and only a short-term occurrence of compound 93 in the culture medium [16]. The ability of Nocardia sp. to transform UA by methylation, by C3 oxidation, and by formation of the enone moiety in the A ring was previously shown. It was noted that the biotransformation process did not depend on the composition of the culture medium used, while the temperature increase (from 28 °C to 36 °C) for actinobacteria cultivation contributed to a 2-fold increase in the reaction rate [127].
The bacterial culture of Streptomyces griseus ATCC 13273 catalyzed hydroxylation and site-selective oxidation of the C29 methyl group of OA (0.04 g/L) to the carboxyl group within 5 days to form 3β-hydroxy-olean-12-ene-28,29-dioic acid (94, 21.9%), 3β,24-dihydroxy-olean-12-ene-28,29-dioic acid (95, 32.7%), and 3β,21β,24-trihydroxy-olean-12-ene-28,29-dioic acid (96, 5.9%). Hydroxylation at C21 was shown to increase the anti-inflammatory activity of OA derivatives [128]. Using the same strain, biotransformation of OA (approximately 0.05 g/L) with the formation of derivatives 94 and 96 was previously described by Y. Zhu et al. [129].
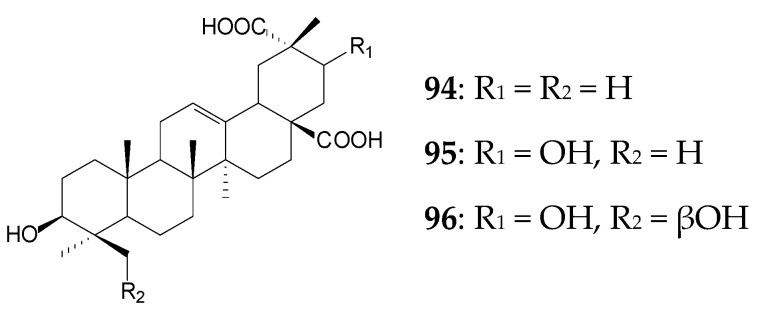
In addition to OA, S. griseus ATCC 13273 cells can also transform UA (0.04 g/L) by catalyzing site-selective oxidation of the C30 methyl group to the carboxyl one and C24-hydroxylation within 3 days to produce 3β-hydroxy-urs-12-ene-28,30-dioic acid (97) and 3β,24-dihydroxy-urs-12-ene-28,30-dioic acid (98), with the product yield exceeding 30.0%. Transformation of UA derivatives 3-oxo-UK (99) and 2α-hydroxy-UA (20, corosolic acid) at a concentration of 0.04 g/L by this actinobacterial strain also occurred by selective C30-oxidation and hydroxylation, that led to a mixture of 3-oxo-urs-12-ene-28,30-dioic acid (100, 24.1%) and 24-hydroxy-3-oxo-urs-12-ene-28,30-dioic acid (101, 45.9%) in the former case and a mixture of 2α,3β-dihydroxy-urs-12-ene-28,30-dioic acid (102, 29.0%) and 2α,3β,24-trihydroxy-urs-12-ene-28,30-dioic acid (103, 15.9%) in the latter case [17].
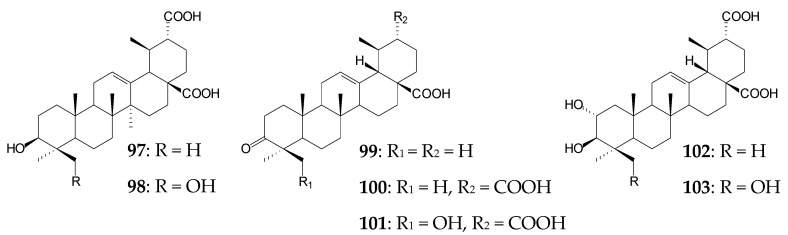
A gram-positive Bacillus megaterium CGMCC 1.1741 was able to transform UA (0.2 g/L), generating the main derivative 1β,11α-dihydroxy-UA (104, 26.87%) and minor derivatives (5.03–13.50%) 3-oxo-urs-12-en-28-oic acid (99), 1β,11α-dihydroxy-3-oxo-urs-12-en-28-oic acid (105), 1β-hydroxy-3-oxo-urs-12-en-28,13-olide (106), and 1β,11α-dihydroxy-3-oxo-urs-12-en -28-O-β-D-glucopyranoside (107) over 4 days. Derivatives 105 and 106 were shown to effectively inhibit LPS-induced NO release in RAW 264.7 cells (IC50 1.71 µmol and 1.24 µmol, respectively) [18], and derivative 99 was shown to inhibit cathepsin L-like rCPB2.8 protease of Leishmania mexicana [130].
Xu et al. (2020) have recently used the tandem biotransformation of oleanane-type pentacyclic triterpenoids using the fungal strain Rhizopus chinensis CICC 40335 and bacterial strains Bacillus subtilis ATCC 6633 and Streptomyces griseus ATCC 13273 [34]. The primary transformation of OA (0.2 g/L) using R. chinensis CICC 40335 occurred within 4 days by the formation of 7β,21β-dihydroxy-OA (54, 53.75%) previously obtained using the fungal strains Mucor rouxii NRRL 1894 [131] and Circinella muscae AS 3.2695 [15]. Further 4-day biotransformation of the obtained compound (54) led to the formation of 7β,21β-dihydroxy-olean-12-en-28-oic acid 3-O-β-D-glucopyranoside (108, 46.5%) using B. subtilis ATCC 6633 cells and a mixture of 7β,21β,29-trihydroxy-OA (109, 26.0%) and 3β,7β,21β-trihydroxy-olean-12-ene-28,29-dioic acid (110, 15.0%) using S. griseus ATCC 13273 cells [34].
Bioconversion of GA (0.2 g/L) using R. chinensis CICC 40335 occurred by selective oxidation with the formation of 7β-hydroxy-GA (61, 77.5%) on day 4 [34]. Note that the C7-hydroxylation process is typical for many cultures, for example, C. muscae AS 3.2695, Rhizopus arrhizus AS 3.2893 [115], and representatives of the genus Cunninghamella [116,119]. Further addition of GA or compound 61 in the culture medium of B. subtilis ATCC 6633 led to the formation of 30-O-β-d-glucopyranoside derivatives (111 (27.5%) and 68 (44.0%), respectively) previously obtained by B. Fan et al. [115]. Assessment of the neuroprotective potential of the obtained OA and GA derivatives revealed that glycosylation significantly contributed to a decrease in the neuroprotective activity of compounds while carboxylation led to a significant increase in the neuroprotective effect of OA derivatives [34].
6. Conclusions
Triterpenoids are secondary metabolites of plants, fungi, marine invertebrates, and algae that are formed during cyclization of an acyclic triterpene squalene [132,133,134,135,136,137,138]. According to the number of cycles, triterpenes and triterpenoids are divided into several groups; the most numerous are pentacyclic triterpenic derivatives [139]. In nature, this group is most widely represented by compounds of oleanane (OA and GA) and ursane (UA) types, which in large quantities can accumulate in various parts of higher plants [39,41,46]. In addition, the biosynthesis of these compounds can be carried out in microbial cells able to catalyze the MVA pathway and to be genetically modified using plant genes [64,68,69,70]. The main difficulties of microbial biosynthesis are generally considered to be complexity and long duration of processes of searching for terpenoid synthesis genes of plants, their isolation, and the preparation of genetically modified microorganisms. The rapid development of bioinformatics methods, sequencing techniques, and de novo DNA synthesis significantly simplified the abovementioned processes and gave a new impetus to research in this field [140]. With the close cooperation of biochemists, microbiologists, and genetic scientists, microbial biosynthesis can become a promising technology for obtaining valuable pentacyclic triterpenoids. The compounds discussed in the review exhibit antitumor, antiviral, hepatoprotective, neuroprotective, and other activities [5,7,42,60,74,82,83]. Despite the wide range of known biological properties, the use of pentacyclic triterpenoids in pharmacology and medicine is limited because of their high hydrophobicity. The solution to this problem might be the synthesis of triterpenic derivatives with increased bioactivity, solubility, and bioavailability [5,82,93,94,141].
Studies of the possibility of obtaining new OA, GA, and UA derivatives by directed biotransformations should be considered a promising area. Over 20 examples of biotransformations of these compounds using fungal and bacterial cultures most often catalyzing hydroxylation have been described since 2013. Less frequently, the literature describes processes of deeper oxidation of triterpenoids as well as their glycosylation, esterification, acetylation, or carboxylation. Biocatalytic formation of triterpenic lactones or their derivatives with fragmented C–C bond was reported only in a few cases using UA [13,121]. In the biotransformation processes employing fungi, the degree of triterpenic acid conversion usually ranges from 2.6% to 77.5%, with an initial concentration of 0.02 g/L to 1.0 g/L, whereas in bacterial transformations, the degree of conversion reaches 27.5‒70.0%, with an initial concentration of 0.04‒0.3 g/L. When analyzed, the data showed that the biotransformation of oleanane and ursane pentacyclic triterpenoids led to derivatives with antioxidant, anti-inflammatory, antiviral, antitumor, antiparasitic, antimicrobial, neuroprotective, and hepatoprotective properties (Table 3). Provided more active development as an interdisciplinary tool, this method of obtaining biologically active compounds and their intermediates seems to be a promising strategy to design new medicinal agents against cancer and neurodegenerative diseases as well as potent antibacterial drugs against antibiotic-resistant pathogenic strains of microorganisms. By combining methods of microbial synthesis of native pentacyclic triterpenoids and their subsequent microbial transformations into bioavailable compounds, the industrial microbiology could provide a cycle of production of valuable biologically active substances. However, it should be noted that the described microbial catalysts have significant drawbacks. Fungi usually demonstrate mycelial growth type and form spores and mycotoxins, whereas few bacterial catalysts described are mainly represented by species, with their individual strains being pathogens. In this context, it is essential to conduct further in-depth studies of the processes of biological transformation of pentacyclic triterpenoids and to search for new nonpathogenic bacterial strains able to carry out highly effective synthesis of triterpenic derivatives with pronounced biological activities.
Author Contributions
All authors made equal contributions to preparing this manuscript. Conceptualization, I.B.I., V.V.G., and N.A.L.; writing—original draft preparation, V.V.G. and N.A.L.; writing—review and editing, I.B.I., V.V.G., and N.A.L.; funding acquisition, I.B.I. and V.V.G. All authors have read and agreed to the published version of the manuscript.
Funding
The work was funded by the Russian Foundation for Basic Research (grant No. 20-34-90104), by the Russian Science Foundation (grant number 18-14-00140), and by the Russian Federation Ministry of Education and Science (State Assignments AAAA-A19-119112290008-4, AAAA-A18-118030790037-7, and AAAA-A19-119112290010-7).
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Calixto J.B. The role of natural products in modern drug discovery. An. Acad. Bras. Ciênc. 2019;91:e20190105. doi: 10.1590/0001-3765201920190105. [DOI] [PubMed] [Google Scholar]
- 2.Kumar D., Dubey K.K. Hybrid approach for transformation for betulin (an anti-HIV molecule) In: Gupta P., Pandey A., editors. New and Future Developments in Microbial Biotechnology and Bioengineering. Elsevier BV; Amsterdam, The Netherlands: 2019. pp. 193–203. [DOI] [Google Scholar]
- 3.Duric K., Kovac-Besovic E., Niksic H., Sofic E. Antibacterial activity of methanolic extracts, decoction and isolated triterpene products from different parts of birch, Betula pendula, Roth. J. Plant Stud. 2013;2:61–70. doi: 10.5539/jps.v2n2p61. [DOI] [Google Scholar]
- 4.Song N.Y., Cho J.G., Im D., Lee D.Y., Wu Q., Seo W.D., Kang H.C., Lee Y.H., Baek N.I. Triterpenoids from Fragaria ananassa calyx and their inhibitory effects on melanogenesis in B16-F10 mouse melanoma cells. Nat. Prod. Res. 2013;27:2219–2223. doi: 10.1080/14786419.2013.805330. [DOI] [PubMed] [Google Scholar]
- 5.Zou L.-W., Dou T.-Y., Wang P., Lei W., Weng Z.-M., Hou J., Wang D.-D., Fan Y.-M., Zhang W.-D., Ge G., et al. Structure-activity relationships of pentacyclic triterpenoids as potent and selective inhibitors against human carboxylesterase 1. Front. Pharmacol. 2017;8:435. doi: 10.3389/fphar.2017.00435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zhang J., Liu F., Zhang X. Inhibition of proliferation of SGC7901 and BGC823 human gastric cancer cells by ursolic acid occurs through a caspase-dependent apoptotic pathway. Med Sci. Monit. 2019;25:6846–6854. doi: 10.12659/MSM.916740. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 7.Feng J., Yi X., Huang W., Wang Y., He X. Novel triterpenoids and glycosides from durian exert pronounced anti-inflammatory activities. Food Chem. 2018;241:215–221. doi: 10.1016/j.foodchem.2017.08.097. [DOI] [PubMed] [Google Scholar]
- 8.Alho D.P.S., Salvador J.A.R., Cascante M., Marin S. Synthesis and antiproliferative activity of novel heterocyclic glycyrrhetinic acid derivatives. Molecules. 2019;24:766. doi: 10.3390/molecules24040766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chouaïb K., Hichri F., Nguir A., Daami-Remadi M., Elie N., Touboul D., Ben Jannet H., Hamza M.A. Semi-synthesis of new antimicrobial esters from the natural oleanolic and maslinic acids. Food Chem. 2015;183:8–17. doi: 10.1016/j.foodchem.2015.03.018. [DOI] [PubMed] [Google Scholar]
- 10.Grishko V.V., Galaiko N.V., Tolmacheva I.A., Kucherov I.I., Eremin V.F., Boreko E.I., Savinova O.V., Slepukhin P.A. Functionalization, cyclization and antiviral activity of A-secotriterpenoids. Eur. J. Med. Chem. 2014;83:601–608. doi: 10.1016/j.ejmech.2013.12.058. [DOI] [PubMed] [Google Scholar]
- 11.Wu P.P., Zhang B.J., Cui X.P., Yang Y., Jiang Z.Y., Zhou Z.H., Zhong Y.Y., Mai Y.Y., Ouyang Z., Chen H.S., et al. Synthesis and biological evaluation of novel ursolic acid analogues as potential α-glucosidase inhibitors. Sci. Rep. 2017;7:45578. doi: 10.1038/srep45578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Shah S.A.A., Tan H.L., Sultan S., Faridz M.A.B.M., Shah M.A.B.M., Nurfazilah S., Hussain M. Microbial-catalyzed biotransformation of multifunctional triterpenoids derived from phytonutrients. Int. J. Mol. Sci. 2014;15:12027–12060. doi: 10.3390/ijms150712027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fu S.B., Yang J.-S., Cui J.L., Sun D.-A. Biotransformation of ursolic acid by Syncephalastrum racemosum CGMCC 3.2500 and anti-HCV activity. Fitoterapia. 2013;86:123–128. doi: 10.1016/j.fitote.2013.02.007. [DOI] [PubMed] [Google Scholar]
- 14.Ma Y., Liu J.M., Chen R., An X.Q., Dai J.G. Microbial transformation of glycyrrhetinic acid and potent neural anti-inflammatory activity of the metabolites. Chin. Chem. Lett. 2017;28:1200–1204. doi: 10.1016/j.cclet.2017.02.019. [DOI] [Google Scholar]
- 15.Yan S., Lin H., Huang H.-L., Yang M., Xu B., Chen G.-T. Microbial hydroxylation and glycosidation of oleanolic acid by Circinella muscae and their anti-inflammatory activities. Nat. Prod. Res. 2018;33:1849–1855. doi: 10.1080/14786419.2018.1477150. [DOI] [PubMed] [Google Scholar]
- 16.Ludwig B., Geib D., Haas C., Steingroewer J., Bley T., Muffler K., Ulber R. Whole-cell biotransformation of oleanolic acid by free and immobilized cells of Nocardia iowensis: Characterization of new metabolites. Eng. Life Sci. 2015;15:108–115. doi: 10.1002/elsc.201400121. [DOI] [Google Scholar]
- 17.Xu S.H., Zhang C., Wang W.W., Yu B.Y., Zhang J. Site-selective biotransformation of ursane triterpenes by Streptomyces griseus ATCC 13273. RSC Adv. 2017;7:20754–20759. doi: 10.1039/C7RA01811H. [DOI] [Google Scholar]
- 18.Zhang C., Xu S.-H., Ma B.-L., Wang W.-W., Yu B., Zhang J. New derivatives of ursolic acid through the biotransformation by Bacillus megaterium CGMCC 1.1741 as inhibitors on nitric oxide production. Bioorg. Med. Chem. Lett. 2017;27:2575–2578. doi: 10.1016/j.bmcl.2017.03.076. [DOI] [PubMed] [Google Scholar]
- 19.Grishko V.V., Tarasova E.V., Ivshina I.B. Biotransformation of betulin to betulone by growing and resting cells of the actinobacterium Rhodococcus rhodochrous IEGM 66. Process. Biochem. 2013;48:1640–1644. doi: 10.1016/j.procbio.2013.08.012. [DOI] [Google Scholar]
- 20.Tarasova E.V., Grishko V.V., Ivshina I.B. Cell adaptations of Rhodococcus rhodochrous IEGM 66 to betulin biotransformation. Process. Biochem. 2017;52:1–9. doi: 10.1016/j.procbio.2016.10.003. [DOI] [Google Scholar]
- 21.Laskin A.I., Grabowich P., Fried J., Meyers C.D.L. Transformations of eburicoic acid. V. Cleavage of ring A by the fungus Glomerella fusarioides. J. Med. Chem. 1964;7:406–409. doi: 10.1021/jm00334a003. [DOI] [PubMed] [Google Scholar]
- 22.Canonica L., Jommi G., Pagnoni U.M., Pelizzoni F., Ranzi B.M., Scolastico C. Microbiological oxidation of triterpenoids. I. 7β-Hydroxyglycyrrhetic acid. Gazz. Chim. Ital. 1966;96:820–831. [Google Scholar]
- 23.Canonica L., Ferrari M., Jommi G., Pagnoni U.M., Pelizzoni F., Ranzi B.M., Maroni S., Nencini G., Salvatori T. Microbiological oxidation of triterpenoids. II. 15α-Hydroxyglycyrrhetic and 7β,15α-dihydroxyglycyrrhetic acids. Gazz. Chim. Ital. 1967;97:1032–1051. [Google Scholar]
- 24.Hikino H., Nabetani S., Takemoto T. Microbial transformation of oleanolic acid. Yakugaku Zasshi. 1969;89:809–813. doi: 10.1248/yakushi1947.89.6_809. [DOI] [PubMed] [Google Scholar]
- 25.Ferrari M., Pagnoni U.M., Pelizzoni F., Ranzi B.M., Salvatori T. Microbiological oxidation of triterpenoids. III. Behavior of 18α-glycyrrhetic, liquiritic, and 18α-liquiritic acids. Gazz. Chim. Ital. 1969;99:848–862. [Google Scholar]
- 26.Sakano K.-I., Ohshima M. Structures of conversion products formed from 18β-glycyrrhetinic acid by Streptomyces sp. G-20. Agric. Biol. Chem. 1986;50:763–766. doi: 10.1080/00021369.1986.10867460. [DOI] [Google Scholar]
- 27.Sakano K.-I., Ohshima M. Microbial conversion of 18β-glycyrrhetinic acid and 22α-hydroxy-18β-glycyrrhetinic acid by Chainia antibiotic. Agric. Biol. Chem. 1986;50:1239–1245. doi: 10.1080/00021369.1986.10867532. [DOI] [Google Scholar]
- 28.Collins D.O., Ruddock P.L.D., De Grasse J.C., Reynolds W.F., Reese P.B. Microbial transformation of cadina-4,10(15)-dien-3-one, aromadendr-1(10)-en-9-one and methyl ursolate by Mucor plumbeus ATCC 4740. Phytochemistry. 2002;59:479–488. doi: 10.1016/S0031-9422(01)00486-1. [DOI] [PubMed] [Google Scholar]
- 29.Cheng Z., Yu B.-Y., Cordell G.A., Qiu S.X. Biotransformation of quinovic acid glycosides by microbes: Direct conversion of the ursane to the oleanane triterpene skeleton by Nocardia sp. NRRL 5646. Org. Lett. 2004;6:3163–3165. doi: 10.1021/ol048787b. [DOI] [PubMed] [Google Scholar]
- 30.Parra A., Rivas F., Garcia-Granados A., Martinez A. Microbial transformation of triterpenoids. Mini Rev. Org. Chem. 2009;18:307–320. doi: 10.2174/157019309789371569. [DOI] [Google Scholar]
- 31.Bhatti H.N., Khera R.A. Biotransformations of diterpenoids and triterpenoids: A review. J. Asian Nat. Prod. Res. 2013;16:70–104. doi: 10.1080/10286020.2013.846908. [DOI] [PubMed] [Google Scholar]
- 32.Mutafova B., Fernandes P., Mutafov S., Berkov S., Pavlov A. Microbial transformations of plant secondary metabolites. In: Pavlov A., Bley T., editors. Bioprocessing of Plant In Vitro Systems. Springer Science and Business Media LLC; New York, NY, USA: 2018. pp. 85–124. [DOI] [Google Scholar]
- 33.Azerad R. Microbial transformations of pentacyclic triterpenes. In: Patel R.N., editor. Green Biocatalysis. John Wiley & Sons, Inc.; Hoboken, NJ, USA: 2016. pp. 675–714. [DOI] [Google Scholar]
- 34.Xu S.H., Chen H.L., Fan Y., Xu W., Zhang J. Application of tandem biotransformation for biosynthesis of new pentacyclic triterpenoid derivatives with neuroprotective effect. Bioorg. Med. Chem. Lett. 2020;30:126947. doi: 10.1016/j.bmcl.2019.126947. [DOI] [PubMed] [Google Scholar]
- 35.Grishko V.V., Nogovitsina Y.M., Ivshina I.B. Bacterial transformation of terpenoids. Russ. Chem. Rev. 2014;83:323–342. doi: 10.1070/RC2014v083n04ABEH004396. [DOI] [Google Scholar]
- 36.Hill R.A., Connolly J.D. Triterpenoids. Nat. Prod. Rep. 2012;29:780–818. doi: 10.1039/c2np20027a. [DOI] [PubMed] [Google Scholar]
- 37.Thimmappa R., Geisler K., Louveau T., O’Maille P., Osbourn A. Triterpene Biosynthesis in plants. Annu. Rev. Plant Biol. 2014;65:225–257. doi: 10.1146/annurev-arplant-050312-120229. [DOI] [PubMed] [Google Scholar]
- 38.Wu H., Li G., Liu S., Liu D., Chen G., Hu N., Suo Y., You J. Simultaneous determination of six triterpenic acids in some Chinese medicinal herbs using ultrasound-assisted dispersive liquid–liquid microextraction and high-performance liquid chromatography with fluorescence detection. J. Pharm. Biomed. Anal. 2015;107:98–107. doi: 10.1016/j.jpba.2014.10.031. [DOI] [PubMed] [Google Scholar]
- 39.Moldoveanu S.C., Scott W.A. Analysis of four pentacyclic triterpenoid acids in several bioactive botanicals with gas and liquid chromatography and mass spectrometry detection. J. Sep. Sci. 2015;39:324–332. doi: 10.1002/jssc.201501041. [DOI] [PubMed] [Google Scholar]
- 40.Peragón J. Time course of pentacyclic triterpenoids from fruits and leaves of olive tree (Olea europaea L.) cv. Picual and cv. Cornezuelo during ripening. J. Agric. Food Chem. 2013;61:6671–6678. doi: 10.1021/jf401495t. [DOI] [PubMed] [Google Scholar]
- 41.Kalani K., Agarwal J., Alam S., Khan F.F., Pal A., Srivastava S.K. In silico and in vivo anti-malarial studies of 18β-glycyrrhetinic acid from Glycyrrhiza glabra. PLoS ONE. 2013;8:e74761. doi: 10.1371/journal.pone.0074761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chen H.-J., Kang S.-P., Lee I.-J., Lin Y.-L. Glycyrrhetinic acid suppressed NF-κB activation in TNF-α-induced hepatocytes. J. Agric. Food Chem. 2014;62:618–625. doi: 10.1021/jf405352g. [DOI] [PubMed] [Google Scholar]
- 43.Li J., Lee Y.S., Choi J.S., Sung H.Y., Kim J.K., Lim S.S., Kang Y.H. Roasted licorice extracts dampen high glucose-induced mesangial hyperplasia and matrix deposition through blocking Akt activation and TGF-β signaling. Phytomedicine. 2010;17:800–810. doi: 10.1016/j.phymed.2010.02.004. [DOI] [PubMed] [Google Scholar]
- 44.Ko B.S., Jang J.S., Hong S.M., Sung S.R., Lee J.E., Lee M.Y., Jeon W.K., Park S. Changes in components, glycyrrhizin and glycyrrhetinic acid, in raw Glycyrrhiza uralensis Fisch, modify insulin sensitizing and insulinotropic actions. Biosci. Biotechnol. Biochem. 2007;71:1452–1461. doi: 10.1271/bbb.60533. [DOI] [PubMed] [Google Scholar]
- 45.Woźniak Ł., Skąpska S., Marszałek K. Ursolic acid—A pentacyclic triterpenoid with a wide spectrum of pharmacological activities. Molecules. 2015;20:20614–20641. doi: 10.3390/molecules201119721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Guinda A., Rada M., Delgado T., Gutiérrez-Adánez P., Castellano J.M. Pentacyclic triterpenoids from olive fruit and leaf. J. Agric. Food Chem. 2010;58:9685–9691. doi: 10.1021/jf102039t. [DOI] [PubMed] [Google Scholar]
- 47.Romero C., García A., Medina E., Ruíz-Méndez M.V., De Castro A., Brenes M. Triterpenic acids in table olives. Food Chem. 2010;118:670–674. doi: 10.1016/j.foodchem.2009.05.037. [DOI] [Google Scholar]
- 48.Pensec F., Pączkowski C., Grabarczyk M., Woźniak A., Bénard-Gellon M., Bertsch C., Chong J., Szakiel A. Changes in the triterpenoid content of cuticular waxes during fruit ripening of eight grape (Vitis vinifera) cultivars grown in the upper rhine valley. J. Agric. Food Chem. 2014;62:7998–8007. doi: 10.1021/jf502033s. [DOI] [PubMed] [Google Scholar]
- 49.Wang C.M., Chen H.T., Li T.C., Weng J.H., Jhan Y.L., Lin S.X., Chou C.H. The role of pentacyclic triterpenoids in the allelopathic effects of Alstonia scholaris. J. Chem. Ecol. 2014;40:90–98. doi: 10.1007/s10886-013-0376-y. [DOI] [PubMed] [Google Scholar]
- 50.Alvarado H.L., Abrego G., Garduño-Ramirez M.L., Clares B., García M.L., Calpena A.C. Development and validation of a high-performance liquid chromatography method for the quantification of ursolic/oleanic acids mixture isolated from Plumeria obtusa. J. Chromatogr. B. 2015;983:111–116. doi: 10.1016/j.jchromb.2015.01.009. [DOI] [PubMed] [Google Scholar]
- 51.Hương N.T.N., Hung T., Dẹp T.T. A study on morphogenesis of roots of Panax stipuleanatus HT Tsai et KM Feng in vitro and preliminary determination of oleanolic acid in roots. Vietnam. J. Biotechnol. 2016;14:49–54. doi: 10.15625/1811-4989/14/1/9291. [DOI] [Google Scholar]
- 52.Yamamoto E.S., Campos B.L.S., Laurenti M.D., Lago J.H.G., Grecco S.D.S., Corbett C.E.P., Passero L.F.D. Treatment with triterpenic fraction purified from Baccharis uncinella leaves inhibits Leishmania (Leishmania) amazonensis spreading and improves Th1 immune response in infected mice. Parasitol. Res. 2014;113:333–339. doi: 10.1007/s00436-013-3659-x. [DOI] [PubMed] [Google Scholar]
- 53.Falev D.I., Kosyakov D.S., Ul’Yanovskii N.V., Ovchinnikov D.V. Rapid simultaneous determination of pentacyclic triterpenoids by mixed-mode liquid chromatography–tandem mass spectrometry. J. Chromatogr. A. 2020;1609:460458. doi: 10.1016/j.chroma.2019.460458. [DOI] [PubMed] [Google Scholar]
- 54.Jang S.-E., Jeong J.-J., Hyam S.R., Han M.J., Kim D.-H. Ursolic acid isolated from the seed of Cornus officinalis ameliorates colitis in mice by inhibiting the binding of lipopolysaccharide to toll-like receptor 4 on macrophages. J. Agric. Food Chem. 2014;62:9711–9721. doi: 10.1021/jf501487v. [DOI] [PubMed] [Google Scholar]
- 55.Zhang S., Sun Y., Sun Z., Wang X., You J., Suo Y. Determination of triterpenic acids in fruits by a novel high performance liquid chromatography method with high sensitivity and specificity. Food Chem. 2014;146:264–269. doi: 10.1016/j.foodchem.2013.09.056. [DOI] [PubMed] [Google Scholar]
- 56.Fu Q., Zhang L., Cheng N., Jia M., Zhang Y. Extraction optimization of oleanolic and ursolic acids from pomegranate (Punica granatum L.) flowers. Food Bioprod. Process. 2014;92:321–327. doi: 10.1016/j.fbp.2012.12.006. [DOI] [Google Scholar]
- 57.Oancea I.A., Van Staden J., Koos F., Oancea E., Ungureanu E.-M. Electrochemical detection of ursolic acid from spruce (Picea abies) essential oils using modified amperometric microsensors. Anal. Lett. 2019;52:2214–2226. doi: 10.1080/00032719.2019.1605373. [DOI] [Google Scholar]
- 58.Choudhary A., Mittal A.K., Radhika M., Tripathy D., Chatterjee A., Banerjee U.C., Singh I.P. Two new stereoisomeric antioxidant triterpenes from Potentilla fulgens. Fitoterapia. 2013;91:290–297. doi: 10.1016/j.fitote.2013.09.008. [DOI] [PubMed] [Google Scholar]
- 59.Li H.H., Su M.H., Yao D.H., Zeng B.Y., Chang Q., Wang W., Xu J. Anti-hepatocellular carcinoma activity of tormentic acid derived from suspension cells of Eriobotrya japonica (Thunb.) Lindl. Plant Cell Tissue Organ Cult. 2017;130:427–433. doi: 10.1007/s11240-017-1221-8. [DOI] [Google Scholar]
- 60.Wang Z.W., Wang J., Luo J., Kong L. α-Glucosidase inhibitory triterpenoids from the stem barks of Uncaria laevigata. Fitoterapia. 2013;90:30–37. doi: 10.1016/j.fitote.2013.07.005. [DOI] [PubMed] [Google Scholar]
- 61.Wu X.D., He J., Li X.Y., Dong L.B., Gong X., Gao X., Song L.D., Li Y., Peng L.Y., Zhao Q. Triterpenoids and steroids with cytotoxic activity from Emmenopterys henryi. Planta Med. 2013;79:1356–1361. doi: 10.1055/s-0033-1350645. [DOI] [PubMed] [Google Scholar]
- 62.Zhu M., Wang C., Sun W., Zhou A., Wang Y., Zhang G., Zhou X., Huo Y., Li C. Boosting 11-oxo-β-amyrin and glycyrrhetinic acid synthesis in Saccharomyces cerevisiae via pairing novel oxidation and reduction system from legume plants. Metab. Eng. 2018;45:43–50. doi: 10.1016/j.ymben.2017.11.009. [DOI] [PubMed] [Google Scholar]
- 63.Krivoruchko A., Nielsen J. Production of natural products through metabolic engineering of Saccharomyces cerevisiae. Curr. Opin. Biotechnol. 2015;35:7–15. doi: 10.1016/j.copbio.2014.12.004. [DOI] [PubMed] [Google Scholar]
- 64.Lu C., Zhang C., Zhao F., Li D., Lu W. Biosynthesis of ursolic acid and oleanolic acid in Saccharomyces cerevisiae. AIChE J. 2018;64:3794–3802. doi: 10.1002/aic.16370. [DOI] [Google Scholar]
- 65.Zhao Y., Fan J., Wang C., Feng X., Li C. Enhancing oleanolic acid production in engineered Saccharomyces cerevisiae. Bioresour. Technol. 2018;257:339–343. doi: 10.1016/j.biortech.2018.02.096. [DOI] [PubMed] [Google Scholar]
- 66.Dai Z., Wang B., Liu Y., Shi M., Wang D., Zhang X., Liu T., Huang L., Zhang X. Producing aglycons of ginsenosides in bakers’ yeast. Sci. Rep. 2014;4:3698. doi: 10.1038/srep03698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Fukushima E.O., Seki H., Esawai S., Suzuki M., Ohyama K., Saito K., Muranaka T. Combinatorial biosynthesis of legume natural and rare triterpenoids in engineered yeast. Plant Cell Physiol. 2013;54:740–749. doi: 10.1093/pcp/pct015. [DOI] [PubMed] [Google Scholar]
- 68.Dale M.P., Moses T., Johnston E.J., Rosser S.J. A systematic comparison of triterpenoid biosynthetic enzymes for the production of oleanolic acid in Saccharomyces cerevisiae. PLoS ONE. 2020;15:e0231980. doi: 10.1371/journal.pone.0231980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Li D., Wu Y., Wei P., Gao X., Li M., Zhang C., Zhou Z., Lu W. Metabolic engineering of Yarrowia lipolytica for heterologous oleanolic acid production. Chem. Eng. Sci. 2020;218:115529. doi: 10.1016/j.ces.2020.115529. [DOI] [Google Scholar]
- 70.Dai Z., Liu Y., Sun Z., Wang D., Qu G., Ma X., Fan F., Zhang L., Li S., Zhang X. Identification of a novel cytochrome P450 enzyme that catalyzes the C-2α hydroxylation of pentacyclic triterpenoids and its application in yeast cell factories. Metab. Eng. 2019;51:70–78. doi: 10.1016/j.ymben.2018.10.001. [DOI] [PubMed] [Google Scholar]
- 71.Moses T., Pollier J., Almagro L., Buyst D., Van Montagu M., Pedreño M.A., Martins J.C., Thevelein J.M., Goossens A. Combinatorial biosynthesis of sapogenins and saponins in Saccharomyces cerevisiae using a C-16 hydroxylase from Bupleurum falcatum. Proc. Natl. Acad. Sci. USA. 2014;111:1634–1639. doi: 10.1073/pnas.1323369111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Moses T., Thevelein J.M., Goossens A., Pollier J. Comparative analysis of CYP93E proteins for improved microbial synthesis of plant triterpenoids. Phytochemistry. 2014;108:47–56. doi: 10.1016/j.phytochem.2014.10.002. [DOI] [PubMed] [Google Scholar]
- 73.Zhang G., Cao Q., Liu J., Liu B., Li J., Li C. Refactoring β-amyrin synthesis in Saccharomyces cerevisiae. AIChE J. 2015;61:3172–3179. doi: 10.1002/aic.14950. [DOI] [Google Scholar]
- 74.Kannan S., Sathasivam G., Marudhamuthu M. Decrease of growth, biofilm and secreted virulence in opportunistic nosocomial Pseudomonas aeruginosa ATCC 25619 by glycyrrhetinic acid. Microb. Pathog. 2019;126:332–342. doi: 10.1016/j.micpath.2018.11.026. [DOI] [PubMed] [Google Scholar]
- 75.Jyoti A., Zerin T., Kim T.-H., Hwang T.-S., Jang W.S., Nam K.-W., Song H.-Y. In vitro effect of ursolic acid on the inhibition of Mycobacterium tuberculosis and its cell wall mycolic acid. Pulm. Pharmacol. Ther. 2015;33:17–24. doi: 10.1016/j.pupt.2015.05.005. [DOI] [PubMed] [Google Scholar]
- 76.Tohmé M., Giménez M., Peralta A., Colombo M., Delgui L.R. Ursolic acid: A novel antiviral compound inhibiting rotavirus infection in vitro. Int. J. Antimicrob. Agents. 2019;54:601–609. doi: 10.1016/j.ijantimicag.2019.07.015. [DOI] [PubMed] [Google Scholar]
- 77.Kumar A., Choudhir G., Shukla S.K., Sharma M., Tyagi P., Bhushan A., Rathore M. Identification of phytochemical inhibitors against main protease of COVID-19 using molecular modeling approaches. J. Biomol. Struct. Dyn. 2020:1–11. doi: 10.1080/07391102.2020.1772112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Sampangi-Ramaiah M.H., Vishwakarma R., Shaanker R.U. Molecular docking analysis of selected natural products from plants for inhibition of SARS-CoV-2 main protease. Curr. Sci. 2020;118:1087–1092. doi: 10.18520/cs/v118/i7/1087-1092. [DOI] [Google Scholar]
- 79.Villar V.H., Vögler O., Barceló F., Gómez-Florit M., Martínez-Serra J., Obrador-Hevia A., Martín-Broto J., Ruíz-Gutierrez V., Alemany R. Oleanolic and maslinic acid sensitize soft tissue sarcoma cells to doxorubicin by inhibiting the multidrug resistance protein MRP-1, but not P-glycoprotein. J. Nutr. Biochem. 2014;25:429–438. doi: 10.1016/j.jnutbio.2013.12.003. [DOI] [PubMed] [Google Scholar]
- 80.Shan J., Xuan Y., Zhang Q., Zhu C., Liu Z., Zhang S. Ursolic acid synergistically enhances the therapeutic effects of oxaliplatin in colorectal cancer. Protein Cell. 2016;7:571–585. doi: 10.1007/s13238-016-0295-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Jing B., Liu M., Yang L., Cai H.Y., Chen J.B., Li W., Kou X., Wu Y.Z., Qin D.J., Zhou L., et al. Characterization of naturally occurring pentacyclic triterpenes as novel inhibitors of deubiquitinating protease USP7 with anticancer activity in vitro. Acta Pharmacol. Sin. 2018;39:492–498. doi: 10.1038/aps.2017.119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Yu F., Wang Q., Zhang Z., Peng Y., Qiu Y., Shi Y., Zheng Y., Xiao S., Wang H., Huang X., et al. Development of oleanane-type triterpenes as a new class of HCV entry inhibitors. J. Med. Chem. 2013;56:4300–4319. doi: 10.1021/jm301910a. [DOI] [PubMed] [Google Scholar]
- 83.Zhou J., Cai W., Jin M., Xu J., Wang Y., Xiao Y., Hao L., Wang B., Zhang Y., Han J., et al. 18β-Glycyrrhetinic acid suppresses experimental autoimmune encephalomyelitis through inhibition of microglia activation and promotion of remyelination. Sci. Rep. 2015;5:13713. doi: 10.1038/srep13713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Ayub A., Begum S., Ali S.N., Siddiqui B.S. Triterpenoids from the aerial parts of Lantana camara. J. Asian Nat. Prod. Res. 2017;21:141–149. doi: 10.1080/10286020.2017.1408595. [DOI] [PubMed] [Google Scholar]
- 85.Lu M.Y., Liao Z.X., Ji L.J., Sun H.F. Triterpenoids of Chrysosplenium carnosum. Fitoterapia. 2013;85:119–124. doi: 10.1016/j.fitote.2013.01.006. [DOI] [PubMed] [Google Scholar]
- 86.Xu Y.Q., Zhang J.H., Yang X.S. Corosolic acid induces potent anti-cancer effects in CaSki cervical cancer cells through the induction of apoptosis, cell cycle arrest and PI3K/Akt signalling pathway. Bangladesh J. Pharmacol. 2016;11:453. doi: 10.3329/bjp.v11i2.26793. [DOI] [Google Scholar]
- 87.Kaweetripob W., Mahidol C., Prawat H., Ruchirawat S. Lupane, friedelane, oleanane, and ursane triterpenes from the stem of Siphonodon celastrineus Griff. Phytochemistry. 2013;96:404–417. doi: 10.1016/j.phytochem.2013.09.027. [DOI] [PubMed] [Google Scholar]
- 88.Liang C., Ding Y., Song S.B., Kim J.A., Nguyen M.C., Ma J.Y., Kim Y.H., Cuong N.M. Oleanane-triterpenoids from Panax stipuleanatus inhibit NF-κB. J. Ginseng Res. 2013;37:74–79. doi: 10.5142/jgr.2013.37.74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Yan M., Zhu Y., Zhang H.J., Jiao W.H., Han B.N., Liu Z.X., Qiu F., Chen W.S., Lin H.W. Anti-inflammatory secondary metabolites from the leaves of Rosa laevigata. Bioorg. Med. Chem. 2013;21:3290–3297. doi: 10.1016/j.bmc.2013.03.018. [DOI] [PubMed] [Google Scholar]
- 90.Kang H., Ku S.K., Kim J., Chung J., Kim S.C., Zhou W., Na M., Bae J.S. Anti-vascular inflammatory effects of pentacyclic triterpenoids from Astilbe rivularis in vitro and in vivo. Chem. Interact. 2017;261:127–138. doi: 10.1016/j.cbi.2016.11.014. [DOI] [PubMed] [Google Scholar]
- 91.Mosquera C., Panay A.J., Montoya G. Pentacyclic triterpenes from Cecropia telenitida can function as inhibitors of 11β-hydroxysteroid dehydrogenase type 1. Molecules. 2018;23:1444. doi: 10.3390/molecules23061444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Yang F., Dong X., Yin X., Wang W., You L., Ni J. Radix Bupleuri: A review of traditional uses, botany, phytochemistry, pharmacology, and toxicology. BioMed Res. Int. 2017;2017:7597596. doi: 10.1155/2017/7597596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Huang L.R., Luo H., Yang X.S., Chen L., Zhang J.X., Wang D.P., Hao X.J. Enhancement of anti-bacterial and anti-tumor activities of pentacyclic triterpenes by introducing exocyclic α,β-unsaturated ketone moiety in ring A. Med. Chem. Res. 2014;23:4631–4641. doi: 10.1007/s00044-014-1031-z. [DOI] [Google Scholar]
- 94.Liang S., Li M., Yu X., Jin H., Zhang Y., Zhang L., Zhou D., Xiao S. Synthesis and structure-activity relationship studies of water-soluble β-cyclodextrin-glycyrrhetinic acid conjugates as potential anti-influenza virus agents. Eur. J. Med. Chem. 2019;166:328–338. doi: 10.1016/j.ejmech.2019.01.074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Lee J.Y., Choi J.K., Jeong N.-H., Yoo J., Ha Y.S., Lee B., Choi H., Park P.-H., Shin T.-Y., Kwon T.K., et al. Anti-inflammatory effects of ursolic acid-3-acetate on human synovial fibroblasts and a murine model of rheumatoid arthritis. Int. Immunopharmacol. 2017;49:118–125. doi: 10.1016/j.intimp.2017.05.028. [DOI] [PubMed] [Google Scholar]
- 96.Ishida T., Miki I., Tanahashi T., Yagi S., Kondo Y., Inoue J., Kawauchi S., Nishiumi S., Yoshida M., Maeda H., et al. Effect of 18β-glycyrrhetinic acid and hydroxypropyl γcyclodextrin complex on indomethacin-induced small intestinal injury in mice. Eur. J. Pharmacol. 2013;714:125–131. doi: 10.1016/j.ejphar.2013.06.007. [DOI] [PubMed] [Google Scholar]
- 97.Wiemann J., Heller L., Csuk R. Targeting cancer cells with oleanolic and ursolic acid derived hydroxamates. Bioorg. Med. Chem. Lett. 2016;26:907–909. doi: 10.1016/j.bmcl.2015.12.064. [DOI] [PubMed] [Google Scholar]
- 98.Yoon Y., Lim J.W., Kim J., Kim Y., Chun K.H. Discovery of ursolic acid prodrug (NX-201): Pharmacokinetics and in vivo antitumor effects in PANC-1 pancreatic cancer. Bioorg. Med. Chem. Lett. 2016;26:5524–5527. doi: 10.1016/j.bmcl.2016.10.008. [DOI] [PubMed] [Google Scholar]
- 99.Mendes V.I.S., Bartholomeusz G.A., Ayres M., Gandhi V., Salvador J.A.R. Synthesis and cytotoxic activity of novel A-ring cleaved ursolic acid derivatives in human non-small cell lung cancer cells. Eur. J. Med. Chem. 2016;123:317–331. doi: 10.1016/j.ejmech.2016.07.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Xu B., Wu G.R., Zhang X.Y., Yan M.M., Zhao R., Xue N.N., Fang K., Wang H., Chen M., Guo W.B., et al. An overview of structurally modified glycyrrhetinic acid derivatives as antitumor agents. Molecules. 2017;22:924. doi: 10.3390/molecules22060924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Ayeleso T.B., Matumba M.G., Mukwevho E. Oleanolic acid and its derivatives: Biological activities and therapeutic potential in chronic diseases. Molecules. 2017;22:1915. doi: 10.3390/molecules22111915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Mlala S., Oyedeji A.O., Gondwe M., Oyedeji O. Ursolic acid and its derivatives as bioactive agents. Molecules. 2019;24:2751. doi: 10.3390/molecules24152751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Hodon J., Borkova L., Pokorny J., Kazakova A., Urban M. Design and synthesis of pentacyclic triterpene conjugates and their use in medicinal research. Eur. J. Med. Chem. 2019;182:111653. doi: 10.1016/j.ejmech.2019.111653. [DOI] [PubMed] [Google Scholar]
- 104.Isah M.B., Ibrahim M.A., Mohammed A., Aliyu A.B., Masola B., Coetzer T.H.T. A systematic review of pentacyclic triterpenes and their derivatives as chemotherapeutic agents against tropical parasitic diseases. Parasitology. 2016;143:1219–1231. doi: 10.1017/S0031182016000718. [DOI] [PubMed] [Google Scholar]
- 105.Ren Y., Kinghorn A.D. Natural product triterpenoids and their semi-synthetic derivatives with potential anticancer activity. Planta Med. 2019;85:802–814. doi: 10.1055/a-0832-2383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Parida P.K., Sau A., Ghosh T., Jana K., Biswas K., Raha S., Misra A.K. Synthesis and evaluation of triazole linked glycosylated 18β-glycyrrhetinic acid derivatives as anticancer agents. Bioorg. Med. Chem. Lett. 2014;24:3865–3868. doi: 10.1016/j.bmcl.2014.06.054. [DOI] [PubMed] [Google Scholar]
- 107.Ghosh S. Triterpene structural diversification by plant cytochrome P450 enzymes. Front. Plant Sci. 2017;8:1886. doi: 10.3389/fpls.2017.01886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Martinez A., Rivas F., Perojil A., Parra A., Garcia-Granados A., Fernandez-Vivas A. Biotransformation of oleanolic and maslinic acids by Rhizomucor miehei. Phytochemistry. 2013;94:229–237. doi: 10.1016/j.phytochem.2013.05.011. [DOI] [PubMed] [Google Scholar]
- 109.Djerassi C., Henry J.A., Lemin A.J., Rios T., Thomas G.H. Terpenoids. XXIV. The structure of the cactus triterpene queretaroic acid. J. Am. Chem. Soc. 1956;78:3783–3787. doi: 10.1021/ja01596a060. [DOI] [Google Scholar]
- 110.Kinoshita K., Yang Y., Koyama K., Takahashi K., Nishino H. Inhibitory effect of some triterpenes from cacti on 32Pi-incorporation into phospholipids of HeLa cells promoted by 12-O-tetradecanoylphorbol-13-acetate. Phytomedicine. 1999;6:73–77. doi: 10.1016/S0944-7113(99)80038-5. [DOI] [PubMed] [Google Scholar]
- 111.Fujii Y., Hirosue S., Fujii T., Matsumoto N., Agematu H., Arisawa A. Hydroxylation of oleanolic acid to queretaroic acid by cytochrome P450 from Nonomuraea recticatena. Biosci. Biotechnol. Biochem. 2006;70:2299–2302. doi: 10.1271/bbb.60126. [DOI] [PubMed] [Google Scholar]
- 112.Martinez A., Perojil A., Rivas F., Parra A., Garcia-Granados A., Fernández-Vivas A. Biotransformation of oleanolic and maslinic methyl esters by Rhizomucor miehei CECT 2749. Phytochemistry. 2015;117:500–508. doi: 10.1016/j.phytochem.2015.07.020. [DOI] [PubMed] [Google Scholar]
- 113.Sun H., Liu T., Shen Y.-J., Zhang L.-M., Wang M. Preparation and crystal structure of 15α-hydroxyl-oleanolic acid. Jiegou Huaxue. 2010;29:1789–1801. [Google Scholar]
- 114.Gong T., Zheng L., Zhen X., He H.-X., Zhu H.-X., Zhu P. Microbial transformation of oleanolic acid by Trichothecium roseum. J. Asian Nat. Prod. Res. 2014;16:383–386. doi: 10.1080/10286020.2014.884564. [DOI] [PubMed] [Google Scholar]
- 115.Fan B., Jiang B., Yan S., Xu B., Huang H., Chen G.-T. Anti-inflammatory 18β-glycyrrhetinin acid derivatives produced by biocatalysis. Planta Med. 2019;85:56–61. doi: 10.1055/a-0662-0296. [DOI] [PubMed] [Google Scholar]
- 116.Qin Y.J., Feng B., Song X.B., Zhou W.B., Yu H.S., Zhao L.L., Yu L.Y., Ma B.P. Biotransformation of glycyrrhetinic acid by Cunninghamella blakesleeana. Chin. J. Nat. Med. 2010;8:373–381. doi: 10.3724/SP.J.1009.2010.00373. [DOI] [Google Scholar]
- 117.Maatooq G.T., Marzouk A.M., Gray A.I., Rosazza J.P. Bioactive microbial metabolites from glycyrrhetinic acid. Phytochemistry. 2010;71:262–270. doi: 10.1016/j.phytochem.2009.09.014. [DOI] [PubMed] [Google Scholar]
- 118.He C.J., Yang Y.M., Wu K.Y. Microbial transformation of glycyrrhetinic acid by Colletotrichum lini AS3.4486. Adv. Mater. Res. 2015;1120:877–881. doi: 10.4028/www.scientific.net/AMR.1120-1121.877. [DOI] [Google Scholar]
- 119.Choudhary M.I., Siddiqui Z.A., Nawaz S.A. Atta-Ur-Rahman Microbial transformation of 18β-glycyrrhetinic acid by Cunninghamella elegans and Fusarium lini, and lipoxygenase inhibitory activity of transformed products. Nat. Prod. Res. 2009;23:507–513. doi: 10.1080/14786410500463536. [DOI] [PubMed] [Google Scholar]
- 120.Huang F.X., Yang W.Z., Ye F., Tian J.Y., Hu H.B., Feng L.M., Guo D.A., Ye M. Microbial transformation of ursolic acid by Syncephalastrum racemosum (Cohn) Schroter AS 3.264. Phytochemistry. 2012;82:56–60. doi: 10.1016/j.phytochem.2012.06.020. [DOI] [PubMed] [Google Scholar]
- 121.Fu S., Meng Q., Yang J., Tu J., Sun D.-A. Biocatalysis of ursolic acid by the fungus Gliocladium roseum CGMCC 3.3657 and resulting anti-HCV activity. RSC Adv. 2018;8:16400–16405. doi: 10.1039/C8RA01217B. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Zhang S.S., He L.S., Zhao Y.M., Fu S., Liu D.L., Yu Z.H., Liu B.C. Three new triterpenoids transformed from ursolic acid by Mucor spinosus AS3.3450 and their cytotoxicity. Phytochem. Lett. 2019;32:33–37. doi: 10.1016/j.phytol.2019.04.019. [DOI] [Google Scholar]
- 123.Zhang J., Cheng Z.H., Yu B.Y., Cordell G.A., Qiu S.X. Novel biotransformation of pentacyclic triterpenoid acids by Nocardia sp. NRRL 5646. Tetrahedron Lett. 2005;46:2337–2340. doi: 10.1016/j.tetlet.2005.01.155. [DOI] [Google Scholar]
- 124.Huang D., Ding Y., Li Y., Zhang W., Fang W.-S., Chen X. Anti-tumor activity of a 3-oxo derivative of oleanolic acid. Cancer Lett. 2006;233:289–296. doi: 10.1016/j.canlet.2005.03.019. [DOI] [PubMed] [Google Scholar]
- 125.Funari C.S., De Almeida L., Passalacqua T.G., Martinez I., Ambrósio D.L., Cicarelli R.M.B., Silva D.H.S., Graminha M.A.S. Oleanonic acid from Lippia lupulina (Verbenaceae) shows strong in vitro antileishmanial and antitrypanosomal activity. Acta Amaz. 2016;46:411–416. doi: 10.1590/1809-4392201600204. [DOI] [Google Scholar]
- 126.Krivoruchko A.V., Kuyukina M.S., Ivshina I.B. Advanced Rhodococcus biocatalysts for environmental biotechnologies. Catalysts. 2019;9:236. doi: 10.3390/catal9030236. [DOI] [Google Scholar]
- 127.Leipold D., Wünsch G., Schmidt M., Bart H.-J., Bley T., Neuhaus H.E., Bergmann H., Richling E., Muffler K., Ulber R. Biosynthesis of ursolic acid derivatives by microbial metabolism of ursolic acid with Nocardia sp. strains—Proposal of new biosynthetic pathways. Process. Biochem. 2010;45:1043–1051. doi: 10.1016/j.procbio.2010.03.013. [DOI] [Google Scholar]
- 128.Xu S.H., Wang W.W., Zhang C., Liu X.F., Yu B.Y., Zhang J. Site-selective oxidation of unactivated C–H sp 3 bonds of oleanane triterpenes by Streptomyces griseus ATCC 13273. Tetrahedron. 2017;73:3086–3092. doi: 10.1016/j.tet.2017.04.036. [DOI] [Google Scholar]
- 129.Zhu Y.Y., Qian L.W., Zhang J., Liu J.H., Yu B. New approaches to the structural modification of olean-type pentacylic triterpenes via microbial oxidation and glycosylation. Tetrahedron. 2011;67:4206–4211. doi: 10.1016/j.tet.2011.04.055. [DOI] [Google Scholar]
- 130.De Sousa L.R.F., Silva J.A.D., Vieira P.C., Costa M.B., Dos Santos M.L., Sbardelotto A.B., Pessoa C.D.Ó., De Moraes M.O., Menezes A.C.S. Chemical constituents of the stem bark of Vochysia thyrsoidea Pohl. (Vochysiaceae) and evaluation of their cytotoxicity and inhibitory activity against cathepsins B and K. Química Nova. 2014;37:288–292. doi: 10.5935/0100-4042.20140049. [DOI] [Google Scholar]
- 131.Capel C.S., De Souza A.C.D., De Carvalho T.C., De Sousa J.P.B., Ambrósio S.R., Martins C.H.G., Cunha W.R., Hernández-Galán R., Furtado N.A.J.C. Biotransformation using Mucor rouxii for the production of oleanolic acid derivatives and their antimicrobial activity against oral pathogens. J. Ind. Microbiol. Biotechnol. 2011;38:1493–1498. doi: 10.1007/s10295-010-0935-y. [DOI] [PubMed] [Google Scholar]
- 132.Chianese G., Golin-Pacheco S.D., Taglialatela-Scafati O., Collado J.A., Munoz E., Appendino G., Pollastro F. Bioactive triterpenoids from the caffeine-rich plants guayusa and maté. Food Res. Int. 2019;115:504–510. doi: 10.1016/j.foodres.2018.10.005. [DOI] [PubMed] [Google Scholar]
- 133.Sit N.W., Chan Y., Lai S., Lim L., Looi G., Tay P., Tee Y., Woon Y., Khoo K., Ong H. In vitro antidermatophytic activity and cytotoxicity of extracts derived from medicinal plants and marine algae. J. Mycol. Med. 2018;28:561–567. doi: 10.1016/j.mycmed.2018.07.001. [DOI] [PubMed] [Google Scholar]
- 134.Su H.G., Peng X.R., Shi Q.Q., Huang Y.J., Zhou L., Qiu M.H. Lanostane triterpenoids with anti-inflammatory activities from Ganoderma lucidum. Phytochemistry. 2020;173:112256. doi: 10.1016/j.phytochem.2019.112256. [DOI] [PubMed] [Google Scholar]
- 135.Tian M., Zhao P.-J., Li G., Zhang K.-Q. In depth natural product discovery from the basidiomycetes Stereum species. Microorganisms. 2020;8:1049. doi: 10.3390/microorganisms8071049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Ying Y.M., Yu H.F., Tong C.P., Shan W.G., Zhan Z.J. Spiroinonotsuoxotriols A and B, two highly rearranged triterpenoids from Inonotus obliquus. Org. Lett. 2020;22:3377–3380. doi: 10.1021/acs.orglett.0c00866. [DOI] [PubMed] [Google Scholar]
- 137.Shi Q.Q., Huang Y.J., Su H.G., Gao Y., Lu S.Y., Peng X.R., Li X.N., Zhou L., Qiu M.H. Structurally diverse lanostane triterpenoids from medicinal and edible mushroom Ganoderma resinaceum Boud. Bioorg. Chem. 2020;100:103871. doi: 10.1016/j.bioorg.2020.103871. [DOI] [PubMed] [Google Scholar]
- 138.Nursid M., Marraskuranto E., Chasanah E. Cytotoxicity and apoptosis induction of sea cucumber Holothuria atra extracts. Pharmacogn. Res. 2019;11:41. doi: 10.4103/pr.pr_3_18. [DOI] [Google Scholar]
- 139.Xu R., Fazio G.C., Matsuda S.P.T. On the origins of triterpenoid skeletal diversity. Phytochemistry. 2004;65:261–291. doi: 10.1016/j.phytochem.2003.11.014. [DOI] [PubMed] [Google Scholar]
- 140.Chubukov V., Mukhopadhyay A., Petzold C.J., Keasling J.D., Martin H.G. Synthetic and systems biology for microbial production of commodity chemicals. NPJ Syst. Biol. Appl. 2016;2:16009. doi: 10.1038/npjsba.2016.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Heller L., Schwarz S., Perl V., Köwitsch A., Siewert B., Csuk R. Incorporation of a Michael acceptor enhances the antitumor activity of triterpenoic acids. Eur. J. Med. Chem. 2015;101:391–399. doi: 10.1016/j.ejmech.2015.07.004. [DOI] [PubMed] [Google Scholar]