

ABSTRACT
Enteroaggregative Escherichia coli (EAEC) is a common diarrhoeagenic human pathogen, isolated from patients in both developing and industrialized countries, that is becoming increasingly resistant to many frontline antibiotics. In this study, we screened 50 E. coli strains from children presenting with diarrhea at the outpatients clinic of Assiut University Children’s Hospital, Egypt. We show that all of these isolates were resistant to multiple classes of antibiotics and identified two as being typical EAEC strains. Using whole genome sequencing, we determined that both isolates carried, amongst others, blaCTX-M and blaTEM antibiotic resistance genes, as well as many classical EAEC virulence determinants, including the transcriptional regulator, AggR. We demonstrate that the expression of these virulence determinants is dependent on AggR, including aar, which encodes for a repressor of AggR, Aar. Since biofilm formation is the hallmark of EAEC infection, we examined the effect of Aar overexpression on both biofilm formation and AggR-dependent gene expression. We show that whilst Aar has a minimal effect on AggR-dependent transcription it is able to completely disrupt biofilm formation, suggesting that Aar affects these two processes differently. Taken together, our results suggest a model for the induction of virulence gene expression in EAEC that may explain the ubiquity of EAEC in both sick and healthy individuals.
KEYWORDS: EAEC, antibiotic resistance, virulence, bacterial gene regulation, genome sequencing
Introduction
Diarrhoeagenic Escherichia coli strains are important human pathogens, which cause considerable morbidity and mortality around the globe, particularly amongst infants and children in developing countries. These pathogens are classified into different pathotypes, which include enteroaggregative E. coli (EAEC), enterohaemorrhagic E. coli (EHEC), enteroinvasive E. coli (EIEC), enteropathogenic E. coli (EPEC), enterotoxigenic E. coli (ETEC) and diffuse adhering E. coli (DAEC), based on their disease characteristics and specific adherence patterns [1]. Enteroaggregative Escherichia coli (EAEC) is a commonly isolated human pathogen that is responsible for causing mucoid diarrhea in patients from both industrialized and developing countries [2–5]. EAEC has been shown to elicit travelers’ diarrhea, pediatric diarrhea, impairment of pediatric growth and cognition, and even extra-intestinal infections, such as urinary tract infections and septicemia [4,6–11]. In addition, EAEC strains have been the cause of a number of serious diarrhea outbreaks throughout the world, with the food-borne outbreak caused by the Shiga-toxin-producing EAEC O104:H4 strain in Germany, resulting in 54 deaths [12–16].
EAEC pathogenesis proceeds by the colonization of the human intestinal mucosa followed by the secretion of toxins, such as plasmid-encoded toxin (Pet) and enteroaggregative heat-stable toxin (EAST-1). This, coupled with the resultant inflammation, is thought to lead to diarrhea and disease [17,18]. Typical EAEC strains carry the plasmid-encoded AggR transcription regulator protein, a member of the AraC-XylS family of transcription factors [19,20]. AggR activates the expression of many genes thought to be required for pathogenesis, for example the attachment adherence fimbriae (AAF) required for colonization, the anti-aggregation protein dispersin (Aap), and its dedicated type I secretion system (T1SS) [21–24]. Additionally, expression of AggR is up-regulated by AggR itself [25] and its activity is reported to be down-regulated by the Aar protein (AggR-activated regulator), whose expression is also induced by AggR [24,26,27]. Thus, there is a complex interplay between AggR and Aar to control EAEC virulence [26,28].
Much of what we understand about EAEC virulence comes from research with archetypal EAEC strains, such as EAEC strains 042 and 17–2 [29]. However, it is becoming clear that, in the environment, EAEC strains are extremely heterogeneous and many virulence determinants are not present in all strains [5,16]. For example, both EAEC 042 and 17–2 possess large virulence plasmids (i.e. pAA2 and pAA, respectively) and carry EAEC-associated genes encoding AggR, Aap and EAST-1 (astA). However, EAEC 042 possesses AAF Type II fimbriae (aafDA and afaB-aafCB), whilst EAEC 17–2 expresses AAF Type I fimbriae (aggDCBA) and lacks the Pet cytotoxin [5,19,21,30]. Like many enteric pathogens, EAEC strains are increasingly resistant to many clinically used antibiotics [31–33] and, as EAEC is often isolated from asymptomatic individuals [5,16], there is a need to understand how EAEC is tolerated by their human hosts but causes disease in certain individuals.
In Egypt, EAEC is increasingly recognized as an emerging enteric pathogen, showing high levels of antibiotic resistance. However, in-depth molecular characterization of Egyptian EAEC isolates has been limited to only a few studies [34–38]. In order to examine both the prevalence of antibiotic resistance and the occurrence of EAEC, we screened E. coli strains from infants and children with diarrhea from Assiut, Egypt. Alarmingly, most strains were resistant to many classes of antibiotics. Amongst these, we found two apparently typical EAEC strains that we characterized, using whole genome sequencing, identifying their virulence determinants, and investigating their regulation by AggR. We show that feed-forward activation of AggR expression by AggR is conserved, but that Aar-mediated repression is not. Our results lead to a model for the triggering of virulence in typical EAEC strains.
Materials and methods
Bacterial strains, plasmids, primers and growth conditions
The bacterial strains, plasmids and promoter fragments used in this study are listed in Table S1. The oligodeoxynucleotide primers used to amplify and mutate the various DNA fragments are listed in Table S2. Standard procedures for PCR, cloning and DNA manipulation were used throughout [39]. All promoter DNA fragments used in this study are flanked by EcoRI and HindIII and the DNA sequence of each fragment is numbered from the base adjacent to the HindIII site. Base substitutions are defined by the position of the base altered and the substituted base introduced. Cells were routinely grown in Lysogeny Broth (LB medium) at 37ºC with shaking. To measure promoter activities, fragments were cloned into the low copy number broad host range lac expression vectors, pRW50 [40] and pRW224 [41] and maintained with 15 μg ml−1 tetracycline. To examine the effect of aggR and aar expression, cells were transformed with various pBAD derivatives (Table S1), which were maintained in cells with 100 µg ml−1 ampicillin. AggR and/or Aar expression, using pBAD vectors, was induced by the addition of 0.2% w/v arabinose in the medium, where appropriate [22].
Isolation and characterization of Egyptian E. coli strains
In total, 113 stool samples were collected from infants and children, whose age ranged between 2 months and 5 years, presenting with diarrhea to the outpatients clinic of Assiut University Children’s Hospital in 2016. Ethical approval was granted by the Medical School Ethical Review Board before sample collection proceeded. Cases enrolled in this study had diarrhea, characterized by frequent watery stools (>3 times/day), with or without blood or mucus. Children and infants with severe protein energy malnutrition (either Marasmus or Kwashiorkor), or who had received antibiotics within the last 72 hours, were excluded from the study. Only one stool specimen from each case was tested, with isolation and identification of E. coli carried out at the Medical Research Center, Faculty of Medicine, Assiut University. All E. coli strains were tested for susceptibility to a range of antimicrobial agents, using the Kirby-Bauer disc diffusion method [42], which was interpreted according to the CLSI 2014 [43]. The antimicrobial discs (Hi-Media, India) contained the following antibiotics: imipenem (10 µg), meropenem (10 µg), trimethoprim/sulfamethoxazole (5 µg), cefaclor (30 µg), ceftriaxone (30 µg), ampicillin (10 µg), ciprofloxacin (5 µg), oxytetracycline (30 µg), amoxicillin (25 µg), norfloxacin: (10 µg), tobramycin: (10 µg) and amikacin (30 µg). To identify EAEC-associated genes, isolates were screened for aggR, aap and the CVD432 marker sequence (which encodes part of the EAEC Aat T1SS), using PCR and the primers detailed in Table S2.
Complete genome sequencing and analysis
Complete genome sequencing of EAEC strains E36 and E42 was carried out using Illumina sequencing by Microbes NG (https://microbesng.com/). Plated cultures of each isolate were inoculated into a cryopreservative (Microbank™, Pro-Lab Diagnostics UK). 10 to 20 µl of this suspension were lysed with 120 µL of TE buffer containing lysozyme (final concentration 0.1 mg mL−1) and RNase A (ITW Reagents, Barcelona, Spain) (final concentration 0.1 mg mL−1), incubated for 25 min at 37°C. Proteinase K (VWR Chemicals, Ohio, USA) (final concentration 0.1 mg mL−1) and SDS (Sigma-Aldrich, Missouri, USA) (final concentration 0.5% v/v) were added and incubated for 5 min at 65°C. Genomic DNA was purified using an equal volume of SPRI beads and resuspended in EB buffer (Qiagen, Germany). DNA was quantified with the Quant-iT dsDNA HS kit (ThermoFisher Scientific) assay in an Eppendorf AF2200 plate reader (Eppendof UK Ltd, UK). Genomic DNA libraries were prepared using the Nextera XT Library Prep Kit (Illumina, San Diego, USA) following the manufacturer’s protocol with the following modifications: 2 ng of DNA were used as input, and PCR elongation time was increased to 1 min from 30 s. DNA quantification and library preparation were carried out on a Hamilton Microlab STAR automated liquid handling system (Hamilton Bonaduz AG, Switzerland). Pooled libraries were quantified using the Kapa Biosystems Library Quantification Kit for Illumina on a Roche light cycler 96 qPCR machine. Libraries were sequenced with the Illumina HiSeq using a 250bp paired end protocol.
EAEC strain 17–2 was sequenced using the enhanced sequencing option from MicrobesNG, which utilizes both the Illumina and Oxford Nanopore Technologies (ONT). A broth culture was pelleted out and the pellet was resuspended in the cryopreservative of a Microbank™ (Pro-Lab Diagnostics UK, United Kingdom) tube and stored in the tube. Approximately 2 × 109 cells were used for high molecular weight DNA extraction using Nanobind CCB Big DNA Kit (Circulomics, Maryland, USA). DNA was quantified with the Qubit dsDNA HS assay in a Qubit 3.0 (Invitrogen) Eppendof UK Ltd, UK). Long read genomic DNA libraries were prepared with the Oxford Nanopore SQK-LSK109 kit with Native Barcoding EXP-NBD104/114 (ONT, UK), using 400–500 ng of HMW DNA. Twelve to twenty-four barcoded samples were pooled together into a single sequencing library and loaded on a FLO-MIN106 (R.9.4 or R.9.4.1) flow cell in a GridION (ONT, UK). Illumina reads were adapter trimmed using Trimmomatic 0.30 with a sliding window quality cutoff of Q15 [44]. Genome assembly was performed using Unicycler v0.4.0 [45] and contigs were annotated using Prokka 1.11 [46]. This Whole Genome Shotgun project has been deposited at DDBJ/ENA/GenBank with the sequence data for EAEC strains E36, E42 and 17–2 under the accession numbers JACEFX000000000, JACEFW000000000 and JACEFV000000000, respectively.
Bioinformatic analysis of genome sequences
Draft genomes were visualized using Artemis [47], comparisons between EAEC genomes were examined using the CGView Server (http://stothard.afns.ualberta.ca/cgview_server/) [48], the Basic Local Alignment Search Tool (BLAST) at NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi) and the Artemis Comparison Tool (ACT) [49]. Representations of genome organization were drawn using the CGView Server [48], ACT [49] and DNAPlotter [50]. E. coli sequence types were determined using MLST 2.0 [51], bacterial serotyping was determined using SerotypeFinder 2.0 [52], plasmid replicons were detected using PlasmidFinder 2.1 [53], antibiotic resistance gene analysis used ResFinder 3.2 [54], and virulence gene analysis was performed using VirulenceFinder 2.0 [55] with the online software from the Center for Genomic Epidemiology (http://www.genomicepidemiology.org/). Insertion sequences were identified using ISfinder (https://www-is.biotoul.fr/blast/resultat.php) [56]. The phylogenetic analysis of EAEC strains E36 and E42 was carried out by recreating the phylogenetic tree from Dunne et al. [57], using the 35 genomes of Escherichia and Shigella listed in (Figure 1), the three EAEC genomes generated in this study, and EAEC O104:H4 strain C227-11 (accession number AFST00000000) [58]. Automated annotation was performed using Prokka v1.12 [46]. A pangenome analysis was performed using Roary v3.13.0 [59] to determine the sizes of the core genomes and pangenomes. A whole-genome phylogeny was reconstructed from the core genome alignment using RAxML 8.2.4 [60].
Figure 1.
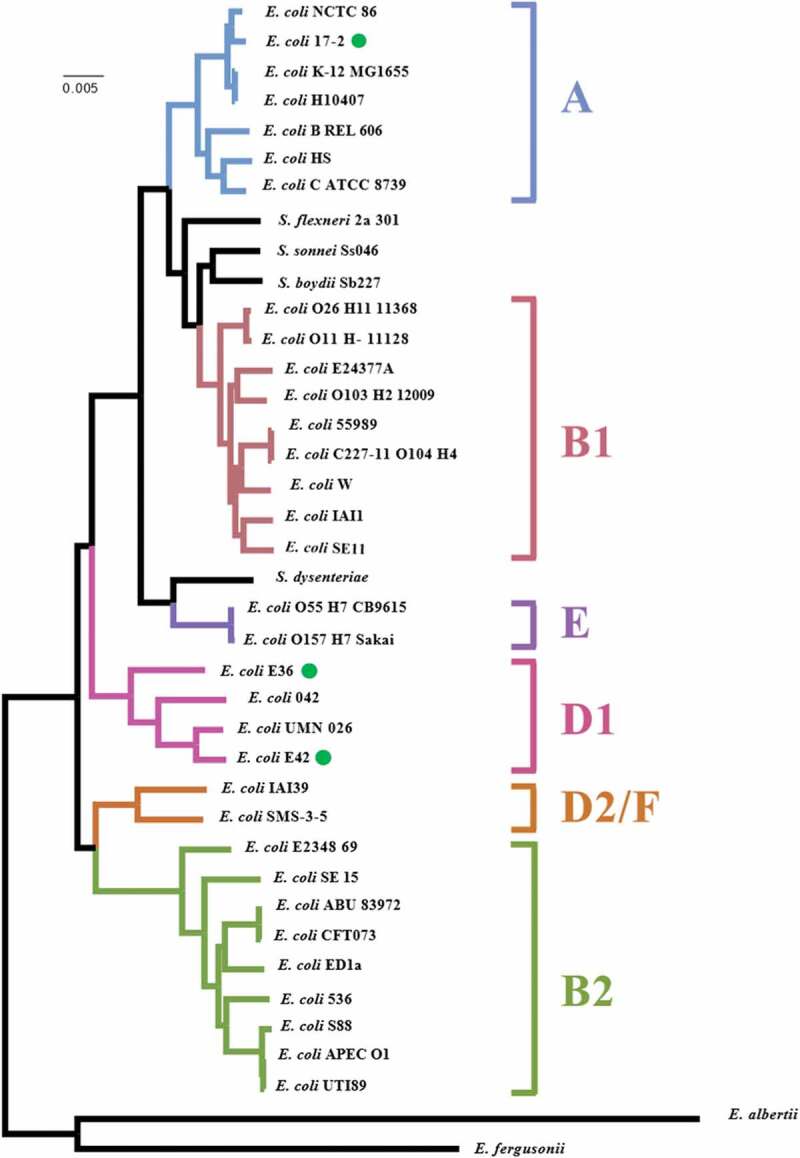
Phylogenetic analysis of EAEC strains E36 and E42. The figure shows a phylogenetic tree of E. coli strains highlighting the position of EAEC strains E36, E42 and 17–2 (green dots). The phylogenetic tree was reconstructed from Dunne et al. [57], using RAxML (version 8.2.4) based on maximum likelihood analysis. Phylogenetic groups and labeling is similar to the reference phylogenetic tree in [57] and EAEC O104:H4 strain C227-11 (accession number AFST00000000) [58] has also been included
Promoter fragment and plasmid construction
The promoter fragments aggR90, aar100, aatP100 and aatP98 were amplified by PCR using the primer pairs listed in Table S2, with EAEC 042, EAEC 17–2, EAEC E36 or EAEC E42 genomic DNA as template. All DNA fragments are flanked by EcoRI and HindIII sites to facilitate cloning into pRW50 and pRW224 to generate lacZ transcriptional fusions [40,41], and sequences are numbered from the HindIII site. Point mutations were introduced into fragments using megaprimer PCR [61] and constructs were verified by Sanger DNA sequencing.
To generate the various pBAD30/aar plasmids used in this study (Table S1), the aar gene was amplified from EAEC 042 and EAEC E36 DNA by PCR using primers aar XbaI up, aar E36 XbaI up or aar* XbaI up with primers aar SphI down or aar E36 SphI down, to generate each aar or aar* PCR products (Tables S1 and S2). Note that aar* derivatives carry the strong RBS from the pET20b expression vector (Novagen). Purified PCR products were restricted with XbaI and SphI and then cloned into pBAD30, cut with the same enzymes, thus, placing the expression of aar derivatives under the control of the arabinose inducible paraBAD promoter [62]. To generate the pBAD/aggR/aar series of vectors (Table S1), purified aar and aar* PCR products were cloned downstream of aggR in pBAD/aggR, again using XbaI and SphI, to generate synthetic aggR-aar operons, which allow the concurrent expression of AggR and Aar, by induction with 0.2% w/v arabinose.
Assays of promoter activity
To assay the expression from promoter derivatives cloned into the lac expression vectors pRW50 and pRW224, E. coli K-12 BW25113 Δlac strain was transformed with each construct and β-galactosidase activity was measured, as described in our previous work [27,63]. AggR was expressed from pBAD/aggR, which carries EAEC 042 aggR cloned downstream of the arabinose inducible promoter, paraBAD[22]. Cells were grown in LB medium at 37°C with shaking to mid-logarithmic phase (OD650 = 0.4–0.6) and 0.2% w/v arabinose was included in the medium to induce AggR expression, where appropriate. β-galactosidase activities are expressed as nmol of ONPG hydrolyzed min−1 mg−1 dry cell mass and each activity is the average of three independent determinations.
Quantitative biofilm formation assays
Quantitative biofilm formation assays were carried out as in our previous work [5]. Overnight EAEC cultures, grown in LB medium, were sub-cultured (1 in 100) into 5 ml of DMEM (Dulbecco’s Modified Eagle Medium) high glucose (Sigma) and incubated at 37°C for 1 hour with shaking. 150 µl of each culture was pipetted into a microtiter plate in triplicate, which was sealed with a Breath Easy gas permeable membrane (Sigma), and then incubated overnight statically at 37°C. After ~16-17 hrs the spent media was removed and 150 µl of 0.1% (w/v) crystal violet was added to each well and left at 4°C for 30 minutes. The crystal violet solution was removed, the plate washed thoroughly with water, excess liquid removed and 150 µl of ethanol/acetone solution (80 ml ethanol and 20 ml acetone) was added. The plate was left on a shaker for 30 minutes at room temperature and the absorbance was measured at 595 nm by a Labsystems Multiskan MS plate reader (Thermo Fisher Scientific Inc).
Results and discussion
Isolation and characterization of E. coli strains from Egyptian children with diarrhea
In total, 50 E. coli strains were isolated from infants and children, presenting with diarrhea at the outpatient clinic of Assiut University Children’s Hospital, and their susceptibility against various antimicrobials tested. Results in Table 1 show that all were resistant to the cephalosporin antibiotics, cefaclor (100%) and ceftriaxone (100%), and the vast majority of isolates were resistant to trimethoprim/sulfamethoxazole (94%) and ampicillin (98%). Furthermore, four isolates (E16, E28, E35 and E36) were resistant to all tested antibiotics (Table S3), highlighting the multidrug resistance phenotype of many of the E. coli strains isolated during this study.
Table 1.
Antimicrobial susceptibility profile of 50 E. coli strains isolated from children with diarrhea from Assiut Children’s Hospital, Egypt
| Antibiotic | Resistant n (%) |
Sensitive n (%) |
Intermediate a n (%) |
|---|---|---|---|
| Imipenem | 16 (32) | 30 (60) | 4 (8) |
| Meropenem | 14 (28) | 27 (54) | 9 (18) |
| Trimethoprim/sulfamethoxazole | 47 (94) | 2 (4) | 1 (2) |
| Cefaclor | 50 (100) | 0 (0) | 0 (0) |
| Ceftriaxone | 50 (100) | 0 (0) | 0 (0) |
| Ampicillin | 49 (98) | 1 (2) | 0 (0) |
| Ciprofloxacin | 24 (48) | 13 (26) | 13 (26) |
| Oxytetracycline | 29 (58) | 14 (28) | 7 (14) |
| Amoxicillin | 27 (54) | 12(24) | 11 (22) |
| Norfloxacin | 22 (44) | 21(42) | 7 (14) |
| Tobramycin | 34 (68) | 1 (2) | 15 (30) |
| Amikacin | 22 (44) | 13 (26) | 15 (30) |
aIntermediate resistance refers to strains above the point of antibiotic susceptibility but below the resistant breakpoint as defined by the [43].
To determine if any of these isolates were EAEC in character, strains were screened for the EAEC-associated genes, aggR and aap, as well as the CVD432 marker sequence, which encodes part of the EAEC Aat T1SS [10]. Two strains, E36 and E42, possessed aggR, aap and CVD432, identifying them as typical EAEC isolates. To verify this, and to determine what antibiotic resistance genes and virulence determinants they carried, the genome of each of these strains was sequenced, together with the genome of the prototypical EAEC strain, 17–2, as a reference (Table S1). Analysis indicated that the genomes of strains E36 and E42 were similar in size, with E42 containing slightly more coding sequences (i.e. 5046 vs 4912) (Table 2). Both strains were found to belong to E. coli phylogenetic group D, like EAEC 042, with E36 belonging to sequence type ST38 and E42 to ST1380 (Table 2; Figure 1). EAEC 17–2, which possessed a slightly smaller genome, was E. coli phylogenetic group A and sequence type ST10 (Table 2; Figure 1) [64]. For each strain, a number of plasmid replicons were detected, consistent with each strain possessing a number of plasmids [53]. Due to the draft nature of the E36 and E42 genome assemblies, each plasmid replicon was found on separate contigs and, therefore, it is not immediately clear how many plasmids each isolate possesses. However, analysis of EAEC 17–2 (Table 2), which possess a single large virulence plasmid (pAA) [65], revealed similar IncFIB and IncFII replicons on the same large 81 kbp contig (Fig. S1) [53]. As both of these replicons often occur together on other IncF plasmids [53,66], it is likely that the genomes of E36 and E42 contain a similar large virulence plasmid, bearing both IncF replicons. Interestingly, comparison of plasmid pAA2 from EAEC 042 [67] with our pAA sequence from EAEC 17–2, indicated that these two plasmids were very different, with regions of homology confined mainly to EAEC virulence determinants (see below) and insertion sequences (Fig. S1). This is consistent with the proposition that a conserved plasmid backbone may not exist for pAA plasmids, and that EAEC virulence determinants have integrated into many different plasmids to produce the EAEC pathotype [64].
Table 2.
Genome analysis of EAEC isolates E36, E42, 17–2, 042 and C227-11
| EAEC E36 | EAEC E42 | EAEC 17–2 | EAEC 042 | EAEC C227-11 | |
|---|---|---|---|---|---|
| Genome size | 5,410,238 bp | 5,290,086 bp | 5,185,323 bp | 5,355,323 bp | 5542971bp |
| G/C % | 50.3% | 50.47% | 50.63% | 50.56% h | 50.71% h |
| Number of contigs | 150 | 149 | 9 | N/A | 37 |
| Genes (CDS) | 5046 | 4912 | 4924 | 4921 | 5431 |
| Phylotype | D | D | A | D | B1 |
| Sequence Type a | ST38 | ST1380 | ST10 | ST414 | ST678 |
| Serotype b | O?:H30 | O17:H18 | O3:H2 | O44:H18 | O104:H4 |
| Plasmid replicons c | IncFIB, IncFII, IncQ1 | IncFIB, IncFII, IncI1-I(γ) | IncFIB, IncFII | IncFIC(FII) | IncFIB, IncFII, IncI1-I(γ), IncQ1 |
| Antibiotic resistance genes d | aadA1, aph(3ʹ’)-Ib, aph(3ʹ)-Ia, aph(6)-Id, blaCTX-M-14b, blaTEM-1B, dfrA1, mdfA, sul2, tetA | blaCTX-M-15, blaTEM-1B, mdfA, gyrA pS83A | aadA1, dfrA1, mdfA, sul2, | aadA1, catA1, mdfA, sul1, tetA |
aph(3ʹ’)-Ib, aph(6)-Id, blaCTX-M-15, blaTEM-1B, dfrA7, sul1, sul2, mdfA, tetA, gyrA pS83A |
| Virulence genes e,f | orf3, orf4, aap, aar, aatA, agg3B, agg3C, agg3D, agg5A, aggR, capU, air, eilA, gadB, chuA, fyuA, irp2, iss | orf3, orf4, aaiC, aap, aatA, agg4A, agg4C, agg4D, aggR, capU, air, eilA, gadA, gadB, chuA, iss, lpfA, sepA | orf3, orf4, aaiC, aap, aar, aatA, aggA, aggB, aggC, aggD, aggR, capU, astA, gadA, gadB, iha, iss, fyuA, irp2, iucC, iutA, papA, papC, sat | orf3, orf4, aaiC, aap, aar, aatA, aafA, aafB, aafC, aafD, aggR, capU, astA, air, eilA, gadA, gadB, chuA, fyuA, irp2, lpfA, mchB, mchC, mchF, mcmA, pet, pic | orf3, orf4, aaiC, aap, aar aatA, aggA, aggB, aggC, aggD, aggR, capU, gadA, gadB, iha, fyuA irp2, iucC, iutA, lpfA, mchB, mchC, mchF, neuC, pic, sepA, sigA, stx2A, stx2B |
Antibiotic resistance genes carried by EAEC strains E36 and E42
In the genome of EAEC E36, we detected various antibiotic resistance genes (Table 2), which can result in aminoglycoside resistance (aadA1, aph(3ʹ’)-Ib, aph(3ʹ)-Ia and aph(6)-Id), tetracycline resistance (tetA), β-lactam resistance (blaCTX-M-14b and blaTEM-1B), trimethoprim resistance (dfrA1), macrolide resistance (mdfA) and sulfonamide resistance (sul2). This is in line with the observed multidrug resistance phenotype of EAEC E36 (Table S3). For EAEC E42, we detected resistance genes for β-lactams (blaCTX-M-15 and blaTEM-1B), macrolides (mdfA) and nalidixic acid/ciprofloxacin (gyrA pS83A). Note that the extended spectrum β-lactamases (ESBL) type CTX-M and TEM found in both strains are often associated with EAEC isolates and the sequence types to which each strain belongs (i.e. ST38 and ST1380) [31–33,68].
Interestingly, in EAEC E36 the sul2, aph(3ʹ’)-Ib and aph(6)-Id resistance genes were found on the same 4996 bp contig as the IncQ1 plasmid replicon, indicating that they were likely plasmid encoded (Fig. S2). The organization of this replicon and resistance genes is identical to that of the broad-host range plasmid pRSF100 (Fig. S2) [53,69]. Due to the draft nature of the EAEC E36 genome sequence, it is unclear whether its ESBL genes are plasmid-borne, however, the tetA, tetR and blaTEM-1B genes are located on the same 3907 bp contig (Fig. S2), indicating that they would be co-inherited if plasmid encoded. In contrast, both ESBL genes from EAEC E42 (i.e. blaCTX-M-15, blaTEM-1B) were found on the same large contig as the IncI1-I(γ) plasmid replicon (Fig. S3), indicating that these genes are carried by the same plasmid. Indeed, IncI plasmids, isolated from animals and humans, have been shown to carry both blaCTX-M-15 and blaTEM-1 ESBLs, including the EAEC O104:H4 German outbreak strain C227-11 (Table 2) [58,70–72]. Furthermore, the entire sequence of this contig was 99% identical to a large resistance plasmid, pEC_Bactec (92,970 bp) [72], which was isolated in the U.K. from a horse, and carries a Tn3-like transposon which contains both ESBL genes (Fig. S3).
Virulence genes carried by EAEC strains E36 and E42
Comparison of the E36 and E46 genome sequences with the EAEC 042 chromosome (Figure 2) and pAA2 plasmid (Figure 3) confirmed that both are typical EAEC strains (Table 2), possessing the AggR master regulator, which is very similar to AggR from EAEC strains 042 and 17–2 (Fig. S4) [19,21,67]. The EAEC E36 and 17–2 genomes also encode Aar, which can repress AggR mediated-activation [26,73] (Figure 3 and S4). However, Aar was absent from the genome of EAEC E42.
Figure 2.
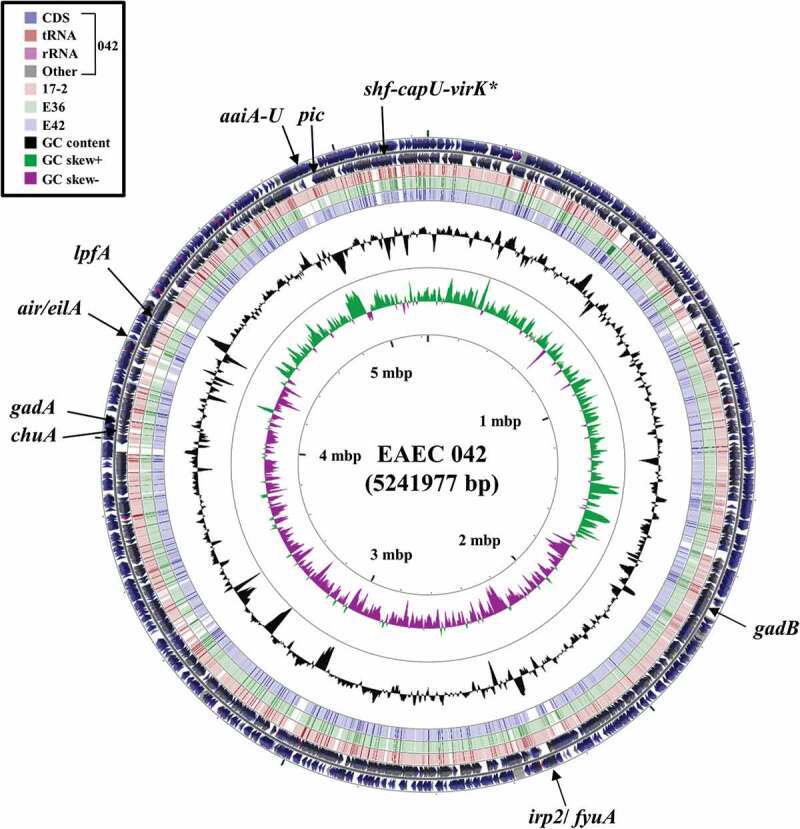
Comparison of the EAEC 042 chromosome with the genomes of EAEC strains 17–2, E36 and E42. The figure shows the comparison of the EAEC 042 chromosome with the genomes of EAEC 17–2, E36 and E42 using GCview (http://stothard.afns.ualberta.ca/cgview_server/) [48]. The outer two rings display the genes and features of the EAEC 042 chromosome (FN5554766.1) on both strands, with selected genes labeled [67]. The brown, green and blue rings illustrate the BLAST results when the genome sequences of EAEC 17–2, E36 and E42, respectively, are compared to the EAEC 042 genome, with shaded regions indicating synteny. The inner two rings display GC content (black) and GC skew (dark green and purple) for the EAEC 042 chromosome. The shf-capU-virK locus is starred as this region is found on both the EAEC 042 chromosome and pAA2 plasmid [67]
Figure 3.
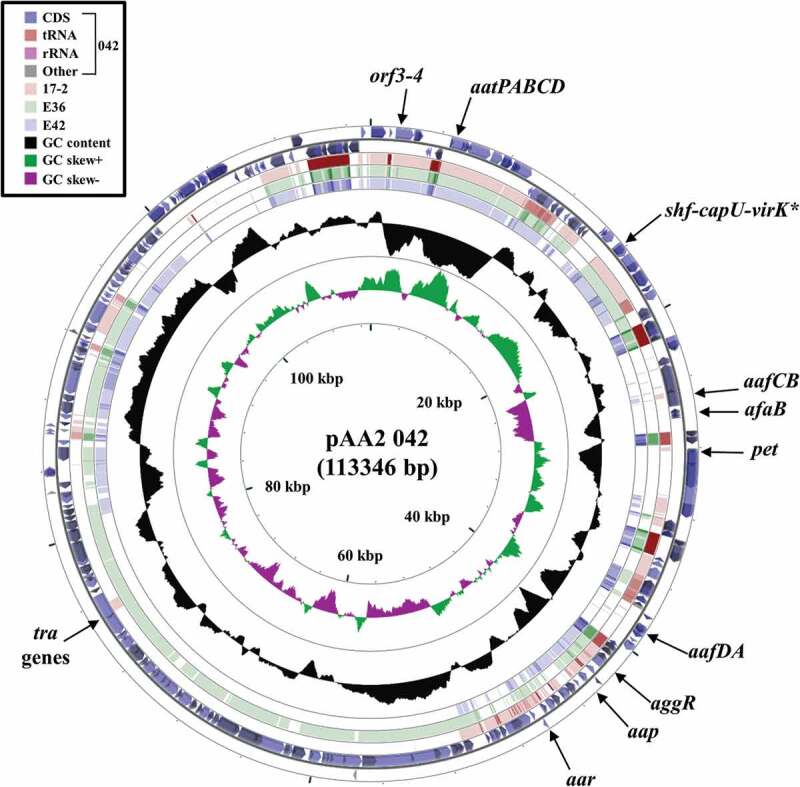
Comparison of the EAEC 042 plasmid pAA2 with the genomes of EAEC strains 17–2, E36 and E42. The figure shows the comparison of the EAEC 042 pAA2 plasmid with the genomes of EAEC 17–2, E36 and E42 using GCview (http://stothard.afns.ualberta.ca/cgview_server/) [48]. The outer two rings display the genes and features of pAA2 (FN554767.1) on both strands, with selected genes labeled [67]. The brown, green and blue rings illustrate BLAST results when the genome sequences of EAEC 17–2, E36 and E42, respectively, are compared to pAA2. The inner two rings display GC content (black) and GC skew (dark green and purple) for pAA. The shf-capU-virK locus is starred as this region is found on both the EAEC 042 chromosome and pAA2 [67]
Both EAEC E36 and E42 possess characteristic EAEC AAF fimbriae genes, which are central to the attachment of EAEC strains to human epithelial cells [10], with E36 carrying AAF/V (agg3DCB-agg5A) and E42 AAF/IV (agg4DCA) type fimbriae (Table 2) [74,75]. Our analysis also confirmed that E36, E42 and 17–2 possess the anti-aggregation protein, dispersin (aap) [22], the Aat T1SS required for dispersin secretion (aatA) [23], orf3 and orf4 that encode proteins that are involved in biofilm formation [24] and capU, encoding a glycosyl transferase that forms an operon with shf and virK in EAEC 042 (Table 2; Figures 2 and 3) [76]. Like EAEC 042, EAEC E42 and 17–2 also carry the genes encoding the Aai type VI secretion system (T6SS) (aaiC) [77]. As all of these genes are induced by AggR in EAEC 042, it is likely that they also form part of the AggR regulon in EAEC E36, E42 and 17–2 [24,27]. Note that, in EAEC E36, all these genes (with the exception of capU) are located on two large contigs, which carried either the IncFIB plasmid replicon and/or plasmid conjugation genes (i.e. tra genes), indicating that they are plasmid-encoded, as is the case for typical EAEC strains [10]. As expected for strain EAEC 17–2, all AggR-regulated genes, with the exception of aaiC and capU, were found on the single large contig relating to the EAEC 17–2 virulence plasmid, pAA.
In addition to potential AggR-regulated virulence determinants, EAEC E36 and E42 both carry air (encoding the enteroaggregative immunoglobulin repeat protein), which is an accessory adhesin of EAEC 042 [78], eilA, which encodes the regulator of air expression [78], various glutamate decarboxylases genes (gadA/gadB) involved in acid resistance [79], and iss/bor, which encodes a lipoprotein involved in increased serum survival [80] (Table 2; Figure 2). Being E. coli phylogenetic group D, like EAEC 042, both E36 and E42 carry the chuA heme-binding protein [67,81], whilst only E36 possesses the Yersiniabactin uptake system (ipr2/fyuA) found in EAEC 042 and 17–2 [67] (Table 2; Figure 2). In addition, EAEC E42 possesses the genes encoding the lpfA (long polar fimbriae) adhesin [82] and the Shigella extracellular protein A autotransporter toxin, sepA [83] (Table 2). Thus, it is clear that both EAEC E36 and E42 possess many genes associated with EAEC pathogenicity, as well as additional ones found in other pathogenic Enterobacteriaceae.
Characterization of AggR-dependent promoters from different EAEC strains
Previously we predicted and characterized many AggR-dependent promoters from EAEC strain 042, identifying both the DNA site for AggR and the associated −10 promoter element [27]. Our studies showed that AggR binds to the consensus sequence 5ʹ-WWWWWWWTATC-3ʹ (where W = A/T) and functional sites are located 21 to 23 bp upstream of the −10 promoter hexamer element, which is recognized by the RNA polymerase σ subunit [27,84]. Comparison of the DNA sequences upstream of the transcript units encoding virulence genes in E36 (aagR, aatP, aar, aap, orf3 and agg3D) and E42 (aagR, aatP, aap, orf3, agg4D and aaiA) (Table 2) identified similar promoters (Fig. S5), suggesting that all these genes formed part of the AggR regulon in these organisms.
To confirm our predictions, we focused on the aatP promoter region, as this promoter has not been previously characterized. During infection, EAEC produce the anti-aggregation protein dispersin (Aap) and its dedicated T1SS (AatPABCD), which aids in the presentation of AAF fimbriae for binding to human epithelial cells [22,23]. Thus, to examine the regulation of the aatP promoter, the DNA upstream of aatP was amplified by PCR from EAEC strains 042, E36 and E42, generating the aatP100 042, aatP100 E36 and aatP100 E42 promoter fragments, respectively (Table S1). Each fragment was cloned into the low copy number lacZ expression vector pRW50 [40] to generate a lac transcriptional fusion and plasmid constructs were transferred into the Δlac E. coli K-12 strain, BW25113. To investigate the effect of AggR, host cells also carried plasmid pBAD/aggR, which encodes AggR from EAEC 042 expressed from an arabinose-inducible promoter, or empty pBAD24 vector (Table S1) [22]. Cells were grown in LB medium to mid-logarithmic phase, either with or without AggR induction by arabinose, and β-galactosidase activities were determined as a proxy for promoter activity. Results illustrated in Figure 4 show that expression from each aatP100 construct was induced by AggR, confirming that the expression of the Aat T1SS in E36 and E42 is AggR-dependent. Note that, for both the E36 and E42 aatP promoters, there is a clear increase in AggR-independent activity (Figure 4), indicating that DNA sequence differences at these promoters affect expression (Fig. S5B). Furthermore, our assignments for the AggR-binding site and the −10 promoter element at the aatP promoter were confirmed by mutational analysis. Thus, point mutations, introduced into the predicted AggR-binding site and −10 element of the 042 aatP promoter fragment greatly decreased AggR-dependent induction (Fig. S6).
Figure 4.
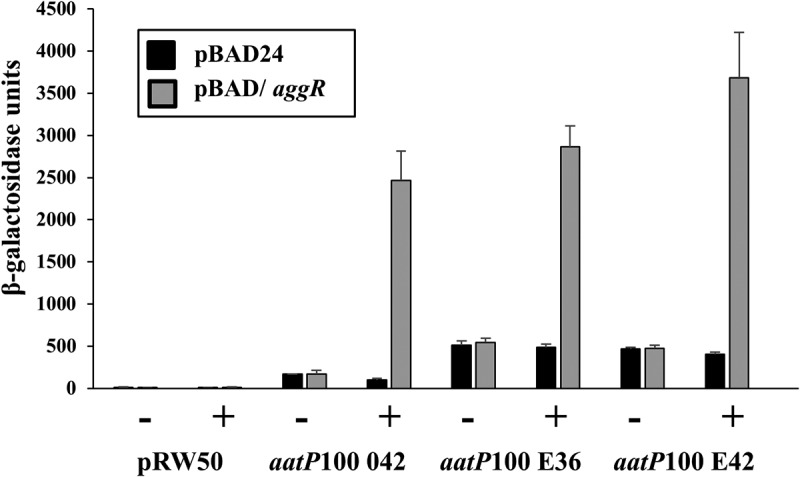
Activation of the aatP promoter from EAEC strains 042, E36 and E42. The figure illustrates measured β-galactosidase activities in E. coli K-12 BW25113 ∆lac cells, containing pRW50 carrying aatP100 promoter fragments from EAEC strains 042, E36 and E42. Cells also carried either pBAD/aggR (gray bars) or pBAD24 (black bars), and were grown in LB medium with (+) or without (-) 0.2% arabinose. β-galactosidase activities are expressed as nmol of ONPG hydrolyzed min−1 mg−1 dry cell mass. Each activity is the average of three independent determinations and standard deviations are shown for all data points
Since AggR and Aar proteins are central to the regulation of EAEC virulence, we chose to examine the aggR and aar promoters in more detail. AggR expression in EAEC 042 is auto-activated [25], and alignment of the DNA upstream of aggR from EAEC E36, E42 and 17–2, with that from EAEC 042 [25], revealed polymorphisms in both the AggR-binding sites and the promoter −10 elements (Fig. S5). Hence, aggR promoter DNA from EAEC E36 and E42 was amplified using PCR to generate the aggR90 E36 and aggR90 E42 promoter fragments, which were cloned to generate plasmid-borne lac transcriptional fusions (Table S1). As controls, similar promoter fragments were generated from EAEC 042 and 17–2 (i.e. aggR90 042 and aggR90 17–2). Each construct was then transferred into BW25113 cells, carrying either pBAD/aggR or pBAD24. β-galactosidase activities were measured, as before and data in Figure 5 show that expression from each aggR promoter fragment was substantially increased by arabinose in cells containing pBAD/aggR, but not with pBAD24 alone. This indicates that each fragment carries an AggR-activated promoter. Note that the basal AggR-independent expression differs from promoter to promoter, with the aggR promoter from EAEC 17–2 showing the highest background levels (Figure 5).
Figure 5.
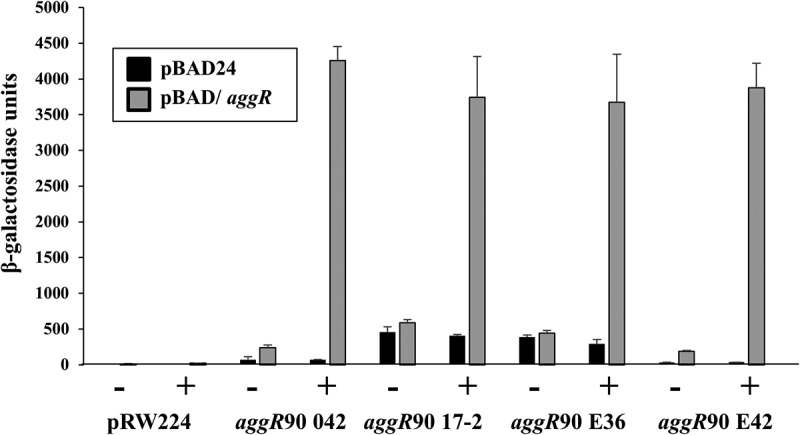
Activation of the aggR promoter from EAEC strains 042, 17–2, E36 and E42. The figure illustrates measured β-galactosidase activities in E. coli K-12 BW25113 ∆lac cells, containing pRW224 carrying aggR90 promoter fragments from EAEC strains 042, 17–2, E36 and E42. Cells also carried either pBAD/aggR (gray bars) or pBAD24 (black bars) and were grown in LB medium in the presence (+) or absence (-) of 0.2% arabinose. β-galactosidase activities are expressed as nmol of ONPG hydrolyzed min−1 mg−1 dry cell mass. Each activity is the average of three independent determinations and standard deviations are shown for all data points
Although expression of the aar repressor protein has been examined by transcriptomic analysis, the aar promoter region had not been characterized [24,27] (Fig. S5). Therefore, the DNA upstream of aar was amplified from EAEC strains 042 and E36, to generate the aar100 042 and aar100 E36 promoter fragments, which were then cloned to generate lac transcriptional fusions. The β-galactosidase activity of BW25113 cells, carrying these constructs, with pBAD/aggR or pBAD24, was then measured and data in Figure 6 show that promoter activity from both aar promoters is increased by AggR, particularly for EAEC E36. Interestingly, for both aar constructs substantial expression is observed in the absence of AggR. To confirm our promoter predictions, point mutations were introduced into the AggR-binding site and −10 promoter element of the aar100 042 promoter fragment. Fig. S7 shows that these substitutions substantially decrease AggR-dependent and AggR-independent promoter activity, confirming our assignment of the key promoter elements controlling aar expression.
Figure 6.
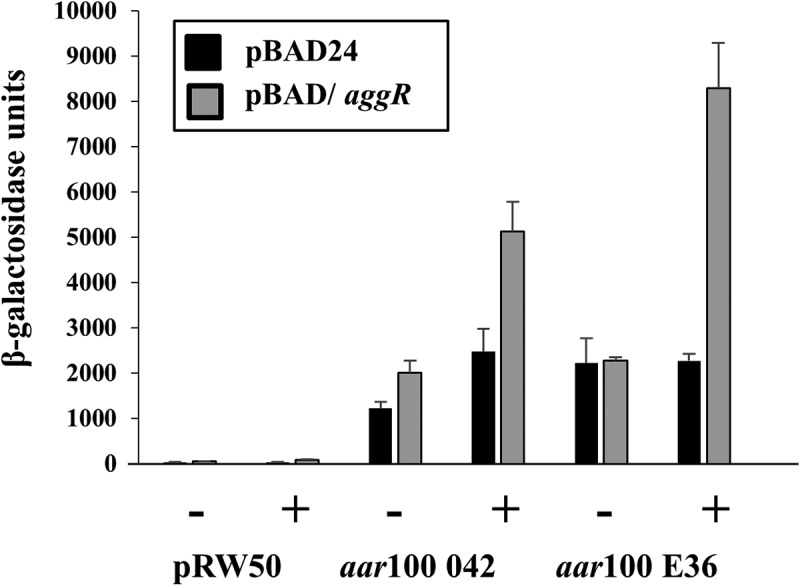
Activity of the aar promoter from EAEC strains 042 and E36. The figure illustrates measured β-galactosidase activities in E. coli K-12 BW25113 ∆lac cells, containing pRW50 carrying aar100 promoter fragments from EAEC strains 042 and E36. Cells also carried either pBAD/aggR (gray bars) or pBAD24 (black bars) and were grown in LB medium in the presence (+) or absence (-) of 0.2% arabinose. β-galactosidase activities are expressed as nmol of ONPG hydrolyzed min−1 mg−1 dry cell mass. Each activity is the average of three independent determinations and standard deviations are shown for all data points
The effect of Aar expression on EAEC biofilm formation
A characteristic of EAEC strains is their capacity to form biofilms on epithelial cells or abiotic surfaces [85,86]. Therefore, we investigated the ability of EAEC strains E36, E42, 042 and 17–2 to form biofilms on plastic microtitre plates when grown in DMEM high glucose medium, which induces AggR-mediated biofilm formation [5,24,27,85]. Results presented in Figure 7a show that EAEC E36 and E42, along with EAEC 17–2, produce biofilm, but to a lesser extent than EAEC 042. Such differences have been observed before for different EAEC isolates [5,85] and may reflect the ability of different AAF fimbrial types to adhere to abiotic surfaces.
Figure 7.
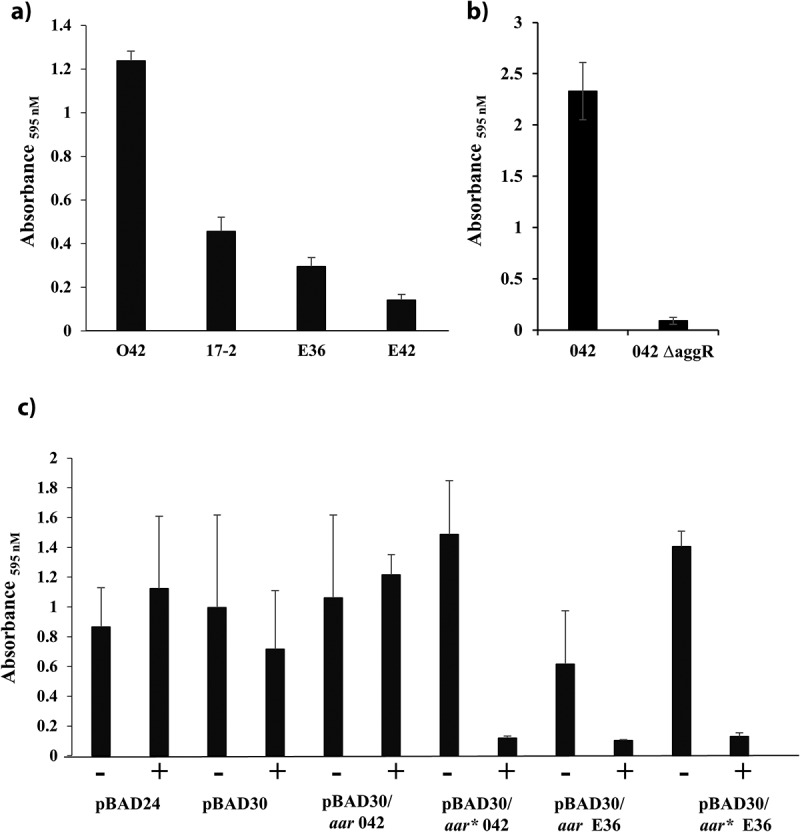
Expression of Aar inhibits biofilm formation in EAEC strain 042. A) The panel shows crystal violet biofilm formation assays, which monitor the ability of EAEC strains 042, 17–2, E36 and E42 to form biofilms on plastic microtitre plates, when grown in DMEM high glucose. B) The panel shows biofilm formation assays, monitoring the ability of EAEC 042 and EAEC 042 ΔaggR to form biofilms on plastic microtitre plates, when grown in DMEM high glucose. C) The panel shows the biofilm formed by EAEC strain 042 carrying either pBAD24, pBAD30, pBAD30/aar 042, pBAD30/aar* 042, pBAD30/aar E36 or pBAD30/aar* E36. Cells were grown in DMEM high glucose in presence (+) or absence (-) of 0.2% (w/v) arabinose. In all panels, the data displayed are representative experiments, with each value being an average of eight replicate samples
Biofilm formation in EAEC 042 is dependent on AggR and the induction of the AggR-regulon (Figure 7b) [85,86]. Thus, to investigate the effect of Aar overexpression on biofilm formation, DNA fragments, carrying the aar coding sequence from EAEC strains 042 and E36, were amplified and cloned into the arabinose inducible vector pBAD30 (Table S1; Fig. S8) to generate pBAD30/aar 042 and pBAD30/aar E36. Both constructs were transferred into EAEC 042 and the biofilm assays were repeated with arabinose added to induce Aar expression. Results in Figure 7c show that induction of E36 Aar decreased EAEC 042 biofilm production, whilst induction of 042 Aar expression had little or no effect. Comparison of the E36 and 042 aar sequences (Figs. S8A and B) identified the ribosome binding site (RBS) as a likely cause of the difference, since the 042 aar RBS corresponds poorly to the consensus. Hence we replaced the aar RBS for both the 042 and the E36 genes with the strong RBS from the pET20b expression vector to generate the pBAD30/aar* 042 and pBAD30/aar* E36 vectors (Table S1; Fig. S8). Biofilm formation of EAEC 042, carrying these new constructs, was then assayed, as before. Results in Figure 7c show that arabinose induction of both 042 and E36 Aar, carrying the improved RBS, completely disrupts biofilm formation. Thus, we conclude that the inability of pBAD30/aar 042 to interfere with biofilm formation is likely due to the poor RBS of the EAEC 042 aar gene, resulting in lower levels of Aar.
Direct effects of Aar on expression from AggR-dependent promoters
As Aar inhibits transcription activation by AggR [26,73], we used our simple two-plasmid system to examine the direct effect of Aar on AggR-dependent activation at various EAEC promoters. To achieve this, we took the pBAD/aggR plasmid (which carries EAEC 042 aggR) and cloned the DNA encoding either EAEC 042 or E36 aar downstream of aggR, to generate a synthetic operon containing both genes (i.e. pBAD/aggR/aar 042 and pBAD/aggR/aar E36) (Fig. S8). We also made constructs, which carry the strong RBS from pET20b (i.e. pBAD/aggR/aar* 042 and pBAD/aar* E36) (Fig. S8). These plasmids, as well as pBAD/aggR, were transferred into BW25113 cells, which carried a fusion of the aggR promoter from EAEC 042 or E36 fused to lacZ, as before. Cells were grown in LB medium, with 0.2% arabinose included to induce co-expression of AggR and Aar or expression of AggR alone. Surprisingly, results in Figure 8 show that AggR-dependent expression from the 042 aggR promoter was only marginally inhibited by Aar expression, whilst that from the E36 aggR promoter was completely unaffected, regardless of the aar-containing construct used. As this may be a peculiarity of the aggR promoter, we also examined expression from the AggR-dependent fimbrial promoters, afaB100 and agg4D100 [27]. Results in Figure 8 show that induction of Aar expression only decreased AggR-dependent activity from each promoter by ~2-fold. Thus, we conclude that, although Aar expression has a considerable effect on biofilm formation, it only has a minor direct effect on expression from AggR-dependent promoters.
Figure 8.
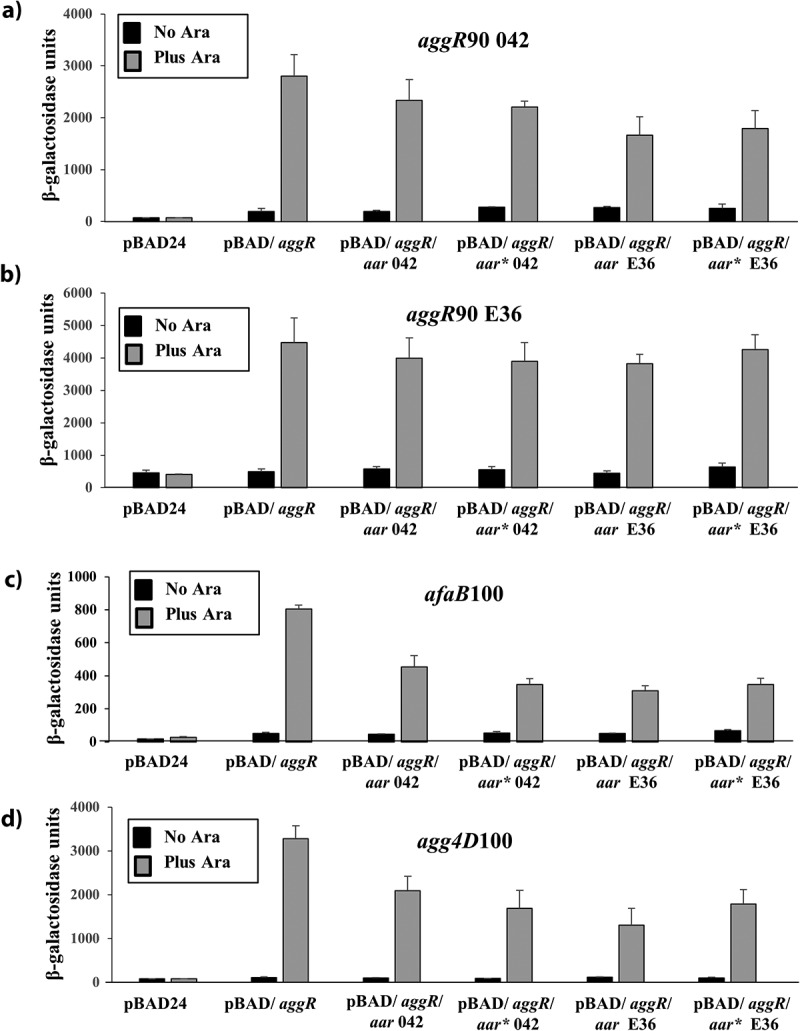
The effects of Aar expression on AggR-dependent activation of target promoters. The figure shows β-galactosidase activities measured in E.coli K-12 BW25113 cells, containing lacZ expression vectors carrying either the A) aggR90 042, B) aggR90 E36, C) afaB100 or D) agg4D100 promoter fragments. In panels A) and B), promoter fragments were cloned into pRW224 and in C) and D), they were cloned into pRW50. Cells also carry either pBAD24, pBAD/aggR, pBAD/aggR/aar 042, pBAD/aggR/aar* 042, pBAD/aggR/aar E36 or pBAD/aggR/aar* E36. Cells were grown in LB medium in the presence or absence of 0.2% (w/v) arabinose. β-galactosidase activities are expressed as nmol of ONPG hydrolyzed min−1 mg−1 dry cell mass. Each activity is the average of three independent determinations and standard deviations are shown for all data points
Conclusions
Since their discovery, the gold standard for characterizing EAEC has been the ability of cells to adhere to HEp-2 cells in a “stacked brick” aggregative (AA) phenotype [87], with typical EAEC strains possessing aggR and atypical strains lacking it [20]. Recently, typical EAEC strains have been further defined molecularly as E. coli strains carrying aggR and a complete AAF gene cluster or a functional CS22 ETEC colonization factor operon [88]. Thus, it is clear that AggR-dependent activation of virulence determinant expression is the hallmark of typical EAEC strains. Our data from two new EAEC strains from Assiut, Egypt, suggest that a feed-forward activation mechanism is shared between strains from different sources. The mechanism, illustrated in (Figure 9) suggests that virulence, triggered either by a specific signal or by random fluctuations in AggR levels, follows this feed-forward activation of AggR expression. In any strain, the probability of switching into the virulent state will depend on the “basal” level of AggR, which will depend on AggR-independent activity of the aggR promoter, and also Aar levels, which, in turn, depend on AggR-independent aar promoter expression, and also the translation of Aar from the aar gene messenger. Our data suggest that these parameters vary from one strain to another and this may well explain the prevalence of EAEC in asymptomatic individuals: hence, the onset of virulence could well be a rare event. Note that aar is not present in all EAEC strains and appears to be an elaboration to the feed-forward mechanism, operating ubiquitously both pre – and post-triggering. Boisen et al [88]. recently presented the most comprehensive survey to-date of genome sequences from nearly 100 E. coli strains, collected from diverse geographic settings. Our findings, reported here, are consistent with the view of Boisen et al. [88] of ongoing random shuffling and redistribution of different genetic determinants, which results in massive mosaic variation in EAEC strains.
Figure 9.
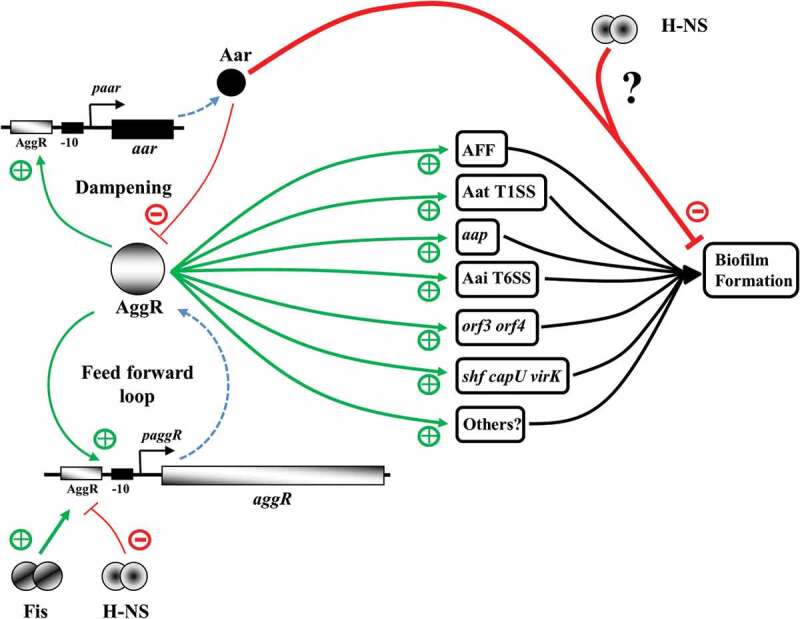
The AggR-dependent virulence switch: a feed forward loop with dampening. In the absence of triggering, AggR-independent expression from the aggR and aar promoters maintains a balanced level of AggR and Aar, keeping the virulence switch in an off state. AggR expression is controlled positively (+) by Fis and repressed (-) by H-NS [25]. Triggering of the AggR-dependent virulence switch, either by a specific signal or a stochastic event, leads to the increased expression of AggR, amplifying the response (i.e. a feed forward loop), and the production of specific virulence determinants involved in host attachment, biofilm formation and protein secretion. Aar expression is also increased, inhibiting AggR-dependent transcription activation, dampening the AggR feed forward loop, and interfering with biofilm formation, possibly in concert with H-NS at specific promoters. The virulence switch will be reset by the removal of the triggering signal and Aar-mediated repression of AggR. Green arrows denote activation (+), red lines repression (-), and dotted blue arrows indicate the processes of transcription and translation for aggR and aar.
Here, we isolated Escherichia coli strains from infants and children, presenting with diarrhea at the outpatients clinic of Assiut University Children’s Hospital. Worryingly, all isolates were resistant to multiple classes of antibiotics and four were resistant to all antibiotics tested, highlighting the possibility that treatment with frontline antibiotics, if necessary, would likely fail to resolve many of these infections. Furthermore, all isolates were resistant to the cephalosporin antibiotics, cefaclor and ceftriaxone, suggesting that the carriage of extended spectrum β-lactamases (ESBL) is wide spread [89,90]. In support of this, genome analysis of E36 and E42 indicated that each strain carried two types of ESBLs (i.e. blaCTX-M and blaTEM) (Table 2). We also detected other antibiotic resistance genes carried by the E36 and E42 strains (Table 2) and these likely account for their multidrug resistance phenotype. Interestingly, E36 was one of the few isolates resistant to the carbapenem antibiotics, imipenem and meropenem, (Table 1 and S1), and yet we did not detect any obvious carbapenem resistance genes in its genome. As carbapenem resistance can be due to a variety of mechanisms [90,91], it is possible that we failed to detect such genes due to the draft nature of our genome assemblies. Further studies will be required to determine the mechanism of carbapenem resistance in this strain.
In many studies, EAEC strains have been isolated from asymptomatic individuals [5,16]. Like E36, E42 and many pathogenic EAEC strains, 17–2 possesses a battery of AggR-regulated virulence determinants and toxins, such as the EAST-1 toxin (astA) [30] and the Sat secreted autotransporter toxin (Table 2) [92]. EAEC 17–2 belongs to the ST10 clonal complex, which is a major sequence type complex associated with EAEC and linked to disease causation [64]. Thus, EAEC 17–2 possesses all the characteristics of a pathogenic EAEC strain and yet, in a volunteer study, 17–2 failed to elicit diarrhea in adult volunteers [29] and, in another, only one adult out of 19 produced diarrhea [65]. Like our Egyptian EAEC strains, EAEC 17–2 was isolated from an infant with diarrhea [93]. As immune protection to EAEC strains, such as 17–2, may be acquired early in life, it has been suggested that virulence should not be determined in adult volunteers [64,65]. Thus, it is clear that, for EAEC infections, many factors contribute to whether disease occurs, such as bacterial genotype and, importantly, the immune status, nutritional status and physical status of the infected individual [29]. It is plausible that disease is only observed in naive individuals, or when a new or particularly potent combination of virulence genes occurs, as was observed for the Shiga-toxin-producing EAEC O104:H4 outbreak strain in Germany (Table 2) [12–16,58]. Given the importance of the precise levels of AggR and Aar in cells [28], we suggest that stochastic cell-to-cell variation influences infectivity as much as genotype.
Supplementary Material
Acknowledgments
This work was generously supported by a studentship from the Egyptian Ministry of Higher Education (Cultural Affairs and Missions Sector) and the Grant Office from the Medical School, Assiut University to RA, a Darwin Trust of Edinburgh PhD studentship to MY and BBSRC research grant BB/R017689/1 to DFB and SJWB. We thank MicrobesNG and Emily Jane Richardson for sequencing, genome annotation and helpful suggestions for the analysis of this data.
Funding Statement
This work was supported by the Biotechnology and Biological Sciences Research Council [BB/R017689/1]; Egyptian Ministry of Higher Education (Cultural Affairs and Missions Sector) and the Grant Office from the Medical School, Assiut University [No Number]; Darwin Trust of Edinburgh PhD studentship [No Number].
Author Contributions
DFB, SJWB, EA, EH, NHA and ED conceived and designed the research project. RA, MY, SJE, GSC, FS, REG, NHA and DFB performed the experiments. DFB, SJWB and RA analysed the data and wrote the manuscript with input from all authors.
Disclosure statement
The authors declare no conflict of interest.
Supplementary material
Supplemental data for this article can be accessed here.
References
- [1].Kaper JB, Nataro JP, Mobley HL.. Pathogenic Escherichia coli. Nat Rev Microbiol. 2004;2(2):123–140. [DOI] [PubMed] [Google Scholar]
- [2].Nataro JP, Mai V, Johnson J, et al. Diarrheagenic Escherichia coli infection in Baltimore, Maryland, and New Haven, Connecticut. Clin Infect Dis. 2006;43:402–407. [DOI] [PubMed] [Google Scholar]
- [3].Wilson A, Evans J, Chart H, et al. Characterisation of strains of enteroaggregative Escherichia coli isolated during the infectious intestinal disease study in England. Eur J Epidemiol. 2001;17(12):1125–1130. [DOI] [PubMed] [Google Scholar]
- [4].Okeke IN, Lamikanra A, Czeczulin J, et al. Heterogeneous virulence of Enteroaggregative Escherichia coli strains isolated from children in Southwest Nigeria. J Infect Dis. 2000;181(1):252–260. [DOI] [PubMed] [Google Scholar]
- [5].Franca FL, Wells TJ, Browning DF, et al. Genotypic and phenotypic characterisation of enteroaggregative Escherichia coli from children in Rio de Janeiro, Brazil. PLoS One. 2013;8:e69971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Adachi JA, Jiang ZD, Mathewson JJ, et al. Enteroaggregative Escherichia coli as a major Etiologic agent in Traveler’s Diarrhea in 3 regions of the world. Clin Infect Dis. 2001;32(12):1706–1709. . [DOI] [PubMed] [Google Scholar]
- [7].Durrer P, Zbinden R, Fleisch F, et al. Intestinal infection due to Enteroaggregative Escherichia coli among human immunodeficiency virus–infected persons. J Infect Dis. 2000;182(5):1540–1544. . [DOI] [PubMed] [Google Scholar]
- [8].Olesen B, Scheutz F, Andersen RL, et al. Enteroaggregative Escherichia coli O78:H10, the cause of an outbreak of urinary tract infection. J Clin Microbiol. 2012;50(11):3703–3711. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Herzog K, Engeler Dusel J, Hugentobler M, et al. Diarrheagenic enteroaggregative Escherichia coli causing urinary tract infection and bacteremia leading to sepsis. Infection. 2014;42(2):441–444. . [DOI] [PubMed] [Google Scholar]
- [10].Harrington SM, Dudley EG, Nataro JP. Pathogenesis of enteroaggregative Escherichia coli infection. FEMS Microbiol Lett. 2006;254(1):12–18. [DOI] [PubMed] [Google Scholar]
- [11].Steiner TS, Lima AA, Nataro JP, et al. Enteroaggregative Escherichia coli produce intestinal inflammation and growth impairment and cause interleukin-8 release from intestinal epithelial cells. J Infect Dis. 1998;177(1):88–96. [DOI] [PubMed] [Google Scholar]
- [12].Itoh Y, Nagano I, Kunishima M, et al. Laboratory investigation of enteroaggregative Escherichia coli O untypeable:H10 associated with a massive outbreak of gastrointestinal illness. J Clin Microbiol. 1997;35(10):2546–2550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Harada T, Hiroi M, Kawamori F, et al. A food poisoning diarrhea outbreak caused by enteroaggregative Escherichia coli serogroup O126: h27in Shizuoka, Japan. Jpn J Infect Dis. 2007;60:154–155. [PubMed] [Google Scholar]
- [14].Frank C, Werber D, Cramer JP, et al. Epidemic profile of Shiga-toxin–producing Escherichia coli O104:H4 outbreak in Germany. N Engl J Med. 2011;365(19):1771–1780. . [DOI] [PubMed] [Google Scholar]
- [15].Boisen N, Melton-Celsa AR, Scheutz F, et al. Shiga toxin 2a and Enteroaggregative Escherichia coli – a deadly combination. Gut Microbes. 2015;6(4):272–278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Estrada-Garcia T, Navarro-Garcia F. Enteroaggregative Escherichia coli pathotype: a genetically heterogeneous emerging foodborne enteropathogen. FEMS Immunol Med Microbiol. 2012;66(3):281–298. [DOI] [PubMed] [Google Scholar]
- [17].Henderson IR, Hicks S, Navarro-Garcia F, et al. Involvement of the EnteroaggregativeEscherichia coli plasmid-encoded toxin in causing human intestinal damage. Infect Immun. 1999;67(10):5338–5344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Savarino SJ, Fasano A, Robertson DC, et al. Enteroaggregative Escherichia coli elaborate a heat-stable enterotoxin demonstrable in an in vitro rabbit intestinal model. J Clin Invest. 1991;87(4):1450–1455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Nataro JP, Yikang D, Yingkang D, et al. AggR, a transcriptional activator of aggregative adherence fimbria I expression in enteroaggregative Escherichia coli. J Bacteriol. 1994;176(15):4691–4699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Sarantuya J, Nishi J, Wakimoto N, et al. Typical enteroaggregative Escherichia coli is the most prevalent pathotype among E. coli strains causing diarrhea in Mongolian children. J Clin Microbiol. 2004;42(1):133–139. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Elias WP Jr., Czeczulin JR, Henderson IR, et al. Organization of biogenesis genes for aggregative adherence fimbria II defines a virulence gene cluster in enteroaggregative Escherichia coli. J Bacteriol. 1999;181(6):1779–1785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Sheikh J, Czeczulin JR, Harrington S, et al. A novel dispersin protein in enteroaggregative Escherichia coli. J Clin Invest. 2002;110(9):1329–1337. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Nishi J, Sheikh J, Mizuguchi K, et al. The export of coat protein from Enteroaggregative Escherichia coli by a specific ATP-binding cassette transporter system. J Biol Chem. 2003;278(46):45680–45689. . [DOI] [PubMed] [Google Scholar]
- [24].Morin N, Santiago AE, Ernst RK, et al. Characterization of the AggR regulon in Enteroaggregative Escherichia coli. Infect Immun. 2013;81(1):122–132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Morin N, Tirling C, Ivison SM, et al. Autoactivation of the AggR regulator of enteroaggregative Escherichia coli in vitro and in vivo. FEMS Immunol Med Microbiol. 2010;58(3):344–355. [DOI] [PubMed] [Google Scholar]
- [26].Santiago AE, Ruiz-Perez F, Jo NY, et al. A large family of antivirulence regulators modulates the effects of transcriptional activators in gram-negative pathogenic bacteria. PLoS Pathog. 2014;10:e1004153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Yasir M, Icke C, Abdelwahab R, et al. Organization and architecture of AggR-dependent promoters from enteroaggregative Escherichia coli. Mol Microbiol. 2019;111(2):534–551. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Mickey AS, Nataro JP. Dual function of Aar, a member of the new AraC negative regulator family, in Escherichia coli gene expression. Infect Immun. 2020;88(6). DOI: 10.1128/IAI.00100-20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Nataro JP, Deng Y, Cookson S, et al. Heterogeneity of enteroaggregative Escherichia coli virulence demonstrated in volunteers. J Infect Dis. 1995;171(2):465–468. . [DOI] [PubMed] [Google Scholar]
- [30].Veilleux S, Holt N, Schultz BD, et al. Escherichia coli EAST1 toxin toxicity of variants 17-2 and O 42. Comp Immunol Microbiol Infect Dis. 2008;31:567–578. [DOI] [PubMed] [Google Scholar]
- [31].Zhang R, Gu DX, Huang YL, et al. Comparative genetic characterization of Enteroaggregative Escherichia coli strains recovered from clinical and non-clinical settings. Sci Rep. 2016;6(1):24321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Guiral E, Mendez-Arancibia E, Soto SM, et al. CTX-M-15–producing Enteroaggregative Escherichia coli as cause of travelers’ Diarrhea. Emerg Infect Dis. 2011;17(10):1950–1953. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Chattaway MA, Jenkins C, Ciesielczuk H, et al. Evidence of evolving Extraintestinal Enteroaggregative Escherichia coli ST38 Clone. Emerg Infect Dis. 2014;20(11):1935–1937. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Ali MM, Ahmed SF, Klena JD, et al. Enteroaggregative Escherichia coli in diarrheic children in Egypt: molecular characterization and antimicrobial susceptibility. J Infect Dev Ctries. 2014;8(05):589–596. [DOI] [PubMed] [Google Scholar]
- [35].Sanders JW, Putnam SD, Gould P, et al. Diarrheal illness among deployed U.S. military personnel during Operation Bright Star 2001–Egypt. Diagn Microbiol Infect Dis. 2005;52(2):85–90. . [DOI] [PubMed] [Google Scholar]
- [36].Riddle MS, Halvorson HA, Shiau D, et al. Acute gastrointestinal infection, respiratory illness, and noncombat injury among US military personnel during Operation Bright Star 2005, in Northern Egypt. J Travel Med. 2007;14(6):392–401. . [DOI] [PubMed] [Google Scholar]
- [37].Meheissen M, Header D, Abdelaty K. Phylogenetic and pathotype analysis of Escherichia coli stool isolates from Egyptian patients with inflammatory bowel disease. Germs. 2019;9(4):172–181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Ahmed SF, Shaheen HI, Abdel-Messih IA, et al. The epidemiological and clinical characteristics of diarrhea associated with enteropathogenic, enteroaggregative and diffuse-adherent Escherichia coli in Egyptian children. J Trop Pediatr. 2014;60(5):397–400. . [DOI] [PubMed] [Google Scholar]
- [39].Sambrook J, Russell DW. Molecular cloning: a laboratory manual. 3rd ed. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 2001. [Google Scholar]
- [40].Lodge J, Fear J, Busby S, et al. Broad host range plasmids carrying the Escherichia coli lactose and galactose operons. FEMS Microbiol Lett. 1992;74(2–3):271–276. [DOI] [PubMed] [Google Scholar]
- [41].Islam MS, Bingle LE, Pallen MJ, et al. Organization of the LEE1 operon regulatory region of enterohaemorrhagic Escherichia coli O157: H7 O157:H7 and activation by GrlA. Mol Microbiol. 2011;79(2):468–483. [DOI] [PubMed] [Google Scholar]
- [42].Bauer AW, Kirby WM, Sherris JC, et al. Antibiotic susceptibility testing by a standardized single disk method. Am J Clin Pathol. 1966;45:493–496. [PubMed] [Google Scholar]
- [43].Clinical_and_Laboratory_Standards_Institute_(CLSI) . Performance standards for antimicrobial susceptibility testing; Twenty-Fourth informational supplement, CLSI Document M100-S24, Wayne. 2014;34(1) [Google Scholar]
- [44].Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. 2014;30(15):2114–2120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Wick RR, Judd LM, Gorrie CL, et al. Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput Biol. 2017;13(6):e1005595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014;30(14):2068–2069. [DOI] [PubMed] [Google Scholar]
- [47].Rutherford K, Parkhill J, Crook J, et al. Artemis: sequence visualization and annotation. Bioinformatics. 2000;16(10):944–945. . [DOI] [PubMed] [Google Scholar]
- [48].Grant JR, Stothard P. The CGView Server: a comparative genomics tool for circular genomes. Nucleic Acids Res. 2008;36(Web Server):W181–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Carver TJ, Rutherford KM, Berriman M, et al. ACT: the Artemis Comparison Tool. Bioinformatics. 2005;21(16):3422–3423. [DOI] [PubMed] [Google Scholar]
- [50].Carver T, Thomson N, Bleasby A, et al. DNAPlotter: circular and linear interactive genome visualization. Bioinformatics. 2009;25(1):119–120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Larsen MV, Cosentino S, Rasmussen S, et al. Multilocus sequence typing of total-genome-sequenced bacteria. J Clin Microbiol. 2012;50(4):1355–1361. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Joensen KG, Tetzschner AM, Iguchi A, et al. Rapid and easy in Silico serotyping of Escherichia coli isolates by use of whole-genome sequencing data. J Clin Microbiol. 2015;53(8):2410–2426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].Carattoli A, Zankari E, Garcia-Fernandez A, et al. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother. 2014;58:3895–3903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Zankari E, Hasman H, Cosentino S, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012;67:2640–2644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Joensen KG, Scheutz F, Lund O, et al. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J Clin Microbiol. 2014;52(5):1501–1510. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Zhang Z, Schwartz S, Wagner L, et al. A greedy algorithm for aligning DNA sequences. J Comput Biol. 2000;7(1–2):203–214. [DOI] [PubMed] [Google Scholar]
- [57].Dunne KA, Chaudhuri RR, Rossiter AE, et al. Sequencing a piece of history: complete genome sequence of the original Escherichia coli strain. Microb Genom. 2017;3:mgen000106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Rasko DA, Webster DR, Sahl JW, et al. Origins of the E. coli strain causing an outbreak of Hemolytic–Uremic Syndrome in Germany. N Engl J Med. 2011;365(8):709–717. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Page AJ, Cummins CA, Hunt M, et al. Roary: rapid large-scale prokaryote pan genome analysis. Bioinformatics. 2015;31(22):3691–3693. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30(9):1312–1313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Sarkar G, Sommer SS. The “megaprimer” method of site-directed mutagenesis. Biotechniques. 1990;8:404–407. [PubMed] [Google Scholar]
- [62].Guzman LM, Belin D, Carson MJ, et al. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J Bacteriol. 1995;177(14):4121–4130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Browning DF, Lee DJ, Wolfe AJ, et al. The Escherichia coli K-12 NarL and NarP proteins insulate the nrf promoter from the effects of integration host factor. J Bacteriol. 2006;188(21):7449–7456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Okeke IN, Wallace-Gadsden F, Simons HR, et al. Multi-locus sequence typing of enteroaggregative Escherichia coli isolates from Nigerian children uncovers multiple lineages. PLoS One. 2010;5(11):e14093. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Nataro JP, Deng Y, Maneval DR, et al. Aggregative adherence fimbriae I of enteroaggregative Escherichia coli mediate adherence to HEp-2 cells and hemagglutination of human erythrocytes. Infect Immun. 1992;60(6):2297–2304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Villa L, Garcia-Fernandez A, Fortini D, et al. Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J Antimicrob Chemother. 2010;65(12):2518–2529. [DOI] [PubMed] [Google Scholar]
- [67].Chaudhuri RR, Sebaihia M, Hobman JL, et al. Complete genome sequence and comparative metabolic profiling of the prototypical enteroaggregative Escherichia coli strain 042. PLoS One. 2010;5(1):e8801. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Ho PL, Lo WU, Lai EL, et al. Clonal diversity of CTX-M-producing, multidrug-resistant Escherichia coli from rodents. J Med Microbiol. 2015;64(2):185–190. . [DOI] [PubMed] [Google Scholar]
- [69].Scholz P, Haring V, Wittmann-Liebold B, et al. Complete nucleotide sequence and gene organization of the broad-host-range plasmid RSF1010. Gene. 1989;75:271–288. [DOI] [PubMed] [Google Scholar]
- [70].Timofte D, Maciuca IE, Evans NJ, et al. Detection and molecular characterization of Escherichia coli CTX-M-15 and Klebsiella pneumoniae SHV-12 beta-lactamases from bovine mastitis isolates in the United Kingdom. Antimicrob Agents Chemother. 2014;58(2):789–794. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Zhang L, Lu X, Zong Z. The emergence of blaCTX-M-15-carrying Escherichia coli of ST131 and new sequence types in Western China. Ann Clin Microbiol Antimicrob. 2013;12(1):35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [72].Smet A, Van Nieuwerburgh F, Vandekerckhove TT, et al. Complete nucleotide sequence of CTX-M-15-plasmids from clinical Escherichia coli isolates: insertional events of transposons and insertion sequences. PLoS One. 2010;5(6):e11202. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Santiago AE, Yan MB, Tran M, et al. A large family of anti-activators accompanying XylS/AraC family regulatory proteins. Mol Microbiol. 2016;101:314–332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Jonsson R, Struve C, Boisen N, et al. Novel aggregative adherence fimbria variant of enteroaggregative Escherichia coli. Infect Immun. 2015;83(4):1396–1405. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Boisen N, Struve C, Scheutz F, et al. New adhesin of enteroaggregative Escherichia coli related to the Afa/Dr/AAF family. Infect Immun. 2008;76(7):3281–3292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [76].Fujiyama R, Nishi J, Imuta N, et al. The shf gene of a Shigella flexneri homologue on the virulent plasmid pAA2 of enteroaggregative Escherichia coli 042 is required for firm biofilm formation. Curr Microbiol. 2008;56:474–480. [DOI] [PubMed] [Google Scholar]
- [77].Dudley EG, Thomson NR, Parkhill J, et al. Proteomic and microarray characterization of the AggR regulon identifies a pheU pathogenicity island in enteroaggregative Escherichia coli. Mol Microbiol. 2006;61:1267–1282. [DOI] [PubMed] [Google Scholar]
- [78].Sheikh J, Dudley EG, Sui B, et al. EilA, a HilA-like regulator in enteroaggregative Escherichia coli. Mol Microbiol. 2006;61(2):338–350. [DOI] [PubMed] [Google Scholar]
- [79].Lund P, Tramonti A, De Biase D. Coping with low pH: molecular strategies in neutralophilic bacteria. FEMS Microbiol Rev. 2014;38:1091–1125. [DOI] [PubMed] [Google Scholar]
- [80].Johnson TJ, Wannemuehler YM, Nolan LK. Evolution of the iss gene in Escherichia coli. Appl Environ Microbiol. 2008;74:2360–2369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of theEscherichia coli Phylogenetic Group. Appl Environ Microbiol. 2000;66(10):4555–4558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Toma C, Higa N, Iyoda S, et al. The long polar fimbriae genes identified in Shiga toxin-producing Escherichia coli are present in other diarrheagenic E. coli and in the standard E. coli collection of reference (ECOR) strains. Res Microbiol. 2006;157(2):153–161. [DOI] [PubMed] [Google Scholar]
- [83].Benjelloun-Touimi Z, Sansonetti PJ, Parsot C. SepA, the major extracellular protein of Shigella flexneri: autonomous secretion and involvement in tissue invasion. Mol Microbiol. 1995;17(1):123–135. [DOI] [PubMed] [Google Scholar]
- [84].Browning DF, Godfrey RE, Richards KL, et al. Exploitation of the Escherichia coli lac operon promoter for controlled recombinant protein production. Biochem Soc Trans. 2019;47:755–763. [DOI] [PubMed] [Google Scholar]
- [85].Sheikh J, Hicks S, Dall’Agnol M, et al. Roles for Fis and YafK in biofilm formation by enteroaggregative Escherichia coli. Mol Microbiol. 2001;41(5):983–997. [DOI] [PubMed] [Google Scholar]
- [86].Boisen N, Hansen AM, Melton-Celsa AR, et al. The Presence of the pAA Plasmid in the German O104: h4Shiga O104:H4 Shiga Toxin Type 2a (Stx2a)-producing Stx2a)–Producing Enteroaggregative Escherichia coli strain promotes the translocation of Stx2a across an Epithelial cell Monolayer. J Infect Dis. 2014;210(12):1909–1919. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].Nataro JP, Kaper JB, Robins-Browne R, et al. Patterns of adherence of diarrheagenic Escherichia coli to HEp-2 cells. Pediatr Infect Dis J. 1987;6(9):829–831. [DOI] [PubMed] [Google Scholar]
- [88].Boisen N, Østerlund MT, Joensen KG, et al. Redefining enteroaggregative Escherichia coli (EAEC): genomic characterization of epidemiological EAEC strains. PLoS Negl Trop Dis. 2020;14(9):e0008613. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [89].Oteo J, Perez-Vazquez M, Campos J. Extended-spectrum [beta]-lactamase β-lactamase producing Escherichia coli: changing epidemiology and clinical impact. Curr Opin Infect Dis. 2010;23(4):320–326. [DOI] [PubMed] [Google Scholar]
- [90].Tooke CL, Hinchliffe P, Bragginton EC, et al. beta-Lactamases and beta-Lactamase Inhibitors in the 21st Century. J Mol Biol. 2019;431(18):3472–3500. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [91].Chetri S, Bhowmik D, Paul D, et al. AcrAB-TolC efflux pump system plays a role in carbapenem non-susceptibility in Escherichia coli. BMC Microbiol. 2019;19:210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Guyer DM, Henderson IR, Nataro JP, et al. Identification of sat, an autotransporter toxin produced by uropathogenic Escherichia coli. Mol Microbiol. 2000;38:53–66. [DOI] [PubMed] [Google Scholar]
- [93].Vial PA, Robins-Browne R, Lior H, et al. Characterization of enteroadherent-aggregative Escherichia coli, a putative agent of diarrheal disease. J Infect Dis. 1988;158(1):70–79. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.