

Physical activity produces analgesia through supraspinal, spinal, and peripheral mechanisms. Currently, no exercise mode has proven to be superior to others for analgesic effects.
Keywords: Pain, Exercise, Hyperalgesia, Analgesia, Animal models, Central mechanisms, Peripheral mechanisms
Abstract:
Physical activity has become a first-line treatment in rehabilitation settings for individuals with chronic pain. However, research has only recently begun to elucidate the mechanisms of exercise-induced analgesia. Through the study of animal models, exercise has been shown to induce changes in the brain, spinal cord, immune system, and at the site of injury to prevent and reduce pain. Animal models have also explored beneficial effects of exercise through different modes of exercise including running, swimming, and resistance training. This review will discuss the central and peripheral mechanisms of exercise-induced analgesia through different modes, intensity, and duration of exercise as well as clinical applications of exercise with suggestions for future research directions.
1. Introduction
Physical inactivity has become a major health concern due to its role in increasing all-cause mortality and promoting conditions such as obesity, cardiovascular disease, diabetes, cancer, dementia, and depression.9,54,87,119,122,129,155,161 Physical inactivity also increases the risk of developing chronic pain, with individuals with lower amounts of physical activity reporting more incidence of musculoskeletal pain.88,89 Furthermore, evidence-based practice guidelines recommend exercise with moderate to strong evidence for a variety of chronic pain conditions such low back pain, osteoarthritis, and fibromyalgia.8,17,18,20,21,126 It is recognized that an acute bout of physical activity or exercise can increase pain in individuals with chronic pain.43,90 Animal models have been developed to model these phenomena, show multiple underlying mechanisms, and have been reviewed elsewhere.92,140 Thus, although an acute bout of exercise can increase pain in individuals with chronic pain and animal models, regular physical activity and exercise can both prevent and alleviate chronic pain.
Although exercise is one of the main treatment approaches for these chronic pain conditions, there are still many questions surrounding exercise that need to be resolved. Specifically, questions still exist regarding appropriate prescription of exercise type, duration, intensity, and volume for chronic pain populations. Similarly, although exercise is typically prescribed for treatment of chronic pain, we do not fully understand the underlying mechanisms of exercise-induced pain relief. Thus, there has been a push to understand the underlying mechanisms of exercise-induced analgesia through use of animal models of exercise, which allow for a better understanding of exercise's effects on the brain, spinal cord, immune system, and periphery to produce analgesia. Elucidating these mechanisms could aid in more effective prescription and dosing of exercise for chronic pain, as well as development of new pharmacological targets for pain relief. The purpose of this review is to explore the mechanisms of exercise-induced analgesia in animal models for both pain prevention and alleviation.
2. Models of exercise-induced analgesia
Exercise can be performed aerobically or anaerobically and can be used with shortening (concentric), lengthening (eccentric) or static (isometric) contractions. Eccentric exercises produce muscle damage and have been used to model acute muscle pain in animals,4,61,114,151,152 whereas concentric-based exercises do not produce muscle damage or pain.74,116,153 The most commonly studied form of exercise in animal models is aerobic conditioning exercises such as treadmill running, swimming, or running wheel activity. Only one group has studied resistance training effects on pain and uses a single bout of concentric exercise.56,58 Thus, the majority of studies in animal models use aerobic exercise to examine mechanisms of analgesia.
Exercise-induced analgesia has been studied in multiple ways using animal models. One approach examined effects in animals by testing changes in response to painful stimuli immediately after a single exercise bout, which is comparable to studies in human subjects examining exercise-induced hypoalgesia to a single bout of exercise. This paradigm generally results in a short-lasting analgesic response (<30 minutes).127 More commonly, animal studies examine the effects of repeated exercise bouts over a longer period and are comparable to clinical treatment of individuals with chronic pain with a regular exercise program.59,88,89 These animal studies apply the repetitive exercise program either before or after a painful insult known to produce long-lasting hyperalgesia. Training sessions conducted before a painful insult explore the ability of exercise to prevent the onset of long-lasting pain, whereas training after onset of hyperalgesia will study exercises ability to alleviate pain.
The earliest work that demonstrated exercise-induced analgesia in animals was seen after short bouts of a forced cold-water swim. These studies demonstrated that swimming for as little as 3 minutes reduced pain related behaviors in response to tail shock and thermal tail-flick tests.13–15,24,37,62,105,120,160 Although these early studies demonstrated exercise-induced analgesia, it is hard to tease out analgesic effects due to the exercise paradigm as opposed to stress-induced analgesia by the cold water.22 Thus, current research has moved away from cold-water swim tests and towards less stressful exercise models. Currently, exercise models for animals use forced treadmill running, forced swimming, forced resistance training, and voluntary running wheels placed in cages. Forced treadmill running and swimming are the most commonly used exercise paradigms because they allow for control over intensity and duration of physical activity. However, these exercise models have demonstrated the ability to increase biomarkers of stress.36 Again, stress itself can produce analgesia through activation of opioid and serotonergic systems thus confounding results from these studies.25,37,164 A way to avoid this is through the use of voluntary running wheels placed in cages, which allows the animal to exercise without a stressful component. With that being said, the majority of exercise models show analgesia in pain-free animals and prevent or reverse hyperalgesia in animal models of neuropathic, inflammatory, and noninflammatory muscle pain.
Neuropathic pain models use numerous methods to induce hyperalgesia. These models produce injury to peripheral nerves or spinal cord, or is induced by diabetes. These pain models produce both mechanical and thermal paw hyperalgesia, which last for weeks to months. Exercise protocols that start either after or before the induction of neuropathic pain show a reduction in hyperalgesia when compared to sedentary animals (Tables 1 and 2). For example, repetitive swimming protocols showed that 50, 60, or 90 minutes of swimming 5 days a week reversed both mechanical and thermal hyperalgesia after onset of training compared to sedentary animals in animal models of peripheral nerve injury.3,28,86,150 Similar reductions in mechanical and thermal hyperalgesia in animal models of neuropathic pain are seen in animal studies using treadmills for exercise after induction of neuropathic pain. Treadmill protocols using repeated bouts of 10, 20, 30, or 60 minutes ranging from 3 to 7 days a week have been shown to reduce hyperalgesia if started before or after the induction of nerve injury5,10–12,26–29,31,34,35,50,63,72,73,80,81,85,96,97,101,102,131,135,149,154,156,162,165 (Fig. 1A). Furthermore, initiation of exercise either 1 week or 3 weeks after nerve injury produced analgesia 2 weeks after initiation of training,149 suggesting exercise is effective regardless of when the injury occurs (Fig. 1B). Greater intensity of treadmill training (10 m/min vs 16 m/min) produced greater analgesia, whereas differences in frequency of training (3 d/wk vs 5 d/wk) did not alter analgesic effects149 (Fig. 1B). By contrast, treadmill training 2 weeks before injury combined with 2 weeks after injury produced the greatest analgesic effects compared with solely training either before or after injury.11 Interestingly, treadmill training to exhaustion during each session blocked the long-term analgesic effects of exercise and resulted in hyperalgesia that was more severe than sedentary nerve-injured animals.35 This suggests that intensity and volume of exercise could be a key factor in production of analgesia with higher intensity and volume producing more analgesic effects while too much intensity could prove detrimental. Finally, in exercise models where animals are trained solely before induction of neuropathic pain mixed results are seen with voluntary wheel running for 6 weeks and treadmill training for 2 weeks blunting the development of mechanical hyperalgesia,11,65 whereas other studies with treadmill training for 2 or 3 weeks before injury saw no effect.131,156 Interestingly, in experimental models where animals are given access to running wheels after induction of neuropathic pain, some studies show positive analgesic effects,65,67,124,163 whereas others fail to show any pain relief.137,159 Overall, the majority of research illustrates that various exercise models and paradigms have proven successful in preventing and alleviating neuropathic pain.
Table 1.
Therapeutic effects of exercise.
| Reference | Pain model | Animals | Exercise type | Pretraining exercise | Initiation of exercise | Exercise duration + Intensity | Effective for pain |
|---|---|---|---|---|---|---|---|
| Allen et al.1 | OA—MIA | Male rats | Treadmill | N/A | PID 10 | 12–16 m/min for 30 , 4 d/wk for 5 wks | Yes for mechanical sensitivity, weightbearing symmetry, and ongoing pain |
| Almeida et al.3 | Neuropathic—PSNL | Male mice | Swimming | 1 min for 5 d | PID 7 | 10 min on days 1–3, duration increased by 10 min every 3 sessions till 50 min reached, 5 d/wk for 5 wks | Yes for mechanical and thermal sensitivity |
| Arbat-Plana et al.5 | Neuropathic—SNSR | Female rats | Treadmill | N/A | PID 3 | 18 m/min for 60 min, 5 d/wk for 2 wks | Yes for mechanical |
| Bement et al.6 | Muscle -acidic saline | Male rats | Treadmill | 3.05 m/min for 5 min for 3 d | PID 0 | 6.1 m/min for 15 min on days 1–2, 30 min on days 3–5 | Yes for muscle and mechanical sensitivity |
| Benson et al.7 | EAE | Female mice | VWR | 60 min access daily for 1 wk | PID 1 | 60 min access daily for 3 wks | Yes for mechanical sensitivity |
| Bobinski et al.10 | Neuropathic—SC | Male mice | Treadmill | 10 m/min for 10 min for 6 d | PID 3 | 10 m/min for 30 min, 5 d/wk for 2 wks | Yes for mechanical sensitivity |
| Bobinski et al.11 | Neuropathic—SC | Male mice | Treadmill | 10 m/min for 10 min for 6 d | PID 3 | 10 m/min for 30 min, 5 d/wk for 2 wks | Yes for mechanical and thermal sensitivity |
| Bobinski et al.12 | Neuropathic—SC | Male mice | Treadmill | 10 m/min for 10 min for 6 d | PID 3 | 5 d/wk for 2 wk, 30 min at 10 m/min | Yes for mechanical sensitivity |
| Chen et al.28 | Neuropathic—CCI | Male rats | Treadmill or swimming | Treadmill: 20 m/min for 15 min, for 3 d swimming: N/A | 1 day before injury | Graded exercise protocols, 5 d/wk for 6 wk | Both exercise protocols effective for mechanical and thermal sensitivity |
| Chen et al.27 | Diabetic neuropathy | Male rats | Treadmill | N/A | Not stated | 20 m/min for 60 min, 7 d/wk for 8 wks | Yes for mechanical and thermal sensitivity |
| Chen et al.29 | Incision | Male rats | Treadmill | 18 m/min for 15 min for 3 d | PID 8 | 18 m/min for 55 min, 5 d/wk for 4 wks | Yes for mechanical sensitivity |
| Chen et al.30 | Incision | Male rats | Treadmill | 12 m/min for 15 min for 2 d | PID 8 | 18 m/min for 55 min, 5 d/wk for 4 wks, | Yes for mechanical sensitivity |
| Chen et al.26 | Diabetic neuropathy | Male rats | Treadmill | 10 m/min for 15 min for 3 d | PID 3 | Wk 1–2: 20 m/min for 30 min, wk 3–4: 25 m/min for 60 min, 7 d/wk for 4 wks | Yes for mechanical and thermal sensitivity |
| Chhaya et al.31 | Neuropathic—SCI | Female rats | Treadmill | N/A | PID 5 | Started at 5 m/min and increased speed till 14 m/min for 20 min, 5 d/wk for 4 wks | Yes for mechanical sensitivity |
| Chuganji et al.33 | Cast | Male rats | Treadmill | N/A | PID 1 | 15 m/min for 30 min, 5 d/wk for 8 wks | Yes for mechanical sensitivity |
| Cobianchi et al.35 | Neuropathic—CCI | Male mice | Treadmill | 21 m/min till exhaustion or 60 min, 5 d/wk for 2 wks | PID 3 | 21 m/min till exhaustion or 60 min, 5d or 5 d/wk for 7 wks | Yes for mechanical sensitivity after 5 d of training, no for >5 d of training |
| Cobianchi et al.34 | Neuropathic—SNSR | Female rats | Treadmill | N/A | PID 3 | 19 m/min for 60 min for 5 d | Yes for mechanical sensitivity |
| Cormier et al.38 | OA—MIA | Male rats | VWR | 7 or 21 d | PID 1 | 24 hr access 7 d/wk for 3 wks | Yes for weightbearing symmetry with 21 days prior but not 7 |
| de Azambuja et al.46 | Glutamate-induced pain | Male rats | Swimming | 15–20 min for 6 d | PID1 | 30 min for 5 d | No for muscle sensitivity |
| Detloff et al.50 | Neuropathic—SCI | Female rats | FWR | N/A | PID 5 | 14 m/min for 20 min, 5 d/wk for 5 wks | Yes for mechanical, no for thermal sensitivity |
| Detloff et al.50 | Neuropathic—SCI | Female rats | FWR | N/A | PID 14 or 28 | 14 m/min for 20 min, 5 d/wk for 5 wks | No for mechanical sensitivity |
| Gong et al.63 | Neuropathic—SNI | Male rat pups | Treadmill | N/A | PID 11 | Days 1–3: 5 m/min for 10 min, days 4–6: 8 m/min for 20 min, days 8–13: 10 m/min for 30 min, days: 15–20 15 m/min for 30 min, 6 d/wk for 3 wks | Yes for mechanical sensitivity |
| Grace et al.65 | Neuropathic—CCI | Male rats | VWR | N/A | PID 1 or 14 | 24 hr access 7 d/wk for 6 or 11 wks | Yes for mechanical sensitivity; PID 1 = PID 14 |
| Groover et al.67 | Diabetic neuropathy | Male mice | VWR | N/A | PID 1 | 24 hr access 7 d/wk for 12 wk | Yes for mechanical and visceral sensitivity |
| Huang et al.72 | Neuropathic—CCI | Male rats | Treadmill | N/A | PID 8 | 14–16 m/min at 8% incline for 30 min, 7 d/wk for 3 wks | Yes for mechanical and thermal sensitivity |
| Hutchinson et al.73 | Neuropathic—SCI | Female rats | Treadmill | 12 m/min for 1 wk | PID 4 | 11–13 m/min for 20–25 min, 5 d/wk for 7 wks | Yes for mechanical sensitivity |
| Ishikawa et al.76 | Inflammatory—kaolin + carrageenan injection | Male rats | Stimulated quad contractions | N/A | PID 1 | 20 min for 6 d | Yes for knee pressure pain thresholds and paw mechanical sensitivity |
| Kami et al.80 | Neuropathic—CCI | Male mice | Treadmill | Wk 1: 5 d, 7 m/min for 10 min wk 2: 5 d, 7 m/min for 20–60 min | PID 2 | 7 m/min for 60 min for 5 d | Yes for mechanical and thermal sensitivity |
| Kami et al.81 | Neuropathic—CCI | Male mice | Treadmill | Wk 1: 5 d, 7 m/min for 10 min wk 2: 5 d, 7 m/min for 20–60 min | PID 2 | 7 m/min for 60 min for 5 d | Yes for mechanical and thermal sensitivity |
| Korb et al.85 | Neuropathic—SNSR | Male rats | Treadmill | N/A | PID 7 | Wk 1: 9 m/min for 20–50 min, wk 2–4: 9 m/min for 60 min, 5 d/wk for 4 wks | Yes for mechanical sensitivity |
| Kuphal et al.86 | Neuropathic—PSNL | Male rats and mice | Swimming | Rats: 90 min for 13 d mice: 30 min for 5 d | PID 1 | Rats: 90 min for 25 d mice: 30 min for 6 d | Yes for thermal sensitivity for both rats and mice |
| Lopez-Alvarez et al.96 | Neuropathic—SNSR | Female rats | Treadmill | 19 m/min for 60 min for 1 d | PID 3 | 19 m/min for 60 min, 5 d/wk for 1 or 2 wks | Yes for mechanical and thermal sensitivity, 1 wk = 2 wk of training |
| Lopez-Alvarez et al.97 | Neuropathic—SNSR | Female rats | Treadmill | 19 m/min for 60 min for 1 d | PID 3 | 19 m/min for 60 min for 12 d | Yes for mechanical and thermal sensitivity |
| Luan et al.101 | IDD | Male rats | Treadmill | N/A | PID 14 | Wk 1: 9 m/min for 20 min, Wk 2: 11 m/min for 30 min, Wk 3–8: 13 m/min for 40 min, 7 d/wk for 1–8 wks | Yes for mechanical sensitivity |
| Ma et al.103 | Diabetic neuropathy | Male rats | Treadmill | N/A | PID 1 | Wk 1–2: 5 m/min at 10% incline for 10 min, wk 3–5: 10 m/min at 10% incline for 10 min, 4 d/wk for 5 wks | Yes for mechanical sensitivity |
| Ma et al.102 | Diabetic neuropathy | Male rats | Treadmill | N/A | PID 1 | Wk 1–2: 5 m/min at 10% incline for 10 min, wk 3–5: 10 m/min at 10% incline for 10 min, 4 d/wk for 5 wks | Yes for mechanical sensitivity |
| Martins et al.106 | CRPS | Male mice | Swimming | N/A | PID 7 | 30 min for 5 d | Yes for mechanical sensitivity |
| Mifflin et al.110 | EAE | Male and female mice | VWR | 1 hr access for 3d | PID 4 | 1 hr access 7 d/wk until disease onset | Yes for mechanical sensitivity in females but not males |
| Morimoto et al.112 | Cast | Male rats | Treadmill | 12 m/min for 30 min for 3 d | PID 3 | 12 m/min for 30 min, 3 d/wk for 2 wks | Yes for mechanical sensitivity |
| Parent-Vachon et al.124 | Neuropathic—SNI | Female mice | VWR | N/A | PID 14 | 24 hr access 7 d/wk for 6 wks | Yes for mechanical sensitivity |
| Pitcher et al.125 | Inflammatory—CFA | Male rats | VWR | N/A | PID 3 | 2 hr/d access 4 d/wk for 3 wks | Yes for weightbearing asymmetry and thermal sensitivity |
| Ross et al.128 | I/R | Male mice | Treadmill | N/A | PID 0 | 13 m/min for 45 min for 1 d | No for mechanical sensitivity |
| Safakhah et al.131 | Neuropathic—CCI | Male rats | Treadmill | 10 m/min for 10 min for 5 d | PID 5 | 16 m/min for 30 min, 5 d/wk for 3 wks | Yes for mechanical and thermal sensitivity |
| Sanford et al.132 | Chronic stress | Female rats | VWR | N/A | PID 1 | 24 hr access 6 d/wk for 3 wks | Yes for visceral sensitivity |
| Shankarappa et al.135 | Diabetic neuropathy | Male rats | Treadmill | 18 m/min for 60 min for 5 d | PID 1 | 18 m/min for 60 min, 5 d/wk for 10 wks | Yes for mechanical sensitivity |
| Sharma et al.136 | Muscle -acidic saline | Female mice | Treadmill | 13 m/min for 3d (no duration given) | PID 5 | Wk 1: 13 m/min for 30 min, Wk 2: 14 m/min for 40 min, Wk 3: 15–16 m/min for 45 min, 5 d/wk for 3 wks | Yes for muscle and mechanical sensitivity |
| Sheahan et al.137 | Neuropathic—SNI | Male mice | VWR | N/A | PID 8–10 | 2 or 12 hr access, 5-6 d/wk 2 wks | No for mechanical sensitivity |
| Smith et al.146 | Overuse muscle injury | Female rats | Treadmill | N/A | Initiated during wk 5 of overuse injury model | 23 m/min for 60 min, 5 d/wk for 6 wks | No for mechanical sensitivity |
| Stagg et al.149 | Neuropathic—SNL | Male rats | Treadmill | 18 m/min for 10 min, 2 d/wk for 2 wks | PID 7 or 21 | 14–16 m/min for 30 min, 3 or 5 d/wk for 5 wks; OR 10 or 16 m/min for 30 min, 5 d/wk for 5 wks | Yes for mechanical and thermal sensitivity; PID 7 = 21; 3 d/wk = 5 d/wk; 16 m/min: yes, 10 m/min: no |
| Sun et al.150 | TNT | Male rats | Swimming | N/A | PID 7 | Wk 1: 10–50 min, Wk 2–5: 60 min, 5 d/wk for 5 wks, | Yes for mechanical sensitivity |
| Tsai et al.154 | Neuropathic—CCI | Male rats | Treadmill | N/A | PID 6 | 14–16 m/min for 30 min at 0 or 8% incline, 7 d/wk for 3 wks | Yes for mechanical and thermal sensitivity 8% > 0% incline |
| Wakaizumi et al.156 | Neuropathic—PSNL | Male mice | Treadmill | None or 6 or 12 m/min for 60 min 5 d/wk for 2 wks | PID 1 | 6 or 12 m/min for 60 min 5 d/wk for 1 or 2 wks, | Yes for mechanical and thermal sensitivity (6 m/min > 12 m/min) |
| Whitehead et al.159 | Neuropathic—CCI | Male rats | VWR | 60 min access for 1 wk | PID 2–3 | 60 min access for 7 or 18 d | No for mechanical sensitivity |
| Yamaoka et al.162 | Neuropathic—PSNL | Female rats | Treadmill | N/A | PID 1 | 20 m/min at 10° incline for 10 min, 5 d/wk 6 wks | Yes for mechanical and thermal sensitivity |
| Ye et al.163 | Neuropathic—ART | Male mice | VWR | N/A | PID 0 | 2 hr access 5 d/wk for 13 wks | Yes for mechanical and thermal sensitivity |
| Yoon et al.165 | Diabetic neuropathy | Male rats | Treadmill | 5 m/min for 20 min for 2 d | PID 7 | 10 m/min for 60 min 5 d/wk for 6 wks | Yes for mechanical sensitivity |
Table for effects of exercise on pain when exercise is initiated after induction of pain model.
ART, antiretroviral therapy; CCI, chronic constriction injury; CFA, complete Freund's adjuvant; CPRS, complex regional pain syndrome; EAE, experimental autoimmune encephalomyelitis; FWR, forced wheel running; I/R, ischemia and reperfusion; IDD, intervertebral disk degeneration; m/min, meters per min; MIA, monosodium iodoacetate; N/A, not applicable; OA, osteoarthritis; PID, postinjury day; PSNL, peripheral sciatic nerve ligation; SC, sciatic crush; SCI, spinal cord injury; SNI, spared nerve injury; SNSR, sciatic nerve resection and repair; TNT, tibial neuroma transposition; VWR, voluntary wheel running.
Table 2.
Preventative effects of exercise.
| Reference | Pain model | Animals | Exercise type | Exercise Duration + Intensity | Duration of pretraining | Effective for pain |
|---|---|---|---|---|---|---|
| Bobinski et al.11 | Neuropathic –SC | Male mice | Treadmill | 10 m/min for 30 min, 5 d/wk | 2 wk | Yes for mechanical, no for thermal sensitivity |
| Brito et al.16 | Muscle -acidic saline | Male and female mice | VWR | 24 hr access 7 d/wk | 8 wk | Yes for muscle and mechanical sensitivity |
| de Azambaja et al.45 | Inflammatory-carrageenan | Male rats | Swimming | 40 min, 5 d/wk, 4% BW attached to animals' body | 10 wk | Yes for muscle sensitivity |
| Grace et al.65 | Neuropathic—CCI | Male rats | VWR | 24 hr access 7 d/wk | 6 wk | Yes for mechanical sensitivity |
| Kuphal et al.86 | Inflammatory-formalin | Male rats | Swimming | 90 min | 9 d | No for phase 1 pain response, yes for phase 2 pain response |
| Leung et al.91 | Muscle—activity-induced | Male and female mice | VWR | 24 hr access 7 d/wk | 8 wk | Yes for muscle and mechanical sensitivity |
| Lima et al.92 | Muscle—activity-induced | Male and female mice | VWR | 24 hr access | 5 d | Yes for mechanical, no for muscle sensitivity |
| Martins et al.107 | Glutamate-induced pain | Male mice | Swimming | 30 min, 5 d/wk | 1–2 wk | Yes for mechanical sensitivity 2 wk > 1 wk |
| Ross et al.128 | Muscle—I/R | Male mice | VWR | 24 hr access | 2 d | Yes for mechanical sensitivity |
| Sabharwal et al.130 | Muscle—activity-induced | Male and female mice | VWR | 24 hr access 7 d/wk | 5 d or 8 wk | 5 d: yes for mechanical, no for muscle sensitivity; 8 wk: yes for mechanical and muscle sensitivity |
| Safakhah et al.131 | Neuropathic—CCI | Male rats | Treadmill | 16 m/min for 30 min 5 d/wk | 3 wk | No for mechanical and thermal sensitivity |
| Sheahan et al.137 | Inflammatory-formalin | Male mice | VWR | 2 or 12 hr/d access, 5–6 d/wk | 1–4 wk | No for nocifensive behaviors |
| Sluka et al.142 | Muscle—acidic saline or activity-induced | Male mice | VWR | 24 hr access 7 d/wk | 5 d or 8 wk | 5 d: yes for mechanical, no for muscle sensitivity; 8 wk: yes for mechanical and muscle sensitivity |
| Wakaizumi et al.156 | Neuropathic—PSNL | Male mice | Treadmill | 6 or 12 m/min for 60 min, 5 d/wk | 2 wk | No for mechanical or thermal sensitivity |
Table for effects of exercise on pain when exercise is performed before induction of pain model.
CCI, chronic constriction injury; I/R, ischemia and reperfusion; m/min, meters per min; PSNL, peripheral sciatic nerve ligation; SC, sciatic crush; VWR, voluntary wheel running.
Figure 1.
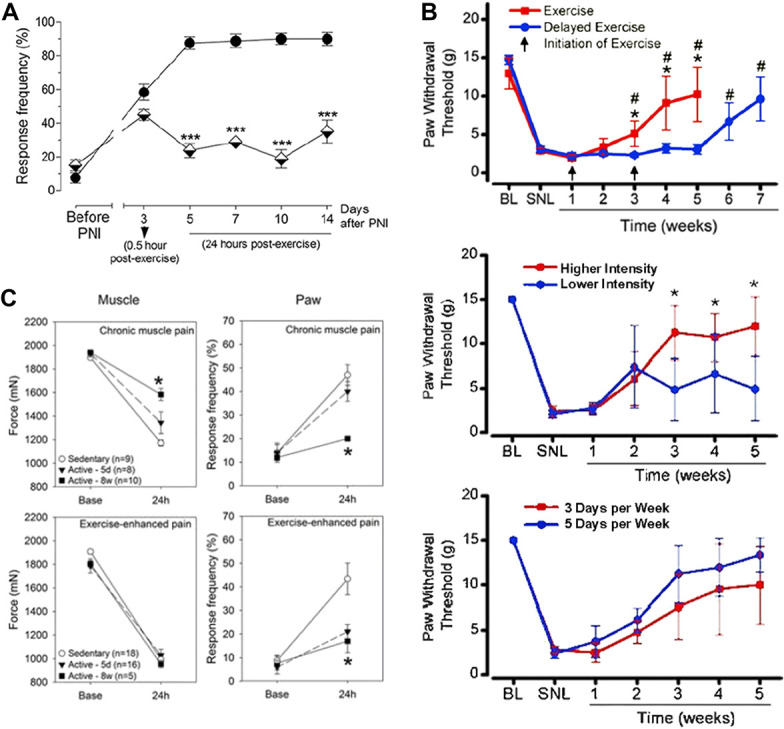
Exercise-induced analgesia. (A) Treadmill exercise after peripheral nerve injury (PNI) results in decreased mechanical hypersensitivity in as little as 2 training sessions. Filled circles represent PNI/Sedentary group and half-filled triangles represent PNI/Exercised group. Reproduced with permission from 12. (B) Exercise training resulted in reductions in hypersensitivity 2 weeks after induction of training regardless if 1 or 3 weeks passed between injury and initiation of training. Higher intensity of treadmill training resulted in a greater analgesic effect. However, no effect seen on degree of analgesia when treadmill training was conducted either 3 or 5 days a week. Reproduced with permission from 142 (C) Eight weeks of physical activity resulted in prevention of hyperalgesia both at muscle and paw after induction of noninflammatory muscle pain. Reproduced with permission from 149.
In models of noninflammatory muscle pain, hyperalgesia is induced through a dual stimulus model of acid injections into the gastrocnemius muscle, which are administered 5 days apart. This pain model results in long-lasting widespread hyperalgesia without tissue damage.111,141 In this model, exercise produces analgesia when performed as a preventative tool before muscle insult or as a therapeutic tool when started after the muscle insult (Tables 1 and 2). Development of mechanical hyperalgesia measured at the paw, ie, secondary hyperalgesia, is completely blocked in animals granted access to running wheels for 5 days or 8 weeks before induction of model.93,130,142 Further blockade of muscle hyperalgesia, ie, primary hyperalgesia, is only seen in animals granted access to running wheels for 8 weeks before induction of the model suggesting longer duration of training is needed to block primary hyperalgesia than secondary hyperalgesia16,91,130 (Fig. 1C). It is worth noting that distance run by animals with free access to running wheels is variable ranging between 0.2 and 5 km/d; yet, there is no correlation between distance run and degree of analgesia.65,91,142 These data suggest that there is a minimal threshold of activity necessary to protect against development of hyperalgesia. Similarly, treadmill training starting after induction of muscle pain caused a transient reversal of muscle pain immediately after the training session, whereas 3 weeks of training resulted in a complete reversal of the hyperalgesia.6,136 Thus, exercise can be effective for producing analgesic effects in models of noninflammatory muscle pain.
Exercise has also shown to be beneficial for pain relief in less widely studied pain models including osteoarthritis,1,38 incisional pain model,29,30 inflammatory pain model,45,76,86,125 complex regional pain syndrome,106 intervertebral disk degeneration (IDD),101 immobilization-induced pain,33,112 chronic stress-induced pain,132 ischemia and reperfusion,128 glutamate-induced pain,107 and experimental autoimmune encephalomyelitis7,110 (Tables 1 and 2). Although behavioral data are important for demonstrating the analgesic effects of exercise, understanding the mechanisms through which physical activity is producing these effects will be beneficial for future development of therapeutic targets for pain reduction. Consequently, the rest of this review covers the central, peripheral, and neuroimmune changes that occur to produce exercise-induced analgesia.
3. Central mechanism of exercise-induced analgesia
3.1. Endogenous opioids
One of the most widely studied central mechanisms of exercise-induced analgesia is the role of endogenous opioids in producing the analgesic effects. Early work showed that in healthy animals, systemic administration of naloxone, an opioid receptor antagonist, blocks the acute analgesic effects of short bouts of swimming.13,24,62,120,137,160 More recently, these behavioral effects have been replicated in exercise models of treadmill running, voluntary wheel running and resistance training as well as in pain models of noninflammatory muscle pain, neuropathic pain, arthritic pain, and glutamate-induced pain.1,6,16,47,58,93,106,107,147 Specifically, 8 weeks of voluntary running wheel activity blocked the production of hyperalgesia in a model of noninflammatory muscle pain. Naloxone administered systemically or directly into the periaqueductal gray (PAG) or the rostral ventral medial medulla (RVM), 2 brain locations important in endogenous opioid system, blocked these analgesic effects, whereas systemic injection of naloxone methiodide, a peripherally acting opioid antagonist, had no effect suggesting the central but not peripheral effects of exercise on mediating the endogenous opioid system16 (Fig. 2A). Similarly, the analgesic effects of treadmill running for neuropathic pain were reversed with either subcutaneous or intracerebroventricular injection of naloxone but not naloxone methiodide.149 Also, treadmill training after induction of knee osteoarthritis caused decrease in tactile sensitivity, improvement in weightbearing symmetry, and reduction of ongoing pain, and these analgesic exercise effects were reversed with systemic naloxone.1 Finally, 5 days of voluntary running wheel activity prevented the development of hyperalgesia in an activity-induced muscle pain model in wild-type but not mu-opioid receptor knockout mice demonstrating exercise produces its analgesic effects through activation of mu-opioid receptors.93
Figure 2.

Central mechanisms of exercise-induced analgesia. (A) Systemic naloxone but not naloxone methiodide reversed analgesic effects of exercise. Naloxone delivered directly into periaqueductal gray (PAG) or rostroventromedial medulla (RVM) also reversed the analgesic effects of exercise. Reproduced with permission from 16. (B) Treadmill training for 5 weeks resulted in increases in beta endorphin and met-enkephalin in PAG. Reproduced with permission from 149. (C) Two weeks of treadmill training reversed the increases in SERT in RVM seen after neuropathic injury. Reproduced with permission from 10. (D) Eight weeks of physical activity blocked increases in p-NR1 in RVM after induction of noninflammatory muscle pain. Reproduced with permission from 142. (E) Swimming reduced neuropathic injury increases in NGF and BDNF in lumbar DRG. Reproduced with permission from 3. BDNF, brain-derived neurotrophic factor; NGF, nerve growth factor.
Studies have also shown changes in the endogenous opioid system due to exercise in several locations. Increases in beta-endorphin and met-enkephalin, 2 endogenous opioid peptides, were found in both the PAG and RVM after 5 weeks of treadmill running and in the PAG and hypothalamus after 8 weeks of treadmill training, whereas increases in beta-endorphin in cerebrospinal fluid were found to be elevated for up to 48 hours after a single exercise bout33,70,149 (Fig. 2B). Also, increased opioid peptides met-enkephalin, leu-enkephalin, and beta-endorphin were found in frontal cortex of female rats given access to running wheels for 4 weeks after nerve injury.124 Similarly, increases in enkephalin were found in lumbar dorsal root ganglia (DRG) after 6 weeks of treadmill training.165 Finally, both forced and voluntary running activity for 7 days increased mu-opioid receptor expression in the hippocampus. Interestingly, when animals were subjected to forced or voluntary running activity for 45 sessions over a 9-week span, the increases in mu-opioid receptor were absent.47 This suggests that repeated activation of the opioid system by exercise can produce a compensatory downregulation of opioid receptor expression, similar to that seen in chronic administration of exogenous opioids such as morphine.2 In parallel, behavior studies show that chronic, but not acute, running wheel exposure in uninjured animals decreases analgesic effects of morphine, a mu-opioid agonist, and U50,488H, a kappa opioid agonist, and increases in withdrawal-like symptoms after administration of opioid receptor antagonists.40,82,143 Thus, current research suggests that exercise-induced analgesia is mediated by endogenous opioid activity.
3.2. Serotonergic system
The serotonergic system has been widely studied for its role in both the development and alleviation of chronic pain. Not surprisingly, exercise has been shown to produce its analgesic effects through modulation of the serotonergic system. Serotonin (5-HT) is elevated in the brainstem after a single 60-minute bout of swimming exercise and in the dorsal horn of the lumbar spinal cord after 15 minutes of treadmill running in pain-free rats.51,60 When rats swam 6 days a week for 4 weeks, increased levels of 5-HT were found in both the brainstem and parieto-occipital cortex and these increased levels were still significantly higher than sedentary controls one week after cessation of exercise.51 Similarly, in pain-free animals, increased levels of 5-HT were found in the cerebellum and midbrain after 8 weeks of treadmill training, whereas increased levels of 5-HT immunoreactivity were found in RVM after 4 weeks of treadmill training.19,85 Interestingly, after peripheral nerve injury, increased levels of 5-HT immunoreactivity were found bilaterally in the lumbosacral ventral horn of the spinal cord after 4 weeks of treadmill training but not in animals that underwent sham nerve injury.85 Also, animals with the peripheral nerve injury did not see increases in 5-HT immunoreactivity in the RVM, suggesting exercise could potentially produce different central nervous system effects depending on injury state of animal.85
In the RVM, increases in serotonin are analgesic with extracellular levels of serotonin controlled by the serotonin transporter (SERT).75,95 The SERT is increased, whereas 5-HT is decreased, in the RVM and brainstem after induction of neuropathic pain.10 Two weeks of treadmill training after induction of neuropathic pain reversed the increases in SERT and decreases in 5-HT with a corresponding reduction in mechanical hyperalgesia10 (Fig. 2C). Animals pretreated with ρ-chlorophenylalanine, a 5-HT synthesis inhibitor, blocked brainstem increases of 5-HT and prevented the analgesic effect of treadmill exercise in animals with neuropathic pain, repeated stimulated muscle contractions in pain-free animals, and 5 days of swimming exercise before acetic acid pain.10,69,107 Similar to the neuropathic pain model, there are increases in SERT in the RVM after induction of a noninflammatory muscle pain model, which is prevented by 8 weeks of prior voluntary running wheel activity.16 Finally, after induction of the noninflammatory pain model, blockade of SERT in the RVM, by microinjection of the selective serotonin reuptake inhibitor fluoxetine, reversed hyperalgesia showing modulation of SERT alters pain behaviors.16,93 Together, these data suggest that exercise-induced release of serotonin as well as changes in central serotonergic system is critical to the analgesic effects of regular exercise.
Recent evidence has also suggested an interactive role between the opioid and serotonergic systems to produce exercise-induced analgesia. Serotonin neurons in the RVM receive inputs from PAG opioid neurons and blockade of serotonin receptors in the PAG or spinal cord reduces the analgesic effects of morphine.25,83,134 Increases in SERT occur in the RVM after induction of an activity-induced pain model and 5 days of voluntary wheel running before induction of the model can block the SERT increases.93 However, physically active animals pretreated with systemic naloxone show greater SERT immunoreactivity in the RVM compared with physically active animals receiving vehicle. Similarly, physically active mu-opioid receptor knockout mice showed increased SERT immunoreactivity in RVM compared with physically active wild-type animals. Together, these data suggest that activation of opioid receptors during exercise is crucial for reducing expression of SERT in the brainstem to promote exercise-induced analgesia.
3.3. Endocannabinoids
Activation of the endocannabinoid system has also been attributed to the analgesic effects of exercise. Endocannabinoid receptors are located in the PAG, RVM, and dorsal horn of the spinal cord, all of which are locations known to be important for pain modulation.71,109,157 There are 2 naturally occurring ligands, anandamide (AEA) and 2-arachidonylglycerol (2-AG), that bind to the endocannabinoid CB1 and CB2 receptors to produce analgesic effects. In both human and animal studies, acute bouts of exercise increase circulating levels of endocannabinoids, which contributes to the feeling of “runner's high” associated with exercise.39,55–57,84,148 Endocannabinoid-mediated exercise-induced analgesia has been found after exercise in both healthy animals and animals subjected to glutamate-induced pain.56,57,107 In healthy animals, a single bout of treadmill training or resistance training increased nociceptive threshold to mechanical stimuli, which was blocked by pretreatment with systemic, intrathecal, or intracerebroventricular injections of CB1 and CB2 inverse agonists, AM251 and AM630, suggesting the endocannabinoid system is working at multiple locations to produce exercise-induced analgesia.56,57 Also, the antinociceptive response could be prolonged through systemic, intrathecal, or intracerebroventricular pretreatment with endocannabinoid metabolizing enzyme inhibitors. Similarly, 30 minutes of swimming for 1 week before intraplantar glutamate injection produced an antinociceptive effect, which was blocked by pretreatment with intrathecal, but not intraplantar, administration of a CB1 antagonist (AM281).107 Finally, increased expression of CB1 receptors were found in the dorsolateral and ventrolateral PAG after a single bout of treadmill training or resistance training and injection of AM251 into the PAG before exercise reduced this upregulation.56,57 Interestingly, similar to the serotonin-opioid interaction for producing analgesia, evidence has suggested a similar relationship between the endocannabinoid and opioid systems23,39,115; however, this relationship has not been studied in animal models of exercise-induced analgesia. Thus, evidence suggests there are supraspinal and spinal actions of exercise-induced analgesia involving the endocannabinoid system.
3.4. NDMA receptor alterations
The N-methyl-d-aspartate (NMDA) receptor is an excitatory glutamate receptor which has been implicated in the development of hyperalgesia. Increases in phosphorylation of the NR1 subunit (p-NR1) of the NMDA receptor in the RVM are crucial for the development of chronic muscle hyperalgesia. In models of noninflammatory and activity-induced muscle pain, there is increased p-NR1 expression in the RVM, and downregulation or pharmacological blockade of NR1 in the RVM prevents development of hyperalgesia41,42,139 showing a role for NMDA receptors in chronic muscle pain. More recent work shows that physical activity blocks the increases in p-NR1 in the RVM after muscle injury142 (Fig. 2D). Similarly, an hour of running daily for a week prevented increases in p-NR1 expression in the superficial dorsal horn in a model of experimental autoimmune encephalomyelitis.7 Finally, treadmill training was able to reduce increased NR1 expression in the lumbar spinal cord resulting from an incisional pain model.29 This suggests that exercise reduces excitatory transmission by preventing phosphorylation of the NMDA receptors in the central nervous system.
3.5. Noradrenergic system
The noradrenergic system also plays a role in the analgesic effects of exercise. Receptors for catecholamines (α1, α2, β2-adrenergic receptors) are found in locations known to mediate pain such as the PAG, locus coeruleus, dorsal raphe and spinal cord DRG.117 The noradrenergic system is activated during exercise to increase release of catecholamines.52 Early studies show that short bouts of swimming produced analgesia is potentiated by systemic pretreatment with norepinephrine, clonidine (α2-adrenergic agonist), or desipramine (norepinephrine reuptake inhibitor) when compared to swimming alone, whereas systemic or intracerebroventricular pretreatment with α1-or α2-adrenergic receptor antagonists prevented the swim-induced analgesia.14,15,121 Similarly, systemic pretreatment with α2, α2-A, or α2-C adrenergic receptor antagonists blocked exercise-induced analgesia from a single bout of either treadmill training or resistance training; there was no effect when administered intrathecally or intracerebroventricularly.48 Furthermore, in α2-A/C adrenergic receptor knockout mice, treadmill training failed to produce any antinociceptive effects to swimming or treadmill training.48 Finally, after induction of a neuropathic pain model, regular treadmill training prevented the downregulation α1 and β2 receptors in the dorsal horn of the spinal cord. Exercise was also shown to upregulate α1 and β2 receptors in the locus coeruleus and dorsal raphe. Systemic inhibition of the β2 receptors with butoxamine before exercise training completely blocked the analgesic effects of exercise. Furthermore, neuropathic injury-induced increases in activated microglia in the locus coeruleus were reduced with exercise; however, this effect was blocked by butoxamine treatment.97 This work demonstrates the role of the noradrenergic system in producing exercises' analgesic effects. Future work will need to be done to further determine location of action of the noradrenergic system in the production of exercise-induced analgesia.
3.6. Role of adenosine receptors
There has been a small amount of research into the role of adenosine and its receptor subtypes, A1 and A2A, in the role of exercise-induced analgesia. One study found swimming exercise for one week before intraplantar glutamate injection prevented glutamate-induced nociception.107 When these animals were pretreated with an A1 adenosine receptor antagonist, 1,3-dipropyl-8-cyclopentylxanthine (DPCPX), either intrathecally or intraplantarly, the antinociception effects of the swimming exercise was prevented. Similarly, swimming exercise mitigated mechanical hyperalgesia in an animal model of complex regional pain syndrome, but systemic pretreatment with caffeine (adenosine receptor antagonist) or an A1 receptor antagonist (DPCPX), but not an A2A receptor antagonist (ZM241385), blocked these analgesic effects.106 Furthermore, systemic pretreatment with an adenosine degradation inhibitor (erythro-9-(2-hydroxy-3nonyl) adenine; EHNA) potentiated exercise-induced analgesia. Together, these data suggest that exercise produces analgesia through A1 adenosine receptors both centrally and peripherally.
3.7. Changes in spinal cord and dorsal root ganglia
There have been multiple studies that have examined a number of different neurotransmitters, intracellular messengers, transcription factors, and growth factors in the spinal cord or DRG in a variety of animal models of exercise-induced analgesia. Neuropathic injury increases the intracellular messenger PLCγ-1 and the transcription factor CREB in the dorsal horn of the spinal cord, and increases both nerve growth factor (NGF) and brain-derived neurotrophic factor in spinal cord and DRG.3 Regular swimming (5 days/wk for 5 weeks) or 2 weeks of treadmill running not only prevented the development of hyperalgesia, but also returned all these measures (PLCg-1, CREB, NGF, and brain-derived neurotrophic factor) to normal uninjured levels3,12 (Fig. 2E). Similarly, 2 weeks of swimming prevented the increased intracellular messenger phosphorylated-PKA in the lumbar spinal cord after intraplantar glutamate injection,107 a single bout of treadmill training blocked the increase in c-FOS expression in the spinal cord after intraplantar formalin injection,94 and 3 weeks of treadmill training before induction of paw incisional pain model reduced the amount of phosphorylated-p38 in the spinal cord.64 Also, in a model of knee osteoarthritis there was an increase in calcitonin gene-related peptide in the dorsal horn of the lumbar spinal cord in laminae I–VI. Six days of electrically stimulated quadriceps contractions starting at onset of knee osteoarthritis completely blocked this increase.76 In addition, in models of diabetic neuropathy, there are increases in TRPV1 channels, phosphorylated-p38, phosphorylated m-TOR and its downstream signals S6K1 and 4E-BP1, and inflammatory cytokines IL-1β, IL-6, and TNFα in lumbar DRG.102,165 Regular treadmill training starting at the onset of diabetes was able to block the increases in TRPV1 channels, intracellular messengers, and inflammatory cytokines. Finally, neuropathic injury decreases the inhibitory neurotransmitter gamma-aminobutyric acid (GABA) and glutamic acid decarboxylase (which converts glutamate to GABA) in the dorsal horn of the spinal cord, which is prevented by regular treadmill running 60 minutes per day 5 days a week.80 Thus, this research suggests that exercise works to reverse increases in pronociceptive intracellular messengers, transcription factors, and growth factors while restoring antinociceptive mediators in the spinal cord.
Electrophysiology studies have found increased excitability of spinal cord dorsal horn neurons after injection of NGF into the multifidus muscles over 5 days. Swimming for 5 days between NGF injections blocked increases in measures of neuronal excitability in the spinal cord.46 Namely, the exercise blocked increases in the number of neurons responding to deep tissue stimulation, the receptive field size, and the resting activity compared to sedentary animals. There are also increases in DRG neurons excitability after induction of neuropathic pain with increased resting membrane potential, promoted spike frequency and spike counts, and decreased threshold potential in lumbar DRG.163 Voluntary running wheel activity for 2 hours a day, 5 days a week, during model induction completely blocked all increases in DRG excitability measures. Finally, in a pain model of diabetic peripheral neuropathy, treadmill training for 60 minutes, 5 days a week, delayed the onset of mechanical hypersensitivity. In sedentary animals, a >2.5 increase in calcium channel current density was measured at lumbar DRG.135 In animals that exercised, this increase in calcium channel current density was completely blocked. This research suggests that exercise is also working at the level of the spinal cord and DRG to produce its analgesic effects through altering electrophysiological changes that occur due to injury.
4. Neuroimmune mechanisms of exercise-induced analgesia
Alterations in the immune system by exercise have been studied both in the central nervous system and locally at the site of injury. Centrally, immune responses are modulated through activity of microglial cells and astrocytes, which can alter their morphology resulting in production of different levels of pro-inflammatory and anti-inflammatory cytokines. Specifically, microglia and astrocytes in the spinal cord will transform into a hypertrophic state after sciatic nerve injury resulting in an activated state that will increase pro-inflammatory cytokine production.3,7,12,31,35,63,65,81 In parallel, increases in pro-inflammatory cytokines TNF-α, IL-1β, and IL-6 and decreases in anti-inflammatory cytokines IL-4, IL-5, and IL-1ra have been reported in the RVM, spinal cord, and DRG after induction of sciatic nerve injury.10–12,63,65,165 Spinal cord increases in hyperactive microglia and astrocytes are blocked by treadmill running or swimming both before and after sciatic nerve injury3,7,12,31,35,65,81 (Fig. 3A). Furthermore, animals that exercised had reduced expression of pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6) and increases in anti-inflammatory cytokines (IL-4, IL-1ra, IL-5, and IL-10)10–12,63,65,165 (Fig. 3B). Finally, in IL-4 knockout mice or mice given an IL-4 antibody during treadmill training, exercise-induced analgesia in response to a sciatic nerve injury was completely blocked, suggesting that the increases in anti-inflammatory cytokines are part of the mechanism through which exercise protects and reverses the development of hyperalgesia.12
Figure 3.

Immune system mechanisms of exercise-induced analgesia. (A) Treadmill training for 2 weeks after neuropathic injury reduced hyperactive astrocytes (GFAP) and microglia (Iba-1) in lumbar spinal cord dorsal horns. Reproduced with permission from 12. (B) Treadmill training after sciatic nerve injury increased anti-inflammatory cytokines in spinal cord and sciatic nerve. Reproduced with permission from 12. (C) Physical activity increased percentage of M2 macrophages in gastrocnemius muscle (F4/80 macrophage stain, CD206 M2 macrophage stain, CD11c M1 macrophage stain). Reproduced with permission from 91. (D) Pretreatment with IL-10 blocked hyperalgesia in response to noninflammatory muscle pain. Reproduced with permission from 91.
Exercise also modulates the immune response in the periphery at the site of injury. A model of knee osteoarthritis found increased total number of macrophages in the synovium of the arthritic knee. The increase in macrophages was attenuated by electrically stimulated quadriceps contractions starting at the onset of knee osteoarthritis.76 Also, treadmill training blocked the increases in pro-inflammatory cytokines (TNF-α, IL-1β, and IL-6), increases in chemokine CCL2,65 and decreases in anti-inflammatory cytokines (IL-4, IL1ra, and IL-10) that are found in the sciatic nerve after induction of neuropathic pain.11,12,26,28,72,154 Furthermore, sciatic nerve injury was found to alter macrophage distribution in the sciatic nerve. Macrophages are immune cells that have different phenotypes depending on their cytokine release profile: M1 macrophages release pro-inflammatory cytokines, whereas M2 macrophages release anti-inflammatory cytokines.113 There is an increased proportion of M1 to M2 macrophages in the sciatic nerve after injury.12,65 Regular treadmill training before or after injury increased the number of M2 vs M1 macrophages, increased serum IL-10, and decreased IL-1β and CCL2 in the serum and sciatic nerve.12,65,72 Similarly, physically active, uninjured mice show an increased proportion of M2 macrophages in the muscle91 (Fig. 3C). In a noninflammatory muscle pain model, the analgesia produced by 8 weeks of running wheel activity is prevented by systemic or intramuscular blockade of IL-10 receptors, whereas administration of IL-10 systemically or into the muscle mimicked the analgesic effects similar to exercise91 (Fig. 3D). Also, in an ischemic model of muscle pain, there was an increase in IL-1β levels in the muscle which was blocked with prior exercise.128 Furthermore, blockade of IL-1 receptors prevented the exercise-induced analgesia, whereas treatment with IL-1 resulted in decreases in pain behaviors.128 Finally, in a pain model of IDD, there was an increase in IL-1β, TNFα, and leptin in the multifidus muscle, all of which were decreased by voluntary running wheel activity.77 Thus, this research suggests that exercise can alter macrophage phenotype distribution and subsequent pro-inflammatory and anti-inflammatory cytokine profiles in the periphery at the site of injury to promote analgesia.
5. Peripheral mechanisms of exercise-induced analgesia
There have been a few studies on exercise-induced changes in the periphery that can lead to exercise's analgesic effects. In the paw, after sciatic nerve injury, there are increases in nerve fiber sprouting from adjacent areas into the denervated areas resulting in mechanical hyperalgesia both at the medial and lateral test sites of the paw.96 Treadmill running for 60 minutes a day decreased the amount of nerve sprouting and reinnervation of plantar skin from the medial paw into the denervated lateral paw resulting in decreased responses to mechanical stimuli. Also, tibial nerve injury resulted in increasing plasma levels of leptin and decreased levels of adiponectin with a subsequent mechanical hyperalgesia.150 Swimming exercise 5 days a week reversed mechanical hyperalgesia and normalized circulating leptin and adiponectin levels. Systemic administration of leptin after swimming sessions blocked the analgesic effects of exercise, whereas administration of adiponectin alone was able to decrease mechanical hyperalgesia. Finally, a model of IDD results in degenerative changes in the nucleus pulposus and anulus fibrosus. Daily treadmill training initiated after onset of the IDD model resulted in increased cell counts in the nucleus pulposus and anulus fibrosus, as well as increased markers of cell proliferation in the intervertebral disk and the adjacent epiphyseal cartilage when compared to sedentary controls.101 Thus, these data suggest that alterations in the periphery at or away from the site of injury can also impact pain behaviors and exercise can help to mitigate any pronociceptive changes at these sites.
6. Clinical implications
From a clinical standpoint, this research on animal models of exercise-induced analgesia could provide the basis for new clinical studies to improve exercise prescription guidelines for humans. First, exercise works to both prevent onset of pain and resolve pain after injury. Currently, 58/64 (90%) articles published on the topic of exercise-induced analgesia demonstrate positive effects of exercise in as little as one bout of activity. Also, this research demonstrates that specificity of exercise might not be as important as the act of exercise itself. Exercise-induced analgesia is demonstrated in exercise models of running, swimming, and resistance training. Similarly, in human studies, different types of exercise programs have been applied to chronic pain patients with no difference in outcomes seen between the different exercise programs.68,79,133 Also, both aerobic and resistance training exercise programs are listed as exercise recommendations for low back pain, osteoarthritis, and fibromyalgia.8,17,18,20,21,126 This lack of specificity of exercise is possibly a result of the multitude of mechanisms through which exercise can reduce pain. Therefore, exercise prescription should be dependent upon patient preferences, therapist training, available equipment, cost, and safety.133 Furthermore, there is evidence to suggest that duration and intensity of exercise can impact the degree of exercise-induced analgesia. Higher amounts of duration and intensity resulted in a greater degree of analgesia; however, too much intensity was found to have detrimental effects, suggesting a possible inverted U-shaped curve when it comes to exercise intensity and pain relief.35,94,149 Thus, dosing of exercise needs to be done appropriately to maximize pain relief while limiting negative side effects. Finally, as previously noted individuals with chronic pain may fail to receive pain relief from an acute bout of exercise or may suffer from exercise-induced pain.127 This increase in pain with an acute bout exercise could be a barrier to exercise adherence. This can be managed through appropriate dosing of exercise to decrease exercise-induced symptom flares, or through treatments that reduce activity-induced pain. Our recent clinical trial shows that transcutaneous electrical nerve stimulation (TENS) significantly reduces activity-induced pain when compared to placebo or no-TENS in women with fibromyalgia,44 suggesting TENS could be an effective treatment to manage activity-induced pain. Another approach could be to use psychological and educational interventions to decrease fear-avoidance behavior surrounding activity and catastrophic thinking regarding exercise-induced hyperalgesia.104,118,127
7. Future directions
Although work has repeatedly shown the analgesic effects of exercise, the underlying mechanism for this analgesia has only recently begun to be elucidated. Continued research into the mechanisms through which exercise can prevent and reduce pain will help develop intervention targets for treating chronic pain. Specifically, research into the mechanisms through which exercise prevents pain vs reduces pain could reveal different mechanisms. Also, we do not know if the neurological and immune response to exercise is the same in pain-free animals as compared to animals after injury. Human studies have suggested that overweight individuals see greater increases in pro-inflammatory cytokines (IL-6, IL-8, and TNFα) compared with lean individuals after acute bouts of exercise.32 Thus, future work needs to study the impact of comorbidity factors on the analgesic mechanisms of exercise. Although studies have shown that all modes of exercise are beneficial, a majority of the research in animal models has been on aerobic-based exercise (running and swimming) as opposed to strength training which is typically performed in rehabilitation settings. Although all exercise modes have beneficial effects, we do not know if aerobic and resistance training-based exercise programs are producing analgesia through the same mechanisms. Similarly, we do not know the adequate amount of dose or minimal effective dose for these exercise programs. Thus, further research needs to be conducted on other forms of exercise as well as on different dosing of exercise to improve exercise prescription in chronic pain patients. Finally, nearly all exercise-induced analgesia research has been conducted in populations using only male animals. Recent research has called for inclusion of female animals in all experiments due to sexual differences found in hyperalgesia mechanisms and pain phenotypes.66,78,98–100,123,145–147,158 Because mechanistic differences in pain development have been seen between the sexes, we cannot assume the same mechanistic effects of exercise-induced analgesia between males and females. Of the research that has used both sexes while studying exercise-induced analgesia, mixed results have been found, with some finding sexual differences in exercise-induced analgesia110, whereas others have not.93,139 Therefore, future studies need to include both sexes in their experiments to determine if there are possible sex differences in the mechanisms of exercise-induced analgesia. The incorporation of females into animal studies is also crucial because chronic pain conditions are more prevalent in women.53 By only including male animals, the generalizability to the population of individuals who suffer from chronic pain conditions is limited. All of this research would help improve utilization, adherence, and prescription of exercise for chronic pain patients.
Disclosures
The authors have no conflicts of interest to declare.
Acknowledgements
Research supported by National Institutes of Health AR061371, AR06371-S1, AR073187, GM067795, and Foundation of Physical Therapy Research Promotion of Doctoral Studies (PODS I).
Footnotes
Sponsorships or competing interests that may be relevant to content are disclosed at the end of this article.
References
- [1].Allen J, Imbert I, Havelin J, Henderson T, Stevenson G, Liaw L, King T. Effects of treadmill exercise on advanced osteoarthritis pain in rats. Arthritis Rheumatol 2017;69:1407–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Allouche S, Noble F, Marie N. Opioid receptor desensitization: mechanisms and its link to tolerance. Front Pharmacol 2014;5:280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Almeida C, DeMaman A, Kusuda R, Cadetti F, Ravanelli MI, Queiroz AL, Sousa TA, Zanon S, Silveira LR, Lucas G. Exercise therapy normalizes BDNF upregulation and glial hyperactivity in a mouse model of neuropathic pain. PAIN 2015;156:504–13. [DOI] [PubMed] [Google Scholar]
- [4].Alvarez P, Levine JD, Green PG. Eccentric exercise induces chronic alterations in musculoskeletal nociception in the rat. Eur J Neurosc 2010;32:819–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Arbat-Plana A, Cobianchi S, Herrando-Grabulosa M, Navarro X, Udina E. Endogenous modulation of TrkB signaling by treadmill exercise after peripheral nerve injury. Neuroscience 2017;340:188–200. [DOI] [PubMed] [Google Scholar]
- [6].Bement MK, Sluka KA. Low-intensity exercise reverses chronic muscle pain in the rat in a naloxone-dependent manner. Arch Phys Med Rehabil 2005;86:1736–40. [DOI] [PubMed] [Google Scholar]
- [7].Benson C, Paylor JW, Tenorio G, Winship I, Baker G, Kerr BJ. Voluntary wheel running delays disease onset and reduces pain hypersensitivity in early experimental autoimmune encephalomyelitis (EAE). Exp Neurol 2015;271:279–90. [DOI] [PubMed] [Google Scholar]
- [8].Bidonde J, Busch AJ, Schachter CL, Overend TJ, Kim SY, Goes SM, Boden C, Foulds HJ. Aerobic exercise training for adults with fibromyalgia. Cochrane Database Syst Rev 2017;Cd012700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Biswas A, Oh PI, Faulkner GE, Bajaj RR, Silver MA, Mitchell MS, Alter DA. Sedentary time and its association with risk for disease incidence, mortality, and hospitalization in adults: a systematic review and meta-analysis. Ann Intern Med 2015;162:123–32. [DOI] [PubMed] [Google Scholar]
- [10].Bobinski F, Ferreira TAA, Cordova MM, Dombrowski PA, da Cunha C, Santo C, Poli A, Pires RGW, Martins-Silva C, Sluka KA, Santos ARS. Role of brainstem serotonin in analgesia produced by low-intensity exercise on neuropathic pain after sciatic nerve injury in mice. PAIN 2015;156:2595–606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Bobinski F, Martins DF, Bratti T, Mazzardo-Martins L, Winkelmann-Duarte EC, Guglielmo LG, Santos AR. Neuroprotective and neuroregenerative effects of low-intensity aerobic exercise on sciatic nerve crush injury in mice. Neuroscience 2011;194:337–48. [DOI] [PubMed] [Google Scholar]
- [12].Bobinski F, Teixeira JM, Sluka KA, Santos ARS. Interleukin-4 mediates the analgesia produced by low-intensity exercise in mice with neuropathic pain. PAIN 2018;159:437–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Bodnar RJ, Kelly DD, Spiaggia A, Ehrenberg C, Glusman M. Dose-dependent reductions by naloxone of analgesia induced by cold-water stress. Pharmacol Biochem Behav 1978;8:667–72. [DOI] [PubMed] [Google Scholar]
- [14].Bodnar RJ, Mann PE, Stone EA. Potentiation of cold-water swim analgesia by acute, but not chronic desipramine administration. Pharmacol Biochem Behav 1985;23:749–52. [DOI] [PubMed] [Google Scholar]
- [15].Bodnar RJ, Merrigan KP, Sperber E. Potentiation of cold-water swim analgesia and hypothermia by clonidine. Pharmacol Biochem Behav 1983;19:447–51. [DOI] [PubMed] [Google Scholar]
- [16].Brito RG, Rasmussen LA, Sluka KA. Regular physical activity prevents development of chronic muscle pain through modulation of supraspinal opioid and serotonergic mechanisms. Pain Rep 2017;2:e618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Brosseau L, Taki J, Desjardins B, Thevenot O, Fransen M, Wells GA, Mizusaki Imoto A, Toupin-April K, Westby M, Álvarez Gallardo IC, Gifford W, Laferrière L, Rahman P, Loew L, De Angelis G, Cavallo S, Shallwani SM, Aburub A, Bennell KL, Van der Esch M, Simic M, McConnell S, Harmer A, Kenny GP, Paterson G, Regnaux J-P, Lefevre-Colau M-M, McLean L. The Ottawa panel clinical practice guidelines for the management of knee osteoarthritis. Part three: aerobic exercise programs. Clin Rehabil 2017;31:612–24. [DOI] [PubMed] [Google Scholar]
- [18].Brosseau L, Taki J, Desjardins B, Thevenot O, Fransen M, Wells GA, Mizusaki Imoto A, Toupin-April K, Westby M, Álvarez Gallardo IC, Gifford W, Laferrière L, Rahman P, Loew L, De Angelis G, Cavallo S, Shallwani SM, Aburub A, Bennell KL, Van der Esch M, Simic M, McConnell S, Harmer A, Kenny GP, Paterson G, Regnaux J-P, Lefevre-Colau M-M, McLean L. The Ottawa panel clinical practice guidelines for the management of knee osteoarthritis. Part two: strengthening exercise programs. Clin Rehabil 2017;31:596–611. [DOI] [PubMed] [Google Scholar]
- [19].Brown BS, Payne T, Kim C, Moore G, Krebs P, Martin W. Chronic response of rat brain norepinephrine and serotonin levels to endurance training. J Appl Physiol Respir Environ Exerc Physiol 1979;46:19–23. [DOI] [PubMed] [Google Scholar]
- [20].Busch AJ, Webber SC, Brachaniec M, Bidonde J, Bello-Haas VD, Danyliw AD, Overend TJ, Richards RS, Sawant A, Schachter CL. Exercise therapy for fibromyalgia. Curr Pain Headache Rep 2011;15:358–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Busch AJ, Webber SC, Richards RS, Bidonde J, Schachter CL, Schafer LA, Danyliw A, Sawant A, Dal Bello-Haas V, Rader T, Overend TJ. Resistance exercise training for fibromyalgia. Cochrane Database Syst Rev 2013;Cd010884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Butler RK, Finn DP. Stress-induced analgesia. Prog Neurobiol 2009;88:184–202. [DOI] [PubMed] [Google Scholar]
- [23].Butler RK, Rea K, Lang Y, Gavin AM, Finn DP. Endocannabinoid-mediated enhancement of fear-conditioned analgesia in rats: opioid receptor dependency and molecular correlates. PAIN 2008;140:491–500. [DOI] [PubMed] [Google Scholar]
- [24].Carmody J, Cooper K. Swim stress reduces chronic pain in mice through an opioid mechanism. Neurosc Lett 1987;74:358–63. [DOI] [PubMed] [Google Scholar]
- [25].Carruba MO, Nisoli E, Garosi V, Sacerdote P, Panerai AE, Da Prada M. Catecholamine and serotonin depletion from rat spinal cord: effects on morphine and footshock induced analgesia. Pharmacol Res 1992;25:187–94. [DOI] [PubMed] [Google Scholar]
- [26].Chen YW, Chiu CC, Hsieh PL, Hung CH, Wang JJ. Treadmill training combined with insulin suppresses diabetic nerve pain and cytokines in rat sciatic nerve. Anesth Analg 2015;121:239–46. [DOI] [PubMed] [Google Scholar]
- [27].Chen YW, Hsieh PL, Chen YC, Hung CH, Cheng JT. Physical exercise induces excess hsp72 expression and delays the development of hyperalgesia and allodynia in painful diabetic neuropathy rats. Anesth Analg 2013;116:482–90. [DOI] [PubMed] [Google Scholar]
- [28].Chen YW, Li YT, Chen YC, Li ZY, Hung CH. Exercise training attenuates neuropathic pain and cytokine expression after chronic constriction injury of rat sciatic nerve. Anesth Analg 2012;114:1330–7. [DOI] [PubMed] [Google Scholar]
- [29].Chen YW, Lin MF, Chen YC, Hung CH, Tzeng JI, Wang JJ. Exercise training attenuates postoperative pain and expression of cytokines and N-methyl-D-aspartate receptor subunit 1 in rats. Reg Anesth Pain Med 2013;38:282–8. [DOI] [PubMed] [Google Scholar]
- [30].Chen YW, Tzeng JI, Lin MF, Hung CH, Wang JJ. Forced treadmill running suppresses postincisional pain and inhibits upregulation of substance P and cytokines in rat dorsal root ganglion. J Pain 2014;15:827–34. [DOI] [PubMed] [Google Scholar]
- [31].Chhaya SJ, Quiros-Molina D, Tamashiro-Orrego AD, Houle JD, Detloff MR. Exercise-induced changes to the macrophage response in the dorsal root ganglia prevent neuropathic pain after spinal cord injury. J Neurotrauma 2019;36:877–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Christiansen T, Bruun JM, Paulsen SK, Olholm J, Overgaard K, Pedersen SB, Richelsen B. Acute exercise increases circulating inflammatory markers in overweight and obese compared with lean subjects. Eur J Appl Physiol 2013;113:1635–42. [DOI] [PubMed] [Google Scholar]
- [33].Chuganji S, Nakano J, Sekino Y, Hamaue Y, Sakamoto J, Okita M. Hyperalgesia in an immobilized rat hindlimb: effect of treadmill exercise using non-immobilized limbs. Neurosci Lett 2015;584:66–70. [DOI] [PubMed] [Google Scholar]
- [34].Cobianchi S, Casals-Diaz L, Jaramillo J, Navarro X. Differential effects of activity dependent treatments on axonal regeneration and neuropathic pain after peripheral nerve injury. Exp Neurol 2013;240:157–67. [DOI] [PubMed] [Google Scholar]
- [35].Cobianchi S, Marinelli S, Florenzano F, Pavone F, Luvisetto S. Short- but not long-lasting treadmill running reduces allodynia and improves functional recovery after peripheral nerve injury. Neuroscience 2010;168:273–87. [DOI] [PubMed] [Google Scholar]
- [36].Contarteze RV, Manchado Fde B, Gobatto CA, De Mello MA. Stress biomarkers in rats submitted to swimming and treadmill running exercises. Comp Biochem Physiol A Mol Integr Physiol 2008;151:415–22. [DOI] [PubMed] [Google Scholar]
- [37].Cooper K, Carmody J. The characteristics of the opioid-related analgesia induced by the stress of swimming in the mouse. Neurosci Lett 1982;31:165–70. [DOI] [PubMed] [Google Scholar]
- [38].Cormier J, Cone K, Lanpher J, Kinens A, Henderson T, Liaw L, Bilsky EJ, King T, Rosen CJ, Stevenson GW. Exercise reverses pain-related weight asymmetry and differentially modulates trabecular bone microarchitecture in a rat model of osteoarthritis. Life Sci 2017;180:51–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Crombie KM, Brellenthin AG, Hillard CJ, Koltyn KF. Endocannabinoid and opioid system interactions in exercise-induced hypoalgesia. Pain Med 2018;19:118–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].D'Anci KE, Gerstein AV, Kanarek RB. Long-term voluntary access to running wheels decreases kappa-opioid antinociception. Pharmacol Biochem Behav 2000;66:343–6. [DOI] [PubMed] [Google Scholar]
- [41].Da Silva LF, Desantana JM, Sluka KA. Activation of NMDA receptors in the brainstem, rostral ventromedial medulla, and nucleus reticularis gigantocellularis mediates mechanical hyperalgesia produced by repeated intramuscular injections of acidic saline in rats. J Pain 2010;11:378–87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Da Silva LF, Walder RY, Davidson BL, Wilson SP, Sluka KA. Changes in expression of NMDA-NR1 receptor subunits in the rostral ventromedial medulla modulate pain behaviors. PAIN 2010;151:155–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Dailey DL, Keffala VJ, Sluka KA. Do cognitive and physical fatigue tasks enhance pain, cognitive fatigue, and physical fatigue in people with fibromyalgia? Arthritis Care Res 2015;67:288–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Dailey DL, Vance CGT, Rakel BA, Zimmerman MB, Embree J, Merriwether EN, Geasland KM, Chimenti R, Williams JM, Golchha M, Crofford LJ, Sluka KA. Transcutaneous electrical nerve stimulation reduces movement-evoked pain and fatigue: a randomized, controlled trial. Arthritis Rheumatol 2020;72:824–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].de Azambuja G, Botasso Gomes B, Messias LHD, Aquino BM, Jorge CO, Manchado-Gobatto FB, Oliveira-Fusaro MCG. Swimming physical training prevented the onset of acute muscle pain by a mechanism dependent of PPARγ receptors and CINC-1. Neuroscience 2020;427:64–74. [DOI] [PubMed] [Google Scholar]
- [46].de Azambuja G, Hortscht U, Hoheisel U, Oliveira Fusaro MC, Mense S, Treede RD. Short-term swimming exercise attenuates the sensitization of dorsal horn neurons in rats with NGF-induced low back pain. Eur J Pain 2018;22:1409–18. [DOI] [PubMed] [Google Scholar]
- [47].de Oliveira MS, da Silva Fernandes MJ, Scorza FA, Persike DS, Scorza CA, da Ponte JB, de Albuquerque M, Cavalheiro EA, Arida RM. Acute and chronic exercise modulates the expression of MOR opioid receptors in the hippocampal formation of rats. Brain Res Bull 2010;83:278–83. [DOI] [PubMed] [Google Scholar]
- [48].de Souza GG, Duarte ID, de Castro Perez A. Differential involvement of central and peripheral alpha2 adrenoreceptors in the antinociception induced by aerobic and resistance exercise. Anesth Analg 2013;116:703–11. [DOI] [PubMed] [Google Scholar]
- [49].Detloff MR, Quiros-Molina D, Javia AS, Daggubati L, Nehlsen AD, Naqvi A, Ninan V, Vannix KN, McMullen MK, Amin S, Ganzer PD, Houlé JD. Delayed exercise is ineffective at reversing aberrant nociceptive afferent plasticity or neuropathic pain after spinal cord injury in rats. Neurorehabil Neural Repair 2016;30:685–700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Detloff MR, Smith EJ, Quiros Molina D, Ganzer PD, Houlé JD. Acute exercise prevents the development of neuropathic pain and the sprouting of non-peptidergic (GDNF- and artemin-responsive) c-fibers after spinal cord injury. Exp Neurol 2014;255:38–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Dey S, Singh RH, Dey PK. Exercise training: significance of regional alterations in serotonin metabolism of rat brain in relation to antidepressant effect of exercise. Physiol Behav 1992;52:1095–9. [DOI] [PubMed] [Google Scholar]
- [52].Dunn AL, Reigle TG, Youngstedt SD, Armstrong RB, Dishman RK. Brain norepinephrine and metabolites after treadmill training and wheel running in rats. Med Sci Sports Exerc 1996;28:204–9. [DOI] [PubMed] [Google Scholar]
- [53].Fillingim RB, King CD, Ribeiro-Dasilva MC, Rahim-Williams B, Riley JL., III Sex, gender, and pain: a review of recent clinical and experimental findings. J Pain 2009;10:447–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [54].Fischer CP, Berntsen A, Perstrup LB, Eskildsen P, Pedersen BK. Plasma levels of interleukin-6 and C-reactive protein are associated with physical inactivity independent of obesity. Scand J Med Sci Sports 2007;17:580–7. [DOI] [PubMed] [Google Scholar]
- [55].Fuss J, Steinle J, Bindila L, Auer MK, Kirchherr H, Lutz B, Gass P. A runner's high depends on cannabinoid receptors in mice. Proc Natl Acad Sci U S A 2015;112:13105–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Galdino G, Romero T, Silva JF, Aguiar D, Paula AM, Cruz J, Parrella C, Piscitelli F, Duarte I, Di Marzo V, Perez A. Acute resistance exercise induces antinociception by activation of the endocannabinoid system in rats. Anesth Analg 2014;119:702–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Galdino G, Romero TR, Silva JF, Aguiar DC, de Paula AM, Cruz JS, Parrella C, Piscitelli F, Duarte ID, Di Marzo V, Perez AC. The endocannabinoid system mediates aerobic exercise-induced antinociception in rats. Neuropharmacology 2014;77:313–24. [DOI] [PubMed] [Google Scholar]
- [58].Galdino GS, Duarte ID, Perez AC. Participation of endogenous opioids in the antinociception induced by resistance exercise in rats. Braz J Med Biol Res 2010;43:906–9. [DOI] [PubMed] [Google Scholar]
- [59].Geneen LJ, Moore RA, Clarke C, Martin D, Colvin LA, Smith BH. Physical activity and exercise for chronic pain in adults: an overview of Cochrane Reviews. Cochrane Database Syst Rev 2017;CD011279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Gerin C, Teilhac JR, Smith K, Privat A. Motor activity induces release of serotonin in the dorsal horn of the rat lumbar spinal cord. Neurosci Lett 2008;436:91–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [61].Gibson W, Arendt-Nielsen L, Taguchi T, Mizumura K, Graven-Nielsen T. Increased pain from muscle fascia following eccentric exercise: animal and human findings. Exp Brain Res 2009;194:299–308. [DOI] [PubMed] [Google Scholar]
- [62].Girardot MN, Holloway FA. Cold water stress analgesia in rats: differential effects of naltrexone. Physiol Behav 1984;32:547–55. [DOI] [PubMed] [Google Scholar]
- [63].Gong X, Chen Y, Fu B, Jiang J, Zhang M. Infant nerve injury induces delayed microglial polarization to the M1 phenotype, and exercise reduces delayed neuropathic pain by modulating microglial activity. Neuroscience 2017;349:76–86. [DOI] [PubMed] [Google Scholar]
- [64].Gong X, Jiang J, Zhang M. Exercise preconditioning reduces neonatal incision surgery-induced enhanced hyperalgesia via inhibition of P38 mitogen-activated protein kinase and IL-1β, TNF-α release. Int J Dev Neurosci 2016;52:46–54. [DOI] [PubMed] [Google Scholar]
- [65].Grace PM, Fabisiak TJ, Green-Fulgham SM, Anderson ND, Strand KA, Kwilasz AJ, Galer EL, Walker FR, Greenwood BN, Maier SF, Fleshner M, Watkins LR. Prior voluntary wheel running attenuates neuropathic pain. PAIN 2016;157:2012–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Gregory NS, Gibson-Corley K, Frey-Law L, Sluka KA. Fatigue-enhanced hyperalgesia in response to muscle insult: induction and development occur in a sex-dependent manner. PAIN 2013;154:2668–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].Groover AL, Ryals JM, Guilford BL, Wilson NM, Christianson JA, Wright DE. Exercise-mediated improvements in painful neuropathy associated with prediabetes in mice. PAIN 2013;154:2658–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Henry SM, Van Dillen LR, Ouellette-Morton RH, Hitt JR, Lomond KV, DeSarno MJ, Bunn JY. Outcomes are not different for patient-matched versus nonmatched treatment in subjects with chronic recurrent low back pain: a randomized clinical trial. Spine J 2014;14:2799–810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [69].Hoffmann P, Skarphedinsson JO, Delle M, Thoren P. Electrical stimulation of the gastrocnemius muscle in the spontaneously hypertensive rat increases the pain threshold: role of different serotonergic receptors. Acta Physiol Scand 1990;138:125–31. [DOI] [PubMed] [Google Scholar]
- [70].Hoffmann P, Terenius L, Thorén P. Cerebrospinal fluid immunoreactive beta-endorphin concentration is increased by voluntary exercise in the spontaneously hypertensive rat. Regul Pept 1990;28:233–9. [DOI] [PubMed] [Google Scholar]
- [71].Hohmann AG, Tsou K, Walker JM. Cannabinoid suppression of noxious heat-evoked activity in wide dynamic range neurons in the lumbar dorsal horn of the rat. J Neurophysiol 1999;81:575–83. [DOI] [PubMed] [Google Scholar]
- [72].Huang PC, Tsai KL, Chen YW, Lin HT, Hung CH. Exercise combined with ultrasound attenuates neuropathic pain in rats associated with downregulation of IL-6 and TNF-alpha, but with upregulation of IL-10. Anesth Analg 2017;124:2038–44. [DOI] [PubMed] [Google Scholar]
- [73].Hutchinson KJ, Gomez-Pinilla F, Crowe MJ, Ying Z, Basso DM. Three exercise paradigms differentially improve sensory recovery after spinal cord contusion in rats. Brain 2004;127:1403–14. [DOI] [PubMed] [Google Scholar]
- [74].Iguchi M, Shields RK. Quadriceps low-frequency fatigue and muscle pain are contraction-type-dependent. Muscle Nerve 2010;42:230–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [75].Inase M, Nakahama H, Otsuki T, Fang JZ. Analgesic effects of serotonin microinjection into nucleus raphe magnus and nucleus raphe dorsalis evaluated by the monosodium urate (MSU) tonic pain model in the rat. Brain Res 1987;426:205–11. [DOI] [PubMed] [Google Scholar]
- [76].Ishikawa K, Kajiwara Y, Sakamoto J, Sasaki R, Goto K, Honda Y, Kataoka H, Okita M. Low-intensity muscle contraction exercise following the onset of arthritis improves hyperalgesia via reduction of joint inflammation and central sensitization in the spinal cord in a rat model. Neurosci Lett 2019;706:18–23. [DOI] [PubMed] [Google Scholar]
- [77].James G, Millecamps M, Stone LS, Hodges PW. Dysregulation of the inflammatory mediators in the multifidus muscle after spontaneous intervertebral disc degeneration SPARC-null mice is ameliorated by physical activity. Spine 2018;43:E1184–e1194. [DOI] [PubMed] [Google Scholar]
- [78].Ji Y, Murphy AZ, Traub RJ. Sex differences in morphine-induced analgesia of visceral pain are supraspinally and peripherally mediated. Am J Physiol Regul Integr Comp Physiol 2006;291:R307–314. [DOI] [PubMed] [Google Scholar]
- [79].Jull G, Falla D, Treleaven J, Hodges P, Vicenzino B. Retraining cervical joint position sense: the effect of two exercise regimes. J Orthop Res 2007;25:404–12. [DOI] [PubMed] [Google Scholar]
- [80].Kami K, Taguchi Ms S, Tajima F, Senba E. Improvements in impaired GABA and GAD65/67 production in the spinal dorsal horn contribute to exercise-induced hypoalgesia in a mouse model of neuropathic pain. Mol Pain 2016;12:1744806916629059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Kami K, Taguchi S, Tajima F, Senba E. Histone acetylation in microglia contributes to exercise-induced hypoalgesia in neuropathic pain model mice. J Pain 2016;17:588–99. [DOI] [PubMed] [Google Scholar]
- [82].Kanarek RB, Gerstein AV, Wildman RP, Mathes WF, D'Anci KE. Chronic running-wheel activity decreases sensitivity to morphine-induced analgesia in male and female rats. Pharmacol Biochem Behav 1998;61:19–27. [DOI] [PubMed] [Google Scholar]
- [83].Kiefel JM, Cooper ML, Bodnar RJ. Serotonin receptor subtype antagonists in the medial ventral medulla inhibit mesencephalic opiate analgesia. Brain Res 1992;597:331–8. [DOI] [PubMed] [Google Scholar]
- [84].Koltyn KF, Brellenthin AG, Cook DB, Sehgal N, Hillard C. Mechanisms of exercise-induced hypoalgesia. J Pain 2014;15:1294–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [85].Korb A, Bonetti LV, da Silva SA, Marcuzzo S, Ilha J, Bertagnolli M, Partata WA, Faccioni-Heuser MC. Effect of treadmill exercise on serotonin immunoreactivity in medullary raphe nuclei and spinal cord following sciatic nerve transection in rats. Neurochem Res 2010;35:380–9. [DOI] [PubMed] [Google Scholar]
- [86].Kuphal KE, Fibuch EE, Taylor BK. Extended swimming exercise reduces inflammatory and peripheral neuropathic pain in rodents. J Pain 2007;8:989–97. [DOI] [PubMed] [Google Scholar]
- [87].Lahart IM, Metsios GS, Nevill AM, Carmichael AR. Physical activity, risk of death and recurrence in breast cancer survivors: a systematic review and meta-analysis of epidemiological studies. Acta Oncol 2015;54:635–54. [DOI] [PubMed] [Google Scholar]
- [88].Landmark T, Romundstad P, Borchgrevink PC, Kaasa S, Dale O. Associations between recreational exercise and chronic pain in the general population: evidence from the HUNT 3 study. PAIN 2011;152:2241–7. [DOI] [PubMed] [Google Scholar]
- [89].Landmark T, Romundstad PR, Borchgrevink PC, Kaasa S, Dale O. Longitudinal associations between exercise and pain in the general population--the HUNT pain study. PLoS One 2013;8:e65279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Lannersten L, Kosek E. Dysfunction of endogenous pain inhibition during exercise with painful muscles in patients with shoulder myalgia and fibromyalgia. PAIN 2010;151:77–86. [DOI] [PubMed] [Google Scholar]
- [91].Leung A, Gregory NS, Allen LA, Sluka KA. Regular physical activity prevents chronic pain by altering resident muscle macrophage phenotype and increasing interleukin-10 in mice. PAIN 2016;157:70–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Lima LV, Abner TSS, Sluka KA. Does exercise increase or decrease pain? Central mechanisms underlying these two phenomena. J Physiol 2017;595:4141–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [93].Lima LV, DeSantana JM, Rasmussen LA, Sluka KA. Short-duration physical activity prevents the development of activity-induced hyperalgesia through opioid and serotoninergic mechanisms. PAIN 2017;158:1697–710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [94].Liu HY, Yang K, Guo Y, Potenza MN, Zhao Y. The effects of different intensities of exercise on behavioral and molecular pain-related reactions in rats: implications for medication development. Yale J Biol Med 2017;90:5–13. [PMC free article] [PubMed] [Google Scholar]
- [95].Llewelyn MB, Azami J, Roberts MH. The effect of modification of 5-hydroxytryptamine function in nucleus raphe magnus on nociceptive threshold. Brain Res 1984;306:165–70. [DOI] [PubMed] [Google Scholar]
- [96].Lopez-Alvarez VM, Modol L, Navarro X, Cobianchi S. Early increasing-intensity treadmill exercise reduces neuropathic pain by preventing nociceptor collateral sprouting and disruption of chloride cotransporters homeostasis after peripheral nerve injury. PAIN 2015;156:1812–25. [DOI] [PubMed] [Google Scholar]
- [97].Lopez-Alvarez VM, Puigdomenech M, Navarro X, Cobianchi S. Monoaminergic descending pathways contribute to modulation of neuropathic pain by increasing-intensity treadmill exercise after peripheral nerve injury. Exp Neurol 2018;299:42–55. [DOI] [PubMed] [Google Scholar]
- [98].Loyd DR, Morgan MM, Murphy AZ. Sexually dimorphic activation of the periaqueductal gray–rostral ventromedial medullary circuit during the development of tolerance to morphine in the rat. Eur J Neurosci 2008;27:1517–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [99].Loyd DR, Murphy AZ. Sex differences in the anatomical and functional organization of the periaqueductal gray-rostral ventromedial medullary pathway in the rat: a potential circuit mediating the sexually dimorphic actions of morphine. J Comp Neurol 2006;496:723–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].Loyd DR, Wang X, Murphy AZ. Sex differences in -opioid receptor expression in the rat midbrain periaqueductal gray are essential for eliciting sex differences in morphine analgesia. J Neurosci 2008;28:14007–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Luan S, Wan Q, Luo H, Li X, Ke S, Lin C, Wu Y, Wu S, Ma C. Running exercise alleviates pain and promotes cell proliferation in a rat model of intervertebral disc degeneration. Int J Mol Sci 2015;16:2130–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [102].Ma X, Liu S, Liu D, Wang Q, Li H, Zhao Z. Exercise intervention attenuates neuropathic pain in diabetes via mechanisms of mammalian target of rapamycin (mTOR). Arch Physiol Biochem 2020;126:41–8. [DOI] [PubMed] [Google Scholar]
- [103].Ma XQ, Qin J, Li HY, Yan XL, Zhao Y, Zhang LJ. Role of exercise activity in alleviating neuropathic pain in diabetes via inhibition of the pro-inflammatory signal pathway. Biol Res Nurs 2019;21:14–21. [DOI] [PubMed] [Google Scholar]
- [104].Malfliet A, Kregel J, Meeus M, Cagnie B, Roussel N, Dolphens M, Danneels L, Nijs J. Applying contemporary neuroscience in exercise interventions for chronic spinal pain: treatment protocol. Braz J Phys Ther 2017;21:378–87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [105].Marek P, Mogil JS, Sternberg WF, Panocka I, Liebeskind JC. N-methyl-D-aspartic acid (NMDA) receptor antagonist MK-801 blocks non-opioid stress-induced analgesia. II. Comparison across three swim-stress paradigms in selectively bred mice. Brain Res 1992;578:197–203. [DOI] [PubMed] [Google Scholar]
- [106].Martins DF, Mazzardo-Martins L, Soldi F, Stramosk J, Piovezan AP, Santos AR. High-intensity swimming exercise reduces neuropathic pain in an animal model of complex regional pain syndrome type I: evidence for a role of the adenosinergic system. Neuroscience 2013;234:69–76. [DOI] [PubMed] [Google Scholar]
- [107].Martins DF, Siteneski A, Ludtke DD, Dal-Secco D, Santos ARS. High-intensity swimming exercise decreases glutamate-induced nociception by activation of G-protein-coupled receptors inhibiting phosphorylated protein kinase A. Mol Neurobiol 2017;54:5620–31. [DOI] [PubMed] [Google Scholar]
- [108].Mazzardo-Martins L, Martins DF, Marcon R, Dos Santos UD, Speckhann B, Gadotti VM, Sigwalt AR, Guglielmo LG, Santos AR. High-intensity extended swimming exercise reduces pain-related behavior in mice: involvement of endogenous opioids and the serotonergic system. J Pain 2010;11:1384–93. [DOI] [PubMed] [Google Scholar]
- [109].Meng ID, Manning BH, Martin WJ, Fields HL. An analgesia circuit activated by cannabinoids. Nature 1998;395:381–3. [DOI] [PubMed] [Google Scholar]
- [110].Mifflin KA, Yousuf MS, Thorburn KC, Huang J, Perez-Munoz ME, Tenorio G, Walter J, Ballanyi K, Drohomyrecky PC, Dunn SE, Kerr BJ. Voluntary wheel running reveals sex-specific nociceptive factors in murine experimental autoimmune encephalomyelitis. PAIN 2019;160:870–81. [DOI] [PubMed] [Google Scholar]
- [111].Miranda A, Peles S, Rudolph C, Shaker R, Sengupta JN. Altered visceral sensation in response to somatic pain in the rat. Gastroenterology 2004;126:1082–9. [DOI] [PubMed] [Google Scholar]
- [112].Morimoto A, Winaga H, Sakurai H, Ohmichi M, Yoshimoto T, Ohmichi Y, Matsui T, Ushida T, Okada T, Sato J. Treadmill running and static stretching improve long-lasting hyperalgesia, joint limitation, and muscle atrophy induced by cast immobilization in rats. Neurosci Lett 2013;534:295–300. [DOI] [PubMed] [Google Scholar]
- [113].Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol 2008;8:958–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [114].Murase S, Terazawa E, Queme F, Ota H, Matsuda T, Hirate K, Kozaki Y, Katanosaka K, Taguchi T, Urai H, Mizumura K. Bradykinin and nerve growth factor play pivotal roles in muscular mechanical hyperalgesia after exercise (delayed-onset muscle soreness). J Neurosci 2010;30:3752–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [115].Navarro M, Chowen J, Rocío A Carrera M, del Arco I, Villanúa MA, Martin Y, Roberts AJ, Koob GF, de Fonseca FR. CB1 cannabinoid receptor antagonist-induced opiate withdrawal in morphine-dependent rats. Neuroreport 1998;9:3397–402. [DOI] [PubMed] [Google Scholar]
- [116].Newham DJ, Mills KR, Quigley BM, Edwards RH. Pain and fatigue after concentric and eccentric muscle contractions. Clin Sci (Lond) 1983;64:55–62. [DOI] [PubMed] [Google Scholar]
- [117].Nicholas AP, Hökfelt T, Pieribone VA. The distribution and significance of CNS adrenoceptors examined with in situ hybridization. Trends Pharmacol Sci 1996;17:245–55. [DOI] [PubMed] [Google Scholar]
- [118].Nijs J, Lluch Girbés E, Lundberg M, Malfliet A, Sterling M. Exercise therapy for chronic musculoskeletal pain: innovation by altering pain memories. Man Ther 2015;20:216–20. [DOI] [PubMed] [Google Scholar]
- [119].Nocon M, Hiemann T, Müller-Riemenschneider F, Thalau F, Roll S, Willich SN. Association of physical activity with all-cause and cardiovascular mortality: a systematic review and meta-analysis. Eur J Cardiovasc Prev Rehabil 2008;15:239–46. [DOI] [PubMed] [Google Scholar]
- [120].O'Connor P, Chipkin RE. Comparisons between warm and cold water swim stress in mice. Life Sci 1984;35:631–9. [DOI] [PubMed] [Google Scholar]
- [121].Oluyomi AO, Hart SL. Alpha-adrenoceptor involvement in swim stress-induced antinociception in the mouse. J Pharm Pharmacol 1990;42:778–84. [DOI] [PubMed] [Google Scholar]
- [122].Paffenbarger RS, Jr, Lee IM, Leung R. Physical activity and personal characteristics associated with depression and suicide in American college men. Acta Psychiatr Scand Suppl 1994;377:16–22. [DOI] [PubMed] [Google Scholar]
- [123].Paige C, Maruthy GB, Mejia G, Dussor G, Price T. Spinal inhibition of P2XR or p38 signaling disrupts hyperalgesic priming in male, but not female, mice. Neuroscience 2018;385:133–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [124].Parent-Vachon M, Beaudry F, Carrier D, Di Cristo G, Vachon P. The effects of exercise on pain and reproductive performance in female pregnant mice with neuropathic pain. Biol Res Nurs 2019;21:500–9. [DOI] [PubMed] [Google Scholar]
- [125].Pitcher MH, Tarum F, Rauf IZ, Low LA, Bushnell C. Modest amounts of voluntary exercise reduce pain- and stress-related outcomes in a rat model of persistent hind limb inflammation. J Pain 2017;18:687–701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [126].Qaseem A, Wilt TJ, McLean RM, Forciea MA. Clinical guidelines committee of the American college of P. Noninvasive treatments for acute, subacute, and chronic low back pain: a clinical practice guideline from the American college of physicians. Ann Intern Med 2017;166:514–30. [DOI] [PubMed] [Google Scholar]
- [127].Rice D, Nijs J, Kosek E, Wideman T, Hasenbring MI, Koltyn K, Graven-Nielsen T, Polli A. Exercise-induced hypoalgesia in pain-free and chronic pain populations: state of the art and future directions. J Pain 2019;20:1249–66. [DOI] [PubMed] [Google Scholar]
- [128].Ross JL, Queme LF, Lamb JE, Green KJ, Ford ZK, Jankowski MP. Interleukin 1beta inhibition contributes to the antinociceptive effects of voluntary exercise on ischemia/reperfusion-induced hypersensitivity. PAIN 2018;159:380–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [129].Rovio S, Kåreholt I, Helkala E-L, Viitanen M, Winblad B, Tuomilehto J, Soininen H, Nissinen A, Kivipelto M. Leisure-time physical activity at midlife and the risk of dementia and Alzheimer's disease. Lancet Neurol 2005;4:705–11. [DOI] [PubMed] [Google Scholar]
- [130].Sabharwal R, Rasmussen L, Sluka KA, Chapleau MW. Exercise prevents development of autonomic dysregulation and hyperalgesia in a mouse model of chronic muscle pain. PAIN 2016;157:387–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [131].Safakhah HA, Moradi Kor N, Bazargani A, Bandegi AR, Gholami Pourbadie H, Khoshkholgh-Sima B, Ghanbari A. Forced exercise attenuates neuropathic pain in chronic constriction injury of male rat: an investigation of oxidative stress and inflammation. J Pain Res 2017;10:1457–66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [132].Sanford MT, Yeh JC, Mao JJ, Guo Y, Wang Z, Zhang R, Holschneider DP, Rodriguez LV. Voluntary exercise improves voiding function and bladder hyperalgesia in an animal model of stress-induced visceral hypersensitivity: a multidisciplinary approach to the study of urologic chronic pelvic pain syndrome research network study. Neurourol Urodyn 2020;39:603–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [133].Saragiotto BT, Maher CG, Yamato TP, Costa LOP, Menezes Costa LC, Ostelo RWJG, Macedo LG. Motor control exercise for chronic non-specific low-back pain. Cochrane Database Syst Rev 2016;CD012004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [134].Schul R, Frenk H. The role of serotonin in analgesia elicited by morphine in the periaqueductal gray matter (PAG). Brain Res 1991;553:353–7. [DOI] [PubMed] [Google Scholar]
- [135].Shankarappa SA, Piedras-Rentería ES, Stubbs EB., Jr Forced-exercise delays neuropathic pain in experimental diabetes: effects on voltage-activated calcium channels. J Neurochem 2011;118:224–36. [DOI] [PubMed] [Google Scholar]
- [136].Sharma NK, Ryals JM, Gajewski BJ, Wright DE. Aerobic exercise alters analgesia and neurotrophin-3 synthesis in an animal model of chronic widespread pain. Phys Ther 2010;90:714–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [137].Sheahan TD, Copits BA, Golden JP, Gereau RW. Voluntary exercise training: analysis of mice in uninjured, inflammatory, and nerve-injured pain states. PLoS One 2015;10:e0133191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [138].Shyu BC, Andersson SA, Thoren P. Endorphin mediated increase in pain threshold induced by long-lasting exercise in rats. Life Sci 1982;30:833–40. [DOI] [PubMed] [Google Scholar]
- [139].Sluka KA, Danielson J, Rasmussen L, DaSilva LF. Exercise-induced pain requires NMDA receptor activation in the medullary raphe nuclei. Med Sci Sports Exerc 2012;44:420–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [140].Sluka KA, Frey-Law L, Hoeger Bement M. Exercise-induced pain and analgesia? Underlying mechanisms and clinical translation. PAIN 2018;159(suppl 1):S91–s97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [141].Sluka KA, Kalra A, Moore SA. Unilateral intramuscular injections of acidic saline produce a bilateral, long-lasting hyperalgesia. Muscle Nerve 2001;24:37–46. [DOI] [PubMed] [Google Scholar]
- [142].Sluka KA, O'Donnell JM, Danielson J, Rasmussen LA. Regular physical activity prevents development of chronic pain and activation of central neurons. J Appl Physiol 2013;114:725–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [143].Smith MA, Yancey DL. Sensitivity to the effects of opioids in rats with free access to exercise wheels: mu-opioid tolerance and physical dependence. Psychopharmacology (Berl) 2003;168:426–34. [DOI] [PubMed] [Google Scholar]
- [144].Smith TTG, Barr-Gillespie AE, Klyne DM, Harris MY, Amin M, Paul RW, Cruz GE, Zhao H, Gallagher S, Barbe MF. Forced treadmill running reduces systemic inflammation yet worsens upper limb discomfort in a rat model of work-related musculoskeletal disorders. BMC musculoskeletal disorders 2020;21:57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [145].Sorge RE, LaCroix-Fralish ML, Tuttle AH, Sotocinal SG, Austin JS, Ritchie J, Chanda ML, Graham AC, Topham L, Beggs S, Salter MW, Mogil JS. Spinal cord Toll-like receptor 4 mediates inflammatory and neuropathic hypersensitivity in male but not female mice. J Neurosci 2011;31:15450–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [146].Sorge RE, Mapplebeck JCS, Rosen S, Beggs S, Taves S, Alexander JK, Martin LJ, Austin J-S, Sotocinal SG, Chen D, Yang M, Shi XQ, Huang H, Pillon NJ, Bilan PJ, Tu Y, Klip A, Ji R-R, Zhang J, Salter MW, Mogil JS. Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Nat Neurosci 2015;18:1081–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [147].Sorge RE, Totsch SK. Sex differences in pain. J Neurosci Res 2016;95:1271–81. [DOI] [PubMed] [Google Scholar]
- [148].Sparling PB, Giuffrida A, Piomelli D, Rosskopf L, Dietrich A. Exercise activates the endocannabinoid system. Neuroreport 2003;14:2209–11. [DOI] [PubMed] [Google Scholar]
- [149].Stagg NJ, Mata HP, Ibrahim MM, Henriksen EJ, Porreca F, Vanderah TW, Philip Malan T., Jr Regular exercise reverses sensory hypersensitivity in a rat neuropathic pain model: role of endogenous opioids. Anesthesiology 2011;114:940–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [150].Sun L, Lv Y, Tian J, Yu T, Niu F, Zhang X, Du D. Regular swimming exercise attenuated neuroma pain in rats: involvement of leptin and adiponectin. J Pain 2019;20:1112–24. [DOI] [PubMed] [Google Scholar]
- [151].Taguchi T, Matsuda T, Tamura R, Sato J, Mizumura K. Muscular mechanical hyperalgesia revealed by behavioural pain test and c-Fos expression in the spinal dorsal horn after eccentric contraction in rats. J Physiol 2005;564:259–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [152].Taguchi T, Sato J, Mizumura K. Augmented mechanical response of muscle thin-fiber sensory receptors recorded from rat muscle-nerve preparations in vitro after eccentric contraction. J Neurophysiol 2005;94:2822–31. [DOI] [PubMed] [Google Scholar]
- [153].Taylor L. Effects of concentric and eccentric contractions on exercise-induced muscle injury, inflammation, and serum IL-6. J Sports Sci Med 2004;3:226–3. [PMC free article] [PubMed] [Google Scholar]
- [154].Tsai K-L, Huang P-C, Wang L-K, Hung C-H, Chen Y-W. Incline treadmill exercise suppresses pain hypersensitivity associated with the modulation of pro-inflammatory cytokines and anti-inflammatory cytokine in rats with peripheral nerve injury. Neurosci Lett 2017;643:27–31. [DOI] [PubMed] [Google Scholar]
- [155].Tuomilehto J, Lindström J, Eriksson JG, Valle TT, Hämäläinen H, Ilanne-Parikka P, Keinänen-Kiukaanniemi S, Laakso M, Louheranta A, Rastas M, Salminen V, Uusitupa M, Finnish Diabetes Prevention Study G. Prevention of type 2 diabetes mellitus by changes in lifestyle among subjects with impaired glucose tolerance. New Engl J Med 2001;344:1343–50. [DOI] [PubMed] [Google Scholar]
- [156].Wakaizumi K, Kondo T, Hamada Y, Narita M, Kawabe R, Narita H, Watanabe M, Kato S, Senba E, Kobayashi K, Kuzumaki N, Yamanaka A, Morisaki H, Narita M. Involvement of mesolimbic dopaminergic network in neuropathic pain relief by treadmill exercise: a study for specific neural control with Gi-DREADD in mice. Mol Pain 2016;12:1744806916681567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [157].Walker JM, Huang SM, Strangman NM, Tsou K, Sañudo-Peña MC. Pain modulation by release of the endogenous cannabinoid anandamide. Proc Natl Acad Sci U S A 1999;96:12198–203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [158].Wang X, Traub RJ, Murphy AZ. Persistent pain model reveals sex difference in morphine potency. Am J Physiol Regul Integr Comp Physiol 2006;291:R300–306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [159].Whitehead RA, Lam NL, Sun MS, Sanchez J, Noor S, Vanderwall AG, Petersen TR, Martin HB, Milligan ED. Chronic sciatic neuropathy in rat reduces voluntary wheel-running activity with concurrent chronic mechanical allodynia. Anesth Anal 2017;124:346–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [160].Willow M, Carmody J, Carroll P. The effects of swimming in mice on pain perception and sleeping time in response to hypnotic drugs. Life Sci 1980;26:219–24. [DOI] [PubMed] [Google Scholar]
- [161].Wolin KY, Yan Y, Colditz GA, Lee IM. Physical activity and colon cancer prevention: a meta-analysis. Br J Cancer 2009;100:611–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [162].Yamaoka S, Oshima Y, Horiuchi H, Morino T, Hino M, Miura H, Ogata T. Altered gene expression of RNF34 and PACAP possibly involved in mechanism of exercise-induced analgesia for neuropathic pain in rats. Int J Mol Sci 2017;18:1962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [163].Ye H, Du X, Hua Q. Effects of voluntary exercise on antiretroviral therapy-induced neuropathic pain in mice. J Physiol Sci 2018;68:521–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [164].Yesilyurt O, Seyrek M, Tasdemir S, Kahraman S, Deveci MS, Karakus E, Halici Z, Dogrul A. The critical role of spinal 5-HT7 receptors in opioid and non-opioid type stress-induced analgesia. Eur J Pharmacol 2015;762:402–10. [DOI] [PubMed] [Google Scholar]
- [165].Yoon H, Thakur V, Isham D, Fayad M, Chattopadhyay M. Moderate exercise training attenuates inflammatory mediators in DRG of Type 1 diabetic rats. Exp Neurol 2015;267:107–14. [DOI] [PubMed] [Google Scholar]