

Abstract
The molecular mechanism underlying photoperiodic response in seasonal breeding animals such as the Japanese quail, red jungle fowl, sheep, mouse, and hamster involves thyroid-stimulating hormone beta subunit (TSHβ) mRNA expression in the pars tuberalis stimulated by the extension in day length. Furthermore, this mechanism is regulated by eyes absent 3 (Eya3) in mammals. Even in birds, the expression of both TSHβ and EYA3 is induced in the pars tuberalis by the extension in day length; however, the relationship between the two genes is unknown. To clarify the function of EYA3 in quail photoperiodism, in the present study, we performed mRNA structure analysis of the Japanese quail EYA3 mRNA using reverse transcription-polymerase chain reaction (RT-PCR) and Southern blot analysis. The results revealed that there are four types of splice variants within regions of exons 7, 8, and 9 of quail EYA3 mRNA. Among the four splice variants of quail EYA3, the splice variant containing exon 7 was expressed in the pars tuberalis on the first long day, when quails were transferred from the short-day condition to the long-day condition. The results indicate that EYA3 splice variant containing exon 7 is involved in the photoperiodic response of the pars tuberalis in the Japanese quail.
Keywords: EYA3, Japanese quail, photoperiodism, splice variant
Introduction
The seasonal change in day length results in various physiological responses in animals and plants, and this phenomenon is called photoperiodism. Photoperiodism in animals involves seasonal breeding, molting, hair replacement, hibernation, and migration. The molecular mechanism underlying photoperiodic response in seasonal breeding organisms, such as the Japanese quail, red jungle fowl, sheep, mouse, and hamster, involves expression of the thyroid-stimulating hormone (TSH)β in the pars tuberalis (PT) stimulated by the extension in day length (Nakao et al., 2008; Ono et al., 2008; Dardente et al., 2010; Hanon et al., 2010; Masumoto et al., 2010). TSH secreted by the PT induces the expression of type 2 deiodinase, which is an enzyme converting thyroxine into triiodothyronine (Nakao et al., 2008). Triiodothyronine is synthesized locally in the mediobasal hypothalamus byty pe 2 deiodinase, which increases the secretion of gonadotropin releasing hormone that then induces the secretion of both follicle-stimulating hormone and luteinizing hormone, which are involved in the development of gonads (Yoshimura et al., 2003; Watanabe et al., 2004). Furthermore, it is clear that the expression of TSHβ mRNA encoding TSH in the PT of mouse and sheep is regulated by eyes absent 3 (Eya3) genes, which are transcriptional coactivators preceding TSHβ mRNA expression (Dardente et al., 2010; Masumoto et al., 2010). In quail, both TSHβ and EYA3 expression is induced, like in mammals, by long-day stimulation (Nakao et al., 2008); however, the mechanism of EYA3 protein triggering TSHβ mRNA expression in the quail PT is unclear.
Eya is reported to be necessary for the formation of the eyes in Drosophila melanogaster; the Eya gene family includes Eya1-Eya4 in mammals (Jemc and Rebay, 2007). Eya3 includes two domains, namely, the Eya domain (ED) and Eya domain 2 (ED2); in the C terminal, ED is highly conserved among the Eya family members (Xu et al., 1997b; Zimmerman et al., 1997). The ED interacts with sine oculis homeobox (SIX) and dachshund (DACH) proteins (Ohto et al., 1999; Ikeda et al., 2002; Li et al., 2003) and has tyrosine phosphatase activity (Li et al., 2003; Rayapureddi et al., 2003; Tootle et al., 2003). In the N-terminal region, Eyas have threonine phosphatase activity (Okabe et al., 2009; Sano and Nagata, 2011). Furthermore, Eya1, Eya2, and Eya4 that contain proline, serine, and threonine-rich (P/S/T-rich) regions include the ED2 domain and two MAPK phosphorylation consensus sites, which are important for the transactivation function that the activation of the RAS/MAPK pathway potentiates in the transcriptional output of EYA (Xu et al., 1997a; Silver et al., 2003). Thus, the Eya family members possess both transcriptional coactivator function and tyrosine/threonine phosphatase function, and theyare involved in embryogenesis and organogenesis. However, the function of quail EYA3 in the photoperiodic response is not well understood. In the present study, to clarify a function of Eya3 in quail photoperiodism, we performed molecular structure analysis of Eya3 mRNA.
Materials and Methods
Animals and Tissue Collection
Japanese quails (4-weeks-old) were obtained from Motoki Corporation, Saitama, Japan. The quails were maintained under the short-day (SD) condition (6 h light and 18 h dark) for 4 weeks, and then divided into two groups: one group was maintained under SD conditions and the other group was transferred to long-day (LD) conditions (20 h light and 4 h dark). Samples of the mediobasal hypothalamus including the PT were collected at 16 h after dawn of SD or on the first long day (1LD). From the 1LD group, the retina, heart, spleen, lungs, liver, kidneys, testes, gall bladder, skeletal muscle, ovaries, and skin were collected after decapitation and stored at −80°C until further analysis. Food and water were provided ad libitum. All procedures were conducted in accordance with the provision for animal welfare of the Nippon Veterinary and Life Science University (approval number: 30S-50).
RNA Isolation and cDNA Synthesis
The total RNA from each tissue was extracted using TRIzol (Thermo Fisher Scientific, Waltman, MA, USA) and reverse transcribed at 50°C for 60 min with Super ScriptIII RT (Thermo Fisher Scientific). The polymerase chain reaction (PCR) for cDNA cloning of the coding region of EYA3, and reverse transcription-PCR (RT-PCR) and Southern blotting were performed with Ex Taq (TAKARA, Tokyo, Japan). The PCR was performed for 30 cycles at 96°C for 10 s, 64°C for 20 s, 72°C for 2 min 30 s (cDNA cloning), 94°C for 10 s, 52.9°C for 20 s, and 72°C for 1 min (RT-PCR and Southern blotting). The sequence of primers is shown in Table 1. The amplified cDNA fragments were ligated to the pGEM-T EasyVector (Promega, Madison, WI, USA) and transformed into Escherichia coli. The plasmid DNA was extracted from the bacteria and purified using an alkaline lysis method. The sequence of the cDNA was analyzed using the ABI PRISM 310 Genetic Analyzer (Thermo Fisher Scientific, Tokyo, Japan) with the BigDye Terminator v3.1 sequence kit (Thermo Fisher Scientific).
Table 1. Primers used for PCR.
| cDNA | Sequence (5′-3′) | |
|---|---|---|
| qEYA3* | Forward | GAATCATGGAAGAGCCACAAGATTTACCC |
| Reverse | GTCGACGTTACAGGAAGTCTAACTCGAGGG | |
| qEYA3** | Forward | ATCGGAAACAATGAGCCCTTAAC |
| Reverse | ACAGAGGGAAAAGGAAAGC | |
| Exon 7† | Forward | GGTGCACTGTGGCCAGGTAT |
| Reverse | AACCCTGTAGTGCAGGTAAGAACAC |
Applied for cDNA cloning.
Applied for RT-PCR Southern blot.
Applied for labelled DNA fragment for Southern blot.
RT-PCR
PCR was performed to amplify regions from exons 6 to 10 of EYA3 mRNA under the 1LD condition. The PCR products from the retina, heart, spleen, lungs, liver, kidneys, testes, gall bladder, skeletal muscle, ovaries, and skin were separated using 1.5% agarose electrophoresis. The digital images of electrophoresis gels were acquired using Molecular Imager FX (Bio-Rad, Hercules, CA, USA).
Southern Blotting
The PCR products were separated using 1.5% agarose electrophoresis, and then blotted onto the Hybond-N+ membrane (GE Healthcare, Chicago, IL, USA) after alkaline denaturation for 16 h. The blotted membranes were fixed by UV crosslinking. Labeling of DNA probes with alkaline phosphatase and hybridization were performed using the AIkPhos Direct Labelling and Detection System (GE Healthcare) according to the manufacturer's protocol. The membranes were hybridized with alkaline phosphatase-labeled DNA fragments derived from the exon 7 region (92 bp) of EYA3 cDNA at 55°C for 16 h. After washing twice with primarywash buffer (2M urea, 0.1% SDS, 150 mM NaCl, 1 mM MgCl2, 0.2% blocking reagent, and 50 mM phosphate buffer; pH 7.0) at 55°C for 10 min, the membranes were washed twice with secondary wash buffer (2M NaCl, 2 mM MgCl2, and 1M Tris-HCl buffer; pH 9.5) at 25°C for 5 min. After treating the membranes with CDP-Star Detection Reagent (GE Healthcare), an Eya3 mRNA splice variant containing exon 7 was detected using the LumiCube chemiluminescence analyzer (Liponics, Inc., Tokyo Japan) and quantified using JustTLC image analysis software (Sweday, Södra Sandby, Sweden).
Statistical Analysis
All data were analyzed using the Mann–WhitneyU test with the statistics software GraphPad Prism version 4 (GraphPad Software, USA). Statistical significance was set at a P-value of less than 0.05.
Results
Identification of EYA3Splice Variants in Quail
There were four splice variants (exons 6, 7, 8, 9, and 10; exons 6, 8, 9, and 10; exons 6, 7, and 10; and exons 6 and 10) in the domain from exons 6 to 10 of EYA3. Multiple sequence alignments showed the exon 7 region of Eya3 is not present in mouse and sheep (Fig. 1). In the four Eya3 splice variants, the ED in the C-terminal domain, and Cys-57, Tyr-78, His-80, and Tyr-93 were well conserved in Eya3 (Fig. 1).
Fig. 1.

Comparison of amino acid sequences of Eya3 in chicken, quail, mouse, and sheep. Asterisks indicate identical amino acids. The arrowheads indicate potential residues, Cys-56, Tyr-77, His-79, and Tyr-90, involved in the threonine–phosphatase activity of mouse Eya3. The black circles indicate proline, serine, and threonine in exon 7. Boxed region indicates the Eya domain (ED). Bold numbers indicate the number of exons, referenced from the chicken ensemble database.
Tissue Distribution of EYA3Splice Variant mRNA in Quail
Because the exon 7, 8, and 9 regions of EYA3 mRNA were spliced out, the level of EYA3 mRNA splice variant containing exons 7, 8, and 9 in the PT, eye, heart, spleen, lungs, liver, kidneys, gallbladder, skeletal muscle, testes, ovary, and skin was determined using RT-PCR analysis. The splice variants containing exons 6, 8, 9, and 10 (489 bp), lacking exon 7, were observed in all the tissues. Furthermore, a low expression of spliced variants of exons 7, 8, and 9 was also observed in all the tissues. In contrast, a splice variant containing exon 7 was expressed onlyin the PT and skeletal muscle. Notably, the expression of a splice variant containing exons 6, 7, 8, 9, and 10 was higher in the PT than in the other tissues under the 1LD condition (Fig. 2).
Fig. 2.
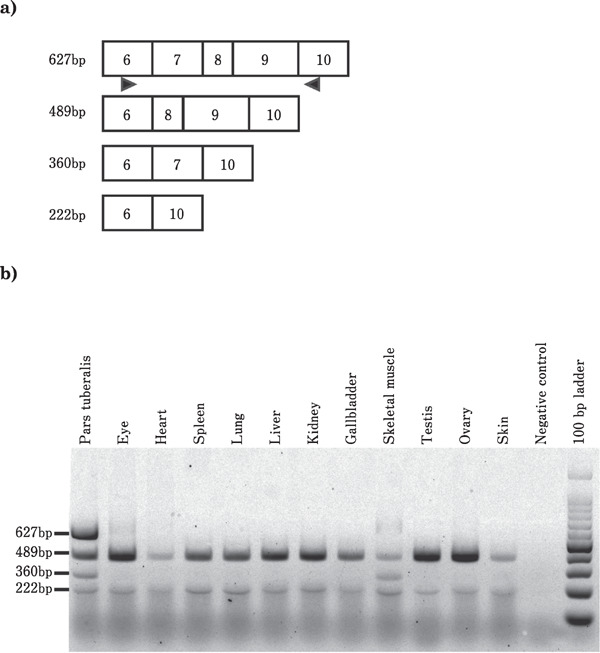
Expression profiles of EYA3 mRNA splice variants in quail tissues analyzed by RT-PCR. PCR was performed to amplif yregions from exons 6 to 10 of EYA3 mRNA under the 1LD condition. a: Schematic representation of the cDNA structure of EYA3 mRNA splice variants. Boxed region indicates the exon. The numerals in the box indicate the number of exons. The 627 bp, 489 bp, 360 bp, and 222 bp indicate amplified cDNA size. The arrowheads show the position of primers used for PCR. b: Tissue distribution of splice variants determined using 1.5% gel electrophoresis. bp indicates base pair.
Expression of EYA3Splice Variants was Day Length-dependent in the Quail PT
Splice variants of EYA3 in the PT under SD and 1LD conditions determined by RT-PCR and Southern blotting showed that a 627 bp variant containing exons 6, 7, 8, 9, and 10 and a 360 bp variant containing exons 6, 7, and 10 were significantly elevated under the 1LD condition compared with that under the SD condition (p<0.01, n=5) (Fig. 3).
Fig. 3.
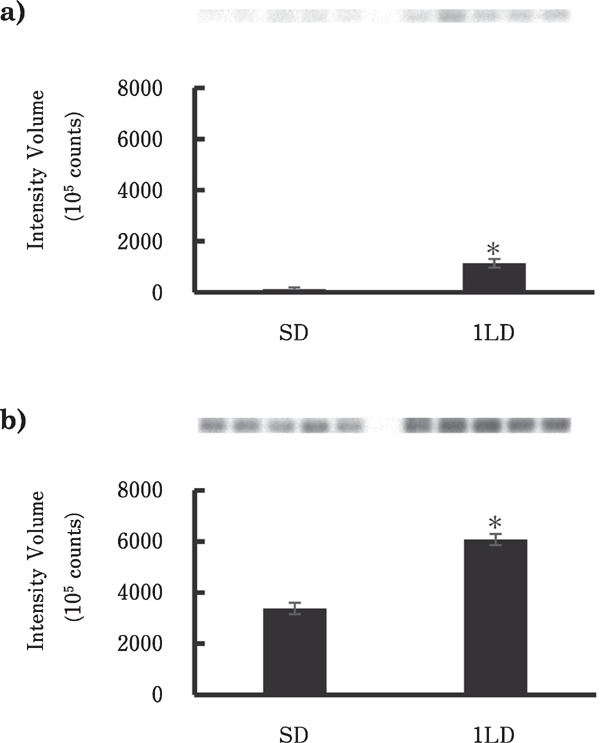
EYA3 mRNA containing exon 7 in the quail pars tuberalis under the SD and 1LD conditions. PCR was performed to amplify regions from exons 6 to 10 of EYA3 mRNA. The primer position is shown in Fig. 2 and the probe for Southern blotting was 92 bp of EYA3 exon 7. Each bar represents the mean±SEM (n=5). The upper panel of the graph shows the signal containing exon 7 of EYA3 detected by chemiluminescence. SD and 1LD indicate short daycondition and first long daycondition, respectively. a: Expression of exons 6, 7, and 10 (360 bp) under the SD and 1LD conditions. b: Expression of exons 6, 7, 8, 9, and 10 (627 bp) under the SD and 1LD conditions. Asterisks indicate significant difference (P<0.01).
Discussion
Here, the structural analysis of EYA3 mRNA elucidated the function of EYA3 in the photoperiodic response of the Japanese quail. There are four types of EYA3 mRNA splice variants. Interestingly, exon 7, which is expressed in quail, is not present in mouse and sheep. A splice variant of EYA3 containing exon 7 might participate in the function of the encoded protein in the photoperiodic response of quail. In the C-terminal of EYA3, the ED is conserved among the four splice variants of EYA3. Residues Cys-56, Tyr-76, His-79, and Tyr-90 in mouse Eya3 are known to be essential for the threonine-phosphatase activity (Sano and Nagata, 2011). In quail, residues Cys-57, Tyr-78, His-80, and Tyr-93 were well conserved among the four splice variants of EYA3. Taken together, the four splice variants of EYA3 have conserved sequences that function as transcriptional coactivators and tyrosine and threonine phosphatases; however, their functions in the quail are unknown.
The tissue distribution of EYA3 mRNA including exon7 demonstrated that the expression of mRNA with exon 7 spliced out was widespread and was similar to the expression pattern of mouse Eya3 (Zimmerman et al., 1997); however, the variant containing exon 7 was only expressed in the PT and skeletal muscle. In particular, the mRNA expression of EYA3 containing exons 7, 8, and 9 in the PT has been known to playa central role in photoperiodic control, and its expression was higher than that in the skeletal muscle. Moreover, comparative expression analysis of EYA3 in the PT under the SD and 1LD conditions demonstrated that EYA3 including exon 7 was the splice variant of the 1LD-specific gene. The amino acid sequence that exon 7 encodes corresponds to none of the domains that are necessary for the threonine phosphatase activity of Eya3 (Sano and Nagata, 2011), and it involves ED and ED2, which are specific to the Eya family members (Zimmerman et al., 1997). In contrast, the amino acid sequence of exon 7 was rich in proline, serine, and threonine, which is similar to the P/S/T-rich domain of the Eya family members, and might be involved in the transactivation function (Xu et al., 1997a; Silver et al., 2003).
The photoperiodic control in mammals involves TSH, which is regulated by Eya3 expression before TSHβ expression (Dardente et al., 2010; Masumoto et al., 2010). In quail, the peak expression of EYA3 was almost concomitant with TSHβ expression under the 1LD; EYA3 expression decreased after the second long day under the 2-week-long day condition (Nakao et al., 2008). To clarify how the transcriptional activity of TSHβ mRNA is affected by EYA3 under different long photoperiod stimuli, it is necessary to perform future studies on the subcellular localization of EYA3 and TSHβ based on in situ hybridization and the homoor/and heterotypic dimerization-driven activation of EYA3 protein.
In conclusion, we identified four types of splice variants of Japanese quail EYA3 mRNA and showed that the splice variants containing exon 7 were expressed in the PT by the first long daystimulation. The exon 7 of EYA3 is considered to be an important key element for elucidating the action of EYA3 in the quail photoperiodic response. Detailed analysis of exon 7 is important to clarify the role of EYA3 in the photoperiodic response.
Acknowledgments
This work was supported in part by JSPS KAKENHI (Grant Number JP22780266).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dardente H, Wyse CA, Birnie MJ, Dupré SM, Loudon AS, Lincoln GA, Hazlerigg DG. A molecular switch for photoperiod responsiveness in mammals. Current Biology, 20: 2193-2198. 2010. [DOI] [PubMed] [Google Scholar]
- Hanon EA, Routledge K, Dardente H, Masson-Pévet M, Morgan PJ, Hazlerigg DG. Effect of photoperiod on the thyroid-stimulating hormone neuroendocrine system in the European hamster (Cricetus cricetus). Journal of Neuroendocrinology, 22: 51-55. 2010. [DOI] [PubMed] [Google Scholar]
- Ikeda K, Watanabe Y, Ohto H, Kawakami K. Molecular interaction and synergistic activation of a promoter by Six, Eya, and Dach proteins mediated through CREB binding protein. Molecular and Cellular Biology, 22: 6759-6766. 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jemc J, Rebay I. The eyes absent family of phosphotyrosine phosphatases: properties and roles in developmental regulation of transcription. Annual Review of Biochemistry, 76: 513-538. 2007. [DOI] [PubMed] [Google Scholar]
- Li X, Oghi KA, Zhang J, Krones A, Bush KT, Glass CK, Nigam SK, Aggarwal AK, Maas R, Rose DW, Rosenfeld MG. Eya protein phosphatase activity regulates Six1-Dach-Eya transcriptional effects in mammalian organogenesis. Nature, 426: 247-254. 2003. [DOI] [PubMed] [Google Scholar]
- Masumoto KH, Ukai-Tadenuma M, Kasukawa T, Nagano M, Uno KD, Tsujino K, Horikawa K, Shigeyoshi Y, Ueda HR. Acute induction of Eya3 by late-night light stimulation triggers TSHβ expression in photoperiodism. Current Biology, 20: 2199-2206. 2010. [DOI] [PubMed] [Google Scholar]
- Nakao N, Ono H, Yamamura T, Anraku T, Takagi T, Higashi K, Yasuo S, Katou Y, Kageyama S, Uno Y, Kasukawa T, Iigo M, Sharp PJ, Iwasawa A, Suzuki Y, Sugano S, Niimi T, Mizutani M, Namikawa T, Ebihara S, Ueda HR, Yoshimura T. Thyrotrophin in the pars tuberalis triggers photoperiodic response. Nature, 452: 317-322. 2008. [DOI] [PubMed] [Google Scholar]
- Ohto H, Kamada S, Tago K, Tominaga SI, Ozaki H, Sato S, Kawakami K. Cooperation of six and eya in activation of their target genes through nuclear translocation of Eya. Molecular and cellular biology, 19: 6815-6824. 1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okabe Y, Sano T, Nagata S. Regulation of the innate immune response by threonine-phosphatase of Eyes absent. Nature, 460: 520-524. 2009. [DOI] [PubMed] [Google Scholar]
- Ono H, Hoshino Y, Yasuo S, Watanabe M, Nakane Y, Murai A, Ebihara S, Korf HW, Yoshimura T. Involvement of thyrotropin in photoperiodic signal transduction in mice. Proceedings of the National Academy of Sciences of the United States of America, 105: 18238-18242. 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rayapureddi JP, Kattamuri C, Steinmetz BD, Frankfort BJ, Ostrin EJ, Mardon G, Hegde RS. Eyes absent represents a class of protein tyrosine phosphatases. Nature, 426: 295-298. 2003. [DOI] [PubMed] [Google Scholar]
- Sano T, Nagata S. Characterization of the threonine-phosphatase of mouse eyes absent 3. FEBS Letters, 585: 2714-2719. 2011. [DOI] [PubMed] [Google Scholar]
- Silver SJ, Davies EL, Doyon L, Rebay I. Functional dissection of eyes absent reveals new modes of regulation within the retinal determination gene network. Molecular and Cellular Biology, 23: 5989-5999. 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tootle TL, Silver SJ, Davies EL, Newman V, Latek RR, Mills IA, Selengut JD, Parlikar BE, Rebay I. The transcription factor Eyes absent is a protein tyrosine phosphatase. Nature, 426: 299-302. 2003. [DOI] [PubMed] [Google Scholar]
- Watanabe M, Yasuo S, Watanabe T, Yamamura T, Nakao N, Ebihara S, Yoshimura T. Photoperiodic regulation of type 2 deiodinase gene in Djungarian hamster: possible homologies between avian and mammalian photoperiodic regulation of reproduction. Endocrinology, 145: 1546-1549. 2004. [DOI] [PubMed] [Google Scholar]
- Xu PX, Cheng J, Epstein JA, Maas RL. Mouse Eya genes are expressed during limb tendon development and encode a transcriptional activation function. Proceedings of the National Academy of Sciences of the United States of America, 94: 11974-11979. 1997. a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu PX, Woo I, Her H, Beier DR, Maas RL. Mouse Eya homologues of the Drosophila eyes absent gene require Pax6 for expression in lens and nasal placode. Development, 124: 219-231. 1997. b. [DOI] [PubMed] [Google Scholar]
- Yoshimura T, Yasuo S, Watanabe M, Iigo M, Yamamura T, Hirunagi K, Ebihara S. Light-induced hormone conversion of T4 to T3 regulates photoperiodic response of gonads in birds. Nature, 426: 178-181. 2003. [DOI] [PubMed] [Google Scholar]
- Zimmerman JE, Bui QT, Steingrímsson E, Nagle DL, Fu W, Genin A, Spinner NB, Copeland NG, Jenkins NA, Bucan M, Bonini NM. Cloning and characterization of two vertebrate homologs of the Drosophila eyes absent gene. Genome Research, 7: 128-141. 1997. [DOI] [PubMed] [Google Scholar]