

Abstract
Mycobacterium tuberculosis (Mtb) is the deadliest bacterial pathogen in the world. An estimated one-third of humans harbor Mtb in a dormant state. These asymptomatic, latent infections impede tuberculosis eradication due to the long-term potential for reactivation. Dormant Mtb has reduced enzymatic activity, but hydrolases that remain active facilitate pathogen survival. We targeted Mtb esterases, a diverse set of enzymes in the serine hydrolase family, and studied their activities using both activity-based probes (ABPs) and fluorogenic esterase substrates. These small-molecule probes revealed functional esterases in active, dormant, and reactivating cultures. Using ABPs, we identified five esterases that remained active in dormant Mtb, including LipM (Rv2284), LipN (Rv2970c), CaeA (Rv2224c), Rv0183, and Rv1683. Three of these, CaeA, Rv0183, and Rv1683, were catalytically active in all three culture conditions. Fluorogenic probes additionally revealed LipH (Rv1399c), Culp1 (Rv1984c), and Rv3036c esterase activity in dormant and active cultures. Esterases with persistent activity are potential diagnostic biomarkers or therapeutic targets for Mtb-infected individuals with latent or active tuberculosis.
Keywords: Mycobacterium tuberculosis, esterase, chemical biology, proteomics, fluorescent
Graphical Abstract

Mycobacterium tuberculosis (Mtb) is the causative agent of tuberculosis (TB), the world’s most lethal single-agent infectious disease.1 An estimated 2 billion people harbor latent TB, an asymptomatic form of the disease. In latent TB, Mtb slows its metabolism and converts to a dormant, nonreplicating state.2,3 Dormant Mtb is phenotypically drug-tolerant, which contributes to the prolonged, multidrug treatment required to cure TB.6 Furthermore, common TB screening methods do not distinguish between latent and active infections.2,3 Approximately 10% of latently infected individuals will transition to active TB.4
To investigate latent TB biology in vitro, researchers subject bacilli to stresses encountered in the human host, including hypoxia, nutrient deprivation, reactive nitrogen species, and acidic pH.5 Mtb adapts to these stresses by converting to a nonreplicating state that mimics latent infection. Dormant Mtb has decreased metabolism,6 phenotypic drug resistance,5,7 and a substantially altered transcriptome6,8,9 and proteome.10,11
Survival in dormancy and during reactivation relies on enzymes that process fatty acids as an energy source.7,12−14 These enzymes—including esterases and the lipid-preferring lipase subclass—are ideal drug targets due to their roles in Mtb persistence.15−18 Esterase regulation in dormancy has been inferred primarily from transcriptomic8,9,13 and proteomic10,11 data. However, those studies cannot reveal whether or not an esterase is catalytically active. The activity of serine hydrolases, including esterases, can be post-translationally regulated.19,20 Three Mtb esterases—LipE, LipN, and cutinase-like protein (Culp) 2—contain acetylated lysines, which could influence their activity.21 LipI and LipN can be phosphorylated at their catalytic serine, which could be a mechanism for regulating catalytic activity.22 Overall, it is unclear which of the approximately 40 Mtb esterases are active in dormant and reactivating Mtb.
Functional enzymes can be detected and identified in Mtb using activity-based probes (ABPs), which are irreversible, active-site targeted small molecules (Figure 1).23,24 Thus far, two studies have identified mycobacterial esterases through activity-based protein profiling (ABPP). Wenk and co-workers used an ABP to identify the targets of tetrahydrolipstatin (THL), a lipase and fatty acid synthase inhibitor with anti-Mtb activity.25 They found 14 THL targets in normoxic (replicating) and hypoxic (dormant) cultures of Mycobacterium bovis (BCG), a close genetic relative of Mtb. Interestingly, they did not identify two validated mycobacterial targets of THL: Culp626 or LipY.27
Figure 1.
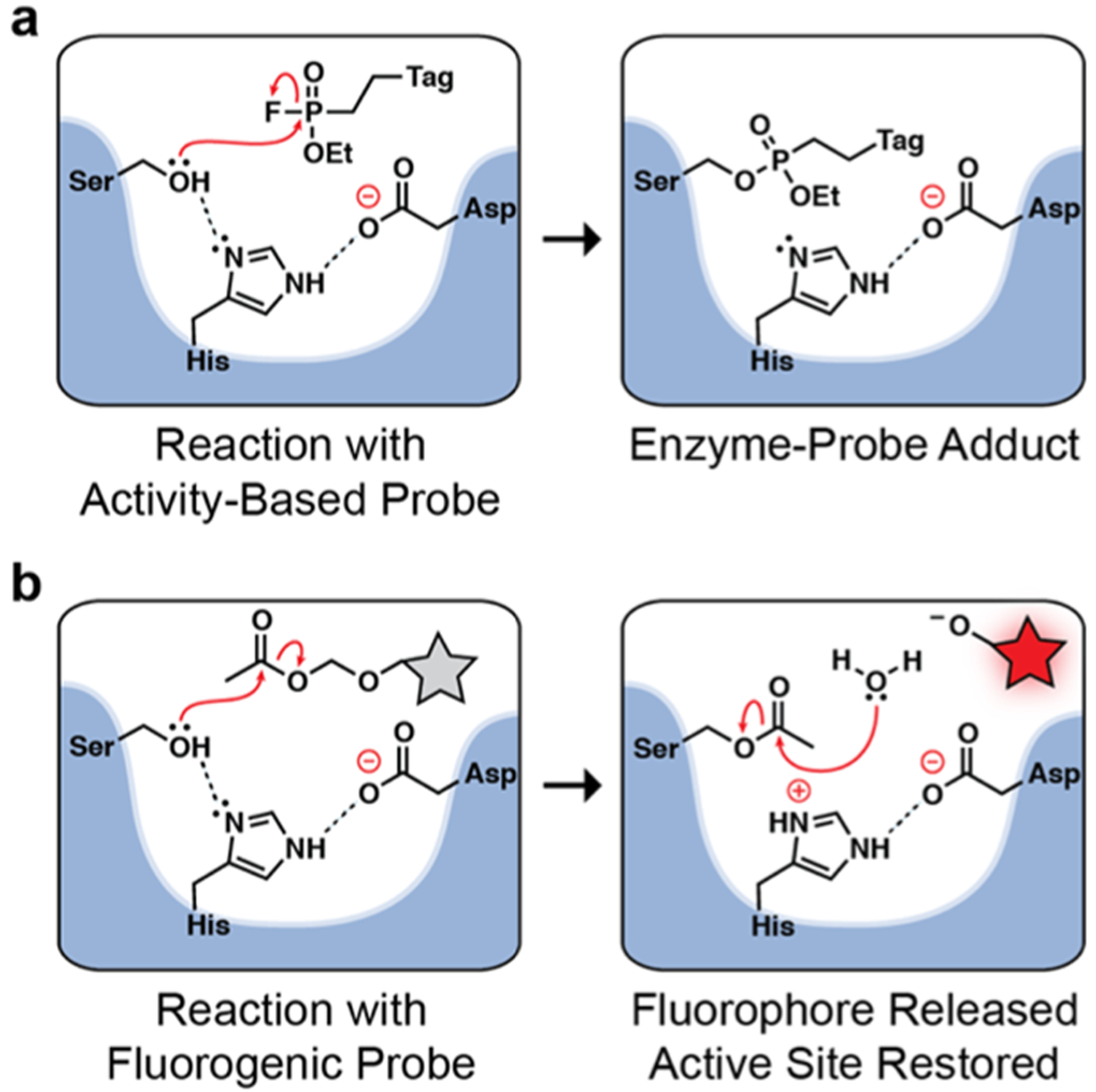
Small-molecule probes for revealing active Mtb esterases. (a) ABPs form a covalent bond with an active site serine for irreversible serine hydrolase labeling. ABPs contain a tag for direct detection or for affinity enrichment (i.e., for identification by mass spectrometry-based proteomics). (b) Fluorogenic esterase probes are substrates that produce fluorescence upon serine-mediated hydrolysis. Following probe cleavage, the active site is restored.
In 2016, Grundner et al. used a serine-reactive fluorophosphonate (FP) ABP to identify serine hydrolases in dormant and replicating Mtb cultures.19 They identified 78 proteins with serine hydrolase activity of the 186 putative Mtb serine hydrolases (see Table S128). They also demonstrated that Mtb serine hydrolase abundance and activity are poorly correlated. Unlike Wenk et al., they found Culp6 in both normoxic and hypoxic cultures. Similar to the prior study, they did not find LipY, a triacylglycerol (TAG) lipase that is transcriptionally up-regulated in dormant Mtb.8,13
The Mtb esterases and lipases are members of a diverse group of serine hydrolases that cannot be identified using a single small-molecule probe. With FP probes, even the linker can influence probe reactivity.29 As a complement to ABPs, we previously developed fluorogenic probes for detecting Mtb hydrolases in a native polyacrylamide gel electrophoresis (PAGE) assay.30−33 Unlike the stoichiometric (1:1) labeling of ABPs, fluorogenic probes are hydrolyzed to produce an amplifiable fluorescent signal that correlates with enzyme activity (Figure 1b).24 We varied both the fluorescent reporter32 and the masking group33 to increase the diversity of enzymes we could detect. Most recently,33 we described a fluorogenic lipase substrate, DDAO-OME, which revealed Culp1 activity in Mtb—an esterase missing from the two prior ABPP studies.
In the present study, we used chemically diverse ABPs and fluorogenic probes to obtain a more comprehensive list of Mtb esterases that retain activity under replicating and nonreplicating conditions. For the first time, we identified esterases that are functional during the early hours of reactivation from dormancy, a key pathophysiological transition. We used desthiobiotin-FP, an ABP, to identify 22 esterases in replicating, normoxic Mtb—the most comprehensive hit list to date. Of these esterases, 5 retained activity in hypoxia, and 3 were active during reactivation. We used our fluorogenic probes to confirm esterase functionality and to detect esterases that were missed by ABPs (e.g., Rv3036c). We propose that esterases with persistent activity in dormancy and reactivation are valuable targets for new diagnostic assays or therapeutics.
RESULTS AND DISCUSSION
TAMRA-FP Reveals Serine Hydrolase Activity Is Down-regulated in Dormancy.
We initiated our research using red-fluorescent TAMRA-FP (see Figure S1 for structure) to detect serine hydrolases in replicating, normoxic cultures of Mtb mc26020 (an auxotrophic strain of H37Rv33). TAMRA-FP labeling was reduced by pre-incubating the lysate with covalent serine hydrolase inhibitors, demonstrating that active enzyme was required for probe labeling (Figure 2). Methyl arachidonyl FP (MAFP), a serine hydrolase inhibitor with an FP warhead, blocked nearly all TAMRA-FP labeling. Diethyl-p-nitrophenyl phosphate (E-600, a carboxyl esterase inhibitor), phenylmethanesulfonyl fluoride (PMSF, a serine protease inhibitor), and THL each inhibited some of the serine hydrolases. No two inhibitors targeted the same set of enzymes, highlighting the diversity of the Mtb serine hydrolases. We also treated lysates with ebselen, a cysteine-reactive allosteric inhibitor of the antigen 85 (Ag85) complex.34,35 Due to ebselen’s specificity, we concluded that the region indicated with a star corresponds to Ag85-associated serine hydrolase activity (e.g., Ag85A, Ag85B, and/or Ag85C).
Figure 2.
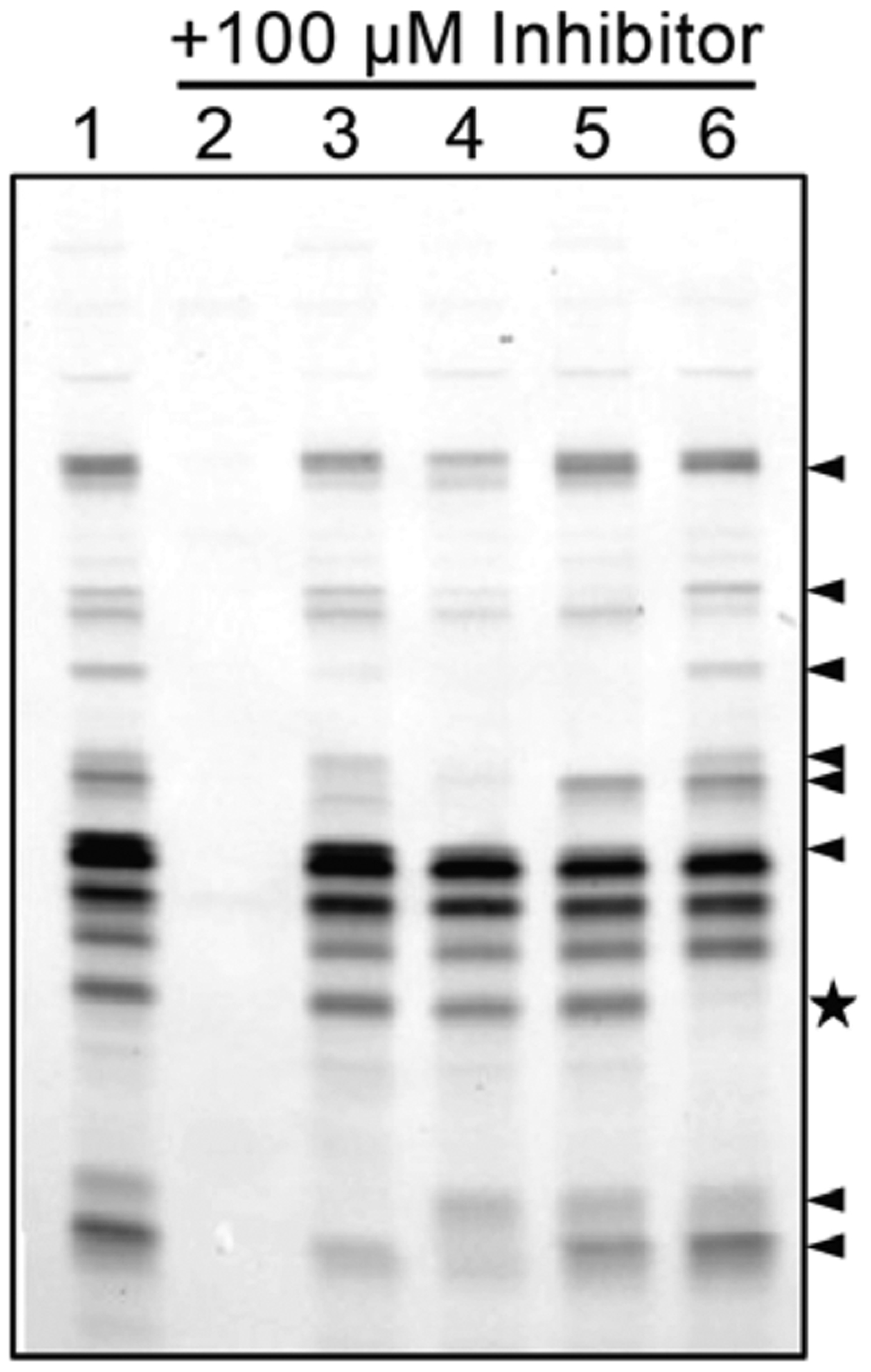
TAMRA-FP enables detection of active serine hydrolases in replicating Mtb. To inhibit activity, lysates were pretreated with various inhibitors or DMSO (vehicle control) prior to TAMRA-FP labeling. Lanes: 1, DMSO; 2, MAFP; 3, PMSF; 4, E-600; 5, THL; 6, ebselen. Arrowheads highlight some of the differences in banding between PMSF, E-600, and THL inhibition. The star highlights the likely location of the antigen 85 complex. Lysates were resolved on a 12% Bis-Tris gel in MOPS buffer.
Next, we induced Mtb dormancy using two different models: carbon starvation36 and hypoxia.37 Carbon starvation was induced by culturing Mtb in unsupplemented medium for 5 weeks, as described by Hung and co-workers.36 Hypoxia was induced in standing flasks using a hypoxia chamber (1% O2 and 5% CO2) for 2 weeks. We allowed hypoxic cultures of dormant Mtb to reactivate for 4.5 h using two different reaeration conditions. In one, we reaerated in rich growth medium. In the other, we reaerated under carbon starvation to encourage the bacteria to utilize stored lipids (i.e., TAG).13 We chose 4.5 h as our time point to observe rapid changes in serine hydrolase activity during initial reactivation. We collected cells from each condition, treated the resulting lysates with TAMRA-FP, and resolved the samples by SDS-PAGE.
TAMRA-FP labeling revealed substantial down-regulation of serine hydrolase activity in the hypoxic (lane 2) and carbon-starved (lane 5) cultures compared to the normoxic culture (lane 1) (Figure 3). Lysates from carbon-starved Mtb displayed a more pronounced decrease in serine hydrolase activity than the hypoxic culture. After reaeration from hypoxia in rich medium (lane 3), some serine hydrolase activity was restored. By comparison, changes in activity were undetectable for reactivation under carbon starvation (lane 4), which could reflect a slower recovery from dormancy. Overall, the TAMRA-FP probe confirmed that serine hydrolase activities were modulated in response to environmental changes.
Figure 3.
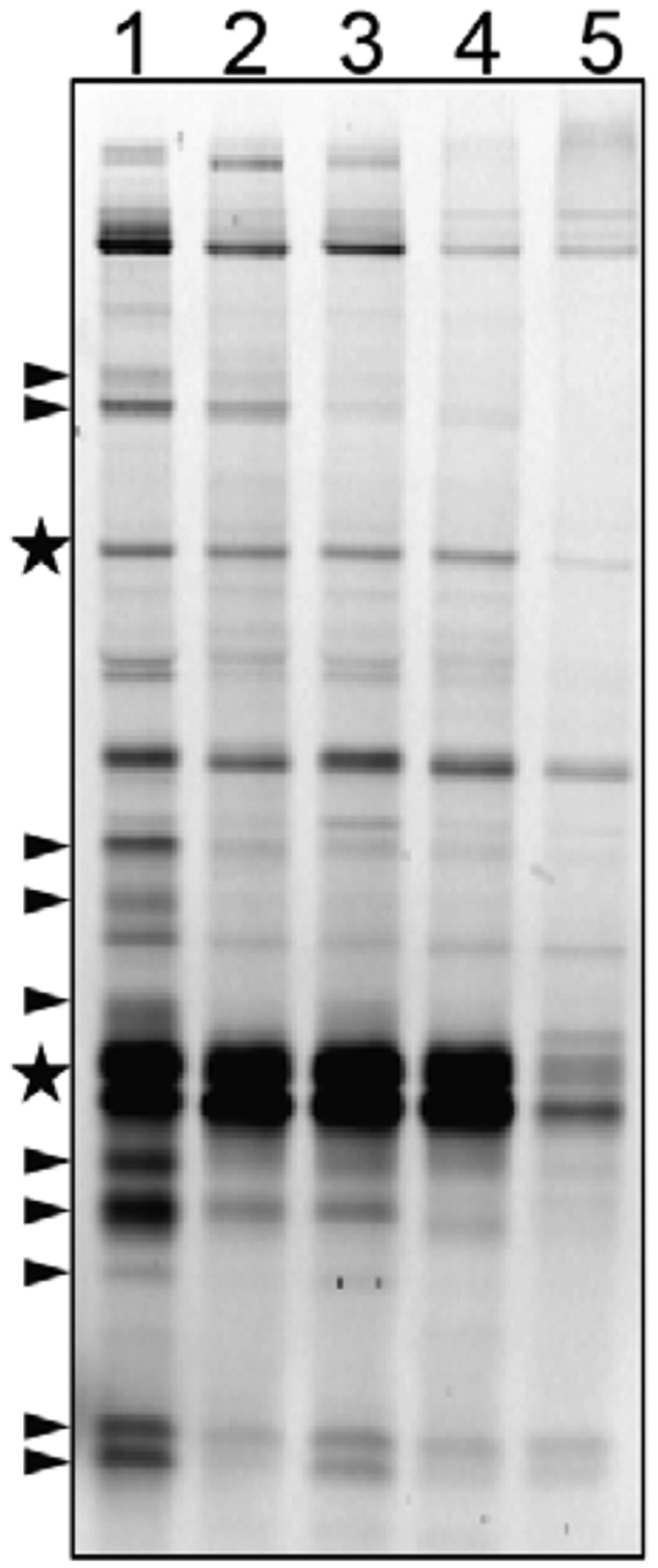
Serine hydrolase activity is reduced in dormancy. Lysates from normoxic, dormant, and reaerated cultures were treated with TAMRA-FP and then resolved by SDS-PAGE with XT-MES buffer. Lanes: 1, normoxic lysates; 2, hypoxic lysates; 3, reaerated lysates, reactivated from hypoxia in rich growth medium; 4, reaerated lysates, reactivated from hypoxia under carbon starvation; 5, carbon-starved lysates. The arrowheads highlight some of the bands with decreased activity in dormancy. The stars indicate bands that have substantially decreased activity under carbon starvation but not hypoxia.
Desthiobiotin-FP Facilitates Active Serine Hydrolase Identification.
ABPs are powerful tools for enriching subsets of the proteome for mass spectrometry-based identification. We labeled lysates with desthiobiotin-FP to identify Mtb serine hydrolases active in replicating, dormant, and reactivating conditions. We induced dormancy by hypoxia (2 weeks) and reactivated cultures by reaeration (4.5 h) under carbon starvation.8,13 We evaluated our labeling and enrichment by Western blot, probing with an anti-biotin antibody (Figure S2). Although we observed endogenously biotinylated proteins, as previously described,38 there were additional biotinylated proteins in the desthiobiotin-FP-labeled samples. Biotinylated proteins were isolated by streptavidin affinity chromatography and digested with trypsin for liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis. LC-MS/MS analysis was performed by the University of California (UC)—Davis Proteomics Core (see the Supporting Information for details). Positive hits were selected on the basis of two or more distinct peptide hits per protein found in at least two of the three replicates.
We identified 183 distinct proteins, including 52 serine hydrolases (Table S2). The identified serine hydrolases ranged in functional annotation, including intermediary metabolism/respiration (26 proteins), lipid metabolism (11 proteins), cell wall/cell processes (7 proteins), conserved hypotheticals (6 proteins, including 2 putative esterases), and virulence/detoxification/adaptation (2 proteins). It is worth noting that the genome of Mtb is only partially annotated, and esterases fall into more than one of these categories. We identified fewer serine hydrolases in dormancy and reactivation compared to normoxic lysates (Figure 4), consistent with prior studies.19,39
Figure 4.
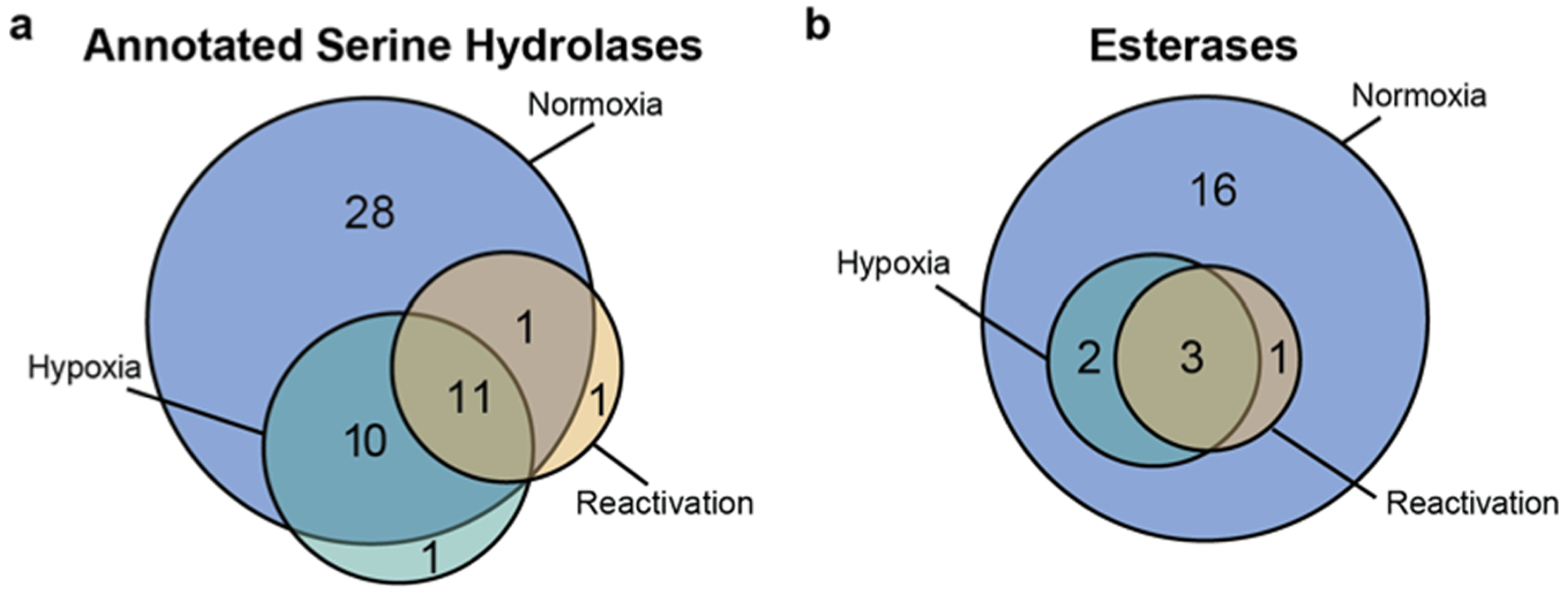
Diagrams of the number of ABPP hits associated with normoxic, hypoxic, and reactivated cultures: (a) distribution of the 52 enzymes bioinformatically annotated as serine hydrolases (for a list of annotated Mtb serine hydrolases, see Table S1); (b) distribution of the 22 active esterases identified in each condition (for a curated list of Mtb esterases, see Table S4).
Some Mtb Esterases Retain Activity in Dormancy.
We narrowed our focus to known esterases and lipases (see Table S4). We then compared our findings with desthiobiotin-FP to prior studies that used a THL-alkyne (Wenk)39 or an alkyne-PEG-FP (Grundner)19 (Table 1). Overall, Wenk identified fewer esterases because THL has a narrow reactivity profile. Many of the esterases we identified in our enrichment overlap with those found by Grundner et al. Their alkyne-PEG-FP probe revealed three esterases missed in our study (Culp6, Rv0421c, and Rv3591c), whereas we detected eight esterases missed by them (Culp1, Culp4, LipG, LipO, LipR, LipT, LipW, and Rv1191). The differential detection is possibly due to the alkyl chain of desthiobiotin-FP being a better lipid mimetic than the PEG of the alkyne-PEG-FP probe.29 This is supported by the fact that LipG, LipO, and LipW were also detected by Wenk et al. using a THL analogue. In Grundner’s work, they used a slightly different method to induce hypoxia, which could also contribute to our different results.
Table 1.
Active Mtb Esterases Revealed by ABPPa
| THL-alkyneb | alkyne-PEG-FPc | desthiobiotin-FP | |||||
|---|---|---|---|---|---|---|---|
| Rv Number | Esterase | N | H | N | H | N | H |
| Rv2224c | CaeA | X | X | X | X | X | X |
| Rv1984c | Culp1 | X | |||||
| Rv2301 | Culp2 | X | X | ||||
| Rv3452 | Culp4 | X | |||||
| Rv3802c | Culp6 | X | X | ||||
| Rv1923 | LipD | X | X | X | |||
| Rv3775 | LipE | X | X | ||||
| Rv0646c | LipG | X | X | ||||
| Rv1399c | LipH | X | X | X | |||
| Rv1400c | LipI | X | X | X | |||
| Rv1497 | LipL | X | X | X | |||
| Rv2284 | LipM | X | X | X | X | X | X |
| Rv2970c | LipN | X | X | X | X | X | |
| Rv1426c | LipO | X | X | ||||
| Rv3084 | LipR | X | |||||
| Rv2045c | LipT | X | |||||
| Rv3203 | LipV | X | X | X | |||
| Rv0217c | LipW | X | X | ||||
| Rv0045c | Rv0045c | X | X | ||||
| Rv0183 | Rv0183 | X | X | X | X | ||
| Rv0421c | Rv0421c | X | X | ||||
| Rv0774c | Rv0774c | X | X | ||||
| Rv1191 | Rv1191 | X | |||||
| Rv1683 | Rv1683 | X | X | X | X | ||
| Rv3591c | Rv3591c | X | X | ||||
| Total hits: | 10 | 2 | 17 | 9 | 22 | 5 | |
An “×” indicates that the esterase was identified in the corresponding condition for each study (N, normoxic; H, hypoxic). Esterases highlighted in green were found in all three studies in both N and H. Esterases highlighted in blue were uniquely identified in this study.
Wenk et al. performed ABPP in M. bovis (BCG) with a THL analogue, and their proteins are listed as the Mtb (H37Rv) orthologue.39
There were only two esterases identified by all three studies in both normoxic and hypoxic cultures: CaeA and LipM. This suggests that CaeA and LipM are potential therapeutic targets for both latent and active TB. In all three ABPP studies, LipD, LipI, and LipV were associated with replicating cultures, and these esterases could be used to distinguish active TB from latent disease.
Esterases that remain active in dormant Mtb could facilitate survival during dormancy. In all three ABPP studies, no esterases were found to be uniquely active during dormancy. We found five esterases that retained catalytic activity in dormancy: CaeA, LipM, LipN, Rv0183, and Rv1683. We identified only four esterases in Mtb undergoing reactivation from hypoxia: CaeA, LipR, Rv0183, and Rv1683. LipR was not identified in our hypoxia samples, and it is possible that LipR activity is rapidly restored during reactivation.
ABPP with desthiobiotin-FP enabled us to identify five esterases that were missed by prior studies: Culp1, Culp4, LipR, LipT, and Rv1191. Culp1 and Culp4 are notable findings because they are both functionally linked to Mtb dissemination.40,41 Immunization with recombinant Culp1 provides partial protection against TB,42 and supplementing the BCG vaccine with Culp1 and other antigens enhances the vaccine’s efficacy in mice.43 Furthermore, both Culp1 and Culp4 are being explored as potential TB immunodiagnostic biomarkers, with Culp4 having the greatest sensitivity and specificity of the examined enzymes.44
Our identification of CaeA, Rv0183, and Rv1683 in dormant, reactivating, and active cultures suggests to us that these esterases are worth further investigation as diagnostic or therapeutic targets for all stages of TB disease. CaeA is a cell wall-associated dual-function esterase45 and protease.46 It is required for full virulence in a mouse model of TB45 and is transcriptionally up-regulated under nutrient starvation.9 Rv0183 is a cell wall-associated monoacylglycerol (MAG) lipase that likely degrades host-cell lipids.47,48 MAG lipase inhibitors—including ones specific for Rv0183—are being pursued as antimicrobials.16−18 Rv1683 is annotated as a bifunctional long-chain acyl-CoA synthase and lipase, but has not yet been biochemically characterized. Rv1683 is essential for growth in vitro49 and is found in the cell membrane.50 In addition to their persistent activities, CaeA, Rv0183, and Rv1683 are accessible targets due to their localization.
Like Grundner and Wenk, we did not identify LipY by mass spectrometry-based proteomics. In over a dozen proteomic studies, LipY was identified only once.51 There are a few possible reasons why LipY eludes detection. LipY could be inactive, absent, or in low abundance—explanations that seem inconsistent with LipY’s transcriptional up-regulation in dormancy.8,13 We identified an additional explanation: LipY encodes only three tryptic cut sites. The first two sites are in the proline-glutamic acid (PE) domain, and the third is in a linker sequence. In Mtb, both the PE domain and the linker are proteolytically removed prior to secretion of the C-terminal lipase domain.52 Thus, these peptides would be obtained only from unprocessed, cytosolic LipY. We treated purified, recombinant LipY with trypsin and confirmed that the two peptides detected by MS are in the cleaved region (Figure S3).
Fluorogenic Probes Are Complementary to ABPP and Illuminate Rv3036c Activity.
Desthiobiotin-FP and alkyne-PEG-FP have the same reactive warhead but react with different Mtb esterases (Table 1). This is consistent with our prior observation that fluorogenic esterase probes are not “one size fits all” substrates.32,33 Thus, we further evaluated Mtb esterase activity in native protein gel-resolved lysates. Esterase activity patterns were revealed using a diverse set of fluorogenic esterase probes (see Figure S4 for structures). We used a new fluorogenic probe, 2′,7′-dichlorofluorescein diacetyoxymethyl ether (DCF-AME), and our previously validated probes (C2- masked DDAO-7-AME32 and C8-masked DDAO-7-OME33) for these studies.
DCF-AME and DDAO-7-AME revealed similar Mtb esterase activity patterns in normoxic lysates, but DCF-AME revealed a greater number of esterases (Figure 5). DDAO-7-OME revealed fewer esterases than DCF-AME and DDAO-7-AME but detected a band that we previously identified as Culp1.33 We excised additional fluorescent esterase bands for identification by MS-based proteomics and identified LipV, Rv3036c, Rv0045c, and LipH.
Figure 5.
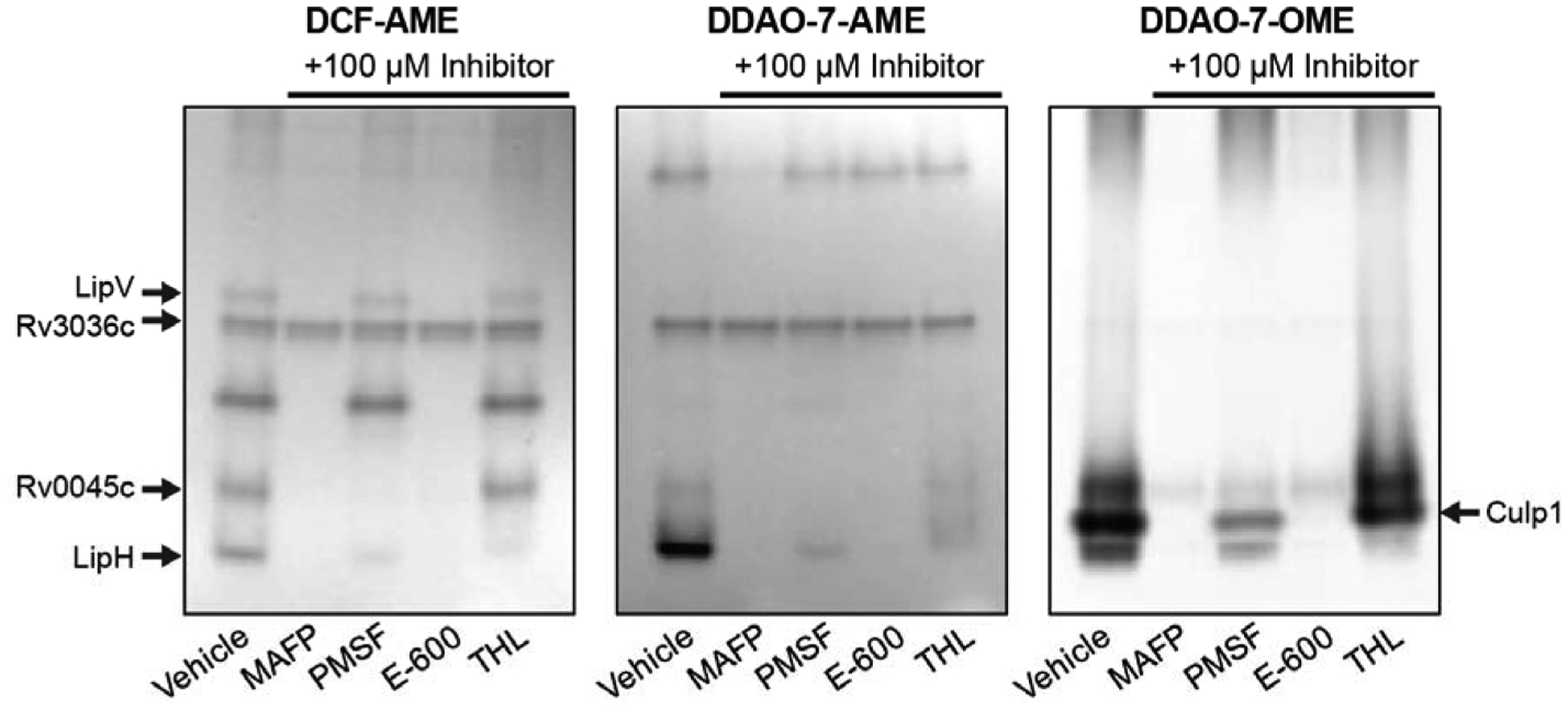
Fluorogenic probes detect active Mtb esterases. Lysates were pre-incubated with 100 μM of the indicated inhibitor for 1 h at room temperature and then resolved by native PAGE. Gels were soaked in 5 μM probe (5–10 min) and imaged on a fluorescence scanner. Esterases identified by mass spectrometry are indicated (see Table S5 for peptide hits).
We pre-incubated lysates with serine hydrolase inhibitors and found that some esterases were completely inhibited while others were partially inhibited (Figure 5). As in the TAMRA-FP labeling experiment (Figure 2), MAFP inhibited the greatest number of esterases, followed by E-600. PMSF and THL each inhibited only a few esterases.
These results highlight the power of using fluorogenic probes with a native PAGE assay for fast inhibitor screens.
Because MAFP did not inhibit all of the active esterases ingel, we analyzed samples treated with TAMRA-FP for residual activity with DCF-AME (Figure 6). Bright DCF fluorescence was observed for one enzyme, which we excised and identified as Rv3036c. Rv3036c is a cell wall-associated, secreted esterase that prefers two-carbon substrates. It has a noncanonical Ser-XX-Lys active site motif, which could affect its reactivity with FPs.53 Notably, this esterase retained activity under nonreplicating conditions (see Figure 7).
Figure 6.
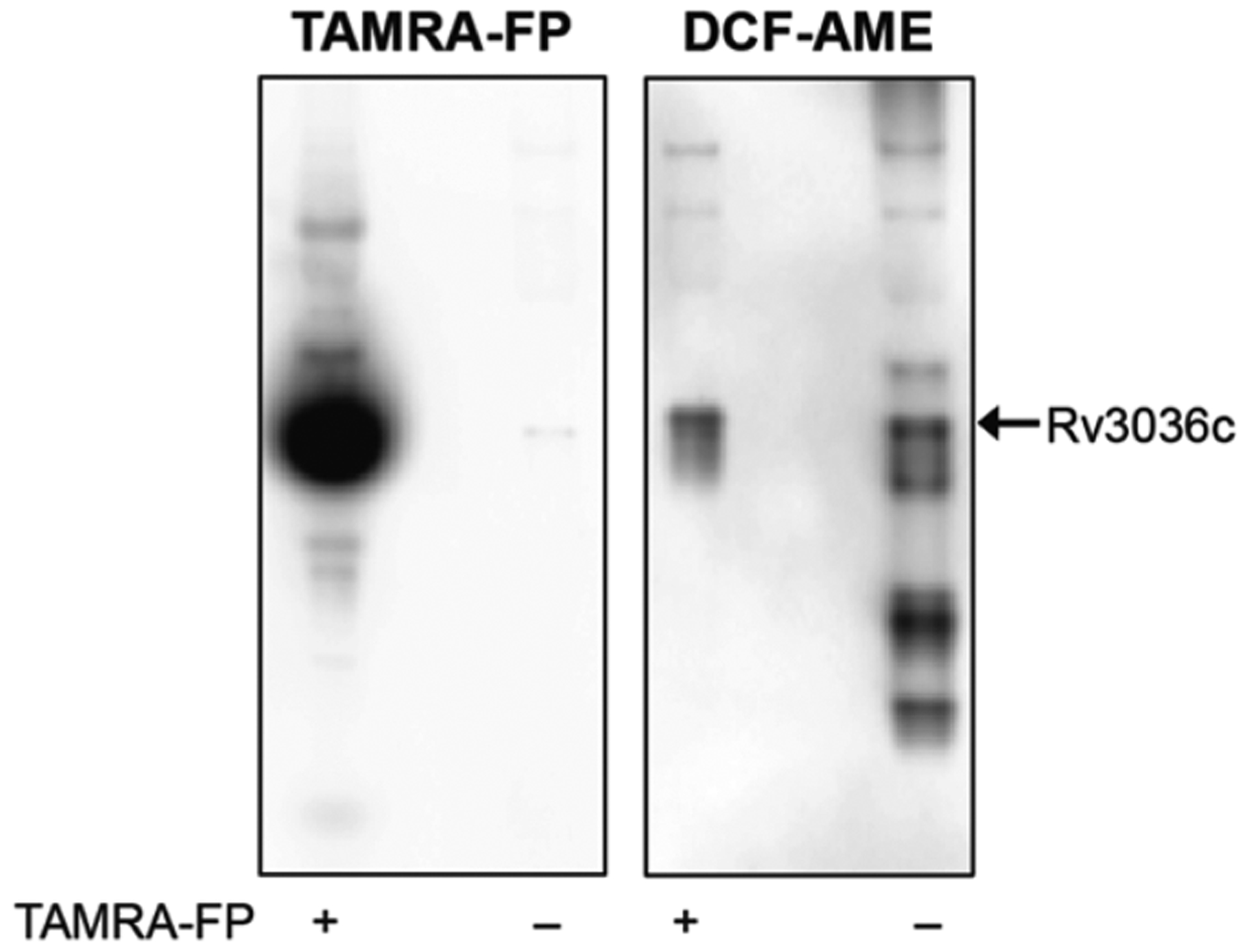
Fluorophosphonate ABPs and fluorogenic probes reveal different active esterases in native gel-resolved lysates. After TAMRA imaging (left image), the gel was incubated with 5 μM DCF-AME and imaged again (right image). A fluorescent band containing Rv3036c, a secreted esterase with a noncanonical active-site motif, was not labeled by TAMRA-FP but hydrolyzed DCF-AME.
Figure 7.
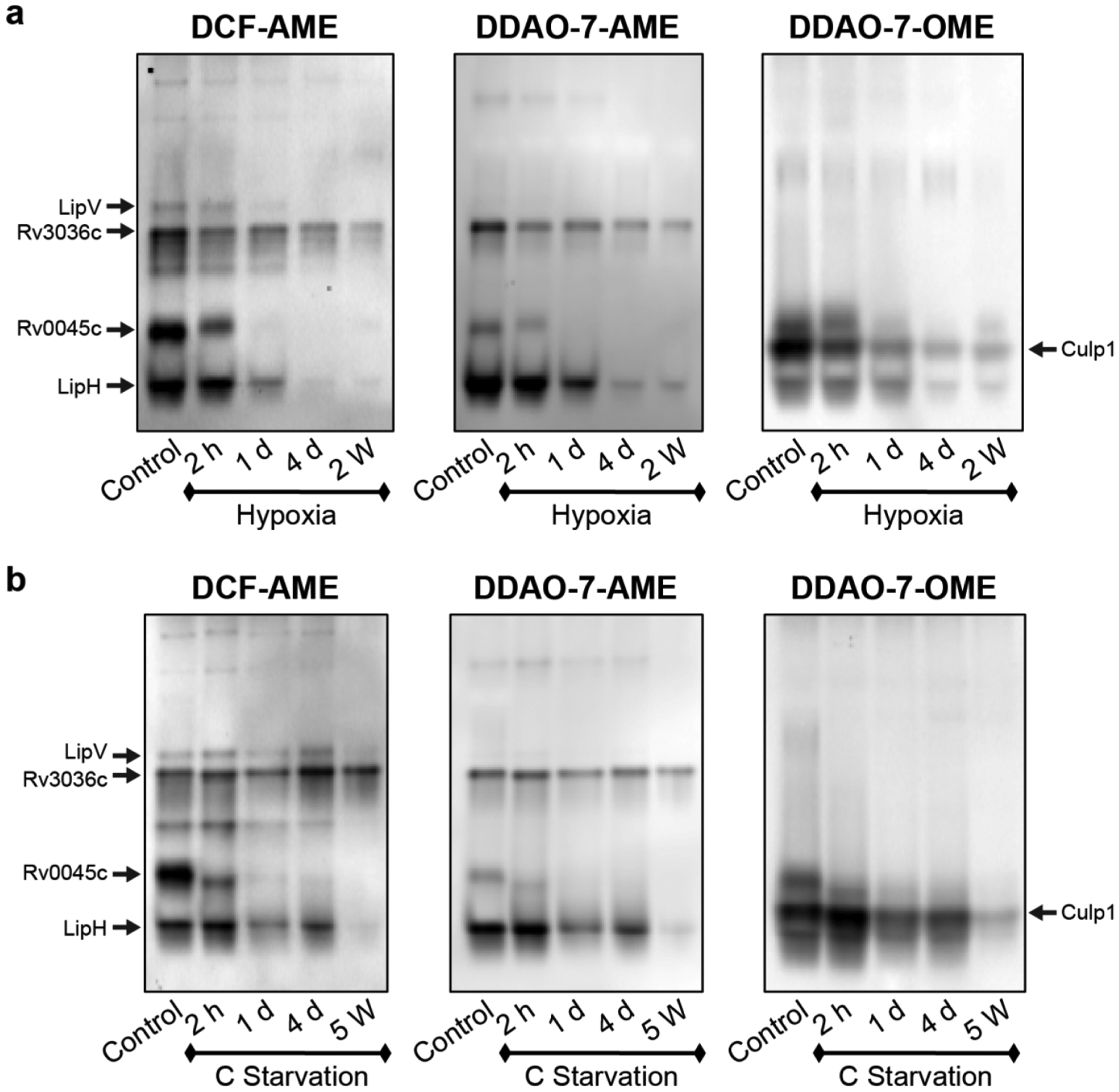
Mtb esterase activity is down-regulated in a time-dependent manner under hypoxia (a) or carbon starvation (b). Lysates were resolved by native PAGE, and the gels were incubated with the indicated probe before fluorescence imaging. Active esterase bands were excised and identified using mass spectrometry-based proteomics, as indicated.
Fluorogenic Probes Reveal Time-Dependent Down-regulation of Esterase Activity.
Our ABPP experiments required large amounts of protein (~1 mg) for target identification, whereas profiling with fluorogenic probes in a native PAGE assay used substantially less (1–10 μg). The smaller sample amount enabled us to evaluate time-dependent changes in Mtb esterase activity during dormancy induced by hypoxia or carbon starvation (Figure 7). We prepared lysates from cultures harvested after 2 h, 1 day, 4 days, or 2 weeks in hypoxia. We selected these time points because prior studies found that Mtb undergoes an initial hypoxic response followed by an enduring hypoxic response, which is a substantial, stable response that occurs by day 4.54,55 For comparison, we harvested similar time points for Mtb subjected to carbon starvation.
In both models, the decrease in esterase activity was time-dependent. In the hypoxia model, esterase activity decreased by 2 h, with substantial down-regulation by 1 day (Figure 7a). We observed further loss of activity by 4 days that remained absent through 2 weeks. Under carbon starvation, esterase activity was reduced by 2 h, with further down-regulation by 1 day (Figure 7b). The 5-week carbon-starved culture showed minimal esterase activity.
We observed persistent activity in dormancy from an excised band containing LipH peptides and from Culp1. These two esterases were identified only in replicating cultures by ABPP (Table 1), highlighting an advantage of using amplifiable fluorogenic substrates to detect enzymes.
Other esterase activities correlated well between our ABPP and fluorogenic probe-based studies. Our fluorogenic probes confirmed that LipV and Rv0045c were undetectable at 2 weeks in hypoxia-induced dormant Mtb.
In conclusion, we used serine hydrolase ABPs and fluorogenic esterase probes to identify catalytically active esterases in replicating, dormant, and reactivating Mtb. These chemical tools revealed that many esterase activities were down-regulated in dormancy. However, ABPs and fluorogenic probes together enabled us to identify eight esterases that retained activity in dormancy: CaeA, Culp1, LipH, LipM, LipN, Rv1683, Rv0183, and Rv3036c. We suggest that these esterases are worth investigating further as either diagnostic or therapeutic targets. Functional CaeA, Rv1683, Rv0183, and LipR were also present in recently reactivated Mtb. We directly compared ABPP and fluorogenic probe-based profiling for evaluating Mtb esterase activity across metabolic states. Overall, ABPP worked well for enrichment and identification of serine hydrolases, but fluorogenic probe-based profiling was more sensitive for detecting esterases with persistent activities and required 50–100-fold less protein.
METHODS
Mycobacterial Culture Conditions.
Cells were thawed from frozen stocks (stored at −80 °C in 6% glycerol). Mtb mc26020 (ΔlysA and ΔpanCD mutant)56 was cultured in 7H9 medium (7H9 broth, 0.5% glycerol, 0.05% Tween 80, 10% OADC) supplemented with 80 μg/mL lysine, 24 μg/mL pantothenate, and 0.2% casamino acids. Cultures were grown at 37 °C with 75 rpm shaking and handled as BSL-2 pathogens. Cells were collected by centrifugation, and the pellets were stored at −30 °C.
Culturing Mtb in Hypoxic Conditions.
Cultures were grown to an OD600 between 0.5 and 1.0 and then diluted to an OD600 of 0.4 in 7H9 complete medium (see above). Cultures (200 mL) were grown at 37 °C in standing flasks with (0.2 μm filtered) vented caps in a Heracell 150i incubator (Thermo Scientific) at 1% O2 and 5% CO2. Cultures were harvested by centrifugation (5 min, 2800g, 4 °C) after 2 h, 24 h, 4 days, or 2 weeks. Methylene blue (1.5 μg/mL) decolorization in control cultures indicated oxygen depletion between 9 and 14 days, as previously reported.9,57
Culturing Mtb Using a Carbon Starvation Model.
A modified version of the carbon starvation protocol developed by Grant et al. was used to induce dormancy.36 Cultures were grown in 7H9 complete medium until an OD600 between 0.6 and 1.5 was reached. Cells were harvested by centrifugation (5 min, 2800g, 4 °C) and washed twice with PBS containing 0.05% tyloxapol. Cells were resuspended at an OD600 of 0.2 in carbon starvation medium (7H9 broth base with 0.05% tyloxapol) and grown as standing cultures in sealed 1 L bottles (200 mL of culture per bottle) at 37 °C. Cultures were harvested by centrifugation (5 min, 2800g, 4 °C) after 2 h, 24 h, 4 days, or 5 weeks. Grant et al. reported that cultures become dormant by 2 weeks but that the respiration rate of Mtb under starvation conditions continues to decline until a plateau is reached by 5 weeks.36,58
Preparation of Reaeration Samples.
Cultures were grown under hypoxia as described above. After 2 weeks, cells were harvested by centrifugation (5 min, 2800g, 4 °C) and washed twice with PBS containing 0.05% tyloxapol. The pellets were resuspended in 50 mL of carbon starvation medium supplemented with 80 μg/mL lysine and 24 μg/mL pantothenate or in 7H9 complete medium and then grown at 37 °C in air.
Preparation of Lysates for In-Gel Analysis with Fluorogenic Probes.
Cell pellets were thawed and resuspended in lysis buffer [50 mM Tris (pH 7.5 at 4 °C), 200 mM NaCl, 0.5 mM CaCl2, 0.5 mM MgCl2, and 0.2% Triton X-100]. Cells were lysed by mechanical disruption on a BioSpec Beadbeater (3 × 1 min pulses with rests on ice) using 0.1 mm zirconia/silica beads (BioSpec Products) in screw-cap tubes. Insoluble debris and beads were pelleted by centrifugation (13000g, 4 °C, 10 min), and the clarified supernatant was transferred to a clean tube. Samples were further clarified by centrifugation (15700g, 4 °C, 5 min) to remove residual insoluble material. Lysates were sterilized by filtration through a 0.2 μm PVDF membrane (Pall Acrodisc 13 mm syringe filter).
In-Gel Detection of Active Mtb Serine Hydrolases Using an Activity-Based Probe.
For studies comparing dormancy, reactivation, and actively growing samples, lysates (15 μg total protein) were incubated with 10 μM ActivX TAMRA-FP (Fisher, PI88318) for 1 h at room temperature. Samples were boiled for 5 min in SDS-PAGE loading buffer, and insoluble material was removed by centrifugation. Proteins were resolved on a 10% Bis-Tris gel with XT-MES buffer at 180 V for 1 h at 4 °C, and TAMRA fluorescence was detected (λex 488 nm, 580 BP 30 nm) using a Typhoon 9400 Imager (GE Healthcare).
For inhibitor studies, lysates (2.5 μg total protein) were pre-incubated with 100 μM of inhibitor (MAFP, PMSF, E-600, THL, or ebselen) or DMSO (vehicle control) for 45 min at room temperature prior to treatment with 0.25 μM TAMRA-FP for 3 h. The samples were boiled with SDS-PAGE loading buffer and separated on a 12% Bis-Tris gel with MOPS buffer at 180 V for 75 min at 4 °C. TAMRA fluorescence was detected (λex 488 nm, 580 BP 30 nm) using a Typhoon 9400 Imager (GE Healthcare).
Identification of Active Mtb Serine Hydrolases Using Desthiobiotin-FP.
Mtb mc26020 lysates (n = 3 each condition) were concentrated to a volume of 45 μL using a 10 kDa MWCO centrifugal filter (Amicon). For normoxic and reactivation lysates, 750 μg of total protein was used. For hypoxic lysates, 500 μg was used due to the low protein content of those samples. The lysates were labeled with 100 μM ActivX desthiobiotin-FP (Fisher, PI88317) for 1 h at room temperature. Samples were enriched for biotinylated proteins using High Capacity Streptavidin Agarose Resin (Thermo Scientific), and labeled proteins were digested with trypsin, as described in the Supporting Information. Peptides were submitted to the UC—Davis Proteomics Core and analyzed by LC-MS/MS.
Western Blot Analysis.
Denatured protein samples were resolved on a 10% Bis-Tris polyacrylamide gel and transferred to a PVDF membrane. Biotinylated proteins were detecting by probing with anti-biotin-HRP (Jackson ImmunoResearch; 1:25,000 dilution). Chemiluminescence was detected on a MyECL imager (Thermo Scientific).
In-Gel Detection of Active Mtb Esterases Using Fluorogenic Probes.
For inhibitor competition experiments, lysates (2 μg of total protein) were pre-incubated with 100 μM inhibitor (MAFP, PMSF, E-600, or THL) for 1 h at room temperature. For all other experiments, lysates contained 8 μg of total protein. The lysates were resolved by native PAGE as previously described.33 Gels were equilibrated in 10 mM HEPES (pH 7.3), incubated with 5 μM probe for 5–10 min, and then imaged on a Typhoon 9400 Imager (DCF detection, λex 488 nm, 520 BP 40 nm; DDAO detection, λex 633 nm, 670 BP 30 nm).
Select bands were excised and submitted to the OHSU Proteomics Shared Resource for protein tryptic digestion, peptide extraction, and LC-MS/MS analysis, as described in the Supporting Information. See Table S5 for peptide identifications.
Supplementary Material
ACKNOWLEDGMENTS
Funding for this research was generously provided by the Knight Cancer Institute at OHSU, the Collins Medical Trust, and the Medical Research Foundation of Oregon. S.R.L. was supported by an NIH T32 training grant (T32-AI07472). We are grateful to I. Carter-O’Connell (OHSU) and B. Phinney (UC—Davis Proteomics Core) for helpful discussions. G. Simon of Abide Therapeutics generously shared their list of serine hydrolases identified in Mtb by PFAM notation.
Footnotes
Supporting Information
The Supporting Information is available free of charge on the ACS Publications website at DOI: 10.1021/acsinfecdis.6b00135.
Synthesis of DCF-AME, expression and purification of LipY, detailed methods for the streptavidin enrichment of desthiobiotin-FP-labeled proteins and their proteomic identification, and supplementary figures and tables (PDF)
Known and predicted serine hydrolases used in this study; detected proteins and their unique spectral counts for each sample; functional annotation of all identified serine hydrolases (XLSX)
Raw data files, results, peak lists, and search engine files for ABPP LC-MS/MS experiments have been deposited with the ProteomeXchange via MassIVE. The PX identifier is PXD005127. The MassIVE accession number is MSV000080238.
The authors declare no competing financial interest.
REFERENCES
- (1).(2015) Global Tuberculosis Report 2015, World Health Organization, Geneva, Switzerland. [Google Scholar]
- (2).Barry CE 3rd, Boshoff HI, Dartois V, Dick T, Ehrt S, Flynn J, Schnappinger D, Wilkinson RJ, and Young D (2009) The spectrum of latent tuberculosis: rethinking the biology and intervention strategies. Nat. Rev. Microbiol 7, 845–855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (3).Esmail H, Barry CE 3rd, and Wilkinson RJ (2012) Understanding latent tuberculosis: the key to improved diagnostic and novel treatment strategies. Drug Discovery Today 17, 514–521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (4).Tufariello JM, Chan J, and Flynn JL (2003) Latent tuberculosis: mechanisms of host and bacillus that contribute to persistent infection. Lancet Infect. Dis 3, 578–590. [DOI] [PubMed] [Google Scholar]
- (5).Nathan C, and Barry CE 3rd. (2015) TB drug development: immunology at the table. Immunol. Rev 264, 308–318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (6).Shi L, Sohaskey CD, Kana BD, Dawes S, North RJ, Mizrahi V, and Gennaro ML (2005) Changes in energy metabolism of Mycobacterium tuberculosis in mouse lung and under in vitro conditions affecting aerobic respiration. Proc. Natl. Acad. Sci. U. S. A 102, 15629–15634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (7).Deb C, Lee CM, Dubey VS, Daniel J, Abomoelak B, Sirakova TD, Pawar S, Rogers L, and Kolattukudy PE (2009) A novel in vitro multiple-stress dormancy model for Mycobacterium tuberculosis generates a lipid-loaded, drug-tolerant, dormant pathogen. PLoS One 4, e6077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (8).Galagan JE, Minch K, Peterson M, Lyubetskaya A, Azizi E, Sweet L, Gomes A, Rustad T, Dolganov G, Glotova I, Abeel T, Mahwinney C, Kennedy AD, Allard R, Brabant W, Krueger A, Jaini S, Honda B, Yu WH, Hickey MJ, Zucker J, Garay C, Weiner B, Sisk P, Stolte C, Winkler JK, Van de Peer Y, Iazzetti P, Camacho D, Dreyfuss J, Liu Y, Dorhoi A, Mollenkopf HJ, Drogaris P, Lamontagne J, Zhou Y, Piquenot J, Park ST, Raman S, Kaufmann SH, Mohney RP, Chelsky D, Moody DB, Sherman DR, and Schoolnik GK (2013) The Mycobacterium tuberculosis regulatory network and hypoxia. Nature 499, 178–183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (9).Betts JC, Lukey PT, Robb LC, McAdam RA, and Duncan K (2002) Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol. Microbiol 43, 717–731. [DOI] [PubMed] [Google Scholar]
- (10).Schubert OT, Ludwig C, Kogadeeva M, Zimmermann M, Rosenberger G, Gengenbacher M, Gillet LC, Collins BC, Rost HL, Kaufmann SH, Sauer U, and Aebersold R (2015) Absolute proteome composition and dynamics during dormancy and resuscitation of Mycobacterium tuberculosis. Cell Host Microbe 18, 96–108. [DOI] [PubMed] [Google Scholar]
- (11).Gopinath V, Raghunandanan S, Gomez RL, Jose L, Surendran A, Ramachandran R, Pushparajan AR, Mundayoor S, Jaleel A, and Kumar RA (2015) Profiling the proteome of Mycobacterium tuberculosis during dormancy and reactivation. Mol. Cell. Proteomics 14, 2160–2176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (12).Daniel J, Deb C, Dubey VS, Sirakova TD, Abomoelak B, Morbidoni HR, and Kolattukudy PE (2004) Induction of a novel class of diacylglycerol acyltransferases and triacylglycerol accumulation in Mycobacterium tuberculosis as it goes into a dormancy-like state in culture. J. Bacteriol 186, 5017–5030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (13).Deb C, Daniel J, Sirakova TD, Abomoelak B, Dubey VS, and Kolattukudy PE (2006) A novel lipase belonging to the hormone-sensitive lipase family induced under starvation to utilize stored triacylglycerol in Mycobacterium tuberculosis. J. Biol. Chem 281, 3866–3875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (14).Low KL, Rao PS, Shui G, Bendt AK, Pethe K, Dick T, and Wenk MR (2009) Triacylglycerol utilization is required for regrowth of in vitro hypoxic nonreplicating Mycobacterium bovis bacillus Calmette-Guerin. J. Bacteriol 191, 5037–5043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (15).Côtes K, Bakala N’Goma JC, Dhouib R, Douchet I, Maurin D, Carrière F, and Canaan S (2008) Lipolytic enzymes in Mycobacterium tuberculosis. Appl. Microbiol. Biotechnol 78, 741–749. [DOI] [PubMed] [Google Scholar]
- (16).Saravanan P, Dubey VK, and Patra S (2012) Potential selective inhibitors against Rv0183 of Mycobacterium tuberculosis targeting host lipid metabolism. Chem. Biol. Drug Des 79, 1056–1062. [DOI] [PubMed] [Google Scholar]
- (17).Point V, Malla RK, Diomande S, Martin BP, Delorme V, Carriere F, Canaan S, Rath NP, Spilling CD, and Cavalier JF (2012) Synthesis and kinetic evaluation of cyclophostin and cyclipostins phosphonate analogs as selective and potent inhibitors of microbial lipases. J. Med. Chem 55, 10204–10219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (18).Point V, Malla RK, Carriere F, Canaan S, Spilling CD, and Cavalier JF (2013) Enantioselective inhibition of microbial lipolytic enzymes by nonracemic monocyclic enolphosphonate analogues of cyclophostin. J. Med. Chem 56, 4393–4401. [DOI] [PubMed] [Google Scholar]
- (19).Ortega C, Anderson LN, Frando A, Sadler NC, Brown RW, Smith RD, Wright AT, and Grundner C (2016) Systematic survey of serine hydrolase activity in Mycobacterium tuberculosis defines changes associated with persistence. Cell Chem. Biol 23, 290–298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (20).Bachovchin DA, and Cravatt BF (2012) The pharmacological landscape and therapeutic potential of serine hydrolases. Nat. Rev. Drug Discovery 11, 52–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (21).Xie L, Wang X, Zeng J, Zhou M, Duan X, Li Q, Zhang Z, Luo H, Pang L, Li W, Liao G, Yu X, Li Y, Huang H, and Xie J (2015) Proteome-wide lysine acetylation profiling of the human pathogen Mycobacterium tuberculosis. Int. J. Biochem. Cell Biol 59, 193–202. [DOI] [PubMed] [Google Scholar]
- (22).Prisic S, Dankwa S, Schwartz D, Chou MF, Locasale JW, Kang C-M, Bemis G, Church GM, Steen H, and Husson RN (2010) Extensive phosphorylation with overlapping specificity by Mycobacterium tuberculosis serine/threonine protein kinases. Proc. Natl. Acad. Sci. U. S. A 107, 7521–7526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (23).Cravatt BF, Wright AT, and Kozarich JW (2008) Activity-based protein profiling: from enzyme chemistry to proteomic chemistry. Annu. Rev. Biochem 77, 383–414. [DOI] [PubMed] [Google Scholar]
- (24).Baruch A, Jeffery DA, and Bogyo M (2004) Enzyme activity—it’s all about image. Trends Cell Biol. 14, 29–35. [DOI] [PubMed] [Google Scholar]
- (25).Ravindran MS, Rao SPS, Cheng X, Shukla A, Cazenave Gassiot A, Yao SQ, and Wenk MR (2014) Targeting lipid esterases in mycobacteria grown under different physiological conditions using activity-based profiling with tetrahydrolipstatin (THL). Mol. Cell. Proteomics 13, 435–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (26).Parker SK, Barkley RM, Rino JG, and Vasil ML (2009) Mycobacterium tuberculosis Rv3802c encodes a phospholipase/thioesterase and is inhibited by the antimycobacterial agent tetrahydrolipstatin. PLoS One 4, e4281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (27).Delorme V, Diomande SV, Dedieu L, Cavalier JF, Carriere F, Kremer L, Leclaire J, Fotiadu F, and Canaan S (2012) MmPPOX inhibits Mycobacterium tuberculosis lipolytic enzymes belonging to the hormone-sensitive lipase family and alters mycobacterial growth. PLoS One 7, e46493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (28).Simon G (2015) Re: Abide Contact Submission (Levine SR, Ed.) [Google Scholar]
- (29).Kidd D, Liu Y, and Cravatt BF (2001) Profiling serine hydrolase activities in complex proteomes. Biochemistry 40, 4005–4015. [DOI] [PubMed] [Google Scholar]
- (30).Beatty KE, Williams M, Carlson BL, Swarts BM, Warren RM, van Helden PD, and Bertozzi CR (2013) Sulfatase-activated fluorophores for rapid discrimination of mycobacterial species and strains. Proc. Natl. Acad. Sci. U. S. A 110, 12911–12916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (31).Smith EL, Bertozzi CR, and Beatty KE (2014) An expanded set of fluorogenic sulfatase activity probes. ChemBioChem 15, 1101–1105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (32).Tallman KR, and Beatty KE (2015) Far-red fluorogenic probes for esterase and lipase detection. ChemBioChem 16, 70–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (33).Tallman KR, Levine SR, and Beatty KE (2016) Profiling esterases in Mycobacterium tuberculosis using far-red fluorogenic substrates. ACS Chem. Biol 11, 1810–1815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (34).Favrot L, Grzegorzewicz AE, Lajiness DH, Marvin RK, Boucau J, Isailovic D, Jackson M, and Ronning DR (2013) Mechanism of inhibition of Mycobacterium tuberculosis antigen 85 by ebselen. Nat. Commun 4, 2748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (35).Favrot L, Lajiness DH, and Ronning DR (2014) Inactivation of the Mycobacterium tuberculosis antigen 85 complex by covalent, allosteric inhibitors. J. Biol. Chem 289, 25031–25040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (36).Grant SS, Kawate T, Nag PP, Silvis MR, Gordon K, Stanley SA, Kazyanskaya E, Nietupski R, Golas A, Fitzgerald M, Cho S, Franzblau SG, and Hung DT (2013) Identification of novel inhibitors of nonreplicating Mycobacterium tuberculosis using a carbon starvation model. ACS Chem. Biol 8, 2224–2234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (37).Wayne LG (2001) In vitro model of hypoxically induced nonreplicating persistence of Mycobacterium tuberculosis. Methods Mol. Med 54, 247–269. [DOI] [PubMed] [Google Scholar]
- (38).Duckworth BP, Geders TW, Tiwari D, Boshoff HI, Sibbald PA, Barry CE 3rd, Schnappinger D, Finzel BC, and Aldrich CC (2011) Bisubstrate adenylation inhibitors of biotin protein ligase from Mycobacterium tuberculosis. Chem. Biol 18, 1432–1441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (39).Ravindran MS, Rao SP, Cheng X, Shukla A, Cazenave Gassiot A, Yao SQ, and Wenk MR (2014) Targeting lipid esterases in mycobacteria grown under different physiological conditions using activity-based profiling with tetrahydrolipstatin (THL). Mol. Cell. Proteomics 13, 435–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (40).Vir P, Gupta D, Agarwal R, and Verma I (2014) Interaction of alveolar epithelial cells with CFP21, a mycobacterial cutinase-like enzyme. Mol. Cell. Biochem 396, 187–199. [DOI] [PubMed] [Google Scholar]
- (41).Schué M, Maurin D, Dhouib R, Bakala N’Goma JC, Delorme V, Lambeau G, Carriere F, and Canaan S (2010) Two cutinase-like proteins secreted by Mycobacterium tuberculosis show very different lipolytic activities reflecting their physiological function. FASEB J. 24, 1893–1903. [DOI] [PubMed] [Google Scholar]
- (42).Grover A, Ahmed MF, Verma I, Sharma P, and Khuller GK (2006) Expression and purification of the Mycobacterium tuberculosis complex-restricted antigen CFP21 to study its immunoprophylactic potential in mouse model. Protein Expression Purif. 48, 274–280. [DOI] [PubMed] [Google Scholar]
- (43).Kalra M, Grover A, Mehta N, Singh J, Kaur J, Sable SB, Behera D, Sharma P, Verma I, and Khuller GK (2007) Supplementation with RD antigens enhances the protective efficacy of BCG in tuberculous mice. Clin. Immunol 125, 173–183. [DOI] [PubMed] [Google Scholar]
- (44).Brust B, Lecoufle M, Tuaillon E, Dedieu L, Canaan S, Valverde V, and Kremer L (2011) Mycobacterium tuberculosis lipolytic enzymes as potential biomarkers for the diagnosis of active tuberculosis. PLoS One 6, e25078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (45).Lun S, and Bishai WR (2007) Characterization of a novel cell wall-anchored protein with carboxylesterase activity required for virulence in Mycobacterium tuberculosis. J. Biol. Chem 282, 18348–18356. [DOI] [PubMed] [Google Scholar]
- (46).Rengarajan J, Murphy E, Park A, Krone CL, Hett EC, Bloom BR, Glimcher LH, and Rubin EJ (2008) Mycobacterium tuberculosis Rv2224c modulates innate immune responses. Proc. Natl. Acad. Sci. U. S. A 105, 264–269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (47).Cotes K, Dhouib R, Douchet I, Chahinian H, de Caro A, Carriere F, and Canaan S (2007) Characterization of an exported monoglyceride lipase from Mycobacterium tuberculosis possibly involved in the metabolism of host cell membrane lipids. Biochem. J 408, 417–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (48).Dhouib R, Laval F, Carriere F, Daffe M, and Canaan S (2010) A monoacylglycerol lipase from Mycobacterium smegmatis involved in bacterial cell interaction. J. Bacteriol 192, 4776–4785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (49).Sassetti CM, Boyd DH, and Rubin EJ (2003) Genes required for mycobacterial growth defined by high density mutagenesis. Mol. Microbiol 48, 77–84. [DOI] [PubMed] [Google Scholar]
- (50).Malen H, Pathak S, Softeland T, de Souza GA, and Wiker HG (2010) Definition of novel cell envelope associated proteins in Triton X-114 extracts of Mycobacterium tuberculosis H37Rv. BMC Microbiol. 10, 132-2180-2110-2132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (51).Kruh NA, Troudt J, Izzo A, Prenni J, and Dobos KM (2010) Portrait of a pathogen: the Mycobacterium tuberculosis proteome in vivo. PLoS One 5, e13938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (52).Daleke MH, Cascioferro A, de Punder K, Ummels R, Abdallah AM, van der Wel N, Peters PJ, Luirink J, Manganelli R, and Bitter W (2011) Conserved Pro-Glu (PE) and Pro-Pro-Glu (PPE) protein domains target LipY lipases of pathogenic mycobacteria to the cell surface via the ESX-5 pathway. J. Biol. Chem 286, 19024–19034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (53).Chen L, Dang G, Deng X, Cao J, Yu S, Wu D, Pang H, and Liu S (2014) Characterization of a novel exported esterase Rv3036c from Mycobacterium tuberculosis. Protein Expression Purif. 104C, 50–56. [DOI] [PubMed] [Google Scholar]
- (54).Park HD, Guinn KM, Harrell MI, Liao R, Voskuil MI, Tompa M, Schoolnik GK, and Sherman DR (2003) Rv3133c/dosR is a transcription factor that mediates the hypoxic response of Mycobacterium tuberculosis. Mol. Microbiol 48, 833–843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (55).Rustad TR, Harrell MI, Liao R, and Sherman DR (2008) The enduring hypoxic response of Mycobacterium tuberculosis. PLoS One 3, e1502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (56).Sambandamurthy VK, Derrick SC, Jalapathy KV, Chen B, Russell RG, Morris SL, and Jacobs WR Jr. (2005) Long-term protection against tuberculosis following vaccination with a severely attenuated double lysine and pantothenate auxotroph of Mycobacterium tuberculosis. Infect. Immun 73, 1196–1203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (57).Bryk R, Gold B, Venugopal A, Singh J, Samy R, Pupek K, Cao H, Popescu C, Gurney M, Hotha S, Cherian J, Rhee K, Ly L, Converse PJ, Ehrt S, Vandal O, Jiang X, Schneider J, Lin G, and Nathan C (2008) Selective killing of nonreplicating mycobacteria. Cell Host Microbe 3, 137–145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- (58).Loebel RO, Shorr E, and Richardson HB (1933) The influence of foodstuffs upon the respiratory metabolism and growth of human tubercle bacilli. J. Bacteriol 26, 139–166. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.