

Abstract
Treatment of osteoarthritis (OA) is mainly symptomatic by alleviating pain to postpone total joint replacement. Bone morphogenetic protein 7 (BMP7) is a candidate morphogen for experimental OA treatment that favorably alters the chondrocyte and cartilage phenotype. Intra-articular delivery and sustained release of a recombinant growth factor for treating OA are challenging, whereas the use of peptide technology potentially circumvents many of these challenges. In this study, we screened a high-resolution BMP7 peptide library and discovered several overlapping peptide sequences from two regions in BMP7 with nanomolar bioactivity that attenuated the pathological OA chondrocyte phenotype. A single exposure of OA chondrocytes to peptides p[63−82] and p[113−132] ameliorated the OA chondrocyte phenotype for up to 8 days, and peptides were bioactive on chondrocytes in OA synovial fluid. Peptides p[63−82] and p[113−132] required NKX3-2 for their bioactivity on chondrocytes and provoke changes in SMAD signaling activity. The bioactivity of p[63−82] depended on specific evolutionary conserved sequence elements common to BMP family members. Intra-articular injection of a rat medial meniscal tear (MMT) model with peptide p[63−82] attenuated cartilage degeneration. Together, this study identified two regions in BMP7 from which bioactive peptides are able to attenuate the OA chondrocyte phenotype. These BMP7-derived peptides provide potential novel disease-modifying treatment options for OA.
Keywords: BMP7, peptides, osteoarthritis, chondrocyte phenotype, hypertrophy
Graphical abstract
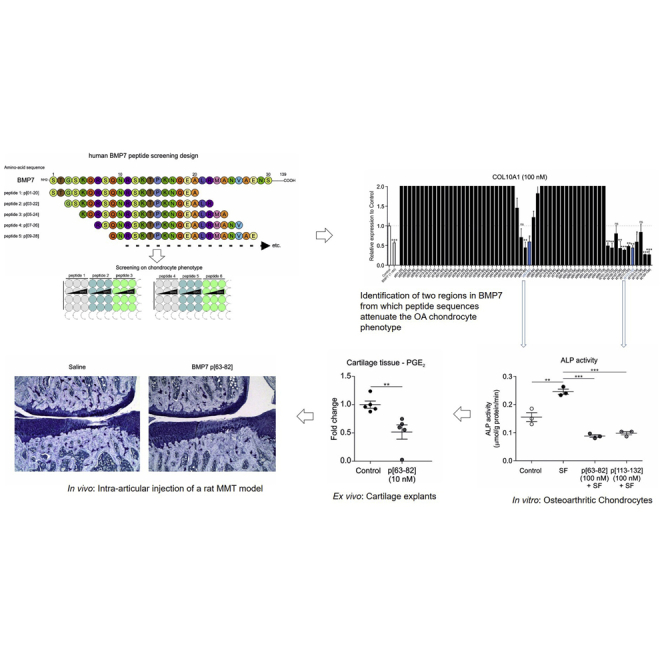
BMP7 is a candidate morphogen for experimental osteoarthritis treatment that favorably alters the chondrocyte and cartilage phenotype. This study discovered two regions in BMP7 from which bioactive peptides are able to attenuate the OA chondrocyte phenotype. These BMP7-derived peptides provide potential novel disease-modifying treatment options for OA.
Introduction
Osteoarthritis (OA) is the most common degenerative joint disorder worldwide and presents with degradation of articular cartilage, leading to loss of joint mobility and function, accompanied by chronic pain.1 Biochemically, OA is characterized by uncontrolled synthesis of extracellular matrix-degrading enzymes, such as aggrecanases (a disintegrin and metalloprotease with thrombospondin motifs [ADAMTSs]) and matrix metalloproteinases (MMPs), resulting in the active breakdown of the cartilage tissue matrix.2 Important risk factors for developing OA are joint overloading (misalignment, obesity, work related); diabetes; aging; articular cartilage damage due to, e.g., trauma;3 and others. The analogy between endochondral ossification and OA progression has been widely recognized, and many of the cartilage-degrading enzymes that are secreted by hypertrophic chondrocytes in the growth plate are also involved in OA development.4, 5, 6 This places, next to the local joint inflammatory condition,7 the chondrocyte/cartilage differentiation status central in the progression or even cause of OA.8 Current treatments for OA are mainly symptomatic by alleviating pain and interfering with the cartilage-degenerative processes to postpone total joint replacement.
A number of growth factor-based treatment options for OA are currently under investigation.9 One growth factor that modifies the chondrocyte and cartilage differentiation status is bone morphogenetic protein 7 (BMP7; also called osteogenic protein 1 [OP-1]). Studies addressing the disease-modifying properties of BMP7 showed that it decreases MMP13 expression in interleukin (IL)-1β-exposed chondrocytes,10 stimulates proteoglycan synthesis in OA chondrocytes,11 counteracts inflammatory cytokines (e.g., IL-1β), and induces an overall anabolic response in healthy chondrocytes.12,13 Intra-articular (IA) administration of BMP7 protects against OA development in rabbits14 and delays the progression of OA in rats.15 A phase 1 clinical trial has been completed for BMP7 in end-stage OA patients and reported no serious adverse events after the intra-articular injection of BMP7.16 In concert with these reports, our previous work17 unveiled that BMP7 suppresses the pathological chondrocyte phenotype associated with OA8 via NK3 Homeobox 2 (NKX3-2) and by inhibiting chondrocyte catabolism and hypertrophy.
Challenges that come with intra-articular delivery of a recombinant growth factor for treating OA are its formulation for sustained release from a drug carrier, stability in the hydrolytic and proteolytic OA synovial fluid (SF) environment,18, 19, 20 and high production costs of the recombinant growth factor. Mimicking the growth factor-initiated OA chondrocyte biological responses with the use of peptides is a promising area that was recently applied to growth hormone/somatostatin, preventing cartilage degradation in a rat model for OA.21 The biomolecular synthesis of short linear peptides is relatively straightforward, and in general, peptides are less susceptible to conformational inactivation as compared to recombinant growth factors. Consequently, the use of peptide technology potentially circumvents many of the challenges associated with full-length recombinant growth factors.22 Peptides from BMP7 have previously been reported to support osteogenesis23, 24, 25 and prevent fibrosis in acute and chronic kidney injury.26 Whether peptides derived from BMP7 are able to suppress the pathological OA chondrocyte phenotype27 was unknown. Although bioactive peptides from BMP7 have been reported, the action of BMPs and BMP-derived peptides is highly cell-type dependent.28 Therefore, a screening was performed of an overlapping sequential BMP7 peptide library to discover peptide sequences with a bioactivity that specifically attenuates the pathological OA chondrocyte phenotype. We postulate that the potential identification of a peptide from BMP7 will hold the future promise to be more compatible, and biochemically modifiable, for incorporation into macromolecular sustained release systems that may potentially be used in the intra-articular treatment of OA.
Results
Peptide sequences derived from BMP7 that attenuate the OA chondrocyte phenotype
To investigate whether BMP7 harbors potential peptide sequences that improve the OA chondrocyte phenotype, a peptide library was designed from the mature 139 amino acid-long human BMP7 sequence (Table S2). The peptide library was designed as 20-mer peptides with 2 amino acid intervals (18 amino acid overlap) between individual peptides and with all cysteine residues substituted by serine residues to avoid uncontrolled oxidation of cysteine groups. This yielded 61 individual peptides covering the complete mature human BMP7 sequence (Figure 1A). The peptide library was screened using a pool of primary human OA articular chondrocytes (OA-HACs) from 18 individual donors. Prior to pooling, individual OA-HAC isolates were tested for their responsiveness to BMP7 (Figure S1). The screening was conducted by exposing the OA-HAC pool for 24 h to a concentration series (1, 10, 100, or 1,000 nM) of each peptide from the library. The chondrocyte response to each condition was established by measuring a set of OA chondrocyte phenotype genes in which their expression was previously shown to significantly improve by BMP7 treatment.27 This is defined as reduced expression of collagen type X alpha 1 chain (COL10A1), alkaline phosphatase (ALPL), Runt-related transcription factor 2 (RUNX2), ADAMTS5, MMP13, prostaglandin-endoperoxide synthase 2 or cyclooxygenase-2 (COX-2), and interleukin 6 (IL6) and increased expression of SRY-box transcription factor 9 (SOX9), COL2A1, and NKX3-2. Library screening data of the 100-nM condition for COL10A1 expression are presented in Figure 1B (for other genes and peptide concentrations, see Figure S2) and show that the majority of the peptides worsened the chondrocyte phenotype and provoked chondrocyte hypertrophy (COL10A1 expression). However, peptides derived from the central region of BMP7, as well as from the C terminus, exhibited bioactivity that improved the expression of the predefined OA chondrocyte phenotype gene set. Representative peptides inducing the above gene expression characteristics in OA-HACs were selected from both regions for further investigation. These were peptide p[63−82] and p[113−132] from the library, covering amino acids 63−82 and 113−132 of the mature BMP7 amino acid sequence. The complete quantitative real-time PCR data for peptides p[63−82] and p[113−132] (Figure 2) show that these peptides induced expression of SOX9, COL2A1, and NKX3-2 (Figure 2A) and inhibited expression of RUNX2, COL10A1, ALPL, MMP13, ADAMTS5, COX-2, and IL6 (Figures 2B and 2C). It is noteworthy that these peptides exhibited bioactivity in the low nanomolar range. For the two candidate peptides, we validated data using an independent cohort of three OA-HACs exposed to 100 nM of p[63−82] or p[113−132] or a combination of both peptides. Gene expression analyses of COL10A1, ALPL, RUNX2, COL2A1, SOX9, NKX3-2, ADAMTS5, MMP13, COX-2, and IL6 expression showed similar responses as above (Figure S3). Peptide p[63−82] increased the sulfated glycosaminoglycan (sGAG) content of OA-HAC cultures (Figure 2D), and both peptides caused a significant decrease in alkaline phosphatase (ALP) activity (Figure 2E) and prostaglandin E2 (PGE2) levels (Figure 2F) in these cultures. To investigate sequence specificity of the bioactivity of peptides p[63−82] and p[113−132], scrambled version of these peptides were tested, and neither one of them was able to recapitulate the bioactivity that was observed for candidate peptides p[63−82] and p[113−132] on the expression of COL2A1, RUNX2, COL10A1, and COX-2 (Figure 2G). Taken together, two regions in BMP7 were identified delivering peptide sequences that are able to improve the OA chondrocyte phenotype.
Figure 1.

BMP7 peptide library screening on primary human OA articular chondrocytes (OA-HACs)
(A) Schematic representation of the design of the human BMP7 peptide library. (B) COL10A1 mRNA expression in samples from the library screening on a pool of 18 OA-HACs individually exposed for 24 h to the peptides (100 nM) in the library. Data are shown as compared to the control condition (white bar) and full-length BMP7 (1 nM; gray bar). The two blue bars represent candidate peptides p[63−82] and p[113−132]. Data from these two peptides showed significantly downregulated COL10A1 expression but also showed reduced expression of RUNX2, ALPL, COX-2, IL-6, ADAMTS5, and MMP13 and increased expression of SOX9, COL2A1, and NKX3-2 in all tested peptide concentrations. Error bars represent mean ± SEM, and statistical significance for peptide condition or BMP7 versus control condition as determined by unpaired two-tailed Student’s t test is represented as ∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001; NS, not significant. Data for other measured mRNAs and peptide concentrations are provided in Figure S2.
Figure 2.

Bioactivity of BMP7-derived peptides p[63−82] and p[113−132] on OA-HACs
Complete peptide library screening quantitative real-time PCR dataset for peptides p[63−82] and p[113−132] is presented. (A) SOX9, COL2A1, and NKX3-2 mRNA expression in the 18 OA-HAC pool exposed to 1,000, 100, 10, or 1 nM of peptide p[63−82] and p[113−132] is shown and presented relative to control (black bars) and alongside full-length BMP7 (white bars). (B) Similar to (A) but for chondrocyte hypertrophy-associated genes RUNX2, COL10A1, and ALPL. (C) Similar to (A) but for cartilage extracellular matrix remodeling-associated genes MMP13 and ADAMTS5 and inflammation-related genes COX-2 and IL-6. (D) Glycosaminoglycan (GAG) content (normalized to total protein content) on an independent cohort of three individual OA-HAC donors treated with 100 nM peptides p[63−82] and p[113−132] for 24 h. Corresponding gene expression data are shown in Figure S3. (E) Alkaline phosphatase (ALP) activity (normalized to total protein content) in similar samples from (D). (F) Prostaglandin E2 (PGE2) levels in culture supernatant in similar samples from (D). (G) In the same OA-HAC pool of 18 OA-HAC donors from the peptide library screening, sequence dependency of peptides p[63−82] and p[113−132] was determined by exposing the OA-HAC pool to scrambled versions of peptides p[63−82] and p[113−132] (100 nM, 24 h). Expression of COL2A1, RUNX2, COL10A1, and COX-2 mRNAs is shown. Error bars represent mean ± SEM, and statistical significance for peptide conditions or BMP7 versus control condition as determined by unpaired two-tailed Student’s t test is represented as ∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001.
BMP7-derived peptides retain bioactivity on HACs cultured in OA SF
To further corroborate the potential OA-protective properties of peptides p[63−82] and p[113−132], we mimicked the OA environment by exposing non-OA-HACs to OA SF. Culturing non-OA-HACs in the presence of OA SF reduced their SOX9, COL2A1, and NKX3-2 expression (Figure 3A), whereas expression of RUNX2, COL10A1, ALPL, MMP13, ADAMTS5, COX-2, and IL6 was induced by OA SF (Figures 3B and 3C). This was accompanied by increased ALP activity and PGE2 secretion (Figures 3E and 3F). The decline of SOX9 and COL2A1 expression was counteracted to some extent by peptides p[63−82] and p[113−132] (Figure 3A), and expression of NKX3-2 was induced by the peptides to levels higher than in the control condition (Figure 3A). The OA SF-induced expression of RUNX2, COL10A1, ALPL, MMP13, ADAMTS5, COX-2, and IL6 was, without exception, mitigated by peptides p[63−82] and p[113−132] (Figures 3B and 3C). sGAG content of HAC cultures was unaltered by the peptides (Figure 3D). Retainment of peptide p[63−82] and p[113−132] bioactivity on HACs in an OA SF environment was also confirmed for ALP activity (Figure 3E) and PGE2 secretion (Figure 3F). Data demonstrate that bioactivity of peptide p[63−82] and p[113−132] on HACs is to a certain extent compatible with OA SF.
Figure 3.

Bioactivity of peptides p[63−82] and p[113−132] on OA articular chondrocytes in OA synovial fluid (SF)
OA-HACs (n = 3 individual donors, tested in triplicate) were exposed to 100 nM peptide p[63−82] or p[113−132] in the presence of OA SF (20% [v/v]; equal ratio mix from five individual donors). Control is 20% (v/v) 0.9% NaCl. Cells were cultured in these conditions for 24 h and analyzed for mRNA expression of indicated genes (normalized for 28S rRNA expression and relative to control condition). (A) Expression of SOX9, COL2A1, and NKX3-2 mRNAs. (B) Expression of RUNX2, COL10A1, and ALPL mRNAs. (C) Expression of MMP13, ADAMTS5, COX-2, and IL-6 mRNAs. (D) GAG content (normalized to total protein content) was determined in the same conditions. (E) ALP enzyme activity in cell lysates of the same conditions was determined and normalized for total protein content. (F) PGE2 levels in culture supernatants of the same conditions. Error bars represent mean ± SEM, and statistical significance for peptide conditions or control versus SF condition as determined by unpaired two-tailed Student’s t test is represented as ∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001.
Duration of chondrocyte phenotype modulation by BMP7-derived peptides
The duration of chondrocyte phenotypic change induced by candidate peptides p[63−82] and p[113−132] was determined by exposing OA-HACs to a single dose of peptide or continuously keeping the peptides present during OA-HAC culture. In both cases, the phenotype of the OA-HACs was analyzed every other day during a 10-day follow-up. Non-OA-HACs were used as a reference for chondrocyte phenotype status. Continuous exposure of OA-HACs to peptide p[63−82] or p[113−132] by repeated addition of the peptide to the culture media every other day resulted in an overtime increasing mRNA expression of COL2A1 levels (Figure 4A). A reciprocal response was observed for COL10A1 gene expression, which steadily decreased over time (Figure 4B). Inhibition of MMP13 and COX-2 expression was continuously observed during the treatment period (Figures 4C and 4D). Follow-up of the single-dose treatment of OA-HACs with either peptide p[63−82] or p[113−132] revealed inhibition of COL10A1, MMP13, and COX-2 gene expression up to and including day 8, reaching similar levels of expression of these genes in non-OA-HACs. At day 10, expression of COL10A1, MMP13, and COX-2 in the single-dose condition was indistinguishable from non-treated OA HACs. This timing was similar for COL2A1, although the action of peptide p[63−82] was not detectable anymore from day 8 onward. Together, data show that a single exposure of OA-HACs to peptides p[63−82] or p[113−132] leads to detectable changes in the chondrocyte gene expression phenotype up to 8 days.
Figure 4.

Duration of chondrocyte phenotype modulation by BMP7-derived peptides
A pool of OA-HACs (n = 3 donors) was cultured in the presence or absence of 100 nM peptides p[63−82] or p[113−132] and compared to a pool of human articular chondrocytes from a non-OA source. The medium was changed every 2 days. Peptides were added at the start of the experiment and subsequently at every medium change (multiple) or only at the start of the experiment and not during every subsequent medium change (single). At days 0, 2, 4, 6, 8, and 10 in culture, samples were harvested and analyzed for the expression of the indicated mRNAs by quantitative real-time PCR (normalized for 28S rRNA expression) (A) COL2A1 mRNA expression. (B) COL10A1 mRNA expression. (C) MMP13 mRNA expression. (D) COX-2 mRNA expression. In graphs, error bars represent mean ± SEM; data are presented relative to the “control OA” condition of each time point. Statistical differences were calculated to control conditions. Error bars represent mean ± SEM, and statistical significance for peptide conditions or control non-OA versus control OA condition as determined by unpaired two-tailed Student’s t test is represented as ∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001.
Molecular characterization of peptide bioactivity
A chondrocyte phenotypic consequence of exposure to peptides p[63−82] or p[113−132] is an overall attenuation of hypertrophy. Previously, we17,27 and others29,30 recognized NKX3-2 (BAPX1) as an important negative regulator of chondrocyte hypertrophy, and the hypertrophy-inhibiting action of BMP7 on OA-HACs is, at least in part, mediated via NKX3-2.27 Taking into consideration that the ability to induce NKX3-2 expression was one of the screening criteria for the identification of the here-described, BMP7-derived peptides (Figures 2 and S2), we next investigated whether NKX3-2 is involved in the bioactivity of peptides p[63−82] and p[113−132]. By means of small interfering (si)RNA transfection, NKX3-2 expression was reduced in a pool of OA-HACs. Knockdown of NKX3-2 expression was confirmed (Figure 5A), and in concert with our previous findings,27 expression of chondrocyte hypertrophy-associated genes was sharply induced (Figure 5A). Treatment of control conditions (scrambled siRNA) with peptide p[63−82] or p[113−132]confirmed the peptides’ chondrocyte hypertrophy-inhibiting bioactivity (Figure 5A). OA-HACs with NKX3-2 knockdown and treated with either peptide p[63−82] or p[113−132] did not show rescue of the hypertrophy-inducing consequences of abrogated NKX3-2 expression (Figure 5A). This suggests that the chondrocyte hypertrophy-inhibiting bioactivity of these peptides is NKX3-2 dependent.
Figure 5.

Molecular characterization of peptides p[63−82] and p[113−132]
(A) A pool of OA-HACs (n = 4 donors) was transfected with a scrambled siRNA or with an NKX3-2 siRNA (100 nM siRNA) and cultured for 24 h. Cultures were then exposed to peptide p[63−82] or p[113−132] (100 nM peptide). Samples were harvested 24 h later and analyzed for the expression of the indicated mRNAs (normalized to 28S rRNA). (B) SW1353 chondrocytic cells were transfected with BRE (SMAD1/5/8 reporter) or CAGA12 (SMAD2/3 reporter) firefly luciferase-reporter plasmids. A CMV-Gaussia plasmid was cotransfected as a transfection control. Cells were then cultured for 24 h and subsequently exposed to control, p[63−82], or p[113−132] for 8 h. Firefly and Gaussia bioluminescence were measured in cells and medium (respectively), and relative light units (RLUs) of firefly luciferase were normalized for the Gaussia luciferase signal. The normalized firefly luciferase signal of the control condition was set at 1, and peptide conditions were calculated relative to the control condition (FL/GL). Error bars represent mean ± SEM; statistical significance for peptide conditions versus siScrambled or siNKX3-2 (A) or control condition (B) as determined by unpaired two-tailed Student’s t test is represented as ∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001. (C) An alanine scanning library of peptide p[63−82] was synthesized, and the pool of 18 OA-HACs originally used for the full library scanning (Figure 1) was exposed to individual peptides from the alanine scanning library (100 nM peptide) for 24 h. An alanine-scanning bioactivity score (sum of the fold change gene expression of COL10A1, ALPL, MMP13, and COL2A1 plus NKX3-2 of the alanine-substituted peptides versus the wild-type p[63−82] peptide) was calculated for each alanine substitution in the amino acid sequence of peptide p[63−82] and is represented by the height of each letter on the y axis (data in Figure S4). (D) Multiple sequence alignment of the human BMP7 p[63−82] peptide with the homologous region in BMP7 of other species.
Since peptides p[63−82] and p[113−132] are BMP7 derived, and transcription-repressing activity of NKX3-2 depends on BMP signaling,31 we next determined whether these peptides can modulate SMAD (small mothers against decapentaplegic) transcriptional activity. Activity of the BRE reporter (SMAD1/5/8 dependent32) and the CAGA12 reporter (SMAD3 dependent33) was measured in SW1353 chondrocytes under influence of either one of the peptides. Data showed that both peptides were able to induce the activity of the BRE reporter, whereas CAGA12-reporter activity was diminished (Figure 5B). This indicates that BMP7-derived peptides p[63−82] and p[113−132] can alter SMAD signaling activity in chondrocytes.
We aimed to further understand the sequence dependency of the peptide bioactivity. Peptide p[63−82] was derived from the central region within BMP7, and since this yielded a limited series of bioactive peptides (Figures 1B and S2), we further focused our analyses on peptide p[63−82]. To determine which amino acids are important for the bioactivity of peptide p[63−82], an alanine-scanning peptide library was synthesized, based on systematic alanine substitution of consecutive amino acids in the peptide p[63−82] sequence (Table S1). This alanine-scanning library was then screened by exposing an OA-HAC pool (the same pool that was used for the full library scanning in Figure 1) for 24 h to 100 nM of each peptide in the alanine-scanning library. The bioactivity of each peptide was determined by measuring gene expression of COL10A1, ALPL, MMP13, COL2A1, and NKX3-2 (Figure S4). Changes in the bioactivity of the alanine-scanning library peptides were combined into a “bioactivity score” of each individual substituted amino acid in the parental p[63−82] peptide (Figure 5C). From the bioactivity score, it became evident that Y3A, G7A, E8A, F11A, Y16A, and M17A represent alanine substitutions with an important impact on peptide p[63−82] bioactivity. From these substitutions, G7A had the largest impact on the chondrocyte phenotype-modulating action of the peptide, with a reciprocal response on chondrocyte hypertrophy (Figure S4). The region in BMP7 from which peptide p[63−82] was derived and the G7 position in it are evolutionary highly conserved, as determined by sequence alignment of BMP7 sequences from various animal species (Figure 5D).
Collectively, these data demonstrate that NKX3-2 is important for the bioactivity of peptides p[63−82] and p[113−132] on HACs and that the peptides led to the activation of BRE- and inhibition of CAGA12-reporter activity in chondrocytic cells. The identity of specific amino acids with the p[63−82] peptide sequence is important for its bioactivity, with the glycine (G) residue at position 7 playing an important role in p[63−82] peptide bioactivity on HACs.
Peptide activity on OA knee joint tissues
To determine whether peptide p[63−82] has the potency to alter chondrocyte behavior in cartilage tissue, rather than isolated HACs, cartilage biopsies from OA knees were cultured in the presence of peptide p[63−82], alongside its scrambled version and full-length BMP7. In agreement with our observations in isolated OA-HACs, we found that peptide p[63−82] was specifically able to induce COL2A1 while inhibiting COL10A1 and COX-2 expression in cartilage explants (Figure 6A). This action was comparable to full-length recombinant BMP7. The scrambled version of peptide p[63−82] lacked this activity. Treatment of cartilage explants with BMP7 or peptide p[63−82] had no inhibitory effect on sGAG release (Figure 6B).
Figure 6.

Peptide p[63−82] activity on OA knee joint tissues
Human OA knee joint tissues were cultured for 24 h in the presence of BMP7, peptide p[63−82], or the scrambled version of peptide p[63−82] and compared to control conditions. (A) Cartilage tissues (n = 7 individual donors) were exposed to shown conditions and analyzed for COL2A1, COL10A1, and COX-2 mRNA expression by quantitative real-time PCR (normalized for 28S rRNA expression and relative to control conditions). (B) GAG release in medium from cartilage tissues (n = 5 individual donors) was determined, and data were calculated relative to the control condition. (C) PGE2 levels in culture supernatant from synovium (n = 5 donors). (D) PGE2 levels in culture supernatant from cartilage tissues from (B) (n = 5 donors). (E) PGE2 levels in culture supernatant from IPFP (n = 5 donors). (F) PGE2 levels in culture supernatant from the meniscus. Synovium, IPFP, and meniscus tissues were from the same 5 donors as the cartilage tissues in (B) and (C). PGE2 levels in conditions were calculated relative to controls for each donor. In graphs, error bars represent mean ± SEM; data are presented relative to the control condition for each time point. Statistical differences (unpaired two-tailed Student’s t test) were calculated to control condition: ∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001.
Intra-articularly administered molecules for the treatment of OA are expected to get in contact with multiple intra-articular tissues. In this context, it should be considered that besides a chondrocyte hypertrophy-modulating bioactivity, peptide p[63−82] also influences the inflammatory behavior of OA-HACs (Figures 2, 3, and 4). Therefore, we next tested the anti-inflammatory potential of peptide p[63−82] on cartilage, synovium, infra-patellar fat pad (IPFP), and meniscus explants from knee OA patients. PGE2 levels in culture supernatants of synovial explants were not significantly changed by the peptide nor by BMP7 (Figure 6C). However, PGE2 levels from cartilage, IPFP, and meniscus were reduced by peptide p[63−82] and to a similar extent as by BMP7 (Figures 6D−6F). Together, these data indicate that peptide p[63−82] is able to alter the phenotype of the chondrocyte in cartilage tissue and ex vivo reduces the inflammatory status of knee OA tissues by reducing PGE2 levels.
Attenuation of medial meniscal tear (MMT)-induced cartilage damage
We next tested whether peptide p[63−82] has the potency to delay the progression of trauma-induced cartilage degeneration in the rat MMT model.34,35 MMT damage was unilaterally initiated. Starting at 1 week post-MMT surgery, rats were two times per week intra-articularly injected with saline, 100 ng peptide p[63−82] in saline, or 100 ng scrambled peptide p[63−82] in saline. At 4 weeks post-MMT surgery, rats were sacrificed for histopathological scoring of the MMT knee joints. Visual assessment of frontal sections of the knee joints demonstrated the loss of toluidine blue staining and chondrocyte death, as well as frequent development of a dominant full-thickness cartilage defect in the lateral compartment of the medial tibia plateau in the saline-injected group (Figure 7A, left panel). Histology of the peptide p[63−82]-injected group revealed a certain loss of toluidine blue-staining intensity and decreased articular chondrocyte cellularity in the lateral aspect of the medial tibia plateau. However, cartilage-degenerative changes were less apparent in the p[63−82] group (Figure 7A, middle panel). In contrast, histology of the group injected with scrambled peptide p[63−82] was characterized by diminished toluidine blue staining and lower chondrocyte cellularity at the lateral side of the medial tibia plateau, as well as cartilage degeneration (Figure 7A, right panel). Histopathology scoring35 revealed degenerative changes of the medial tibia articular cartilage in the saline-injected group, with increasing signs of cartilage degeneration from zone 3 (inside) toward zone 1 (outside). The femoral compartment also developed cartilage degeneration but less pronounced. This compartment is also more variable in its degenerative changes.35 There were no statistically significant differences between cartilage-degeneration scores of the tibia or femur in MMT rats injected with saline or scrambled p[63−82] in any of the 3 zones (Figures 7B and 7C). Neither was there a significant difference between saline and scrambled p[63−82] in the total cartilage-degeneration scores (Figures 7B and 7C). Histopathology scoring of the group injected with the bioactive p[63−82] peptide revealed significantly lower medial tibia cartilage-degeneration scores in zone 1 (the most severely affected zone in the MMT model) (Figure 7B) and a trend toward a statistically significant lower medial femur cartilage-degeneration score in zone 1, as compared to the saline group (Figure 7C). Cartilage-degeneration scores in zones 2 and 3 were generally low, as is known for the rat MMT model.35 Combined (total) tibia and femur cartilage-degeneration scores of MMT rats injected with the bioactive p[63−82] peptide were also significantly lower than the saline group (Figures 7B and 7C). A comparison of cartilage-degeneration scores between groups injected with p[63−82] or scrambled p[63−82] did not reveal differences that were statistically significant (Figures 7B and 7C). The scores for depth ratio of tibia cartilage degeneration followed a similar pattern as the tibia cartilage-degeneration scores, with a significantly lower depth ratio in the p[63−82] group compared to saline in zone 1, as well as the total depth ratio score (Figure 7D). In line with these attenuated cartilage-degenerative aspects, the total joint score35 was significantly lower in the group injected with p[63−82] as compared to the saline-injected group (Figure 7E). The average joint score of the group injected with scrambled p[63−82] peptide was in between the values for saline and p[63−82] peptide but was not statistically different compared to either one of the saline or p[63−82] conditions. Other scored histopathological items can be found in Figure S5. Together, these data demonstrate that the p[63−82] peptide, compared to saline control, is effective in reducing cartilage-degenerative changes in the rat MMT model.
Figure 7.

BMP7-derived peptide p[63−82] in rat MMT model
The potency of BMP7 peptide p[63−82] to delay the progression of trauma-induced cartilage degeneration was tested in the rat MMT model. 1 week post-MMT-surgery, rats were two times per week intra-articularly injected with saline, 100 ng peptide p[63−82] in saline, or 100 ng scrambled peptide p[63−82] in saline (10 rats per group). All injection volumes were 50 μL. At 4 weeks post-MMT surgery, rats were sacrificed for histopathological scoring of the MMT knee joints. (A) Representative micrographs of toluidine blue-stained sections of medial aspects of the MMT knee joints. Conditions are indicated. (B) Medial tibia cartilage-degeneration scores. (C) Medial femur cartilage-degeneration scores. (D) Medial tibia cartilage-degeneration depth ratios. Scores and ratios are shown for zones 1, 2, and 3 individually.35 The total scores/ratio are cumulative for zone 1, 2, and 3. (E) Total joint scores. Individual data points represent individual rats. Error bars represent mean ± SEM. Statistical differences were calculated between groups (Mann-Whitney U test), and ∗p < 0.05, ∗∗p < 0.01, and ∗∗∗p < 0.001. Other data from this MMT experiment are shown in Figure S5.
Discussion
Here, we report on the first high-resolution BMP peptide library screening for the identification of peptide sequences derived from BMP7 that attenuate the pathological chondrocyte phenotype associated with OA. Previous attempts to develop peptides derived from BMP family members were based on low-resolution library screening23,24 or focused on mimicking the canonical ligand-receptor interaction.23,26,36 The action of BMPs and BMP-derived peptides is cell-type dependent.28 Therefore, our peptide library screening was not based on a priori knowledge on BMP ligand epitopes or activation of receptor signaling but on the identification of BMP7-derived peptide bioactivity capable of attenuating the articular chondrocyte cell phenotype associated with OA.8,37
Although a majority of the peptides in the library induced an unfavorable chondrocyte hypertrophy-provoking cellular response, peptides from the central and C-terminal regions of BMP7 were active in altering the OA chondrocyte phenotype in a similar fashion as full-length BMP7 on this cell type. The origin of this region-dependent functional separation of BMP7 peptide bioactivity remains unclear at this point. Earlier work reports on a dominant domain in mature BMP2, BMP7, and BMP9 from which peptides are able to support osteogenesis,23,24,38,39 promote chondrogenesis,40 and prevent kidney fibrosis.26 In these cases, peptide sequences were derived from the C-terminal part of the parental BMPs. This region contains the knuckle epitope, which is involved in BMP signaling via the type II BMP receptor (BMPR-II).41 The bioactivity of peptides from the knuckle domain can probably be attributed to the binding of the peptides to the BMPR-II, although knuckle peptide-driven activation of ALK3 (a type I receptor) has also been reported.26 The C-terminal BMP7-derived peptides identified in the present work (represented by p[113−132]) are located in this knuckle domain and overlap with previously reported bioactive peptides from this region of different BMP family members. This indicates that peptides derived from the BMP7 C-terminal domain are likely universally active on a diversity of cell types.
The central region of BMP7 harbors the wrist epitope. This wrist epitope is a key domain shared by BMP protein species and is involved in binding and activating the type IA BMP receptor (BMPR-IA).41 The here-identified bioactive peptides with OA chondrocyte phenotype-modulating activity derived from the central region of BMP7 (represented by p[63−82]) are only partially located within this wrist domain. A peptide covering the complete wrist domain of BMP9 was recently reported to activate BMP signaling in a cell-type-dependent manner.28 However, BMP7-derived peptides present in our library and homologous to the previously described BMP9 wrist domain peptide (p[69−88]−p[79−98]) did not act favorably in altering the OA chondrocyte phenotype. Instead, these and C-terminally adjacent peptides showed induced chondrocyte hypertrophy, which is bioactivity that we were not seeking in this study. A potentially unique aspect of the p[63−82] peptide is the notion that its C-terminal part covers a portion of the wrist domain, whereas its N-terminal part is located within the CXGXC motif of BMP proteins. The CXGXC motif is a major part of the ten-membered cystine knot that is involved in the homodimerization of BMP family members and essential for their action as a receptor ligand.42 Mutation of the G in CXGXC motifs abrogates efficient folding of the domain, impairing dimer formation, as shown previously for the G residue in the CXGXC motif of the human chorionic gonadotropin protein.43 In keeping with this notion, the alanine substitution for which we observed the largest impact on the bioactivity of peptide p[63−82] was G7A, which is the G in the CXGXC motif in the peptide. Although potential disulfide-bridge formation of the peptide with BMP subunits is excluded due to the substitution of all cysteine residues to serine in the library, the major impact of the G7A substitution on peptide p[63−82] function provides a basis for the involvement of functional dimerization of BMP subunits as a potential mechanism for the peptide’s action.
A temporary exposure of OA chondrocytes to the peptides leads to a modulation of the cellular phenotype for up to 8 days. The peptides also provoke a general hypertrophy-suppressive response in other chondrocytic cell models (SW1353, C28I2, ATDC5; data not shown) and activated SMAD1/5/8-dependent BRE-reporter activity while reducing the activity of the SMAD2/3-dependent CAGA12 reporter in SW1353 cells. This is unexpected, since activation of SMAD1/5/8 is generally associated with an OA chondrocyte phenotype and its terminal differentiation.44 In contrast to its clear hypertrophy-suppressive action on OA chondrocytes,27 BMP7 has previously been shown to activate SMAD1/5/8 signaling45,46 and inhibit SMAD3 activity.47 How signaling downstream of different BMPs (e.g., BMP2, BMP4, BMP7, BMP9, etc.) differentiates to BMP-specific target gene expression is largely elusive. As a consequence, it remains to be determined how the action of both peptides alters SMAD-dependent signaling responses related to the chondrocyte phenotype. Our data also reveal that the bioactivity of peptides p[63−82] and p[113−132] depends on the presence of NKX3-2, a well-known BMP-dependent31 transcriptional regulator for chondrocyte hypertrophy.29 This recapitulates the molecular mechanism we previously identified for the BMP7-specific attenuation of the (OA) chondrocyte phenotype.17,27
The BMP7-driven favorable alterations in the (OA) chondrocyte phenotype not only encompass changes in chondrocyte hypertrophy but are additionally accompanied, among others, by dampening of the chondrocyte’s inflammatory status.10,27,48 This is highlighted by BMP7-driven attenuation of chondrocyte COX-2, IL-6, and PGE2 levels in the present work, with similar actions for peptides p[63−82] and p[113−132]. BMP7 has previously also been reported as a morphogen with anti-inflammatory properties in other tissues and cell types like kidney,49 heart,50 blood vessels,50 and macrophages.51 The observation that these peptides attenuate PGE2 release from intra-articular tissue other than cartilage (IPFP, meniscus, and not significantly from synovium) is in line with the previously reported tissue-wide anti-inflammatory actions of BMP7.
When applied intra-articularly, OA disease-modifying molecules will have to reach their target tissue via the SF. Regardless of the method of delivery, peptides should thus be able to survive the OA SF environment to a certain extent. In this respect, it is noteworthy that we found that peptides p[63−82] and p[113−132] are able to ameliorate, at least in part, the negative effects of OA SF on the chondrocyte phenotype. Confirming bioactivity in an in vivo intra-articular environment, we could indeed demonstrate that the progression of cartilage degeneration in the rat MMT model was attenuated by peptide p[63−82] when compared to injection with saline. Histopathology scores of the group injected with a scrambled version of peptide p[63−82] were never significantly different from the group injected with the bioactive peptide p[63−82]. However, no statistically significant differences between scores of the group injected with saline or the scrambled p[62−83] peptide were found either. The average histopathology scores of the scrambled peptide group were mostly in between the scores from the bioactive p[63−83] peptide and the saline group. Together, this indicates that the scrambled p[62−82] peptide may have some influence on cartilage degeneration in this model, but only the bioactive p[62−83] peptide had the potency to harness a significant inhibiting action on cartilage degeneration. The rat MMT model has been used before to determine the potency of a broad-spectrum MMP inhibitor34 and the efficacy of fibroblast growth factor (FGF)18.52 Although the MMT model is generally regarded as a rapidly progressive model for traumatic OA, peptide p[62−83] was able to significantly improve the total joint score in this model to a similar extent as broad-spectrum MMP inhibition34 or MMP13 inhibition.53 Besides the attenuated progression of structural cartilage degeneration detected in the present work, pre-clinical follow-up work should address whether the intra-articular administration of peptide p[62−83] dampens the functional consequences of traumatic OA models by determining gait and load bearing.
In conclusion, this study reports on the first high-resolution peptide library screening of a BMP. The articular chondrocyte phenotypic screening identified two regions in BMP7 from which bioactive peptide sequences are able to attenuate the OA chondrocyte phenotype. The p[62−83] peptide spans the conserved CXGXC motif in BMP7 and reaches into its wrist domain with its exact biomolecular mechanism of action to be dissected. These BMP7-derived peptides may represent novel lead molecules for the development of a disease-modifying treatment of OA.
Materials and methods
Cell and tissue culture
Cartilage was obtained from total knee arthroplasty of end-stage (K&L grades 3−4) OA patients and from resected knee cartilage from cartilage repair procedures of non-OA patients. Medical ethical permission was received from the Maastricht University Medical Center Medical Ethical Committee (approval number 2017-0183). Chondrocytes were isolated using collagenase as described earlier.27 The HACs were cultured until passage two in DMEM/F12 (Life Technologies), 10% fetal calf serum (FCS; Sigma-Aldrich), 1% antibiotic/antimycotic (Life Technologies), and 1% non-essential amino acids (Life Technologies) in a humidified atmosphere at 37°C, 5% CO2. SW1353 cells (ATCC HTB-94, short tandem repeat [STR] profiled)54 were cultured in identical conditions. For experiments, cells were plated in technical triplicates at 30,000 cells/cm2. BMP7 (R&D Systems) was used at 1 nM. OA SF was perioperatively collected from OA patients (same medical ethics statement [2017-0183] as for cartilage above) and was centrifuged to remove cells/debris. For experiments, an SF pool from five OA patients was prepared (equal volume ratios) and applied on cells in a 20% (v/v) concentration. For RNAi, scrambled and NKX3-2 siRNA duplexes (Table S1) were custom made (Eurogentec). Transfection (100 nM siRNA) in HACs was performed using HiPerFect (QIAGEN) according to the manufacturer’s protocol. The BRE reporter,32 CAGA12 reporter,33 or cytomegalovirus (CMV)-Gaussia as transfection control55 were transfected (100 ng/μL) in SW1353 cells using FuGENE (Promega) according to the manufacturer’s protocol. The bioluminescent readout was performed by lysing cells in Passive Lysis Buffer (Promega) and measuring luciferase activity using the Berthold TriStar2 LB 942 Modular Multimode Microplate Reader using the Luciferase Assay System for firefly luciferase (BRE and CAGA12) (Promega) and the Gaussia Luciferase Kit (New England Biolabs). Cartilage from femoral condyles and tibial plateaus, synovial tissue, the inner parts of the IPFP, or the inner parts of the meniscus (carefully avoiding to obtain synovial tissue present at the outer edges of the meniscus) obtained from OA patients (same medical ethical statement [2017-0183] as above for cartilage) were cut into small pieces, washed thoroughly with 0.9% NaCl (Sigma-Aldrich) three times, and cultured in suspension for 24 h in a concentration of 100 mg tissue/mL in DMEM-F12 low glucose, supplemented with 1% insulin-transferrin-selenite (ITS; Invitrogen) and 1% antibiotic/antimycotic.56 After overnight culture, the medium was replaced with peptides or BMP7 for 24 h, after which tissue explants were snap frozen in liquid nitrogen, or conditioned medium was harvested and stored at −80°C.
Peptides
The peptide library was designed from the mature 139-amino acid-long human BMP7 sequence (https://www.ncbi.nlm.nih.gov/protein/4502427; NCBI reference sequence NP_001710). The alanine-scanning library was designed from the peptide sequence covering BMP7 amino acid position 63−82 (p[63−82] from the library). Scrambled sequences were designed from p[63−82] and p[113−132]. All peptides were custom designed and synthesized (Pepscan, Lelystad, the Netherlands) and purified to >90% purity. Peptide sequence identity was confirmed by mass spectrometry. To prevent potential oxidation of free cysteines, all cysteine residues in peptides were substituted for serine residues. Peptide sequences are shown in Table S2.
Gene expression analysis
For the peptide-screening and alanine-scanning experiments, cDNA was prepared using the Cells-to-Ct kit (Invitrogen) according to the manufacturer’s protocol. For the other experiments, cells or homogenized tissues (Mikro-Dismembrator; Braun Biotech International) were lysed in TRIzol (Life Technologies) and RNA isolation and cDNA synthesis were performed as described earlier.27,57 Quantitative real-time PCR was performed using Takyon qPCR Master Mix plus blue for SYBR Green (Eurogentec). A CFX96 Real-Time PCR Detection System (Bio-Rad) was used for amplification: initial denaturation 95°C for 10 min, followed by 40 cycles of amplification (denaturing 15 s at 95°C and annealing 1 min at 60°C). Validated primer sequences are shown in Table S3. Data were analyzed using the standard curve method, mRNA expression was normalized to a reference gene (28S rRNA), and gene expression was calculated as fold change as compared to control conditions.
sGAG assay
The sGAG content was measured using a modified dimethyl methylene blue (DMB) assay.58 The absorbance of samples was read at 540 and 595 nm using a spectrophotometer (Multiskan FC; Thermo Fisher Scientific). GAG concentrations were calculated using a standard curve of chondroitin sulfate (Sigma-Aldrich). GAGs were normalized for total protein content with a bicinchoninic acid assay (BCA) assay (Sigma-Aldrich).
ALP activity assay
Cells were lysed in 1.5 M Tris-HCl (pH 9.0) and 2% (v/v) Triton X-100 and homogenized by sonication (Soniprep 150 MSE). Insoluble material was removed by centrifugation (5 min; 13,000 × g; 4°C). Total protein concentration was determined by a BCA assay. ALP enzyme activity in time was measured by ALP-dependent enzymatic conversion of p-nitrophenyl phosphate to p-nitrophenol in buffer containing 1.5 M Tris-HCl (pH 9.0), 1 mM ZnCl2, 1 mM MgCl2, and 7.5 mM p-nitrophenyl phosphate. Substrate conversion was spectrophotometrically quantified at 405 nm, and p-nitrophenol concentrations were determined via a p-nitrophenol calibration series. Values were normalized to total protein concentration, and ALP enzyme activity was calculated in units (1 U = 1 μmol/g/min).
PGE2 ELISA
PGE2 levels were determined in the culture supernatant. PGE2 concentration was determined by a standardized enzyme immunoassay (EIA) according to the manufacturer’s protocol (Cayman Chemical).
Animal study
The study was conducted via a contract research project at Bolder BioPATH (Boulder, CO, USA); study designs and animal usage were approved by their Institutional Animal Care and Use Committee (IACUC) prior to study initiation (IACUC protocols BBP13-029 and BBP12-004). Animal care, including room, cage, and equipment sanitation, conformed to accepted guidelines cited in the Guide for the Care and Use of Laboratory Animals (the Guide; 2011) and the applicable Bolder BioPATH standard operating procedures (SOPs). Male Lewis rats (n = 48; ∼283 g; Envigo RMS, Indianapolis, IN, USA) underwent a unilateral MMT on study day 0, as established earlier by Bendele et al.59,60 Rats were intra-articularly injected on days 7, 10, 14, 17, 21, and 24 with saline (0.9% NaCl, 50 μL/injection/animal), peptide p[63−82] (100 ng [in 50 μL saline]/injection/animal), or scrambled peptide p[63−82] (100 ng [in 50 μL saline]/injection/animal). The animals were euthanized for necropsy 28 days post-surgery.59,60 Right (surgery) knees from all animals were collected, trimmed of muscle (patella removed), and placed in 10% neutral-buffered formalin and then transferred to 70% ethanol for processing for microscopy. Operated joints were cut into two approximately equal halves in the frontal plane and embedded in paraffin. One section was cut from each knee and stained with toluidine blue. The worst-case scenario for the two halves on each slide was determined and used for evaluation. The tissues were analyzed microscopically by a veterinary pathologist as described earlier35 (Tables S4–S13). An example of the predominant sites where lesions formed in this model in the tibial and femoral cartilage is indicated in Figure S6.
Statistics
Statistical significance for in vitro and ex vivo experiments presented in Figures 1, 2, 3, 4, 5, and 6 was determined by two-tailed Student’s t tests using GraphPad PRISM 5.0 (La Jolla, CA, USA). With the consideration of small sample sizes (Figures 1, 2, 3, 4, and 5, n = 3 samples; Figure 6, n = 5 samples), normal distribution of input data was assumed. For histopathology from the animal study, peptide p[63−82], scrambled peptide p[63−82], and saline groups were compared using a Mann-Whitney U test, as not all input data passed the D’Agostino-Pearson omnibus normality tests. Significance for all tests was set at p ≤0.05. Error bars in graphs represent mean ± standard error of the mean.
Acknowledgments
The authors thank Eke van Zwol for her contributions to Figure 6. This work was financially supported by grants from the Dutch Arthritis Association (13-2-201, 15-3-403, and LLP14) and Stichting De Weijerhorst.
Author contributions
Substantial contributions to research design, M.M.J.C., E.G.J.R., G.v.d.A., N.K.A.P.W., L.W.v.R., P.J.E., and T.J.M.W.; substantial contributions to the acquisition of samples, M.M.J.C., E.G.J.R., G.v.d.A., N.K.A.P.W., J.S., D.A.M.S., A.C., and T.J.M.W.; substantial contributions to analysis, M.M.J.C., E.G.J.R., G.v.d.A., N.K.A.P.W., J.S., D.A.M.S., A.C., and T.J.M.W.; substantial contributions to the interpretation of data, M.M.J.C., E.G.J.R., G.v.d.A., N.K.A.P.W., J.S., D.A.M.S., A.C., P.J.E., L.W.v.R., and T.J.M.W.; drafting the paper, M.M.J.C., E.G.J.R., G.v.d.A., and T.J.M.W.; revising paper critically, M.M.J.C., E.G.J.R., G.v.d.A., N.K.A.P.W., J.S., D.A.M.S., A.C., P.J.E., L.W.v.R., and T.J.M.W.; approval of the submitted and final versions, M.M.J.C., E.G.J.R., G.v.d.A., N.K.A.P.W., J.S., D.A.M.S., A.C., P.J.E., L.W.v.R., and T.J.M.W. All authors have read and approved the final submitted manuscript.
Declaration of interests
The study sponsors had no involvement in study design, collection, analysis, and interpretation of data; the writing of the manuscript; or the decision to submit the manuscript for publication. M.M.J.C. and T.J.M.W. are inventors on patents WO2017178251 and WO2017178253 (owned by Chondropeptix). P.J.E., L.W.v.R., and T.J.M.W. are shareholders in Chondropeptix and are CMO, CDO, and CSO of Chondropeptix, respectively. The other authors declare no competing interests.
Footnotes
Supplemental information can be found online at https://doi.org/10.1016/j.omtm.2021.03.009.
Supplemental information
References
- 1.Loeser R.F., Goldring S.R., Scanzello C.R., Goldring M.B. Osteoarthritis: a disease of the joint as an organ. Arthritis Rheum. 2012;64:1697–1707. doi: 10.1002/art.34453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Sofat N. Analysing the role of endogenous matrix molecules in the development of osteoarthritis. Int. J. Exp. Pathol. 2009;90:463–479. doi: 10.1111/j.1365-2613.2009.00676.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Felson D.T., Lawrence R.C., Dieppe P.A., Hirsch R., Helmick C.G., Jordan J.M., Kington R.S., Lane N.E., Nevitt M.C., Zhang Y. Osteoarthritis: new insights. Part 1: the disease and its risk factors. Ann. Intern. Med. 2000;133:635–646. doi: 10.7326/0003-4819-133-8-200010170-00016. [DOI] [PubMed] [Google Scholar]
- 4.Dreier R. Hypertrophic differentiation of chondrocytes in osteoarthritis: the developmental aspect of degenerative joint disorders. Arthritis Res. Ther. 2010;12:216. doi: 10.1186/ar3117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Tchetina E.V. Developmental mechanisms in articular cartilage degradation in osteoarthritis. Arthritis (Egypt) 2011;2011:683970. doi: 10.1155/2011/683970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.van der Kraan P.M., van den Berg W.B. Chondrocyte hypertrophy and osteoarthritis: role in initiation and progression of cartilage degeneration? Osteoarthritis Cartilage. 2012;20:223–232. doi: 10.1016/j.joca.2011.12.003. [DOI] [PubMed] [Google Scholar]
- 7.Berenbaum F., van den Berg W.B. Inflammation in osteoarthritis: changing views. Osteoarthritis Cartilage. 2015;23:1823–1824. doi: 10.1016/j.joca.2015.09.012. [DOI] [PubMed] [Google Scholar]
- 8.Ji Q., Zheng Y., Zhang G., Hu Y., Fan X., Hou Y., Wen L., Li L., Xu Y., Wang Y., Tang F. Single-cell RNA-seq analysis reveals the progression of human osteoarthritis. Ann. Rheum. Dis. 2019;78:100–110. doi: 10.1136/annrheumdis-2017-212863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hunter D.J. Pharmacologic therapy for osteoarthritis--the era of disease modification. Nat. Rev. Rheumatol. 2011;7:13–22. doi: 10.1038/nrrheum.2010.178. [DOI] [PubMed] [Google Scholar]
- 10.Im H.J., Pacione C., Chubinskaya S., Van Wijnen A.J., Sun Y., Loeser R.F. Inhibitory effects of insulin-like growth factor-1 and osteogenic protein-1 on fibronectin fragment- and interleukin-1beta-stimulated matrix metalloproteinase-13 expression in human chondrocytes. J. Biol. Chem. 2003;278:25386–25394. doi: 10.1074/jbc.M302048200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Stöve J., Schneider-Wald B., Scharf H.P., Schwarz M.L. Bone morphogenetic protein 7 (bmp-7) stimulates proteoglycan synthesis in human osteoarthritic chondrocytes in vitro. Biomed. Pharmacother. 2006;60:639–643. doi: 10.1016/j.biopha.2006.09.001. [DOI] [PubMed] [Google Scholar]
- 12.Chubinskaya S., Otten L., Soeder S., Borgia J.A., Aigner T., Rueger D.C., Loeser R.F. Regulation of chondrocyte gene expression by osteogenic protein-1. Arthritis Res. Ther. 2011;13:R55. doi: 10.1186/ar3300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chubinskaya S., Hurtig M., Rueger D.C. OP-1/BMP-7 in cartilage repair. Int. Orthop. 2007;31:773–781. doi: 10.1007/s00264-007-0423-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hayashi M., Muneta T., Ju Y.J., Mochizuki T., Sekiya I. Weekly intra-articular injections of bone morphogenetic protein-7 inhibits osteoarthritis progression. Arthritis Res. Ther. 2008;10:R118. doi: 10.1186/ar2521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sekiya I., Tang T., Hayashi M., Morito T., Ju Y.J., Mochizuki T., Muneta T. Periodic knee injections of BMP-7 delay cartilage degeneration induced by excessive running in rats. J. Orthop. Res. 2009;27:1088–1092. doi: 10.1002/jor.20840. [DOI] [PubMed] [Google Scholar]
- 16.Hunter D.J., Pike M.C., Jonas B.L., Kissin E., Krop J., McAlindon T. Phase 1 safety and tolerability study of BMP-7 in symptomatic knee osteoarthritis. BMC Musculoskelet. Disord. 2010;11:232. doi: 10.1186/1471-2474-11-232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Caron M.M., Emans P.J., Cremers A., Surtel D.A., Coolsen M.M., van Rhijn L.W., Welting T.J. Hypertrophic differentiation during chondrogenic differentiation of progenitor cells is stimulated by BMP-2 but suppressed by BMP-7. Osteoarthritis Cartilage. 2013;21:604–613. doi: 10.1016/j.joca.2013.01.009. [DOI] [PubMed] [Google Scholar]
- 18.Lohmander L.S., Hoerrner L.A., Lark M.W. Metalloproteinases, tissue inhibitor, and proteoglycan fragments in knee synovial fluid in human osteoarthritis. Arthritis Rheum. 1993;36:181–189. doi: 10.1002/art.1780360207. [DOI] [PubMed] [Google Scholar]
- 19.Mantle D., Falkous G., Walker D. Quantification of protease activities in synovial fluid from rheumatoid and osteoarthritis cases: comparison with antioxidant and free radical damage markers. Clin. Chim. Acta. 1999;284:45–58. doi: 10.1016/s0009-8981(99)00055-8. [DOI] [PubMed] [Google Scholar]
- 20.Tchetverikov I., Lohmander L.S., Verzijl N., Huizinga T.W., TeKoppele J.M., Hanemaaijer R., DeGroot J. MMP protein and activity levels in synovial fluid from patients with joint injury, inflammatory arthritis, and osteoarthritis. Ann. Rheum. Dis. 2005;64:694–698. doi: 10.1136/ard.2004.022434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Montjean R., Escaich S., Paolini R., Carelli C., Pirson S., Neutelings T., Henrotin Y., Vêtu C. REG-O3 chimeric peptide combining growth hormone and somatostatin sequences improves joint function and prevents cartilage degradation in rat model of traumatic knee osteoarthritis. PLoS ONE. 2020;15:e0231240. doi: 10.1371/journal.pone.0231240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Liu Q., Jia Z., Duan L., Xiong J., Wang D., Ding Y. Functional peptides for cartilage repair and regeneration. Am. J. Transl. Res. 2018;10:501–510. [PMC free article] [PubMed] [Google Scholar]
- 23.Kirkwood K., Rheude B., Kim Y.J., White K., Dee K.C. In vitro mineralization studies with substrate-immobilized bone morphogenetic protein peptides. J. Oral Implantol. 2003;29:57–65. doi: 10.1563/1548-1336(2003)029<0057:IVMSWS>2.3.CO;2. [DOI] [PubMed] [Google Scholar]
- 24.Chen Y., Webster T.J. Increased osteoblast functions in the presence of BMP-7 short peptides for nanostructured biomaterial applications. J. Biomed. Mater. Res. A. 2009;91:296–304. doi: 10.1002/jbm.a.32246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kim H.K., Lee J.S., Kim J.H., Seon J.K., Park K.S., Jeong M.H., Yoon T.R. Bone-forming peptide-2 derived from BMP-7 enhances osteoblast differentiation from multipotent bone marrow stromal cells and bone formation. Exp. Mol. Med. 2017;49:e328. doi: 10.1038/emm.2017.40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sugimoto H., LeBleu V.S., Bosukonda D., Keck P., Taduri G., Bechtel W., Okada H., Carlson W., Jr., Bey P., Rusckowski M. Activin-like kinase 3 is important for kidney regeneration and reversal of fibrosis. Nat. Med. 2012;18:396–404. doi: 10.1038/nm.2629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Caron M.M.J., Emans P.J., Surtel D.A.M., van der Kraan P.M., van Rhijn L.W., Welting T.J.M. BAPX-1/NKX-3.2 acts as a chondrocyte hypertrophy molecular switch in osteoarthritis. Arthritis Rheumatol. 2015;67:2944–2956. doi: 10.1002/art.39293. [DOI] [PubMed] [Google Scholar]
- 28.Tong Z., Guo J., Glen R.C., Morrell N.W., Li W. A Bone Morphogenetic Protein (BMP)-derived Peptide Based on the Type I Receptor-binding Site Modifies Cell-type Dependent BMP Signalling. Sci. Rep. 2019;9:13446. doi: 10.1038/s41598-019-49758-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Provot S., Kempf H., Murtaugh L.C., Chung U.I., Kim D.W., Chyung J., Kronenberg H.M., Lassar A.B. Nkx3.2/Bapx1 acts as a negative regulator of chondrocyte maturation. Development. 2006;133:651–662. doi: 10.1242/dev.02258. [DOI] [PubMed] [Google Scholar]
- 30.Yamashita S., Andoh M., Ueno-Kudoh H., Sato T., Miyaki S., Asahara H. Sox9 directly promotes Bapx1 gene expression to repress Runx2 in chondrocytes. Exp. Cell Res. 2009;315:2231–2240. doi: 10.1016/j.yexcr.2009.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kim D.W., Lassar A.B. Smad-dependent recruitment of a histone deacetylase/Sin3A complex modulates the bone morphogenetic protein-dependent transcriptional repressor activity of Nkx3.2. Mol. Cell. Biol. 2003;23:8704–8717. doi: 10.1128/MCB.23.23.8704-8717.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Korchynskyi O., ten Dijke P. Identification and functional characterization of distinct critically important bone morphogenetic protein-specific response elements in the Id1 promoter. J. Biol. Chem. 2002;277:4883–4891. doi: 10.1074/jbc.M111023200. [DOI] [PubMed] [Google Scholar]
- 33.Dennler S., Itoh S., Vivien D., ten Dijke P., Huet S., Gauthier J.M. Direct binding of Smad3 and Smad4 to critical TGF beta-inducible elements in the promoter of human plasminogen activator inhibitor-type 1 gene. EMBO J. 1998;17:3091–3100. doi: 10.1093/emboj/17.11.3091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Janusz M.J., Bendele A.M., Brown K.K., Taiwo Y.O., Hsieh L., Heitmeyer S.A. Induction of osteoarthritis in the rat by surgical tear of the meniscus: Inhibition of joint damage by a matrix metalloproteinase inhibitor. Osteoarthritis Cartilage. 2002;10:785–791. doi: 10.1053/joca.2002.0823. [DOI] [PubMed] [Google Scholar]
- 35.Gerwin N., Bendele A.M., Glasson S., Carlson C.S. The OARSI histopathology initiative - recommendations for histological assessments of osteoarthritis in the rat. Osteoarthritis Cartilage. 2010;18(Suppl 3):S24–S34. doi: 10.1016/j.joca.2010.05.030. [DOI] [PubMed] [Google Scholar]
- 36.Saito A., Suzuki Y., Ogata Si., Ohtsuki C., Tanihara M. Activation of osteo-progenitor cells by a novel synthetic peptide derived from the bone morphogenetic protein-2 knuckle epitope. Biochim. Biophys. Acta. 2003;1651:60–67. doi: 10.1016/s1570-9639(03)00235-8. [DOI] [PubMed] [Google Scholar]
- 37.Ripmeester E.G.J., Timur U.T., Caron M.M.J., Welting T.J.M. Recent Insights into the Contribution of the Changing Hypertrophic Chondrocyte Phenotype in the Development and Progression of Osteoarthritis. Front. Bioeng. Biotechnol. 2018;6:18. doi: 10.3389/fbioe.2018.00018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bergeron E., Senta H., Mailloux A., Park H., Lord E., Faucheux N. Murine preosteoblast differentiation induced by a peptide derived from bone morphogenetic proteins-9. Tissue Eng. Part A. 2009;15:3341–3349. doi: 10.1089/ten.TEA.2009.0189. [DOI] [PubMed] [Google Scholar]
- 39.Suzuki Y., Tanihara M., Suzuki K., Saitou A., Sufan W., Nishimura Y. Alginate hydrogel linked with synthetic oligopeptide derived from BMP-2 allows ectopic osteoinduction in vivo. J. Biomed. Mater. Res. 2000;50:405–409. doi: 10.1002/(sici)1097-4636(20000605)50:3<405::aid-jbm15>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- 40.Renner J.N., Kim Y., Liu J.C. Bone morphogenetic protein-derived peptide promotes chondrogenic differentiation of human mesenchymal stem cells. Tissue Eng. Part A. 2012;18:2581–2589. doi: 10.1089/ten.TEA.2011.0400. [DOI] [PubMed] [Google Scholar]
- 41.Miyazono K., Kamiya Y., Morikawa M. Bone morphogenetic protein receptors and signal transduction. J. Biochem. 2010;147:35–51. doi: 10.1093/jb/mvp148. [DOI] [PubMed] [Google Scholar]
- 42.Groppe J., Greenwald J., Wiater E., Rodriguez-Leon J., Economides A.N., Kwiatkowski W., Affolter M., Vale W.W., Izpisua Belmonte J.C., Choe S. Structural basis of BMP signalling inhibition by the cystine knot protein Noggin. Nature. 2002;420:636–642. doi: 10.1038/nature01245. [DOI] [PubMed] [Google Scholar]
- 43.Kinoshita K., Kusunoki M., Miyai K. Analysis of the three dimensional structure of the CXGXC motif in the CMGCC and CAGYC regions of alpha-and beta-subunits of human chorionic gonadotropin: importance of glycine residue (G) in the motif. Endocr. J. 2006;53:51–58. doi: 10.1507/endocrj.53.51. [DOI] [PubMed] [Google Scholar]
- 44.van der Kraan P.M., Blaney Davidson E.N., Blom A., van den Berg W.B. TGF-beta signaling in chondrocyte terminal differentiation and osteoarthritis: modulation and integration of signaling pathways through receptor-Smads. Osteoarthritis Cartilage. 2009;17:1539–1545. doi: 10.1016/j.joca.2009.06.008. [DOI] [PubMed] [Google Scholar]
- 45.Chang H.M., Cheng J.C., Leung P.C. Theca-derived BMP4 and BMP7 down-regulate connexin43 expression and decrease gap junction intercellular communication activity in immortalized human granulosa cells. J. Clin. Endocrinol. Metab. 2013;98:E437–E445. doi: 10.1210/jc.2012-3851. [DOI] [PubMed] [Google Scholar]
- 46.Huang X., Zhong L., Post J.N., Karperien M. Co-treatment of TGF-β3 and BMP7 is superior in stimulating chondrocyte redifferentiation in both hypoxia and normoxia compared to single treatments. Sci. Rep. 2018;8:10251. doi: 10.1038/s41598-018-27602-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Luo D.D., Phillips A., Fraser D. Bone morphogenetic protein-7 inhibits proximal tubular epithelial cell Smad3 signaling via increased SnoN expression. Am. J. Pathol. 2010;176:1139–1147. doi: 10.2353/ajpath.2010.090459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Fan Z., Chubinskaya S., Rueger D.C., Bau B., Haag J., Aigner T. Regulation of anabolic and catabolic gene expression in normal and osteoarthritic adult human articular chondrocytes by osteogenic protein-1. Clin. Exp. Rheumatol. 2004;22:103–106. [PubMed] [Google Scholar]
- 49.Li R.X., Yiu W.H., Wu H.J., Wong D.W.L., Chan L.Y.Y., Lin M., Leung J.C.K., Lai K.N., Tang S.C.W. BMP7 reduces inflammation and oxidative stress in diabetic tubulopathy. Clin. Sci. (Lond.) 2015;128:269–280. doi: 10.1042/CS20140401. [DOI] [PubMed] [Google Scholar]
- 50.Aluganti Narasimhulu C., Singla D.K. The Role of Bone Morphogenetic Protein 7 (BMP-7) in Inflammation in Heart Diseases. Cells. 2020;9:280. doi: 10.3390/cells9020280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Singla D.K., Singla R., Wang J. BMP-7 Treatment Increases M2 Macrophage Differentiation and Reduces Inflammation and Plaque Formation in Apo E-/- Mice. PLoS ONE. 2016;11:e0147897. doi: 10.1371/journal.pone.0147897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Moore E.E., Bendele A.M., Thompson D.L., Littau A., Waggie K.S., Reardon B., Ellsworth J.L. Fibroblast growth factor-18 stimulates chondrogenesis and cartilage repair in a rat model of injury-induced osteoarthritis. Osteoarthritis Cartilage. 2005;13:623–631. doi: 10.1016/j.joca.2005.03.003. [DOI] [PubMed] [Google Scholar]
- 53.Baragi V.M., Becher G., Bendele A.M., Biesinger R., Bluhm H., Boer J., Deng H., Dodd R., Essers M., Feuerstein T. A new class of potent matrix metalloproteinase 13 inhibitors for potential treatment of osteoarthritis: Evidence of histologic and clinical efficacy without musculoskeletal toxicity in rat models. Arthritis Rheum. 2009;60:2008–2018. doi: 10.1002/art.24629. [DOI] [PubMed] [Google Scholar]
- 54.Gebauer M., Saas J., Sohler F., Haag J., Söder S., Pieper M., Bartnik E., Beninga J., Zimmer R., Aigner T. Comparison of the chondrosarcoma cell line SW1353 with primary human adult articular chondrocytes with regard to their gene expression profile and reactivity to IL-1β. Osteoarthritis Cartilage. 2005;13:697–708. doi: 10.1016/j.joca.2005.04.004. [DOI] [PubMed] [Google Scholar]
- 55.Groot A.J., Habets R., Yahyanejad S., Hodin C.M., Reiss K., Saftig P., Theys J., Vooijs M. Regulated proteolysis of NOTCH2 and NOTCH3 receptors by ADAM10 and presenilins. Mol. Cell. Biol. 2014;34:2822–2832. doi: 10.1128/MCB.00206-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Timur U.T., Caron M.M.J., Bastiaansen-Jenniskens Y.M., Welting T.J.M., van Rhijn L.W., van Osch G.J.V.M., Emans P.J. Celecoxib-mediated reduction of prostanoid release in Hoffa’s fat pad from donors with cartilage pathology results in an attenuated inflammatory phenotype. Osteoarthritis Cartilage. 2018;26:697–706. doi: 10.1016/j.joca.2018.01.025. [DOI] [PubMed] [Google Scholar]
- 57.Caron M.M., Emans P.J., Surtel D.A., Cremers A., Voncken J.W., Welting T.J., van Rhijn L.W. Activation of NF-κB/p65 facilitates early chondrogenic differentiation during endochondral ossification. PLoS ONE. 2012;7:e33467. doi: 10.1371/journal.pone.0033467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Farndale R.W., Sayers C.A., Barrett A.J. A direct spectrophotometric microassay for sulfated glycosaminoglycans in cartilage cultures. Connect. Tissue Res. 1982;9:247–248. doi: 10.3109/03008208209160269. [DOI] [PubMed] [Google Scholar]
- 59.Bendele A.M. Animal models of osteoarthritis. J. Musculoskelet. Neuronal Interact. 2001;1:363–376. [PubMed] [Google Scholar]
- 60.Bendele A.M. Animal models of osteoarthritis in an era of molecular biology. J. Musculoskelet. Neuronal Interact. 2002;2:501–503. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.