

Abstract
In Alzheimer's disease, amyloid‐β (Aβ) accumulates as insoluble plaques in the brain and deposits in blood vessel walls as cerebral amyloid angiopathy (CAA). The severity of CAA correlates with the degree of cognitive decline in dementia. The distribution of Aβ in the walls of capillaries and arteries in CAA suggests that Aβ is deposited in the perivascular pathways by which interstitial fluid drains from the brain. Soluble Aβ from the extracellular spaces of gray matter enters the basement membranes of capillaries and drains along the arterial basement membranes that surround smooth muscle cells toward the leptomeningeal arteries. The motive force for perivascular drainage is derived from arterial pulsations combined with the valve effect of proteins present in the arterial basement membranes. Physical and biochemical changes associated with arteriosclerosis, aging and possession of apolipoprotein E4 genotype lead to a failure of perivascular drainage of soluble proteins, including Aβ. Perivascular cells associated with arteries and the lymphocytes recruited in the perivenous spaces contribute to the clearance of Aβ. The failure of perivascular clearance of Aβ may be a major factor in the accumulation of Aβ in CAA and may have significant implications for the design of therapeutics for the treatment of Alzheimer's disease.
Keywords: Alzheimer's disease, amyloid β, cerebral amyloid angiopathy, interstitial fluid neuroimmunology, perivascular drainage
Anatomy of the Cerebral Vasculature
Arteries penetrate the cerebral cortex perpendicular to the pial surface 25. As they enter the cortex from the subarachnoid space, arteries gain a complete layer of pia mater, separating the subarachnoid space from the perivascular Virchow‐Robin spaces 95. Macrophages recruited in part from the pool of peripheral monocytes reside within these perivascular spaces 6. Cortical arteries are, in fact, arterioles as they do not have elastic laminae, but they possess a tunica media, with multiple layers of smooth muscle, separated by a spiral basement membrane (Figure 1A). Basement membranes are thin (∼150 nm) sheets of glycoproteins and proteoglycans that provide functional and structural support to the vessel 94. The larger proteins, type IV collagen and laminins, are linked by integrins and nidogen and bound by chondroitin and heparan sulfate proteoglycans 66, 93. As the penetrating arteries give rise to smaller arterioles, the layer of pial cells becomes incomplete, leading to an absence of both pia and perivascular spaces at the capillary level 95. Cerebral capillaries have an endothelium with tight junctions, lined by an endothelial basement membrane that is fused with an astroglial basement membrane (Figure 1B). Pericytes are contractile cells completely surrounded by basement membrane 28, 35, 45. Cerebral veins do not possess a smooth muscle layer and have very little basement membrane. Perivascular spaces around the vein are confluent with the subpial space and only small numbers of pial cells are associated with the vessel wall 81. Lymphocytes pass into the brain across the venous endothelium and dilate perivenous Virchow‐Robin spaces 6, 26, 84. The recruitment of neutrophils requires G protein‐coupled receptor‐dependent activation of β2 integrins and binding to ICAM‐1 present on the endothelium 34. The crawling of neutrophils upon the venous endothelium is mediated by endothelial ICAM‐1 and ICAM‐2 and neutrophil LFA‐1 and Mac‐1 34.
Figure 1.
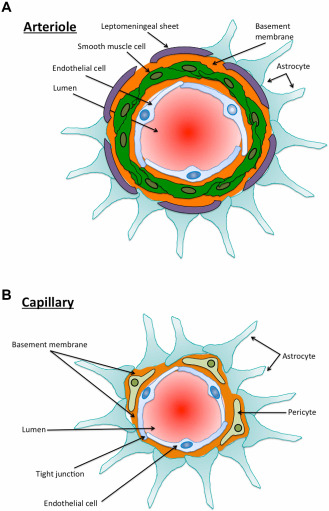
A. Diagrammatic representation of a cross section through a cerebral cortical arteriole. There are no elastic laminae, but there is a tunica media with smooth muscle cells (green) and basement membrane (orange) interposed between the smooth muscle cells. The astrocyte end‐feet (light blue) abut upon the leptomeningeal sheet that reflects upon the artery as it penetrates the brain from the subarachnoid space. B. Diagrammatic representation of a capillary in the cerebral cortex. The endothelial cells are connected by tight junctions. On the abluminal border of the endothelial cells there is a basement membrane that results from the fusion of the basement membranes of the endothelium and the astrocyte end‐feet. Pericytes are completely surrounded by basement membrane.
Perivenous Infiltrates of Lymphocytes: Significance for Immunological Aspects of Neurodegenerative Diseases
Taking into account the anatomy of brain vessels, immune cells need to penetrate two distinct barriers in their way from the blood stream to the brain parenchyma: the vascular wall with its basement membranes (first step of neuroinflammation) and the glia limitans which also includes a separate basement membrane (second step of neuroinflammation). The vascular and the glial basement membranes differ in their content of laminin isoforms 71. As the glial basement membrane is anchored to dystroglycan, which is cleaved by matrix metalloproteases (MMP) 2 and 9 1, blockage of MMP using BB‐94 blocks the penetration of the glia limitans and restricts immune cells to perivascular spaces 76. The first step is mainly driven by CCL‐2 76, which is crucial for the recruitment of Ly‐6ChiCCR2 + monocytes, for example, after irradiation 53. Thus, while recruitment of immune cells into perivascular spaces requires release of CCL2, additional signals are required for the induction of MMPs. While the latter causes intraparenchymal infiltration and neurological symptoms, the former may be regarded as a form immune surveillance.
Cerebral Capillaries and Arteries Host the Pathways for Perivascular Drainage of Interstitial Fluid (ISF ISF)
The pathways for perivascular drainage of interstitial fluid and solutes have been identified both in experimental animals 13 and in humans 61, 85. The pathway for the bulk flow drainage of ISF and solutes from the brain has been characterized in detail by injecting 0.5 μL of fluorescent dextran of 3–10 kDa or ovalbumin 49 kDa into the gray matter of the mouse 13 (Figure 2A). Drainage of solutes from the extracellular spaces appears to be very fast, occurring within 5–15 minutes. By 5 minutes after injection, tracers had spread diffusely through the extracellular spaces and entered the walls of blood vessels. Confocal microscopy showed that tracers were co‐localized with laminin in the basement membranes of capillaries and in the basement membranes of the tunica media of arteries 13. Microinfusion studies using 14C‐labeled glucose and d‐lactate demonstrate their perivascular drainage from the inferior colliculus to the arteries at the base of the brain 4.
Figure 2.

A. Diagrammatic representation of perivascular clearance of solutes, normally entering the walls (basement membranes) of capillaries and then arteries, but not of veins. B. Cross section through a human artery at the surface of the brain. The tissue has been immunostained for collagen IV (a marker of basement membranes, shown here in blue), smooth muscle cells (immunostaining for smooth muscle actin, shown in green) and amyloid‐β (Aβ), using anti‐4G8 antibody, shown here in red. Note the presence of Aβ surrounding the smooth muscle cells, leaving the endothelial basement membrane (blue) free of Aβ. There is some Aβ accumulating at the surface of the brain, with the collagen IV present in glia limitans. Confocal microscopy. Scale bar: 20 μm.
Within 5 minutes of injections, in the arterial walls, the tracers were selectively located in the basement membranes between the smooth muscle cells in the tunica media and not in the endothelial basement membranes nor in the outer basement membrane at the interface between the artery and the extracellular matrix. By 3 h after injection, the tracers had disappeared from the extracellular spaces of the gray matter and from the basement membranes in the walls of capillaries and arteries. At this time, tracers were observed in the perivascular macrophages within the ipsilateral parenchyma and in the leptomeninges 13. At 24 h after injection, the tracers were present in the perivascular macrophages adjacent to the ipsilateral intracerebral arteries and around leptomeningeal arteries on the surface of the brain 13.
Particles of 1000 nm size do not enter capillary or arterial basement membranes and remain confined to the injection site, whereas particles of 20 nm enter the basement membranes of capillaries and the Virchow‐Robin perivascular spaces, where particulate matter can be observed for up to 2 years 13, 43. The exact capacity of the cerebrovascular basement membranes for clearance is still unclear and requires an in vivo experimental approach using a range of nanoparticulate sizes 30.
Because of the limited sensitivity of fluorescent markers injected in low volumes into the ISF, it was not possible to determine whether they reach cervical lymph nodes. However, older studies using radioactive tracers show that they drain along the tunica media and the tunica adventitia of the major cerebral arteries, through the base of the skull to deep cervical lymph nodes 72. The perivascular drainage of ISF appears to be separated from the cerebrospinal fluid (CSF), as only 10%–15% of tracers injected into the deep gray matter of the brain can be identified in the CSF 72.
Motive forces for the perivascular drainage of interstitial fluid
It is difficult to investigate experimentally the motive force for perivascular drainage of ISF and solutes from the brain, as the timescale is fast and the pathways are inaccessible and small (100–150 nm) 36. However, the observation that perivascular transport only occurs in living animals and ceases immediately after cardiac arrest suggests that pulsations in artery walls may generate the motive force for the transport of ISF and solutes out of the brain 4. Mathematical models indicate that perivascular transport of ISF and solutes may be driven by the contrary (reflection) waves that follow each pulse wave 67, 80. A valve‐like action is required in order to prevent reflux during the passage of the major pulse wave along the vessel wall 67. As the route of drainage is within basement membranes 13, it is possible that the valve action results from changes in the conformation of the basement membrane or from different biochemical interactions. During cardiac systole and the pulse wave, the arterial internal elastic lamina and basement membranes are stretched as the vessels expand 83. During diastole, the vessel wall expands allowing the contrary wave to pass back along the artery wall and the basement membranes to also expand. These rhythmical changes in the dimensions and conformation of the basement membranes may provide the necessary valve action to ensure that ISF is driven along the artery walls toward leptomeningeal arteries 67. Mathematical fluid flow modeling demonstrates that peristaltic contractions in the walls of arteries, comparable to the pulse contractions, are enough to drive ISF along perivascular drainage pathways 80.
A recent detailed transcardial Doppler analysis of the blood flow through the circle of Willis demonstrated that mean flow velocity and the pulsatility index are reduced with age alone and decrease further with disease 63. This suggests that aging vessels become stiff, less elastic and arteriosclerotic, particularly in humans, interfering with perivascular drainage of ISF and solutes in elderly individuals 82. Administering the calcium channel antagonist nimodipine did not affect the pattern or the rate of perivascular drainage of soluble dextrans 5. In this study, nimodipine was co‐infused with the tracer, resulting in a local vasodilation, rather than a general systemic effect on the pulsatile blood flow.
Cerebral Amyloid Angiopathy (CAA): Failure of Perivascular Drainage of Amyloid‐β (Aβ)
Aβ accumulates in the human brain with age and in Alzheimer's disease (AD) as insoluble plaques in brain parenchyma and in the walls of blood vessel as CAA 55. In both humans and AD transgenic mouse models, it is mainly the more soluble Aβ40 that is deposited in the vessel walls 64, whereas the less soluble Aβ42 is found mainly in the senile plaques 23. Raised levels of soluble Aβ in the cerebral cortex and the severity of CAA appear to correlate better with the severity of cognitive decline and dementia than does the number of insoluble plaques of Aβ in brain parenchyma 48, 57, 79.
In familial forms of AD and in transgenic mouse models of AD, Aβ is overproduced or there is production of aberrant forms of Aβ resulting in the accumulation of Aβ in the brain and blood vessel walls 91. However, there is little evidence for overproduction of Aβ in the more common sporadic form of AD. It is, therefore, likely that accumulation of Aβ in the brain and in the vessel walls in AD is caused by failure of elimination of Aβ from the aging brain.
A number of different mechanisms and pathways for the elimination of Aβ from the brain have been identified in experimental animals and in humans. Enzymes such as the catabolic peptidase, neprilysin, and the metalloproteinase, insulin‐degrading enzyme, degrade Aβ in brain parenchyma 54, 70. Neprilysin activity declines significantly with age in the mouse hippocampus 27. Upregulation of neprilysin in aged SwAPP transgenic mice does not lower the level of toxic Aβ oligomers and does not improve cognition, despite the removal of Aβ plaques from the brain 51. Peripherally administered neprilysin reduce levels of Aβ in the blood, but not in the brain 39.
Antibodies against Aβ in the serum remove Aβ from the brain 22 and microglia degrade Aβ in vivo and in vitro, in mice and humans immunized against the Aβ peptides 2, 88. Patrolling monocytes and perivascular macrophages regulate the severity of CAA via mechanisms involving scavenger receptor A, class B type 1 and CD36 47, 52, 58, 75. Astrocytes and smooth muscle cells also contribute to the uptake and degradation of Aβ in a process that may be mediated by the expression of aquaporin‐4 40, 62. Aβ is absorbed into the blood by low‐density lipoprotein receptor‐related protein‐1 (LRP) and P‐glycoprotein, with α2‐macroglobulin and apolipoprotein E also acting upon these pathways of clearance 7, 21, 68. However, LRP‐1 expression in endothelial cells and the effectiveness of this pathway are reduced in older mice and possibly in AD 7. Concomitant with enzymatic, cellular and transendothelial transport, soluble Aβ is also cleared by the perivascular bulk flow drainage of ISF 86.
The predominant distribution of Aβ in the walls of large cortical and leptomeningeal arteries in CAA support the hypothesis that Aβ is deposited in the perivascular pathways by which interstitial fluid normally drains from the brain 14 (Figure 2B). Soluble tracers, including Aβ40, injected into the ISF of the adult mouse brain distribute along basement membranes in the same pattern as Aβ deposition in human CAA 37. Increased severity of vascular load of Aβ42 is observed following anti‐Aβ immunotherapy experiments in mice and humans, suggesting that Aβ solubilized from parenchymal plaques drains along and becomes entrapped within perivascular clearance pathways 10. Other amyloid species, including cystatin, transthyretin and ABri 50, also accumulate in the vessel walls of the brain, indicating impairment of ISF drainage pathways as a common factor in the pathogensesis of CAA regardless of the amyloidogenic peptide involved. Such accumulations are often associated with alternations in basement membranes and precede endogenous CAA development in the TGF‐β1 transgenic mice 89, 90.
Factors that Disrupt Perivascular Drainage of Aβ from the Brain
Age
Age is a major risk factor for both AD and cerebrovascular disease (CVD). CVD encompasses a number of changes in the cerebral blood vessels of aging humans. There is progressive fibrosis of the intima and media in major cerebral arteries and flattening of internal elastic laminae, reflecting the generalized stiffening and loss of elasticity of artery walls with age (arteriosclerosis) 12. Focal plaques of fibrosis and lipid deposition result in atherosclerosis, and arteries may be occluded by thrombi and emboli, leading to infarction and poor vascular perfusion 8. In addition to directly affecting cerebral perfusion, CVD may contribute to AD dementia by altering the efficiency of perivascular clearance of Aβ from the elderly brain. Reduction in the amplitude of arterial pulsations is predicted to reduce the motive force driving perivascular clearance of Aβ, leading to its accumulation 67. This hypothesis is supported by experimental findings that solute clearance along basement membranes is decreased in aged mice 36, 37 and when perfusion was impaired by occlusion of the middle cerebral artery 33. Histological observations also confirm that thrombotic occlusion of cortical arteries is associated with deposition of Aβ in the walls of the capillaries served by those arteries, suggesting that Aβ is entrapped in capillary walls that form the proximal portions of the perivascular interstitial fluid drainage pathways 92.
Aging is also associated with changes in the biochemical composition of basement membranes in both humans and experimental models. Age‐dependent downregulation in collagen IV and laminin and upregulation of perlecan and fibronectin are observed in the blood vessels of mouse brains 36, 37. Small‐diameter vessels from AD patients show a decrease in the amount of collagen IV, compared to aged‐matched controls 17, while levels of heparan sulfate proteoglycans are increased 8, 69. Laminin, nidogens and collagen IV inhibit aggregation of Aβ and destabilize pre‐formed fibrils of Aβ 11. Heparan sulfate proteoglycans such as fibronectin and perlecan accelerate Aβ aggregation through high‐affinity interactions 15, 19. This age‐related shift toward increased expression of pro‐amyloidogenic basement membrane proteins may promote CAA by increasing the affinity of the vessel wall for Aβ. CAA deposition may further impair perivascular drainage pathways 2, 37, leading to a feedforward impairment in the elimination of not only Aβ but also other toxic solutes from the brain that may contribute to cognitive decline in AD.
Apolipoprotein E (ApoE) genotype
ApoE is the major carrier of cholesterol in the brain, regulating lipid homeostasis and is expressed by astrocytes, smooth muscle cells and neurons 46. The ApoE gene codes for three different alleles (ε2, 3, 4). Possession of one or two copies of ApoE4 increases the risk of developing AD and reduces the median age of onset 3, 16, 74, 77. Further, deposition of Aβ in capillary walls and increased CAA severity are observed in the brains of humans and transgenic mice expressing human apoE4 31, 60, 74.
ApoE genotype influences the efficiency of most of the Aβ clearance mechanisms in the brain, including enzymatic 18, 24, microglial/monocytic 78, 96 and transendothelial 20. Perivascular drainage pathways are also affected by the expression of ApoE4. Immunocytochemical and Western blot analysis of basement membranes demonstrated that levels of laminin and collagen IV were higher, while fibronectin expression was lower, in the brains of 3‐month old targeted replacement (TR) mice expressing the human APOE4 (TRE4) allele compared to those expressing the human APOE3 (TRE3) allele and wild‐type controls 41. In 16‐month‐old mice, laminin and collagen IV levels were unchanged between wild‐type and TRE3 mice but lower in the blood vessels of TRE4 mice. The pattern of distribution of Aβ40 following injection into the hippocampus was significantly disrupted in the blood vessels of TRE4 mice, as early as 3 months of age 38. In 16‐month‐old TRE4 mice, soluble Aβ40 aggregated within the periarterial drainage pathways, suggesting that APOE4 is associated with a significant risk of failure of clearance of Aβ40. Interestingly, although ApoE co‐localized with laminin in the blood vessels of both TRE3 and TRE4 mice, no co‐localization was noted between ApoE and deposits of Aβ in the vessels of either TRE3 or TRE4 mice 38. Thus, the role of ApoE in the clearance of Aβ along perivascular pathways remains to be fully elucidated.
Immune complexes
Anti‐Aβ immunotherapy trials for the treatment of AD in humans were introduced following successful studies in transgenic mice demonstrating that insoluble Aβ plaques were removed from the brain following immunization of mice with Aβ42 65. However, despite the clearance of plaques in patients with AD 29, 49, 56 active immunization significantly increased arterial CAA load in both transgenic animals and in humans 56, 87. As these vascular Aβ deposits were composed principally of Aβ42, it was hypothesized that Aβ solubilized from Aβ42‐positive plaques became trapped in the perivascular drainage pathway 59, 82. Both active 9, 32 and passive 41, 42 immunotherapy trials have encountered side effects comprising focal abnormalities in the cerebral white matter on imaging. The pathophysiology underlying these side effects is as yet unclear but it may be caused by the effects of immune complexes in relation to the cerebral vasculature.
In a mouse model of immune complex formation in the brain, the antigen (ovalbumin), IgG and complement C3 were located in basement membranes of artery walls 24 h after challenge with antigen 73. It is likely that the presence of immune complexes in the walls of arteries will be associated with significant disturbance of the periarterial drainage pathways.
Implications for Therapy of AD
Increasing evidence suggests that cerebrovasculature health is important for both the prevention and the treatment of AD. Risk factors for the development of cardiovascular disease, such as diabetes, hypertension and hypercholesterolemia, also increase the risk of AD 44. Maintaining the health of cerebral vessels by preventing or minimizing the impact of such modifiable risk factors may therefore help to promote efficient clearance of Aβ across the blood–brain barrier and along perivascular pathways of the aging brain. Moreover, as indicated by findings from the anti‐Aβ immunization trials, the success of Aβ‐based therapies is strongly dependent on the robustness of cerebral blood vessels and such factors must be taken into consideration in the design of new therapeutics for the treatment of AD.
Conclusions
Perivascular drainage along cerebrovascular basement membranes is a major route by which Aβ is eliminated from the brain. Theoretical and experimental models support the hypothesis that efficient drainage of Aβ along perivascular pathways is dependent on the biochemical composition of the basement membrane and the force of arterial pulsations. Factors that affect cerebrovascular health, such as age and APOE genotype, alter both the structure of the blood vessels and the expression of the basement membrane proteins such that the efficiency of perivascular drainage of Aβ is reduced. As increasing amounts of Aβ become entrapped within the drainage pathways, it causes damage to the underlying vasculature, further reducing the functionality of the vessel and creating a feedforward mechanism by which increasing amounts of Aβ accumulate as CAA. Finally, diffusion of soluble Aβ and ISF through brain tissue is blocked by insoluble Aβ in the extracellular spaces, levels of soluble Aβ and other metabolites in brain parenchyma rise and dementia ensues.
Acknowledgments
We wish to thank Alzheimer's Research UK and Rosetrees Trust for funding work that was presented here. We are grateful to Professors Johannes Attems, Rajesh Kalaria, the Newcastle Brain Tissue Resource Brain Bank and to the patients and their relatives for the human tissue.
The copyright line for this article was changed on June 19, 2014 after original online publication.
References
- 1. Agrawal S, Anderson P, Durbeej M, van Rooijen N, Ivars F, Opdenakker G, Sorokin LM (2006) Dystroglycan is selectively cleaved at the parenchymal basement membrane at sites of leukocyte extravasation in experimental autoimmune encephalomyelitis. J Exp Med 203:1007–1019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Arbel‐Ornath M, Hudry E, Eikermann‐Haerter K, Hou S, Gregory JL, Zhao L et al (2013) Interstitial fluid drainage is impaired in ischemic stroke and Alzheimer's disease mouse models. Acta Neuropathol 126:353–364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Ashford JW (2004) APOE genotype effects on Alzheimer's disease onset and epidemiology. J Mol Neurosci 23:157–165. [DOI] [PubMed] [Google Scholar]
- 4. Ball KK, Cruz NF, Mrak RE, Dienel GA (2010) Trafficking of glucose, lactate, and amyloid‐beta from the inferior colliculus through perivascular routes. J Cereb Blood Flow Metab 30:162–176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Barua NU, Bienemann AS, Hesketh S, Wyatt MJ, Castrique E, Love S, Gill SS (2012) Intrastriatal convection‐enhanced delivery results in widespread perivascular distribution in a pre‐clinical model. Fluids Barriers CNS 9:2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Bechmann I, Priller J, Kovac A, Bontert M, Wehner T, Fernandez Klett F et al (2001) Immune surveillance of mouse brain perivascular spaces by blood‐borne macrophages. Eur J Neurosci 14:1651–1658. [DOI] [PubMed] [Google Scholar]
- 7. Bell RD, Sagare AP, Friedman AE, Bedi GS, Holtzman DM, Deane R, Zlokovic BV (2007) Transport pathways for clearance of human Alzheimer's amyloid beta‐peptide and apolipoproteins E and J in the mouse central nervous system. J Cereb Blood Flow Metab 27:909–918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Berzin TM, Zipser BD, Rafii MS, Kuo‐LeBlanc V, Yancopoulos GD, Glass DJ et al (2000) Agrin and microvascular damage in Alzheimer's disease 2. Neurobiol Aging 21:349–355. [DOI] [PubMed] [Google Scholar]
- 9. Boche D, Nicoll JA (2008) The role of the immune system in clearance of Abeta from the brain. Brain Pathol 18:267–278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Boche D, Zotova E, Weller RO, Love S, Neal JW, Pickering RM et al (2008) Consequence of Abeta immunization on the vasculature of human Alzheimer's disease brain. Brain 131(Pt 12):3299–3310. [DOI] [PubMed] [Google Scholar]
- 11. Bronfman FC, Alvarez A, Morgan C, Inestrosa NC (1998) Laminin blocks the assembly of wild‐type A beta and the Dutch variant peptide into Alzheimer's fibrils. Amyloid 5:16–23. [DOI] [PubMed] [Google Scholar]
- 12. Brown WR, Thore CR (2011) Review: cerebral microvascular pathology in ageing and neurodegeneration. Neuropathol Appl Neurobiol 37:56–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Carare RO, Bernardes‐Silva M, Newman TA, Page AM, Nicoll JA, Perry VH, Weller RO (2008) Solutes, but not cells, drain from the brain parenchyma along basement membranes of capillaries and arteries: significance for cerebral amyloid angiopathy and neuroimmunology. Neuropathol Appl Neurobiol 34:131–144. [DOI] [PubMed] [Google Scholar]
- 14. Carare RO, Hawkes CA, Jeffrey M, Kalaria RN, Weller RO (2013) Cerebral amyloid angiopathy, prion angiopathy, CADASIL and the spectrum of protein elimination‐failure angiopathies (PEFA) in neurodegenerative disease with a focus on therapy. Neuropathol Appl Neurobiol 39:593–611. [DOI] [PubMed] [Google Scholar]
- 15. Castillo GM, Ngo C, Cummings J, Wight TN, Snow AD (1997) Perlecan binds to the beta‐amyloid proteins (A beta) of Alzheimer's disease, accelerates A beta fibril formation, and maintains A beta fibril stability. J Neurochem 69:2452–2465. [DOI] [PubMed] [Google Scholar]
- 16. Chalmers K, Wilcock GK, Love S (2003) APOE epsilon 4 influences the pathological phenotype of Alzheimer's disease by favouring cerebrovascular over parenchymal accumulation of A beta protein. Neuropathol Appl Neurobiol 29:231–238. [DOI] [PubMed] [Google Scholar]
- 17. Christov A, Ottman J, Hamdheydari L, Grammas P (2008) Structural changes in Alzheimer's disease brain microvessels. Curr Alzheimer Res 5:392–395. [DOI] [PubMed] [Google Scholar]
- 18. Cook DG, Leverenz JB, McMillan PJ, Kulstad JJ, Ericksen S, Roth RA et al (2003) Reduced hippocampal insulin‐degrading enzyme in late‐onset Alzheimer's disease is associated with the apolipoprotein E‐epsilon4 allele. Am J Pathol 162:313–319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Cotman SL, Halfter W, Cole GJ (2000) Agrin binds to á‐amyloid (Aá), accelerates Aá fibril formation, and is localized to Aá deposits in Alzheimer's disease brain. Mol Cell Neurosci 15:183–198. [DOI] [PubMed] [Google Scholar]
- 20. Deane R, Sagare A, Hamm K, Parisi M, Lane S, Finn MB et al (2008) apoE isoform‐specific disruption of amyloid beta peptide clearance from mouse brain. J Clin Invest 118:4002–4013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Deane R, Wu Z, Sagare A, Davis J, Du Yan S, Hamm K et al (2004) LRP/amyloid beta‐peptide interaction mediates differential brain efflux of Abeta isoforms. Neuron 43:333–344. [DOI] [PubMed] [Google Scholar]
- 22. DeMattos RB, Bales KR, Cummins DJ, Dodart JC, Paul SM, Holtzman DM (2001) Peripheral anti‐A beta antibody alters CNS and plasma A beta clearance and decreases brain A beta burden in a mouse model of Alzheimer's disease 20. Proc Natl Acad Sci USA 98:8850–8855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Dickson DW, Farlo J, Davies P, Crystal H, Fuld P, Yen SHC (1988) A double‐labeling immunohistochemical study of senile plaques. Am J Pathol 132:86–101. [PMC free article] [PubMed] [Google Scholar]
- 24. Du J, Chang J, Guo S, Zhang Q, Wang Z (2009) ApoE 4 reduces the expression of Abeta degrading enzyme IDE by activating the NMDA receptor in hippocampal neurons. Neuroscience Lett 464:140–145. [DOI] [PubMed] [Google Scholar]
- 25. Duvernoy HM, Delon S, Vannson JL (1981) Cortical blood vessels of the human brain 5. Brain Res Bull 7:519–579. [DOI] [PubMed] [Google Scholar]
- 26. Engelhardt B, Wolburg H (2004) Mini‐review: transendothelial migration of leukocytes: through the front door or around the side of the house? Eur J Immunol 34:2955–2963. [DOI] [PubMed] [Google Scholar]
- 27. Farris W, Schutz SG, Cirrito JR, Shankar GM, Sun X, George A et al (2007) Loss of neprilysin function promotes amyloid plaque formation and causes cerebral amyloid angiopathy. Am J Pathol 171:241–251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Fernandez‐Klett F, Offenhauser N, Dirnagl U, Priller J, Lindauer U (2010) Pericytes in capillaries are contractile in vivo, but arterioles mediate functional hyperemia in the mouse brain. Proc Natl Acad Sci USA 107:22290–22295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Ferrer I, Boada RM, Sanchez Guerra ML, Rey MJ, Costa‐Jussa F (2004) Neuropathology and pathogenesis of encephalitis following amyloid‐beta immunization in Alzheimer's disease 49. Brain Pathol 14:11–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Foley CP, Nishimura N, Neeves KB, Schaffer CB, Olbricht WL (2012) Real‐time imaging of perivascular transport of nanoparticles during convection‐enhanced delivery in the rat cortex. Ann Biomed Eng 40:292–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Fryer JD, Simmons K, Parsadanian M, Bales KR, Paul SM, Sullivan PM, Holtzman DM (2005) Human apolipoprotein E4 alters the amyloid‐beta 40:42 ratio and promotes the formation of cerebral amyloid angiopathy in an amyloid precursor protein transgenic model 1. J Neurosci 25:2803–2810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Gilman S, Koller M, Black RS, Jenkins L, Griffith SG, Fox NC et al (2005) Clinical effects of Abeta immunization (AN1792) in patients with AD in an interrupted trial. Neurology 64:1553–1562. [DOI] [PubMed] [Google Scholar]
- 33. Go AS, Mozaffarian D, Roger VL, Benjamin EJ, Berry JD, Borden WB et al (2013) Heart disease and stroke statistics—2013 update: a report from the American Heart Association. Circulation 127:e6–e245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Gorina R, Lyck R, Vestweber D, Engelhardt B (2014) beta2 integrin‐mediated crawling on endothelial ICAM‐1 and ICAM‐2 Is a prerequisite for transcellular neutrophil diapedesis across the inflamed blood‐brain barrier. J Immunol 192:324–337. [DOI] [PubMed] [Google Scholar]
- 35. Graeber MB, Streit WJ, Buringer D, Sparks DL, Kreutzberg GW (1992) Ultrastructural location of major histocompatibility complex (MHC) class II positive perivascular cells in histologically normal human brain 80. J Neuropathol Exp Neurol 51:303–311. [DOI] [PubMed] [Google Scholar]
- 36. Hawkes CA, Gatherer M, Sharp MM, Dorr A, Yuen HM, Kalaria R et al (2013) Regional differences in the morphological and functional effects of aging on cerebral basement membranes and perivascular drainage of amyloid‐beta from the mouse brain. Aging Cell 12:224–236. [DOI] [PubMed] [Google Scholar]
- 37. Hawkes CA, Hartig W, Kacza J, Schliebs R, Weller RO, Nicoll JA, Carare RO (2011) Perivascular drainage of solutes is impaired in the ageing mouse brain and in the presence of cerebral amyloid angiopathy. Acta Neuropathol 121:431–443. [DOI] [PubMed] [Google Scholar]
- 38. Hawkes CA, Sullivan PM, Hands S, Weller RO, Nicoll JA, Carare RO (2012) Disruption of arterial perivascular drainage of amyloid‐beta from the brains of mice expressing the human APOE epsilon4 Allele. PLoS ONE 7:e41636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Henderson SJ, Andersson C, Narwal R, Janson J, Goldschmidt TJ, Appelkvist P et al (2014) Sustained peripheral depletion of amyloid‐beta with a novel form of neprilysin does not affect central levels of amyloid‐beta. Brain 137:553–564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Iliff JJ, Wang M, Liao Y, Plogg BA, Peng W, Gundersen GA et al (2012) A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid beta. Sci Transl Med 4:147ra11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Kaufer D, Gandy S (2009) APOE {epsilon}4 and bapineuzumab: infusing pharmacogenomics into Alzheimer disease therapeutics. Neurology 73:2052–2053. [DOI] [PubMed] [Google Scholar]
- 42. Kerchner GA, Boxer AL (2010) Bapineuzumab. Expert Opin Biol Ther 10:1121–1130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Kida S, Steart PV, Zhang ET, Weller RO (1993) Perivascular cells act as scavengers in the cerebral perivascular spaces and remain distinct from pericytes, microglia and macrophages. Acta Neuropathol (Berl) 85:646–652. [DOI] [PubMed] [Google Scholar]
- 44. Kivipelto M, Solomon A (2008) Alzheimer's disease—the ways of prevention. J Nutr Health Aging 12:89S–94S. [DOI] [PubMed] [Google Scholar]
- 45. Krueger M, Bechmann I (2010) CNS pericytes: concepts, misconceptions, and a way out. Glia 58:1–10. [DOI] [PubMed] [Google Scholar]
- 46. Li WH, Tanimura M, Luo CC, Datta S, Chan L (1988) The apolipoprotein multigene family: biosynthesis, structure, structure‐function relationships, and evolution. J Lipid Res 29:245–271. [PubMed] [Google Scholar]
- 47. Lifshitz V, Weiss R, Levy H, Frenkel D (2013) Scavenger receptor A deficiency accelerates cerebrovascular amyloidosis in an animal model. J Mol Neurosci 50:198–203. [DOI] [PubMed] [Google Scholar]
- 48. Lue LF, Kuo YM, Roher AE, Brachova L, Shen Y, Sue L et al (1999) Soluble amyloid beta peptide concentration as a predictor of synaptic change in Alzheimer's disease. Am J Pathol 155:853–862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Masliah E, Hansen L, Adame A, Crews L, Bard F, Lee C et al (2005) Abeta vaccination effects on plaque pathology in the absence of encephalitis in Alzheimer disease. Neurology 64:129–131. [DOI] [PubMed] [Google Scholar]
- 50. Mead S, James‐Galton S, Revesz T, Bala Doshi R, Harwood G, Lee Pan E et al (2000) Familial British dementia with amyloid angiopathy. Early clinical, neuropsychological and imaging findings. Brain 123:975–991. [DOI] [PubMed] [Google Scholar]
- 51. Meilandt WJ, Cisse M, Ho K, Wu T, Esposito LA, Scearce‐Levie K et al (2009) Neprilysin overexpression inhibits plaque formation but fails to reduce pathogenic Abeta oligomers and associated cognitive deficits in human amyloid precursor protein transgenic mice. J Neurosci 29:1977–1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Michaud JP, Bellavance MA, Prefontaine P, Rivest S (2013) Real‐time in vivo imaging reveals the ability of monocytes to clear vascular amyloid beta. Cell Rep 5:646–653. [DOI] [PubMed] [Google Scholar]
- 53. Mildner A, Schmidt H, Nitsche M, Merkler D, Hanisch UK, Mack M et al (2007) Microglia in the adult brain arise from Ly‐6ChiCCR2+ monocytes only under defined host conditions. Nat Neurosci 10:1544–1553. [DOI] [PubMed] [Google Scholar]
- 54. Miners JS, Baig S, Palmer J, Palmer LE, Kehoe PG, Love S (2008) Abeta‐degrading enzymes in Alzheimer's disease. Brain Pathol 18:240–252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Miyakawa T, Shimoji A, Kuramoto R, Higuchi Y (1982) The relationship between senile plaques and cerebral blood vessels in Alzheimer's disease and senile dementia. Virchows Arch (Cell Pathol) 40:121–129. [DOI] [PubMed] [Google Scholar]
- 56. Nicoll JA, Wilkinson D, Holmes C, Steart P, Markham H, Weller RO (2003) Neuropathology of human Alzheimer disease after immunization with amyloid‐beta peptide: a case report. Nat Med 9:448–452. [DOI] [PubMed] [Google Scholar]
- 57. Nicoll JA, Yamada M, Frackowiak J, Mazur‐Kolecka B, Weller RO (2004) Cerebral amyloid angiopathy plays a direct role in the pathogenesis of Alzheimer's disease. Pro‐CAA position statement. Neurobiol Aging 25:589–597. [DOI] [PubMed] [Google Scholar]
- 58. Park L, Zhou J, Zhou P, Pistick R, El Jamal S, Younkin L et al (2013) Innate immunity receptor CD36 promotes cerebral amyloid angiopathy. Proc Natl Acad Sci USA 110:3089–3094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Patton RL, Kalback WM, Esh CL, Kokjohn TA, Van Vickle GD, Luehrs DC et al (2006) Amyloid‐beta peptide remnants in AN‐1792‐immunized Alzheimer's disease patients: a biochemical analysis. Am J Pathol 169:1048–1063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Premkumar DR, Cohen DL, Hedera P, Friedland RP, Kalaria RN (1996) Apolipoprotein E‐epsilon4 alleles in cerebral amyloid angiopathy and cerebrovascular pathology associated with Alzheimer's disease. Am J Pathol 148:2083–2095. [PMC free article] [PubMed] [Google Scholar]
- 61. Preston SD, Steart PV, Wilkinson A, Nicoll JA, Weller RO (2003) Capillary and arterial cerebral amyloid angiopathy in Alzheimer's disease: defining the perivascular route for the elimination of amyloid beta from the human brain. Neuropathol Appl Neurobiol 29:106–117. [DOI] [PubMed] [Google Scholar]
- 62. Prior R, Wihl G, Urmoneit B (2000) Apolipoprotein E, smooth muscle cells and the pathogenesis of cerebral amyloid angiopathy: the potential role of impaired cerebrovascular A beta clearance. Ann N Y Acad Sci 903:180–186. [DOI] [PubMed] [Google Scholar]
- 63. Roher AE, Garami Z, Tyas SL, Maarouf CL, Kokjohn TA, Belohlavek M et al (2011) Transcranial Doppler ultrasound blood flow velocity and pulsatility index as systemic indicators for Alzheimer's disease. Alzheimer's Dement 7:445–455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Roher AE, Kuo YM, Esh C, Knebel C, Weiss N, Kalback W et al (2003) Cortical and leptomeningeal cerebrovascular amyloid and white matter pathology in Alzheimer's disease. Mol Med 9:112–122. [PMC free article] [PubMed] [Google Scholar]
- 65. Schenk D, Barbour R, Dunn W, Gordon G, Grajeda H, Guido T et al (1999) Immunization with amyloid‐beta attenuates Alzheimer‐disease‐like pathology in the PDAPP mouse. Nature 400:173–177. [DOI] [PubMed] [Google Scholar]
- 66. Schittny JC, Yurchenco PD (1989) Basement membranes: molecular organization and function in development and disease. Curr Opin Cell Biol 1:983–988. [DOI] [PubMed] [Google Scholar]
- 67. Schley D, Carare‐Nnadi R, Please CP, Perry VH, Weller RO (2006) Mechanisms to explain the reverse perivascular transport of solutes out of the brain. J Theor Biol 238:962–974. [DOI] [PubMed] [Google Scholar]
- 68. Shibata M, Yamada S, Kumar SR, Calero M, Bading J, Frangione B et al (2000) Clearance of Alzheimer's amyloid‐ss(1‐40) peptide from brain by LDL receptor‐related protein‐1 at the blood‐brain barrier. J Clin Invest 106:1489–1499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Shimizu H, Ghazizadeh M, Sato S, Oguro T, Kawanami O (2009) Interaction between beta‐amyloid protein and heparan sulfate proteoglycans from the cerebral capillary basement membrane in Alzheimer's disease. J Clin Neurosci 16:277–282. [DOI] [PubMed] [Google Scholar]
- 70. Shirotani K, Tsubuki S, Iwata N, Takaki Y, Harigaya W, Maruyama K et al (2001) Neprilysin degrades both amyloid beta peptides 1‐40 and 1‐42 most rapidly and efficiently among thiorphan‐ and phosphoramidon‐sensitive endopeptidases 98. J Biol Chem 276:21895–21901. [DOI] [PubMed] [Google Scholar]
- 71. Sixt M, Engelhardt B, Pausch F, Hallmann R, Wendler O, Sorokin LM (2001) Endothelial cell laminin isoforms, laminins 8 and 10, play decisive roles in T cell recruitment across the blood‐brain barrier in experimental autoimmune encephalomyelitis. J Cell Biol 153:933–946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Szentistvanyi I, Patlak CS, Ellis RA, Cserr HF (1984) Drainage of interstitial fluid from different regions of rat brain 32. Am J Physiol 246(6 Pt 2):F835–F844. [DOI] [PubMed] [Google Scholar]
- 73. Teeling JL, Carare RO, Glennie MJ, Perry VH (2012) Intracerebral immune complex formation induces inflammation in the brain that depends on Fc receptor interaction. Acta Neuropathol 124:479–490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Thal DR, Larionov S, Abramowski D, Wiederhold KH, Van Dooren T, Yamaguchi H et al (2007) Occurrence and co‐localization of amyloid beta‐protein and apolipoprotein E in perivascular drainage channels of wild‐type and APP‐transgenic mice. Neurobiol Aging 28:1221–1230. [DOI] [PubMed] [Google Scholar]
- 75. Thanopoulou K, Fragkouli A, Stylianopoulou F, Georgopoulos S (2010) Scavenger receptor class B type I (SR‐BI) regulates perivascular macrophages and modifies amyloid pathology in an Alzheimer mouse model. Proc Natl Acad Sci USA 107:20816–20821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Toft‐Hansen H, Buist R, Sun XJ, Schellenberg A, Peeling J, Owens T (2006) Metalloproteinases control brain inflammation induced by pertussis toxin in mice overexpressing the chemokine CCL2 in the central nervous system. J Immunol 177:7242–7249. [DOI] [PubMed] [Google Scholar]
- 77. Utter S, Tamboli IY, Walter J, Upadhaya AR, Birkenmeier G, Pietrzik CU et al (2008) Cerebral small vessel disease‐induced apolipoprotein E leakage is associated with Alzheimer disease and the accumulation of amyloid beta‐protein in perivascular astrocytes. J Neuropathol Exp Neurol 67:842–856. [DOI] [PubMed] [Google Scholar]
- 78. Vitek MP, Brown CM, Colton CA (2009) APOE genotype‐specific differences in the innate immune response. Neurobiol Aging 30:1350–1360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Wang J, Dickson DW, Trojanowski JQ, Lee VM (1999) The levels of soluble versus insoluble brain Abeta distinguish Alzheimer's disease from normal and pathologic aging 1. Exp Neurol 158:328–337. [DOI] [PubMed] [Google Scholar]
- 80. Wang P, Olbricht WL (2011) Fluid mechanics in the perivascular space. J Theor Biol 274:52–57. [DOI] [PubMed] [Google Scholar]
- 81. Weller RO (2005) Microscopic morphology and histology of the human meninges. Morphologie 89:22–34. [DOI] [PubMed] [Google Scholar]
- 82. Weller RO, Boche D, Nicoll JA (2009) Microvasculature changes and cerebral amyloid angiopathy in Alzheimer's disease and their potential impact on therapy. Acta Neuropathol 118:87–102. [DOI] [PubMed] [Google Scholar]
- 83. Weller RO, Djuanda E, Yow HY, Carare RO (2009) Lymphatic drainage of the brain and the pathophysiology of neurological disease. Acta Neuropathol 117:1–14. [DOI] [PubMed] [Google Scholar]
- 84. Weller RO, Engelhardt B, Phillips MJ (1996) Lymphocyte targeting of the central nervous system: a review of afferent and efferent CNS‐immune pathways. Brain Pathol 6:275–288. [DOI] [PubMed] [Google Scholar]
- 85. Weller RO, Massey A, Newman TA, Hutchings M, Kuo YM, Roher AE (1998) Cerebral amyloid angiopathy: amyloid beta accumulates in putative interstitial fluid drainage pathways in Alzheimer's disease. Am J Pathol 153:725–733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Weller RO, Subash M, Preston SD, Mazanti I, Carare RO (2008) Perivascular drainage of amyloid‐beta peptides from the brain and its failure in cerebral amyloid angiopathy and Alzheimer's disease. Brain Pathol 18:253–266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Wilcock DM, Colton CA (2009) Immunotherapy, vascular pathology, and microhemorrhages in transgenic mice. CNS Neurol Disord Drug Targets 8:50–64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Wilcock DM, Rojiani A, Rosenthal A, Levkowitz G, Subbarao S, Alamed J et al (2004) Passive amyloid immunotherapy clears amyloid and transiently activates microglia in a transgenic mouse model of amyloid deposition 1. J Neurosci 24:6144–6151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Wyss‐Coray T, Lin C, von Euw D, Masliah E, Mucke L, Lacombe P (2000) Alzheimer's disease‐like cerebrovascular pathology in transforming growth factor‐beta 1 transgenic mice and functional metabolic correlates 1. Ann NY Acad Sci 903:317–323. [DOI] [PubMed] [Google Scholar]
- 90. Wyss‐Coray T, Lin C, Sanan DA, Mucke L, Masliah E (2000) Chronic overproduction of transforming growth factor‐beta1 by astrocytes promotes Alzheimer's disease‐like microvascular degeneration in transgenic mice. Am J Pathol 156:139–150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Yamada M, Naiki H (2012) Cerebral amyloid angiopathy. Prog Mol Biol Transl Sci 107:41–78. [DOI] [PubMed] [Google Scholar]
- 92. Weller RO, Yow HY, Preston SD, Mazanti I, Nicoll JA (2002) Cerebrovascular disease is a major factor in the failure of elimination of Abeta from the aging human brain: implications for therapy of Alzheimer's disease. Ann N Y Acad Sci 977:162–168. [DOI] [PubMed] [Google Scholar]
- 93. Yurchenco PD, Cheng YS, Schittny JC (1990) Heparin modulation of laminin polymerization. J Biol Chem 265:3981–3991. [PubMed] [Google Scholar]
- 94. Yurchenco PD, Schittny JC (1990) Molecular architecture of basement membranes. FASEB J 4:1577–1590. [DOI] [PubMed] [Google Scholar]
- 95. Zhang ET, Inman CB, Weller RO (1990) Interrelationships of the pia mater and the perivascular (Virchow‐Robin) spaces in the human cerebrum. J Anat 170:111–123. [PMC free article] [PubMed] [Google Scholar]
- 96. Zhao L, Lin S, Bales KR, Gelfanova V, Koger D, Delong C et al (2009) Macrophage‐mediated degradation of beta‐amyloid via an apolipoprotein E isoform‐dependent mechanism. J Neurosci 29:3603–3612. [DOI] [PMC free article] [PubMed] [Google Scholar]