

Abstract
In the central nervous system, nuclear factor erythroid-2-related factor 2 (Nrf2) protects neurons from oxidant injury, thereby ameliorating neurodegeneration. We explored the key circular RNAs (circRNAs) and long non-coding RNAs (lncRNAs) involved in Nrf2-induced neuroprotection. We used microarrays to examine the circRNAs (DEcircRNAs), lncRNAs (DElncRNAs) and mRNAs (DEmRNAs) differentially expressed between Nrf2 (+/+) and Nrf2 (-/-) mice and identified DEcircRNA/DElncRNA-miRNA-DEmRNA interaction networks. In total, 197 DEcircRNAs, 685 DElncRNAs and 356 DEmRNAs were identified in prefrontal cortical tissues from Nrf2 (-/-) mice. The expression patterns of selected DEcircRNAs (except for mmu_circ_0003404) and DElncRNAs in qRT-PCR analyses were generally consistent with the microarray analysis results. Functional annotation of the DEmRNAs in the DEcircRNA/DElncRNA-miRNA-DEmRNA networks indicated that five non-coding RNAs (mmu_circ_0000233, ENSMUST00000204847, NONMMUT024778, NONMMUT132160 and NONMMUT132168) may contribute to Nrf2 activity, with the help of mmu_circ_0015035 and NONMMUT127961. The results also revealed that four non-coding RNAs (cicRNA.20127, mmu_circ_0012936, ENSMUST00000194077 and NONMMUT109267) may influence glutathione metabolism. Additionally, 44 DEcircRNAs and 7 DElncRNAs were found to possess coding potential. These findings provide clues to the molecular pathways through which Nrf2 protects neurons.
Keywords: Nrf2, circular RNA, long non-coding RNA
INTRODUCTION
Nuclear factor erythroid-2-related factor 2 (Nrf2/NFE2L2) is a pleiotropic transcription factor, and studies in mouse models have indicated that Nrf2 influences the susceptibility to diseases ranging from neurodegenerative disorders to endotoxin-induced septic shock [1–3]. Nrf2 belongs to a subset of basic leucine-zipper genes sharing a conserved CNC (cap ‘n’ collar) structural domain [4]. The mammalian CNC family includes four closely related proteins (p45-NFE2, Nrf1, Nrf2 and Nrf3) and two distantly related proteins (Bach1 and Bach2) that function as heterodimeric transcription factors by pairing with other basic leucine-zipper proteins, including the small-Mafs [4]. In the central nervous system, Nrf2 protects against oxidative stress, improves mitochondrial function, inhibits the unfolded protein responses, promotes autophagy, etc [5–10].
Non-coding RNAs (ncRNAs), including circular RNAs (circRNAs), long non-coding RNAs (lncRNAs) and microRNAs (miRNAs), are the dominant products of eukaryotic transcription, and function directly at the RNA level [11–13]. NcRNAs not only are involved in cell proliferation, apoptosis, differentiation, metabolism and other physiological processes, but also contribute to the pathogenesis of diseases [14–18]. LncRNAs are endogenous linear ncRNAs greater than 200 nucleotides in length. CircRNAs lack 5' caps and 3' tails, and thus are resistant to degradation by RNA exonuclease and are much more stable than linear isoforms [19]. CircRNAs and lncRNAs may share certain mechanisms of target gene regulation, including binding to miRNAs as competitive endogenous RNAs (ceRNAs) [20, 21], altering gene transcription, binding to proteins and encoding peptides/proteins [22].
In previous studies, we examined the effects of Nrf2-related circRNAs and lncRNAs on the brain by detecting the altered expression of circRNAs, lncRNAs and mRNAs in different brain areas in Nrf2-null (Nrf2 (-/-)) mice, including substantia nigra, corpus striatum and hippocampus [23–25]. The prefrontal cortex has often been called the command and control center of the brain, and is mainly responsible for cognitive control, especially the ability to orchestrate thought and action in accordance with internal goals [26]. Thus, in this study, we selected the prefrontal cortex to continue this series of studies in Nrf2 (-/-) mice. First, we performed microarray analyses to identify differentially expressed circRNAs (DEcircRNAs), lncRNAs (DElncRNAs) and mRNAs (DEmRNAs) between prefrontal cortex tissues from Nrf2 (-/-) and Nrf2 (+/+) mice. Then, we constructed DEcircRNA/DElncRNA-miRNA-DEmRNA networks and performed Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses to annotate the functions of the DEmRNAs in the networks. Additionally, we evaluated the coding potential of the DEcircRNAs/DElncRNAs, the expression correlations between the DEcircRNAs/DElncRNAs and their host mRNAs, and the intersections between the DEcircRNAs/DElncRNAs/DEmRNAs in different brain areas. Our study has provided clues into the molecular pathways whereby Nrf2 protects neurons.
RESULTS
DEcircRNAs, DElncRNAs and DEmRNAs between the prefrontal cortexes of Nrf2 (-/-) and Nrf2 (+/+) mice
After identifying the genotypes of the mice (Supplementary Figure 1), we performed microarray analyses to detect DEcircRNAs, DElncRNAs and DEmRNAs between the prefrontal cortex of Nrf2 (-/-) and Nrf2 (+/+) mice. The raw data have been deposited in the Gene Expression Omnibus database (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE162315). In total, 197 DEcircRNAs with p-value < 0.05 and fold-changes (FCs) > 1.5 were detected in the prefrontal cortex, of which 103 up-regulated and 94 down-regulated in Nrf2 (-/-) mice compared with Nrf2 (+/+) mice. Among them, cicRNA.17816 and cicRNA.24872 were the most significantly up- and down-regulated genes, respectively (Table 1). Additionally, 685 DElncRNAs (281 up-regulated and 404 down-regulated in Nrf2 (-/-) mice) were identified, of which NONMMUT132168 and ENSMUST00000195276 were the most significantly up- and down-regulated genes, respectively (Table 2). Moreover, 356 DEmRNAs were detected, of which 87 up- and 269 down-regulated in Nrf2 (-/-) mice. Scatter plots (Figure 1A–1C), volcano diagrams (Figure 1D–1F) and hierarchical clustering analyses (Figure 1G–1I) were used to depict the DEcircRNAs, DElncRNAs and DEmRNAs between the prefrontal cortex of Nrf2 (-/-) and Nrf2 (+/+) mice.
Table 1. Top 10 up- and down-regulated DEcircRNAs in the prefrontal cortex of Nrf2 (-/-) mice.
| circRNA | chrom | Foldchange | Regulation |
| cicRNA.17816 | chr1 | 2.900872 | up |
| cicRNA.2059 | chr17 | 2.751136 | up |
| cicRNA.22457 | chr4 | 2.491297 | up |
| cicRNA.25732 | chr2 | 2.194124 | up |
| cicRNA.27857 | chr19 | 2.177916 | up |
| cicRNA.10101 | chr12 | 2.130815 | up |
| mmu_circ_0000233 | chr11 | 2.049909 | up |
| cicRNA.14293 | chr10 | 2.034235 | up |
| cicRNA.12176 | chr11 | 2.023146 | up |
| cicRNA.24048 | chr3 | 2.015869 | up |
| cicRNA.24872 | chr2 | 36.804170 | down |
| cicRNA.14175 | chr10 | 5.765533 | down |
| cicRNA.21621 | chr5 | 4.358284 | down |
| cicRNA.20127 | chr8 | 4.081641 | down |
| mmu_circ_0002377 | chr10 | 3.477857 | down |
| cicRNA.16195 | chr1 | 3.138005 | down |
| cicRNA.9167 | chr13 | 2.993984 | down |
| cicRNA.2029 | chr17 | 2.849510 | down |
| cicRNA.13394 | chr11 | 2.782886 | down |
| cicRNA.6318 | chr15 | 2.727061 | down |
DEcircRNAs, differentially expressed circRNAs.
Table 2. Top 10 up- and down-regulated DElncRNAs in the prefrontal cortex of Nrf2 (-/-) mice.
| lncRNA | chrom | Foldchange | Regulation |
| NONMMUT132168 | chr9 | 6.486807 | up |
| NR_038039 | chr10 | 6.417579 | up |
| NONMMUT127961 | chr7 | 5.724120 | up |
| NONMMUT132160 | chr9 | 5.327114 | up |
| NONMMUT077500 | chr1 | 4.428169 | up |
| NR_045161 | chr13 | 3.619127 | up |
| NONMMUT122421 | chr6 | 3.489428 | up |
| NONMMUT132149 | chr9 | 3.395637 | up |
| NONMMUT125225 | chr7 | 3.084584 | up |
| NONMMUT083894 | chr11 | 3.048123 | up |
| ENSMUST00000195276 | chr4 | 13.210466 | down |
| NONMMUT118467 | chr5 | 11.100098 | down |
| NONMMUT114173 | chr3 | 10.866240 | down |
| NONMMUT132224 | chr9 | 10.746878 | down |
| ENSMUST00000194077 | chr3 | 10.500292 | down |
| NONMMUT105493 | chr19 | 9.502475 | down |
| NONMMUT000100 | chr1 | 8.942628 | down |
| NONMMUT124473 | chr6 | 8.569627 | down |
| NONMMUT106900 | chr2 | 8.519742 | down |
| NONMMUT076758 | chr1 | 8.509335 | down |
DElncRNAs, differentially expressed lncRNAs.
Figure 1.

Expression profiles of circRNAs, lncRNAs and mRNAs in prefrontal cortex tissues from Nrf2 (-/-) and Nrf2 (+/+) mice. (A–C) Scatter plots showing the expression variation of circRNAs (A), lncRNAs (B) and mRNAs (C) between Nrf2 (-/-) and Nrf2 (+/+) prefrontal cortex tissues. The values of the X and Y axes in the scatter plots are the normalized signal values of the samples (log2-scaled). The red (up-regulated) and blue (down-regulated) points represent the DEcircRNAs (A), DElncRNAs (B) and DEmRNAs (C) with more than FCs > 1.5 between Nrf2 (-/-) and Nrf2 (+/+) prefrontal cortex tissues. (D–F) Volcano plots showing the differential expression of circRNAs (D), lncRNAs (E) and mRNAs (F) between Nrf2 (-/-) and Nrf2 (+/+) prefrontal cortex tissues. The vertical lines correspond to 1.5-fold upregulation and downregulation, and the horizontal line represents a p-value of 0.05. The red (up-regulated) and blue (down-regulated) points represent the DEcircRNAs (D), DElncRNAs (E) and DEmRNAs (F) with statistical significance. g1: group 1, refers to the Nrf2 (+/+) group. g2: group 2, refers to the Nrf2 (-/-) group. (G–I) Hierarchical clustering analyses were performed to depict the DEcircRANs (G), DElncRNAs (H) and DEmRNAs (I) in Nrf2 (-/-) prefrontal cortex compared with Nrf2 (+/+) prefrontal cortex tissues. The clustering analysis was used to group samples based on their expression values so that the relationships among samples could be predicted. ‘Red’ denotes high relative expression, and ‘green’ denotes low relative expression.
Quantitative real-time polymerase chain reaction (qRT-PCR) validation of DEcircRNA and DElncRNA expression
Considering that DEcircRNAs and DElncRNAs with higher FCs and lower p-values were more likely to be regulated by Nrf2, and that higher expression may enable more accurate microarray detection, we selected DEcircRNAs and DElncRNAs with relatively high FCs, high expression values and low p-value to validate the accuracy of the microarray data using qRT-PCR. We used different screening criteria to screen different RNAs. For DElncRNAs, those with p-values < 0.015, FCs > 2.8 or < 0.15, and Standardized Expression Levels (SELs) > 1 were selected. For DEcircRNAs included in “circBase”, those with p-values < 0.05, FCs >1.7 or < 0.55, and SELs > 1 were selected. For DEcircRNAs not included in “circBase”, those with p-values < 0.01, FCs >2.5 or < 0.25, and SELs > 1 were selected.
Subsequently, qRT-PCR was used to screen primers, and only six DEcircRNAs and eight DElncRNAs were found to have suitable primers. Thus, three up-regulated DEcircRNAs (mmu_circ_0000233, mmu_circ_0015035 and mmu_circ_0003404), three down-regulated DEcircRNAs (mmu_circ_0012936, mmu_circ_0008393 and cicRNA.20127), six up-regulated DElncRNAs (NONMMUT132168, NONMMUT127961, NONMMUT132160, NR_045161, ENSMUST00000204847 and NONMMUT024778) and two down-regulated DElncRNAs (NONMMUT109267 and ENSMUST00000194077) with relatively high FCs, high expression values and low p-values were selected to validate the accuracy of the microarray data using qRT-PCR. As shown in Figure 2, the qRT-PCR results for all the DElncRNAs and DEcircRNAs except for mmu_circ_0003404 were consistent with the microarray results.
Figure 2.

Validation of the DEcircRNAs and DElncRNAs. The levels of six DEcircRNAs and eight DElncRNAs were validated in prefrontal cortex samples from Nrf2 (-/-) and Nrf2 (+/+) mice using qRT-PCR and normalized to the internal reference gene (β-actin). (A) The qRT-PCR results for five of the verified circRNAs (all but mmu_circ_0003404) were consistent with the microarray results. (B) The qRT-PCR results for all eight verified lncRNAs were consistent with the microarray results. All experiments were replicated three times. The deep red column indicates the FCs of the DEcircRNAs/DElncRNAs determined through microarray analyses; the blue column indicates the expression FCs of the DEcircRNAs/DElncRNAs determined through qRT-PCR experiments. The presented values are the means ± standard deviations. Values > 1 indicate up-regulated DEcircRNAs/DElncRNAs, and values < 1 indicate down-regulated DEcircRNAs/DElncRNAs. *p < 0.05, **p < 0.01.
Construction of the DEcircRNA/DElncRNA-miRNA-DEmRNA crosstalk networks
Twelve of the ncRNAs validated by qRT-PCR (all but mmu_circ_0003404 and NR_045161) were found to be ceRNAs for miRNA targets. When we constructed DEcircRNA/DElncRNA-miRNA-DEmRNA crosstalk networks (Figure 3), we noted that mmu_circ_0000233, ENSMUST00000204847, NONMMUT024778, NONMMUT132160 and NONMMUT132168 shared miRNA response elements (MREs) with four up-regulated DEmRNAs (HSD11B1, P2RX3, TCF7L1 and CHP2) (Figures 3, 4C); mmu_circ_0015035 and NONMMUT127961 also shared MREs with HSD11B1, P2RX3 and TCF7L1 (Figures 3, 4C); and cicRNA.20127, mmu_circ_0012936, ENSMUST00000194077 and NONMMUT109267 shared miRNAs with three down-regulated DEmRNAs (GSTM1, GSTM2 and MGST1) (Figures 3, 4D).
Figure 3.

DEcircRNA/DElncRNA-miRNA-DEmRNA crosstalk network. Through linear regression model analysis and seed sequence matching method based on gene expression, five DEcircRNAs and seven DElncRNAs were established as a regulatory network of miRNA sponge adsorbents. (A–E) The network of five verified circRNAs (mmu_circ_0000233 (A), mmu_circ_0015035 (B), mmu_circ_0012936 (C), mmu_circ_0008393 (D) and cicRNA.20127 (E)). (F–L) The network of seven verified lncRNAs (NONMMUT132168 (F), NONMMUT127961 (G), NONMMUT132160 (H), ENSMUST00000204847 (I), NONMMUT024778 (J), NONMMUT109267 (K) and ENSMUST00000194077 (L)). Red rhombi: up-regulated DEcircRNAs. Green rhombi: down-regulated DEcircRNAs. Red squares: up-regulated DElncRNAs. Green squares: down-regulated DElncRNAs. Gray V: miRNAs. Red circles: up-regulated DEmRNAs. Green circles: down-regulated DEmRNAs.
Figure 4.

Predicted functions of the DEcircRNAs/DElncRNAs involved in the DEcircRNA/DElncRNA-miRNA-DEmRNA network. (A, B) The top 30 most significantly enriched GO (A) and KEGG (B) pathways of the DEmRNAs that were targets of the five DEcircRNAs (mmu_circ_0000233, mmu_circ_0015035, mmu_circ_0012936, mmu_circ_0008393 and cicRNA.20127) and seven DElncRNAs (NONMMUT132168, NONMMUT127961, NONMMUT132160, ENSMUST00000204847, NONMMUT024778, NONMMUT109267 and ENSMUST00000194077). The X-axis displays the enrich factor in the GO and KEGG pathways and the Y-axis displays the GO and KEGG pathways. The color scale depicts the p-values. The sizes of the nodes represent the counts of genes enriched in the GO and KEGG pathways. (C, D) Diagram of the various functions of the DEcircRNAs/DElncRNAs.
Functional annotation of DEmRNAs that were targets of DEcircRNAs and DElncRNAs
GO and KEGG pathway analyses were employed to annotate the functions of the DEmRNAs in the DEcircRNA/DElncRNA-miRNA-DEmRNA network. In the GO analysis, the Rough endoplasmic reticulum (GO: 0005791, p = 9.090e-05) and Lymphocyte chemotaxis (GO: 0048247, p = 4.203e-06) were the most significant cellular component and biological process, respectively (Figure 4A). According to the KEGG pathway analysis, the top 30 most significantly enriched pathways for the DEmRNAs included Glutathione metabolism (KEGG: mmu00480, p = 4.435e-03), Amyotrophic lateral sclerosis (ALS) (KEGG: mmu05014, p = 5.047e-03), Apoptosis (KEGG: mmu04210, p = 2.211e-02), Wnt signaling pathway (KEGG: mmu04310, p = 3.932e-02) and Calcium signaling pathway (KEGG: mmu04020, p = 6.673e-02) (Figure 4B).
We found that some DEmRNAs (HSD11B1, P2RX3, TCF7L1 and CHP2) that shared MREs with mmu_circ_0000233, mmu_circ_0015035, ENSMUST00000204847, NONMMUT024778, NONMMUT132160, NONMMUT132168 or NONMMUT127961 were enriched in the Rough endoplasmic reticulum (GO: 0005791), Amyotrophic lateral sclerosis (ALS) (KEGG: mmu05014), Apoptosis (KEGG: mmu04210), Wnt signaling pathway (KEGG: mmu04310) or Calcium signaling pathway (KEGG: mmu04020). These findings indicated that circRNAs and lncRNAs both have wide-ranging physiological functions and may co-regulate certain target genes (Figure 4C). We also noted that GSTM1 (sharing miRNAs with mmu_circ_0012936 and NONMMUT109267), GSTM2 (sharing miRNAs with cicRNA.20127 and ENSMUST00000194077) and MGST1 (sharing miRNAs with mmu_circ_0012936 and ENSMUST00000194077) were enriched in the pathway of Glutathione metabolism (KEGG: mmu00480) (Figure 4D).
Coding potential of DEcircRNAs and DElncRNAs
Among the 197 DEcircRNAs and 685 DElncRNAs, we found that 44 DEcircRNAs and 7 DElncRNAs had scores > 0 in the Coding-Non-Coding Index [27], suggesting that they may have coding potential (Supplementary Table 1).
Expression correlation analysis between DEcircRNAs/DElncRNAs and their host mRNAs
We next evaluated whether the levels of the DEcircRNAs/DElncRNAs correlated with those of their host mRNAs, in order to determine whether the DEcircRNAs/DElncRNAs altered the expression of their host genes. The levels of 38% of the DEcircRNAs (74 of 197) correlated with the levels of their host mRNAs, with 70% correlating positively and 30% correlating negatively (Figure 5A and Supplementary Table 2). After we removed 527 intergenic DElncRNAs without maternal genes, we found that the levels of 35% of the DElncRNAs (55 of 158) correlated with the levels of their host mRNAs, with 67% correlating positively and 33% correlating negatively (Figure 5B and Supplementary Table 2).
Figure 5.
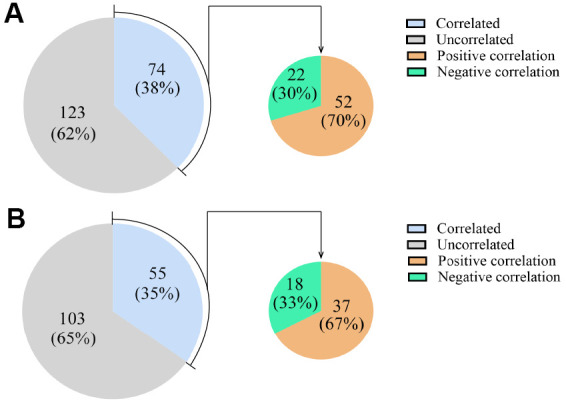
Expression correlations between DEcircRNAs/DElncRNAs and their host mRNAs. (A) The expression correlations between DEcircRNAs and their host mRNAs. (B) The expression correlations between DElncRNAs and their host mRNAs. The blue area represents the DEcircRNAs/DElncRNAs that correlated with their host mRNAs as a percentage of all the DEcircRNAs/DElncRNAs. The gray area represents the remaining DEcircRNAs/DElncRNAs. The orange area represents the DEcircRNAs/DElncRNAs that correlated positively with their host mRNAs, and the green area represents the DEcircRNAs/DElncRNAs that correlated negatively with their host mRNAs, as percentages of the DEcircRNAs/DElncRNAs that correlated with their host mRNAs.
Comparison of DEcircRNAs, DElncRNAs and DEmRNAs among the prefrontal cortex, substantia nigra, corpus striatum and hippocampus
Next, we determined the shared DEcircRNAs, DElncRNAs and DEmRNAs among the prefrontal cortex, substantia nigra, corpus striatum and hippocampus, in order to identify common regulatory mechanisms of Nrf2 in these areas. Surprisingly, few circRNAs/lncRNAs were consistently regulated by Nrf2 in all four brain regions. Based on their differential expression between Nrf2 (-/-) and Nrf2 (+/+) mice, no circRNAs/lncRNAs were consistently regulated by Nrf2 in the prefrontal cortex and substantia nigra; only one down-regulated ncRNA (mmu_circ_0010156) was consistently regulated by Nrf2 in the prefrontal cortex, corpus striatum and hippocampus; only one up-regulated ncRNA (ENSMUST00000144818) was consistently regulated by Nrf2 in the prefrontal cortex and corpus striatum; and only two up-regulated ncRNAs (cicRNA.4014 and NR_027831) and one down-regulated ncRNA (ENSMUST00000145790) were consistently regulated by Nrf2 in the prefrontal cortex and hippocampus (Figure 6A, 6B).
Figure 6.

The intersections of DEcircRNAs, DElncRNAs and DEmRNAs in the prefrontal cortex (cor), substantia nigra (SN), corpus striatum (CPU) and hippocampus (hip). (A–D) Venn diagrams of up-regulated DEcircRNAs/DElncRNAs (A), down-regulated DEcircRNAs/DElncRNAs (B), up-regulated DEmRNAs (C) and down-regulated DEmRNAs (D) were showed respectively.
While it was surprising that there were so few intersecting DEcircRNAs/DElncRNAs in different brain areas, it was even more interesting that there were few intersecting DEmRNAs. Based on their differential expression between Nrf2 (-/-) and Nrf2 (+/+) mice, only one up-regulated mRNA (THNSL2) and one down-regulated mRNA (MYCT1) were consistently regulated by Nrf2 in the prefrontal cortex and substantia nigra; only one up-regulated mRNA (RBM45) and one down-regulated mRNA (NFE2L2) were consistently regulated by Nrf2 in the prefrontal cortex, corpus striatum and hippocampus; only two up-regulated mRNAs (PRSS56 and SLC29A4), and eleven down-regulated mRNAs (GBP11, FAS, GSTM3, LYZ1, CYBB, CHRNB1, GM14548, GSTM2, C3, PLAC8, and DSG1C) were consistently regulated by Nrf2 in the prefrontal cortex and corpus striatum; and only two up-regulated mRNAs (TEX11 and FAM181A), and twelve down-regulated mRNAs (NAPSA, CYCT, AOX1, SFMBT2, OMD, MRC1, FOXC2, MS4A6B, RIDA, CD163, GSTM1, and MS4A6D) were consistently regulated by Nrf2 in the prefrontal cortex and hippocampus (Figure 6C, 6D). These results indicated that Nrf2 may not have the same function in different brain areas.
DISCUSSION
The prefrontal cortex is responsible for cognition, emotion, pain and behavior management, and its function is disrupted in various neurodegenerative diseases [28, 29]. There is abundant evidence that insufficient Nrf2 signaling is a major cause of nervous system disorders, particularly neurodegenerative diseases [8, 30–32]. We hypothesized that Nrf2 could exert its neuroprotective effects by altering the expression of circRNAs and lncRNAs. Although circRNAs and lncRNAs differ significantly in shape and size, both are expressed in tissue- and spatiotemporal-specific patterns and regulate their target genes by similar mechanisms [33–36]. Since the ceRNA hypothesis was proposed [37], both circRNAs and lncRNAs have been reported to function as miRNA sponges that alter the stability or translation of mRNAs [21, 38]. Thus, we used microarray and bioinformatics analyses to detect DEcircRNAs and DElncRNAs between the prefrontal cortex tissues of Nrf2 (-/-) and Nrf2 (+/+) mice.
In this study, five DEcircRNAs (cicRNA.20127, mmu_circ_0000233, mmu_circ_0008393, mmu_circ_0012936 and mmu_circ_0015035) and seven DElncRNAs (ENSMUST00000194077, ENSMUST00000204847, NONMMUT024778, NONMMUT109267, NONMMUT127961, NONMMUT132160 and NONMMUT132168) were verified by qRT-PCR and used to construct DEcircRNA/DElncRNA-miRNA-DEmRNA crosstalk networks. Then GO and KEGG pathway analyses were employed to predict their functions. Five of these ncRNAs (mmu_circ_0000233, ENSMUST00000204847, NONMMUT024778, NONMMUT132160 and NONMMUT132168) shared MREs with four up-regulated DEmRNAs (HSD11B1, P2RX3, TCF7L1 and CHP2). HSD11B1 was enriched in the Rough endoplasmic reticulum (GO: 0005791, an organelle that synthesizes new proteins); P2RX3 was enriched in the Rough endoplasmic reticulum (GO: 0005791) and the Calcium signaling pathway (KEGG: mmu04020, an important ion pathway involved in neuronal synaptic signal transmission); TCF7L1 was enriched in the Wnt signaling pathway (KEGG: mmu04310, a key pathway for cell development); and CHP2 was enriched in Amyotrophic lateral sclerosis (ALS) (KEGG: mmu05014, a neurodegenerative disease), Apoptosis (KEGG: mmu04210), Wnt signaling pathway (KEGG: mmu04310) and Calcium signaling pathway (KEGG: mmu04020). Thus, the five aforementioned ncRNAs may contribute to the effects of Nrf2 on cognitive function by regulating protein synthesis, cell degeneration, neuronal development and intersynaptic signal transduction. We also found that mmu_circ_0015035 and NONMMUT127961 could be ceRNAs for HSD11B1, P2RX3 and TCF7L1, suggesting that mmu_circ_0015035 and NONMMUT127961 may assist the above five ncRNAs in regulating of cognitive function in the prefrontal cortex.
In addition, four ncRNAs (cicRNA.20127, mmu_circ_0012936, ENSMUST00000194077 and NONMMUT109267) were found to be ceRNAs for GSTM1, GSTM2 and MGST1. These three DEmRNAs were significantly down-regulated in the prefrontal cortex in Nrf2 (-/-) mice and were enriched in the pathway of Glutathione metabolism (KEGG: mmu00480). GSTM1 and GSTM2 encode glutathione S-transferases, which mainly participate in the phase II detoxification of endogenous oxidative stress products and exogenous electrophilic chemical compounds [39]. Reduced GSTs activity has been observed in multiple brain regions and ventricular cerebrospinal fluid from Alzheimer’s disease patients shortly after death [40]. Furthermore, the cytoprotective effects of Nrf2 are thought to be due to its strong induction of enzymes involved in glutathione synthesis and recycling [41]. MGST1 is a member of the Membrane-Associated Proteins in Eicosanoid and Glutathione metabolism family, an ancient and diverse family thought to predate the cytosolic family of GSTs [42]. MGST1 detoxifies reactive intermediates such as metabolic electrophile intermediates and lipophilic hydroperoxides through its glutathione-dependent transferase and peroxidase activities [43]. Thus, cicRNA.20127, mmu_circ_0012936, ENSMUST00000194077 and NONMMUT109267 may facilitate the effects of Nrf2 on cognitive function by regulating glutathione metabolism.
Interestingly, the results of the GO analysis included immune-related function such as Lymphocyte chemotaxis (GO: 0048247). The central nervous system lacks a classical lymphatic drainage system; however, the discovery of the lymphatic system in the central nervous system in 2015 suggested that central nervous system diseases may be associated with immune system disorders [44–46]. Our results support the notion that the lymphatic system is present in the brain, and indicate that its activity may be influenced by the Nrf2 pathway. However, the mechanisms whereby immune cells enter and exit the central nervous system remain poorly understood.
Although circRNAs and lncRNAs commonly alter gene expression as ceRNAs, when they are present at low levels they may not impact their target miRNAs [47]. CircRNAs and lncRNAs may function through other mechanisms, such as binding with proteins [48, 49] or being translated into proteins/peptides [50, 51]. Interestingly, we identified 44 DEcircRNAs and 7 DElncRNAs with coding potential, in keeping with the view that some ncRNAs may also be mRNAs.
In addition, some reports have indicated that circRNAs and lncRNAs can alter the expression of their host mRNAs [33]. Cis-acting circRNAs and lncRNAs regulate gene expression in a manner dependent on the location of their own transcription sites, at varying distances from their targets in the linear genome [52, 53]. Thus, we analyzed the expression correlations between DEcircRNAs/DElncRNAs and their corresponding host mRNAs. Our results suggested that only a small percentage of DEcircRNAs/DElncRNAs (38% of DEcircRNAs and 35% of DElncRNAs) altered the expression of their host mRNAs. These results were consistent with previous reports indicating that the expression of many circRNAs and lncRNAs is independent of the expression of their related mRNA isoforms [54, 55]. However, other studies have demonstrated that circRNAs are typically generated at the cost of their canonical mRNA isoforms [56, 57].
Among the DEcircRNAs that were correlated with their host mRNAs, only 30% exhibited negative correlations, suggesting that many DEcircRNAs activate transcription. CircRNA, especially exon-intron circRNAs, have been reported to bind specific ally with U1 small nuclear ribonucleoprotein RNA and promote the transcription of their parental genes in cis, although they may also influence other loci in trans [58, 59]. Likewise, the non-random positioning of lncRNAs implies that they are functionally associated with their nearby genes [60]. Cis-acting lncRNAs have been demonstrated to activate, repress or otherwise alter the expression of their target genes through various mechanisms [52]. In the present study, among the DElncRNAs that were associated with their host mRNAs, 67% exhibited positive correlations and 33% negative correlations.
We also determined the shared DEcircRNAs, DElncRNAs and DEmRNAs in four brain regions: prefrontal cortex, substantia nigra, corpus striatum and hippocampus. Interestingly, few circRNAs/lncRNAs were consistently regulated by Nrf2 in these four brain regions, indicating that Nrf2 may not have the same function in different brain areas. Recent studies have demonstrated that alters the expression of not only antioxidant genes, but also genes involved in autophagy, intermediate metabolism, stem cell quiescence and unfolded protein reactions [9]. Therefore, the function of Nrf2 is complex. Moreover, our research suggested that Nrf2 activity may be tissue-specific, which has not been reported before and is worth studying in the future.
In conclusion, our study demonstrated that four DEcircRNAs (mmu_circ_0000233, mmu_circ_0015035, cicRNA.20127 and mmu_circ_0012936) and seven DElncRNAs (ENSMUST00000204847, NONMMUT024778, NONMMUT132160, NONMMUT132168, NONMMUT127961, ENSMUST00000194077 and NONMMUT109267) may contribute to the effects of Nrf2 on cognitive function by regulating protein synthesis, cell degeneration, neuronal development, intersynaptic signal transduction and glutathione metabolism. Our study has laid the foundation for future investigations into the molecular pathways through which Nrf2 protects neurons against oxidative stress and exerts other functions.
MATERIALS AND METHODS
Nrf2-knockout mice and ethics
Adult male Nrf2 (+/+) and Nrf2 (-/-) mice (25-30 g, 3-4 months, n = 3) on an ICR background were kindly provided by Professor Chun-Yan Li (Department of Neurology, Second Hospital of Hebei Medical University, Shijiazhuang, China) for this study. PCR amplification of genomic DNA from tails was used to determine the genotypes of the mice. All the mice were sacrificed using an overdose of an isoflurane/oxygen mixture (Huazhong Haiwei Gene Technology Co, Ltd, Beijing, China, 021400). Prefrontal cortex tissues were surgically obtained from each mouse, and immediately homogenized for the extraction of total RNA (TRIzol, Invitrogen, Carlsbad, CA, USA, 15596026).
The animal experiments in this study complied with the regulations of the Animal Welfare Act and the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, revised 1996) and were approved by the ethics committee of Hebei Medical University (IACUC-Hebmu-Glp-2016017).
Microarray analysis
Six mice (Nrf2 (+/+) group, n = 3; Nrf2 (-/-) group, n = 3) were anesthetized, and their prefrontal cortex tissues were collected and rapidly stored in liquid nitrogen. Total RNA was extracted and purified using an RNeasy Mini Kit (Cat. #74106, QIAGEN, GmbH, Germany) in accordance with the manufacturer’s instructions. Shanghai Biotechnology Corporation (Shanghai, China) performed the quality control for the gene analyses by evaluating the total RNA with a NanoDrop ND-2000 spectrophotometer and an Agilent Bioanalyzer 2100 (Agilent Technologies, Inc., Santa Clara, CA, USA). Only RNA samples that met the quality control standards (RNA integrity number ≥ 7.0 and 28S/18S ≥ 0.7) were included in the subsequent microarray experiments. The included samples were amplified and labeled with a Low Input Quick Amp Labeling Kit, One-Color (Cat. #5190-2305, Agilent Technologies) in accordance with the manufacturer’s instructions. The labeled complementary RNA was purified with an RNeasy Mini Kit (Cat. #74106, QIAGEN).
An SBC Mouse (4*180 K) ceRNA microarray (Shanghai Biotechnology Corporation) was used to detect three types of RNA (circRNA, lncRNA and mRNA). Each slide was hybridized with 1.65 μg of Cy3-labeled complementary RNA using a Gene Expression Hybridization Kit (Cat. #5188-5242, Agilent Technologies) in a Hybridization Oven (Cat. #G2545A, Agilent Technologies) according to the manufacturer’s instructions. After 17 hours of hybridization, the slides were washed in staining dishes (Cat. #121, Thermo Fisher Scientific, Waltham, MA, USA) with a Gene Expression Wash Buffer Kit (Cat. # 5188-5327, Agilent Technologies) and then scanned on an Agilent Microarray Scanner (Cat. #G2565CA, Agilent Technologies) using the default settings. Data were extracted with Feature Extraction software 10.7 (Agilent Technologies). The raw data were normalized with the Quantile algorithm and LIMMA packages in RStudio (http://www.rstudio.com/). DEcircRNAs, DElncRNAs and DEmRNAs with p-values < 0.05 and FCs > 1.5 were selected through scatter plots and volcano diagrams using the ‘pheatmap’ package in R language. The data were also visualized through hierarchical clustering analyses.
qRT-PCR validation
Total RNA was isolated from Nrf2 (-/-) and Nrf2 (+/+) prefrontal cortex tissues using Trizol reagent (Thermo Fisher Scientific, Wilmington, DE, USA) according to the instruction manual, and the RNA quality was examined using 1% agarose gel electrophoresis. Then, the total RNA (2 μg) was reverse-transcribed to cDNA using an M-MLV First Strand Kit (Thermo Fisher Scientific Inc., 00341186) with random primers. The levels of six DEcircRNAs and eight lncRNAs were assessed via qRT-PCR using SuperReal PreMix Plus (SYBR Green) (TIANGEN, China, FP205) on an Illumina Eco PCR machine (Illumina, San Diego, CA, USA). The primers are displayed in Table 3. The qRT-PCR reaction conditions were as follows: an initial denaturation step of 15 min at 95° C, followed by 40 cycles of 15 s at 95° C, 20 s at 55° C, and 20 s at 72° C, and a final step of 5 min at 72° C. For normalization, β-actin was selected as an internal control. All experiments were performed in triplicate. The comparative cycle threshold (2-ΔΔCT) method was used to calculate the relative FCs.
Table 3. The primers used in qRT-PCR experiments.
| Transcript ID | Primers |
| mmu_circ_0000233 | Forward: 5′ ATCGCCACGCTACCCTCC 3′ |
| Reverse: 5′ CCAGCCTTTCCCCATCCA 3′ | |
| mmu_circ_0015035 | Forward: 5′ TTCAGAAGAACAAAGATTACTAC 3′ |
| Reverse: 5′ CACTGGAACAGGTGAGATG 3′ | |
| mmu_circ_0003404 | Forward: 5′ TCAACCCAGACCCCAAGT 3′ |
| Reverse: 5′ TCCTGAGAGTCCACAGAGAAAT 3′ | |
| mmu_circ_0012936 | Forward: 5′ ATCGTATGGCTTGCCTTC 3′ |
| Reverse: 5′ GGGGTTCCCTTCTGTGTC 3′ | |
| mmu_circ_0008393 | Forward: 5′ CAAAACTAAGTGAAAGGC 3′ |
| Reverse: 5′ GAAAGTCCAGACCATTG 3′ | |
| cicRNA.20127 | Forward: 5′ CTCCCGCCTGAACTACC 3′ |
| Reverse: 5′ CACAAACAGCCAACCATC 3′ | |
| NONMMUT132168 | Forward: 5' CTCATAGACTAAACTTTCACCTT 3' |
| Reverse: 5' CCCCTCAAACTAACCATC 3' | |
| NONMMUT127961 | Forward: 5' AGAAGGCACTGGACTGGCTC 3' |
| Reverse: 5' GAGTTGGAGTTCCGCTGTGAG 3' | |
| NONMMUT132160 | Forward: 5' CACTGTGTGTCGCTTGT 3' |
| Reverse: 5' GTGACTACTGATAGAGAGGG 3' | |
| NR_045161 | Forward: 5' CACATAGAAGCCACACCCA 3' |
| Reverse: 5' AGTCCTCCACCTCGCCT 3' | |
| ENSMUST00000204847 | Forward: 5' TGACCACTTGGGGGAAAT 3' |
| Reverse: 5' AAGAGGCTGCTTAGAAAAACA 3' | |
| NONMMUT024778 | Forward: 5' CCACTGTTTTGTCGGGGT 3' |
| Reverse: 5' CTGGGTGTTGGGGCTATT 3' | |
| NONMMUT109267 | Forward: 5' TGCGATGTGTTCTGAGTT 3' |
| Reverse: 5' TGTGTCCTGCTGTTCTTT 3' | |
| ENSMUST00000194077 | Forward: 5' ATCATTTTCTCTTATCAT 3' |
| Reverse: 5' TTACTCTCACTTTCTCTC 3' | |
| β-actin | Forward: 5' TCATCACTATTGGCAACGAGCGGT 3' |
| Reverse: 5' GTGTTGGCATAGAGGTCTTTACG 3' |
qRT-PCR, quantitative real-time polymerase chain reaction.
Construction of ceRNA networks based on DEcircRNA/DElncRNA-miRNA-DEmRNA interactions
DEcircRNAs and DElncRNAs verified by qRT-PCR were selected for the construction of interaction networks by Shanghai Biotechnology Corporation. First, a DEcircRNA/DElncRNA-DEmRNA co-expression analysis was performed based on the Pearson correlation coefficients between DEmRNA and DEcircRNA/DElncRNA levels in the SBC Mouse (4*180K) ceRNA Array analysis. Genes with Pearson correlation coefficients > 0.81 and p-values < 0.05 were recommended for further analysis. Then, DEcircRNA/DElncRNA-miRNA-DEmRNA interaction networks were constructed using miRanda, and binding sites with relatively high scores (≥ 140) were identified. Finally, the networks were visualized using Cytoscape_v3.7.0 (http://www.cytoscape.org/).
Functional annotation of DEmRNAs that were targets of DEcircRNAs and DElncRNAs
GO and KEGG pathway enrichment analyses were conducted for the target DEmRNAs of miRNAs sponged by the DEcircRNAs/DElncRNAs in the prefrontal cortex tissues. The analyses were performed using the clusterProfiler of R/bioconductor (http://enrich.shbio.com/index/ga.asp) to determine the potential functions of the DEcircRNAs and DElncRNAs.
Coding potential of DEcircRNAs and DElncRNAs
The Coding-Non-Coding Index (https://github.com/www-bioinfo-org/CNCI) [27], which distinguishes between coding and noncoding transcripts based on their triplet base composition, was used to predict the protein coding potential of the DEcircRNAs and DElncRNAs (The full-length sequences of some of the DEcircRNAs have yet to be confirmed, so the sequences within 300 base pairs around the cut site were used to predict their coding potential).
Expression correlation analysis between DEcircRNAs/DElncRNAs and their host mRNAs
Pearson correlation coefficients were used to determine the expression correlations between DEcircRNAs/DElncRNAs and mRNAs encoded by the same parental genes. DEcircRNA/DElncRNA-host mRNA pairs with p-values < 0.05 were retained.
Comparison of DEcircRNAs, DElncRNAs and DEmRNAs among the prefrontal cortex, substantia nigra, corpus striatum and hippocampus
DEcircRNAs, DElncRNAs and DEmRNAs in the substantia nigra, corpus striatum and hippocampus were screened in our previous studies of Nrf2 (-/-) and Nrf2 (+/+) mice [23–25]. The shared DEcircRNAs, DElncRNAs and DEmRNAs among these areas and the prefrontal cortex were determined using the Bioinformatics and Evolutionary Genomics tool (http://bioinformatics.psb.ugent.be/webtools/Venn/).
Statistical analysis
All data were analyzed using SPSS 22.0 software (IBM). The means and standard deviations were determined, and independent-samples t-test were performed. P-values < 0.05 was considered significant.
Supplementary Material
ACKNOWLEDGMENTS
We appreciate Professor Chun-Yan Li for supplying the experimental animals.
Abbreviations
- Nrf2/NFE2L2
nuclear factor erythroid-2-related factor 2
- CNC
cap ‘n’collar
- ncRNA
non-coding RNA
- circRNA
circular RNA
- lncRNA
long non-coding RNA
- miRNA
microRNA
- ceRNA
competitive endogenous RNA
- DEcircRNAs
differentially expressed circRNAs
- DElncRNAs
differentially expressed lncRNAs
- DEmRNAs
differentially expressed mRNAs
- FC
fold change
- PCR
polymerase chain reaction
- MRE
miRNA response element
- ALS
Amyotrophic lateral sclerosis
- GSTs
glutathione S-transferases
Footnotes
AUTHOR CONTRIBUTIONS: Dan-Dan Geng and Lei Wang conceived the study. Yan-Jing Gao, Run-Jiao Zhang and Qing Liu contributed materials and performed the experiments. Shao-Guang Sun, Mao-Yang Qi and Yue Wang analyzed the data. Yan-Jing Gao contributed significantly to the writing of the manuscript. All authors read and approved the final manuscript.
CONFLICTS OF INTEREST: The authors declare that they have no conflicts of interest.
FUNDING: This study was supported by National Natural Science Foundation of China (81670273), High-level Talent Foundation of Hebei Province (A2016002073) and Hebei Provincial Natural Science Foundation Precision Medicine Joint Fund Cultivation Project (C2020206037, H2020206299). The funding body had no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
REFERENCES
- 1.Chan K, Lu R, Chang JC, Kan YW. NRF2, a member of the NFE2 family of transcription factors, is not essential for murine erythropoiesis, growth, and development. Proc Natl Acad Sci U S A. 1996; 93:13943–48. 10.1073/pnas.93.24.13943 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Venugopal R, Jaiswal AK. Nrf1 and Nrf2 positively and c-Fos and Fra1 negatively regulate the human antioxidant response element-mediated expression of NAD(P)H:quinone oxidoreductase1 gene. Proc Natl Acad Sci U S A. 1996; 93:14960–65. 10.1073/pnas.93.25.14960 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Itoh K, Chiba T, Takahashi S, Ishii T, Igarashi K, Katoh Y, Oyake T, Hayashi N, Satoh K, Hatayama I, Yamamoto M, Nabeshima Y. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem Biophys Res Commun. 1997; 236:313–22. 10.1006/bbrc.1997.6943 [DOI] [PubMed] [Google Scholar]
- 4.Kensler TW, Wakabayashi N, Biswal S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu Rev Pharmacol Toxicol. 2007; 47:89–116. 10.1146/annurev.pharmtox.46.120604.141046 [DOI] [PubMed] [Google Scholar]
- 5.Shih AY, Johnson DA, Wong G, Kraft AD, Jiang L, Erb H, Johnson JA, Murphy TH. Coordinate regulation of glutathione biosynthesis and release by Nrf2-expressing glia potently protects neurons from oxidative stress. J Neurosci. 2003; 23:3394–406. 10.1523/JNEUROSCI.23-08-03394.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Vargas MR, Pehar M, Cassina P, Beckman JS, Barbeito L. Increased glutathione biosynthesis by Nrf2 activation in astrocytes prevents p75NTR-dependent motor neuron apoptosis. J Neurochem. 2006; 97:687–96. 10.1111/j.1471-4159.2006.03742.x [DOI] [PubMed] [Google Scholar]
- 7.Shih AY, Li P, Murphy TH. A small-molecule-inducible Nrf2-mediated antioxidant response provides effective prophylaxis against cerebral ischemia in vivo. J Neurosci. 2005; 25:10321–35. 10.1523/JNEUROSCI.4014-05.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zweig JA, Caruso M, Brandes MS, Gray NE. Loss of NRF2 leads to impaired mitochondrial function, decreased synaptic density and exacerbated age-related cognitive deficits. Exp Gerontol. 2020; 131:110767. 10.1016/j.exger.2019.110767 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tonelli C, Chio II, Tuveson DA. Transcriptional regulation by Nrf2. Antioxid Redox Signal. 2018; 29:1727–45. 10.1089/ars.2017.7342 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bartolini D, Dallaglio K, Torquato P, Piroddi M, Galli F. Nrf2-p62 autophagy pathway and its response to oxidative stress in hepatocellular carcinoma. Transl Res. 2018; 193:54–71. 10.1016/j.trsl.2017.11.007 [DOI] [PubMed] [Google Scholar]
- 11.Jeck WR, Sharpless NE. Detecting and characterizing circular RNAs. Nat Biotechnol. 2014; 32:453–61. 10.1038/nbt.2890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Wu XC, Wang SH, Ou HH, Zhu B, Zhu Y, Zhang Q, Yang Y, Li H. The NmrA-like family domain containing 1 pseudogene Loc344887 is amplified in gallbladder cancer and promotes epithelial-mesenchymal transition. Chem Biol Drug Des. 2017; 90:456–63. 10.1111/cbdd.12967 [DOI] [PubMed] [Google Scholar]
- 13.Gullett JM, Chen Z, O’Shea A, Akbar M, Bian J, Rani A, Porges EC, Foster TC, Woods AJ, Modave F, Cohen RA. MicroRNA predicts cognitive performance in healthy older adults. Neurobiol Aging. 2020; 95:186–94. 10.1016/j.neurobiolaging.2020.07.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gao J, Xu W, Wang J, Wang K, Li P. The role and molecular mechanism of non-coding RNAs in pathological cardiac remodeling. Int J Mol Sci. 2017; 18:608. 10.3390/ijms18030608 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chen LY, Wang L, Ren YX, Pang Z, Liu Y, Sun XD, Tu J, Zhi Z, Qin Y, Sun LN, Li JM. The circular RNA circ-ERBIN promotes growth and metastasis of colorectal cancer by miR-125a-5p and miR-138-5p/4EBP-1 mediated cap-independent HIF-1α translation. Mol Cancer. 2020; 19:164. 10.1186/s12943-020-01272-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhang D, Ni N, Wang Y, Tang Z, Gao H, Ju Y, Sun N, He X, Gu P, Fan X. CircRNA-vgll3 promotes osteogenic differentiation of adipose-derived mesenchymal stem cells via modulating miRNA-dependent integrin α5 expression. Cell Death Differ. 2021; 28:283–302. 10.1038/s41418-020-0600-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shen S, Yao T, Xu Y, Zhang D, Fan S, Ma J. CircECE1 activates energy metabolism in osteosarcoma by stabilizing c-Myc. Mol Cancer. 2020; 19:151. 10.1186/s12943-020-01269-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Li K, Xu J, Luo Y, Zou D, Han R, Zhong S, Zhao Q, Mang X, Li M, Si Y, Lu Y, Li P, Jin C, et al. Panoramic transcriptome analysis and functional screening of long noncoding RNAs in mouse spermatogenesis. Genome Res. 2021; 31:13–26. 10.1101/gr.264333.120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Qu S, Yang X, Li X, Wang J, Gao Y, Shang R, Sun W, Dou K, Li H. Circular RNA: a new star of noncoding RNAs. Cancer Lett. 2015; 365:141–48. 10.1016/j.canlet.2015.06.003 [DOI] [PubMed] [Google Scholar]
- 20.Hansen TB, Jensen TI, Clausen BH, Bramsen JB, Finsen B, Damgaard CK, Kjems J. Natural RNA circles function as efficient microRNA sponges. Nature. 2013; 495:384–88. 10.1038/nature11993 [DOI] [PubMed] [Google Scholar]
- 21.Liu L, Wang H, Yu Y, Zeng B, Rao X, Chen J, Zhou C, Zheng P, Pu J, Yang L, Zhang H, Wei H, Xie P. Microbial regulation of a lincRNA-miRNA-mRNA network in the mouse hippocampus. Epigenomics. 2020; 12:1377–87. 10.2217/epi-2019-0307 [DOI] [PubMed] [Google Scholar]
- 22.Wesselhoeft RA, Kowalski PS, Anderson DG. Engineering circular RNA for potent and stable translation in eukaryotic cells. Nat Commun. 2018; 9:2629. 10.1038/s41467-018-05096-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yang JH, Zhang RJ, Lin JJ, Cao MC, Wang Q, Cui HX, Sun SG, Wang L. The differentially expressed circular RNAs in the substantia nigra and corpus striatum of Nrf2-knockout mice. Cell Physiol Biochem. 2018; 50:936–51. 10.1159/000494478 [DOI] [PubMed] [Google Scholar]
- 24.Liu J, Xu Y, Kang Y, Cao S, Shi G, Cui H, Sun S, Wang L. The aberrantly expressed long non-coding RNA in the substantia nigra and corpus striatum of Nrf2-knockout mice. J Neurochem. 2017; 143:65–75. 10.1111/jnc.14141 [DOI] [PubMed] [Google Scholar]
- 25.Zhang RJ, Li Y, Liu Q, Gao YJ, Du J, Ma J, Sun SG, Wang L. Differential expression profiles and functional prediction of circular RNAs and long non-coding RNAs in the hippocampus of Nrf2-knockout mice. Front Mol Neurosci. 2019; 12:196. 10.3389/fnmol.2019.00196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Miller EK, Cohen JD. An integrative theory of prefrontal cortex function. Annu Rev Neurosci. 2001; 24:167–202. 10.1146/annurev.neuro.24.1.167 [DOI] [PubMed] [Google Scholar]
- 27.Sun L, Luo H, Bu D, Zhao G, Yu K, Zhang C, Liu Y, Chen R, Zhao Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013; 41:e166. 10.1093/nar/gkt646 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Labus J, Röhrs KF, Ackmann J, Varbanov H, Müller FE, Jia S, Jahreis K, Vollbrecht AL, Butzlaff M, Schill Y, Guseva D, Böhm K, Kaushik R, et al. Amelioration of Tau pathology and memory deficits by targeting 5-HT7 receptor. Prog Neurobiol. 2021; 197:101900. 10.1016/j.pneurobio.2020.101900 [DOI] [PubMed] [Google Scholar]
- 29.Ji MH, Lei L, Gao DP, Tong JH, Wang Y, Yang JJ. Neural network disturbance in the medial prefrontal cortex might contribute to cognitive impairments induced by neuroinflammation. Brain Behav Immun. 2020; 89:133–44. 10.1016/j.bbi.2020.06.001 [DOI] [PubMed] [Google Scholar]
- 30.Rojo AI, Pajares M, Rada P, Nuñez A, Nevado-Holgado AJ, Killik R, Van Leuven F, Ribe E, Lovestone S, Yamamoto M, Cuadrado A. NRF2 deficiency replicates transcriptomic changes in Alzheimer’s patients and worsens APP and TAU pathology. Redox Biol. 2017; 13:444–51. 10.1016/j.redox.2017.07.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Skibinski G, Hwang V, Ando DM, Daub A, Lee AK, Ravisankar A, Modan S, Finucane MM, Shaby BA, Finkbeiner S. Nrf2 mitigates LRRK2- and α-synuclein-induced neurodegeneration by modulating proteostasis. Proc Natl Acad Sci USA. 2017; 114:1165–70. 10.1073/pnas.1522872114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Silva-Palacios A, Ostolga-Chavarría M, Zazueta C, Königsberg M. Nrf2: molecular and epigenetic regulation during aging. Ageing Res Rev. 2018; 47:31–40. 10.1016/j.arr.2018.06.003 [DOI] [PubMed] [Google Scholar]
- 33.Xu K, Zhang Y, Xiong W, Zhang Z, Wang Z, Lv L, Liu C, Hu Z, Zheng YT, Lu L, Hu XT, Li J. CircGRIA1 shows an age-related increase in male macaque brain and regulates synaptic plasticity and synaptogenesis. Nat Commun. 2020; 11:3594. 10.1038/s41467-020-17435-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cai H, Li Y, Niringiyumukiza JD, Su P, Xiang W. Circular RNA involvement in aging: an emerging player with great potential. Mech Ageing Dev. 2019; 178:16–24. 10.1016/j.mad.2018.11.002 [DOI] [PubMed] [Google Scholar]
- 35.Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer M, Loewer A, Ziebold U, Landthaler M, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013; 495:333–38. 10.1038/nature11928 [DOI] [PubMed] [Google Scholar]
- 36.Li Z, Zhu X, Huang S. Extracellular vesicle long non-coding RNAs and circular RNAs: biology, functions and applications in cancer. Cancer Lett. 2020; 489:111–20. 10.1016/j.canlet.2020.06.006 [DOI] [PubMed] [Google Scholar]
- 37.Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the rosetta stone of a hidden RNA language? Cell. 2011; 146:353–58. 10.1016/j.cell.2011.07.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chen W, Lin J, Li B, Cao S, Li H, Zhao J, Liu K, Li Y, Li Y, Sun S. Screening and functional prediction of differentially expressed circRNAs in proliferative human aortic smooth muscle cells. J Cell Mol Med. 2020; 24:4762–72. 10.1111/jcmm.15150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Li B, Zhou J, Zhang G, Wang Y, Kang L, Wu J, Chen J, Guan H. Relationship between the altered expression and epigenetics of GSTM3 and age-related cataract. Invest Ophthalmol Vis Sci. 2016; 57:4721–32. 10.1167/iovs.16-19242 [DOI] [PubMed] [Google Scholar]
- 40.Lovell MA, Gabbita SP, Markesbery WR. Increased DNA oxidation and decreased levels of repair products in Alzheimer’s disease ventricular CSF. J Neurochem. 1999; 72:771–76. 10.1046/j.1471-4159.1999.0720771.x [DOI] [PubMed] [Google Scholar]
- 41.Telorack M, Meyer M, Ingold I, Conrad M, Bloch W, Werner S. A glutathione-Nrf2-thioredoxin cross-talk ensures keratinocyte survival and efficient wound repair. PLoS Genet. 2016; 12:e1005800. 10.1371/journal.pgen.1005800 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kelner MJ, Diccianni MB, Yu AL, Rutherford MR, Estes LA, Morgenstern R. Absence of MGST1 mRNA and protein expression in human neuroblastoma cell lines and primary tissue. Free Radic Biol Med. 2014; 69:167–71. 10.1016/j.freeradbiomed.2014.01.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Schaffert CS. Role of MGST1 in reactive intermediate-induced injury. World J Gastroenterol. 2011; 17:2552–57. 10.3748/wjg.v17.i20.2552 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Schiller M, Ben-Shaanan TL, Rolls A. Neuronal regulation of immunity: why, how and where? Nat Rev Immunol. 2021; 21:20–36. 10.1038/s41577-020-0387-1 [DOI] [PubMed] [Google Scholar]
- 45.Aspelund A, Antila S, Proulx ST, Karlsen TV, Karaman S, Detmar M, Wiig H, Alitalo K. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J Exp Med. 2015; 212:991–99. 10.1084/jem.20142290 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Louveau A, Smirnov I, Keyes TJ, Eccles JD, Rouhani SJ, Peske JD, Derecki NC, Castle D, Mandell JW, Lee KS, Harris TH, Kipnis J. Structural and functional features of central nervous system lymphatic vessels. Nature. 2015; 523:337–41. 10.1038/nature14432 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Denzler R, Agarwal V, Stefano J, Bartel DP, Stoffel M. Assessing the ceRNA hypothesis with quantitative measurements of miRNA and target abundance. Mol Cell. 2014; 54:766–76. 10.1016/j.molcel.2014.03.045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Du WW, Zhang C, Yang W, Yong T, Awan FM, Yang BB. Identifying and characterizing circRNA-protein interaction. Theranostics. 2017; 7:4183–91. 10.7150/thno.21299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ferrè F, Colantoni A, Helmer-Citterich M. Revealing protein-lncRNA interaction. Brief Bioinform. 2016; 17:106–16. 10.1093/bib/bbv031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Liang WC, Wong CW, Liang PP, Shi M, Cao Y, Rao ST, Tsui SK, Waye MM, Zhang Q, Fu WM, Zhang JF. Translation of the circular RNA circβ-catenin promotes liver cancer cell growth through activation of the Wnt pathway. Genome Biol. 2019; 20:84. 10.1186/s13059-019-1685-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wang J, Zhu S, Meng N, He Y, Lu R, Yan GR. ncRNA-encoded peptides or proteins and cancer. Mol Ther. 2019; 27:1718–25. 10.1016/j.ymthe.2019.09.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Gil N, Ulitsky I. Regulation of gene expression by cis-acting long non-coding RNAs. Nat Rev Genet. 2020; 21:102–17. 10.1038/s41576-019-0184-5 [DOI] [PubMed] [Google Scholar]
- 53.Yang F, Fang E, Mei H, Chen Y, Li H, Li D, Song H, Wang J, Hong M, Xiao W, Wang X, Huang K, Zheng L, Tong Q. Cis-acting circ-CTNNB1 promotes β-catenin signaling and cancer progression via DDX3-mediated transactivation of YY1. Cancer Res. 2019; 79:557–71. 10.1158/0008-5472.CAN-18-1559 [DOI] [PubMed] [Google Scholar]
- 54.Salzman J, Chen RE, Olsen MN, Wang PL, Brown PO. Cell-type specific features of circular RNA expression. PLoS Genet. 2013; 9:e1003777. 10.1371/journal.pgen.1003777 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Engreitz JM, Haines JE, Perez EM, Munson G, Chen J, Kane M, McDonel PE, Guttman M, Lander ES. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature. 2016; 539:452–55. 10.1038/nature20149 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Salzman J, Gawad C, Wang PL, Lacayo N, Brown PO. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS One. 2012; 7:e30733. 10.1371/journal.pone.0030733 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ashwal-Fluss R, Meyer M, Pamudurti NR, Ivanov A, Bartok O, Hanan M, Evantal N, Memczak S, Rajewsky N, Kadener S. circRNA biogenesis competes with pre-mRNA splicing. Mol Cell. 2014; 56:55–66. 10.1016/j.molcel.2014.08.019 [DOI] [PubMed] [Google Scholar]
- 58.Li Z, Huang C, Bao C, Chen L, Lin M, Wang X, Zhong G, Yu B, Hu W, Dai L, Zhu P, Chang Z, Wu Q, et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat Struct Mol Biol. 2015; 22:256–64. 10.1038/nsmb.2959 [DOI] [PubMed] [Google Scholar]
- 59.Zlotorynski E. Non-coding RNA: circular RNAs promote transcription. Nat Rev Mol Cell Biol. 2015; 16:206. 10.1038/nrm3967 [DOI] [PubMed] [Google Scholar]
- 60.Luo S, Lu JY, Liu L, Yin Y, Chen C, Han X, Wu B, Xu R, Liu W, Yan P, Shao W, Lu Z, Li H, et al. Divergent lncRNAs regulate gene expression and lineage differentiation in pluripotent cells. Cell Stem Cell. 2016; 18:637–52. 10.1016/j.stem.2016.01.024 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.