

Abstract
The engineering of living cells and microbes is ushering in a new era of cancer therapy. Due to recent microbiome studies indicating the prevalence of bacteria within the human body and specifically in tumor tissue, bacteria have generated significant interest as potential targets for cancer therapy. Notably, a multitude of empirical studies over the past decades have demonstrated that administered bacteria home and grow in tumors due to reduced immune surveillance of tumor necrotic cores. Given their specificity for tumors, bacteria present a unique opportunity to be engineered as intelligent delivery vehicles for cancer therapy with synthetic biology techniques. In this review, we discuss the history, current state, and future challenges associated with using bacteria as a cancer therapy.
Introduction
Synthetic biology has produced numerous examples of genetic circuits that produce complex, dynamic behaviors in single-cells or across populations including switches, oscillators, sensors, filters, counters, and recorders [1–9]. While most genetic circuits have been engineered in a controlled laboratory environment as a proof of principle, a growing number of these circuits are being implemented for environmental, material, and medical applications. Because of the realization that bacteria exist in a multitude of tissue types previously thought to be sterile [10–12], an emerging focus of synthetic biology is to design bacteria to supplant natural niches to tackle various diseases including inflammation [13,14], infections [15–17], metabolic disorders [18,19], and cancer [20–22]. With synthetic biology’s level of cellular control, bacteria can be programmed as living medicines that sense their environment and produce agents that diagnose and treat diseases.
Bacteria cancer therapy: a brief history
One of the earliest reports of using bacteria as a cancer therapy dates back to 1891, when Dr. William Coley started to inject live cultures of Streptococcus bacteria that led to remissions of inoperable bone and soft tissue sarcomas in patients [23]. Largely due to the dangers associated with infection from the live bacteria before the widespread availability of antibiotics, the clinical use of this therapeutic approach went undeveloped. The therapeutic effect generated by bacteria is thought to be mediated by two key principles. First, a multitude of studies has shown that many bacteria selectively colonize tumor cores through leaky vasculature upon systemic administration, primarily due to reduced immune surveillance in the tumor’s hypoxic and acidic environment [24–32]. Second, while inside of tumors, bacteria compete with cancer cells for nutrients, produce toxins, and stimulate a local immune response [32,33]. Over the past century, several bacteria have been explored for cancer therapy such as studies using attenuated Clostridium novyi spores that demonstrated tumor regression in mice, dogs, and eventually a human patient [27,34]. While these studies have shown some promising results, further development in enhancing efficacy while retaining safety profiles are needed. To date, the only example of a common bacteria cancer therapy in clinics remains to be the use of Mycobacterium bovis (BCG therapy) for the treatment of high-risk non-muscle-invasive bladder cancer [35,36].
Since finding a bacteria species with the ideal properties to treat cancers is challenging, the focus has shifted to genetic engineering of bacteria with enhanced safety and efficacy profiles for cancer therapy. Escherichia coli and Salmonella typhimurium are emerging as the leading and most frequently investigated engineered bacteria due to their ease in genetic manipulation. Importantly, various genetic engineering tools allow for genetic attenuations such as a reduction in strain virulence and endotoxicity [37–40]. For example, knockout of the msbB gene, involved in terminal myristoylation of lipid A, resulted in viable strains that reduce TNF-α induction 104-fold in S. typhimurium. Further knockouts of purI and xyl genes prevented metabolism of purine and xylose, reducing pathogenicity. A strain with all of these attenuations (VNP20009) was tested in a Phase I metastatic melanoma trial, where a maximum tolerated a dose of 3 × 108 intravenously administered bacteria was seen to cause no significant grade toxicities [41,42]. While safety was largely established, no efficacy was observed in these trials with bacteria alone, and thus engineering bacteria to locally produce therapeutics in tumors has now become the focal point of the bacteria cancer therapy approach.
Current research and future directions
One of the primary advantages of bacteria therapies is local therapeutic delivery (Figure 1), which allows for a reduction in toxicities associated with systemic drug delivery [43,44]. For example, chemotherapies including 5-fluorouracil for the treatment of advanced and metastatic colorectal cancer cause cytotoxicity to healthy cells, leading to dose reduction, delays and discontinuation of therapies which ultimately limits therapeutic efficacy [45–47]. Additionally, patient tumors can be resistant to standard of care treatments [48], necessitating alternative therapeutic approaches. To increase efficacy, combination cytotoxic therapy is employed as an effective approach such as fluorouracil, leucovorin, and irinotecan (FOLFIRI) and other combinations (FOLFOX, XELOX, FOLFOXIRI) [49–51]. However, systemic delivery of combination chemo- and immunotherapy lead to significantly higher toxicities than single agents alone [52,53]. Since bacteria selectively colonize tumors, this targeted and controlled approach can deliver therapeutics that can be toxic systemically, such as bacterial toxins, and improve efficacy and safety profiles. Various therapeutics can be made from bacteria to target a broad range of anti-tumor mechanisms such as membrane-damaging, apoptosis, protein synthesis inhibition, and innate/adaptive immune stimulation. Notably, cancer immunotherapy has witnessed a renaissance in the past decade with the development of novel engineered immune cell therapies and antibodies that target immune checkpoints to activate antitumor immunity [54]. Combining immunotherapy and microbial synthetic biology, bacteria have recently been engineered to deliver immunostimulants directly into the core of a growing tumor — locally priming antitumor immune cells to systemically attack primary tumor and disseminated metastases [55,56]. Delivery of such therapeutics by engineered bacteria in situ will enhance ‘on-tumor’ targeting specificity to elicit superior antitumor efficacy with less toxicity than current systemic anticancer treatment regimens.
Figure 1. Approach for the development of bacterial cancer therapies.
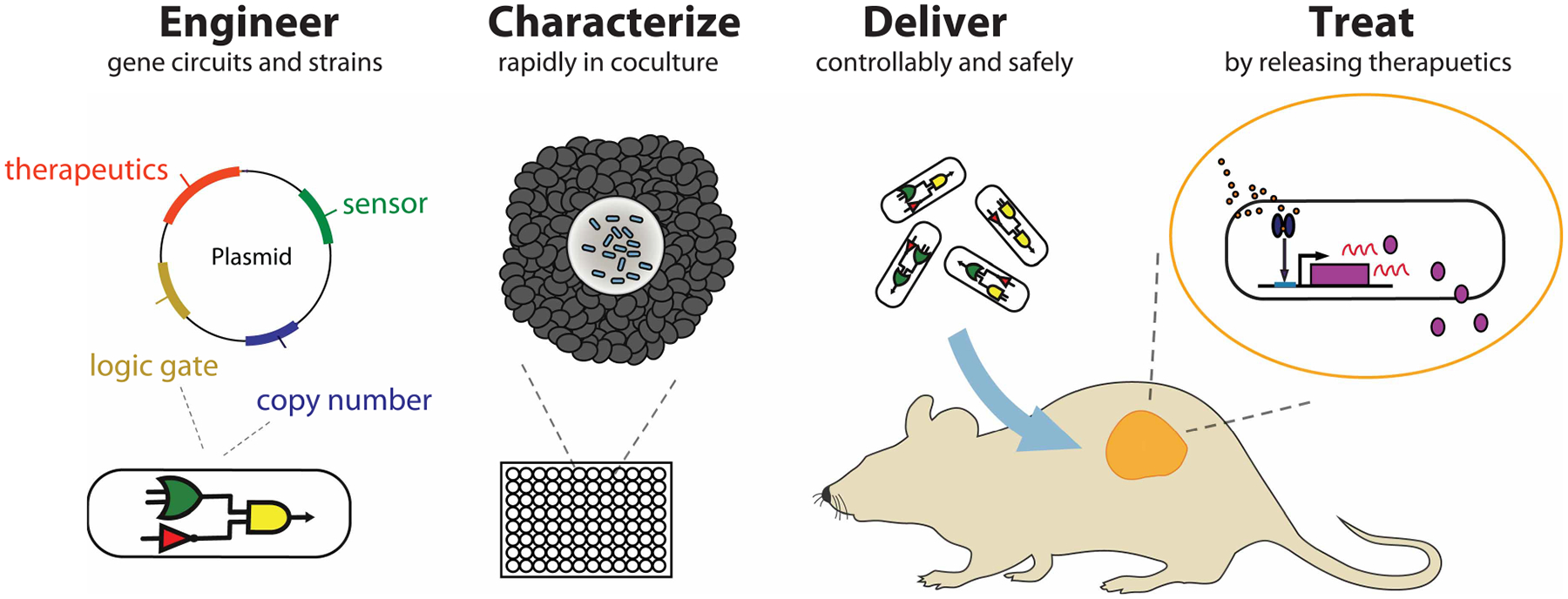
Bacteria are engineered guided by synthetic biology design and introduction of plasmid or modifying genomes. In vitro coculture systems can be used to rapidly characterize and develop effective engineered bacteria. To safely deliver therapeutics to the tumor, bacteria can sense and respond to a tumor microenvironment using synthetic gene circuits. Therapeutics are locally released including potentially toxic molecules such as bacterial toxins and potent immune stimulators.
The production of potentially toxic therapeutic cargo has necessitated further control over the expression and delivery of therapeutics. New synthetic biology approaches are being developed that enable bacteria to controllably sense and respond to tumor environments (Figure 1), which can be used for enhancing safety and efficacy. For example, genetic circuits were recently reported enabling bacteria to sense bacterial population density using quorum sensing to trigger gene expressions in vivo [15,57]. In addition, quorum sensing has been coupled with bacterial self-lysis, effectively releasing therapeutics and maintaining a low population in tumors [58]. This engineered strain increased survival in a liver metastasis model when orally delivered, while also improving safety profiles compared with constitutively producing therapeutic strains. The release of the therapeutic can be also achieved in many other ways such as secretion, passive cell death, or phagocytosis by host cells in the body [32]. Additionally, triggering of the production of therapeutics or release can be connected to an array of environmental cues that may mark a tumor microenvironment such as hypoxia or high lactate levels [19,21,59,60]. These sensing and actuating activities will affect surrounding cancer cells in a heterogeneous manner and can be used to further optimize therapeutic efficacy.
Although there has been significant development of engineered bacteria as a cancer therapy, the vast majority of studies incrementally study one bacteria strain, one therapeutic payload, one genetic circuit, in an animal model. However, the ability to create engineered therapies far outpaces the throughput of animal-based testing, thus creating a major bottleneck to the progress of engineered microbial and cell therapies [61,62]. Coculture of bacteria with mammalian cells provides a powerful platform to study the performance of engineered cells in vitro (Figure 1). Coculture systems have traditionally relied on physical compartmentalization of microbes and cells by porous membranes for a relatively short time scale to prevent bacterial overgrowth in culture [61]. This approach is simple to execute, but lacks spatial orientation and contact between the cells seen in vivo. The role of 3D interactions between cells has been shown to create strikingly distinct responses to drugs compared with traditional monolayers [63]. To this end, 3D models including the use of multicellular spheroids, tumor cylindroids, tumor-on-a-chip, and organoids have been introduced to recapitulate spatial orientation in relation to interaction with bacteria [64–67]. Microfluidics has also been employed to allow bacterial population control over a longer time period by washing out excess bacteria [58,66]. These approaches enabled quantitative measurement of bacterial penetration to tumor tissues in a controlled environment and provided valuable insights such as the role of chemotaxis, motility, and tumor heterogeneity [57,66,67]. The majority of these systems have tended to have low throughput and used over relatively short time windows due to complexity, population instability, and limitation to types of cells used in the platform. Furthermore, long-term analysis of bacteria circuit dynamics is not commonly assessed, which is important for creating new active sense and respond circuitry [61,68]. One example to address this challenge is a recent development of high-throughput coculture system that allows for long-term growth of bacteria in the necrotic core of tumor spheroids, recapitulating bacterial colonization and therapeutic outcomes of tumors in vivo [69]. Utilizing this system, therapeutic candidates exhibiting significant tumor reduction were identified that demonstrated superior efficacy to previously characterized therapeutics in animal models. Integration of such in vitro platforms will enable rapid characterization of novel therapeutics, gene circuits, strains, and cell lines for clinical translation of engineered bacterial therapy.
While there has been progress in the use of synthetic biology and coculture systems to develop bacteria for cancer therapy, the precise mechanism of tumor colonization is still not well understood. The role of bacterial motility has been suggested to contribute to an active colonization mechanism with contradicting results thus far. Some studies have shown overexpression of flagellum regulator flhDC can improve bacterial accumulation by up to five times compared with the wildtype bacteria in vitro [70], while others have demonstrated non-motile bacterial spores or non-motile cloning strains are also capable of localizing and germinating within the tumor [34,71]. Furthermore, the role of immune response to bacterial targeting to tumor tissue remains unclear. One report has demonstrated that pro-inflammatory cytokine production upon bacterial systemic introduction, specifically TNF-α, triggers an influx of blood into tumors that carries bacteria together and important for achieving effective colonization [26]. On the other hand, another study has shown that reduction in TNF-α by using msbB deleted S. typhimurium did not compromise its tumor-targeting capability [38]. These types of mechanistic studies on bacterial colonization are critical to better understand and design future bacteria therapies (Figure 1).
Comparison of colonization mechanism and therapeutic effect between bacterial species can accelerate the development of effective therapy. For example, BCG is delivered through the intravesical route and attaches to the urothelium layer in the bladder [72], and seem to disappear quickly from the tissue without persistence [73]. E. coli has also been engineered to attach to the colorectal cancer epithelial cells after oral delivery [74]. Listeria, when injected intraperitoneally, can accumulate in tumors and are thought to be brought by infected myeloid-derived suppressor cells [75,76]. However, how these compare to the mechanism of selective colonization of bacteria in tumor necrotic cores is not well known. In addition, the extent of which tumors can be colonized via various delivery routes (intratumoral, intravenous, intraperitoneal, and oral) and which cancers can be accessed is still under exploration as well. One study reported more abundant growth of attenuated Listeria monocytogenes in metastasis compared with the primary pancreatic tumor upon intraperitoneal delivery [75]. Lastly, while it is widely believed that the inflammatory reaction to BCG mediates innate immune response, leading to antitumor adaptive immunity [77], the majority of attempts to improve upon the current BCG therapy have been unsuccessful for treatment of non-muscle-invasive bladder cancer [77]. Current research on combination treatment strategy with other standard-of-care therapies and understanding interpatient response variability may provide novel insight to improving bacteria cancer therapies [78–81].
Increasing knowledge of the microbiota that resides in the human body has led to a developing area of research in using microbes for therapy [82–88]. Future research may utilize and engineer these indigenous microbes for personalized therapy, by taking advantage of unique individual microbiota compositions [89]. To develop such next-generation microbial therapies, the novel design of engineered bacteria, development of platforms to study these microbes, and understanding mechanisms of bacterial colonization of various species can provide valuable insights towards clinical translation. With the fast-paced design of novel gene circuits to control bacteria’s ability to sense and respond to physiological conditions, the diverse capabilities of engineered bacteria may offer new therapeutic paradigms to treating a wide range of diseases.
Summary.
Bacterial cancer therapy offers unique advantages over traditional approaches including localized drug delivery, and intelligent sensing and responding module controlled by synthetic biology technologies.
Future efforts on enhancing safety and efficacy, developing characterization platforms, and understanding mechanisms of action, can accelerate clinical translation of bacterial therapies.
Funding
This work was supported in part by the Honjo International Foundation Scholarship (T.H.), R01GM069811 (T.D.), Bonnie J. Addario Lung Cancer Foundation Young Investigators Team Award (T.D.), Emerson Collective (T.D.), and DoD Era of Hope Scholar Award (BC160541) (T.D.).
Abbreviation
- FOLFIRI
fluorouracil, leucovorin, and irinotecan
Footnotes
Competing Interests
The Authors declare that there are no competing interests associated with the manuscript.
References
- 1.Gardner TS, Cantor CR and Collins JJ (2000) Construction of a genetic toggle switch in Escherichia coli. Nature 403, 339–342 10.1038/35002131 [DOI] [PubMed] [Google Scholar]
- 2.Elowitz MB and Leibler S (2000) A synthetic oscillatory network of transcriptional regulators. Nature 403, 335–338 10.1038/35002125 [DOI] [PubMed] [Google Scholar]
- 3.Basu S, Gerchman Y, Collins CH, Arnold FH and Weiss R (2005) A synthetic multicellular system for programmed pattern formation. Nature 434, 1130–1134 10.1038/nature03461 [DOI] [PubMed] [Google Scholar]
- 4.Friedland AE, Lu TK, Wang X, Shi D, Church G and Collins JJ (2009) Synthetic gene networks that count. Science 324, 1199–1202 10.1126/science.1172005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Danino T, Mondragón-Palomino O, Tsimring L and Hasty J (2010) A synchronized quorum of genetic clocks. Nature 463, 326–330 10.1038/nature08753 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sheth RU, Yim SS, Wu FL and Wang HH (2017) Multiplex recording of cellular events over time on CRISPR biological tape. Science 358, 1457–1461 10.1126/science.aao0958 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Farzadfard F and Lu TK (2014) Synthetic biology. Genomically encoded analog memory with precise in vivo DNA writing in living cell populations. Science 346, 1256272 10.1126/science.1256272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Park M, Patel N, Keung AJ and Khalil AS (2019) Engineering epigenetic regulation using synthetic read-write modules. Cell 176, 227–238.e20 10.1016/j.cell.2018.11.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tabor JJ, Salis HM, Simpson ZB, Chevalier AA, Levskaya A, Marcotte EM et al. (2009) A synthetic genetic edge detection program. Cell 137, 1272–1281 10.1016/j.cell.2009.04.048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Païssé S, Valle C, Servant F, Courtney M, Burcelin R, Amar J et al. (2016) Comprehensive description of blood microbiome from healthy donors assessed by 16S targeted metagenomic sequencing. Transfusion 56, 1138–1147 10.1111/trf.13477 [DOI] [PubMed] [Google Scholar]
- 11.Dickson RP, Erb-Downward JR and Huffnagle GB (2013) The role of the bacterial microbiome in lung disease. Expert Rev. Respir. Med 7, 245–257 10.1586/ers.13.24 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Urbaniak C, Gloor GB, Brackstone M, Scott L, Tangney M, Reid G et al. (2016) The microbiota of breast tissue and its association with breast cancer. Appl. Environ. Microbiol 82, 5039–5048 10.1128/AEM.01235-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Riglar DT, Giessen TW, Baym M, Kerns SJ, Niederhuber MJ, Bronson RT et al. (2017) Engineered bacteria can function in the mammalian gut long-term as live diagnostics of inflammation. Nat. Biotechnol 35, 653–658 10.1038/nbt.3879 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Daeffler KN-M, Galley JD, Sheth RU, Ortiz-Velez LC, Bibb CO, Shroyer NF et al. (2017) Engineering bacterial thiosulfate and tetrathionate sensors for detecting gut inflammation. Mol. Syst. Biol 13, 923 10.15252/msb.20167416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hwang IY, Koh E, Wong A, March JC, Bentley WE, Lee YS et al. (2017) Engineered probiotic Escherichia coli can eliminate and prevent Pseudomonas aeruginosa gut infection in animal models. Nat. Commun 8, 15028 10.1038/ncomms15028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mao N, Cubillos-Ruiz A, Cameron DE and Collins JJ (2018) Probiotic strains detect and suppress cholera in mice. Sci. Transl. Med 10, eaao2586 10.1126/scitranslmed.aao2586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liu Y, Bai P, Woischnig AK, Charpin-El Hamri G, Ye H, Folcher M et al. (2018) Immunomimetic designer cells protect mice from MRSA infection. Cell 174, 259–270.e11 10.1016/j.cell.2018.05.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Teixeira AP and Fussenegger M (2017) Synthetic biology-inspired therapies for metabolic diseases. Curr. Opin. Biotechnol 47, 59–66 10.1016/j.copbio.2017.06.004 [DOI] [PubMed] [Google Scholar]
- 19.Isabella VM, Ha BN, Castillo MJ, Lubkowicz DJ, Rowe SE, Millet YA et al. (2018) Development of a synthetic live bacterial therapeutic for the human metabolic disease phenylketonuria. Nat. Biotechnol 36, 857–864 10.1038/nbt.4222 [DOI] [PubMed] [Google Scholar]
- 20.Roybal KT, Rupp LJ, Morsut L, Walker WJ, McNally KA, Park JS et al. (2016) Precision tumor recognition by T cells with combinatorial antigen-sensing circuits. Cell 164, 770–779 10.1016/j.cell.2016.01.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Anderson JC, Clarke EJ, Arkin AP and Voigt CA (2006) Environmentally controlled invasion of cancer cells by engineered bacteria. J. Mol. Biol 355, 619–627 10.1016/j.jmb.2005.10.076 [DOI] [PubMed] [Google Scholar]
- 22.Zhou S, Gravekamp C, Bermudes D and Liu K (2018) Tumour-targeting bacteria engineered to fight cancer. Nat. Rev. Cancer 18, 727–743 10.1038/s41568-018-0070-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Coley WB II (1891) Contribution to the knowledge of sarcoma. Ann. Surg 14, 199–220 10.1097/00000658-189112000-00015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chien T, Doshi A and Danino T (2017) Advances in bacterial cancer therapies using synthetic biology. Curr. Opin. Syst. Biol 5, 1–8 10.1016/j.coisb.2017.05.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Brader P, Stritzker J, Riedl CC, Zanzonico P, Cai S, Burnazi EM et al. (2008) Escherichia coli Nissle 1917 facilitates tumor detection by positron emission tomography and optical imaging. Clin. Cancer Res 14, 2295–2302 10.1158/1078-0432.CCR-07-4254 [DOI] [PubMed] [Google Scholar]
- 26.Leschner S, Westphal K, Dietrich N, Viegas N, Jablonska J, Lyszkiewicz M et al. (2009) Tumor invasion of Salmonella enterica serovar Typhimurium is accompanied by strong hemorrhage promoted by TNF-α. Plos One 4, e6692 10.1371/journal.pone.0006692 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Roberts NJ, Zhang L, Janku F, Collins A, Bai R-Y, Staedtke V et al. (2014) Intratumoral injection of Clostridium novyi-NT spores induces antitumor responses. Sci. Transl. Med 6, 249ra111 10.1126/scitranslmed.3008982 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.St Jean AT, Zhang M and Forbes NS (2008) Bacterial therapies: completing the cancer treatment toolbox. Curr. Opin. Biotechnol 19, 511–517 10.1016/j.copbio.2008.08.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Taniguchi SI, Fujimori M, Sasaki T, Tsutsui H, Shimatani Y, Seki K et al. (2010) Targeting solid tumors with non-pathogenic obligate anaerobic bacteria. Cancer Sci 101, 1925–1932 10.1111/j.1349-7006.2010.01628.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Xiang S, Fruehauf J and Li CJ (2006) Short hairpin RNA–expressing bacteria elicit RNA interference in mammals. Nat. Biotechnol 24, 697–702 10.1038/nbt1211 [DOI] [PubMed] [Google Scholar]
- 31.Zhao M, Yang M, Li X-M, Jiang P, Baranov E, Li S et al. (2005) Tumor-targeting bacterial therapy with amino acid auxotrophs of GFP-expressing Salmonella typhimurium. Proc. Natl Acad. Sci. U.S.A 102, 755–760 10.1073/pnas.0408422102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Forbes NS (2010) Engineering the perfect (bacterial) cancer therapy. Nat. Rev. Cancer 10, 785–794 10.1038/nrc2934 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Tsung K and Norton JA (2006) Lessons from Coley’s toxin. Surg. Oncol 15, 25–28 10.1016/j.suronc.2006.05.002 [DOI] [PubMed] [Google Scholar]
- 34.Dang LH, Bettegowda C, Huso DL, Kinzler KW and Vogelstein B (2001) Combination bacteriolytic therapy for the treatment of experimental tumors. Proc. Natl Acad. Sci. U.S.A 98, 15155–15160 10.1073/pnas.251543698 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Redelman-Sidi G, Glickman MS and Bochner BH (2014) The mechanism of action of BCG therapy for bladder cancer—a current perspective. Nat. Rev. Urol 11, 153–162 10.1038/nrurol.2014.15 [DOI] [PubMed] [Google Scholar]
- 36.Morales A, Eidinger D and Bruce AW (2002) Intracavitary bacillus Calmette–Guerin in the treatment of superficial bladder tumors. J. Urol 167, 891–894. discussion 893–895 10.1016/S0022-5347(02)80294-4 [DOI] [PubMed] [Google Scholar]
- 37.Low KB, Ittensohn M, Luo X, Zheng LM, King I and Pawelek JM (2004) Construction of VNP20009: a novel, genetically stable antibiotic-sensitive strain of tumor-targeting Salmonella for parenteral administration in humans. Methods Mol. Med 90, 47–60 10.1385/1-59259-429-8:47 [DOI] [PubMed] [Google Scholar]
- 38.Low KB, Ittensohn M, Le T, Platt J, Sodi S, Amoss M et al. (1999) Lipid A mutant Salmonella with suppressed virulence and TNFα induction retain tumor-targeting in vivo. Nat. Biotechnol 17, 37–41 10.1038/5205 [DOI] [PubMed] [Google Scholar]
- 39.Clairmont C, Lee KC, Pike J, Ittensohn M, Low KB, Pawelek J et al. (2000) Biodistribution and genetic stability of the novel antitumor agent VNP20009, a genetically modified strain of Salmonella typhimurium. J. Infect. Dis 181, 1996–2002 10.1086/315497 [DOI] [PubMed] [Google Scholar]
- 40.Luo X, Li Z, Lin S, Le T, Ittensohn M, Bermudes D et al. (2001) Antitumor effect of VNP20009, an attenuated Salmonella, in murine tumor models. Oncol Res 12, 501–508 10.3727/096504001108747512 [DOI] [PubMed] [Google Scholar]
- 41.Toso JF, Gill VJ, Hwu P, Marincola FM, Restifo NP, Schwartzentruber DJ et al. (2002) Phase I study of the intravenous administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J. Clin. Oncol 20, 142–152 10.1200/JCO.2002.20.1.142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Cunningham C and Nemunaitis J (2001) A phase I trial of genetically modified Salmonella typhimurium expressing cytosine deaminase (TAPET-CD, VNP20029) administered by intratumoral injection in combination with 5-fluorocytosine for patients with advanced or metastatic cancer. Protocol no: CL-017. Version: April 9, 2001. Hum. Gene Ther 12, 1594–1596 [PubMed] [Google Scholar]
- 43.Eccles SA and Welch DR (2007) Metastasis: recent discoveries and novel treatment strategies. Lancet 369, 1742–1757 10.1016/S0140-6736(07)60781-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Guan X (2015) Cancer metastases: challenges and opportunities. Acta Pharm. Sin. B 5, 402–418 10.1016/j.apsb.2015.07.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Braun MS and Seymour MT (2011) Balancing the efficacy and toxicity of chemotherapy in colorectal cancer. Ther. Adv. Med. Oncol 3, 43–52 10.1177/1758834010388342 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Fang L, Jiang Y, Yang Y, Zheng Y, Zheng J, Jiang H et al. (2016) Determining the optimal 5-FU therapeutic dosage in the treatment of colorectal cancer patients. Oncotarget 7, 81880–81887 10.18632/oncotarget.11980 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Grenon NN and Chan J (2009) Managing toxicities associated with colorectal cancer chemotherapy and targeted therapy: a new guide for nurses. Clin. J. Oncol. Nurs 13, 285–296 10.1188/09.CJON.285-296 [DOI] [PubMed] [Google Scholar]
- 48.Hammond WA, Swaika A and Mody K (2016) Pharmacologic resistance in colorectal cancer: a review. Ther. Adv. Med. Oncol 8, 57–84 10.1177/1758834015614530 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.DeVita VT Jr, and Chu E (2008) A history of cancer chemotherapy. Cancer Res 68, 8643–8653 10.1158/0008-5472.CAN-07-6611 [DOI] [PubMed] [Google Scholar]
- 50.Devita VT Jr, Serpick AA and Carbone PP (1970) Combination chemotherapy in the treatment of advanced Hodgkin’s disease. Ann. Intern. Med 73, 881–895 10.7326/0003-4819-73-6-881 [DOI] [PubMed] [Google Scholar]
- 51.Fakih MG (2015) Metastatic colorectal cancer: current state and future directions. J. Clin. Oncol 33, 1809–1824 10.1200/JCO.2014.59.7633 [DOI] [PubMed] [Google Scholar]
- 52.Larkin J, Chiarion-Sileni V, Gonzalez R, Grob JJ, Cowey CL, Lao CD et al. (2015) Combined nivolumab and ipilimumab or monotherapy in untreated melanoma. N. Engl. J. Med 373, 23–34 10.1056/NEJMoa1504030 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Michot JM, Bigenwald C, Champiat S, Collins M, Carbonnel F, Postel-Vinay S et al. (2016) Immune-related adverse events with immune checkpoint blockade: a comprehensive review. Eur. J. Cancer 54, 139–148 10.1016/j.ejca.2015.11.016 [DOI] [PubMed] [Google Scholar]
- 54.Pardoll DM (2012) The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 12, 252–264 10.1038/nrc3239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Chowdhury S, Castro S, Hinchliffe TE, Coker C, Arpaia N and Danino T (2019) Programmable bacteria induce durable tumor regression and systemic antitumor immunity. Nat. Med 25, 1057–1063 10.1038/s41591-019-0498-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Zheng JH, Nguyen VH, Jiang SN, Park SH, Tan W and Hong SH et al. (2017) Two-step enhanced cancer immunotherapy with engineered Salmonella typhimurium secreting heterologous flagellin. Sci. Transl. Med 9, eaak9537 10.1126/scitranslmed.aak9537 [DOI] [PubMed] [Google Scholar]
- 57.Swofford CA, Van Dessel N and Forbes NS (2015) Quorum-sensing Salmonella selectively trigger protein expression within tumors. Proc. Natl Acad. Sci. U.S.A 112, 3457–3462 10.1073/pnas.1414558112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K et al. (2016) Synchronized cycles of bacterial lysis for in vivo delivery. Nature 536, 81–85 10.1038/nature18930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Deyneko IV, Kasnitz N, Leschner S, Weiss S and Allen IC (2016) Composing a tumor specific bacterial promoter. PLoS One 11, e0155338 10.1371/journal.pone.0155338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Yu B, Yang M, Shi L, Yao Y, Jiang Q, Li X et al. (2012) Explicit hypoxia targeting with tumor suppression by creating an “obligate” anaerobic Salmonella typhimurium strain. Sci. Rep 2, 436. 10.1038/srep00436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Goers L, Freemont P and Polizzi KM (2014) Co-culture systems and technologies: taking synthetic biology to the next level. J. R. Soc. Interface 11, 20140065 10.1098/rsif.2014.0065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Riglar DT and Silver PA (2018) Engineering bacteria for diagnostic and therapeutic applications. Nat. Rev. Microbiol 16, 214–225 10.1038/nrmicro.2017.172 [DOI] [PubMed] [Google Scholar]
- 63.Fang Y and Eglen RM (2017) Three-dimensional cell cultures in drug discovery and development. SLAS Discov 22, 456–472 10.1177/1087057117696795 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Osswald A, Sun Z, Grimm V, Ampem G, Riegel K, Westendorf AM et al. (2015) Three-dimensional tumor spheroids for in vitro analysis of bacteria as gene delivery vectors in tumor therapy. Microb. Cell Fact 14, 199. 10.1186/s12934-015-0383-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Clevers H (2016) Modeling development and disease with organoids. Cell 165, 1586–1597 10.1016/j.cell.2016.05.082 [DOI] [PubMed] [Google Scholar]
- 66.Brackett EL, Swofford CA and Forbes NS (2016) Microfluidic device to quantify the behavior of therapeutic bacteria in three-dimensional tumor tissue. Methods Mol. Biol 1409, 35–48 10.1007/978-1-4939-3515-4_4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Kasinskas RW and Forbes NS (2006) Salmonella typhimurium specifically chemotax and proliferate in heterogeneous tumor tissue in vitro. Biotechnol. Bioeng 94, 710–721 10.1002/bit.20883 [DOI] [PubMed] [Google Scholar]
- 68.Kim HJ, Li H, Collins JJ and Ingber DE (2016) Contributions of microbiome and mechanical deformation to intestinal bacterial overgrowth and inflammation in a human gut-on-a-chip. Proc. Natl Acad. Sci. U.S.A 113, E7–E15 10.1073/pnas.1522193112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Harimoto T, Singer ZS, Velazquez OS, Zhang J, Castro S, Hinchliffe TE et al. (2019) Rapid screening of engineered microbial therapies in a 3D multicellular model. Proc. Natl Acad. Sci. U.S.A 116, 9002–9007 10.1073/pnas.1820824116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Raman V, Van Dessel N, O’Connor OM and Forbes NS (2019) The motility regulator flhDC drives intracellular accumulation and tumor colonization of Salmonella. J. Immunother. Cancer 7, 44 10.1186/s40425-018-0490-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Zhang H-Y, Man J-H, Liang B, Zhou T, Wang C-H, Li T et al. (2010) Tumor-targeted delivery of biologically active TRAIL protein. Cancer Gene Ther 17, 334–343 10.1038/cgt.2009.76 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Bevers RF, Kurth KH and Schamhart DHJ (2004) Role of urothelial cells in BCG immunotherapy for superficial bladder cancer. Br. J. Cancer 91, 607–612 10.1038/sj.bjc.6602026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Biot C, Rentsch CA, Gsponer JR, Birkhäuser FD, Jusforgues-Saklani H, Lemaître F et al. (2012) Preexisting BCG-specific T cells improve intravesical immunotherapy for bladder cancer. Sci. Transl. Med 4, 137ra72 10.1126/scitranslmed.3003586 [DOI] [PubMed] [Google Scholar]
- 74.Ho CL, Tan HQ, Chua KJ, Kang A, Lim KH, Ling KL et al. (2018) Engineered commensal microbes for diet-mediated colorectal-cancer chemoprevention. Nat. Biomed. Eng 2, 27–37 10.1038/s41551-017-0181-y [DOI] [PubMed] [Google Scholar]
- 75.Quispe-Tintaya W, Chandra D, Jahangir A, Harris M, Casadevall A, Dadachova E et al. (2013) Nontoxic radioactive Listeriaat is a highly effective therapy against metastatic pancreatic cancer. Proc. Natl Acad. Sci. U.S.A 110, 8668–8673 10.1073/pnas.1211287110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Chandra D, Jahangir A, Quispe-Tintaya W, Einstein MH and Gravekamp C (2013) Myeloid-derived suppressor cells have a central role in attenuated Listeria monocytogenes-based immunotherapy against metastatic breast cancer in young and old mice. Br. J. Cancer 108, 2281–2290 10.1038/bjc.2013.206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Pettenati C and Ingersoll MA (2018) Mechanisms of BCG immunotherapy and its outlook for bladder cancer. Nat. Rev. Urol 15, 615–625 10.1038/s41585-018-0055-4 [DOI] [PubMed] [Google Scholar]
- 78.Kamat AM, Colombel M, Sundi D, Lamm D, Boehle A, Brausi M et al. (2017) BCG-unresponsive non-muscle-invasive bladder cancer: recommendations from the IBCG. Nat. Rev. Urol 14, 244–255 10.1038/nrurol.2017.16 [DOI] [PubMed] [Google Scholar]
- 79.Kamat AM, Li R, O’Donnell MA, Black PC, Roupret M, Catto JW et al. (2018) Predicting response to intravesical bacillus Calmette–Guerin immunotherapy: are we there yet? A systematic review. Eur. Urol 73, 738–748 10.1016/j.eururo.2017.10.003 [DOI] [PubMed] [Google Scholar]
- 80.Nepple KG, Lightfoot AJ, Rosevear HM, O’Donnell MA and Lamm DL, Bladder Cancer Genitourinary Oncology Study Group (2010) Bacillus Calmette–Guerin with or without interferon α−2b and megadose versus recommended daily allowance vitamins during induction and maintenance intravesical treatment of nonmuscle invasive bladder cancer. J. Urol 184, 1915–1919 10.1016/j.juro.2010.06.147 [DOI] [PubMed] [Google Scholar]
- 81.Shore ND, Boorjian SA, Canter DJ, Ogan K, Karsh LI, Downs TM et al. (2017) Intravesical rAd-IFNα/Syn3 for patients with high-grade, bacillus Calmette–Guerin-refractory or relapsed non-muscle-invasive bladder cancer: a phase II randomized study. J. Clin. Oncol 35, 3410–3416 10.1200/JCO.2017.72.3064 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Gopalakrishnan V, Helmink BA, Spencer CN, Reuben A and Wargo JA (2018) The influence of the gut microbiome on cancer, immunity, and cancer immunotherapy. Cancer Cell 33, 570–580 10.1016/j.ccell.2018.03.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.García-González AP, Ritter AD, Shrestha S, Andersen EC, Yilmaz LS and Walhout AJM (2017) Bacterial metabolism affects the C. elegans response to cancer chemotherapeutics. Cell 169, 431–441.e8 10.1016/j.cell.2017.03.046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Jin C, Lagoudas GK, Zhao C, Bullman S, Bhutkar A, Hu B et al. (2019) Commensal microbiota promote lung cancer development via γδ T cells. Cell 176, 998–1013.e16 10.1016/j.cell.2018.12.040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Donia MS, Cimermancic P, Schulze CJ, Wieland Brown LC, Martin J, Mitreva M et al. (2014) A systematic analysis of biosynthetic gene clusters in the human microbiome reveals a common family of antibiotics. Cell 158, 1402–1414 10.1016/j.cell.2014.08.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Scott TA, Quintaneiro LM, Norvaisas P, Lui PP, Wilson MP, Leung K-Y et al. (2017) Host-microbe co-metabolism dictates cancer drug efficacy in C. elegans. Cell 169, 442–456.e18 10.1016/j.cell.2017.03.040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Landry BP and Tabor JJ (2017) Engineering diagnostic and therapeutic gut bacteria. Microbiol. Spectr 5 10.1128/microbiolspec.BAD-0020-2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Gopalakrishnan V, Spencer CN, Nezi L, Reuben A, Andrews MC, Karpinets TV et al. (2018) Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science 359, 97–103 10.1126/science.aan4236 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Lam KN, Alexander M and Turnbaugh PJ (2019) Precision medicine goes microscopic: engineering the microbiome to improve drug outcomes. Cell Host Microbe 26, 22–34 10.1016/j.chom.2019.06.011 [DOI] [PMC free article] [PubMed] [Google Scholar]