

Abstract
Bifidobacteria have attracted significant attention because they provide health-promoting effects in the human gut. In this review, we present a current overview of the three-dimensional structures of bifidobacterial proteins involved in carbohydrate uptake, degradation, and metabolism. As predominant early colonizers of the infant's gut, distinct bifidobacterial species are equipped with a panel of transporters and enzymes specific for human milk oligosaccharides (HMOs). Interestingly, Bifidobacterium bifidum and Bifidobacterium longum possess lacto-N-biosidases with unrelated structural folds to release the disaccharide lacto-N-biose from HMOs, suggesting the convergent evolution of this activity from different ancestral proteins. The crystal structures of enzymes that confer the degradation of glycans from the mucin glycoprotein layer provide a structural basis for the utilization of this sustainable nutrient in the gastrointestinal tract. The utilization of several plant dietary oligosaccharides has been studied in detail, and the prime importance of oligosaccharide-specific ATP-binding cassette (ABC) transporters in glycan utilisations by bifidobacteria has been revealed. The structural elements underpinning the high selectivity and roles of ABC transporter binding proteins in establishing competitive growth on preferred oligosaccharides are discussed. Distinct ABC transporters are conserved across several bifidobacterial species, e.g. those targeting arabinoxylooligosaccharide and α-1,6-galactosides/glucosides. Less prevalent transporters, e.g. targeting β-mannooligosaccharides, may contribute to the metabolic specialisation within Bifidobacterium. Some bifidobacterial species have established symbiotic relationships with humans. Structural studies of carbohydrate-utilizing systems in Bifidobacterium have revealed the interesting history of molecular coevolution with the host, as highlighted by the early selection of bifidobacteria by mucin and breast milk glycans.
Keywords: ABC transport proteins, bifidobacterium, carbohydrate metabolism, glycoside hydrolase, host–microbe interactions, human gut microbes
Introduction
The human gut microbiota (HGM) is a key determinant of the host's health [1–3]. Members of the Bifidobacterium genus have attracted significant attention as health-promoting ‘probiotic’ HGM members [4]. Bifidobacteria colonise the guts of social insects (e.g. honey bees) and mammals [5–7], which is supported by co-evolutionary adaptation to these niches, notably, the human gut [8]. Bifidobacteria are saccharolytic, relying on the metabolism of dietary and/or host-derived carbohydrates. Together with other key glycan degraders e.g. Bacteroidaceae, bifidobacteria have maintained symbiosis with their hosts during hominid evolution over 15 million years [9]. The central role of glycan catabolism in this successful adaptation [10,11] is consistent with the abundance of Bacteroides and Bifidobacterium [12]. Polysaccharide utilization loci (PULs) that encode TonB-dependent oligosaccharide transporters, transcriptional regulators and one or several outer-membrane attached and periplasmic carbohydrate-active enzymes (CAZymes) that target a specific glycan have been proposed to promote human colonisation by Bacteroides [13,14]. TonB transporters consist of an integral membrane pore (SusC) and extracellular lipid-anchored glycan-binding protein (SusD) that confers oligosaccharide capture and internalization via SusC [15]. Similarly, genes for carbohydrate utilization and ATP-binding cassette (ABC) transporters are often colocalised in bifidobacteria [16]. ABC transporters consist of dimeric transmembrane domains that form the translocation pore, two cytoplasmic nucleotide-binding domains that energise the uptake, and an extracellular MalE-type substrate-binding protein (SBP, or solute-binding protein) for high specificity and affinity ligand-capture [17]. This review summarizes the current knowledge and recent advances in the bifidobacterial carbohydrate uptake and metabolism apparatus with a focus on structural studies. The evolutionary origins of this protein machinery are also surmised based on structural comparisons of homologs.
Catabolism of digestible carbohydrates
The allosteric l-lactate dehydrogenase, which catalyses the last step of glycolysis, from Bifidobacterium longum aM101-2 was the first to be studied from this genus [18]. The structure of 1 : 1 complex of R-state (relaxed, high substrate affinity) and T-state (tense, no substrate affinity) tetramers from a single crystal was reported (Figure 1A) [19,20]. Binding of the allosteric activator, fructose 1,6-bisphosphate, triggers a quaternary structural change of the tetramer, and the substrate affinity is controlled by helix sliding between subunits. Interestingly, bifidobacteria adopt a unique fermentation pathway called the ‘bifid’ shunt [21]. The key bifid shunt enzyme is phosphoketolase, which catalyses phosphorolytic cleavage of fructose 6-phosphate or xylulose 5-phosphate to aldose phosphate, acetyl phosphate, and H2O. The crystal structures of phosphoketolases from B. longum JCM 1217 and Bifidobacterium breve 203 were determined [22–24] (Figure 1B). Phosphoketolase is dependent on thiamine diphosphate coenzyme and catalyses dehydration and phosphorylation reactions. The reaction intermediates were captured by crystal structures, providing the structural basis for this unique catalysis [24].
Figure 1. Enzymes for fermentation and digestible carbohydrates.

(A) Allosteric l-lactate dehydrogenase from B. longum aM101-2 (PDB: 1LTH). R-state (green, high substrate affinity) and T-state (cyan, no substrate affinity) tetramers are packed in a single crystal. NADH, FBP, and oxamate (substrate analogue) are shown as yellow, blue, and magenta spheres, respectively. (B) Phosphoketolase from B. breve 203. Left panel, the overall dimer structure (PDB: 3AHC) is shown with thiamine diphosphate (ThDP) as yellow spheres. Right panel, the active site is shown as a composite of α,β-dihydroxyethyl ThDP (yellow, PDB: 3AHD) and 2-acetyl-ThDP (green, PDB: 3AHE) intermediates and phosphate (PDB: 3AHF). (C) GH13 sucrose phosphorylase from B. adolescentis DSM20083. Left panel, overall structure (PDB: 2GDU) is shown with sucrose as yellow spheres. Right panel, the active site is shown as a superimposition of a sucrose complex (yellow, PDB: 2GDU) and a covalent glucosyl-enzyme intermediate (green, PDB: 2GDV). The catalytic acid/base residue (Glu232) was mutated to glutamine to obtain the complex with sucrose. (D) The overall structure of GH32 β-fructofuranosidase from B. longum KN29.1 is shown with β-fructofuranose as yellow spheres (PDB: 3PIJ).
The first structure of a bifidobacterial glycoside hydrolase (GH) assigned in the Carbohydrate-Active enZymes (CAZy) database [25] was the GH13 sucrose phosphorylase from Bifidobacterium adolescentis DSM20083 (Figure 1C) [26]. The reaction mechanism of this enzyme, involving the covalent enzyme-glucosyl intermediate formation and nucleophilic attack by phosphate, was proposed [27]. Concerning the metabolism of fructose-containing carbohydrates, the crystal structure of the GH32 β-fructofuranosidase from B. longum KN29.1, which has a typical GH32-type catalytic domain with a five-bladed β-propeller fold, was reported (Figure 1D) [28]. This enzyme releases fructose residues from sucrose, 1-kestose, nystose, inulin, and raffinose in vitro. Inulin-type fructans are abundant storage polysaccharides and common dietary fibres. The putative intracellular localisation of the β-fructofuranosidase suggests that inulin oligomers are the likely physiologically relevant substrates of this enzyme.
Import and degradation of host-derived glycans
Common metabolic pathway of HMOs and mucin glycans in infant gut-associated bifidobacteria
Several Bifidobacterium species, e.g. B. longum subsp. longum (B. longum), B. longum subsp. infantis (B. infantis), B. breve, and Bifidobacterium bifidum are prevalent in infant's guts [6]. Bifidobacterium, which is the most abundant genus of the infant gut microbiota, is associated with beneficial effects on infant health [29]. The symbiotic relationship between humans and infant gut-associated bifidobacteria is mediated through mother's milk oligosaccharides (human milk oligosaccharides, HMOs) [30]. HMOs are complex oligosaccharides composed of galactose (Gal), glucose (Glc), N-acetylglucosamine (GlcNAc), l-fucose (Fuc), and sialic acid (Neu5Ac) [31]. Lacto-N-biose I (Galβ1-3GlcNAc, LNB) is an abundant HMO building block and one of the key units of the symbiosis. The phosphorylase gene that catalyses the cleavage of LNB into α-galactose 1-phosphate and GlcNAc was initially isolated from B. longum JCM 1217, and this enzyme became the founding member of the GH112 family [32]. Because this enzyme is also highly active on galacto-N-biose (Galβ1-3GalNAc, GNB), it was termed GNB/LNB phosphorylase (GLNBP). GNB is a major core disaccharide unit of O-glycoproteins, which are present in human milk fat globules and mucins in gastrointestinal mucus (described below) [33]. The GH112 GLNBP gene is ubiquitous in infant gut-associated bifidobacteria that can grow on LNB [29,34]. The three-dimensional structure determination of GLNBP provided a hint for a possible evolutionary origin of this enzyme [35]. The overall structure and domain architecture of GLNBP are similar to those of GH42 β-galactosidases that are widely distributed in infant gut-associated bifidobacteria (Figure 2A) [36]. Interestingly, the catalytic components of GLNBP at the centre of the catalytic domain are superimposed with those of GH42 β-galactosidases: The phosphate-binding site and the catalytic acid (proton donor) of GLNBP (inverting phosphorylase) are positioned near the nucleophile and the acid/base residues of GH42 β-galactosidase (retaining hydrolase), respectively. Therefore, a possible evolutionary relationship between these galactoside-cleaving enzymes was suggested [35]. The catalytic domain of GLNBP was suggested to undergo a closure motion upon binding of phosphate, which involved a large and unusual deformation of the (β/α)8 barrel fold. Such an intricate adaptation to the substrate by a deformation of the (β/α)8 fold instead of variations in the loops is a unique case of molecular evolution.
Figure 2. Enzymes and the transporter solute binding protein in the LNB/GNB pathway of B. longum JCM 1217.
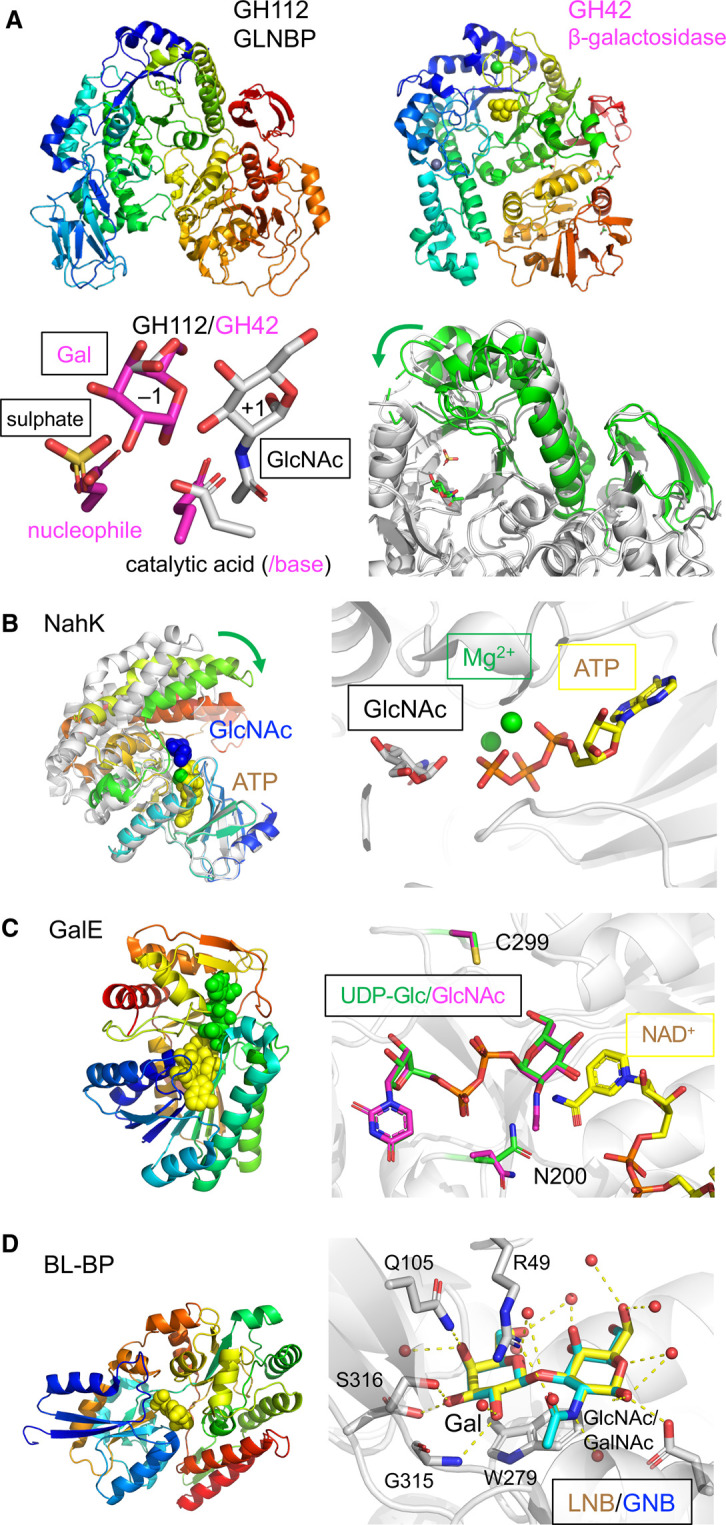
(A) The GH112 GLNBP (upper left, PDB: 2ZUS) and the GH42 β-galactosidase (upper right, PDB: 1KWK) with galactose shown as yellow spheres. Bottom left panel, superimposition of the active sites of the GH112 GLNBP (white, composite of PDB: 2ZUV and 2ZUW) and the GH42 β-galactosidase (magenta, PDB: 1KWK). Bottom right panel, the closing motion of the (β/α)8 barrel of GLNBP is shown as a superimposition of open (white, PDB: 2ZUS) and closed (green, PDB: 2ZUV) states. (B) N-acetylhexosamine 1-kinase NahK. Left panel, the closing motion of NahK is shown with open (white, PDB: 4WH3) and closed (rainbow colour, PDB: 4OCJ) states. ATP, Mg2+, and GlcNAc are shown as yellow, green and blue spheres, respectively. Right panel, the active site is shown as a composite of GlcNAc (white sticks) and ATP (yellow sticks) plus Mg2+ ions (green spheres). (C) UDP-glucose 4-epimerase GalE. Left panel, overall structure (PDB: 6K0H) is shown with NAD+ and UDP-Glc as yellow and green spheres, respectively. Right panel, the active site is shown as a superimposition of the complex structures with UDP-Glc (green) and UDP-GlcNAc (magenta). NAD+ is shown as yellow sticks. (D) The transporter GNB/LNB-binding protein (GL-BP). Left panel, overall structure (PDB: 2Z8D) is shown with LNB as yellow spheres. Right panel, the substrate-binding site is shown as a superimposition of the complex structures of LNB (yellow, PDB: 2Z8D) and GNB (cyan, PDB: 2Z8E).
Along with GLNBP, the gene cluster for LNB/GNB metabolism is composed of an ABC transporter and three intracellular enzymes that convert the cleavage products of GLNBP into glycolysis precursors: N-acetylhexosamine 1-kinase (NahK), UDP-glucose 4-epimerase (GalE), and UDP-glucose–hexose-1-phosphate uridylyltransferase (GalT) [37]. NahK phosphorylates GlcNAc or GalNAc while galactokinase in the Leloir pathway (a common galactose metabolism route) is specific for galactose [37]. Remarkably, the crystal structure of NahK was not structurally similar to galactokinase that also phosphorylates the sugars at the anomeric C1 position [38]. Note that the first report of NahK structure wrongly designates its name as ‘N-acetylhexosamine 1-phosphate kinase’. A large open-close conformational change in NahK was observed, and two Mg2+ in the ATP-binding site facilitate the catalysis (Figure 2B) [39,40]. GalE catalyses the interconversion of gluco- and galacto-hexoses (C4 epimerization) linked to UDP. Crystal structures of GalE from B. longum were determined in a complex with UDP-Glc and UDP-GlcNAc (Figure 2C) [41]. GalE has broad substrate specificity due to the large pocket formed by Cys299 and the swing-in/out motion of Asn200. Interestingly, GalE from B. longum exhibits functional and structural similarities to the corresponding enzyme from the human host [42]. The bifidobacterial GalT belongs to a minor class of the GalT family (class II) [43], which remains structurally uncharacterized. The SBP of the ABC transporter in the LNB/GNB pathway was also characterized in detail [44]. Since this SBP was highly specific for LNB and GNB, it was termed the GNB/LNB-binding protein (GL-BP). GL-BP has a similar fold with the maltose binding protein (SBP fold), and both LNB as well as GNB are extensively recognized by the protein (Figure 2D) [44].
Divergent strategies of HMO utilization
Bifidobacteria, which generally possess fewer extracellular enzymes than Bacteroides, rely on ABC transporters to import dietary oligosaccharides. The infant gut-associated B. infantis and B. breve use ABC transporters for uptake and subsequent intracellular digestion of HMOs [29,45]. B. infantis JCM 1222 has two ABC transporters for uptake of the major HMOs, fucosyllactose (FL). The prevalent FL transporter-2 has a broader specificity than the less common counterpart [46]. The crystal structures of the FL transporter-2 SBP complexed with 2’-FL and 3-FL revealed the recognition of a disaccharide Fuc-[Gal/Glc] motif, with the Fuc unit residing at the bottom of the binding cleft (Figure 3A). These findings rationalise the basis for the dual transporter specificity.
Figure 3. Enzymes and SBP for HMO utilization by infant gut-associated bifidobacteria.
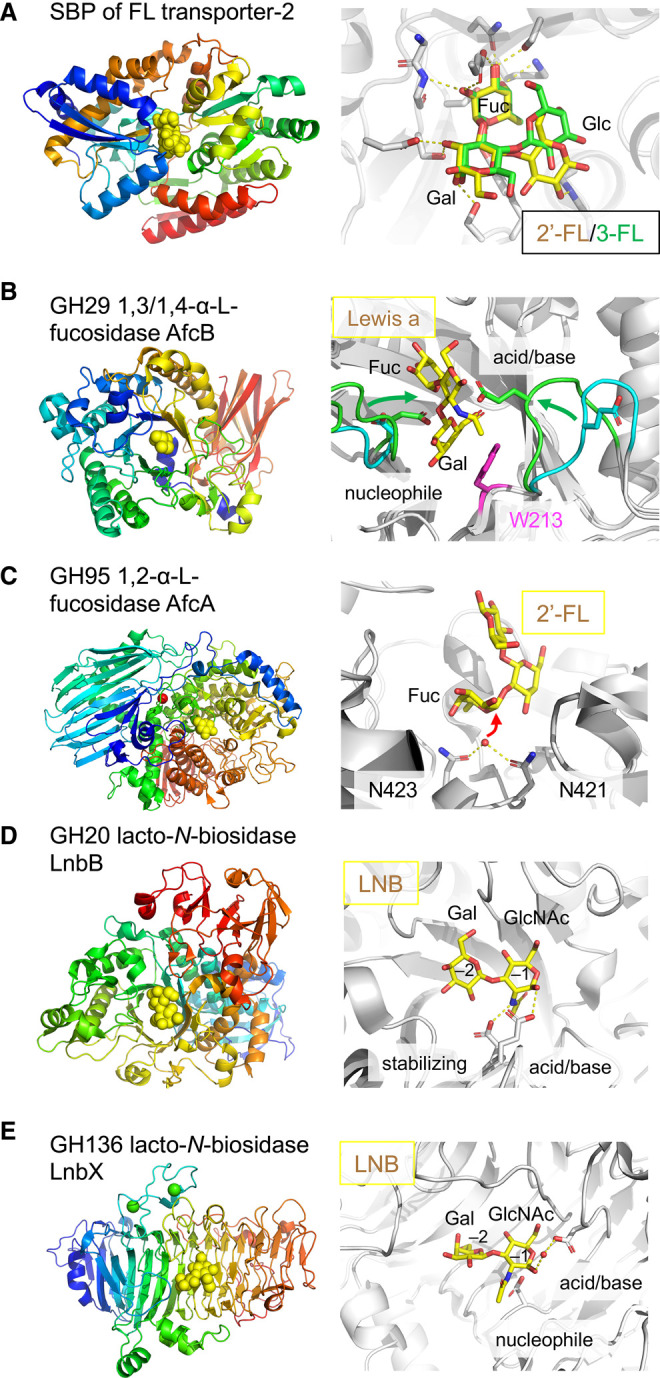
(A) Fucosyllactose binding protein (SBP) of the FL transporter-2 from B. infantis JCM 1222. Left panel, overall structure (PDB: 6HUR) shown with 2′-fucosyllactose (2′-FL) as yellow spheres. Right panel, the substrate-binding site is shown as a superimposition of the complex structures with 2′-fucosyllactose (2′-FL, yellow, PDB: 6HUR) and 3-fucosyllactose (3-FL, green, PDB: 6HUS). (B) GH29 1,3/1,4-α-L-fucosidase AfcB from B. infantis ATCC 15697. Left panel, overall structure (PDB: 3UET) is shown with fucose as yellow spheres. Right panel, the induced-fit motion of the active site loops shown as a superimposition of apo (cyan, PDB: 3MO4) and complex (green, PDB: 3UET) structures with Lewis a trisaccharide (yellow sticks). The side chains of the catalytic residues (green sticks) were modelled from the D172A/E217A double mutant structure. The aromatic stacking platform for Gal (Trp213, magenta) is also shown. (C) GH95 1,2-α-L-fucosidase AfcA from B. bifidum JCM 1254. Left panel, overall structure (PDB: 2EAC) is shown with deoxyfuconojirimycin as yellow spheres. Right panel, the active site is shown as a composite of the 2′-FL complex (PDB: 2EAD) and the wild-type enzyme (PDB: 2EAC). 2′-FL, two catalytic asparagine residues, and the nucleophilic water are shown as yellow sticks, white sticks, and a red sphere, respectively. (D) GH20 lacto-N-biosidase LnbB from B. bifidum JCM 1254. Left panel, overall structure (PDB: 4H04) is shown with LNB as yellow spheres. Right side, the active site showing LNB. (E) GH136 lacto-N-biosidase LnbX from B. longum JCM 1217. Left panel, overall structure (PDB: 5GQF) is shown with LNB as yellow spheres. Right side, the active site showing LNB. The side chains of the catalytic residues are shown in (D) and (E).
The intracellular GH29 1,3/1,4-α-L-fucosidase in B. infantis ATCC15697 (AfcB) effectively cleaves the 3-fucosyl and 4-fucosyl substitutions of HMOs [47]. The crystal structure of AfcB revealed a prominent galactose-binding site that contributes to selective binding of the Lewis a and x epitopes in HMOs or other host glycans (Figure 3B) [48]. AfcB exhibits unique induced-fit loop motions to position the catalytic nucleophile and acid/base residues [48].
Compared with the two infant gut-associated species described above, B. longum and B. bifidum adopt an ‘extracellular digestion’ strategy [49]. In particular, B. bifidum strains behave as altruistic members in the gut microbiota ecosystem by sharing HMO degradants produced by their extracellular glycosidases [50,51]. For example, the GH95 1,2-α-l-fucosidase from B. bifidum JCM 1254 (AfcA) specifically cleaves the α-1,2-linked fucosyl residues of HMOs or related host glycans [52]. The overall structure of AfcA (anomer-inverting GH, Figure 3C) exhibited a striking resemblance to inverting glycoside phosphorylases of GH65 and GH94 [53]. The active site of AfcA has a unique feature involving two asparagine residues that were suggested to activate the water nucleophile [53]. For the degradation of LNB-containing (type 1) HMOs, B. bifidum has a key extracellular enzyme called lacto-N-biosidase (LnbB), which belongs to GH20 and hydrolyses the central β1,3 bond of lacto-N-tetraose to produce LNB and lactose [54]. The structure of LnbB (Figure 3D) differ from canonical monosaccharide-releasing GH20 exo-β-N-acetylhexosaminidases [55] by having an extra –2 subsite that allows the accommodation of the nonreducing galactosyl in LNB. The N-acetyl group of GlcNAc adopted a distorted conformation to be poised for nucleophilic attack on the anomeric C1 atom, consistent with the substrate-assisted mechanism in GH20.
B. longum has a special adaptation among infant gut-associated bacteria because it is also widely distributed in the adult and elderly human gut [56]. The recently discovered extracellular lacto-N-biosidase from B. longum JCM 1217 (LnbX) is not homologous to GH20 LnbB and thus represents the founding member of GH136 [57]. LnbX adopts a β-helix fold (Figure 3E) and a retaining mechanism [58]. The active site pocket architecture of LnbX is distinct from other polymer-acting β-helix endo-glycosidases, which have a long cleft. Although the evolutionary origin of LnbX is enigmatic, the protein fold has significant structural similarity to tailspike proteins from bacteriophages, which occasionally possess carbohydrate-binding ability [58]. GH136 LnbX is inactive unless it is coexpressed with an adjacent gene. The homologous lacto-N-biosidase from Eubacterium ramulus indicated that GH136 enzymes are likely to occur either as heterodimers or as two-domain enzymes, with the second domain/subunit adopting a previously unknown helical fold and contributing to the active site architecture [59].
Mucin degradation
The mucus layer barrier that protects the epithelium serves also as a sustainable nutrient supply for distinct bacteria via its mucin glycoproteins [60]. Long-term coevolution was likely established due to a mutually beneficial relationship between the human host and the mucin-adherent microbiota [61]. Strains of B. longum, B. bifidum, and B. breve possess glycosidases targeting O-linked mucin glycans [62,63]. The three-dimensional structures of two bifidobacterial mucin-degrading enzymes have been reported. The extracellular GH101 endo-α-N-acetylgalactosaminidase from B. longum JCM 1217 (EngBF) releases the α-linked GNB disaccharide from the Ser or Thr residues in the core 1 units (or T-antigen) [64], which allows for GNB catabolism through the LNB/GNB pathway. The catalytic domain of EngBF has a partially broken (β/α)8 barrel similar to GH13 α-amylases (Figure 4A) [65]. Two active site tryptophans stack onto GNB by an induced-fit motion [66]. The second mucin-degrading enzyme structure was determined for the GH129 α-N-acetylgalactosaminidase from B. bifidum JCM 1254 (NagBb) [67], which releases the α-linked GalNAc from the Tn antigen [68]. The catalytic domains of NagBb and GH101 enzymes are similar (Figure 4B). Recognition of the GalNAc is supported by Ca2+ and stacking-platform of a tryptophan.
Figure 4. Enzymes for mucin degradation.

(A) GH101 endo-α-N-acetylgalactosaminidase EngBF from B. longum JCM 1217. Left panel, overall structure (PDB: 2ZXQ). Right panel, the active site showing GNB modelled by automated docking (yellow sticks). The two tryptophan residues (magenta) form aromatic stacking interactions with the disaccharide. (B) GH129 α-N-acetylgalactosaminidase NagBb from B. bifidum JCM 1254. Left panel, overall structure (PDB: 5WZN) is shown with GalNAc and Ca2+ as yellow and green spheres. GH101 EngBF (grey) is overlayed to show the structural similarity. Right panel, the active site showing GalNAc (yellow sticks), Ca2+ (green sphere), waters, and protein ligands of Ca2+. The aromatic stacking platform for GalNAc (Trp398, magenta) is also shown. The side chains of the catalytic residues are also shown in (A) and (B).
Import and degradation of plant glycans
Exposure to complex glycans from vegetables and fruits triggers a pronounced expansion of the infant HGM diversity during weaning [69]. The abundance of bifidobacteria decreases after weaning, but plant dietary glycan utilizers remain relatively abundant in adults. Distinct bifidobacteria, e.g. Bifidobacterium animalis subsp. lactis (B. lactis) and B. longum subsp. longum, are commonly used commercial probiotics. This section focuses on the bifidobacterial proteins that target abundant dietary plant saccharides.
Common pathways for the utilization of plant dietary saccharides
Arabinoxylan is a major dietary fibre from cereal cell walls. Only a few primary degraders from Bacteroides [70], Roseburia intestinalis [71] and Eubacterium can degrade xylan. Bifidobacteria have a conserved genetic locus that confers growth on arabinoxylan oligomers (arabinoxylooligosaccharides, AXOS). This locus is common in the B. longum, B. adolescentis, and Bifidobacterium pseudolongum (that includes B. lactis) groups [72]. The AXOS locus, which encodes an ABC transporter, esterases, arabinofuranosidases, xylosidases and xylose catabolism enzymes, was shown to be transcriptionally upregulated in B. lactis grown on XOS [73]. The binding protein (BlAXBP) of this transporter has a preference for a xylotetraose (Kd ≈ 100 nM) and terminal nonreducing end arabinosyl substitutions [72]. Both arabinoxylobiose and arabinoxylotriose were bound with Kd ≈ 100 nM. The crystal structures of BlAXBP in complex with xylotriose, xylotetraose, 32-α-l-arabinofuranosyl-xylobiose, and 23-α-l-arabinofuranosyl-xylotriose assume a canonical SBP fold (cluster B-I [74]) comprising two domains connected with a tripartite hinge region. The binding site of BlAXBP accommodates a xylotetraose backbone and a flexible lid-loop adjusts its conformation to bind different ligands (Figure 5A). The most densely recognized xylosyl is sandwiched between two tryptophans and recognized by Asp386 acid and His199, which define subsite 1 that is occupied in all structures (Figure 5B,C). Cavities flanking subsites 1 or 2 allow binding of C2 or C3-arabinosyl substituents. The AXOS backbone can be accommodated in two opposite directionalities, which expands the range of captured AXOS. Altogether, these features contribute to the versatility of AXOS capture.
Figure 5. SBPs for abundant plant-derived oligosaccharides by B. animalis subsp. lactis Bl-04.

(A) The overall structure of the arabinoxylooligosaccharide binding protein BlAXBP in complex with xylotetraose (PDB: 3ZKK). (B) and (C) Recognition of arabinoxylotriose (PDB: 4C1T) and arabinoxylobiose (PDB: 4CIU), respectively, with the amino acid residues coloured according to the domain colour as in (A). The arabinosyl sidechains (orange) can be accommodated in cavities in the binding protein, and the structure view in (C) is tilted approximately 90° for clarity. (D) The overall structure of the SBP BlG16BP that mediates the capture of both α-1,6-linked galactosides and glucosides in bound form with the preferred ligand panose (PDB: 4ZZE). (E,F) Details of binding panose and raffinose (PDB: 4ZS9), respectively. The recognition motif of BlG16BP is the disaccharide unit at positions 1 and 2, and the open binding site allows the accommodation of different monosaccharide units at position 3 (Glc and Fru at position 3 are shown in orange).
Raffinose family oligosaccharides from soybean and isomaltooligosaccharides from starch degradation, honey, or dextran breakdown are abundant in the human gut. Both oligosaccharide classes share a [Gal/Glc]α1,6Glc motif and promote the growth of bifidobacteria, except for B. bifidum [75]. The most transcriptionally upregulated locus during the growth of B. lactis [73] on both α1,6-glycosides is conserved in other bifidobacteria except B. bifidum, consistent with the observed growth profiles [75]. This locus encodes a transcriptional regulator, an α-galactosidase [76], a GH13_31 α-1,6-glucosidase [77], and an ABC transporter. Uptake of oligosaccharides sharing a [Gal/Glc]α1,6Glc via the ABC transporter [75] was corroborated by the crystal structures of the associated α1,6-glycoside binding protein (BlG16BP) in complex with raffinose (Galα1,6Glcα1,2βFru) and panose (Glcα1,6Glcα1,4Glc) (Figure 5D–F). Compared to AXOS binding in a cleft, the binding site of BlG16BP features a deep pocket [75]. Asp394 confers the dual recognition of galactosyl or glucosyl units at subsite 1 via hydrogen bonds to either the axial or equatorial C4-OH, respectively (Figure 5E,F). The C-terminal domain harbours an aromatic stacking platform and Asp394 and His395 that recognise the saccharide unit at position 1. The recognition of the glucosyl at subsite 2 in both panose and raffinose is identical, whereas the lack of direct polar bonds at subsite 3 is consistent with the plasticity at this site. B. lactis outcompeted Bacteroides ovatus after 18 h of growth on raffinose in a 50 : 50 starting mixed culture [75], which underscores the role of ABC transporters in conferring competitive growth on preferred substrates.
Specialization of distinct bifidobacteria on certain plant glycans
β-Mannans are structural and storage plant polysaccharides, which are heavily used as food additives. Primary degraders of mannan include Bacteroides ovatus [78] and R. intestinalis [79]. Mannan utilisation is not as widespread in Bifidobacterium as arabinoxylan or α1,6-glycosides discussed above. A predicted cell-attached GH26 mannanase from B. adolescentis [80] and a secreted GH5 mannanase from B. lactis have been biochemically characterized [81]. The growth of B. lactis ATCC 27673 on konjac glucomannan and galactomannan from carob bean gum has been reported [82]. Proteomic analysis revealed the upregulation of a GH5 mannanase, a putative β-mannosidase, a β-glucosidase, as well as an ABC transporter with two homologous SBPs during growth on mannan [82]. The affinity of one of these binding proteins (BlMnBP1) to mannobiose was 2500-fold higher compared to the second (BlMnBP2). Otherwise, both proteins had similar binding profiles with a preference for a mannotriose and binding of galactomannooligosaccharides with a galactosyl at the reducing mannosyl unit (Kd ≈ 2 μM), which is consistent with the growth on galactomannan. The structures of BlMnBP1 and BlMnBP2 in complex with mannobiose/mannotriose and mannotriose, respectively [82], revealed similar binding sites (Figure 6A,B). The mannosyl specificity is mediated by hydrogen bonds to the axial C2-OH at subsites 1 and 2, whereas a spacious cavity beyond subsite 3 accommodates galactosyl sidechains (Figure 6C). The most striking difference is the substitution of an asparagine (Asn63) that is hydrogen-bonded to the mannosyl at subsite 2 in BlMnBP1 to glycine in BlMnBP2 (Figure 6B). Mutational analyses supported the role of this residue in substrate binding. This study highlighted the differentiation of SBP specificities via duplication and one or several mutations that alter the affinity/specificity to distinct substrates.
Figure 6. Transporter SBPs and enzymes of specialized utilization for plant-derived glycans by B. animalis subsp. lactis.

(A) Structure of the mannooligosaccharide binding protein from B. animalis subsp. lactis ATCC 27673 (BlMnBP1) in complex with mannotriose (PDB: 6I5V). (B) Differences in the binding site of the two homologous binding proteins BlMnBP1 and BlMnBP2 (PDB: 6FUV) that display high and low binding affinity to mannobiose, respectively. The amino acid residues are coloured according to the domain colour in (A). BlMnBP1/BlMnBP2 amino acid identities and numbering are shown. Asn63 is present only in BlMnBP1 and interacts with the mannosyl at subsite 2. (C) Cross-sectional view of the electrostatic surface of BlMnBP1 cocrystallised with mannopentaose (PDB: 6I5W) reveals additional space and cavities beyond subsite +3, consistent with the binding profile of larger or galactosyl-decorated oligosaccharides. (D) and (E) show the overall structure of the binding protein Bal6GBP (PDB: 6H0H) in complex with β1,6-galactoside and the detailed recognition of this ligand, respectively. (F) Comparison of the active site subsite –1 of the recently discovered α-l-arabinopyranosidase BlArap42B (PDB: 5XB7) and the β-1,6/1,3-β-galactosidase BlGal42A (PDB: 4UNI) in complex with galactose. A key difference is the substitution of a histidine that recognizes the C6-OH with a hydrogen bond in BlGal42A with a tryptophan residue that makes a steric clash with the C6-OH and instead is likely to stack onto the arabinopyranose that lacks this group.
The role of oligosaccharide transporters has been also investigated in the specialization of B. lactis Bl-04 towards β1,6-galactooligosaccharides. The preferred β1,6-galactobiose conferred rapid and immediate growth of B. lactis Bl-04, while the less preferred β1,3 and β1,4 isomers resulted in several hours-long lag phases. The galactooligosaccharide locus in this strain comprises an ABC transporter and a GH42 β-galactosidase (BlGH42A) [73], which was shown to prefer β1,6-galactobiose, thus defining a new specificity in GH42 [83]. This enzyme had comparable catalytic efficiencies (within 5-fold) towards the β1,3- and the β1,4-galactobiose compared to the preferred β1,6-isomer. Galactose binding at subsite -1 induced a conformational change in the loop region Asn202-Gln206, allowing the formation of a hydrogen bond to the C4-OH of the bound galactose.
In contrast to the enzymatic promiscuity of BlGH42A, the ABC transporter's SBP (Bal6GBP) is strikingly selective towards β1,6-galactobiose (Kd = 92 nM), with 300- and 1630-fold lower affinities towards the β1,3- and β1,4-linked isomers, respectively [84]. The structure of Bal6GBP was determined in complex with β1,6-galactobiose (Figure 6D). The β1,6-galactobiose ligand is recognized by 12 potential hydrogen bonds, with all the OH-groups of the nonreducing galactosyl (subsite 1) being engaged in hydrogen bonds (Figure 6E). Interestingly, galactosyl binding at subsite 1 is conserved in Bal6GBP and the B. longum LNB binding protein (GL-BP) (Figures 2D and 6E). The substitution of Trp63 in Bal6GBP with Arg49 in GL-BP from B. longum correlates with the change in specificity at position 2 from Gal in Bal6GBP to GlcNAc in GL-BP.
The catabolism of plant-derived bioactive glycosides by the HGM exerts an important impact on human health. The mechanisms of this metabolic facet remain largely unexplored except in a few studies e.g. in Lactobacillus acidophilus [85]. Recently, a new GH42 α-l-arabinopyranosidase from B. lactis (BlArap42B) was discovered [86]. BlArap42B releases α-l-arabinopyranoside from bioactive plant glycosides, e.g. paenolide or ginsenoside Rb2. This specificity reflects the structural similarity between α-l-arabinopyranoside and β-d-galactopyranoside. The only difference is the loss of C6-OH in the former. Indeed, the space occupied by the C6-OH in β-galactosidases is blocked with the indole side chain of Trp358 in BlArap42B (Figure 6F). These active site differences revealed the signatures underpinning the diversification of specificities with GH42.
β-Arabinooligosaccharides
The diverse human diet promotes the evolution of unique pathways in bifidobacteria to specialise on less common plant-derived carbohydrates. B. longum JCM 1217 possesses a gene cluster encoding the utilisation of β-linked arabinofuranooligosaccharides [87]. β-Arabinoligosaccharides are present in plant cell wall components called hydroxyproline-rich glycoproteins, including expansin and lectins from edible plants. The utilisation system comprises an extracellular enzyme that releases β-1,2-linked arabinofuranose disaccharide (β-Ara2; GH121 β-l-arabinobiosidase HypBA2), which is internalised by an ABC transporter and an intracellular enzyme that cleaves the disaccharide into monomers (GH127 β-l-arabinofuranosidase HypBA1) [88]. The catalytic domain of GH121 HypBA2 adopts an (α/α)6 barrel fold (Figure 7A), similar to several GH families [89]. Three acidic (glutamate and aspartate) residues in the active site pocket are essential for catalysis. The SBP of the ABC transporter (β-arabinobiose-binding protein) exclusively binds β-Ara2 via extensive interactions, and its dynamic feature was investigated by computational simulation (Figure 7B) [90]. The crystal structure of the retaining GH127 HypBA1 revealed an unprecedented ‘cysteine glycoside hydrolase’, which uses a cysteine residue as the catalytic nucleophile (Figure 7C) [91]. Although its catalytic domain has a typical (α/α)6 barrel fold, the catalytic site has a unique architecture composed of a Zn2+ coordinated by (Cys)3-Glu. Cys417 is closely positioned to the anomeric C1 of β-l-arabinofuranose and identified as the possible nucleophile. A detailed mechanistic study using a cyclophellitol-derived inhibitor revealed the covalent glycosyl-enzyme intermediate structure and the itinerary of the catalytic reaction of HypBA1 [92].
Figure 7. Enzymes and SBP for β-arabinooligosaccharides utilization by B. longum JCM 1217.

(A) Overall structure of GH121 β-l-arabinobiosidase HypBA2 (PDB: 6M5A). (B) β-Arabinobiose binding protein. Left panel, overall structure (PDB: 6LCE) showing β-Ara2 as yellow spheres. Middle panel, the substrate-binding site showing β-Ara2 (yellow sticks). Right panel, closing motion is shown by the superimposition of the liganded structure (rainbow colour) with the open ligand-free state (white) calculated by a molecular dynamics simulation. (C) GH127 β-l-arabinofuranosidase HypBA1 (PDB: 3WKX). Left panel, overall structure showing β-l-arabinofuranose as yellow spheres. Right panel, the active site structure. The covalent intermediate structure with a cyclophellitol-derived inhibitor (PDB: 7DIF) is shown. Cys417 and Glu322 are the catalytic nucleophile and the acid/base residues.
Conclusions
The crystal structures of other bifidobacterial glycosidases were also reported: GH3 β-glucosidase from B. longum KACC 91563 for saponin metabolism [93], GH5 β-mannosidase from B. longum NCC2705 for N-glycan utilization [94], and GH42 β1,6-galactosidase [95] and GH43 exo-β1,3-galactanase from B. bifidum S17 [96] for degradation of β-galactoside-containing carbohydrates in the human diet. Here we presented examples of the structural studies performed to date on carbohydrate-related enzymes and transporter binding proteins from the Bifidobacterium genus. However, considering their surprisingly broad carbohydrate utilisation capabilities, our structural and biochemical insights into these catabolic pathways remain limited; thus, further investigation of such enzymes and transport proteins is required. Bifidobacteria have been highly successful in establishing symbiotic relationships with different hosts, most notably humans. Therefore, deciphering the structure-based mechanisms of the protein components is instrumental for revealing the interesting molecular coevolution history between bifidobacteria and their hosts.
Perspectives
Importance to the field: A considerable expansion of pathways targeting host-derived or dietary plant glycans is observed in Bifidobacterium. Although bifidobacteria are considered secondary degraders with few predicted extracellular glycosidases, they are equipped with an arsenal of ABC transporters that are crucial for competitive growth on diverse oligosaccharides.
Summary of current thinking: Although some of the transporters of bifidobacteria have been broadly shared within the genus, others have evolved to drive specialization within specific groups. The ability of substrate-binding proteins to capture ligands with high affinity and specificity is likely to be important for efficient cross-feeding.
Future directions: Bifidobacteria have been widely used as commercial probiotics, but the molecular mechanisms responsible for their symbiosis with humans and associated health benefits remain underexplored. Insight into the species- and/or strain-specific metabolic signatures will be instrumental for designing personalized mechanism-based dietary interventions to target distinct bifidobacteria and promote human health.
Acknowledgements
S.F. acknowledges helpful discussions with Drs. Takatoshi Arakawa and Chihaya Yamada.
Abbreviations
- ABC
ATP-binding cassette
- AXOS
arabinoxylooligosaccharides
- CAZymes
carbohydrate-active enzymes
- FL
fucosyllactose
- GH
glycoside hydrolase
- GNB
galacto-N-biose
- HGM
human gut microbiota
- HMOs
human milk oligosaccharides
- LNB
lacto-N-biose I
- SBP
substrate- or solute-binding protein
Competing Interests
The authors declare that there are no competing interests associated with the manuscript.
Funding
S.F. acknowledges funding from JSPS-KAKENHI under Grant Numbers 19H00929, 19K05789, 15H02443, 15F15091, 26660083, and 24380053. M.A.H acknowledges funding from the Independent Research Fund Denmark Natural Sciences (FNU) Research Project 2 under Grant 4002-00297.
Open Access Statement
Open access for this article was enabled by the participation of the University of Tokyo in an all-inclusive Read & Publish pilot with Portland Press and the Biochemical Society under a transformative agreement with USACO.
Author Contributions
Both authors contributed to the writing and editing of this manuscript.
References
- 1.Clemente, J.C., Ursell, L.K., Parfrey, L.W. and Knight, R. (2012) The impact of the gut microbiota on human health: an integrative view. Cell 148, 1258–1270 10.1016/j.cell.2012.01.035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Belkaid, Y. and Hand, T.W. (2014) Role of the microbiota in immunity and inflammation. Cell 157, 121–141 10.1016/j.cell.2014.03.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sonnenburg, J.L. and Bäckhed, F. (2016) Diet-microbiota interactions as moderators of human metabolism. Nature 535, 56–64 10.1038/nature18846 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.McFarland L, V., Evans, C.T. and Goldstein, E.J.C. (2018) Strain-specificity and disease-specificity of probiotic efficacy: a systematic review and meta-analysis. Front. Med. 5, 124 10.3389/fmed.2018.00124 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Turroni, F., van Sinderen, D. and Ventura, M. (2011) Genomics and ecological overview of the genus Bifidobacterium. Int. J. Food Microbiol. 149, 37–44 10.1016/j.ijfoodmicro.2010.12.010 [DOI] [PubMed] [Google Scholar]
- 6.Mattarelli, P., Biavati, B., Holzapfel, W. and Wood, B.J. (2018) The Bifidobacteria and Related Organisms: Biology, Taxonomy, Applications, 1st edn, Academic Press, Cambridge, MA [Google Scholar]
- 7.Milani, C., Mangifesta, M., Mancabelli, L., Lugli, G.A., James, K., Duranti, S.et al. (2017) Unveiling bifidobacterial biogeography across the mammalian branch of the tree of life. ISME J. 11, 2834–2847 10.1038/ismej.2017.138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wong, C.B., Odamaki, T. and Xiao, J.Z. (2020) Insights into the reason of human-residential bifidobacteria (HRB) being the natural inhabitants of the human gut and their potential health-promoting benefits. FEMS Microbiol. Rev. 44, 369–385 10.1093/femsre/fuaa010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Moeller, A.H., Caro-Quintero, A., Mjungu, D., Georgiev A, V., Lonsdorf E, V., Muller, M.N.et al. (2016) Cospeciation of gut microbiota with hominids. Science 353, 380–382 10.1126/science.aaf3951 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Briggs, J.A., Grondin, J.M. and Brumer, H. (2021) Communal living: glycan utilization by the human gut microbiota. Environ. Microbiol. 23, 15–35 10.1111/1462-2920.15317 [DOI] [PubMed] [Google Scholar]
- 11.Tamura, K. and Brumer, H. (2021) Glycan utilization systems in the human gut microbiota : a gold mine for structural discoveries. Curr. Opin. Struct. Biol. 68, 26–40 10.1016/j.sbi.2020.11.001 [DOI] [PubMed] [Google Scholar]
- 12.Arumugam, M., Raes, J., Pelletier, E., Le Paslier, D., Yamada, T., Mende, D.R.et al. (2011) Enterotypes of the human gut microbiome. Nature 473, 174–180 10.1038/nature09944 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Grondin, J.M., Tamura, K., Déjean, G., Abbott, D.W. and Brumer, H. (2017) Polysaccharide utilization loci: Fueling microbial communities. J. Bacteriol. 199, e00860-16 10.1128/JB.00860-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ndeh, D. and Gilbert, H.J. (2018) Biochemistry of complex glycan depolymerisation by the human gut microbiota. FEMS Microbiol. Rev. 42, 146–164 10.1093/femsre/fuy002 [DOI] [PubMed] [Google Scholar]
- 15.Glenwright, A.J., Pothula, K.R., Bhamidimarri, S.P., Chorev, D.S., Baslé, A., Firbank, S.J.et al. (2017) Structural basis for nutrient acquisition by dominant members of the human gut microbiota. Nature 541, 407–411 10.1038/nature20828 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Parche, S., Amon, J., Jankovic, I., Rezzonico, E., Beleut, M., Barutçu, H.et al. (2006) Sugar transport systems of Bifidobacterium longum NCC2705. J. Mol. Microbiol. Biotechnol. 12, 9–19 10.1159/000096455 [DOI] [PubMed] [Google Scholar]
- 17.Beek J, T., Guskov, A. and Slotboom, D.J. (2014) Structural diversity of ABC transporters. J. Gen. Physiol. 143, 419–435 10.1085/jgp.201411164 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Iwata, S. and Ohta, T. (1993) Molecular basis of allosteric activation of bacterial l-lactate dehydrogenase. J. Mol. Biol. 230, 21–27 10.1006/jmbi.1993.1122 [DOI] [PubMed] [Google Scholar]
- 19.Iwata, S., Yoshida, S. and Ohta, T. (1994) A regular 1:1 complex of two allosteric states in the single crystal of L-lactate dehydrogenase from Bifidobacterium longum. J. Mol. Biol. 236, 958–959 10.1006/jmbi.1994.1202 [DOI] [PubMed] [Google Scholar]
- 20.Iwata, S., Kamata, K., Yoshida, S., Minowa, T. and Ohta, T. (1994) T and R states in the crystals of bacterial L-lactate dehydrogenase reveal the mechanism for allosteric control. Nat. Struct. Biol. 1, 176–185 10.1038/nsb0394-176 [DOI] [PubMed] [Google Scholar]
- 21.Pokusaeva, K., Fitzgerald, G.F. and Van Sinderen, D. (2011) Carbohydrate metabolism in Bifidobacteria. Genes Nutr. 6, 285–306 10.1007/s12263-010-0206-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Takahashi, K., Tagami, U., Shimba, N., Kashiwagi, T., Ishikawa, K. and Suzuki, E.I. (2010) Crystal structure of Bifidobacterium longum phosphoketolase: key enzyme for glucose metabolism in Bifidobacterium. FEBS Lett. 584, 3855–3861 10.1016/j.febslet.2010.07.043 [DOI] [PubMed] [Google Scholar]
- 23.Suzuki, R., Kim, B.-J., Shibata, T., Iwamoto, Y., Katayama, T., Ashida, H.et al. (2010) Overexpression, crystallization and preliminary X-ray analysis of xylulose-5-phosphate/fructose-6-phosphate phosphoketolase from Bifidobacterium breve. Acta Crystallogr. Sect. F. Struct. Biol. Cryst. Commun. 66, 941–943 10.1107/S1744309110023845 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Suzuki, R., Katayama, T., Kim, B.J., Wakagi, T., Shoun, H., Ashida, H.et al. (2010) Crystal structures of phosphoketolase: thiamine diphosphate-dependent dehydration mechanism. J. Biol. Chem. 285, 34279–34287 10.1074/jbc.M110.156281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lombard, V., Golaconda Ramulu, H., Drula, E., Coutinho, P.M. and Henrissat, B. (2014) The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 42, D490–D495 10.1093/nar/gkt1178 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sprogøe, D., Van Den Broek, L.A.M., Mirza, O., Kastrup, J.S., Voragen, A.G.J., Gajhede, M.et al. (2004) Crystal structure of sucrose phosphorylase from Bifidobacterium adolescentis. Biochemistry. 43, 1156–1162 10.1021/bi0356395 [DOI] [PubMed] [Google Scholar]
- 27.Mirza, O., Skov, L.K., Sprogøe, D., Van Den Broek, L.A.M., Beldman, G., Kastrup, J.S.et al. (2006) Structural rearrangements of sucrose phosphorylase from Bifidobacterium adolescentis during sucrose conversion. J. Biol. Chem. 281, 35576–35584 10.1074/jbc.M605611200 [DOI] [PubMed] [Google Scholar]
- 28.Bujacz, A., Jedrzejczak-Krzepkowska, M., Bielecki, S., Redzynia, I. and Bujacz, G. (2011) Crystal structures of the apo form of β-fructofuranosidase from Bifidobacterium longum and its complex with fructose. FEBS J. 278, 1728–1744 10.1111/j.1742-4658.2011.08098.x [DOI] [PubMed] [Google Scholar]
- 29.Sakanaka, M., Gotoh, A., Yoshida, K., Odamaki, T., Koguchi, H., Xiao, J.Z.et al. (2020) Varied pathways of infant gut-associated Bifidobacterium to assimilate human milk oligosaccharides: Prevalence of the gene set and its correlation with bifidobacteria-rich microbiota formation. Nutrients. 12, 71 10.3390/nu12010071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Fushinobu, S. (2010) Unique sugar metabolic pathways of bifidobacteria. Biosci. Biotechnol. Biochem. 74, 2374–2384 10.1271/bbb.100494 [DOI] [PubMed] [Google Scholar]
- 31.Urashima, T., Fukuda, K., Kitaoka, M., Ohnishi, M., Terabayashi, T. and Kobata, A. (2010) Milk Oligosaccharides, Nova Science Publishers, New York [Google Scholar]
- 32.Kitaoka, M., Tian, J. and Nishimoto, M. (2005) Novel putative galactose operon involving lacto-N-biose phosphorylase in Bifidobacterium longum. Appl. Environ. Microbiol. 71, 3158–3162 10.1128/AEM.71.6.3158-3162.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Brockhausen, I. and Stanley, P. (2017) Chapter 10 O-GalNAc Glycans. In Essentials of Glycobiology (Varki A, ed.), 3rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY: [PubMed] [Google Scholar]
- 34.Kiyohara, M., Tachizawa, A., Nishimoto, M., Kitaoka, M., Ashida, H. and Yamamot, K. (2009) Prebiotic effect of lacto-N-biose I on bifidobacterial growth. Biosci. Biotechnol. Biochem. 73, 1175–1179 10.1271/bbb.80697 [DOI] [PubMed] [Google Scholar]
- 35.Hidaka, M., Nishimoto, M., Kitaoka, M., Wakagi, T., Shoun, H. and Fushinobu, S. (2009) The crystal structure of galacto-N-biose/lacto-N-biose I phosphorylase. A large deformation of a tim barrel scaffold. J. Biol. Chem. 284, 7273–7283 10.1074/jbc.M808525200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ambrogi, V., Bottacini, F., O'Sullivan, J., O'Connell Motherway, M., Linqiu, C., Schoemaker, B.et al. (2019) Characterization of GH2 and GH42 β-galactosidases derived from bifidobacterial infant isolates. AMB Express 9, 9 10.1186/s13568-019-0735-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nishimoto, M. and Kitaoka, M. (2007) Identification of N-acetylhexosamine 1-kinase in the complete lacto-N-biose I/galacto-N-biose metabolic pathway in Bifidobacterium longum. Appl. Environ. Microbiol. 73, 6444–6449 10.1128/AEM.01425-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wang, K.C., Lyu, S.Y., Liu, Y.C., Chang, C.Y., Wu, C.J. and Li, T.L. (2014) Insights into the binding specificity and catalytic mechanism of N-acetylhexosamine 1-phosphate kinases through multiple reaction complexes. Acta Crystallogr. Sect. D Biol. Crystallogr. 70, 1401–1410 10.1107/S1399004714004209 [DOI] [PubMed] [Google Scholar]
- 39.Sato, M., Arakawa, T., Nam, Y.W., Nishimoto, M., Kitaoka, M. and Fushinobu, S. (2015) Open-close structural change upon ligand binding and two magnesium ions required for the catalysis of N-acetylhexosamine 1-kinase. Biochim. Biophys. Acta 1854, 333–340 10.1016/j.bbapap.2015.01.011 [DOI] [PubMed] [Google Scholar]
- 40.Zhao, Y., She, N., Ma, Y., Wang, C. and Cao, Z. (2018) A description of enzymatic catalysis in N-acetylhexosamine 1-kinase: concerted mechanism of two-magnesium-ion-assisted GlcNAc phosphorylation, flexibility behavior of lid motif upon substrate recognition, and water-assisted GlcNAc-1-P release. ACS Catal. 8, 4143–4159 10.1021/acscatal.8b00006 [DOI] [Google Scholar]
- 41.Nam, Y.-W., Nishimoto, M., Arakawa, T., Kitaoka, M. and Fushinobu, S. (2019) Structural basis for broad substrate specificity of UDP-glucose 4-epimerase in the human milk oligosaccharide catabolic pathway of Bifidobacterium longum. Sci. Rep. 9, 11081 10.1038/s41598-019-47591-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Fushinobu, S. (2021) Molecular evolution and functional divergence of UDP-hexose 4-epimerases. Curr. Opin. Chem. Biol. 61, 53–62 10.1016/j.cbpa.2020.09.007 [DOI] [PubMed] [Google Scholar]
- 43.De Bruyn, F., Beauprez, J., Maertens, J., Soetaert, W. and De Mey, M. (2013) Unraveling the leloir pathway of Bifidobacterium bifidum: Significance of the uridylyltransferases. Appl. Environ. Microbiol. 79, 7028–7035 10.1128/AEM.02460-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Suzuki, R., Wada, J., Katayama, T., Fushinobu, S., Wakagi, T., Shoun, H.et al. (2008) Structural and thermodynamic analyses of solute-binding protein from Bifidobacterium longum specific for core 1 disaccharide and lacto-N-biose I. J. Biol. Chem. 283, 13165–13173 10.1074/jbc.M709777200 [DOI] [PubMed] [Google Scholar]
- 45.Sela, D.A. and Mills, D.A. (2010) Nursing our microbiota: molecular linkages between bifidobacteria and milk oligosaccharides. Trends Microbiol. 18, 298–307 10.1016/j.tim.2010.03.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Sakanaka, M., Hansen, M.E., Gotoh, A., Katoh, T., Yoshida, K., Odamaki, T.et al. (2019) Evolutionary adaptation in fucosyllactose uptake systems supports bifidobacteria-infant symbiosis. Sci. Adv. 5, eaaw7696 10.1126/sciadv.aaw7696 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Sela, D.A., Garrido, D., Lerno, L., Wu, S., Tan, K., Eom, H.J.et al. (2012) Bifidobacterium longum subsp. infantis ATCC 15697 α-fucosidases are active on fucosylated human milk oligosaccharides. Appl. Environ. Microbiol. 78, 795–803 10.1128/AEM.06762-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Sakurama, H., Fushinobu, S., Hidaka, M., Yoshida, E., Honda, Y., Ashida, H.et al. (2012) 1,3-1,4-α-L-Fucosynthase that specifically introduces lewis a/x antigens into type-1/2 chains. J. Biol. Chem. 287, 16709–16719 10.1074/jbc.M111.333781 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Katayama, T. (2016) Host-derived glycans serve as selected nutrients for the gut microbe: human milk oligosaccharides and bifidobacteria. Biosci. Biotechnol. Biochem. 80, 621–632 10.1080/09168451.2015.1132153 [DOI] [PubMed] [Google Scholar]
- 50.Gotoh, A., Katoh, T., Sakanaka, M., Ling, Y., Yamada, C., Asakuma, S.et al. (2018) Sharing of human milk oligosaccharides degradants within bifidobacterial communities in faecal cultures supplemented with Bifidobacterium bifidum. Sci. Rep. 8, 13958 10.1038/s41598-018-32080-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Turroni, F., Duranti, S., Milani, C., Lugli, G.A., van Sinderen, D. and Ventura, M. (2019) Bifidobacterium bifidum: a key member of the early human gut microbiota. Microorganisms 7, 544 10.3390/microorganisms7110544 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Katayama, T., Sakuma, A., Kimura, T., Makimura, Y., Hiratake, J., Sakata, K.et al. (2004) Molecular cloning and characterization of Bifidobacterium bifidum 1,2-α-L-fucosidase (AfcA), a novel inverting glycosidase (glycoside hydrolase family 95). J. Bacteriol. 186, 4885–4893 10.1128/JB.186.15.4885-4893.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Nagae, M., Tsuchiya, A., Katayama, T., Yamamoto, K., Wakatsuki, S. and Kato, R. (2007) Structural basis of the catalytic reaction mechanism of novel 1,2-α-L-fucosidase from Bifidobacterium bifidum. J. Biol. Chem. 282, 18497–18509 10.1074/jbc.M702246200 [DOI] [PubMed] [Google Scholar]
- 54.Wada, J., Ando, T., Kiyohara, M., Ashida, H., Kitaoka, M., Yamaguchi, M.et al. (2008) Bifidobacterium bifidum lacto-N-biosidase, a critical enzyme for the degradation of human milk oligosaccharides with a type 1 structure. Appl. Environ. Microbiol. 74, 3996–4004 10.1128/AEM.00149-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ito, T., Katayama, T., Hattie, M., Sakurama, H., Wada, J., Suzuki, R.et al. (2013) Crystal structures of a glycoside hydrolase family 20 lacto-N-biosidase from Bifidobacterium bifidum. J. Biol. Chem. 288, 11795–11806 10.1074/jbc.M112.420109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Odamaki, T., Bottacini, F., Kato, K., Mitsuyama, E., Yoshida, K., Horigome, A.et al. (2018) Genomic diversity and distribution of Bifidobacterium longum subsp. longum across the human lifespan. Sci. Rep. 8, 85 10.1038/s41598-017-18391-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sakurama, H., Kiyohara, M., Wada, J., Honda, Y., Yamaguchi, M., Fukiya, S.et al. (2013) Lacto-N-biosidase encoded by a novel gene of Bifidobacterium longum subspecies longum shows unique substrate specificity and requires a designated chaperone for its active expression. J. Biol. Chem. 288, 25194–25206 10.1074/jbc.M113.484733 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Yamada, C., Gotoh, A., Sakanaka, M., Hattie, M., Stubbs, K.A., Katayama-Ikegami, A.et al. (2017) Molecular insight into evolution of symbiosis between breast-fed infants and a member of the human gut microbiome Bifidobacterium longum. Cell Chem. Biol. 24, 515–524.e5 10.1016/j.chembiol.2017.03.012 [DOI] [PubMed] [Google Scholar]
- 59.Pichler, M.J., Yamada, C., Shuoker, B., Alvarez-Silva, C., Gotoh, A., Leth, M.L.et al. (2020) Butyrate producing colonic clostridiales metabolise human milk oligosaccharides and cross feed on mucin via conserved pathways. Nat. Commun. 11, 3285 10.1038/s41467-020-17075-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Bell, A. and Juge, N. (2020) Mucosal glycan degradation of the host by the gut microbiota. Glycobiology in press 10.1093/glycob/cwaa097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Neish, A.S. (2009) Microbes in gastrointestinal health and disease. Gastroenterology 136, 65–80 10.1053/j.gastro.2008.10.080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Tailford, L.E., Crost, E.H., Kavanaugh, D. and Juge, N. (2015) Mucin glycan foraging in the human gut microbiome. Front. Genet. 6, 81 10.3389/fgene.2015.00081 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.González-Morelo, K.J., Vega-Sagardía, M. and Garrido, D. (2020) Molecular insights into O-linked glycan utilization by gut microbes. Front. Microbiol. 11, 591568 10.3389/fmicb.2020.591568 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Fujita, K., Oura, F., Nagamine, N., Katayama, T., Hiratake, J., Sakata, K.et al. (2005) Identification and molecular cloning of a novel glycoside hydrolase family of core 1 type O-glycan-specific endo-α-N-acetylgalactosaminidase from Bifidobacterium longum. J. Biol. Chem. 280, 37415–37422 10.1074/jbc.M506874200 [DOI] [PubMed] [Google Scholar]
- 65.Suzuki, R., Katayama, T., Kitaoka, M., Kumagai, H., Wakagi, T., Shoun, H.et al. (2009) Crystallographic and mutational analyses of substrate recognition of endo-alpha-N-acetylgalactosaminidase from Bifidobacterium longum. J. Biochem. 146, 389–398 10.1093/jb/mvp086 [DOI] [PubMed] [Google Scholar]
- 66.Gregg, K.J., Suits, M.D.L., Deng, L., Vocadlo, D.J. and Boraston, A.B. (2015) Structural analysis of a family 101 glycoside hydrolase in complex with carbohydrates reveals insights into its mechanism. J. Biol. Chem. 290, 25657–25669 10.1074/jbc.M115.680470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Sato, M., Liebschner, D., Yamada, Y., Matsugaki, N., Arakawa, T., Wills, S.S.et al. (2017) The first crystal structure of a family 129 glycoside hydrolase from a probiotic bacterium reveals critical residues and metal cofactors. J. Biol. Chem. 292, 12126–12138 10.1074/jbc.M117.777391 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Kiyohara, M., Nakatomi, T., Kurihara, S., Fushinobu, S., Suzuki, H., Tanaka, T.et al. (2012) α-N-acetylgalactosaminidase from infant-associated bifidobacteria belonging to novel glycoside hydrolase family 129 is implicated in alternative mucin degradation pathway. J. Biol. Chem. 287, 693–700 10.1074/jbc.M111.277384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Koropatkin, N.M., Cameron, E.A. and Martens, E.C. (2012) How glycan metabolism shapes the human gut microbiota. Nat. Rev. Microbiol. 10, 323–335 10.1038/nrmicro2746 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Rogowski, A., Briggs, J.A., Mortimer, J.C., Tryfona, T., Terrapon, N., Lowe, E.C.et al. (2015) Glycan complexity dictates microbial resource allocation in the large intestine. Nat. Commun. 6, 7481 10.1038/ncomms8481 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Leth, M.L., Ejby, M., Workman, C., Ewald, D.A., Pedersen, S.S., Sternberg, C.et al. (2018) Differential bacterial capture and transport preferences facilitate co-growth on dietary xylan in the human gut. Nat. Microbiol. 3, 570–580 10.1038/s41564-018-0132-8 [DOI] [PubMed] [Google Scholar]
- 72.Ejby, M., Fredslund, F., Vujicic-Zagar, A., Svensson, B., Slotboom, D.J. and Abou Hachem, M. (2013) Structural basis for arabinoxylo-oligosaccharide capture by the probiotic Bifidobacterium animalis subsp. lactis Bl-04. Mol. Microbiol. 90, 1100–1112 10.1111/mmi.12419 [DOI] [PubMed] [Google Scholar]
- 73.Andersen, J.M., Barrangou, R., Hachem, M.A., Lahtinen, S.J., Goh, Y.J., Svensson, B.et al. (2013) Transcriptional analysis of oligosaccharide utilization by Bifidobacterium lactis Bl-04. BMC Genomics 14, 312 10.1186/1471-2164-14-312 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Scheepers, G.H., Lycklama A Nijeholt, J.A. and Poolman, B. (2016) An updated structural classification of substrate-binding proteins. FEBS Lett. 590, 4393–4401 10.1002/1873-3468.12445 [DOI] [PubMed] [Google Scholar]
- 75.Ejby, M., Fredslund, F., Andersen, J.M., Žagar, A.V., Henriksen, J.R., Andersen, T.L.et al. (2016) An ATP binding cassette transporter mediates the uptake of α-(1,6)-linked dietary oligosaccharides in Bifidobacterium and correlates with competitive growth on these substrates. J. Biol. Chem. 291, 20220–20231 10.1074/jbc.M116.746529 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Fredslund, F., Abou Hachem, M., Jonsgaard Larsen, R., Gerd Sørensen, P., Coutinho, P.M., Lo Leggio, L.et al. (2011) Crystal structure of α-galactosidase from Lactobacillus acidophilus NCFM: Insight into tetramer formation and substrate binding. J. Mol. Biol. 412, 466–480 10.1016/j.jmb.2011.07.057 [DOI] [PubMed] [Google Scholar]
- 77.Møller, M.S., Fredslund, F., Majumder, A., Nakai, H., Poulsen, J.C.N., Lo, L.L.et al. (2012) Enzymology and structure of the GH13_31 glucan 1,6-α-glucosidase that confers isomaltooligosaccharide utilization in the probiotic Lactobacillus acidophilus NCFM. J. Bacteriol. 194, 4249–4259 10.1128/JB.00622-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Bagenholm, V., Reddy, S.K., Bouraoui, H., Morrill, J., Kulcinskaja, E., Bahr, C.M.et al. (2017) Galactomannan catabolism conferred by a polysaccharide utilization locus of Bacteroides ovatus: Enzyme synergy and crystal structure of a β-mannanase. J. Biol. Chem. 292, 229–243 10.1074/jbc.M116.746438 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.La Rosa, S.L., Leth, M.L., Michalak, L., Hansen, M.E., Pudlo, N.A., Glowacki, R.et al. (2019) The human gut Firmicute Roseburia intestinalis is a primary degrader of dietary β-mannans. Nat. Commun. 10, 905 10.1038/s41467-019-08812-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Kulcinskaja, E., Rosengren, A., Ibrahim, R., Kolenová, K. and Stålbrand, H. (2013) Expression and characterization of a Bifidobacterium adolescentis beta-mannanase carrying mannan-binding and cell association motifs. Appl. Environ. Microbiol. 79, 133–140 10.1128/AEM.02118-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Morrill, J., Kulcinskaja, E., Sulewska, A.M., Lahtinen, S., Stålbrand, H., Svensson, B.et al. (2015) The GH5 1,4-β-mannanase from Bifidobacterium animalis subsp. lactis Bl-04 possesses a low-affinity mannan-binding module and highlights the diversity of mannanolytic enzymes. BMC Biochem. 16, 26 10.1186/s12858-015-0055-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ejby, M., Guskov, A., Pichler, M.J., Zanten, G.C., Schoof, E., Saburi, W.et al. (2019) Two binding proteins of the ABC transporter that confers growth of Bifidobacterium animalis subsp. lactis ATCC27673 on β-mannan possess distinct manno-oligosaccharide-binding profiles. Mol. Microbiol. 112, 114–130 10.1111/mmi.14257 [DOI] [PubMed] [Google Scholar]
- 83.Viborg, A.H., Fredslund, F., Katayama, T., Nielsen, S.K., Svensson, B., Kitaoka, M.et al. (2014) A β1-6/β1-3 galactosidase from Bifidobacterium animalis subsp. lactis Bl-04 gives insight into sub-specificities of β-galactoside catabolism within Bifidobacterium. Mol. Microbiol. 94, 1024–1040 10.1111/mmi.12815 [DOI] [PubMed] [Google Scholar]
- 84.Theilmann, M.C., Fredslund, F., Svensson, B., Lo Leggio, L. and Abou Hachem, M. (2019) Substrate preference of an ABC importer corresponds to selective growth on β-(1,6)-galactosides in Bifidobacterium animalis subsp. lactis. J. Biol. Chem. 294, 11701–11711 10.1074/jbc.RA119.008843 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Theilmann, M.C., Goh, Y.J., Nielsen, K.F., Klaenhammer, T.R., Barrangou, R. and Hachem, M.A. (2017) Lactobacillus acidophilus metabolizes dietary plant glucosides and externalizes their bioactive phytochemicals. MBio 8, e01421-17 10.1128/mBio.01421-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Viborg, A.H., Katayama, T., Arakawa, T., Abou Hachem, M., Lo Leggio, L., Kitaoka, M.et al. (2017) Discovery of α-L-arabinopyranosidases from human gut microbiome expands the diversity within glycoside hydrolase family 42. J. Biol. Chem. 292, 21092–21101 10.1074/jbc.M117.792598 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Fujita, K., Sakamoto, S., Ono, Y., Wakao, M., Suda, Y., Kitahara, K.et al. (2011) Molecular cloning and characterization of a β-L-arabinobiosidase in Bifidobacterium longum that belongs to a novel glycoside hydrolase family. J. Biol. Chem. 286, 5143–5150 10.1074/jbc.M110.190512 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Fujita, K., Takashi, Y., Obuchi, E., Kitahara, K. and Suganuma, T. (2014) Characterization of a novel β-L-arabinofuranosidase in Bifidobacterium longum: Functional elucidation of a DUF1680 protein family member. J. Biol. Chem. 289, 5240–5249 10.1074/jbc.M113.528711 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Saito, K., Viborg, A.H., Sakamoto, S., Arakawa, T., Yamada, C., Fujita, K.et al. (2020) Crystal structure of β-L-arabinobiosidase belonging to glycoside hydrolase family 121. PLoS One 15, e0231513 10.1371/journal.pone.0231513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Miyake, M., Terada, T., Shimokawa, M., Sugimoto, N., Arakawa, T., Shimizu, K.et al. (2020) Structural analysis of β-L-arabinobiose-binding protein in the metabolic pathway of hydroxyproline-rich glycoproteins in Bifidobacterium longum. FEBS J. 287, 5114–5129 10.1111/febs.15315 [DOI] [PubMed] [Google Scholar]
- 91.Ito, T., Saikawa, K., Kim, S., Fujita, K., Ishiwata, A., Kaeothip, S.et al. (2014) Crystal structure of glycoside hydrolase family 127 β-L-arabinofuranosidase from Bifidobacterium longum. Biochem. Biophys. Res. Commun. 447, 32–37 10.1016/j.bbrc.2014.03.096 [DOI] [PubMed] [Google Scholar]
- 92.McGregor, N.G.S., Coines, J., Borlandelli, V., Amaki, S., Artola, M., Nin-Hill, A.et al. (2020) Cysteine nucleophiles in glycosidase catalysis: application of a covalent β-l-arabinofuranosidase inhibitor. Angew. Chemie - Int. Ed. in press 10.1002/anie.202013920 [DOI] [PubMed] [Google Scholar]
- 93.Yan, S., Wei, P., Chen, Q., Chen, X., Wang, S., Li, J.et al. (2018) Functional and structural characterization of a β-glucosidase involved in saponin metabolism from intestinal bacteria. Biochem. Biophys. Res. Commun. 496, 1349–1356 10.1016/j.bbrc.2018.02.018 [DOI] [PubMed] [Google Scholar]
- 94.Cordeiro, R.L., Pirolla, R.A.S., Persinoti, G.F., Gozzo, F.C., de Giuseppe, P.O. and Murakami, M.T. (2019) N-glycan utilization by bifidobacterium gut symbionts involves a specialist β-mannosidase. J. Mol. Biol. 431, 732–747 10.1016/j.jmb.2018.12.017 [DOI] [PubMed] [Google Scholar]
- 95.Godoy, A.S., Camilo, C.M., Kadowaki, M.A., Muniz, H.D., Espirito Santo, M., Murakami, M.T.et al. (2016) Crystal structure of β1→6-galactosidase from Bifidobacterium bifidum S17: trimeric architecture, molecular determinants of the enzymatic activity and its inhibition by α-galactose. FEBS J. 283, 4097–4112 10.1111/febs.13908 [DOI] [PubMed] [Google Scholar]
- 96.Godoy, A.S., De Lima, M.Z.T., Camilo, C.M. and Polikarpov, I. (2016) Crystal structure of a putative exo-β-1,3-galactanase from Bifidobacterium bifidum S17. Acta Crystallogr. Sect. Struct. Biol. Commun. 72, 288–293 10.1107/S2053230X16003617 [DOI] [PMC free article] [PubMed] [Google Scholar]