

Abstract
Africa is the continent with the greatest genetic diversity among humans and the level of diversity is further enhanced by incorporating non-majority groups, which are often understudied. Many of today’s minority populations historically practiced foraging lifestyles, which were the only subsistence strategies prior to the rise of agriculture and pastoralism, but only a few groups practicing these strategies remain today. Genomic investigations of Holocene human remains excavated across the African continent show that the genetic landscape was vastly different compared to today’s genetic landscape and that many groups that today are population isolate inhabited larger regions in the past. It is becoming clear that there are periods of isolation among groups and geographic areas, but also genetic contact over large distances throughout human history in Africa. Genomic information from minority populations and from prehistoric remains provide an invaluable source of information on the human past, in particular deep human population history, as Holocene large-scale population movements obscure past patterns of population structure. Here we revisit questions on the nature and time of the radiation of early humans in Africa, the extent of gene-flow among human populations as well as introgression from archaic and extinct lineages on the continent.
Introduction
Africa has been identified as the place of origin of Homo sapiens, evident by the highest genetic diversity and deepest splits being found among people from this continent. The fact that the earliest fossil finds of early humans have been found in Africa, including northern (dating to about 300 000 years old), southern (a 260 000-year-old individual) and eastern Africa (about 195 000–160 000 years old, 1–7) further points to Africa’s importance for early human development. The African fossil record of early humans displays a large morphological diversity and possible coexistence with now-extinct lineages of other Homo species or forms (2,5–15). The complex early history of humans in Africa poses questions on how our species emerged, the deep population structure and its origin, and whether there has been a genetic exchange of deeply structured and/or archaic populations with modern humans in Africa. With more genomes becoming available from diverse African groups as well as efforts in ancient DNA sequencing, we are gradually gaining more information on African diversity and these genomic data can reveal unknown ancestries or genetic make-up by providing information before migration or admixture events and can give indications of population continuity.
Hunting and gathering were the lifestyle of all humans prior to the Neolithic transition and emergence of agriculture and pastoralism. Most hunter-gatherer groups have been replaced or incorporated in expanding agriculturalist and pastoralist populations (16), only a few extant groups continue to practice this way of life and can be found scattered in various regions across the globe, including Africa. Today, most remaining practicing hunter-gatherer populations occupy geographical areas that are not suitable for agriculture or grazing, such as rainforests and deserts. As a consequence, the number of hunter-gatherer groups (and practicing individuals) has been reduced drastically in the last few thousand years, due to the expansion and increase in population size of agriculturalists and pastoralists (16–21).
In Africa, hunter-gatherer groups can be found in the south and east and in the Congo Basin. In southern Africa, the San hunter-gatherers and the Khoekhoe herders, who adopted a pastoralist lifestyle after contact with east African pastoralists (22), inhabit arid regions unsuitable for agriculture. They are collectively named Khoe–San, in correspondence to speaking Khoisan languages, which comprise ‘click’-rich languages belonging to five linguistic families that are otherwise unrelated (23). In eastern Africa, there are various populations that practice, or have until recently, a hunter-gatherer lifestyle. East African hunter-gatherers (EAHG) have been shown to be more closely genetically related to each other than to other hunter-gatherer populations in Africa (24–27). Furthermore, several hunter-gatherer populations inhabit the rainforests of equatorial Africa, such as the Biaka, Baka, Bakola, Bedzan, Batwa, Twa and Mbuti. These rainforest hunter-gatherer (RHG) groups have adopted the languages of neighboring agricultural populations (27,28). The rapid expansion in the last 5000 years, as well as other recent migrations in Africa obscure patterns of deep population structure in sub-Saharan Africa (20) and much of sub-equatorial Africa, is today populated by people of western African descent speaking Bantu languages.
It is now increasingly recognized that the human population in Africa was stratified prior to the migrations of humans Out-of-Africa some 80 000 years ago (ya), which rejects the idea of a single panmictic human population in eastern Africa that expands into southern and western Africa and out from Africa (20,29,30). To investigate deep population structure, that goes beyond 100 000 years, it is essential to investigate populations from a wide geographic range that potentially represent descendants of groups living in the area prior to new groups arriving in the second half of the Holocene. Hunter-gatherer populations and DNA samples from ancient individuals can provide insight into deeper population history that is not obscured by the confounding factors of recent large-scale migrations. Recent admixture can, for example, lead to an underestimation of split times (31). Ancient DNA also offers the possibility of unearthing genetic diversity that is lost in the modern gene-pool. With the rapid development in genome sequencing technologies, the number of whole-genome sequences available from modern-day hunter-gatherers and genome-wide information from African archeological samples has increased rapidly in recent years, despite the fact that much of Africa has not been extensively surveyed by archaeologists and that DNA does not preserve particularly well in the hot and humid climate. Since the publication of the first ancient genome from east Africa in 2015 (32), genome-wide information from several ancient African samples was successfully retrieved, with samples spanning the entire continent and even African islands; the oldest samples, from north Africa, date to 15 000 years (31,33–40).
Hunter-gatherer Population Structure
The high genetic diversity in Africa is a result of a deep population structure, likely shaped by periods of isolation, possibly driven by climatic oscillations (30,41). Various measurements of genetic diversity show the most extreme values in Africa, such as comparatively high allelic richness, heterozygosity as well as short runs of homozygosity (42). African genomes, on average, harbor the most divergent genetic lineages among all humans, with the exception of lineages tracing to Neanderthals and Denisovans (19–21,42). African hunter-gatherer groups have previously been shown to be the most genetically diverse contemporary populations, carry the most basal uniparental markers (24,43–46), and harbor the deepest autosomal branches (18,25,29,31,42,47–50).
The genetic relationship between the various contemporary regional hunter-gatherer groups can be modeled with isolation-by-distance, revealing that hunter-gatherer populations were connected, potentially in overlapping areas and inhabited larger geographic areas (35,40,51). There are indications of gene-flow between Khoe–San and other African hunter-gatherer populations after they split from each other (27,35,42,52), suggesting that the genetic exchange persisted until the Holocene. For instance, there appear to have been a clinal link between southern African San groups and eastern African hunter-gatherers, where prehistoric individuals from e.g. current-day Malawi display affinities to both San groups and to eastern African hunter-gatherers (35,38), with some San-like ancestry, detected in ancient individuals excavated in eastern Africa (40). Today, the eastern African hunter-gatherers display low Ne due to a small census size (27,53).
The contraction of once wide-spread, overlapping hunter-gatherer populations, which led to the scattered populations we know today, left a mark in their genomes. For instance, many hunter-gatherer populations do not display an effective population size (Ne) increase in the Holocene, contrasting to many other groups (35,40,42). Today the RHG exhibits lower Ne than surrounding agriculturalists (54). The southern African Khoe–San have repeatedly been shown to display the highest genetic diversity in global comparisons (18,29,31,42,48). This is caused by a large Ne of the Khoe–San for the majority of human history, as well as admixture from non-Khoe–San populations (18,19,42). The lineage leading to current-day Khoe–San populations likely inhabited a larger geographical region than the distribution of contemporary southern African hunter-gatherers (35,40). It has been hypothesized that the ancestors of the Khoe–San were most likely the only inhabitants of southern Africa for most of the region’s prehistory (31,35). Ne starts to differ between Khoe–San and all other groups some 300 000–200 000 ya (42,52), demonstrating that there was population structure at that time, followed by a population decline that affected populations differently. For instance, the San and RHG maintained a larger Ne than other African populations prior to 60 000 ya, but all groups, including the Khoe–San and RHG, display population decline in this time period (42,52).
Populations from north Africa are often exempted when discussing deep African history as they show mainly Eurasian ancestry, with a modest ancestry component assigned to sub-Saharan Africa (55,56). However, recent ancient DNA results from Moroccan fossil samples dating back 15 000 years provide insights into hunter-gatherers from north Africa. These samples derive most of their ancestry from a non-African like source (best match: Natufian) but also derive one-third of their ancestry from sub-Saharan Africa (37). The sub-Saharan ancestry component appears as a mix of east and west African ancestry but there are no clear sources for these ancestries, rather the ancestry is likely from an unsampled population that is related to both modern-day west and east Africans. These individuals from Morocco along with younger samples (7000 and 5000 years old) show decreasing sub-Saharan ancestry over time (36,37), a trend which has also been observed in Egypt (34). This pattern likely arises from isolation in the Maghreb since the Upper Paleolithic (36). Whether the non-African ancestry derives from an early back-admixture event or if there is long-standing gene-flow between north Africa and non-African populations has not yet been resolved.
Models of Human Evolution and Population Divergence Time Estimates
It is common to model the evolutionary history of humans as a bifurcating tree and to estimate divergence times between the branches. Trees are a simplification and they miss some features of human population history, such as gene-flow, but they are useful as models in order to understand the relations among groups and the relative divergences among groups. The estimates for a specific event typically vary due to the methods and the assumptions of the models, the scaling parameters (e.g. mutation rate and generation time), and the set of individuals and populations used for the comparisons (57). Alternatively, human evolution can be represented with metapopulation models (58), though such modeling for addressing questions of deep human history is still rare (59).
Figure 1 gives an overview of the range of estimates of deep divergence events in human population history based on high-coverage autosomal genomes from a range of studies (see 57 for the underlying data). Note that estimates based on MSMC and MSMC2 methods (60,61) are more recent than estimates based on e.g. genealogical concordance, site-frequency based methods or approximate Bayesian computation analyses (after rescaling with the same mutation rate), potentially due to the fact that the former methods use the midpoint of cross-coalescences as the divergence time estimate. However, the rank order of divergence times does not differ much between the different approaches, which points to consistent population topology across the methods. The divergence between the ancestors of the Khoe–San and the ancestors of the rest of modern humans is estimated to between 340 000 and 200 000 ya (31,42,49,62), and with younger estimates (160 000–90 000) based on the MSMC cross-coalescence approach (19,21,52,63). The next event assuming a simplified bifurcating tree is a divergence between the RHG ancestors and the ancestors of the rest of modern humans (minus the Khoe–San); the estimates vary from 350 000 to 70 000 ya (19,21,31,42,52,62–64) but are generally more recent than the Khoe–San divergence. Eastern African groups, including hunter-gatherers, such as the Hadza and the Sandawe, point to divergences from all other African groups, including western Africans, at ~140 000–70 000 ya (31,42,52).
Figure 1.
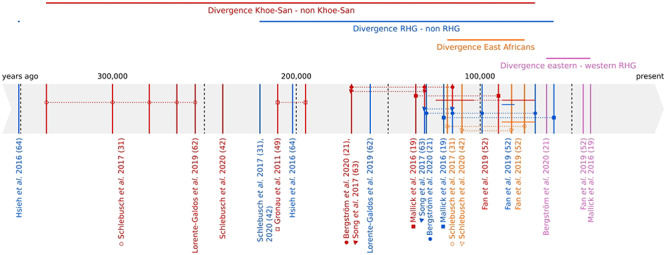
Schematic overview of divergence times inferred from whole-genomes. Estimates of the divergence between Khoe-San and other populations are represented in red; between rainforest hunter-gatherers and other populations in blue; between east African hunter-gatherers and other populations in orange; and between eastern and western rainforest hunter-gatherers in pink. Point estimates from the same study and for the same event are identified by a symbol and are connected by dotted lines; a given study can give several point estimates because e.g. of different comparative populations or different methods. In the case of Song et al. (63), estimates were given as ranges, which are represented by a continuous line.
Most studies find a bifurcation as the deepest divergence among humans (but see next section for the potential impact of unknown groups). For instance, mtDNA and complete genomes repeatedly find the deepest divergence among humans being between Khoe–San and all other groups (20,21,31,42,49). However, some studies point at alternatives, for instance, a very deep divergence involving RHGs (64) or a trifurcation as the deepest divergence, partly based on genetic data from 8000 years old human remains from west Africa (Shum Laka), between Khoe–San, west Africans and RHGs (39). The 8000 years old Shum Laka individual interestingly showed both west African and RHG ancestry, and if we estimate the divergence time for this individual, it shows divergence time to the Khoe–San of 260 000–350 000 years, and 120 000–220 000 years to west Africans and RHGs (Fig. 2), solidly placing the 8000 years old Shum Laka individual as an individual on the non-Khoe–San branch, related to west Africans and RHGs (as a result of admixture or shared ancestry).
Figure 2.
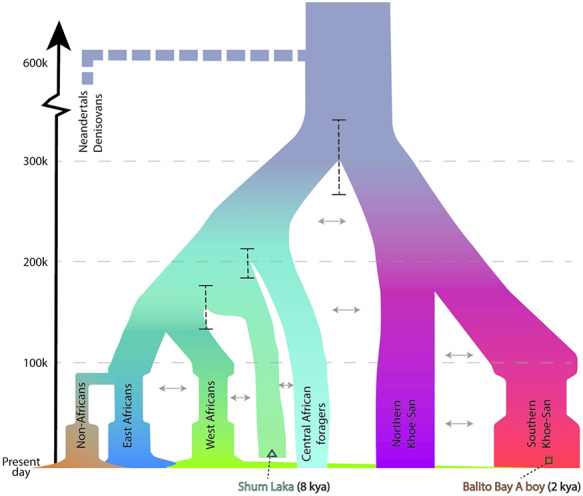
Demographic model of African history and estimated divergences. Summary of population split times, hierarchy and population sizes (29,31,39,42,54). Vertical dashed black lines indicate confidence intervals of population split times estimated using the ‘TT method’ (84). To estimate the divergence times between the Shum Laka individual, we download the genome sequence data (39) and used the ‘TT’ method to compute divergence times. The ‘TT’ method uses sample configurations of variants across the genome in two individuals in a population divergence model. Independent of the drift in each daughter population and given a mutation rate, this approach estimates the number of generations since a population divergence for a pair of individuals.
Although not as well studied, the estimates for the divergence between northern and southern San have a large range as well, from ~170 000 to ~30 000 ya (19,31,52). This possibly reflects the fact that an isolation-by-distance model might be a better representation of the relationships between these populations (51). The common origin of western and eastern RHG has been demonstrated (65) and the divergence between western and eastern RHG has been estimated to ~60 000–40 000 ya (19,21,52). Note that all these estimates are based on MSMC/MSMC2, which typically provide more shallow divergence time estimates. See Figures 1 and 2 for an overview of the human population topology and divergence time estimates.
Introgression from Extinct Lineages
There has been a resurging interest in the question of archaic admixture in Africa. With the recovery of the first genome-wide data from Neanderthals and Denisovans (66,67), admixture from these archaic groups has become evident in non-Africans. With new methods that do not necessarily require a reference genome from the introgressing lineage (62,68–71) and better genome data, it is now possible to investigate the question of introgression from extinct lineages in Africa. Given the rich human and hominin history in Africa, the lineage leading to modern-day humans has the potential to interact and also mix with hominin groups that separated from our lineage far back in time, perhaps on par with the divergence of Neanderthals and Denisovans some 600 000 ya. It is also likely that distinct modern human groups existed, possibly diverging a few hundred thousand years ago or more recently, that later disappeared for some reason. Such populations could have mixed with human groups that did contribute substantially to today’s people in Africa. We can refer to this latter phenomenon as introgression from ‘ghost’ populations, and the former case as ‘archaic’ introgression. One way to separate these two introgression processes is to define ‘archaic’ introgression as involving a hominin group separated from the human lineage before the deepest divergence among modern humans, some 300 000 ya, and ‘ghost’ introgression involving extinct anatomically modern groups that separated around 300 000 ya or more recently.
Since the publication of the Neanderthal genome (67), the Neanderthal admixture into human populations is continuously being unraveled, which also has bearing on the understanding of the demographic history of African populations. The west African Yoruba have often been used as an unadmixed reference population to estimate global Neanderthal admixture proportions (67), although they have been shown to carry small levels of Neanderthal ancestry (16,72). A new method that eliminates the need for an unadmixed modern source showed that the Neanderthal ancestry proportion in Africans was larger than previously thought (IBDmix, 72). Chen et al. (72) found on average 17 Mb of Neanderthal sequence per individual of which 94% is shared with non-Africans. Bergström et al. (21) also find archaic variants from Neanderthal and Denisovans in west Africans that are absent in non-Africans, reflecting the larger genetic variation found in Africa. The Neanderthal ancestry in modern Africans can be explained by back migration from an ancestral European-like source into Africa (72), a scenario which is supported by other studies that found Neanderthal ancestry proportions to correlate with Eurasian ancestry proportions (21,32,73,74).
Even before the first Neanderthal genome was published, signals of introgression from deeply branching populations had already been detected in African populations (68,75). A potential explanation for this observation would be that these other populations are now extinct and their genetic material only lives on as traces in contemporary populations. With the publication of the Neanderthal and Denisovan genomes (66,67), it became evident that while Neanderthals and Denisovans can account for all of the archaic admixture in non-Africans, they cannot account for the signals observed in African populations. With the absence of representative reference genomes, archaic and ghost introgression is often identified as divergent regions in the genome that contain strongly linked ancestral variants and/or show deep coalescence times. The sequences identified in this way might stem from an unsampled extinct ‘ghost’ population, or represent patterns arising from some alternative complex demography, that include deep stratification among past groups. However, with the analyses presented this far, it is difficult to differentiate between deep structure, real introgression from archaic and/or ghost populations, and statistical artifacts (76,77).
Admixture from ‘ghost’ populations and even ‘archaic groups’ has been suggested to explain particular patterns of variation in various African populations (31,35,39,62,68–71,75,78–82) (Table 1). It has been hypothesized that interbreeding with archaic populations happened frequently but at low levels (80). Especially west Africans have often been identified to harbor archaic ancestry. This signal has been shown to come from one or more populations that split from the ancestors of modern human approximately at the same time or slightly earlier than the Neanderthal split (35,39,62,68–71,75,81). Another signal, or an alternative explanation to the archaic introgression, has been proposed for west Africans, with introgression from a ghost population which split around 300 000 ya (31,39). The admixture signal from an archaic or ghost population in west Africa is consistent with the non-symmetrical relationship of the Khoe–San populations to west and east African populations (35). Furthermore, the differences in archaic ancestry proportions across Africa indicate that population structure can already have been established at the time of introgression, or alternatively, this observation can be a consequence of gene-flow from non-Africans (72). Some studies have also suggested that the introgressing sequences come from a population with a large Ne, which hints at it being a structured population itself (69,71). Another possibility is that the large Ne is shaped by multiple introgression events from divergent lineages, which are hard to distinguish without archaic reference sequences. Unfortunately, many studies of archaic or ghost introgression in Africa focus on a few populations and/or use only one method for inference, so that the effect of the identified archaic or ghost introgression is not yet comparable across all major branches of modern humans in a systematic way.
Table 1.
Overview archaic and ‘ghost’ introgression
| Publication | Split time of source population | Admixture recipient | Modern admixed populations | Introgression time | Admixture proportion (%) | Method |
|---|---|---|---|---|---|---|
| Plagnol and Wall (2006), (68) | Yoruba | 5 | S* | |||
| Wall et al. (2009), (75) | Yoruba | S* | ||||
| Wall et al. (2019), (82) | ancestors of KhoeSan | African populations included in the GenomeAsia 100K project | LD-based | |||
| Speidel et al. (2019), (70) | Yoruba | Relate | ||||
| Hsieh et al. (2016), (80) | 1.08 mya | Biaka, Baka | 9048 ya | S* | ||
| Durvasula and Sankararaman (2020), (71) | 625 kya | Yoruba, Esan, Gambian, Mende | 43 kya | 11 | CSFS | |
| Hammer et al. (2011), (78) | 700 kya | Biaka Pygmy, Mbuti Pygmy and San | 35 kya | 2 | S* | |
| Skoglund et al. (2017), (35) | Pre South African/other Africans split | Mende | 13 | qpgraph | ||
| Yoruba | 9 | |||||
| Ragsdale and Gravel (2019), (81) | 460−540 kya | Pre-split OOA branch | YRI | 90−160 kya | 4.7–9.2 | Multi-population two-locus diversity statistics |
| CEU, CHB | 1.9–6.6 | |||||
| Lipson et al. (2020), (39) | ~KhoeSan/RHG split | Ancestral west African branch | Lemande, Yoruba, Mende, Bantu | 10 | qpgraph | |
| Mende | Additional 3 | |||||
| Mota (ancient east African) | 29 | |||||
| ~Neanderthal/H. sapiens split | Ancestral west African lineage | Lemande, Yoruba, Mende, Bantu | 2 | |||
| Mende | Additional 1 | |||||
| Lorente-Galdos et al. (2019), (62) | 528 kya | KhoeSan | 3.8 | ABC-DL | ||
| Mbuti | 3.9 | |||||
| Mandenka | 5.8 | |||||
| Lachance et al. (2012), (79) | 1.2-1.3 mya | Hadza, Sandawe, WRHG | Predating the split of the populations | S* | ||
| Schlebusch et al. (2017), (31) | Pre ancient south African/ancient east African split | Yoruba | 31 | qpgraph | ||
| Hey et al. (2018), (69) | 850 kya | Ancestral branch of Yoruba, Baka, Hadza and Sandawe | Yoruba, Baka, Hadza, Sandawe | IMa3 |
The table gives an overview of all studies that have found archaic admixture or admixture from a ‘ghost’ source in African populations to date. ‘Admixture recipient’ shows which branch experienced the introgression event while ‘modern admixed populations’ show the populations that were investigated and showed the admixture signal.
Interestingly, many studies identified a fairly recent time for the introgression from extinct lineages in Africa, with introgression events even after the split from non-African populations (71,78,80,81), hinting at survival of archaic human populations until relatively recently in time. This contrasts with observations from the fossil record, where most of the morphological diversity disappeared before 100 000 ya.
Future Outlook
One of the questions that are still unresolved is how Homo sapiens arose. While it is clear that human origin, or emergence, is an evolutionary process over a relatively long time, we can narrow down the question of how humans originated by setting some time-limits. Starting from the present and going backward in time, the deepest divergence among humans—some 300 000 years ago—can be viewed as lower bound for the existence of fully developed (likely both behavioral and cognitive) modern humans (but not necessarily anatomically modern humans) simply because the descendant groups (that live today) of this first divergence certainly are modern humans. The human Neandertal/Denisovan divergence of some 650 000 years into the past can then be viewed as an upper bound of the development of unique features of Homo sapiens. In other words, by using these constraints, we can ask the question how humans developed unique characteristics from the time of the common ancestor of Neandertals/Denisovans till the starting time of human stratification observed today. Such unique characteristics may be governed by genetic changes, including frequency changes, and potentially epigenetic changes. There are various models of the origin of our species in Africa. Some of these include versions of (i) a single origin that expands and replaces all other hominin populations, (ii) an African multiregional model, where several African groups (likely geographically separated) are deeply stratified, possibly by isolation and (iii) range expansion from one group/area, but with some regional continuity and/or archaic introgression (83). The diverse morphology from fossil finds, material culture and genetics are hinting against a purely single origin of modern Homo sapiens in Africa (5,29,30), but possibly for emergence from a scattered diverse population with sporadic gene-flow (30,58). However, it is unclear how a model of a structured metapopulation can both uphold sufficient barriers to gene-flow to cause stratification while allowing for genetic variants important to modern human development to spread among populations simultaneously.
Disentangling the potentially complex early history of humans is a challenging task as later events, including various forms of gene-flow and migration, can distort genetic signals. More complex models, more diverse data and better statistical tools will allow us to look beyond simple bifurcating models and facilitate disentangling the intricate demography of African populations and inform on the models of the origin of our species as outlined in the previous paragraph. With efforts to gather more data, we still see a bias in data sampling; southern Africa in particular is currently overrepresented, both for modern and ancient genomic data. More genomic data from understudied regions and potential improvements in analyzing ancient DNA will refine our geographic and temporal understanding of African prehistory and allow us to decipher the early events.
Acknowledgements
This work was supported by the Knut and Alice Wallenberg Foundation [to M.J.], the Swedish Research Council [642-2013-8019 to M.J.] and SciLifeLab [postdoctoral fellowship to N.H.].
Conflict of interest statement. None declared.
Contributor Information
Nina Hollfelder, Human Evolution, Department of Organismal Biology, Uppsala University, Norbyvägen 18C, 75236 Uppsala, Sweden.
Gwenna Breton, Human Evolution, Department of Organismal Biology, Uppsala University, Norbyvägen 18C, 75236 Uppsala, Sweden.
Per Sjödin, Human Evolution, Department of Organismal Biology, Uppsala University, Norbyvägen 18C, 75236 Uppsala, Sweden.
Mattias Jakobsson, Human Evolution, Department of Organismal Biology, Uppsala University, Norbyvägen 18C, 75236 Uppsala, Sweden; Palaeo-Research Institute, University of Johannesburg, Physical, Cnr Kingsway & University Roads, Auckland Park, Johannesburg 2092, South Africa; SciLifeLab, Stockholm and Uppsala, Entrance C11, BMC, Husargatan 3, 752 37 Uppsala, Sweden.
References
- 1. Day, M.H. (1969) Omo human skeletal remains. Nature, 222, 1135. [DOI] [PubMed] [Google Scholar]
- 2. Grün, R., Brink, J.S., Spooner, N.A., Taylor, L., Stringer, C.B., Franciscus, R.G. and Murray, A.S. (1996) Direct dating of Florisbad hominid. Nature, 382, 500–501. [DOI] [PubMed] [Google Scholar]
- 3. White, T.D., Asfaw, B., DeGusta, D., Gilbert, H., Richards, G.D., Suwa, G. and Howell, F.C. (2003) Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature, 423, 742–747. [DOI] [PubMed] [Google Scholar]
- 4. McDougall, I., Brown, F.H. and Fleagle, J.G. (2008) Sapropels and the age of hominins Omo I and II, Kibish, Ethiopia. J. Hum. Evol., 55, 409–420. [DOI] [PubMed] [Google Scholar]
- 5. Stringer, C. (2016) The origin and evolution of Homo sapiens. Philos. Trans. R. Soc. B Biol. Sci., 371, 20150237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Hublin, J.-J., Ben-Ncer, A., Bailey, S.E., Freidline, S.E., Neubauer, S., Skinner, M.M., Bergmann, I., Le Cabec, A., Benazzi, S., Harvati, K. et al. (2017) New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens. Nature, 546, 289–292. [DOI] [PubMed] [Google Scholar]
- 7. Richter, D., Grün, R., Joannes-Boyau, R., Steele, T.E., Amani, F., Rué, M., Fernandes, P., Raynal, J.-P., Geraads, D., Ben-Ncer, A. et al. (2017) The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age. Nature, 546, 293–296. [DOI] [PubMed] [Google Scholar]
- 8. Berger, L.R. and Parkington, J.E. (1995) A new pleistocene hominid-bearing locality at Hoedjiespunt, South Africa. Am. J. Phys. Anthropol., 98, 601–609. [DOI] [PubMed] [Google Scholar]
- 9. Rightmire, G.P. (2009) Middle and later Pleistocene hominins in Africa and Southwest Asia. Proc. Natl. Acad. Sci., 106, 16046–16050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Gunz, P., Bookstein, F.L., Mitteroecker, P., Stadlmayr, A., Seidler, H. and Weber, G.W. (2009) Early modern human diversity suggests subdivided population structure and a complex out-of-Africa scenario. Proc. Natl. Acad. Sci., 106, 6094–6098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Harvati, K., Stringer, C., Grün, R., Aubert, M., Allsworth-Jones, P. and Folorunso, C.A. (2011) The later Stone Age calvaria from Iwo Eleru, Nigeria: morphology and chronology. PLoS One, 6, e24024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Dusseldorp, G., Lombard, M. and Wurz, S. (2013) Pleistocene Homo and the updated Stone Age sequence of South Africa. Pleistocene Homo and the updated Stone Age sequence of South Africa. S. Afr. J. Sci., 109, 1–7. [Google Scholar]
- 13. Berger, L.R., Hawks, J., Dirks, P.H.G.M., Elliott, M. and Roberts, E.M. (2017) Homo naledi and Pleistocene hominin evolution in subequatorial Africa. elife, 6, e24234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Dirks, P.H.G.M., Roberts, E.M., Hilbert-Wolf, H., Kramers, J.D., Hawks, J., Dosseto, A., Duval, M., Elliott, M., Evans, M., Grün, R. et al. (2017) The age of Homo naledi and associated sediments in the Rising Star Cave. South Africa. Elife, 6, e24231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Grün, R., Pike, A., McDermott, F., Eggins, S., Mortimer, G., Aubert, M., Kinsley, L., Joannes-Boyau, R., Rumsey, M., Denys, C. et al. (2020) Dating the skull from Broken Hill, Zambia, and its position in human evolution. Nature, 580, 372–375. [DOI] [PubMed] [Google Scholar]
- 16. Gurdasani, D., Carstensen, T., Tekola-Ayele, F., Pagani, L., Tachmazidou, I., Hatzikotoulas, K., Karthikeyan, S., Iles, L., Pollard, M.O., Choudhury, A. et al. (2015) The African genome variation project shapes medical genetics in Africa. Nature, 517, 327–332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Excoffier, L. and Schneider, S. (1999) Why hunter-gatherer populations do not show signs of Pleistocene demographic expansions. Proc. Natl. Acad. Sci., 96, 10597–10602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Kim, H.L., Ratan, A., Perry, G.H., Montenegro, A., Miller, W. and Schuster, S.C. (2014) Khoisan hunter-gatherers have been the largest population throughout most of modern-human demographic history. Khoisan hunter-gatherers have been the largest population throughout most of modern-human demographic history. Nat. Commun., 5, 1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Mallick, S., Li, H., Lipson, M., Mathieson, I., Gymrek, M., Racimo, F., Zhao, M., Chennagiri, N., Nordenfelt, S., Tandon, A. et al. (2016) The Simons genome diversity project: 300 genomes from 142 diverse populations. Nature, 538, 201–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Schlebusch, C.M. and Jakobsson, M. (2018) Tales of human migration, admixture, and selection in Africa. Annu. Rev. Genomics Hum. Genet., 19, 405–428. [DOI] [PubMed] [Google Scholar]
- 21. Bergström, A., McCarthy, S.A., Hui, R., Almarri, M.A., Ayub, Q., Danecek, P., Chen, Y., Felkel, S., Hallast, P., Kamm, J. et al. (2020) Insights into human genetic variation and population history from 929 diverse genomes. Science, 367, eaay5012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Breton, G., Schlebusch, C.M., Lombard, M., Sjödin, P., Soodyall, H. and Jakobsson, M. (2014) Lactase persistence alleles reveal partial east African ancestry of southern African Khoe pastoralists. Curr. Biol., 24, 852–858. [DOI] [PubMed] [Google Scholar]
- 23. Güldemann, T. and Fehn, A. (2014) Beyond’khoisan’: historical relations in the kalahari basin. In Current Issues in Linguistic Theory. Amsterdam, John Benjamins Publishing Company, Vol. 330, pp. 1–41. [Google Scholar]
- 24. Tishkoff, S.A., Gonder, M.K., Henn, B.M., Mortensen, H., Knight, A., Gignoux, C., Fernandopulle, N., Lema, G., Nyambo, T.B., Ramakrishnan, U. et al. (2007) History of click-speaking populations of Africa inferred from mtDNA and Y chromosome genetic variation. Mol. Biol. Evol., 24, 2180–2195. [DOI] [PubMed] [Google Scholar]
- 25. Henn, B.M., Gignoux, C.R., Jobin, M., Granka, J.M., Macpherson, J.M., Kidd, J.M., Rodríguez-Botigué, L., Ramachandran, S., Hon, L. and Brisbin, A. (2011) Hunter-gatherer genomic diversity suggests a southern African origin for modern humans. Proc. Natl. Acad. Sci., 108, 5154–5162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Pickrell, J.K., Patterson, N., Barbieri, C., Berthold, F., Gerlach, L., Güldemann, T., Kure, B., Mpoloka, S.W., Nakagawa, H., Naumann, C. et al. (2012) The genetic prehistory of southern Africa. Nat. Commun., 3, 1–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Scheinfeldt, L.B., Soi, S., Lambert, C., Ko, W.-Y., Coulibaly, A., Ranciaro, A., Thompson, S., Hirbo, J., Beggs, W., Ibrahim, M. et al. (2019) Genomic evidence for shared common ancestry of east African hunting-gathering populations and insights into local adaptation. Proc. Natl. Acad. Sci., 116, 4166–4175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Bahuchet, S. (2012) Changing language, remaining Pygmy. Hum. Biol., 84, 11–43. [DOI] [PubMed] [Google Scholar]
- 29. Schlebusch, C.M., Skoglund, P., Sjödin, P., Gattepaille, L.M., Hernandez, D., Jay, F., Li, S., De Jongh, M., Singleton, A., Blum, M.G.B. et al. (2012) Genomic variation in seven Khoe-San groups reveals adaptation and complex African history. Science, 338, 374–379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Scerri, E.M.L., Thomas, M.G., Manica, A., Gunz, P., Stock, J.T., Stringer, C., Grove, M., Groucutt, H.S., Timmermann, A., Rightmire, G.P. et al. (2018) Did our species evolve in subdivided populations across Africa, and why does it matter? Trends Ecol. Evol., 33, 582–594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Schlebusch, C.M., Malmström, H., Günther, T., Sjödin, P., Coutinho, A., Edlund, H., Munters, A.R., Vicente, M., Steyn, M., Soodyall, H. et al. (2017) Southern African ancient genomes estimate modern human divergence to 350,000 to 260,000 years ago. Science, 358, 652–655. [DOI] [PubMed] [Google Scholar]
- 32. Gallego Llorente, M., Jones, E.R., Eriksson, A., Siska, V., Arthur, K.W., Arthur, J.W., Curtis, M.C., Stock, J.T., Coltorti, M., Pieruccini, P. et al. (2015) Ancient Ethiopian genome reveals extensive Eurasian admixture throughout the African continent. Science, 350, 820–822. [DOI] [PubMed] [Google Scholar]
- 33. Rodríguez-Varela, R., Günther, T., Krzewińska, M., Storå, J., Gillingwater, T.H., MacCallum, M., Arsuaga, J.L., Dobney, K., Valdiosera, C., Jakobsson, M. et al. (2017) Genomic analyses of pre-European conquest human remains from the Canary Islands reveal close affinity to modern north Africans. Curr. Biol., 27, e5, 3396–3402. [DOI] [PubMed] [Google Scholar]
- 34. Schuenemann, V.J., Peltzer, A., Welte, B., van Pelt, W.P., Molak, M., Wang, C.-C., Furtwängler, A., Urban, C., Reiter, E., Nieselt, K. et al. (2017) Ancient Egyptian mummy genomes suggest an increase of sub-Saharan African ancestry in post-roman periods. Nat. Commun., 8, 15694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Skoglund, P., Thompson, J.C., Prendergast, M.E., Mittnik, A., Sirak, K., Hajdinjak, M., Salie, T., Rohland, N., Mallick, S., Peltzer, A. et al. (2017) Reconstructing prehistoric African population structure. Cell, 171, 59–71.e21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Fregel, R., Méndez, F.L., Bokbot, Y., Martín-Socas, D., Camalich-Massieu, M.D., Santana, J., Morales, J., Ávila-Arcos, M.C., Underhill, P.A., Shapiro, B. et al. (2018) Ancient genomes from north Africa evidence prehistoric migrations to the Maghreb from both the Levant and Europe. Proc. Natl. Acad. Sci., 115, 6774–6779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. van de Loosdrecht, M., Bouzouggar, A., Humphrey, L., Posth, C., Barton, N., Aximu-Petri, A., Nickel, B., Nagel, S., Talbi, E.H., El Hajraoui, M.A. et al. (2018) Pleistocene north African genomes link near eastern and sub-Saharan African human populations. Science, 360, 548–552. [DOI] [PubMed] [Google Scholar]
- 38. Prendergast, M.E., Lipson, M., Sawchuk, E.A., Olalde, I., Ogola, C.A., Rohland, N., Sirak, K.A., Adamski, N., Bernardos, R., Broomandkhoshbacht, N. et al. (2019) Ancient DNA reveals a multistep spread of the first herders into sub-Saharan Africa. Science, 364, eaaw6275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Lipson, M., Ribot, I., Mallick, S., Rohland, N., Olalde, I., Adamski, N., Broomandkhoshbacht, N., Lawson, A.M., López, S., Oppenheimer, J. et al. (2020) Ancient west African foragers in the context of African population history. Nature, 577, 665–670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Wang, K., Goldstein, S., Bleasdale, M., Clist, B., Bostoen, K., Bakwa-Lufu, P., Buck, L.T., Crowther, A., Dème, A., McIntosh, R.J. et al. (2020) Ancient genomes reveal complex patterns of population movement, interaction, and replacement in sub-Saharan Africa. Sci. Adv., 6, eaaz0183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Scerri, E.M.L., Drake, N.A., Jennings, R. and Groucutt, H.S. (2014) Earliest evidence for the structure of Homo sapiens populations in Africa. Quat. Sci. Rev., 101, 207–216. [Google Scholar]
- 42. Schlebusch, C.M., Sjödin, P., Breton, G., Günther, T., Naidoo, T., Hollfelder, N., Sjöstrand, A.E., Xu, J., Gattepaille, L.M., Vicente, M. et al. (2020) Khoe-san genomes reveal unique variation and confirm the deepest population divergence in Homo sapiens. Mol. Biol. Evol., 37, 2944–2954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Behar, D.M., Villems, R., Soodyall, H., Blue-Smith, J., Pereira, L., Metspalu, E., Scozzari, R., Makkan, H., Tzur, S., Comas, D. et al. (2008) The dawn of human matrilineal diversity. Am. J. Hum. Genet., 82, 1130–1140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Naidoo, T., Schlebusch, C.M., Makkan, H., Patel, P., Mahabeer, R., Erasmus, J.C. and Soodyall, H. (2010) Development of a single base extension method to resolve Y chromosome haplogroups in sub-Saharan African populations. Investig. Genet., 1, 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Barbieri, C., Vicente, M., Rocha, J., Mpoloka, S.W., Stoneking, M. and Pakendorf, B. (2013) Ancient substructure in early mtDNA lineages of southern Africa. Am. J. Hum. Genet., 92, 285–292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Schlebusch, C.M., Lombard, M. and Soodyall, H. (2013) MtDNA control region variation affirms diversity and deep sub-structure in populations from southern Africa. BMC Evol. Biol., 13, 56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Li, J.Z., Absher, D.M., Tang, H., Southwick, A.M., Casto, A.M., Ramachandran, S., Cann, H.M., Barsh, G.S., Feldman, M., Cavalli-Sforza, L.L. et al. (2008) Worldwide human relationships inferred from genome-wide patterns of variation. Science, 319, 1100–1104. [DOI] [PubMed] [Google Scholar]
- 48. Tishkoff, S., Reed, F., Friedlaender, F. and Ehret, C. (2009) The genetic structure and history of Africans and African Americans. Science, 324, 1035–1044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Gronau, I., Hubisz, M.J., Gulko, B., Danko, C.G. and Siepel, A. (2011) Bayesian inference of ancient human demography from individual genome sequences. Nat. Genet., 43, 1031–1034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Veeramah, K.R., Wegmann, D., Woerner, A., Mendez, F.L., Watkins, J.C., Destro-Bisol, G., Soodyall, H., Louie, L. and Hammer, M.F. (2012) An early divergence of KhoeSan ancestors from those of other modern humans is supported by an ABC-based analysis of autosomal resequencing data. Mol. Biol. Evol., 29, 617–630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Vicente, M., Jakobsson, M., Ebbesen, P., Schlebusch, C.M. and Heyer, E. (2019) Genetic affinities among southern Africa hunter-gatherers and the impact of admixing farmer and herder populations. Mol. Biol. Evol., 36, 1849–1861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Fan, S., Kelly, D.E., Beltrame, M.H., Hansen, M.E.B., Mallick, S., Ranciaro, A., Hirbo, J., Thompson, S., Beggs, W., Nyambo, T. et al. (2019) African evolutionary history inferred from whole genome sequence data of 44 indigenous African populations. Genome Biol., 20, 82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Gopalan, S., Berl, R.E.W., Belbin, G., Gignoux, C.R., Feldman, M.W., Hewlett, B.S. and Henn, B.M. (2019) Hunter-gatherer genomes reveal diverse demographic trajectories following the rise of farming in east Africa. bioRxiv, 517730. [Google Scholar]
- 54. Patin, E., Siddle, K.J., Laval, G., Quach, H., Harmant, C., Becker, N., Froment, A., Regnault, B., Lemee, L., Gravel, S. et al. (2014) The impact of agricultural emergence on the genetic history of African rainforest hunter-gatherers and agriculturalists. Nat. Commun., 5, 3163. [DOI] [PubMed] [Google Scholar]
- 55. Jakobsson, M., Scholz, S.W., Scheet, P., Gibbs, J.R., VanLiere, J.M., Fung, H.-C., Szpiech, Z.A., Degnan, J.H., Wang, K., Guerreiro, R. et al. (2008) Genotype, haplotype and copy-number variation in worldwide human populations. Nature, 451, 998–1003. [DOI] [PubMed] [Google Scholar]
- 56. Henn, B.M., Botigue, L.R., Gravel, S., Wang, W., Brisbin, A., Byrnes, J.K., Fadhlaoui-Zid, K., Zalloua, P.A., Moreno-Estrada, A., Bertranpetit, J. et al. (2012) Genomic ancestry of north Africans supports back-to-Africa migrations. PLoS Genet., 8, e1002397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Breton, G. (2020) Human demographic history: insights on the human past based on genomes from southern through Central Africa. Human Evolution, Department of Organismal Biology, Biology, Disciplinary Domain of Science and Technology, Uppsala University, Acta Universitatis Upsaliensis, Uppsala.
- 58. Scerri, E.M.L., Chikhi, L. and Thomas, M.G. (2019) Beyond multiregional and simple out-of-Africa models of human evolution. Nat. Ecol. Evol., 3, 1370–1372. [DOI] [PubMed] [Google Scholar]
- 59. Mazet, O., Rodríguez, W., Grusea, S., Boitard, S. and Chikhi, L. (2016) On the importance of being structured: instantaneous coalescence rates and human evolution—lessons for ancestral population size inference? Heredity, 116, 362–371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Schiffels, S. and Durbin, R. (2014) Inferring human population size and separation history from multiple genome sequences. Nat. Genet., 46, 919–925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Malaspinas, A.-S., Westaway, M.C., Muller, C., Sousa, V.C., Lao, O., Alves, I., Bergström, A., Athanasiadis, G., Cheng, J.Y., Crawford, J.E. et al. (2016) A genomic history of aboriginal Australia. Nature, 538, 207–214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Lorente-Galdos, B., Lao, O., Serra-Vidal, G., Santpere, G., Kuderna, L.F.K., Arauna, L.R., Fadhlaoui-Zid, K., Pimenoff, V.N., Soodyall, H., Zalloua, P. et al. (2019) Whole-genome sequence analysis of a pan African set of samples reveals archaic gene flow from an extinct basal population of modern humans into sub-Saharan populations. Genome Biol., 20, 77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Song, S., Sliwerska, E., Emery, S. and Kidd, J.M. (2017) Modeling human population separation history using physically phased genomes. Genetics, 205, 385–395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Hsieh, P.H., Veeramah, K.R., Lachance, J., Tishkoff, S.A., Wall, J.D., Hammer, M.F. and Gutenkunst, R.N. (2016) Whole-genome sequence analyses of western central African Pygmy hunter-gatherers reveal a complex demographic history and identify candidate genes under positive natural selection. Genome Res., 26, 279–290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Patin, E., Laval, G., Barreiro, L.B., Salas, A., Semino, O., Santachiara-Benerecetti, S., Kidd, K.K., Kidd, J.R., Van der Veen, L., Hombert, J.-M. et al. (2009) Inferring the demographic history of African farmers and Pygmy hunter–gatherers using a multilocus resequencing data set. PLoS Genet., 5, e1000448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Reich, D., Green, R.E., Kircher, M., Krause, J., Patterson, N., Durand, E.Y., Viola, B., Briggs, A.W., Stenzel, U., Johnson, P.L.F. et al. (2010) Genetic history of an archaic hominin group from Denisova cave in Siberia. Nature, 468, 1053–1060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Green, R.E., Krause, J., Briggs, A.W., Maricic, T., Stenzel, U., Kircher, M., Patterson, N., Li, H., Zhai, W., Fritz, M.H.Y. et al. (2010) A draft sequence of the neandertal genome. Science, 328, 710–722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Plagnol, V. and Wall, J.D. (2006) Possible ancestral structure in human populations. PLoS Genet., 2, e105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Hey, J., Chung, Y., Sethuraman, A., Lachance, J., Tishkoff, S., Sousa, V.C. and Wang, Y. (2018) Phylogeny estimation by integration over isolation with migration models. Mol. Biol. Evol., 35, 2805–2818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Speidel, L., Forest, M., Shi, S. and Myers, S.R. (2019) A method for genome-wide genealogy estimation for thousands of samples. Nat. Genet., 51, 1321–1329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Durvasula, A. and Sankararaman, S. (2020) Recovering signals of ghost archaic introgression in African populations. Sci. Adv., 6, eaax5097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Chen, L., Wolf, A.B., Fu, W., Li, L. and Akey, J.M. (2020) Identifying and interpreting apparent Neanderthal ancestry in African individuals. Cell, 180, 677–687.e16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Wang, S., Lachance, J., Tishkoff, S.A., Hey, J. and Xing, J. (2013) Apparent variation in Neanderthal admixture among African populations is consistent with gene flow from non-African populations. Genome Biol. Evol., 5, 2075–2081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Hollfelder, N., Schlebusch, C.M., Günther, T., Babiker, H., Hassan, H.Y. and Jakobsson, M. (2017) Northeast African genomic variation shaped by the continuity of indigenous groups and Eurasian migrations. PLoS Genet., 13, e1006976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Wall, J.D., Lohmueller, K.E. and Plagnol, V. (2009) Detecting ancient admixture and estimating demographic parameters in multiple human populations. Mol. Biol. Evol., 26, 1823–1827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Eriksson, A. and Manica, A. (2012) Effect of ancient population structure on the degree of polymorphism shared between modern human populations and ancient hominins. Proc. Natl. Acad. Sci., 109, 13956–13960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Chikhi, L., Rodríguez, W., Grusea, S., Santos, P., Boitard, S. and Mazet, O. (2018) The IICR (inverse instantaneous coalescence rate) as a summary of genomic diversity: insights into demographic inference and model choice. Heredity, 120, 13–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Hammer, M.F., Woerner, A.E., Mendez, F.L., Watkins, J.C. and Wall, J.D. (2011) Genetic evidence for archaic admixture in Africa. Proc. Natl. Acad. Sci. USA, 108, 15123–15128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Lachance, J., Vernot, B., Elbers, C.C., Ferwerda, B., Froment, A., Bodo, J.-M., Lema, G., Fu, W., Nyambo, T.B., Rebbeck, T.R. et al. (2012) Evolutionary history and adaptation from high-coverage whole-genome sequences of diverse African hunter-gatherers. Cell, 150, 457–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Hsieh, P.H., Woerner, A.E., Wall, J.D., Lachance, J., Tishkoff, S.A., Gutenkunst, R.N. and Hammer, M.F. (2016) Model-based analyses of whole-genome data reveal a complex evolutionary history involving archaic introgression in central African Pygmies. Genome Res., 26, 291–300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Ragsdale, A.P. and Gravel, S. (2019) Models of archaic admixture and recent history from two-locus statistics. PLoS Genet., 15, e1008204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Wall, J.D., Ratan, A., Stawiski, E. and GenomeAsia 100K Consortium (2019) Identification of African-specific admixture between modern and archaic humans. Am. J. Hum. Genet., 105, 1254–1261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Henn, B.M., Steele, T.E. and Weaver, T.D. (2018) Clarifying distinct models of modern human origins in Africa. Curr. Opin. Genet. Dev., 53, 148–156. [DOI] [PubMed] [Google Scholar]
- 84. Sjödin, P., McKenna, J. and Jakobsson, M. (2020) Estimating divergence times from DNA sequences. Genetics, in press. [DOI] [PMC free article] [PubMed] [Google Scholar]