

TO THE EDITOR:
Sickle cell disease (SCD) is a devastating disease characterized by a single-nucleotide mutation in the β-globin chain, encoding the production of an abnormal type of hemoglobin (Hb): hemoglobin S (HbS). HbS polymerizes upon deoxygenation, causing red blood cells (RBC) to sickle. Sickled RBCs are poorly deformable, which leads to vasoocclusion and hemolytic anemia. In addition, increased red cell adhesion, endothelial dysfunction, inflammation, oxidative stress, hemostatic activation, and rheological abnormalities all contribute to the complex pathophysiology of SCD.1 Among the factors that influence sickling are RBC metabolic intermediates, in particular, levels of 2,3-diphosphyglycerate (2,3-DPG) and adenosine triphosphate (ATP). 2,3-DPG is produced in the Rapoport-Luebering shunt, a unique RBC-specific glycolytic bypass, and serves as an important regulator of oxygen affinity of Hb. The increased intracellular 2,3-DPG levels lower oxygen affinity, thereby promoting polymerization of HbS upon deoxygenation and, hence, sickling.2 ATP is critical for maintaining RBC membrane integrity and deformability, and ∼50% of the cell’s ATP is generated in the last step of glycolysis catalyzed by pyruvate kinase (PK). Decreased levels of ATP have been reported in SCD mice,3 and ATP depletion has been associated with an increased number of irreversibly sickled cells.4 Interestingly, these metabolic changes are strongly reminiscent of key metabolic changes observed in PK deficiency, a rare inherited glycolytic enzymopathy that is caused by mutations in the PKLR gene and that is associated with nonspherocytic hemolytic anemia.5 PK deficiency results in decreased levels of ATP, whereas retrograde accumulation of glycolytic intermediates leads to increased levels of 2,3-DPG.6 Mitapivat is an allosteric activator of PK and currently is in phase 3 clinical trials for PK deficiency (#NCT02476916, #NCT03548220, #NCT03559699, #NCT03853798) and in phase 1 trial for SCD (#NCT4000165). This small molecule directly targets PK by binding in a pocket at the dimer-dimer interface, resulting in enhanced activity of both wild-type and mutant PK.7 Both phase 1 and 2 studies in healthy volunteers and patients who were PK deficient demonstrated glycolytic pathway activation upon treatment with mitapivat and confirmed safety and efficacy.8,9 In this study, we investigated key properties of PK in SCD, and the effect of ex vivo treatment with mitapivat on PK properties, metabolic features, and sickling behavior.
Whole blood from patients with SCD and healthy controls was collected according to protocol and approved by the Ethical Committee of UMC Utrecht (17-450/M and 17-392/M). Unless stated otherwise, blood was collected in EDTA. Routine hematological parameters were measured using the Cell-Dyn Sapphire (Abbott Diagnostics). Fetal Hb and HbS levels were measured by high-performance liquid chromatography (Tosoh G8) during routine visits to the outpatient clinic. PK and hexokinase (HK) activity measurements, PK protein levels, and thermostability were performed on RBCs purified from whole blood using microcrystalline cellulose-α-cellulose.10 PK thermostability was measured on lysates after 1 hour of incubation (53°C) and expressed as percentage residual enzymatic activity (final substrate concentration, 5 mM) after incubation.6 PK protein levels were determined by Mesoscale Assay (MesoScale Discovery) as described.6 Quantitative analysis of ATP and 2,3-DPG was performed as described.6 Sickling behavior was assessed on whole blood, standardized to a fixed RBC count of 200 × 06 per 5 mL Oxy-iso by oxygen gradient ektacytometry on the Laser Optical Rotational Red Cell Analyzer (Lorrca; RR Mechatronics) with oxygenscan add-on.11,12 The point of sickling (PoS) indicates the specific pO2 at which RBCs start to sickle. For ex vivo treatment with mitapivat, purified RBCs were incubated for 24 hours at 37°C in the presence or absence of 10 µM mitapivat as described.6 After 24 hours, PK activity and ATP levels (CellTiterGlo; Promega) were measured. Oxygen gradient ektacytometry, ATP/2,3-DPG levels, p50 (Hemox analyzer TCS), and PK thermostability6 measurements (final substrate concentration, 0.5 mM) were performed on buffy coat–depleted whole blood (0.2 hematocrit), collected in citrate-phosphate-dextrose solution with adenine, and incubated for 22 hours in AGAM buffer,6 in the presence or absence of 50 or 100 µM mitapivat. Incubation was performed overnight (18 hours) at 4°C, after which samples were allowed to recover at room temperature (1 hour), and 37°C (3 hours).
Baseline patient characteristics, RBC features, and PK features of 22 patients with SCD and 10 healthy controls are shown in Table 1. Reticulocytosis in patients with SCD was accompanied by increased activity of HK, but PK activity, which is also age related, was found to be decreased, as reflected by a decreased PK/HK ratio13,14 (Table 1; Figure 1A). This deficiency of PK was accompanied by a substantial decrease in PK thermostability in patients with SCD (Table 1; Figure 1B) and a decreased ATP/2,3-DPG ratio (Table 1; Figure 1C). At the same time, PK protein levels were increased (Table 1), likely because of the relatively young age of the RBC population (positive correlation with reticulocyte count, r = 0.655, P < .0001, and inverse correlation with Hb, r = −0.785, P < .0001). Together, these findings imply that enzyme activity and stability, as well as red cell metabolism, is compromised in SCD, similar to what is observed in PK-deficient patients.6 PK thermostability inversely correlated with reticulocytes, that is, patients displaying the lowest degree of PK stability had the highest reticulocyte count (R = −0.797, P < .0001). This suggests that a lower degree of PK stability is associated with more severe anemia, which is underlined by the correlation with Hb (r = 0.731, P < .0001). In addition, PK thermostability also inversely correlated with PoS, indicating that decreased PK thermostability is associated with sickling at a higher pO2 (r = −0.559, P = .007; Figure 1D). Notably, this association between PK thermostability and clinical severity was not observed in patients with SCD followed after starting HU therapy: Despite significant decreases in PoS of 16% and 19% compared with baseline after 3 and 6 months, respectively (supplemental Figure 1A, available on the Blood Web site), PK thermostability remained unchanged (baseline: 51%, 3 months, 54%, 6 months, 55%; supplemental Figure 1B). This indicates that the clinical benefit of HU is independent of PK properties. We next investigated if PK activity from patients with SCD could be enhanced by mitapivat and whether this would be associated with an improvement of RBC metabolism and functional properties. Upon ex vivo treatment with mitapivat, an increase in PK activity was seen in purified RBCs of patients and controls, with a mean increase of 129% in patients with SCD (range, 113% to 148%), and 144% in controls (range, 111% to 188%; Figure 1E). Accordingly, mean ATP levels increased 131% in SCD patients (range, 123% to 139%), and 133% in controls (range, 101% to 155%; Figure 1F). This shows that mitapivat can bind PK from patients with SCD like healthy controls, and that this small molecule activator can elicit an allosteric response upon binding, resulting in enhanced production of ATP. Further evidence for functional improvement was obtained from a series of experiments on buffy coat–depleted whole blood from both nontransfused and regularly transfused patients with SCD (blood collected just before transfusion). This showed that ex vivo treatment with mitapivat considerably restored PK thermostability (79% residual activity compared with 32% for vehicle control; Figure 1G) and led to a 25% increase in ATP/2,3-DPG ratio (Figure 1H). This was mainly due to a 17% decrease in 2,3-DPG levels (supplemental Figure 2A), whereas the 5% increase in ATP was nonsignificant under these conditions (supplemental Figure 2B). In line with the decrease in 2,3-DPG, there was 5% decrease in p50 (Figure 1I), which was accompanied by a functional improvement of SCD RBCs, as illustrated by a significant 9% decrease in PoS compared with untreated samples (Figure 1K, representative curve depicted in Figure 1J). In addition, we observed a modest but significant 2% increase in RBC deformability at normoxic conditions (EImax), indicative of improved RBC membrane health of SCD RBCs treated with mitapivat (Figure 1L). Importantly, comparable results were obtained for nontransfused and transfused patients, and there was no statistical difference in response between these 2 groups (supplemental Figure 3A-E).
Table 1.
Baseline characteristics of patients with SCD and controls
| Patients with SCD* (n = 22) | Controls (n = 10) | |
|---|---|---|
| Patient-related features | ||
| Sex, female, n (%) | 9 (40.9) | 9 (90.0) |
| Age, y | 23 (7-57) | 48 (26-63) |
| α-Thalassemia, n (%) | 3 (13.6) | N/D |
| Splenectomy, n (%) | 1 (4.5) | N/D |
| HU, n (%) | 13 (59.1) | N/D |
| Chronic transfusion, n (%) | 0 (0.0) | N/D |
| Red cell features | ||
| HbF, % | 7.4 (0.6-28.8) | 0.5 (0.4-0.6) |
| HbS, % | 81.3 (59.4-89.3) | 0.0 (0.0-0.0) |
| Hb, g/dL | 9.4 (7.1-12.3) | 14.5 (12.3-17.4) |
| Reticulocytes, % | 9.1 (4.0-21.9) | 1.4 (0.7-2.3) |
| RBC count, 1012/L | 2.9 (2.2-4.4) | 4.7 (3.7-5.4) |
| MCV, fl | 82.3 (69.4-125) | 92.7 (83.8-105) |
| MCHC, g/dL | 34.9 (33.4-38.2) | 34.4 (31.9-37.5) |
| PK features | ||
| PK activity, U/gHb | 13.7 (7.5-22.4)† | 7.9 (5.3-8.7) |
| HK activity, U/gHb | 2.6 (1.2-4.9)† | 0.8 (0.5-1.0) |
| PK/HK ratio | 5.1 (2.5-9.9)† | 8.9 (7.4-14.0) |
| PK-thermostability, % residual activity | 54.9 (35.8.2-80.8)† | 76.3 (72.6-80.9) |
| PK protein, AU/gHb | 146 (105-176)† | 102 (78-124) |
| PKLR mutations | None detected | N/D |
| Metabolic features* | ||
| 2,3-DPG, 10−3 μg/gHb | 4.6 (3.5-6.3)† | 3.3 (2.8-4.1) |
| ATP, 10−3 μg/gHb | 2.0 (1.9-3.0) | 2.0 (1.6-2.5) |
| ATP/2,3-DPG ratio | 0.44 (0.34-0.67)‡ | 0.57 (0.46-0.81) |
Numbers represent median (range) unless stated otherwise.
HU, hydroxyurea; MCHC, mean corpuscular hemoglobin concentration; MCV, mean corpuscular volume; N/D, not determined.
Metabolic features were obtained from 14/22 patients with SCD.
P < .0001.
P < .01.
Figure 1.
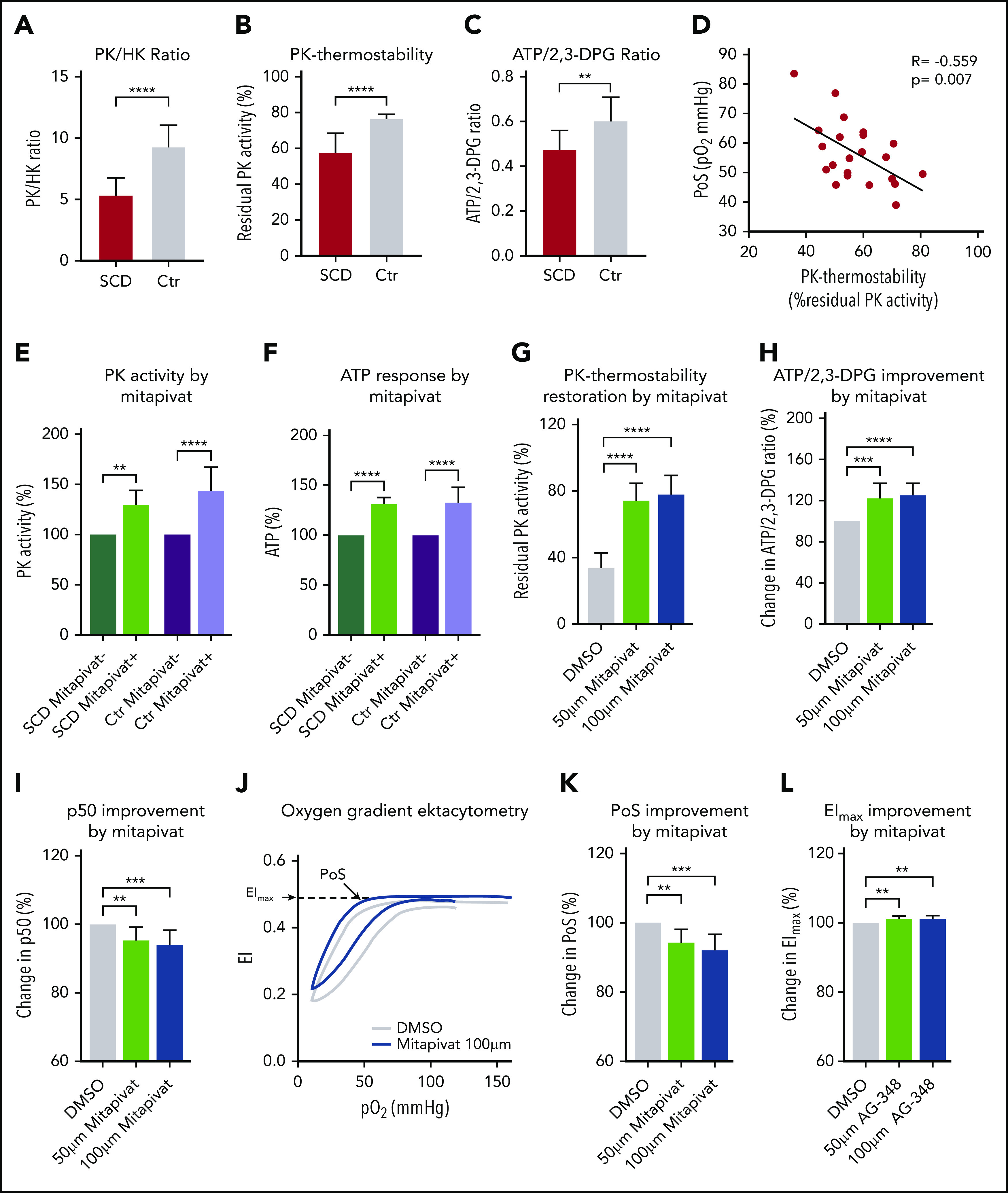
PK activity, thermostability, and cellular metabolism is compromised in RBCs from patients with SCD and correlates with markers of clinical severity. (A) PK/HK ratio, as a means to evaluate PK activity in the presence of high number of reticulocytes, is decreased in RBCs from patients with SCD (red, N = 22) compared with healthy controls (Ctr; gray, N = 10), indicating a deficiency of PK. (B-C) The deficiency of PK is accompanied by a decrease in PK thermostability in RBCs of patients with SCD (N = 22) compared with controls (N = 10) (B), as well as a decrease in ATP/2,3-DPG ratio in patients with SCD (N = 14) compared with healthy controls (N = 10) (C). (D) PK thermostability correlates with PoS (pO2 at which RBCs start to sickle) in patients with SCD (N = 22), a novel potential biomarker of clinical severity determined by oxygen gradient ektacytometry.20 (E-F) Ex vivo treatment of purified SCD RBCs with mitapivat increases PK activity and ATP levels. PK activation and ATP response (%) were assessed from purified RBCs of patients with SCD (green, N = 7) and 14 healthy controls (Ctr; purple, N = 14) after 24 hours’ incubation in the presence (light green and light purple) or absence (dark green and dark purple) of 10 μM mitapivat. An increase in PK activity is seen in patients with SCD (mean increase, 129%; range, 113% to 148%) and controls (mean increase, 144%; range, 111% to 188%) (E). This increase is accompanied by an increase in ATP response in patients with SCD (mean increase, 131%; range, 123% to 139%) and controls (mean increase, 133%; range, 101% to 155%) (F). (G-L) Ex vivo treatment of buffy coat–depleted whole blood with mitapivat restores PK thermostability, increases ATP/2,3-DPG ratio, decreases p50 and PoS, and increases membrane health. PK thermostability (% residual PK activity), after 22 hours’ incubation in the presence of 50 μM (light green) and 100 μM (dark blue) mitapivat, is restored to 79% (100 μM) compared with vehicle controls (32%; dimethyl sulfoxide [DMSO] control; light gray, N = 11) (G), causing a 25% increase in ATP/2,3-DPG ratio (N = 11) (H) and a 5% decrease in p50 (N = 11) (I). (J) Representative oxygen gradient ektacytometry curve shows improved PoS and EImax after incubation in the presence of 100 μM mitapivat (dark blue) compared with vehicle control (light gray). (K-L) PoS and EImax of 9 patients with SCD improve with 9% and 2%, respectively, after 22 hours’ incubation in the presence of 100 μM (dark blue) mitapivat compared with vehicle controls (light gray). Error bars represent standard deviation. ****P < .0001; ***P < .001; **P < .01.
In summary, we report on a novel finding in SCD RBCs: PK is less stable and less active compared with healthy control RBCs. The mechanism involved is yet to be determined but could involve altered intracellular redox state and subsequent inhibition of the enzyme through oxidation of cysteine residues.15,16 Defective PK function and, hence, compromised RBC metabolism therefore could contribute to the complex pathophysiology of SCD. Notably, the importance of PK function in SCD is underlined by a case report on a combination of PK deficiency and sickle cell trait causing an acute sickling syndrome.17 Our findings furthermore demonstrate that ex vivo treatment with mitapivat restores PK activity and PK thermostability, reduces 2,3-DPG levels, decreases p50, and subsequently reduces sickling behavior. Targeting PK by mitapivat treatment therefore represents a potential novel therapeutic option for SCD,18,19 by itself or, because HU treatment does not affect PK properties, synergistically with HU treatment or transfusion. Clinical trials are warranted to investigate whether mitapivat treatment will ameliorate clinical features, such as hemolysis, vaso-occlusive episodes, and anemia.
Supplementary Material
The online version of this article contains a data supplement.
Acknowledgments
The authors sincerely thank the patients who participated in this study.
This research has been funded, in part, by Agios Pharmaceuticals Inc and the Eurostars Program (SCOOP, ESTAR18105).
Footnotes
For original data, please e-mail the corresponding author at r.vanwijk@umcutrecht.nl.
Authorship
Contribution: The concept and design of the study were developed by M.A.E.R., S.v.S., L.D., C.K., E.J.v.B., and R.v.W.; M.A.E.R., J.B., B.A.v.O., P.A.K., and C.K. developed methodology; M.A.E.R., J.B., B.A.v.O., P.A.K., V.C., H.K., and H.M. acquired data; M.A.E.R., J.B., B.A.v.O., P.A.K., V.C., H.K., H.M., L.D., C.K., and R.v.W. analyzed and interpreted data; M.A.E.R. and E.J.v.B. provided patient data; M.A.E.R. and R.v.W. wrote the manuscript; and J.B., B.A.v.O., S.v.S., P.A.K., H.M., R.E.G.S., G.P., L.D., C.K., and E.J.v.B. reviewed and/or revised the manuscript.
Conflict-of-interest disclosure: E.J.v.B. and R.v.W. are consultants for Agios Pharmaceuticals. The remaining authors declare no competing financial interests.
Correspondence: Richard van Wijk, Central Diagnostic Laboratory–Research, University Medical Center Utrecht, Room G.03.550, PO Box 85500, 3508 GA Utrecht, The Netherlands; e-mail: r.vanwijk@umcutrecht.nl.
REFERENCES
- 1.Piel FB, Steinberg MH, Rees DC. Sickle cell disease. N Engl J Med. 2017;376(16):1561-1573. [DOI] [PubMed] [Google Scholar]
- 2.Charache S, Grisolia S, Fiedler AJ, Hellegers AE. Effect of 2,3-diphosphoglycerate on oxygen affinity of blood in sickle cell anemia. J Clin Invest. 1970;49(4):806-812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Banerjee T, Kuypers FA. Reactive oxygen species and phosphatidylserine externalization in murine sickle red cells. Br J Haematol. 2004;124(3):391-402. [DOI] [PubMed] [Google Scholar]
- 4.Jensen M, Shohet SB, Nathan DG. The role of red cell energy metabolism in the generation of irreversibly sickled cells in vitro. Blood. 1973;42(6):835-842. [PubMed] [Google Scholar]
- 5.Grace RF, Zanella A, Neufeld EJ, et al. Erythrocyte pyruvate kinase deficiency: 2015 status report. Am J Hematol. 2015;90(9):825-830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Rab MAE, van Oirschot BA, Kosinski PA, et al. AG-348 (Mitapivat), an allosteric activator of red blood cell pyruvate kinase, increases enzymatic activity, protein stability, and ATP levels over a broad range of PKLR genotypes. Haematologica. 2021;106(1):238-249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kung C, Hixon J, Kosinski PA, et al. AG-348 enhances pyruvate kinase activity in red blood cells from patients with pyruvate kinase deficiency. Blood. 2017;130(11):1347-1356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Yang H, Merica E, Chen Y, et al. Phase 1 single- and multiple-ascending-dose randomized studies of the safety, pharmacokinetics, and pharmacodynamics of AG-348, a first-in-class allosteric activator of pyruvate kinase R, in healthy volunteers. Clin Pharmacol Drug Dev. 2019;8(2):246-259. [DOI] [PubMed] [Google Scholar]
- 9.Grace RF, Rose C, Layton DM, et al. Safety and efficacy of mitapivat in pyruvate kinase deficiency. N Engl J Med. 2019;381(10):933-944. [DOI] [PubMed] [Google Scholar]
- 10.Beutler E. Red cell metabolism. A manual of biochemical methods. 3rd ed. Orlando: Grune & Stratton; 1984. [Google Scholar]
- 11.Rab MAE, van Oirschot BA, Bos J, et al. Rapid and reproducible characterization of sickling during automated deoxygenation in sickle cell disease patients. Am J Hematol. 2019;94(5):575-584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rab MAE, Kanne CK, Bos J, et al. Methodological aspects of the oxygenscan in sickle cell disease: a need for standardization. Am J Hematol. 2020;95(1):E5-E8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bianchi P, Fermo E, Glader B, et al. ; with the endorsement of EuroBloodNet, the European Reference Network in Rare Hematological Diseases . Addressing the diagnostic gaps in pyruvate kinase deficiency: consensus recommendations on the diagnosis of pyruvate kinase deficiency. Am J Hematol. 2019;94(1):149-161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Al-Samkari H, Addonizio K, Glader B, et al. The pyruvate kinase (PK) to hexokinase enzyme activity ratio and erythrocyte PK protein level in the diagnosis and phenotype of PK deficiency. Br J Haematol. 2021;192(6):1092-1096. [DOI] [PubMed] [Google Scholar]
- 15.Sprengers ED, van Berkel TJ, Koster JF, Staal GE. Influence of the rebox state of glutathione upon pyruvate kinase in the intact erythrocyte. Clin Chim Acta. 1977;80(3):495-502. [DOI] [PubMed] [Google Scholar]
- 16.Anastasiou D, Poulogiannis G, Asara JM, et al. Inhibition of pyruvate kinase M2 by reactive oxygen species contributes to cellular antioxidant responses. Science. 2011;334(6060):1278-1283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Alli N, Coetzee M, Louw V, et al. Sickle cell disease in a carrier with pyruvate kinase deficiency. Hematology. 2008;13(6):369-372. [DOI] [PubMed] [Google Scholar]
- 18.Nardo-Marino A, Brousse V, Rees D. Emerging therapies in sickle cell disease. Br J Haematol. 2020;190(2):149-172. [DOI] [PubMed] [Google Scholar]
- 19.Salinas Cisneros G, Thein SL. Recent advances in the treatment of sickle cell disease. Front Physiol. 2020;11:435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Rab MAE, Kanne CK, Bos J, et al. Oxygen gradient ektacytometry-derived biomarkers are associated with vaso-occlusive crises and correlate with treatment response in sickle cell disease. Am J Hematol. 2021;96(1):E29-E32. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.