

ABSTRACT
Palmitoylation is the most common post-translational lipid modification in the brain; however, the role of palmitoylation and palmitoylating enzymes in the nervous system remains elusive. One of these enzymes, Zdhhc5, has previously been shown to regulate synapse plasticity. Here, we report that Zdhhc5 is also essential for the formation of excitatory, but not inhibitory, synapses both in vitro and in vivo. We demonstrate in vitro that this is dependent on the enzymatic activity of Zdhhc5, its localization at the plasma membrane and its C-terminal domain, which has been shown to be truncated in a patient with schizophrenia. Loss of Zdhhc5 in mice results in a decrease in the density of excitatory hippocampal synapses accompanied by alterations in membrane capacitance and synaptic currents, consistent with an overall decrease in spine number and silent synapses. These findings reveal an important role for Zdhhc5 in the formation and/or maintenance of excitatory synapses.
KEY WORDS: Hippocampus, Lipid, Palmitate, Palmitoyl acyltransferase, Palmitoylation, Synapse
Summary: The plasma membrane-associated Zdhhc5 enzyme enhances excitatory synapse formation in vitro and in vivo through motifs at its C-terminal domain.
INTRODUCTION
Palmitoylation is a reversible post-translational lipid modification that anchors proteins to specialized membrane domains, and can critically impact protein stability, trafficking and function (Aicart-Ramos et al., 2011; Hannoush and Sun, 2010; Linder and Deschenes, 2007; Resh, 2006; Salaun et al., 2010). Palmitoylation is mediated by a family of 24 Zdhhc enzymes (Braschi et al., 2019), and growing evidence suggests that Zdhhc enzymes are important for proper brain development and function. Indeed, palmitoylation is the most common lipid modification in the brain (Fukata and Fukata, 2010) and has been shown to regulate numerous neuronal processes, including neurite outgrowth, axon pathfinding, filopodial formation, and spine development, maintenance, pruning and plasticity (Arstikaitis et al., 2008; Gauthier-Campbell et al., 2004; El-Husseini and Bredt, 2002; Kato et al., 2000; Kutzleb et al., 1998; Laux et al., 2000; Ueno, 2000; Brigidi et al., 2015; Thomas et al., 2012; Hayashi et al., 2005; Greaves and Chamberlain, 2011; Kang et al., 2004, 2008; Shah et al., 2019; Shimell et el., 2019). Of note, 9 of the 24 Zdhhc enzymes are associated with disorders of the brain (reviewed in Zaręba-Kozioł et al., 2018), and bioinformatics analysis has demonstrated that while 10% of all gene products are modified by palmitoylation, 41% of all synaptic proteins (Sanders et al., 2015) are substrates for palmitoylation, further amplifying the potential role for palmitoylation in synapse biology.
Our laboratory has previously shown an important role for one of these enzymes, Zdhhc5, in regulating the plasticity of synaptic connections in the hippocampus (Brigidi et al., 2015). In this study, the dynamic trafficking of Zdhhc5 enabled differential palmitoylation of its substrates, providing one means by which Zdhhc5 function can be regulated. Zdhhc5 can be stabilized at the synaptic membrane through its association with its accessory protein, Golga7b (Woodley and Collins, 2019) as well as Fyn kinase (Brigidi et al., 2015), by inhibiting Zdhhc5–AP2µ interactions and clathrin-mediated endocytosis. Binding to PSD-95 (also known as DLG4) via its C-terminal PDZ-binding domain further stabilizes Zdhhc5 at the membrane (Brigidi et al., 2015). Interestingly, a de novo mutation in Zdhhc5 has been reported in a patient with schizophrenia that introduces a premature stop codon at residue 648 (E648), resulting in the loss of the last 68 amino acids of Zdhhc5 (Fromer et al., 2014), including the PDZ-binding motif (Li et al., 2010; Brigidi et al., 2015). These studies reveal that Zdhhc5 is closely associated with a number of binding partners that control its subcellular localization and function.
In the present study we demonstrate, both in vitro and in vivo, that Zdhhc5 (Braschi et al., 2019) can regulate the formation and/or maintenance of excitatory, but not inhibitory, synapses. Moreover, we show that this depends on the enzymatic activity of Zdhhc5, its plasma membrane localization and its C-terminal domain. This, coupled with the findings showing that Zdhhc5 substrates are localized to different subcellular domains (Li et al., 2012; Thomas et al., 2012; Brigidi et al., 2015; Kokkola et al., 2011) and the fact that Zdhhc5 localization can be dynamically regulated (Brigidi et al., 2015), suggests that Zdhhc5 may play a key role in the regulation of excitatory synapses during development by changing its subcellular localization in response to external cues.
RESULTS AND DISCUSSION
Zdhhc5 promotes excitatory synapse formation and regulates spine stability
To determine whether Zdhhc5 is involved in dendritic arborization and/or the formation of excitatory and inhibitory synapses, hippocampal neurons were transfected with eGFP plus the indicated constructs and immunostained for PSD-95 and gephyrin [faithful markers of excitatory and inhibitory synapses, respectively (Brigidi et al., 2015; Shah et al., 2019; Shimell et al., 2019); masking shown in Fig. S1A,B]. While Zdhhc5 knockdown did not impact dendritic length (Fig. 1A), complexity (Fig. 1B) or the density of gephyrin puncta (Fig. 1C,D), it did significantly reduce the density of PSD-95 puncta (Fig. 1C,D; see Fig. S1B for the creation of masks to quantify synaptic marker density). This is consistent with our previous finding showing that Zdhhc5 is primarily localized to excitatory synapses (57% of Zdhhc5 colocalizes with PSD-95, while 31% colocalizes with gephyrin; Brigidi et al., 2015). We confirmed that the reduction in PSD-95 puncta reflected a decrease in bona fide excitatory synapses by quantifying the density of colocalized PSD-95 and VGlut1 (also known as SLC17A7) puncta (Fig. 1E). A reduction in the density of dendritic spines, and specifically a decrease in the density of stubby and mushroom spines, was also observed in Zdhhc5 knockdown neurons (Fig. 1F,G).
Fig. 1.

Zdhhc5 regulates the density of excitatory synapses and mature spines. (A,B) Transfection with Zdhhc5 shRNA with or without WT Zdhhc5R had no effect on total dendritic length (A) or complexity (B) in cultured hippocampal neurons (13 DIV). (C) Representative images of PSD-95 and gephyrin immunolabeling with GFP cell fill. Scale bar: 5 µm. (D,E) Zdhhc5 shRNA significantly reduced the density of the excitatory postsynaptic marker PSD-95 (D) as well as colocalized PSD-95 and VGlut-1 puncta (E), with no changes in the density of the inhibitory postsynaptic marker, gephyrin (D). These effects were reversed by co-transfection with Zdhhc5R. n=40 neurons per condition, >3 cultures. (F,G) Zdhhc5 shRNA significantly reduced the density of total spines, specifically resulting from a reduction in stubby and mushroom spines. n=40 neurons per condition, 3 cultures. (H) Left, pseudocolored maximum intensity projected time-lapse images of GCaMP6f fluorescence from a 2 min acquisition showing the locations of miniature spine Ca2+ events (white arrowheads) imaged in TTX (1 μM; 15 DIV). Scale bar: 5 µm. Right: GCaMP6f ΔF/F0 traces from representative single spine ROIs. (I–K) Zdhhc5 knockdown decreased the density of active spines (I), but not the amplitude (J) or number of Ca2+ events per spine (K). n=11–14 neurons per condition, 3 cultures. Results are mean±s.e.m. with individual data points shown. *P<0.05, **P<0.01, ***P<0.001 (one-way ANOVA with Tukey's post hoc test).
We next measured miniature spine Ca2+ transients with GCaMP6f to determine whether Zdhhc5 knockdown alters synaptic activity (Reese and Kavalali, 2015; Sinnen et al., 2016). The density of active spines was significantly reduced in cells expressing Zdhhc5 shRNA (Fig. 1I). However, no changes were observed in the amplitude of spine Ca2+ events (Fig. 1H,J) or the mean number of Ca2+ events at each active spine (Fig. 1H,K), indicating that Zdhhc5 may regulate excitatory synapse density without altering other properties of basal synaptic transmission.
Zdhhc5 domains important for regulating synapse density
Zdhhc5 has an extended C-terminal tail (∼70% of the total protein length) that contains a PDZ-binding motif (Kim and Sheng, 2004; Feng and Zhang, 2009; Li et al., 2010; Brigidi et al., 2015), as well as phosphorylation, GlcNAcylation and palmitoylation sites that regulate internalization and activity (Brigidi et al., 2015; Woodley and Collins, 2019; Plain et al., 2020). To investigate the mechanism by which Zdhhc5 regulates synapse formation and/or maintenance, we tested the ability of several Zdhhc5 mutants (expression levels shown in Fig. S2) to rescue the Zdhhc5 knockdown-mediated decrease in excitatory synapse density. As Zdhhc5 membrane localization is activity regulated and plays an important role in synapse plasticity (Brigidi et al., 2015), we also examined the membrane localization of each mutant.
None of the Zdhhc5 mutant constructs impacted the density of gephyrin puncta (Fig. 2A,B). While expression of shRNA-resistant Zdhhc5R rescued the Zdhhc5 knockdown-mediated decrease in PSD-95 puncta density (Fig. 1C-E; Fig. 2A,B), expression of the enzymatically-dead mutant, Zdhhs5R (in which the cysteine residue in the catalytic DHHC domain was changed to serine) did not, indicating that the enzymatic activity of Zdhhc5 is required for the formation and/or maintenance of excitatory synapses (Fig. 2A–C). Surface levels of Zdhhs5R were similar to wild-type (WT) Zdhhc5R, suggesting that the inability of Zdhhs5R to rescue the Zdhhc5 knockdown phenotype was due to its lack of enzymatic function and not changes in surface expression (Fig. 3A,B).
Fig. 2.
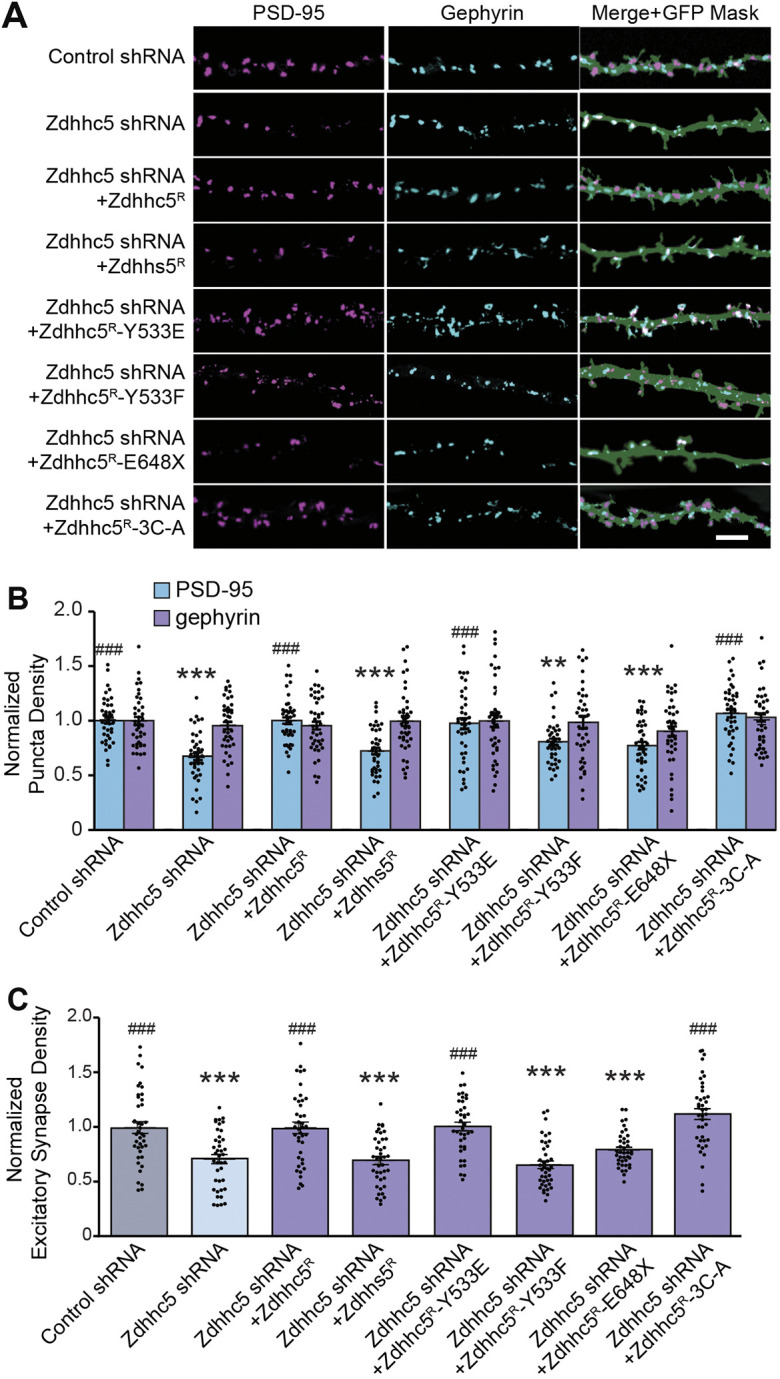
Zdhhc5 regulates excitatory synapse density via its enzymatic activity, plasma membrane localization, and C-terminal domain. (A) Representative confocal images of rat hippocampal neurons transfected at 10 DIV with eGFP plus the indicated constructs and imaged at 13 DIV. Scale bar: 5 µm. (B,C) The normalized density of PSD-95-positive puncta (B) and colocalized PSD-95 and VGlut1 puncta (C) was significantly decreased in cells expressing Zdhhc5 shRNA. Synapse density was rescued by the Zdhhc5 Y533E (phospho-mimetic) and the 3C-A (palmitoylation-dead) mutants, but not the Zdhhs5 (enzymatically dead), Zdhhc5 Y533F (phospho-dead) or E648X (C-terminal truncation) mutants. The density of gephyrin-positive puncta remained unchanged. n=40 neurons per condition, >3 cultures. Results are mean±s.e.m. with individual data points shown. **P<0.01, ***P<0.001 compared to control shRNA PSD-95; ###P<0.001 compared to Zdhhc5 shRNA PSD-95 (one-way ANOVA; Tukey's post hoc test).
Fig. 3.

Zdhhc5 turnover is mediated by the Y533 and C-terminal palmitoylation sites. (A,B) HEK293T cells transfected with sGFP-tagged Zdhhc5 constructs were biotinylated and lysates immunoprecipitated with neutravidin-coated beads to isolate all surface proteins and blots probed with anti-GFP to identify surface biotinylated Zdhhc5 mutants. The control represents a non-transfected sample and R182A (the only extracellular site of biotinylation) a biotinylation control. The Y533E and 3C-A mutants were significantly more localized to the plasma membrane, while the Y533F mutant exhibited reduced surface localization. n=3 blots, 3 separate cultures. Results are mean±s.e.m. with individual data points shown. *P<0.05, **P<0.01, ***P<0.001 (one-way ANOVA with Tukey's post hoc test). (C–E) Representative pseudocolored confocal images of neurons transfected with sGFP-tagged Zdhhc5 constructs following FRAP of a single dendritic spine. Spines were photobleached at 0 s within a 1 µm² ROI (yellow circle) (C). Y533E and 3C-A mutants exhibit significantly reduced FRAP recovery curves (D), while the Y533E, Y533F, and 3C-A mutants have significantly reduced plateau values (relative fluoresence fraction within the ROI at the 5 min time point (E). n=12–17 neurons, >3 cultures. Results are mean±s.e.m. with individual data points shown in E. **P<0.01, ***P<0.001 [two-way ANOVA (D) or one-way ANOVA (E) with Tukey's post hoc test]. Two-way ANOVA revealed a significant interaction for all parameters [interaction: F(290)=1.268, P=0.0018; construct: F(5)=427.1, P<0.0001; time: F(58)=19.96), P<0.001].
We next expressed a C-terminal truncated Zdhhc5 identified in a patient with schizophrenia (Fromer et al., 2014), which results in a premature stop codon that deletes the final 68 amino acids of Zdhhc5 (Zdhhc5R E648X), including the PDZ-binding domain. Although surface expression of Zdhhc5R E648X was similar to WT Zdhhc5 (Fig. 3A,B), Zdhhc5R E648X did not rescue the synaptic phenotype observed upon Zdhhc5 knockdown (Fig. 2A–C). This deficit is likely due to reduced Zdhhc5 association with PDZ domain-containing proteins, such as PSD-95, that are known to be critical for activity induced spine plasticity (Brigidi et al., 2015).
Our laboratory has previously demonstrated that phosphorylation of tyrosine 533 (Y533) immobilizes Zdhhc5 at the synaptic membrane by occluding Zdhhc5 binding to the endocytic protein AP2µ (encoded by Ap2m1) (Brigidi et al., 2015). We observed a significant reduction in the surface localization of the Zdhhc5 phospho-dead mutant (Zdhhc5R Y533F) (Fig. 3A,B), which also failed to rescue Zdhhc5 knockdown-mediated decrease in excitatory synapse density (Fig. 2A–C). In contrast, while the phospho-mimetic Zdhhc5R Y533E mutant was significantly more localized to the plasma membrane (Fig. 3A,B), the density of excitatory synapses in cells expressing Zdhhc5R Y533E was indistinguishable from cells expressing WT Zdhhc5, suggesting that Zdhhc5 surface expression is necessary but not sufficient to promote synapse formation and/or maintenance. Recent work by Hao and colleagues have also shown that phosphorylation of tyrosine residue Y91 near the DHHC domain can decrease Zdhhc5 activity, further underscoring the role of tyrosine phosphorylation in Zdhhc5 function (Hao et al., 2020).
Previous work has shown that disrupting the palmitoylation of Zdhhc5 on cysteine residues 236, 237 and 245, can impede endocytosis and increase Zdhhc5 surface localization (Woodley and Collins, 2019; but see Ko et al., 2019). We therefore used a palmitoylation-defective Zdhhc5R 3C-A mutant to further explore the role of Zdhhc5 membrane localization in regulating synapse formation. As predicted, surface localization of Zdhhc5R 3C-A was significantly increased when compared with WT Zdhhc5 (Fig. 3A,B), yet no changes were observed in the density of excitatory synapses in cells expressing Zdhhc5R 3C-A (Fig. 2A,B).
Together, this demonstrates that the palmitoylation of protein(s) by Zdhhc5 at nascent synaptic membranes is required for the maturation or maintenance of synaptic connections. Decreased Zdhhc5 surface expression reduces excitatory synapse density; however, increased Zdhhc5 localization to the plasma membrane does not appear to be sufficient to promote synapse formation or maintenance. This may be due to alterations in substrate interaction and/or recruitment, as the mutants that display the greatest surface localization have alterations in amino acid residues known to be important in these processes (Brigidi et al., 2015; Woodley and Collins, 2019).
We next investigated the mobility of Zdhhc5 mutants within postsynaptic spines using fluorescence recovery after photobleaching (FRAP). Analysis of FRAP curves demonstrated a significant decrease in the mobility of Zdhhc5 Y533E and 3C-A mutants (Fig. 3C,D), whereas analysis of the mobile faction at just the 5 min time point demonstrated a significant decrease in mobility of Zdhhc5 Y533E, 3C-A and Y533F mutants (Fig. 3E). While the decrease in Zdhhc5 Y533F surface expression suggests more Y533F in the recycling endosome pool, the concomitant decrease in mobility in spines may be accounted for by a decrease in lateral mobility along the membrane.
Zdhhc5 regulates excitatory synapse density in vivo
To determine whether Zdhhc5 is similarly important for the formation and/or maintenance of excitatory synapses in vivo, we analyzed Zdhhc5 gene-trap (Zdhhc5-GT) mice (Li et al., 2010), in which Zdhhc5 protein is not detected (Fig. 4A). Using electron microscopy, we compared the density of synapses in the stratum radiatum of the hippocampus. Excitatory (asymmetric) and inhibitory (symmetric) synapses were identified based on their morphological characteristics (Gray, 1959; Fig. 4B). To ensure accuracy, samples were also immunolabeled with PSD-95 to identify excitatory synapses (validated in Liu and Cheng, 2016; Mills et al., 2017; Fig. 4B). We observed a significant decrease in the density of PSD-95-positive asymmetric excitatory synapses, but no change in the density of PSD-95-negative symmetric inhibitory synapses in Zdhhc5-GT samples compared to controls (Fig. 4C). No differences in active zone length were observed (Fig. 4D).
Fig. 4.

Zdhhc5 regulates excitatory synapse density in vivo. (A) Western blots of cortical (Ctx) and hippocampal (Hpc) lysates from Zdhhc5-GT (gene trapped) mice and age-matched littermate controls showing the absence of Zdhhc5 expression in Zdhhc5-GT mice. (B) Representative immunogold-electron microscopy images showing excitatory symmetric synapses (black arrows) and inhibitory asymmetric synapses (white arrows). Scale bar: 500 nm. (C) Zdhhc5-GT mice exhibit a significant reduction in the density of excitatory synapses, with no change in inhibitory synapses. n=3 mice per genotype. **P<0.01 [two-way ANOVA, significant interaction between genotype and synapse type, F(1,8)=8.753, P=0.0182, Bonferonni's test post hoc, n=3 mice per genotype]. (D) There is no significant difference in the active zone between Zdhhc5-GT and WT littermates. (E) CA1 pyramidal neurons from acute Zdhhc5-GT brain sections exhibit lower capacitance than pyramidal neurons from WT mice (n=24 WT neurons; 4 mice and 31 Zdhhc5-GT neurons; 5 mice) *P=0.0127 [unpaired two-tailed t-test, t(53)=2.579]. (F,G) There is no significant difference between WT and Zdhhc5-GT mice in either resistance (F) or holding current (G). (H–J) There is no significant difference of sEPSC frequency (H,I), sEPSC amplitude (J,K), or paired-pulse ratio (L). (M) CA1 pyramidal neurons from Zdhhc5-GT brain slices exhibit a significantly higher AMPAR to NMDAR current ratio than WT slices. n=12 WT neurons; 3 mice and 13 Zdhhc5-GT neurons; 3 mice. *P=0.0363 [unpaired two-tailed t-test, t(23)=2.224]. Insets in L, J and M show representative current traces from each experiment. All data is from comparisons of Zdhhc5-GT with age-matched littermate controls and is shown as mean±s.e.m.
To determine whether the observed decrease in excitatory synapse density results in functional deficits, whole-cell voltage-clamp recordings were made in CA1 pyramidal neurons. The membrane properties revealed a lower capacitance in Zdhhc5-GT neurons compared to WT neurons (Fig. 4E), with no change in membrane resistance or holding current (Fig. 4F,G). As Zdhhc5 did not impact dendritic length or complexity in vitro (Fig. 1A,B), and Zdhhc5-GT mice have a lower density of excitatory synapses (Fig. 4C), this reduced capacitance may be due to fewer spines resulting in less total membrane. The lack of change in the resistance (4F) and holding current (4G) when voltage-clamped at −70 mV suggests that the composition of the channels open in the cells at this potential is unchanged and the cells do not have altered excitability.
Zdhhc5-GT and WT littermates revealed no difference in the frequency or the amplitude of spontaneous excitatory postsynaptic currents (sEPSCs) (Fig. 4H–K), despite our finding that Zdhhc5-GT mice have a decreased number of excitatory synapses. These observations could not be attributed to compensatory increases in pre-synaptic glutamate release in Zdhhc5-GT mice, as we found no significant difference in the paired pulse ratio (PPR) compared to WT littermates (Fig. 4L). However, we observed a significantly higher α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor (AMPAR) to N-methyl-D-aspartate receptor (NMDAR) ratio (denoted A/N ratio) in Zdhhc5-GT mice (Fig. 4M), without any changes in the amplitude of AMPAR sEPSCs (Fig. 4J,K). These data may therefore reveal a selective decrease in the number of silent synapses (which contain NMDARs, but lack AMPARs; Isaac et al., 1995; Liao et al., 1995; Kerchner and Nicoll, 2008) in Zdhhc5-GT WT neurons. This is consistent with our observations that Zdhhc5 is required for spine maturation and stabilization, and would indicate that immature (silent) synapses might be eliminated before reaching maturity.
The findings of this study demonstrate that Zdhhc5, previously linked to learning and memory (Li et al., 2010) and schizophrenia (Schizophrenia Working Group of the Psychiatric Genomics, 2014), regulates excitatory synapse formation via the interplay of its palmitoylation activity, its C-terminal domain and its surface localization. Two of the key neuropathological findings of schizophrenia are gray matter loss (reviewed in Vita et al., 2012), and changes in hippocampal dendritic spines (Steen et al., 2006; Kolomeets et al., 2005), while disruptions to excitatory/inhibitory balance underlie a number of other neuropathologies. Our study contributes to the understanding of the role of Zdhhc5 in the hippocampus as a key component for learning and memory.
MATERIALS AND METHODS
Animals
All experimental procedures and housing conditions were approved by the UBC Animal Care Committee and were in accordance with the Canadian Council on Animal Care (CCAC) guidelines. Zdhhc5-GT mice were obtained from Don Hilgemann (UT Southwestern, TX, USA). The mice were originally generated by Li et al., (2010) from an embryonic stem cell line (RRD533, strain 129/Ola) with an insertional mutation in Zdhhc5 from the International Gene Trap Consortium (Skarnes et al., 2004; Nord et al., 2006). A gene-trapping vector, pGT11xf, introduced an in-frame fusion between the 5′ exons of the trapped gene and a reporter.
Cell cultures
Hippocampi were isolated from embryonic day (E)18 Sprague-Dawley rats as previously described (Xie et al., 2000) and plated at a density of 130 cells per mm2. Neurons were transfected with Lipofectamine 2000 (Thermo Fisher Scientific, 11668019) at 10 days in vitro (DIV) following the manufacturer's recommendations and used for experiments at 13 DIV. HEK293T cells [ATCC (ACS-4500)] were transfected using Lipofectamine 2000 according to the manufacturer's recommendations. HEK293T cells were transfected at ∼70–80% confluency and incubated for 48 h before harvesting for biochemistry.
cDNA constructs
Control shRNA, Zdhhc5 shRNA and HA–Zdhhc5 were kind gifts from Richard Huganir (Johns Hopkins U., Baltimore, USA). shRNA resistant HA–Zdhhc5 (Zdhhc5R) was generated as previously described (Brigidi et al., 2014). To make sGFP (Kremers et al., 2007) Zdhhc5 constructs (sGFP-Zdhhc5-WT; -R182A; -E648X; -Y533E; -Y533F and Zdhhs5), the coding sequences of mouse Zdhhc5 WT, R182A, E648X, Y533E, Y533F and Zddhs5 were amplified from HA–Zdhhc5 constructs used previously (Brigidi et al., 2015) using the following primers: Fwd, 5′-CCGGCGAATTCTATGCCCGCAGAGTCTG-3′; Rev, 5′-GCCGGGGATCCTCACACAGAAATCTC-3′ for -WT, -R182A, -Y553E, -Y533F and Zdhhs5, or 5′-GCCGGGGATCCTCACTCTGAGACACCAGA-3′ for -E648X. Fragments were then cut with EcoRI and BamHI and ligated into the multiple cloning site of the sGFP-C1 vector (Addgene #22881). To make sGFP–Zdhhc5-3A, the coding sequence of mouse Zdhhc5-WT was amplified with Gibson mutagenic primers to create two fragments incorporating the 3A mutations that were then joined using the NEB Gibson reaction mix (Fragment 1 primers: Fwd, 5′-GATCTCGAGCTCAAGCTTCGAA-3′; Rev, 5′-ACTGGCGAGGACACGGCTAACGTTGTTAGCGGCGCCATTGGTGAA-3′; Fragment 2 primers, Fwd, 5′-ATGGCGCCGCTAACAACGTTAGCCGTGTCCTCGCCAGTTCTCCAGCA-3′; Rev, 5′-TGATCAGTTATCTAGATCCGGTGG-3′). The joined Zdhhc5 3A fragment was then digested with EcoRI and BamHI and ligated into the sGFP-C1 vector.
Immunocytochemistry
Immunocytochemistry experiments were performed as previously reported (Sun and Bamji, 2011). Briefly, cultured neurons were fixed in 4% paraformaldehyde and 4% sucrose, permeabilized with 0.1% Triton-X, and blocked in 10% goat serum for 1 h at room temperature. Primary antibodies were diluted in 1% goat serum and applied to neurons overnight at 4°C. Secondary antibodies were also diluted in 1% goat serum, and applied to neurons for 1 h at room temperature. Coverslips were mounted with Prolong Gold (Thermo Fisher Scientific, P36930). Primary antibodies were as follows: PSD-95 (mouse monoclonal, IgG2a isotype, 1:500, Abcam, ab2723), gephyrin (mouse monoclonal, IgG1 isotype, 1:500, Synaptic Systems, 147 011), VGlut1 (guinea pig, 1:500, EMD Millipore, AB5905). Secondary antibodies were as follows: goat-anti-mouse IgG2a Alexa Fluor 568 (Life Technologies, A21134), goat-anti-mouse IgG1 Alexa Fluor 647 (Life Technologies, A21240), goat-anti-guinea pig Alexa Fluor 633 (Life Technologies, A21105), goat anti-mouse IgG Alexa Fluor 568 (Life Technologies, A11019), goat anti-mouse IgG Alexa Fluor 633 (Life Technologies, A21050).
Immunogold electron microscopy
Samples from mouse CA1 hippocampus were processed as described previously (Mills et al., 2017). Brains were sliced into 250 µm-thick sections with a vibratome, and pieces of CA1 hippocampus (<1 mm in all dimensions) were dissected from slices and cryoprotected in 30% glycerol overnight at 4°C. Samples were plunge-frozen in liquid ethane at −170°C in an EM cryopreparation chamber (Leica) and transferred to a 1.5% uranyl acetate solution in 100% methanol, kept at −90°C in a Leica EM AFS for 30 h. The temperature was gradually increased and samples infiltrated with HM-20 acrylic resin (Electron Microscopy Sciences, Hatfield, PA, USA). Samples were set up in capsules containing pure resin and polymerized under UV light for 24 h. Tissue sections were cut at 85 nm using a Diatome diamond knife and a Leica ultramicrotome. Sections were collected on 300-mesh, formvar coated nickel grids (Electron Microscopy Sciences).
Post-embedding immunostaining was performed on the EM grids as described previously (Mills et al., 2017). Grids were rinsed with distilled water and immersed in a bead of Tris-buffered saline with 0.1% Tween 20 (TTBS) with 0.1% Triton-X, 0.1% sodium borohydride and 50 mM glycine. Nonspecific binding was blocked with 2% BSA in TTBS with 0.1% Triton-X. Primary antibody against PSD-95 (rabbit, Frontier Institute, Af628; RRID: AB_2571611, 1:100) were diluted in 2% BSA in TTBS with 0.1% Triton-X. Grids were immersed in 15 µl beads of diluted primary antibody overnight at room temperature in a humidified chamber. The next day, grids were rinsed in TTBS with 0.1% Triton-X. Secondary antibodies were diluted in 2% BSA in TTBS with 0.1% Triton-X and 0.05% polyethylene glycol (PEG). Girds were immersed in 15 µl beads of secondary antibody (Electron Microscopy Sciences, goat-anti-rabbit-IgG 15 nm, cat. no. 25112) for 1.5 h. Grids were rinsed in TTBS with 0.1% Triton-X, in Milli-Q H2O and dried. Grids were then lightly counterstained with 2% uranyl acetate and Reynold's lead citrate. Images were collected at 23,000× magnification on a Tecnai G2 Spirit transmission electron microscope (FEI Company, Eindhoven, the Netherlands). To quantify excitatory and inhibitory synapse density in the CA1 hippocampus, the number of PSD-95-positive and -negative synapses was quantified within a 2500 µm2 region of the hippocampus. All images were acquired and analyzed by a researcher who was blind to the genotype of each mouse.
Fluorescence recovery after photobleaching
sGFP–Zdhhc5 and sGFP–Zdhhc5 mutant construct puncta localized to dendritic spines (identified using mCherry cell fill) within 100 µm of the cell body were imaged every second for 5 min after photobleaching using a Zeiss LSM780 confocal microscope. An ∼1 µm diameter circular region of interest was used to photobleach the Zdhhc5, and a 1 µm diameter circular region of interest was placed both on the cell body for a reference and a region of background for background subtraction. Analysis was carried out using easyFRAP (Rapsomaniki et al., 2012) and data exported to Prism Software (GraphPad) for data visualization, analysis and plateau value generation.
Immunoblotting
Western blotting was performed as previously described (Brigidi et al., 2014; Sun and Bamji, 2011). HEK293T cells were homogenized in an ice-cold lysis buffer containing 1% IGEPAL CA-630 (Sigma), 50 mM Tris-HCl, pH7.5, 150 mM NaCl and 10% glycerol, supplemented with PMSF solution and a protease inhibitor cocktail with ethylenediaminetetraacetic acid (Roche). Brain tissue was homogenized in ice-cold RIPA buffer (Thermo Fisher Scientific, CAT# 89900). Proteins were cleared by centrifugation at 14,000 g for 30 min at 4°C. Proteins were separated by SDS-PAGE and probed with antibodies against Zdhhc5 (1:1000; Sigma Prestige, HPA014670), HA (1:1000; Sigma, H9658), and GAPDH (1:1000; Abcam, ab9484). Bands were visualized using enhanced chemiluminescence (Pierce Biotechnology) on a C-DiGit Chemiluminescence Western Blot Scanner (LI-COR).
Biotinylation assay
Biotinylation experiments were performed as previously described (Shimell et al., 2019). Briefly, neurons in 10 cm dishes were nucleofected with indicated constructs and experiments were carried out at 13 DIV. Neurons were washed with ice cold PBS-CM (0.1 mM CaCl2 and 1 mM MgCl2 in 1× PBS, pH 8) and incubated for 30 min with 0.5 mg/ml NHS-SS-Biotin in ice cod PBS-CM at 4°C with gentle rocking. After incubation, cells were washed once with PBS-CM and the unbound biotin quenched via two 8 min incubations with quenching buffer (20 mM glycine in PBS-CM). Lysis was performed using mechanical scraping in lysis buffer (1% IGEPAL-CA630 and 1mM PMSF with Roche Complete protease inhibitor tablet) and subsequently spun down at 500 g for 5 min at 4°C. Samples were vortexed, run through a 26 1/2-gauge syringe three times, and nutated at 4°C for 30 min. After nutation, samples were spun down at 16,100 g for 30 min at 4°C to clear the lysate. The cell lysate was then quantified for protein using a BCA assay kit (Thermo Fisher Scientific) as per the manufacturer's instructions. 10 mg of each whole-cell lysate was then combined with SDS-sample buffer (50 mM Tris-HCl, 2% SDS, 10% glycerol, 14.5 mM EDTA and 0.02% bromophenol blue with 1% β-mercaptoethanol), boiled for 5 min at 95°C and stored at −20°C as the input sample. 100–200 mg of the remaining protein sample was added to a 50 ml 50% slurry of Neutravidin-conjugated agarose beads (Thermo Fisher Scientific) that was pre-washed three times in lysis buffer. Each sample was then brought to a total volume of 500 ml with lysis buffer and nutated at 4°C overnight. The following day beads were pelleted and washed seven times using centrifugation (500 g for 3 min). Elution of the beads was performed using 40 ml of SDS-sample buffer with 100mM DTT. Samples were boiled at 90°C for 5 min and then run on a western blot with the whole-cell lysates.
Ca2+ imaging with GCaMP6f
Cultured hippocampal neurons were transfected on 12 DIV with pCAG-GCaMP6f (kind gift from Mark Dell'Acqua, University of Coloroado Denver, Denver, USA; 0.2 μg), and either control shRNA (1 μg), Zdhhc5 shRNA (1 μg), or Zdhhc5 shRNA (1 μg) and WT HA–Zdhhc5R (1 μg). At 15 DIV, neurons were incubated at 20°C in an artificial CSF (aCSF) imaging medium containing the following (in mM): 135 NaCl, 5 KCl, 25 HEPES pH 7.4, 10 glucose, 3 CaCl2, and 0.001 tetrodotoxin (TTX). Images were acquired using a Zeiss LSM 880 AxioObserver Airyscan microscope with a Plan-Apochromat 63×/1.4 Oil DIC M27 objective and a 0.71 μs dwell time (488 nm laser) using AiryScan Fast mode. Single z-plane images of the proximal dendritic arbor covering a 133.5×133.5 μm field of view were acquired at 3.8 Hz for 2 min.
To measure the number and amplitude of Ca2+ events, the time-lapse image was processed using the ‘Delta F up’ plugin from the ImageJ Cookbook T-functions application and then maximum intensity projected to create a binary mask of Ca2+ event locations, which were then converted into regions of interest (ROIs). Any events that occurred in presumed axons or the soma were removed. The mean GCaMP6f fluorescence within each ROI was measured. A baseline of 10 frames was established for each ROI and used to calculate the ΔF/F0. Ca2+ events were counted when they surpassed a threshold of a 200% increase in fluorescence over baseline. The mean number of events per spine was calculated as the total number of active spines divided by the total number of Ca2+ events. Ca2+ events per μm were calculated as the total number of active spines (classified as spines with one or more Ca2+ event) divided by total dendrite length, which was measured using the ‘NeuronJ’ plugin in ImageJ (Meijering et al., 2004).
Slice preparation and electrophysiology
Male 2-month-old Zdhhc5-GT (Li et al., 2010) and WT control mice were deeply anesthetized with isofluorane vapor, decapitated and the brain rapidly removed. Acute sagittal brain slices (300 µm) containing medial dorsal hippocampus were cut on a vibratome (Leica VT1000) in ice-cold artificial cerebrospinal fluid (aCSF; with 0.5 mM CaCl2 and 2.5 mM MgCl2) equilibrated with 95% O2 and 5% CO2. Slices were transferred to a holding chamber with aCSF at 35°C containing (in mM): 125 NaCl, 2.5 KCl, 25 NaHCO3, 1.25 NaH2PO4, 1 MgCl2, 2 CaCl2, 10 glucose, pH 7.3–7.4, 305–310 mosmol l−1 for 45 min then maintained at room temperature. In the recording chamber, slices were equilibrated for 10 min while being continuously superfused at room temperature with oxygenated aCSF at 1–2 ml/min containing picrotoxin (50 μM, Tocris Bioscience, MO, USA) to block GABAA receptor-mediated inhibitory responses.
Pipettes (3–5 MΩ) were made from borosilicate glass capillaries on a Narishigi micropipette puller (Narishige International, East Meadow, NY, USA). Whole cell patch-clamp recording was performed with a multiclamp-900 amplifier and pClamp 10 software (Axon Instruments, CA, USA) digitized at 20 kHz and filtered at 10 kHz (sEPSCs were filtered at 1 kHz with a detection threshold of −8 pA). CA1 pyramidal neurons were maintained voltage-clamped at −70 mV for sEPSC, PPR and the AMPAR:NMDAR ratio (A/N ratio; except that the voltage was switched to +40 mV to measure NMDAR current) experiments with internal solution containing in mM: 120 cesium methane-sulfonate (CH3O3SCs), 5 NaCl, 1.1 EGTA, 4 MgATP, 0.3 NaGTP, 5 QX-314 Cl, 10 tetraethylammonium chloride (TEA), 10 HEPES pH 7.25, osmolarity 290 mOsm. Series resistance was <20 mΩ and uncompensated. To evoke synaptic currents, stimuli (100 μs duration) were delivered with a glass electrode (2–3 MΩ) filled with aCSF placed <300 µm from the recorded cell to stimulate the Schaffer collateral pathway. PPRs were calculated by dividing the amplitude of the second EPSC by the amplitude of the first, and increasing the interpulse interval by 50 ms to a maximum of 500 ms and repeated twice for each cell. For A/N ratio, the AMPAR component was the average peak amplitude of four evoked EPSCs at −70 mV. The NMDAR component was the average of four EPSC evoked at the same stimulation intensity and clamped at +40 mV measured 75 ms after stimulation to eliminate the possibility of contamination by AMPAR.
Image acquisition, analysis and quantification
Confocal images were obtained on either an Olympus Fluoview FV1000 inverted laser scanning confocal microscope or a Zeiss 880 laser scanning confocal microscope using a 60× or 63× imaging objective, respectively. For synapse density analyses, confocal images were subjectively thresholded using ImageJ software (by an observer who was blind to the condition). Puncta were identified as a fluorescence cluster with an area between 0.05 and 3 μm2. Puncta area and the integrated density (the product of the area and the mean gray value) were then determined using ImageJ. An ImageJ colocalization plugin was used to assess the colocalization between VGlut1 and PSD-95 puncta (http://rsb.info.nih.gov/ij/plugins/colocalization.html). Points of colocalization were defined as regions of >4 pixels in size with a >50 intensity ratio between the two channels. The length of each dendrite was measured using the ‘NeuronJ’ ImageJ plugin (Meijering et al., 2004), and puncta density was calculated by computing the number of puncta in each image divided by the length of dendrite in each image. Sholl analysis was performed using an ImageJ plugin (Ferreira et al., 2014). Reconstruction and analysis of dendritic spine type was measured using NeuronStudio computational software (Rodriguez et al., 2008).
Statistical analysis
All data values are expressed as the mean±s.e.m.. For imaging experiments, the n numbers shown refer to the number of cells used per condition over three separate cultures, with the exception of Fig. 3A,B (FRAP), where N refers to the number of spines and is specified within figure legends. All electrophysiological data were analyzed using Clampfit 10.4 and Graphpad Prism 5 and are presented as the mean±s.e.m. of n=number of neurons from a minimum of three animals per genotype. Statistical significance was determined by unpaired, two-tailed Student's t-test, one-way or two-way ANOVA with Bonferroni's post hoc test using Prism where indicated. Statistical significance was assumed when P<0.05. All figures were generated using Photoshop CS6 and/or Illustrator CS6 software (Adobe Systems, Inc.).
Supplementary Material
Acknowledgements
Some of the text and figures in this paper were included in Andrea Kathleen Globa's PhD thesis in for the Graduate Program in Neuroscience at the University of British Columbia (2017).
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: S.X.B.; Methodology: J.J.S., A.G., M.D.S., A.R.W.; Validation: J.J.S., A.G., M.D.S., A.R.W., N.M.; Formal analysis: J.J.S., A.G., M.D.S., A.R.W., N.M.; Investigation: J.J.S., A.G., M.D.S., A.R.W., N.M.; Resources: L.A.R., S.X.B.; Data curation: J.J.S., A.G., N.M.; Writing - original draft: J.J.S., A.G., S.X.B.; Writing - review & editing: J.J.S., L.A.R., S.X.B.; Visualization: J.J.S., A.R.W., S.X.B.; Supervision: L.A.R., S.X.B.; Project administration: L.A.R., S.X.B.; Funding acquisition: L.A.R., S.X.B.
Funding
This work was supported by grants from Canadian Institutes of Health Research (CIHR; FDN-159907 to S.X.B; FDN-143210 to L.A.R.). Deposited in PMC for immediate release.
Peer review history
The peer review history is available online at https://journals.biologists.com/jcs/article-lookup/DOI/10.1242/jcs.254276
References
- Aicart-Ramos, C., Valero, R. A. and Rodriguez-Crespo, I. (2011). Protein palmitoylation and subcellular trafficking. Biochim. Biophys. Acta (BBA) Biomembranes 1808, 2981-2994. 10.1016/j.bbamem.2011.07.009 [DOI] [PubMed] [Google Scholar]
- Arstikaitis, P., Gauthier-Campbell, C., Carolina Gutierrez Herrera, R., Huang, K., Levinson, J. N., Murphy, T. H., Kilimann, M. W., Sala, C., Colicos, M. A., El-Husseini, A.et al. (2008). Paralemmin-1, a modulator of filopodia induction is required for spine maturation. Mol. Biol. Cell 19, 2026-2038. 10.1091/mbc.e07-08-0802 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braschi, B., Denny, P., Gray, K., Jones, T., Seal, R., Tweedie, S., Yates, B. and Bruford, E. (2019). Genenames.org: the HGNC and VGNC resources in 2019. Nucleic Acids Res. 47, D786-D792. 10.1093/nar/gky930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brigidi, G. S., Sun, Y., Beccano-Kelly, D., Pitman, K., Mobasser, M., Borgland, S. L., Milnerwood, A. J. and Bamji, S. X. (2014). Palmitoylation of δ-catenin by DHHC5 mediates activity-induced synapse plasticity. Nat. Neurosci. 17, 522-532. 10.1038/nn.3657 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brigidi, G. S., Santyr, B., Shimell, J., Jovellar, B. and Bamji, S. X. (2015). Activity-regulated trafficking of the palmitoyl-acyl transferase DHHC5. Nat. Commun. 6, 8200. 10.1038/ncomms9200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Husseini, A. E.-D. and Bredt, D. S. (2002). Protein palmitoylation: a regulator of neuronal development and function. Nat. Rev. Neurosci. 3, 791-802. 10.1038/nrn940 [DOI] [PubMed] [Google Scholar]
- Feng, W. and Zhang, M. (2009). Organization and dynamics of PDZ-domain-related supramodules in the postsynaptic density. Nat. Rev. Neurosci. 10, 87-99. 10.1038/nrn2540 [DOI] [PubMed] [Google Scholar]
- Ferreira, T. A., Blackman, A. V., Oyrer, J., Jayabal, S., Chung, A. J., Watt, A. J., Sjöström, P. J. and van Meyel, D. J. (2014). Neuronal morphometry directly from bitmap images. Nat. Methods 11, 982-984. 10.1038/nmeth.3125 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fromer, M., Pocklington, A. J., Kavanagh, D. H., Williams, H. J., Dwyer, S., Gormley, P., Georgieva, L., Rees, E., Palta, P., Ruderfer, D. M.et al. (2014). De novo mutations in schizophrenia implicate synaptic networks. Nature 506, 179-184. 10.1038/nature12929 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukata, Y. and Fukata, M. (2010). Protein palmitoylation in neuronal development and synaptic plasticity. Nat. Rev. Neurosci. 11, 161-175. 10.1038/nrn2788 [DOI] [PubMed] [Google Scholar]
- Gauthier-Campbell, C., Bredt, D. S., Murphy, T. H. and El-Husseini, A. E.-D. (2004). Regulation of dendritic branching and filopodia formation in hippocampal neurons by specific acylated protein motifs. Mol. Biol. Cell 15, 2205-2217. 10.1091/mbc.e03-07-0493 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray, E. G. (1959). Axo-somatic and axo-dendritic synapses of the cerebral cortex: an electron microscope study. J. Anat. 93, 420-433. [PMC free article] [PubMed] [Google Scholar]
- Greaves, J. and Chamberlain, L. H. (2011). Differential palmitoylation regulates intracellular patterning of SNAP25. J. Cell Sci. 124, 1351-1360. 10.1242/jcs.079095 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hannoush, R. N. and Sun, J. (2010). The chemical toolbox for monitoring protein fatty acylation and prenylation. Nat. Chem. Biol. 6, 498-506. 10.1038/nchembio.388 [DOI] [PubMed] [Google Scholar]
- Hao, J.-W., Wang, J., Guo, H., Zhao, Y.-Y., Sun, H.-H., Li, Y.-F., Lai, X.-Y., Zhao, N., Wang, X., Xie, C.et al. (2020). CD36 facilitates fatty acid uptake by dynamic palmitoylation-regulated endocytosis. Nat. Commun. 11, 4765. 10.1038/s41467-020-18565-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi, T., Rumbaugh, G. and Huganir, R. L. (2005). Differential regulation of AMPA receptor subunit trafficking by palmitoylation of two distinct sites. Neuron 47, 709-723. 10.1016/j.neuron.2005.06.035 [DOI] [PubMed] [Google Scholar]
- Isaac, J. T. R., Nicoll, R. A. and Malenka, R. C. (1995). Evidence for silent synapses: implications for the expression of LTP. Neuron 15, 427-434. 10.1016/0896-6273(95)90046-2 [DOI] [PubMed] [Google Scholar]
- Kang, R., Swayze, R., Lise, M. F., Gerrow, K., Mullard, A., Honer, W. G. and El-Husseini, A. (2004). Presynaptic trafficking of synaptotagmin I is regulated by protein palmitoylation. J. Biol. Chem. 279, 50524-50536. 10.1074/jbc.M404981200 [DOI] [PubMed] [Google Scholar]
- Kang, R., Wan, J., Arstikaitis, P., Takahashi, H., Huang, K., Bailey, A. O., Thompson, J. X., Roth, A. F., Drisdel, R. C., Mastro, R.et al. (2008). Neural palmitoyl-proteomics reveals dynamic synaptic palmitoylation. Nature 456, 904-909. 10.1038/nature07605 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kato, H., Allen, N. D., Emson, P. C. and Kiyama, H. (2000). GAP-43 N-terminal translocation signal targets β-galactosidase to developing axons in a pan-neuronal transgenic mouse line. Dev. Brain Res. 121, 109-112. 10.1016/S0165-3806(00)00019-5 [DOI] [PubMed] [Google Scholar]
- Kerchner, G. A. and Nicoll, R. A. (2008). Silent synapses and the emergence of a postsynaptic mechanism for LTP. Nat. Rev. Neurosci. 9, 813-825. 10.1038/nrn2501 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, E. and Sheng, M. (2004). PDZ domain proteins of synapses. Nat. Rev. Neurosci. 5, 771-781. 10.1038/nrn1517 [DOI] [PubMed] [Google Scholar]
- Kremers, G.-J., Goedhart, J., van den Heuvel, D. J., Gerritsen, H. C. and Gadella, T. W. J. (2007). Improved green and blue fluorescent proteins for expression in bacteria and mammalian cells. Biochemistry 46, 3775-3783. 10.1021/bi0622874 [DOI] [PubMed] [Google Scholar]
- Ko, P.-J., Woodrow, C., Dubreuil, M. M., Martin, B. R., Skouta, R., Bassik, M. C. and Dixon, S. J. (2019). A Zdhhc5-GOLGA7 protein Acyltransferase complex promotes nonapoptotic cell death. Cell Chem. Biol. 26, 1716-1724.e9. 10.1016/j.chembiol.2019.09.014 [DOI] [PubMed] [Google Scholar]
- Kokkola, T., Kruse, C., Roy-Pogodzik, E.-M., Pekkinen, J., Bauch, C., Hönck, H.-H., Hennemann, H. and Kreienkamp, H.-J. (2011). Somatostatin receptor 5 is palmitoylated by the interacting Zdhhc5 palmitoyltransferase. FEBS Lett. 585, 2665-2670. 10.1016/j.febslet.2011.07.028 [DOI] [PubMed] [Google Scholar]
- Kolomeets, N. S., Orlovskaya, D. D., Rachmanova, V. I. and Uranova, N. A. (2005). Ultrastructural alterations in hippocampal mossy fiber synapses in schizophrenia: a postmortem morphometric study. Synapse 57, 47-55. 10.1002/syn.20153 [DOI] [PubMed] [Google Scholar]
- Kutzleb, C., Sanders, G., Yamamoto, R., Wang, X., Lichte, B., Petrasch-Parwez, E. and Kilimann, M. W. (1998). Paralemmin, a prenyl-palmitoyl-anchored phosphoprotein abundant in neurons and implicated in plasma membrane dynamics and cell process formation. J. Cell Biol. 143, 795-813. 10.1083/jcb.143.3.795 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laux, T., Fukami, K., Thelen, M., Golub, T., Frey, D. and Caroni, P. (2000). GAP43, MARCKS, and CAP23 modulate PI(4,5)P2 at plasmalemmal rafts, and regulate cell cortex actin dynamics through a common mechanism. J. Cell Biol. 149, 1455-1472. 10.1083/jcb.149.7.1455 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Y., Hu, J., Höfer, K., Wong, A. M. S., Cooper, J. D., Birnbaum, S. G., Hammer, R. E. and Hofmann, S. L. (2010). DHHC5 interacts with PDZ domain 3 of post-synaptic density-95 (PSD-95) protein and plays a role in learning and memory. J. Biol. Chem. 285, 13022-13031. 10.1074/jbc.M109.079426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Y., Martin, B. R., Cravatt, B. F. and Hofmann, S. L. (2012). DHHC5 protein palmitoylates flotillin-2 and is rapidly degraded on induction of neuronal differentiation in cultured cells. J. Biol. Chem. 287, 523-530. 10.1074/jbc.M111.306183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao, D., Hessler, N. A. and Malinow, R. (1995). Activation of postsynaptically silent synapses during pairing-induced LTP in CA1 region of hippocampal slice. Nature 375, 400-404. 10.1038/375400a0 [DOI] [PubMed] [Google Scholar]
- Linder, M. E. and Deschenes, R. J. (2007). Palmitoylation: policing protein stability and traffic. Nat. Rev. Mol. Cell Biol. 8, 74-84. 10.1038/nrm2084 [DOI] [PubMed] [Google Scholar]
- Liu, X. B. and Cheng, H. J. (2016). Dual-labeling immuno-electron microscopic method for identifying pre- and postsynaptic profiles in Mammalian brain. In Neuromethods, Vol. 115, pp. 21-33. Humana Press Inc.. 10.1007/7657_2015_78. [DOI] [Google Scholar]
- Meijering, E., Jacob, M., Sarria, J.-C. F., Steiner, P., Hirling, H. and Unser, M. (2004). Design and validation of a tool for neurite tracing and analysis in fluorescence microscopy images. Cytometry A 58A, 167-176. 10.1002/cyto.a.20022 [DOI] [PubMed] [Google Scholar]
- Mills, F., Globa, A. K., Liu, S., Cowan, C. M., Mobasser, M., Phillips, A. G., Borgland, S. L. and Bamji, S. X. (2017). Cadherins mediate cocaine-induced synaptic plasticity and behavioral conditioning. Nat. Neurosci. 20, 540-549. 10.1038/nn.4503 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nord, A. S., Chang, P. J., Conklin, B. R., Cox, A. V., Harper, C. A., Hicks, G. G., Huang, C. C., Johns, S. J., Kawamoto, M., Liu, S.et al. (2006). The international gene trap consortium website: a portal to all publicly available gene trap cell lines in mouse. Nucleic Acids Res. 34, D642-D648. 10.1093/nar/gkj097 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plain, F., Howie, J., Kennedy, J., Brown, E., Shattock, M. J., Fraser, N. J. and Fuller, W. (2020). Control of protein palmitoylation by regulating substrate recruitment to a zDHHC-protein acyltransferase. Commun. Biol. 3, 411. 10.1038/s42003-020-01145-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rapsomaniki, M. A., Kotsantis, P., Symeonidou, I.-E., Giakoumakis, N.-N., Taraviras, S. and Lygerou, Z. (2012). easyFRAP: an interactive, easy-to-use tool for qualitative and quantitative analysis of FRAP data. Bioinformatics 28, 1800-1801. 10.1093/bioinformatics/bts241 [DOI] [PubMed] [Google Scholar]
- Reese, A. L. and Kavalali, E. T. (2015). Spontaneous neurotransmission signals through store-driven Ca2+ transients to maintain synaptic homeostasis. eLife 4, e09262. 10.7554/eLife.09262 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Resh, M. D. (2006). Palmitoylation of ligands, receptors, and intracellular signaling molecules. Sci. STKE 2006, re14. 10.1126/stke.3592006re14 [DOI] [PubMed] [Google Scholar]
- Rodriguez, A., Ehlenberger, D. B., Dickstein, D. L., Hof, P. R. and Wearne, S. L. (2008). Automated three-dimensional detection and shape classification of dendritic spines from fluorescence microscopy images. PLoS ONE, 3, e1997. 10.1371/journal.pone.0001997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salaun, C., Greaves, J. and Chamberlain, L. H. (2010). The intracellular dynamic of protein palmitoylation. J. Cell Biol. 191, 1229-1238. 10.1083/jcb.201008160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders, S. S., Martin, D. D. O., Butland, S. L., Lavallée-Adam, M., Calzolari, D., Kay, C., Yates, J. R., III and Hayden, M. R. (2015). Curation of the mammalian palmitoylome indicates a pivotal role for palmitoylation in diseases and disorders of the nervous system and cancers. PLoS Comput. Biol. 11, e1004405. 10.1371/journal.pcbi.1004405 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schizophrenia Working Group of the Psychiatric Genomics Consortium. (2014). Biological insights from 108 schizophrenia-associated genetic loci. Nature 511, 421-427. 10.1038/nature13595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shah, B. S., Shimell, J. J. and Bamji, S. X. (2019). Regulation of dendrite morphology and excitatory synapse formation by zDHHC15. J. Cell Sci. 132, jcs230052. 10.1242/jcs.230052 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimell, J. J., Shah, B. S., Cain, S. M., Thouta, S., Kuhlmann, N., Tatarnikov, I., Jovellar, D. B., Brigidi, G. S., Kass, J., Milnerwood, A. J.et al. (2019). The X-linked intellectual disability gene Zdhhc9 is essential for dendrite outgrowth and inhibitory synapse formation. Cell Rep. 29, 2422-2437.e8. 10.1016/j.celrep.2019.10.065 [DOI] [PubMed] [Google Scholar]
- Sinnen, B. L., Bowen, A. B., Gibson, E. S. and Kennedy, M. J. (2016). Local and use-dependent effects of β-Amyloid oligomers on NMDA receptor function revealed by optical quantal analysis. J. Neurosci. 36, 11532-11543. 10.1523/JNEUROSCI.1603-16.2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skarnes, W. C., von Melchner, H., Wurst, W., Hicks, G., Nord, A. S., Cox, T., Young, S. G., Ruiz, P., Soriano, P., Tessier-Lavigne, M.et al. (2004). A public gene trap resource for mouse functional genomics. Nat. Genet. 36, 543-544. 10.1038/ng0604-543 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steen, R. G., Mull, C., McClure, R., Hamer, R. M. and Lieberman, J. A. (2006). Brain volume in first-episode schizophrenia: systematic review and meta-analysis of magnetic resonance imaging studies. Br. J. Psychiatry 188, 510-518. 10.1192/bjp.188.6.510 [DOI] [PubMed] [Google Scholar]
- Sun, Y. and Bamji, S. X. (2011). β-Pix modulates actin-mediated recruitment of synaptic vesicles to synapses. J. Neurosci. 31, 17123-17133. 10.1523/JNEUROSCI.2359-11.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas, G. M., Hayashi, T., Chiu, S.-L., Chen, C.-M. and Huganir, R. L. (2012). Palmitoylation by DHHC5/8 targets GRIP1 to dendritic endosomes to regulate AMPA-R trafficking. Neuron 73, 482-496. 10.1016/j.neuron.2011.11.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ueno, K. (2000). Involvement of fatty acid synthase in axonal development in mouse embryos. Genes Cells 5, 859-869. 10.1046/j.1365-2443.2000.00369.x [DOI] [PubMed] [Google Scholar]
- Vita, A., De Peri, L., Deste, G. and Sacchetti, E. (2012). Progressive loss of cortical gray matter in schizophrenia: a meta-analysis and meta-regression of longitudinal MRI studies. Transl. Psychiatry 2, e190. 10.1038/tp.2012.116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodley, K. T. and Collins, M. O. (2019). S-acylated Golga7b stabilises DHHC5 at the plasma membrane to regulate cell adhesion. EMBO Rep. 20, e47472. 10.15252/embr.201847472 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie, C., Markesbery, W. R. and Lovell, M. A. (2000). Survival of hippocampal and cortical neurons in a mixture of MEM+ and B27-supplemented neurobasal medium. Free Radic. Biol. Med. 28, 665-672. 10.1016/s0891-5849(99)00268-3 [DOI] [PubMed] [Google Scholar]
- Zaręba-Kozioł, M., Figiel, I., Bartkowiak-Kaczmarek, A. and Włodarczyk, J. (2018). Insights into protein S-palmitoylation in synaptic plasticity and neurological disorders: potential and limitations of methods for detection and analysis. Fron. Mol. Neurosci. 11, 175. 10.3389/fnmol.2018.00175 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.