

Abstract
Drug addiction is a complex, persistent, and chronically relapsing neurological disorder exacerbated by acute and chronic stress. It is well known that the dynorphin/kappa opioid receptor (KOR) system regulates stress perception and responsivity, while the mesolimbic dopamine system plays a role in reward and reinforcement associated with alcohol and substance use disorders. Interestingly, the dopamine and dynorphin/KOR systems are highly integrated in mesolimbic areas, with KOR activation leading to inhibition of dopamine release, further altering the perception of reinforcing and aversive stimuli. Chronic or repeated exposure to stress or drugs potentiates KOR function ultimately contributing to a hypodopaminergic state. This hypodopaminergic state is one of the hallmarks of hyperkatifeia, defined as the hypersensitivity to emotional distress that is exacerbated during drug withdrawal and abstinence. The relationship between stress and drug addiction is bidirectional; repeated/chronic stress promotes pro-addictive behaviors, and repeated cycles of drug exposure and withdrawal, across various drug classes, produces stress. Neuroadaptations driven by this bidirectional relationship ultimately influence the perception of the reinforcing value of rewarding stimuli. In this chapter, we address the involvement of the dopamine and dynorphin/KOR systems and their interactions in shaping reinforcement value processing after drug and stress exposure, as well as a combinatorial impact of both drugs and stress.
Keywords: Alcohol use disorder, Dopamine, Dynorphins, Kappa opioid receptor, Nucleus accumbens, Substance use disorder
1. Introduction
The dynorphin/kappa opioid receptor (KOR) system is a main player involved in regulating stress responsivity, mood, and reinforcement learning. Previous literature suggests that KOR and its interaction with dopamine modulates motivated behaviors as well as aversion and negative affective states – all of which contribute to the role of KOR in drug addiction (Margolis and Karkhanis 2019). Modulation of the KOR system shapes drug-seeking behaviors at various stages of the drug addiction cycle (Wee and Koob 2010; Chavkin and Koob 2016). Because the KOR is coupled to inhibitory G-proteins and potassium channels, KOR activation ultimately inhibits neuronal function, thereby reducing neurotransmission of various neurotransmitters such as dopamine, serotonin, glutamate, and GABA (Heijna et al. 1990; Hill and Brotchie 1995, 1999; Tao and Auerbach 2002; Grilli et al. 2009; Li et al. 2012; Karkhanis et al. 2016a). Although KOR activation inhibits neurotransmitter release, the ultimate behavioral outcome of reduced neuronal activity in a single location depends on the circuit in which KOR is involved. In this review, we focus on dynorphin/KOR system-mediated regulation of dopamine in the mesolimbic pathway and the resulting behavioral impact on stress responsivity, drug self-administration, and stress-induced development of drug addiction. We extend the discussion to how increased function of the dynorphin/KOR system and its interactions with the dopamine system alter the hedonic state of the animal. This results in hyperkatifeia, defined as the hypersensitivity to emotional distress leading to negative affective symptoms seen during drug withdrawal and abstinence.
2. KOR Interactions with the Dopamine System
The modulatory control of the dynorphin/KOR system over dopamine in mesolimbic areas has several behavioral implications including development of a negative affective state and changes in drug self-administration. Using electron microscopy immunohistochemistry, Svingos et al. (2001) showed that among neuronal synapses in the nucleus accumbens (NAc), KOR and dopamine transporter (DAT) are colocalized in presynaptic structures, as well as on the plasma membrane of vesicles within terminals. These data suggest that KOR is located on dopamine neuron terminals and potentially regulate dopamine transmission directly. In this section, we will discuss the interaction of KOR with various dopamine receptors and transporter proteins.
2.1. KOR-Mediated Regulation of Dopamine Release
The activation of KOR leads to inhibition of dopamine release in the NAc core and shell (Heijna et al. 1990; Karkhanis et al. 2016a; Rose et al. 2016a; Siciliano et al. 2016; Melchior and Jones 2017). This KOR activation reduces overall extracellular dopamine levels (Di Chiara and Imperato 1988; Spanagel et al. 1992; Zhang et al. 2004; Chefer et al. 2006; Fuentealba et al. 2006; Gehrke et al. 2008; Karkhanis et al. 2016b) as well as decreases salient stimulus evoked tonic and phasic dopamine release (Karkhanis et al. 2016b; Rose et al. 2016a; Siciliano et al. 2016; Melchior and Jones 2017). A recent study showed that the inhibitory effect of KOR activation on dopamine release is potentiated in the caudal compared to rostral NAc shell (Pirino et al. 2020). These changes in release ultimately occur as a result of augmented K+ and attenuated Ca2+ conductance, resulting in cell hyperpolarization, decreased cell firing, and blockade of neurotransmitter release (Dhawan et al. 1996).
Interestingly, there are sex differences in KOR regulation of dopamine release. Particularly, KOR activation-mediated inhibition of dopamine release is blunted in females compared to males (Abraham et al. 2018; Conway et al. 2019). This sex difference is likely driven by two mechanisms: first, dopamine synthesis is greater in female compared to male rats (Conway et al. 2019); second, estradiol increases phosphorylation of G-protein receptor protein kinase 2, thereby reducing G-protein signaling and ultimately attenuating KOR control over dopamine release in mice (Abraham et al. 2018). Notably, mRNA of dynorphin-precursor (Pdyn) and KOR genes (Oprk1) were equivalent in male and female rats (Conway et al. 2019), suggesting that the availability of KOR and dynorphins may not be different between the two sexes but rather dopamine availability and gonadal hormones may mediate differential intracellular signaling further, promoting the observed differences in magnitude of dopamine release. For further review on sex differences in KOR function, see Chartoff and Mavrikaki (2015).
2.2. KOR-Mediated Regulation of Dopamine Uptake
The DAT is predominantly involved in regulating the reuptake of dopamine from the synapse. Interestingly, co-immunoprecipitation, bioluminescence resonance energy transfer (BRET), and Fröster resonance energy transfer (FRET) data suggest that the DAT and KOR may physically interact, with KOR activation increasing the number of DAT-KOR complexes (Kivell et al. 2014). Because KOR and DAT are colocalized on dopamine terminals, potentially forming physical complexes, it is likely that KOR activation alters DAT function thereby influencing dopamine uptake. Indeed, DAT and KOR have been shown to functionally interact; salvinorin A, a selective KOR agonist, increases DAT function resulting in enhanced rate of dopamine reuptake in an ERK 1/2 dependent manner (Kivell et al. 2014). However, this increase in DAT function may be transient since repeated agonist administration resulted in decreased rate of dopamine reuptake in the NAc (Thompson et al. 2000). This decrease in dopamine uptake rate has also been observed using ex vivo fast scan cyclic voltammetry with the partial KOR agonist nalmefene (Rose et al. 2016b), but not with full KOR agonists (Ebner et al. 2010; Ehrich et al. 2015; Hoffman et al. 2016). These differences could be due to differential intracellular mechanisms of action of the partial agonist nalmefene and full agonists such as U50,488. Furthermore, nalmefene also acts as an antagonist at the mu and delta opioid receptors (Bart et al. 2005). Thus far, there have been no studies showing mu or delta opioid receptors on dopamine terminals, making it unlikely that inhibition or activation of these receptors would result in any changes in DAT function (rate of uptake). Conversely, inhibition of KOR using the antagonist norbinaltorphimine (norBNI) augmented dopamine uptake rate measured by quantifying extraction fraction (Ed, an indirect measure of dopamine uptake) with no-net flux microdialysis (Chefer et al. 2006; Azocar et al. 2019). Overall, these studies suggest that KOR activation modulates DAT function, although this occurs in a manner that is sensitive to acute versus repeated KOR activation, or rather the homeostatic state of the animal.
2.3. KOR-Mediated Regulation of D2/D3 Receptors
Behavioral studies examining interactions between the KOR and membrane bound dopamine receptors have shown synergistic effects of coactivating these receptors. Specifically, acute coadministration of U69,593, a KOR agonist, and quinpirole, a D2R/D3R agonist, resulted in increased dynorphin mRNA expression in the dorsal striatum (Perreault et al. 2007), while no changes were seen with quinpirole alone (Jiang and Wang 1991; Engber et al. 1992; You et al. 1994; Perreault et al. 2007) or U69,593 alone (Tzaferis and McGinty 2001; Perreault et al. 2007). Two days after repeated U69,593 injections not only increased dynorphin mRNA levels (Perreault et al. 2007) but also reduced D2R density in the dorsal striatum. In contrast, 10 days after repeated U69,593 administration, D2R density in the NAc increased (Izenwasser et al. 1998). Repeated KOR activation has also attenuated D2R function on presynaptic dopamine terminals (Acri et al. 2001; Fuentealba et al. 2006), likely due to a decrease in D2R density as observed by Izenwasser et al. (1998), ultimately resulting in elevated extracellular levels of dopamine (Fuentealba et al. 2006). However, when D2R and KOR are repeatedly activated concomitantly, there is an increase in D2R density (Perreault et al. 2007), which may subsequently reduce dopamine levels. Studies have also shown differential changes in locomotor behavior following combinatorial KOR and D2R/D3R activation and activation of each receptor alone. For example, administration of U69,593 or low dose of quinpirole (0.05 mg/kg, considered to target presynaptic D2/D3 autoreceptors) decreased locomotor activity, whereas high dose quinpirole (0.5 mg/kg, considered to target postsynaptic D2/D3 receptors) increased locomotor activity (Perreault et al. 2006). Coadministration of high dose quinpirole and U69,593 further enhanced locomotor activity (Perreault et al. 2006; Escobar et al. 2017) suggesting that postsynaptic D2R and D3R functionally interact with KOR. Together these studies suggest that KOR activation alone versus KOR and D2R/D3R combined activation results in distinct neurochemical and neurobiological changes ultimately affecting behavior. Overall, the interaction between KOR and D2Rs is complex, and more research is necessary to pinpoint the mechanism driving these interactions.
3. Behavioral Effects of KOR Activation in Humans and Animal Models
In humans, KOR activation leads to analgesia, psychotomimesis, sedation, and dysphoria (MacLean et al. 2013; Pande et al. 1996; Pfeiffer et al. 1986; for review see Wadenberg 2003), and similar effects are seen in animals (Shippenberg and Herz 1986; Katoh et al. 1990; Todtenkopf et al. 2004; McLaughlin et al. 2006a). Direct infusions of KOR agonists into the NAc (Bals-Kubik et al. 1989) and systemic KOR agonist administration (Land et al. 2008) have been shown to promote conditioned place aversion or block conditioned place preference (CPP). Furthermore, the aversive effects of exposure to forced swim or foot-shock stress, as reflected by the development of conditioned aversions to odors or places paired with stress, were blocked by KOR antagonist treatment and were absent in Pdyn−/− mice (Land et al. 2008), suggesting that dynorphins and KOR are crucial in forming context-stress associations. Interestingly, however, recent studies have shown that KOR activation results in opposing behavioral outcomes – both negative and positive affective responses. Particularly, activation of KOR in the rostral NAc shell stimulated orofacial hedonic responses to sucrose and CPP; in contrast, activation of KOR in the caudal NAc shell inhibited these orofacial hedonic responses and promoted conditioned place aversion (Castro and Berridge 2014). These differences are likely driven by differential effects of KOR activation on dopamine release as mentioned above (Pirino et al. 2020). Topographical differences were also reflected in operant optical self-stimulation of dynorphinergic neurons; mice exhibited enhanced self-stimulation of dynorphinergic neurons in the dorsal NAc shell but not the ventral shell. This effect in the dorsal NAc shell was reversed in the presence of norBNI (Al-Hasani et al. 2015). Furthermore, selective photostimulation of dynorphinergic neurons in the dorsal NAc shell promoted place preference while stimulation of the neurons in the ventral shell promoted place avoidance (Al-Hasani et al. 2015). Such topographically dependent variations in behavioral outcomes are likely driven by differential and cell-specific KOR expression along the rostro-caudal or dorso-ventral axis.
In addition to topographical effects of KOR activation, direction of study outcomes is also dose- and time-dependent. For example, relatively low doses of U50,488 (0.3–3.0 mg/kg) administered daily within 10 min of conditioning blocked ethanol-induced CPP (Matsuzawa et al. 1999; Logrip et al. 2009); conversely, daily administration of U50,488 (10 mg/kg) 90 min before conditioning potentiated ethanol-induced CPP (Sperling et al. 2010). Similarly, when KOR agonists were acutely administered before intracranial self-stimulation (ICSS) testing, ICSS thresholds dose dependently increased, suggesting a KOR-induced aversive and anhedonic-like state (Todtenkopf et al. 2004; Carlezon et al. 2006; Ebner et al. 2010). This anhedonic/dysphoric state enhances reward sensitivity; when salvinorin A is administered 15 to 60 min before cocaine, ICSS thresholds decreased. In contrast, 24 h after salvinorin A administration ICSS thresholds increased in the presence of cocaine, suggesting decreased sensitivity to rewarding stimuli (Chartoff et al. 2016). The temporal effects of KOR activation may be a result of differential temporal dynamics of KOR interaction with various proteins that regulate dopamine transmission, potentially affecting perceived reward value.
KOR agonists are also known to produce pro-anxiety and pro-depressive effects while antagonists produce anxiolytic and antidepressive effects (for review, see Van’t Veer and Carlezon 2013). For example, in the forced swim test, a classic test used to screen potential antidepressants (Porsolt et al. 1977; Detke et al. 1995), systemic administration of U69,593 in rats promotes immobility (a sign the animal is no longer trying to escape), whereas intracerebroventricular administration of norBNI diminishes immobility behavior. These results suggest that KOR agonists increase depressive-like behaviors while antagonists have antidepressant-like effects (Mague et al. 2003; Carr et al. 2010). Similarly, in the learned helplessness model of behavioral depression both intracerebroventricular and intra-NAc administration of norBNI decreased escape failures, suggesting that norBNI had an anti-depressant-like effect (Newton et al. 2002; Shirayama et al. 2004). Likewise, KOR antagonists produced anxiolytic-like effects in multiple assays, including the elevated plus maze (Knoll et al. 2007; Huang et al. 2016), fear-potentiated startle (Knoll et al. 2007), novelty induced hypophagia (Knoll et al. 2007; Carr and Lucki 2010), and defensive burying tests (Carr and Lucki 2010). For further review, see Hang et al. (2015).
Behavioral effects of KOR activation, including analgesia, anxiety, depression, and anhedonia are sex-dependent (for review see Chartoff and Mavrikaki 2015). While the KOR agonist U50,488 dose-dependently augmented ICSS responding in both male and female rats, female rats, independent of estrous cycle phase, were significantly less sensitive to its reward-decreasing effects (Russell et al. 2014). Another study showed that this effect in females was not due to circulating gonadal hormones (Conway et al. 2019). Although rodent studies have shown no differences in gene expression of the KOR and Pdyn genes in the NAc or VTA (Conway et al. 2019), human PET imaging studies of the whole brain revealed less KOR availability in females compared to males (Vijay et al. 2016). It is possible that active KOR availability is in fact different even though gene expression is comparable between the two sexes in preclinical models; similar PET imaging studies in preclinical models have not been completed. Ultimately, further research is needed in order to understand the mechanisms that drive these sex differences and how they may affect clinical translation of KOR-targeted medications.
4. Involvement of KOR in Addiction Processing and Stress
The dynorphin/KOR system has been shown to interact bidirectionally with stress and drug exposure. Some studies even suggest that acute and chronic exposure to stress drive opposing changes in levels of dynorphins. Moreover, numerous studies have shown a direct link between chronic drug exposure and KOR function in the NAc. In general, repeated exposure to various drug classes such as alcohol, opioids, and stimulants produces a hyperfunctioning KOR system (Wee and Koob 2010). In this section, we will discuss the interaction between the dynorphin/KOR system and exposure to (1) alcohol; (2) other substances of misuse; and (3) stress. Lastly, we tie these three areas of research together and explain the role of the dynorphin/KOR system in stress-induced development of addiction.
4.1. KOR and Alcohol Use Disorder
Based on preclinical evidence, the United States Food and Drug administration approved naltrexone, a nonselective opioid receptor antagonist, as a treatment option for alcohol use disorder (Fairbanks et al. 2020). In humans with moderate to heavy drinking experience, PET imaging with the selective KOR ligand [11C]-LY2795050 showed that occupancy of the KOR by naltrexone is associated with naltrexone-mediated reduction in alcohol drinking and craving, suggesting that alcohol use changes dynorphin/KOR system function, which potentially is involved in the clinical response of reduced consumption (de Laat et al. 2019, 2020). Particularly, augmented craving before naltrexone administration was correlated with higher KOR occupancy by naltrexone, which suggests greater KOR availability (de Laat et al. 2020). Given the interaction between the dynorphin/KOR and dopamine systems, it is possible that both are involved in the progression of alcohol use disorder. Indeed, preclinically, acute systemic administration of ethanol elevates extracellular levels of dynorphins (Marinelli et al. 2006) and dopamine (Di Chiara and Imperato 1988; Yim and Gonzales 2000). Interestingly, however, dopamine levels peak roughly 20 min post injection (Di Chiara and Imperato 1988; Yim and Gonzales 2000), while levels of dynorphins peak at roughly 30–40 min (Marinelli et al. 2006) when dopamine levels are descending. These studies indicate that ethanol may cause both acute and persistent changes in the dynorphin/KOR and dopamine systems. Indeed, changes in KOR function have been observed using various ethanol exposure paradigms across species (Karkhanis et al. 2016a; Siciliano et al. 2016). Using a chronic intermittent ethanol (CIE) exposure model in adult male mice, KOR function was potentiated, which was demonstrated by greater inhibition of tonic and phasic dopamine release in CIE-exposed mice compared to air-exposed controls (Karkhanis et al. 2016a; Siciliano et al. 2016; Melchior and Jones 2017). Monkeys with a history of excessive ethanol consumption also exhibited augmented KOR-mediated inhibition of dopamine release (Siciliano et al. 2016). Interestingly, levels of dynorphins were observed to be diminished following CIE exposure in mice (Rose et al. 2016a). These studies collectively suggest that repeated ethanol exposure and withdrawal cycles may result in prolonged activation of the dynorphin/KOR system even though these cycles of exposure acutely increase levels of dynorphins. The repeated elevations in levels of dynorphins may eventually lead to overall reduction in availability of dynorphins, which may further enhance KOR function at the receptor. More research is necessary to understand this relationship between availability of dynorphins and KOR function.
As mentioned before, an acute ethanol challenge in naïve mice and rats elevates extracellular levels of dopamine (Yim and Gonzales 2000; Karkhanis et al. 2014, 2016a). In contrast, an acute ethanol challenge during acute withdrawal following CIE exposure decreases extracellular levels of dopamine, an effect reversed by pretreatment with a KOR antagonist (Karkhanis et al. 2016a). These data suggest that CIE-mediated increase in KOR function ultimately alters the reinforcing value of ethanol. Together, the augmented control of KOR over dopamine at baseline and the blunting effects of ethanol-induced elevation in dopamine suggest that the dynorphin/KOR system may also contribute to the CIE exposure-dependent enhancement in ethanol consumption and anxiety-like/compulsive behaviors (Anderson et al. 2016; Rose et al. 2016b). Indeed, the CIE-associated augmentation in ethanol consumption and marble burying (compulsive) behavior in mice was reversed with KOR blockade (Rose et al. 2016b).
Ethanol-induced changes in the dynorphin/KOR system extend to other mesolimbic regions such as the bed nucleus of the stria terminalis (BNST). For example, KOR blockade via direct infusion of the KOR antagonist norBNI into the BNST decreased both binge drinking and sucrose consumption in mice exposed to drinking-in-the-dark (DID), a model of binge-like ethanol consumption (Haun et al. 2020). Moreover, systemic administration of norBNI, as well as DREADD-based inhibition of dynorphinergic neurons into the central amygdala, also reduced binge drinking in the DID model (Anderson et al. 2019). Together, these studies show that heavy and moderate ethanol exposure enhances ethanol consumption due to, at least in part, augmented KOR activity in the mesolimbic pathway. Neurochemical analysis in the NAc suggests that these behavioral changes are likely driven by enhanced KOR-mediated inhibitory control over dopamine transmission. The switch from dopamine-exciting effects of ethanol after acute exposure to the dopamine-depleting effects of ethanol after chronic exposure is likely a result of this augmented KOR control over dopamine. Ultimately, these neuroadaptations, summarized in Fig. 1, not only result in temporary negative affect but also promote prolonged hyperkatifeia, maintaining the animal in a negative emotion state.
Fig. 1.
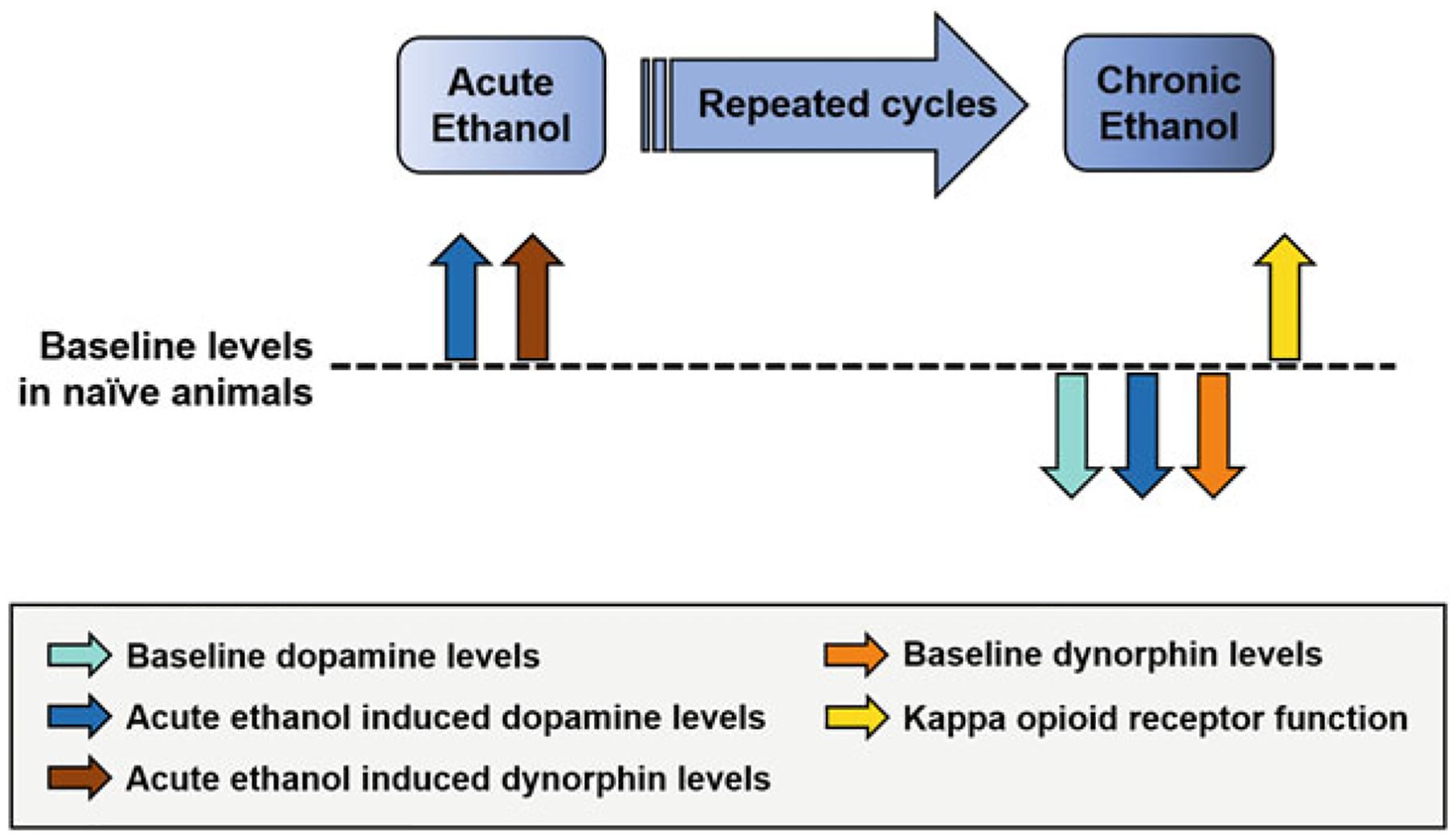
Impact of acute and chronic ethanol exposure on dopamine and dynorphin/KOR systems. In naïve animals, acute ethanol results in elevations in extracellular levels of dopamine (blue arrow) and dynorphins (brown arrow). Repeated cycles of ethanol exposure and withdrawal result in attenuated baseline dopamine (aqua arrow) and dynorphins (orange arrow) levels and augmented KOR function (yellow arrow). Subsequent acute ethanol challenge following chronic ethanol exposure during acute withdrawal results in a reduction in extracellular levels of dopamine (dark blue arrow). The reduced baseline dopamine levels and reduction in dopamine in response to acute ethanol challenge suggest that repeated cycles of ethanol exposure and withdrawal produce a hypodopaminergic state and ethanol tolerance, respectively. These effects may be driven by augmented KOR function or diminished levels of dynorphins. Current literature does not identify the order of changes in dopamine, dynorphins, and KOR. However, it is important to recognize that both the dopamine and dynorphin/KOR systems are integrated, leading to neuroadaptations that may bidirectionally affect each system
The observed ethanol-induced changes in dopamine and KOR function are both sex-specific and dependent on age of ethanol exposure; findings reported thus far have been in male adult rodents. In this subsection, we will discuss the impact of ethanol exposure during adolescence on the dynorphin/KOR system. Adolescent intermittent ethanol (AIE) exposure during early adolescence (PD 25–45) followed by prolonged forced abstinence potentiated KOR function as measured by its inhibitory control over dopamine transmission in adult female rats (Spodnick et al. 2020). Interestingly, this effect was not observed in male rats. On the contrary, KOR function was attenuated following prolonged forced abstinence in male rats exposed to AIE during late adolescence (PD 45–65), an effect not observed in female rats exposed during late adolescents (Spodnick et al. 2020). These sex differences suggest that KOR function in males and females is profoundly distinct and dependent on the age of ethanol exposure. While measurements of KOR mRNA levels in male animals across perinatal development indicate that KOR mRNA reaches adult levels by early adolescence (Georges et al. 1998), hardly any studies exist examining the ontogeny of the female KOR system. From a developmental perspective, it is important to note that the inhibitory control of KOR over dopamine release increases between embryonic day 17 through PD 7 and then decreases between PD 7 and 21 in male rats to the adult levels, suggesting that the KOR system is matured at the beginning of adolescence (De Vries et al. 1990). However, the relationship between the dynorphin/KOR and dopamine systems is complex, especially during the developmental adolescence stage since both systems mature at distinct ages. While the dynorphin/KOR system appears to mature by PD 21, the dopamine system continues to mature into young adulthood, albeit in a sex-dependent manner. For instance, studies of dopaminergic neural markers suggest that the female dopaminergic system steadily progresses toward levels typical of adulthood throughout adolescence, whereas studies of dopaminergic firing in males follow an inverted-U-shaped trajectory of development in which firing peaks at PD 42–48 and then declines toward adult levels (Prakash and Wurst 2006; Alavian et al. 2008; Katunar et al. 2009; McCutcheon and Marinelli 2009; Marinelli and McCutcheon 2014). Thus, it is possible that the impact of ethanol on the dopamine and KOR systems is highly dependent on age. KOR regulation of dopamine transmission may also differ between sexes since maturation of the dopamine system is delayed in males. These differences likely drive the differential effects of adolescent intermittent ethanol exposure on KOR function following forced abstinence. More systematic research is necessary to pinpoint exact mechanisms that drive these age- and sex-specific neuroadaptations.
Based on the preclinical literature, at least in adulthood, KOR has emerged as a promising therapeutic target to treat alcohol use disorder. Naltrexone, an antagonist of the opioid receptors with higher affinity for the mu opioid receptor over delta and kappa opioid receptors, is currently approved by the Food and Drug Administration for treating alcohol use disorder. It functions by blunting the positive reinforcing effects of alcohol; however, its effectiveness is moderate in the clinic (Ashenhurst et al. 2012). Most preclinical studies used specific KOR antagonists that were long-acting; the complex mechanisms of action and interactions of these drugs deem them problematic in humans. In fact, phase I clinical trial of JDTic failed due to cardio-toxic effects (Buda et al. 2015). JDTic activates the cJun N-terminal kinase (JNK), similar to norBNI, ultimately inactivating the KOR (Melief et al. 2010, 2011; Buda et al. 2015; Schattauer et al. 2017). Though norBNI can no longer be detected in plasma levels after 1 day, the behavioral effects of norBNI are persistent for at least 21 days (Horan et al. 1992; Bruchas et al. 2007b; Munro et al. 2012), with one study showing effects for at least 85 days (Potter et al. 2011). KOR antagonists that are not long-lasting and do not target the JNK pathway may be safer therapeutic alternatives. Thus, recent clinical studies have begun examining the effects of short-acting KOR antagonists such as aticaprant (developmental codes JNJ-67953964, CERC-501, and LY-2456302) (Page et al. 2019). CERC-501 (now aticaprant) has been shown to reverse anxiety-like behaviors resulting from chronic ethanol exposure as well as block stress-induced ethanol reinstatement (Domi et al. 2018). LY2444296, an analog of aticaprant, has also been effective in reducing ethanol consumption in rodent studies (Anderson et al. 2019). Positive results from preclinical studies using short-acting KOR antagonists have prompted human studies and clinical trials of aticaprant (NCT02800928, NCT02218736, NCT01913535, NCT02641028).
4.2. KOR and Substance Use Disorder
Similar to alcohol, exposure to other drugs such as stimulants alters the dynorphin/KOR system, making the KOR a prime cellular target for therapeutics to treat substance use disorder. For instance, cocaine exposure elevates levels of dynorphins (Sivam 1989; Smiley et al. 1990), with repeated injections of cocaine (5 injections, 30 mg/kg, i.p., 6 h apart) increasing immunoreactivity to content of dynorphins to approximately 270% of the control in the NAc 8 h after the last cocaine injection (Smiley et al. 1990). In a similar study, repeated injections of cocaine (20 mg/kg, i. p.) once per day for 4 days resulted in augmented levels of dynorphins in the striatum, which were persistent 4 days post-cocaine administration (Sivam 1989). Likewise, Pdyn mRNA and dynorphin expression levels in the striatum were elevated in rodents after non-contingent cocaine exposure (Daunais et al. 1993; Spangler et al. 1993, 1996; Schlussman et al. 2003, 2005; Sun et al. 2020). Repeated cocaine administration reduced synthesis of dynorphins due, at least in part, to augmented ΔFosB expression and attenuated CREBS (Nestler et al. 2001). With respect to KOR, in situ U69,593-induced [35S]GTPγS binding in VTA of rats increased 24 h into withdrawal after chronic cocaine exposure, suggesting greater KOR function in an acute withdrawal state (Piras et al. 2010). It is possible that the augmented KOR function is a resulting neuroadaptation due to attenuated availability of dynorphins observed during drug withdrawal. In humans with a recent history of cocaine use, as well as cocaine-overdose victims, dynorphin mRNA and KOR receptor binding was increased in postmortem striatal tissue, also suggesting an upregulation of the KOR system (Hurd and Herkenham 1993; Staley et al. 1997; Mash and Staley 1999). After a 3-day cocaine binge (average 1,500 mg of smoked cocaine), PET imaging revealed that cocaine-dependent individuals had reduced KOR agonist binding by approximately 20%, suggesting an elevation in levels (Martinez et al. 2019). Together, these studies suggest that cocaine exposure results in an upregulation of the dynorphin/KOR system. The contingency factor of cocaine self-administration, timing of the last cocaine exposure, and withdrawal state seem to be critical in driving changes in the dynorphin/KOR system, particularly modulating levels of dynorphins. Alterations in levels of dynorphins likely promote upregulation in KOR function.
Interestingly, both KOR agonists and antagonists are efficacious at decreasing drug-taking behaviors in preclinical models of substance use (for review see Karkhanis et al. 2017), with both agonists and antagonists showing similar effects. It is very important to understand the link between KOR activation and drug-taking/seeking behaviors and how the behavioral output of KOR activation is time-dependent. KOR agonists may modulate drug-seeking behaviors through diminishing the rewarding effects of cocaine by KOR activation (Shippenberg et al. 2007). This would make the effects of KOR agonists most beneficial early in cocaine use since KOR activation averts the dopamine-elevating properties of drugs. In several preclinical studies, the beneficial effects of KOR activation are time-dependent. For example, administering cocaine 15 or 60 min after systemic KOR agonist administration led to profound or slight decreases, respectively, in ICSS threshold, suggesting an increased reward valuation of cocaine; however, when cocaine was administered 24 h after KOR activation, ICSS threshold increased, suggesting reduced reward valuation (Chartoff et al. 2016). Interestingly, McLaughlin et al. (2006a) observed that activating KOR 15 min before conditioning blocked cocaine CPP in mice, whereas activation 60 min prior to conditioning potentiated cocaine CPP, confirmed by Ehrich et al. (2014). Although the ICSS and CPP data suggest contrasting findings at the 15-min timepoint, but similar results at 60-min timepoint, these differences could be due to the differential aspects of behavioral responses of the two paradigms. It is possible that this contrasting result is due to KOR activation and cocaine administration occurring in close temporal proximity (15 min), thus the cocaine injection would have a negative association, further leading to blockade of CPP. Ultimately, however, given these time-dependent differences, timing of KOR activation with respect to cocaine administration likely have differential effects on the perceived reward value of cocaine.
On the other hand, preclinical behavioral studies have consistently shown that KOR antagonists do not block the rewarding effects of drugs as KOR agonists do; instead, KOR antagonists block the stress-induced potentiation of drug reward, stress-induced reinstatement, and escalation of drug consumption (for review see Bruchas et al. 2010). After animals self-administered cocaine on a long access paradigm (6 h/day with unlimited infusions, 0.5 mg/kg/inf), administration of norBNI does not alter the amount of cocaine consumed on a fixed ratio one schedule of reinforcement. However, when conditions were switched to a progressive ratio schedule, norBNI treated animals exhibited lower breakpoints, suggesting reduced motivation to self-administer cocaine (Wee et al. 2009). These data suggest that KOR blockade selectively reduces cocaine seeking (appetitive behavior) but not cocaine consumption. Interestingly, only animals with a history of extended cocaine exposure were sensitive to norBNI effects, suggesting an importance of drug history; animals that self-administered on a short access procedure (1 h/day with unlimited infusions, 0.5 mg/kg/inf) were unaffected by norBNI (Wee et al. 2009). These data indicate that longer sessions with cocaine promote KOR function, while shorter access sessions either have no effect or the opposite effect on KOR function. Indeed, long access (6 h/day) and short access (1–2 h/day) to cocaine have differing effects of cocaine potency at the dopamine transporter (DAT). Long access leads to reduced inhibition of dopamine uptake at the DAT by cocaine, while short access increases the effect of cocaine on the DAT (Calipari et al. 2013). As discussed earlier, KOR and DAT interact with each other (Kivell et al. 2014); therefore, cocaine may result in complex changes in the two proteins and how these proteins interact, which could be dependent on the amount of cocaine exposure. Interestingly, cocaine potency at the DAT tracks with motivational state of the animal measured by cocaine seeking/motivation to perform even at greater response requirements to maintain blood cocaine levels, but it does not track or predict consumption (Siciliano and Jones 2017). Given the interaction between KOR and DAT, KOR potentially contributes to this effect on motivated behavior. This motivational state of the animal is likely crucial for the antagonist to be effective in reducing drug-seeking behaviors.
Research thus far has shown that KOR antagonists reduce drug-seeking behaviors (for review see Karkhanis et al. 2017). However, as mentioned earlier, most rodent studies to date have used long-acting antagonists that have complex actions, which have increased interest in developing short-acting KOR antagonists to study more discrete changes with KOR blockade. The short-acting KOR antagonist LY244296 was recently shown to reduce escalation of cocaine intake on an extended access paradigm (fixed ratio one, 18 h/day) in rats (Valenza et al. 2020). Additionally, responding decreased when re-exposed to cocaine after 2 days of abstinence in animals treated with LY2444296 compared to vehicle-treated animals (Valenza et al. 2020). Animals in these studies may be sensitive to the behavioral effects of KOR antagonists due to an upregulation of KOR after repeated cocaine exposure (Spangler et al. 1993; Unterwald et al. 1994). Clinical studies have shown that the KOR antagonist LY2456302 is safe to administer in early-abstinent cocaine-dependent individuals (Reed et al. 2018). This study examined the effects of only one dose of LY2456302 (10 mg) for 4 consecutive days, which did not alter subjective cocaine craving (Reed et al. 2018). Overall, illicit substances affect multiple brain mechanisms leading to various neuroadaptations; the ones related to the dynorphin/KOR and dopamine systems are summarized in Fig. 2. For example, as discussed in this section, although cocaine primarily targets the DAT, repeated exposure to cocaine affects both dynorphin/KOR and dopamine systems, perhaps due to repeated cycles of withdrawal, which likely promotes stress. Preclinical studies have shown promising results for the use of KOR antagonists for substance use disorder, with clinical studies showing that short-acting KOR antagonists are safe in cocaine-dependent individuals. However, further clinical research is necessary to fully understand the effects of KOR antagonism on cocaine craving and taking throughout various phases of the addiction cycle.
Fig. 2.
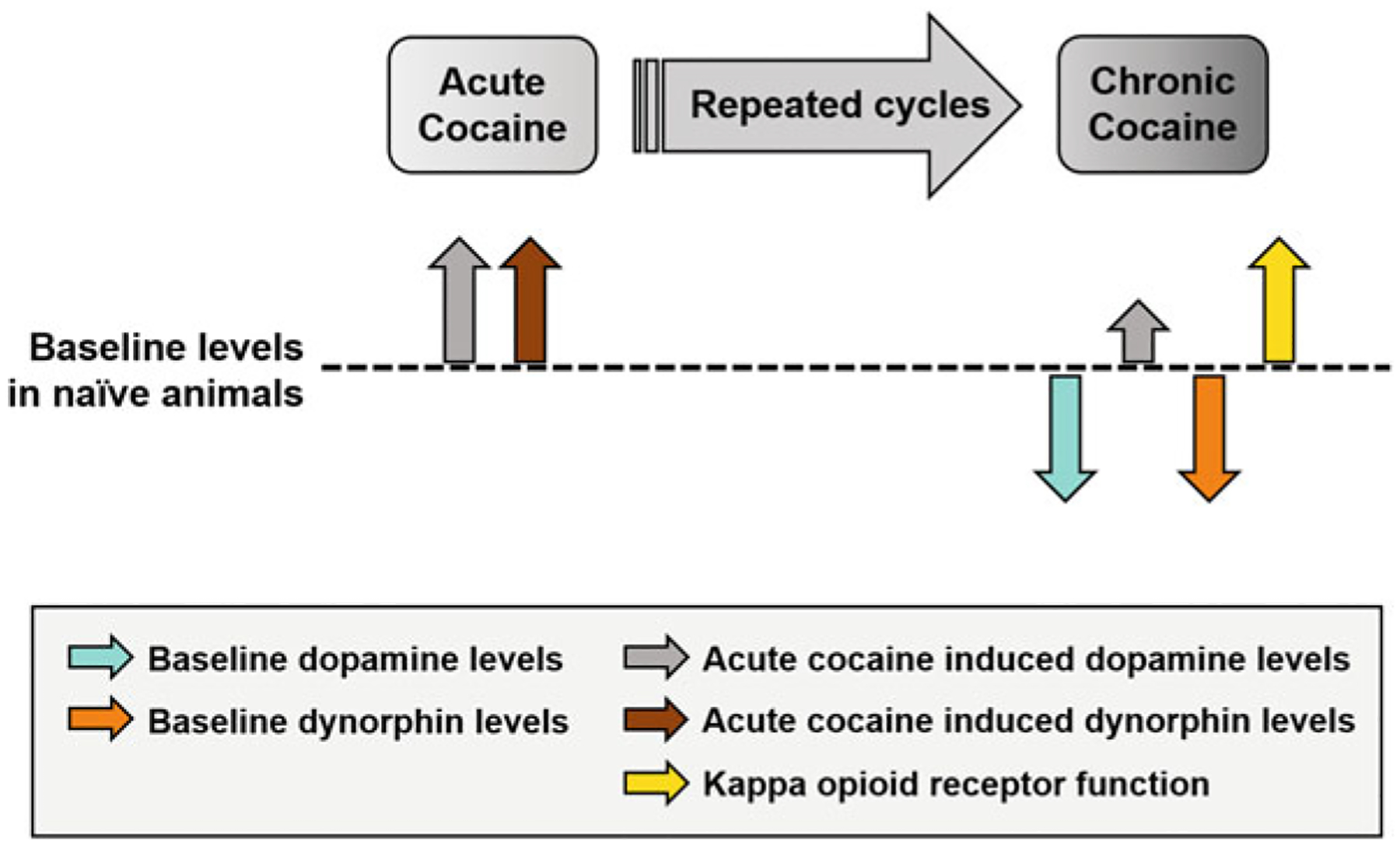
Impact of acute and chronic cocaine exposure on dopamine and dynorphin/KOR systems. In naïve animals, acute cocaine results in elevations in extracellular levels of dopamine (gray arrow) and dynorphins (brown arrow). Repeated cycles of contingent cocaine exposure and withdrawal result in attenuated baseline dopamine (aqua arrow) and dynorphins (orange arrow) levels and augmented KOR function (yellow arrow). Subsequent acute cocaine challenge following chronic cocaine exposure results in an increase in dopamine levels (gray arrow), albeit the difference from baseline is smaller than in cocaine naïve animals. The reduced baseline dopamine levels and a reduction in dopamine in response to acute cocaine challenge suggest that repeated cycles of cocaine exposure and withdrawal produce a hypodopaminergic state and cocaine tolerance, respectively. It is important to note that the chronic cocaine effects summarized here are based on long daily sessions of operant self-administration. It is possible that cocaine-induced changes in the dopamine system produce profound neuroadaptive alterations in the dynorphin/KOR system as a result of the interactions between the two neuromodulatory systems
4.3. Acute and Chronic Stress Impact on Dynorphin/KOR System
Both acute and chronic stress exposure affect dopamine transmission; acute stressors such as foot shock, tail pinch, and single social defeat stress exposure elevate dopamine levels in the NAc (Rougé-Pont et al. 1993; Kalivas and Duffy 1995; Lapiz et al. 2003; Holly et al. 2015), while chronic or repeated stress exposure attenuates tonic dopamine levels in the NAc (Scheggi et al. 2002; Karkhanis et al. 2016b). The initial dopamine response to acute stress is likely driven by arousal, and the later subsequent decrease in dopamine may be related to neural adaptations or coping failure at a behavioral level (Cabib and Puglisi-Allegra 1991; Pani et al. 2000). Given the interaction between dopamine and dynorphin/KOR systems, the neuroadaptive changes may be associated with changes in the dynorphin/KOR system (see Margolis and Karkhanis 2019 for review). In this section, we discuss the interaction between stress and KOR, particularly examining the impact of stress on the dynorphin/KOR system and ultimately exploring the impact that KOR may have on stress-related drug-seeking behaviors. Both acute and chronic/repeated stress exposure induce changes in the dynorphin/KOR system, affecting ligand levels as well as receptor function. Acute stress, such as acute psychological stress (Takahashi et al. 1990), electric tail or foot shock (Watkins et al. 1992; Menendez et al. 1993; Land et al. 2008), forced swim stress (McLaughlin et al. 2003; Shirayama et al. 2004), immobilization (Shirayama et al. 2004), and social defeat stress (McLaughlin et al. 2006a; Donahue et al. 2015) elevate levels of dynorphins. On the other hand, the impact of prolonged or repeated stress on levels of dynorphins is inconsistent; for example, we and others have shown that prolonged adolescent social isolation and repeated exposure to social defeat stress attenuated tissue levels of dynorphins in the NAc (Donahue et al. 2015; Karkhanis et al. 2016b). In contrast, Bérubé et al. (2013) show augmented accumbal levels of dynorphins in a subgroup of animals found to be vulnerable to social defeat (animals quick to assume a subordinate position) with no change in resilient, or less subordinate, rats. Despite discrepancies in the literature regarding ligand levels, KOR expression has been shown to increase after chronic stress exposure. For example, KOR immunoreactivity was augmented after repeated swim stress (Bruchas et al. 2007a), and KOR activity in the NAc was enhanced following prolonged adolescent social isolation (Karkhanis et al. 2016b). In the subsequent text, we will discuss the impact of various types of chronic stress exposure on KOR-mediated regulation of behavioral outcomes and dopamine transmission.
Repeated social defeat stress episodes increase ICSS thresholds, indicating anhedonic-like behavior. Interestingly, this effect is dampened in mice lacking KOR specifically on dopamine neuron terminals (Donahue et al. 2015), suggesting that KOR-mediated changes in dopamine transmission are essential for stress-induced anhedonia. Furthermore, blockade of the KOR on second and third exposure to social defeat stress reduced stress-induced immobility and defeat posture (McLaughlin et al. 2006b). Similarly, KOR blockade in mice before social defeat stress exposure prevented the development of anhedonia and social avoidance behaviors (Williams et al. 2018). In rats, repeated restraint stress attenuated social investigation and preference behaviors in a KOR-dependent manner (Varlinskaya et al. 2018). Particularly, this stress paradigm also decreased palatable tastant intake in both adult and adolescent rats (Anderson et al. 2013). Interestingly, however, while acute pharmacological KOR activation promoted social avoidance and taste aversion to a palatable substance in stress-naïve adult rats, stress-exposed rats showed a dampened taste aversion and no change in social avoidance compared to controls (Anderson et al. 2013; Varlinskaya et al. 2018). Moreover, KOR antagonists blocked ethanol-induced conditioned taste aversion in stress-exposed but not stress-naïve adult rats (Anderson et al. 2013). Interestingly, conditioned taste aversion is not blocked by KOR inhibition in restraint stress-exposed adolescents as it is in restraint stress-exposed adults (Anderson et al. 2013), suggesting that stress-associated conditioned taste aversion is mediated via a mechanism other than KOR in adolescents. Together, these studies show that repeated KOR activation due to stress is imperative to induce anhedonic-like behaviors and stress responsivity. However, it is important to note that the effects of KOR modulation are largely dependent on age and homeostatic state of the animal.
Overall, these studies show that while acute activation of KOR induces stress, acute and chronic stress differentially affect the dynorphin/KOR system, with acute stress exposure elevating levels of dynorphins and chronic/repeated stress attenuating levels of dynorphins but augmenting functional responsivity of the dynorphin/KOR system (Fig. 3). This biphasic response of KOR may be driven by presynaptic and postsynaptic neuroadaptations occurring from repeated stress exposure. Effects of stress on the dynorphin/KOR system are also dependent on age of exposure to stress.
Fig. 3.
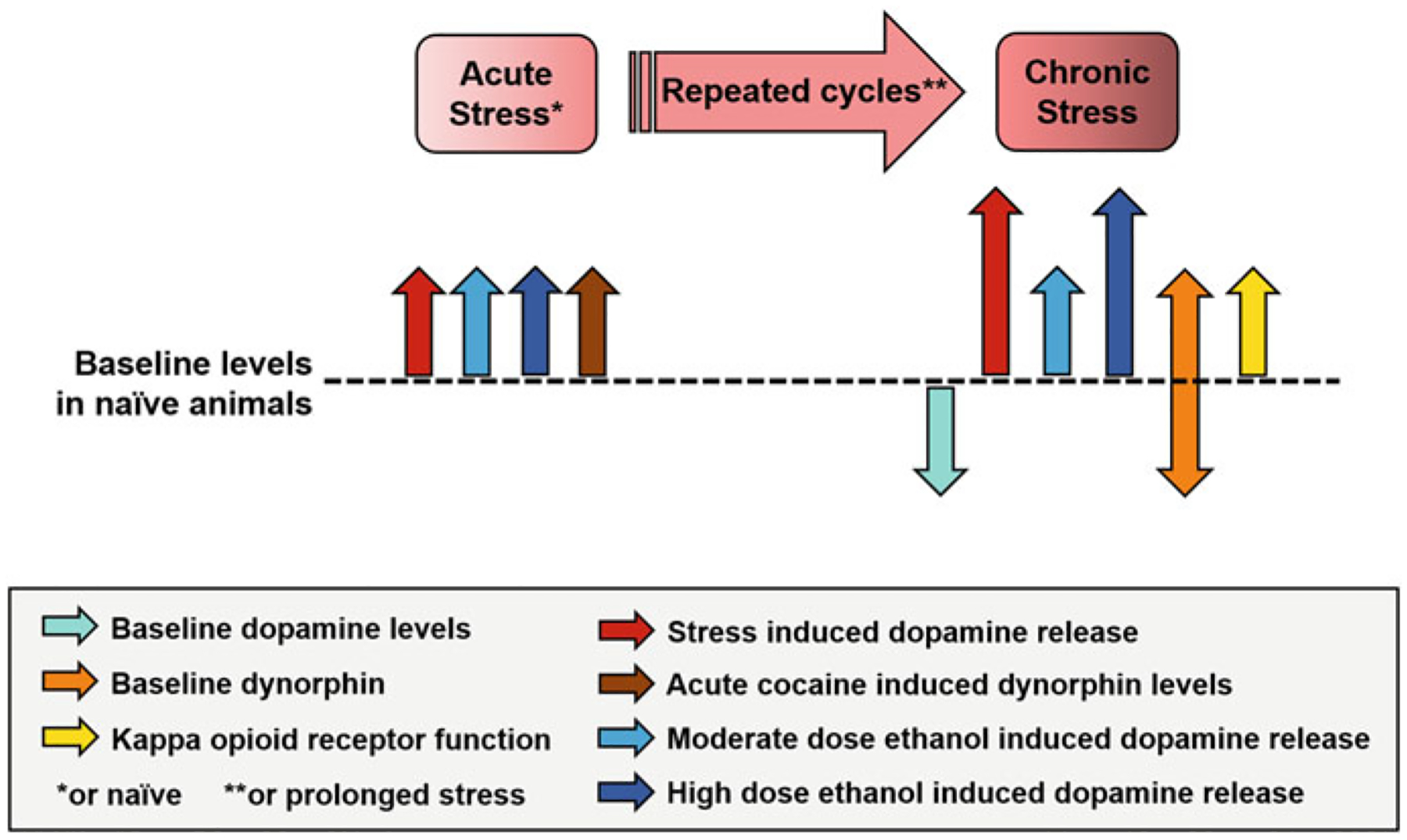
Impact of acute and chronic stress exposure on dopamine and dynorphin/KOR systems. In naïve animals, acute stress results in elevations in extracellular levels of dopamine (red arrow) and dynorphins (brown arrow). Acute systemic ethanol administration at moderate (1 g/kg; light blue arrow) and high doses (2 g/kg; dark blue arrow) result in comparable elevations in extracellular levels of dopamine. Repeated cycles of stress exposure or prolonged stress exposure attenuates baseline dopamine levels (aqua arrow); however, reports on levels of dynorphins (orange arrow) are mixed with some studies showing an elevation and others showing a reduction. Interestingly, all studies report an elevation in KOR function (yellow arrow). Dopamine response to subsequent stress (red arrow) and high dose of ethanol (2 g/kg; dark blue arrow) exposure is exacerbated in chronic stress-exposed animals. A subsequent moderate ethanol dose (1 g/kg; light blue) elicits a dopamine response comparable to stress naïve animals, but this response is augmented with KOR blockade (see text). Because stress affects both the dopamine and dynorphin/KOR systems, the resulting interactions are complex. Ultimately, however, these neuroadaptations promote pro-addictive behaviors
4.4. Stress-Induced Development of Drug Addiction
The dynorphin/KOR system plays an integral role in stress responsivity and resultant drug-related behaviors. Phenotypes of depression and anxiety can be induced by stressors in humans (Kessler 1997), with stress exposure in animal models resulting in similar depression- and anxiety-like behaviors (for review see Chavkin and Koob 2016). These effects can be attributed, at least in part, to KOR decreasing dopamine system function in mesocorticolimbic areas involved in reinforcement circuitry (Margolis and Karkhanis 2019 for review). Substance use after stress exposure, as well as stress related to drug withdrawal, plays a role in drug craving and relapse (Sinha 2007). One theoretical framework proposed that the effects of KOR activation on drug self-administration is dependent on the hedonic state of the animal, which changes based on stress history (see Bruchas et al. 2010 for review). A reward such as cocaine would increase the hedonic state to a smaller degree in non-stressed animals compared to stress-exposed animals since a stressed animal is likely in a dysphoric-like state resulting from KOR hyperfunction. This would contribute to a drug reward having a larger positive valence in stress-exposed animals, increasing drug reinforcement and subsequent taking.
One connection between stress and addiction identified in humans thus far includes a human variant on the KOR gene (OPRK1, rs6989250 C>G), which is associated with increased stress-induced cocaine craving and risk of relapse (Xu et al. 2013). Functional MRI results also showed that individuals with this OPRK1 variant had increased activity in the limbic and midbrain regions during stress and drug-related cues (Xu et al. 2013). OPRK1 variants have also been associated with increased risk of opioid addiction (SNP G36 T, Gerra et al. 2007) and alcoholism (multiple SNPs, Xuei et al. 2006). Findings from Martinez et al. (2019) support this relationship between stress and increased cocaine-taking behaviors in humans. Subjects with cocaine use disorder were subject to a cold pressor task, which induces a stress response, prior to self-administering cocaine in a choice procedure. Results showed that subjects who made more stress-induced cocaine choices had greater KOR availability in the striatum as determined by PET scanning (Martinez et al. 2019).
Several preclinical stress models exhibit pro-addictive behaviors. In the following section it will be important to consider (1) the type of stressor, (2) the stage of addiction (acquisition, maintenance, relapse), as well as (3) the outcome measure when trying to understand the association between stress, KOR system activation, and development or exacerbation of an addiction-like phenotype.
Repeated forced swim stress and repeated social defeat stress potentiate cocaine CPP and nicotine CPP, both blocked by pretreating animals with norBNI (Jackson et al. 2013; McLaughlin et al. 2003; Nygard et al. 2016). Repeated forced swim stress has also been shown to elevate motivation to self-administer cocaine on a threshold procedure, which was attenuated after KOR inhibition via norBNI (Groblewski et al. 2015). Genetic disruption of KOR or dynorphin (KOR −/− or Pdyn −/−) has been shown to block cocaine and nicotine CPP (McLaughlin et al. 2003; Nygard et al. 2016). Stress-induced reinstatement, a self-administration model of relapse-like behavior, is another way the role of the KOR system in modulating stress-induced drug and alcohol seeking can be examined. For example, norBNI prevented stress-induced cocaine reinstatement in rats exposed to cold-water forced swim stressor (Polter et al. 2014, 2017). Another KOR antagonist JDTic reduced foot shock-induced reinstatement (Beardsley et al. 2005). These studies indicate that KOR antagonists are capable of reducing drug-seeking behavior after stress exposure. KOR antagonists have been shown to prevent not only stress-induced reinstatement of cocaine (Beardsley et al. 2005; Carey et al. 2007; Redila and Chavkin 2008) but also stress-induced reinstatement of heroin (Sedki et al. 2015) and ethanol (Sperling et al. 2010) without any effects on drug-primed reinstatement. These results suggest a conserved role of the KOR in inducing stress-dependent reinstatement of various drugs of abuse.
Prolonged stress exposure during adolescence, such as the adolescent social isolation model discussed earlier, promotes anxiety-like behavior, ethanol drinking, cocaine consumption, cocaine seeking, and a hypodopaminergic state (Yajie Ding et al. 2005; Karkhanis et al. 2014; Skelly et al. 2015; Rose et al. 2016a; Fosnocht et al. 2019). These studies have linked augmented ethanol consumption and changes in the dopamine system to an increase in KOR system function (Karkhanis et al. 2016b). For example, adolescent socially isolated (aSI) rats exhibited greater ethanol intake and ethanol preference compared to adolescent group housed (aGH) rats. This augmented ethanol intake and preference was reversed by norBNI administration selectively in aSI animals, suggesting a greater inhibitory control of KOR over dopamine (Karkhanis et al. 2016b). Indeed, direct examination of KOR function using ex vivo fast scan cyclic voltammetry showed hyper-responsivity of KOR to the agonist U50,488, as the inhibitory effects of KOR activation on dopamine release were observed to be significantly greater in aSI compared to aGH rats (Karkhanis et al. 2016b). Rats exposed to aSI also showed lower baseline extracellular dopamine levels in the NAc compared to aGH rats; KOR blockade with norBNI resulted in an increase in the extracellular dopamine levels, matching those in aGH rats, suggesting that augmented KOR function contributes to the low-dopamine state (Karkhanis et al. 2016b). Furthermore, dopamine response to ethanol (1 g/kg, i.p.) was greater in the presence of KOR blockade selectively in aSI rats, indicating that the KOR function was increased following aSI exposure and dampened the dopamine-elevating effects of ethanol at moderate doses (Karkhanis et al. 2016b). In summary, stress-mediated changes in KOR function not only promote anxiety-like and depressive-like behaviors but also alter reward value and motivation, thus increasing vulnerability to develop addictive behaviors, particularly drug consumption.
5. Conclusion
The literature reviewed here support that the dynorphin/KOR system plays a primary role in stress responsivity and shapes drug-seeking behaviors at various stages of the drug addiction cycle, partially through modulation of the mesolimbic dopamine system. Exposure to stress is a well-recognized risk factor for exacerbation of drug use and relapse during abstinence. Repeated cycles of drug exposure and withdrawal also induce stress, further affecting KOR signaling and dopamine transmission. The interaction between the two systems shapes the overall behavioral outcome and reward perception. For example, prior stress exposure promotes a hypodopaminergic state, characterized by low tonic levels of dopamine; KOR hyperfunctioning influences this state, as KOR activation leads to inhibition of dopamine release and KOR inhibition mediated reversal of low extracellular levels of dopamine. The augmented KOR function also affects perception of reward value by its influence on the mesolimbic dopamine signaling. For example, chronic stress and drug exposure produce an anhedonic state, driven by high KOR function in combination with low synaptic levels of dopamine; subsequent exposure to drugs during this state exacerbates the reinforcing value of the drug mostly due to larger changes in dopamine concentrations. Blockade of KOR signaling reverses anhedonia by elevating synaptic levels of dopamine and thus changing perceived reinforcing value of drugs. ln summary, the development of addictive behaviors and negative affect occur as a result of an orchestrated interaction between the dynorphin/KOR and dopamine systems.
Contributor Information
Paige M. Estave, Department of Physiology and Pharmacology, Wake Forest School of Medicine, Winston-Salem, NC, USA
Mary B. Spodnick, Department of Psychology, Developmental Exposure Alcohol Research Center, Center for Developmental and Behavioral Neuroscience, Binghamton University – SUNY, Binghamton, NY, USA
Anushree N. Karkhanis, Department of Psychology, Developmental Exposure Alcohol Research Center, Center for Developmental and Behavioral Neuroscience, Binghamton University – SUNY, Binghamton, NY, USA
References
- Abraham AD, Schattauer SS, Reichard KL, Cohen JH, Fontaine HM, Song AJ, Johnson SD, Land BB, Chavkin C (2018) Estrogen regulation of GRK2 inactivates kappa opioid receptor signaling mediating analgesia, but not aversion. J Neurosci 38:8031–8043 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Acri JB, Thompson AC, Shippenberg T (2001) Modulation of pre- and postsynaptic dopamine D2 receptor function by the selective kappa-opioid receptor agonist U69593. Synapse 39:343–350 [DOI] [PubMed] [Google Scholar]
- Alavian KN, Scholz C, Simon HH (2008) Transcriptional regulation of mesencephalic dopaminergic neurons: the full circle of life and death. Mov Disord 23:319–328 [DOI] [PubMed] [Google Scholar]
- Al-Hasani R, McCall JG, Shin G, Gomez AM, Schmitz GP, Bernardi JM, Pyo C-O, Park SI, Marcinkiewcz CM, Crowley NA, Krashes MJ, Lowell BB, Kash TL, Rogers JA, Bruchas MR (2015) Distinct subpopulations of nucleus accumbens dynorphin neurons drive aversion and reward. Neuron 87:1063–1077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson RI, Agoglia AE, Morales M, Varlinskaya EI, Spear LP (2013) Stress, kappa manipulations, and aversive effects of ethanol in adolescent and adult male rats. Neuroscience 249:214–222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson RI, Lopez MF, Becker HC (2016) Stress-induced enhancement of ethanol intake in C57BL/6J mice with a history of chronic ethanol exposure: involvement of kappa opioid receptors. Front Cell Neurosci 10:45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson RI, Lopez MF, Griffin WC, Haun HL, Bloodgood DW, Pati D, Boyt KM, Kash TL, Becker HC (2019) Dynorphin-kappa opioid receptor activity in the central amygdala modulates binge-like alcohol drinking in mice. Neuropsychopharmacology 44:1084–1092 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashenhurst JR, Bujarski S, Ray LA (2012) Delta and kappa opioid receptor polymorphisms influence the effects of naltrexone on subjective responses to alcohol. Pharmacol Biochem Behav 103:253–259 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azocar VH, Sepúlveda G, Ruiz C, Aguilera C, Andrés ME, Fuentealba JA (2019) The blocking of kappa-opioid receptor reverses the changes in dorsolateral striatum dopamine dynamics during the amphetamine sensitization. J Neurochem 148:348–358 [DOI] [PubMed] [Google Scholar]
- Bals-Kubik R, Herz A, Shippenberg TS (1989) Evidence that the aversive effects of opioid antagonists and κ-agonists are centrally mediated. Psychopharmacology 98:203–206 [DOI] [PubMed] [Google Scholar]
- Bart G, Schluger JH, Borg L, Ho A, Bidlack JM, Kreek MJ (2005) Nalmefene induced elevation in serum prolactin in normal human volunteers: partial kappa opioid agonist activity? Neuropsychopharmacology 30:2254–2262 [DOI] [PubMed] [Google Scholar]
- Beardsley PM, Howard JL, Shelton KL, Carroll FI (2005) Differential effects of the novel kappa opioid receptor antagonist, JDTic, on reinstatement of cocaine-seeking induced by footshock stressors vs cocaine primes and its antidepressant-like effects in rats. Psychopharmacology 183:118–126 [DOI] [PubMed] [Google Scholar]
- Bérubé P, Laforest S, Bhatnagar S, Drolet G (2013) Enkephalin and dynorphin mRNA expression are associated with resilience or vulnerability to chronic social defeat stress. Physiol Behav 122:237–245 [DOI] [PubMed] [Google Scholar]
- Bruchas MR, Land BB, Aita M, Xu M, Barot SK, Li S, Chavkin C (2007a) Stress-induced p38 mitogen-activated protein kinase activation mediates κ-opioid-dependent dysphoria. J Neurosci 27:11614–11623 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruchas MR, Yang T, Schreiber S, Defino M, Kwan SC, Li S, Chavkin C (2007b) Long-acting kappa opioid antagonists disrupt receptor signaling and produce noncompetitive effects by activating c-Jun N-terminal kinase. J Biol Chem 282:29803–29811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruchas MR, Land BB, Chavkin C (2010) The dynorphin/kappa opioid system as a modulator of stress-induced and pro-addictive behaviors. Brain Res 1314:44–55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buda JJ, Carroll FI, Kosten TR, Swearingen D, Walters BB (2015) A double-blind, placebo-controlled trial to evaluate the safety, tolerability, and pharmacokinetics of single, escalating oral doses of JDTic. Neuropsychopharmacology 40:2059–2065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cabib S, Puglisi-Allegra S (1991) Genotype-dependent effects of chronic stress on apomorphine-induced alterations of striatal and mesolimbic dopamine metabolism. Brain Res 542:91–96 [DOI] [PubMed] [Google Scholar]
- Calipari ES, Ferris MJ, Zimmer BA, Roberts DCS, Jones SR (2013) Temporal pattern of cocaine intake determines tolerance vs sensitization of cocaine effects at the dopamine transporter. Neuropsychopharmacology 38:2385–2392 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carey AN, Borozny K, Aldrich JV, McLaughlin JP (2007) Reinstatement of cocaine place-conditioning prevented by the peptide kappa-opioid receptor antagonist arodyn. Eur J Pharmacol 569:84–89 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlezon WA, Béguin C, DiNieri JA, Baumann MH, Richards MR, Todtenkopf MS, Rothman RB, Ma Z, Lee DY-W, Cohen BM (2006) Depressive-like effects of the kappa-opioid receptor agonist salvinorin A on behavior and neurochemistry in rats. J Pharmacol Exp Ther 316:440–447 [DOI] [PubMed] [Google Scholar]
- Carr GV, Lucki I (2010) Comparison of the kappa-opioid receptor antagonist DIPPA in tests of anxiety-like behavior between Wistar Kyoto and Sprague Dawley rats. Psychopharmacology 210:295–302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carr GV, Bangasser DA, Bethea T, Young M, Valentino RJ, Lucki I (2010) Antidepressant-like effects of κ-opioid receptor antagonists in Wistar Kyoto rats. Neuropsychopharmacology 35:752–763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castro DC, Berridge KC (2014) Opioid hedonic hotspot in nucleus accumbens shell: mu, delta, and kappa maps for enhancement of sweetness “liking” and “wanting”. J Neurosci 34:4239–4250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chartoff EH, Mavrikaki M (2015) Sex differences in kappa opioid receptor function and their potential impact on addiction. Front Neurosci 9. 10.3389/fnins.2015.00466/full. Accessed 25 July 2020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chartoff EH, Ebner SR, Sparrow A, Potter D, Baker PM, Ragozzino ME, Roitman MF (2016) Relative timing between kappa opioid receptor activation and cocaine determines the impact on reward and dopamine release. Neuropsychopharmacology 41:989–1002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chavkin C, Koob GF (2016) Dynorphin, dysphoria, and dependence: the stress of addiction. Neuropsychopharmacology 41:373–374 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chefer VI, Zapata A, Shippenberg TS, Bungay PM (2006) Quantitative no-net-flux microdialysis permits detection of increases and decreases in dopamine uptake in mouse nucleus accumbens. J Neurosci Methods 155:187–193 [DOI] [PubMed] [Google Scholar]
- Conway SM, Puttick D, Russell S, Potter D, Roitman MF, Chartoff EH (2019) Females are less sensitive than males to the motivational- and dopamine-suppressing effects of kappa opioid receptor activation. Neuropharmacology 146:231–241 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daunais JB, Roberts DCS, McGinty JF (1993) Cocaine self-administration increases preprodynorphin, but not c-fos, mRNA in rat striatum. Neuroreport 4:543–546 [DOI] [PubMed] [Google Scholar]
- de Laat B, Goldberg A, Shi J, Tetrault JM, Nabulsi N, Zheng M-Q, Najafzadeh S, Gao H, Kapinos M, Ropchan J, O’Malley SS, Huang Y, Morris ED, Krishnan-Sarin S (2019) The kappa opioid receptor is associated with naltrexone-induced reduction of drinking and craving. Biol Psychiatry 86:864–871 [DOI] [PubMed] [Google Scholar]
- de Laat B, Nabulsi N, Huang Y, O’Malley SS, Froehlich JC, Morris ED, Krishnan-Sarin S (2020) Occupancy of the kappa opioid receptor by naltrexone predicts reduction in drinking and craving. Mol Psychiatry [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Vries TJ, Hogenboom F, Mulder AH, Schoffelmeer AN (1990) Ontogeny of mu-, delta- and kappa-opioid receptors mediating inhibition of neurotransmitter release and adenylate cyclase activity in rat brain. Brain Res Dev Brain Res 54:63–69 [DOI] [PubMed] [Google Scholar]
- Detke MJ, Rickels M, Lucki I (1995) Active behaviors in the rat forced swimming test differentially produced by serotonergic and noradrenergic antidepressants. Psychopharmacology 121:66–72 [DOI] [PubMed] [Google Scholar]
- Dhawan BN, Cesselin F, Raghubir R, Reisine T, Bradley PB, Portoghese PS, Hamon M (1996) International Union of Pharmacology. XII. Classification of opioid receptors. Pharmacol Rev 48:567–592 [PubMed] [Google Scholar]
- Di Chiara G, Imperato A (1988) Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc Natl Acad Sci U S A 85:5274–5278 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding Y, Kang L, Li B, Ma L (2005) Enhanced cocaine self-administration in adult rats with adolescent isolation experience. Pharmacol Biochem Behav 82:673–677 [DOI] [PubMed] [Google Scholar]
- Domi E, Barbier E, Augier E, Augier G, Gehlert D, Barchiesi R, Thorsell A, Holm L, Heilig M (2018) Preclinical evaluation of the kappa-opioid receptor antagonist CERC-501 as a candidate therapeutic for alcohol use disorders. Neuropsychopharmacology 43:1805–1812 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donahue RJ, Landino SM, Golden SA, Carroll FI, Russo SJ, Carlezon WA (2015) Effects of acute and chronic social defeat stress are differentially mediated by the dynorphin/kappa-opioid receptor system. Behav Pharmacol 26:654–663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebner SR, Roitman MF, Potter DN, Rachlin AB, Chartoff EH (2010) Depressive-like effects of the kappa opioid receptor agonist salvinorin A are associated with decreased phasic dopamine release in the nucleus accumbens. Psychopharmacology 210:241–252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehrich JM, Phillips PEM, Chavkin C (2014) Kappa opioid receptor activation potentiates the cocaine-induced increase in evoked dopamine release recorded in vivo in the mouse nucleus accumbens. Neuropsychopharmacology 39:3036–3048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehrich JM, Messinger DI, Knakal CR, Kuhar JR, Schattauer SS, Bruchas MR, Zweifel LS, Kieffer BL, Phillips PEM, Chavkin C (2015) Kappa opioid receptor-induced aversion requires p38 MAPK activation in VTA dopamine neurons. J Neurosci 35:12917–12931 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Engber TM, Boldry RC, Kuo S, Chase TN (1992) Dopaminergic modulation of striatal neuropeptides: differential effects of D1 and D2 receptor stimulation on somatostatin, neuropeptide Y, neurotensin, dynorphin and enkephalin. Brain Res 581:261–268 [DOI] [PubMed] [Google Scholar]
- Escobar AP, González MP, Meza RC, Noches V, Henny P, Gysling K, España RA, Fuentealba JA, Andrés ME (2017) Mechanisms of kappa opioid receptor potentiation of dopamine D2 receptor function in quinpirole-induced locomotor sensitization in rats. Int J Neuropsychopharmacol 20:660–669 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fairbanks J, Umbreit A, Kolla BP, Karpyak VM, Schneekloth TD, Loukianova LL, Sinha S (2020) Evidence-based pharmacotherapies for alcohol use disorder: clinical pearls. Mayo Clin Proc 95:1964–1977 [DOI] [PubMed] [Google Scholar]
- Fosnocht AQ, Lucerne KE, Ellis AS, Olimpo NA, Briand LA (2019) Adolescent social isolation increases cocaine seeking in male and female mice. Behav Brain Res 359:589–596 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuentealba JA, Gysling K, Magendzo K, Andrés ME (2006) Repeated administration of the selective kappa-opioid receptor agonist U-69593 increases stimulated dopamine extracellular levels in the rat nucleus accumbens. J Neurosci Res 84:450–459 [DOI] [PubMed] [Google Scholar]
- Gehrke BJ, Chefer VI, Shippenberg TS (2008) Effects of acute and repeated administration of salvinorin A on dopamine function in the rat dorsal striatum. Psychopharmacology 197:509–517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Georges F, Normand E, Bloch B, Le Moine C (1998) Opioid receptor gene expression in the rat brain during ontogeny, with special reference to the mesostriatal system: an in situ hybridization study. Dev Brain Res 109:187–199 [DOI] [PubMed] [Google Scholar]
- Gerra G, Leonardi C, Cortese E, D’Amore A, Lucchini A, Strepparola G, Serio G, Farina G, Magnelli F, Zaimovic A, Mancini A, Turci M, Manfredini M, Donnini C (2007) Human kappa opioid receptor gene (OPRK1) polymorphism is associated with opiate addiction. Am J Med Genet B Neuropsychiatr Genet 144B:771–775 [DOI] [PubMed] [Google Scholar]
- Grilli M, Neri E, Zappettini S, Massa F, Bisio A, Romussi G, Marchi M, Pittaluga A (2009) Salvinorin A exerts opposite presynaptic controls on neurotransmitter exocytosis from mouse brain nerve terminals. Neuropharmacology 57:523–530 [DOI] [PubMed] [Google Scholar]
- Groblewski PA, Zietz C, Willuhn I, Phillips PEM, Chavkin C (2015) Repeated stress exposure causes strain-dependent shifts in the behavioral economics of cocaine in rats. Addict Biol 20:297–301 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hang A, Wang Y, He L, Liu J (2015) The role of the dynorphin/κ opioid receptor system in anxiety. Acta Pharmacol Sin 36:783–790 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haun HL, Griffin WC, Lopez MF, Becker HC (2020) Kappa opioid receptors in the bed nucleus of the stria terminalis regulate binge-like alcohol consumption in male and female mice. Neuropharmacology 167:107984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heijna MH, Padt M, Hogenboom F, Portoghese PS, Mulder AH, Schoffelmeer AN (1990) Opioid receptor-mediated inhibition of dopamine and acetylcholine release from slices of rat nucleus accumbens, olfactory tubercle and frontal cortex. Eur J Pharmacol 181:267–278 [DOI] [PubMed] [Google Scholar]
- Hill MP, Brotchie JM (1995) Modulation of glutamate release by a kappa-opioid receptor agonist in rodent and primate striatum. Eur J Pharmacol 281:R1–R2 [DOI] [PubMed] [Google Scholar]
- Hill MP, Brotchie JM (1999) Control of glutamate release by calcium channels and kappa-opioid receptors in rodent and primate striatum. Br J Pharmacol 127:275–283 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman AF, Spivak CE, Lupica CR (2016) Enhanced dopamine release by dopamine transport inhibitors described by a restricted diffusion model and fast-scan cyclic voltammetry. ACS Chem Neurosci 7:700–709 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holly EN, DeBold JF, Miczek KA (2015) Increased mesocorticolimbic dopamine during acute and repeated social defeat stress: modulation by corticotropin releasing factor receptors in the ventral tegmental area. Psychopharmacology 232:4469–4479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horan P, Taylor J, Yamamura HI, Porreca F (1992) Extremely long-lasting antagonistic actions of nor-binaltorphimine (nor-BNI) in the mouse tail-flick test. J Pharmacol Exp Ther 260:1237–1243 [PubMed] [Google Scholar]
- Huang P, Tunis J, Parry C, Tallarida R, Liu-Chen L-Y (2016) Synergistic antidepressant-like effects between a kappa opioid antagonist (LY2444296) and a delta opioid agonist (ADL5859) in the mouse forced swim test. Eur J Pharmacol 781:53–59 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hurd YL, Herkenham M (1993) Molecular alterations in the neostriatum of human cocaine addicts. Synapse 13:357–369 [DOI] [PubMed] [Google Scholar]
- Izenwasser S, Acri JB, Kunko PM, Shippenberg T (1998) Repeated treatment with the selective kappa opioid agonist U-69593 produces a marked depletion of dopamine D2 receptors. Synapse 30:275–283 [DOI] [PubMed] [Google Scholar]
- Jackson KJ, McLaughlin JP, Carroll FI, Damaj MI (2013) Effects of the kappa opioid receptor antagonist, norbinaltorphimine, on stress and drug-induced reinstatement of nicotine-conditioned place preference in mice. Psychopharmacology (Berl) 226. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3821839/. Accessed 9 Sept 2020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang HK, Wang JY (1991) The effects of dopamine D-1 and D-2 receptor subtype agonists on nigrostriatal opioid dynorphin and enkephalin immunostaining in 6-hydroxydopamine lesioned rats. Chin J Physiol 34:413–425 [PubMed] [Google Scholar]
- Kalivas PW, Duffy P (1995) Selective activation of dopamine transmission in the shell of the nucleus accumbens by stress. Brain Res 675:325–328 [DOI] [PubMed] [Google Scholar]
- Karkhanis AN, Locke JL, McCool BA, Weiner JL, Jones SR (2014) Social isolation rearing increases nucleus accumbens dopamine and norepinephrine responses to acute ethanol in adulthood. Alcohol Clin Exp Res 38:2770–2779 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karkhanis AN, Huggins KN, Rose JH, Jones SR (2016a) Switch from excitatory to inhibitory actions of ethanol on dopamine levels after chronic exposure: role of kappa opioid receptors. Neuropharmacology 110:190–197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karkhanis AN, Rose JH, Weiner JL, Jones SR (2016b) Early-life social isolation stress increases kappa opioid receptor responsiveness and downregulates the dopamine system. Neuropsychopharmacology 41:2263–2274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karkhanis A, Holleran KM, Jones SR (2017) Chapter Three - Dynorphin/kappa opioid receptor signaling in preclinical models of alcohol, drug, and food addiction. In: Thiele TE (ed) International review of neurobiology. The role of neuropeptides in addiction and disorders of excessive consumption, pp 53–88. Academic Press. http://www.sciencedirect.com/science/article/pii/S0074774217301393. Accessed 8 Aug 2019 [DOI] [PubMed] [Google Scholar]
- Katoh A, Nabeshima T, Kameyama T (1990) Behavioral changes induced by stressful situations: effects of enkephalins, dynorphin, and their interactions. J Pharmacol Exp Ther 253:600–607 [PubMed] [Google Scholar]
- Katunar MR, Saez T, Brusco A, Antonelli MC (2009) Immunocytochemical expression of dopamine-related transcription factors Pitx3 and Nurr1 in prenatally stressed adult rats. J Neurosci Res 87:1014–1022 [DOI] [PubMed] [Google Scholar]
- Kessler RC (1997) The effects of stressful life events on depression. Annu Rev Psychol 48:191–214 [DOI] [PubMed] [Google Scholar]
- Kivell B, Uzelac Z, Sundaramurthy S, Rajamanickam J, Ewald A, Chefer V, Jaligam V, Bolan E, Simonson B, Annamalai B, Mannangatti P, Prisinzano TE, Gomes I, Devi LA, Jayanthi LD, Sitte HH, Ramamoorthy S, Shippenberg TS (2014) Salvinorin A regulates dopamine transporter function via a kappa opioid receptor and ERK1/2-dependent mechanism. Neuropharmacology 86:228–240 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knoll AT, Meloni EG, Thomas JB, Carroll FI, Carlezon WA (2007) Anxiolytic-like effects of kappa-opioid receptor antagonists in models of unlearned and learned fear in rats. J Pharmacol Exp Ther 323:838–845 [DOI] [PubMed] [Google Scholar]
- Land BB, Bruchas MR, Lemos JC, Xu M, Melief EJ, Chavkin C (2008) The dysphoric component of stress is encoded by activation of the dynorphin kappa-opioid system. J Neurosci 28:407–414 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lapiz MDS, Fulford A, Muchimapura S, Mason R, Parker T, Marsden CA (2003) Influence of postweaning social isolation in the rat on brain development, conditioned behavior, and neurotransmission. Neurosci Behav Physiol 33:13–29 [DOI] [PubMed] [Google Scholar]
- Li C, Pleil KE, Stamatakis AM, Busan S, Vong L, Lowell BB, Stuber GD, Kash TL (2012) Presynaptic inhibition of GABA release in the BNST by kappa opioid receptor signaling. Biol Psychiatry 71:725–732 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Logrip ML, Janak PH, Ron D (2009) Blockade of ethanol reward by the kappa opioid receptor agonist U50,488H. Alcohol 43:359–365 [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacLean KA, Johnson MW, Reissig CJ, Prisinzano TE, Griffiths RR (2013) Dose-related effects of salvinorin A in humans: dissociative, hallucinogenic, and memory effects. Psychopharmacology 226:381–392 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mague SD, Pliakas AM, Todtenkopf MS, Tomasiewicz HC, Zhang Y, Stevens WC, Jones RM, Portoghese PS, Carlezon WA (2003) Antidepressant-like effects of kappa-opioid receptor antagonists in the forced swim test in rats. J Pharmacol Exp Ther 305:323–330 [DOI] [PubMed] [Google Scholar]
- Margolis EB, Karkhanis AN (2019) Dopaminergic cellular and circuit contributions to kappa opioid receptor mediated aversion. Neurochem Int 129:104504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marinelli M, McCutcheon JE (2014) Heterogeneity of dopamine neuron activity across traits and states. Neuroscience 282:176–197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marinelli PW, Lam M, Bai L, Quirion R, Gianoulakis C (2006) A microdialysis profile of dynorphin A1–8 release in the rat nucleus accumbens following alcohol administration. Alcohol Clin Exp Res 30:982–990 [DOI] [PubMed] [Google Scholar]
- Martinez D, Slifstein M, Matuskey D, Nabulsi N, Zheng M-Q, Lin S, Ropchan J, Urban N, Grassetti A, Chang D, Salling M, Foltin R, Carson RE, Huang Y (2019) Kappa-opioid receptors, dynorphin, and cocaine addiction: a positron emission tomography study. Neuropsychopharmacology 44:1720–1727 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mash DC, Staley JK (1999) D3 dopamine and kappa opioid receptor alterations in human brain of cocaine-overdose victims. Ann N Y Acad Sci 877:507–522 [DOI] [PubMed] [Google Scholar]
- Matsuzawa S, Suzuki T, Misawa M, Nagase H (1999) Different roles of mu-, delta- and kappa-opioid receptors in ethanol-associated place preference in rats exposed to conditioned fear stress. Eur J Pharmacol 368:9–16 [DOI] [PubMed] [Google Scholar]
- McCutcheon JE, Marinelli M (2009) Age matters. Eur J Neurosci 29:997–1014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLaughlin JP, Marton-Popovici M, Chavkin C (2003) Kappa opioid receptor antagonism and prodynorphin gene disruption block stress-induced behavioral responses. J Neurosci 23:5674–5683 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLaughlin JP, Land BB, Li S, Pintar JE, Chavkin C (2006a) Prior activation of kappa opioid receptors by U50,488 mimics repeated forced swim stress to potentiate cocaine place preference conditioning. Neuropsychopharmacology 31:787–794 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLaughlin JP, Li S, Valdez J, Chavkin TA, Chavkin C (2006b) Social defeat stress-induced behavioral responses are mediated by the endogenous kappa opioid system. Neuropsychopharmacology 31:1241–1248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melchior JR, Jones SR (2017) Chronic ethanol exposure increases inhibition of optically targeted phasic dopamine release in the nucleus accumbens core and medial shell ex vivo. Mol Cell Neurosci 85:93–104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melief EJ, Miyatake M, Bruchas MR, Chavkin C (2010) Ligand-directed c-Jun N-terminal kinase activation disrupts opioid receptor signaling. Proc Natl Acad Sci U S A 107:11608–11613 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melief EJ, Miyatake M, Carroll FI, Béguin C, Carlezon WA, Cohen BM, Grimwood S, Mitch CH, Rorick-Kehn L, Chavkin C (2011) Duration of action of a broad range of selective κ-opioid receptor antagonists is positively correlated with c-Jun N-terminal kinase-1 activation. Mol Pharmacol 80:920–929 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menendez L, Andres-Trelles F, Hidalgo A, Baamonde A (1993) Involvement of spinal kappa opioid receptors in a type of footshock induced analgesia in mice. Brain Res 611:264–271 [DOI] [PubMed] [Google Scholar]
- Munro TA, Berry LM, Van’t Veer A, Béguin C, Carroll FI, Zhao Z, Carlezon WA, Cohen BM (2012) Long-acting κ opioid antagonists nor-BNI, GNTI and JDTic: pharmacokinetics in mice and lipophilicity. BMC Pharmacol 12:5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nestler EJ, Barrot M, Self DW (2001) ΔFosB: a sustained molecular switch for addiction. Proc Natl Acad Sci 98:11042–11046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newton SS, Thome J, Wallace TL, Shirayama Y, Schlesinger L, Sakai N, Chen J, Neve R, Nestler EJ, Duman RS (2002) Inhibition of cAMP response element-binding protein or dynorphin in the nucleus accumbens produces an antidepressant-like effect. J Neurosci 22:10883–10890 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nygard SK, Hourguettes NJ, Sobczak GG, Carlezon WA, Bruchas MR (2016) Stress-induced reinstatement of nicotine preference requires dynorphin/kappa opioid activity in the basolateral amygdala. J Neurosci 36:9937–9948 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Page S, Mavrikaki MM, Lintz T, Puttick D, Roberts E, Rosen H, Carroll FI, Carlezon WA, Chartoff EH (2019) Behavioral pharmacology of novel kappa opioid receptor antagonists in rats. Int J Neuropsychopharmacol 22:735–745 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pande AC, Pyke RE, Greiner M, Wideman GL, Benjamin R, Pierce MW (1996) Analgesic efficacy of enadoline versus placebo or morphine in postsurgical pain. Clin Neuropharmacol 19:451–456 [DOI] [PubMed] [Google Scholar]
- Pani L, Porcella A, Gessa GL (2000) The role of stress in the pathophysiology of the dopaminergic system. Mol Psychiatry 5:14–21 [DOI] [PubMed] [Google Scholar]
- Perreault ML, Graham D, Bisnaire L, Simms J, Hayton S, Szechtman H (2006) Kappa-opioid agonist U69593 potentiates locomotor sensitization to the D2/D3 agonist quinpirole: pre- and postsynaptic mechanisms. Neuropsychopharmacology 31:1967–1981 [DOI] [PubMed] [Google Scholar]
- Perreault ML, Graham D, Scattolon S, Wang Y, Szechtman H, Foster JA (2007) Cotreatment with the kappa opioid agonist U69593 enhances locomotor sensitization to the D2/D3 dopamine agonist quinpirole and alters dopamine D2 receptor and prodynorphin mRNA expression in rats. Psychopharmacology 194:485–496 [DOI] [PubMed] [Google Scholar]
- Pfeiffer A, Brantl V, Herz A, Emrich HM (1986) Psychotomimesis mediated by kappa opiate receptors. Science 233:774–776 [DOI] [PubMed] [Google Scholar]
- Piras AP, Zhou Y, Schlussman SD, Ho A, Kreek MJ (2010) Acute withdrawal from chronic escalating-dose binge cocaine administration alters kappa opioid receptor stimulation of [35S] GTPγS binding in the rat ventral tegmental area. Neuroscience 169:751–757 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pirino B, Spodnick M, Gargiulo A, Curtis G, Barson J, Karkhanis A (2020) Kappa-opioid receptor-dependent changes in dopamine and anxiety-like or approach-avoidance behavior occur differentially across the nucleus accumbens shell rostro-caudal axis. Neuropharmacology 181:108341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polter AM, Bishop RA, Briand LA, Graziane NM, Pierce RC, Kauer JA (2014) Poststress block of kappa opioid receptors rescues long-term potentiation of inhibitory synapses and prevents reinstatement of cocaine seeking. Biol Psychiatry 76:785–793 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polter AM, Barcomb K, Chen RW, Dingess PM, Graziane NM, Brown TE, Kauer JA (2017) Constitutive activation of kappa opioid receptors at ventral tegmental area inhibitory synapses following acute stress. elife 6:e23785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porsolt RD, Le Pichon M, Jalfre M (1977) Depression: a new animal model sensitive to antidepressant treatments. Nature 266:730–732 [DOI] [PubMed] [Google Scholar]
- Potter DN, Damez-Werno D, Carlezon WA, Cohen BM, Chartoff EH (2011) Repeated exposure to the κ-opioid receptor agonist salvinorin A modulates extracellular signal-regulated kinase and reward sensitivity. Biol Psychiatry 70:744–753 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prakash N, Wurst W (2006) Genetic networks controlling the development of midbrain dopaminergic neurons. J Physiol 575:403–410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Redila VA, Chavkin C (2008) Stress-induced reinstatement of cocaine seeking is mediated by the kappa opioid system. Psychopharmacology 200:59–70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reed B, Butelman ER, Fry RS, Kimani R, Kreek MJ (2018) Repeated administration of opra kappa (LY2456302), a novel, short-acting, selective KOP-r antagonist, in persons with and without cocaine dependence. Neuropsychopharmacology 43:739–750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rose JH, Karkhanis AN, Chen R, Gioia D, Lopez MF, Becker HC, McCool BA, Jones SR (2016a) Supersensitive kappa opioid receptors promotes ethanol withdrawal-related behaviors and reduce dopamine signaling in the nucleus accumbens. Int J Neuropsychopharmacol 19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rose JH, Karkhanis AN, Steiniger-Brach B, Jones SR (2016b) Distinct effects of nalmefene on dopamine uptake rates and kappa opioid receptor activity in the nucleus accumbens following chronic intermittent ethanol exposure. Int J Mol Sci 17:1216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rougé-Pont F, Piazza PV, Kharouby M, Le Moal M, Simon H (1993) Higher and longer stress-induced increase in dopamine concentrations in the nucleus accumbens of animals predisposed to amphetamine self-administration. A microdialysis study. Brain Res 602:169–174 [DOI] [PubMed] [Google Scholar]
- Russell SE, Rachlin AB, Smith KL, Muschamp J, Berry L, Zhao Z, Chartoff EH (2014) Sex differences in sensitivity to the depressive-like effects of the kappa opioid receptor agonist U-50488 in rats. Biol Psychiatry 76:213–222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schattauer SS, Land BB, Reichard KL, Abraham AD, Burgeno LM, Kuhar JR, Phillips PEM, Ong SE, Chavkin C (2017) Peroxiredoxin 6 mediates Gαi protein-coupled receptor inactivation by cJun kinase. Nat Commun 8. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5622097/. Accessed 30 Sept 2020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheggi S, Leggio B, Masi F, Grappi S, Gambarana C, Nanni G, Rauggi R, De Montis MG (2002) Selective modifications in the nucleus accumbens of dopamine synaptic transmission in rats exposed to chronic stress. J Neurochem 83:895–903 [DOI] [PubMed] [Google Scholar]
- Schlussman SD, Zhang Y, Yuferov V, LaForge KS, Ho A, Kreek MJ (2003) Acute “binge” cocaine administration elevates dynorphin mRNA in the caudate putamen of C57BL/6J but not 129/J mice. Brain Res 974:249–253 [DOI] [PubMed] [Google Scholar]
- Schlussman SD, Zhou Y, Bailey A, Ho A, Kreek MJ (2005) Steady-dose and escalating-dose “binge” administration of cocaine alter expression of behavioral stereotypy and striatal preprodynorphin mRNA levels in rats. Brain Res Bull 67:169–175 [DOI] [PubMed] [Google Scholar]
- Sedki F, Eigenmann K, Gelinas J, Schouela N, Courchesne S, Shalev U (2015) A role for kappa-, but not mu-opioid, receptor activation in acute food deprivation-induced reinstatement of heroin seeking in rats. Addict Biol 20:423–432 [DOI] [PubMed] [Google Scholar]
- Shippenberg TS, Herz A (1986) Differential effects of mu and kappa opioid systems on motivational processes. NIDA Res Monogr 75:563–566 [PubMed] [Google Scholar]
- Shippenberg TS, Zapata A, Chefer VI (2007) Dynorphin and the pathophysiology of drug addiction. Pharmacol Ther 116:306–321 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shirayama Y, Ishida H, Iwata M, Hazama G-I, Kawahara R, Duman RS (2004) Stress increases dynorphin immunoreactivity in limbic brain regions and dynorphin antagonism produces antidepressant-like effects. J Neurochem 90:1258–1268 [DOI] [PubMed] [Google Scholar]
- Siciliano CA, Jones SR (2017) Cocaine potency at the dopamine transporter tracks discrete motivational states during cocaine self-administration. Neuropsychopharmacology 42:1893–1904 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siciliano CA, Calipari ES, Yorgason JT, Lovinger DM, Mateo Y, Jimenez VA, Helms CM, Grant KA, Jones SR (2016) Increased presynaptic regulation of dopamine neurotransmission in the nucleus accumbens core following chronic ethanol self-administration in female macaques. Psychopharmacology 233:1435–1443 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinha R (2007) The role of stress in addiction relapse. Curr Psychiatry Rep 9:388–395 [DOI] [PubMed] [Google Scholar]
- Sivam SP (1989) Cocaine selectively increases striatonigral dynorphin levels by a dopaminergic mechanism. J Pharmacol Exp Ther 250:818–824 [PubMed] [Google Scholar]
- Skelly MJ, Chappell AE, Carter E, Weiner JL (2015) Adolescent social isolation increases anxiety-like behavior and ethanol intake and impairs fear extinction in adulthood: possible role of disrupted noradrenergic signaling. Neuropharmacology 97:149–159 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smiley PL, Johnson M, Bush L, Gibb JW, Hanson GR (1990) Effects of cocaine on extrapyramidal and limbic dynorphin systems. J Pharmacol Exp Ther 253:938–943 [PubMed] [Google Scholar]
- Spanagel R, Herz A, Shippenberg TS (1992) Opposing tonically active endogenous opioid systems modulate the mesolimbic dopaminergic pathway. Proc Natl Acad Sci 89:2046–2050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spangler R, Unterwald EM, Kreek MJ (1993) “Binge” cocaine administration induces a sustained increase of prodynorphin mRNA in rat caudate-putamen. Brain Res Mol Brain Res 19:323–327 [DOI] [PubMed] [Google Scholar]
- Spangler R, Ho A, Zhou Y, Maggos CE, Yuferov V, Kreek MJ (1996) Regulation of kappa opioid receptor mRNA in the rat brain by ‘binge’ pattern cocaine administration and correlation with preprodynorphin mRNA. Mol Brain Res 38:71–76 [DOI] [PubMed] [Google Scholar]
- Sperling RE, Gomes SM, Sypek EI, Carey AN, McLaughlin JP (2010) Endogenous kappa-opioid mediation of stress-induced potentiation of ethanol-conditioned place preference and self-administration. Psychopharmacology 210:199–209 [DOI] [PubMed] [Google Scholar]
- Spodnick MB, Amirault RT, Towner TT, Varlinskaya EI, Spear LP, Karkhanis AN (2020) Adolescent intermittent ethanol exposure effects on kappa opioid receptor mediated dopamine transmission: sex and age of exposure matter Brain Sci 10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Staley JK, Rothman RB, Rice KC, Partilla J, Mash DC (1997) Kappa2 opioid receptors in limbic areas of the human brain are upregulated by cocaine in fatal overdose victims. J Neurosci 17:8225–8233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun H, Luessen DJ, Kind KO, Zhang K, Chen R (2020) Cocaine self-administration regulates transcription of opioid peptide precursors and opioid receptors in rat caudate putamen and prefrontal cortex. Neuroscience 443:131–139 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svingos AL, Chavkin C, Colago EE, Pickel VM (2001) Major coexpression of kappa-opioid receptors and the dopamine transporter in nucleus accumbens axonal profiles. Synapse, 42 (3):185–192 [DOI] [PubMed] [Google Scholar]
- Takahashi M, Senda T, Tokuyama S, Kaneto H (1990) Further evidence for the implication of a kappa-opioid receptor mechanism in the production of psychological stress-induced analgesia. Jpn J Pharmacol 53:487–494 [DOI] [PubMed] [Google Scholar]
- Tao R, Auerbach SB (2002) Opioid receptor subtypes differentially modulate serotonin efflux in the rat central nervous system. J Pharmacol Exp Ther 303:549–556 [DOI] [PubMed] [Google Scholar]
- Thompson AC, Zapata A, Justice JB, Vaughan RA, Sharpe LG, Shippenberg TS (2000) Kappa-opioid receptor activation modifies dopamine uptake in the nucleus accumbens and opposes the effects of cocaine. J Neurosci 20:9333–9340 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todtenkopf MS, Marcus JF, Portoghese PS, Carlezon WA (2004) Effects of kappa-opioid receptor ligands on intracranial self-stimulation in rats. Psychopharmacology 172:463–470 [DOI] [PubMed] [Google Scholar]
- Tzaferis JA, McGinty JF (2001) Kappa opioid receptor stimulation decreases amphetamine-induced behavior and neuropeptide mRNA expression in the striatum. Brain Res Mol Brain Res 93:27–35 [DOI] [PubMed] [Google Scholar]
- Unterwald EM, Rubenfeld JM, Kreek MJ (1994) Repeated cocaine administration upregulates kappa and mu, but not delta, opioid receptors. Neuroreport 5:1613–1616 [DOI] [PubMed] [Google Scholar]
- Valenza M, Windisch KA, Butelman ER, Reed B, Kreek MJ (2020) Effects of kappa opioid receptor blockade by LY2444296 HCl, a selective short-acting antagonist, during chronic extended access cocaine self-administration and re-exposure in rat. Psychopharmacology 237:1147–1160 [DOI] [PubMed] [Google Scholar]
- Van’t Veer A, Carlezon WA (2013) Role of kappa-opioid receptors in stress and anxiety-related behavior. Psychopharmacology 229:435–452 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varlinskaya EI, Spear LP, Diaz MR (2018) Stress alters social behavior and sensitivity to pharmacological activation of kappa opioid receptors in an age-specific manner in Sprague Dawley rats. Neurobiol Stress 9:124–132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vijay A, Wang S, Worhunsky P, Zheng M-Q, Nabulsi N, Ropchan J, Krishnan-Sarin S, Huang Y, Morris ED (2016) PET imaging reveals sex differences in kappa opioid receptor availability in humans, in vivo. Am J Nucl Med Mol Imaging 6:205–214 [PMC free article] [PubMed] [Google Scholar]
- Wadenberg M-LG (2003) A review of the properties of spiradoline: a potent and selective kappa-opioid receptor agonist. CNS Drug Rev 9:187–198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watkins LR, Wiertelak EP, Maier SF (1992) Kappa opiate receptors mediate tail-shock induced antinociception at spinal levels. Brain Res 582:1–9 [DOI] [PubMed] [Google Scholar]
- Wee S, Koob GF (2010) The role of the dynorphin-kappa opioid system in the reinforcing effects of drugs of abuse. Psychopharmacology 210:121–135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wee S, Orio L, Ghirmai S, Cashman JR, Koob GF (2009) Inhibition of kappa opioid receptors attenuated increased cocaine intake in rats with extended access to cocaine. Psychopharmacology 205:565–575 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams AV, Laman-Maharg A, Armstrong CV, Ramos-Maciel S, Minie VA, Trainor BC (2018) Acute inhibition of kappa opioid receptors before stress blocks depression-like behaviors in California mice. Prog Neuro-Psychopharmacol Biol Psychiatry 86:166–174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu K, Seo D, Hodgkinson C, Hu Y, Goldman D, Sinha R (2013) A variant on the kappa opioid receptor gene (OPRK1) is associated with stress response and related drug craving, limbic brain activation and cocaine relapse risk. Transl Psychiatry 3:e292–e292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xuei X, Dick D, Flury-Wetherill L, Tian H-J, Agrawal A, Bierut L, Goate A, Bucholz K, Schuckit M, Nurnberger J, Tischfield J, Kuperman S, Porjesz B, Begleiter H, Foroud T, Edenberg HJ (2006) Association of the κ-opioid system with alcohol dependence. Mol Psychiatry 11:1016–1024 [DOI] [PubMed] [Google Scholar]
- Yim HJ, Gonzales RA (2000) Ethanol-induced increases in dopamine extracellular concentration in rat nucleus accumbens are accounted for by increased release and not uptake inhibition. Alcohol 22:107–115 [DOI] [PubMed] [Google Scholar]
- You ZB, Herrera-Marschitz M, Nylander I, Goiny M, O’Connor WT, Ungerstedt U, Terenius L (1994) The striatonigral dynorphin pathway of the rat studied with in vivo microdialysis--II. Effects of dopamine D1 and D2 receptor agonists. Neuroscience 63:427–434 [DOI] [PubMed] [Google Scholar]
- Zhang Y, Butelman ER, Schlussman SD, Ho A, Kreek MJ (2004) Effect of the kappa opioid agonist R-84760 on cocaine-induced increases in striatal dopamine levels and cocaine-induced place preference in C57BL/6J mice. Psychopharmacology 173:146–152 [DOI] [PubMed] [Google Scholar]