

Abstract
Chemotherapy-induced cognitive impairment (CICI) is often reported as a neurotoxic side effect of chemotherapy. Although CICI has emerged as a significant medical problem, meaningful treatments are not currently available due to a lack of mechanistic understanding underlying CICI pathophysiology. Using the platinum-based chemotherapy cisplatin as a model for CICI, we show here that cisplatin suppresses nicotinamide adenine dinucleotide (NAD+) levels in the adult female mouse brain in vivo and in human cortical neurons derived from induced pluripotent stem cells in vitro. Increasing NAD+ levels through nicotinamide mononucleotide (NMN) administration prevented cisplatin-induced abnormalities in neural progenitor proliferation, neuronal morphogenesis, and cognitive function without affecting tumor growth and anti-tumor efficacy of cisplatin. Mechanistically, cisplatin inhibited expression of the NAD+ biosynthesis rate-limiting enzyme nicotinamide phosphoribosyl transferase (Nampt). Selective restoration of Nampt expression in adult-born neurons was sufficient to prevent cisplatin-induced defects in dendrite morphogenesis and memory function. Taken together, our findings suggest that aberrant Nampt-mediated NAD+ metabolic pathways may be a key contributor in cisplatin-induced neurogenic impairments, thus causally leading to memory dysfunction. Therefore, increasing NAD+ levels could represent a promising and safe therapeutic strategy for cisplatin-related neurotoxicity.
Keywords: Cisplatin, cognitive impairment, Nampt, NAD+, adult hippocampal neurogenesis
Introduction
Chemotherapy-induced cognitive impairment (CICI) comprises adverse neurotoxic effects that consists of subtle to moderate impairments in several cognitive domains, including working memory, processing speed and executive functioning. While this symptomatology may be derived from exacerbated stress, anxiety and depression stemming from cancer diagnosis, these cognitive dysregulation (also called “chemobrain” or “chemofog”) can persist for months or years following chemotherapy. While CICI has been reported in up to 75% of patients treated with chemotherapy, a subset of survivors (15%–25%) experience measurably persistent cognitive decline, thus representing a significant public health concern (1). However, meaningful treatments are not currently available due to a lack of mechanistic understanding underlying CICI pathophysiology.
Along with the prefrontal cortex, white matter and other brain structures, the hippocampus is a brain region important for memory formation that is significantly affected by CICI. Notably, the hippocampus is one of a few brain regions where adult-born neurons derived from neural stem/progenitor cells (NPCs) are constantly generated throughout life in most mammals, including humans (2,3). This process, called adult hippocampal neurogenesis, represents a remarkable example of brain plasticity that plays a pivotal role for healthy learning and memory function (4,5). Emerging evidence shows that NPCs are particularly vulnerable to the neurotoxic effects of chemotherapy, which results in significant reductions of neurogenesis (6–8), and may contribute to CICI (9). However, the molecular pathways that underlie CICI are still largely unknown.
Proper brain function is highly dependent upon controlled energy metabolism. Nicotinamide adenine dinucleotide (NAD+) is an important metabolite involved in genomic stability, cell division and cell survival. Declines in cellular NAD+ levels and its rate-limiting enzyme, nicotinamide phosphoribosyltransferase (Nampt), play a pathogenic role in age-related diseases (10,11). Importantly, these age-related impairments are attenuated by increasing NAD+ through NAD+ precursors such as nicotinamide mononucleotide (NMN) or nicotinamide riboside. Both compounds have been reported as promising therapeutic compounds to delay aging, extend the lifespan, and improve cognition (10). Notably, chemotherapy detrimentally alters brain function similar to advanced aging as evidenced by reduced neurogenesis, increased neuroinflammation (astrocyte and microglia activation) and memory dysfunction (7,8,12). In particular, chemotherapy accelerates biological aging as reported in a subset of breast cancer patients (13,14). These observations suggest that CICI and brain aging may share common pathophysiological mechanisms mediating cognitive impairment. Therefore, we sought to determine whether chemotherapy causes cognitive dysfunction through depletion of NAD+ metabolism, while investigating if increasing NAD+ levels has beneficial therapeutic effects in preventing CICI.
Materials and Methods
Mouse husbandry
All experiments were performed on 3–4 month old female C57BL/6J mice (Jackson Laboratory) unless noted otherwise. Nampt OXf/f mice were kindly provided by Dr. Joseph Baur at the University of Pennsylvania, and were generated as previously described (15). To generate adult-born neuron specific overexpression of Nampt mice, Ascl1-CreERT2 mice were purchased from the Jackson Laboratory (Stock # 012882) (16). Briefly, Nampt OXf/f mice were crossed with Ascl1-CreERT2 mice to produce Nampt OXf/+;Ascl1-CreERT2 (cNampt OX) and WT;Ascl1-CreERT2 mice (control). All mice were housed in standard cages under a 12-hour light/dark cycle with water and food ad libitum. The animal experiments were conducted in accordance with National Institutes of Health guidance on the care and use of laboratory animals. All procedures were approved by the Mayo Clinic Institutional Animal Care and Use Committee (IACUC protocol # A00005043 and A00004190).
Cisplatin and NMN administration
To induce cognitive dysfunction in mice, we used the platinum-based compound cisplatin (Fresenius Kabi, Cat# 100351), which has been reported to accumulate in significant concentrations in patient brains (17). Cisplatin is used to treat central nervous system (CNS) cancers such as neuroblastoma, germ cell tumors and primary CNS neoplasms. In addition, it is also a widely used compound to treat non-CNS cancers including ovarian, testicular, lung, breast, and bladder cancer. Briefly, female mice at age of 3–4 months were treated with daily injections of cisplatin (2.3 mg/kg/day, i.p.) or vehicle for 5 consecutive days followed by 5 days without injections. This treatment regimen (5 injection days followed by 5 non-injection days) is indicative of 1 cycle of treatment and mice were administered 3 to 4 cycles as appropriate for each experiment objective (Fig. 1A). This cisplatin regimen is known to be comparable to clinical treatment for cancer patients (18–21), and therefore widely used for rodent models of CICI as well as chemotherapy-induced neuropathic pain (CIPN) (22). Specifically, this dose regimen showed antitumor efficacy, while still inducing significant neurotoxicity to dorsal root ganglion and hippocampal neurons, leading to cisplatin-induced peripheral neuropathy and cognitive impairments in mice, respectively (23).
Figure 1. Distribution of cisplatin in adult mouse brain.
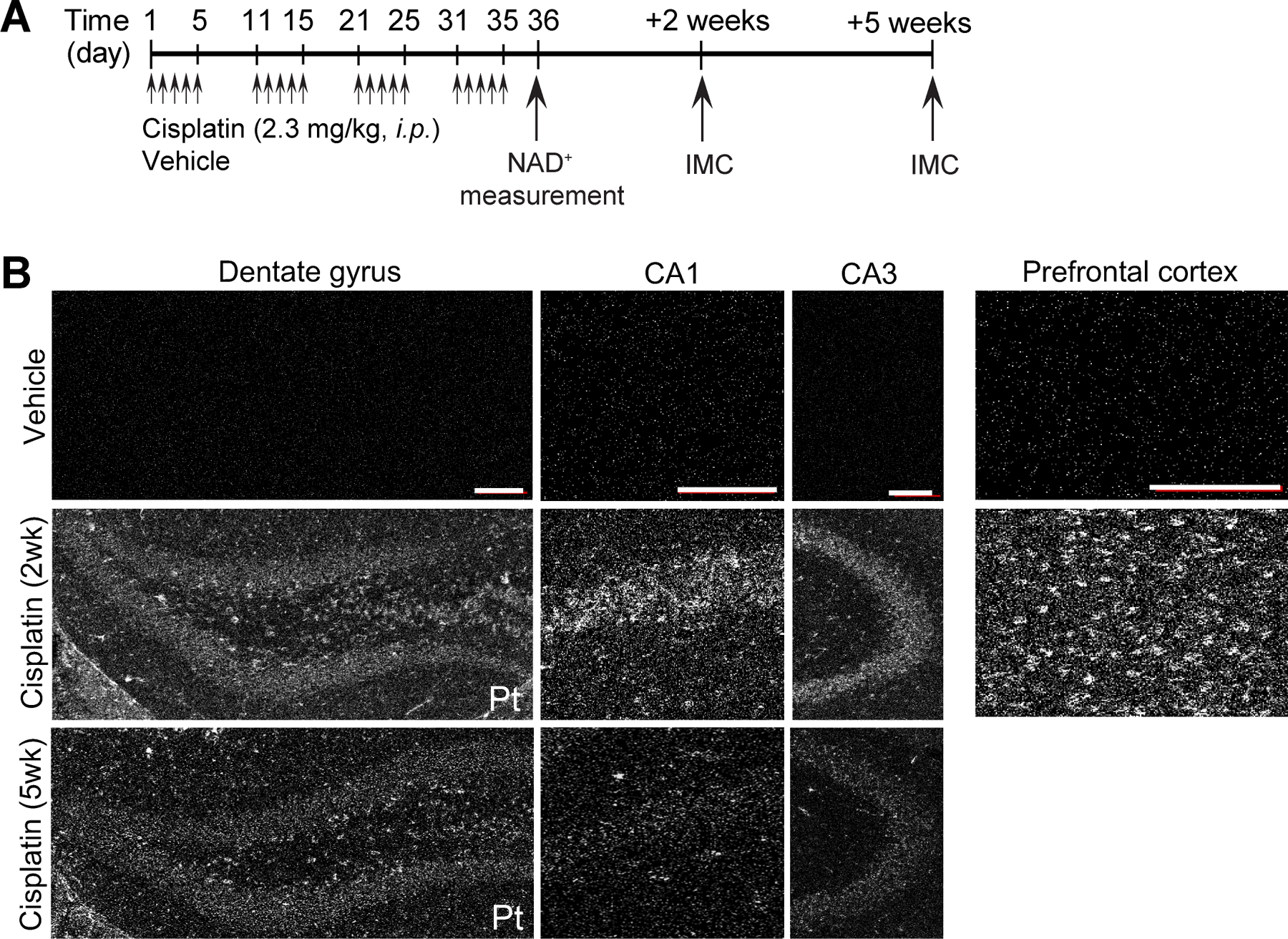
A, Schematic diagram of experimental design for cisplatin or vehicle administration. B, Representative platinum (Pt) distribution images of hippocampus sub-regions (dentate gyrus, CA1 and CA3) and prefrontal cortex from vehicle and cisplatin-treated mice using Imaging Mass Cytometry (IMC). Adult female mice were intraperitoneally injected with vehicle or cisplatin, and sacrificed at 2 and 5 weeks after the final vehicle or cisplatin injection. All scale bars: 100 μm for dentate gyrus, CA1, CA3, and prefrontal cortex.
To test the effect of NMN in cisplatin-induced cognitive impairments as presented in Fig. 2B, mice were pre-treated with NMN (250 mg/kg/day, i.p., Sigma-Aldrich, Cat# N-3501) 4 hr prior to cisplatin administration for 4 cycles. While cisplatin-treated mice exhibited significant decreases in body weight (Supplementary Fig. S1A), this did not meet moribund criteria by institution guidelines.
Figure 2. NMN prevents cisplatin-induced impairments in cognitive function.
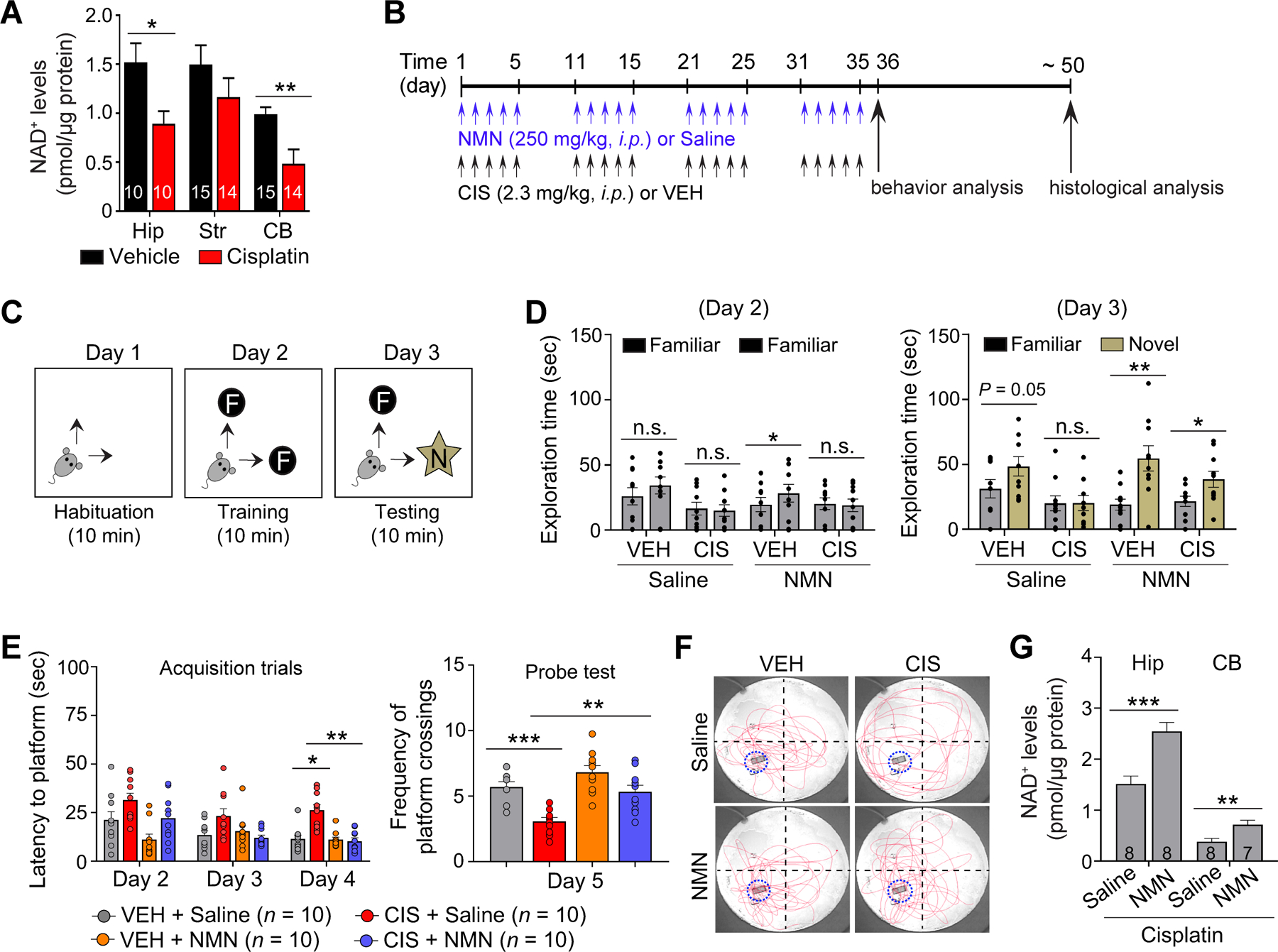
A, Reduction in cellular NAD+ levels by cisplatin administration in adult mouse hippocampus (Hip), striatum (Str), and cerebellum (CB). B, Schematic diagram of experimental design for NMN and cisplatin administrations, and follow up behavior and cellular analysis presented in Fig. 2, and 3A and B. C, Schematic diagrams of the novel object recognition (NOR) task. D, Left: Time exploring the familiar objects on Day 2. While there was a slight increase in mice treated with NMN alone, there were no major differences detected in time spent exploring the two familiar objects, indicating a lack of location preference. Right: Time exploring the objects in familiar and novel locations on Day 3 was recorded. n = 9–10 mice. E, Average latency to find the hidden platform during training, and average number of target zone crossings where the hidden platform was previously located during the probe test in Morris water maze (MWM) test. F, Representative swimming paths during the probe test (Day 5). G, Increase in cellular NAD+ levels by NMN in combination with cisplatin treatment in the adult mouse hippocampus (Hip) and cerebellum (CB). The number associated with bar graphs indicates the number of mice tested. Circles within each bar in graphs represent an individual mouse. Data represent mean ± SEM. Unpaired, two-tailed Student’s t-test (G) with Welch’s correction (A), paired, two-tailed Student’s t-test (D), and two-way ANOVA followed by Tukey’s post-hoc corrections (F). *: P < 0.05, **: P < 0.01, ***: P < 0.001, n.s.: not significant.
Measurement of NAD+ levels
To assess NAD+ levels in the hippocampus, striatum and cerebellum (Fig. 2A), these brain tissues were freshly extracted (~10–25 mg/tissue sample) without undergoing perfusion with iced saline or PFA prior to brain dissection, snap frozen in dry ice and stored at −80°C until processed. NAD+ levels were determined by using the commercial colorimetric NAD+/NADH Quantification kit (Bio Vision, Cat#: K337) according to the manufacturer’s instructions. Absorbance was measured at 450 nm using a spectrometer (Molecular Devices, Spectra Max3).
Behavior Analyses
Mice were transferred from the housing room to acclimate to the behavior facility holding area for 1 hr before testing. Testing equipment was sanitized with 70% ethanol between trials.
Novel Object Recognition (NOR) test:
NOR is a relatively fast and efficient means for testing different phases of learning and memory in rodent animals (24). The main advantage of the test is that it relies on rodents’ natural proclivity for exploring novel objects and environments. NOR testing was performed across 3 consecutive days (Fig. 2C), where on Day 1 mice were placed in an empty arena (40 × 40 × 40 cm chamber) to freely explore and habituate to the environment for 10 min and, then placed back in their home cage. On Day 2 (training phase), two identical objects (familiar objects) were placed at the bottom of the arena diagonally opposed to each other (e.g. one in the NW corner and one in the SE corner; Fig. 2C), and animals were placed in the arena to explore the objects for 10 min. On Day 3 (testing phase), one of the objects was replaced by a new object (novel object), and animals were allowed to explore for 10 minutes. The time spent exploring each object was recorded by EthoVision XT 10 (Noldus,Inc) video tracking software. The total amount of time explored on the each object with the nose was recorded and analyzed. The amount of time taken to explore the novel object in relation to the familiar object provides an index of recognition memory. As depicted in Fig. 2D, NOR results were presented as described previously (25–27).
Morris Water Maze (MWM):
MWM is a classic behavioral procedure mostly used with rodents to study spatial learning and memory. The MWM has proven to be a robust and reliable test that is strongly correlated with hippocampal-dependent cognitive function (28,29). The arena setting contains a round pool (120 cm diameter and 90 cm deep) filled with water mixed with non-fat dry milk power to make opaque water. A video camera (supported by EthoVision XT by Noldus, Inc video tracking program) was set up directly above the water pool to ensure the entire maze is within the camera’s field of view. The maze was divided into four quadrants of equal dimensions with each designated as N, S, E, W quadrants, respectively. Each quadrant had a visible spatial cue above it to facilitate spatial memory function. A submerged escape platform (30 cm high) was placed in the center of the SE quadrant. During each testing day, the water temperature was maintained between 25 °C to 28 °C. As shown in Supplementary Fig. S1B, the test was conducted in 6 consecutive days consisting of visible platform days (Day 1 and Day 6), training days (Days 2–4) and a memory probe day (Day 5). Each test day, mice would undergo 4 swimming trials per day for a duration of 2 min/trial, and interspaced with a 2–5 min interval between each trial, totaling 4 trials/day. Each trial commenced following releasing each test mouse from each quadrant release point (NE, SE, SW, NW). Mice that failed to find the location of the visible or submerged (hidden) escape platform in each trial, were manually guided to the platform for 30 seconds to learn the platform location. Briefly, on Day 1 for habituation, the escape platform was placed slightly above the water level with an orange flag pole to signal its location, thus mice were able to see and learn the platform location to escape the water. On Days 2–4, mice underwent acquisition training for spatial learning to find the hidden escape platform, which was set 1 inch below the water level so mice were trained to rely on spatial cues to learn to find the submerged platform. The latency, which is the time spent to find the platform, was recorded for further data analysis. If mice failed to find the target in 2 min, their latency was recorded as 2 min and they were guided to the platform and allowed to stay there for 30 sec. On Day 5 for the memory probe test, the escape platform was removed to determine whether mice relied on spatial cues and learned to memorize the position of the hidden platform. The frequency of crossing the platform zone and latency to find the target platform zone was recorded for data analysis. On Day 6 for visual test, the escape platform was set slightly above the water level and signaled by flag of its location. The latency to find this visible platform was recorded.
Perfusion Process and Immunostaining
Mice were anesthetized by a standard dosing regimen of ketamine (100 mg/kg), xylazine (10 mg/kg), and acepromazine (3 mg/kg) cocktail (i.p. injection at 0.1 ml/10 g body weight), and perfused transcardially through the left ventricle with cold 0.1 M phosphate buffered saline at pH 7.4 followed by a phosphate-buffered solution of 4% paraformaldehyde (PFA). Brains were post-fixed with 4% PFA overnight and 24 h later placed into 30% sucrose. Brains were sectioned and stored in antifreeze solution at −20°C. Coronal sections (40 μm in thickness) from anterior to posterior through the entire brain were prepared in serial order and processed for histological analysis. For MCM2 and Nestin immunostaining in Fig. 3A and B, and Nampt, doublecortin (DCX) and NeuN immunostaining in Supplementary Fig. S5A and S5D, an antigen retrieval procedure was carried out using a microwave as previously described (30,31). Briefly, citrate buffer (1.8 mM citric acid, 8.2 mM tri-sodium citrate) was pre-heated for 5 min at maximum power. Sections were then placed in hot citrate buffer and incubated for another 7 min. Sections were then allowed to cool at room-temperature in citrate buffer for 1 hour. Immunostaining was performed with the primary antibodies followed by secondary antibodies. DAPI (Molecular Probes, Cat# D1306) was used for counterstaining. Antibodies used in this study are listed in Supplemental Table 1. Images were acquired on a Zeiss LSM 780 single-photon confocal system using a multi-track configuration.
Figure 3. NMN prevents cisplatin-induced defects in adult hippocampal neurogenesis in vivo.
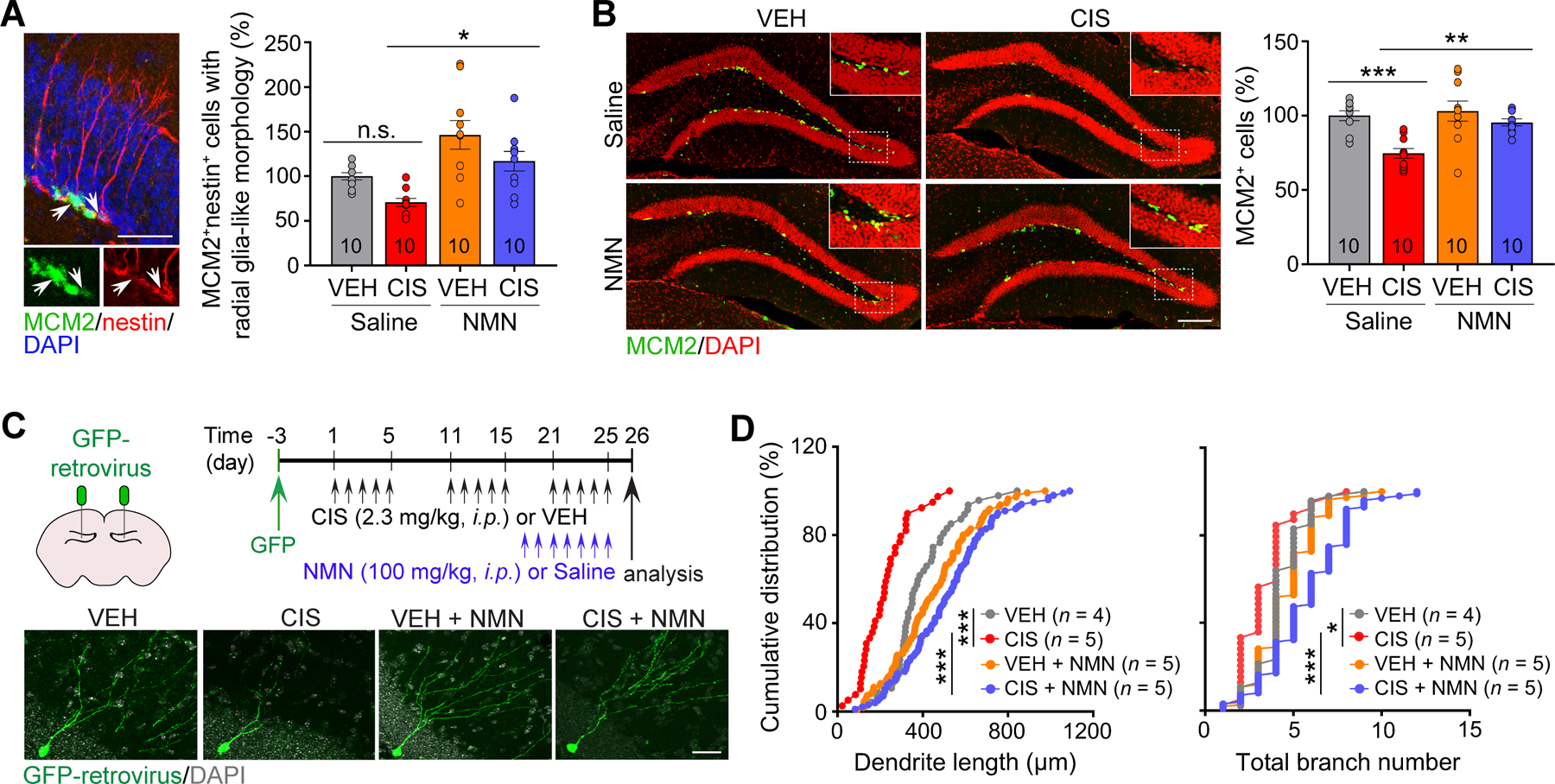
A, Representative images of MCM2 (a proliferating cell marker), nestin (a neural stem cell marker) and DAPI counter staining, and a summary of radial glia-like neural stem cell (RGL) quantification in the adult sub-granular zone (SGZ). Arrows point to MCM2+nestin+ RGLs. Scale bars: 50 μm. B, Representative images of MCM2 and DAPI counter staining, and a summary of intermediate progenitor cell (IPC) quantification in the adult SGZ. Scale bar: 200 μm. The number associated with bar graphs indicates the number of mice tested. Circles within each bar in graphs represent an individual mouse. Data represent mean ± SEM. C, Schematic representation of the intra-hippocampal retroviral injection (Upper left) and experimental design timeline (Upper right). Lower panels: Representative images of GFP+ newborn neurons (green) and DAPI staining (gray). Scale bar: 50 μm. D, Cumulative distribution plots of total dendrite length and branch number of adult-born neurons. Each symbol represents data from a single GFP+ neuron. Two-way ANOVA followed by Tukey’s post-hoc corrections (A and B). Kolmogorov-Smirnov test (D). *: P < 0.05, **: P < 0.01, ***: P < 0.001, n.s.: not significant.
Retrovirus Production, Stereotaxic Surgery, and Dendrite Analysis
Engineered self-inactivating murine onco-retroviruses were used to express GFP selectively in proliferating neural progenitors as previously described (31). Briefly, high titers of engineered retroviruses were produced by co-transfection of retroviral vectors and vesicular stomatitis virus G (VSVG) into HEK293gp cells followed by ultra-centrifugation of viral supernatant. To visualize dendrite morphology of adult-born neurons (Fig. 3C, upper left), adult female mice at 3–4 months of age were anaesthetized and retroviruses were stereotaxically injected into the dentate gyrus at 4 sites (0.5 μl per site at 0.25 μl/min) with the following coordinates (in mm): posterior = 2 from Bregma, lateral = ± 1.6, ventral = 2.5; posterior = 3 from Bregma, lateral = ± 2.6, ventral = 3.2. At 3 days post-retroviral injection when adult-born NPCs mature into post-mitotic neurons, mice were treated with cisplatin for 3-cycles followed by NMN daily treatment 7 days prior to completion of cisplatin administration (Fig. 3C; upper right). One day after completion of NMN and cisplatin administration, total dendritic length and branch numbers of GFP+ adult-born neurons were analyzed. Briefly, three-dimensional (3-D) reconstructions of the dendritic processes of each GFP+ neuron were made from Z-series stacks of confocal images (Zeiss 780). The projection images were semi-automatically traced with NIH Image J using the Neuron J plugin. The total dendritic length and branch number of individual GFP+ neurons were subsequently analyzed (31). Summaries of total dendritic length and branch number of each individual neuron are shown in cumulative distribution plots (Fig. 3D right). To induce Cre recombinase (Fig. 6C), mixed male and female Nampt OXf/+ (heterozygous) mice of 3–4 months of age were stereotaxically injected with the pCAG-GFP-IRES-Cre (Addgene plasmid #48201) retrovirus to produce overexpression of Nampt in adult-born neurons of the dentate gyrus. The pCAG-GFP (Addgene plasmid #16664) retrovirus was used as control. At 3 days post-retroviral injection, mice were treated with cisplatin for 3 cycles and sacrificed and brains were processed 1 day after the last cisplatin injection for dendrite morphological analysis.
Figure 6. Nampt overexpression selectively in adult-born neurons is sufficient to prevent cisplatin-induced impairments in dendrite morphogenesis and memory function.
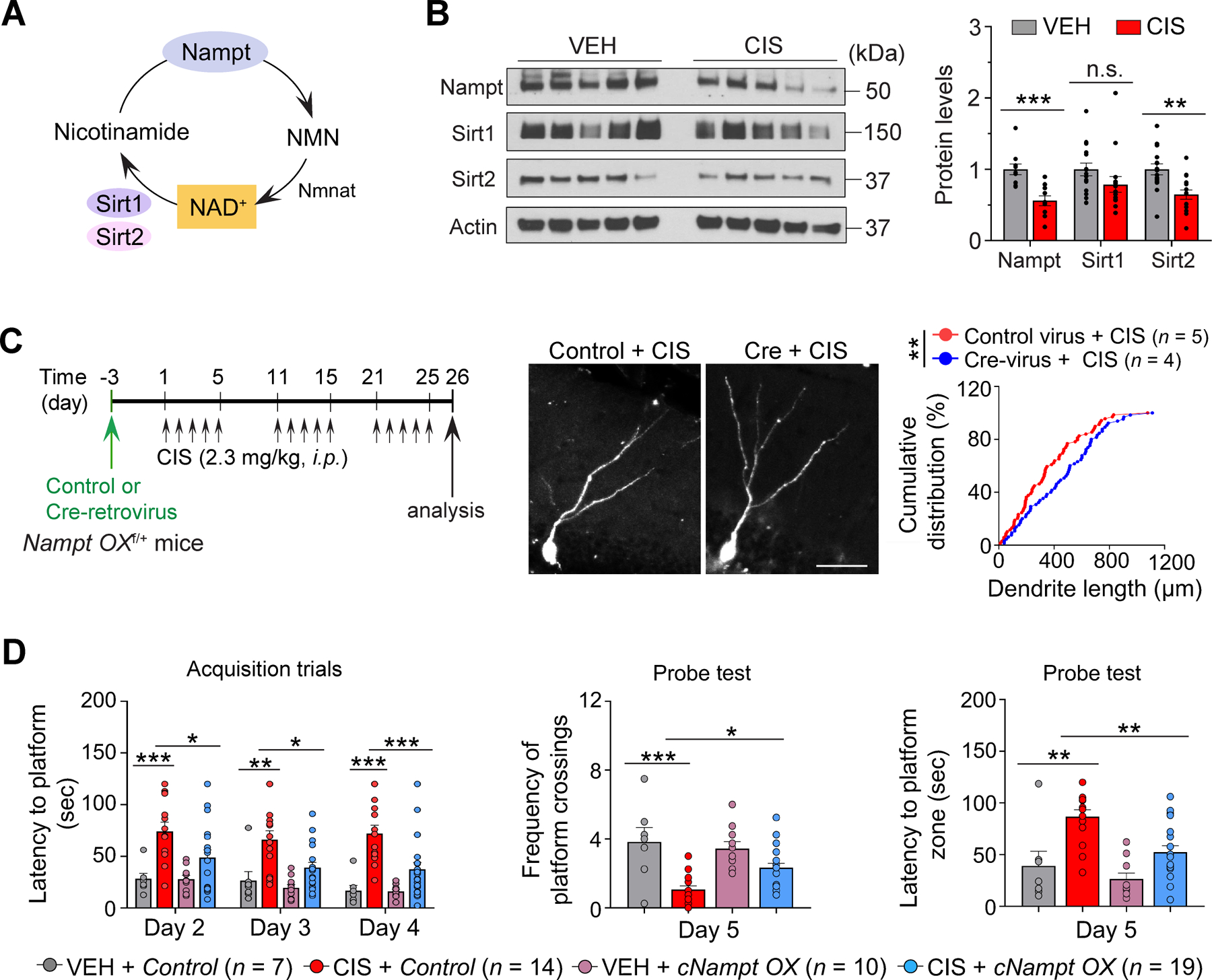
A, Nicotinamide phosphoribosyltransferase (Nampt)-mediated nicotinamide adenine dinucleotide (NAD+) biosynthetic pathways. Nampt converts nicotinamide to nicotinamide mononucleotide (NMN), which is subsequently converted into NAD+. NAD+ serves as a co-substrate for NAD+-dependent Sirtuin enzymes such as Sirt1 and Sirt2. Nicotinamide mononucleotide adenylyltransferase (Nmnat) catalyzes NAD+ synthesis. B, Representative western blots and densitometry quantification in adult mouse hippocampus lysates. n = 10 mice/group for Nampt, n = 15 mice/group for Sirt1 and Sirt2. C, Left: Schematic diagram of experimental design for the Cre- or control-retroviral injection and cisplatin administration in adult floxed Nampt overexpression mice (Nampt OXf/+). Right: Representative images of GFP+ aborn neurons and cumulative distribution plots of total dendrite length of adult-born neurons in each group. Scale bar: 50 μm. Each symbol represents data GFP+ neurons. D, Water maze test; Average latency to find the hidden platform during the training periods (left), average number of target zone crossings where the hidden platform was previously located (middle) and latency to reach platform zone during the probe test (right). Circles within each bar in graphs represent an individual mouse. Data represent mean ± SEM. Unpaired, two-tailed Student’s t-test with Welch’s correction (B), Kolmogorov-Smirnov test (C) and two-way ANOVA followed by Tukey’s post-hoc correction (D). *: P < 0.05, **: P < 0.01, ***: P < 0.001. n.s.: not significant.
Breast Cancer Cell Lines
All breast cancer cell lines were obtained from ATCC and were provided by J.R. Hawse (Mayo Clinic) for this study. All cell lines were routinely tested for Mycoplasma using the Mycoplasma Detection Kit (SouthernBiotech, Cat# 13100–01) and have been confirmed to be negative. All cells were grown in DMEM/F12 media (Gibco, Cat #11330032) supplemented with 10% FBS (HyClone, Cat# SH30071.03) and 1% penicillin–streptomycin (Gibco, Cat# 15070063) and cultured at 37°C and 5% CO2. Cells were utilized at low passage and were cultured for no more than 2 months from time of thawing to experimentation.
Cell Viability Analysis
To test the effect of NMN on tumor growth, we performed an in vitro cell viability analysis using 9 different breast cancer cell lines and two ovarian cancer patient-derived xenografts (PDX) 3D models that has demonstrated strong correlation with clinical platinum-sensitivity in patients (32).
For breast cancer cell line studies (Supplementary Fig. S4A and S4B), ~1,000 cells/well were plated in 96 well tissue culture plates in replicates of 8 and allowed to adhere overnight. Cells were subsequently treated with vehicle control or indicated concentrations of NMB and allowed to proliferate for 7 days. Following treatment, cells were lysed and cell viability was determined using CellTiter-Glo® Luminescent Cell Viability Assays according to the manufacturer’s protocol. Vehicle treated cells were set to 100% viability and values from NMN treatments are depicted relative to controls.
For PDX studies (Supplementary Fig. S4C), ovarian cancer ex vivo 3D culture was performed on fresh PDX tissue resected from mice. Tumors (1 to 1.5 grams) were first minced with surgical shears and partially digested using a MACS Tumor Dissociation Kit (Miltenyi Biotec Inc., Cat# 130-095-929) with gentleMACS C Tubes in an Octo Dissociator for 1 hr at 37°C. Cell aggregates were washed twice with MACS buffer and seeded on 96-well low-binding plates at 1 to 5×104 cells per well in DMEM with 10% FBS and 1% Pen/Strep. Cells were allowed to acclimate for 24 hrs before exposure to one of three conditions: cisplatin (0, 1.5, 3, 6.25, 12.5, 25 and 50 μM), NMN (0, 31.25, 62.5,125, 250, 500 and 1000 μM), or both in combination. RealTime-Glo MT Cell Viability reagent (Promega, Cat# G9711) was added 72 hrs after plating and cells were imaged for viability ~120 hrs after plating.
Xenograft Tumor Models
To test the in vivo effects of NMN and cisplatin on tumor growth (Fig. 5), MDA-MB-231 cells (triple-negative) and MCF-7 ERα+ cells (estrogen receptor-alpha) were injected orthotopically into 4–5 months-old CB17/Icr-Prkdcscid SCID female mice (Charles River, Strain Code: 236). Briefly, cells with a concentration of ~1 × 106 cells in 100μL + 100μL of matrigel (Corning, Cat# 356231) were injected into the mammary fat pads of 4–5 month-old CB17 SCID female mice. For mice injected with MCF-7 cells, 17β-estradiol (0.54 mg/90-day release; Innovative Research of America, Cat# NE-121) pellets were implanted into the nape of the neck using a trochar to induce tumor growth. Once the average tumor volume for all animals reached approximately 50 mm3, mice were treated with 4 cycles of 5 daily i.p. injections of cisplatin (2.3 mg/kg/day) concurrently with NMN (250 mg/kg/day) or saline, followed by a 5-day rest without injections.
Figure 5. No impact of NMN on tumor growth or anti-tumor activity of cisplatin.
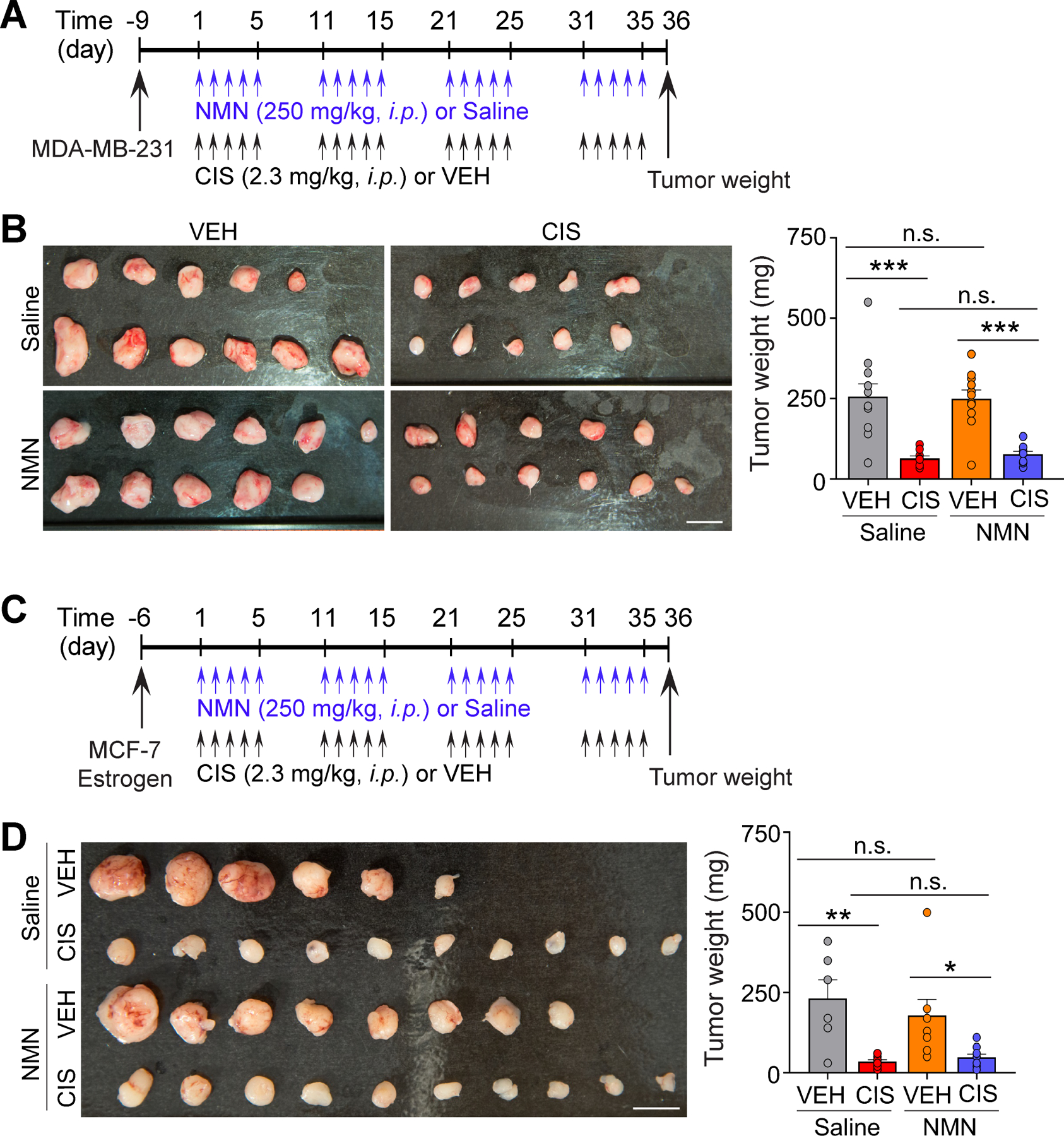
A, Schematic diagram of experimental design. B, MDA-MB-231 xenograft tumors harvested from mice and comparison of tumor weights in each group. C, Diagram of experimental design. n = 10–11 mice. D, MCF-7 xenograft tumors harvested from mice and comparison of tumor weights in each group. Scale bars: 1 cm. Circles within each bar in graphs represent an individual mouse. n = 6–10 mice. Data represents mean ± SEM. Two-way ANOVA followed by Tukey’s post-hoc corrections (B and D). *: P < 0.05, **: P < 0.01, ***: P < 0.001, n.s.: not significant.
Statistical Analysis
All statistical analyses were performed using GraphPad Prism 8. A two-tailed unpaired or paired student’s t-test, one or two-way ANOVA, and Dunnett’s or Tukey’s post-hoc test for multiple comparisons were used, as appropriate for each experiment. For dendrite analysis in Fig. 3C and Fig. 6C, the Kolmogorov-Smirnov test was performed. Statistical significance was defined as P ≤ 0.05 (*P < 0.05, **P < 0.01, ***P < 0.001). P > 0.05 were defined as statistically not significant (n.s.). Two outliers on Fig. 2D (Day 2) and Fig. 2G were identified via GraphPad Prism Grubbs outlier test, and these values were removed. All exact P values, detailed statistical analysis information, and animal sources, group size, and gender of the mouse models are provided in the Supplemental Table 2. All experiments and data analyses were performed in a blinded fashion.
See Supplementary Information for additional methods.
Results
Cisplatin penetrates the blood–brain barrier
We established an in vivo model of CICI using female mice administered cisplatin (Fig. 1A), a platinum-based compound widely used to treat various cancers including ovarian, testicular, lung, and breast cancer. Given that cisplatin has been reported to induce cognitive dysfunction in cancer survivors (33), understanding the mechanisms of how cisplatin induces neurotoxicity will be critical for development of therapies to prevent or treat CICI. To explore whether cisplatin permeates the brain, we performed Imaging Mass Cytometry (IMC) to visualize platinum distribution in the hippocampus and prefrontal cortex from mice treated with cisplatin or vehicle. We have chosen the hippocampus and the prefrontal cortex because both structures are essential for cognitive function and are reported to be more vulnerable to chemotherapy in humans (34–37). As shown in Fig. 1B, we detected platinum in the brains of mice treated with cisplatin at 2 and 5 weeks following the last cisplatin injection, thus demonstrating that cisplatin penetrates the blood–brain barrier and persists long after cessation of cisplatin administration.
Preventative effects of NMN on cisplatin-induced cognitive impairments in adult female mice
Declines in NAD+ have emerged as a molecular hallmark of age-related diseases (10). Given the similarity between CICI and brain aging, we tested if cisplatin alters cellular NAD+ levels in the adult female mouse brain. We observed that NAD+ is significantly reduced by cisplatin in the hippocampus and cerebellum (Fig. 2A). This result led us to test whether increasing NAD+ levels could rescue cisplatin-induced cognitive deficits. NAD+ precursor NMN is known to increase hippocampal NAD+ levels, promote NSC proliferation (11), and improve memory function in aged mice (38). Adult female mice were then pre-treated with NMN at the daily dose of 250 mg/kg i.p., followed by cisplatin treatment four hours later in each of 4 cycles (Fig. 2B). Body weight (BW) assessment did not find detrimental effects of NMN, although cisplatin-treated mice displayed significant decreases in BW (Supplementary Fig. S1A). We next performed novel object recognition (NOR; Fig. 2C) testing and the Morris water maze (MWM; Supplementary Fig. S1B), both of which assess memory and are strongly affected by hippocampal impairments. In addition, cisplatin has been reported to impair memory function in these tests (8,39). Our NOR results showed that while there was a slight increase in mice treated with NMN alone, there were no significant differences detected in time spent exploring the two familiar objects (Day 2; Fig. 2D, left), indicating a lack of location exploration preference. However, when a familiar object was replaced by a novel object during test Day 3, cisplatin treated mice spent less time exploring the novel object, relative to vehicle-treated mice, indicating that cisplatin impairs memory function. Notably, when NMN was given to vehicle and cisplatin-treated mice, both groups significantly increased exploration of the novel object (Fig. 2D, right), suggesting that NMN prevents cisplatin-induced deficits in recognition memory.
To strengthen our results and avoid reliance on a single memory test such as NOR, we performed the MWM which is a more nuanced memory assay. The MWM test showed that cisplatin-treated mice had longer latencies to find the hidden platform during acquisition training (Day 2–4), suggesting that cisplatin-treated mice displayed learning impairments. However, NMN administration prior to cisplatin-treatment significantly reduced escape latencies on Day 4 (Fig. 2E, left). During the probe test (Day 5), cisplatin-treated mice exhibited significantly fewer target platform zone crossings (Fig. 2E, right and Fig. 2F) and longer latencies to reach the target platform zone (Supplementary Fig. S1C) compared to vehicle-treated mice, indicating spatial memory dysfunction by cisplatin. Importantly, NMN in cisplatin-treated mice showed greater crossing frequencies in the target platform zone and shorter escape latencies when compared to mice given cisplatin, suggesting a neuroprotective role for NMN in cisplatin-induced spatial memory dysfunction in female mice. Notably, there were no differences in swim speed or visual function (Supplementary Fig. S1D and E, respectively), confirming that the effects of NMN or cisplatin are on learning and memory rather than physical or visual function. Furthermore, we found that the observed neuroprotective effects of NMN on memory function is correlated with increased levels of NAD+ in the hippocampus and cerebellum of cisplatin treated mice (Fig. 2G).
Neuroprotective effects of NMN on cisplatin-induced defects in neurogenesis in adult female mice in vivo.
Significant reduction of hippocampal neurogenesis by cisplatin and other chemotherapeutics is a common pathological feature that may contribute to cognitive dysfunction (6–8). As an initial step of neurogenesis, radial glia-like neural stem cells (RGLs) in the subgranular zone (SGZ) produce intermediate progenitor cells (IPCs), which give rise to neuroblasts in the adult mouse dentate gyrus (DG; Supplementary Fig. S2A). Following completion of behavior testing, we analyzed the effects of cisplatin and NMN on RGLs defined by co-expressing MCM2 (a proliferative marker) and nestin (a neural stem cell marker) cells with radial processes. IPCs were defined by MCM2 expression. Our quantitative analysis showed a decreased trend in RGL population in cisplatin-treated female mice, while this reduction was significantly restored by NMN administration (Fig. 3A). Similarly, MCM2-expressing IPCs were significantly reduced by cisplatin, which was prevented by NMN administration (Fig. 3B). There were no obvious sex differences found as cisplatin induced reductions in RGL and IPC populations (Supplementary Fig. S2B). We further analyzed if cisplatin affects neuronal morphogenesis of adult-born neurons and whether NMN can reverse this abnormal process. Adult female mice were stereotaxically injected with a GFP-expressing retrovirus to visualize adult-born neuron morphology. At 3 days post-retroviral injection when adult-born NPCs mature into post-mitotic neurons, mice were treated with cisplatin for 3-cycles followed by NMN daily treatment 7 days prior to completion of cisplatin administration (Fig. 3C). We show that cisplatin significantly decreases total dendritic length and branch numbers of GFP+ adult-born neurons compared to vehicle administration. Remarkably, the cisplatin-induced reduction in dendrite length and branch number was fully recovered by NMN (Fig. 3D). Taken together, these findings suggest a therapeutic effect of NMN in cisplatin-induced impairments in NPC proliferation, neuronal morphogenesis and memory function in adult female mice.
Neuroprotective effects of NMN in cisplatin-induced abnormality in neuronal morphogenesis in human cortical neurons derived from iPSCs in vitro.
To further assess the effects of cisplatin and NMN in human neurons, we generated excitatory cortical neurons derived from human induced pluripotent stem cells (iPSCs; Fig. 4A, B, and Supplementary Fig. S3A). Cortical human neurons that were exposed to cisplatin displayed significant suppression of neuronal NAD+ levels at the lowest dose (0.01 μM; Fig. 4C) without significantly affecting cell viability (Supplementary Fig. S3B), indicating that neuronal NAD+ reductions were not due to decreased cell numbers by cisplatin. Intriguingly, while NMN alone does not affect cell viability, pre-treatment with NMN significantly prevents cisplatin-induced reduction in cell survival (Supplementary Fig. S3C). Morphological analysis also showed that cisplatin dramatically suppressed total neurite length in human neurons in a dose-dependent fashion (Supplementary Fig. S3D). More importantly, these abnormal phenotypes were ameliorated by NMN at doses ranging from 250–1000 μM, with the greatest efficacy at 500 μM (Fig. 4D). Notably, NMN administration without cisplatin treatment significantly promoted neurite outgrowth, suggesting that neuronal morphogenesis is highly dependent on neuronal NAD+ levels. Collectively, our results confirm a neuroprotective role for NMN in cisplatin-induced defects in cell viability and neuronal morphogenesis in human cortical neurons.
Figure 4. Neuroprotective effects of NMN in cisplatin-induced abnormality in neuronal morphogenesis in human cortical neurons derived from iPSCs in vitro.
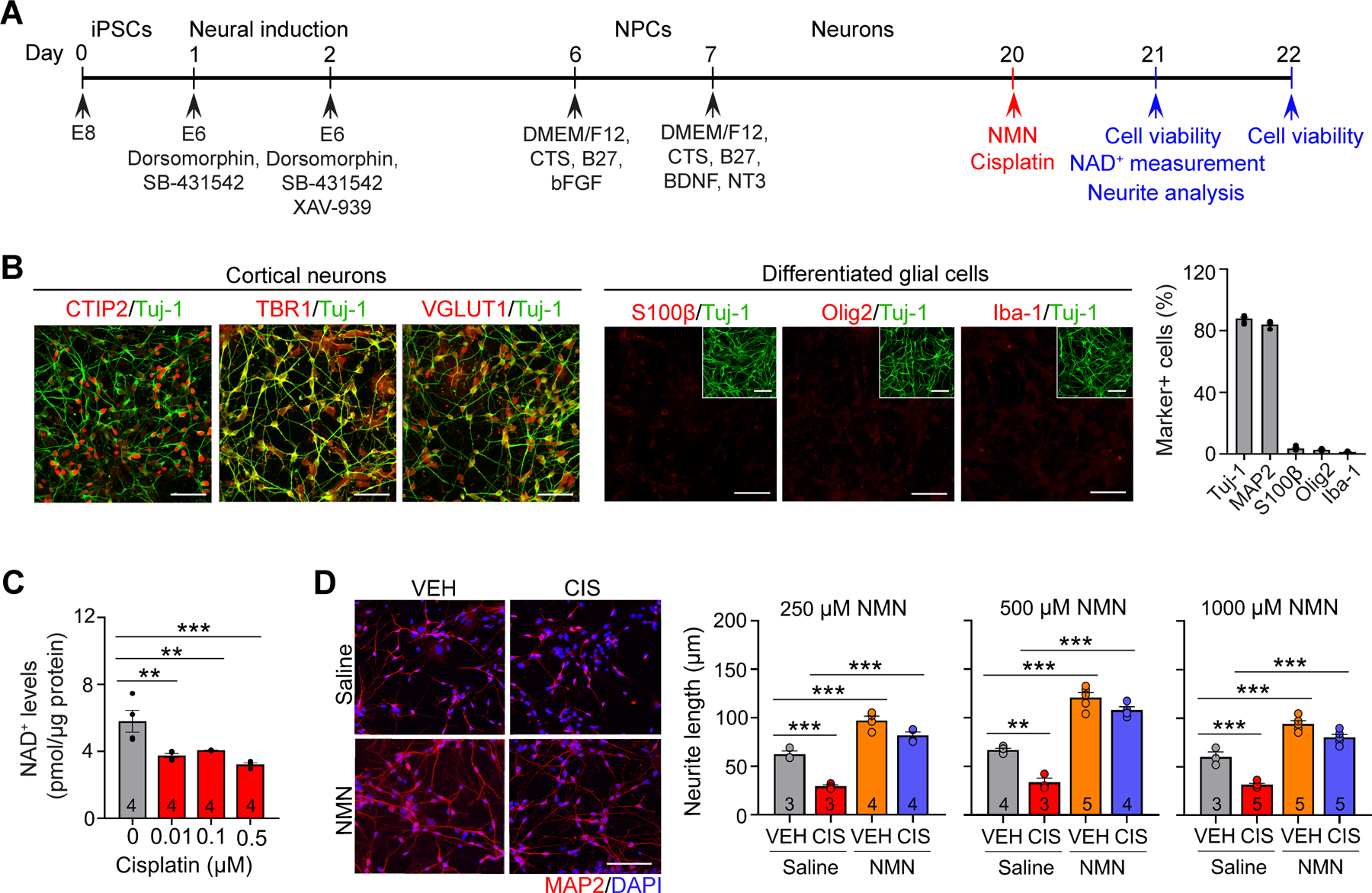
A, Schematic diagram of the differentiation procedure. B, Representative confocal images for Tuj-1 (neuron markers), CTIP2 and TBR1 (cortical neuron markers), VGLUT1 (excitatory neuron marker), S100β (astrocyte marker), Olig2 (oligodendrocyte marker) and Iba-1 (microglia marker). Note that the majority of iPSCs were differentiated into cortical excitatory neurons, but not glial cells. All scale bars: 50 μm. C, Reduction in neuronal NAD+ levels by cisplatin administration in human cortical neurons. n = 4 wells/each treatment group. D, Left: Representative images of MAP2 (neuron marker) and DAPI (nuclei marker). Scale bar: 50 μm. Right: Quantification of the neurite outgrowth in each treatment group. Each circle represents an individual well. Data represents mean ± SEM. One-way ANOVA followed by Dunnett’s (C) and two-way ANOVA followed by Tukey’s post-hoc corrections (D). **: P < 0.01, ***: P < 0.001.
No impact of NMN on tumor growth or anti-tumor activity of cisplatin.
As we aimed to identify NMN as a potential compound to prevent CICI, an important consideration is whether NMN inadvertently promotes tumor growth or interferes with the efficacy of cisplatin. We did not find cell viability differences in 9 breast cancer cell lines when exposed to our NMN doses (0-to-1000 μM NMN), indicating that NMN does not promote tumor growth in these cell lines (Supplementary Fig. S4A and S4B). Similarly, NMN does not promote tumor growth or impair cisplatin’s anti-tumor activity in two ovarian cancer PDX 3D models (Supplementary Fig. S4C). To further test the effects of NMN in vivo, triple negative (MDA-MB-231; Fig. 5A and B) and estrogen receptor positive (MCF-7; Fig. 5C and D) breast cancer cell lines were implanted orthotopically into adult severe combined immunodeficiency female mice followed by NMN or cisplatin administration. While cisplatin administration reduced tumor sizes, there were no significant tumor size changes by NMN treatment in either vehicle-treated mice or cisplatin-treated mice. Our results suggest that NMN neither promotes tumor growth nor interrupts the anti-tumor activity of cisplatin, ensuring NMN safety at the effective dose utilized for behavior memory tests.
Genetic Nampt overexpression selectively in adult-born neurons is sufficient to prevent cisplatin-induced impairments in dendrite morphogenesis and memory function.
We next asked how cisplatin suppresses cellular NAD+ levels to potentially impair adult hippocampal neurogenesis and consequent memory function. Given that Nampt is the rate-limiting enzyme in NAD+ biosynthesis (Fig. 6A), we assessed if cisplatin inhibits Nampt levels in the adult mouse hippocampus. Western blot analysis showed that Nampt expression was suppressed by cisplatin to about 50% of vehicle-treated mice (Fig. 6B). Notably, we detected reduction of Nampt expression exclusively in hippocampal neurons (Supplementary Fig. S5A). In addition, NAD+-dependent Sirtuin enzyme, Sirt2, expression was significantly reduced by cisplatin without significantly changing Sirt1 levels (Fig. 6B). Therefore, these data indicate that cisplatin reduces expression of Nampt, and NAD+-dependent Sirt2 in the adult female mouse hippocampus.
Genetic deletion of Nampt or Sirt2 is known to impair NPC proliferation, synaptic plasticity and memory function (40,41). To determine if reduction in Nampt levels is responsible for cisplatin’s aberrant effects on neurogenesis, we reasoned that sustained Nampt levels may be able to prevent cisplatin-induced neurogenic deficits. To selectively increase Nampt levels within adult-born neurons, adult floxed Nampt overexpressing mice (Nampt OXf/+ mice) were stereotaxically injected in the hippocampal DG with a co-expressing Cre-recombinase and GFP-retrovirus (Cre-retrovirus) or GFP-retrovirus that does not express Cre-recombinase (Control-retrovirus), followed by 3-cycles of cisplatin injections (Fig. 6C, left). Morphological assessment at 1-day post cisplatin administration demonstrated an increase in total dendrite length of Cre-expressing adult-born neurons compared to controls (Fig. 6C, right). These results suggest that Nampt overexpression promotes adult-born neuron dendrite outgrowth in cisplatin-treated mice in a cell-autonomous manner. We then examined whether selectively increasing Nampt levels within adult-born neurons is sufficient to prevent cisplatin-induced memory impairment by generating conditional Nampt OXf/+;Ascl1-CreERT2 (cNampt OX) and WT;Ascl1-CreERT2 mice (Control; Supplementary Fig. S5B–S5E). We selected Ascl1-CreERT2 mice because the majority of Ascl1-expressing neural progenitors in the adult DG mature into adult dentate granule neurons expressing NeuN within 30 days after tamoxifen administration (16), thus minimizing confounding glial contribution to behavior function derived from progenitors. As expected, cisplatin-treated mice exhibited longer latencies to reach the hidden platform during acquisition trials, fewer platform crossings and longer latencies to reach the target platform zone during the probe test when compared to vehicle-treated controls (Fig. 6D; left, center and right, respectively and Supplementary Fig. S5F–S5H). In contrast, cisplatin-treated cNampt OX mice showed decreased latencies to find the hidden platform, increased platform crossings and decreased latencies to reach the target platform zone relative to cisplatin-treated controls. Interestingly, cNampt OX mice did not show a complete recovery from cisplatin’s effects. A potential explanation is that the adult-born neuron population comprises a small portion (approximately 6%) of the whole dentate granule neuron population in the adult hippocampus (42). Therefore, we expect that Nampt overexpression selectively within adult-born neurons will only partially increase NAD+ levels at the whole-tissue level, thus incompletely preventing the learning and memory impairments induced by cisplatin. Nonetheless, these results suggest that selective sustained expression of Nampt in adult-born neurons is sufficient to prevent cisplatin-induced impairment in spatial learning and memory function, demonstrating critical causative roles of Nampt and adult-born neurons that are detrimentally affected by cisplatin.
Discussion
Here we identify that disruption in Nampt-mediated NAD+ metabolism may be a contributor in cisplatin-induced impairments on neurogenesis and memory function. While early studies of methotrexate-induced memory impairment attributed glial dysregulation and neuroinflammation as contributors to CICI (12), our study extends a new causative role for adult-born neurons underlying cisplatin-induced memory impairment. Furthermore, we identify Nampt as an essential enzyme that mediates cisplatin-induced NAD+ loss, indicating that Nampt-NAD+ dysregulation results in CICI. Notably, the Nampt-mediated NAD+ metabolic pathway is known to play a role in cancer pathogenesis (43). However, we did not find any detrimental impact of NMN on tumor growth and cisplatin’s anti-tumor activity, ensuring that NMN is a safe and promising compound for CICI. In addition, since other chemotherapies are associated with cognitive dysfunction (12), it is of importance to determine whether the neurotoxic effect of these drugs are mediated through dysregulated NAD+ metabolism. If this is the case, NMN could be a general therapeutic option to abate the neurotoxicity associated with other chemotherapies. Moreover, given that CICI is commonly reported among cancer patients with breast cancer, we focused our study on the ameliorative effects of NMN on cisplatin-CICI in female mice. However, whether NMN may have similar neuroprotective effects in male mice remains an open question. Nonetheless, Nampt-NAD+ metabolism is actively being developed as a pharmacotherapy against aging and age-related diseases, and NMN is currently undergoing clinical trials to improve age-related metabolic dysfunction. Therefore, our prophylactic strategies to prevent NAD+ loss using NMN supplementation during cisplatin-chemotherapy is a promising therapeutic strategy that is rapidly and safely applicable to prevent cisplatin-induced neurotoxicity and improve quality of life for cancer patients.
Supplementary Material
Statement of significance.
Increasing NAD+ through nicotinamide mononucleotide supplementation offers a potential therapeutic strategy to safely prevent cisplatin-induced cognitive impairments, thus providing hope for improved quality of life in cancer survivors.
Acknowledgements
We would like to thank Dr. Edurado Chini, Dr. Jann Sarkaria, Dr. Nathan LeBrasseur, and Dr. Anthony Windebank for their helpful suggestions, and Dr. Zachary T. Resch at Center for Regenerative Medicine Biotrust at Mayo Clinic for generating iPSC line. This work was supported by NIH (R01CA242158, R01AG058560), Mayo Clinic Breast Cancer SPORE (P50CA116201), Regenerative Medicine Minnesota (RMM091718 DS 005), Eagles 5th District Cancer Telethon Funds to M.H. Jang, Mayo Clinic SPORE in Ovarian Cancer (CA136393) to S.J. Weroha, and the 2019 The Bosarge Family Foundation-Waun Ki Hong Scholar Award for Regenerative Cancer Medicine from the American Association for Cancer Research (19-40-60-OLIV) to A. Oliveros.
Footnotes
Conflict of Interest: The authors have declared that no conflict of interest exists.
References
- 1.Lange M, Joly F, Vardy J, Ahles T, Dubois M, Tron L, et al. Cancer-related cognitive impairment: an update on state of the art, detection, and management strategies in cancer survivors. Ann Oncol 2019;30:1925–40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, et al. Neurogenesis in the adult human hippocampus. Nat Med 1998;4:1313–7 [DOI] [PubMed] [Google Scholar]
- 3.Moreno-Jimenez EP, Flor-Garcia M, Terreros-Roncal J, Rabano A, Cafini F, Pallas-Bazarra N, et al. Adult hippocampal neurogenesis is abundant in neurologically healthy subjects and drops sharply in patients with Alzheimer’s disease. Nat Med 2019;25:554–60 [DOI] [PubMed] [Google Scholar]
- 4.Sahay A, Scobie KN, Hill AS, O’Carroll CM, Kheirbek MA, Burghardt NS, et al. Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature 2011;472:466–70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Clelland CD, Choi M, Romberg C, Clemenson GD Jr., Fragniere A, Tyers P, et al. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science 2009;325:210–3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gong X, Schwartz PH, Linskey ME, Bota DA. Neural stem/progenitors and glioma stem-like cells have differential sensitivity to chemotherapy. Neurology 2011;76:1126–34 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rendeiro C, Sheriff A, Bhattacharya TK, Gogola JV, Baxter JH, Chen H, et al. Long-lasting impairments in adult neurogenesis, spatial learning and memory from a standard chemotherapy regimen used to treat breast cancer. Behav Brain Res 2016;315:10–22 [DOI] [PubMed] [Google Scholar]
- 8.Chiu GS, Maj MA, Rizvi S, Dantzer R, Vichaya EG, Laumet G, et al. Pifithrin-mu Prevents Cisplatin-Induced Chemobrain by Preserving Neuronal Mitochondrial Function. Cancer Res 2017;77:742–52 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wigmore P The effect of systemic chemotherapy on neurogenesis, plasticity and memory. Curr Top Behav Neurosci 2013;15:211–40 [DOI] [PubMed] [Google Scholar]
- 10.Lautrup S, Sinclair DA, Mattson MP, Fang EF. NAD(+) in Brain Aging and Neurodegenerative Disorders. Cell Metab 2019;30:630–55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Stein LR, Imai S. Specific ablation of Nampt in adult neural stem cells recapitulates their functional defects during aging. EMBO J 2014;33:1321–40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Gibson EM, Nagaraja S, Ocampo A, Tam LT, Wood LS, Pallegar PN, et al. Methotrexate Chemotherapy Induces Persistent Tri-glial Dysregulation that Underlies Chemotherapy-Related Cognitive Impairment. Cell 2019;176:43–55 e13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sanoff HK, Deal AM, Krishnamurthy J, Torrice C, Dillon P, Sorrentino J, et al. Effect of cytotoxic chemotherapy on markers of molecular age in patients with breast cancer. J Natl Cancer Inst 2014;106:dju057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Carroll JE, Van Dyk K, Bower JE, Scuric Z, Petersen L, Schiestl R, et al. Cognitive performance in survivors of breast cancer and markers of biological aging. Cancer 2019;125:298–306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Frederick DW, Davis JG, Davila A Jr., Agarwal B, Michan S, Puchowicz MA, et al. Increasing NAD synthesis in muscle via nicotinamide phosphoribosyltransferase is not sufficient to promote oxidative metabolism. J Biol Chem 2015;290:1546–58 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kim EJ, Leung CT, Reed RR, Johnson JE. In vivo analysis of Ascl1 defined progenitors reveals distinct developmental dynamics during adult neurogenesis and gliogenesis. J Neurosci 2007;27:12764–74 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nakagawa H, Fujita T, Kubo S, Tokiyoshi K, Yamada M, Kanayama T, et al. Difference in CDDP penetration into CSF between selective intraarterial chemotherapy in patients with malignant glioma and intravenous or intracarotid administration in patients with metastatic brain tumor. Cancer Chemother Pharmacol 1996;37:317–26 [DOI] [PubMed] [Google Scholar]
- 18.Le X, Hanna EY. Optimal regimen of cisplatin in squamous cell carcinoma of head and neck yet to be determined. Ann Transl Med 2018;6:229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Szturz P, Wouters K, Kiyota N, Tahara M, Prabhash K, Noronha V, et al. Low-Dose vs. High-Dose Cisplatin: Lessons Learned From 59 Chemoradiotherapy Trials in Head and Neck Cancer. Front Oncol 2019;9:86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cavaletti G, Marzorati L, Bogliun G, Colombo N, Marzola M, Pittelli MR, et al. Cisplatin-induced peripheral neurotoxicity is dependent on total-dose intensity and single-dose intensity. Cancer 1992;69:203–7 [DOI] [PubMed] [Google Scholar]
- 21.van der Hoop RG, van der Burg ME, ten Bokkel Huinink WW, van Houwelingen C, Neijt JP. Incidence of neuropathy in 395 patients with ovarian cancer treated with or without cisplatin. Cancer 1990;66:1697–702 [DOI] [PubMed] [Google Scholar]
- 22.Ta LE, Low PA, Windebank AJ. Mice with cisplatin and oxaliplatin-induced painful neuropathy develop distinct early responses to thermal stimuli. Mol Pain 2009;5:9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhou W, Kavelaars A, Heijnen CJ. Metformin Prevents Cisplatin-Induced Cognitive Impairment and Brain Damage in Mice. PLoS One 2016;11:e0151890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Broadbent NJ, Gaskin S, Squire LR, Clark RE. Object recognition memory and the rodent hippocampus. Learn Mem 2010;17:5–11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.McGinley LM, Kashlan ON, Bruno ES, Chen KS, Hayes JM, Kashlan SR, et al. Human neural stem cell transplantation improves cognition in a murine model of Alzheimer’s disease. Sci Rep 2018;8:14776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Horiguchi M, Hannaway KE, Adelekun AE, Jayathilake K, Meltzer HY. Prevention of the phencyclidine-induced impairment in novel object recognition in female rats by co-administration of lurasidone or tandospirone, a 5-HT(1A) partial agonist. Neuropsychopharmacology 2012;37:2175–83 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Cruz-Sanchez A, Dematagoda S, Ahmed R, Mohanathaas S, Odenwald N, Arruda-Carvalho M. Developmental onset distinguishes three types of spontaneous recognition memory in mice. Sci Rep 2020;10:10612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Clark RE, Broadbent NJ, Squire LR. The hippocampus and spatial memory: findings with a novel modification of the water maze. J Neurosci 2007;27:6647–54 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Vorhees CV, Williams MT. Morris water maze: procedures for assessing spatial and related forms of learning and memory. Nat Protoc 2006;1:848–58 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hussaini SM, Jun H, Cho CH, Kim HJ, Kim WR, Jang MH. Heat-induced antigen retrieval: an effective method to detect and identify progenitor cell types during adult hippocampal neurogenesis. J Vis Exp 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jang MH, Bonaguidi MA, Kitabatake Y, Sun J, Song J, Kang E, et al. Secreted frizzled-related protein 3 regulates activity-dependent adult hippocampal neurogenesis. Cell Stem Cell 2013;12:215–23 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Shuford S, Wilhelm C, Rayner M, Elrod A, Millard M, Mattingly C, et al. Prospective Validation of an Ex Vivo, Patient-Derived 3D Spheroid Model for Response Predictions in Newly Diagnosed Ovarian Cancer. Sci Rep 2019;9:11153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chovanec M, Vasilkova L, Setteyova L, Obertova J, Palacka P, Rejlekova K, et al. Long-Term Cognitive Functioning in Testicular Germ-Cell Tumor Survivors. Oncologist 2018;23:617–23 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Dumas JA, Makarewicz J, Schaubhut GJ, Devins R, Albert K, Dittus K, et al. Chemotherapy altered brain functional connectivity in women with breast cancer: a pilot study. Brain Imaging Behav 2013;7:524–32 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kesler SR, Kent JS, O’Hara R. Prefrontal cortex and executive function impairments in primary breast cancer. Arch Neurol 2011;68:1447–53 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cheng H, Li W, Gong L, Xuan H, Huang Z, Zhao H, et al. Altered resting-state hippocampal functional networks associated with chemotherapy-induced prospective memory impairment in breast cancer survivors. Sci Rep 2017;7:45135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kesler S, Janelsins M, Koovakkattu D, Palesh O, Mustian K, Morrow G, et al. Reduced hippocampal volume and verbal memory performance associated with interleukin-6 and tumor necrosis factor-alpha levels in chemotherapy-treated breast cancer survivors. Brain Behav Immun 2013;30 Suppl:S109–16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tarantini S, Valcarcel-Ares MN, Toth P, Yabluchanskiy A, Tucsek Z, Kiss T, et al. Nicotinamide mononucleotide (NMN) supplementation rescues cerebromicrovascular endothelial function and neurovascular coupling responses and improves cognitive function in aged mice. Redox Biol 2019;24:101192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Oz M, Nurullahoglu Atalik KE, Yerlikaya FH, Demir EA. Curcumin alleviates cisplatin-induced learning and memory impairments. Neurobiol Learn Mem 2015;123:43–9 [DOI] [PubMed] [Google Scholar]
- 40.Stein LR, Wozniak DF, Dearborn JT, Kubota S, Apte RS, Izumi Y, et al. Expression of Nampt in hippocampal and cortical excitatory neurons is critical for cognitive function. J Neurosci 2014;34:5800–15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wang G, Li S, Gilbert J, Gritton HJ, Wang Z, Li Z, et al. Crucial Roles for SIRT2 and AMPA Receptor Acetylation in Synaptic Plasticity and Memory. Cell Rep 2017;20:1335–47 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Cameron HA, McKay RD. Adult neurogenesis produces a large pool of new granule cells in the dentate gyrus. J Comp Neurol 2001;435:406–17 [DOI] [PubMed] [Google Scholar]
- 43.Garrido A, Djouder N. NAD(+) Deficits in Age-Related Diseases and Cancer. Trends Cancer 2017;3:593–610 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.