

Abstract
Arbuscular mycorrhiza (AM) are mutualistic interactions formed between soil fungi and plant roots. AM symbiosis is a fundamental and widespread trait in plants with the potential to sustainably enhance future crop yields. However, improving AM fungal association in crop species requires a fundamental understanding of host colonisation dynamics across varying agronomic and ecological contexts. To this end, we demonstrate the use of betalain pigments as in vivo visual markers for the occurrence and distribution of AM fungal colonisation by Rhizophagus irregularis in Medicago truncatula and Nicotiana benthamiana roots. Using established and novel AM-responsive promoters, we assembled multigene reporter constructs that enable the AM-controlled expression of the core betalain synthesis genes. We show that betalain colouration is specifically induced in root tissues and cells where fungal colonisation has occurred. In a rhizotron setup, we also demonstrate that betalain staining allows for the noninvasive tracing of fungal colonisation along the root system over time. We present MycoRed, a useful innovative method that will expand and complement currently used fungal visualisation techniques to advance knowledge in the field of AM symbiosis.
Arbuscular mycorrhiza are mutualistic interactions formed between soil fungi and plant roots. This study presents the MycoRed system, which uses red plant pigments derived from beetroot to reveal how fungi establish symbiosis with living legume and wild tobacco roots.
Introduction
Arbuscular mycorrhiza (AM) fungi of the subphylum Glomeromycotina are soil fungi that engage in symbiosis with land plants [1]. Symbiotic associations with AM fungi date back to over 400 million years ago and can be formed by 70% to 72% of extant land plant species [2–4]. AM fungi are obligate biotrophs that receive all their carbon intake from the plant, which is estimated at up to 20% of the plant’s photosynthate [5]. In exchange, the fungus assists the plant with the acquisition of mineral nutrients, mainly phosphorus, whose availability in soils is often a limiting factor for plant growth [6]. Phosphorus contribution through the mycorrhizal pathway can be very high, and, in some instances, can account for the entire phosphorus consumption of a plant [7]. During AM symbiosis, fungal hyphae form dichotomously branched structures, named arbuscules, within root cortex cells. Hyphal extension and arbuscule formation are accompanied by the de novo extension of a specialised plant cell membrane that separates the fungal hyphae from the plant cytoplasm [8]. In order to accommodate arbuscule formation, plant cells undergo a series of changes in gene expression to aid the establishment of symbiosis [9]. Examples of such AM-induced genes include MtPT4 and MtBCP1 in the model legume Medicago truncatula. MtPT4 encodes a phosphate transporter, belonging to the phosphate transporter 1 (PHT1) subfamily, which is exclusively expressed in arbuscule-containing cells [10]. MtPT4 localises to the plant cell membrane surrounding arbuscules and participates in the acquisition of phosphate released by the fungus during symbiosis. MtBCP1 encodes a member of the blue copper protein (BCP) family and is also specifically expressed in regions of the root hosting arbuscule development during AM symbiosis [11]. MtBCP1 expression is strongest in arbuscule-containing cells but can additionally be observed in adjacent cortical cells [11].
The study of AM symbiotic processes involves the detection, visualisation, and quantification of fungal colonisation. Current techniques rely on the specific staining of fungal cell walls through fast, simple, and cost-effective procedures. Commonly employed methods include the use of trypan blue [12], cotton blue and Sudan IV [13], acid fuchsin [14], ink–vinegar [15], or fluorescein-labelled wheat germ agglutinin (WGA) [16]. All of these methods are destructive, requiring the excision and chemical treatment of roots that are typically visualised with light or fluorescent microscopy. On the other hand, nondestructive methods for detection and visualisation of AM symbiosis offer a number of significant research opportunities but often rely on specialised equipment or are only available for certain species. For example, the foliar accumulation of blumenol-derived metabolites can function as a quantitative proxy for AM colonisation in a number of crop and model plants, with potential applications in field-based quantitative trait locus (QTL) mapping of AM fungi-related genes [17]. However, blumenol accumulation is not visible, and its detection requires specialised extraction and quantification steps. Furthermore, some cereal crops and species of the Liliaceae and Fabaceae naturally produce apocarotenoid yellow pigments in roots upon mycorrhizal colonisation [18,19], which has been useful, for example, for the identification of maize mutants affected in symbiotic interaction [20]. To date, however, the use of natural pigments as visual markers of AM symbiosis is limited to select species within these families and has not been implemented in other crops and model plants.
Betalains are naturally occurring tyrosine-derived water-soluble pigments, comprising yellow to orange betaxanthins and red to purple betacyanins. Betalains have a number of bio-industrial applications, as natural food colourants and antioxidants [21], and as biosensors [22–26]. Betalains were first discovered in plants, where they are unique to the flowering plant order Caryophyllales [27], but have also been reported in the proteobacterium Gluconacetobacter diazotrophicus [28] and the fungi Amanita [29] and Hygrocybe [30]. The betalain biosynthesis pathway in Caryophyllales consists only of 3 main enzymatic steps with additional glycosylation and spontaneous chemical reactions (Fig 1). Initially, tyrosine undergoes an hydroxylation reaction to 3,4-dihydroxy-ʟ-phenylalanine (ʟ-DOPA) catalysed by members of the CYP76AD family of cytochrome P450 enzymes (CYP76AD1/5/6 in Beta vulgaris) [31,32]. ʟ-DOPA is then cleaved by the action of the ʟ-DOPA-4,5-dioxygenase (DODA) enzyme to form a 4,5-secodopa intermediate that spontaenously cyclises to betalamic acid [33]. Betalamic acid is the central betalain chromophore and can spontaneously condense with amino groups to produce yellow betaxanthins [34]. Alternatively, betalamic acid can spontaneously condense with cyclo-dihydroxyphenylalanine (cyclo-DOPA) to produce the red betacyanin—betanidin. Cyclo-DOPA is obtained via oxidation of ʟ-DOPA, catalysed by CYP76AD1 [35]. Glycosylation of betacyanins is common and can occur at either the betanidin [36] or cyclo-DOPA stages [37], with the latter catalysed by the enzyme cyclo-DOPA 5-O-glucosyltransferase (cDOPA5GT). Betanin is the glycosylated form of betanidin.
Fig 1. The betalain biosynthetic pathway.
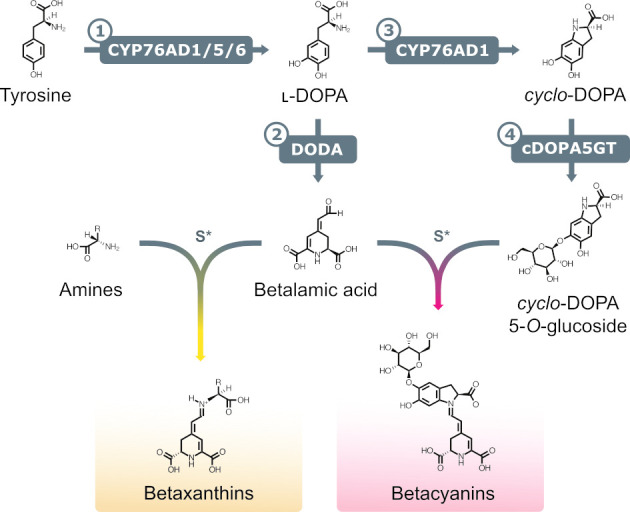
A simplified schematic representation of the main enzymatic and spontaneous reactions leading to the formation of red/purple betacyanins and yellow betaxanthins. Enzymatic steps: (1) tyrosine hydroxylation to ʟ-DOPA catalysed by CYP76AD cytochrome P450 enzymes; (2) cleavage of ʟ-DOPA to form betalamic acid by DODA; (3) ʟ-DOPA oxidation to cyclo-DOPA by CYP76AD1; and (4) cyclo-DOPA glycosylation to cyclo-DOPA 5-O-glucoside by the enzyme cDOPA5GT. S*, spontaneous reaction. Betacyanins are represented as a molecule of betanin. cDOPA5GT, cyclo-DOPA 5-O-glucosyltransferase; cyclo-DOPA, cyclo-dihydroxyphenylalanine; DODA, ʟ-DOPA-4,5-dioxygenase; ʟ-DOPA, 3,4-dihydroxy-ʟ-phenylalanine.
Elucidation of these core enzymatic steps has enabled the engineering of the betalain biosynthetic pathway in a wide range of heterologous hosts, including microbes such as Saccharomyces cerevisiae [38]; plant model organisms like Arabidopsis thaliana, Nicotiana tabacum, and Petunia hybrida [31,39,40]; and a diversity of crops such as Oryza sativa (rice), Solanum lycopersicum (tomato), Solanum tuberosum (potato), and Solanum melongena (aubergine) [40,41]. Betalains have been used as biosensors in a number of heterologous contexts to report increased production of metabolites, such as tyrosine, dopamine, and ʟ-DOPA in Escherichia coli and Nicotiana benthamiana [23,24,26], to measure metabolic flux between competing pathways in the synthesis of benzylisoquinoline alkaloids (BIAs) in S. cerevisiae [22] and for the detection of copper by heavy metal–resistant bacteria in bioremediation processes [25]. In plants, specific promoters have been successfully used to target betalain production in specific tissues such as fruits and seed endosperm [40–42]. Expressing the betalain biosynthetic genes under the control of the AtYUC4 promoter in A. thaliana resulted in pigment production in tissues likely to present auxin biosynthesis activity [42]. Similarly, the use of the DR5 synthetic auxin-responsive promoter in O. sativa calli allowed for easier selection of transformed calli in in vitro transformation protocols [42]. The use of betalains as in vivo reporters offer a number of advantages: (1) the relative simplicity of betalain biosynthesis; (2) the potential for heterologous betalain expression in phylogenetically diverse hosts; and (3) the ease of betalain visualisation and quantification.
Here, we present MycoRed, a betalain-based in vivo and noninvasive reporter system for the occurrence and progression of AM symbiosis in roots of the two model species M. truncatula and N. benthamiana. We leveraged known AM-responsive genes from M. truncatula to identify orthologous N. benthamiana promoters that are similarly responsive to AM fungal colonisation. Heterologous expression of betalain biosynthesis genes specifically driven by AM-responsive promoters effectively tracked AM colonisation dynamics in both species. Collectively, our work demonstrates the efficacy of betalain pigments as reliable in vivo visual markers for the previously inaccessible dynamic tracing of AM symbiosis within root systems, thereby providing a valuable resource for the plant–microbe research community.
Results
Betalains can be used to visualise AM fungus colonisation in living Medicago truncatula roots
To establish a reporter system that allows for the noninvasive visualisation of AM fungal colonisation in roots, we explored the use of betalain biosynthesis genes under the control of AM symbiosis–specific plant promoters. In M. truncatula, PT4 and BCP1 are specifically expressed during root colonisation by AM fungi and are often used as markers to quantify the extent of AM symbiosis [10,11]. We generated T-DNA constructs harbouring CYP76AD1, the enzyme catalysing steps 1 and 3 of the betalain biosynthetic pathway (Fig 1), driven by either MtPT4 or MtBCP1 symbiotic-specific promoters.The T-DNA also carried genes encoding the two additional enzymes needed to produce betalains: DODA and cDOPA5GT (steps 2 and Fig 1) under 35S and Ubi10 constitutive promoters, respectively. We decided to use different promoters in order to avoid transcriptional gene silencing induced by sequence repeat. We refer to these multigene vectors as MtPT4-p1 and MtBCP1-p1 (Fig 2A). Next, we generated M. truncatula composite plants expressing these constructs in hairy roots. Resultant MtPT4-p1 and MtBCP1-p1 transgenic roots did not display visible pigments prior to AM inoculation. After 4 weeks of colonisation with the AM fungus Rhizophagus irregularis, AM symbiosis marker genes were transcriptionally induced, and the roots produced visibly red-coloured root (Figs 2B–2G and S1). We did not observe any pigment production or AM marker gene induction in mock conditions for MtPT4-p1 or MtBCP1-p1 composite roots. Dissection and microscopy of pigmented and nonpigmented root fragments revealed that betacyanin production consistently colocalised or was adjacent to arbuscule-containing cells both in MtPT4-p1 and MtBCP1-p1 roots (Fig 3). Pigment presence often extended beyond arbuscule-containing cells in the inner cortex and was also observed in the endodermis, the pericycle, the stele, and, in some cases, adjacent cortical cells, which represent root tissue layers that did not show any intracellular fungal structures (Fig 3). To confirm that the employed promoter fragments were indeed associated with intracellular fungal structures, we generated MtPT4 and MtBCP1 GUS fusions and expressed them in roots of M. truncatula composite plants (S2 Fig). Staining pattern after fungal colonisation showed that prominent GUS substrate accumulation was limited to inner cortical cells colonised by the fungus, as previously reported [10,11]. Therefore, red betacyanin distribution extends to cells beyond those with promoter activity but still specifically labels colonised tissues and is thus a promising candidate for the in vivo visualisation of AM symbiosis processes.
Fig 2. Betalains can be produced in Medicago truncatula roots as a response to AM fungi colonisation.
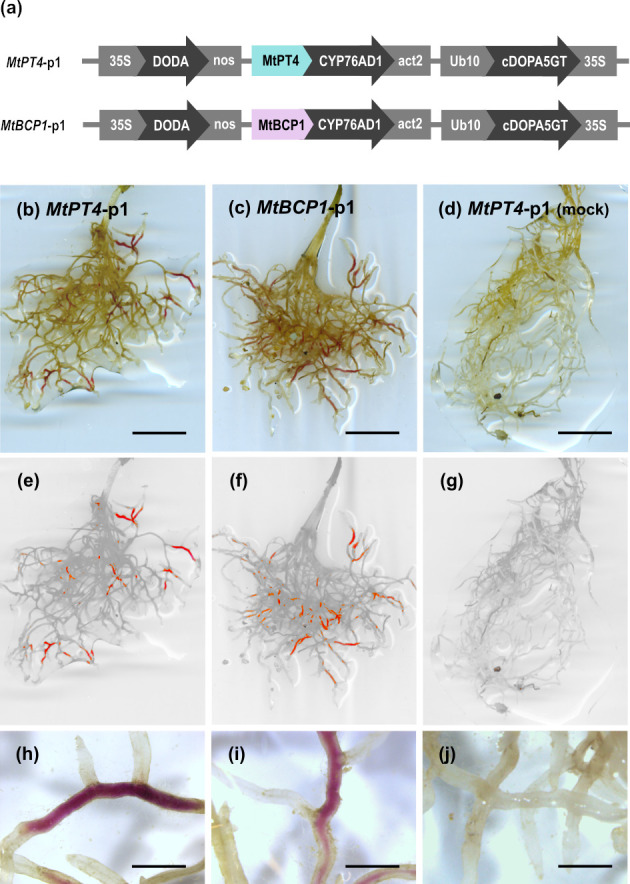
(a) Schematic of the multigene vectors constructed for inducible betalain expression in M. truncatula roots where only the first gene of the betalain biosynthesis pathway is controlled by AM symbiosis–specific promoters. Expression of MtPT4-p1 (b, e, and h) and MtBCP1-p1 (c, f, and i) in roots of M. truncatula 4 weeks after inoculation with Rhizophagus irregularis. (d, g, and j) Example of MtPT4-p1 expressing root system mock inoculated with autoclaved R. irregularis. (e, f, g) Root system images filtered for red colouring only. Scale bar (b, c, and d), 1 cm; scale bar (h, i, and j), 1.5 mm. AM, arbuscular mycorrhiza.
Fig 3. Betalain accumulation is observed at different intensities in different tissue layers and appears higher in the endodermal cells adjacent to arbuscule-containing cortical cells.

Root sections of Medicago truncatula expressing MtPT4-p1 (a and c) and MtBCP1-p1 (b and d) 4 weeks after inoculation with Rhizophagus irregularis. (c and d) Left: Betacyanin pigments are visible in red; right: WGA-FITC staining of fungal structures in blue. Open arrows mark internal hyphae, and filled arrows signal cells containing arbuscules. Scale bar, 100 μm. WGA, wheat germ agglutinin.
Identification and validation of AM symbiosis marker genes in Nicotiana benthamiana
Transiently transformed M. truncatula hairy roots display varying degrees of transgene expression that could impact reporter intensity and functionality. To avoid this, we sought to establish betalains as reporters of AM symbiosis in stable transgenics. Here, we explored the classic model species N. benthamiana, which has been extensively used to develop methods of heterologous betalain production [23,31,43]. We first identified homologues of MtPT4 (NbPT5b; Niben101scf02726g00004.1) and MtBCP1 (NbBCP1b; Niben101Scf07438g04015.1) in N. benthamiana via BLASTP against the proteins from the most recent genome sequence (v1.01, Solgenomics) and confirmed the homology of the N. benthamiana to the M. truncatula loci, by phylogenetic analysis of the BCP and PT orthogroups (S3 Fig). To confirm inducibility of the NbPT5b and NbBCP1b genes during mycorrhizal symbiosis, we investigated their expression at different time points on colonised and non-colonised roots of N. benthamiana with R. irregularis (Fig 4A). We used the R. irregularis β-tubulin (RiBTub) gene as a fungal biomass marker to confirm successful colonisation. Expression of NbPT5b and NbBCP1b increased after 2 weeks and showed significantly elevated transcript levels 3 to 4 weeks postinoculation (Fig 4A). We therefore cloned promoter regions up to −1,068 bp and −1,231 bp upstream of the start of NbPT5b and NbBCP1b coding sequences, respectively (S1 Data). To address tissue specificity of NbPT5b and NbBCP1b promoters, we generated promoter–GUS fusions for expression in N. benthamiana. Stable N. benthamiana transformants expressing NbPT5b-GUS and NbBCP1b-GUS fusions displayed a GUS staining pattern specific to root areas colonised by R. irregularis (Fig 4B and 4C). In both cases, GUS staining was stronger in cells containing arbuscules but could also be observed in adjacent cells at varying intensities (Fig 4B and 4C). Non-colonised areas were consistently free of staining in NbPT5b-GUS and NbBCP1b-GUS roots. Altogether, the results support the use of NbPT5b and NbBCP1b promoters for the construction of a betalain-based AM symbiosis reporter system in N. benthamiana.
Fig 4. Expression of NbPT5b and NbBCP1b genes is induced under colonisation with Rhizophagus irregularis in Nicotiana benthamiana.
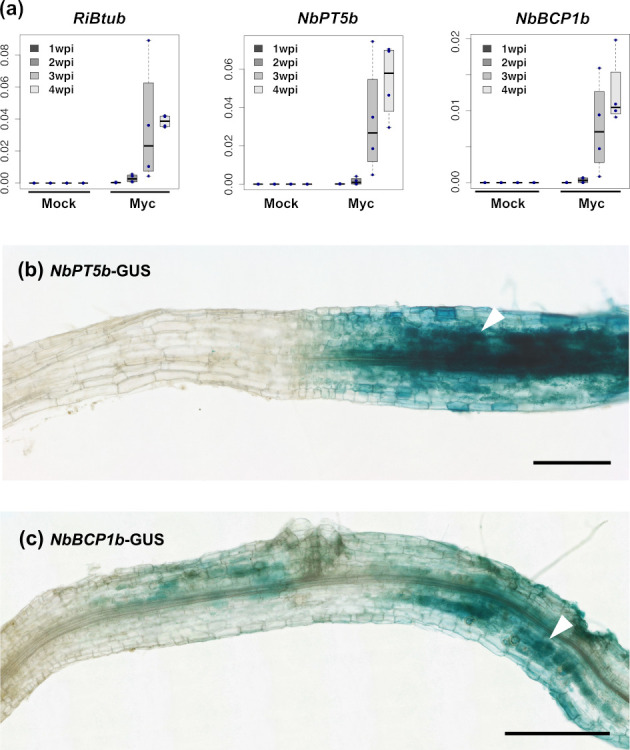
(a) qRT-PCR analysis of NbPT5b and NbBCP1b shows that gene expression is induced 2 wpi with R. irregularis and increases with time and degree of colonisation. Y axes indicate expression levels relative to N. benthamiana’s elongation factor 1 alpha (NbEF). Expression of RiBTub is used as a fungal biomass marker for root colonisation. Raw qRT-PCR values can be found in S3 Data. (b and c) GUS staining of NbPT5b-GUS and NbBCP1b-GUS expressing N. benthamiana roots 4 weeks after inoculation with R. irregularis. GUS activity can only be observed in root areas colonised by R. irregularis and is more predominant in arbuscule-containing cells (white arrows). Scale bar, 500 μm. qRT-PCR, quantitative real-time polymerase chain reaction; RiBTub, R. irregularis β-tubulin; wpi, weeks postinoculation.
Betalains visualise AM fungus colonisation in living Nicotiana benthamiana roots
We then generated multigene betalain reporter constructs for stable transformation in N. benthamiana. A first generation of vectors placed CYP76AD1 under the control of the NbPT5b or NbBCP1b promoters, with DODA and cDOPA5GT driven by 35S and Ubi10 constitutive promoters, respectively. Hereafter, we refer to these multigene vectors with one AM fungal colonisation-specific promoter as NbPT5b-p1 and NbBCP1b-p1 (Fig 5A). We generated 16 independent transgenic N. benthamiana NbPT5b-p1 lines and 12 independent NbBCP1b-p1 lines. NbPT5b-p1 and NbBCP1b-p1 plants produced betalains in the roots upon colonisation with R. irregularis (Fig 5B and 5C). Betalain production was not visible in mock conditions for NbPT5b-p1 or NbBCP1b-p1 plants inoculated with autoclaved R. irregularis (S4 Fig). Dissection and microscopy of pigmented and nonpigmented root fragments revealed betalain presence colocalised with fungal colonisation structures (Fig 5D–5G).
Fig 5. Betalains can be produced in Nicotiana benthamiana roots as a response to AM fungi colonisation.

(a) Schematic of the multigene vectors constructed for inducible betalain expression in N. benthamiana roots where only the first gene of the betalain biosynthesis pathway is controlled by AM symbiosis–specific promoters. Expression of NbPT5b-p1 (b) and NbBCP1b-p1 (c) in whole root systems of N. benthamiana T1 plants 4 weeks after inoculation with Rhizophagus irregularis. Right: Root images are filtered for red colour. (d–g) Confocal microscopy of red NbPT5b-p1 (d and e) and NbBCP1b-p1 (f and g) transgenic root sections which were cut and stained with WGA-FITC to visualise fungal structures. (d and f) colour-filtered for red; (e and g) FITC green fluorescence. Scale bar (b and c), 1 cm; scale bar (d–g), 100 μm. AM, arbuscular mycorrhiza; WGA, wheat germ agglutinin.
To assess the extent to which red pigment co-occurs with intraradical fungal structures, we divided NbPT5b-p1 and NbBCP1b-p1 T1 plant root systems into red-pigmented and nonpigmented fragments and ink stained them separately for the presence of fungal structures (S5 Fig). We then imaged the stained root fragments and divided each root image into sections of similar length for the scoring of fungal presence. For both NbPT5b-p1 and NbBCP1b-p1 root systems, we found that all of the betacyanin-positive root fragments contained fungal structures and were extensively colonised (Table 1). White root segments of the NbPT5b line were overall much less likely to harbour fungal structures (max. 22.9% intraradical hyphae) compared to NbBCP1b (88.2%). Furthermore, the extent of fungal structures within individual white root segments was also much lower in NbPT5b than in NbBCP1b roots (4.8% versus 46.4%)(Table 1). This suggests that NbPT5b-driven CYP76AD1 more reliably labels root segments containing fungal structures. Red root sections predominantly displayed colonisation by arbuscules and hyphae, and, to a lesser extent, vesicles, suggesting that accumulation of pigment in N. benthamiana is also associated with fully developed AM symbiosis but may decline at late stages (Table 1).
Table 1. Quantification of fungal structures observed in roots of Nicotiana benthamiana expressing NbPT5b-p1 and NbBCP1-p1 4 wpi with Rhizophagus irregularis.
| Genotype | Pigmented | N | Percent of colonised root fragments (%) | Average length of root fragments colonised (%) | ||||
|---|---|---|---|---|---|---|---|---|
| IH | A | V | %IHC | %AC | %VC | |||
| NbPT5b-p1 | + | 45 | 100.0 | 100.0 | 97.8 | 77.9 ± 3.5 | 74.7 ± 3.9 | 62.3 ± 4.1 |
| - | 35 | 22.9 | 17.1 | 14.3 | 4.8 ± 2.2 | 3.0 ± 1.4 | 1.1 ± 0.5 | |
| NbBCP1b-p1 | + | 31 | 100.0 | 100.0 | 100.0 | 93.1 ± 2.4 | 86.6 ± 3.4 | 78.5 ± 4.0 |
| - | 34 | 88.2 | 70.6 | 67.6 | 46.4 ± 5.7 | 33.6 ± 5.2 | 21.4 ± 4.3 | |
We cut and divided roots into pigmented (red rows) and nonpigmented (noncoloured rows) fragments for ink staining. N refers to the total number of root fragments analysed for each condition. Table shows the number of root fragments containing fungal structures and the average extent of colonisation by these structures over the length of the root fragment. Error is shown as standard error.
%AC, percentage of arbuscule colonisation; %IHC, percentage of internal hyphae colonisation; %VC, percentage of vesicle colonisation; A, arbuscules; IH, internal hyphae; V, vesicles; wpi, weeks postinoculation.
Stable expression of NbPT5b-p1 and NbBCP1b-p1 can cause shoot developmental defects in N. benthamiana
Surprisingly, many NbPT5b-p1 and NbBCP1b-p1 transgenic lines displayed vegetative developmental defects of varying severity (S6 Fig). Specifically, 13 out of 16 NbPT5b-p1 and 11 out of 12 NbBCP1-p1 lines were affected with moderate to severe defects. Aberrant phenotypes were not visible in early stages of development but became prominent during maturation and flowering. Affected plants were smaller and displayed altered leaf morphology, ranging from curvy leaf edges to acute deformation of leaf shape and thickness (S6 Fig). We also observed defects in flower morphology, including stunted or absent perianth, and, in more severe cases, missing floral reproductive organs (S6 Fig). Only plants with mild to no phenotypic alterations were able to set seed, although seed set was not abundant. To address whether these phenotypes were linked to the expression of constitutive or AM-specific promoter–driven betalain genes, we performed semiquantitative reverse transcription PCR (RT-PCR) on abnormal leaves of NbPT5b-p1 transgenic plants. As expected, we detected 35S promoter-driven DODA expression and cDOPA5GT in all analysed T0 lines, but, unexpectedly, DODA expression was increased in severely affected lines (S6 Fig). Surprisingly, we detected expression of AM-responsive PT5b promoter-driven CYP76AD1 in T0 lines with light and moderate developmental defects (S6 Fig). Expression intensity of CYP76AD1 appeared to decrease with phenotype severity and was barely detectable in lines displaying acute developmental defects. T1 seedlings from various NbPT5b-p1 transformant lines developed smaller leaves and shorter roots than wild-type (WT) seedlings (S7 Fig). T1 plants were nevertheless able to grow to adult stage, flower, and set seed. In addition to developmental defects, we also detected unanticipated accumulation of betanin by HPLC in leaves of T1 plants that descended from NbPT5b-p1 lines with mild to no phenotypic alterations (S8 Fig). Leaf betanin accumulation arose in both inoculated and mock conditions (S8 Fig). In summary, we found that expression constructs with 2 constitutive biosynthesis genes frequently resulted in both developmental aberrations and unexpected aerial betalain expression.
Expression of all betalain synthesis genes under AM symbiosis–specific promoters ameliorates defective phenotypes in N. benthamiana
To avoid the developmental defects associated with the constitutive expression of betalain genes in N. benthamiana, we sought to express all 3 betalain biosynthesis genes under AM symbiosis–specific promoters. Here, a second generation of vectors placed CYP76AD1, DODA, and cDOPA5GT under the control of either the NbPT5b or NbBCP1 promoters. Hereafter, we refer to these multigene vectors as NbPT5b-p3 and NbBCP1b-p3 (Fig 6A). To test their performance, we generated 11 independent transgenic NbPT5b-p3 lines and 7 NbBCP1b-p3 lines and grew them with R. irregularis spores in a rhizotron setup. Root system pigmentation patterns of the NbPT5b-p3 and NbBCP1b-p3 reporter constructs performed similarly to NbPT5b-p1 and NbBCP1b-p1 (S9 Fig). Colonisation extent and fungal structures frequency in inoculated root systems did not differ between WT and NbPT5b-p3 and NbBCP1b-p3 reporter lines (S10 Fig). Once again, plants grown in mock conditions did not harbour any observable colouration despite being grown in non-sterile substrate, further suggesting that reporter expression is not activated by the presence of other microorganisms. This is also supported by the absence of red pigmentation upon inoculation of reporter lines with spores of the oomycete root pathogen Phytophthora palmivora (S11 Fig).
Fig 6. Betalain production in Nicotiana benthamiana roots as a response to AM fungi colonisation via expression of the entire betalain pathway under AM symbiosis–specific promoters.

(a) Schematic of the multigene vectors constructed for inducible betacyanin expression in N. benthamiana roots with the 3 betalain biosynthetic genes controlled by the NbPT5b or the NbBCP1 promoters. Expression of NbPT5b-p3 (b and c) and NbBCP1-p3 (d and e) in roots of N. benthamiana 4 weeks after inoculation with Rhizophagus irregularis. Betacyanin production was not visible in mock conditions for any NbPT5b-p3 or NbBCP1b-p3 roots. (c and e) are a magnification of the areas delimited by the dashed squares. Scale bar (b and d), 500 μm; scale bar (c and e), 100 μm. AM, arbuscular mycorrhiza; Ar, arbuscules; en, endodermis; ep, epidermis; ih, internal hyphae; v, vesicle.
Dissection and microscopy of pigmented and nonpigmented NbPT5b-p3 and NbBCP1b-p3 root fragments inoculated with R. irregularis revealed once more that betalain presence colocalised with fungal colonisation structures (Fig 6). Arbuscule formation could be observed in all cortex cell layers in N. benthamiana. In NbPT5b-p3 and NbBCP1b-p3 roots, betalain pigmentation was confined to single cells that were clearly distinguishable. Pigmentation was, however, not restricted to arbuscule-containing cells and could also be detected in adjacent cells with no apparent arbuscule structures including cells of the endodermis (Fig 6). Similarly, some arbuscule-containing cells in the proximity of red pigmented cells appeared unpigmented both in NbPT5b-p3 and NbBCP1b-p3 roots (Fig 6). Importantly, only 1 out of the 11 lines in the NbPT5b-p3 genotype and 2 out of the 7 lines in the NbBCP1b-p3 genotype displayed the developmental defects seen with NbPT5b-p1 and NbBCP1b-p1 reporter constructs. Finally, the T1 generations derived from NbPT5b-p3 and NbBCP1b-p3 lines did not exhibit any discernible pigmentation in shoot tissues. Therefore, the expression of all betalain biosynthetic pathway genes under AM-specific promoters ameliorates the incidence of developmental alterations and nonspecific betalain production in N. benthamiana plants.
We also confirmed the functionality of constructs where all betalain synthesis genes are driven by arbusculated cell specific promoters in M. truncatula. We used the MtPT4 promoter to drive the expression of betalain genes and included a constitutively expressed DsRed marker to facilitate selection of transformed hairy roots. Betalain colouration was produced in colonised roots and was associated with arbuscule presence (S12 Fig). However, once again, staining was not restricted to arbuscule-containing cells of the inner cortex and could be mainly observed in internal tissue layers like the endodermis, pericycle, and stele (S12 Fig). M. truncatula Doesn’t Make Infections 3 (dmi3) mutants fail to progress in intraradical AM fungal colonisation and are unable to engage in symbiosis [44,45]. Therefore, reporter expression in dmi3 mutants should not harbour any colouration regardless of inoculum presence. Notably, red pigmentation was not observed in composite roots of M. truncatula dmi3 mutants (S13 Fig). The efficiency of betalain accumulation as a marker of AM colonisation with MtPT4 was similar to that observed for NbPT5b in N. benthamiana plants (S14 Fig and S1 Table).
Promoter-controlled betalain biosynthesis allows for dynamic tracing of root colonisation processes
Expression of the betalain biosynthesis genes under the control of NbPT5b and NbBCP1b promoters in N. benthamiana also allowed for the noninvasive imaging of AM fungal root colonisation in root systems. We grew NbPT5b-p3 and NbBCP1b-p3 transgenic N. benthamiana lines in a vertical rhizotron-based setup with R. irregularis spores that maintained root system architecture by separating roots from the substrate with a porous black cloth. This also allowed for nondisruptive imaging on a flatbed scanner and subsequent image analysis using a selective colour filter for red pigment. At 52 days postinoculation (dpi), plants had developed significant root stretches with red pigmentation in both NbPT5b-p3 and NbBCP1b-p3 lines that was easily discriminated by computer imaging (Fig 7).
Fig 7. Red pigment distribution in root systems of NbBCP1b-p3 and NbPT5b-p3 Nicotiana benthamiana plants colonised by Rhizophagus irregularis.

Images were taken at 52 dpi. (a and c) are reflective light images, and (b, d, and e) are filtered for red colouring only. (e) High magnification image of (d) showing varying red colouration in parallel running roots. Scale bar = 1.3 cm. dpi, days postinoculation.
To assess AM colonisation dynamics over time, we also acquired consecutive images of intact NbBCP1b-p3 and NbPT5b-p3 rhizotrons at different days. This revealed increasing red pigment development within the root systems (Figs 8 and S15). Betalain pigmentation served as a reliable and long-lasting indicator of AM colonisation, as we did not observe root stretches that displayed red pigmentation and later lost it. Together, these data demonstrate that red pigmentation effectively traces fungal colonisation over time allowing for the dynamic and noninvasive assessment of root sections where the AM-responsive promoters have been activated.
Fig 8. NbBCP1b promoter-controlled betalain biosynthesis allows for dynamic tracing of root colonisation processes in Nicotiana benthamiana.

Transgenic NbBCP1-p3 plants grown in a rhizotron setup supplied with Rhizophagus irregularis spore inoculum imaged over time. (a) Reflective light images, (b) represent the same images filtered for red/magenta hues, and (c) is a magnification of the area delimited by the dashed square over time. Scale bar, 1 cm. dpi, days postinoculation.
Discussion
AM symbiosis is a fundamental and widespread trait in plants that greatly expands the root surface area for nutrient uptake. AM symbiosis is consequently a key agronomic trait in the drive to enhance future crop yields through environmentally sustainable mechanisms. However, enhancing AM fungal association in crop species requires the tools to understand its fundamental dynamics across varying agronomic and ecological contexts. To this end, we demonstrate the use of betalain pigments as in vivo visual markers for the occurrence and distribution of AM fungal colonisation in the roots of M. truncatula and N. benthamiana. We have generated multigene vectors in which AM colonisation specific plant promoters control the expression of core betalain synthesis enzymes in the production of betalain pigments. We show that AM-specific promoter-controlled betalain pigmentation is a powerful macroscopic tool to report and trace fungal colonisation in vivo along the root (Fig 9).
Fig 9. Betalains can be used as markers for AM colonisation in plant roots.

Left: Red pigmentation is easily observable in whole plant root systems. Right: Red pigmentation is most prominent in colonised tissues as well as in adjacent tissue layers.
Macroscopically, betalain colouration was specifically limited to regions of the root colonised by R. irregularis in both M. truncatula and N. benthamiana, as we detected few to no false positives. Furthermore, no betalain pigmentation was detected in mutant lines of M. truncatula impaired in AM symbiosis. Quantification of fungal structures revealed that pigmented root fragments were extensively colonised, showing arbuscule-containing cells over the majority of the root length for all tested promoters. We also observed some fungal structures in a low percentage of nonpigmented root fragments. In these cases, arbuscule colonisation was restricted over the length of the analysed root fragments, with the exception of NbBCP1b expressing N. benthamiana roots that exhibited a greater degree of unpigmented yet colonised root fragments. Thus, reporter ability to document the totality of colonisation events depends on the promoter chosen for vector construction. It appears that the NbBCP1b promoter-containing constructs are not equally activated in all colonisation events, or perhaps pigment accumulation becomes evident only at certain stages of colonisation. Promoter sequences of PT4 homologues such as NbPT5b were able to report the majority of colonisation events in the root system and are therefore preferred for reporting total root colonisation. A future solution to document total AM colonisation could involve the establishment of systems whereby the betalain biosynthesis genes are activated by transactivators, which could be then driven by promoters that are active at early, main, and late AM fungal colonisation stages [46–48]. Overall, betalain pigmentation effectively reports fungal colonisation with very little to no error and allows for simple selection of colonised root areas.
Microscopy of pigmented tissues revealed that betalain accumulation extended beyond arbusculated cells and into adjacent cell layers in both M. truncatula and N. benthamiana. In M. truncatula, betalain pigmentation was most prominent in the endodermis and pericycle cells adjoining arbusculated inner cortical cells (Fig 3). In N. benthamiana, we also observed betalain pigmentation in the endodermis and pericycle cells, yet pigmentation was more strongly retained in cortical cells (Fig 6). However, colouration in N. benthamiana was sometimes present in non-arbusculated cells adjacent to arbusculated cortical cells, consistent with previously observed GUS staining patterns. These extended patterns occur even when using constructs where all three biosynthesis enzymes are driven by AM-specific promoters and cannot therefore be attributed to constitutive gene expression artefacts. Such pigment accumulation in non-arbusculated cells may be the result of a former colonisation or of moderate degree of pigment migration. Betalains are water-soluble pigments and, in native betalain-pigmented species, are produced in the cytoplasm and then stored in vacuoles [49], although the mechanisms responsible for the intracellular transport of betalains are unknown. As small water-soluble compounds, betalains may have the potential to move symplastically through root plasmodesmata, although this remains unproven. Transient expression of betalains in leaves of N. benthamiana leads to macroscopically well delimited pigmented areas [23,31,43], but cellular migration across the boundary tissue has not been studied. Unpigmented cells harbouring arbuscules could also be a result of mechanical disruption during the sectioning of root tissues for microscopy, where cells that previously contained betalains lose pigmentation after being sliced open. Nevertheless, the betalain reporter system remains a highly effective marker of AM colonisation, especially at the macroscopic level.
The betalain biosynthetic pathway has been previously constitutively expressed in several plant species, including plants of the Solanaceae, with no report of observable developmental defects [31,40]. Yet, in our study, expression of betalain producing constructs where only CYP76AD1 was under the control of AM-specific promoters led to developmental defects in N. benthamiana lines. Affected plants displayed dwarf phenotypes, altered leaf, and flower morphology and were unable to set seed (S6 Fig). Our findings could be explained by a mechanism whereby constitutive expression of either, or both, DODA and cDOPA5GT in absence of CYP76AD1 expression cause developmental abnormality. Three observations are consistent with this hypothesis: (1) expression of constructs containing all three biosynthesis enzymes under symbiosis specific promoters substantially decreased the number of affected plants; (2) DODA expression appeared enhanced in leaves of severely affected plants; and (3) expression of CYP76AD1 was detected in leaves of plants with mild defects and seemed to decrease with phenotype severity. We speculate that, in absence of ʟ-DOPA, the DODA enzyme could be promiscuously acting on alternative N. benthamiana substrates, with negative developmental consequences. This effect is alleviated by the presence of CYP76AD1, which provides the DODA enzyme with an abundance of its native substrate, ʟ-DOPA, resulting in the production of inert betalain pigmentation. Constitutive DODA expression may therefore create a shoot developmental conflict that can be partially mitigated by compensatory expression of CYP76AD1. Our selection process could therefore have been biased towards T0 plants with a degree of escaped CYP76AD1 expression, which would also explain the presence of betanin in a number of T1 plants descending from CYP76AD1 shoot-expressing lines (S9 Fig). Further experimentation is required to support this hypothesis, but in any case, developmental defects and vegetative betanin expression can be avoided when all three biosynthesis enzymes are driven by AM symbiosis–specific promoters.
The most powerful application of our betalain-based AM reporter lies in its ability to noninvasively document colonisation in fully developed root systems over time (Figs 8 and S15). This will facilitate answering important questions in the field of AM symbiosis and its potential for agronomic improvement. These include understanding the dynamics of root system colonisation, including the time difference between lateral root emergence and its colonisation, or the differential colonisation susceptibility of different root orders [50]. Colonisation can be assessed in scenarios where plants compete with each other [51] or with shoot pests [52], different nutrient regimes, CO2 concentration, temperatures, soil structures [53], and other abiotic factors. A second important application is in the survey of induced plant genetic variation or fungal natural genetic variation that impacts on root system colonisation. Here, its use is only limited by the transformability of the plant species, and an important future step will be to test our approach in monocot crops, some of which have already been engineered to express betalain pigments in grains [41]. Finally, a third application derives from the ability to bulk collect betalain-pigmented root sections produced under AM fungal colonisation, especially when there is very little overall colonisation as is common in some symbiosis signalling pathway mutants or when colonisation is restricted to specific roots. Here, stage-specific early or late promoters will unlock targeted transcriptomic, proteomic, or metabolomic analysis of colonised roots without the dilution effect of non-colonised tissues. Such enrichment strategies may, for example, help in understanding communication mechanisms between AM fungi and plants. Targeted sampling of pigmented roots will also simplify low-throughput microscopy such as cryo-electron microscopy and drastically improve the signal-to-noise ratio inherent in current sampling processes. Selective sampling at red-to-white transition zones may additionally aid time-lapse microscopy of expanding fungal colonisation arrays within roots.
In summary, betalains are plant pigments with a strong potential as visual markers for the study of physiological and developmental processes in plants and microorganisms. Here, we have expanded the application of these versatile pigments to add to an ever increasing range of betalain-based technologies as reporters of AM fungi colonisation in plant roots. MycoRed complements currently used fungal visualisation techniques and constitutes a powerful tool that will be of great value for the plant–microbe research community to advance knowledge in the field of AM symbiosis.
Materials and methods
Plant material and growth conditions
M. truncatula plants were grown in dried industrial sand at 21°C day temperature, 19°C night temperature, 65% humidity, and a 16-hour photoperiod at 350 μmoles·m−2·s−1 light intensity. N. benthamiana plants belong to a standard laboratory line maintained by selfing. N. benthamiana plants were grown in F2 soil (Levington, Frimley, United Kingdom) in controlled greenhouse facilities maintained at 25°C day temperature, 15°C night temperature, ambient humidity, and a 16-hour photoperiod with 125 w·m−2 of supplementary light. Watering was performed via dripping on capillary mats for 2 to 4 minutes every 6 hours.
Seed material for N. benthamiana MycoRed reporter lines (NbPT5b-p3 and NbBCP1b-p3) are available for shipping. Requests can be sent to corresponding author Dr. Sebastian Schornack.
Plant inoculation with R. irregularis
M. truncatula and N. benthamiana plants were transferred to sand and inoculated with a commercial R. irregularis crude inoculum (consisting of oil-dry substrate, pieces of colonised maize roots, and R. irregularis spores and external hyphae) at a concentration of 1:20 (v/v) inoculum:sand. Plants were watered 3 times per week with 1x Long-Ashton solution (20x concentrate: potassium nitrate, 40 mM; calcium nitrate, 40 mM; monosodium phosphate, 15 mM; magnesium sulfate, 30 mM; manganese sulfate, 0.1 mM; copper sulfate, 0.01 mM; zinc sulfate, 0.01 mM; boric acid, 0.5 mM; sodium chloride, 1 mM; ammonium heptamolybdate, 0.7 μM; ethylenediaminetetraacetic acid ferric sodium salt, 0.6 mM).
RNA extraction and gene expression analysis
N. benthamiana 2-week-old seedlings were inoculated with R. irregularis, and roots were harvested once a week for 4 weeks after inoculation. M. truncatula MtPT4-p1 hairy roots were sampled 4 weeks after inoculation. Mock control plants were inoculated with autoclaved R. irregularis. Total RNA from roots was extracted using the Qiagen RNeasy Mini Kit according to manufacturer’s instructions (QIAGEN, Hilden, Germany). RNA quantity and quality were assessed by NanoDrop (Thermo Fisher Scientific, Waltham, Massachusetts, United States of America) and agarose gel electrophoresis. cDNA libraries were prepared using the iScript cDNA Synthesis Kit (Bio-Rad, Watford, UK) according to manufacturer’s recommendations. Quantitative real-time PCR was performed on 1:20 diluted cDNA in combination with the Roche LightCycler 480 SYBR Green I Master Mix (Roche, Basel, Switzerland) and the oligonucleotide primers listed in S2 Table. Three technical replicates were performed per sample. Transcript levels from N. benthamiana were normalised using N. benthamiana’s elongation factor 1 alpha (NbEF; Niben101Scf04639g06007.1) and the ΔCt method as previously described [54]. Transcript levels from M. truncatula hairy roots were normalised using M. truncatula’s phosphatidylinositol 3- and 4-kinase gene belonging to the ubiquitin family (MtUBQ; Medtr3g091400) [55]. Expression of R. irregularis’ β-tubulin (RiBTub; XM_025314309.1) and elongation factor 1 alpha (RiEF; XM_025321412.1) were used as fungal biomass markers for root colonisation.
Promoter identification and isolation
Homologue colonisation marker genes in N. benthamiana were identified by BLASTP on NCBI with standard parameters using orthologous M. truncatula loci as bait. To confirm the homology of BCP1 and PT loci in N. benthamiana and M. trucatula, orthogroups for BCP and PT gene families were retrieved from phytozome, aligned by codon using MAFFT, and analysed by FastTree. Trees were post-processed to include FastTree support values and to mask monophyletic tips. Leaf tissue from N. benthamiana and M. truncatula A17 plants was snap frozen in liquid nitrogen and stored at −80°C. Frozen tissue was ground to a fine powder using 5-mm glass beads and a Tissue Lyser II homogeniser (QIAGEN). Genomic DNA extraction was performed on up to 100-mg ground tissue using the QIAGEN DNeasy Plant Mini Kit according to the manufacturer’s specifications. Resulting genomic DNA was used to amplify 5′ flanking regions for M. truncatula’s PT4 and BCP1 gene and N. benthamiana’s PT5b and BCP1b genes by PCR using Phusion High-Fidelity DNA polymerase (Thermo Fisher Scientific), with the oligonucleotide primers listed in S2 Table. Primers were designed to include a region of 836 bp upstream the start codon for MtPT4, 1,108 bp for MtBCP1, 1,068 bp for NbPT5b (Niben101Scf02726g00004.1), and 1,231 bp for NbBCP1b (Niben101Scf07438g04015.1). Amplification length for N. benthamiana promoter sequences was based on published promoter sizes in M. truncatula and suitable oligo binding sites [10,11]. PCR products were ligated into the pBlueScript SK cloning vector using T4 DNA ligase (New England Biolabs, Hitchin, UK), and plasmids were sequenced to confirm gene identity. Cloned sequences can be found in S1 Data.
Generation of vectors for plant transformation
The construction of multigene vectors containing the betalain biosynthetic genes under the control of AM-specific promoters was carried out using Golden Gate cloning [56]. Cloning components were acquired from the MoClo Tool Kit [57,58] and the MoClo Plant Parts Kit [59] (Addgene) except for the 1,327 bp A. thaliana Ubiquitin 10 promoter (Ubi10), which was provided by Dr Nicola Patron (Earlham Institute, Norwich, UK). Isolated MtPT4, MtBCP1, NbPT5b, and NbBCP1b promoter regions were cloned into the pICH41295 level 0 accepting vector for promoter + 5′ UTR modules. Cloning proceeded through level 0, level 1, and level 2 modules using the one-pot, one-step type IIS restriction–mediated cloning reaction. GUS reporter vectors were constructed with an eGFP–GUS fusion gene placed under the control of the cloned arbuscular-responsive specific promoters and also included a Basta resistance gene under the nos promoter and terminator. Previously generated plasmids containing the betalain biosynthetic genes were used for the construction of betacyanin producing multigene vectors [23]. DODAα1 and CYP76AD1 sequences were originally isolated from B. vulgaris cDNA libraries, whereas the cDOPA5GT sequence was obtained from flowers of Mirabilis jalapa. Gene sequences had been previously domesticated to remove BsaI and BpiI restriction enzyme sites considering codon usage in N. benthamiana. The first set of multigene vectors created for hairy root transformation of M. truncatula and stable transformation of N. benthamiana contained BvCYP76AD1 under the control of the arbuscular-responsive specific promoters (MtPT4, MtBCP1, NbPT5b, or NbBCP1b), BvDODAα1 and MjcDOPA5GT under constitutive promoters, and included a Basta resistance gene under the nos promoter and terminator. We refer to these multigene vectors as MtPT4-p1, MtBCP1-p1, NbPT5b-p1, and NbBCP1b-p1, respectively (Figs 2A and 5A). The second set of multigene vectors created for root transformation of M. truncatula and stable transformation of N. benthamiana contained BvDODAα1, BvCYP76AD1, and MjcDOPA5GT genes all under the control of the arbuscular-responsive specific promoters (MtPT4, NbPT5b, and NbBCP1b) and included a DSRed fluorescent marker in the M. truncatula vector or a Basta resistance gene in N. benthamiana vectors both under the nos promoter and terminator. We refer to these multigene vectors as MtPT4-p3, NbPT5b-p3, and NbBCP1b-p3, respectively (Figs S12A and 6A). Level 0 and level 1 vectors were confirmed by Sanger sequencing of the full inserts. Level 2 vectors were confirmed by Sanger sequencing of insert boundaries and by diagnostic digestion with restriction enzymes. MycoRed reporter vectors and level 0 plasmids used for their construction are listed in S3 Table. All vectors are available through Addgene (Cambridge, Massachusetts, USA).
Hairy root transformation of M. truncatula
Hairy root transformation of M. truncatula was performed according to the previously described Agrobacterium rhizogenes–mediated method with modifications [60]. Multigene constructs were transformed into the A. rhizogenes ARQUA1 strain and grown at 28°C in LB media supplemented with antibiotics (carbenicillin 12.5 mg/L, kanamycin 50 mg/L) until reaching an OD600 of approximately 1.5. Cultures were then brought to a final OD600 of approximately 1 in sterile distilled water. M. truncatula seeds were sterilised with 96% sulphuric acid for 5 minutes and pure sodium hypochlorite for 5 minutes, both steps followed by 5 to 10 washes with sterile distilled water. After a 2-day stratification process at 4°C, seeds were germinated upside down on square dishes containing wet whatman paper overnight in the dark at 20°C. Hypocotyl tips were then cut, and seedlings were suspended in A. rhizogenes cultures with acetosyringone (0.2 μM). Seed coats were removed, and seedlings were moved to solid mycorrhisation media (potassium nitrate, 80 mg/L; magnesium sulfate, 731 mg/L; potassium chloride, 65 mg/L; monopotassium phosphate, 4.8 mg/L; calcium nitrate, 288 mg/L; manganese (II) chloride, 6 mg/L; boric acid, 1.5 mg/L; zinc sulfate, 2.65 mg/L; sodium molybdate, 0.0024 mg/L; copper sulfate, 0.13 mg/L; potassium iodide, 0.75 mg/L; ethylenediaminetetraacetic acid ferric sodium salt, 8 mg/L; glycine, 3 mg/L; myoinositol, 50 mg/L; nicotinic acid, 0.5 mg/L; pyridoxine hydrochloride, 0.1 mg/L; thiamine hydrochloride, 0.1 mg/L; phytagel, 4 g/L; and pH 5.5) to grow vertically for 7 days at 20°C and indirect lighting. Newly formed roots were then sectioned in half, and seedlings were transferred to mycorrhisation media supplemented with cefotaxime (250 μg/mL) and covered with Whatman’s filter paper. Plants were grown vertically for 4 weeks at 20°C and indirect lighting to allow for transgenic root development. Plants were then transferred to sand and subjected to fungal colonisation with R. irregularis. For constructs containing DsRed, all non-transformed roots were cut off before acclimation and inoculation. DSRed fluorescence was detected with a Leica M165 FC fluorescence dissecting stereomicroscope (Leica Biosystems, Wetzlar, Germany) using a filter with excitation and emission wavelengths of 510 to 560 nm and 590 to 650 nm, respectively.
Stable transformation of N. benthamiana
N. benthamiana stable transformation was performed according to the previously described A. tumefaciens–mediated method with modifications [61]. Fully developed leaves of 4-week-old N. benthamiana plants were agro-infiltrated with A. tumefaciens carrying the constructs of interest according to previously described methods [61]. All constructs were transformed into the Agrobacterium tumefaciens GV3101 strain and grown at 28°C in LB media supplemented with antibiotics (gentamycin 25 mg/L, rifampicin 60 mg/L, kanamycin 50 mg/L) until reaching an OD600 of approximately 1.5. Cultures were then brought to a final OD600 of approximately 0.3 in infiltration media (10 mM MgCl2, 0.2 mM acetosyringone, 10 mM MES at pH 5.6). Infiltration was performed on the abaxial surface of young expanding leaves of 4-week-old N. benthamiana plants. Three days after infiltration, full leaves were excised and cut into 1 to 2 cm2 leaf squares. Leaf explants were sterilised in 70% ethanol for 5 minutes, 20% sodium hypochlorite supplemented with Tween 20 (0.1%) for 20 minutes, and washed thoroughly with sterile water. Sterilised leaf explants were then transferred adaxial side up to fresh selection media (1X Murashige and Skoog basal salt mixture, 1X Gamborg’s B5 vitamins, 1% sucrose, 0.59 g/L MES, 2.0 mg/L BAP, 0.05 mg/L NAA, 0.4% Agargel, pH 5.7) supplemented with cefotaxime (500 mg/L), timentin (320 mg/L), and phosphinothricin (2 mg/L). Explants were subcultured onto new fresh media weekly until the appearance of first shoots, which were excised and planted in rooting media (½ Murashige and Skoog basal salt mixture, 0.5% sucrose, 0.25% Gelrite, 0.05mg/L NAA, pH 5.8) supplemented with augmentin (500 mg/L), timentin (320 mg/L), and phosphinotricin (2 mg/L). Approximately after 2 weeks, transgenic plantlets with growing root systems were transferred to jars with sterile peat blocks. Transgenic plants were transplanted to the glasshouse when fully developed and allowed to grow until flowering and harvesting of seeds.
RT-PCR gene expression analysis of N. benthamiana stable lines
N. benthamiana leaf tissue was snap frozen in liquid nitrogen and stored at −80°C. Frozen tissue was ground to a fine powder using 5 mm glass beads and a Tissue Lyser II homogeniser (QIAGEN). RNA extraction was performed using the Concert Plant RNA Reagent (Invitrogen, Carlsbad, California, USA) followed by the TURBO DNA-free kit (Ambion, Carlsbad, California, USA) to remove DNA. RNA concentration was quantified by Nanodrop, and RNA integrity was assessed by agarose gel electrophoresis. cDNA libraries were prepared using BioScript Reverse Transcriptase (Bioline Reagents, London, UK) and oligo dT primers, according to manufacturer’s recommendations. RT-PCR was performed on a 1:10 cDNA dilution, using the KAPA 2G Fast DNA polymerase kit (KAPA Biosystems, Wilmington, Massachusetts, USA) and the oligonucleotides specified in S2 Table.
Histochemical staining for GUS activity
Transgenic M. truncatula and N. benthamiana roots carrying the MtPT4::eGFP:GUS, MtBCP1::eGFP:GUS, NbPT5b1::eGFP:GUS, and NbBCP1b::eGFP:GUS sequences were harvested 4 weeks after inoculation and treated with 80% acetone at −20°C for 30 minutes. Roots were then washed with PBS for 5 minutes and incubated in a staining solution containing 100 mM sodium phosphate pH 7.0, 5 mM potassium ferrocyanide, 5 mM potassium ferricyanide, and 2 mM 5-bromo-4-chloro-3-indoxyl-β-D-glucuronid acid (X-gluc) for 3 hours at 37°C. Roots were then washed twice with PBS and prepared for sectioning. M. truncatula roots were cast in paraffin wax, sectioned to a 5-μm width with a HM 340E rotary microtome (Thermo Fisher Scientific), and observed with an Olympus BX41 microscope (Olympus, Tokyo, Japan). N. benthamiana roots were embedded in 4% low melting point agarose, sectioned to a 100-μm width with a Leica VT1200S vibratome (Leica Biosystems) and imaged with a Keyence VHX-5000 microscope (Keyence, Osaka, Japan).
Staining and quantification of fungal structures
Ink staining of fungal structures was performed in M. truncatula and N. benthamiana roots as previously described [15]. In brief, roots were incubated with 10% potassium hydroxide at 95°C for 5 minutes and washed 3 times with sterile deionised water. Roots were then incubated in a solution of 5% pen ink (Sheaffer, Providence, Rhode Island, USA) and 5% acetic acid at 95°C for 2 minutes, washed with 5% acetic acid and then 3 times with sterile deionised water. N. benthamiana roots were additionally incubated with ClearSee [62] for 20 seconds and washed 3 times with sterile deionised water. Stained root fragments were then mounted in parallel on a glass slide and imaged with a Keyence VHX-5000 digital microscope (Keyence). Each root image was divided into equally distant sections by digitally applying a 0.75 mm × 0.75 mm grid, and presence or absence of fungal structures (internal hyphae, arbuscules, and vesicles) were recorded in each square for each root. The length of root fragment colonised was calculated as the percentage of the number of squares containing fungal structures from the total number of squares in each root. Results were averaged for each genotype and condition. For wheat germ agglutinin-FITC conjugate (WGA-FITC) staining in M. truncatula, roots were incubated in 1 mg/mL of WGA-FITC conjugate (Sigma-Aldrich, Saint Louis, Missouri, USA) in potassium phosphate buffer 0.1 M (pH 7.2) 10 minutes in the dark, rinsed several times in potassium phosphate buffer 0.1 M (pH 7.2), and mounted on glass slides. For staining in N. benthamiana, roots were incubated in WGA-FITC conjugate in potassium phosphate buffer 0.1 M (pH 7.2) supplemented with sucrose (3%) for 1 hour in the dark.
Dissection and microscopy of roots for betalain visualisation
M. truncatula roots expressing MtPT4-p1, MtBCP1-p1, and MtPT4-p3 were harvested 4 weeks after inoculation. Roots were hand-sectioned and mounted in potassium phosphate buffer 0.1 M (pH 7.2) on glass slides. Sections were imaged using a Zeiss Axioimager microscope (Zeiss, Oberkochen, Germany) with brightfield and UV light. N. benthamiana roots expressing NbPT5b-p1 and NbBCP1b-p1 were harvested 4 weeks after inoculation, embedded in 4% agarose supplemented with sucrose (3%) and sectioned to a 50-μm width with a Leica VT1200S vibratome (Leica Biosystems). Sections were then mounted on a glass slide and stained with WGA-FITC conjugate as previously described. Root sections were visualised with a Leica TCS SP8 confocal laser scanning microscope equipped with a Leica DFC 7000T camera and HyD detectors (Leica Biosystems). N. benthamiana roots expressing NbPT5b-p3 and NbBCP1b-p3 were harvested 4 weeks after inoculation, embedded in 4% agarose, and sectioned to a 100-μm width with a Leica VT1200S vibratome (Leica Biosystems). Sections were then mounted on a glass slide and imaged with a VHX-7000 Keyence digital microscope (Keyence). Root images were loaded into GIMP 2.8.2. Red colouring was visualised by adjusting Hue/Saturation via Colors menu. A settings file is provided as S2 Data.
Betalain extraction and detection using HPLC
Extraction and liquid chromatography of betalains in N. benthamiana leaf samples were performed as previously described [43]. In brief, leaf tissue samples were snap frozen in liquid nitrogen and grinded to a fine powder using mortar and pestle. Betalains were extracted overnight at 4°C in 80% aqueous methanol with 50 mM ascorbic acid with a volume of 1 mL extraction buffer per 50 mg leaf tissue. After extraction, samples were clarified twice by centrifuging at 12,000 g for 5 minutes and collecting the supernatants. Liquid chromatography analysis was performed using a Thermo Fisher Scientific Accela HPLC autosampler (Thermo Fisher Scientific) and pump system incorporating a photodiode array detector. Betalains were separated using a Luna Omega column (100 Å, 5 μm, 4.6 × 150 mm) from Phenomenex (Torrance, California, USA) under the following conditions: 3 minutes, 0% B; 3 to 19 minutes, 0% to 75% B; and 7 minutes, 0% B where mobile phase A was 0.1% formic acid in 1% acetonitrile, and solvent B was 100% acetonitrile, and at a flow rate of 500 μL/min. The betacyanin betanin was quantified since it has been shown to be the predominant pigment arising from the combined expression of DODA, CYP76AD1, and cDOPA5GT [23]. Betanin was detected by UV/VIS absorbance at a wavelength of 540 nm. Identification and quantification of betanin were carried out using a commercially available B. vulgaris extract (Tokyo Chemical Industry UK, Oxford, UK).
Rhizotron
The rhizotron consists of a 10 cm × 10 cm square petri dish where one of the sides of the base of the petri dish was removed to allow shoots to grow out. Ten-day-old N. benthamiana seedlings were placed on the base of the petri dish, covered with a coarse woven black cloth layer, followed by a layer of sand containing AM fungal spore inoculum. The lid was fixed using transparent adhesive tape. Rhizotrons were placed in black plastic bags and incubated in upright position over 52 days. For imaging, the plastic bag was removed, and rhizotrons were placed on a flatbed scanner. Images were loaded into GIMP 2.8.2. Red colouring was visualised by adjusting Hue/Saturation via Colors menu. A settings file is provided as S2 Data.
Phytophthora infection assay
N. benthamiana seeds were sterilised for 5 minutes in 70% ethanol, followed by 5 minutes in 2% sodium hypochlorite solution, and washed 5 times with sterile water. The seeds were germinated on half-strength Murashige & Skoog medium including B5 vitamins (Duchefa Biochemie, Haarlem, the Netherlands) with 1% bactoagar, pH 5.7, and allowed to grow for 17 days at 25°C under a 16-hour photoperiod. Roots of 17-day-old seedlings were inoculated with 10 μL zoospore solution from P. palmivora LILI [63], which had been transformed anew with a pTOR-tdTomato fluorescent reporter provided by Dr Stephen Whisson (The James Hutton Institute, Dundee, UK) following the protocol described by Evangelisti and colleagues [64]. Zoospores were harvested as previously described [65] and diluted to the concentration of 20,000 zoospores/mL (200 zoospores/seedling). Control (mock) plants were inoculated with 10 μL of sterile water. Inoculated seedlings were grown at 25°C under a 24 hours photoperiod for 9 days. Inoculated seedlings were imaged using Leica M165FC microscope for visualising the extent of infection by P. palmivora and an Epson Perfection Flatbed Scanner (Epson UK, Hemel Hempstead, UK) using default settings and a resolution of 600 dots per inch for full-colour images to record betalain pigmentation.
Supporting information
Expression of RiBTub and RiEF is used as a fungal biomass marker for root colonisation. Gene expression was analysed by qRT-PCR 4 wpi. Mock roots were inoculated with autoclaved R. irregularis inoculum. Error bars represent standard errors of 3 biological replicates. Statistical p-value (p) was obtained via unpaired 2-sampled Student t test. Data underlying this figure can be found in S3 Data. AM, arbuscular mycorrhiza; qRT-PCR, quantitative real-time polymerase chain reaction; RiBTub, R. irregularis β-tubulin; RiEF, R. irregularis longation factor 1-alpha; wpi, weeks postinoculation.
(PDF)
GUS staining of Medicago truncatula roots expressing MtPT4::GUS (a and b) and MtBCP1::GUS (c and d). GUS staining confirms promoter expression is limited to single cells in typically arbusculated tissue layers (inner cortex). Scale bar, 100 μm.
(PDF)
Phylogenetic analysis of the PT and BCP1 orthogroups containing the (a) MtPT4 and NbPT5b homologues and (b) MtBCP1 and NbBCP1b homologues, respectively.
(PDF)
Six-week old Nicotiana benthamiana T1 plants from NbPT5b-p1 (a–d) and NbBCP1-p1 (e–h) expressing lines. (a, b, e, and f) Plants 4 wpi with Rhizophagus irregularis. (c, d, g, and h) Plants descending from same lines mock inoculated with autoclaved R. irregularis inoculum. Scale bar, 1 cm. wpi, weeks after inoculation.
(PDF)
Ink staining of Nicotiana benthamiana roots expressing NbPT5b-p1 (a–d) and NbBCP1b-p1 (e–h) after inoculation with Rhizophagus irregularis. (a, b, e, and f) Root fragments that displayed betalain colouration before ink staining are represented by a red “+” sign. (c, d, g, and h) Root fragments that displayed no colouration before ink staining are represented by a grey “−”sign. (b, d, f, and h) are amplified images of the area delimited by the dashed squares. Scale bar, 1 mm.
(PDF)
Figure shows examples of T0 plants that varied in the severity of observable developmental defects and were phenotypically classified as severe (S; a, d, and e), moderate (M; b, f, and g), and light (L; c). Only plants with a light phenotype were able to set seeds, although not abundantly. Defects affected overall plant size and leaf and flower morphology. Severely affected plants displayed thicker leaves with strong deformation commonly showing a bifurcated shape (d) and flowers lacking reproductive organs (e). Moderately affected plants displayed altered leaf elongation with curvy edges (f) and flowers with a shorter (or lacking) perianth (g). Light phenotypes included slight leaf deformation, and flowers with shorter perianth but typical overall plant growth (c). (h) Semiquantitative RT-PCR analysis of betalain biosynthetic transgene expression in leaves of N. benthamiana T0 stable NbPT5b-p1 transformants. Line 18 displayed light developmental defects and was able to set seed, line 6 displayed moderate developmental defects, and lines 1 and 15 displayed severe developmental defects. NbEF1α is used as housekeeping control. Positive control (+) is NbPT5b-p1 plasmid DNA. Negative control (−) is water. −RT stands for minus reverse transcriptase control generated from line 18. Original gel images can be found in S4 Data. RT-PCR, reverse transcription PCR.
(PDF)
NbPT5b-p1 lines developed shorter roots and smaller leaves, which also appeared lightly pigmented in some cases. NbPT5b-p1 lines 18, 19, and 23, and the NbBCP1b-p1 line 32, were able to produce pigments upon colonisation with R. irregularis. NbPT5b-p1 line 23 did not produce any pigments upon colonisation.
(PDF)
(a) Some T1 plants show pigmentation in the leaves at varying degrees. (b) Liquid chromatography of pigmented leaves and controls. Pigmented leaves of NbPT5b-p1 expressing plants both in inoculated (myc) and non-inoculated (mock) conditions showed a peak at the expected retention time for betanin (red dotted line). Betanin standard is a commercially available Beta vulgaris extract and displays 2 peaks corresponding to the 2 betanin isomers commonly present in beet extracts. NbPT5b::GUS leaf tissue of same age T1 plants was used as negative control. Data underlying this figure can be found in S3 Data.
(PDF)
Root systems of NbPT5b-p3 (a–d) and NbBCP1b-p3 (e–h) Nicotiana benthamiana plants colonised by Rhizophagus irregularis. Images were taken at 52 dpi. (a, b, e, and f) Plants inoculated with R. irregularis. (c, d, g, and h) Plants descending from the same lines mock inoculated with autoclaved inoculum. (a, c, e, and g) are reflective light images, and (b, d, f, and h) are filtered for red colouring only. Scale bar, 1.3 cm. dpi, days postinoculation.
(PDF)
N. benthamiana plants were inoculated with Rhizophagus irregularis and ink stained 4 wpi. (a) Colonisation extent in N. benthamiana WT versus lines expressing NbBCP1b-p3 (BCP1-19 and BCP1-24) and NbPT5b-p3 (PT5b-16 and PT5b-21). Colonisation extent is calculated as the percentage of root containing arbuscules, vesicles, or internal hyphae over the total root system. (b) Frequency of arbuscules and vesicles recorded in colonised roots of N. benthamiana WT versus lines expressing NbBCP1b-p3 and NbPT5b-p3. Frequency is calculated as the percentage of root containing arbuscules or vesicles over the extent of root colonised by R. irregularis. Individual points represent individual plants. Error bars represent standard errors. Data underlying this figure can be found in S3 Data. AM, arbuscular mycorrhiza; wpi, weeks after inoculation; WT, wild-type.
(PDF)
A total of 16 transgenic betalain reporter lines per construct were grown on petridishes: 8 were not infected (mock), and 8 were infected with red fluorescent P. palmivora LILI-td zoospores. Plates were subsequently imaged under an epifluorescent microscope at 2 dpi (a) to document pathogen colonisation on roots and via flatbed scanning at 2 dpi (b) and 9 dpi (c). Application of a red filtering did not show any betalain pigment development. Scale bar, 50 mm. dpi, days postinfection.
(PDF)
(a) Schematic of the multigene vector constructed for inducible betalain expression in M. truncatula roots with the 3 betalain biosynthetic genes controlled by the MtPT4 promoter. (b–e) Expression of MtPT4-p3 in roots 4 wpi with Rhizophagus irregularis led to pigment accumulation in roots. (f–i) Mock inoculated roots expressing MtPT4-p3 did not show any pigment production. (b and f) Images taken under reflective light, and (c and g) are filtered for red colouring only. (d and h) Close up root images in bright field, and (e and i) and filtered to observe DSRed fluorescence. (j) Root section of M. truncatula expressing MtPT4-p3. Betalain accumulation correlates with arbuscule formation but can be observed mainly in the endodermal layer adjacent to arbuscule-containing cortical cells, pericycle, and steele. Open arrows mark internal hyphae and filled arrows signal cells containing arbuscules. Scale bar (b and f), 1 cm; scale bar (e and i), 1.5 mm; scale bar (j), 150 μm. AM, arbuscular mycorrhiza; wpi, weeks after inoculation.
(PDF)
(a and b) Example of an A17 M. truncatula root system expressing MtPT4-p3 able to produce betalains upon inoculation. (c and d) Example of dmi3 M. truncatula root system expressing MtPT4-p3. (a and c) Images taken under reflective light, and (b and d) are filtered for red colouring only. Scale bar, 1 cm. wpi, weeks after inoculation.
(PDF)
(a) Schematic of the fungal visualisation and quantification process of Medicago truncatula hairy roots after inoculation with Rhizophagus irregularis: (1) M. truncatula transgenic hairy roots express the DSRed marker and can be selected through detection of fluorescence under 510–560 nm; (2) DSRed positive roots are cut and divided into betalain producing and nonproducing root fragments; (3) pigmented and nonpigmented root fragments are ink stained separately (betalain colouration fades upon incubation with potassium hydroxide during the ink staining process); and (4) ink-stained root fragments are mounted in glass slides and imaged for visualisation and quantification of fungal structures. (b) Example of MtPT4-p3 pigmented root fragments showing fungal colonisation. (c) Example of MtPT4-p3 nonpigmented uncolonised root fragments. Scale bar, 100 μm.
(PDF)
Transgenic NbPT5b-p3 plants grown in a rhizotron setup supplied with Rhizophagus irregularis spore inoculum imaged over time. (a) Reflective light images, and (b) represent the same images filtered for red/magenta hues. Scale bar, 1 cm. dpi, days postinoculation.
(PDF)
We selected roots for DSRed fluorescence and then cut and divided them in pigmented (red rows) and nonpigmented (noncoloured rows) fragments for ink staining. N refers to the total number of root fragments analysed for each condition. Table shows the number of root fragments containing fungal structures and the average extent of colonisation by these structures over the length of the root fragment. Error shown as standard error. %AC, percentage of arbuscule colonisation; %IHC, percentage of internal hyphae colonisation; %VC, percentage of vesicle colonsation; A, arbuscules; IH, internal hyphae; V, vesicles; wpi, weeks postinoculation.
(PDF)
(PDF)
All vectors are available to obtain free of charge through Addgene. Plasmid maps and sequence files can be downloaded in the Addgene website with the catalogue numbers provided.
(PDF)
All sequences were selected immediately upstream the start codon of the gene.
(PDF)
(TXT)
(PDF)
Acknowledgments
We would like to thank Liron Shenhav for maintenance of N. benthamiana plants, Bo Xu and Edouard Evangelisti for help with wax casting, Nicola Patron for providing Ubi10 promoter, Stephen Whisson for providing pTOR-tdTomato reporter, Edouard Evangelisti and Sabine Brumm for imaging, and Philip Carella for comments on the manuscript. P. palmivora research is covered by the Department for Environment, Food and Rural Affairs’ (Defra) plant health license 114614/208745/4.
Abbreviations
- AM
arbuscular mycorrhiza
- BCP
blue copper protein
- BIA
benzylisoquinoline alkaloid
- cDOPA5GT
cyclo-DOPA 5-O-glucosyltransferase
- cyclo-DOPA
cyclo-dihydroxyphenylalanine
- dmi3
Doesn’t Make Infections 3
- DODA
ʟ-DOPA-4,5-dioxygenase
- dpi
days postinoculation
- ʟ-DOPA
3,4-dihydroxy-ʟ-phenylalanine
- PHT1
phosphate transporter 1
- QTL
quantitative trait locus
- RiBTub
R. irregularis β-tubulin
- RT-PCR
reverse transcription PCR
- WGA
wheat germ agglutinin
- WT
wild-type
- X-gluc
5-bromo-4-chloro-3-indoxyl-β-D-glucuronid acid
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by an iCASE BBSRC-DTP (RG88096) sponsored by Coca-Cola to S.F.B & A.T., Gatsby Charitable Foundation (GAT3395/GLD) and Royal Society (UF110073, UF160413) to S.S., and a Natural Environmental Research Council Independent Research Fellowship (NE/K009303) to S.F.B. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Spatafora JW, Chang Y, Benny GL, Lazarus K, Smith ME, Berbee ML, et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia. 2016;108(5):1028–46. doi: 10.3852/16-042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Parniske M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat Rev Microbiol. 2008;6:763–75. doi: 10.1038/nrmicro1987 [DOI] [PubMed] [Google Scholar]
- 3.Brundrett MC, Tedersoo L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018;220(4):1108–15. doi: 10.1111/nph.14976 [DOI] [PubMed] [Google Scholar]
- 4.Genre A, Lanfranco L, Perotto S, Bonfante P. Unique and common traits in mycorrhizal symbioses. Nat Rev Microbiol. 2020;18:649–60. doi: 10.1038/s41579-020-0402-3 [DOI] [PubMed] [Google Scholar]
- 5.Bago B, Pfeffer PE, Shachar-Hill Y. Carbon metabolism and transport in arbuscular mycorrhizas. Plant Physiol. 2000;124:949–57. doi: 10.1104/pp.124.3.949 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Holford ICR. Soil phosphorus: Its measurement, and its uptake by plants. Aust J Soil Res. 1997;35(2):227–39. [Google Scholar]
- 7.Smith SE, Smith FA, Jakobsen I. Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiol. 2003;133(1):16–20. doi: 10.1104/pp.103.024380 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Harrison MJ. Signaling in the Arbuscular Mycorrhizal Symbiosis. Annu Rev Microbiol. 2005;59(1):19–42. doi: 10.1146/annurev.micro.58.030603.123749 [DOI] [PubMed] [Google Scholar]
- 9.Franken P, Requena N. Analysis of gene expression in arbuscular mycorrhizas: new approaches and challenges. New Phytol. 2001;150(3):517–23. [Google Scholar]
- 10.Harrison MJ, Dewbre GR, Liu J. A phosphate transporter from Medicago truncatula involved in the acquisition of phosphate released by arbuscular mycorrhizal fungi. Plant Cell. 2002;14(10):2413–29. doi: 10.1105/tpc.004861 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hohnjec N, Vieweg MF, Pühler A, Becker A, Küster H. Overlaps in the transcriptional profiles of Medicago truncatula roots inoculated with two different Glomus fungi provide insights into the genetic program activated during arbuscular mycorrhiza. Plant Physiol. 2005;137(4):1283–301. doi: 10.1104/pp.104.056572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Phillips JM, Hayman DS. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br Mycol Soc. 1970;55(1):158–IN18. [Google Scholar]
- 13.Nicolson TH. Mycorrhiza in the Gramineae. Trans Br Mycol Soc. 1959;42(4):421–IN3. [Google Scholar]
- 14.Gerdemann JW. Relation of a large soil-borne spore to phycomycetous mycorrhizal infections. Mycologia. 1955;47(5):619–32. [Google Scholar]
- 15.Vierheilig H, Coughlan AP, Wyss U, Piché Y. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal fungi. Appl Environ Microbiol. 1998;64(12):5004–7. doi: 10.1128/AEM.64.12.5004-5007.1998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bonfante-Fasolo P, Faccio A, Perotto S, Schubert A. Correlation between chitin distribution and cell wall morphology in the mycorrhizal fungus Glomus versiforme. Mycol Res. 1990;94(2):157–65. [Google Scholar]
- 17.Wang M, Schäfer M, Li D, Halitschke R, Dong C, McGale E, et al. Blumenols as shoot markers of root symbiosis with arbuscular mycorrhizal fungi. Elife. 2018;7. doi: 10.7554/eLife.37093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Klingner A, Bothe H, Wray V, Marner FJ. Identification of a yellow pigment formed in maize roots upon mycorrhizal colonization. Phytochemistry. 1995;38(1):53–5. [Google Scholar]
- 19.Klingner A, Hundeshagen B, Kernebeek H, Bothe H. Localization of the yellow pigment formed in roots of gramineous plants colonized by arbuscular fungi. Vol. 185, Protoplasma. Springer-Verlag; 1995. [Google Scholar]
- 20.Paszkowski U, Jakovleva L, Boller T. Maize mutants affected at distinct stages of the arbuscular mycorrhizal symbiosis. Plant J. 2006;47(2):165–73. doi: 10.1111/j.1365-313X.2006.02785.x [DOI] [PubMed] [Google Scholar]
- 21.Gandía-Herrero F, Cabanes J, Escribano J, García-Carmona F, Jiménez-Atiénzar M. Encapsulation of the most potent antioxidant betalains in edible matrixes as powders of different colors. J Agric Food Chem. 2013;61(18):4294–302. doi: 10.1021/jf400337g [DOI] [PubMed] [Google Scholar]
- 22.DeLoache WC, Russ ZN, Narcross L, Gonzales AM, Martin VJJ, Dueber JE. An enzyme-coupled biosensor enables (S)-reticuline production in yeast from glucose. Nat Chem Biol. 2015;11(7):465–71. doi: 10.1038/nchembio.1816 [DOI] [PubMed] [Google Scholar]
- 23.Timoneda A, Sheehan H, Feng T, Lopez-Nieves S, Maeda HA, Brockington S. Redirecting primary metabolism to boost production of tyrosine-derived specialised metabolites in planta. Sci Rep. 2018;8(1):17256. doi: 10.1038/s41598-018-33742-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lin YK, Yeh YC. Dual-signal microbial biosensor for the detection of dopamine without inference from other catecholamine neurotransmitters. Anal Chem. 2017;89(21):11178–82. doi: 10.1021/acs.analchem.7b02498 [DOI] [PubMed] [Google Scholar]
- 25.Chen PH, Lin C, Guo KH, Yeh YC. Development of a pigment-based whole-cell biosensor for the analysis of environmental copper. RSC Adv. 2017;7(47):29302–5. [Google Scholar]
- 26.Chou YC, Shih CI, Chiang CC, Hsu CH, Yeh YC. Reagent-free DOPA-dioxygenase colorimetric biosensor for selective detection of L-DOPA. Sens Actuators B. 2019;297:126717. [Google Scholar]
- 27.Brockington SF, Walker RH, Glover BJ, Soltis PS, Soltis DE. Complex pigment evolution in the Caryophyllales. New Phytol. 2011;190(4):854–64. doi: 10.1111/j.1469-8137.2011.03687.x [DOI] [PubMed] [Google Scholar]
- 28.Contreras-Llano LE, Guerrero-Rubio MA, Lozada-Ramírez JD, García-Carmona F, Gandía-Herrero F. First betalain-producing bacteria break the exclusive presence of the pigments in the plant kingdom. MBio. 2019;10(2):e00345–19. doi: 10.1128/mBio.00345-19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Musso H. The pigments of fly agaric, Amanita muscaria. Tetrahedron. 1979;35(24):2843–53. [Google Scholar]
- 30.Babos M, Halász K, Zagyva T, Zöld-Balogh Á, Szego D, Bratek Z. Preliminary notes on dual relevance of ITS sequences and pigments in Hygrocybe taxonomy. Persoonia. 2011;26:99–107. doi: 10.3767/003158511X578349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Polturak G, Breitel D, Grossman N, Sarrion-Perdigones A, Weithorn E, Pliner M, et al. Elucidation of the first committed step in betalain biosynthesis enables the heterologous engineering of betalain pigments in plants. New Phytol. 2016;210(1):269–83. doi: 10.1111/nph.13796 [DOI] [PubMed] [Google Scholar]
- 32.Sunnadeniya R, Bean A, Brown M, Akhavan N, Hatlestad G, Gonzalez A, et al. Tyrosine hydroxylation in betalain pigment biosynthesis is performed by cytochrome P450 enzymes in beets (Beta vulgaris). PLoS ONE. 2016;11(2):e0149417. doi: 10.1371/journal.pone.0149417 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Christinet L, Burdet FX, Zaiko M, Hinz U, Zrÿd J-P. Characterization and functional identification of a novel plant 4,5-extradiol dioxygenase involved in betalain pigment biosynthesis in Portulaca grandiflora. Plant Physiol. 2004;134(1):265–74. doi: 10.1104/pp.103.031914 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gandía-Herrero F, Escribano J, García-Carmona F. Betaxanthins as pigments responsible for visible fluorescence in flowers. Planta. 2005;222(4):586–93. doi: 10.1007/s00425-005-0004-3 [DOI] [PubMed] [Google Scholar]
- 35.Hatlestad GJ, Sunnadeniya RM, Akhavan N. a, Gonzalez A, Goldman IL, McGrath JM, Lloyd AM. The beet R locus encodes a new cytochrome P450 required for red betalain production. Nat Genet. 2012;44(7):816–20. doi: 10.1038/ng.2297 [DOI] [PubMed] [Google Scholar]
- 36.Vogt T, Grimm R, Strack D. Cloning and expression of a cDNA encoding betanidin 5-O-glucosyltransferase, a betanidin- and flavonoid-specific enzyme with high homology to inducible glucosyltransferases from the Solanaceae. Plant J. 1999;19(5):509–19. doi: 10.1046/j.1365-313x.1999.00540.x [DOI] [PubMed] [Google Scholar]
- 37.Sasaki N, Wada K, Koda T, Kasahara K, Adachi T, Ozeki Y. Isolation and characterization of cDNAs encoding an enzyme with glucosyltransferase activity for cyclo-DOPA from four o’clocks and feather cockscombs. Plant Cell Physiol. 2005;46(4):666–70. doi: 10.1093/pcp/pci064 [DOI] [PubMed] [Google Scholar]
- 38.Grewal PS, Modavi C, Russ ZN, Harris NC, Dueber JE. Bioproduction of a betalain color palette in Saccharomyces cerevisiae. Metab Eng. 2018;45:180–8. doi: 10.1016/j.ymben.2017.12.008 [DOI] [PubMed] [Google Scholar]
- 39.Harris NN, Javellana J, Davies KM, Lewis DH, Jameson PE, Deroles SC, et al. Betalain production is possible in anthocyanin-producing plant species given the presence of DOPA-dioxygenase and L-DOPA. BMC Plant Biol. 2012;12(1):34. doi: 10.1186/1471-2229-12-34 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Polturak G, Grossman N, Vela-Corcia D, Dong Y, Nudel A, Pliner M, et al. Engineered gray mold resistance, antioxidant capacity, and pigmentation in betalain-producing crops and ornamentals. Proc Natl Acad Sci U S A. 2017;114(34):9062–7. doi: 10.1073/pnas.1707176114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tian Y, Fu X, Yang Z, Wang B, Gao J, Wang M, et al. Metabolic engineering of rice endosperm for betanin biosynthesis. New Phytol. 2019;225(5):1915–22. doi: 10.1111/nph.16323 [DOI] [PubMed] [Google Scholar]
- 42.He Y, Zhang T, Sun H, Zhan H, Zhao Y. A reporter for noninvasively monitoring gene expression and plant transformation. Hortic Res. 2020;7(1):1–6. doi: 10.1038/s41438-020-00390-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sheehan H, Feng T, Walker-Hale N, Lopez-Nieves S, Pucker B, Guo R, et al. Evolution of L-DOPA 4,5-dioxygenase activity allows for recurrent specialisation to betalain pigmentation in Caryophyllales. New Phytol. 2019;227(3):914–29. doi: 10.1111/nph.16089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lévy J, Bres C, Geurts R, Chalhoub B, Kulikova O, Duc G, et al. A putative Ca2+ and calmodulin-dependent protein kinase required for bacterial and fungal symbioses. Science. 2004;303(5662):1361–4. doi: 10.1126/science.1093038 [DOI] [PubMed] [Google Scholar]
- 45.Mitra RM, Gleason CA, Edwards A, Hadfield J, Downie JA, Oldroyd GED, et al. A Ca2+/calmodulin-dependent protein kinase required for symbiotic nodule development: Gene identification by transcript-based cloning. Proc Natl Acad Sci U S A. 2004;101(13):4701–5. doi: 10.1073/pnas.0400595101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Takeda N, Sato S, Asamizu E, Tabata S, Parniske M. Apoplastic plant subtilases support arbuscular mycorrhiza development in Lotus japonicus. Plant J. 2009;58(5):766–77. doi: 10.1111/j.1365-313X.2009.03824.x [DOI] [PubMed] [Google Scholar]
- 47.Xue L, Klinnawee L, Zhou Y, Saridis G, Vijayakumar V, Brands M, et al. AP2 transcription factor CBX1 with a specific function in symbiotic exchange of nutrients in mycorrhizal Lotus japonicus. Proc Natl Acad Sci U S A. 2018;115(39):E9239–46. doi: 10.1073/pnas.1812275115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Floss DS, Gomez SK, Park HJ, MacLean AM, Müller LM, Bhattarai KK, et al. A transcriptional program for arbuscule degeneration during AM symbiosis is regulated by MYB1. Curr Biol. 2017;27(8):1206–12. doi: 10.1016/j.cub.2017.03.003 [DOI] [PubMed] [Google Scholar]
- 49.Chen N, Yu Z-H, Xiao X-G. Cytosolic and nuclear co-localization of betalain biosynthetic enzymes in tobacco suggests that betalains are synthesized in the cytoplasm and/or nucleus of betalainic plant cells. Front Plant Sci. 2017;8:831. doi: 10.3389/fpls.2017.00831 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Gutjahr C, Casieri L, Paszkowski U. Glomus intraradices induces changes in root system architecture of rice independently of common symbiosis signaling. New Phytol. 2009;182(4):829–37. doi: 10.1111/j.1469-8137.2009.02839.x [DOI] [PubMed] [Google Scholar]
- 51.Whiteside MD, Werner GDA, Caldas VEA, van’t Padje A, Dupin SE, Elbers B, Bakker M, Wyatt GAK, Klein M, Hink MA, Postma M, Vaitla B, Noë R, Shimizu TS, West SA, Kiers ET. Mycorrhizal fungi respond to resource inequality by moving phosphorus from rich to poor patches across networks. Curr Biol. 2019;29(12):2043-2050.e8. doi: 10.1016/j.cub.2019.04.061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Charters MD, Sait SM, Field KJ. Aphid herbivory drives asymmetry in carbon for nutrient exchange between plants and an arbuscular mycorrhizal fungus. Curr Biol. 2020;30(10):1801–8. doi: 10.1016/j.cub.2020.02.087 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Rillig MC, Mummey DL. Mycorrhizas and soil structure. New Phytol. 2006;171(1):41–53. doi: 10.1111/j.1469-8137.2006.01750.x [DOI] [PubMed] [Google Scholar]
- 54.Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, Speleman F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002;3(7):1–12. doi: 10.1186/gb-2002-3-7-research0034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kakar K, Wandrey M, Czechowski T, Gaertner T, Scheible WR, Stitt M, et al. A community resource for high-throughput quantitative RT-PCR analysis of transcription factor gene expression in Medicago truncatula. Plant Methods. 2008;4(1):18. doi: 10.1186/1746-4811-4-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Engler C, Kandzia R, Marillonnet S. A one pot, one step, precision cloning method with high throughput capability. PLoS ONE. 2008;3(11):e3647. doi: 10.1371/journal.pone.0003647 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Weber E, Engler C, Gruetzner R, Werner S, Marillonnet S. A modular cloning system for standardized assembly of multigene constructs. PLoS ONE. 2011;6(2):e16765. doi: 10.1371/journal.pone.0016765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Werner S, Engler C, Weber E, Gruetzner R, Marillonnet S. Fast track assembly of multigene constructs using Golden Gate cloning and the MoClo system. Bioeng Bugs. 2012;3(1):38–43. doi: 10.4161/bbug.3.1.18223 [DOI] [PubMed] [Google Scholar]
- 59.Engler C, Youles M, Gruetzner R, Ehnert T-M, Werner S, Jones JDG, et al. A golden gate modular cloning toolbox for plants. ACS Synth Biol. 2014;3(11):839–43. doi: 10.1021/sb4001504 [DOI] [PubMed] [Google Scholar]
- 60.Boisson-Dernier A, Chabaud M, Garcia F, Bécard G, Rosenberg C, Barker DG. Agrobacterium rhizogenes-transformed roots of Medicago truncatula for the study of nitrogen-fixing and endomycorrhizal symbiotic associations. Mol Plant Microbe Interact. 2001;14(6):695–700. doi: 10.1094/MPMI.2001.14.6.695 [DOI] [PubMed] [Google Scholar]
- 61.Sparkes IA, Runions J, Kearns A, Hawes C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat Protoc. 2006;1(4):2019–25. doi: 10.1038/nprot.2006.286 [DOI] [PubMed] [Google Scholar]
- 62.Kurihara D, Mizuta Y, Sato Y, Higashiyama T. ClearSee: A rapid optical clearing reagent for whole-plant fluorescence imaging. Development. 2015;142(23):4168–79. doi: 10.1242/dev.127613 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Torres GA, Sarria GA, Varon F, Coffey MD, Elliott ML, Martinez G. First report of bud rot caused by Phytophthora palmivora on African oil palm in Colombia. Plant Dis. 2010;94(9):1163–3. doi: 10.1094/PDIS-94-9-1163A [DOI] [PubMed] [Google Scholar]
- 64.Evangelisti E, Yunusov T, Shenhav L, Schornack S. N-acetyltransferase AAC(3)-I confers gentamicin resistance to Phytophthora palmivora and Phytophthora infestans. BMC Microbiol. 2019;19(1):265. doi: 10.1186/s12866-019-1642-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Evangelisti E, Shenhav L, Yunusov T, Le Naour-Vernet M, Rink P, Schornack S. Hydrodynamic shape changes underpin nuclear rerouting in branched hyphae of an oomycete pathogen. MBio. 2019;10(5):e01516–19. doi: 10.1128/mBio.01516-19 [DOI] [PMC free article] [PubMed] [Google Scholar]