

Abstract
Simple Summary
Glioblastoma (GB) is the most aggressive brain cancer in humans. Patient survival outcomes have remained dismal despite intensive research over the past 50 years, with a median overall survival of only 14.6 months. We highlight the critical role of the renin–angiotensin system (RAS) on GB cancer stem cells and the tumor microenvironment which, in turn, influences cancer stem cells in driving tumorigenesis and treatment resistance. We present recent developments and underscore the need for further research into the GB tumor microenvironment. We discuss the novel therapeutic targeting of the RAS using existing commonly available medications and utilizing model systems to further this critical investigation.
Abstract
Glioblastoma (GB) is an aggressive primary brain tumor. Despite intensive research over the past 50 years, little advance has been made to improve the poor outcome, with an overall median survival of 14.6 months following standard treatment. Local recurrence is inevitable due to the quiescent cancer stem cells (CSCs) in GB that co-express stemness-associated markers and components of the renin–angiotensin system (RAS). The dynamic and heterogeneous tumor microenvironment (TME) plays a fundamental role in tumor development, progression, invasiveness, and therapy resistance. There is increasing evidence showing the critical role of the RAS in the TME influencing CSCs via its upstream and downstream pathways. Drugs that alter the hallmarks of cancer by modulating the RAS present a potential new therapeutic alternative or adjunct to conventional treatment of GB. Cerebral and GB organoids may offer a cost-effective method for evaluating the efficacy of RAS-modulating drugs on GB. We review the nexus between the GB TME, CSC niche, and the RAS, and propose re-purposed RAS-modulating drugs as a potential therapeutic alternative or adjunct to current standard therapy for GB.
Keywords: glioblastoma, renin–angiotensin system, pluripotent stem cells, organoids, cancer stem cells, cancer stem cell niche, tumor microenvironment
1. Introduction
Glioblastoma (GB), the most common and most aggressive primary brain cancer in humans, is classified as a WHO grade IV astrocytoma, and is characterized by microvascular proliferation and central necrosis [1]. Primary GB arises de novo and accounts for 90% of cases with a predilection for older individuals, while secondary GB arises from low-grade astrocytoma and affects younger patients [2]. GB has been categorized into four distinct molecular subtypes: classical, mesenchymal, neural, and proneural [3], although other studies have only identified classical, mesenchymal, and proneural subtypes [4]. The classical subtype includes amplification or mutation of epidermal growth factor receptor (EGFR), the mesenchymal subtype includes deletions of the 17q11.2 region containing the gene NF1, and the proneural subtype is characterized by high levels of platelet-derived growth factor receptor α (PDGFRα) expression and point mutations in isocitrate dehydrogenase 1 (IDH1) and p53 [3].
Various genetic or epigenetic changes may affect the prognosis of GB patients including IDH mutations and O6-methylguanine-DNA methyltransferase (MGMT) methylation status. GB may be divided into IDH-wild-type and IDH-mutant tumors. IDH is an enzyme that catalyzes oxidative decarboxylation of isocitrate to 2-oxoglutarate. The most common mutation in GB affects IDH1 with a single amino acid missense mutation at arginine 132 which is replaced by histidine [5]. IDH-wild-type GB is more common, tends to arise de novo, and is generally more aggressive with a worse prognosis than IDH-mutant GB. By contrast, IDH-mutant GB is predominantly observed in secondary GB and is associated with a better prognosis [6]. The current standard treatment for GB involves maximal safe surgical resection with adjuvant chemotherapy and radiotherapy, known as the Stupp protocol [7]. Temozolomide, an alkylating agent, is used as first-line chemotherapy for GB with its efficacy related to the methylation status of the MGMT promoter [8]. MGMT methylation is associated with an improved overall survival in GB patients [9]. Despite this intensive treatment, tumor recurrence in GB patients is inevitable with an overall median survival time of 14.6 months with a range of 12–14 months which has not changed since the introduction of the Stupp protocol in 2005 [10,11].
We reviewed the dynamic relationship between the tumor microenvironment (TME), the RAS, and cancer stem cells (CSCs) in GB. We speculate that RAS-modulating drugs may offer a potential therapeutic alternative or adjunct to current standard therapy. Further functional and epidemiological studies are required to investigate the efficacy of RAS-targeting drugs in the treatment of GB.
2. GB Tumor Microenvironment
The GB tumor microenvironment (TME) is highly heterogeneous and consists of cancer cells and non-cancer cells. Non-cancer cell types include immune cells, such as tumor-associated macrophages (TAMs), resident glial cells, peripheral macrophages, endothelial cells, pericytes, astrocytes, CSCs, fibroblasts, and other components such as the extracellular matrix (ECM) [12]. Given the rarity of extracranial metastasis from GB [13], it appears that GB development requires the unique intracerebral microenvironment inclusive of the blood–brain barrier (BBB) [14]. The TME, with emphasis on glioma-associated microglia/macrophages, pericytes, and reactive astrocytes, is increasingly recognized to play a critical role in GB development and progression [15]. The idea that cytokines, growth factors, chemokines, inflammatory mediators, and remodeling enzymes are involved in intra- and inter-cellular communications within the TME is not novel [16]. Additionally, constant communication between GB cells and the surrounding TME [14] is facilitated by extracellular vesicles that expedite bi-directional cross-talk within the TME [12,17].
Anatomically distinct regions of the TME, known as tumor niches, are thought to contain CSCs and play a fundamental role in the regulation of metabolism, immune surveillance, survival, invasion, and self-maintenance with the renin–angiotensin system (RAS) playing a critical role [15,18,19]. The GB TME may consist of several distinct tumor niches including the hypoxic tumor niche, the perivascular or angiogenic tumor niche, and the vascular-invasive tumor niche. The perivascular niche contains CSCs in close juxtaposition with the abnormal angiogenic vasculature and provides a supportive environment for CSC growth, maintenance, and survival. The vasculature in the hypoxic tumor niche is either non-functional or has regressed, leading to areas of necrosis that are surrounded by rows of hypoxic palisading tumor cells [20]. The vascular-invasive tumor niche contains tumor cells co-opted with normal blood vessels that migrate deep into the brain parenchyma [20].
GB is highly vascular and is characterized by extensive neovascularization and pathological angiogenesis predominantly induced by vascular endothelial growth factor (VEGF), which is produced by tumor cells, CSCs, and immune cells [21,22]. Other angiogenic factors, such as transforming growth factor-β1 (TGF-β1), platelet-derived growth factor-BB, and fibroblast growth factor-2, may also play a role in the pathological angiogenesis [23,24]. In addition to endothelial proliferation, bone marrow-derived endothelial and pericyte progenitor cells may be recruited and incorporated into the growing vessels [25]. There is also evidence that CSCs may be involved in neovascularization by differentiating into endothelial cells or pericytes in GB [26,27,28]. Increased VEGF expression also fosters an immunosuppressive microenvironment that enables tumors, including GB, to evade host immune surveillance [29]. The abnormal vasculature in GB includes dilated and leaky vessels and glomeruloid microvascular proliferation in which endothelial cells and pericytes form poorly organized vascular structures, which effectively disrupt the BBB, leading to cerebral edema. In addition, the blood–brain tumor barrier (BBTB) hinders drug delivery to the tumor [30].
The BBB is a highly specialized, selectively permeable barrier between the brain and the systemic blood supply that helps to maintain homeostasis of the cerebral microenvironment. The structure of the BBB includes endothelial cells with tight junctions, adherens junctions, astrocytes, pericytes, and the basement membrane [31]. The BBB plays several fundamental roles, including supplying the brain with essential nutrients, such as oxygen and glucose, mediating the efflux of waste products, facilitating the movement of nutrients and plasma proteins, and restricting toxins into the central nervous system (CNS) [32]. Disruption of the BBB and its tight regulation of the cerebral microenvironment leads to increased blood vessel permeability with plasma and fluid leakage into the tumor tissue causing cerebral edema and raised interstitial and intracranial pressure [33]. The combination of abnormal vasculature in GB and the disruption of the BBB leads to impaired blood flow and reduced oxygen delivery within the tumor [34]. Microvascular thrombosis may also occur causing occlusion of the blood vessels, further promoting intra-tumoral hypoxia, leading to pseudo-palisading necrosis [35]. Hypoxia is also a consequence of increased oxygen diffusion distance due to the fact of tumor growth and expansion [34], which may, in and of itself, be a key regulator of tumor cell survival, stemness, and immune surveillance in the TME [36,37,38]. Hypoxia also sustains tumor cell proliferation, invasiveness, and contributes to chemotherapy and radiotherapy resistance. This occurs via inhibition of free radicals, which reduces the efficacy of radiotherapy [39], and through upregulation of the multi-drug resistance gene, MDR1/ABCB1, which reduces chemotherapy effectiveness. Hypoxia-inducible factor-1 (HIF-1) and HIF-2 mediate the response to hypoxia on a molecular level in GB [40] and may potentially modify CSCs [41]. The GB microenvironmental niche also consists of pseudo-palisading glioma cells that upregulate HIF proteins, inducing expression of factors, such as VEGF and interleukin 8 (IL-8), which are implicated in tumor cell survival, metabolism, invasion, and angiogenesis. The resultant cross-talk releases pro-inflammatory signals from the areas of necrosis in the hypoxic tumor niche into the surrounding TME, promoting immunosuppression, and angiogenesis [42].
Immune cells, including circulating monocytes, neutrophils, and myeloid-derived suppressor cells (MDSCs), are another source of angiogenic factors. In ovarian cancer, MDSCs increase CSC characteristics by increasing microRNA-101 expression, which induces the expression of stemness genes [43]. It is interesting to speculate that MDSCs also regulate the stemness of CSCs within the GB TME via this mechanism (Figure 1). These cells may enter the brain as a result of breakdown of the BBB in GB and the production of tumor-derived chemokines and cytokines, contributing to the immunosuppressive GB TME [44,45,46]. TAMs are the dominant immune cell population in GB and may include resident microglial cells and peripheral macrophages [47,48]. Traditionally, TAMs have been defined as either anti-tumoral M1/Th1 (classical-activated macrophages) or pro-tumoral M2/Th2 (alternative-activated macrophages) phenotypes. M1 macrophages foster the inflammatory response by secreting pro-inflammatory cytokines such as IL-12, tumor necrosis factor-α (TNF-α), CXCL-10, and interferon-γ (IFN-γ) and produce high levels of nitric oxide synthase to exert anti-tumor cell activity (Figure 1). M2 macrophages, on the other hand, play a key immunosuppressive function by secreting anti-inflammatory cytokines, such as IL-10, IL-13, and IL-4, and express abundant arginase-1, mannose receptor CD206, and scavenger receptors to promote tumor progression [49,50,51]. The release of TGF-β by TAMs has been shown to induce matrix metalloproteinase 9 (MMP9) and, thus, increase CSC invasiveness [52]. A more recent study has demonstrated that the TAM population is in a constant state of transition or plasticity between the two phenotypes and that M1 phenotype expression may be enhanced by TME changes or therapeutic interventions [51]. Resident microglia are present within the brain, but it is the recruitment of peripheral macrophages to the GB TAM pool, in particular, that may mediate tumor phagocytosis with disruption of the signal regulatory protein α receptor (SIRP-α)–CD47 axis. This facilitates immune evasion because the antiphagocytic “don’t eat me” surface protein CD47 is upregulated, which binds to SIRP-α on phagocytic cells to inhibit phagocytosis [53]. However, even in the absence of macrophages, resident microglia may be transformed into effector cells of tumor cell phagocytosis, in response to anti-CD47 blockade [54]. In models of pancreatic ductal adenocarcinoma, for example, RP-182 may selectively induce conformational switching of the mannose receptor CD206, which is expressed on the M2 TAM phenotype, ultimately reprogramming M2-like TAMs into an anti-tumor M1-like phenotype [55]. The immunosuppressive phenotype of TAMs may be controlled by long-chain fatty acid metabolism, and chemical inhibitors targeting this metabolic pathway may block TAM polarization in vitro and tumor growth in vivo [56]. GB-derived exosomes may reprogram M1 macrophages to M2 macrophages and condition M2 macrophages to become strongly immunosuppressive TAMs [57].
Figure 1.
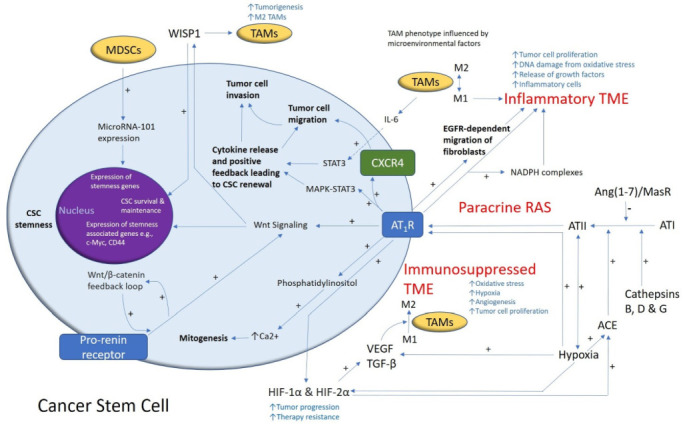
A schema demonstrating the role of the renin–angiotensin system (RAS) and its convergent signaling pathways in the glioblastoma tumor microenvironment (TME) and cancer stem cells (CSCs). A cancer stem cell (with the cytoplasm depicted in light blue and the nucleus in purple) residing within the glioblastoma TME. Angiotensin II (ATII), the physiologically active end-product of the paracrine RAS, activates ATII receptor 1 (AT1R) leading to increased tumor cell proliferation, oxidative stress, hypoxia and angiogenesis, and inflammation—the hallmarks of cancer. This contributes to an inflammatory TME by increasing the number of inflammatory cells, partly by increasing the number of NADPH complexes, leading to tumor cell proliferation, DNA damage from oxidative stress, and release of growth factors. AT1R also activates phosphatidylinositol signaling, which increases cytosolic Ca2+ to promote mitogenesis. Hypoxia increases paracrine RAS activity by upregulating angiotensin-converting enzyme (ACE) and the expression of hypoxia-inducible factor 1α (HIF-1α) and HIF-2α, which increase tumor progression and treatment resistance. HIF-1α, HIF-2α, and hypoxia increase the expression of vascular endothelial growth factor (VEGF) which increases angiogenesis. AT1R, via MAPK-STAT3 signaling, contributes to a cytokine release that leads to CSC renewal. C-X-C chemokine receptor type 4 (CXCR4) promotes tumor cell migration and invasion. AT1R signaling and the prorenin receptor, which act in a feedback loop with Wnt/β-catenin, increase Wnt signaling which promotes CSC stemness by upregulating stemness-associated markers. Myeloid-derived suppressor cells (MDSCs) promote CSC characteristics by increasing microRNA-101 expression that induces expression of stemness-related genes in CSCs. The Ang(1–7)/MasR axis opposes the ACE/ATII/AT1R axis. Cathepsins B, D, and G act as bypass loops for the RAS. Under the influence of the TME, polarization of tumor-associated macrophages (TAMs)—immune cells that are located within the TME—changes from the M1 to M2 phenotype. M2 TAMs induce the proliferation of CSCs via interleukin 6 (IL-6)-induced activation of STAT3, leading to cytokine release and positive feedback contributing to CSC renewal. Glioblastoma CSCs secrete Wnt-induced signaling protein 1 (WISP1), which facilitates a pro-tumor TME by promoting the survival of CSCs and M2 TAMs, and also promotes CSC maintenance. Abbreviations: ATI, angiotensin I; AT2R, ATII receptor 2; Ang(1–7), angiotensin 1–7; ATIII, angiotensin III; MAPK, mitogen-activated protein kinase. Figure modified and reproduced with permission from the J Histochem Cytochem [19].
3. Glioblastoma Cancer Stem Cells
The CSC concept proposes that a small distinct population of cells within a tumor with self-renewal capability are responsible for driving tumorigenesis [58,59]. These CSCs may be defined as stem cell-like cells within a tumor that also have the capacity for proliferation and multi-potency. This may be regarded as a functional definition insofar as CSCs may be characterized through the generation of serially transplantable tumors that faithfully recapitulate the parent tumor [60]. There is marked intra- and inter-tumoral heterogeneity including, differing numbers of highly tumorigenic CSCs [61]. Such heterogeneity may be best explained by a combination of different models of cancer, including the stochastic model (also known as the clonal evolution model), the CSC concept of cancer (also known as the hierarchical model of cancer), and the concept of plasticity [62,63].
The traditional model of cancer is predicated on the stochastic model of carcinogenesis which proposes that cancer cells are derived from normal cells that acquire genetic and/or epigenetic mutations resulting in typically unidirectional transitions from benign to malignant cells. These malignant tumor cells have unrestricted division capacities and their high mutation rates increase the likelihood of successive generations of cloned cells being adapted to the selection pressures of the tumor site. However, the stochastic model does not fully account for all aspects of cancer biology including tumor recurrence following treatment [64].
In contrast, the CSC concept of cancer proposes that CSCs contribute to carcinogenesis, invasion, metastasis, therapy resistance, and recurrence [65,66]. CSCs divide asymmetrically into non-tumorigenic cancer cells, which form the bulk of a tumor, and identical highly tumorigenic but less abundant CSCs, which sit at the apex of the cellular hierarchy [67]. CSCs have been postulated to originate from non-malignant stem cells or progenitor cells [66] or dedifferentiated cancer cells [68]. The overlap between the stochastic model and the CSC concept may be explained by the concept of cellular plasticity whereby cancer cells may reversibly transition between stem-like and non-stem-like cell states [69]. This process of transition may be driven by embryonic stem cell (ESC)-associated regulatory networks and may be affected by the dynamic TME including the CSC niche [70]. Moreover, certain cancer cells may de-differentiate and re-enter the CSC pool, thus regaining the capacity for tumorigenesis and clonal expansion [71].
CSCs have been found in many different cancer types, including myeloid leukemia [72], pancreatic cancer [73], breast cancer [74], oral cavity squamous cell carcinoma (SCC) [75,76,77], primary [78] and metastatic [79] cutaneous SCC, primary [80] and metastatic [81] colon adenocarcinoma, metastatic malignant melanoma [82,83], and GB [84]. The aggressive nature of GB and its resistance to conventional therapy has been attributed to the presence of CSCs [85] that were first postulated in human brain tumors, identified by their expression of the neural stem cell surface marker CD133 [86]. Stem-like neural precursor cells responsible for the growth and recurrence in serial transplantations were identified in GB [87]. The presence of such quiescent CSCs is well-supported in the literature and the interaction of such cells with the ECM and TME factors, including TGF-β and hypoxia, may contribute to their resistance to conventional therapy [88] (Figure 1). There is evidence that CSCs may be stimulated to differentiate into endothelial cells by activating Notch1 signaling [89] and may be associated with induction of cytokines, MMPs, and adhesion proteins in the TME [90].
A crucial function of stem cells is self-renewal, for which the Notch, Sonic hedgehog, and Wnt signaling pathways may be essential [91] (Figure 1). GB expresses a number of stemness-associated markers including cell surface markers (CD133, CD15, A2B5, and L1CAM), cytoskeletal proteins (nestin), transcription factors (SOX2, NANOG, and OCT4), post-transcriptional factors (Musashi1), and polycomb transcriptional suppressors (Bmi1 and Ezh2) [85]. There is also evidence of plasticity and bi-directional interconversion between CSCs and cancer cells [92]. In a landmark study, pluripotent stem cells were formed from reprogrammed mouse embryonic and adult fibroblasts by the addition of transcription factors OCT4, SOX2, c-MYC, and KLF4 [93]. These factors, in addition to NANOG, which are expressed by ESCs, have been identified in GB [84]. The capacity of GB cells for perpetual self-renewal may rely on the contribution from transcription factors such as OCT4 and SOX2 [85]. SOX2 is highly expressed in GB [84] and may play a key role in maintaining plasticity for bi-directional cellular conversion in GB [94]. Moreover, silencing of SOX2 inhibits tumor proliferation in GB [95] and, thus, it may be a potential therapeutic target in the treatment of GB [96]. Another potential therapeutic target involves the JAK–STAT3 signaling pathway which is also associated with the self-renewal capacity of GB. Inhibition of this pathway may impede the migratory and invasive potential of GB by decreasing activation of the transcription factor STAT3 and, thus, reducing the levels of MMPs and associated invadopodia activity [97]. In addition, STAT3 binding to the Notch1 promoter inhibits this signaling pathway and may impede the maintenance of glioma stem-like cells while reducing the expression of glioma stem cell markers CD133, SOX2, and nestin [98] (Figure 1).
4. The Renin–Angiotensin System and Convergent Signaling Pathways in Glioblastoma
The RAS has been proposed to play an important role in the TME [19] in various cancer types, including lung cancer, through its effect on tumor cells, non-malignant cells, hypoxia, angiogenesis, and the inflammatory response [99]. The RAS is a complex physiological system and has a multitude of interactions with many different convergent signaling pathways that operate in carcinogenesis, some of which lie outside the scope of this article.
Classically, the RAS regulates blood pressure and electrolyte and fluid homeostasis involving primarily the renal, cardiovascular, and endocrine systems [100]. The RAS pathway is composed of multiple steps culminating in the formation of the main effector hormone, angiotensin II (ATII) [101]. Activity of this key homeostatic system in the CNS is well documented [102]. In this review article, RAS inhibition broadly refers to inhibition of any of the components of the RAS, reducing its downstream effects.
Angiotensinogen, primarily synthesized in the liver by hepatocytes, is cleaved by renin, to form angiotensin I (AT1) [103]. Angiotensinogen is synthesized and secreted by astrocytes and is converted to several neuroactive peptides [104,105]. Angiotensinogen is also produced within neurons, which can secrete or retain it intracellularly. These neuroactive peptides bind their respective receptors within the local microenvironment to induce receptor signaling by different cell types [104,105]. Renin is physiologically derived from the juxtaglomerular apparatus in the kidneys and its release is tightly regulated by macula densa and local baroreceptors [106]. Renin is formed by the binding of prorenin to the prorenin receptor (PRR) [107] and is also catalyzed by enzymes such as cathepsins B, D, and G [108,109,110,111]. ATI is converted to ATII by angiotensin-converting enzyme (ACE), also known as ACE1, which is primarily found in the lungs [112]. ATII binds to ATII receptor 1 (AT1R) and ATII receptor 2 (AT2R) [113]. ATII binding to AT1R causes MAPK–STAT3 activation [114] and phosphatidylinositol signaling, which increases cytosolic Ca2+ and effects mitogenesis [115]. AT1R signaling increases RAS activity in the TME, and the formation of NF-κB and TGF-β1 which promotes cellular proliferation, inflammation, and angiogenesis [116]. AT2R activation by ATII inhibits cellular growth and enhances apoptosis [116]. ATII can be further converted into angiotensin III (ATIII), and then angiotensin IV (ATIV) by aminopeptidase-A (AP-A) and aminopeptidase-N (AP-N), respectively. ATIV binds to ATII receptor 4 (AT4R), and in high concentrations, may bind to AT1R. Angiotensin (1–7) (Ang(1–7)) is produced by the cleavage of either ATI by neutral endopeptidase (NEP) or ATII by ACE2, an isoform of ACE. Ang(1–7) binds to Mas receptors (MasRs) [117,118]. ATI may also be cleaved by ACE2 to form Ang(1–9), which can be cleaved by ACE1 and is converted to Ang(1–7), which in addition to binding to MasRs, can also bind to AT2R with low affinity, and Mas-related-G protein coupled receptors (MrgDs) [119]. MrgDs are a recently discovered component of the RAS [102], and their role in the GB TME is yet to be defined. Lastly, the primary ligand for MrgDs is almandine, an Ang(1–7) analog formed by decarboxylation of Ang(1–7) [102] (Figure 2).
Figure 2.
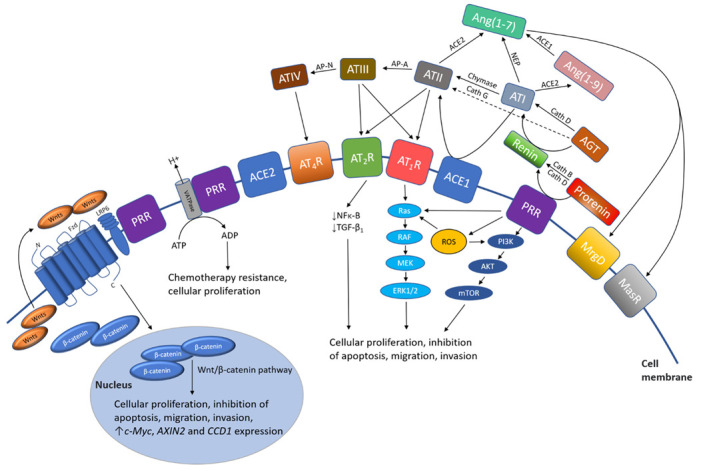
A schema showing the effect of the renin–angiotensin system (RAS) and its convergent signaling pathways on the tumor microenvironment to influence cellular proliferation, invasiveness, and cell survival in cancer development. The RAS interacts with downstream pathways, such as the Ras/RAF/MEK/ERK (light blue) pathway and the PI3K/AKT/mTOR (dark blue) pathway, and the upstream Wnt/β-catenin pathway (intermediate blue) that influence cellular proliferation, migration, inhibition of apoptosis, migration, and invasion (see text). PRR, pro-renin receptor; LRP6, low-density lipoprotein receptor-related protein; Fzd, frizzled receptor; Cath G, cathepsin G; Cath B, cathepsin B; Cath D, cathepsin D; ACE1, angiotensin-converting enzyme 1; ACE2, angiotensin-converting enzyme 2; ADP, adenosine diphosphate; AGT, angiotensinogen; ATP, adenosine triphosphate; Ang(1–7), angiotensin (1–7); Ang(1–9), angiotensin (1–9); AP-A, aminopeptidase-A; NEP, neutral endopeptidase; AP-N, aminopeptidase-N; ATI, angiotensin I; ATII, angiotensin II; ATIII, angiotensin III; ATIV, angiotensin IV; AT1R, angiotensin II receptor 1; AT2R, angiotensin II receptor 2; AT4R, angiotensin II receptor 4; MrgD, Mas-related-G protein coupled receptor; MasR, Mas receptor; mTOR, mammalian target of rapamycin; NF-κB, nuclear factor kappa B; TGF-β1, transforming growth factor-β1; V-ATPase, vacuolar H+-adenosine triphosphate.
Key components of the RAS are also activated in CNS diseases [101]. Renin, and its precursor prorenin, are expressed variably in neurons, astrocytes, oligodendrocytes, and microglia in different regions of the brain [120,121]. PRR is widely distributed in different organs throughout the body including the brain, eyes, and immune system [122]. ACE1 is expressed in areas of the brain involved in blood pressure control and homeostasis including the choroid plexus, organum vasculosum of the lamina terminalis, subfornical organ, and area postrema [104]. ACE2 is found in the endothelium of the brain in various regions including the cortex and brainstem [123]. ACE2 contributes to the neuroprotective ACE2/Ang(1–7)MasR signaling axis by converting ATII to Ang(1–7) which is a ligand for MasR [124].
The RAS, as a constituent of the TME, is involved in several hallmarks of cancer, including angiogenesis, hypoxia, and tumor cell proliferation [125]. Components of the RAS are expressed in different types of cancer including colon adenocarcinoma [126] and malignant melanoma [127]. RAS components are also expressed by CSCs in oral cavity SCC [128,129], renal clear cell carcinoma [130], primary [131], and metastatic [132], cutaneous SCC, metastatic colon adenocarcinoma [133], metastatic malignant melanoma [82,83], and GB [134]. In GB, PRR, AT1R, and AT2R are co-expressed with stemness-associated markers [134]. PRR is highly expressed in GB compared with lower-grade gliomas; this higher expression of PRR in higher-grade glioma is notable as the Wnt/β-catenin signaling pathway is implicated in the self-renewal of stem cells [135] (Figure 1).
The Wnt/β-catenin signaling pathway, which sits downstream of the RAS, is implicated in tumor initiation in several cancer types [136]. In brief, this pathway results in active β-catenin translocating into the nucleus, upregulating the expression of oncogenes such as c-Myc, AXIN2, and CCND1 [136]. PRR is a component of the Wnt receptor complex and acts as an adapter between vacuolar H+-adenosine triphosphate (V-ATPase) and low-density lipoprotein receptor-related protein 6. V-ATPase, a proton pump, is essential for cellular acidification and is involved in the mechanism for β-catenin activation [137]. This process facilitates binding of Wnts to their respective Wnt receptor complex [138]. Further, PRR promotes brain cancers via the Wnt/β-catenin signaling pathway, and in addition to being a membrane receptor, exists in the cytoplasm and increases the protein expression of Wnt2 within glioma cells [135]. This evidence underscores the PRR as a potential oncoprotein via Wnt/β-catenin pathway-related carcinogenesis [136], which influences cell stemness [139], tumorigenesis, and cellular proliferation [140,141]. Renin is expressed in GB and may contribute to the mechanisms of neovascularization in GB [142]. Furthermore, downregulation of the Ang(1–7)/MAS signaling axis by podocalyxin results in enhanced GB cell invasion and proliferation [143]. Finally, bypass loops of the RAS involving various cathepsins that may also contribute to the proliferative activity in GB, for example, cathepsin G coverts ATI to AII and from AGT directly to ATII, which binds to AT1R, to promote cancer progression [144,145,146]. GB CSCs have been shown to secrete Wnt-induced signaling protein 1 (WISP1) that promotes the survival of both the CSCs and M2 TAMs to promote a pro-TME [147] (Figure 1).
Other related signaling pathways, such as the PI3K/AKT/mammalian target of rapamycin (mTOR) and Ras/RAF/MEK/ERK pathways within the GB TME, downstream to the RAS, are activated via AT1R and PRR signal transduction. MAPK/ERK signaling is activated upon binding of renin or prorenin to PRR, and this upregulates ERK1/2 in various cell types including neurons [148]. ERK1/2 activation induces TGF-β1 formation and cellular proliferation, both of which influence cancer development [136]. Supporting this is the fact that silencing of PRR downregulates expression of ERK1/2, AKT, and NF-κB [149]. Additionally, PRR activation leads to the production of reactive oxygen species, which activates both the PI3K/AKT/mTOR and Ras/RAF/MEK/ERK pathways (Figure 2). It is interesting to speculate that both pathways operate in conjunction with the RAS and Wnt/β-catenin to influence proliferation, survival, stemness, and invasiveness of CSCs within the GB TME.
The use of RAS inhibitors (RASis) in the treatment of cancer may mitigate the cytotoxic treatment-related adverse effects experienced by cancer patients to improve their overall quality of life [150]. A meta-analysis of 17 observational studies by Shen et al. [123] show RASis are associated with a reduced risk of cancer [151]. A prospective population-based study also shows long-term (>3 years) administration of RASis is associated with a decreased risk of cancer in patients with a DD genotype, which is associated with high levels of ACE and, thus, increased RAS activity. This is relevant as increased levels of ATII caused by elevated RAS activity promotes cancer progression by its actions on AT1R [152]. Other epidemiological studies have shown a protective benefit of RASis against colorectal cancer [153,154] and an overall reduced risk of cancer [155]. RASis have also been shown to improve the overall survival of patients with aggressive non-metastatic pancreatic ductal adenocarcinoma [156]. Although current data remain inconclusive, RASis appear to be broadly protective against cancer [157].
A retrospective study analyzing clinical data from 810 patients enrolled in two large multicenter studies investigating the role of two drugs targeting the RAS combined with statins in GB, shows no benefit in overall survival [158]. A recent trial on repurposing multiple drugs in combination with temozolomide, including two drugs that affect the RAS (i.e., captopril and celecoxib) for patients with GB, observed maintenance of good quality of life [159]. Captopril, an ACE inhibitor, and celecoxib, which inhibits cyclocoxygenase-2, reduce RAS activity [19]. In addition, RASis, in combination with bevacizumab, improve survival in patients with GB [160], although there is no overall survival benefit of this VEGF inhibitor as a monotherapy for de novo or recurrent GB [161]. PRR may be a critical biomarker and a therapeutic target for the treatment of GB with its connections to V-ATPase function [162], and the Wnt/β-catenin, MAPK/ERK, and PI3K/AKT/mTOR pathways [135,136,149,163] (Figure 1). Several other steps of the RAS pathway can potentially be targeted [164]. The effects of a novel approach, targeting the RAS, its bypass loops, and converging pathways simultaneously using multiple repurposed drugs on the quality of life and progression-free survival in GB patients are currently being investigated in a clinical trial [165]. Therapeutic options may be facilitated by augmenting the compensatory mechanisms of the RAS [136,164,165,166].
5. Recent Developments
Recent technological breakthroughs in generating human cerebral organoids [167] from pluripotent cells, combined with genetic engineering [168], mass spectroscopic proteomics [169], and next generation gene sequencing tools [170], allow more detailed investigation into the GB TME, and the role of the RAS in this niche. Cerebral organoids have been shown to more faithfully recapitulate the temporal and spatial aspects of the developing brain [171,172]. Vascularized cerebral organoids have been developed by utilizing ectopic expression of human ETS variant 2 in engineered ESCs to form a vascular-like network in organoids akin to endothelial cells [173]. In addition, VEGF has been used to induce blood vessel-like structures in cerebral organoids expressing markers associated with the BBB, namely, CD31 and claudin-5 [174]. In addition, human umbilical vein endothelial cells have been used to develop cerebral organoids with a well-developed tubular vascular structure. In another notable development, choroid plexus-like organoids modeled cerebrospinal fluid production with a selective barrier akin to the BBB, which may be used to model the BBTB in the GB TME [175,176,177]. Using RNA sequencing, moreover, GB cerebral organoid models have been shown to best mimic the cellular states and plasticity found in the GB TME compared to gliospheres, tumor organoids, and orthotopic patient-derived xenografts [177].
6. Conclusions
Despite intensive research into the biology and treatment of GB, the prognosis of patients with GB remains dismal. Understanding the heterogeneity of the tumor–host microenvironment in GB, the role of RAS and CSCs, and mapping salient interactions on a cellular level employing techniques, such as single-cell RNA sequencing, may lead to the discovery of potential therapeutic targets [178]. Cerebral and GB organoids represent an exciting yet relatively cost-effective way to delineate relevant signaling pathways within the GB TME, including the RAS, and provide models for developing and testing drug screening and therapeutic targets including RASis [179].
Author Contributions
M.O. and E.J.K. drafted the manuscript. T.M., A.H.K., S.T.T. and A.C.W. critically revised the manuscript. All authors commented on and approved the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. S.T.T. is an inventor of the patents Cancer Diagnosis and Therapy (PCT/NZ2015/050108; AUS/2012302419; JAP/2017528398; US/0281472), Cancer Therapeutic (PCT/NZ2018/050006), and Novel Pharmaceutical Compositions for Cancer Therapy (PCT/NZ2019/050087).
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Louis D.N., Perry A., Reifenberger G., Von Deimling A., Figarella-Branger D., Cavenee W.K., Ohgaki H., Wiestler O.D., Kleihues P., Ellison D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016;131:803–820. doi: 10.1007/s00401-016-1545-1. [DOI] [PubMed] [Google Scholar]
- 2.Crespo I., Vital A.L., Gonzalez-Tablas M., Patino Mdel C., Otero A., Lopes M.C., De Oliveira C., Domingues P., Orfao A., Tabernero M.D. Molecular and Genomic Alterations in Glioblastoma Multiforme. Am. J. Pathol. 2015;185:1820–1833. doi: 10.1016/j.ajpath.2015.02.023. [DOI] [PubMed] [Google Scholar]
- 3.Verhaak R.G., Hoadley K.A., Purdom E., Wang V., Qi Y., Wilkerson M.D., Miller C.R., Ding L., Golub T., Mesirov J.P., et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell. 2010;17:98–110. doi: 10.1016/j.ccr.2009.12.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wang Q., Hu B., Hu X., Kim H., Squatrito M., Scarpace L., DeCarvalho A.C., Lyu S., Li P., Li Y., et al. Tumor Evolution of Glioma-Intrinsic Gene Expression Subtypes Associates with Immunological Changes in the Microenvironment. Cancer Cell. 2017;32:42–56.e46. doi: 10.1016/j.ccell.2017.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Capper D., Zentgraf H., Balss J., Hartmann C., Von Deimling A. Monoclonal antibody specific for IDH1 R132H mutation. Acta Neuropathol. 2009;118:599–601. doi: 10.1007/s00401-009-0595-z. [DOI] [PubMed] [Google Scholar]
- 6.Garrett M., Fujii Y., Osaka N., Ito D., Hirota Y., Sasaki A.T. Emerging Roles of Wild-type and Mutant IDH1 in Growth, Metabolism and Therapeutics of Glioma. In: Debinski W., editor. Gliomas. Exon Publications; Brisbane, Australia: 2021. [DOI] [PubMed] [Google Scholar]
- 7.Stupp R., Mason W.P., Van den Bent M.J., Weller M., Fisher B., Taphoorn M.J., Belanger K., Brandes A.A., Marosi C., Bogdahn U., et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005;352:987–996. doi: 10.1056/NEJMoa043330. [DOI] [PubMed] [Google Scholar]
- 8.Hegi M.E., Diserens A.C., Gorlia T., Hamou M.F., De Tribolet N., Weller M., Kros J.M., Hainfellner J.A., Mason W., Mariani L., et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005;352:997–1003. doi: 10.1056/NEJMoa043331. [DOI] [PubMed] [Google Scholar]
- 9.Binabaj M.M., Bahrami A., ShahidSales S., Joodi M., Joudi Mashhad M., Hassanian S.M., Anvari K., Avan A. The prognostic value of MGMT promoter methylation in glioblastoma: A meta-analysis of clinical trials. J. Cell Physiol. 2018;233:378–386. doi: 10.1002/jcp.25896. [DOI] [PubMed] [Google Scholar]
- 10.Das S., Marsden P.A. Angiogenesis in glioblastoma. N. Engl. J. Med. 2013;369:1561–1563. doi: 10.1056/NEJMcibr1309402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gately L., McLachlan S.A., Dowling A., Philip J. Life beyond a diagnosis of glioblastoma: A systematic review of the literature. J. Cancer Surviv. 2017;11:447–452. doi: 10.1007/s11764-017-0602-7. [DOI] [PubMed] [Google Scholar]
- 12.Simon T., Jackson E., Giamas G. Breaking through the glioblastoma micro-environment via extracellular vesicles. Oncogene. 2020;39:4477–4490. doi: 10.1038/s41388-020-1308-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Awan M., Liu S., Sahgal A., Das S., Chao S.T., Chang E.L., Knisely J.P., Redmond K., Sohn J.W., Machtay M., et al. Extra-CNS metastasis from glioblastoma: A rare clinical entity. Expert Rev. Anticancer Ther. 2015;15:545–552. doi: 10.1586/14737140.2015.1028374. [DOI] [PubMed] [Google Scholar]
- 14.Da Ros M., De Gregorio V., Iorio A.L., Giunti L., Guidi M., De Martino M., Genitori L., Sardi I. Glioblastoma Chemoresistance: The Double Play by Microenvironment and Blood-Brain Barrier. Int. J. Mol. Sci. 2018;19:2879. doi: 10.3390/ijms19102879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Schiffer D., Annovazzi L., Casalone C., Corona C., Mellai M. Glioblastoma: Microenvironment and Niche Concept. Cancers. 2018;11:5. doi: 10.3390/cancers11010005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Balkwill F.R., Capasso M., Hagemann T. The tumor microenvironment at a glance. J. Cell Sci. 2012;125:5591–5596. doi: 10.1242/jcs.116392. [DOI] [PubMed] [Google Scholar]
- 17.Sullivan R., Maresh G., Zhang X., Salomon C., Hooper J., Margolin D., Li L. The Emerging Roles of Extracellular Vesicles As Communication Vehicles within the Tumor Microenvironment and Beyond. Front. Endocrinol. 2017;8:194. doi: 10.3389/fendo.2017.00194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Haznedaroglu I.C., Malkan U.Y. Local bone marrow renin-angiotensin system in the genesis of leukemia and other malignancies. Eur. Rev. Med. Pharmacol. Sci. 2016;20:4089–4111. [PubMed] [Google Scholar]
- 19.Kilmister E.J., Tan S.T. The Role of the Renin-Angiotensin System in the Cancer Stem Cell Niche. J. Histochem. Cytochem. 2021:00221554211026295. doi: 10.1369/00221554211026295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hambardzumyan D., Bergers G. Glioblastoma: Defining Tumor Niches. Trends Cancer. 2015;1:252–265. doi: 10.1016/j.trecan.2015.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bao S., Wu Q., Sathornsumetee S., Hao Y., Li Z., Hjelmeland A.B., Shi Q., McLendon R.E., Bigner D.D., Rich J.N. Stem cell-like glioma cells promote tumor angiogenesis through vascular endothelial growth factor. Cancer Res. 2006;66:7843–7848. doi: 10.1158/0008-5472.CAN-06-1010. [DOI] [PubMed] [Google Scholar]
- 22.Calabrese C., Poppleton H., Kocak M., Hogg T.L., Fuller C., Hamner B., Oh E.Y., Gaber M.W., Finklestein D., Allen M., et al. A perivascular niche for brain tumor stem cells. Cancer Cell. 2007;11:69–82. doi: 10.1016/j.ccr.2006.11.020. [DOI] [PubMed] [Google Scholar]
- 23.Zhang Q., Xiang W., Xue B.Z., Yi D.Y., Zhao H.Y., Fu P. Growth factors contribute to the mediation of angiogenic capacity of glioma-associated mesenchymal stem cells. Oncol. Lett. 2021;21:215. doi: 10.3892/ol.2021.12476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Monteforte A., Lam B., Sherman M.B., Henderson K., Sligar A.D., Spencer A., Tang B., Dunn A.K., Baker A.B. (*) Glioblastoma Exosomes for Therapeutic Angiogenesis in Peripheral Ischemia. Tissue Eng. Part A. 2017;23:1251–1261. doi: 10.1089/ten.tea.2016.0508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Du R., Lu K.V., Petritsch C., Liu P., Ganss R., Passegue E., Song H., Vandenberg S., Johnson R.S., Werb Z., et al. HIF1alpha induces the recruitment of bone marrow-derived vascular modulatory cells to regulate tumor angiogenesis and invasion. Cancer Cell. 2008;13:206–220. doi: 10.1016/j.ccr.2008.01.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wang R., Chadalavada K., Wilshire J., Kowalik U., Hovinga K.E., Geber A., Fligelman B., Leversha M., Brennan C., Tabar V. Glioblastoma stem-like cells give rise to tumour endothelium. Nature. 2010;468:829–833. doi: 10.1038/nature09624. [DOI] [PubMed] [Google Scholar]
- 27.Ricci-Vitiani L., Pallini R., Biffoni M., Todaro M., Invernici G., Cenci T., Maira G., Parati E.A., Stassi G., Larocca L.M., et al. Tumour vascularization via endothelial differentiation of glioblastoma stem-like cells. Nature. 2010;468:824–828. doi: 10.1038/nature09557. [DOI] [PubMed] [Google Scholar]
- 28.Cheng L., Huang Z., Zhou W., Wu Q., Donnola S., Liu J.K., Fang X., Sloan A.E., Mao Y., Lathia J.D., et al. Glioblastoma stem cells generate vascular pericytes to support vessel function and tumor growth. Cell. 2013;153:139–152. doi: 10.1016/j.cell.2013.02.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tamura R., Tanaka T., Akasaki Y., Murayama Y., Yoshida K., Sasaki H. The role of vascular endothelial growth factor in the hypoxic and immunosuppressive tumor microenvironment: Perspectives for therapeutic implications. Med. Oncol. 2019;37:2. doi: 10.1007/s12032-019-1329-2. [DOI] [PubMed] [Google Scholar]
- 30.Van Tellingen O., Yetkin-Arik B., De Gooijer M.C., Wesseling P., Wurdinger T., De Vries H.E. Overcoming the blood-brain tumor barrier for effective glioblastoma treatment. Drug Resist. Updat. 2015;19:1–12. doi: 10.1016/j.drup.2015.02.002. [DOI] [PubMed] [Google Scholar]
- 31.Ballabh P., Braun A., Nedergaard M. The blood-brain barrier: An overview: Structure, regulation, and clinical implications. Neurobiol. Dis. 2004;16:1–13. doi: 10.1016/j.nbd.2003.12.016. [DOI] [PubMed] [Google Scholar]
- 32.Abbott N.J., Patabendige A.A., Dolman D.E., Yusof S.R., Begley D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010;37:13–25. doi: 10.1016/j.nbd.2009.07.030. [DOI] [PubMed] [Google Scholar]
- 33.Bergers G., Song S. The role of pericytes in blood-vessel formation and maintenance. Neuro. Oncol. 2005;7:452–464. doi: 10.1215/S1152851705000232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Monteiro A.R., Hill R., Pilkington G.J., Madureira P.A. The Role of Hypoxia in Glioblastoma Invasion. Cells. 2017;6:45. doi: 10.3390/cells6040045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Rong Y., Durden D.L., Van Meir E.G., Brat D.J. ‘Pseudopalisading’ necrosis in glioblastoma: A familiar morphologic feature that links vascular pathology, hypoxia, and angiogenesis. J. Neuropathol. Exp. Neurol. 2006;65:529–539. doi: 10.1097/00005072-200606000-00001. [DOI] [PubMed] [Google Scholar]
- 36.Seidel S., Garvalov B.K., Wirta V., Von Stechow L., Schanzer A., Meletis K., Wolter M., Sommerlad D., Henze A.T., Nister M., et al. A hypoxic niche regulates glioblastoma stem cells through hypoxia inducible factor 2 alpha. Brain. 2010;133:983–995. doi: 10.1093/brain/awq042. [DOI] [PubMed] [Google Scholar]
- 37.Soeda A., Park M., Lee D., Mintz A., Androutsellis-Theotokis A., McKay R.D., Engh J., Iwama T., Kunisada T., Kassam A.B., et al. Hypoxia promotes expansion of the CD133-positive glioma stem cells through activation of HIF-1alpha. Oncogene. 2009;28:3949–3959. doi: 10.1038/onc.2009.252. [DOI] [PubMed] [Google Scholar]
- 38.Bar E.E., Lin A., Mahairaki V., Matsui W., Eberhart C.G. Hypoxia increases the expression of stem-cell markers and promotes clonogenicity in glioblastoma neurospheres. Am. J. Pathol. 2010;177:1491–1502. doi: 10.2353/ajpath.2010.091021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Guardia G.D.A., Correa B.R., Araujo P.R., Qiao M., Burns S., Penalva L.O.F., Galante P.A.F. Proneural and mesenchymal glioma stem cells display major differences in splicing and lncRNA profiles. NPJ Genom. Med. 2020;5:2. doi: 10.1038/s41525-019-0108-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zagzag D., Lukyanov Y., Lan L., Ali M.A., Esencay M., Mendez O., Yee H., Voura E.B., Newcomb E.W. Hypoxia-inducible factor 1 and VEGF upregulate CXCR4 in glioblastoma: Implications for angiogenesis and glioma cell invasion. Lab. Investig. 2006;86:1221–1232. doi: 10.1038/labinvest.3700482. [DOI] [PubMed] [Google Scholar]
- 41.Bar E.E. Glioblastoma, cancer stem cells and hypoxia. Brain Pathol. 2011;21:119–129. doi: 10.1111/j.1750-3639.2010.00460.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Filatova A., Acker T., Garvalov B.K. The cancer stem cell niche(s): The crosstalk between glioma stem cells and their microenvironment. Biochim. Biophys. Acta. 2013;1830:2496–2508. doi: 10.1016/j.bbagen.2012.10.008. [DOI] [PubMed] [Google Scholar]
- 43.Cui T.X., Kryczek I., Zhao L., Zhao E., Kuick R., Roh M.H., Vatan L., Szeliga W., Mao Y., Thomas D.G., et al. Myeloid-derived suppressor cells enhance stemness of cancer cells by inducing microRNA101 and suppressing the corepressor CtBP2. Immunity. 2013;39:611–621. doi: 10.1016/j.immuni.2013.08.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Feng X., Szulzewsky F., Yerevanian A., Chen Z., Heinzmann D., Rasmussen R.D., Alvarez-Garcia V., Kim Y., Wang B., Tamagno I., et al. Loss of CX3CR1 increases accumulation of inflammatory monocytes and promotes gliomagenesis. Oncotarget. 2015;6:15077–15094. doi: 10.18632/oncotarget.3730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Liang J., Piao Y., Holmes L., Fuller G.N., Henry V., Tiao N., De Groot J.F. Neutrophils promote the malignant glioma phenotype through S100A4. Clin. Cancer Res. 2014;20:187–198. doi: 10.1158/1078-0432.CCR-13-1279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Mi Y., Guo N., Luan J., Cheng J., Hu Z., Jiang P., Jin W., Gao X. The Emerging Role of Myeloid-Derived Suppressor Cells in the Glioma Immune Suppressive Microenvironment. Front. Immunol. 2020;11:737. doi: 10.3389/fimmu.2020.00737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Charles N.A., Holland E.C., Gilbertson R., Glass R., Kettenmann H. The brain tumor microenvironment. Glia. 2011;59:1169–1180. doi: 10.1002/glia.21136. [DOI] [PubMed] [Google Scholar]
- 48.Zhu C., Kros J.M., Cheng C., Mustafa D. The contribution of tumor-associated macrophages in glioma neo-angiogenesis and implications for anti-angiogenic strategies. Neuro. Oncol. 2017;19:1435–1446. doi: 10.1093/neuonc/nox081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Jayasingam S.D., Citartan M., Thang T.H., Mat Zin A.A., Ang K.C., Ch’ng E.S. Evaluating the Polarization of Tumor-Associated Macrophages Into M1 and M2 Phenotypes in Human Cancer Tissue: Technicalities and Challenges in Routine Clinical Practice. Front. Oncol. 2019;9:1512. doi: 10.3389/fonc.2019.01512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Pires-Afonso Y., Niclou S.P., Michelucci A. Revealing and Harnessing Tumour-Associated Microglia/Macrophage Heterogeneity in Glioblastoma. Int. J. Mol. Sci. 2020;21:689. doi: 10.3390/ijms21030689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Pan Y., Yu Y., Wang X., Zhang T. Tumor-Associated Macrophages in Tumor Immunity. Front. Immunol. 2020;11:583084. doi: 10.3389/fimmu.2020.583084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ye X.Z., Xu S.L., Xin Y.H., Yu S.C., Ping Y.F., Chen L., Xiao H.L., Wang B., Yi L., Wang Q.L., et al. Tumor-associated microglia/macrophages enhance the invasion of glioma stem-like cells via TGF-beta1 signaling pathway. J. Immunol. 2012;189:444–453. doi: 10.4049/jimmunol.1103248. [DOI] [PubMed] [Google Scholar]
- 53.Jaiswal S., Jamieson C.H., Pang W.W., Park C.Y., Chao M.P., Majeti R., Traver D., Van Rooijen N., Weissman I.L. CD47 is upregulated on circulating hematopoietic stem cells and leukemia cells to avoid phagocytosis. Cell. 2009;138:271–285. doi: 10.1016/j.cell.2009.05.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hutter G., Theruvath J., Graef C.M., Zhang M., Schoen M.K., Manz E.M., Bennett M.L., Olson A., Azad T.D., Sinha R., et al. Microglia are effector cells of CD47-SIRPalpha antiphagocytic axis disruption against glioblastoma. Proc. Natl. Acad. Sci. USA. 2019;116:997–1006. doi: 10.1073/pnas.1721434116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Jaynes J.M., Sable R., Ronzetti M., Bautista W., Knotts Z., Abisoye-Ogunniyan A., Li D., Calvo R., Dashnyam M., Singh A., et al. Mannose receptor (CD206) activation in tumor-associated macrophages enhances adaptive and innate antitumor immune responses. Sci. Transl. Med. 2020;12:aax6337. doi: 10.1126/scitranslmed.aax6337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wu H., Han Y., Rodriguez Sillke Y., Deng H., Siddiqui S., Treese C., Schmidt F., Friedrich M., Keye J., Wan J., et al. Lipid droplet-dependent fatty acid metabolism controls the immune suppressive phenotype of tumor-associated macrophages. EMBO Mol. Med. 2019;11:e10698. doi: 10.15252/emmm.201910698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Azambuja J.H., Ludwig N., Yerneni S.S., Braganhol E., Whiteside T.L. Arginase-1+ Exosomes from Reprogrammed Macrophages Promote Glioblastoma Progression. Int. J. Mol. Sci. 2020;21:3990. doi: 10.3390/ijms21113990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Reya T., Morrison S.J., Clarke M.F., Weissman I.L. Stem cells, cancer, and cancer stem cells. Nature. 2001;414:105–111. doi: 10.1038/35102167. [DOI] [PubMed] [Google Scholar]
- 59.Clarke M.F., Dick J.E., Dirks P.B., Eaves C.J., Jamieson C.H., Jones D.L., Visvader J., Weissman I.L., Wahl G.M. Cancer stem cells--perspectives on current status and future directions: AACR Workshop on cancer stem cells. Cancer Res. 2006;66:9339–9344. doi: 10.1158/0008-5472.CAN-06-3126. [DOI] [PubMed] [Google Scholar]
- 60.Tang D.G. Understanding cancer stem cell heterogeneity and plasticity. Cell Res. 2012;22:457–472. doi: 10.1038/cr.2012.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Adams J.M., Strasser A. Is tumor growth sustained by rare cancer stem cells or dominant clones? Cancer Res. 2008;68:4018–4021. doi: 10.1158/0008-5472.CAN-07-6334. [DOI] [PubMed] [Google Scholar]
- 62.Rich J.N. Cancer stem cells: Understanding tumor hierarchy and heterogeneity. Medecine. 2016;95:S2–S7. doi: 10.1097/MD.0000000000004764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Najafi M., Mortezaee K., Ahadi R. Cancer stem cell (a)symmetry & plasticity: Tumorigenesis and therapy relevance. Life Sci. 2019;231:116520. doi: 10.1016/j.lfs.2019.05.076. [DOI] [PubMed] [Google Scholar]
- 64.Aponte P.M., Caicedo A. Stemness in Cancer: Stem Cells, Cancer Stem Cells, and Their Microenvironment. Stem Cells Int. 2017;2017:5619472. doi: 10.1155/2017/5619472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Gurel C., Inetas G., Hortu I., Tunc E., Kuscu G.C., Dindaroglu F.C., Sahin O., Buhur A., Oktem G. Cancer and Cancer Stem Cells: New Molecular Perspectives. Crit. Rev. Oncog. 2019;24:99–104. doi: 10.1615/CritRevOncog.2019029628. [DOI] [PubMed] [Google Scholar]
- 66.Clara J.A., Monge C., Yang Y., Takebe N. Targeting signalling pathways and the immune microenvironment of cancer stem cells—A clinical update. Nat. Rev. Clin. Oncol. 2020;17:204–232. doi: 10.1038/s41571-019-0293-2. [DOI] [PubMed] [Google Scholar]
- 67.Bradshaw A., Wickremsekera A., Tan S.T., Peng L., Davis P.F., Itinteang T. Cancer Stem Cell Hierarchy in Glioblastoma Multiforme. Front. Surg. 2016;3:21. doi: 10.3389/fsurg.2016.00021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Eun K., Ham S.W., Kim H. Cancer stem cell heterogeneity: Origin and new perspectives on CSC targeting. BMB Rep. 2017;50:117–125. doi: 10.5483/BMBRep.2017.50.3.222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Plaks V., Kong N., Werb Z. The cancer stem cell niche: How essential is the niche in regulating stemness of tumor cells? Cell Stem Cell. 2015;16:225–238. doi: 10.1016/j.stem.2015.02.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Quail D.F., Taylor M.J., Postovit L.M. Microenvironmental regulation of cancer stem cell phenotypes. Curr. Stem Cell Res. Ther. 2012;7:197–216. doi: 10.2174/157488812799859838. [DOI] [PubMed] [Google Scholar]
- 71.Chaffer C.L., Weinberg R.A. How does multistep tumorigenesis really proceed? Cancer Discov. 2015;5:22–24. doi: 10.1158/2159-8290.CD-14-0788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Zhao C., Chen A., Jamieson C.H., Fereshteh M., Abrahamsson A., Blum J., Kwon H.Y., Kim J., Chute J.P., Rizzieri D., et al. Hedgehog signalling is essential for maintenance of cancer stem cells in myeloid leukaemia. Nature. 2009;458:776–779. doi: 10.1038/nature07737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Li C., Heidt D.G., Dalerba P., Burant C.F., Zhang L., Adsay V., Wicha M., Clarke M.F., Simeone D.M. Identification of pancreatic cancer stem cells. Cancer Res. 2007;67:1030–1037. doi: 10.1158/0008-5472.CAN-06-2030. [DOI] [PubMed] [Google Scholar]
- 74.Al-Hajj M., Wicha M.S., Benito-Hernandez A., Morrison S.J., Clarke M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA. 2003;100:3983–3988. doi: 10.1073/pnas.0530291100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Baillie R., Itinteang T., Yu H.H., Brasch H.D., Davis P.F., Tan S.T. Cancer stem cells in moderately differentiated oral tongue squamous cell carcinoma. J. Clin. Pathol. 2016;69:742–744. doi: 10.1136/jclinpath-2015-203599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Yu H.H., Featherston T., Tan S.T., Chibnall A.M., Brasch H.D., Davis P.F., Itinteang T. Characterization of cancer stem cells in moderately differentiated buccal mucosal squamous cell carcinoma. Front. Surg. 2016;3:46. doi: 10.3389/fsurg.2016.00046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Ram R., Brasch H.D., Dunne J.C., Davis P.F., Tan S.T., Itinteang T. The identification of three cancer stem cell subpopulations within moderately differentiated lip squamous cell carcinoma. Front. Surg. 2017;4:12. doi: 10.3389/fsurg.2017.00012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Koh S.P., Brasch H.D., De Jongh J., Itinteang T., Tan S.T. Cancer stem cell subpopulations in moderately differentiated head and neck cutaneous squamous cell carcinoma. Heliyon. 2019;5:e02257. doi: 10.1016/j.heliyon.2019.e02257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Kilmister E.J., Patel J., Van Schaijik B., Bockett N., Brasch H.D., Paterson E., Sim D., Davis P.F., Roth I.M., Itinteang T., et al. Cancer Stem Cell Subpopulations Are Present Within Metastatic Head and Neck Cutaneous Squamous Cell Carcinoma. Front. Oncol. 2020;10:1091. doi: 10.3389/fonc.2020.01091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Munro M.J., Wickremesekera S.K., Peng L., Marsh R.W., Itinteang T., Tan S.T. Cancer stem cell subpopulations in primary colon adenocarcinoma. PLoS ONE. 2019;14:e0221963. doi: 10.1371/journal.pone.0221963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Humphries H.N., Wickremesekera S.K., Marsh R.W., Brasch H.D., Mehrotra S., Tan S.T., Itinteang T. Characterization of Cancer Stem Cells in Colon Adenocarcinoma Metastasis to the Liver. Front. Surg. 2018;4:76. doi: 10.3389/fsurg.2017.00076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Wickremesekera A.C., Brasch H.D., Lee V.M., Davis P.F., Parker A., Koeck H., Itinteang T., Tan S.T. Cancer stem cell subpopulations in metastatic melanoma to the brain express components of the renin-angiotensin system. J. Cancer Metastasis Treat. 2019;5:62. doi: 10.20517/2394-4722.2019.009. [DOI] [Google Scholar]
- 83.Yoganandarajah V., Patel J., Van Schaijik B., Bockett N., Brasch H.D., Paterson E., Sim D., Davis P.F., Roth I.M., Itinteang T., et al. Identification of Cancer Stem Cell Subpopulations in Head and Neck Metastatic Malignant Melanoma. Cells. 2020;9:324. doi: 10.3390/cells9020324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Bradshaw A., Wickremesekera A., Brasch H.D., Chibnall A.M., Davis P.F., Tan S.T., Itinteang T. Cancer Stem Cells in Glioblastoma Multiforme. Front. Surg. 2016;3:48. doi: 10.3389/fsurg.2016.00048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kalkan R. Glioblastoma Stem Cells as a New Therapeutic Target for Glioblastoma. Clin. Med. Insights. Oncol. 2015;9:95–103. doi: 10.4137/CMO.S30271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Singh S.K., Clarke I.D., Terasaki M., Bonn V.E., Hawkins C., Squire J., Dirks P.B. Identification of a cancer stem cell in human brain tumors. Cancer Res. 2003;63:5821–5828. [PubMed] [Google Scholar]
- 87.Galli R., Binda E., Orfanelli U., Cipelletti B., Gritti A., De Vitis S., Fiocco R., Foroni C., Dimeco F., Vescovi A. Isolation and characterization of tumorigenic, stem-like neural precursors from human glioblastoma. Cancer Res. 2004;64:7011–7021. doi: 10.1158/0008-5472.CAN-04-1364. [DOI] [PubMed] [Google Scholar]
- 88.Tejero R., Huang Y., Katsyv I., Kluge M., Lin J.Y., Tome-Garcia J., Daviaud N., Wang Y., Zhang B., Tsankova N.M., et al. Gene signatures of quiescent glioblastoma cells reveal mesenchymal shift and interactions with niche microenvironment. EBioMedicine. 2019;42:252–269. doi: 10.1016/j.ebiom.2019.03.064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Cui C., Chen X., Liu Y., Cao B., Xing Y., Liu C., Yang F., Li Y., Yang T., Hua L., et al. beta1,4-Galactosyltransferase V activates Notch1 signaling in glioma stem-like cells and promotes their transdifferentiation into endothelial cells. J. Biol. Chem. 2018;293:2219–2230. doi: 10.1074/jbc.RA117.000682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Guichet P.O., Guelfi S., Teigell M., Hoppe L., Bakalara N., Bauchet L., Duffau H., Lamszus K., Rothhut B., Hugnot J.P. Notch1 stimulation induces a vascularization switch with pericyte-like cell differentiation of glioblastoma stem cells. Stem Cells. 2015;33:21–34. doi: 10.1002/stem.1767. [DOI] [PubMed] [Google Scholar]
- 91.Iwadate Y. Plasticity in Glioma Stem Cell Phenotype and Its Therapeutic Implication. Neurol. Med. Chir. 2018;58:61–70. doi: 10.2176/nmc.ra.2017-0089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Safa A.R., Saadatzadeh M.R., Cohen-Gadol A.A., Pollok K.E., Bijangi-Vishehsaraei K. Glioblastoma stem cells (GSCs) epigenetic plasticity and interconversion between differentiated non-GSCs and GSCs. Genes Dis. 2015;2:152–163. doi: 10.1016/j.gendis.2015.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Takahashi K., Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–676. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 94.Berezovsky A.D., Poisson L.M., Cherba D., Webb C.P., Transou A.D., Lemke N.W., Hong X., Hasselbach L.A., Irtenkauf S.M., Mikkelsen T., et al. Sox2 promotes malignancy in glioblastoma by regulating plasticity and astrocytic differentiation. Neoplasia. 2014;16:193–206.e25. doi: 10.1016/j.neo.2014.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Gangemi R.M., Griffero F., Marubbi D., Perera M., Capra M.C., Malatesta P., Ravetti G.L., Zona G.L., Daga A., Corte G. SOX2 silencing in glioblastoma tumor-initiating cells causes stop of proliferation and loss of tumorigenicity. Stem Cells. 2009;27:40–48. doi: 10.1634/stemcells.2008-0493. [DOI] [PubMed] [Google Scholar]
- 96.Garros-Regulez L., Garcia I., Carrasco-Garcia E., Lantero A., Aldaz P., Moreno-Cugnon L., Arrizabalaga O., Undabeitia J., Torres-Bayona S., Villanua J., et al. Targeting SOX2 as a Therapeutic Strategy in Glioblastoma. Front. Oncol. 2016;6:222. doi: 10.3389/fonc.2016.00222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Senft C., Priester M., Polacin M., Schroder K., Seifert V., Kogel D., Weissenberger J. Inhibition of the JAK-2/STAT3 signaling pathway impedes the migratory and invasive potential of human glioblastoma cells. J. Neurooncol. 2011;101:393–403. doi: 10.1007/s11060-010-0273-y. [DOI] [PubMed] [Google Scholar]
- 98.Yahyanejad S., King H., Iglesias V.S., Granton P.V., Barbeau L.M., Van Hoof S.J., Groot A.J., Habets R., Prickaerts J., Chalmers A.J., et al. NOTCH blockade combined with radiation therapy and temozolomide prolongs survival of orthotopic glioblastoma. Oncotarget. 2016;7:41251–41264. doi: 10.18632/oncotarget.9275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Catarata M.J., Ribeiro R., Oliveira M.J., Robalo Cordeiro C., Medeiros R. Renin-Angiotensin System in Lung Tumor and Microenvironment Interactions. Cancers. 2020;12:1457. doi: 10.3390/cancers12061457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Sparks M.A., Crowley S.D., Gurley S.B., Mirotsou M., Coffman T.M. Classical Renin-Angiotensin system in kidney physiology. Compr. Physiol. 2014;4:1201–1228. doi: 10.1002/cphy.c130040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Nakagawa P., Gomez J., Grobe J.L., Sigmund C.D. The Renin-Angiotensin System in the Central Nervous System and Its Role in Blood Pressure Regulation. Curr. Hypertens. Rep. 2020;22:7. doi: 10.1007/s11906-019-1011-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Jackson L., Eldahshan W., Fagan S.C., Ergul A. Within the Brain: The Renin Angiotensin System. Int. J. Mol. Sci. 2018;19:876. doi: 10.3390/ijms19030876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Cassis L.A., Saye J., Peach M.J. Location and regulation of rat angiotensinogen messenger RNA. Hypertension. 1988;11:591–596. doi: 10.1161/01.HYP.11.6.591. [DOI] [PubMed] [Google Scholar]
- 104.Bodiga V.L., Bodiga S. Renin Angiotensin System in Cognitive Function and Dementia. Asian J. Neurosci. 2013;2013:1–18. doi: 10.1155/2013/102602. [DOI] [Google Scholar]
- 105.Grobe J.L., Xu D., Sigmund C.D. An intracellular renin-angiotensin system in neurons: Fact, hypothesis, or fantasy. Physiology. 2008;23:187–193. doi: 10.1152/physiol.00002.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Kopp U.C., DiBona G.F. Neural regulation of renin secretion. Semin. Nephrol. 1993;13:543–551. [PubMed] [Google Scholar]
- 107.Riquier-Brison A.D.M., Sipos A., Prokai A., Vargas S.L., Toma L., Meer E.J., Villanueva K.G., Chen J.C.M., Gyarmati G., Yih C., et al. The macula densa prorenin receptor is essential in renin release and blood pressure control. Am. J. Physiol. Renal. Physiol. 2018;315:F521–F534. doi: 10.1152/ajprenal.00029.2018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Jutras I., Reudelhuber T.L. Prorenin processing by cathepsin B in vitro and in transfected cells. FEBS Lett. 1999;443:48–52. doi: 10.1016/S0014-5793(98)01672-X. [DOI] [PubMed] [Google Scholar]
- 109.Neves F.A., Duncan K.G., Baxter J.D. Cathepsin B is a prorenin processing enzyme. Hypertension. 1996;27:514–517. doi: 10.1161/01.HYP.27.3.514. [DOI] [PubMed] [Google Scholar]
- 110.Naseem R.H., Hedegard W., Henry T.D., Lessard J., Sutter K., Katz S.A. Plasma cathepsin D isoforms and their active metabolites increase after myocardial infarction and contribute to plasma renin activity. Basic Res. Cardiol. 2005;100:139–146. doi: 10.1007/s00395-004-0499-3. [DOI] [PubMed] [Google Scholar]
- 111.Rykl J., Thiemann J., Kurzawski S., Pohl T., Gobom J., Zidek W., Schluter H. Renal cathepsin G and angiotensin II generation. J. Hypertens. 2006;24:1797–1807. doi: 10.1097/01.hjh.0000242404.91332.be. [DOI] [PubMed] [Google Scholar]
- 112.Balyasnikova I.V., Metzger R., Sun Z.L., Berestetskaya Y.V., Albrecht R.F., Danilov S.M. Development and characterization of rat monoclonal antibodies to denatured mouse angiotensin-converting enzyme. Tissue Antigens. 2005;65:240–251. doi: 10.1111/j.1399-0039.2005.00364.x. [DOI] [PubMed] [Google Scholar]
- 113.Nakagawa P., Sigmund C.D. How Is the Brain Renin-Angiotensin System Regulated? Hypertension. 2017;70:10–18. doi: 10.1161/HYPERTENSIONAHA.117.08550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Hashemzehi M., Beheshti F., Hassanian S.M., Ferns G.A., Khazaei M., Avan A. Therapeutic potential of renin angiotensin system inhibitors in cancer cells metastasis. Pathol. Res. Pract. 2020;216:153010. doi: 10.1016/j.prp.2020.153010. [DOI] [PubMed] [Google Scholar]
- 115.Yu C., Tang W., Wang Y., Shen Q., Wang B., Cai C., Meng X., Zou F. Downregulation of ACE2/Ang-(1-7)/Mas axis promotes breast cancer metastasis by enhancing store-operated calcium entry. Cancer Lett. 2016;376:268–277. doi: 10.1016/j.canlet.2016.04.006. [DOI] [PubMed] [Google Scholar]
- 116.Ziaja M., Urbanek K.A., Kowalska K., Piastowska-Ciesielska A.W. Angiotensin II and Angiotensin Receptors 1 and 2-Multifunctional System in Cells Biology, What Do We Know? Cells. 2021;10:381. doi: 10.3390/cells10020381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Bernstein K.E., Koronyo Y., Salumbides B.C., Sheyn J., Pelissier L., Lopes D.H., Shah K.H., Bernstein E.A., Fuchs D.T., Yu J.J., et al. Angiotensin-converting enzyme overexpression in myelomonocytes prevents Alzheimer’s-like cognitive decline. J. Clin. Investig. 2014;124:1000–1012. doi: 10.1172/JCI66541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Elased K.M., Cunha T.S., Marcondes F.K., Morris M. Brain angiotensin-converting enzymes: Role of angiotensin-converting enzyme 2 in processing angiotensin II in mice. Exp. Physiol. 2008;93:665–675. doi: 10.1113/expphysiol.2007.040311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Tetzner A., Gebolys K., Meinert C., Klein S., Uhlich A., Trebicka J., Villacañas Ó., Walther T. G-Protein–Coupled Receptor MrgD Is a Receptor for Angiotensin-(1–7) Involving Adenylyl Cyclase, cAMP, and Phosphokinase A. Hypertension. 2016;68:185–194. doi: 10.1161/HYPERTENSIONAHA.116.07572. [DOI] [PubMed] [Google Scholar]
- 120.Labandeira-Garcia J.L., Rodriguez-Perez A.I., Garrido-Gil P., Rodriguez-Pallares J., Lanciego J.L., Guerra M.J. Brain Renin-Angiotensin System and Microglial Polarization: Implications for Aging and Neurodegeneration. Front. Aging Neurosci. 2017;9:129. doi: 10.3389/fnagi.2017.00129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Xu Q., Jensen D.D., Peng H., Feng Y. The critical role of the central nervous system (pro)renin receptor in regulating systemic blood pressure. Pharmacol. Ther. 2016;164:126–134. doi: 10.1016/j.pharmthera.2016.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Ichihara A., Yatabe M.S. The (pro)renin receptor in health and disease. Nat. Rev. Nephrol. 2019;15:693–712. doi: 10.1038/s41581-019-0160-5. [DOI] [PubMed] [Google Scholar]
- 123.Hamming I., Timens W., Bulthuis M.L., Lely A.T., Navis G., Van Goor H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004;203:631–637. doi: 10.1002/path.1570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Cui C., Xu P., Li G., Qiao Y., Han W., Geng C., Liao D., Yang M., Chen D., Jiang P. Vitamin D receptor activation regulates microglia polarization and oxidative stress in spontaneously hypertensive rats and angiotensin II-exposed microglial cells: Role of renin-angiotensin system. Redox Biol. 2019;26:101295. doi: 10.1016/j.redox.2019.101295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Wegman-Ostrosky T., Soto-Reyes E., Vidal-Millan S., Sanchez-Corona J. The renin-angiotensin system meets the hallmarks of cancer. J. Renin Angiotensin Aldosterone Syst. 2015;16:227–233. doi: 10.1177/1470320313496858. [DOI] [PubMed] [Google Scholar]
- 126.Beitia M., Solano-Iturri J.D., Errarte P., Sanz B., Perez I., Etxezarraga M.C., Loizate A., Asumendi A., Larrinaga G. Altered expression of renin-angiotensin system receptors throughout colorectal adenoma-adenocarcinoma sequence. Int. J. Med. Sci. 2019;16:813–821. doi: 10.7150/ijms.32599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Renziehausen A., Wang H., Rao B., Weir L., Nigro C.L., Lattanzio L., Merlano M., Vega-Rioja A., Del Carmen Fernandez-Carranco M., Hajji N., et al. The renin angiotensin system (RAS) mediates bifunctional growth regulation in melanoma and is a novel target for therapeutic intervention. Oncogene. 2019;38:2320–2336. doi: 10.1038/s41388-018-0563-y. [DOI] [PubMed] [Google Scholar]
- 128.Featherston T., Yu H.H., Dunne J.C., Chibnall A.M., Brasch H.D., Davis P.F., Tan S.T., Itinteang T. Cancer Stem Cells in Moderately Differentiated Buccal Mucosal Squamous Cell Carcinoma Express Components of the Renin-Angiotensin System. Front. Surg. 2016;3:52. doi: 10.3389/fsurg.2016.00052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Itinteang T., Dunne J.C., Chibnall A.M., Brasch H.D., Davis P.F., Tan S.T. Cancer stem cells in moderately differentiated oral tongue squamous cell carcinoma express components of the renin-angiotensin system. J. Clin. Pathol. 2016;69:942–945. doi: 10.1136/jclinpath-2016-203736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Siljee S., Milne B., Brasch H.D., Bockett N., Patel J., Davis P.F., Kennedy-Smith A., Itinteang T., Tan S.T. Expression of Components of the Renin-Angiotensin System by Cancer Stem Cells in Renal Clear Cell Carcinoma. Biomolecules. 2021;11:537. doi: 10.3390/biom11040537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Nallaiah S., Lee V.M.Y., Brasch H.D., De Jongh J., Schaijik B.V., Marsh R., Tan S.T., Itinteang T. Cancer stem cells within moderately differentiated head and neck cutaneous squamous cell carcinoma express components of the renin-angiotensin system. J. Plast. Reconstr. Aesthet. Surg. 2019;72:1484–1493. doi: 10.1016/j.bjps.2018.11.013. [DOI] [PubMed] [Google Scholar]
- 132.Siljee S., Buchanan O., Brasch H.D., Bockett N., Patel J., Paterson E., Purdie G.L., Davis P.F., Itinteang T., Tan S.T. Cancer Stem Cells in Metastatic Head and Neck Cutaneous Squamous Cell Carcinoma Express Components of the Renin-Angiotensin System. Cells. 2021;10:243. doi: 10.3390/cells10020243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Narayanan A., Wickremesekera S.K., Van Schaijik B., Marsh R.W., Brasch H.D., Tan S.T., Itinteang T. Cancer stem cells in liver metastasis from colon adenocarcinoma express components of the renin-angiotensin system. J. Cancer Metastasis Treat. 2019;5:36. doi: 10.20517/2394-4722.2018.77. [DOI] [Google Scholar]
- 134.Bradshaw A.R., Wickremesekera A.C., Brasch H.D., Chibnall A.M., Davis P.F., Tan S.T., Itinteang T. Glioblastoma Multiforme Cancer Stem Cells Express Components of the Renin-Angiotensin System. Front. Surg. 2016;3:51. doi: 10.3389/fsurg.2016.00051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Kouchi M., Shibayama Y., Ogawa D., Miyake K., Nishiyama A., Tamiya T. (Pro)renin receptor is crucial for glioma development via the Wnt/beta-catenin signaling pathway. J. Neurosurg. 2017;127:819–828. doi: 10.3171/2016.9.JNS16431. [DOI] [PubMed] [Google Scholar]
- 136.Wang J., Nishiyama A., Matsuyama M., Wang Z., Yuan Y. The (pro)renin receptor: A novel biomarker and potential therapeutic target for various cancers. Cell Commun. Signal. 2020;18:39. doi: 10.1186/s12964-020-0531-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Nguyen G. Renin, (pro)renin and receptor: An update. Clin. Sci. 2011;120:169–178. doi: 10.1042/CS20100432. [DOI] [PubMed] [Google Scholar]
- 138.Cruciat C.M., Ohkawara B., Acebron S.P., Karaulanov E., Reinhard C., Ingelfinger D., Boutros M., Niehrs C. Requirement of prorenin receptor and vacuolar H+-ATPase-mediated acidification for Wnt signaling. Science. 2010;327:459–463. doi: 10.1126/science.1179802. [DOI] [PubMed] [Google Scholar]
- 139.Zheng H., Ying H., Wiedemeyer R., Yan H., Quayle S.N., Ivanova E.V., Paik J.-H., Zhang H., Xiao Y., Perry S.R., et al. PLAGL2 regulates Wnt signaling to impede differentiation in neural stem cells and gliomas. Cancer Cell. 2010;17:497–509. doi: 10.1016/j.ccr.2010.03.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Pu P., Zhang Z., Kang C., Jiang R., Jia Z., Wang G., Jiang H. Downregulation of Wnt2 and beta-catenin by siRNA suppresses malignant glioma cell growth. Cancer Gene Ther. 2009;16:351–361. doi: 10.1038/cgt.2008.78. [DOI] [PubMed] [Google Scholar]
- 141.Pulvirenti T., Van Der Heijden M., Droms L.A., Huse J.T., Tabar V., Hall A. Dishevelled 2 Signaling Promotes Self-Renewal and Tumorigenicity in Human Gliomas. Cancer Res. 2011;71:7280. doi: 10.1158/0008-5472.CAN-11-1531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Ariza A., Fernandez L.A., Inagami T., Kim J.H., Manuelidis E.E. Renin in glioblastoma multiforme and its role in neovascularization. Am. J. Clin. Pathol. 1988;90:437–441. doi: 10.1093/ajcp/90.4.437. [DOI] [PubMed] [Google Scholar]
- 143.Liu B., Liu Y., Jiang Y. Podocalyxin promotes glioblastoma multiforme cell invasion and proliferation by inhibiting angiotensin-(1-7)/Mas signaling. Oncol. Rep. 2015;33:2583–2591. doi: 10.3892/or.2015.3813. [DOI] [PubMed] [Google Scholar]
- 144.Gopinath S., Malla R., Alapati K., Gorantla B., Gujrati M., Dinh D.H., Rao J.S. Cathepsin B and uPAR regulate self-renewal of glioma-initiating cells through GLI-regulated Sox2 and Bmi1 expression. Carcinogenesis. 2013;34:550–559. doi: 10.1093/carcin/bgs375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Cheng Y.C., Ding Y.M., Hueng D.Y., Chen J.Y., Chen Y. Caffeine suppresses the progression of human glioblastoma via cathepsin B and MAPK signaling pathway. J. Nutr. Biochem. 2016;33:63–72. doi: 10.1016/j.jnutbio.2016.03.004. [DOI] [PubMed] [Google Scholar]
- 146.Koh S.P., Wickremesekera A.C., Brasch H.D., Marsh R., Tan S.T., Itinteang T. Expression of Cathepsins B, D, and G in Isocitrate Dehydrogenase-Wildtype Glioblastoma. Front. Surg. 2017;4:28. doi: 10.3389/fsurg.2017.00028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Tao W., Chu C., Zhou W., Huang Z., Zhai K., Fang X., Huang Q., Zhang A., Wang X., Yu X., et al. Dual Role of WISP1 in maintaining glioma stem cells and tumor-supportive macrophages in glioblastoma. Nat. Commun. 2020;11:3015. doi: 10.1038/s41467-020-16827-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Krop M., Lu X., Danser A.H., Meima M.E. The (pro)renin receptor. A decade of research: What have we learned? Pflugers Arch. 2013;465:87–97. doi: 10.1007/s00424-012-1105-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Arundhathi A., Chuang W.H., Chen J.K., Wang S.E., Shyr Y.M., Chen J.Y., Liao W.N., Chen H.W., Teng Y.M., Pai C.C., et al. Prorenin receptor acts as a potential molecular target for pancreatic ductal adenocarcinoma diagnosis. Oncotarget. 2016 doi: 10.18632/oncotarget.10583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Pinter M., Kwanten W.J., Jain R.K. Renin-Angiotensin System Inhibitors to Mitigate Cancer Treatment-Related Adverse Events. Clin. Cancer Res. 2018;24:3803–3812. doi: 10.1158/1078-0432.CCR-18-0236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Shen J., Huang Y.M., Wang M., Hong X.Z., Song X.N., Zou X., Pan Y.H., Ling W., Zhu M.H., Zhang X.X., et al. Renin-angiotensin system blockade for the risk of cancer and death. J. Renin Angiotensin Aldosterone Syst. 2016;17 doi: 10.1177/1470320316656679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Van der Knaap R., Siemes C., Coebergh J.W., Van Duijn C.M., Hofman A., Stricker B.H. Renin-angiotensin system inhibitors, angiotensin I-converting enzyme gene insertion/deletion polymorphism, and cancer: The Rotterdam Study. Cancer. 2008;112:748–757. doi: 10.1002/cncr.23215. [DOI] [PubMed] [Google Scholar]
- 153.Makar G.A., Holmes J.H., Yang Y.X. Angiotensin-converting enzyme inhibitor therapy and colorectal cancer risk. J. Natl. Cancer Inst. 2014;106:djt374. doi: 10.1093/jnci/djt374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Dai Y.N., Wang J.H., Zhu J.Z., Lin J.Q., Yu C.H., Li Y.M. Angiotensin-converting enzyme inhibitors/angiotensin receptor blockers therapy and colorectal cancer: A systematic review and meta-analysis. Cancer Causes Control. 2015;26:1245–1255. doi: 10.1007/s10552-015-0617-1. [DOI] [PubMed] [Google Scholar]
- 155.Chiang Y.Y., Chen K.B., Tsai T.H., Tsai W.C. Lowered cancer risk with ACE inhibitors/ARBs: A population-based cohort study. J. Clin. Hypertens. 2014;16:27–33. doi: 10.1111/jch.12228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Liu H., Naxerova K., Pinter M., Incio J., Lee H., Shigeta K., Ho W.W., Crain J.A., Jacobson A., Michelakos T., et al. Use of Angiotensin System Inhibitors Is Associated with Immune Activation and Longer Survival in Nonmetastatic Pancreatic Ductal Adenocarcinoma. Clin. Cancer Res. 2017;23:5959–5969. doi: 10.1158/1078-0432.CCR-17-0256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Rosenthal T., Gavras I. Renin-Angiotensin Inhibition in Combating Malignancy: A Review. Anticancer Res. 2019;39:4597–4602. doi: 10.21873/anticanres.13639. [DOI] [PubMed] [Google Scholar]
- 158.Happold C., Gorlia T., Nabors L.B., Erridge S.C., Reardon D.A., Hicking C., Picard M., Stupp R., Weller M., Group E.B.T., et al. Do statins, ACE inhibitors or sartans improve outcome in primary glioblastoma? J. Neurooncol. 2018;138:163–171. doi: 10.1007/s11060-018-2786-8. [DOI] [PubMed] [Google Scholar]
- 159.Kast R.E., Karpel-Massler G., Halatsch M.E. CUSP9* treatment protocol for recurrent glioblastoma: Aprepitant, artesunate, auranofin, captopril, celecoxib, disulfiram, itraconazole, ritonavir, sertraline augmenting continuous low dose temozolomide. Oncotarget. 2014;5:8052–8082. doi: 10.18632/oncotarget.2408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Levin V.A., Chan J., Datta M., Yee J.L., Jain R.K. Effect of angiotensin system inhibitors on survival in newly diagnosed glioma patients and recurrent glioblastoma patients receiving chemotherapy and/or bevacizumab. J. Neurooncol. 2017;134:325–330. doi: 10.1007/s11060-017-2528-3. [DOI] [PubMed] [Google Scholar]
- 161.Ameratunga M., Pavlakis N., Wheeler H., Grant R., Simes J., Khasraw M. Anti-angiogenic therapy for high-grade glioma. Cochrane Database Syst. Rev. 2018;11:CD008218. doi: 10.1002/14651858.CD008218.pub4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Burcklé C., Bader M. Prorenin and its ancient receptor. Hypertension. 2006;48:549–551. doi: 10.1161/01.HYP.0000241132.48495.df. [DOI] [PubMed] [Google Scholar]
- 163.Juillerat-Jeanneret L., Celerier J., Chapuis Bernasconi C., Nguyen G., Wostl W., Maerki H.P., Janzer R.C., Corvol P., Gasc J.M. Renin and angiotensinogen expression and functions in growth and apoptosis of human glioblastoma. Br. J. Cancer. 2004;90:1059–1068. doi: 10.1038/sj.bjc.6601646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Roth I.M., Wickremesekera A.C., Wickremesekera S.K., Davis P.F., Tan S.T. Therapeutic Targeting of Cancer Stem Cells via Modulation of the Renin-Angiotensin System. Front. Oncol. 2019;9:745. doi: 10.3389/fonc.2019.00745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Tan D.C., Roth I.M., Wickremesekera A.C., Davis P.F., Kaye A.H., Mantamadiotis T., Stylli S.S., Tan S.T. Therapeutic Targeting of Cancer Stem Cells in Human Glioblastoma by Manipulating the Renin-Angiotensin System. Cells. 2019;8:1364. doi: 10.3390/cells8111364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Abiodun O.A., Ola M.S. Role of brain renin angiotensin system in neurodegeneration: An update. Saudi J. Biol. Sci. 2020;27:905–912. doi: 10.1016/j.sjbs.2020.01.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Di Lullo E., Kriegstein A.R. The use of brain organoids to investigate neural development and disease. Nat. Rev. Neurosci. 2017;18:573–584. doi: 10.1038/nrn.2017.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Li Y., Muffat J., Omer A., Bosch I., Lancaster M.A., Sur M., Gehrke L., Knoblich J.A., Jaenisch R. Induction of Expansion and Folding in Human Cerebral Organoids. Cell Stem Cell. 2017;20:385–396.e3. doi: 10.1016/j.stem.2016.11.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Pan Y., Lu P., Yin L., Chen K., He Y. Effect of fluoride on the proteomic profile of the hippocampus in rats. Z. Naturforsch. C. J. Biosci. 2015;70:151–157. doi: 10.1515/znc-2014-4158. [DOI] [PubMed] [Google Scholar]
- 170.Adilijiang A., Hirano M., Okuno Y., Aoki K., Ohka F., Maeda S., Tanahashi K., Motomura K., Shimizu H., Yamaguchi J., et al. Next Generation Sequencing-Based Transcriptome Predicts Bevacizumab Efficacy in Combination with Temozolomide in Glioblastoma. Molecules. 2019;24:3046. doi: 10.3390/molecules24173046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Velasco S., Kedaigle A.J., Simmons S.K., Nash A., Rocha M., Quadrato G., Paulsen B., Nguyen L., Adiconis X., Regev A., et al. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature. 2019;570:523–527. doi: 10.1038/s41586-019-1289-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Blue R., Miranda S.P., Gu B.J., Chen H.I. A Primer on Human Brain Organoids for the Neurosurgeon. Neurosurgery. 2020;87:620–629. doi: 10.1093/neuros/nyaa171. [DOI] [PubMed] [Google Scholar]
- 173.Cakir B., Xiang Y., Tanaka Y., Kural M.H., Parent M., Kang Y.J., Chapeton K., Patterson B., Yuan Y., He C.S., et al. Engineering of human brain organoids with a functional vascular-like system. Nat. Methods. 2019;16:1169–1175. doi: 10.1038/s41592-019-0586-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Ham O., Jin Y.B., Kim J., Lee M.O. Blood vessel formation in cerebral organoids formed from human embryonic stem cells. Biochem. Biophys. Res. Commun. 2020;521:84–90. doi: 10.1016/j.bbrc.2019.10.079. [DOI] [PubMed] [Google Scholar]
- 175.Pellegrini L., Bonfio C., Chadwick J., Begum F., Skehel M., Lancaster M.A. Human CNS barrier-forming organoids with cerebrospinal fluid production. Science. 2020;369:aaz5626. doi: 10.1126/science.aaz5626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Krieger T.G., Tirier S.M., Park J., Jechow K., Eisemann T., Peterziel H., Angel P., Eils R., Conrad C. Modeling glioblastoma invasion using human brain organoids and single-cell transcriptomics. Neuro. Oncol. 2020;22:1138–1149. doi: 10.1093/neuonc/noaa091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Pine A.R., Cirigliano S.M., Nicholson J.G., Hu Y., Linkous A., Miyaguchi K., Edwards L., Singhania R., Schwartz T.H., Ramakrishna R., et al. Tumor Microenvironment Is Critical for the Maintenance of Cellular States Found in Primary Glioblastomas. Cancer Discov. 2020;10:964–979. doi: 10.1158/2159-8290.CD-20-0057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Caruso F.P., Garofano L., D’Angelo F., Yu K., Tang F., Yuan J., Zhang J., Cerulo L., Pagnotta S.M., Bedognetti D., et al. A map of tumor-host interactions in glioma at single-cell resolution. Gigascience. 2020;9:giaa109. doi: 10.1093/gigascience/giaa109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Zhang C., Jin M., Zhao J., Chen J., Jin W. Organoid models of glioblastoma: Advances, applications and challenges. Am. J. Cancer Res. 2020;10:2242–2257. [PMC free article] [PubMed] [Google Scholar]