

Abstract
Antisense long non-coding RNAs (antisense lncRNAs), transcribed from the opposite strand of genes with either protein coding or non-coding function, were reported recently to play a crucial role in the process of tumor onset and development. Functionally, antisense lncRNAs either promote or suppress cancer cell proliferation, migration, invasion, and chemoradiosensitivity. Mechanistically, they exert their regulatory functions through epigenetic, transcriptional, post-transcriptional, and translational modulations. Simultaneously, because of nucleotide sequence complementarity, antisense lncRNAs have a special role on its corresponding sense gene. We highlight the functions and molecular mechanisms of antisense lncRNAs in cancer tumorigenesis and progression. We also discuss the potential of antisense lncRNAs to become cancer diagnostic biomarkers and targets for tumor treatment.
Keywords: Antisense lncRNA, Cancer, Transcriptional modulation, Translational control, Target therapy
Background
Protein-coding sequences account for less than 2% of the human genome, whereas most of the remaining regions of both DNA strands have the ability to be transcribed into RNAs. These RNAs cannot be translated into proteins, and are thus termed non-coding RNAs (ncRNAs) [1–3]. NcRNAs were previously considered as non-functional molecules [4]; however, recently, increasing evidence indicates that ncRNAs play an important role in regulating the expression of proteins and modulating various biological processes [5]. Based on their length, ncRNAs are classified into two types, and those RNA molecules that are more than 200 nucleotides are defined as long non-coding RNAs (lncRNAs). Accordingly, the other type is categorized as small non-coding RNAs [6, 7]. LncRNAs can be further classified into several groups, a large proportion of which are antisense lncRNAs, other groups include intergenic lncRNAs, intronic lncRNAs, and bidirectional (or divergent) lncRNAs [8–10]. Antisense lncRNAs are transcribed from the opposite strand of genes which have protein-coding or non-coding function (Fig. 1) [11]. They are defined according to the nearest protein-coding gene position, the same as ncRNAs, and they have no ability to be translated into proteins. Antisense lncRNAs are differentially expressed across different cell types, and regulate the expression of specific genes to modulate different signaling pathways [12]. Interestingly, they can exert their role through cis or trans regulation. Cis-acting antisense lncRNAs modulate the expression of the genes from which they originated by interacting with the promoter region with perfect sequence complementarity, while trans-acting antisense lncRNAs, through imperfect sequence complementarity, affect the expression of other genes [13]. In this review, we will elaborate the roles and molecular mechanisms of antisense lncRNAs in the process of tumorigenesis and tumor progression.
Fig. 1.

According to the original location in relative to its sense transcripts, antisense lncRNA can be further divided to three groups, tail to tail (two transcripts overlap in the 3ʹregion), internal (antisense lncRNA is completely covered by its sense transcripts) and head to head (two transcripts overlap in the 5ʹregion)
Antisense LncRNAs and cancer
Cancer is regarded as a genetic disease, in which mutations of protooncogenes or cancer suppressor genes result in uncontrolled cell growth [14]. Recently, antisense lncRNAs were reported to modulate the expression of genes that play an important role in tumorigenesis and cancer progression [15, 16]. Dysregulation of antisense lncRNAs has been observed in almost all types of cancer, acting as tumor promoters or suppressors. For example, overexpression of CDKN2B-AS1 in hepatocellular carcinoma (HCC) cells promotes tumor growth and metastasis [17]. In non-small cell lung cancer (NSCLC) tissues and cell lines, upregulated NNT-AS1 promotes the proliferation and invasion of cancer cells [18]. In addition, the level of VPS9D1-AS1 is negatively associated with tumor progression and poor prognosis in gastric cancer (GC) [19]. Reduced expression of B3GALT5-AS1 in colon cancer tissues results in cancer cell migration and invasion [20]. There are many ways to identify antisense lncRNAs. RNA sequencing (RNA-seq) analysis is commonly used method to investigate the transcriptome profile of lncRNAs. By quantifiably detecting lncRNAs, upregulated or downregulated lncRNAs can be identified. This method has contributed significantly to the study of antisense lncRNAs [21]. For example, using RNA-seq data from 60 samples collected from 20 patients with HCC, Yang et al. newly assembled 8,603 lncRNAs, 16% of which were antisense lncRNAs. The authors found that antisense lncRNA HAND2-AS1 was the only downregulated lncRNA in portal vein tumor thrombosis (PVTT), suggesting that HAND2-AS1 is associated with cancer metastasis [22]. Antisense lncRNAs do not encode protein, and in most cases, they function upstream of various signaling pathways. Mechanistic investigations revealed that antisense lncRNAs can affect biological process in both the nucleus and cytoplasm, such as epigenetic modulations and translational control. Furthermore, the aberrant expression of antisense lncRNAs is responsible for chemoradioresistance, a major obstacle to cancer therapy [23, 24]. Research on antisense lncRNAs has indicated their potential in therapeutic approaches; therefore, it necessary to summarize the roles and molecular mechanisms of antisense lncRNAs in the process of cancer development and progression. In this review, we present an overview of the main regulatory functions of antisense lncRNAs in different types of cancer types, as well as their potential clinical applications.
Mechanisms of antisense lncRNA activity
Nuclear antisense lncRNAs contribute to the regulation of a large number of genes by either changing the condition of DNA via histone modifications and DNA modifications or recruiting specific factors to the DNA at transcriptional level. Cytoplasmic antisense lncRNAs [25], which are more abundant than nuclear ones, function as regulators of mRNA stability and translation. They can also sponge microRNAs, acting as competing-endogenous RNAs (ceRNAs). Furthermore, cytoplasmic antisense lncRNAs can bind to proteins to alter their half-life.
Epigenetic regulations
Epigenetics is normally defined as heritable changes in gene expression without changes to the DNA sequence. Emerging research shows that antisense lncRNAs exert their role on gene expression through epigenetic modulations, such as DNA methylation and histone modifications. DNA methylation is an epigenetic process of regulating gene expression, and changes in DNA methylation patterns are very important for cancer development [26, 27]. Hypermethylation and hypomethylation of DNA both regulate the expression of oncogenes or tumor suppressors. There is a plethora of evidence linking antisense lncRNAs to the regulation of DNA methylation (Fig. 2a) [28]. For instance, Wu et al. demonstrated that antisense LncRNA DLX6-AS1 is upregulated in liver cancer stem cells (LCSCs) and HCC, in which it functions as a oncogene and promotes the proliferation of LCSCs. Mechanistic studies indicated that downregulation of DLX6-AS1 contributes to a reduction in CADM1 promoter methylation via suppression of DNA methyltransferase 1 (DNMT1), DNMT3a, and DNMT3b, thus elevating CADM1 expression in LCSCs and further inactivating the CADM1-dependent STAT3 signaling pathway [29]. Similarly, in prostate cancer (PCa), highly expressed MCM3AP-AS1 facilitates cancer cell progression by recruiting DNMT1/DNMT3 to the NPY1R promoter, which downregulates NPY1R expression and activates the MAPK pathway [30]. Furthermore, AFAP1-AS1 positively regulates the expression of the AFAP1 protein by negatively regulating CpG island methylation of the AFAP1 promoter in lung cancer [31]. Overexpression of ADAMTS9-AS2 results in the suppression of esophageal cancer development by inducing CDH3 promoter methylation [32].
Fig. 2.

In the nucleus, antisense lncRNAs exert their roles through DNA methylation, histone modification and transcriptional gene regulation. a MCM3AP-AS1 recruits DNMT1/DNMT3 to the promoter of NPY1R, resulting DNA methylation. b FOXD2-AS1 interacts with EZH2 and LSD1 and recruits them to the promoter region of EphB3, respectively guiding H3K27 methylation and H3K4 demethylation. c PAXIP1-AS1 recruits transcription factor ETS1 to the promoter of KIF14 and further affects its transcription
In addition to DNA methylation, chromatin structure and thus gene expression can be influenced by histone modifications [33]. Histone modifications are catalyzed by numerous histone-modifying enzymes, such as histone methyltransferases and histone acetyltransferases (Fig. 2b) [34]. In NSCLC, a high level of AFAP1-AS1 expression correlates with poor clinical outcomes. Mechanistically, AFAP1-AS1 interacts with EZH2, one type of histone methyltransferases, and recruits it to the promoter regions of P21, thus suppressing P21 expression at epigenetic level [35]. In PCa, ZEB1-AS1 interacts with the histone methyltransferase MLL1, a major methyltransferase responsible for the H3K4 modification. In this way, ZEB1-AS1 induces the H3K4me3 histone modification in the ZEB1 promoter region, which activates the expression of ZEB1 [36]. Besides recruiting methyltransferases, antisense lncRNAs can also recruit acetyltransferases. For example, in endometrial cancer, DLX6-AS1 achieves its stimulative function by increasing DLX6 expression via recruiting P300, a protein that can lead to histone acetylation in the DLX6 promoter region [37]. Likewise, AGAP2-AS1 promotes cell growth and inhibits apoptosis in breast cancer (BC) by inducing the histone acetylation in the MYD88 promoter region [38]. In addition, a few antisense lncRNAs have been associated with other histone modifications. In gastric cancer (GC), upregulation of FOXD2-AS1 promotes carcinogenesis by epigenetically silencing EPHB3 via recruiting EZH2 and LSD1, leading to H3K27 methylation and H3K4 demethylation, respectively [39].
Transcriptional modulation
At the transcriptional level, antisense lncRNAs regulate gene expression by recruiting transcription factors to the promoter of a specific gene [40]. Transcription factors play an important role in the process of transcription; they can bind with polymerase II and form a complex to further promote or repress gene expression (Fig. 2c) [40]. For instance, Xu et al. demonstrated that antisense lncRNA PAXIP1-AS1 is highly expressed in glioma and correlated with poor prognosis. Functionally, upregulation of PAXIP1-AS1 promotes migration, invasion, and angiogenesis of cancer cells. Mechanistic investigations indicated that PAXIP1-AS1 recruits the transcription factor ETS1 to the promoter region of KIF14 and further upregulates its expression [41]. Similarly, TMPO-AS1 is upregulated and exerts its oncogenic roles in ovarian cancer (OC). Mechanistically, TMPO-AS1 interacts with E2F6, a transcription factor that binds to the promoter region of LCN2, thus promoting LCN2 transcription [42]. Another antisense lncRNA that exerts its function at the transcriptional level is HOXB-AS1. In glioblastoma (GBM), increased expression of HOXB-AS1 promotes proliferation and induces apoptosis by recruiting transcription factor ILF3 to the promoter regions of HOBX2 and HOBX3 [43].
Antisense LncRNAs acting as ceRNAs
At the post-transcriptional level, numerous antisense lncRNAs serve as regulators of cancers by acting as ceRNAs. ceRNAs are the targets of microRNAs (miRNAs), and interact with miRNAs to further modulate the expression of the specific mRNA targeted by the miRNA (Fig. 3a) [44]. Through this miRNA-mediated method, antisense lncRNAs can affect cancer development.
Fig. 3.

In the cytoplasm, antisense lncRNAs regulate cancer progression through post-trancriptional modulations and translational reprogramming. a the more miR-3184-5p molecules bind to FOXP4-AS1, the less miR-3184-5p molecules interact with FOXP4 mRNA, by this way, FOXP4-AS1 promotes tthe expression of FOXP4. b ZEB1-AS1 recruits AUF1 to ZEB1 mRNA and activates its translation without changing mRNAs level. c PDCD4-AS1 affects PDCD4 mRNA stability by regulating RNA-binding protein HuR binding to mRNA. d SLCO4A1-AS1 interacts with β-catenin and inhibites its phosphorylation
In PCa cells, antisense lncRNA FOXP4-AS1 and its corresponding coding transcript FOXP4 are highly expressed. Upregulation of FOXP4-AS1 correlates with poor prognosis and promotes cancer cell proliferation. Interestingly, FOXP4-AS1 has the binding site for the FOXP4-targeting miRNA, miR-3184-5p. FOXP4-AS1 competes with FOXP4 for binding with miR-3184-5p. As a result, it positively regulates the FOXP4 protein level [45]. ZEB1-AS1 tumor-promoter functions have been confirmed in many types of cancer, and there are two studies indicating that ZEB1-AS1 promotes proliferation and migration of colorectal cancer (CRC) cells by acting as a ceRNA; however, the two targeted-miRNAs are different. Lv et al. found that there is an inverse correlation between ZEB1-AS1 and miR-181a-5p levels in CRC cells. Further research demonstrated that ZEB1-AS1 may function as a molecular sponge for miR-181a-5p [46]. By contrast, miR-101 is reported to function as tumor suppressor by targeting ZEB1 in many types of cancer; therefore, Xiong et al. focused on the mechanism of the miR-101/ZEB1 axis in CRC. Consistently, ZEB1-AS1 knockdown, miR-101 overexpression, and ZEB1 depletion suppressed the proliferation and migration of CRC cells. ZEB1-AS1 functions as a ceRNA for miR-101 and abrogated the silencing of ZEB1 caused by miR-101 [47]. ZEB1-AS1 participates in tumorigenesis and progression in various cancer types, and it is likely that more miRNA targets will be found in the future. Moreover, miR-1253 is the target of FOXC2-AS1 in PCa [48] and TPT1-AS1 acts as a sponge for miR-324-5p in cervical cancer (CC) [49]. It is becoming clear that many antisense LncRNAs exert their roles in cancer via this ceRNA mechanism, thus future works in this area might lead to the development of promising therapeutics.
Translational control by antisense lncRNAs
Regulation of gene expression is not limited to epigenetic and transcriptional regulatory networks, antisense LncRNAs can also regulate gene expression at the translational level. Protein synthesis is controlled by numerous tumor suppressors and oncogenes, making it easy to respond to environmental changes by regulating this process. Antisense lncRNAs are involved in regulating protein synthesis and degradation. Firstly, they can recruit the target of mRNAs so as to affect their translation (Fig. 3b). For instance, in bladder cancer (BCa) cells, Zhao et al. demonstrated that ZEB1-AS1 expression is higher in comparison with that in corresponding normal tissues, which promotes BCa cells migration and invasion. Mechanistically, they found that ZEB1-AS1 upregulates the expression of ZEB1 without increasing its mRNA level. Unexpectedly, it activates the translation of ZEB1 mRNA by recruiting AUF1, which is able to bind to (A + U)-rich elements within 3ʹ-untranslated region (3ʹ-UTR) of target mRNA and promote translation without affecting the mRNA level [50].
Secondly, antisense lncRNAs affect the stability of mRNA by regulating the association of RNA-binding proteins with mRNA (Fig. 3c). PDCD4 is a tumor suppressor in BC, and the expression level of PDCD4 correlates positively with the level of antisense lncRNA PDCD4-AS1. Mechanistically, overexpression of PDCD4-AS1 increases the level of PDCD4 mRNA. To rule out the possibility that PDCD4-AS1 regulates PDCD4 expression at the epigenetic or transcriptional level, researchers quantified the levels of PDCD4 pre-mRNA, which showed that there was no significant change in the level of PDCD4 pre-mRNA in PDCD4-AS1 deleted cells compared with that in the control group. This indicated that PDCD4-AS1 increases the level of PDCD4 mRNA by improving its stability. Additional investigations demonstrated that PDCD4-AS1 promotes PDCD4 mRNA stability by negatively modulating HuR [51].
Likewise, in BC, CERS6-AS1 functions as a cancer promoter by binding to IGF2BP3, which increases the stability of CERS6 mRNA [52]. HOXB-AS1 facilitates cell growth in multiple myeloma by binding to ELAVL1, thus promoting FUT4 mRNA stability [53].
Finally, antisense lncRNAs can affect the level of certain proteins by modulating the process of protein degradation by prolonging or shortening protein half-life (Fig. 3d). For example, SLCO4A1-AS1 was confirmed as a tumor-promoter antisense lncRNA in CRC, in which the level of SLCO4A1-AS1 correlated positively with the level of β-catenin. Further investigations indicated that SLCO4A1-AS1 can interact with β-catenin and increase its stability by inhibiting its phosphorylation [54]. ZFPM2-AS1 expression is higher in GC cells than in normal gastric tissue. By binding to and stabilizing macrophage migration inhibitory factor (MIF), the suppressor of p53 stability, increased levels of ZFPM2-AS1 promote proliferation and suppresses apoptosis of cancer cells [55]. Likewise, FEZF1-AS1 promotes CRC cell proliferation and metastasis through activation of the STAT3 signaling pathway by increasing the stability of the pyruvate kinase 2 (PKM2) [56].
The difference between lncRNAs and antisense lncRNAs
Nucleotide sequence complementarity allows antisense lncRNA to have special effects on their sense gene, thus they are more likely to regulate the expression of their corresponding protein-coding gene, which contrasts with other types of lncRNAs. A good way to find out how antisense lncRNAs affect the growth of cancer cells is to detect the expression of its sense gene.
In prostate cancer, a correlation between the level ZEB1-AS1 and ZEB1 was demonstrated. ZEB1-AS1 recruits histone methyltransferase MLL1 to the promoter region of ZEB1, thus inducing the H3K4me3 modification, and activating ZEB1 transcription [36]. Similarly, ZNF667-AS1 and its sense gene, ZNF667, are downregulated in esophageal squamous cell carcinoma. ZNF667-AS1 affects the expression of ZNF667 via promoter CpG site methylation by recruiting TET1, which can hydrolyze 5′-methylcytosine (5′-mc) to 5′-hydroxymethylcytosine (5′-hmc) [57]. Many histone-modifying enzymes cannot exert their role independently because they lack specific DNA-binding domains, thus a large portion of antisense lncRNAs bind chromatin-modifying enzymes and recruit them to their sense gene [15]. In addition, at the translational level, antisense lncRNAs can directly bind with their sense mRNA and form an RNA duplex, which affects the stability of these targeted mRNAs. For instance, in skin cutaneous carcinoma, TTN-AS1 directly regulates TTN expression by forming a RNA duplex with TTN mRNA [58]. In addition, the overlapping region of UPK1A-AS1 increases the stability of UPK1A mRNA by forming a duplex in lung cancer cells [59].
Antisense lncRNAs in tumorigenesis and progression
Antisense lncRNAs have a crucial effect in the process of tumor development and progression in various cancer types, either acting as oncogenes or tumor suppressors. Interestingly, the function of some antisense lncRNAs depends on the type of cancer, functioning as oncogenic factor in some cancers, while acting a tumor suppressor in other cancer types [60]. In this section, we provide relevant examples of well-established antisense lncRNAs having oncogenic, tumor suppressive, or dual properties (Fig. 4) [61].
Fig. 4.
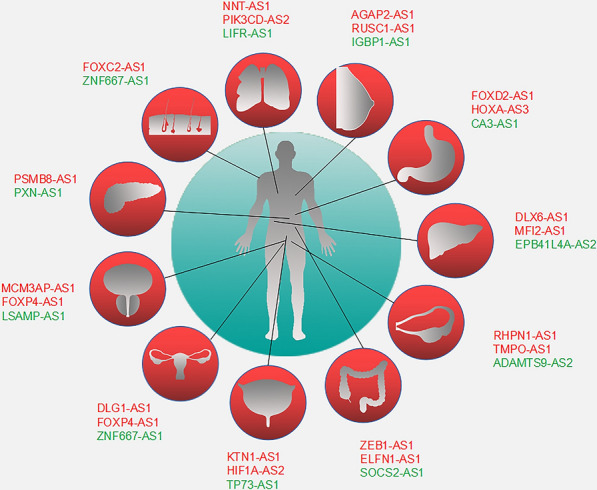
Antisense lncRNAs involve in many types of cancer. Antisense lncRNAs function as oncogene (red) and tumor-suppressor (green) in different cancer types. Clockwise from top left: lung cancer, breast cancer, gastric cancer, liver cancer, ovarian cancer, colorectal cancer, bladder cancer, cervical cancer, prostate cancer, pancreatic cancer, melanoma
Antisense lncRNAs function as oncogenes
In this part, we discuss how antisense lncRNAs promote cancer cell proliferation and migration. Among the numerous oncogenic antisense lncRNAs, we focus on KTN1-AS1 and FOXP4-AS1, whose oncogenic functions have been confirmed in different cancer types. KTN1-AS1 is reported to be highly expressed in six types of cancer. In NSCLC, STAT1-induced upregulation of KTN1-AS1 facilitates cancer cell progression via the miR-23b/DEPDC1 axis [62]. In BCa, KTN1-AS1 knockdown inhibited the proliferation and invasion of cancer cells. Mechanistically, KTN1-AS1 recruits EP300, a histone acetyltransferase, which enriched H3K27Ac in the KTN1 promoter region, thus activating the expression of KTN1 [63]. Furthermore, KTN1-AS1 increases the viability and invasive ability of glioma cells in vitro and in vivo through the KTN1-AS1/miR-505-3p pathway and promotes tumor growth of HCC via the miR-23c/ERBB2IP axis [64, 65].
Antisense lncRNAs forkhead box P4 antisense RNA 1, known as FOXP4-AS1, is significantly overexpressed in approximately 10 types of human cancers. For example, in mantle cell lymphoma, FOXP4-AS1 accelerates the progression of cancer by sponging miR-136-5p to further regulate the downstream target of miR-136-5p, NACC1 [66]. Interestingly, FOXP4-AS1 promotes CC progression by binding with miR-136-5p, the same target microRNA as that in mantle cell lymphoma, indicating the generality of cancer development between different cancer types. Subsequent studies demonstrated that the target of miR-136-5p is not NACC1 but CBX4 in CC [67]. Likewise, in nasopharyngeal carcinoma, FOXP4-AS1 promotes cancer cell proliferation and inhibits apoptosis via the miR-423-5p/STMN1 axis [68]. Moreover, through the miR-3184-5p/FOXP4 axis, FOXP4-AS1 promotes the proliferation of esophageal squamous cell carcinoma cells [69].
Antisense lncRNAs act as tumor suppressors
The expression of some antisense lncRNAs is downregulated in cancer cells, because they inhibit cancer cell proliferation, migration, and invasion. Here, we discuss three antisense lncRNAs, HAND2-AS1, FGF13-AS1, and FGF14-AS2, which exert tumor suppressive roles during the onset and progression of cancer.
Antisense lncRNA HAND2-AS1, transcribed from the opposite strand of HAND2 (encoding heart and neural crest derivatives expressed 2) on chromosome 4q33-34, was first reported to be downregulated in endometrioid endometrial carcinoma (EEC). Its anti-tumorigenic effect is mediated by downregulating NMU, an oncogenic protein in EEC [70]. In GC cells, HAND2-AS1 expression is decreased; however, overexpression of HAND2-AS1 is capable of inhibiting GC cell proliferation and promoting their apoptosis by functioning as a ceRNA that binds with miR-590-3p [71]. Another study demonstrated that HAND2-AS1 can also exert its tumor suppressive role through the miR-769-5p/TCEAL7 axis in GC [72]. In HCC, HAND2-AS1 overexpression reduces the viability and proliferation of cancer cells by sponging miR-300 [73]. Furthermore, in NSCLC cells, HAND2-AS1 represses the proliferation of cancer cells by targeting the PI3K/Akt pathway [74].
The role of FGF13-AS1 in tumors has only been reported in BC, in which it suppresses BC cell proliferation, migration, and invasion by impairing glycolysis and stemness properties. Mechanistically, FGF13-AS1 shortens the half-life of MYC mRNA by interacting with the RNA-binding protein IGF2BPs and further interrupting the interaction between IGF2BPs and MYC mRNA, resulting in the suppressed expression of c-Myc. Simultaneously, downregulated c-Myc transcriptionally inhibits FGF13-AS1, forming a feedback loop [75].
FGF14-AS2 was first identified as a tumor suppressor in BC. Compared with that in adjacent normal tissue, FGF14-AS2 is significantly downregulated in BC tissues [76]. As reported by Jin and coworkers, FGF14-AS2 activates the expression of FGF14 at the post-transcriptional level by functioning as a ceRNA of miR-370-3p in BC [77]. Moreover, it sponges miR-1288-3p, which indirectly controls Ras/ERK signaling, causing inhibition of CRC proliferation [78].
Antisense LncRNAs with dual activity
A few antisense lncRNAs have been reported to play opposite roles in different types of cancer. These inconsistent functions could be partly explained by the wide genetic and phenotypic heterogeneity of tumors, and the different experimental methods and samples used. Herein, three confirmed examples of antisense lncRNAs with divergent roles in tumors are discussed.
A tumor-stimulative role of TP73-AS1 has been reported in various types of solid tumor, including lung, breast, gastric, and hepatic carcinomas. Mechanistically, it exerts its activity on tumor proliferation mostly by functioning as a ceRNA. In lung cancer, TP73-AS1 knockdown inhibited the growth and metastasis of cancer cells through the miR-27b-3p/LAPTM4B axis [79]. In BC, TP73-AS1 sponges miR-200a, indirectly activating the expression of ZEB1 and promoting cell proliferation [80]. Similarly, in HCC, overexpression of TP73-AS1 competes with HMGB1 for miR-200a binding, causing the upregulation of HMGB1, a critical regulator of cell death and survival [81]. However, TP73-AS1 was confirmed to be downregulated in acute myeloid leukemia (AML), which affects the cell proliferation of AML through the miR-21/PTEN axis [82]. A tumor-suppressive role of TP73-AS1 has been also reported in BCa, patients with low TP73-AS1 expression have shorter disease-free survival than patients with high TP73-AS1 expression. Further investigations indicated that TP73-AS1 functions as a tumor suppressor via its role in epithelial-mesenchymal transition (EMT) [83].
ADAMTS9-AS2 has been reported to have either an oncogenic or tumor suppressive function. In GC, ADAMTS9-AS2 acts as a tumor suppressor via its ability to activate NLRP3-mediated pyroptotic cell death through sponging miR-223-3p [84]. In OC, its downregulation correlated with lymph-node metastasis and poor overall survival. ADAMTS9-AS2 inhibits OC progression by regulating the miR-182-5p/FOXF2 axis [85]. However, high ADAMTS9-AS2 expression was observed in tongue squamous cell carcinoma (TSCC), in which it shows an explicitly oncogenic role in tumorigenesis by competing wit̄h miR-600 [86].
Except in myeloid malignancy, LEF1-AS1 has been identified as an oncogene in all cancer types reported to date. In NSCLC, LEF1-AS1 promotes cancer cell proliferation and inhibits their apoptosis by regulating the miR-221/PTEN pathway [87]. Similarly, it functions as a oncogenic factor through the miR-30-5p/SOX9 axis in colon cancer and boosts the proliferation, migration, and invasion of osteosarcoma by increasing the mRNA stability of LEF1 [88, 89]. Nevertheless, downregulation of LEF1-AS1 correlates positively with tumor progression in patients with myelodysplastic syndrome and acute myeloid malignancy, indicating a tumor suppressive role in myeloid malignancy [90].
As summarized in Table 1, we have distilled the conclusions from many studies and present the mechanisms by which antisense lncRNAs affect tumor development and progression.
Table 1.
Antisense LncRNAs act as oncogenes or tumor suppressors in various cancer types
| Antisense LncRNA | Cancer type | Function | Mechanism | Refs |
|---|---|---|---|---|
| MFI2-AS1 | Liver cancer | Oncogene | MFI2-AS1 functions as miR-134 sponge to Upregulate FOXM1 expression | [122] |
| EPB41L4A-AS2 | Tumor suppressor | EPB41L4A-AS2 sponges miR-301a-5p and targets FOXL1 | [123] | |
| PIK3CD-AS2 | Lung cancer | Oncogene | PIK3CD-AS2 suppresses p53 pathway via YBX1 | [124] |
| LIFR-AS1 | Tumor suppressor | LIFR-AS1 regulates miR-942-5p/ZNF471 axis | [125] | |
| ZNFX1-AS1 | Bladder cancer | Oncogene | ZNFX1-AS1 interacts with miR-193a-3p/Syndecan 1 | [126] |
| MAGI2-AS3 | Tumor suppressor | MAGI2-AS3 upregulates TNS1 by sponging miR-31-5p | [127] | |
| RUSC1-AS1 | Breast cancer | Oncogene | RUSC1-AS1 downregulates the expression of CDKN1A and KLF2 | [128] |
| IGBP1-AS1 | Tumor suppressor | IGBP1-AS1 modulates miR-24-1/ZIC3 axis | [129] | |
| ELFN1-AS1 | Colorectal cancer | Oncogene | ELFN1-AS1 acts as a sponge of miR-4644 to increase TRIM44 expression | [130] |
| SOCS2-AS1 | Tumor suppressor | SOCS2-AS1 stabilizes SOCS2 and sponges miR-1264 | [131] | |
| CTBP1-AS2 | Cervical cancer | Oncogene | CTBP1-AS2 upregulates ZNF217 through sponging miR-3163 | [132] |
| ZNF667-AS1 | Tumor suppressor | ZNF667-AS1 counteracts microRNA-93-3p-dependent PEG3 downregulation | [133] | |
| HOXA-AS3 | Gastric cancer | Oncogene | HOXA-AS3 activates NF-κB signaling through miR-29a-3p/LTβR axis | [134] |
| CA3-AS1 | Tumor suppressor | CA3-AS1 sponges miR-93-5p and targets BTG3 | [135] | |
| VPS9D1-AS1 | Prostate cancer | Oncogene | VPS9D1-AS1 sponges miR-4739 to upregulate MEF2D | [136] |
| LSAMP-AS1 | Tumor suppressor | LSAMP-AS1 binds to microRNA-183-5p and upregulates the tumor suppressor DCN | [137] | |
| RHPN1-AS1 | Ovarian cancer | Oncogene | RHPN1-AS1 acts as a ceRNA against miR-596 and upregulating LETM1 | [114] |
| ZNF667-AS1 | Melanoma | Tumor suppressor | ZNF667-AS1 positively regulates MEGF10 | [138] |
| FOXC2-AS1 | Oncogene | FOXC2-AS1 downregulates p15 by recruiting EZH2 | [139] | |
| PSMB8-AS1 | Pancreatic cancer | Oncogene | PSMB8-AS1 modulates miR-382-3p/STAT1/PD-L1 axis | [140] |
| PXN-AS1 | Tumor suppressor | PXN-AS1 acts as a ceRNA of miR-3064 to upregulate PIP4K2B expression | [141] |
Antisense lncRNAs in chemoradioresistance
In addition to surgery, chemotherapy and radiation therapy are the two effective methods to improve the survival rate and prognosis of people with cancer. However, chemoradioresistance represents a major barrier to tumor therapy; therefore, it is necessary to determine the mechanism underlying a tumor chemoradioresistance.
Recently, increasing evidence indicates that the drug-resistant tumor phenotype is regulated by the expression of certain genes [91], and antisense lncRNAs are also reported to be involved in this process. Below, we discuss two antisense lncRNAs whose drug-resistance activities have been determined in some types of cancer, more examples are presented in Table 2.
Table 2.
Antisense LncRNAs are related to drug resistance in cancer
| Antisense LncRNA | Cancer type | Drug | Mechanism | Refs |
|---|---|---|---|---|
| HOXD-AS1 | Cervical cancer | Cisplatin | HOXD-AS1 enhances chemoresistance of cisplatin-resistant cancer cells by modulating miR-130a-3p/ZEB1 axis | [142] |
| DLX6-AS1 | Breast cancer | DLX6-AS1 promotes cisplatin resistance through miR-199b-5p/PXN signaling | [143] | |
| NCK1-AS1 | Osteosarcoma | NCK1-AS1 knockdown enhances Cisplatin sensitivity of cancer cells by regulating miR-137 | [144] | |
| SLC7A11-AS1 | Pancreatic cancer | Gemcitabine | SLC7A11-AS1 promotes Gemcitabine-resistance by Blocking SCF β-TRCP-Mediated Degradation of NRF2 | [145] |
| SBF2-AS1 | Pancreatic cancer | SBF2-AS1 promotes the expression of TWF1 by binding with miR-142-3p to induce gemcitabine resistance | [146] | |
| LOXL1-AS1 | Prostate cancer | Doxorubicin | LOXL1-AS1/miR-let-7a-5p/EGFR-related pathway regulates the doxorubicin resistance | [147] |
| FOXC2-AS1 | Osteosarcoma | FOXC2-AS1 promotes doxorubicin resistance by increasing the expression of FOXC2 | [148] | |
| AFAP1-AS1 | Breast cancer | Trastuzumab | AFAP1-AS1 promotes trastuzumab resistance by binding with AUF1 and activating ERBB2 expression | [149] |
| SBF2-AS1 | Glioblastoma | Temozolomide | SBF2-AS1 enhances chemoresistance to temozolomide by functioning as a ceRNA for miR-151a-3p | [150] |
| ADAMTS9-AS2 | Glioblastoma | ADAMTS9-AS2 promotes Temozolomide Resistance via Upregulating the FUS/MDM2 Ubiquitination Axis | [151] | |
| NR2F1-AS1 | Liver cancer | Oxaliplatin | NR2F1-AS1 regulates oxaliplatin resistance by targeting ABCC1 via miR-363 | [152] |
| DSCAM-AS1 | Breast cancer | Tamoxifen | DSCAM-AS1 enhances Tamoxifen resistance by functioning as a sponge of miR-137 | [153] |
| ADAMTS9-AS2 | Breast cancer | ADAMTS9-AS2 enhances tamoxifen resistance by activating miR-130a-5p | [154] | |
| AFAP1-AS1 | Prostate cancer | Paclitaxel | AFAP1-AS1 modulates the sensitivity of paclitaxel via miR-195-5p/FKBP1A axis | [155] |
| DDX11-AS1 | Esophageal cancer | DDX11-AS1 promotes resistance cancer cells to Paclitaxel by inhibiting TOP2A expression via TAF1 | [156] |
In esophageal squamous cell carcinoma, Liu et al. demonstrated that FXOD2-AS1 overexpression promotes cisplatin resistance through the miR-195/Akt/mTOR axis [92]. In glioma, FOXD2-AS1 functions as a prognostic factor and induces temozolomide resistance in a O(6)-methylguanine-DNA methyltransferase-dependent manner [93]. Meanwhile, FOXD2-AS1 might also contribute to temozolomide resistance in glioma via the miR-98-5p/CPEB4 axis [94]. By promoting STAT3 transcriptional activity, FOXD2-AS1 enhances chemotherapy resistance of laryngeal squamous cell carcinoma [95]. Furthermore, FOXD2-AS1 binds with miR-143, leading to gemcitabine-resistance in BCa [96].
OIP5-AS1 is more likely to function as a ceRNA when playing its role in drug resistance. In osteosarcoma, OIP5-AS1 mediates resistance to doxorubicin by regulating the miR-137-3p/PTN axis [97]. In addition, OIP5-AS1 either modulates the miR-377-3p/FOSL2 signaling pathway or induces the LPAATβ/PI3K/AKT/mTOR signaling pathway by sponging miR-340-5p, thus regulating cisplatin sensitivity [98, 99]. Similarly, in colon cancer, OIP5-AS1 regulates drug-resistance to oxaliplatin by sponging miR-137 [100].
Increasing numbers of studies have focused on the mechanisms by which antisense lncRNAs affect drug sensitivity to cancer, covering various types of chemotherapeutic drugs in different cancers; therefore, providing a new direction to solve this problem.
Similarly, radiation therapy is a very common treatment for many types of cancer, either alone or in combination with other therapeutic methods. The effect greatly depends on the radiosensitivity of the cancer cells. Patients require a higher dose of irradiation when the tumor is resistant to radiation therapy, resulting in more damage to normal tissues. Some studies reported that dysregulation of antisense lncRNAs might be involved in this process. The expression level of certain antisense lncRNAs is different between radioresistant and radiosensitive tumors, indicating that modulation of their expression could improve the radiosensitivity of tumors. In OC, the marked upregulation of FAM83H-AS1 contributes to radioresistance by increasing the stability of HuR, an RNA binding protein that had been reported to regulate radioresistance in multiple cancers [101]. In NSCLC, upregulated SBF2-AS1 reduces the radiosensitivity and apoptosis of cancer cells via regulating the miR-302a/MBNL3 axis [102]. PTPRG-AS1 promotes radioresistance in two cancer types: in nasopharyngeal carcinoma (NPC), PTPRG-AS1 reduces sensitivity to radiotherapy through the miR-194-3p/PRC1 regulatory axis [103]; whereas, under X-ray irradiation, overexpression of PTPRG-AS1 could promote the viability and enhance the radioresistance of NSCLS by modulating the miR-200c-3p/TCF4 axis [104]. In a similar role, TTN-AS1 sponges miR-134-5p to regulate the radiosensitivity of human large intestine cancer cells [105]. It is evident that the modulation of antisense lncRNA expression can be used to improve the radiosensitivity of tumors, providing a new method to solve the problem of radioresistance in cancer.
Antisense LncRNA databases
Online databases are good tools to understand dysregulated lncRNA, simultaneously, these databases can also be used to understand antisense LncRNA. Among the many databases containing information related to lncRNAs, we would like to introduce three particularly useful databases.
NONCODE
NONCODE (http://www.noncode.org/) is an integrated knowledge database dedicated to collecting information regarding noncoding RNA. Recently, it was updated to v6.0. Almost all types of ncRNA (excluding tRNAs and rRNAs) are covered, not only providing basic information, such as the location, sequence, and source, but also advanced information, such as the expression profile and conservation information. In the current version, there are 39 species (16 animals and 23 plants), representing an increase of 22 compared with v5.0. NONCODE has collected a total of 173,112 human lncRNAs, and v6.0 contains updated human lncRNA-cancer relationships, which will help us to explore the roles of lncRNAs in cancer [106].
LncRNADisease
LncRNADisease v2.0 (http://www.rnanut.net/lncrnadisease/) focuses on the relationship between diseases and lncRNAs, collecting experimentally supported lncRNA-disease associations. In comparison with the previous version, LncRNADisease v2.0 has an over 40-fold increase in lncRNA-disease associations. There is a confidence score system to evaluate the reliability of the relationship between a disease and an lncRNA. A score close to 1 represents a strong association. Besides, to further explore the network of lncRNAs with mRNAs and miRNAs, LncRNADisease v2.0 covers 12,207 lncRNA–mRNA and 2368 miRNA–lncRNA regulatory relationships, and an lncRNA–miRNA–mRNA network has also been constructed [107].
LNCipedia
LNCipedia 5 (https://lncipedia.org) contains a total of 56,946 lncRNA genes and 127,802 lncRNA transcripts. Compared with other databases, LNCipedia has an advantage: in the current version, 6% of the genes and 23% of the transcripts are annotated with an official gene symbol, making it more convenient to study lncRNAs. Moreover, in the advanced search, we can choose the class as antisense, making it easier to find dysregulated antisense lncRNAs [108].
Potential applications
Antisense lncRNAs are highly tissue-specific drivers of cancer phenotypes and are identified as crucial regulators associated with tumorigenesis and suppression, showing great potential, not only as biomarkers, but also as therapeutic targets for cancer treatment. Antisense lncRNAs have been found to be involved in all steps of cancer development and progression. First, antisense lncRNAs regulate the proliferation, migration, invasion, and apoptosis of cancer cells, which means they can function as diagnostic biomarkers. Second, the expression levels of some antisense lncRNAs are associated with tumor size and TNM stage; therefore, they could be used to evaluate tumorigenesis and cancer progression. Furthermore, the levels of some antisense lncRNAs correlate with certain prognostic markers, indicating their ability to predict cancer prognosis. For example, in HCC, SOX21-AS1 is a highly expressed antisense LncRNA that acts as an oncogene in cancer cell proliferation and cell cycle progress. Further investigations indicated that the expression level of SOX21-AS1 correlated with tumor size, Edmondson Grade, vascular invasion, and cirrhosis. Kaplan–Meier analysis showed that patients with HCC with high levels of SOX21-AS1 expression had a shorter survival time compared with those with low expression of SOX21-AS1. These results demonstrated that SOX21-AS1 is a potential biomarker for HCC [109]. Likewise, downregulated ZNF385D-AS2 is predictive of poor prognosis of patients with liver cancer [110]. In addition, TMPO-AS1 and FOXC2-AS1 are implicated as biomarkers for PCa [48, 111].
Antisense lncRNAs are differentially expressed in different cancer types and their expression levels are related to tumorigenesis and aggressiveness, making them potential targets for cancer treatment. Targeting antisense lncRNAs and modulating their expression could affect many biological processes of cancer cells. In NSCLC, NNT-AS1 expression is upregulated in cancer cell lines; therefore, to explore the roles of NNT-AS1 in NSCLC, cancer cells were transfected with a small interfering RNA si-(NNT-AS1) and a negative control, si-NC. The results indicated that the migration ability of cancer cells in the si-NNT-AS1 group was suppressed compared with that in the si-NC group. In addition, the invasion ability of cancer cells transfected with si-NNT-AS1 was suppressed compared with that in the si-NC group [112]. Similarly, in BC, HIF1A-AS2 is upregulated, and researchers transfected a short hairpin RNA (shRNA), sh-HIF1A-AS2, into cancer cells to reduce the level of HIF1A-AS2. The results showed that the proliferation capacity of the cancer cells transfected with sh-HIF1A-AS2 was significantly reduced, as were the levels of proliferation marker proteins. Through different ways of targeting antisense lncRNAs and reducing their expression, the growth of cancer cells was suppressed [113]. Meanwhile, this effect also exists in vivo, which further confirms the therapeutic value of targeting antisense lncRNAs. In epithelial ovarian cancer, highly-expressed RHPN1-AS1 was suppressed using an shRNA. Cells were injected into mice and grown for 6 weeks. The results showed that knockdown of RHPN1-AS1 significantly reduced the growth of epithelial ovarian cancer in the xenograft tumor model [114]. In another study, researchers treated cancer cells with lentiviral CRISPR/Cas9 to stably knockout DSCAM-AS1, which inhibited the growth of MCF7 xenograft tumors when compared with the negative control group [115]. In addition, as mentioned above, antisense lncRNAs also have great potential to solve the problems of cancer cell resistance to chemotherapy and radiotherapy.
Challenges to the application of antisense lncRNAs
There are thousands of articles reporting on the relationships between antisense lncRNAs and cancer, providing researchers with a lot of data. These data are the basis for future study; however, similar research sometimes produces conflicting conclusions. For instance, in ovarian cancer, Miao et al. demonstrated that TTN-AS1 expression is decreased in cancer tissues and cells. TTN-AS1 inhibits the cell growth of OC through the miR-15b-5p/FBXW7 axis, as demonstrated in several OC cell lines [116]. However, Liu et al. indicated that a high level of TTN-AS1 is found in OC tissues and cell lines, in which TTN-AS1 promotes the progression of OC by modulating the miR-139-5p/ROCK2 axis, and their samples were mainly obtained from patients with OC [117]. Similarly, three studies on the relationship between LIFR-AS1 and GC reported contradictory results. Their cancer tissues were collected from patients with GC from different areas [118–120]. Ignoring the experimental errors, the different experimental samples might have resulted in the presence of different cancer subtypes, which might have partly contributed to the generation of conflicting results. Therefore, it might be necessary to investigate how different cancer subtypes affect the role of antisense lncRNAs on cancer. By contrast, although we have revealed the mechanisms by which antisense lncRNAs affect the process of cancer development, the present method to detect the expression level of antisense lncRNA is not very useful; therefore, it might be better to identify antisense lncRNA candidates whose expression is easy to monitor. There is still a long way to go to apply these results to clinical practice. Lastly, some antisense lncRNAs have been studied; however, the functions of the majority of these transcripts remain to be determined [121]. Further investigations of antisense lncRNAs will provide more possibilities for cancer diagnostics and therapy.
Conclusion
Growing evidence demonstrates that many antisense lncRNAs are dysregulated in cancer cells. Antisense lncRNAs play a crucial role in tumor onset, progression, chemotherapy responses, and radiotherapy sensitivity by regulating gene and protein expression at epigenetic, transcriptional, post-transcriptional, and translational levels. The close relationship between antisense lncRNAs and cancers mean that antisense lncRNAs have great potential as biomarkers to diagnose cancer, predict prognosis, and as targets for tumor treatment. However, we cannot ignore the difficulty of applying antisense lncRNA-based therapeutic approaches in the clinic. Additional research will provide more hope of finding a cure for cancer.
Acknowledgements
Not applicable.
Abbreviations
- antisense lncRNA
Antisense long non-coding RNA
- ncRNA
Non-coding RNA
- lncRNA
long non-coding RNA
- CDKN2B-AS1
Cyclin-dependent kinase inhibitor 2B antisense RNA 1
- HCC
Hepatocellular carcinoma
- NSCLC
Non-small cell lung cancer
- NNT-AS1
Nicotinamide nucleotide transhydrogenase-antisense 1
- TMPO-AS1
Thymopoietin antisense transcript 1
- DLG1-AS1
DLG1 antisense RNA 1
- PC
Prostate cancer
- CC
Cervical cancer
- DLX6-AS1
Distal-less homeobox 6 antisense RNA 1
- LCSC
Liver cancer stem cell
- CADM1
Cell adhesion molecule 1
- DNMT
Methyltransferase
- STAT3
Signal transducer and activator of transcription 3
- AFAP1-AS1
Actin filament associated protein 1 antisense RNA 1
- ADAMTS9-AS2
ADAM metallopeptidase with thrombospondin type 1 motif, 9 antisense RNA 2
- CDH3
Cadherin 3
- EZH2
Enhancer of Zeste Homolog 2
- ZEB1-AS1
Zinc finger E-box binding homeobox1-antisense RNA 1
- MLL1
Mixed lineage leukaemia protein-1
- H3K4
Histone 3 lysine 4
- AGAP2-AS1
Arf GAP with GTP-binding protein-like domain, Ankyrin repeat and PH domain 2 antisense RNA 1
- BC
Breast cancer
- MyD88
Myeloid differentiation primary response protein 88
- GC
Gastric cancer
- FOXD2-AS1
FOXD2 adjacent opposite strand RNA 1
- EphB3
Ephrin type-B receptor 3
- LSD1
Lysine-specific demethylase 1
- PAXIP1-AS1
PAX-interacting protein 1-antisense RNA1
- ETS1
ETS proto-oncogene 1
- KIF14
Kinesin family member 14
- OC
Ovarian cancer
- E2F6
E2F transcription factor 6
- LCN2
Lipocalin-2
- HOXB-AS1
Homeobox B cluster antisense RNA 1
- GBM
Glioblastoma
- ILF3
Interleukin enhancer-binding factor 3
- ceRNA
Competing endogenous RNA
- miRNA
Micro RNA
- FOXP4-AS1
Forkhead box P4 antisense RNA 1
- CRC
Colorectal cancer
- FOXC2-AS1
Forkhead Box C2 antisense RNA 1
- TPT1-AS1
Tumor protein translationally controlled 1 antisense RNA 1
- BCa
Bladder cancer
- AUF1
AU-rich element RNA-binding factor 1
- PDCD4-AS1
Programmed cell death 4 antisense RNA 1
- MACC1-AS1
Metastasis associated in colon cancer-1 antisense RNA 1
- SLCO4A1-AS1
SLCO4A1 antisense RNA 1
- ZFPM2-AS1
Zinc finger protein multitype 2 antisense RNA 1
- MIF
Migration inhibitory factor
- FEZF1-AS1
FEZ finger zinc 1 antisense 1
- PKM2
Pyruvate kinase 2
- KTN1-AS1
Kinectin 1-Antisense RNA 1
- DEPDC1
DEP domain containing 1
- ERBB2IP
Erbb2 interacting protein
- NACC1
Nucleus accumbens-associated protein 1
- CBX4
Chromobox homolog 4
- STMN1
Stathmin 1
- HAND2-AS1
Heart and neural crest derivatives expressed 2-antisense RNA 1
- KCNT2
Potassium sodium-activated channel subfamily T member 2
- TCEAL7
Transcription Elongation Factor A-like 7
- FGF13-AS1
Fibroblast growth factor 13 antisense RNA 1
- IGF2BPs
Insulin-like growth factor-2 mRNA-binding proteins
- FGF14-AS2
Fibroblast Growth Factor 14 antisense RNA 2
- TP73-AS1
Tumour protein P73 antisense RNA 1
- LAPTM4B
Lysosomal-associated transmembrane protein 4B
- AML
Acute myeloid leukemia
- PTEN
Phosphatase and Tensin Homolog deleted on Chromosome 10
- EMT
Epithelial–mesenchymal transition
- NLRP3
NOD-, LRR- and pyrin domain-containing 3
- OC
Ovarian cancer
- FOXF2
Forkhead box F2
- TSCC
Tongue squamous cell carcinoma
- LEF1-AS1
Lymphoid enhancer-binding factor 1 antisense RNA 1
- SOX9
SRY-Box 9
- CPEB4
Cytoplasmic polyadenylation element binding proteins 4
- PTN
Pleiotrophin
- FOSL2
FOS-like antigen 2
- LPAATβ
Lysophosphatidic acid acyltransferase
- OIP5-AS1
OIP5 antisense RNA 1
- FAM83H-AS1
Family with sequence similarity 83 member H antisense RNA 1
- SBF2-AS1
SET-binding factor 2 antisense RNA 1
- MBNL3
Muscleblind-like 3
- NPC
Nasopharyngeal carcinoma
- PTPRG-AS1
Protein tyrosine phosphatase receptor gamma antisense RNA 1
- PRC1
Protein regulator of cytokinesis 1
- TCF4
Transcription Factor-4
- CUL4B
Cullin 4B
- TTN-AS1
Titin-antisense RNA1
- SOX21-AS1
SOX21 antisense RNA 1
- ZNF385D-AS2
Zinc finger protein 385D antisense RNA 2
- HIF1A-AS2
Hypoxia-inducible factor 1α antisense RNA 2
- PCNA
Proliferating cell nuclear antigen
- FBXW7
F-box with 7 tandem WD40
- ROCK2
Rho-associated protein kinase 2
- LIFR-AS1
Leukemia inhibitory factor receptor antisense RNA 1
Authors’ contributions
BL and JYL made the literature analysis and wrote, discussed and revised the manuscript of this review, WX and JHL made literature search and draft the manuscript of this review, JT, JRW and BL edited the manuscript, ZL, LW and GGY revised the design of the image. All authors read and approved the final manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (81802556), the Hunan Province Natural Science Foundation (2019JJ30039), Huxiang Young Talents Plan Project of Hunan Province (2019RS2015), the New Xiangya Talent Projects of the Third Xiangya Hospital of Central South University (JY201615), the Scientific Projects of Changsha Administration of Science &Technology (kq1901129), and the Scientific Projects of Health Commission of Hunan Province (B2017034, 20201041).
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors agree with the final version of the manuscript and give their consent for its publication.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Slack FJ, Chinnaiyan AM. The role of non-coding RNAs in oncology. Cell. 2019;179(5):1033–1055. doi: 10.1016/j.cell.2019.10.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rinn JL, Chang HY. Genome regulation by long noncoding RNAs. Annu Rev Biochem. 2012;81:145–166. doi: 10.1146/annurev-biochem-051410-092902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Chan JJ, Tay Y. Noncoding RNA: RNA regulatory networks in cancer. Int J Mol Sci. 2018;19(5):1310. doi: 10.3390/ijms19051310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mattick JS, Makunin IV. Non-coding RNA. Hum Mol Genet. 2006;15 Spec No 1:R17–R29. doi: 10.1093/hmg/ddl046. [DOI] [PubMed] [Google Scholar]
- 5.Adams BD, Parsons C, Walker L, Zhang WC, Slack FJ. Targeting noncoding RNAs in disease. J Clin Invest. 2017;127(3):761–771. doi: 10.1172/JCI84424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wilusz JE, Sunwoo H, Spector DL. Long noncoding RNAs: functional surprises from the RNA world. Genes Dev. 2009;23(13):1494–1504. doi: 10.1101/gad.1800909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gibb EA, Brown CJ, Lam WL. The functional role of long non-coding RNA in human carcinomas. Mol Cancer. 2011;10:38. doi: 10.1186/1476-4598-10-38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Latgé G, Poulet C, Bours V, Josse C, Jerusalem G. Natural antisense transcripts: molecular mechanisms and implications in breast cancers. Int J Mol Sci. 2018;19(1):123. doi: 10.3390/ijms19010123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Katayama S, Tomaru Y, Kasukawa T, Waki K, Nakanishi M, Nakamura M, Nishida H, Yap CC, Suzuki M, Kawai J, et al. Antisense transcription in the mammalian transcriptome. Science. 2005;309(5740):1564–1566. doi: 10.1126/science.1112009. [DOI] [PubMed] [Google Scholar]
- 10.Prensner JR, Chinnaiyan AM. The emergence of lncRNAs in cancer biology. Cancer Discov. 2011;1(5):391–407. doi: 10.1158/2159-8290.CD-11-0209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981. doi: 10.1158/0008-5472.CAN-16-2634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zhao S, Zhang X, Chen S, Zhang S. Natural antisense transcripts in the biological hallmarks of cancer: powerful regulators hidden in the dark. J Exp Clin Cancer Res. 2020;39(1):187. doi: 10.1186/s13046-020-01700-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Pelechano V, Steinmetz LM. Gene regulation by antisense transcription. Nat Rev Genet. 2013;14(12):880–893. doi: 10.1038/nrg3594. [DOI] [PubMed] [Google Scholar]
- 14.Bach DH, Lee SK. Long noncoding RNAs in cancer cells. Cancer Lett. 2018;419:152–166. doi: 10.1016/j.canlet.2018.01.053. [DOI] [PubMed] [Google Scholar]
- 15.Magistri M, Faghihi MA, St LGR, Wahlestedt C. Regulation of chromatin structure by long noncoding RNAs: focus on natural antisense transcripts. Trends Genet. 2012;28(8):389–396. doi: 10.1016/j.tig.2012.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Morris KV, Vogt PK. Long antisense non-coding RNAs and their role in transcription and oncogenesis. Cell Cycle. 2010;9(13):2544–2547. doi: 10.4161/cc.9.13.12145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Huang Y, Xiang B, Liu Y, Wang Y, Kan H. LncRNA CDKN2B-AS1 promotes tumor growth and metastasis of human hepatocellular carcinoma by targeting let-7c-5p/NAP1L1 axis. Cancer Lett. 2018;437:56–66. doi: 10.1016/j.canlet.2018.08.024. [DOI] [PubMed] [Google Scholar]
- 18.Cai Y, Dong ZY, Wang JY. LncRNA NNT-AS1 is a major mediator of cisplatin chemoresistance in non-small cell lung cancer through MAPK/Slug pathway. Eur Rev Med Pharmacol Sci. 2018;22(15):4879–4887. doi: 10.26355/eurrev_201808_15624. [DOI] [PubMed] [Google Scholar]
- 19.Chen M, Wu X, Ma W, Zhou Q, Wang X, Zhang R, Wang J, Yang X. Decreased expression of lncRNA VPS9D1-AS1 in gastric cancer and its clinical significance. Cancer Biomark. 2017;21(1):23–28. doi: 10.3233/CBM-170172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wang L, Wei Z, Wu K, Dai W, Zhang C, Peng J, He Y. Long noncoding RNA B3GALT5-AS1 suppresses colon cancer liver metastasis via repressing microRNA-203. Aging (Albany NY) 2018;10(12):3662–3682. doi: 10.18632/aging.101628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sun Y, Li D, Zhang R, Peng S, Zhang G, Yang T, Qian A. Strategies to identify natural antisense transcripts. Biochimie. 2017;132:131–151. doi: 10.1016/j.biochi.2016.11.006. [DOI] [PubMed] [Google Scholar]
- 22.Yang Y, Chen L, Gu J, Zhang H, Yuan J, Lian Q, Lv G, Wang S, Wu Y, Yang YT, et al. Recurrently deregulated lncRNAs in hepatocellular carcinoma. Nat Commun. 2017;8:14421. doi: 10.1038/ncomms14421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Barth DA, Juracek J, Slaby O, Pichler M, Calin GA. lncRNA and mechanisms of drug resistance in cancers of the genitourinary system. Cancers (Basel) 2020;12(8):2148. doi: 10.3390/cancers12082148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Podralska M, Ciesielska S, Kluiver J, van den Berg A, Dzikiewicz-Krawczyk A, Slezak-Prochazka I. Non-coding RNAs in cancer radiosensitivity: MicroRNAs and lncRNAs as regulators of radiation-induced signaling pathways. Cancers (Basel) 2020;12(6):1662. doi: 10.3390/cancers12061662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rashid F, Shah A, Shan G. Long non-coding RNAs in the cytoplasm. Genom Proteom Bioinform. 2016;14(2):73–80. doi: 10.1016/j.gpb.2016.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Meissner A, Mikkelsen TS, Gu H, Wernig M, Hanna J, Sivachenko A, Zhang X, Bernstein BE, Nusbaum C, Jaffe DB, et al. Genome-scale DNA methylation maps of pluripotent and differentiated cells. Nature. 2008;454(7205):766–770. doi: 10.1038/nature07107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell. 2012;150(1):12–27. doi: 10.1016/j.cell.2012.06.013. [DOI] [PubMed] [Google Scholar]
- 28.Morris KV. Long antisense non-coding RNAs function to direct epigenetic complexes that regulate transcription in human cells. Epigenetics-US. 2009;4(5):296–301. doi: 10.4161/epi.4.5.9282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wu DM, Zheng ZH, Zhang YB, Fan SH, Zhang ZF, Wang YJ, Zheng YL, Lu J. Down-regulated lncRNA DLX6-AS1 inhibits tumorigenesis through STAT3 signaling pathway by suppressing CADM1 promoter methylation in liver cancer stem cells. J Exp Clin Cancer Res. 2019;38(1):237. doi: 10.1186/s13046-019-1239-3. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 30.Li X, Lv J, Liu S. MCM3AP-AS1 KD inhibits proliferation, invasion, and migration of PCa Cells via DNMT1/DNMT3 (A/B) methylation-mediated upregulation of NPY1R. Mol Ther Nucleic Acids. 2020;20:265–278. doi: 10.1016/j.omtn.2020.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 31.He J, Wu K, Guo C, Zhou JK, Pu W, Deng Y, Zuo Y, Zhao Y, Liu L, Wei YQ, et al. Long non-coding RNA AFAP1-AS1 plays an oncogenic role in promoting cell migration in non-small cell lung cancer. Cell Mol Life Sci. 2018;75(24):4667–4681. doi: 10.1007/s00018-018-2923-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Liu D, Wu K, Yang Y, Zhu D, Zhang C, Zhao S. Long noncoding RNA ADAMTS9-AS2 suppresses the progression of esophageal cancer by mediating CDH3 promoter methylation. Mol Carcinog. 2020;59(1):32–44. doi: 10.1002/mc.23126. [DOI] [PubMed] [Google Scholar]
- 33.Khalil AM, Guttman M, Huarte M, Garber M, Raj A, Rivea MD, Thomas K, Presser A, Bernstein BE, van Oudenaarden A, et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc Natl Acad Sci U S A. 2009;106(28):11667–11672. doi: 10.1073/pnas.0904715106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Khorasanizadeh S. The nucleosome: from genomic organization to genomic regulation. Cell. 2004;116(2):259–272. doi: 10.1016/S0092-8674(04)00044-3. [DOI] [PubMed] [Google Scholar]
- 35.Yin D, Lu X, Su J, He X, De W, Yang J, Li W, Han L, Zhang E. Long noncoding RNA AFAP1-AS1 predicts a poor prognosis and regulates non-small cell lung cancer cell proliferation by epigenetically repressing p21 expression. Mol Cancer. 2018;17(1):92. doi: 10.1186/s12943-018-0836-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Su W, Xu M, Chen X, Chen N, Gong J, Nie L, Li L, Li X, Zhang M, Zhou Q. Long noncoding RNA ZEB1-AS1 epigenetically regulates the expressions of ZEB1 and downstream molecules in prostate cancer. Mol Cancer. 2017;16(1):142. doi: 10.1186/s12943-017-0711-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zhao H, Xu Q. Long non-coding RNA DLX6-AS1 mediates proliferation, invasion and apoptosis of endometrial cancer cells by recruiting p300/E2F1 in DLX6 promoter region. J Cell Mol Med. 2020;24(21):12572–12584. doi: 10.1111/jcmm.15810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Dong H, Wang W, Mo S, Chen R, Zou K, Han J, Zhang F, Hu J. SP1-induced lncRNA AGAP2-AS1 expression promotes chemoresistance of breast cancer by epigenetic regulation of MyD88. J Exp Clin Cancer Res. 2018;37(1):202. doi: 10.1186/s13046-018-0875-3. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 39.Xu TP, Wang WY, Ma P, Shuai Y, Zhao K, Wang YF, Li W, Xia R, Chen WM, Zhang EB, et al. Upregulation of the long noncoding RNA FOXD2-AS1 promotes carcinogenesis by epigenetically silencing EphB3 through EZH2 and LSD1, and predicts poor prognosis in gastric cancer. Oncogene. 2018;37(36):5020–5036. doi: 10.1038/s41388-018-0308-y. [DOI] [PubMed] [Google Scholar]
- 40.Faghihi MA, Wahlestedt C. Regulatory roles of natural antisense transcripts. Nat Rev Mol Cell Biol. 2009;10(9):637–643. doi: 10.1038/nrm2738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Xu H, Zhao G, Zhang Y, Jiang H, Wang W, Zhao D, Yu H, Qi L. Long non-coding RNA PAXIP1-AS1 facilitates cell invasion and angiogenesis of glioma by recruiting transcription factor ETS1 to upregulate KIF14 expression. J Exp Clin Cancer Res. 2019;38(1):486. doi: 10.1186/s13046-019-1474-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zhao H, Ding F, Zheng G. LncRNA TMPO-AS1 promotes LCN2 transcriptional activity and exerts oncogenic functions in ovarian cancer. FASEB J. 2020;34(9):11382–11394. doi: 10.1096/fj.201902683R. [DOI] [PubMed] [Google Scholar]
- 43.Bi Y, Mao Y, Su Z, Du J, Ye L, Xu F. HOXB-AS1 accelerates the tumorigenesis of glioblastoma via modulation of HOBX2 and HOBX3 at transcriptional and posttranscriptional levels. J Cell Physiol. 2021;236(1):93–106. doi: 10.1002/jcp.29499. [DOI] [PubMed] [Google Scholar]
- 44.Rupaimoole R, Slack FJ. MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nat Rev Drug Discov. 2017;16(3):203–222. doi: 10.1038/nrd.2016.246. [DOI] [PubMed] [Google Scholar]
- 45.Wu X, Xiao Y, Zhou Y, Zhou Z, Yan W. LncRNA FOXP4-AS1 is activated by PAX5 and promotes the growth of prostate cancer by sequestering miR-3184-5p to upregulate FOXP4. Cell Death Dis. 2019;10(7):472. doi: 10.1038/s41419-019-1699-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Lv SY, Shan TD, Pan XT, Tian ZB, Liu XS, Liu FG, Sun XG, Xue HG, Li XH, Han Y, et al. The lncRNA ZEB1-AS1 sponges miR-181a-5p to promote colorectal cancer cell proliferation by regulating Wnt/β-catenin signaling. Cell Cycle. 2018;17(10):1245–1254. doi: 10.1080/15384101.2018.1471317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Xiong WC, Han N, Wu N, Zhao KL, Han C, Wang HX, Ping GF, Zheng PF, Feng H, Qin L, et al. Interplay between long noncoding RNA ZEB1-AS1 and miR-101/ZEB1 axis regulates proliferation and migration of colorectal cancer cells. Am J Transl Res. 2018;10(2):605–617. [PMC free article] [PubMed] [Google Scholar]
- 48.Chen Y, Gu M, Liu C, Wan X, Shi Q, Chen Q, Wang Z. Long noncoding RNA FOXC2-AS1 facilitates the proliferation and progression of prostate cancer via targeting miR-1253/EZH2. Gene. 2019;686:37–42. doi: 10.1016/j.gene.2018.10.085. [DOI] [PubMed] [Google Scholar]
- 49.Jiang H, Huang G, Zhao N, Zhang T, Jiang M, He Y, Zhou X, Jiang X. Long non-coding RNA TPT1-AS1 promotes cell growth and metastasis in cervical cancer via acting AS a sponge for miR-324-5p. J Exp Clin Cancer Res. 2018;37(1):169. doi: 10.1186/s13046-018-0846-8. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 50.Zhao X, Wang D, Ding Y, Zhou J, Liu G, Ji Z. lncRNA ZEB1-AS1 promotes migration and metastasis of bladder cancer cells by post-transcriptional activation of ZEB1. Int J Mol Med. 2019;44(1):196–206. doi: 10.3892/ijmm.2019.4187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Jadaliha M, Gholamalamdari O, Tang W, Zhang Y, Petracovici A, Hao Q, Tariq A, Kim TG, Holton SE, Singh DK, et al. A natural antisense lncRNA controls breast cancer progression by promoting tumor suppressor gene mRNA stability. PLoS Genet. 2018;14(11):e1007802. doi: 10.1371/journal.pgen.1007802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Bao G, Huang J, Pan W, Li X, Zhou T. Long noncoding RNA CERS6-AS1 functions as a malignancy promoter in breast cancer by binding to IGF2BP3 to enhance the stability of CRS6 mRNA. Cancer Med. 2020;9(1):278–289. doi: 10.1002/cam4.2675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Chen R, Zhang X, Wang C. LncRNA HOXB-AS1 promotes cell growth in multiple myeloma via FUT4 mRNA stability by ELAVL1. J Cell Biochem. 2020;121(10):4043–4051. doi: 10.1002/jcb.29573. [DOI] [PubMed] [Google Scholar]
- 54.Yu J, Han Z, Sun Z, Wang Y, Zheng M, Song C. LncRNA SLCO4A1-AS1 facilitates growth and metastasis of colorectal cancer through β-catenin-dependent Wnt pathway. J Exp Clin Cancer Res. 2018;37(1):222. doi: 10.1186/s13046-018-0896-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kong F, Deng X, Kong X, Du Y, Li L, Zhu H, Wang Y, Xie D, Guha S, Li Z, et al. ZFPM2-AS1, a novel lncRNA, attenuates the p53 pathway and promotes gastric carcinogenesis by stabilizing MIF. Oncogene. 2018;37(45):5982–5996. doi: 10.1038/s41388-018-0387-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Bian Z, Zhang J, Li M, Feng Y, Wang X, Zhang J, Yao S, Jin G, Du J, Han W, et al. LncRNA-FEZF1-AS1 promotes tumor proliferation and metastasis in colorectal cancer by regulating PKM2 signaling. Clin Cancer Res. 2018;24(19):4808–4819. doi: 10.1158/1078-0432.CCR-17-2967. [DOI] [PubMed] [Google Scholar]
- 57.Meng W, Cui W, Zhao L, Chi W, Cao H, Wang B. Aberrant methylation and downregulation of ZNF667-AS1 and ZNF667 promote the malignant progression of laryngeal squamous cell carcinoma. J Biomed Sci. 2019;26(1):13. doi: 10.1186/s12929-019-0506-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Wang Y, Li D, Lu J, Chen L, Zhang S, Qi W, Li W, Xu H. Long noncoding RNA TTN-AS1 facilitates tumorigenesis and metastasis by maintaining TTN expression in skin cutaneous melanoma. Cell Death Dis. 2020;11(8):664. doi: 10.1038/s41419-020-02895-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Byun Y, Choi YC, Jeong Y, Yoon J, Baek K. Long noncoding RNA expression profiling reveals upregulation of uroplakin 1A and uroplakin 1A antisense RNA 1 under hypoxic conditions in lung cancer cells. Mol Cells. 2020;43(12):975–988. doi: 10.14348/molcells.2020.0126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Huarte M. The emerging role of lncRNAs in cancer. Nat Med. 2015;21(11):1253–1261. doi: 10.1038/nm.3981. [DOI] [PubMed] [Google Scholar]
- 61.Aprile M, Katopodi V, Leucci E, Costa V. LncRNAs in cancer: from garbage to junk. Cancers (Basel) 2020;12(11):3220. doi: 10.3390/cancers12113220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Liu C, Li X, Hao Y, Wang F, Cheng Z, Geng H, Geng D. STAT1-induced upregulation of lncRNA KTN1-AS1 predicts poor prognosis and facilitates non-small cell lung cancer progression via miR-23b/DEPDC1 axis. Aging (Albany NY) 2020;12(9):8680–8701. doi: 10.18632/aging.103191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Hu X, Xiang L, He D, Zhu R, Fang J, Wang Z, Cao K. The long noncoding RNA KTN1-AS1 promotes bladder cancer tumorigenesis via KTN1 cis-activation and the consequent initiation of Rho GTPase-mediated signaling. Clin Sci (Lond) 2021;135:555–574. doi: 10.1042/CS20200908. [DOI] [PubMed] [Google Scholar]
- 64.Mu Y, Tang Q, Feng H, Zhu L, Wang Y. lncRNA KTN1-AS1 promotes glioma cell proliferation and invasion by negatively regulating miR-505-3p. Oncol Rep. 2020;44(6):2645–2655. doi: 10.3892/or.2020.7821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zhang L, Wang L, Wang Y, Chen T, Liu R, Yang W, Liu Q, Tu K. LncRNA KTN1-AS1 promotes tumor growth of hepatocellular carcinoma by targeting miR-23c/ERBB2IP axis. Biomed Pharmacother. 2019;109:1140–1147. doi: 10.1016/j.biopha.2018.10.105. [DOI] [PubMed] [Google Scholar]
- 66.Tao HF, Shen JX, Hou ZW, Chen SY, Su YZ, Fang JL. lncRNA FOXP4-AS1 predicts poor prognosis and accelerates the progression of mantle cell lymphoma through the miR-423-5p/NACC1 pathway. Oncol Rep. 2020;45(2):469–480. doi: 10.3892/or.2020.7897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Zhao J, Yang T, Li L. LncRNA FOXP4-AS1 is involved in cervical cancer progression via regulating miR-136-5p/CBX4 axis. OncoTargets ther. 2020;13:2347–2355. doi: 10.2147/OTT.S241818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zhong LK, Zhou J, He X, He BF, Zhou XW, Zhu JL, Liu J, Qiu YH. Long non-coding RNA FOXP4-AS1 acts as an adverse prognostic factor and regulates proliferation and apoptosis in nasopharyngeal carcinoma. Eur Rev Med Pharmacol Sci. 2020;24(15):8008–8016. doi: 10.26355/eurrev_202008_22484. [DOI] [PubMed] [Google Scholar]
- 69.Li Y, Li T, Yang Y, Kang W, Dong S, Cheng S. YY1-induced upregulation of FOXP4-AS1 and FOXP4 promote the proliferation of esophageal squamous cell carcinoma cells. Cell Biol Int. 2020;44(7):1447–1457. doi: 10.1002/cbin.11338. [DOI] [PubMed] [Google Scholar]
- 70.Yang X, Wang CC, Lee WYW, Trovik J, Chung TKH, Kwong J. Long non-coding RNA HAND2-AS1 inhibits invasion and metastasis in endometrioid endometrial carcinoma through inactivating neuromedin U. Cancer Lett. 2018;413:23–34. doi: 10.1016/j.canlet.2017.10.028. [DOI] [PubMed] [Google Scholar]
- 71.Yu L, Li H, Li Z, Jia J, Wu Z, Wang M, Li F, Feng Z, Xia H, Gao G. Long non-coding RNA HAND2-AS1 inhibits growth and migration of gastric cancer cells through regulating the miR-590-3p/KCNT2 axis. Oncotargets Ther. 2020;13:3187–3196. doi: 10.2147/OTT.S233256. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 72.Yu L, Luan W, Feng Z, Jia J, Wu Z, Wang M, Li F, Li Z. Long non-coding RNA HAND2-AS1 inhibits gastric cancer progression by suppressing TCEAL7 expression via targeting miR-769-5p. Dig Liver Dis. 2020;53:238–244. doi: 10.1016/j.dld.2020.08.045. [DOI] [PubMed] [Google Scholar]
- 73.Bi HQ, Li ZH, Zhang H. Long noncoding RNA HAND2-AS1 reduced the viability of hepatocellular carcinoma via targeting microRNA-300/SOCS5 axis. Hepatobiliary Pancreat Dis Int. 2020;19(6):567–574. doi: 10.1016/j.hbpd.2020.02.011. [DOI] [PubMed] [Google Scholar]
- 74.Gao T, Dai X, Jiang Y, He X, Yuan S, Zhao P. LncRNA HAND2-AS1 inhibits proliferation and promotes apoptosis of non-small cell lung cancer cells by inactivating PI3K/Akt pathway. 2020. Biosci Rep. 10.1042/BSR20201870. [DOI] [PMC free article] [PubMed]
- 75.Ma F, Liu X, Zhou S, Li W, Liu C, Chadwick M, Qian C. Long non-coding RNA FGF13-AS1 inhibits glycolysis and sternness properties of breast cancer cells through FGF13-AS1/IGF2BPs/Myc feedback loop. Cancer Lett. 2019;450:63–75. doi: 10.1016/j.canlet.2019.02.008. [DOI] [PubMed] [Google Scholar]
- 76.Yang F, Liu YH, Dong SY, Ma RM, Bhandari A, Zhang XH, Wang OC. A novel long non-coding RNA FGF14-AS2 is correlated with progression and prognosis in breast cancer. Biochem Biophys Res Commun. 2016;470(3):479–483. doi: 10.1016/j.bbrc.2016.01.147. [DOI] [PubMed] [Google Scholar]
- 77.Jin Y, Zhang M, Duan R, Yang J, Yang Y, Wang J, Jiang C, Yao B, Li L, Yuan H, et al. Long noncoding RNA FGF14-AS2 inhibits breast cancer metastasis by regulating the miR-370-3p/FGF14 axis. Cell Death Discov. 2020;6:103. doi: 10.1038/s41420-020-00334-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hou R, Liu Y, Su Y, Shu Z. Overexpression of long non-coding RNA FGF14-AS2 inhibits colorectal cancer proliferation via the RERG/Ras/ERK signaling by sponging microRNA-1288-3p. PATHOL ONCOL RES. 2020;26(4):2659–2667. doi: 10.1007/s12253-020-00862-8. [DOI] [PubMed] [Google Scholar]
- 79.Jiang Q, Xing W, Cheng J, Yu Y. Long non-coding RNA TP73-AS1 promotes the development of lung cancer by targeting the miR-27b-3p/LAPTM4B axis. Onco Targets Ther. 2020;13:7019–7031. doi: 10.2147/OTT.S234443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Zou Q, Zhou E, Xu F, Zhang D, Yi W, Yao J. A TP73-AS1/miR-200a/ZEB1 regulating loop promotes breast cancer cell invasion and migration. J Cell Biochem. 2018;119(2):2189–2199. doi: 10.1002/jcb.26380. [DOI] [PubMed] [Google Scholar]
- 81.Li S, Huang Y, Huang Y, Fu Y, Tang D, Kang R, Zhou R, Fan XG. The long non-coding RNA TP73-AS1 modulates HCC cell proliferation through miR-200a-dependent HMGB1/RAGE regulation. J Exp Clin Cancer Res. 2017;36(1):51. doi: 10.1186/s13046-017-0519-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Yuan Z, Li L, Zheng M, Xu J, Wang W. lncRNA TP73-AS1 regulates miR-21/PTEN axis to affect cell proliferation in acute myeloid leukemia. Cancer Biother Radiopharm. 2020;36:268–272. doi: 10.1089/cbr.2019.3142. [DOI] [PubMed] [Google Scholar]
- 83.Tuo Z, Zhang J, Xue W. LncRNA TP73-AS1 predicts the prognosis of bladder cancer patients and functions as a suppressor for bladder cancer by EMT pathway. Biochem Biophys Res Commun. 2018;499(4):875–881. doi: 10.1016/j.bbrc.2018.04.010. [DOI] [PubMed] [Google Scholar]
- 84.Ren N, Jiang T, Wang C, Xie S, Xing Y, Piao D, Zhang T, Zhu Y. LncRNA ADAMTS9-AS2 inhibits gastric cancer (GC) development and sensitizes chemoresistant GC cells to cisplatin by regulating miR-223-3p/NLRP3 axis. Aging (Albany NY) 2020;12(11):11025–11041. doi: 10.18632/aging.103314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Wang A, Jin C, Li H, Qin Q, Li L. LncRNA ADAMTS9-AS2 regulates ovarian cancer progression by targeting miR-182-5p/FOXF2 signaling pathway. Int J Biol Macromol. 2018;120(Pt B):1705–1713. doi: 10.1016/j.ijbiomac.2018.09.179. [DOI] [PubMed] [Google Scholar]
- 86.Li Y, Wan Q, Wang W, Mai L, Sha L, Mashrah M, Lin Z, Pan C. LncRNA ADAMTS9-AS2 promotes tongue squamous cell carcinoma proliferation, migration and EMT via the miR-600/EZH2 axis. Biomed Pharmacother. 2019;112:108719. doi: 10.1016/j.biopha.2019.108719. [DOI] [PubMed] [Google Scholar]
- 87.Xiang C, Zhang Y, Zhang Y, Liu C, Hou Y, Zhang Y. lncRNA LEF1-AS1 promotes proliferation and induces apoptosis of non-small-cell lung cancer cells by regulating miR-221/PTEN signaling. Cancer Manag Res. 2020;12:3845–3850. doi: 10.2147/CMAR.S246422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Sun T, Liu Z, Zhang R, Ma S, Lin T, Li Y, Yang S, Zhang W, Wang Y. Long non-coding RNA LEF1-AS1 promotes migration, invasion and metastasis of colon cancer cells through miR-30-5p/SOX9 axis. Onco Targets Ther. 2020;13:2957–2972. doi: 10.2147/OTT.S232839. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 89.Lu X, Qiao L, Liu Y. Long noncoding RNA LEF1-AS1 binds with HNRNPL to boost the proliferation, migration, and invasion in osteosarcoma by enhancing the mRNA stability of LEF1. J Cell Biochem. 2020;121(10):4064–4073. doi: 10.1002/jcb.29579. [DOI] [PubMed] [Google Scholar]
- 90.Congrains-Castillo A, Niemann FS, Santos DA, Olalla-Saad ST. LEF1-AS1, long non-coding RNA, inhibits proliferation in myeloid malignancy. J Cell Mol Med. 2019;23:3021–3025. doi: 10.1111/jcmm.14152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Yu AM, Ingelman-Sundberg M, Cherrington NJ, Aleksunes LM, Zanger UM, Xie W, Jeong H, Morgan ET, Turnbaugh PJ, Klaassen CD, et al. Regulation of drug metabolism and toxicity by multiple factors of genetics, epigenetics, lncRNAs, gut microbiota, and diseases: a meeting report of the 21(st) International Symposium on Microsomes and Drug Oxidations (MDO) Acta Pharm Sin B. 2017;7(2):241–248. doi: 10.1016/j.apsb.2016.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Liu H, Zhang J, Luo X, Zeng M, Xu L, Zhang Q, Liu H, Guo J, Xu L. Overexpression of the long noncoding RNA FOXD2-AS1 promotes cisplatin resistance in esophageal squamous cell carcinoma through the miR-195/Akt/mTOR axis. Oncol Res. 2020;28(1):65–73. doi: 10.3727/096504019X15656904013079. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 93.Shangguan W, Lv X, Tian N. FoxD2-AS1 is a prognostic factor in glioma and promotes temozolomide resistance in a O(6)-methylguanine-DNA methyltransferase-dependent manner. Korean J Physiol Pharmacol. 2019;23(6):475–482. doi: 10.4196/kjpp.2019.23.6.475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Gu N, Wang X, Di Z, Xiong J, Ma Y, Yan Y, Qian Y, Zhang Q, Yu J. Silencing lncRNA FOXD2-AS1 inhibits proliferation, migration, invasion and drug resistance of drug-resistant glioma cells and promotes their apoptosis via microRNA-98-5p/CPEB4 axis. Aging (Albany NY) 2019;11(22):10266–10283. doi: 10.18632/aging.102455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Li R, Chen S, Zhan J, Li X, Liu W, Sheng X, Lu Z, Zhong R, Chen L, Luo X, et al. Long noncoding RNA FOXD2-AS1 enhances chemotherapeutic resistance of laryngeal squamous cell carcinoma via STAT3 activation. Cell Death Dis. 2020;11(1):41. doi: 10.1038/s41419-020-2232-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.An Q, Zhou L, Xu N. Long noncoding RNA FOXD2-AS1 accelerates the gemcitabine-resistance of bladder cancer by sponging miR-143. Biomed Pharmacother. 2018;103:415–420. doi: 10.1016/j.biopha.2018.03.138. [DOI] [PubMed] [Google Scholar]
- 97.Sun X, Tian C, Zhang H, Han K, Zhou M, Gan Z, Zhu H, Min D. Long noncoding RNA OIP5-AS1 mediates resistance to doxorubicin by regulating miR-137-3p/PTN axis in osteosarcoma. Biomed Pharmacother. 2020;128:110201. doi: 10.1016/j.biopha.2020.110201. [DOI] [PubMed] [Google Scholar]
- 98.Liu L, Wang S. Long non-coding RNA OIP5-AS1 knockdown enhances CDDP sensitivity in osteosarcoma via miR-377-3p/FOSL2 axis. Onco Targets Ther. 2020;13:3853–3866. doi: 10.2147/OTT.S232918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Song L, Zhou Z, Gan Y, Li P, Xu Y, Zhang Z, Luo F, Xu J, Zhou Q, Dai F. Long noncoding RNA OIP5-AS1 causes cisplatin resistance in osteosarcoma through inducing the LPAATβ/PI3K/AKT/mTOR signaling pathway by sponging the miR-340-5p. J Cell Biochem. 2019;120(6):9656–9666. doi: 10.1002/jcb.28244. [DOI] [PubMed] [Google Scholar]
- 100.Liang J, Tian XF, Yang W. Effects of long non-coding RNA Opa-interacting protein 5 antisense RNA 1 on colon cancer cell resistance to oxaliplatin and its regulation of microRNA-137. World J Gastroenterol. 2020;26:1474–1489. doi: 10.3748/wjg.v26.i13.1474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Dou Q, Xu Y, Zhu Y, Hu Y, Yan Y, Yan H. LncRNA FAM83H-AS1 contributes to the radioresistance, proliferation, and metastasis in ovarian cancer through stabilizing HuR protein. Eur J Pharmacol. 2019;852:134–141. doi: 10.1016/j.ejphar.2019.03.002. [DOI] [PubMed] [Google Scholar]
- 102.Yu Z, Wang G, Zhang C, Liu Y, Chen W, Wang H, Liu H. LncRNA SBF2-AS1 affects the radiosensitivity of non-small cell lung cancer via modulating microRNA-302a/MBNL3 axis. Cell Cycle. 2020;19(3):300–316. doi: 10.1080/15384101.2019.1708016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Yi L, Ouyang L, Wang S, Li SS, Yang XM. Long noncoding RNA PTPRG-AS1 acts as a microRNA-194-3p sponge to regulate radiosensitivity and metastasis of nasopharyngeal carcinoma cells via PRC1. J Cell Physiol. 2019;234(10):19088–19102. doi: 10.1002/jcp.28547. [DOI] [PubMed] [Google Scholar]
- 104.Ma Q, Niu R, Huang W, Da L, Tang Y, Jiang D, Xi Y, Zhang C. Long Noncoding RNA PTPRG antisense RNA 1 reduces radiosensitivity of nonsmall cell lung cancer cells via regulating MiR-200c-3p/TCF4. Technol Cancer Res Treat. 2020;19:1079210263. doi: 10.1177/1533033820942615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Zuo Z, Ji S, He L, Zhang Y, Peng Z, Han J. LncRNA TTN-AS1/miR-134–5p/PAK3 axis regulates the radiosensitivity of human large intestine cancer cells through the P21 pathway and AKT/GSK-3β/β-catenin pathway. Cell Biol Int. 2020;44:2284–2292. doi: 10.1002/cbin.11436. [DOI] [PubMed] [Google Scholar]
- 106.Zhao L, Wang J, Li Y, Song T, Wu Y, Fang S, Bu D, Li H, Sun L, Pei D, et al. NONCODEV6: an updated database dedicated to long non-coding RNA annotation in both animals and plants. Nucleic Acids Res. 2021;49(D1):D165–D171. doi: 10.1093/nar/gkaa1046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Bao Z, Yang Z, Huang Z, Zhou Y, Cui Q, Dong D. LncRNADisease 2.0: an updated database of long non-coding RNA-associated diseases. Nucleic Acids RES. 2019;47(D1):D1034–D1037. doi: 10.1093/nar/gky905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Volders PJ, Anckaert J, Verheggen K, Nuytens J, Martens L, Mestdagh P, Vandesompele J. LNCipedia 5: towards a reference set of human long non-coding RNAs. Nucleic Acids Res. 2019;47(D1):D135–D139. doi: 10.1093/nar/gky1031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Wei C, Wang H, Xu F, Liu Z, Jiang R. LncRNA SOX21-AS1 is associated with progression of hepatocellular carcinoma and predicts prognosis through epigenetically silencing p21. Biomed Pharmacother. 2018;104:137–144. doi: 10.1016/j.biopha.2018.05.010. [DOI] [PubMed] [Google Scholar]
- 110.Zhang Z, Wang S, Liu Y, Meng Z, Chen F. Low lncRNA ZNF385D-AS2 expression and its prognostic significance in liver cancer. Oncol Rep. 2019;42(3):1110–1124. doi: 10.3892/or.2019.7238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Huang W, Su X, Yan W, Kong Z, Wang D, Huang Y, Zhai Q, Zhang X, Wu H, Li Y, et al. Overexpression of AR-regulated lncRNA TMPO-AS1 correlates with tumor progression and poor prognosis in prostate cancer. Prostate. 2018;78(16):1248–1261. doi: 10.1002/pros.23700. [DOI] [PubMed] [Google Scholar]
- 112.Shen Q, Jiang Y. LncRNA NNT-AS1 promotes the proliferation, and invasion of lung cancer cells via regulating miR-129-5p expression. Biomed Pharmacother. 2018;105:176–181. doi: 10.1016/j.biopha.2018.05.123. [DOI] [PubMed] [Google Scholar]
- 113.Guo X, Lee S, Cao P. The inhibitive effect of sh-HIF1A-AS2 on the proliferation, invasion, and pathological damage of breast cancer via targeting miR-548c-3p through regulating HIF-1α/VEGF pathway in vitro and vivo. Onco Targets Ther. 2019;12:825–834. doi: 10.2147/OTT.S192377. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 114.Wang J, Ding W, Xu Y, Tao E, Mo M, Xu W, Cai X, Chen X, Yuan J, Wu X. Long non-coding RNA RHPN1-AS1 promotes tumorigenesis and metastasis of ovarian cancer by acting as a ceRNA against miR-596 and upregulating LETM1. Aging (Albany NY) 2020;12(5):4558–4572. doi: 10.18632/aging.102911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Zhang Y, Huang YX, Wang DL, Yang B, Yan HY, Lin LH, Li Y, Chen J, Xie LM, Huang YS, et al. LncRNA DSCAM-AS1 interacts with YBX1 to promote cancer progression by forming a positive feedback loop that activates FOXA1 transcription network. Theranostics. 2020;10(23):10823–10837. doi: 10.7150/thno.47830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Miao S, Wang J, Xuan L, Liu X. LncRNA TTN-AS1 acts as sponge for miR-15b-5p to regulate FBXW7 expression in ovarian cancer. BioFactors. 2020;46:600–607. doi: 10.1002/biof.1622. [DOI] [PubMed] [Google Scholar]
- 117.Liu X, Li Y, Wen J, Qi T, Wang Y. Long non-coding RNA TTN-AS1 promotes tumorigenesis of ovarian cancer through modulating the miR-139-5p/ROCK2 axis. Biomed Pharmacother. 2020;125:109882. doi: 10.1016/j.biopha.2020.109882. [DOI] [PubMed] [Google Scholar]
- 118.Wang HF, Lv JQ, Li HH, Wang W, Lin FQ. High long non-coding LIFR-AS1 expression correlates with poor survival in gastric carcinoma. Eur Rev Med Pharmacol Sci. 2020;24(10):5378–5384. doi: 10.26355/eurrev_202005_21321. [DOI] [PubMed] [Google Scholar]
- 119.Zhao J, Li X, Fu L, Zhang N, Yang J, Cai J. lncRNA LIFR-AS1 inhibits gastric carcinoma cell proliferation, migration and invasion by sponging miR-4698. Mol Med Rep. 2021;23(2):1. doi: 10.3892/mmr.2020.11792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Pan H, Ding Y, Jiang Y, Wang X, Rao J, Zhang X, Yu H, Hou Q, Li T. LncRNA LIFR-AS1 promotes proliferation and invasion of gastric cancer cell via miR-29a-3p/COL1A2 axis. Cancer Cell Int. 2021;21(1):7. doi: 10.1186/s12935-020-01644-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Schmitt AM, Chang HY. Long noncoding RNAs in cancer pathways. Cancer Cell. 2016;29(4):452–463. doi: 10.1016/j.ccell.2016.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Wei Y, Wang Z, Zong Y, Deng D, Chen P, Lu J. LncRNA MFI2-AS1 promotes HCC progression and metastasis by acting as a competing endogenous RNA of miR-134 to upregulate FOXM1 expression. Biomed Pharmacother. 2020;125:109890. doi: 10.1016/j.biopha.2020.109890. [DOI] [PubMed] [Google Scholar]
- 123.Wang YG, Wang T, Shi M, Zhai B. Long noncoding RNA EPB41L4A-AS2 inhibits hepatocellular carcinoma development by sponging miR-301a-5p and targeting FOXL1. J Exp Clin Cancer Res. 2019;38(1):153. doi: 10.1186/s13046-019-1128-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Zheng X, Zhang J, Fang T, Wang X, Wang S, Ma Z, Xu Y, Han C, Sun M, Xu L, et al. The long non-coding RNA PIK3CD-AS2 promotes lung adenocarcinoma progression via YBX1-mediated suppression of p53 pathway. Oncogenesis. 2020;9(3):34. doi: 10.1038/s41389-020-0217-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Wang Q, Wu J, Huang H, Jiang Y, Huang Y, Fang H, Zheng G, Zhou X, Wu Y, Lei C, et al. lncRNA LIFR-AS1 suppresses invasion and metastasis of non-small cell lung cancer via the miR-942-5p/ZNF471 axis. Cancer Cell Int. 2020;20:180. doi: 10.1186/s12935-020-01228-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Wu JP, Zhang GY, Sun XZ. LncRNA ZNFX1-AS1 targeting miR-193a-3p/SDC1 regulates cell proliferation, migration and invasion of bladder cancer cells. Eur Rev Med Pharmacol Sci. 2020;24(9):4719–4728. doi: 10.26355/eurrev_202005_21160. [DOI] [PubMed] [Google Scholar]
- 127.Tang C, Cai Y, Jiang H, Lv Z, Yang C, Xu H, Li Z, Li Y. LncRNA MAGI2-AS3 inhibits bladder cancer progression by targeting the miR-31-5p/TNS1 axis. Aging (Albany NY) 2020;12(24):25547–25563. doi: 10.18632/aging.104162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Hu CC, Liang YW, Hu JL, Liu LF, Liang JW, Wang R. LncRNA RUSC1-AS1 promotes the proliferation of breast cancer cells by epigenetic silence of KLF2 and CDKN1A. Eur Rev Med Pharmacol Sci. 2019;23(15):6602–6611. doi: 10.26355/eurrev_201908_18548. [DOI] [PubMed] [Google Scholar]
- 129.Chen D, Fan Y, Wan F. LncRNA IGBP1-AS1/miR-24-1/ZIC3 loop regulates the proliferation and invasion ability in breast cancer. Cancer Cell Int. 2020;20:153. doi: 10.1186/s12935-020-01214-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Lei R, Feng L, Hong D. ELFN1-AS1 accelerates the proliferation and migration of colorectal cancer via regulation of miR-4644/TRIM44 axis. Cancer Biomark. 2020;27(4):433–443. doi: 10.3233/CBM-190559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Zheng Z, Li X, You H, Zheng X, Ruan X. LncRNA SOCS2-AS1 inhibits progression and metastasis of colorectal cancer through stabilizing SOCS2 and sponging miR-1264. Aging (Albany NY) 2020;12(11):10517–10526. doi: 10.18632/aging.103276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Yang S, Shi F, Du Y, Wang Z, Feng Y, Song J, Liu Y, Xiao M. Long non-coding RNA CTBP1-AS2 enhances cervical cancer progression via up-regulation of ZNF217 through sponging miR-3163. Cancer Cell Int. 2020;20:343. doi: 10.1186/s12935-020-01430-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Li YJ, Yang Z, Wang YY, Wang Y. Long noncoding RNA ZNF667-AS1 reduces tumor invasion and metastasis in cervical cancer by counteracting microRNA-93–3p-dependent PEG3 downregulation. Mol Oncol. 2019;13:2375–2392. doi: 10.1002/1878-0261.12565. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 134.Qu F, Zhu B, Hu YL, Mao QS, Feng Y. LncRNA HOXA-AS3 promotes gastric cancer progression by regulating miR-29a-3p/LTβR and activating NF-κB signaling. Cancer Cell Int. 2021;21(1):118. doi: 10.1186/s12935-021-01827-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Zhang XY, Zhuang HW, Wang J, Shen Y, Bu YZ, Guan BG, Xu F, Dou J. Long noncoding RNA CA3-AS1 suppresses gastric cancer migration and invasion by sponging miR-93–5p and targeting BTG3. Gene Ther. 2020. [DOI] [PubMed]
- 136.Wang X, Chen Q, Wang X, Li W, Yu G, Zhu Z, Zhang W. ZEB1 activated-VPS9D1-AS1 promotes the tumorigenesis and progression of prostate cancer by sponging miR-4739 to upregulate MEF2D. Biomed Pharmacother. 2020;122:109557. doi: 10.1016/j.biopha.2019.109557. [DOI] [PubMed] [Google Scholar]
- 137.Hua X, Liu Z, Zhou M, Tian Y, Zhao PP, Pan WH, Li CX, Huang XX, Liao ZX, Xian Q, et al. LSAMP-AS1 binds to microRNA-183-5p to suppress the progression of prostate cancer by up-regulating the tumor suppressor DCN. EBioMedicine. 2019;50:178–190. doi: 10.1016/j.ebiom.2019.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 138.Yang H, Cai MY, Rong H, Ma LR, Xu YL. ZNF667-AS1, a positively regulating MEGF10, inhibits the progression of uveal melanoma by modulating cellular aggressiveness. J Biochem Mol Toxicol. 2021;35:e22732. doi: 10.1002/jbt.22732. [DOI] [PubMed] [Google Scholar]
- 139.Xu DF, Tao XH, Yu Y, Teng Y, Huang YM, Ma JW, Fan YB. LncRNA FOXC2-AS1 stimulates proliferation of melanoma via silencing p15 by recruiting EZH2. Eur Rev Med Pharmacol Sci. 2020;24(17):8940–8946. doi: 10.26355/eurrev_202009_22835. [DOI] [PubMed] [Google Scholar]
- 140.Zhang H, Zhu C, He Z, Chen S, Li L, Sun C. LncRNA PSMB8-AS1 contributes to pancreatic cancer progression via modulating miR-382-3p/STAT1/PD-L1 axis. J Exp Clin Cancer Res. 2020;39(1):179. doi: 10.1186/s13046-020-01687-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Yan J, Jia Y, Chen H, Chen W, Zhou X. Correction to: Long non-coding RNA PXN-AS1 suppresses pancreatic cancer progression by acting as a competing endogenous RNA of miR-3064 to upregulate PIP4K2B expression. J Exp Clin Cancer Res. 2020;39(1):80. doi: 10.1186/s13046-020-01573-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Chi C, Mao M, Shen Z, Chen Y, Chen J, Hou W. HOXD-AS1 exerts oncogenic functions and promotes chemoresistance in cisplatin-resistant cervical cancer cells. Hum Gene Ther. 2018;29(12):1438–1448. doi: 10.1089/hum.2017.256. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 143.Du C, Wang Y, Zhang Y, Zhang J, Zhang L, Li J. LncRNA DLX6-AS1 contributes to epithelial-mesenchymal transition and cisplatin resistance in triple-negative breast cancer via modulating Mir-199b-5p/Paxillin axis. Cell Transplant. 2020;29:2138922687. doi: 10.1177/0963689720929983. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 144.Cheng Y, Shen X, Zheng M, Zou G, Shen Y. Knockdown Of lncRNA NCK-AS1 regulates cisplatin resistance through modulating miR-137 In osteosarcoma cells. Onco Targets Ther. 2019;12:11057–11068. doi: 10.2147/OTT.S228199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Yang Q, Li K, Huang X, Zhao C, Mei Y, Li X, Jiao L, Yang H. lncRNA SLC7A11-AS1 promotes chemoresistance by blocking SCF(β-TRCP)-mediated degradation of NRF2 in pancreatic cancer. Mol Ther Nucleic Acids. 2020;19:974–985. doi: 10.1016/j.omtn.2019.11.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Hua YQ, Zhu YD, Xie GQ, Zhang K, Sheng J, Zhu ZF, Ning ZY, Chen H, Chen Z, Meng ZQ, et al. Long non-coding SBF2-AS1 acting as a competing endogenous RNA to sponge microRNA-142-3p to participate in gemcitabine resistance in pancreatic cancer via upregulating TWF1. Aging (Albany NY) 2019;11(20):8860–8878. doi: 10.18632/aging.102307. [DOI] [PubMed] [Google Scholar]
- 147.Bai T, Liu Y, Li B. LncRNA LOXL1-AS1/miR-let-7a-5p/EGFR-related pathway regulates the doxorubicin resistance of prostate cancer DU-145 cells. IUBMB Life. 2019;71(10):1537–1551. doi: 10.1002/iub.2075. [DOI] [PubMed] [Google Scholar]
- 148.Zhang CL, Zhu KP, Ma XL. Antisense lncRNA FOXC2-AS1 promotes doxorubicin resistance in osteosarcoma by increasing the expression of FOXC2. Cancer Lett. 2017;396:66–75. doi: 10.1016/j.canlet.2017.03.018. [DOI] [PubMed] [Google Scholar]
- 149.Han M, Gu Y, Lu P, Li J, Cao H, Li X, Qian X, Yu C, Yang Y, Yang X, et al. Exosome-mediated lncRNA AFAP1-AS1 promotes trastuzumab resistance through binding with AUF1 and activating ERBB2 translation. Mol Cancer. 2020;19(1):26. doi: 10.1186/s12943-020-1145-5. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 150.Zhang Z, Yin J, Lu C, Wei Y, Zeng A, You Y. Exosomal transfer of long non-coding RNA SBF2-AS1 enhances chemoresistance to temozolomide in glioblastoma. J Exp Clin Cancer Res. 2019;38(1):166. doi: 10.1186/s13046-019-1139-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Yan Y, Xu Z, Chen X, Wang X, Zeng S, Zhao Z, Qian L, Li Z, Wei J, Huo L, et al. Novel Function of lncRNA ADAMTS9-AS2 in Promoting Temozolomide Resistance in Glioblastoma via Upregulating the FUS/MDM2 Ubiquitination Axis. Front Cell Dev Biol. 2019;7:217. doi: 10.3389/fcell.2019.00217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Huang H, Chen J, Ding CM, Jin X, Jia ZM, Peng J. LncRNA NR2F1-AS1 regulates hepatocellular carcinoma oxaliplatin resistance by targeting ABCC1 via miR-363. J Cell Mol Med. 2018;22(6):3238–3245. doi: 10.1111/jcmm.13605. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 153.Ma Y, Bu D, Long J, Chai W, Dong J. LncRNA DSCAM-AS1 acts as a sponge of miR-137 to enhance Tamoxifen resistance in breast cancer. J Cell Physiol. 2019;234(3):2880–2894. doi: 10.1002/jcp.27105. [DOI] [PubMed] [Google Scholar]
- 154.Shi YF, Lu H, Wang HB. Downregulated lncRNA ADAMTS9-AS2 in breast cancer enhances tamoxifen resistance by activating microRNA-130a-5p. Eur Rev Med Pharmacol Sci. 2019;23(4):1563–1573. doi: 10.26355/eurrev_201902_17115. [DOI] [PubMed] [Google Scholar]
- 155.Leng W, Liu Q, Zhang S, Sun D, Guo Y. LncRNA AFAP1-AS1 modulates the sensitivity of paclitaxel-resistant prostate cancer cells to paclitaxel via miR-195–5p/FKBP1A axis. Cancer Biol Ther. 2020;21:1072–1080. doi: 10.1080/15384047.2020.1829266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Zhang S, Jiang H, Xu Z, Jiang Y, She Y, Huang X, Feng S, Chen W, Chen S, Chen Y, et al. The resistance of esophageal cancer cells to paclitaxel can be reduced by the knockdown of long noncoding RNA DDX11-AS1 through TAF1/TOP2A inhibition. Am J Cancer Res. 2019;9(10):2233–2248. [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.