

ABSTRACT
Animals subjected to dietary restriction (DR) have reduced body size, low fecundity, slower development, lower fat content and longer life span. We identified lamin as a regulator of multiple dietary restriction phenotypes. Downregulation of lmn-1, the single Caenorhabditis elegans lamin gene, increased animal size and fat content specifically in DR animals. The LMN-1 protein acts in the mTOR pathway, upstream of RAPTOR and S6 kinase β1 (S6K), a key component of and target of the mechanistic target of rapamycin (mTOR) complex 1 (mTORC1), respectively. DR excludes the mTORC1 activator RAGC-1 from the nucleus. Downregulation of lmn-1 restores RAGC-1 to the nucleus, a necessary step for the activation of the mTOR pathway. These findings further link lamin to metabolic regulation.
KEY WORDS: Caenorhabditis elegans, Lamin, Dietary restriction, mTOR
Summary: Downregulation of the single C. elegans lamin gene increases animal size and fat content specifically in dietary restricted animals. The lamin protein acts in the mTOR pathway to regulate these phenotypes.
INTRODUCTION
Dietary restriction (DR) is a metabolic intervention with a conserved response among many organisms. It is one of the most effective methods to prolong lifespan as well as health span in many animal models, with beneficial effects in humans. It reduces many age-related pathologies including diabetes and cardiovascular diseases (Colman et al., 2009; Mair and Dillin, 2008).
The mechanistic target of rapamycin (mTOR) is a nutrient sensor that functions as a central regulator of metabolism and physiology. Inhibition of mTOR prolongs the lifespan and improves the healthspan of many model organisms (Johnson et al., 2013; Mannick et al., 2014). Suppression of mTOR is one of the underpinning mechanisms of the beneficiary effects of DR (Papadopoli et al., 2019). By contrast, mTOR signaling is dysregulated in cells harboring disease causing mutations in the human lamin A (LMNA) gene (Chiarini et al., 2019).
Lamins are type V nuclear intermediate filaments that are conserved in metazoan evolution, and are key components of the nuclear lamina. Mutations in the human LMNA gene cause numerous diseases, including metabolic diseases, accelerated aging disorders and muscle diseases (Worman and Bonne, 2007). Caenorhabditis elegans, a free-living nematode with a single lamin gene (lmn-1), is extensively used in the research of lifespan regulating pathways. In addition, it is used in the research of the structure and function of the nuclear lamina (Bank et al., 2011; Link et al., 2018; Turgay et al., 2017; Wiesel et al., 2008).
We identified lamin as the regulator of multiple DR phenotypes. Knockdown of lmn-1 increased fat content and animal size in DR animals, while simultaneous knockdown of RAPTOR [a key component of mTOR complex 1 (mTORC1)] abolished this size increase. Furthermore, lmn-1 knockdown had no impact on size in DR animals lacking S6 kinase β1 (S6K), one of the main targets of the mTOR pathway. Finally, downregulation of lmn-1 enabled the nuclear entry of Ras-related GTP binding C (RAGC-1), which is essential for activation of mTORC1 signaling.
RESULTS
lmn-1 regulates size in DR animals
C. elegans feeding depends on rhythmic contractions (pumping) of the pharynx (Avery and Horvitz, 1990). eat-2 (ad1116)-mutated C. elegans (henceforth denoted eat-2) have a reduced pumping rate, resulting in slower food uptake. This results in longer life span, smaller body length, lower fat content and smaller brood size (Bar et al., 2016; Walker et al., 2005), and these animals thus serve as a suitable model for DR research (Walker et al., 2005). To gain insight to the roles of lamin in aging and metabolism, young eat-2 adults were fed with bacteria expressing lmn-1 double stranded RNA (RNAi), that effectively knocked down LMN-1 (Fig. S1A,B). Surprisingly, downregulation of lmn-1 significantly increased the body size (length and width) of eat-2 animals, but not of controls (Fig. 1A,B, P<0.0001). Since the number of somatic cells in C. elegans is fixed, eat-2 animals are smaller due to smaller cell size (Mörck and Pilon, 2007; Walker et al., 2005). To validate that lamin knockdown rescues cell size, we measured the length of cells expressing red fluorescent protein (RFP) fused to the MYO-2 muscle-specific protein. Downregulation of lmn-1 increased cell length in eat-2 animals (Fig. 1C,D, P<0.0001). This increase was proportional to the increase seen in the entire animal (Fig. 1A,B). These results suggest that cell size changes are responsible for the increase in animal size. Of note, downregulation of lmn-1 did not increase the food intake, as the pumping rate remained unaffected (Fig. 1E).
Fig. 1.

Size reduction in DR worms requires lmn-1 activity. (A) Stereomicroscope images showing eat-2 (ad1116) worms fed for 72 h with lmn-1 (RNAi) or EV Scale bar: 100 μm. (B) Average length of animals from A. n(N2)=93 and n(eat-2)=89 animals. ***P<0.0001 (two-tailed unpaired t-test). (C) Representative microscope images of eat-2 (ad1116) worms expressing myo-2::RFP. Worms were fed with lmn-1 (RNAi) or EV for 72 h. Yellow outline shows cell boundaries. (D) Relative length of muscle cells in N2 and eat-2 worms, both expressing myo-2::RFP, that were fed with lmn-1 (RNAi) or EV for 72 h. n(N2)=24, n(eat-2)=26 worms and totals of 183 and 251 cells, respectively, were used for the analysis. ***P=7.37×10−15 (two-tailed unpaired t-test). (E) Average pumping rate of eat-2 (ad1116) animals fed with lmn-1 (RNAi) or EV for 72 h. n=51. Error bars in all graphs represent mean±s.e.m.
Lamin regulates the reduced fat levels of DR animals
Animals subjected to DR accumulate less fat. This is due to a genetically controlled process, where nutrients are allocated elsewhere (Bar et al., 2016; Palgunow et al., 2012). To test whether lmn-1 regulates fat accumulation in C. elegans, we measured fat levels using Oil Red O staining. Downregulation of lmn-1 significantly increased fat levels in eat-2 but not in control animals (Fig. 2A,B, P<0.0001). These results indicate a role for lmn-1 in regulating fat levels.
Fig. 2.

lmn-1 regulates fat accumulation and sbp-1 transcription in DR animals. (A) Representative stereomicroscope images of Oil Red O staining of control (N2) and eat-2 (ad1116) worms fed with lmn-1 (RNAi) or EV for 72 h. (B) Average levels of relative intensity of Oil Red O staining in eat-2 (ad1116) and in control (N2) animals fed with lmn-1 (RNAi) or EV for 72 h. n(N2)=99, n(eat-2)=75. (C) Representative brightfield (BF) fluorescence microscopy images of control (N2) and eat-2 (ad1116) animals expressing GFP driven by the sbp-1 promoter (sbp-1p::GFP) and fed with either EV or lmn-1 (RNAi). (D) Quantification of C. n(N2)=52, n(eat-2)=60. Error bars in all graphs represent mean±s.e.m. *P<0.01; ***P<0.0001 (two-tailed unpaired t-test). Scale bars: 100 µm.
Lamin regulates the transcription of sterol binding protein 1
Sterol regulatory element-binding protein (SREBP) is a transcription factor required for fatty acid biosynthesis (Eberlé et al., 2004). Sterol-binding protein 1 (SBP-1) is the C. elegans homolog and a positive regulator of lipid storage (Sato, 2010). To test whether lamin regulates sbp-1 transcription, we downregulated lamin in control and eat-2 animals expressing GFP fused to the sbp-1 promoter (sbp-1p::GFP). While sbp-1 transcription levels are relatively low in eat-2 animals (Fig. 2C,D, P<0.0001), downregulation of lmn-1 re-elevated sbp-1 levels, most notably in the gut (Fig. 2C,D, P<0.01). We note that lamin is known to regulate SREBP subcellular localization (Duband-Goulet et al., 2011) and inhibition of mTOR causes SREBP to accumulate at the nuclear envelope, where it is inactive (Peterson et al., 2011). Thus, it is likely that the transcriptional regulation of sbp-1 only partially accounts for the excess fat accumulation in these DR animals.
lmn-1 regulates animal size upstream of the TORC1 complex
atx-2 and gdi-1 are the functional homologs of the tuberous sclerosis complex (TSC) components in C. elegans, which decrease cell size via the inhibition of mTOR pathway (Bar et al., 2016). As both lmn-1 and atx-2 RNAi reverse the DR-induced size reduction (Figs 1A and 3A), we mapped the relationship between these genes. When eat-2 animals were subjected to RNAi against both genes, no additive increase in size was observed (Fig. 3A). This suggests that atx-2 and lmn-1 are part of the same pathway, or at least have the same downstream targets, with respect to body size regulation.
Fig. 3.
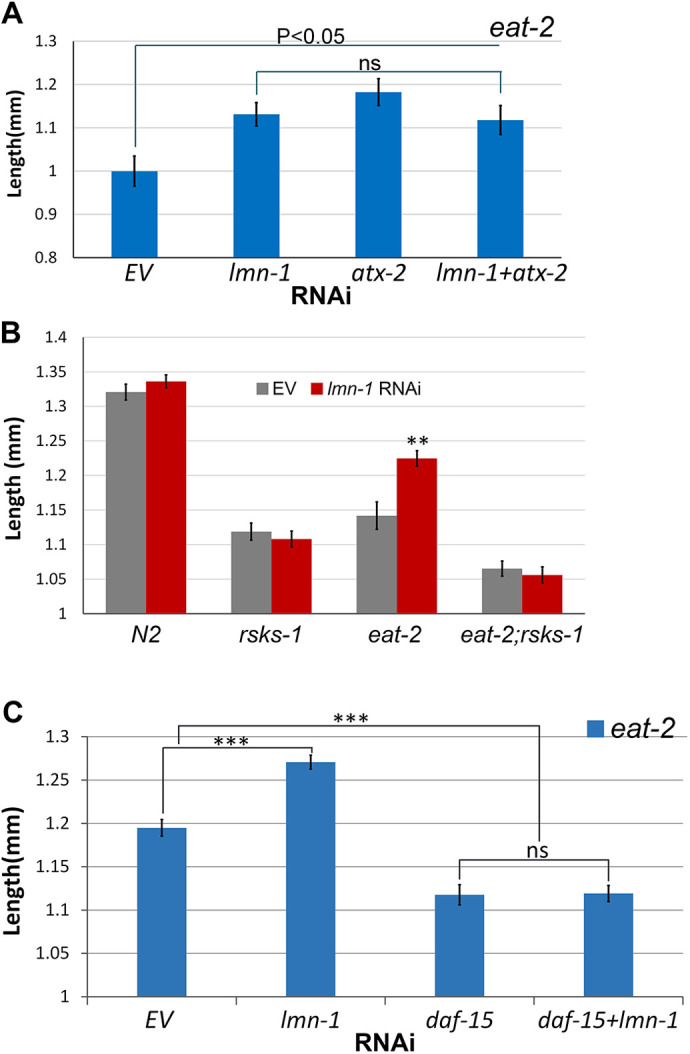
lmn-1 regulates animal size upstream of the mTOR pathway. (A) Average length of eat-2 young adult worms fed with RNAi for either lmn-1, atx-2, both lmn-1 and atx-2 or EV for 72 h. (B) Average length of N2, eat-2, rsk-1 or eat-2;rsk-1 young adult worms fed with RNAi for lmn-1 or EV for 72 h. n(N2)=66, n(rsks-1)=67, n(eat-2)=59, n(eat-2;rsks-1)=54 (C) Average length of eat-2 young adult worms fed with RNAi for either lmn-1, daf-15, both lmn-1 and daf-15 or EV. n(eat-2)=186. Error bars in all graphs represent mean±s.e.m. **P<0.001; ***P<0.0001; ns, not significant (two-tailed unpaired t-test).
ATX-2 regulates the mTOR pathway (Bar et al., 2016; Carmo-Silva et al., 2017; Lastres-Becker et al., 2016), which is dysregulated in some laminopathies. We further investigated the role of lamin in the mTOR pathway. Ribosomal protein S6 kinase β1 (S6K), is a downstream target of mTORC1 (Sfakianos et al., 2018). Its main activity is the phosphorylation of the S6 ribosomal protein, leading to an increase in protein synthesis and cell proliferation. Deletion of rsks-1, the homolog of S6K in C. elegans, resulted in a major decrease in the size of both eat-2 and control worms (Fig. 3B, P<0.0001 and P<0.01, respectively). This size decrease could not be rescued by downregulation of lmn-1, indicating that lamin regulates size upstream of S6K (Fig. 3B). To confirm that the size increase due to lamin knockdown depends directly on mTOR itself, we downregulated lmn-1 together with raptor, a key member of the mTOR complex-1 (mTORC1) (Carriere et al., 2011). Downregulation of daf-15, the C. elegans homolog of RAPTOR, decreased the size of eat-2 animals (Fig. 3C, P<0.0001) and eliminated the lmn-1 (RNAi) size increase (Fig. 3C). Based on these results, we conclude that lamin acts upstream of the mTOR complex-1 to regulate size.
Under nutrient deprivation, the GDI-1:ATX-2 axis acts upstream of Ras homolog enriched in brain (RHEB), to mediated a switch in binding from RHEB-GTP to RHEB-GDP. While RHEB-GTP promotes activation of mTORC1 complex, RHEB-GDP blocks its activity (Bar et al., 2016; Laplante and Sabatini, 2012). This leads to an opposite effect on fed (N2) animals, which are decreased in size due to elimination of RHEB-GTP and reduced activation of mTOR, and increased size in DR animals, due to elimination of RHEB-GDP and reduced inhibition of mTOR (Bar et al., 2016). Indeed, downregulation of rheb-1 in control animals decreased their size (Fig. 4A, P<0.0001); however, downregulation of lmn-1 reversed this effect (Fig. 4A, P<0.0001). Under DR conditions, downregulation of rheb-1 increases their size (Fig. 4B, P<0.0001), while downregulation of both lmn-1 and rheb-1 had no additive effect (Fig. 4B). We conclude that lamin might function downstream of RHEB in regulating the mTOR pathway.
Fig. 4.

Mapping lmn-1 to the mTOR pathway. (A) Average length of N2 young adult worms fed with RNAi for either lmn-1, rheb-1, both lmn-1 and rheb-1 or EV. n(N2)=494. (B) Average length of eat-2 young adult worms fed with RNAi for either lmn-1, rheb-1, both lmn-1 and rheb-1 or EV. n(eat-2)=680. (C) Average length of N2, eat-2, raga-1 or eat-2;raga-1 young adult worms fed with lmn-1 (RNAi) or EV. n(N2)=70, n(raga-1)=92, n(eat-2)=73, n(eat-2;raga-1)=45. (D) Average length of N2, eat-2, daf-2 or eat-2;daf-2 young adult worms fed with lmn-1 (RNAi) or EV. n(N2)=96, n(eat-2)=92, n(daf-2)=72, n(eat-2;daf-2)=51. Experimental conditions for daf-2 and eat-2;daf-2 differed from those for N2 and eat-2 (dashed line; see Materials and Methods for details). (E) Average length of N2, eat-2, daf-16, akt-1, eat-2;daf-16 or eat-2;akt-1 young adult worms fed with lmn-1 (RNAi) or EV. n(N2)=80, n(daf-16)=95, n(akt-1)=115, n(eat-2)=72, n(eat-2;daf-16)=82, n(eat-2;akt-1)=97. (F) Average length of N2, eat-2 or aak-2 young adult worms fed with lmn-1 (RNAi) or EV. n(N2)=50, n(aak-2)=43, n(eat-2)=42, n(eat-2;aak-2)=57. Error bars in all graphs represent mean±s.e.m. *P<0.01; **P<0.001; ***P<0.0001; ns, not significant (two-tailed unpaired t-test).
A key regulator of the mTOR pathway is the Ras-related GTP-binding protein (RAG) complex. Depending on amino acid availability, the subunits RAGA or RAGB bound to GTP [RAG(A/B)-GTP] and RAGC or RAGD bound to GDP [RAG(C/D)-GDP] form a heterodimer that recruits and anchor mTOR to the lysosome (Sancak et al., 2008; Wu et al., 2016), where it can be activated by its upstream regulators. Deletion of raga-1, the C. elegans homolog of RAG(A/B), decreases the size of control animals, and DR conditions further decrease their size (Fig. 4C, P<0.001). Downregulation of lmn-1 in DR worms lacking raga-1 resulted in additional size reduction (Fig. 4C), suggesting that raga-1 is essential for lmn-1 activity in the context of size regulation. The additional size decrease may suggest that lmn-1 has mTOR-independent roles that may affect size.
lmn-1 size regulation is not dependent on the insulin growth factor signaling pathway
Growth factors stimulate cellular growth and proliferation partially via the mTOR pathway. Activated by the growth factors, insulin growth factor receptors (IGFRs) mediate a phosphorylation cascade that eventually promotes mTOR activity (Inoki et al., 2003; Laplante and Sabatini, 2012). Daf-2 is the sole member of the IGFR family in C. elegans. Downregulation of lmn-1 in eat-2 worms that were also mutated in daf-2 resulted in an increase to their size (Fig. 4D, P<0.0001), suggesting that lmn-1 is not dependent on daf-2 for size regulation of eat-2 animals. AKT-1 is triggered by IGF signaling (Laplante and Sabatini, 2012). Once activated, AKT-1 activates the mTOR pathway by inhibiting phosphorylations of the TSC (Inoki et al., 2002; Manning et al., 2002). As expected, deletion of akt-1 resulted in size decrease both in control and eat-2 animals (Fig. 4E, P<0.0001 and P<0.01, respectively). However, downregulation of lmn-1 increased animal size both in control and eat-2 animals (Fig. 4E, P<0.0001), indicating that akt-1 is not required for lmn-1 size regulation.
Adenosine monophosphate-activated protein kinase (AMPK), is a master regulator of cellular energy levels (Inoki et al., 2003). Downregulation of lmn-1 in eat-2 worms also lacking aak-2, the catalytic subunit of AMPK, increased their size (Fig. 4F, P<0.001). DAF-16, a target of AMPK and DAF-2, is a transcription factor that translocates to the nucleus upon multipole stress signals, and inhibits mTORC1 (Robida-Stubbs et al., 2012). Deletion of daf-16 resulted in a size increase of eat-2 animals (Fig. 4E, P<0.001). Downregulation of lmn-1 in eat-2 animals lacking daf-16 resulted in an additional size increase (Fig. 4E, P<0.01). This suggests that daf-16 and lmn-1 regulate the mTOR pathway, at least partially independently.
DR delays age-dependent nuclear envelope deformation
The nuclear lamina acquires structural deformation, including lobulations, membranes invagination and aggregations, in an age-dependent manner (Haithcock et al., 2005). To test the effect of dietary restriction on the nuclear lamina, we imaged animals expressing LMN-1::GFP driven by the lmn-1 promoter at day 2, 4 and 6 of adulthood, and classified the nuclear shape into three groups based on deformation severity (Fig. 5A–G). At day 2 of adulthood, control animals began to show a mild aging phenotype, as more than 30% of the nuclei demonstrated mild abnormalities, such as membrane folding and some lamin foci (class II) (Fig. 5A,G). At day 4 of adulthood this phenotype was aggravated, as animals showed increased lobulations and lamin aggregations (45% class II and III; Fig. 5B,G). This was further aggravated at day 6 as more control animals accumulated nuclear mis-shapes, such as stretched nuclei, nuclear lobulations and increased membrane folding. Overall, more than 50% of the nuclei shifted toward class II and III (Fig. 5C,G). By contrast, DR animals showed a lag in nuclear envelope aging phenotypes. At day 2, most of the nuclei (∼93%) did not exhibit any nuclear deformation as the nuclei were smooth and round (class I), with only a small fraction showing nuclear abnormalities (∼7%) (Fig. 5D,G). Moreover, at day 4 and even 6 of adulthood, most of the nuclei still kept their non-deformed shape (86% and 82%, respectively, in class I; Fig. 5E–G). These changes were accompanied by a mild decrease in total LMN-1 protein levels in control and DR animals with age (Fig. 5H), consistent with previous reports (Haithcock et al., 2005). Based on these results, we conclude that DR conditions delay age-dependent nuclear deformation, as DR nuclei retain their smooth shape and structure even at advanced age. Some DR phenotypes are dependent on AMPK and its downstream targets (Fig. S2; Uno and Nishida, 2016). We tested whether the protective effects of DR on the nuclear morphology depend on this pathway as well. Downregulation of aak-2 resulted in a faster accumulation of nuclear abnormalities (Fig. S2), suggesting this is a regulated process.
Fig. 5.

Dietary restrictions delay age-dependent nuclear deformation. (A–F) Representative microscope images of control (N2) (A–C) and eat-2 (D–F) worms expressing LMN-1::GFP at days 2 (A,D), 4 (B,E) and 6 (C,F) of adulthood. (G) Relative distribution of the three different classes grading (see Materials and Methods) the nuclear morphology changes in N2 and eat-2 animals at days 2, 4 and 6 of adulthood. n(D2)=255 nuclei, n(D4)=228 nuclei, n(D6)=238 nuclei. Note: Error bars in all graphs represent mean±s.e.m. ***P<0.0001 (Fisher exact probability test). (H) Western blot gel image of LMN-1 protein in N2 and eat-2 at days 2, 4 or 6 of adulthood.
Dysregulation of mTOR drives nuclear envelope deformation
Animals with life-extending mutations can, under specific conditions, show a delayed accumulation of nuclear envelope abnormalities (Haithcock et al., 2005; Pérez-Jiménez et al., 2014; Fig. 6A–D,I). By contrast, laminopatic mutations can drive the premature accumulation of such defects (van Tienen et al., 2019). When atx-2 was knocked down in young adult animals, we saw a small, but statistically significant increase, in nuclei showing abnormalities. To aggravate this phenotype and to test the role of atx-2 in nuclear shape regulation, we subjected animals expressing LMN-1::GFP, driven by the lmn-1 promoter, to RNAi downregulation of atx-2 immediately post-hatching. Changes in nuclear morphology were documented every other day, starting at day 1 of adulthood and until day 7. Nuclear morphology was classified into the three class groups as described in the previous section. Already by day 1 of adulthood, nuclei of atx-2 (RNAi) adult animals showed increased lobulations and lamin aggregations at the nuclear envelope (Fig. 6E,I). These age-related phenotypes were much more robust at day 3, as more nuclei shifted to class II and class III (Fig. 6F,I). By day 5, only 7% of the nuclei were smooth (class I) (Fig. 6G,I). By day 7 of adulthood, 75% of the nuclei were fragmented, stretched, and showed invagination of the membrane and severe lobulation (class III), while only a small fraction were in class I (Fig. 6H,I). In line with these findings, accelerated breakdown of the nuclear envelope following atx-2 downregulation was observed in transgenic worms expressing the nuclear envelope protein emr-1 fused to GFP (Fig. S3). We concluded that the mTOR pathway regulates lamin distribution, and that downregulation of atx-2 results in nuclear envelope phenotypes that are similar to those seen in older animals (Haithcock et al., 2005).
Fig. 6.

ATX-2 is required to maintain nuclear structure. (A–H) Representative microscope images of worms expressing LMN-1::GFP fed with either EV (A–D) or atx-2(RNAi) post-hatching (E–H), at day 1 (A,E), day 3 (B,F), day 5 (C,G) and day 7 (D,H) of adulthood. (I) Relative distribution of the three different classes grading the nuclear morphology changes in LMN-1::GFP expressing worms fed with atx-2 (RNAi) or EV at days 1, 3, 5 and 7 of adulthood. n(D1)=228 nuclei, n(D3)=267 nuclei, n(D5)=283 nuclei, n(D7)=262 nuclei. Error bars in all graphs represent mean±s.e.m. ***P<0.0001 (Fisher exact probability test).
lmn-1 regulates RAGC-1 localization to the nucleus
To anchor mTORC1 to the lysosome, RAGC has to enter the nucleus as RAGC-GTP. In the nucleus, RAGC acquires its active RAGC-GDP form and exits back to the cytoplasm (Wu et al., 2016) where it anchors mTORC1 to the lysosome. To follow the expression and localization of RAGC-1, the C. elegans homolog of RAG(C/D), we used CRISPR/Cas9 to fuse a FLAG tag to the endogenous RAGC-1 protein (Fig. 7A). While, in control animals, RAGC-1 was expressed both in the cytoplasm and the nucleus (Fig. 7B), in eat-2 animals, RAGC-1 was mostly excluded from the nucleus [Fig. 7C, empty vector (EV) panel]. We compared the ratio of cytoplasmic RAGC-1 to nuclear RAGC-1 (Fig. 7D). In control animals, as well as control animals subjected to lmn-1 RNAi, this ratio was close to one, indicating equal presence of RAGC-1 in the nuclear and in the cytoplasmic fractions. By contrast, in DR worms, this ratio was at least three times higher than in control animals, possibly indicating a limited shuttling of RAGC-1. If lamin regulates the mTOR pathway through RAGC-1 localization, we would expect downregulation of lmn-1 to restore it to the nucleus. Indeed, following lmn-1 (RNAi), RAGC-1 entered the nucleus, potentially allowing it to acquire its active form (Fig. 7C,D).
Fig. 7.

lmn-1 regulates the entrance of RAGC-1 to the nucleus. (A) Western blot of transgenic ragc-1p::RAGC-1::FLAG in N2 and eat-2 worms. (B,C) Representative confocal images of N2 (B) and eat-2 (C) animals expressing RAGC-1 fused to FLAG-tag (RAGC-1::FLAG) that were fed with lmn-1 (RNAi) or EV for 48 h post-hatching. Scale bar: 10 μm. (D) Relative distribution (cytoplasmic to nuclear) of RAGC-1 in N2 and eat-2 worms expressing RAGC-1 fused to a FLAG tag (tg). n(N2)=19, n(eat-2)=29 worms and totals of 50 and 65 cells, respectively, were used for the analysis. Error bars in all graphs represent mean±s.e.m. **P<0.001; ***P<0.0001 (two-tailed unpaired t-test).
DISCUSSION
Lamin regulates DR
Mutations in LMNA genes are associated with aging disorders as well as metabolic diseases. One example is Hutchinson–Gilford progeria syndrome (HGPS), where lamin mutations result in an accelerated aging disorder and pathological phenotype (Goldman et al., 2004; Gonzalo et al., 2017). A recent study in HGPS mouse models shows the potential of DR in improving life expectancy (Bárcena et al., 2018). Here, we showed that during DR in C. elegans, multiple DR-related phenotypes are regulated by lamin. Downregulation of lmn-1 in DR animals partially rescues their size and fat levels, while having minimal effect on control animals. Two other aspects of DR, fecundity and longevity, could not be properly analyzed in a similar way, as lamin is essential for development and its downregulation also significantly shortens lifespan of both control (N2) and DR animals (Bar et al., 2016). DR is regulated by multiple partially overlapping pathways (Greer and Brunet, 2009). Some of the genes tested here have been shown to be involved in several DR regimes (Bar et al., 2016; Greer and Brunet, 2009). However, other genes and other DR regimes may result in different phenotypic outcomes.
Lamin acts through the mTOR pathway
Multiple studies have shown a crosstalk between lamin genes and the mTOR pathway (Chiarini et al., 2019; Lattanzi et al., 2014). The mTOR pathway is affected by some LMNA mutations, and mTOR inhibitors are successfully used to treat multiple laminopatic mutations in cell lines, in animal models and recently in clinical trials (Pellegrini et al., 2015; Ramos et al., 2012). We explored the regulatory role of lmn-1 in DR using an epistasis assay. Three genes were found to be required for lmn-1 (RNAi) size rescue: rsks-1, encoding S6K, a target of the mTOR pathway; daf-15, encoding the RAPTOR homolog, a core protein of the mTORC1 complex; and raga-1, the homolog of RAG(A/B), an mTOR regulator. We conclude that mTORC1 is essential for lmn-1 size-dependent regulation.
ATX-2 regulates nuclear morphology
As cells age, their nuclei accumulate abnormalities (Haithcock et al., 2005). DR animals accumulated abnormalities slower than control animals (Fig. 5A–G). Activation of the mTOR pathway, mediated by downregulation of the negative regulator atx-2, resulted in a significant increase in nuclear abnormalities (Fig. 6). In mammalian cells, inhibition of mTORC1 affects the nuclear shape through regulation of lipin 1 nuclear localization (Peterson et al., 2011). One possible mechanism of nuclear shape regulation is via post-translational modifications of lamins. Phosphorylation of lamins determines their structural properties and affects signaling (Torvaldson et al., 2015). Insulin, a known mTOR activator, drives lamin phosphorylation during interphase (Friedman and Ken, 1988). It will be interesting to test whether post-translational modifications of lamins and other nuclear envelope proteins might regulate the mTOR pathway under DR.
RAGC-1 exclusion from the nucleus in DR requires an intact nuclear lamina
Lamin directly interacts with the linker of nucleoskeleton and cytoskeleton (LINC) complex and the nuclear pore complex (NPC) (Bone et al., 2014; Guo and Zheng, 2015). The NPC regulates nuclear transport. Several laminopathic mutations disrupt nucleocytoplasmic translocation (Busch et al., 2009; Han et al., 2019; Kelley et al., 2011). Some laminopathies are characterized by failure to exclude proteins from the nucleus. Yes-associated protein (YAP, also known as YAP1) is a sensor and mediator of mechanical cues. Mutations resulting in LMNA-related congenital muscular dystrophy cause the NPC to fail to exclude YAP from the nucleus, contributing to the disease pathology (Owens et al., 2020). A morphological hallmark of the different laminopathies is a deformed nuclear envelope (van Tienen et al., 2019). One plausible mechanism for the impaired nuclear transport in laminopathies is that the altered nuclear shape causes changes of the stretch forces applied on the NPC, a known regulator of the flux (Donnaloja et al., 2019). Nuclear deformations are also found in aged nuclei (Haithcock et al., 2005; Pérez-Jiménez et al., 2014), along with compromised nucleocytoplasmic transportation (D'Angelo et al., 2009). Recently, it was shown that nuclei of aged C. elegans are more susceptible to damaging effects from outside stretch forces (Zuela-Sopilniak et al., 2020). This is likely the result of altered nuclear stiffness and lower elasticity, suggesting that aged nuclei are under constant self-induced stretch. A similar phenomena is observed when the LINC complex is impaired (Zuela-Sopilniak et al., 2020). When nuclei are exposed to outside forces that deform the nuclear envelope morphology, NPC are redistributed and stretched, increasing transcription factor nuclear import (Elosegui-Artola et al., 2017; Hoffman et al., 2020). In mammalian cells, nuclear eccentricity is regulated by nutrients and mTORC1. In C. elegans, DR delays age-related nuclear morphology phenotypes and excludes RAGC-1 from the nucleus, thus inhibiting mTOR. On the other hand, compromising the nuclear envelope by downregulation of lmn-1 restores the level of RAGC-1 in the nucleus, activates mTOR and increases animal size and fat content. This model also explains the increase seen in sbp-1 transcription, which is known to be regulated by mTOR (Fig. 8) (Bar et al., 2016; Li et al., 2011; Peterson et al., 2011).
Fig. 8.

Schematic representation of the NPC model. Diagram showing a possible explanation for these data. The deformed nuclear envelope in fed worms may affect the NPC and allow RAGC-GTP to enter into the nucleus, be exchanged to RAGC-GDP and facilitate the activation of the mTOR pathway. In DR worms, the intact nuclear envelope delays this process. For further details, see text.
Downregulation of lamin likely affects multiple genes and proteins, as the nuclear envelope preferentially interacts with both DNA lamina-associated domains (LADs) and multiple proteins, including transcription factors and chromatin remodelers. We found that in DR animals, the size effect is completely dependent on the RSKS-1, DAF-15 and RAGC-1. Several non-mutually exclusive mechanisms can explain these data. One interpretation of these findings is that lamin, via the NPC and nucleocytoplasmic transport, regulates the mTOR pathway. Under DR, and potentially other types of stress, the nuclear lamina is required to make the NPC less permissive. Stress-dependent post-translational modifications may drive these changes. The less-permissive NPC would prevent passive nuclear entry of RAGC-1, and potentially other proteins. A similar mechanism was demonstrated for biguanides, which regulate the mTOR pathway by preventing RAGC from passing through the NPC (Wu et al., 2016). These findings may have implications on our understanding of how the nuclear lamina regulates gene expression in other situations, including mechanotransduction. Alternatively, disruption of the nuclear lamina is known to affect the localization of multiple nuclear membrane proteins and transcription factors, as well as impact chromatin organization (Heessen and Fornerod, 2007; Markiewicz et al., 2006; Ranade et al., 2019). Downregulation of lamin may thus, directly or indirectly, affect the mTOR pathway through unknown proteins or DNA regulation. Future studies will test whether laminopathic mutations affect the permissibility of the NPC, and whether it is linked to their pathologies.
MATERIALS AND METHODS
C. elegans strains
Strain maintenance and manipulations were performed under standard conditions as previously described (Brenner, 1974). All experiments were performed at 23°C unless described otherwise. The following strains were used: N2 (control); DA116, eat-2(ad1116) II; CF1038, daf-16(mu86) I; YG2227, [eat-2(ad1116) II; daf-16(mu86) I]; RB754, aak-2(ok524) X; YG2223, [eat-2(ad1116) II; aak-2(ok524) X]; RB1206, rsks-1(ok1255) III; YG2229, [eat-2(ad1116) II; rsks-1(1255) III]; CF1041, daf-2(e1370); YG2606, [eat-2(ad1116) II; daf-2(e1370)]; RB759, akt-1(ok525)V; YG2605, [eat-2(ad1116) II; akt-1(ok525)V]; VC222, raga-1(ok386) II; [eat-2(ad1116) II; VC222, raga-1(ok386) II]; [sbp-1p::GFP]; YG2604, {eat-2(ad1116) II; [sbp-1p::GFP]}; YG2607, [ragc-1p::RAGC-1::FLAG]; YG2608, {eat-2(ad1116); YG2607[ragc-1p::RAGC-1::FLAG]}; PD4810, {lmn-1:GFP, ccIs4810 [pJKL380.4 Plmn-1::lmn-1::gfp::lmn-1 3′UTR +myo-2p::MYO-2::RFP]}; LW699, [lmn-1p::lmn-1::gfp::unc-54 3′UTR+ unc-119(+)]. All strains were obtained from the C. elegans Genome Center (CGC), generated using microinjection or obtained using genetic crossing.
Generating the ragc-1p::RAGC-1::FLAG strain using CRISPR-Cas9
ragc-1p::RAGC-1::FLAG, was generated by microinjection using CRISPR-Cas9 as described previously (Friedland et al., 2013), and verified by sequencing and western blotting. A 3x FLAG tag was inserted to the ragc-1 sequence after the ATG start codon using the CRISPR RNA (crRNA) 5′-AATCATCAAAATCCTCGTCA-3′ and the single-stranded oligodeoxynucleotide (ssODN) 5′-taaaccttttcaatttcagaatgGACTACAAGGACCACGACGGAGACTACAAGGACCACGATATCGATTACAAGGACGACGACGACAAGgagtcggatcctgacgaggattttgatgattaccgct-3′, where the start codon is shown in lowercase bold letters, the crRNA site is shown in bold underlined letters, the restriction site is shown in bold underlined italic letters and the 3×FLAG sequence is shown in uppercase. Bases modified to cancel an NGG site are indicated 5′ of the crRNA site.
RNAi experiments
RNAi feeding experiments were performed as described previously (Ahringer, 2006). In brief, nematode growth medium (NGM) plates containing 25 μg/ml ampicillin and 1 mM isopropyl β-d-thiogalactoside (IPTG) were seeded with the appropriate bacteria taken from either the Ahringer library (Kamath and Ahringer, 2003) or the Vidal library (Rual et al., 2004). Controls were placed on feeding plates with an empty L4440 vector (EV). Worms were placed on plates at the appropriate developmental stage and analyzed for various phenotypes. When antibodies or GFP fusions were available, the downregulation efficiency was verified by analysis of the protein level. In other cases, the level of mRNAs of the perturbed gene was analyzed by quantitative PCR, or the animal phenotypes were analyzed by matching the published data.
Size experiments
Animals were synchronized by bleaching (Porta-de-la-Riva et al., 2012) or by allowing them to lay eggs on plates for 6 h. In RNAi experiments, except for daf-15 downregulation and daf-2 mutants, animals were allowed to develop on NGM plates for 48 h until the eat-2 worms reached larval stage 3. Animals were transferred to RNAi plates, and then to fresh RNAi plates every 48 h, to exclude progeny. Unless noted otherwise, size measurements were performed after 72 h of growth on RNAi plates. In the daf-15 downregulation assays, after bleaching, nematodes were placed on daf-15 RNAi plates for 48 h then moved to fresh daf-15 or daf-15+lmn-1 plates. Animals carrying the daf-2 (e1370) mutation were allowed to develop on NGM plates for 96 h at 16°C, until daf-2;eat-2 nematodes reached larval stage 3, then were transferred to fresh RNAi plates for 72 h at 23°C. Imaging was performed using Olympus MVX10 dissecting microscope with a Dino-Eye camera (AnMo Electronics) and the length of the animals from head to tail was measured using ImageJ; 10–50 worms were measured at each time point. These experiments were repeated at least three times. To measure muscle cell length, transgenic animals containing MYO-2 protein fused to RFP were mounted on 2% agarose pads with 2 mM levamisole. Images were acquired with an ORCA-R2 camera (Hamamatsu Photonics) mounted on an Axioplan 2 microscope with a 60× oil lens. Length was measured from vertex to vertex along the muscle cell using ImageJ. At least 20 worms were measured at each time point. P values were calculated using a two-tailed unpaired t-test.
Oil Red O staining
Oil Red O staining and quantification were performed as described previously (O'Rourke et al., 2009). Images were acquired using Nikon Eclipse E200 microscope with an 4× or 10× lens fitted with a Moticam 2300 color camera (Motic). Exposure levels were adjusted for clear staining without saturation, and maintained for all samples. Quantification was performed with the ImageJ package. P values were calculated using a two-sample t-test for unequal variances. Experiments were repeated three times.
sbp-1p::GFP analysis
Animals expressing GFP driven by the sbp-1 promoter of the sbp-1 gene, were synchronized to the fourth larval stage. Animals were placed on feeding plates with lmn-1 (RNAi) for 48 h, then transferred to new RNAi feeding plates for an additional 24 h. Controls were placed on feeding plates with an empty L4440 vector (EV). Each experiment included ∼30 worms. Images were acquired using an Olympus MVX10 microscope with a QImaging Photometric 5MP camera. Quantification of GFP fluorescence intensity was performed using the ImageJ package. P values were calculated using a two-sample t-test for unequal variances.
Immunofluorescence staining
Animals were synchronized by bleaching, and eggs were then laid on lmn-1 RNAi or EV (control) feeding plates. Worms were then allowed to develop for 48 h until eat-2 animals reached larval stage 4. Worms were washed from the plates using M9 and transferred to 1.5 ml tubes. Worms were washed three times with M9 medium (Stiernagle, 2006) and three times with 1× phosphate buffered saline (PBS) containing 0.1% Tween 20 (PBST) to remove excess bacteria. Worms were fixed with freshly made 2% formaldehyde in PBST followed by snap freezing with liquid nitrogen and 10 min incubation at room temperature (RT). Samples were washed three times with PBST and permeabilized using mild sonication (Sonics Vibra-Cell sonicator) with a model CV33 microtip at ∼25% amplification 2×8 s sonication periods. Samples were incubated for 25 min in X2 modified Ruvkun's witches brew (MRWB) (Bettinger et al., 1996; https://www.wormatlas.org/antibodystaining.htm; Yen et al., 2010) followed by three washes with PBST, and incubated for 12.5 min with 10 mM DL-dithiothreitol (DTT). Samples were washed four times with PBST then incubated for 1 h in PBST followed by 10 min treatment with 0.3% H2O2. Samples were then washed three times with PBST and blocked for 1 h with a blocking buffer containing 1% bovine serum albumin (BSA) in PBST. After blocking, samples were incubated overnight at RT, shaking, with a primary mouse anti-FLAG antibody (clone M2; monoclonal; Sigma, CAT # F1804) diluted in blocking buffer (1:500). After four washes for ∼2 h with PBST, samples were incubated with the secondary anti-mouse-Cy3 antibody (Abcam ab97035) diluted in a blocking buffer (1:100) for ∼2 h at RT. After four washes in PBST for 2 h, DNA was stained for 10 min at RT using DAPI (Sigma, Cat# 28718903) diluted in PBST (1:1000), followed by three 10 min washes in PBST. Samples were mounted on glass slides in a drop of mounting medium (Vectashield, CAT# H-1400) and sealed with nail polish. Staining quantification was performed using the ImageJ package. P values were calculated using a two-sample t-test for unequal variances.
Western blot analysis
eat-2 worms were synchronized by bleaching as described above. 120 worms were collected from each sample at the different data points (day 2, day 4 and day 6 of adulthood) for western blotting as previously described (Towbin et al., 1979). Briefly, nematode samples were washed three times in minimal salt buffer M9 and immersed in 100 µl M9 buffer. Nematodes were centrifuged at 375 g for 2 min. Supernatant was removed and the pellet was dissolved in 200 µl RIPA-PI-PMSF lysis buffer (10 mM Tris-HCl pH 8.0, 1 mM EDTA, 0.5 mM EGTA, 1% Triton X-100, 0.1% sodium deoxycholate, 0.1% SDS, 140 mM NaCl), with 1× protease inhibitor (cOmplete protease inhibitor cocktail by Sigma Aldrich) and 1 mM PMSF (Sigma P7626) were added before use. The samples underwent four freezing-thawing cycles in liquid nitrogen and sonication (Sonics Vibra-Cell sonicator with a model CV33 microtip at 34% amplification 4×10 s sonication periods). Samples underwent two freeze-thaw cycles and were centrifuged for 10,600 g for 5 min at 4°C. Supernatant was collected and transferred to a new Eppendorf tube for use. Total protein levels were measured using a bicinchoninic acid (BCA) assay (ThermoFisher Pierce BCA Protein Assay kit, OD measured with BioTek synergy 2 multi-mode multiplate reader). Equal amounts of protein were loaded and samples were run on a 9% SDS polyacrylamide gel, transferred to a nitrocellulose membrane and blotted using anti C. elegans anti-lamin antibody (serum 3932, bleed 6, dilution of 1:1000; a kind gift from Professor Yosef Gruenbaum, The Hebrew University of Jerusalem; Lee et al., 2000; Tzur et al., 2002). Chemiluminescence images were acquired (Vilber Fusion FX).
Real-time PCR analysis
To validate efficient knockdown of genes, DR worms were synchronized by bleaching as described above, and placed on RNAi plates for 48 h. Animals were collected, washed with M9 and total RNA was extracted using Direct-zol RNA Miniprep Plus kit (Zymo, R2071). RNA was converted into cDNA using a High-Capacity RNA-to-cDNA Kit (Applied Biosystems 4387406). Each gene was tested using two primer pairs (Table S1), in triplicate. Real-time PCR was performed in 384-well format on a QuantStudio 12K Flex (Applied Biosystems) using PerfeCTa SYBR Green FastMix PCR Reagent (Quantabio). Similar results were observed across the pairs, and average of the results was used. act-1 and pmp-3 served as reference genes (Hoogewijs et al., 2008; Zhang et al., 2012).
Abnormal nuclear morphology classification
Transgenic control (N2) and eat-2 animals expressing GFP fused to lmn-1 driven by the promoter of the lmn-1 gene and control animals expressing emr-1 fused to GFP under lmn-1 promoter, were synchronized by bleaching. Embryos were placed on feeding plates containing either EV, atx-2 or aak-2 RNAi. Worms were moved to fresh RNAi plates every 48 h. Images were acquired at days 2, 4 and 6 (Figs 6 and 7) or at days 1, 3, 5 and 7 (Fig. 7) of adulthood. Nuclei were counted and grouped into three different classes (class I–III) based on their morphology: class I was nuclei without any nuclear deformation (smooth nuclei); class II was nuclei with mild nuclear abnormalities such as membrane folding and some lamin foci; class III was nuclei with severe nuclear deformation including increased nuclear folding, lobulations and abnormal nuclei shape. Nuclei were counted from the middle part of the worm (excluding head, tail and gonads) and of either hypodermal or muscular origin; neuronal nuclei were excluded. The researcher assessing the phenotype was blind to the experimental conditions. For each time point 8–11 animals were used. A P value was calculated using a Fisher exact probability test.
Microscopy
Fig. 1A was obtained using a Dino-Eye camera mounted on an Olympus MVX10 dissecting microscope. The image Fig. 1C was obtained using an ORCA-R2 camera (Hamamatsu Photonics) mounted on an Axioplan 2 microscope with 20× objective magnification. Images in Fig. 2A were obtained using a Nikon Eclipse E200 microscope with 4× or 10× objective magnifications fitted with a Moticam 2300 color camera (Motic). Images in Fig. 2C (quantification of GFP expression) were recorded using an Olympus MVX10 microscope. Images in Figs 5 and 6 were obtained using an ORCA-R2 camera (Hamamatsu Photonics) mounted on an Axioplan 2 microscope with 100 × objective magnification. Images in Fig. 7B,C, were obtained using a Leica SP5 confocal microscope. Panels A–F of Fig. 5 were obtained at the same exposure, but image levels were adjusted individually to improve feature visibility, as was the case of panels A–H of Fig. 6. The internal panels of Fig. 7B are displayed under identical conditions, as are the internal panels of Fig. 7C. However, the image levels of Fig. 7B and Fig. 7C are not identical.
Statistics
Unless noted otherwise, comparisons were performed using an unpaired two-tailed Student t-test. A minimum of three biological repeats per experiment were performed.
Supplementary Material
Acknowledgements
We thank Professor Yosef Gruenbaum for his help in experiment design and execution; Professor Michal Goldberg for a critical reading of the manuscript; and Gabriel Bonduryansky, Naama Zung and the lab of Yonatan Tzur for technical assistance and suggestions, specifically Hanna Achache and Yisrael Rappaport.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: D.Z.B.; Methodology: C.C.; Validation: C.C.; Data curation: C.C.; Writing - original draft: D.Z.B.; Writing - review & editing: D.Z.B., C.C., S.M.-C.; Visualization: C.C.; Supervision: D.Z.B.; Project administration: S.M.-C.; Funding acquisition: D.Z.B.
Funding
The work was supported by the Israeli Academy of Science (to Professor Yosef Gruenbaum) and the Israeli Science Foundation (grants 654/20 and 632/20 to D.Z.B.). Open access funding provided by Tel Aviv University. Deposited in PMC for immediate release.
Peer review history
The peer review history is available online at https://journals.biologists.com/jcs/article-lookup/doi/10.1242/jcs.258428.
References
- Ahringer, J. (2006). Reverse genetics, WormBook (ed. The C. elegans Research Community); http://www.wormbook.org. 10.1895/wormbook.1.47.1 [DOI] [Google Scholar]
- Avery, L. and Horvitz, H. R. (1990). Effects of starvation and neuroactive drugs on feeding in Caenorhabditis elegans. J. Exp. Zool. 253, 263-270. 10.1002/jez.1402530305 [DOI] [PubMed] [Google Scholar]
- Bank, E. M., Ben-Harush, K., Wiesel-Motiuk, N., Barkan, R., Feinstein, N., Lotan, O., Medalia, O. and Gruenbaum, Y. (2011). A laminopathic mutation disrupting lamin filament assembly causes disease-like phenotypes in Caenorhabditis elegans. Mol. Biol. Cell 22, 2716-2728. 10.1091/mbc.e11-01-0064 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bar, D. Z., Charar, C., Dorfman, J., Yadid, T., Tafforeau, L., Lafontaine, D. L. J. and Gruenbaum, Y. (2016). Cell size and fat content of dietary-restricted Caenorhabditis elegans are regulated by ATX-2, an mTOR repressor. Proc. Natl. Acad. Sci. USA 113, E4620-E4629. 10.1073/pnas.1512156113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bárcena, C., Quirós, P. M., Durand, S., Mayoral, P., Rodríguez, F., Caravia, X. M., Mariño, G., Garabaya, C., Fernández-García, M. T., Kroemer, G.et al. (2018). Methionine restriction extends lifespan in progeroid mice and alters lipid and bile acid metabolism. Cell Rep. 24, 2392-2403. 10.1016/j.celrep.2018.07.089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bettinger, J. C., Lee, K. and Rougvie, A. E. (1996). Stage-specific accumulation of the terminal differentiation factor LIN-29 during Caenorhabditis elegans development. Development 122, 2517-2527. 10.1242/dev.122.8.2517 [DOI] [PubMed] [Google Scholar]
- Bone, C. R., Tapley, E. C., Gorjánácz, M. and Starr, D. A. (2014). The Caenorhabditis elegans SUN protein UNC-84 interacts with lamin to transfer forces from the cytoplasm to the nucleoskeleton during nuclear migration. Mol. Biol. Cell 25, 2853-2865. 10.1091/mbc.e14-05-0971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brenner, S. (1974). The genetics of Caenorhabditis elegans. Genetics 77, 71–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Busch, A., Kiel, T., Heupel, W.-M., Wehnert, M. and Hübner, S. (2009). Nuclear protein import is reduced in cells expressing nuclear envelopathy-causing lamin A mutants. Exp. Cell Res. 315, 2373-2385. 10.1016/j.yexcr.2009.05.003 [DOI] [PubMed] [Google Scholar]
- Carmo-Silva, S., Nobrega, C., Pereira de Almeida, L. and Cavadas, C. (2017). Unraveling the role of Ataxin-2 in metabolism. Trends Endocrinol. Metab. 28, 309-318. 10.1016/j.tem.2016.12.006 [DOI] [PubMed] [Google Scholar]
- Carriere, A., Romeo, Y., Acosta-Jaquez, H. A., Moreau, J., Bonneil, E., Thibault, P., Fingar, D. C. and Roux, P. P. (2011). ERK1/2 phosphorylate Raptor to promote Ras-dependent activation of mTOR complex 1 (mTORC1). J. Biol. Chem. 286, 567-577. 10.1074/jbc.M110.159046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiarini, F., Evangelisti, C., Cenni, V., Fazio, A., Paganelli, F., Martelli, A. M. and Lattanzi, G. (2019). The cutting edge: the role of mTOR signaling in Laminopathies. Int. J. Mol. Sci. 20, 847. 10.3390/ijms20040847 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colman, R. J., Anderson, R. M., Johnson, S. C., Kastman, E. K., Kosmatka, K. J., Beasley, T. M., Allison, D. B., Cruzen, C., Simmons, H. A., Kemnitz, J. W.et al. (2009). Caloric restriction delays disease onset and mortality in rhesus monkeys. Science 325, 201-204. 10.1126/science.1173635 [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Angelo, M. A., Raices, M., Panowski, S. H. and Hetzer, M. W. (2009). Age-dependent deterioration of nuclear pore complexes causes a loss of nuclear integrity in postmitotic cells. Cell 136, 284-295. 10.1016/j.cell.2008.11.037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donnaloja, F., Jacchetti, E., Soncini, M. and Raimondi, M. T. (2019). Mechanosensing at the nuclear envelope by nuclear pore complex stretch activation and its effect in physiology and pathology. Front. Physiol. 10, 896. 10.3389/fphys.2019.00896 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duband-Goulet, I., Woerner, S., Gasparini, S., Attanda, W., Kondé, E., Tellier-Lebègue, C., Craescu, C. T., Gombault, A., Roussel, P., Vadrot, N.et al. (2011). Subcellular localization of SREBP1 depends on its interaction with the C-terminal region of wild-type and disease related A-type lamins. Exp. Cell Res. 317, 2800-2813. 10.1016/j.yexcr.2011.09.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elosegui-Artola, A., Andreu, I., Beedle, A. E. M., Lezamiz, A., Uroz, M., Kosmalska, A. J., Oria, R., Kechagia, J. Z., Rico-Lastres, P., Le Roux, A.-L.et al. (2017). Force triggers YAP nuclear entry by regulating transport across nuclear pores. Cell 171, 1397-1410.e14. 10.1016/j.cell.2017.10.008 [DOI] [PubMed] [Google Scholar]
- Eberlé, D., Hegarty, B., Bossard, P., Ferré, P. and Foufelle, F. (2004). SREBP transcription factors: master regulators of lipid homeostasis. Biochimie 86, 839-848. 10.1016/j.biochi.2004.09.018 [DOI] [PubMed] [Google Scholar]
- Friedland, A. E., Tzur, Y. B., Esvelt, K. M., Colaiácovo, M. P., Church, G. M. and Calarco, J. A. (2013). Heritable genome editing in C. elegans via a CRISPR-Cas9 system. Nat. Methods 10, 741-743. 10.1038/nmeth.2532 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friedman, D. L. and Ken, R. (1988). Insulin stimulates incorporation of 32Pi into nuclear lamins A and C in quiescent BHK-21 cells. J. Biol. Chem. 263, 1103-1106. 10.1016/S0021-9258(19)57270-2 [DOI] [PubMed] [Google Scholar]
- Goldman, R. D., Shumaker, D. K., Erdos, M. R., Eriksson, M., Goldman, A. E., Gordon, L. B., Gruenbaum, Y., Khuon, S., Mendez, M., Varga, R.et al. (2004). Accumulation of mutant lamin A causes progressive changes in nuclear architecture in Hutchinson–Gilford progeria syndrome. Proc. Natl Acad. Sci. USA 101, 8963-8968. 10.1073/pnas.0402943101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzalo, S., Kreienkamp, R. and Askjaer, P. (2017). Hutchinson-gilford progeria syndrome: a premature aging disease caused by LMNA gene mutations. Ageing Res. Rev. 33, 18-29. 10.1016/j.arr.2016.06.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer, E. L. and Brunet, A. (2009). Different dietary restriction regimens extend lifespan by both independent and overlapping genetic pathways in C. elegans. Aging Cell 8, 113-127. 10.1111/j.1474-9726.2009.00459.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo, Y. and Zheng, Y. (2015). Lamins position the nuclear pores and centrosomes by modulating dynein. Mol. Biol. Cell 26, 3379-3389. 10.1091/mbc.E15-07-0482 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haithcock, E., Dayani, Y., Neufeld, E., Zahand, A. J., Feinstein, N., Mattout, A., Gruenbaum, Y. and Liu, J. (2005). Age-related changes of nuclear architecture in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 102, 16690-16695. 10.1073/pnas.0506955102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han, M., Zhao, M., Cheng, C., Huang, Y., Han, S., Li, W., Tu, X., Luo, X., Yu, X., Liu, Y.et al. (2019). Lamin A mutation impairs interaction with nucleoporin NUP155 and disrupts nucleocytoplasmic transport in atrial fibrillation. Hum. Mutat. 40, 310-325. 10.1002/humu.23691 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heessen, S. and Fornerod, M. (2007). The inner nuclear envelope as a transcription factor resting place. EMBO Rep. 8, 914-919. 10.1038/sj.embor.7401075 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman, L. M., Smith, M. A., Jensen, C. C., Yoshigi, M., Blankman, E., Ullman, K. S. and Beckerle, M. C. (2020). Mechanical stress triggers nuclear remodeling and the formation of transmembrane actin nuclear lines with associated nuclear pore complexes. Mol. Biol. Cell 31, 1774-1787. 10.1091/mbc.E19-01-0027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoogewijs, D., Houthoofd, K., Matthijssens, F., Vandesompele, J. and Vanfleteren, J. R. (2008). Selection and validation of a set of reliable reference genes for quantitative sod gene expression analysis in C. elegans. BMC Mol. Biol. 9, 9. 10.1186/1471-2199-9-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inoki, K., Li, Y., Zhu, T., Wu, J. and Guan, K.-L. (2002). TSC2 is phosphorylated and inhibited by Akt and suppresses mTOR signalling. Nat. Cell Biol. 4, 648-657. 10.1038/ncb839 [DOI] [PubMed] [Google Scholar]
- Inoki, K., Zhu, T. and Guan, K.-L. (2003). TSC2 mediates cellular energy response to control cell growth and survival. Cell 115, 577-590. 10.1016/S0092-8674(03)00929-2 [DOI] [PubMed] [Google Scholar]
- Johnson, S. C., Rabinovitch, P. S. and Kaeberlein, M. (2013). mTOR is a key modulator of ageing and age-related disease. Nature 493, 338-345. 10.1038/nature11861 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamath, R. S. and Ahringer, J. (2003). Genome-wide RNAi screening in Caenorhabditis elegans. Methods 30, 313-321. 10.1016/S1046-2023(03)00050-1 [DOI] [PubMed] [Google Scholar]
- Kelley, J. B., Datta, S., Snow, C. J., Chatterjee, M., Ni, L., Spencer, A., Yang, C.-S., Cubeñs-Potts, C., Matunis, M. J. and Paschal, B. M. (2011). The defective nuclear lamina in hutchinson-gilford progeria syndrome disrupts the nucleocytoplasmic ran gradient and inhibits nuclear localization of Ubc9. Mol. Cell. Biol. 31, 3378-3395. 10.1128/MCB.05087-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laplante, M. and Sabatini, D. M. (2012). mTOR signaling in growth control and disease. Cell 149, 274-293. 10.1016/j.cell.2012.03.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lastres-Becker, I., Nonis, D., Eich, F., Klinkenberg, M., Gorospe, M., Kötter, P., Klein, F. A. C., Kedersha, N. and Auburger, G. (2016). Mammalian ataxin-2 modulates translation control at the pre-initiation complex via PI3K/mTOR and is induced by starvation. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 1862, 1558-1569. 10.1016/j.bbadis.2016.05.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lattanzi, G., Ortolani, M., Columbaro, M., Prencipe, S., Mattioli, E., Lanzarini, C., Maraldi, N. M., Cenni, V., Garagnani, P., Salvioli, S.et al. (2014). Lamins are rapamycin targets that impact human longevity: a study in centenarians. J. Cell Sci. 127, 147-157. 10.1242/jcs.133983 [DOI] [PubMed] [Google Scholar]
- Lee, K. K., Gruenbaum, Y., Spann, P., Liu, J. and Wilson, K. L. (2000). C. elegans nuclear envelope proteins emerin, MAN1, lamin, and nucleoporins reveal unique timing of nuclear envelope breakdown during mitosis. Mol. Biol. Cell. 11, 3089-3099. 10.1091/mbc.11.9.3089 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, S., Ogawa, W., Emi, A., Hayashi, K., Senga, Y., Nomura, K., Hara, K., Yu, D. and Kasuga, M. (2011). Role of S6K1 in regulation of SREBP1c expression in the liver. Biochem. Biophys. Res. Commun. 412, 197-202. 10.1016/j.bbrc.2011.07.038 [DOI] [PubMed] [Google Scholar]
- Link, J., Paouneskou, D., Velkova, M., Daryabeigi, A., Laos, T., Labella, S., Barroso, C., Pacheco Piñol, S., Montoya, A., Kramer, H.et al. (2018). Transient and partial nuclear lamina disruption promotes chromosome movement in early meiotic prophase. Dev. Cell 45, 212-225.e7. 10.1016/j.devcel.2018.03.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mair, W. and Dillin, A. (2008). Aging and survival: the genetics of life span extension by dietary restriction. Annu. Rev. Biochem. 77, 727-754. 10.1146/annurev.biochem.77.061206.171059 [DOI] [PubMed] [Google Scholar]
- Mannick, J. B., Del Giudice, G., Lattanzi, M., Valiante, N. M., Praestgaard, J., Huang, B., Lonetto, M. A., Maecker, H. T., Kovarik, J., Carson, S.et al. (2014). mTOR inhibition improves immune function in the elderly. Sci. Transl. Med. 6, 268ra179. 10.1126/scitranslmed.3009892 [DOI] [PubMed] [Google Scholar]
- Manning, B. D., Tee, A. R., Logsdon, M. N., Blenis, J. and Cantley, L. C. (2002). Identification of the tuberous sclerosis complex-2 tumor suppressor gene product tuberin as a target of the phosphoinositide 3-kinase/akt pathway. Mol. Cell 10, 151-162. 10.1016/S1097-2765(02)00568-3 [DOI] [PubMed] [Google Scholar]
- Markiewicz, E., Tilgner, K., Barker, N., van de Wetering, M., Clevers, H., Dorobek, M., Hausmanowa-Petrusewicz, I., Ramaekers, F. C. S., Broers, J. L. V., Blankesteijn, W. M.et al. (2006). The inner nuclear membrane protein emerin regulates β-catenin activity by restricting its accumulation in the nucleus. EMBO J. 25, 3275-3285. 10.1038/sj.emboj.7601230 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mörck, C. and Pilon, M. (2007). Caloric restriction and autophagy in Caenorhabditis elegans. Autophagy 3, 51-53. 10.4161/auto.3418 [DOI] [PubMed] [Google Scholar]
- O'Rourke, E. J., Soukas, A. A., Carr, C. E. and Ruvkun, G. (2009). C. elegans major fats are stored in vesicles distinct from lysosome-related organelles. Cell Metab. 10, 430-435. 10.1016/j.cmet.2009.10.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owens, D. J., Fischer, M., Jabre, S., Moog, S., Mamchaoui, K., Butler-Browne, G. and Coirault, C. (2020). Lamin mutations cause increased YAP nuclear entry in muscle stem cells. Cells 9, 816. 10.3390/cells9040816 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palgunow, D., Klapper, M. and Döring, F. (2012). Dietary restriction during development enlarges intestinal and hypodermal lipid droplets in Caenorhabditis elegans. PLoS ONE 7, e46198. 10.1371/journal.pone.0046198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papadopoli, D., Boulay, K., Kazak, L., Pollak, M., Mallette, F. A., Topisirovic, I. and Hulea, L. (2019). mTOR as a central regulator of lifespan and aging. F1000Research 8, 998. 10.12688/f1000research.17196.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pellegrini, C., Columbaro, M., Capanni, C., D'Apice, M. R., Cavallo, C., Murdocca, M., Lattanzi, G. and Squarzoni, S. (2015). All-trans retinoic acid and rapamycin normalize Hutchinson Gilford progeria fibroblast phenotype. Oncotarget 6, 29914-29928. 10.18632/oncotarget.4939 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pérez-Jiménez, M. M., Rodríguez-Palero, M. J., Ródenas, E., Askjaer, P. and Muñoz, M. J. (2014). Age-dependent changes of nuclear morphology are uncoupled from longevity in Caenorhabditis elegans IGF/insulin receptor daf-2 mutants. Biogerontology 15, 279-288. 10.1007/s10522-014-9497-0 [DOI] [PubMed] [Google Scholar]
- Peterson, T. R., Sengupta, S. S., Harris, T. E., Carmack, A. E., Kang, S. A., Balderas, E., Guertin, D. A., Madden, K. L., Carpenter, A. E., Finck, B. N.et al. (2011). mTOR complex 1 regulates lipin 1 localization to control the SREBP pathway. Cell 146, 408-420. 10.1016/j.cell.2011.06.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porta-de-la-Riva, M., Fontrodona, L., Villanueva, A. and Cerón, J. (2012). Basic Caenorhabditis elegans methods: synchronization and observation. J. Vis. Exp. 64, e4019. 10.3791/4019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramos, F. J., Chen, S. C., Garelick, M. G., Dai, D.-F., Liao, C.-Y., Schreiber, K. H., MacKay, V. L., An, E. H., Strong, R., Ladiges, W. C.et al. (2012). Rapamycin reverses elevated mTORC1 signaling in lamin A/C-deficient mice, rescues cardiac and skeletal muscle function, and extends survival. Sci. Transl. Med. 4, 144ra103. 10.1126/scitranslmed.3003802 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ranade, D., Pradhan, R., Jayakrishnan, M., Hegde, S. and Sengupta, K. (2019). Lamin A/C and Emerin depletion impacts chromatin organization and dynamics in the interphase nucleus. BMC Mol. Cell Biol. 20, 11. 10.1186/s12860-019-0192-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robida-Stubbs, S., Glover-Cutter, K., Lamming, D. W., Mizunuma, M., Narasimhan, S. D., Neumann-Haefelin, E., Sabatini, D. M. and Blackwell, T. K. (2012). TOR signaling and rapamycin influence longevity by regulating SKN-1/Nrf and DAF-16/FoxO. Cell Metab. 15, 713-724. 10.1016/j.cmet.2012.04.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rual, J.-F., Ceron, J., Koreth, J., Hao, T., Nicot, A.-S., Hirozane-Kishikawa, T., Vandenhaute, J., Orkin, S. H., Hill, D. E., van den Heuvel, S.et al. (2004). Toward improving Caenorhabditis elegans phenome mapping with an ORFeome-based RNAi library. Genome Res. 14, 2162-2168. 10.1101/gr.2505604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sancak, Y., Peterson, T. R., Shaul, Y. D., Lindquist, R. A., Thoreen, C. C., Bar-Peled, L. and Sabatini, D. M. (2008). The Rag GTPases bind raptor and mediate amino acid signaling to mTORC1. Science 320, 1496-1501. 10.1126/science.1157535 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato, R. (2010). Sterol metabolism and SREBP activation. Arch. Biochem. Biophys. 501, 177-181. 10.1016/j.abb.2010.06.004 [DOI] [PubMed] [Google Scholar]
- Sfakianos, A. P., Mellor, L. E., Pang, Y. F., Kritsiligkou, P., Needs, H., Abou-Hamdan, H., Désaubry, L., Poulin, G. B., Ashe, M. P. and Whitmarsh, A. J. (2018). The mTOR-S6 kinase pathway promotes stress granule assembly. Cell Death Differ. 25, 1766-1780. 10.1038/s41418-018-0076-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stiernagle, T. (2006). Maintenance of C. elegans. In WormBook (ed. The C. elegans Research Community). WormBook. 10.1895/wormbook.1.101.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torvaldson, E., Kochin, V. and Eriksson, J. E. (2015). Phosphorylation of lamins determine their structural properties and signaling functions. Nucleus 6, 166-171. 10.1080/19491034.2015.1017167 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Towbin, H., Staehelin, T. and Gordon, J. (1979). Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci. USA 76, 4350-4354. 10.1073/pnas.76.9.4350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turgay, Y., Eibauer, M., Goldman, A. E., Shimi, T., Khayat, M., Ben-Harush, K., Dubrovsky-Gaupp, A., Sapra, K. T., Goldman, R. D. and Medalia, O. (2017). The molecular architecture of lamins in somatic cells. Nature 543, 261-264. 10.1038/nature21382 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tzur, Y. B., Hersh, B. M., Horvitz, H. R. and Gruenbaum, Y. (2002). Fate of the nuclear lamina during Caenorhabditis elegans apoptosis. J. Struct. Biol. 137, 146-153. 10.1006/jsbi.2002.4452 [DOI] [PubMed] [Google Scholar]
- Uno, M. and Nishida, E. (2016). Lifespan-regulating genes in C. elegans. NPJ Aging Mech. Dis. 2, 16010. 10.1038/npjamd.2016.10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Tienen, F. H. J., Lindsey, P. J., Kamps, M. A. F., Krapels, I. P., Ramaekers, F. C. S., Brunner, H. G., van den Wijngaard, A. and Broers, J. L. V. (2019). Assessment of fibroblast nuclear morphology aids interpretation of LMNA variants. Eur. J. Hum. Genet. 27, 389-399. 10.1038/s41431-018-0294-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker, G., Houthoofd, K., Vanfleteren, J. R. and Gems, D. (2005). Dietary restriction in C. elegans: from rate-of-living effects to nutrient sensing pathways. Mech. Ageing Dev. 126, 929-937. 10.1016/j.mad.2005.03.014 [DOI] [PubMed] [Google Scholar]
- Wiesel, N., Mattout, A., Melcer, S., Melamed-Book, N., Herrmann, H., Medalia, O., Aebi, U. and Gruenbaum, Y. (2008). Laminopathic mutations interfere with the assembly, localization, and dynamics of nuclear lamins. Proc. Natl. Acad. Sci. USA 105, 180-185. 10.1073/pnas.0708974105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Worman, H. J. and Bonne, G. (2007). “Laminopathies”: a wide spectrum of human diseases. Exp. Cell Res, 313, 2121-2133. 10.1016/j.yexcr.2007.03.028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu, L., Zhou, B., Oshiro-Rapley, N., Li, M., Paulo, J. A., Webster, C. M., Mou, F., Kacergis, M. C., Talkowski, M. E., Carr, C. E.et al. (2016). An ancient, unified mechanism for metformin growth inhibition in C. elegans and Cancer. Cell 167, 1705-1718.e13. 10.1016/j.cell.2016.11.055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yen, K., Le, T. T., Bansal, A., Narasimhan, S. D., Cheng, J.-X. and Tissenbaum, H. A. (2010). A comparative study of fat storage quantitation in nematode Caenorhabditis elegans using label and label-free methods. PLoS ONE 5, e12810. 10.1371/journal.pone.0012810 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, Y., Chen, D., Smith, M. A., Zhang, B. and Pan, X. (2012). Selection of reliable reference genes in Caenorhabditis elegans for analysis of nanotoxicity. PLoS ONE 7, e31849. 10.1371/journal.pone.0031849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zuela-Sopilniak, N., Bar-Sela, D., Charar, C., Wintner, O., Gruenbaum, Y. and Buxboim, A. (2020). Measuring nucleus mechanics within a living multicellular organism: Physical decoupling and attenuated recovery rate are physiological protective mechanisms of the cell nucleus under high mechanical load. Mol. Biol. Cell 31, 1943-1950. 10.1091/mbc.E20-01-0085 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.