

Abstract
The onset and mechanisms underlying neurodegenerative diseases remain uncertain. The main features of neurodegenerative diseases have been related with cellular and molecular events like neuronal loss, mitochondrial dysfunction and aberrant accumulation of misfolded proteins or peptides in specific areas of the brain. The most prevalent neurodegenerative diseases belonging to age-related pathologies are Alzheimer's disease, Huntington's disease, Parkinson's disease and amyotrophic lateral sclerosis. Interestingly, mitochondrial dysfunction has been observed to occur during the early onset of several neuropathological events associated to neurodegenerative diseases. The master regulator of mitochondrial quality control and energetic metabolism is the transcriptional coactivator peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α). Additionally, it has been observed that PGC-1α appears to be a key factor in maintaining neuronal survival and synaptic transmission. In fact, PGC-1α downregulation in different brain areas (hippocampus, substantia nigra, cortex, striatum and spinal cord) that occurs in function of neurological damage including oxidative stress, neuronal loss, and motor disorders has been seen in several animal and cellular models of neurodegenerative diseases. Current evidence indicates that PGC-1α upregulation may serve as a potent therapeutic approach against development and progression of neuronal damage. Remarkably, increasing evidence shows that PGC-1α deficient mice have neurodegenerative diseases-like features, as well as neurological abnormalities. Finally, we discuss recent studies showing novel specific PGC-1α isoforms in the central nervous system that appear to exert a key role in the age of onset of neurodegenerative diseases and have a neuroprotective function in the central nervous system, thus opening a new molecular strategy for treatment of neurodegenerative diseases. The purpose of this review is to provide an up-to-date overview of the PGC-1α role in the physiopathology of neurodegenerative diseases, as well as establish the importance of PGC-1α function in synaptic transmission and neuronal survival.
Key Words: Alzheimer's disease, amyotrophic lateral sclerosis, Huntington's disease, mitochondrial dysfunction, Parkinson's disease, PGC-1α, synaptic function, vascular dementia
Introduction
Neurodegenerative diseases (NDs) are a group of neurological disorders characterized by neuronal loss in specific areas of the central nervous system (CNS) and peripheral nervous system (PNS) that can induce the impairment of vital functions like cognitive alterations, uncontrolled motor activities, and all functions that affect daily tasks related with postural balance, movement, memory, language, breathing, cardiac and social behavior (Dugger and Dickson, 2017). The more common NDs in the elderly population are Alzheimer's disease (AD), Huntington's disease (HD), Parkinson's disease (PD), amyotrophic lateral sclerosis (ALS) and vascular dementia (VaD). They are characterized by a common pathophysiology feature related with aberrant protein aggregation in the brain, e.g. amyloid beta (Aβ), hyperphosphorylated tau, alpha synuclein (α-syn), huntingtin (HTT) and TAR 43 DNA binding protein (Li et al., 2010; Marsh and Blurton-Jones, 2012; Prasad et al., 2019).
Currently, there are no effective therapies for NDs, and the existing treatments are only focused on mitigating the clinical symptoms in patients. The most dramatic example are AD therapies, which have not had any new drugs since 2003 (Cummings et al., 2014). Nowadays, several clinical trials are dedicated to search and find new biomarkers that contribute to the early diagnosis in preclinical or prodromal phases of NDs (Dugger and Dickson, 2017; Durães et al., 2018). It has been described that the key mechanisms of excitotoxicity of misfolded proteins are related with fast and progressive processes of oxidative stress, disruption of cell membranes, Ca2+ dyshomeostasis, mitochondrial dysfunction, synaptic failure and neuronal death (Li et al., 2016; Dugger and Dickson, 2017; Findley et al., 2019).
The master transcriptional coactivator PGC-1α (Peroxisome proliferator-activated receptor gamma coactivator 1-alpha) is the main mediator that coordinates mitochondrial biogenesis, cellular respiration, and energy metabolism (Anderson et al., 2008; Bai and Zhang, 2016; Jesse et al., 2017). Transcriptional function of PGC-1α is mainly regulated by post-translational modifications (PTMs) of NAD-dependent deacetylase sirtuin-1 (SIRT 1) and 5’AMP-activated protein kinase (AMPK), which act as metabolic sensors increasing ATP production and mitochondrial O2 consumption (Anderson et al., 2008; Bai and Zhang, 2016).
Analysis of PGC-1α expression patterns show an increase in protein levels in tissues with a high metabolic demand such as brain, heart, skeletal muscle, liver and brown adipose tissue (BAT) in order to promote gene transcription with important consequences in multiple cellular processes such as gluconeogenesis, insulin response, muscle function, glucose metabolism, angiogenesis, contractile function, energy metabolism, and neuroprotection (Lehman et al., 2000; Wenz, 2011). Particularly in neuronal survival, it has been observed that PGC-1α also participates in maintaining cholinergic (Zhao et al., 2011; Arnold et al., 2014), glutamatergic (Bartley et al., 2015), dopaminergic (Jiang et al., 2016), and GABAergic synapses in different areas of the CNS and PNS such as the cortex, striatum, substantia nigra, hippocampus, forebrain, spinal cord, and cerebellum (Bartley et al., 2015). Upregulation of PGC-1α has been shown to be neuroprotective against oxidative stress and cell damage by improving mitochondrial function, neuronal maintenance, and protein clearance (St-Pierre et al., 2006; Zolezzi and Inestrosa, 2013; Zolezzi et al., 2017). Overall, the aim of this review is to provide a complete picture of the downstream effects associated with the downregulation of PGC-1α in different models as well AD, VaD, HD, PD and ALS thereby providing a new point of view about the putative role of PGC-1α in the maintenance of neuronal function and its role as a potential target for novel pharmacological strategies in the NDs treatments.
Search Strategy and Selection Criteria
Studies cited in this review published from 2000 to 2020 were searched by general medical and science databases (PubMed and Web of Science), using the following keywords “Alzheimer” or “Alzheimer's disease” “Parkinson” or “Parkinson's disease”, “Amyotrophic lateral sclerosis”, “Huntington” or “Huntington's disease”, “Vascular Dementia”, “mitochondrial dysfunction” “PGC-1α isoforms, “brain-specific PGC-1α” for search strategy.
Biological Functions
Canonical PGC-1α and CNS isoforms
PGC-1α is a member of the mammalian family of transcriptional coactivators, along with PGC-1β, and PGC-1-related coactivator (St-Pierre et al., 2003). Initially, PGC-1α was identified as a cold-inducible transcription coactivator of adaptive thermogenesis, but nowadays its well knowing that PGC-1α plays key cellular roles in the regulation of energy metabolism, including gluconeogenesis, insulin sensitivity, mitochondrial biogenesis and ROS detoxification (Puigserver et al., 1998; St-Pierre et al., 2003; Rohas et al., 2007). PGC-1α is a 91 kDa protein encoded by PPARGC1A gene, which spans about 67 kb transcript, including 13 exons located on chromosome 4p15.1 (Esterbauer et al., 1999; Soyal et al., 2012). It has been shown that PGC-1 mRNA expression is markedly elevated upon against a variety of metabolic stimuli e.g., caloric restriction, hypoxia, exercise, as well high energy demands, in several tissues e.g., heart, skeletal muscle, kidney, liver, perirenal adipose tissue and brain (Esterbauer et al., 1999; Rohas et al., 2007). Regulation of transcription and gene expression on PPARGC1A gene are a highly regulated by canonical and non-canonical promoters, alternative splicing, as well PTMs e.g., DNA methylation, acetylation, and phosphorylation (Popov et al., 2017; Soyal et al., 2020).
Increasing evidence have identified novel isoforms of human PPARGC1A transcripts have been identified under control of tissue-specific promoters, such as SNC, liver, BAT, and skeletal (Martínez-Redondo et al., 2015; Soyal et al., 2020). Nowadays, it has been detected in human brain tissue several new brain-specific PGC-1α isoforms under the control of CNS promoter, initiating ~500 kb upstream of canonical promoter (Soyal et al., 2012, 2019). However, the differential physiological and pathological roles of these isoforms in the SNC have not completely understood. Particularly, it has been detected a small truncated 17 kDa PGC-1α isoform with uncertain function, some reports suggest that it should be involved in downregulation of full-length PGC-1α, as well in the onset of some NDs (see below) (Soyal et al., 2019). Actually, it has been suggest that nuclear transcription factors could interact with PGC-1α isoform in the brain to regulated motor function, as well genes related to neurotransmission in the cerebellum, in a sex-dependent brain transcriptional program (Lozoya et al., 2020). Interesting, it has been described that hypoxia, as well polymorphic guanidine thymidine dinucleotide ([GT]n) repeat, can regulated the activation of CNS promoter, in association with several transcription factors e.g., fork head box protein A2 (FOXA2, and hypoxia-inducible factor 1-alpha (Soyal et al., 2012, 2020). Future studies will need to address the mechanisms of regulation between CNS-PGC-1α isoforms, and their specific transcription factors in the brain.
Mechanism of PGC-1α regulation
The mechanisms underlying dynamic modulation of PGC-1α through PTMs have been extensively explored in the past decade. Assembly models suggest that PGC-1α promotes the recruitment of polymerase II, in association with nuclear receptor family transcription factors e.g., PPARs, ERa, and RXRa, acetyl/methyltransferase complexes, as well several coactivators e.g., SRC-1, CRE-binding protein, and CBP/p300, and complexes (Wallberg et al., 2003; Aguilo et al., 2016).
It has been described that nuclear/cytosolic sub-localization of PGC-1α are mediated mainly by cytosolic energy sensors; histone acetyltransferase general control non-repressed 5 protein, NAD+-dependent histone deacetylases silent information regulator 2 homolog 1 (SIRT1), and serine/threonine kinase AMPK (Cantó and Auwerx, 2009). General control non-repressed 5 protein directly acetylates PGC-1α, and negatively regulates its transcriptional activity, resulting in reduced gluconeogenesis, O2 uptake, and mitochondrial biogenesis rates (Lerin et al., 2006; Mutlu and Puigserver, 2020). On the other hand, under conditions of energetic stress, NAD+/NADH ratio increases, promoting SIRT1 activation, as well transcriptional activity of PGC-1, mediated by deacetylating at least one of its 13 acetylated lysine residues, resulting in increased expression in antioxidant proteins e.g., superoxide dismutase (SOD) and glutathione peroxidase (Lehman et al., 2000; Qin et al., 2009). Additionally, AMPK also can modulates PGC-1α/SIRT1 dependent antioxidant program, and mitochondrial homeostasis, in response to disruptions in cellular energy e.g., cellular adenylate charge and mitochondrial reactive oxygen species (Cantó and Auwerx, 2009; Rabinovitch et al., 2017; Figure 1).
Figure 1.
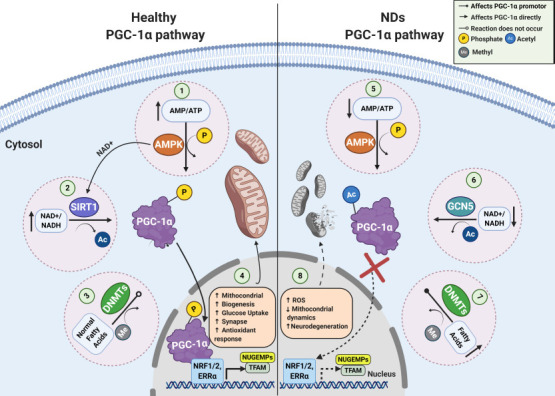
PGC-1α signaling pathways in a normal and pathophysiological context.
In healthy conditions, energy depletion and/or decreased catabolic rates can be sensed by AMPK and SIRT1, promoting the PGC-1α phosphorylation (1) and deacetylation (2), respectively. Additionally, under anti-inflammatory conditions DNMTs activity is downregulated (3), inducing an upregulation of PGC−1α promoter activity, and mitochondrial content as well (4). In NDs, PGC-1α is unable to be deacetylated (5), and phosphorylated (6) to translocate to the nucleus. Also, PGC−1α promoter methylation is upregulated (7), triggering the loss of key antioxidant and mitochondrial genes, imbalance between fusion and fission process, and neuronal death (8). AMPK: AMP-activated kinase; DNMTs: DNA methyltransferases; NDs: neurodegenerative diseases; PGC-1α: peroxisome proliferator-activated receptor gamma coactivator 1-alpha; SIRT1: silent information regulator 2 homolog 1.
Other relevant epigenetic modification that directly affects PGC-1α expression rates is linked to methylation/nucleosome positioning in PGC-1α promoter, which is mediated mainly by DNA methyltransferase (DNMT) isoforms (DNMT1,3A,3B) (Krämer and Handschin, 2019; Yang et al., 2020). Interestingly, upregulation in DNA methylation and nucleosome occupancy of PGC-1α promoter, would be implicated in the pathogenesis of some NDs (see below), obesity, diabetes, and liver disease (Su et al., 2015; Narkar, 2017; Krämer and Handschin, 2019). Elucidation of the mechanisms that regulate PGC-1α and the spatial and temporal identification of all its regulators under conditions of oxidative stress, should uncover promising therapeutic targets for the treatment of metabolic imbalances, and their associated diseases.
PGC-1α and quality control cycle of mitochondria
Quality control cycle of mitochondria depends on several processes that coordinately regulate the number, morphology, integrity of mitochondria. Among them are mitochondrial biogenesis and dynamics, protein turnover, and mitophagy (Celis-Muñoz et al., 2016). Mitochondrial biogenesis depends on the correct performance of mitochondrial/nuclear transcriptional control, which orchestrally coordinates the formation of new functional mitochondria, under several physiological requirements, such as oxidative stress, exercise, synapses, cold, and caloric restriction (Ventura-Clapier et al., 2008; Celis-Muñoz et al., 2016).
It has been described that PGC-1α to reach maximal bioenergetics capacity, acts in association with nuclear respiratory factors, such as nuclear respiratory factors 1 and 2 (NRF-1, NRF-2), as well nuclear hormone receptors, such as estrogen-related receptor alpha (ERRα, NR3B1), peroxisome proliferator-activated receptor gamma (PPAR-γ), and thyroid hormone receptors (Dominy and Puigserver, 2013). The most of mitochondrial genes (~1000) are under the control of nuclear transcriptional complexes to raises the transactivation of nuclear genes encoding mitochondrial proteins e.g., ATP synthase subunits, detoxification proteins, mitochondrial protein import machinery, as well mitochondrial transcription factor A (Schreiber et al., 2004; Rodgers et al., 2008; Celis-Muñoz et al., 2016; Ploumi et al., 2017). Posteriorly into the mitochondrial matrix, mitochondrial transcription factor A binds to the promoter of mitochondrial DNA, and regulates the expression of the rest 13 mitochondrial proteins, coded by circular mitochondrial genome (Gabrielson et al., 2014).
On the other hand, the balance between fusion and fission constitutes a key phenomenon on determining the size, morphology, and distribution of mitochondria. Mitochondrial dynamics are highly coordinated by mitofusin 1, 2 (Mfn1, Mfn2), and mitochondrial dynamin like GTPase for cristae fusion, while dynamin-related protein 1 and mitochondrial fission 1 protein have been related with the constriction and fission of mitochondrial cristae (Celis-Muñoz et al., 2016). Actually, several evidences have postulated that mitochondrial dynamics would be affected by mitochondrial biogenesis, in cross-regulatory circuits (Uittenbogaard and Chiaramello, 2014; Dorn et al., 2015; Song et al., 2015). PGC-1α-deficient mice showed an altered mitochondrial dynamics, including mitochondrial fragmentation and aberrant elongation, as well defects in mitochondrial fusion/fission proteins, such as Mfn1 and dynamin-related protein 1 (Uittenbogaard and Chiaramello, 2014; Dabrowska et al., 2015; Zhang et al., 2020). Improving mitochondrial biogenesis rates may be a potential target for the rectification of diseases associated with mitochondrial morphology defects.
PGC-1α and mitochondrial dysfunction in the CNS
Considering that numerous neuronal mechanisms depends on ATP synthesis and calcium homeostasis, the maintenance of mitochondrial function is essential for key biochemical processes, including Na+/K+ ATPase pump, purinergic receptors activity, and neurotransmission (Ly and Verstreken, 2006; Dabrowska et al., 2015; Godoy et al., 2019). Furthermore, there is increasing evidence indicated that mitochondrial dysfunction might contribute to the pathogenesis of several diseases, such as diabetes, heart, liver and kidney diseases, loss of muscle coordination, aging and several NDs (Dorn et al., 2015; Cai and Tammineni, 2017; Prasad et al., 2019). It has reported that PGC-1α downregulation has associated with mitochondrial dynamics alteration, raises in ROS production, Ca2+ dyshomeostasis, oxidative phosphorylation reduction, and ATP production imbalance (Cantó and Auwerx, 2009; Dabrowska et al., 2015; Martínez-Redondo et al., 2015; Figure 1).
In vitro and in vivo studies have indicated that PGC-1α regulation would involve in the support of multiple neuronal pathways in the brain, including hippocampus, substantia nigra, cortex, and striatum (Li et al., 2004; Cowell et al., 2007; Uittenbogaard and Chiaramello, 2014; Dabrowska et al., 2015). Particularly in neuronal survival, it has been observed that PGC-1α also participates in maintaining cholinergic (Zhao et al., 2011; Arnold et al., 2014), glutamatergic (Bartley et al., 2015), dopaminergic (Jiang et al., 2016), and GABAergic synapses in different areas of the CNS and PNS such as the cortex, striatum, substantia nigra, hippocampus, forebrain, spinal cord, and cerebellum (Bartley et al., 2015). Upregulation of PGC-1α has been shown to be neuroprotective against oxidative stress and cell damage, by improving mitochondrial function, neuroinflammation, protein clearance, and neuronal maintenance (St-Pierre et al., 2006; Zolezzi and Inestrosa, 2013; Zolezzi et al., 2017; Rius-Pérez et al., 2020). It is not possible to conclude a complete explanation for the cellular pathways controlled by PGC-1α during excitotoxicity, but selective in vivo PGC-1α activation could contribute to understanding physiological effects of mitochondrial dysfunction on synaptic network.
Alzheimer's Disease
The etiology of AD has not been completely determined and the discussion is still open. Cholinergic and glutamatergic dysfunction have been associated with the early cognitive impairments observed mainly in the hippocampus, cortex, and amygdala of AD patients (Castellani et al., 2010; Chen and Mobley, 2019). There are two classical histopathological biomarkers detected in AD: intracellular neurofibrillary tangles (Kosik et al., 1986), and extracellular amyloid plaques (Glenner and Wong, 1984) triggered by aberrant misfolding and accumulation of hyperphosphorylated tau protein and Aβ, respectively (Glenner and Wong, 1984; Kosik et al., 1986; Ballard et al., 2011). Currently, there is a large body of evidence indicating that the soluble oligomers of the Aβ peptide are the most neurotoxic agent because they produce the main aspects of AD pathology, e.g. Ca2+ dyshomeostasis and overload, ATP leakage, mitochondrial dysfunction, reticular stress, neuronal loss, synaptic failure and upregulation of the ionotropic channels such as P2XR and NMDAR (Fuentealba et al., 2011; Saez-Orellana et al., 2016, 2018; Sinnen et al., 2016; Godoy et al., 2019; Panes et al., 2020).
PGC-1α is a key mediator in the pathogenesis of AD
Increasing evidence postulate that the impairment of the PGC-1α pathway would be associated with the early loss of synaptic maintenance in AD brain regions like the hippocampus and cortex (Cheng et al., 2012; Bartley et al., 2015; Dong et al., 2020; Singulani et al., 2020; Additional Table 1). In fact, it has been observed in several neuronal Aβ toxicity models and patients with moderate and severe clinical dementia rating, that PGC-1α downregulation is critical for the early onset of the disease and for the defective responses in neuronal circuits of AD patients (Qin et al., 2009; Pedrós et al., 2014; Bartley et al., 2015; Dong et al., 2020; Terada et al., 2020). Additionally, it was shown that the level of PGC-1α markedly decreases in correlation with diminished mitochondrial functionality and nonamyloidogenic processing of APP in Tg2576 AD and APP/PS1 mice (Qin et al., 2009; Pedrós et al., 2014; Dong et al., 2020). Recently, it was found that soluble oligomers of the Aβ peptide were able to induce a loss of the interaction between PGC-1α and SIRT1 by decreasing its nuclear translocation and potentiating the imbalance in mitochondrial dynamics (Panes et al., 2020).
Table 1.
Summary of the main effects of PGC-1α regulation in different neurodegenerative disease models
| Mutations | In vivo/in vitro model | Disease | Main findings | References |
|---|---|---|---|---|
| Human AD (CDR=0-5) APP KM670/671NL (Swe) APP (Swe), PSEN1(deltaE9) | Postmortem AD brain Tg2576transgenic mice (hyperglycemic) APP/PS1transgenic mice |
AD | Decrease in hippocampal PGC-1α expression in function of neuriticplaque Decrease in hippocampal PGC-1α expression in function of Aβ peptide content Decrease in hippocampal PGC-1α expression in function of Nrf1/Nrf2 and OXPHOS downregulation |
Qin et al., 2009; Pedrós et al., 2014; Katsouri et al., 2016; Dong et al., 2020; Panes et al., 2020; Singulani et al., 2020 |
| APP KM670/671NL (Swe) | APP23transgenic mice | PGC-1α overexpression mitigates memory decline, neuronal loss and decreases amyloidogenic pathway | ||
| PS1(M146V), APP (Swe), and tau (P301L) | 3xTg-AD mice | Decrease in hippocampal PGC-1α expression at early stages, preceding AD-pathology | ||
| Wild type | PC-12cells and hippocampal neurons (AβOs treatment) | Decrease in PGC-1α/SIRT1 interaction andPGC-1α expression | ||
| Wild type | BCAS | VaD | Decrease PGC-1α expression inthe CA1 and CA3 area of hippocampus in function of downregulation of key mitochondrial antioxidants genes SOD2, Prx3, Trx2 and GPx1 | Hanet al., 2020 |
| Presymptomatic and symptomatic human HD Hdh(140 CAG repeats) Hdh(111 CAG repeats) |
Postmortem HD brain HD KI mice Immortalized striatal cells |
HD | Decrease of PGC-1α expression incaudate nucleus SNPs (rs17592631, rs2048025 and rs11737023) in PPARGC1A locus in functionof HD onset Downregulation of PGC-1α and mitochondrial genes in MSN Decrease of PGC-1α promoter activity |
Cui et al., 2006; Chaturvedi et al., 2009; Weydt et al., 2009; McConoughey et al., 2010; La Spada, 2012 |
| Hdh(82CAG repeats) | NLS-N171-82Q HD mice | Impaired PGC-1α activation and expression, metabolic disturbances, proteostasis failure, and decrease in mitochondrial genes and mitochondrial DNA inthe striatum | ||
| Hdh(93CAG repeats) | Drosophila model of HD | Decrease of cyt c mRNA levels | ||
| Late-onset sporadicPD and idiopathicPD patients with H&Y stage (R = 0.246, P = 0.020) Human mutantα−syn (A30P/A53T) Human MAOB Mutantα-syn (A53T) |
postmortem PD brain DMSYN PD mice MAO-B transgenic mice |
PD | Decrease of PPARGC1A mRNA expression mediated by hypermethylation of PGC-1α promoter Decrease of full-length and CNS PGC-1α in function of short-length CNS PGC-1α isoform in SNc Decrease of PGC-1α expression mediated by hypermethylation of CpG site DNA (CpG_1 and CpG_17.18) in PPARGC1A locus |
Zheng et al., 2010; Siddiqui et al., 2012; Stevens et al., 2015; Jiang et al., 2016; Soyal et al., 2019; Yang et al., 2020 |
| Mutantparkin (parkinflx/flx) | Stable dox-inducible MAO-B PC12 cells (under oxidative stress) | Decrease of PPARGC1A mRNA expression and aberrant mitochondrial morphology | ||
| ConditionalParkin KO | Downregulation of PGC-1α mediated byα-syn bindingto PGC-1α promoter | |||
| Transcriptional repression of PGC-1α, decline in dopaminergic viability and mitochondrial integrityloss | ||||
| Human sporadicALS (El Escorial criteria) and ALS patients (FUS/TLS mutations) Mutant SOD1 (G93A and G37R) Mutant FusΔNLS |
Postmortem ALS tissues ALS mice Knock-in FusΔNLS/ΔNLS mice |
ALS | Decrease of PPARGC1A mRNA expression in spinal cord, motor cortex and iPSC-derived motoneuronal culture Decrease of PGC-1α expression in function of muscle atrophyand locomotive activity disturbances Decrease of PGC-1α expression and PGC-1α–regulated factors (Tfeb, Sod3 and Bdnf) in lumbar spinal cord primary neurons, andmuscle tissues. |
Da Cruz et al., 2012; Thau et al., 2012; Eschbach et al., 2013; Ladd et al., 2014; Bayer et al., 2017 |
AD: Alzheimer’s disease; ALS: amyotrophic lateral sclerosis; APP: amyloid precursor protein; AβOs: amyloid β oligomers; BCAS: bilateral common carotid artery stenosis; Bdnf: brain-derived neurotrophic factor; CDR: clinical dementia rating; CNS: central nervous system; cyt c: cytochrome c; dox: doxycycline; Fus: fused in sarcoma; GPx: glutathione Peroxidase 1; H&Y stage: Hoehn and Yahr scale; HD: Huntington’s disease; Hdh: mouse Huntington’s disease gene homolog; iPSC: induced pluripotent stem cells; MAOB: monoamine oxidase B; MSN: medium spiny neurons; Nfr: nuclear respiratory factor; NLS: nuclear localization signal; PD: Parkinson’s disease; PGC-1α: peroxisome proliferator-activated receptor gamma coactivator 1-alpha; Prx3: peroxiredoxin 3; PS1: presenilin-1; SIRT1: Sirtuin 1; SNpc: substantia nigra pars compacta; SNPs: single nucleotide polymorphisms; SOD1: superoxide dismutase 1; SOD2: superoxide dismutase 2; Sod3: superoxide dismutase 1; Swe: Swedish; Tfeb: transcription factor EB; TLS: translated in liposarcoma; Trx2: thioredoxin-2; VaD: vascular dementia; α-syn: alpha-synuclein.
Studies in different AD transgenic models have demonstrated that the upregulation or overexpression of PGC-1α was able to mitigate neurodegeneration, Aβ accumulation, and cognitive failure observed in these models (Katsouri et al., 2016; Dong et al., 2020). Effectively, injection of the lentiviral vector human PGC-1α into the hippocampus and cortex areas of APP23 transgenic animals induced a decrease in Aβ production, pyramidal neurodegeneration, impairment of spatial memory, β-secretase transcription, microglial activation, downregulation of neurotrophic factors and the formation of amyloid plaques suggesting an important role of PGC-1α for the control of toxic Aβ species formation (Katsouri et al., 2016). Furthermore, APP/PS1 transgenic mice treated with resveratrol (RSV), a positive modulator of the SIRT1/PGC-1α pathway, showed an improvement in cognitive deficits, learning ability, and spatial memory (Dong et al., 2020). These studies suggest that PGC-1α could have a neuroprotective effect against neuronal loss and synaptic failure observed in AD.
Role of PGC-1α in inhibition/excitation balance in a hippocampal circuit
To understand how PGC-1α can affect synaptic function, it is necessary to evaluate the role of PGC-1α in the maintenance of the inhibition/excitation (I/E) balance in a hippocampal circuit (Cowell et al., 2007; Bartley et al., 2015; Kwakowsky et al., 2018). PGC-1α is highly expressed in GABAergic cell populations in the brain and participates in genetic transcription of the Ca2+ buffer protein parvalbumin in forebrain regions such as the cortex, hippocampus, and striatum (Cowell et al., 2007). Particularly, dysfunction in the GABAergic response (e.g. decrease in GABA currents, downregulation in GABA receptors, and the I/E imbalance) have been implicated in the onset of cognitive dysfunction observed during the early stages of AD, mainly in the hippocampus, superior temporal gyrus, and entorhinal cortex (Bartley et al., 2015; Kwakowsky et al., 2018). Studies in pyramidal CA1 cells of PGC-1α knockout (KO) mice demonstrated an increase in frequency-dependent I/E balance of the synaptic network and impaired nest building of the hippocampus mediated by an increase in the amplitude of inhibitory postsynaptic currents in response to Schaffer collateral stimulation in slices from young adult mice. In fact, PGC-1α KO young adult mice had cognitive alterations associated with an increase in inhibition of GABAergic interneurons of the parvalbumin, triggering a circuit dysfunction in CA1 activation and increasing hippocampal kainate-induced gamma oscillations and behavioral impairments (Cowell et al., 2007; Bartley et al., 2015; Kwakowsky et al., 2018; Figure 2).
Figure 2.
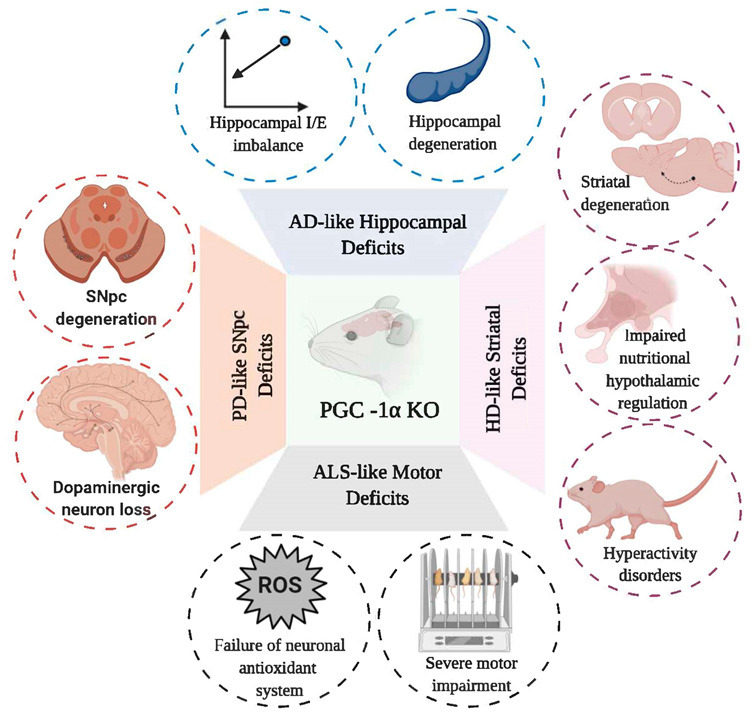
Schematic representation of cellular and behavioral changes observed in several models of PGC-1α KO mice.
PGC-1α deletion can emulate some features of AD, HD, PD and ALS; increasing sensitivity to neuronal loss, antioxidant failure, motor and behavioral abnormalities, and I/E imbalance. AD: Alzheimer's disease; ALS: amyotrophic lateral sclerosis; HD: Huntington's disease; I/E: inhibition-excitation balance; KO: knock out; PD: Parkinson's disease; PGC-1α: peroxisome proliferator-activated receptor gamma coactivator 1-alpha.
Vascular Dementia
VaD is characterized by several neuronal alterations like vascular cognitive deterioration causing a decline in learning, thinking, reasoning and language that is mediated by a progressive hypoperfusion of the cerebral tissues usually seen in patients with heart diseases or cerebral amyloid angiopathy and who have other vascular risk factors such as arterial hypertension, high cholesterol levels, cerebrovascular accidents, diabetes and heart diseases (O’Brien and Thomas, 2015). Clinical diagnosis of VaD is based on neuroimaging criteria such as lacunar strokes, white matter lesions, interruption of the blood-brain barrier, neuroinflammation and endothelial dysfunction of brain structures (Venkat et al., 2015; Cipollini et al., 2019). Traditionally, classic VaD treatments that are used are cholinesterase inhibitors, memantine, galantamine or antihypertensive approaches. Unfortunately, the development of specific treatments for VaD has been hampered by the lack of molecular targets or suitable animal models that can emulate the main features of the disease (Baskys and Hou, 2007; Gorelick et al., 2011). Currently, the early biomarkers that are being studied in patients with VaD are microRNAs (miRNAs), lipocalin 2 and lipid biomarkers such as high-density lipoprotein and low-density lipoprotein (Prabhakar et al., 2017; Cipollini et al., 2019; Llorens et al., 2020).
Emerging evidence of PGC-1α function in VaD
Increasing in vitro and in vivo studies have shown that some compounds or extracts such as RSV, allopregnanolone, Ginkgo biloba extract, and phytoestrogens have an antioxidant effect and enhance metabolic status in VaD patients (Prabhakar et al., 2017; Cipollini et al., 2019). However, there is little data about molecular mechanisms associated with VaD and mitochondrial dysfunction. A recent study in a VaD mouse model induced by bilateral common carotid artery stenosis (BCAS) showed a downregulation in PGC-1α in the CA1 and CA3 area of the hippocampus and a subsequent decrease in the expression of mitochondrial antioxidant genes in the BCAS group (Han et al., 2020; Additional Table 1). Furthermore, the generation of an animal model that overexpresses neuron-specific PGC-1α (nPGC-1α) after BCAS surgery showed an improvement in cognitive function recovery and LTP assessment induced by theta burst stimulation, and also a mitigation in cognitive deficits for learning and memory was observed in the BCAS animals. Additionally, nPGC-1α was able to induce an overexpression of antioxidant mitochondrial genes (e.g. SOD2, Prx3, GPx1, UCP2-5) in the hippocampus after BCAS surgery (Han et al., 2020). This study suggests that PGC-1α could be a potential neuroprotective agent against oxidative stress, energy metabolism imbalance, and neuroinflammation; however, more studies are needed to determine the real impact of PGC-1α function in the synaptic network.
Huntington's Disease
HD is a progressive and irreversible NDs characterized by progressive impairment in cognitive functions (e.g. memory, executive and visuospatial functions), motor functions (e.g. rigidity, chorea, dyskinesia), psychiatric disorders (e.g. irritability, emotional disorders, apathy and psychosis), and metabolic abnormalities (e.g. wasting and altered energy expenditure) (Wilson et al., 2017). HD etiology is associated with an autosomal dominant trait that appears mainly between 30 to 40 years of age triggered by a codon polymorphism expansion of a glutamine residue (CAG) encoded by the huntingtin gene (HTT, also called IT15) located on chromosome 4p16.3 (The Huntington's Disease Collaborative Research Group., 1993; Wilson et al., 2017). Mutant HTT (HTTmut) is able to form intracellular aggregates that can interfere with cellular processes that control gene regulation of energy homeostasis and it can induce a dysregulation in corticoestratial connectivity (Cui et al., 2006; Hervás-Corpión et al., 2018), This has been associated to an overactivity of glutamatergic neurotransmission, a reduction in GABAergic output in basal ganglia, excitotoxic cell injury, progressive neurodegeneration and atrophy of specific areas like the amygdala and striatum which increase thalamo-cortical activity leading to involuntary movements and motor disorders characteristic of the disease (Wilson et al., 2017).
Contribution of PGC-1α in the molecular pathogenesis of HD
Recently, evidence has shown that neurological and psychiatric disorders in HD patients are mediated by a progressive oxidative stress and genetic transcriptional dysregulation of mitochondrial and ribosomal biogenesis against HTTmut toxicity (Cui et al., 2006; Tsunemi and La Spada, 2012; Johri et al., 2013; Jesse et al., 2017). In this context, it has been reported that HTTmut can induce transcriptional downregulation of PGC-1α, induce an inactivation of the mitochondrial biogenesis process, and disrupt mitochondrial function (Cui et al., 2006; Soyal et al., 2012; Jesse et al., 2017; Jodeiri Farshbaf and Ghaedi, 2017). Several animal HD models and HD patients (striatal tissues) have shown that neurodegeneration induced by HTTmut toxicity was mediated by transcriptional dysregulation of PGC-1α (Additional Table 1) (Cui et al., 2006; Chaturvedi et al., 2009; La Spada, 2012; Tsunemi et al., 2012; Intihar et al., 2019).
Interestingly, It was suggested that the mechanism responsible for PGC-1α downregulation in striatal neurons in different HD models would be associated to direct interaction and blocking of HTTmut with the transcriptional activator of PGC-1α CRE-binding protein/TAF4, triggering the behavioral, metabolic, and neuronal disturbances observed in HD (Cui et al., 2006; Chaturvedi et al., 2009). Remarkably, several PGC-1α KO models can emulate the main characteristics of HD, i.e. defective mitochondrial bioenergetics, decreased neurite growth, striatal neurodegeneration, hyperactivity, astrogliosis, impairment in oxidative metabolism, muscle dysfunction, disturbed ribosomal transcription, and a spongiform pattern of lesions in white matter (Johri et al., 2013; Jesse et al., 2017; Figure 2). The nPGC-1α KO model also showed degenerative lesions in the striatum and deep cortical layers, alterations in axonal integrity, and decreases in metabolic programming of hypothalamic neuronal circuits (Ma et al., 2010). The breeding of PGC-1α KO mice with HD KI mice having 140 CAG repeats showed a significant potentiation in stromal neurodegeneration and motor abnormalities in the medial septal nucleus and striatum when compared to HD mice (Cui et al., 2006). Additionally, lentiviral overexpression of PGC-1a in the striatum of R6/2 HD transgenic mice could prevent the neuronal atrophy and dysregulation of axonal transport seen in HD mice (Cui et al., 2006). The breeding of N171-82Q HD transgenic mice with an inducible TRE-PGC-1α mice model was able to decrease aberrant aggregation and turnover of HTTmut, mitigate striatal neurodegeneration, and improve mitochondrial dysfunction observed in HD mice (Tsunemi et al., 2012).
PGC-1α isoforms are associated with early-onset in HD
Interestingly, it has been postulated that “loss of function” of several single nucleotide polymorphism (SNPs) mutations in the PPARGC1A locus that codes for several brain-specific isoforms upstream of exon 2 would be associated with a delay in age of onset of motor symptoms in patients with HD (Weydt et al., 2009; Soyal et al., 2012, 2019). In fact, the MAPS genome scan from HD patients showed several SNPs located in the PPARGC1A haplotypes at chromosomal region 4p16-4p15; rs17592631, rs2048025 and rs11737023, that would be associated with a “protective” genotype for HD incidence (Weydt et al., 2009). Furthermore, 5′-RLM-RACE experiments demonstrated a novel CNS-promoter for PPARGC1A transcripts that code for several dominant brain-specific PGC-1α isoforms that were able to interact with histone acetyltransferase complexes and dominant negative isoforms (Soyal et al., 2012). Moreover, it has been postulated that these truncated isoforms were able to block transcriptional regulation of the nPGC-1α isoform (Soyal et al., 2019). These novel transcripts should be further studied as a function of their interaction domains with mitochondrial cofactors, and we suggest that PGC-1α isoforms could have a protective effect improving neuronal energy imbalance and neurodegeneration.
Parkinson's Disease
PD is a progressive NDs characterized by a gradual loss and degeneration of dopaminergic neurons in the substantia nigra pars compacta (SNpc), which leads to movement and motor disturbances (e.g. tremor at rest, muscle stiffness, bradykinesia, and postural instability) (Emamzadeh and Surguchov, 2018; Kouli et al., 2018), and non-motor symptoms (e.g. depression, sleep disturbances, hyposmia, hallucinations, and dementia) (Martinez-Martin et al., 2011). The conventional histopathological biomarker detected in PD patients is composed of abnormal deposits of α-syn aggregates. The majority of abnormally deposited α-syn aggregates are found in the nigral dopaminergic system, particularly intracellularly in the cell soma and also in the neurological processes, also called Lewy and neurites bodies, respectively, that frequently appear between 55 and 70 years of age (Kalia and Lang, 2015). PD etiology is associated with several risk factors such as environmental factors (e.g. heavy metals, the mitochondrial neurotoxin MPTP, antipsychotic drugs, and antihemetics), family history (e.g. mutations in SNCA, PARK2, PINK1, PARK7 and PARK8 genes), and other NDs such as AD and HD (Emamzadeh and Surguchov, 2018).
Neuroprotective role of PGC-1α in PD progression
Current treatments for PD have been focused primarily on increasing dopamine levels by decreasing oxidative metabolism of dopamine using levodopa, l-DOPA, l-3,4 dihydroxyphenylalanine, dopamine receptor agonists (e.g. amantadine, apomorphine), and also by using mitochondrial monoamine oxidase B (MAO-B) inhibitors (e.g. selegiline and rasagiline) (Emamzadeh and Surguchov, 2018; Kouli et al., 2018). Recently, it has been postulated that one of the mechanisms that could be involved with the onset of PD may be associated with a downregulation of the PGC-1α pathway and decrease of mitochondrial metabolism in PD pathogenesis (Zheng et al., 2010; Youdim and Oh, 2013; Ciron et al., 2015). Indeed, it has been shown that using either MAO inhibitors or PGC-1α/SIRT1 activators (e.g. M30, HLA-20, LSN and RSV) improves mitochondrial and motor function and also provides neuroprotective effects against oxidative stress and mitochondrial toxins in several models of PD (Youdim, 2013; Ferretta et al., 2014; Emamzadeh and Surguchov, 2018; Kouli et al., 2018).
In fact, studies have demonstrated the participation of PGC-1α in neuronal survival of SNpc against mitochondrial neurotoxins associated with PD etiology (St-Pierre et al., 2006; Ciron et al., 2015; Stevens et al., 2015; Jiang et al., 2016; Ye et al., 2017). Using RNA interference to study protein function of PGC-1α in SH-SY5Y cells treated with N-methyl-4-phenylpyridinium ion (MPP+), it was found that PGC-1α ablation exerted a potentiation of mitochondrial dysfunction and cell death in MPP+-induced cells (Ye et al., 2017). The generation of a PGC-1α KO model showed a more sensitive phenotype in substantia nigra and hippocampus against oxidative stressors such as MPTP and kainic acid and had decreased axonal integrity and synaptic network integrity in deep cortical layers and striatum, as well as potentiating striatal lesions and oxidative damage (St-Pierre et al., 2006; Figure 2). α-syn overexpression in SNpc in the PGC-1α KO model also potentiated neurodegeneration, altered mitochondrial ultrastructure, and increased oxidative stress; alterations which were corrected by reestablishing PGC-1α expression (Ciron et al., 2015). Furthermore, conditional PGC-1α KO animals showed a progressive loss of dopaminergic neurons in SNpc, an important decrease in dopamine in the striatum, and subsequent reduction in mitochondrial biogenesis (Jiang et al., 2016). Furthermore, several mutations in the parkin and pink proteins also have been associated with the early loss of mitochondrial quality control and oxidative stress processes detected in PD (Matsuda et al., 2010; Stevens et al., 2015; Ge et al., 2020). In adult conditional parkin KO mice, as well as in PD patients, it was found that mitochondrial dysfunction and dopaminergic loss was associated with PGC-1α downregulation in a PARIS-dependent pathway (Pacelli et al., 2011; Stevens et al., 2015).
Differential role of PGC-1α in dopaminergic neuron survival
Recent studies have revealed that oxidative stress and neurodegeneration of dopaminergic neurons detected in PD models are implicated with several alterations in signaling pathways mediated by PGC1-α regulation (St-Pierre et al., 2006; Siddiqui et al., 2012; Corona and Duchen, 2015; Su et al., 2015; Soyal et al., 2019; Additional Table 1). In cortical neurons of MAO- B transgenic mice, and brain tissues of PD patients, it was shown that α-syn is able to bind with the PGC-1α promoter and inhibit its activity in the nucleus, triggering a PGC-1α downregulation, mitochondrial dysfunction, and cell degeneration (Zheng et al., 2010; Siddiqui et al., 2012). Additionally, in substantia nigra and peripheral blood samples from PD patients, studies found that PPARGC1A downregulation and PD clinical features were directly associated with the increase in non-canonical cytosine hypermethylation of the PGC-1α promoter, and in several CpG sites on the PPARGC1A locus (Su et al., 2015; Yang et al., 2020). Recently, it has been suggested that changes on the expression of CNS-specific PGC-1α (CNS-PGC-1α) full length has an important impact on CNS function, while the overexpression of truncated short isoforms in SNpc, would have deleterious effects related with the age of onset of PD (Yang et al., 2018, 2020; Soyal et al., 2019). Studies in post-mortem tissues from SNpc and globus pallidus of PD patients, indicated that downregulation in full-length PGC-1α isoforms and mitochondrial proteins decreased are associated with dopaminergic neuronal loss and progressive brain disorders observed in PD patients (Weydt et al., 2009; Jiang et al., 2016; Soyal et al., 2019). Taken together, the develop of specific promotor or enhancer of PGC-1a full length expression could be an innovative strategy to improve mitochondrial function for slow down the dopaminergic neurodegeneration in Parkinsonism.
Amyotrophic Lateral Sclerosis
ALS is a motor neuron disorder characterized by a selective loss of motor neurons in the brain, brainstem and spinal cord that leads to a progressive degeneration in upper and lower motor neurons in corticospinal tracts and induces a gradual deficiency in neuromuscular control (e.g. clonus, atrophy, fasciculation, muscle weakness, language problems, respiratory failure and paralysis), which ultimately leads to early death in patients 2–3 years after diagnosis (Muller et al., 2017). Multiparametric neuroimaging studies of ALS patients suggest that the selective anatomical vulnerability pattern in grey matter structures of basal ganglia and commissural white matter tracts could be associated with aberrant accumulation of SOD1 and TARDBP proteins (Bede et al., 2016; Prasad et al., 2019). Older age, gender difference, environment, oxidative stress, disturbances of energy metabolism, and a family history of ALS (e.g. C9ORF72, FUS/TLS, TBK, SOD1 and TARDBP gene mutations) have been recognized as risk factors for ALS onset (Tao and Wu, 2017).
Antioxidant and neuroprotective role of PGC-1α in amyotrophic lateral sclerosis
Increasing evidence in ALS models have shown that oxidative stress and mitochondrial abnormalities in the motor neurons could be associated with an increase on neuronal excitability and Ca2+ dysregulation (Wen et al., 2013; Ragagnin et al., 2019). It has been identified that PGC-1α also participates in: the regulation of gene expression at the neuromuscular junction, increased antioxidant response and increased mitochondrial biogenesis (Wiedemann et al., 1998; Arnold et al., 2014). Interestingly, a downregulation of PPARGC1A expression, and a subsequent decrease in a key mitochondrial genes (e.g. NRF-1, NRF-2, and Tfam), as well changes in the expression of acetylcholine receptor in skeletal muscle tissue were observed in ALS animal models and ALS patients (Eschbach et al., 2013; Ladd et al., 2014; Bayer et al., 2017; Additional Table 1).
Additionally, overexpression of PGC-1α in the SOD1G93A ALS mice model (TgSOD1G93A/PGC-1α) was able to mitigate neuronal death, motor dysfunction, metabolic disturbances and mitochondrial dysfunction in the ventral horn of the lumbar spinal cord of ALS mice (Zhao et al., 2011). Furthermore, breeding transgenic mice with constitutive PGC-1α overexpression in skeletal muscle (MCK-PGC-1α) with transgenic SOD1G37R mice (SOD1G37R/MCK-PGC-1α) was able to improve muscle atrophy, motor activity, mitochondrial dysfunction, and reduced muscle degeneration in SOD1G37R mice; however, it was not able to improve ALS progression or cell survival (Da Cruz et al., 2012; Johri and Beal, 2012). Remarkably, PGC-1α KO transgenic mice showed progressive alterations in motor coordination and downregulation of mRNA levels associated with antioxidant and neuroprotective responses (e.g. Tfeb, Sod3 and Bdnf) (Lucas et al., 2012; Bayer et al., 2017; Figure 2).
Emerging evidence of differential expression of PGC-1α in amyotrophic lateral sclerosis tissues
A biphasic tissue-specific PGC-1α expression has been recently demonstrated in ALS patients, SOD1G93A, and a FusΔNLS/ΔNLS mice models revealing that PPARGC1A mRNA levels was up-regulated in primary brown adipocytes, skeletal muscle and brown fat, whereas non-canonical isoforms of PGC-1α was downregulated in the spinal cord and brainstem of ALS-affected neuronal tissues (Bayer et al., 2017). Interestingly, down-regulation of CNS-PGC-1α expression and mitochondrial PGC-1α-dependent genes (e.g. Tfeb, Sod3 and Bdnf) were reestablished after lactate stimulation (Bayer et al., 2017). Moreover, it has been observed that CNS-PGC-1α isoforms could also exert a key role in the age of onset and gender differences in the incidence of ALS (Eschbach et al., 2013; Pasquinelli et al., 2016). In fact, downregulation of CNG-PARGC1A was more associated in the incidence of male onset of ALS-transgenic mice compared with female ALS-transgenic mice. A similar effect was observed in human ALS (Eschbach et al., 2013). Additionally, an interesting study in ALS patients strongly suggested that antioxidant defense impairment and motor atrophy during intensive training were related with Gly482Ser SNP located in the PPARGC1A genomic locus (Pasquinelli et al., 2016). Further studies about the neuroprotective effect of CNS-PGC-1α isoforms and interacting partners of DNA-binding proteins will be essential for understanding the transcriptional role of PGC-1α in the pathogenesis of ALS onset.
Conclusions
Taken together, the group of NDs reviewed and discussed in this work have been diagnosed traditionally by clinical symptoms; in addition, current pharmacological therapies have failed to mitigate neurodegeneration and anatomic disturbances observed in NDs. Here, we reviewed clear evidence supporting the neuroprotective role of PGC-1α against synaptic failure and neuronal death as a key pathway for the onset of these NDs. Downregulation of PGC-1α is key to understanding the toxicity mechanisms of misfolded proteins that would precede the oxidative stress and progressive neuronal death in specific areas of the brain (Tsunemi and La Spada, 2012; Bayer et al., 2017; Panes et al., 2020; Singulani et al., 2020). Growing knowledge about immunology and molecular biology strategies to develop humanized antibodies that can treat different types of pathologies, associated to the discussed evidence of the key role of PGC-1α isoforms, could be an essential starting point to open new chemical, or especially biological drug therapeutic strategies focused on improving mitochondrial and neuronal function in patients with NDs.
Additional files:
Additional Table 1: Summary of the main effects of PGC-1α regulation in different neurodegenerative disease models.
Additional file 1 (81.9KB, pdf) : Open peer review report 1.
Footnotes
P-Reviewer: Chen XQ; C-Editors: Zhao M, Qiu Y; T-Editor: Jia Y
Funding: This work was supported by Fondecyt 1200908 (to JF) and by the Conicyt 21141247 (to JDP).
Conflicts of interest:The authors declare no conflicts of interest.
Financial support:This work was supported by Fondecyt 1200908 (to JF) and by the Conicyt 21141247 doctoral grant (to JDP).
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open peer reviewer:Xu-Qiao Chen, University of California, USA.
References
- 1.Aguilo F, Li S, Balasubramaniyan N, Sancho A, Benko S, Zhang F, Vashisht A, Rengasamy M, Andino B, Chen CH, Zhou F, Qian C, Zhou MM, Wohlschlegel JA, Zhang W, Suchy FJ, Walsh MJ. Deposition of 5-methylcytosine on enhancer RNAs enables the coactivator function of PGC-1α. Cell Rep. 2016;14:479–492. doi: 10.1016/j.celrep.2015.12.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Anderson RM, Barger JL, Edwards MG, Braun KH, O’Connor CE, Prolla TA, Weindruch R. Dynamic regulation of PGC-1alpha localization and turnover implicates mitochondrial adaptation in calorie restriction and the stress response. Aging Cell. 2008;7:101–111. doi: 10.1111/j.1474-9726.2007.00357.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Anon The Huntington's Disease Collaborative Research Group. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington's disease chromosomes. Cell. 1993;72:971–983. doi: 10.1016/0092-8674(93)90585-e. [DOI] [PubMed] [Google Scholar]
- 4.Arnold AS, Gill J, Christe M, Ruiz R, McGuirk S, St-Pierre J, Tabares L, Handschin C. Morphological and functional remodelling of the neuromuscular junction by skeletal muscle PGC-1α. Nat Commun. 2014;5:3569. doi: 10.1038/ncomms4569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bai W, Zhang X. Nucleus or cytoplasm? The mysterious case of SIRT1's subcellular localization. Cell Cycle. 2016;15:3337–3338. doi: 10.1080/15384101.2016.1237170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ballard C, Gauthier S, Corbett A, Brayne C, Aarsland D, Jones E. Alzheimer's disease. Lancet. 2011;377:1019–1031. doi: 10.1016/S0140-6736(10)61349-9. [DOI] [PubMed] [Google Scholar]
- 7.Bartley AF, Lucas EK, Brady LJ, Li Q, Hablitz JJ, Cowell RM, Dobrunz LE. Interneuron transcriptional dysregulation causes frequency-dependent alterations in the balance of inhibition and excitation in hippocampus. J Neurosci. 2015;35:15276–15290. doi: 10.1523/JNEUROSCI.1834-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Baskys A, Hou AC. Vascular dementia: pharmacological treatment approaches and perspectives. Clin Interv Aging. 2007;2:327–335. [PMC free article] [PubMed] [Google Scholar]
- 9.Bayer H, Lang K, Buck E, Higelin J, Barteczko L, Pasquarelli N, Sprissler J, Lucas T, Holzmann K, Demestre M, Lindenberg KS, Danzer KM, Boeckers T, Ludolph AC, Dupuis L, Weydt P, Witting A. ALS-causing mutations differentially affect PGC-1α expression and function in the brain vs. peripheral tissues. Neurobiol Dis. 2017;97:36–45. doi: 10.1016/j.nbd.2016.11.001. [DOI] [PubMed] [Google Scholar]
- 10.Bede P, Iyer PM, Schuster C, Elamin M, Mclaughlin RL, Kenna K, Hardiman O. The selective anatomical vulnerability of ALS: “disease-defining” and “disease-defying” brain regions. Amyotroph Lateral Scler Frontotemporal Degener. 2016;17:561–570. doi: 10.3109/21678421.2016.1173702. [DOI] [PubMed] [Google Scholar]
- 11.Cai Q, Tammineni P. Mitochondrial aspects of synaptic dysfunction in Alzheimer's disease. J Alzheimers Dis. 2017;57:1087–1103. doi: 10.3233/JAD-160726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cantó C, Auwerx J. PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr Opin Lipidol. 2009;20:98–105. doi: 10.1097/MOL.0b013e328328d0a4. Curr Opin Lipidol 20:98-105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Castellani RJ, Rolston RK, Smith MA. Alzheimer disease. Dis Mon. 2010;56:484–546. doi: 10.1016/j.disamonth.2010.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Celis-Muñoz T, Silva-Grecchi T, A Godoy P, Panes-Fernández J, Barra K, Guzmán L, Fuentealba J. Impact of mitochondrial dysfunction on neurodegenerative diseases: a key step on Alzheimer's disease. J Syst Integr Neurosci. 2016;2:166–173. [Google Scholar]
- 15.Chaturvedi RK, Adhihetty P, Shukla S, Hennessy T, Calingasan N, Yang L, Starkov A, Kiaei M, Cannella M, Sassone J, Ciammola A, Squitieri F, Beal MF. Impaired PGC-1alpha function in muscle in Huntington's disease. Hum Mol Genet. 2009;18:3048–3065. doi: 10.1093/hmg/ddp243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chen XQ, Mobley WC. Exploring the pathogenesis of Alzheimer disease in basal forebrain cholinergic neurons: converging insights from alternative hypotheses. Front Neurosci. 2019;13:446. doi: 10.3389/fnins.2019.00446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cheng A, Wan R, Yang JL, Kamimura N, Son TG, Ouyang X, Luo Y, Okun E, Mattson MP. Involvement of PGC-1α in the formation and maintenance of neuronal dendritic spines. Nat Commun. 2012;3:1250. doi: 10.1038/ncomms2238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cipollini V, Troili F, Giubilei F. Emerging biomarkers in vascular cognitive impairment and dementia: from pathophysiological pathways to clinical application. Int J Mol Sci. 2019;20:2812. doi: 10.3390/ijms20112812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ciron C, Zheng L, Bobela W, Knott GW, Leone TC, Kelly DP, Schneider BL. PGC-1α activity in nigral dopamine neurons determines vulnerability to α-synuclein. Acta Neuropathol Commun. 2015;3:16. doi: 10.1186/s40478-015-0200-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Corona JC, Duchen MR. PPARγ and PGC-1α as therapeutic targets in Parkinson's. Neurochem Res. 2015;40:308–316. doi: 10.1007/s11064-014-1377-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cowell RM, Blake KR, Russell JW. Localization of the transcriptional coactivator PGC-1alpha to GABAergic neurons during maturation of the rat brain. J Comp Neurol. 2007;502:1–18. doi: 10.1002/cne.21211. [DOI] [PubMed] [Google Scholar]
- 22.Cui L, Jeong H, Borovecki F, Parkhurst CN, Tanese N, Krainc D. Transcriptional repression of PGC-1alpha by mutant huntingtin leads to mitochondrial dysfunction and neurodegeneration. Cell. 2006;127:59–69. doi: 10.1016/j.cell.2006.09.015. [DOI] [PubMed] [Google Scholar]
- 23.Cummings JL, Morstorf T, Zhong K. Alzheimer's disease drug-development pipeline: few candidates, frequent failures. Alzheimers Res Ther. 2014;6:37. doi: 10.1186/alzrt269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Da Cruz S, Parone PA, Lopes VS, Lillo C, McAlonis-Downes M, Lee SK, Vetto AP, Petrosyan S, Marsala M, Murphy AN, Williams DS, Spiegelman BM, Cleveland DW. Elevated PGC-1α activity sustains mitochondrial biogenesis and muscle function without extending survival in a mouse model of inherited ALS. Cell Metab. 2012;15:778–786. doi: 10.1016/j.cmet.2012.03.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dabrowska A, Venero JL, Iwasawa R, Hankir MK, Rahman S, Boobis A, Hajji N. PGC-1α controls mitochondrial biogenesis and dynamics in lead-induced neurotoxicity. Aging (Albany NY) 2015;7:629–647. doi: 10.18632/aging.100790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Davies SW, Turmaine M, Cozens BA, DiFiglia M, Sharp AH, Ross CA, Scherzinger E, Wanker EE, Mangiarini L, Bates GP. Formation of neuronal intranuclear inclusions underlies the neurological dysfunction in mice transgenic for the HD mutation. Cell. 1997;90:537–548. doi: 10.1016/s0092-8674(00)80513-9. [DOI] [PubMed] [Google Scholar]
- 27.Dominy JE, Puigserver P. Mitochondrial biogenesis through activation of nuclear signaling proteins. Cold Spring Harb Perspect Biol. 2013;5:a015008. doi: 10.1101/cshperspect.a015008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Dong YT, Cao K, Xiang J, Shan L, Guan ZZ. Silent mating-type information regulation 2 Homolog 1 attenuates the neurotoxicity associated with Alzheimer disease via a mechanism which may involve regulation of peroxisome proliferator-activated receptor gamma coactivator 1-α. Am J Pathol. 2020;190:1545–1564. doi: 10.1016/j.ajpath.2020.03.015. [DOI] [PubMed] [Google Scholar]
- 29.Dorn GW, 2nd, Vega RB, Kelly DP. Mitochondrial biogenesis and dynamics in the developing and diseased heart. Genes Dev. 2015;29:1981–1991. doi: 10.1101/gad.269894.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Dugger BN, Dickson DW. Pathology of neurodegenerative diseases. Cold Spring Harb Perspect Biol. 2017;9:a028035. doi: 10.1101/cshperspect.a028035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Durães F, Pinto M, Sousa E. Old drugs as new treatments for neurodegenerative diseases. Pharmaceuticals (Basel) 2018;11:44. doi: 10.3390/ph11020044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Emamzadeh FN, Surguchov A. Parkinson's disease: biomarkers, treatment, and risk factors. Front Neurosci. 2018;12:612. doi: 10.3389/fnins.2018.00612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Eschbach J, Schwalenstöcker B, Soyal SM, Bayer H, Wiesner D, Akimoto C, Nilsson AC, Birve A, Meyer T, Dupuis L, Danzer KM, Andersen PM, Witting A, Ludolph AC, Patsch W, Weydt P. PGC-1α is a male-specific disease modifier of human and experimental amyotrophic lateral sclerosis. Hum Mol Genet. 2013;22:3477–3484. doi: 10.1093/hmg/ddt202. [DOI] [PubMed] [Google Scholar]
- 34.Esterbauer H, Oberkofler H, Krempler F, Patsch W. Human peroxisome proliferator activated receptor gamma coactivator 1 (PPARGC1) gene: cDNA sequence, genomic organization, chromosomal localization, and tissue expression. Genomics. 1999;62:98–102. doi: 10.1006/geno.1999.5977. [DOI] [PubMed] [Google Scholar]
- 35.Ferretta A, Gaballo A, Tanzarella P, Piccoli C, Capitanio N, Nico B, Annese T, Di Paola M, Dell’aquila C, De Mari M, Ferranini E, Bonifati V, Pacelli C, Cocco T. Effect of resveratrol on mitochondrial function: implications in parkin-associated familiar Parkinson's disease. Biochim Biophys Acta. 2014;1842:902–915. doi: 10.1016/j.bbadis.2014.02.010. [DOI] [PubMed] [Google Scholar]
- 36.Findley CA, Bartke A, Hascup KN, Hascup ER. Amyloid beta-related alterations to glutamate signaling dynamics during Alzheimer's disease progression. ASN Neuro. 2019;11:1759091419855541. doi: 10.1177/1759091419855541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fuentealba J, Dibarrart AJ, Fuentes-Fuentes MC, Saez-Orellana F, Quinones K, Guzman L, Perez C, Becerra J, Aguayo LG. Synaptic failure and adenosine triphosphate imbalance induced by amyloid-beta aggregates are prevented by blueberry-enriched polyphenols extract. J Neurosci Res. 2011;89:1499–1508. doi: 10.1002/jnr.22679. [DOI] [PubMed] [Google Scholar]
- 38.Gabrielson M, Björklund M, Carlson J, Shoshan M. Expression of mitochondrial regulators PGC1α and TFAM as putative markers of subtype and chemoresistance in epithelial ovarian carcinoma. PLoS One. 2014;9:e107109. doi: 10.1371/journal.pone.0107109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ge P, Dawson VL, Dawson TM. PINK1 and Parkin mitochondrial quality control: a source of regional vulnerability in Parkinson's disease. Mol Neurodegener. 2020;15:20. doi: 10.1186/s13024-020-00367-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Glenner GG, Wong CW. Alzheimer's disease: initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem Biophys Res Commun. 1984;120:885–890. doi: 10.1016/s0006-291x(84)80190-4. [DOI] [PubMed] [Google Scholar]
- 41.Godoy PA, Ramírez-Molina O, Fuentealba J. Exploring the role of P2X receptors in Alzheimer's disease. Front Pharmacol. 2019;10:1330. doi: 10.3389/fphar.2019.01330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Gorelick PB, Scuteri A, Black SE, Decarli C, Greenberg SM, Iadecola C, Launer LJ, Laurent S, Lopez OL, Nyenhuis D, Petersen RC, Schneider JA, Tzourio C, Arnett DK, Bennett DA, Chui HC, Higashida RT, Lindquist R, Nilsson PM, Roman GC, et al. Vascular contributions to cognitive impairment and dementia: a statement for healthcare professionals from the american heart association/american stroke association. Stroke. 2011;42:2672–2713. doi: 10.1161/STR.0b013e3182299496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Han B, Jiang W, Liu H, Wang J, Zheng K, Cui P, Feng Y, Dang C, Bu Y, Wang QM, Ju Z, Hao J. Upregulation of neuronal PGC-1α ameliorates cognitive impairment induced by chronic cerebral hypoperfusion. Theranostics. 2020;10:2832–2848. doi: 10.7150/thno.37119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hervás-Corpión I, Guiretti D, Alcaraz-Iborra M, Olivares R, Campos-Caro A, Barco Á, Valor LM. Early alteration of epigenetic-related transcription in Huntington's disease mouse models. Sci Rep. 2018;8:9925. doi: 10.1038/s41598-018-28185-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Intihar TA, Martinez EA, Gomez-Pastor R. Mitochondrial dysfunction in Huntington's disease; interplay between HSF1, p53 and PGC-1α transcription factors. Front Cell Neurosci. 2019;13:103. doi: 10.3389/fncel.2019.00103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Jesse S, Bayer H, Alupei MC, Zügel M, Mulaw M, Tuorto F, Malmsheimer S, Singh K, Steinacker J, Schumann U, Ludolph AC, Scharffetter-Kochanek K, Witting A, Weydt P, Iben S. Ribosomal transcription is regulated by PGC-1alpha and disturbed in Huntington's disease. Sci Rep. 2017;7:8513. doi: 10.1038/s41598-017-09148-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Jiang H, Kang SU, Zhang S, Karuppagounder S, Xu J, Lee YK, Kang BG, Lee Y, Zhang J, Pletnikova O, Troncoso JC, Pirooznia S, Andrabi SA, Dawson VL, Dawson TM. Adult conditional knockout of PGC-1α leads to loss of dopamine neurons. eNeuro 3:ENEURO.0183-16.2016. 2016 doi: 10.1523/ENEURO.0183-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Jodeiri Farshbaf M, Ghaedi K. Huntington's disease and mitochondria. Neurotox Res. 2017;32:518–529. doi: 10.1007/s12640-017-9766-1. [DOI] [PubMed] [Google Scholar]
- 49.Johri A, Beal MF. Muscling in on PGC-1α for improved quality of life in ALS. Cell Metab. 2012;15:567–569. doi: 10.1016/j.cmet.2012.04.015. [DOI] [PubMed] [Google Scholar]
- 50.Johri A, Chandra A, Flint Beal M. PGC-1α, mitochondrial dysfunction, and Huntington's disease. Free Radic Biol Med. 2013;62:37–46. doi: 10.1016/j.freeradbiomed.2013.04.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kalia L V, Lang AE. Parkinson's disease. Lancet. 2015;386:896–912. doi: 10.1016/S0140-6736(14)61393-3. [DOI] [PubMed] [Google Scholar]
- 52.Katsouri L, Lim YM, Blondrath K, Eleftheriadou I, Lombardero L, Birch AM, Mirzaei N, Irvine EE, Mazarakis ND, Sastre M. PPARγ-coactivator-1α gene transfer reduces neuronal loss and amyloid-β generation by reducing β-secretase in an Alzheimer's disease model. Proc Natl Acad Sci. 2016;113:12292–12297. doi: 10.1073/pnas.1606171113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kosik KS, Joachim CL, Selkoe DJ. Microtubule-associated protein tau (tau) is a major antigenic component of paired helical filaments in Alzheimer disease. Proc Natl Acad Sci U S A. 1986;83:4044–4048. doi: 10.1073/pnas.83.11.4044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kouli A, Torsney KM, Kuan WL. Parkinson's Disease: Etiology, Neuropathology, and Pathogenesis. In: Stoker TB, Greenland JC, editors. Parkinson's Disease: Pathogenesis and Clinical Aspects [Internet] Brisbane (AU): Codon Publications; 2018. [PubMed] [Google Scholar]
- 55.Krämer AI, Handschin C. How epigenetic modifications drive the expression and mediate the action of PGC-1α in the regulation of metabolism. Int J Mol Sci. 2019;20:5449. doi: 10.3390/ijms20215449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kwakowsky A, Calvo-Flores Guzmán B, Govindpani K, Waldvogel HJ, Faull RL. Gamma-aminobutyric acid A receptors in Alzheimer's disease: highly localized remodeling of a complex and diverse signaling pathway. Neural Regen Res. 2018;13:1362–1363. doi: 10.4103/1673-5374.235240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.La Spada AR. PPARGC1A/PGC-1α, TFEB and enhanced proteostasis in Huntington disease: defining regulatory linkages between energy production and protein-organelle quality control. Autophagy. 2012;8:1845–1847. doi: 10.4161/auto.21862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ladd AC, Keeney PM, Govind MM, Bennett JP. Mitochondrial oxidative phosphorylation transcriptome alterations in human amyotrophic lateral sclerosis spinal cord and blood. NeuroMolecular Med. 2014;16:714–726. doi: 10.1007/s12017-014-8321-y. [DOI] [PubMed] [Google Scholar]
- 59.Lehman JJ, Barger PM, Kovacs A, Saffitz JE, Medeiros DM, Kelly DP. Peroxisome proliferator-activated receptor gamma coactivator-1 promotes cardiac mitochondrial biogenesis. J Clin Invest. 2000;106:847–856. doi: 10.1172/JCI10268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lerin C, Rodgers JT, Kalume DE, Kim S, Pandey A, Puigserver P. GCN5 acetyltransferase complex controls glucose metabolism through transcriptional repression of PGC-1alpha. Cell Metab. 2006;3:429–438. doi: 10.1016/j.cmet.2006.04.013. [DOI] [PubMed] [Google Scholar]
- 61.Li XJ, Li H, Li S. Clearance of mutant huntingtin. Autophagy. 2010;6:663–664. doi: 10.4161/auto.6.5.12336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Li Y, Sun H, Chen Z, Xu H, Bu G, Zheng H. Implications of GABAergic neurotransmission in Alzheimer's disease. Front Aging Neurosci. 2016;8:31. doi: 10.3389/fnagi.2016.00031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Li Z, Okamoto KI, Hayashi Y, Sheng M. The importance of dendritic mitochondria in the morphogenesis and plasticity of spines and synapses. Cell. 2004;119:873–887. doi: 10.1016/j.cell.2004.11.003. [DOI] [PubMed] [Google Scholar]
- 64.Llorens F, Hermann P, Villar-Piqué A, Diaz-Lucena D, Nägga K, Hansson O, Santana I, Schmitz M, Schmidt C, Varges D, Goebel S, Dumurgier J, Zetterberg H, Blennow K, Paquet C, Baldeiras I, Ferrer I, Zerr I. Cerebrospinal fluid lipocalin 2 as a novel biomarker for the differential diagnosis of vascular dementia. Nat Commun. 2020;11:619. doi: 10.1038/s41467-020-14373-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lozoya OA, Xu F, Grenet D, Wang T, Stevanovic KD, Cushman JD, Jensen P, Hernandez B, Riadi G, Moy SS, Santos JH, Woychik RP. Mutations on a novel brainspecific isoform of PGC1α leads to extensive upregulation of neurotransmitterrelated genes and sexually dimorphic motor deficits in mice. bioRxiv. 2020 doi: https://doi.org/10.1101/2020.09.18.300418. [Google Scholar]
- 66.Lucas EK, Dougherty SE, McMeekin LJ, Trinh AT, Reid CS, Cowell RM. Developmental alterations in motor coordination and medium spiny neuron markers in mice lacking pgc-1α. PLoS One. 2012;7:e42878. doi: 10.1371/journal.pone.0042878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Ly CV, Verstreken P. Mitochondria at the synapse. Neuroscientist. 2006;12:291–299. doi: 10.1177/1073858406287661. [DOI] [PubMed] [Google Scholar]
- 68.Ma D, Li S, Lucas EK, Cowell RM, Lin JD. Neuronal inactivation of peroxisome proliferator-activated receptor γ coactivator 1α (PGC-1α) protects mice from diet-induced obesity and leads to degenerative lesions. J Biol Chem. 2010;285:39087–39095. doi: 10.1074/jbc.M110.151688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Marsh SE, Blurton-Jones M. Examining the mechanisms that link β-amyloid and α-synuclein pathologies. Alzheimers Res Ther. 2012;4:11. doi: 10.1186/alzrt109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Martinez-Martin P, Rodriguez-Blazquez C, Kurtis MM, Chaudhuri KR. The impact of non-motor symptoms on health-related quality of life of patients with Parkinson's disease. Mov Disord. 2011;26:399–406. doi: 10.1002/mds.23462. [DOI] [PubMed] [Google Scholar]
- 71.Martínez-Redondo V, Pettersson AT, Ruas JL. The hitchhiker's guide to PGC-1α isoform structure and biological functions. Diabetologia. 2015;58:1969–1977. doi: 10.1007/s00125-015-3671-z. [DOI] [PubMed] [Google Scholar]
- 72.Matsuda N, Sato S, Shiba K, Okatsu K, Saisho K, Gautier CA, Sou YS, Saiki S, Kawajiri S, Sato F, Kimura M, Komatsu M, Hattori N, Tanaka K. PINK1 stabilized by mitochondrial depolarization recruits Parkin to damaged mitochondria and activates latent Parkin for mitophagy. J Cell Biol. 2010;189:211–221. doi: 10.1083/jcb.200910140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Muller UC, Deller T, Korte M. Not just amyloid: physiological functions of the amyloid precursor protein family. Nat Rev Neurosci. 2017;18:281–298. doi: 10.1038/nrn.2017.29. [DOI] [PubMed] [Google Scholar]
- 74.Mutlu B, Puigserver P. GCN5 acetyltransferase in cellular energetic and metabolic processes. Biochim Biophys Acta Gene Regul Mech. 2021;1864:194626. doi: 10.1016/j.bbagrm.2020.194626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Narkar VA. PGC1α promoter methylation and nucleosome repositioning: insights into exercise and metabolic regulation in skeletal muscle. Endocrinology. 2017;158:2084–2085. doi: 10.1210/en.2017-00439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.O’Brien JT, Thomas A. Vascular dementia. Lancet. 2015;386:1698–1706. doi: 10.1016/S0140-6736(15)00463-8. [DOI] [PubMed] [Google Scholar]
- 77.Pacelli C, De Rasmo D, Signorile A, Grattagliano I, di Tullio G, D’Orazio A, Nico B, Comi G Pietro, Ronchi D, Ferranini E, Pirolo D, Seibel P, Schubert S, Gaballo A, Villani G, Cocco T. Mitochondrial defect and PGC-1α dysfunction in parkin-associated familial Parkinson's disease. Biochim Biophys Acta. 2011;1812:1041–1053. doi: 10.1016/j.bbadis.2010.12.022. [DOI] [PubMed] [Google Scholar]
- 78.Panes J, Godoy P, Silva-Grecchi T, Celis M, Ramírez-Molina O, Gavilan J, Munoz-Montesino C, Castro P, Moraga-Cid G, Yevenes G, Guzmán J, Salisbury J, Trushina E, Fuentealba J. Changes in PGC-1α/SIRT1 signaling impact on mitochondrial homeostasis in amyloid-beta peptide toxicity model: the new pharmacological target. Front Pharmacol. 2020;11:709. doi: 10.3389/fphar.2020.00709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Pasquinelli A, Chico L, Pasquali L, Bisordi C, Lo Gerfo A, Fabbrini M, Petrozzi L, Marconi L, Caldarazzo Ienco E, Mancuso M, Siciliano G. Gly482Ser PGC-1α gene polymorphism and exercise-related oxidative stress in amyotrophic lateral sclerosis patients. Front Cell Neurosci. 2016;10:102. doi: 10.3389/fncel.2016.00102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Pedrós I, Petrov D, Allgaier M, Sureda F, Barroso E, Beas-Zarate C, Auladell C, Pallàs M, Vázquez-Carrera M, Casadesús G, Folch J, Camins A. Early alterations in energy metabolism in the hippocampus of APPswe/PS1dE9 mouse model of Alzheimer's disease. Biochim Biophys Acta. 2014;1842:1556–1566. doi: 10.1016/j.bbadis.2014.05.025. [DOI] [PubMed] [Google Scholar]
- 81.Ploumi C, Daskalaki I, Tavernarakis N. Mitochondrial biogenesis and clearance: a balancing act. FEBS J. 2017;284:183–195. doi: 10.1111/febs.13820. [DOI] [PubMed] [Google Scholar]
- 82.Popov DV, Lysenko EA, Makhnovskii PA, Kurochkina NS, Vinogradova OL. Regulation of PPARGC1A gene expression in trained and untrained human skeletal muscle. Physiol Rep. 2017;5:e13543. doi: 10.14814/phy2.13543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Prabhakar P, Chandra SR, Christopher R. Circulating microRNAs as potential biomarkers for the identification of vascular dementia due to cerebral small vessel disease. Age Ageing. 2017;46:861–864. doi: 10.1093/ageing/afx090. [DOI] [PubMed] [Google Scholar]
- 84.Prasad A, Bharathi V, Sivalingam V, Girdhar A, Patel BK. Molecular mechanisms of TDP-43 misfolding and pathology in amyotrophic lateral sclerosis. Front Mol Neurosci. 2019;12:25. doi: 10.3389/fnmol.2019.00025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Puigserver P, Wu Z, Park CW, Graves R, Wright M, Spiegelman BM. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell. 1998;92:829–839. doi: 10.1016/s0092-8674(00)81410-5. [DOI] [PubMed] [Google Scholar]
- 86.Qin W, Haroutunian V, Katsel P, Cardozo CP, Ho L, Buxbaum JD, Pasinetti GM. PGC-1alpha expression decreases in the Alzheimer disease brain as a function of dementia. Arch Neurol. 2009;66:352–361. doi: 10.1001/archneurol.2008.588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Rabinovitch RC, Samborska B, Faubert B, Ma EH, Gravel S-P, Andrzejewski S, Raissi TC, Pause A, St-Pierre J, Jones RG. AMPK maintains cellular metabolic homeostasis through regulation of mitochondrial reactive oxygen species. Cell Rep. 2017;21:1–9. doi: 10.1016/j.celrep.2017.09.026. [DOI] [PubMed] [Google Scholar]
- 88.Ragagnin AMG, Shadfar S, Vidal M, Jamali MS, Atkin JD. Motor neuron susceptibility in ALS/FTD. Front Neurosci. 2019;13:532. doi: 10.3389/fnins.2019.00532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Rius-Pérez S, Torres-Cuevas I, Millán I, Ortega ÁL, Pérez S. PGC-1α, inflammation, and oxidative stress: an integrative view in metabolism. Oxid Med Cell Longev 2020. 2020 doi: 10.1155/2020/1452696. 1452696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Rodgers JT, Lerin C, Gerhart-Hines Z, Puigserver P. Metabolic adaptations through the PGC-1 alpha and SIRT1 pathways. FEBS Lett. 2008;582:46–53. doi: 10.1016/j.febslet.2007.11.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Rohas LM, St-Pierre J, Uldry M, Jäger S, Handschin C, Spiegelman BM. A fundamental system of cellular energy homeostasis regulated by PGC-1alpha. Proc Natl Acad Sci U S A. 2007;104:7933–7938. doi: 10.1073/pnas.0702683104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Sáez-Orellana F, Fuentes-Fuentes MC, Godoy PA, Silva-Grecchi T, Panes JD, Guzmán L, Yévenes GE, Gavilán J, Egan TM, Aguayo LG, Fuentealba J. P2X receptor overexpression induced by soluble oligomers of amyloid beta peptide potentiates synaptic failure and neuronal dyshomeostasis in cellular models of Alzheimer's disease. Neuropharmacology. 2018;128:366–378. doi: 10.1016/j.neuropharm.2017.10.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Saez-Orellana F, Godoy PA, Bastidas CY, Silva-Grecchi T, Guzman L, Aguayo LG, Fuentealba J. ATP leakage induces P2XR activation and contributes to acute synaptic excitotoxicity induced by soluble oligomers of beta-amyloid peptide in hippocampal neurons. Neuropharmacology. 2016;100:116–123. doi: 10.1016/j.neuropharm.2015.04.005. [DOI] [PubMed] [Google Scholar]
- 94.Schreiber SN, Emter R, Hock MB, Knutti D, Cardenas J, Podvinec M, Oakeley EJ, Kralli A. The estrogen-related receptor α (ERRα) functions in PPARγ coactivator 1α (PGC-1α)-induced mitochondrial biogenesis. Proc Natl Acad Sci U S A. 2004;101:6472–6477. doi: 10.1073/pnas.0308686101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Siddiqui A, Chinta SJ, Mallajosyula JK, Rajagopolan S, Hanson I, Rane A, Melov S, Andersen JK. Selective binding of nuclear alpha-synuclein to the PGC1alpha promoter under conditions of oxidative stress may contribute to losses in mitochondrial function: implications for Parkinson's disease. Free Radic Biol Med. 2012;53:993–1003. doi: 10.1016/j.freeradbiomed.2012.05.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Singulani MP, Pereira CPM, Ferreira AFF, Garcia PC, Ferrari GD, Alberici LC, Britto LR. Impairment of PGC-1α-mediated mitochondrial biogenesis precedes mitochondrial dysfunction and Alzheimer's pathology in the 3xTg mouse model of Alzheimer's disease. Exp Gerontol. 2020;133:110882. doi: 10.1016/j.exger.2020.110882. [DOI] [PubMed] [Google Scholar]
- 97.Sinnen BL, Bowen AB, Gibson ES, Kennedy MJ. Local and use-dependent effects of β-amyloid oligomers on NMDA receptor function revealed by optical quantal analysis. J Neurosci. 2016;36:11532–11543. doi: 10.1523/JNEUROSCI.1603-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Song M, Mihara K, Chen Y, Scorrano L, Dorn GW., 2nd Mitochondrial fission and fusion factors reciprocally orchestrate mitophagic culling in mouse hearts and cultured fibroblasts. Cell Metab. 2015;21:273–286. doi: 10.1016/j.cmet.2014.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Soyal SM, Bonova P, Kwik M, Zara G, Auer S, Scharler C, Strunk D, Nofziger C, Paulmichl M, Patsch W. The expression of CNS-specific PPARGC1A transcripts is regulated by hypoxia and a variable GT repeat polymorphism. Mol Neurobiol. 2020;57:752–764. doi: 10.1007/s12035-019-01731-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Soyal SM, Felder TK, Auer S, Hahne P, Oberkofler H, Witting A, Paulmichl M, Landwehrmeyer GB, Weydt P, Patsch W. A greatly extended PPARGC1A genomic locus encodes several new brain-specific isoforms and influences Huntington disease age of onset. Hum Mol Genet. 2012;21:3461–3473. doi: 10.1093/hmg/dds177. [DOI] [PubMed] [Google Scholar]
- 101.Soyal SM, Zara G, Ferger B, Felder TK, Kwik M, Nofziger C, Dossena S, Schwienbacher C, Hicks AA, Pramstaller PP, Paulmichl M, Weis S, Patsch W. The PPARGC1A locus and CNS-specific PGC-1α isoforms are associated with Parkinson's Disease. Neurobiol Dis. 2019;121:34–46. doi: 10.1016/j.nbd.2018.09.016. [DOI] [PubMed] [Google Scholar]
- 102.St-Pierre J, Drori S, Uldry M, Silvaggi JM, Rhee J, Jager S, Handschin C, Zheng K, Lin J, Yang W, Simon DK, Bachoo R, Spiegelman BM. Suppression of reactive oxygen species and neurodegeneration by the PGC-1 transcriptional coactivators. Cell. 2006;127:397–408. doi: 10.1016/j.cell.2006.09.024. [DOI] [PubMed] [Google Scholar]
- 103.St-Pierre J, Lin J, Krauss S, Tarr PT, Yang R, Newgard CB, Spiegelman BM. Bioenergetic analysis of peroxisome proliferator-activated receptor gamma coactivators 1alpha and 1beta (PGC-1alpha and PGC-1beta) in muscle cells. J Biol Chem. 2003;278:26597–26603. doi: 10.1074/jbc.M301850200. [DOI] [PubMed] [Google Scholar]
- 104.Stevens DA, Lee Y, Kang HC, Lee BD, Lee YI, Bower A, Jiang H, Kang S-U, Andrabi SA, Dawson VL, Shin J-H, Dawson TM. Parkin loss leads to PARIS-dependent declines in mitochondrial mass and respiration. Proc Natl Acad Sci U S A. 2015;112:11696–11701. doi: 10.1073/pnas.1500624112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Su X, Chu Y, Kordower JH, Li B, Cao H, Huang L, Nishida M, Song L, Wang D, Federoff HJ. PGC-1α promoter methylation in Parkinson's disease. PLoS One. 2015;10:e0134087. doi: 10.1371/journal.pone.0134087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Tao QQ, Wu ZY. Amyotrophic lateral sclerosis: precise diagnosis and individualized treatment. Chin Med J (Engl) 2017;130:2269–2272. doi: 10.4103/0366-6999.215323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Terada T, Obi T, Bunai T, Matsudaira T, Yoshikawa E, Ando I, Futatsubashi M, Tsukada H, Ouchi Y. In vivo mitochondrial and glycolytic impairments in patients with Alzheimer disease. Neurology. 2020;94:e1592–1604. doi: 10.1212/WNL.0000000000009249. [DOI] [PubMed] [Google Scholar]
- 108.Tsunemi T, Ashe TD, Morrison BE, Soriano KR, Au J, Roque RAV, Lazarowski ER, Damian VA, Masliah E, La Spada AR. PGC-1α rescues Huntington's disease proteotoxicity by preventing oxidative stress and promoting TFEB function. Sci Transl Med. 2012;4:142ra97. doi: 10.1126/scitranslmed.3003799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Tsunemi T, La Spada AR. PGC-1α at the intersection of bioenergetics regulation and neuron function: from Huntington's disease to Parkinson's disease and beyond. Prog Neurobiol. 2012;97:142–151. doi: 10.1016/j.pneurobio.2011.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Uittenbogaard M, Chiaramello A. Mitochondrial biogenesis: a therapeutic target for neurodevelopmental disorders and neurodegenerative diseases. Curr Pharm Des. 2014;20:5574–5593. doi: 10.2174/1381612820666140305224906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Venkat P, Chopp M, Chen J. Models and mechanisms of vascular dementia. Exp Neurol. 2015;272:97–108. doi: 10.1016/j.expneurol.2015.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Ventura-Clapier R, Garnier A, Veksler V. Transcriptional control of mitochondrial biogenesis: the central role of PGC-1alpha. Cardiovasc Res. 2008;79:208–217. doi: 10.1093/cvr/cvn098. [DOI] [PubMed] [Google Scholar]
- 113.Wallberg AE, Yamamura S, Malik S, Spiegelman BM, Roeder RG. Coordination of p300-mediated chromatin remodeling and TRAP/mediator function through coactivator PGC-1alpha. Mol Cell. 2003;12:1137–1149. doi: 10.1016/s1097-2765(03)00391-5. [DOI] [PubMed] [Google Scholar]
- 114.Wen L, Chen Z, Zhang F, Cui X, Sun W, Geary GG, Wang Y, Johnson DA, Zhu Y, Chien S, Shyy JY. Ca2+/calmodulin-dependent protein kinase kinase β phosphorylation of Sirtuin 1 in endothelium is atheroprotective. Proc Natl Acad Sci U S A. 2013;110:E2420–2427. doi: 10.1073/pnas.1309354110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Wenz T. Mitochondria and PGC-1α in Aging and Age-Associated Diseases. J Aging Res. 2011;2011:810619. doi: 10.4061/2011/810619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Weydt P, Soyal SM, Gellera C, Didonato S, Weidinger C, Oberkofler H, Landwehrmeyer GB, Patsch W. The gene coding for PGC-1alpha modifies age at onset in Huntington's disease. Mol Neurodegener. 2009;4:3. doi: 10.1186/1750-1326-4-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Wiedemann FR, Winkler K, Kuznetsov A V, Bartels C, Vielhaber S, Feistner H, Kunz WS. Impairment of mitochondrial function in skeletal muscle of patients with amyotrophic lateral sclerosis. J Neurol Sci. 1998;156:65–72. doi: 10.1016/s0022-510x(98)00008-2. [DOI] [PubMed] [Google Scholar]
- 118.Wilson H, De Micco R, Niccolini F, Politis M. Molecular imaging markers to track Huntington's disease pathology. Front Neurol. 2017;8:11. doi: 10.3389/fneur.2017.00011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Yang X, Xu S, Qian Y, He X, Chen S, Xiao Q. Hypermethylation of the gene coding for PGC-1α in peripheral blood leukocytes of patients with Parkinson's disease. Front Neurosci. 2020;14:97. doi: 10.3389/fnins.2020.00097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Yang XD, Qian YW, Xu SQ, Wan DY, Sun FH, Chen SD, Xiao Q. Expression of the gene coading for PGC-1α in peripheral blood leukocytes and related gene variants in patients with Parkinson's disease. Parkinsonism Relat Disord. 2018;51:30–35. doi: 10.1016/j.parkreldis.2018.02.037. [DOI] [PubMed] [Google Scholar]
- 121.Ye Q, Chen C, Si E, Cai Y, Wang J, Huang W, Li D, Wang Y, Chen X. Mitochondrial effects of PGC-1alpha silencing in MPP+ treated human SH-SY5Y neuroblastoma cells. Front Mol Neurosci. 2017;10:164. doi: 10.3389/fnmol.2017.00164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Youdim MB, Oh YJ. Promise of neurorestoration and mitochondrial biogenesis in Parkinson's disease with multi target drugs: an alternative to stem cell therapy. Exp Neurobiol. 2013;22:167–172. doi: 10.5607/en.2013.22.3.167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Youdim MBH. Multi target neuroprotective and neurorestorative anti-Parkinson and anti-Alzheimer drugs ladostigil and m30 derived from rasagiline. Exp Neurobiol. 2013;22:1–10. doi: 10.5607/en.2013.22.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Zhang L, Zhang Y, Chang X, Zhang X. Imbalance in mitochondrial dynamics induced by low PGC-1α expression contributes to hepatocyte EMT and liver fibrosis. Cell Death Dis. 2020;11:226. doi: 10.1038/s41419-020-2429-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Zhao W, Varghese M, Yemul S, Pan Y, Cheng A, Marano P, Hassan S, Vempati P, Chen F, Qian X, Pasinetti GM. Peroxisome proliferator activator receptor gamma coactivator-1alpha (PGC-1α) improves motor performance and survival in a mouse model of amyotrophic lateral sclerosis. Mol Neurodegener. 2011;6:51. doi: 10.1186/1750-1326-6-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Zheng B, Liao Z, Locascio JJ, Lesniak KA, Roderick SS, Watt ML, Eklund AC, Zhang-James Y, Kim PD, Hauser MA, Grünblatt E, Moran LB, Mandel SA, Riederer P, Miller RM, Federoff HJ, Wüllner U, Papapetropoulos S, Youdim MB, Cantuti-Castelvetri I, et al. PGC-1α, a potential therapeutic target for early intervention in Parkinson's disease. Sci Transl Med. 2010;2:52ra73. doi: 10.1126/scitranslmed.3001059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Zolezzi JM, Inestrosa NC. Peroxisome proliferator-activated receptors and Alzheimer's disease: hitting the blood-brain barrier. Mol Neurobiol. 2013;48:438–451. doi: 10.1007/s12035-013-8435-5. [DOI] [PubMed] [Google Scholar]
- 128.Zolezzi JM, Santos MJ, Bastias-Candia S, Pinto C, Godoy JA, Inestrosa NC. PPARs in the central nervous system: roles in neurodegeneration and neuroinflammation. Biol Rev Camb Philos Soc. 2017;92:2046–2069. doi: 10.1111/brv.12320. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.