

Abstract
Viruses have evolved precise mechanisms for using the cellular physiological pathways for their perpetuation. These virus-driven biochemical events must be separated in space and time from those of the host cell. In recent years, granular structures, known for over a century for rabies virus, were shown to host viral gene function and were named using terms such as viroplasms, replication sites, inclusion bodies, or viral factories (VFs). More recently, these VFs were shown to be liquid-like, sharing properties with membrane-less organelles driven by liquid–liquid phase separation (LLPS) in a process widely referred to as biomolecular condensation. Some of the best described examples of these structures come from negative stranded RNA viruses, where micrometer size VFs are formed toward the end of the infectious cycle. We here discuss some basic principles of LLPS in connection with several examples of VFs and propose a view, which integrates viral replication mechanisms with the biochemistry underlying liquid-like organelles. In this view, viral protein and RNA components gradually accumulate up to a critical point during infection where phase separation is triggered. This yields an increase in transcription that leads in turn to increased translation and a consequent growth of initially formed condensates. According to chemical principles behind phase separation, an increase in the concentration of components increases the size of the condensate. A positive feedback cycle would thus generate in which crucial components, in particular nucleoproteins and viral polymerases, reach their highest levels required for genome replication. Progress in understanding viral biomolecular condensation leads to exploration of novel therapeutics. Furthermore, it provides insights into the fundamentals of phase separation in the regulation of cellular gene function given that virus replication and transcription, in particular those requiring host polymerases, are governed by the same biochemical principles.
Introduction
Eukaryotic viruses are intracellular parasites in constant coevolution with their host cells. With small genomes and few protein products, they rely on the cellular machinery to execute their life cycle consisting of a series of steps starting with viral entry, and followed by postentry events that lead to the synthesis of viral proteins and genomes, and finally to virion assembly and release, according to particular strategies. These events are orchestrated with exquisite precision, balancing the need to hijack cellular chemical machinery while keeping the cell healthy enough to complete their infectious cycle. These tasks include blocking the innate immune response, preventing apoptosis and generating an adequate environment for genome transcription and replication, for which a physical separation away from the host cytosolic (or nuclear) components is required. Such compartmentalization must be finely regulated to coordinate transcription and replication with nucleocapsid and particle assembly.
To fulfill genome replication, viruses evolved diverse genomic frameworks. In many RNA viruses, such as negative-stranded viruses (NSVs), replication is accomplished by virus-encoded enzymes packaged into the mature viral particle, and which are readily available for initiating viral RNA synthesis upon infection. In those, viral proteins are translated from subgenomic viral mRNAs. In contrast, some positive-sense RNA viruses that do not transcribe subgenomic RNAs translate their viral proteins, including replication enzymes, directly from the messenger-sense genome. Irrespective of specific strategies employed by each viral group to take control of the cell machinery, fundamental questions arise on how de novo synthesized viral proteins and nucleic acids are organized to direct virus multiplication and assembly. Will these diffuse around the cytosol or nuclei? How is genome replication spatiotemporally organized? What mechanisms govern the physical partitioning of gene function and the formation and packaging of nucleocapsids into the virion?
A vast number of complex biochemical processes in the cell can organize into dynamic and finely tuned structures known as membrane-less organelles (MLOs). An increasingly large number of these are reported to be formed by liquid–liquid phase separation (LLPS) of macromolecules, giving place to biomolecular condensates (BMCs) that partake in a wide range of physiological and pathological processes across life kingdoms [1–3]. Examples of BMCs are stress granules (SGs), nucleoli, P granules, Cajal bodies, among others extensively reviewed [4–8].
At the time MLOs were being described as liquid-like structures, viral replication was also found to take place within granular structures with dynamic properties evoking LLPS. The scope of this review is to show that this appears to be a widespread phenomenon in virus life cycles, describe some examples, and provide a general biochemical background for LLPS and biomolecular condensation in connection with viral perpetuation. In addition, we discuss how these mechanisms operate for viral gene function and other aspects of virus biology that benefit from these dynamic structures.
Replication sites: Different names, similar entities
A common feature observed during the course of infection by many viruses, whether they replicate in the cytoplasm or nucleus of the infected cells, is the formation of electrondense structures. These structures have been referred to as viral factories (VFs), viral inclusions, inclusion bodies (IBs), replication organelles, viral replication compartments (VRCs), transcription–replication complexes, virosomes, or viroplasms. One of the first characterization of VFs was provided for Poxviruses, enveloped DNA viruses that replicate exclusively in the cytoplasm [9]. Poxvirus factories are sites where protein synthesis, viral mRNA transcription, and DNA replication take place and are gradually enwrapped by rough endoplasmic reticulum (ER) membranes, later dispersed as viral assembly starts [10].
Nuclear-replicating double-stranded (ds) DNA viruses share common strategies including the formation of VRCs that drive a profound remodeling of subnuclear compartments, such as the promyelocitic leukemia nuclear bodies, Cajal bodies, and nucleoli [11–13]. For example, during cytomegalovirus infection, VRCs coalesce to form a single compartment that may occupy most of the nuclear space [13]. These viruses can also interact with DNA damage response (DDR) machinery, frequently hijacking DNA repair and replication factors for their own benefit [14].
Factories formed by Rotavirus, a member of the Reoviridae family that is associated with viral gastroenteritis in young children and infants worldwide, have been extensively characterized. Transcription and replication of the viral segmented dsRNA genome, as well as packaging of the newly synthesized pregenomic RNA, take place into membrane-less cytosolic electron-dense inclusions termed viroplasms. These complex structures are composed of genomic dsRNAs packed together with viral and cellular proteins, including ER chaperones [15], lipid droplets-associated proteins [16], and ribonuclear proteins [17], and are organized in concentric layers [18].
NSVs are enveloped and contain either one (nonsegmented) or several single-stranded RNA segments. The viral genome is enwrapped by multiple copies of the nucleoprotein (N) forming viral ribonucleoprotein (vRNP) complexes, which associate with the viral RNA–dependent RNA polymerase (vRdRp). Almost all NSV replicate in the cytoplasm of the infected cells, where they form VFs typically involving at least N protein and the viral polymerase and, in some cases, additional viral proteins, such as, for example, the phosphoprotein of nonsegmented NSV (nsNSV), which is an essential cofactor of the polymerase [19]. They have also been shown to colocalize with a number of host proteins including those involved in cellular mRNA metabolism, ribosomal subunit proteins, SG proteins, translation initiation factors, and key factors of the interferon pathway among others [20–23].
Replication sites confined to membranous structures
A hallmark of positive-sense RNA viruses is the association of viral replication with an extensive rearrangement of cellular membranes including formation of invaginated vesicles or spherules, or double-membrane vesicles (DMVs). For example, within the Flaviviridae family, dengue virus and Zika virus build up a scaffold of single-membrane invaginated vesicles in the ER, which are connected to the cytosol by small pores [24,25]. They are assumed to be the site of genome replication, as they contain dsRNA replication intermediates and viral replicase complex proteins [26]. Hepatitis C virus, in turn, creates a membranous web of DMVs, which protrude from the ER and incorporate viral RNA and replicase proteins [24].
DMV-type VFs have also been described for the distantly related Picornaviridae, Arteriviridae, and Coronaviridae families [27–29]. Recent analyses of severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2)-infected cells have revealed that DMVs, which contain RNA, are connected to the cytosol through crown-shaped pores, which would provide a transport route for newly synthesized viral genomes and messenger RNAs into the cytoplasm [30,31]. Altogether, the extensive ER rearrangement organized by many positive single-stranded RNA viruses to facilitate viral replication is not limited to animal viruses, but also applies to members of the Bromoviridae and Tombusviridae families of plant viruses that generate vesicle-like membrane invaginations (spherules) where cellular proteins are recruited and replication takes place [32,33].
A number of positive-stranded RNA viruses, particularly SARS-CoV2 [34] and Flaviviruses, including hepatitis C virus, dengue virus, Zika virus, West Nile virus, and Japanese encephalitis virus [35], have been reported to co-opt lipid droplets as an auxiliary platform for viral replication. Viral recruitment of lipid droplets, which are dynamic cytoplasmic organelles containing neutral lipids and proteins, has been proposed to assist the RNA polymerase activity [36] as well as to sustain scaffolding for genome encapsidation process [37].
Overall, viral proteins synthetized in the cytosol must come together to form viral replication sites, which require physical and functional separation from host cell processes. What is their exact nature, and how is their assembly and disassembly regulated?
Biomolecular condensation and liquid–liquid phase separation
Principles of LLPS
Under certain solvent conditions, and above a determined threshold concentration, a homogenous polymer–solvent mixture can separate or “demix” into 2 phases, one enriched in the polymer at the expense of partial depletion of the same polymer from the diluted phase, leading to the formation of liquid-like droplets coexisting with the dilute surrounding. This well-known principle in synthetic polymer chemistry [38] applies to biological macromolecules, mainly proteins and nucleic acids, the key players in BMCs. In simple and general thermodynamic terms, the condensed phase is a lower entropy state (molecules have less freedom) than the homogeneous mixture or diluted phase. Thus, phase separation may only take place if the confinement-associated entropic cost is counterbalanced by favorable macromolecular interactions. This scenario is possible in poor solvent conditions, i.e., scarce polymer solubility and polymer–polymer interactions being more energetically favorable than polymer–solvent interactions. In other words, under phase separation conditions, the polymer or macromolecule has more affinity for itself (or other macromolecules) than for the solvent (Fig 1A). In the context of the highly crowded environment of the cell, corresponding to protein concentrations of ca 150 mg/mL [39], some proteins are often above their solubility. This supersaturation state is prone to rapid changes triggered by different effectors giving rise to phase separation [40].
Fig 1. Overview of LLPS, phase diagram, and modulation.
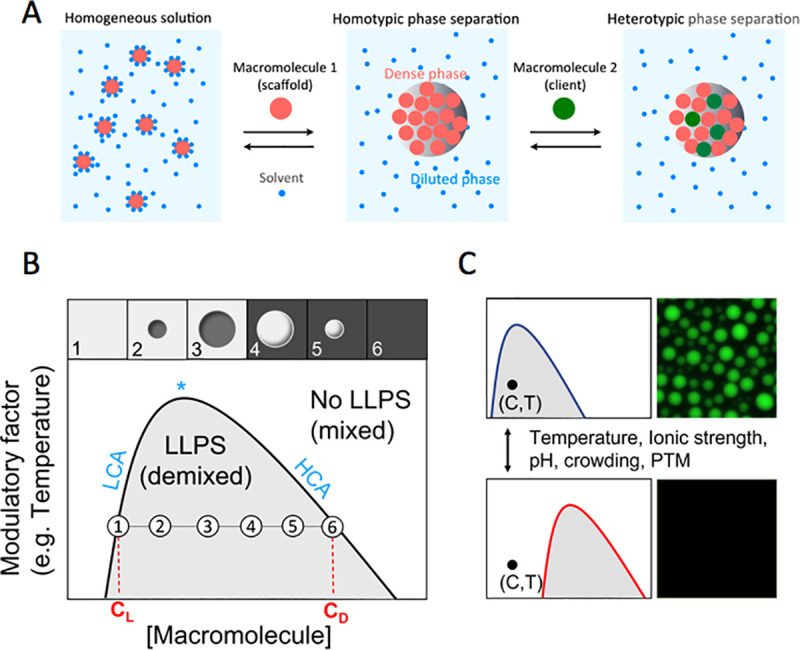
(A) Under certain conditions, a solvated macromolecule (scaffold) undergo homotypic LLPS and concentrate in a distinct liquid compartment (dense phase). One or more additional macromolecules (clients) can partition into the new phase through heterotypic LLPS. (B) A phase diagram describes the phase behavior of a binary (macromolecule and solvent) or multicomponent system (at least 2 macromolecules) as a function of macromolecular concentration or any other physicochemical factor that may modulate its condensation tendency. Here, we present a concentration vs temperature phase diagram for a binary system. A phase boundary (black curve, known as binodal) defines whether the system is in a 1-phase regime (mixed solution) or in a 2-phase regime (demixed solution). All coordinate pairs of concentration and temperature that lie beneath the phase boundary (gray) give rise to LLPS. The phase boundary maximum is the critical point (star), above which a homogeneous solution is seen at any macromolecular concentration. The critical point divides the phase boundary in 2 segments known as low concentration arm (LCA, left) and high concentration arm (HCA, right). The LCA defines the concentration of the diluted or light phase (CL), whereas the HCA defines the concentration of the dense phase (CD). Increasing total concentration in the light phase above the concentration threshold only changes the relative volumes between phases (i.e., droplets become larger at the expense of the diluted phase; see Fig 3). The top panel of (B) illustrates this phenomenon: (1) macromolecule concentration at threshold, no LLPS. At higher concentration, small droplets form (2) growing in size (3) as concentration increases. Eventually, the volume of the dense phase is higher than the diluted phase, so surface tension dictates the formation of diluted droplets surrounded by dense phase (4). After this inversion boundary, increasing concentration decreases the diluted droplets size (5) until a 1-phase regime of dense solution only is achieved (6). (C) Modulatory effectors such as PTMs or pH operate by altering the forces that drive droplet formation, thus changing the phase boundary. All of these modulatory effects may act in favor or against LLPS, depending on the nature of the interactions involved. For instance, addition of a negative charge by phosphorylation has the potential to engage components in electrostatic attractive or repulsive forces. C,T, concentration, temperature; HCA, high concentration arm; LCA, low concentration arm; LLPS, liquid–liquid phase separation; PTM, posttranslational modification.
A demixing process consisting of 1 polymer or macromolecular component and solvent is referred to as homotypic, whereas 2 or more components are referred to as heterotypic LLPS (Fig 1A). Hallmarks of LLPS are the formation of spherical liquid droplets, which increase in size upon increasing component concentration in the surrounding phase. These droplets are dynamic, they fuse and coalesce, are frequently reversible, and are studied using a broad range of methods [41,42].
Characteristics of the macromolecules and interactions involved
Proteins involved in LLPS usually exhibit a subset of the following features: (i) intrinsic disorder; (ii) multivalence; (iii) modularity; (iv) nucleic acid binding; (v) oligomeric nature; and (vi) dynamic conformation. Viral proteomes are known to be overabundant in intrinsically disordered proteins (IDPs), or regions (IDRs), providing functional flexibility and genome economy for interfering with cellular processes in favor of their infectious cycle [43]. Within IDPs, the so-called “low complexity regions” (LCRs) stand out as highly disordered polypeptides, some of them exhibiting a strong tendency for homotypic LLPS [44]. Low complexity refers to a high relative abundance of a small subset of the 20 amino acids; consequently, no significant folding or persistent secondary structure is possible. Folding inability may be due to the low abundance of aromatic and hydrophobic amino acids making it difficult for a hydrophobic core to be formed. Multivalent, low affinity (≤0.2 kcal/mol) interactions between certain amino acids drive LLPS in LCRs. They often involve aromatic (F, Y, W), polar (Q, N, S), or charged residues (D, E, R, K) that engage in π–π stacking or π–cation [45], dipolar [46], or complementary charge-based interactions [45,47]. On the other hand, multidomain modular proteins include a variety of examples with strong tendency to LLPS. These are composed of multiple folded domains separated by disordered linkers. They often form discrete and stable oligomers, which are an important driving force for LLPS by increasing avidity and lowering the entropic cost of confinement [5].
Although cellular condensates such as the nucleolus include a diversity of components [48], many proteins can undergo homotypic LLPS spontaneously without other components [45,47,49]. These molecules are referred to as scaffolds, which are able to recruit clients, components that do not form LLPS spontaneously but are a fundamental part of the condensate [5]. A combination of ratios between scaffold and clients, high or low binding affinities and very importantly high valence, dictate the compositional control of BMCs [5].
A large number of the condensates that make up MLOs involve RNA as a central player. These include SGs, processing bodies (PBs) [50], paraspeckles [51], and the nucleolus [52], among others, grouped together as ribonucleoprotein granules [53,54]. In these, RNA binding proteins and RNAs act as essential drivers for the condensate, since its polyanion nature confers multiple binding sites, i.e., high valency. Although less studied, similar considerations apply to DNA, as an anionic multivalent polymer capable of multiple weak electrostatic interactions driving BMC, in particular those associated to transcriptional control [55]. In line with this, emerging evidence highlights the importance of an interplay between chromatin organization, chromatin binding, and LLPS in the formation of transcription factories [56].
Modulation of BMCs assembly and disassembly
According to ideal polymer LLPS theory [57], an equilibrium exists between condensate droplets and the dilute phase. Thus, cellular droplet formation and dissolution can be achieved by varying macromolecular concentration above or below a concentration threshold (Fig 1B). The higher the concentration threshold, the lower the tendency for a macromolecule to condensate (Fig 1C). In turn, condensation tendency is governed by the sum of attractive and repulsive forces involved in macromolecular interactions. Thus, cellular control of droplet formation and dissolution can also be achieved by enabling/disabling interactions and modulating their affinities, either by covalent or noncovalent modifications on the scaffold itself (posttranslational modifications (PTMs), oligomerization state, ligands, or cosolutes) or through physicochemical changes in the milieu (temperature, ionic strength, osmolarity, pH, and crowding). All these modulatory effectors may act in favor or against LLPS depending on the nature of the interactions involved [58–60]. For instance, PTMs such as phosphorylation or methylation may affect the charge state or binding properties of a protein, thus imposing drastic effects on its phase-separating behavior [61].
Material properties
While many condensates exhibit highly dynamic liquid-like properties, others behave as less-dynamic, noncoalescent gels or even functional solids [62]. Thus, BMCs span a spectrum of viscoelastic properties suited to fulfill diverse functions that depend on markedly different diffusional rates, such as enhancing enzymatic activities or transiently storing biomolecules. Furthermore, LLPS is frequently mediated by metastable elements that may alternatively follow irreversible aggregation routes that derive in the formation of amorphous aggregates or regular fibrils, a phenomenon known as droplet maturation, aging, or liquid-to-solid transition [63,64]. As the function of BMCs depends on their material properties, cellular maintenance of droplet fluidity relies on ATP-consuming processes that may involve chaperones, disaggregases, and RNA-helicases [65–67].
Viral factories as dynamic liquid-like entities
Cumulative evidence suggests that assembly of VFs is often regulated by multivalent interactions that allow gradual incorporation of specific components [68]. Here, we describe some of the most representative examples of VFs that have been characterized to date as being liquid-like and potentially LLPS-driven.
Negative-stranded RNA viruses
nsNSVs are classified within the order Mononegavirales, which includes important human and animal pathogens such as rabies virus (RABV), measles virus (MeV), and Ebola virus (EBOV) [69]. Most of these viruses may utilize an endocytic pathway to enter cells and release their ribonucleoprotein complex (RNP) into the cytosol. All nsNSVs share a common genomic structure and code for 5 to 11 proteins depending on the virus species. The vRNP consists of the viral RNA genome encapsidated by N and associated with the vRdRp (L) and the nonenzymatic phosphoprotein (P). For some nsNSVs, additional viral proteins are also associated with the vRNP, such as the processivity factor M2-1 of pneumoviruses, or the equivalent transcription enhancer VP30 and the VP24 of filoviruses. Mononegavirales N proteins share structural characteristics; they present 2 globular domains with the RNA bound in a central groove [70]. P proteins (VP35 for filoviruses) differ significantly in sequence, size, and oligomeric state. However, a modular and dynamic architecture of a tight oligomeric domain flanked by IDRs is conserved (reviewed in [71,72]).
Initial observations of cytoplasmic viral condensates formed during nsNSV infections come from RABV. The viral genome is approximately 12 kb long and encodes 5 proteins: N, P, L, a matrix protein (M), and a glycoprotein (G), gene order being 3′-N-P-M-G-L-5′ in the viral RNA [73]. First studies of rabies infections by histological analyses of infected tissues revealed cytoplasmic inclusions, called Negri bodies (NBs) after their discovery in 1903 by Adelchi Negri [74]. NBs recruit all the viral replication machinery together with M and cellular proteins such as the focal adhesion kinase FAK, the chaperone Hsp70, and the eukaryotic cytosolic chaperonines CCTα and CCTγ [75–77]. NBs were shown to be involved in transcription, replication, and viral assembly and provided the first evidence that VFs are liquid-like and could be generated by LLPS in vivo [76,78]. Indeed, NBs display characteristics similar to LLPS-driven MLOs since they are small and spherical during early times after infection, they fuse and become more heterogeneous over time, and they can be dissolved by hypotonic shock [78]. Viral nucleocapsids are ejected from these condensates, possibly due to a decrease in their solubility within NBs, and are transported further away along the microtubule network to form new VFs [78]. Alternatively, vRNPs may outflow the NBs through the formation of a double membrane that surrounds the factories in late stages of infection, allowing direct budding of virions [73].
Shortly after the evidence that NBs are liquid-like VFs, condensates displaying similar properties were described for vesicular stomatitis virus (VSV), MeV, and human metapneumovirus [79–81]. MeV condensates, as for other nsNSVs, are composed of viral RNA, N, P, and L viral proteins. In addition, they contain the viral nonstructural C protein, which is involved in counteracting the host cell immune system [82], as well as the WD repeat-containing protein 5 (WDR5) host protein [23]. Interestingly, MeV condensates evolve from liquid to gel-like structures as they mature over time during infection, suggesting that physical properties may change to fulfill the viral replication cycle [80]. Another example illustrating the relevance of LLPS in nsNSV life cycle is respiratory syncytial virus (RSV), the main cause of bronchiolitis in infants worldwide. In RSV-infected cells, condensates are formed where viral RNA and N, P, and L proteins concentrate together with the viral transcription antiterminator M2-1. Only found in the Pneumoviridae family, M2-1 interacts with P and RNA [83] and plays a key role as an elongation factor, to enable synthesis of full-length viral mRNAs [84]. Termed IBs, these condensates have been characterized as sites where replication and transcription occur [85]. Interestingly, it has been shown that M2-1 and viral mRNA are transiently located in dynamic subcompartments within IBs, named inclusion body–associated granules (IBAGs), which exclude N, P, and L and genomic RNA. In vitro studies demonstrated that upon disassembly of IBAGs, M2-1 and mRNA are released from IBs, suggesting that M2-1 directs viral mRNAs to the cytosol for translation [85]. IBs also recruit M to the vRNP complex during infection, possibly inhibiting viral transcription to facilitate virion assembly and packaging [86].
Overall, nsNSV factories display properties of liquid organelles and concentrate viral RNA and at least N, P, and L viral proteins [87], but how proteins interact with each other and trigger VF assembly may differ along virus families. In vitro systems revealed the minimal requirements for the formation of these condensates. In the case of RABV, MeV, and RSV, coexpression of N and P proteins in transfected cells are necessary and sufficient to form spherical inclusions [78,80,85,88,89]. In the case of VSV, L forms inclusions by itself, but all 3 L, N, and P are required to trigger cytoplasmic phase separation [79], and for EBOV, the expression of N protein is sufficient for the generation of inclusions in transfected cells [90].
In contrast to other nsNSVs, Borna disease virus (BDV), a member of the Bornaviridae family, replicates noncytopathically in the nucleus and establishes persistent infections. The membrane-less BDV factories contain viral RNA and N and P proteins and are assembled within the nucleus in close association with host chromatin [91]. Photobleaching techniques to evaluate BDV components that make up these spherical nuclear inclusions revealed that P protein is mobile and shuttles between inclusions, suggesting LLPS properties [92].
Influenza virus
RNA viruses that replicate in the nucleus include members of the Orthomyxoviridae family, such as influenza A virus (IAV), a pathogen of worldwide impact. Following uncoating, IAV nucleocapsids are imported into the nucleus where viral mRNAs are transcribed and viral genome is replicated. Newly produced vRNPs, exported to the cytoplasm, come together and accumulate in membrane-less foci (vRNP hotspots) that can fuse together as they are transported to the plasma membrane, the site of virus assembly [93]. Cellular Rab11a-containing endosomes are thought to serve as platforms for the trafficking of vRNPs to the plasma membrane via the microtubule network [94]. These vRNP hotspots have been recently described as displaying characteristics of liquid-like organelles in terms of shape, dynamics, ability to deform, and reactivity to physiological changes and have been proposed to increase vRNA concentration at specific sites to facilitate the early stages of viral genome packaging [95].
Retroviruses
Retroviruses positive-sense single-strand RNA genome is retrotranscribed to a DNA molecule that is integrated into the host genome. The integrated provirus serves as template for the cellular RNA polymerase II (Pol II)-directed viral RNA synthesis, and viral assembly occurs at the plasma membrane, where proteins associated with dimers of genomic RNA condense to form immature budding particles [96]. LLPS has been implicated in different events of the human immunodeficiency virus type 1 (HIV-1) cycle. Viral infection was shown to rearrange intranuclear compartments and induce clustering of viral DNA, viral RNA, and host proteins in large nuclear foci, which originate in the absence of chromosomal integration and might represent viral reservoirs or a way to escape from the innate immune response [97,98]. Likewise, in vitro and in cellulo experiments have revealed that the HIV-1 nucleocapsid protein (NC) displays the ability to drive liquid-like condensates. NC is a small, basic nucleic acid binding protein derived from the Gag polyprotein precursor that wraps the viral RNA. NC bears intrinsically disordered prion-like domains (PrLDs) and 2 conserved Cys3His zinc-finger motifs. In vitro experiments showed that HIV-1 NC protein condensed into spherical assemblies in the presence of crowding agents or cell homogenates. NC condensates exhibited properties of LLPS droplets, including shape, fluidity, rapid internal diffusion, and the ability to fuse and coalesce, and required an intact Cys3His motif [99]. Treatment with Zn2+ ejectors caused nuclear relocalization of NC and viral RNA and inhibition of virus release. Moreover, a common Zn2+-dependent LLPS-based mechanism of retroviral assembly has been suggested that would also impact on the ability of HIV-1 to regulate the assembly and disassembly of SGs during infection [99].
DNA viruses
Herpes simplex virus 1 (HSV-1) was the first dsDNA virus hypothesized to perform replication in the context of LLPS, based on the fact that HSV-1 replication compartments, which recruit cellular RNA-Polymerase II, are spherical and fuse upon contact, and also on the fact that many HSV-1 proteins are predicted to be highly disordered [100–102]. However, a quantitative analysis suggested that the diffusion kinetics of these VRCs compared to the surrounding nucleoplasm would not be consistent with liquid-like properties [103]. Nevertheless, additional emerging evidence appears to support the involvement of proteins from HSV-1 and other members of the Herpesviridae family in the formation of BMCs. These include the HSV-1 transcription factor (TF) ICP4 [102] and HSV-1 UL11, the smallest conserved tegument protein among herpesviruses [104]. HSV-1 UL11 is an IDP, binds RNA, and undergoes LLPS in vitro, strongly suggesting that LLPS could be implicated in the assembly of the viral tegument layer located between the nucleocapsid and the lipid envelope [104]. The Epstein–Barr virus proteins EBNA2 and EBNALP, with roles in viral and cellular gene transcription, have been shown to mediate the formation of liquid-like condensates at superenhancer sites of cellular genes through their IDRs [105]. Another interesting example is the viral latency-associated nuclear antigen (LANA) from Kaposi’s sarcoma-associated herpesvirus, which associates with the viral genome to form dynamic LANA-nuclear bodies implicated in episome maintenance, through a mechanism partially mediated by LLPS [106].
Human papillomavirus (HPV) infection leads to the formation of the so-called replication foci or HPV E1/E2 foci, which contain the viral helicase E1 and the E2 master regulator and recruit DDR proteins [107]. HPV16 E2 can associate into ionic strength-dependent and readily reversible insoluble oligomers with both the E7 oncoprotein and with DNA [108,109]. While emerging evidence supports that compartmentalization of superenhancers is regulated by phase separation [110,111], formation of superenhancer-like elements has been postulated as a novel mechanism of HPV-16 integration [112], highlighting the potential role of LLPS in viral oncogenesis.
Nucleoprotein and RNA-driven LLPS in SARS-CoV-2
The SARS-CoV-2 nucleocapsid (NCoV2) protein encapsulates and packages the approximately 30 kb viral RNA genome into the 80 to 100 nm membrane-enveloped virion and regulates viral gene transcription [113]. It has globular RNA binding and dimerization domains, flanked by 2 IDRs and a linker between them. Its modularity, multivalence, flexibility, RNA binding capacity, and IDRs point NCoV2 as a candidate for LLPS. Indeed, several groups showed that it undergoes cooperative LLPS upon binding to RNA [114–119]. Heterotypic LLPS occurs at an optimal RNA length and concentration, above which the process is inhibited [119,118], a phenomenon known as reentrant phase separation, related to the ratio of binding sites [120]. There appears to be some sequence preferences for binding but no evident sequence specificity for LLPS [115,116,118,119], as expected for a protein that binds throughout the entire genome. Interestingly, structural features of certain regions of the genomic RNA drive condensation while other regions dissolve it [117]. Moreover, high density condensates of N-RNA in cells recruit the RNA-dependent RNA polymerase, supporting the role of LLPS-based viral condensation in SARS-CoV-2 transcription and replication [115]. The membrane protein (M) promotes heterotypic LLPS with N, and 3-component mixtures of N, M, and RNA form condensates with mutually exclusive compartments containing N-M or N-RNA. Structures described as annular were observed, with the M protein coating the outer layer of the N-RNA droplets [119].
Phosphorylation at the central IDR of NCoV2 is required for transcription at the replication transcription complex [121]. While unphosphorylated N forms gel-like structures, phosphorylation leads to liquid-like droplets, showing how the material properties of the condensate can be modulated by PTM [114,119]. Thus, unmodified N leads to a structured oligomer more competent for nucleocapsid assembly, whereas the liquid-like condensate formed by the phosphorylated protein leads to a condensate better suited for viral RNA synthesis activity [119]. Another possible layer of modulation arises from the biphasic triggering/dissolving effect of ATP on N-RNA LLPS, where ATP binds at an RNA site with affinities in the range of cellular concentrations of the nucleotide [122], suggesting a role for ATP in the uncoating, localization, and packing of the RNA genome [122].
SGs are at the crossroads between the viral infection and host factors and are modulated by viruses to maximize replication efficiency [123]. NCoV2 was found to phase separate in vitro with human RNA binding proteins prone to LLPS (TDP-43, FUS, hnRNPA2), all associated with SG formation, suggesting a possible mechanism to co-opt host proteins [116]. N was also reported to be associated with SGs within cells [115,119] and was shown to interact with Ras-GTPase-activating protein SH3-domain-binding protein (G3BP) and disrupt SG assembly through its IDR1 [124,119]. Further, N blocks the interaction of G3BP1 with SG-related proteins, and the domains of N important for phase separation with G3BP and SG disassembly are required for viral production [124]. N-RNA-driven LLPS was also shown to recruit TK1 and IKK complex, both key kinases in the NF-kB signaling pathway, producing a NF-κB hyperactivation, proposed as a possible route to dysfunctional inflammatory response [125]. Despite most of the work on SARS CoV2 condensation has been obtained by in vitro experiments with recombinant and pure components, N condensation through LLPS was suggested to take place during infection, and a natural mutation (R203K/G204R) resulting in a nucleotide polymorphism in 37% of 100,000 genome sequences analyzed was associated with higher propensity of N to undergo LLPS and a more pronounced effect on interferon inhibition [126]. Finally, gallocatechin gallate (GCG), a polyphenol obtained from green tea, and which interferes with viral N-RNA complexes of other viruses, was found to inhibit SARS-CoV-2 replication likely through impairment of LLPS of N [126].
Biochemical advantages of LLPS for viral factory formation, function, and fate
Viruses must replicate their genomes, transcribe their genes into mRNA using own or host enzymes, and rely on the cellular machinery for translation of viral proteins. The condensation of viral molecules into dynamic structures seems to be nature’s answer for these processes to be separated spatiotemporally. The most widely described VF assemblies to date are those of nsNSVs. We can picture how this might operate in the light of how their formation, function, and fate benefit from the chemistry behind LLPS. Fundamental features are at least in part shared even with viruses displaying different genome architecture and life cycles. We hypothesize a plausible set of sequential events from a biochemical perspective.
Upon entry, nsNSVs use the structural proteins carried within the infective virion for initial transcription. As protein levels increase and genome replication progresses, accumulated protein and RNA scaffolds nucleate and condense, recruiting viral and most likely several cellular clients. Additionally, the flexibility of the incoming vRNP and associated proteins could contribute to nucleate the LLPS process. A basic principle of LLPS is that the concentration of components within a condensate is constant, and increasing the concentration of components in the surrounding dilute phase leads to an increase in size of the condensate (Fig 1B). Further, a consequence of the transcription polarity of nsNSVs and their highly conserved gene arrangement is that those proteins encoded at the 3′ end of the genome will be produced first and at higher levels, with the late product invariably being the RNA-dependent RNA polymerase L (Fig 2A). While initial amounts of the vRdRp drive initial transcription and replication at minimum levels in the dilute cytosolic phase, the newly translated polymerase would enter at a later stage as client to the growing viral condensates.
Fig 2. A general integrative model for condensation of VFs based on NSVs.

(A) Simplified depiction of nsNSV genome structure. The viral transcription mechanism ensures a gradient of protein expression, with higher abundance of N, encoded at the 3′ region, and lower abundance of the large polymerase, invariably encoded at the 5′ end of nsNSV genomes. (B) Model depicting the formation of cytosolic VFs along viral infection. The schema stresses the amplification effect on RNA and protein synthesis caused by the self-primed formation of condensates (see main text). Toward the end of the infectious cycle, the immature nucleocapsids (proto-nucleocapsids) protrude from the VF and are transported along the cytoskeleton network to the assembly site [73]. The principle behind LLPS indicate that as the concentration of components increase, a partition into a newer denser phase takes place, where the concentration within the droplets remain constant, and further increase in the concentration of the components in the surrounding milieu cause an increment of the size of the condensates. The condensation event causes an abrupt increase of the effective local concentration of RNA and proteins, the drivers of the LLPS, which results in a substantial amplification of transcription and genome replication in the VF. LLPS, liquid–liquid phase separation; nsNSV, nonsegmented NSV; NSV, negative-stranded virus; RdRp, RNA-dependent RNA polymerase; VF, viral factory.
As the condensates grow in size at the expense of the accumulation of components, transcription and replication rates would increase at least 1 order of magnitude, as a consequence of the concentration/condensation process [127] (Fig 2B). Such a mass action phenomenon largely increases the effective local concentration of both the polymerase, its nucleocapsid template and the NTP substrates and cofactors, maximizing RNA synthesis, capping, and methylation, resulting in transcription amplification [128]. This idea is supported by changes from linear to exponential increase of RNA synthesis in Paramyxoviruses [81,129,130]. Another possible and not mutually exclusive explanation is that de novo synthesis of vRdRp from the incoming template correlates with the change from a linear to an exponential vRNA accumulation [130]. The enlargement in RNA length is equivalent to an increment in concentration as multiple binding sites become available, added to the increase in RNA transcript molecules per se, both acting as LLPS scaffolds that further rise the size of the VF. At a certain level of transcript concentration and possibly through mechanism involving compositional/stoichiometric control, mature transcripts exit to the cytosol for further translation and protein accumulation into the condensates, generating a self-priming amplification cycle that maximize growth of the VFs to structures of up to 20 μm2 hosting a large amount of encapsidated genomes [78]. In addition to a size-limiting mechanism, the number of VFs is also limited to a few per cell [78]. At this point, capsid/genome packages of the approximate size of the virion are ejected from the factories and transported via cytoskeleton to the membrane, with the participation of the matrix (M) protein [131,132], to bridge and interact with the intracellular domains of the viral membrane glycoproteins, which were synthesized in the ER, before exiting the cell [133,132]. In the case of nsNSVs, packages ejected from the VFs include the N-encapsidated genome and the structural proteins that will be part of the virion (L, polymerase cofactor/s and M protein) (referred to as “proto-nucleocapsids” in Fig 2B). Besides the concentration of the macromolecules, compositional control, confinement, and environmental factors, PTMs should also be considered as an additional key layer of control over formation and disassembly of the viral condensates, particularly if modifying host enzymes are likely present in the condensate [134].
It should be noted that the whole process requires that the interactions within the VF are weak, dynamic, and reversible, all properties intrinsic to LLPS condensates. Some of the components of replication complexes form stoichiometrically defined species in solution based on specific interactions, in equilibrium with the free components (Fig 3). Interactions that hold the condensate together are, on the other hand, weak, transient, and of low specificity. Thus, the droplet or condensate itself is not an ordered complex, but rather a different liquid phase that includes stoichiometric complexes interacting weakly with the other components, including cellular factors (Fig 3).
Fig 3. Nature of interactions in a viral condensate.
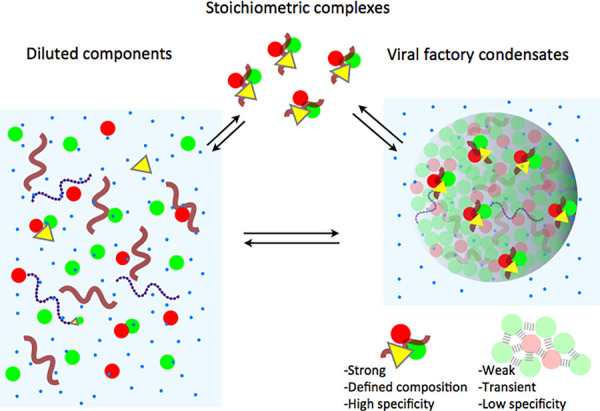
Complexes of different molecularity between the replication/transcription machinery exist in solution below their respective association constant. Above that level, defined stoichiometric complexes are formed, which are also present within the dense phase, coexisting with excess free components, particularly those acting as scaffold or drivers for LLPS. These complexes are held together by strong and specific interaction. Conversely, the interactions holding the condensate are weak and transient, required for a modulated assembly/disassembly, and display low specificity. These low affinity interactions take place only in the condensate, where concentrations can be a few orders of magnitude higher than those in the diluted cytosolic phase. LLPS, liquid–liquid phase separation.
Conclusions and prospects
Virus genomes encode a higher proportion of disordered proteins than eukaryotic or bacterial genomes, which often interact with a number of cellular partners, implying multiple weak and nonspecific interactions. Viruses also require a number of cellular proteins for their perpetuation and the ability of viral proteins to establish multiple dynamic and weak interactions linked to the formation of phase-separated condensates provide a mechanism for as yet unidentified host proteins partitioning together with the viral machinery. The validation of the liquid nature of VFs has been already established, and the reconstitution of condensates from its essential components in vitro emerges as a fundamental tool to understand the underlying mechanisms that can be ultimately probed by reverse genetics and a battery of techniques being developed for investigating BMCs in cellulo. These systems not only provide insights into gene function across viral families but also contribute to the understanding of fundamental mechanisms of LLPS-based biomolecular condensation. They also provide valuable models for the understanding of BMC in cellular transcription and replication, manifested in changes in chromatin and the condensed nature of superenhancers. Finally, LLPS-associated pathological processes emerge as novel therapeutic targets, which definitely involves a new approach for antiviral discovery, complementing classical drug design and screening [135,136].
Acknowledgments
We are grateful to Gabriela Noval for critical reading of the manuscript.
Funding Statement
This work was supported by Argentine National Agency for the Promotion of Science and Technology (ANPCyT, PICT-2019-2019-03295 to Gonzalo de Prat-Gay). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Kim J, Lee H, Lee HG, Seo PJ. Get closer and make hotspots: liquid-liquid phase separation in plants. EMBO Rep. 2021;22(5):e51656. doi: 10.15252/embr.202051656 ; PMCID: PMC8097390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fromm SA, Kamenz J, Noldeke ER, Neu A, Zocher G, Sprangers R. In vitro reconstitution of a cellular phase-transition process that involves the mRNA decapping machinery. Angew Chem Int Ed Engl. 2014;53(28):7354–9. doi: 10.1002/anie.201402885 ; PMCID: PMC4320757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Azaldegui CA, Vecchiarelli AG, Biteen JS. The emergence of phase separation as an organizing principle in bacteria. Biophys J. 2021;120(7):1123–38. doi: 10.1016/j.bpj.2020.09.023 ; PMCID: PMC8059088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wheeler RJ, Hyman AA. Controlling compartmentalization by non-membrane-bound organelles. Philos Trans R Soc Lond B Biol Sci. 2018;373(1747):20170193. doi: 10.1098/rstb.2017.0193 ; PMCID: PMC5904305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Banani SF, Lee HO, Hyman AA, Rosen MK. Biomolecular condensates: organizers of cellular biochemistry. Nat Rev Mol Cell Biol. 2017;18(5):285–98. doi: 10.1038/nrm.2017.7 ; PMCID: PMC7434221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Shin Y, Brangwynne CP. Liquid phase condensation in cell physiology and disease. Science. 2017;357(6357):eaaf4382. doi: 10.1126/science.aaf4382 . [DOI] [PubMed] [Google Scholar]
- 7.Mitrea DM, Kriwacki RW. Phase separation in biology; functional organization of a higher order. Cell Commun Signal. 2016;14:1. doi: 10.1186/s12964-015-0125-7 ; PMCID: PMC4700675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ismail H, Liu X, Yang F, Li J, Zahid A, Dou Z, et al. Mechanisms and regulation underlying membraneless organelle plasticity control. J Mol Cell Biol. 2021;Apr 29:mjab028. doi: 10.1093/jmcb/mjab028 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Moss B. Poxvirus DNA replication. Cold Spring Harb Perspect Biol. 2013;5(9):a010199. doi: 10.1101/cshperspect.a010199 ; PMCID: PMC3753712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Schramm B, Locker JK. Cytoplasmic organization of POXvirus DNA replication. Traffic. 2005;6(10):839–46. doi: 10.1111/j.1600-0854.2005.00324.x . [DOI] [PubMed] [Google Scholar]
- 11.Everett RD. The spatial organization of DNA virus genomes in the nucleus. PLoS Pathog. 2013;9(6):e1003386. doi: 10.1371/journal.ppat.1003386 ; PMCID: PMC3694854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hidalgo P, Gonzalez RA. Formation of adenovirus DNA replication compartments. FEBS Lett. 2019;593(24):3518–30. doi: 10.1002/1873-3468.13672 . [DOI] [PubMed] [Google Scholar]
- 13.Strang BL. Viral and cellular subnuclear structures in human cytomegalovirus-infected cells. J Gen Virol. 2015;96(Pt 2):239–52. doi: 10.1099/vir.0.071084-0 pntd.0009788. [DOI] [PubMed] [Google Scholar]
- 14.Charman M, Weitzman MD. Replication Compartments of DNA Viruses in the Nucleus: Location, Location, Location. Viruses. 2020;12(2):151. doi: 10.3390/v12020151 ; PMCID: PMC7077188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Maruri-Avidal L, Lopez S, Arias CF. Endoplasmic reticulum chaperones are involved in the morphogenesis of rotavirus infectious particles. J Virol. 2008;82(11):5368–80. doi: 10.1128/JVI.02751-07 ; PMCID: PMC2395214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cheung W, Gill M, Esposito A, Kaminski CF, Courousse N, Chwetzoff S, et al. Rotaviruses associate with cellular lipid droplet components to replicate in viroplasms, and compounds disrupting or blocking lipid droplets inhibit viroplasm formation and viral replication. J Virol. 2010;84(13):6782–98. doi: 10.1128/JVI.01757-09 ; PMCID: PMC2903253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dhillon P, Tandra VN, Chorghade SG, Namsa ND, Sahoo L, Rao CD. Cytoplasmic Relocalization and Colocalization with Viroplasms of Host Cell Proteins, and Their Role in Rotavirus Infection. J Virol. 2018;92(15):e00612–8. doi: 10.1128/JVI.00612-18 ; PMCID: PMC6052293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Garces Suarez Y, Martinez JL, Torres Hernandez D, Hernandez HO, Perez-Delgado A, Mendez M, et al. Nanoscale organization of rotavirus replication machineries. Elife. 2019;8:e42906. doi: 10.7554/eLife.42906 ; PMCID: PMC6692110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Whelan SPJ. Viral Replication Strategies. In: Knipe DM, Howley PM, editors. Fields Virology. 6th ed. Philadelphia: Lippincott, Williams & Wilkins; 2013. p. 105–126. [Google Scholar]
- 20.Davies KA, Chadwick B, Hewson R, Fontana J, Mankouri J, Barr JN. The RNA Replication Site of Tula Orthohantavirus Resides within a Remodelled Golgi Network. Cells. 2020;9(7):1569. doi: 10.3390/cells9071569 ; PMCID: PMC7408811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Wu X, Qi X, Liang M, Li C, Cardona CJ, Li D, et al. Roles of viroplasm-like structures formed by nonstructural protein NSs in infection with severe fever with thrombocytopenia syndrome virus. FASEB J. 2014;28(6):2504–16. doi: 10.1096/fj.13-243857 . [DOI] [PubMed] [Google Scholar]
- 22.Baird NL, York J, Nunberg JH. Arenavirus infection induces discrete cytosolic structures for RNA replication. J Virol. 2012;86(20):11301–10. doi: 10.1128/JVI.01635-12 ; PMCID: PMC3457164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ma D, George CX, Nomburg JL, Pfaller CK, Cattaneo R, Samuel CE. Upon Infection, Cellular WD Repeat-Containing Protein 5 (WDR5) Localizes to Cytoplasmic Inclusion Bodies and Enhances Measles Virus Replication. J Virol. 2018;92(5):e01726–17. doi: 10.1128/JVI.01726-17 ; PMCID: PMC5809725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Neufeldt CJ, Cortese M, Acosta EG, Bartenschlager R. Rewiring cellular networks by members of the Flaviviridae family. Nat Rev Microbiol. 2018;16(3):125–42. doi: 10.1038/nrmicro.2017.170 ; PMCID: PMC7097628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Cortese M, Goellner S, Acosta EG, Neufeldt CJ, Oleksiuk O, Lampe M, et al. Ultrastructural Characterization of Zika Virus Replication Factories. Cell Rep. 2017;18(9):2113–23. doi: 10.1016/j.celrep.2017.02.014 ; PMCID: PMC5340982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ci Y, Shi L. Compartmentalized replication organelle of flavivirus at the ER and the factors involved. Cell Mol Life Sci. 2021;Apr 12:1–16. doi: 10.1007/s00018-020-03576-x ; PMCID: PMC8041242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Belov GA, Nair V, Hansen BT, Hoyt FH, Fischer ER, Ehrenfeld E. Complex dynamic development of poliovirus membranous replication complexes. J Virol. 2012;86(1):302–12. doi: 10.1128/JVI.05937-11 ; PMCID: PMC3255921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Knoops K, Barcena M, Limpens RW, Koster AJ, Mommaas AM, Snijder EJ. Ultrastructural characterization of arterivirus replication structures: reshaping the endoplasmic reticulum to accommodate viral RNA synthesis. J Virol. 2012;86(5):2474–87. doi: 10.1128/JVI.06677-11 ; PMCID: PMC3302280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Knoops K, Kikkert M, Worm SH, Zevenhoven-Dobbe JC, van der Meer Y, Koster AJ, et al. SARS-coronavirus replication is supported by a reticulovesicular network of modified endoplasmic reticulum. PLoS Biol. 2008;6(9):e226. doi: 10.1371/journal.pbio.0060226 ; PMCID: PMC2535663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wolff G, Limpens R, Zevenhoven-Dobbe JC, Laugks U, Zheng S, de Jong AWM, et al. A molecular pore spans the double membrane of the coronavirus replication organelle. Science. 2020;369(6509):1395–8. doi: 10.1126/science.abd3629 ; PMCID: PMC7665310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Klein S, Cortese M, Winter SL, Wachsmuth-Melm M, Neufeldt CJ, Cerikan B, et al. SARS-CoV-2 structure and replication characterized by in situ cryo-electron tomography. Nat Commun. 2020;11(1):5885. doi: 10.1038/s41467-020-19619-7 ; PMCID: PMC7676268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Diaz A, Wang X. Bromovirus-induced remodeling of host membranes during viral RNA replication. Curr Opin Virol. 2014;9:104–10. doi: 10.1016/j.coviro.2014.09.018 . [DOI] [PubMed] [Google Scholar]
- 33.Nagy PD, Feng Z. Tombusviruses orchestrate the host endomembrane system to create elaborate membranous replication organelles. Curr Opin Virol. 2021;48:30–41. doi: 10.1016/j.coviro.2021.03.007 . [DOI] [PubMed] [Google Scholar]
- 34.Pagliari F, Marafioti MG, Genard G, Candeloro P, Viglietto G, Seco J, et al. ssRNA Virus and Host Lipid Rearrangements: Is There a Role for Lipid Droplets in SARS-CoV-2 Infection? Front Mol Biosci. 2020;7:578964. doi: 10.3389/fmolb.2020.578964 ; PMCID: PMC7579428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Monson EA, Trenerry AM, Laws JL, Mackenzie JM, Helbig KJ. Lipid droplets and lipid mediators in viral infection and immunity. FEMS Microbiol Rev. 2021. Jan 29:fuaa066. doi: 10.1093/femsre/fuaa066 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Dias SSG, Soares VC, Ferreira AC, Sacramento CQ, Fintelman-Rodrigues N, Temerozo JR, et al. Lipid droplets fuel SARS-CoV-2 replication and production of inflammatory mediators. PLoS Pathog. 2020;16(12):e1009127. doi: 10.1371/journal.ppat.1009127 ; PMCID: PMC7773323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Samsa MM, Mondotte JA, Iglesias NG, Assuncao-Miranda I, Barbosa-Lima G, Da Poian AT, et al. Dengue virus capsid protein usurps lipid droplets for viral particle formation. PLoS Pathog. 2009;5(10):e1000632. doi: 10.1371/journal.ppat.1000632 ; PMCID: PMC2760139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Flory PJ. Fundamental principles of condensation polymerization. Chem Rev. 1946;39:137–97. doi: 10.1021/cr60122a003 [DOI] [PubMed] [Google Scholar]
- 39.Ellis RJ. Macromolecular crowding: an important but neglected aspect of the intracellular environment. Curr Opin Struct Biol. 2001;11(1):114–9. doi: 10.1016/s0959-440x(00)00172-x . [DOI] [PubMed] [Google Scholar]
- 40.Alberti S, Dormann D. Liquid-Liquid Phase Separation in Disease. Annu Rev Genet. 2019;53:171–94. doi: 10.1146/annurev-genet-112618-043527 . [DOI] [PubMed] [Google Scholar]
- 41.Alberti S, Gladfelter A, Mittag T. Considerations and Challenges in Studying Liquid-Liquid Phase Separation and Biomolecular Condensates. Cell. 2019;176(3):419–34. doi: 10.1016/j.cell.2018.12.035 ; PMCID: PMC6445271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Mitrea DM, Chandra B, Ferrolino MC, Gibbs EB, Tolbert M, White MR, et al. Methods for Physical Characterization of Phase-Separated Bodies and Membrane-less Organelles. J Mol Biol. 2018;430(23):4773–805. doi: 10.1016/j.jmb.2018.07.006 ; PMCID: PMC6503534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Xue B, Williams RW, Oldfield CJ, Goh GK, Dunker AK, Uversky VN. Viral disorder or disordered viruses: do viral proteins possess unique features? Protein Pept Lett. 2010;17(8):932–51. doi: 10.2174/092986610791498984 . [DOI] [PubMed] [Google Scholar]
- 44.Brocca S, Grandori R, Longhi S, Uversky V. Liquid-Liquid Phase Separation by Intrinsically Disordered Protein Regions of Viruses: Roles in Viral Life Cycle and Control of Virus-Host Interactions. Int J Mol Sci. 2020;21(23):9045. doi: 10.3390/ijms21239045 ; PMCID: PMC7730420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Nott TJ, Petsalaki E, Farber P, Jervis D, Fussner E, Plochowietz A, et al. Phase transition of a disordered nuage protein generates environmentally responsive membraneless organelles. Mol Cell. 2015;57(5):936–47. doi: 10.1016/j.molcel.2015.01.013 ; PMCID: PMC4352761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Das RK, Ruff KM, Pappu RV. Relating sequence encoded information to form and function of intrinsically disordered proteins. Curr Opin Struct Biol. 2015;32:102–12. doi: 10.1016/j.sbi.2015.03.008 ; PMCID: PMC4512920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Mitrea DM, Cika JA, Stanley CB, Nourse A, Onuchic PL, Banerjee PR, et al. Self-interaction of NPM1 modulates multiple mechanisms of liquid-liquid phase separation. Nat Commun. 2018;9(1):842. doi: 10.1038/s41467-018-03255-3 ; PMCID: PMC5827731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Stenstrom L, Mahdessian D, Gnann C, Cesnik AJ, Ouyang W, Leonetti MD, et al. Mapping the nucleolar proteome reveals a spatiotemporal organization related to intrinsic protein disorder. Mol Syst Biol. 2020;16(8):e9469. doi: 10.15252/msb.20209469 ; PMCID: PMC7397901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Elbaum-Garfinkle S, Kim Y, Szczepaniak K, Chen CC, Eckmann CR, Myong S, et al. The disordered P granule protein LAF-1 drives phase separation into droplets with tunable viscosity and dynamics. Proc Natl Acad Sci U S A. 2015;112(23):7189–94. doi: 10.1073/pnas.1504822112 ; PMCID: PMC4466716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Riggs CL, Kedersha N, Ivanov P, Anderson P. Mammalian stress granules and P bodies at a glance. J Cell Sci. 2020;133(16):jcs242487. doi: 10.1242/jcs.242487 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Fox AH, Nakagawa S, Hirose T, Bond CS. Paraspeckles: Where Long Noncoding RNA Meets Phase Separation. Trends Biochem Sci. 2018;43(2):124–35. doi: 10.1016/j.tibs.2017.12.001 . [DOI] [PubMed] [Google Scholar]
- 52.Lafontaine DLJ, Riback JA, Bascetin R, Brangwynne CP. The nucleolus as a multiphase liquid condensate. Nat Rev Mol Cell Biol. 2021;22(3):165–82. doi: 10.1038/s41580-020-0272-6 . [DOI] [PubMed] [Google Scholar]
- 53.Nielsen FC, Hansen HT, Christiansen J. RNA assemblages orchestrate complex cellular processes. BioEssays. 2016;38(7):674–81. doi: 10.1002/bies.201500175 ; PMCID: PMC5084767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Mittag T, Parker R. Multiple Modes of Protein-Protein Interactions Promote RNP Granule Assembly. J Mol Biol. 2018;430(23):4636–49. doi: 10.1016/j.jmb.2018.08.005 ; PMCID: PMC6204294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Serebreni L, Stark A. Insights into gene regulation: From regulatory genomic elements to DNA-protein and protein-protein interactions. Curr Opin Cell Biol. 2021;70:58–66. doi: 10.1016/j.ceb.2020.11.009 . [DOI] [PubMed] [Google Scholar]
- 56.Rippe K. Liquid-Liquid Phase Separation in Chromatin. Cold Spring Harb Perspect Biol. 2021. Jun 14:a040683. doi: 10.1101/cshperspect.a040683 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Rubinstein M, Colby RH. Polymer physics. Oxford University Press; 2003. ISBN-10: 0-19-852059-X [Google Scholar]
- 58.Cinar H, Fetahaj Z, Cinar S, Vernon RM, Chan HS, Winter RHA. Temperature, Hydrostatic Pressure, and Osmolyte Effects on Liquid-Liquid Phase Separation in Protein Condensates: Physical Chemistry and Biological Implications. Chemistry. 2019;25(57):13049–69. doi: 10.1002/chem.201902210 . [DOI] [PubMed] [Google Scholar]
- 59.Ruff KM, Dar F, Pappu RV. Ligand effects on phase separation of multivalent macromolecules. Proc Natl Acad Sci U S A. 2021;118(10):e2017184118. doi: 10.1073/pnas.2017184118 ; PMCID: PMC7958451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zhou Q, Usluer S, Zhang F, Lenard AJ, Bourgeois BMR, Madl T. ATP regulates RNA-driven cold inducible RNA binding protein phase separation. Protein Sci. 2021;30(7):1438–53. doi: 10.1002/pro.4123 ; PMCID: PMC8197425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Bratek-Skicki A, Pancsa R, Meszaros B, Van Lindt J, Tompa P. A guide to regulation of the formation of biomolecular condensates. FEBS J. 2020;287(10):1924–35. doi: 10.1111/febs.15254 . [DOI] [PubMed] [Google Scholar]
- 62.Woodruff JB, Hyman AA, Boke E. Organization and Function of Non-dynamic Biomolecular Condensates. Trends Biochem Sci. 2018;43(2):81–94. doi: 10.1016/j.tibs.2017.11.005 . [DOI] [PubMed] [Google Scholar]
- 63.Patel A, Lee HO, Jawerth L, Maharana S, Jahnel M, Hein MY, et al. A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation. Cell. 2015;162(5):1066–77. doi: 10.1016/j.cell.2015.07.047 . [DOI] [PubMed] [Google Scholar]
- 64.Ray S, Singh N, Kumar R, Patel K, Pandey S, Datta D, et al. alpha-Synuclein aggregation nucleates through liquid-liquid phase separation. Nat Chem. 2020;12(8):705–16. doi: 10.1038/s41557-020-0465-9 . [DOI] [PubMed] [Google Scholar]
- 65.Brangwynne CP, Mitchison TJ, Hyman AA. Active liquid-like behavior of nucleoli determines their size and shape in Xenopus laevis oocytes. Proc Natl Acad Sci U S A. 2011;108(11):4334–9. doi: 10.1073/pnas.1017150108 ; PMCID: PMC3060270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Kroschwald S, Maharana S, Mateju D, Malinovska L, Nuske E, Poser I, et al. Promiscuous interactions and protein disaggregases determine the material state of stress-inducible RNP granules. elife. 2015;4:e06807. doi: 10.7554/eLife.06807 ; PMCID: PMC4522596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Hubstenberger A, Noble SL, Cameron C, Evans TC. Translation repressors, an RNA helicase, and developmental cues control RNP phase transitions during early development. Dev Cell. 2013;27(2):161–73. doi: 10.1016/j.devcel.2013.09.024 ; PMCID: PMC3869996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Su JM, Wilson MZ, Samuel CE, Ma D. Formation and Function of Liquid-Like Viral Factories in Negative-Sense Single-Stranded RNA Virus Infections. Viruses. 2021;13(1):126. doi: 10.3390/v13010126 ; PMCID: PMC7835873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Amarasinghe GK, Arechiga Ceballos NG, Banyard AC, Basler CF, Bavari S, Bennett AJ, et al. Taxonomy of the order Mononegavirales: update 2018. Arch Virol. 2018;163(8):2283–94. doi: 10.1007/s00705-018-3814-x ; PMCID: PMC6076851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Luo M, Terrell JR, McManus SA. Nucleocapsid Structure of Negative Strand RNA Virus. Viruses. 2020;12(8):835. doi: 10.3390/v12080835 ; PMCID: PMC7472042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Cardone C, Caseau CM, Pereira N, Sizun C. Pneumoviral Phosphoprotein, a Multidomain Adaptor-Like Protein of Apparent Low Structural Complexity and High Conformational Versatility. Int J Mol Sci. 2021;22(4):1537. doi: 10.3390/ijms22041537 ; PMCID: PMC7913705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Gerard FC, Ribeiro Ede A Jr, Leyrat C, Ivanov I, Blondel D, Longhi S, et al. Modular organization of rabies virus phosphoprotein. J Mol Biol. 2009;388(5):978–96. doi: 10.1016/j.jmb.2009.03.061 . [DOI] [PubMed] [Google Scholar]
- 73.Nikolic J, Lagaudriere-Gesbert C, Scrima N, Blondel D, Gaudin Y. Structure and Function of Negri Bodies. Adv Exp Med Biol. 2019;1215:111–27. doi: 10.1007/978-3-030-14741-9_6 . [DOI] [PubMed] [Google Scholar]
- 74.Kristensson K, Dastur DK, Manghani DK, Tsiang H, Bentivoglio M. Rabies: interactions between neurons and viruses. A review of the history of Negri inclusion bodies. Neuropathol Appl Neurobiol. 1996;22(3):179–87. . [PubMed] [Google Scholar]
- 75.Fouquet B, Nikolic J, Larrous F, Bourhy H, Wirblich C, Lagaudriere-Gesbert C, et al. Focal adhesion kinase is involved in rabies virus infection through its interaction with viral phosphoprotein P. J Virol. 2015;89(3):1640–51. doi: 10.1128/JVI.02602-14 ; PMCID: PMC4300764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Lahaye X, Vidy A, Pomier C, Obiang L, Harper F, Gaudin Y, et al. Functional characterization of Negri bodies (NBs) in rabies virus-infected cells: Evidence that NBs are sites of viral transcription and replication. J Virol. 2009;83(16):7948–58. doi: 10.1128/JVI.00554-09 ; PMCID: PMC2715764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Zhang J, Wu X, Zan J, Wu Y, Ye C, Ruan X, et al. Cellular chaperonin CCTgamma contributes to rabies virus replication during infection. J Virol. 2013;87(13):7608–21. doi: 10.1128/JVI.03186-12 ; PMCID: PMC3700271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Nikolic J, Le Bars R, Lama Z, Scrima N, Lagaudriere-Gesbert C, Gaudin Y, et al. Negri bodies are viral factories with properties of liquid organelles. Nat Commun. 2017;8(1):58. doi: 10.1038/s41467-017-00102-9 ; PMCID: PMC5498545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Heinrich BS, Maliga Z, Stein DA, Hyman AA, Whelan SPJ. Phase Transitions Drive the Formation of Vesicular Stomatitis Virus Replication Compartments. mBio. 2018;9(5):e02290–17. doi: 10.1128/mBio.02290-17 ; PMCID: PMC6123442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Zhou Y, Su JM, Samuel CE, Ma D. Measles Virus Forms Inclusion Bodies with Properties of Liquid Organelles. J Virol. 2019;93(21):e00948–19. doi: 10.1128/JVI.00948-19 ; PMCID: PMC6803276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Cifuentes-Muñoz N, Branttie J, Slaughter KB, Dutch RE. Human Metapneumovirus Induces Formation of Inclusion Bodies for Efficient Genome Replication and Transcription. J Virol. 2017. Nov 30;91(24):e01282–17. doi: 10.1128/JVI.01282-17 ; PMCID: PMC5709606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ito M, Iwasaki M, Takeda M, Nakamura T, Yanagi Y, Ohno S. Measles virus nonstructural C protein modulates viral RNA polymerase activity by interacting with host protein SHCBP1. J Virol. 2013;87(17):9633–42. doi: 10.1128/JVI.00714-13 ; PMCID: PMC3754092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Tran TL, Castagné N, Dubosclard V, Noinville S, Koch E, Moudjou M, et al. The respiratory syncytial virus M2-1 protein forms tetramers and interacts with RNA and P in a competitive manner. J Virol. 2009. Jul;83(13):6363–74. doi: 10.1128/JVI.00335-09 ; PMCID: PMC2698528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Fearns R, Collins PL. Role of the M2-1 transcription antitermination protein of respiratory syncytial virus in sequential transcription. J Virol. 1999. Jul;73(7):5852–64. doi: 10.1128/JVI.73.7.5852-5864.1999 ; PMCID: PMC112646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Rincheval V, Lelek M, Gault E, Bouillier C, Sitterlin D, Blouquit-Laye S, et al. Functional organization of cytoplasmic inclusion bodies in cells infected by respiratory syncytial virus. Nat Commun. 2017;8(1):563. doi: 10.1038/s41467-017-00655-9 ; PMCID: PMC5601476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Ghildyal R, Mills J, Murray M, Vardaxis N, Meanger J. Respiratory syncytial virus matrix protein associates with nucleocapsids in infected cells. J Gen Virol. 2002;83(Pt 4):753–7. doi: 10.1099/0022-1317-83-4-753 . [DOI] [PubMed] [Google Scholar]
- 87.Nevers Q, Albertini AA, Lagaudriere-Gesbert C, Gaudin Y. Negri bodies and other virus membrane-less replication compartments. Biochim Biophys Acta, Mol Cell Res. 2020;1867(12):118831. doi: 10.1016/j.bbamcr.2020.118831 ; PMCID: PMC7442162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.García J, García-Barreno B, Vivo A, Melero JA. Cytoplasmic inclusions of respiratory syncytial virus-infected cells: formation of inclusion bodies in transfected cells that coexpress the nucleoprotein, the phosphoprotein, and the 22K protein. Virology. 1993. Jul;195(1):243–7. doi: 10.1006/viro.1993.1366 . [DOI] [PubMed] [Google Scholar]
- 89.Galloux M, Risso-Ballester J, Richard CA, Fix J, Rameix-Welti MA, Eleouet JF. Minimal Elements Required for the Formation of Respiratory Syncytial Virus Cytoplasmic Inclusion Bodies In Vivo and In Vitro. mBio. 2020;11(5):e01202–20. doi: 10.1128/mBio.01202-20 ; PMCID: PMC7512546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Miyake T, Farley CM, Neubauer BE, Beddow TP, Hoenen T, Engel DA. Ebola Virus Inclusion Body Formation and RNA Synthesis Are Controlled by a Novel Domain of Nucleoprotein Interacting with VP35. J Virol. 2020;94(16):e02100–19. doi: 10.1128/JVI.02100-19 ; PMCID: PMC7394894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Matsumoto Y, Hayashi Y, Omori H, Honda T, Daito T, Horie M, et al. Bornavirus closely associates and segregates with host chromosomes to ensure persistent intranuclear infection. Cell Host Microbe. 2012;11(5):492–503. doi: 10.1016/j.chom.2012.04.009 . [DOI] [PubMed] [Google Scholar]
- 92.Charlier CM, Wu YJ, Allart S, Malnou CE, Schwemmle M, Gonzalez-Dunia D. Analysis of borna disease virus trafficking in live infected cells by using a virus encoding a tetracysteine-tagged p protein. J Virol. 2013;87(22):12339–48. doi: 10.1128/JVI.01127-13 ; PMCID: PMC3807882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Chou YY, Heaton NS, Gao Q, Palese P, Singer RH, Lionnet T. Colocalization of different influenza viral RNA segments in the cytoplasm before viral budding as shown by single-molecule sensitivity FISH analysis. PLoS Pathog. 2013;9(5):e1003358. doi: 10.1371/journal.ppat.1003358 ; PMCID: PMC3649991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Lakdawala SS, Fodor E, Subbarao K. Moving On Out: Transport and Packaging of Influenza Viral RNA into Virions. Annu Rev Virol. 2016;3(1):411–27. doi: 10.1146/annurev-virology-110615-042345 . [DOI] [PubMed] [Google Scholar]
- 95.Alenquer M, Vale-Costa S, Etibor TA, Ferreira F, Sousa AL, Amorim MJ. Influenza A virus ribonucleoproteins form liquid organelles at endoplasmic reticulum exit sites. Nat Commun. 2019;10(1):1629. doi: 10.1038/s41467-019-09549-4 ; PMCID: PMC6456594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Jouvenet N, Simon SM, Bieniasz PD. Visualizing HIV-1 assembly. J Mol Biol. 2011;410(4):501–11. doi: 10.1016/j.jmb.2011.04.062 ; PMCID: PMC3144478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Rensen E, Mueller F, Scoca V, Parmar JJ, Souque P, Zimmer C, et al. Clustering and reverse transcription of HIV-1 genomes in nuclear niches of macrophages. EMBO J. 2021;40(1):e105247. doi: 10.15252/embj.2020105247 ; PMCID: PMC7780146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Scoca V, Di Nunzio F. Membraneless organelles restructured and built by pandemic viruses: HIV-1 and SARS-CoV-2. J Mol Cell Biol. 2021;Mar 24:mjab020. doi: 10.1093/jmcb/mjab020 ; PMCID: PMC8083626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Monette A, Niu M, Chen L, Rao S, Gorelick RJ, Mouland AJ. Pan-retroviral Nucleocapsid-Mediated Phase Separation Regulates Genomic RNA Positioning and Trafficking. Cell Rep. 2020;31(3):107520. doi: 10.1016/j.celrep.2020.03.084 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Chang L, Godinez WJ, Kim IH, Tektonidis M, de Lanerolle P, Eils R, et al. Herpesviral replication compartments move and coalesce at nuclear speckles to enhance export of viral late mRNA. Proc Natl Acad Sci U S A. 2011;108(21):E136–44. doi: 10.1073/pnas.1103411108 ; PMCID: PMC3102408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Tomer E, Cohen EM, Drayman N, Afriat A, Weitzman MD, Zaritsky A, et al. Coalescing replication compartments provide the opportunity for recombination between coinfecting herpesviruses. FASEB J. 2019. Aug;33(8):9388–403. doi: 10.1096/fj.201900032R ; PMCID: PMC6662979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Seyffert M, Georgi F, Tobler K, Bourqui L, Anfossi M, Michaelsen K, et al. The HSV-1 Transcription Factor ICP4 Confers Liquid-Like Properties to Viral Replication Compartments. Int J Mol Sci. 2021;22(9):4447. doi: 10.3390/ijms22094447 ; PMCID: PMC8123221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.McSwiggen DT, Hansen AS, Teves SS, Marie-Nelly H, Hao Y, Heckert AB, et al. Evidence for DNA-mediated nuclear compartmentalization distinct from phase separation. Elife. 2019;8:e47098. doi: 10.7554/eLife.47098 ; PMCID: PMC6522219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Metrick CM, Koenigsberg AL, Heldwein EE. Conserved Outer Tegument Component UL11 from Herpes Simplex Virus 1 Is an Intrinsically Disordered, RNA-Binding Protein. mBio. 2020;11(3):e00810–20. doi: 10.1128/mBio.00810-20 ; PMCID: PMC7403781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Peng Q, Wang L, Qin Z, Wang J, Zheng X, Wei L, et al. Phase Separation of Epstein-Barr Virus EBNA2 and Its Coactivator EBNALP Controls Gene Expression. J Virol. 2020;94(7):e01771–19. doi: 10.1128/JVI.01771-19 ; PMCID: PMC7081900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Vladimirova O, De Leo A, Deng Z, Wiedmer A, Hayden J, Lieberman PM. Phase separation and DAXX redistribution contribute to LANA nuclear body and KSHV genome dynamics during latency and reactivation. PLoS Pathog. 2021;17(1):e1009231. doi: 10.1371/journal.ppat.1009231 ; PMCID: PMC7943007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Sakakibara N, Mitra R, McBride AA. The papillomavirus E1 helicase activates a cellular DNA damage response in viral replication foci. J Virol. 2011;85(17):8981–95. doi: 10.1128/JVI.00541-11 ; PMCID: PMC3165833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Smal C, Wetzler DE, Dantur KI, Chemes LB, Garcia-Alai MM, Dellarole M, et al. The human papillomavirus E7-E2 interaction mechanism in vitro reveals a finely tuned system for modulating available E7 and E2 proteins. Biochemistry. 2009;48(50):11939–49. doi: 10.1021/bi901415k . [DOI] [PubMed] [Google Scholar]
- 109.Ferreiro DU, Lima LM, Nadra AD, Alonso LG, Goldbaum FA, de Prat-Gay G. Distinctive cognate sequence discrimination, bound DNA conformation, and binding modes in the E2 C-terminal domains from prototype human and bovine papillomaviruses. Biochemistry. 2000;39(47):14692–701. doi: 10.1021/bi001694r . [DOI] [PubMed] [Google Scholar]
- 110.Cho WK, Spille JH, Hecht M, Lee C, Li C, Grube V, et al. Mediator and RNA polymerase II clusters associate in transcription-dependent condensates. Science. 2018;361(6400):412–5. doi: 10.1126/science.aar4199 ; PMCID: PMC6543815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Sabari BR, Dall’Agnese A, Boija A, Klein IA, Coffey EL, Shrinivas K, et al. Coactivator condensation at super-enhancers links phase separation and gene control. Science. 2018;361(6400):eaar3958. doi: 10.1126/science.aar3958 ; PMCID: PMC6092193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Dooley KE, Warburton A, McBride AA. Tandemly Integrated HPV16 Can Form a Brd4-Dependent Super-Enhancer-Like Element That Drives Transcription of Viral Oncogenes. mBio. 2016;7(5):e01446–16. doi: 10.1128/mBio.01446-16 ; PMCID: PMC5021809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Masters PS. Coronavirus genomic RNA packaging. Virology. 2019;537:198–207. doi: 10.1016/j.virol.2019.08.031 ; PMCID: PMC7112113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Carlson CR, Asfaha JB, Ghent CM, Howard CJ, Hartooni N, Safari M, et al. Phosphoregulation of Phase Separation by the SARS-CoV-2 N Protein Suggests a Biophysical Basis for its Dual Functions. Mol Cell. 2020;80(6):1092–103. e4. doi: 10.1016/j.molcel.2020.11.025 PMCID: PMC7677695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Savastano A, Ibanez de Opakua A, Rankovic M, Zweckstetter M. Nucleocapsid protein of SARS-CoV-2 phase separates into RNA-rich polymerase-containing condensates. Nat Commun. 2020;11(1):6041. doi: 10.1038/s41467-020-19843-1 ; PMCID: PMC7699647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Perdikari TM, Murthy AC, Ryan VH, Watters S, Naik MT, Fawzi NL. SARS-CoV-2 nucleocapsid protein phase-separates with RNA and with human hnRNPs. EMBO J. 2020;39(24):e106478. doi: 10.15252/embj.2020106478 ; PMCID: PMC7737613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Iserman C, Roden CA, Boerneke MA, Sealfon RSG, McLaughlin GA, Jungreis I, et al. Genomic RNA Elements Drive Phase Separation of the SARS-CoV-2 Nucleocapsid. Mol Cell. 2020;80(6):1078–91. e6. doi: 10.1016/j.molcel.2020.11.041 ; PMCID: PMC7691212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Cubuk J, Alston JJ, Incicco JJ, Singh S, Stuchell-Brereton MD, Ward MD, et al. The SARS-CoV-2 nucleocapsid protein is dynamic, disordered, and phase separates with RNA. Nat Commun. 2021;12(1):1936. doi: 10.1038/s41467-021-21953-3 ; PMCID: PMC8007728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Lu S, Ye Q, Singh D, Cao Y, Diedrich JK, Yates JR 3rd, et al. The SARS-CoV-2 nucleocapsid phosphoprotein forms mutually exclusive condensates with RNA and the membrane-associated M protein. Nat Commun. 2021;12(1):502. doi: 10.1038/s41467-020-20768-y ; PMCID: PMC7820290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Banerjee PR, Milin AN, Moosa MM, Onuchic PL, Deniz AA. Reentrant Phase Transition Drives Dynamic Substructure Formation in Ribonucleoprotein Droplets. Angew Chem Int Ed Engl. 2017;56(38):11354–9. doi: 10.1002/anie.201703191 ; PMCID: PMC5647147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Wu CH, Chen PJ, Yeh SH. Nucleocapsid phosphorylation and RNA helicase DDX1 recruitment enables coronavirus transition from discontinuous to continuous transcription. Cell Host Microbe. 2014;16(4):462–72. doi: 10.1016/j.chom.2014.09.009 ; PMCID: PMC7104987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Dang M, Li Y, Song J. ATP biphasically modulates LLPS of SARS-CoV-2 nucleocapsid protein and specifically binds its RNA-binding domain. Biochem Biophys Res Commun. 2021;541:50–5. doi: 10.1016/j.bbrc.2021.01.018 ; PMCID: PMC7808732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Eiermann N, Haneke K, Sun Z, Stoecklin G, Ruggieri A. Dance with the Devil: Stress Granules and Signaling in Antiviral Responses. Viruses. 2020;12(9):984. doi: 10.3390/v12090984 ; PMCID: PMC7552005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Luo L, Li Z, Zhao T, Ju X, Ma P, Jin B, et al. SARS-CoV-2 nucleocapsid protein phase separates with G3BPs to disassemble stress granules and facilitate viral production. Sci Bull (Beijing). 2021;66(12):1194–204. doi: 10.1016/j.scib.2021.01.013 ; PMCID: PMC7816596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Wu Y, Ma L, Cai S, Zhuang Z, Zhao Z, Jin S, et al. RNA-induced liquid phase separation of SARS-CoV-2 nucleocapsid protein facilitates NF-kappaB hyper-activation and inflammation. Signal Transduct Target Ther. 2021;6(1):167. doi: 10.1038/s41392-021-00575-7 ; PMCID: PMC8065320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Zhao M, Yu Y, Sun LM, Xing JQ, Li T, Zhu Y, et al. GCG inhibits SARS-CoV-2 replication by disrupting the liquid phase condensation of its nucleocapsid protein. Nat Commun. 2021;12(1):2114. doi: 10.1038/s41467-021-22297-8 ; PMCID: PMC8035206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Peeples W, Rosen MK. Mechanistic dissection of increased enzymatic rate in a phase-separated compartment. Nat Chem Biol. 2021;17(6):693–702. doi: 10.1038/s41589-021-00801-x . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Wei MT, Chang YC, Shimobayashi SF, Shin Y, Strom AR, Brangwynne CP. Nucleated transcriptional condensates amplify gene expression. Nat Cell Biol. 2020;22(10):1187–96. doi: 10.1038/s41556-020-00578-6 . [DOI] [PubMed] [Google Scholar]
- 129.Noton SL, Fearns R. Initiation and regulation of paramyxovirus transcription and replication. Virology. 2015. May(479–480):545–54. doi: 10.1016/j.virol.2015.01.014 ; PMCID: PMC4424093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Plumet S, Duprex WP, Gerlier D. Dynamics of viral RNA synthesis during measles virus infection. J Virol. 2005. Jun;79(11):6900–8. doi: 10.1128/JVI.79.11.6900-6908.2005 ; PMCID: PMC1112129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Riedel C, Hennrich AA, Conzelmann KK. Components and Architecture of the Rhabdovirus Ribonucleoprotein Complex. Viruses. 2020;12(9):959. doi: 10.3390/v12090959 ; PMCID: PMC7552012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Kolesnikova L, Nanbo A, Becker S, Kawaoka Y. Inside the Cell: Assembly of Filoviruses. Curr Top Microbiol Immunol. 2017;411:353–80. doi: 10.1007/82_2017_15 . [DOI] [PubMed] [Google Scholar]
- 133.El Najjar F, Schmitt AP, Dutch RE. Paramyxovirus glycoprotein incorporation, assembly and budding: a three way dance for infectious particle production. Viruses. 2014. Aug 7;6(8):3019–54. doi: 10.3390/v6083019 ; PMCID: PMC4147685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Richard CA, Rincheval V, Lassoued S, Fix J, Cardone C, Esneau C, et al. RSV hijacks cellular protein phosphatase 1 to regulate M2-1 phosphorylation and viral transcription. PLoS Pathog. 2018;14(3):e1006920. doi: 10.1371/journal.ppat.1006920 ; PMCID: PMC5847313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Risso-Ballester J, Galloux M, Cao J, Le Goffic R, Hontonnou F, Jobart-Malfait A, et al. A condensate-hardening drug blocks RSV replication in vivo. Nature. 2021;595(7868):596–9. doi: 10.1038/s41586-021-03703-z . [DOI] [PubMed] [Google Scholar]
- 136.Wang S, Dai T, Qin Z, Pan T, Chu F, Lou L, et al. Targeting liquid-liquid phase separation of SARS-CoV-2 nucleocapsid protein promotes innate antiviral immunity by elevating MAVS activity. Nat Cell Biol. 2021;23(7):718–32. doi: 10.1038/s41556-021-00710-0 . [DOI] [PubMed] [Google Scholar]