

ABSTRACT
The hospital-acquired pathogen Acinetobacter baumannii possesses a complex cell envelope that is key to its multidrug resistance and virulence. The bacterium, however, lacks many canonical enzymes that build the envelope in model organisms. Instead, A. baumannii contains a number of poorly annotated proteins that may allow alternative mechanisms of envelope biogenesis. We demonstrated previously that one of these unusual proteins, ElsL, is required for maintaining a characteristic short rod shape and for withstanding antibiotics that attack the septal cell wall. Curiously, ElsL is composed of a leaderless YkuD-family domain usually found in secreted, cell wall-modifying l,d-transpeptidases (LDTs). Here, we show that, rather than being an LDT, ElsL is actually a new class of cytoplasmic l,d-carboxypeptidase (LDC) that provides a critical step in cell wall recycling previously thought to be missing from A. baumannii. Absence of ElsL impairs cell wall integrity, morphology, and intrinsic resistance due to buildup of murein tetrapeptide precursors, toxicity of which is bypassed by preventing muropeptide recycling. Multiple pathways in the cell become sites of vulnerability when ElsL is inactivated, including l,d-cross-link formation, cell division, and outer membrane lipid homoeostasis, reflecting its pleiotropic influence on envelope physiology. We thus reveal a novel class of cell wall-recycling LDC critical to growth and homeostasis of A. baumannii and likely many other bacteria.
KEYWORDS: Acinetobacter; l,d-carboxypeptidase; antibiotic resistance; cell wall recycling; morphology; peptidoglycan
INTRODUCTION
The Gram-negative pathogen Acinetobacter baumannii is a significant cause of health care-associated infections, including ventilator-associated pneumonia, bloodstream infections, urinary tract infections, and sepsis (1, 2). A. baumannii strains show widespread multidrug resistance, limiting the number of therapies effective against such infections (3). This problem is compounded by the insufficient pipeline of new antibiotics active against Gram-negative pathogens (4). Reflecting the urgency of this threat, the World Health Organization has ranked A. baumannii as a pathogen of highest priority for research and development of new antibiotics (4).
The distinct cell envelope of A. baumannii is a key potential target for new treatments, but many aspects of its synthesis and control are not understood. Building the critical peptidoglycan (PG) cell wall layer, in particular, appears to involve several unconventional and poorly defined strategies. A number of proteins integral to the canonical pathways of septal PG synthesis, cell separation, and PG recycling have no homologs in the microorganism (5). For instance, A. baumannii lacks an ortholog of Escherichia coli LdcA or Vibrio cholerae LdcV, l,d-carboxypeptidases (LDCs) necessary for reusing peptides derived from old PG for synthesis of new cell wall (6, 7).
In addition to lacking canonical enzymes, the pathogen encodes several proteins that are implicated in PG homeostasis based on domain annotations but whose actual functions remain mysterious (5). Prominent among this group are two YkuD-domain proteins, which we have named ElsL (ACX60_RS03475) and LdtAb (ACX60_RS05685) (5) (referred to as LdtK and LdtJ in reference 8). YkuD domains are found in secreted enzymes that modify the cell wall for a variety of purposes in other organisms (9). For example, many YkuD-domain proteins carry out l,d-transpeptidase (LDT) reactions that generate 3-3 cell wall cross-links. These bonds are usually less abundant than the 4-3 cross-links catalyzed by dd-transpeptidases (penicillin binding proteins [PBPs]) but may reinforce the wall against envelope stress (9). Unlike PBPs, LDTs are generally insensitive to β-lactam antibiotics (with the exception of carbapenems), and thus LDTs have been implicated in contributing to drug resistance (10, 11). LdtAb, but not ElsL, was recently found to be essential for 3-3 cross-links (8). Interestingly, ElsL does not contain a detectable secretion signal or additional domains such as PG binding motifs that would target the protein to the cell wall (5). Functional predictions based on identification of conserved domains in ElsL are therefore limited. A few leads were obtained based on genome-wide profiling of antibiotic susceptibility phenotypes (5). These studies found that ElsL deficiency is closely related to malfunction of the Rod system, the multiprotein PG synthetic machinery responsible for cell elongation. Mutations affecting ElsL and the Rod system both caused hypersensitivity to the same subset of β-lactam antibiotics as well as dramatic loss of the bacterium’s characteristic short rod shape (5). ElsL mutation was also associated with increased outer membrane (OM) shedding (8). The role ElsL plays in the cell and its link to Rod system function and the OM remain unknown.
In this paper, we have determined the function of ElsL in A. baumannii cell wall synthesis. We show that ElsL defines a novel, noncanonical class of cytoplasmic LDC essential to cell wall recycling. We have identified critical perturbations to cell wall metabolism and integrity that occur in the absence of this function. We also delineated the complete network of genetic interactions with elsL, revealing multiple cellular pathways that are affected by its inactivation.
RESULTS
ElsL and LdtAb have opposing effects on l,d-cross-link formation in A. baumannii.
To determine the effect of the two A. baumannii YkuD-family proteins, ElsL and LdtAb, on modifying the cell wall, we analyzed deletion mutants using two assays that detect PG remodeling: (i) metabolic cell wall labeling using the fluorescent precursor HCC-amino d-alanine (HADA), incorporation of which depends on an exchange reaction catalyzed by transpeptidases, including LDTs (12), and (ii) analysis of cell wall muropeptide composition by ultraperformance liquid chromatography-mass spectrometry (UPLC-MS) (13).
Wild-type (WT) bacteria incubated with HADA incorporated the label throughout their cell walls, with the population of dividing cells showing increased signal at the midcell (Fig. 1A and B). Deletion of ldtAb resulted in a dramatic loss of signal along the side-wall of cells that was reversed by reintroducing the gene in trans (Fig. 1A to D). In contrast, deletion of elsL, while causing loss of the short rod shape, caused no decrease in HADA incorporation (Fig. 1A and B). These phenotypes confirm previous findings with a related fluorescent label (8). The ΔldtAb phenotype also resembled that seen with E. coli completely lacking its 6 LDT paralogs (14).
FIG 1.
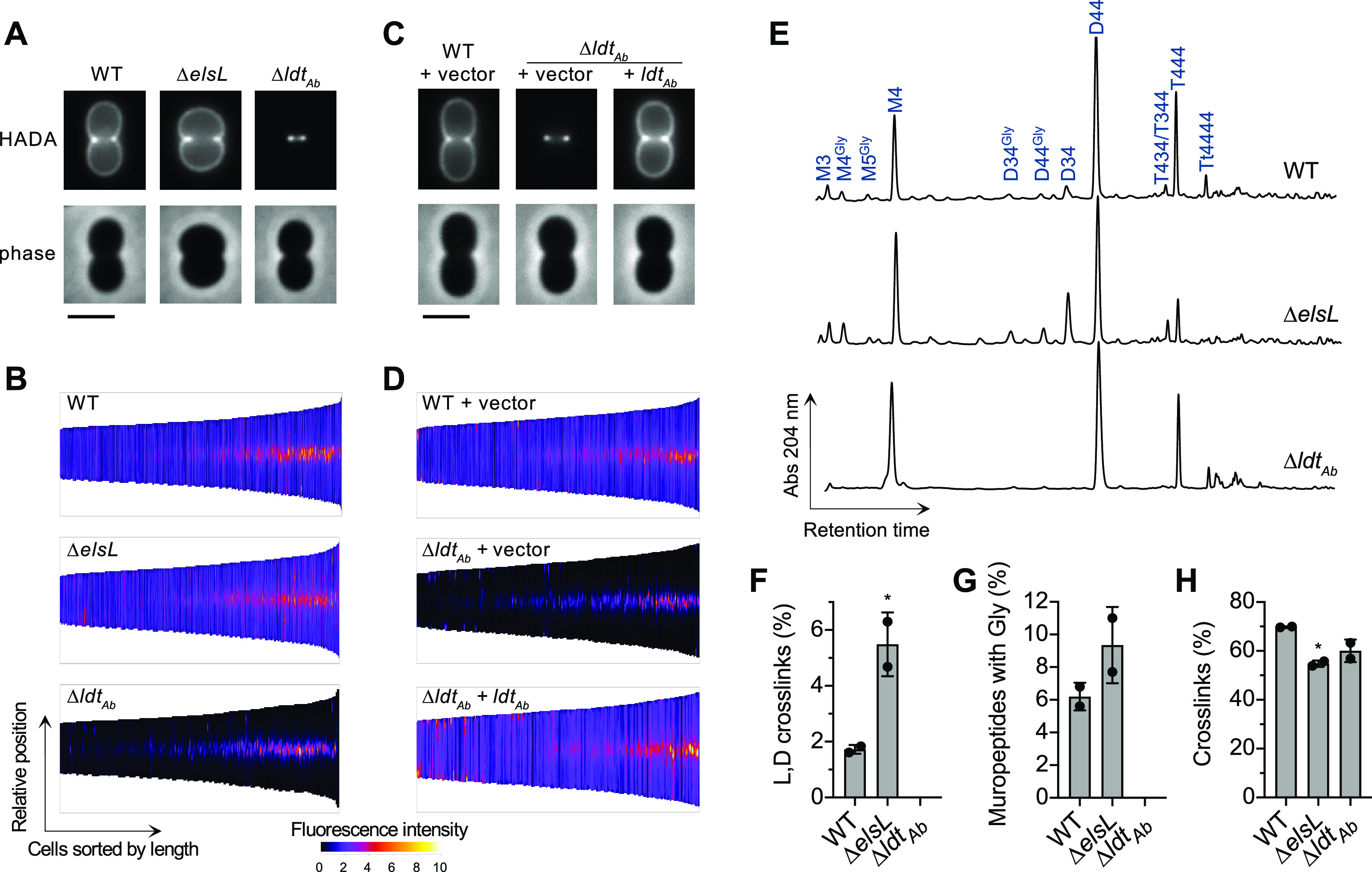
ElsL and LdtAb have opposing effects on l,d-cross-link formation. (A and C) Cells of the indicated A. baumannii strains were metabolically labeled with HADA and imaged by fluorescence and phase-contrast microscopy. Representative cells are shown. Scale bar, 2 μm. (B and D) Demographic representation of the cellular distribution and intensity of the HADA label. Cells (n ≥ 227 with each strain) were ordered according to their length and stacked by aligning cell midpoints. Fluorescence intensity along the medial axis of each cell is displayed as a heat map. (E) Muropeptide profile analysis. Major muropeptides and those showing differences between the WT and mutant are labeled. “M” indicates monomeric muropeptide; “D,” “T,” and “Tt” indicate dimeric, trimeric, or tetrameric cross-linked muropeptides, respectively; number(s) indicate the residue length of the stem peptide(s). “Gly” indicates that the terminal residue is a glycine. (F to H) The percentage of l,d-cross-links, glycine-containing muropeptides, and total cross-linked muropeptides were quantified. Bars show the mean ± standard deviation (SD) (n = 2 biological replicates). *, P < 0.05 in unpaired t test comparing ΔelsL versus WT (F and G) or in one-way analysis of variance (ANOVA) with Dunnett’s multiple-comparison test comparing each mutant to the WT (H).
Cell wall muropeptide profiling revealed changes consistent with the metabolic labeling experiments. The WT profile consisted of major and minor muropeptide peaks similar to those reported previously (Fig. 1E) (15–18). Deletion of ldtAb caused complete loss of LDT-generated muropeptides, including the D34 (l,d-cross-linked) dimer (Fig. 1E and F) and muropeptides containing a terminal glycine (fourth position) residue from d-alanine exchange (Fig. 1E and G), consistent with prior findings (8). In stark contrast, the ΔelsL strain showed an increase in the l,d-cross-linked D34 muropeptide compared to the WT (Fig. 1E and F; Fig. S1). Muropeptides with terminal glycines also appeared to increase in the ΔelsL mutant (Fig. 1E), although their overall levels were not significantly different from the WT based on unpaired t test results (Fig. 1G). The biological significance of these glycine residues in A. baumannii or other organisms (19, 20) is not well defined. Despite the increase in 3-3-cross-links, the ΔelsL cell wall showed lower total cross-linkage (including 4-3 as well as 3-3 bonds; Fig. 1H). Taken together, these data confirm that LdtAb is the LDT responsible for alternative cross-linking within the A. baumannii cell wall and indicate that ElsL is not a canonical LDT. Rather, ElsL has an alternative function that influences both 4-3 and 3-3 cross-link formation as well as cell shape.
UPLC/MS analysis of A. baumannii cell wall. Muropeptide fragments isolated from ΔelsL sacculi were identified by MS (table below chromatograms). Bold values in the table indicate the protonated state of the muropeptide more abundantly detected in the UPLC/MS analysis. The corresponding peaks were labelled (blue numbers) according to their muropeptide identity in the ΔelsL UPLC chromatogram (bottom chromatogram; same as that in Fig. 1E). The reference WT UPLC chromatogram is also shown (top chromatogram; same as that in Fig. 1E). Download FIG S1, PDF file, 0.3 MB (357.3KB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Analysis of fluorescent protein fusions supports the prediction that ElsL function is cytosolic. Fusion of ElsL to a monomeric superfolder green fluorescent protein (GFP) (msGFP2 [21]) caused a diffuse signal throughout the cell interior, consistent with cytoplasmic localization (Fig. 2A). An analogous LdtAb-msGFP2 fusion, in contrast, showed peripheral fluorescent patches consistent with its predicted localization in the periplasm (Fig. 2B). This difference in localization occurred despite both gene fusions resulting in similarly efficient levels of intact chimeric proteins (Fig. S2A). Expression of elsL-msGFP2 reversed the shape defect of ΔelsL bacteria, resulting in short rods with maximal width matching that of the WT, indicating that the fusion protein was functional (Fig. 2C). With a cytoplasmic location partitioned away from the sacculus, ElsL may thus act as an LDC, an alternative or additional activity seen with some YkuD-family proteins (22, 23). The logical extension of this prediction is that ElsL is the missing-link cytoplasmic LDC allowing recycling of imported cell wall fragments in A. baumannii.
FIG 2.

An ElsL-msGFP fusion shows diffuse cytoplasmic localization, while LdtAb-msGFP localizes to the cell periphery. (A and B) Fluorescent micrographs showing representative WT cells harboring the plasmid-borne gene fusions of msGFP2 to elsL (A) or ldtAb (B). Scale bar, 2 μm. (C) The elsL-msGFP2 fusion reverses the morphology defect of ΔelsL. Cells were imaged by phase-contrast microscopy, and cell width was measured by image analysis. Bars show median values (n ≥ 118). *, P < 0.0001 in Kruskal-Wallis test.
Western blot analysis of fusion protein levels. WT or ΔelsL bacteria containing the indicated IPTG-regulated construct were cultured with the noted IPTG dose, and proteins were analyzed after blotting. The top image in each panel shows a representative immunoblot using the indicted primary antibody. The bottom image shows the total protein levels from the same blot determined by SYPRO ruby staining prior to immunodetection. Location of molecular weight markers (BioRad Precision Plus Protein dual color) are indicated to the left of each blot. –, no insert (vector control). NA, not analyzed. (A) Detection of msGFP2 fusion proteins. (B and C) Detection of 3×FLAG epitope-tagged proteins. Arrowheads indicate the position of the indicated fusion protein (ElsLFLAG in panel C). Download FIG S2, PDF file, 0.9 MB (904.3KB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
elsL and ldtAb have interconnected aggravating genetic interactions.
As a parallel approach to identify leads on ElsL function, we mapped its full network of genetic interactions throughout the genome, which ultimately enabled a series of tests of the above-described hypothesis. To this end, we performed comparative transposon insertion sequencing (Tn-seq) analysis using ElsL+ and ElsL– strains. A dense mariner transposon library was constructed in the ΔelsL background, and colony growth of the resulting double mutants was quantified in massively parallel fashion by measuring Illumina sequencing read abundance corresponding to every member of the library (Materials and Methods). This growth was then compared to that of the matching single (ElsL+) transposon mutants within a control mariner library generated previously in the WT (5). Potential aggravating interactions were identified as genes for which knockout showed growth dependence on ElsL (i.e., a low ratio of ΔelsL/control read counts), with significance determined by permutation test. Examination of such interactions should illuminate the pathways in which ElsL functions.
A large number of genes showed potential aggravating interactions with elsL (Fig. 3A, left; Data Set S1). Several of these mapped to cell wall synthesis, cell division, and envelope stability pathways. For example, the nonessential cell division-associated genes minCD, blhA (5, 24), and ACX60_RS13190 (structural maintenance of chromosomes family [5]) showed substantially lower Tn-seq read counts when mutated in combination with ΔelsL (Fig. 3A and B). A similar result was seen with zapA, a cell division locus linked to RS13190, albeit without passing the 5% false-discovery rate cutoff (Fig. 3A and B). In addition, nearly all mla genes, which play a role in lipid transport between the OM and inner membrane (IM) (25, 26), had Tn-seq read counts dependent on intact elsL (Fig. 3A and B). Notably, ldtAb also showed a prominent growth defect dependent on ΔelsL (Fig. 3A and B), indicating that knockout of both YkuD-family proteins was poorly tolerated in A. baumannii, as suggested previously (8). To interrogate this last result, we generated a parallel mariner library in the ΔldtAb strain and analyzed its genome-wide interactions (Data Set S2). While many fewer potential negative interactions were identified with ΔldtAb than with ΔelsL, the strongest hit was elsL (Fig. 3A, right; Fig. 3B; Data Set S2), supporting the notion that the two genes have an aggravating interaction.
FIG 3.
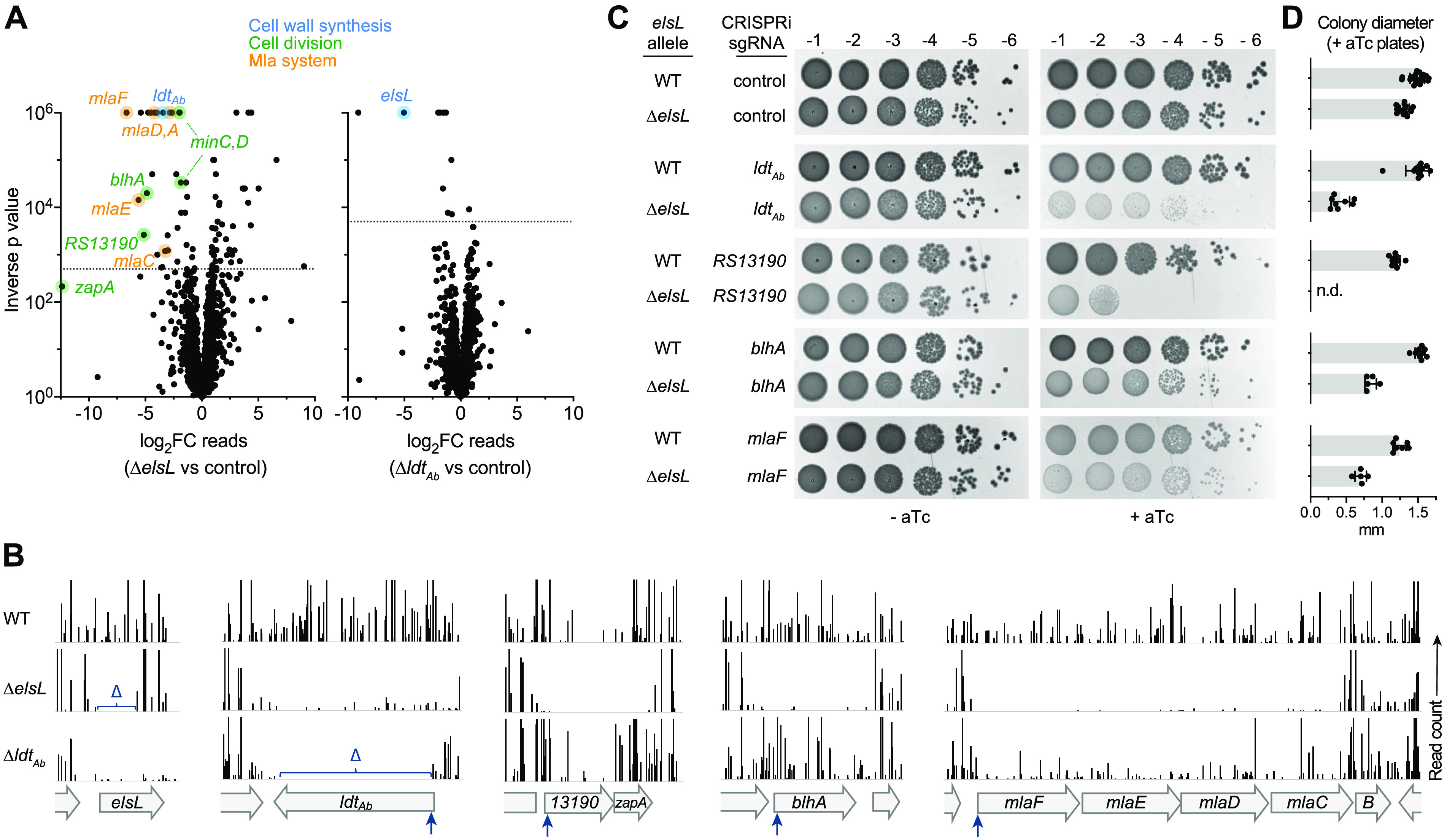
Tn-seq reveals aggravating genetic interactions with elsL and ldtAb. (A) Tn-seq genetic interaction analysis. Volcano plot shows the ratio of Tn-seq read counts mapped to genes in the mutant mariner transposon library (ΔelsL or ΔldtAb) compared to the control transposon library (WT). Dotted horizontal lines indicate a false-discovery rate (q value) of 0.05. Hits described in the text are color-highlighted according to the indicated pathway. (B) Tracks show Tn-seq read counts at each insertion site at different chromosomal loci within the indicated mariner library. Bars represent the normalized read count, and vertical arrows indicate the position targeted by CRISPRi. Δ indicates a deleted region. (C) Validation of aggravating genetic interactions with ΔelsL via targeted CRISPRi and colony formation tests. WT or ΔelsL strains harboring aTc-inducible dcas9 and the indicated sgRNA were serially diluted (10-fold) and spotted on LB agar medium supplemented with 0 or 50 ng/mL aTc. Colonies resulting after overnight 37°C incubation were imaged. (D) Diameters of colonies from triplicate aTc plates at 10−6 or higher dilution were measured by image analysis. Bars show the mean ± SD (n ≥ 5 colonies). P < 0.0001 in unpaired t tests comparing ΔelsL to WT with each sgRNA. n.d., colonies not detected and statistical test not performed.
Tn-seq genetic interactions with ΔelsL. Download Data Set S1, XLSX file, 0.9 MB (915.2KB, xlsx) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Tn-seq genetic interactions with ΔldtAb. Download Data Set S2, XLSX file, 0.6 MB (648.8KB, xlsx) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
We used CRISPR interference (CRISPRi) (27) to validate the negative genetic interactions between elsL and the strongest hits within each pathway (ldtAb, RS13190-zapA, blhA, and mlaC-F). Chimeric single guide RNAs (sgRNAs) were designed to target the 5′ end of each locus (Fig. 3B, blue arrows) and were introduced into WT and ΔelsL strains harboring anhydrotetracycline (aTc)-inducible dCas9. CRISPRi allows efficient knockdown of operons (28); RS13190-zapA and mlaC-F are therefore each likely to be corepressed in this strategy. The effect of knockdown on colony growth was compared with two parallel controls—(i) growth in the absence of dCas9 inducer (– aTc) and (ii) growth with a control, nontargeting sgRNA (27). CRISPRi of each locus in the WT strain resulted in colony growth that was at or near control levels (Fig. 3C and D). In contrast, CRISPRi in ΔelsL caused greatly amplified growth defects. Silencing RS13190 in the ΔelsL background completely blocked colony formation (Fig. 3C and D), indicative of strong genetic aggravation. Other knockdowns in ΔelsL resulted in significantly reduced colony growth compared to controls (Fig. 3C and D). To test whether these growth defects reflect aggravating interactions with elsL, we compared them to those expected from a null multiplicative model based on the growth of single-lesion strains (Materials and Methods). Each double-lesion strain (CRISPRi + ΔelsL) had significantly lower growth than expected from the null model (Table S1). Together, these results indicate that elsL interacts negatively with LDT, cell division, and OM lipid transport pathways.
Quantification of genetic interactions with ΔelsL. Download Table S1, PDF file, 0.02 MB (23KB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Aggravating interactions frequently arise if the underlying genes function in parallel or redundant pathways (29). This scenario could explain the basis for the aggravating interaction between elsL and nonessential cell division genes. Mutation of blhA (24) and silencing of RS13190-zapA (Fig. S3A) each impair cell division and cause a mode of growth that is more heavily dependent on cell elongation compared to the WT. Loss of elsL causes loss of short rod shape and is phenotypically linked to malfunction of the cell elongation machinery (5) (Fig. 2C). Blocking genes of each type in combination may thus result in cells being unable to divide or elongate efficiently. This is supported by microscopy of ΔelsL CRISPRiRS13190-zapA, which showed large, irregular spheroid morphology rather than the filaments seen with CRISPRiRS13190-zapA alone (Fig. S3A). These data are consistent with synthetic lethality occurring due to an inability of cells to continue growing their cell wall laterally as a way to compensate for deficient or delayed cell division. Defective elongational growth machinery could also affect how septal PG synthesis is initiated (30, 31), hypersensitizing to inhibition of a parallel cell division pathway. In addition to parallel pathways, aggravating interactions may arise when one gene product limits the toxicity generated by the absence of the other (32–34). We considered this alternative model in examining the mechanism of the elsL-ldtAb genetic interaction.
Analysis of elsL genetic interactions with RS13190-zapA and ldtAb. (A) elsL deficiency blocks elongated phenotype caused by RS13190-zapA knockdown. WT or ΔelsL bacteria harboring dcas9 and the indicated sgRNA were cultured in the presence of aTc (50 ng/mL) and imaged in log phase by phase-contrast microscopy. Scale bar, 4 μm. (B) elsL and ldtAb are synthetic lethal on LB0 medium. Genetic interaction was analyzed by CRISPRi knockdown of ldtAb in the ΔelsL strain background. Colonies were grown on LB0 agar plates and imaged as in Fig. 3C. (C) Muropeptide profile analysis of the ΔelsL ΔldtAb strain compared to single mutants and WT analyzed in parallel. The muropeptide profiles of the single mutants and WT are the same as those shown in Fig. 1E and are included for comparison with ΔelsL ΔldtAb. Muropeptides are labelled as in Fig. 1E. (D) Percentage of crosslinked muropeptides determined from muropeptide profiles (Materials and Methods). The first 3 samples are the same as those shown in Fig. 1H and are included for comparison with ΔelsL ΔldtAb. Bars show the mean ± SD. Download FIG S3, PDF file, 2.1 MB (2.1MB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Synthetic lethal relationship between elsL and ldtAb.
To investigate the elsL-ldtAb genetic relationship, we combined the two deletions (Materials and Methods) and identified conditions that further aggravate its growth phenotype. The ΔelsL ΔldtAb double mutant formed small, translucent colonies, identical to the phenotype of the ΔelsL CRISPRildtAb strain (Fig. 4A). Since both genes affect the cell wall, we examined growth with low osmolarity medium (LB without NaCl, “LB0”). LB0 medium dramatically aggravated the ΔelsL ΔldtAb growth defect, with colony formation completely blocked at dilutions beyond 10−2, indicating synthetic lethality (Fig. 4A). A similar result was obtained with ΔelsL CRISPRildtAb (Fig. S3B). Consistent with the colony phenotypes, ΔelsL ΔldtAb showed lower viable counts during liquid culture compared to the single mutants and WT, with low osmolarity amplifying the defect (Fig. 4B). The ΔelsL ΔldtAb sacculus showed compositional defects that reflected the combination of each single lesion, with low overall cross-linking and absence of LDT-mediated muropeptides (Fig. S3C and D). The detrimental effect of lacking both genes also manifested in defective cell shape, with the double mutant forming enlarged spheroids in LB and irregular, bloated shapes with frequent blebs and lysis in LB0 (Fig. 4C). Notably, the ΔelsL single mutant also had viability and shape defects compared to the WT that were exacerbated by low osmolarity, while ΔldtAb was unaffected (Fig. 4B and C), underlining the importance of ElsL to physiology and stress resistance. Together, these results indicate that elsL and ldtAb have a synthetic lethal relationship, with the elsL defect, in particular, making cells sensitive to conditions of increased cell wall stress.
FIG 4.

Synthetic lethal interaction between elsL and ldtAb. (A) WT A. baumannii or the indicated deletion mutant were serially diluted and spotted on LB (top) or LB0 (bottom) agar medium, and the resulting colonies were imaged as in Fig. 3. (B) The indicated strains were cultured to saturation in LB and diluted in LB or LB0, and viable counts were determined over time. Data points show the geometric mean ± SD (n = 3 biological replicates). Where not visible, error bars are within the confines of the symbol. (C) The indicated strains (columns) cultured in LB or LB0 (rows) were imaged by phase-contrast microscopy. Arrowheads indicate blebs; the arrow indicates an example of a lysed cell lacking dense phase contrast. Scale bar, 4 μm.
Suppression of elsL-ldtAb synthetic lethality by blocking cell wall muropeptide recycling.
To illuminate ElsL function and identify the source of ΔelsL toxicity, we exploited the synthetic lethality of ΔelsL ΔldtAb and isolated suppressor mutants reversing its major growth defect. Large, opaque colonies forming from ΔelsL ΔldtAb on solid LB or LB0 medium were purified and their mutations mapped (Materials and Methods). Of 22 distinct suppressors identified using this strategy, 21 mapped to 2 genes functioning in PG recycling, ampG (the muropeptide permease) and mpl (murein peptide-UDP-MurNAc ligase) (35) (Fig. 5A). In each case, the mutation was a predicted null allele. Representative mutants showed enhanced colony growth with both LB and LB0 (Fig. 5B), as did an independently constructed, in-frame deletion of ampG (Fig. 5C). The remaining suppressor, which also enhanced colony growth relative to its parent (Fig. 5B), mapped to ACX60_RS13100, which we have named ltgF (lytic transglycosylase determining fosfomycin susceptibility, as explained below). This locus encodes a predicted lytic transglycosylase having an MltD-like catalytic domain and a large C-terminal region with multiple LysM repeats resembling the PG-anchoring domain of autolysins (Fig. 5D) (36). The mutant allele had an insertion sequence (IS) between these two regions. As a predicted PG turnover enzyme, ltgF may have an important role in PG recycling like the other sites of suppression. Supporting this idea, the Tn-seq antibiotic susceptibility profile (phenotypic signature) (5) of ltgF closely correlates with those of canonical PG-recycling genes (Fig. 5E, Fig. S4A). In-frame deletion of ltgF in ATCC 17978 (Fig. 5F and G) as well as transposon mutation in a different strain background (AB5075, Fig. S4C, D) each caused hypsersusceptibility to fosfomycin, a mark of defective cell wall recycling (37, 38). The susceptibility defect was akin to that seen with recycling-blocked ampG mutants tested in parallel (Fig. 5F and G, Fig. S4C and D). Among predicted lytic transglycosylases in A. baumannii, LtgF is the only one with a Tn-seq phenotypic signature showing significant positive correlation with cell wall recycling (Fig. S4B) and characterized by significant hypersensitivity to fosfomycin (5). The above-described results support a role for ltgF in PG turnover and recycling. The findings altogether reveal that the ΔelsL ΔldtAb growth defect is suppressed by blocking cell wall recycling at key points—its earliest steps (generation of anhydromuropeptides or their import) or at the step of peptide reuse (Fig. 5I, left, bold steps).
FIG 5.

Blocking early cell wall-recycling proteins or the Mpl peptide-recycling ligase suppresses ΔelsL ΔldtAb synthetic lethality. (A) Listed are mutations identified by whole-genome sequencing in derivatives of ΔelsL ΔldtAb allowing enhanced colony formation on LB or LB0 agar medium. The protein effect is listed in the case of substitutions; in all other cases, the mutation and the corresponding nucleotide (nt) position relative to the gene start are listed. IS, insertion sequence. (B and C) The indicated spontaneous mutant and the ΔelsL ΔldtAb parent strain (B), or the indicated deletion mutants and WT control (C) were serially diluted and spotted on LB (left) or LB0 (right) agar medium, and the resulting colonies were imaged as in Fig. 3. (D) Schematic of LtgF protein. IS indicates the location affected by the ISAba1 suppressor mutation in ltgF. Rectangles and brackets indicate predicted domains from CDD or SignalP. CDD E values are listed; E values of the LysM domains were below 1e-5. (E) LtgF and canonical cell wall-recycling genes have correlated phenotypic signatures. The heat map shows the Pearson correlation coefficient (r) measuring relatedness of the Tn-seq phenotypic signatures (5) of each gene with that of ltgF. *, P < 0.05. (F and G). The susceptibility to fosfomycin (32 μg/mL) was determined by CFE assay. Bars (F) show the geometric mean ± SD (n = 3 biological replicates). *, P < 0.0001 in unpaired t tests comparing each mutant to the WT. Representative colonies are shown in G. (H) Direction of the Tn-seq genetic interaction between elsL and cell wall-recycling genes depends on the component being recycled. The heat map shows the fold change in Tn-seq read counts of the indicated gene in the ΔelsL mariner library versus that in the WT control library (Data Set S1). (I) Model for the role of ElsL in A. baumannii cell wall recycling. (Left) PG is degraded by lytic transglycosylases (e.g., LtgF) and endopeptidases to release anhydro-MurNAc-containing products. Anhydro-MurNAc-tetrapeptides imported by AmpG are processed to tripeptide form by ElsL. After removal of the PG amino sugars, the tripeptide is reused via ligation to UDP-MurNAc by Mpl, followed by addition of d-Ala-d-Ala to result in UDP-MurNAc-pentapeptide and ultimately lipid II. In the absence of ElsL, in contrast, tetrapeptides are reused by Mpl, resulting in synthesis of aberrant lipid II tetrapeptide (right).
Phenotypes of LtgF (ACX60_RS13100) are closely related to those of cell wall-recycling genes. (A) Tn-seq phenotypic signature of ltgF clusters with those of cell wall-recycling genes. Each row is the Tn-seq phenotypic signature of the indicated gene. The heat map shows normalized Tn-seq fitness in Z-scored units across a panel of distinct drug treatments (columns). Phenotypic signature data are from reference 5. (B) Analysis of correlations between lytic transglycosylase gene and cell wall-recycling gene phenotypic signatures. The heat map shows the Pearson correlation coefficient (r) measuring the relatedness of the Tn-seq phenotypic signatures (5) of the indicated lytic transglycosylase gene (columns) with that of each PG recycling gene (rows). *, P < 0.05. Lytic transglycosylase genes were identified based on functional annotations (5). Gene names are shown when available; otherwise, the locus tag number following ACX60_ is shown. An additional gene, mltG, had no phenotypic signature due to lack of transposon insertions and was not analyzed. (C and D) Disruption of ltgF in strain AB5075 results in hypersusceptibility to fosfomycin. (C) AB5075 WT and the indicated mutant were serially diluted and spotted on solid medium without or with drug, and the resulting colonies were imaged as in Fig. 3. (D) Growth of strains in panel C was monitored during culture in microplates with or without fosfomycin (75 μg/mL). Data points show the geometric mean ± SD (dotted bands) from n = 2 independent cultures. Where not visible, the SD is within the confines of the symbol. (E) ldtAb genetic interactions with cell wall-recycling genes are not dependent on the component being recycled. The heat map shows the fold change in Tn-seq read counts of the indicated gene in the ΔldtAb mariner library versus the WT control library. Download FIG S4, PDF file, 0.5 MB (495.6KB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Reexamination of our genetic interaction Tn-seq data revealed that blocking the same cell wall-recycling steps identified as suppressors of ΔelsL ΔldtAb lethality also alleviated the mild colony growth defect of ΔelsL. For instance, transposon mutations in ltgF and cell wall-recycling enzymes acting on muropeptide or peptide substrates (Fig. 5I) each led to slightly enhanced growth within the ΔelsL library compared to the control (Fig. 5H). In contrast, mutation of other recycling steps dedicated to MurNAc sugar recycling had the opposite effect, decreasing ΔelsL growth compared to the control (Fig. 5H). A clear differential effect of blocking different branches of the cell wall recycling circuit on growth was not observed with the ΔldtAb library (Fig. S4E). These results point to ElsL deficiency and peptide reuse as the key sources of synthetic toxicity that are bypassed by cell wall recycling block.
The suppressor analysis clearly implicates ElsL in cell wall recycling. Integrated with our genetic interaction, muropeptide composition, and localization data, these results lend strong support for the above-described model that ElsL is the cell wall-recycling LDC in A. baumannii. The deleterious effects of ElsL deficiency can thus be explained as a consequence of aberrant buildup of its tetrapeptide substrates. Processing of these intermediates by Mpl and other enzymes would lead to generation of lipid-II tetrapeptide and, ultimately, incorporation of tetrapeptide stems into the cell wall (Fig. 5I, right). These stems are toxic dead-ends, as they cannot be used as donors in cross-link formation by PBPs (39). This would account for the decrease in overall cross-links seen with ΔelsL (Fig. 1E and H). Toxicity would be limited to some extent by LdtAb, since LDTs use tetrapeptides as donors (6); this would account for the elevated level of l,d-cross-links seen with ΔelsL (Fig. 1F). Absent LdtAb, however, the uncrosslinked tetrapeptide stems would compromise cell wall integrity, accounting for the aggravated phenotypes (Fig. 4). Preventing tetrapeptide reuse by suppressor mutations would efficiently bypass this toxic pathway.
Block in cell wall recycling suppresses ΔelsL shape and susceptibility defects.
We interrogated the ElsL LDC model through parallel genetic and biochemical approaches. First, given the ability of muropeptide recycling block to enhance ΔelsL Tn-seq fitness (Fig. 5H), we tested the degree to which such a block also alleviates ΔelsL morphological and antibiotic hypersusceptibility phenotypes. Blocking cell wall recycling by ampG mutation completely restored a short rod shape to cells lacking ElsL, with median cell width returning to the WT value (Fig. 6A and B). ΔelsL bacteria are hypersusceptible to antibiotics that attack division septum PG synthesis, such as sulbactam (5). ampG deletion also completely reversed this defect. While ΔelsL was unable to form colonies on solid medium with low doses of sulbactam, ΔelsL ΔampG grew efficiently, restoring the MIC to a WT level (1 μg/mL; Fig. 6C). This effect extended to another treatment predicted to target septal PG synthesis, the broad-spectrum combination drug piperacillin-tazobactam (40). Specific block of divisional PG synthesis by piperacillin-tazobactam was supported by the dramatic elongation of WT cells in the presence of the drug at levels at or below the MIC (Fig. S5A) and by the marked hypersusceptibility of an elongation-defective Rod system mutant, Δpbp2 (Fig. S5B). ΔelsL similarly showed increased susceptibility to piperacillin-tazobactam, which was reversed by reintroducing the WT allele (Fig. S5B and C). Unlike the WT, ΔelsL did not form elongated filaments in the presence of piperacillin-tazobactam but instead grew as enlarged spheroids (Fig. S5A), similar to genetic interaction results (Fig. S3A) and consistent with the notion that elsL mutation impairs cell wall elongation. Notably, hypersusceptibility of ΔelsL to piperacillin-tazobactam was also fully reversed by removing cell wall recycling through deletion of ampG (Fig. 6D). Together, these results are consistent with ElsL deficiency owing its multiple toxic phenotypes to recycling of aberrant cell wall intermediates.
FIG 6.

Blocking cell wall recycling suppresses morphology and antibiotic hypersusceptibility defects of the ΔelsL mutant. (A and B) ampG deletion restores the rod shape in the ΔelsL strain. Cells were imaged by phase-contrast microscopy, and cell width was measured by image analysis (A). Lines show median values (n ≥ 102). *, P < 0.0001 in the Kruskal-Wallis test. Representative cells are shown in panel B. Scale bar, 2 μm. (C and D) ampG deletion reverses ΔelsL hypersusceptibility to divisome-targeting β-lactams. Susceptibility to sulbactam (C) and piperacillin-tazobactam (D) was measured by CFE assay. Data points show the geometric mean ± SD (n = 3 biological replicates).
Responses of A. baumannii WT and mutants to piperacillin-tazobactam indicate that the treatment targets divisional cell wall synthesis. (A) Piperacillin-tazobactam, like sulbactam, causes WT cells to grow as filaments, consistent with targeting divisome PG synthesis. ΔelsL cells do not grow as filaments in the presence of piperacillin-tazobactam. Cells were grown without (untreated) or with piperacillin-tazobactam (treated, dose indicated) and were imaged by phase-contrast microscopy. Scale bar, 10 μm. (B and C) ElsL and PBP2 mutants show similar hypersusceptibility to piperacillin-tazobactam. Susceptibility was measured by CFE assay. Data points show the geometric mean ± SD (n = 3 biological replicates). Download FIG S5, PDF file, 1.3 MB (1.3MB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
ElsL deficiency causes accumulation of aberrant cytosolic tetrapeptides and disrupts the synthesis of pentapeptide precursors.
We further tested the potential recycling LDC function of ElsL by measuring its effect on levels of PG precursors in vivo. Canonical recycling LDCs act on tetrapeptide-containing products derived from turnover of the cell wall (6, 7). The resulting tripeptides lead to generation of the key cytosolic building block UDP-MurNAc pentapeptide (UDP-M5) (Fig. 5I, left); aberrant reuse of unprocessed tetrapeptides, in contrast, would lead to the potentially toxic UDP-MurNAc tetrapeptide (UDP-M4) (Fig. 5I, right). UDP-M4 should therefore show higher levels in ElsL– versus ElsL+ A. baumannii if the protein functions as an LDC. To test this prediction, we analyzed UDP-linked muramyl-peptides within the bacterial strains via UPLC-MS. In contrast to ElsL+ strains, which had UDP-M5 without detectable UDP-M4 (Fig. 7A, WT and ΔldtAb), ElsL– mutants showed abundant UDP-M4 with a concomitant decrease in UDP-M5 (Fig. 7A, ΔelsL and ΔelsL ΔldtAb), consistent with tetrapeptide substrate buildup due to an absence of LDC function. Analysis of isogenic AmpG– strains revealed the likely mechanism of suppression of blocking cell wall recycling. Compared to AmpG+, AmpG– variants showed dramatically reduced amounts of UDP-M4 that were now below the level of UDP-M5 (from de novo biosynthesis) in each strain (Fig. 7A, ΔelsL ΔampG and ΔelsL ΔldtAb ΔampG). AmpG deletion therefore prevents the toxic tetrapeptide products from being reused for PG synthesis, explaining how recycling mutations bypass the toxic phenotypes of ElsL deficiency.
FIG 7.
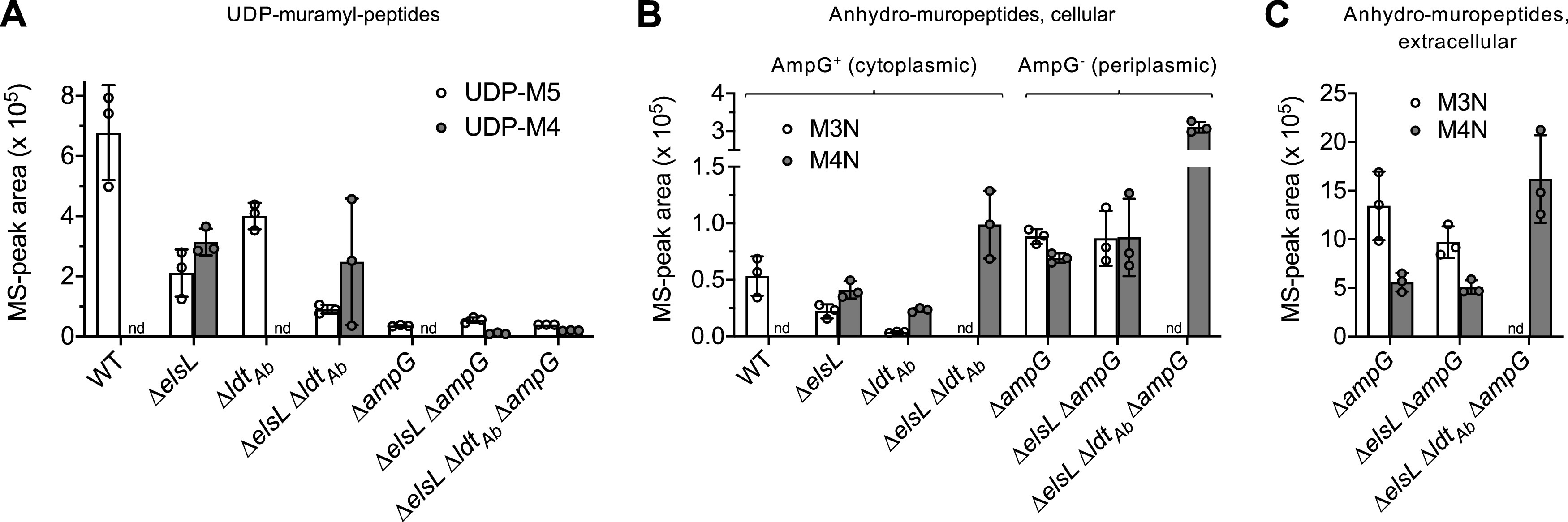
ElsL absence causes accumulation of cytoplasmic tetrapeptide recycling intermediates and a reduction in pentapeptide precursor synthesis that is bypassed by ampG deletion. (A) UDP-muramyl-peptides (UDP-M5, UDP-M4) were detected by MS in the cellular fraction from cultures of the indicated strains. (B and C) Anhydro-muropeptides (M3N and M4N) were detected in the intracellular (B) and extracellular (supernatant, C) fractions by MS from cultures of the indicated strains. In panel B, anhydro-muropeptides detected in AmpG+ cultures are imported and thus reflect cytoplasmic levels; detection of these PG turnover products in AmpG– cultures, in which they are not efficiently imported, thus reflects levels in the periplasm. Bars show the mean ± SD (n = 3 biological replicates). nd, not detected.
Examination of anhydromuropeptides, the turnover-derived fragments that are recycled into UDP-linked precursors, further supports the critical function of ElsL as a recycling LDC. In the WT, anhydromurotripeptide (M3N) but not anhydromurotetrapeptide (M4N) was detected in the cellular fraction (Fig. 7B). In contrast, ΔelsL had a buildup of M4N (Fig. 7B), consistent with lack of a recycling LDC. The AmpG permease is required for cytosolic import of anhydromuropeptides (35); detection of these products in AmpG– cultures thus reflects a periplasmic or extracellular, rather than cytoplasmic, locale. In contrast to the results with AmpG+ strains, elsL deletion did not affect periplasmic (Fig. 7B) or extracellular (Fig. 7C) levels of M4N in the ΔampG background (compare ΔelsL ΔampG to ΔampG). These results are consistent with ElsL providing only a cytoplasmic LDC function. We note that some M3N was detected in ΔelsL, despite the strain lacking the putative recycling LDC; in addition, an M4N increase and M3N decrease (versus WT) was observed with ΔldtAb, despite the presence of the putative LDC (Fig. 7B). In each of these cases, the levels of these anhydromuropeptides can be explained as reflecting the compositionally altered cell walls from which they derive (e.g., the ΔldtAb wall lacks tripeptide stems, while ΔelsL accumulates them; Fig. 1E). In the case of ΔelsL ΔldtAb, the large increase in M4N and undetectable M3N (Fig. 7B and C) are consistent with the combined effect of no cytosolic LDC activity and altered cell wall composition.
ElsL has LDC activity paralleling that of E. coli LdcA.
Three additional results support the model that ElsL is the missing LDC in A. baumannii. First, ΔelsL defects are reversed by LdcA, the canonical LDC from E. coli. To show this, each protein was 3×-FLAG-tagged and expressed in ΔelsL under IPTG control, and IPTG concentrations yielding equivalent protein levels were determined (50 μM with ElsLFLAG; 500 μM with LdcAFLAG; Fig. 8A; Fig. S2B). Using these conditions, both enzymes completely reversed the sulbactam hypersusceptibility of ΔelsL (Fig. 8B). At lower expression levels, LdcA caused partial complementation, while full complementation was still seen with ElsL (Fig. 8A and B; compare ldcAFLAG 50 with elsLFLAG 0 μM IPTG [leaky expression]). These results confirm that the ΔelsL defect is due to deficient LDC activity and suggest that A. baumannii cell wall recycling has evolved to be most efficient with the ElsL YkuD-family LDC.
FIG 8.
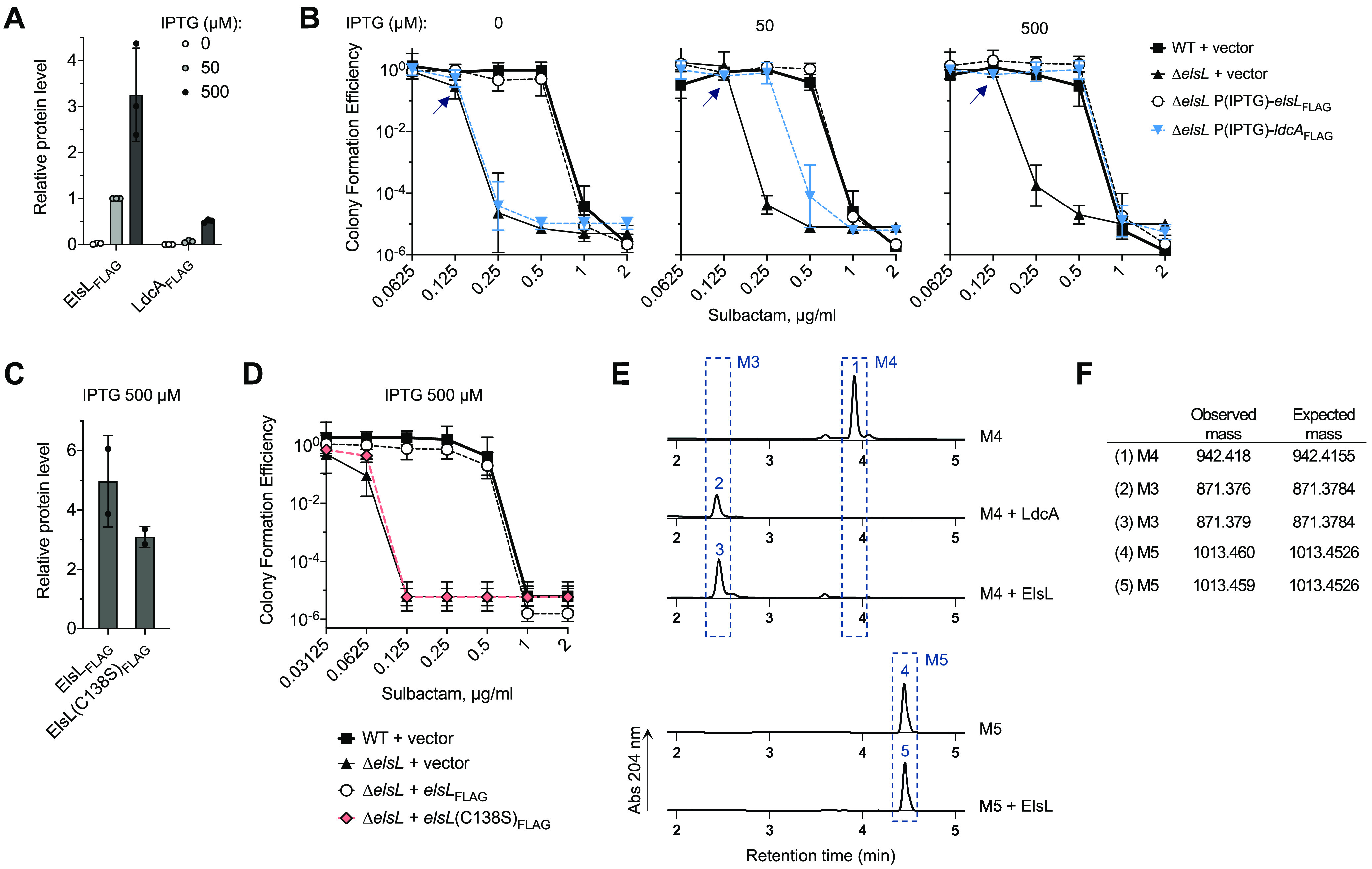
ElsL is the missing-link PG recycling l,d-carboxypeptidase in A. baumannii. (A) Western blot quantification of ElsLFLAG and LdcAFLAG. ΔelsL bacteria with inducible expression of each enzyme by P(IPTG) were cultured with the indicated IPTG concentration. Data points show protein level (relative to ElsLFLAG, IPTG 50 μM) determined from image analysis. Bars show the mean ± SD (n = 3 biological replicates). (B) E. coli LdcA reverses the ΔelsL sulbactam hypersusceptibility defect. Susceptibility was determined by CFE assay. Plates contained the indicated sulbactam and IPTG concentrations. Data points show the geometric mean ± SD (n = 3 biological replicates). Arrows denote that colonies formed by ΔelsL + vector on 0.125 μg/mL sulbactam medium were pinpoint-sized. (C) Western blot quantification of ElsLFLAG in ΔelsL bacteria with the indicated P(IPTG)-inducible allele cultured in the presence of 500 μM IPTG. Data points show the protein level (relative to ElsLFLAG, IPTG 50 μM) as in panel A, except n = 2. (D) Intrinsic sulbactam resistance conferred by ElsL depends on an active site cysteine. Susceptibility to sulbactam was determined by CFE as in panel B. (E and F) ElsL has LDC activity in vitro. Shown are UPLC chromatograms of the reaction products following incubation of substrate (M4, murotetrapeptide, top; or M5, muropentapeptide, bottom) with the indicated purified enzyme (E). Numbered peaks were identified based on retention time and confirmed by MS (F). Minor peaks, which were trace muropeptide contaminants arising during manual HPLC purification of M4 and M5 substrates from C. crescentus PG (Materials and Methods), were identified as M5Gly (at minute ∼3.6) and M2 (at minute ∼4.1).
Second, ElsL activity depends on a cysteine residue that aligns with the conserved catalytic cysteine of YkuD-family LDTs (41, 42). ElsL harboring a mutation at this site (C138S) lost all ability to confer intrinsic sulbactam resistance to ΔelsL when expressed in trans (Fig. 8D), despite high-level induction with 500 μM IPTG (Fig. 8C; Fig. S2C).
Third, purified ElsL has LDC activity in vitro. ElsL and E. coli LdcA, a positive control for LDC activity, were both purified using a C-terminal His tag and assayed in parallel. LDCs cleave between the d-alanine and the meso-diaminopimelic acid only in disaccharide tetrapeptides (M4) and not in disaccharide pentapeptides (M5); we thus used these two muropeptides to assay LDC activity. Similar to LdcA, ElsL completely cleaved the M4 substrate to the disaccharide tripeptide (M3) (Fig. 8E and F). ElsL had no activity on M5 (Fig. 8E and F). ElsL thus possesses selective LDC activity. We conclude that ElsL is a novel type of cell wall-recycling enzyme using a cytoplasmic YkuD-family domain to catalyze the key LDC reaction.
The ElsL LDC family includes orthologs in diverse bacteria.
Search of a database of ∼14,000 representative bacterial genomes (Materials and Methods) revealed predicted cytoplasmic ElsL orthologs in a range of different organisms (Data Set S3). Orthologs were found mainly within the Proteobacteria, with the largest representation by diverse members of the Gamma- and Betaproteobacteria classes (624 and 101, respectively, out of 788 total hits identified). This included the intracellular pathogens Legionella pneumophila and Coxiella burnetii (43). A total of 5/21 gammaproteobacterial and 2/6 betaproteobacterial orders did not contain ElsL orthologs, suggesting that while relatively widespread, the enzyme may have been lost or replaced by other LDCs in those cases. ElsL orthologs were also present in all orders of the Verrucomicrobia, including the human gut microbiota member Akkermansia muciniphila. In sum, a large number of diverse bacterial species appear to have evolved to use the cytoplasmic ElsL-class LDC for recycling of the cell wall.
ElsL orthologs. Download Data Set S3, XLSX file, 0.07 MB (71.1KB, xlsx) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
DISCUSSION
We report here the identification of a novel class of cytoplasmic LDC that enables cell wall recycling in A. baumannii. This class uses a cysteine-containing catalytic domain usually found in LDT proteins. ElsL is the founding member of this family, and orthologs were identified in a range of other bacteria. ElsL deficiency causes a number of phenotypes: (i) loss of rod shape, (ii) impaired growth, (iii) hypersensitivity to septal cell wall-targeting antibiotics, and (iv) cell wall structural defects, including increased 3-3 cross-links with reduced overall cross-linkage. Most of these phenotypes are aggravated by mutating the enzyme necessary for 3-3 cross-link formation (LdtAb) or by low osmolarity, while they are suppressed by blocking muropeptide recycling. Each of these features is explained by the model that ElsL is the LDC that processes turnover-derived muropeptides in A. baumannii, with toxic ΔelsL phenotypes caused by the deleterious tetrapeptides that accumulate absent this processing. Analysis of cell wall precursors supported this model and showed that tetrapeptide-containing biosynthetic intermediates increase in ΔelsL bacteria at the expense of the mature pentapeptide building blocks (Fig. 7). When incorporated as part of nascent cell wall, these tetrapeptides cannot be used as donors in 4-3 bond formation by PBPs and thus diminish overall cell wall cross-linkage (Fig. 1H). Periplasmic LDTs, however, can act on tetrapeptides, likely mitigating their toxic effects and explaining the synthetic lethality of the double elsL ldtAb mutant (Fig. 4). ElsL has LDC activity in vitro, catalyzing the removal of the terminal d-Ala from disaccharide tetrapeptides but not pentapeptides (Fig. 8E), solidifying its critical role in muropeptide recycling.
The highly similar phenotypes resulting from loss of ElsL and loss of Rod system proteins suggest a possible relationship between recycling LDC activity and the ability of cell wall elongation to proceed normally in A. baumannii, although the exact basis for the linked phenotypes remains to be elucidated. The shared phenotypes include loss of rod morphology as well as highly correlated antibiotic susceptibility signatures defined by marked hypersensitivity to division-targeting β-lactams (Fig. S5B) (5). In addition, the ΔelsL growth defect was aggravated by mutation of nonessential cell division genes (Fig. 3), which mirrors the effect of antibiotics blocking division (Fig. 6). Further, ΔelsL was unable to form elongated filaments under conditions of impaired division (Fig. S3A and S5A). Together, these results are consistent with the model that the Rod system is defective when cells lack ElsL, resulting in a heavy reliance on divisome PG synthesis and hypersusceptibility to inhibition of this process. Given its diffuse cytoplasmic location, the recycling LDC is almost certainly not a requisite component of the Rod complex. An alternative possibility is that accumulation of tetrapeptide intermediates is directly or indirectly responsible for Rod system failure. That aberrant intermediates may have selective effects on a particular PG biosynthetic machine is supported by previous work with E. coli showing that PG precursor composition can determine a cell’s preference for lateral versus septal cell wall growth (44–46). For instance, increasing the levels of a d,d-carboxypeptidase, which cleaves the terminal d-alanine on pentapeptide subunits to yield tetrapeptides, caused rod-shaped WT cells to grow as spheres (45) and allowed filamentous pbp3 hypomorphs to shorten and divide more efficiently (44). These findings are consistent with the model that the Rod system prefers to act on PG with pentapeptide subunits and does not use tetrapeptides (or their tripeptide derivatives) efficiently (44, 45). The opposite may be true of the divisome, in which the main transpeptidase, PBP3, is thought to prefer or require tripeptides, originating from tetrapeptides, as the acceptor muropeptide (47, 48). Interestingly, dramatic Rod– phenotypes are also seen with mutation of A. baumannii dacC (5), a predicted PG hydrolase that may also modulate the balance of cell wall peptide substrates in ways promoting lateral cell wall growth. The idea that cell wall subunit levels determine Rod system versus divisome activity may also contribute to the mitigation of toxicity in ΔelsL cells by LdtAb, in agreement with what was proposed in reference 6. While this mitigation is likely due to provision of alternative, supportive cross-links, an additional possibility is that LdtAb directly or indirectly generates tripeptide substrates in ΔelsL that are needed for optimal cell division. Further work is required to understand the mechanisms by which alterations to cell wall stem peptides in A. baumannii drives the function (and malfunction) of specific PG biosynthesis enzymes.
Our genetic interaction analysis revealed an additional interdependent relationship between ElsL and the Mla lipid transport pathway. The Mla proteins mediate retrograde transport of phospholipids from the OM to the inner membrane, facilitating lipid asymmetry in the OM (25, 49). The system may also prevent excessive loss of lipids in the form of OM vesicles (OMVs) (50, 51). Interestingly, an elsL mutant sheds larger amounts of OMVs than the WT (8). It is thus possible that membrane lipid loss contributes to the aggravating phenotype of the elsL mla double mutant. Another possibility is that combining an impaired cell wall (due to low cross-linkage, ElsL–) with a compromised OM (Mla–) leads to synergistic failure of envelope mechanical integrity (52), manifesting as an aggravated growth defect. In sum, a variety of pathways, including those determining alternative cross-link formation, cell division, and OM homeostasis, become indispensable in the absence of ElsL function, underlining the importance of this key enzyme to multiple facets of A. baumannii envelope biogenesis.
The novel ElsL class of cell wall-recycling LDC identified in this work uses a cysteine catalytic residue, contrasting it from that of the two described recycling LDC enzymes, LdcA and LdcV. LdcA, found in E. coli, Pseudomonas aeruginosa, and several other organisms, is a serine peptidase that uses a Ser-His-Glu catalytic triad (53), while LdcV, identified recently in Vibrio cholerae, Aeromonas hydrophila and Proteus mirabilis, belongs to the LAS superfamily of zinc-dependent metalloproteases that use an active site zinc ion coordinated by histidines (6, 54). While all three LDC classes enable muropeptide recycling, they may differ in their relative preferences for the variety of potential tetrapeptide-containing substrates derived from the cell wall (7), although this remains to be determined. The identification of a distinct, noncanonical class of LDC related to LDTs has two important implications. First, the finding helps illuminate the muropeptide recycling pathway in many bacteria, such as A. baumannii, that lack a homolog of the previously identified recycling LDCs (35). Lack of a known LDC homolog in such bacteria has been explained previously by the possibility that they bypass the need for a recycling LDC due to low cell wall tetrapeptide content or high Mpl selectivity against tetrapeptides (35). While these mechanisms may be true in many cases, an alternative explanation is that a range of organisms possess a nonclassical LDC (e.g., of the ElsL family), allowing the typical peptide recycling loop to be fully closed. Second, the reliance of A. baumannii on two YkuD-domain proteins for maintaining the cell wall points to a unique potential vulnerability in this highly resistant pathogen. Simultaneously targeting the catalytic cysteines in both ElsL and LdtAb could exploit the synthetic lethality of the two proteins to potently weaken the sacculus of the pathogen and inhibit growth. Copper (14) and carbapenem antibiotics (55), known inhibitors of LDT proteins, are candidates for such a strategy.
In conclusion, we have identified a new family of LDC critical to muropeptide recycling in A. baumannii and likely many other organisms. Our combined genetic and biochemical interrogation of ElsL function provided insights into the role of the protein in recycling as well as its pleiotropic influence on numerous pathways important to envelope integrity and growth. Future work will examine the mechanisms by which changes to cell wall precursors in cells showing altered expression of ElsL or other muropeptide-modifying enzymes lead to preferential action/inaction of specific PG synthesis machines, which will likely shed light on how cell wall synthesis and cell proliferation are coordinated in A. baumannii. Furthermore, the vulnerabilities exposed by ElsL inhibition, precursor buildup, and their network of toxic effects present opportunities for the rational development of improved antimicrobials aimed at controlling this intractable pathogen.
MATERIALS AND METHODS
Bacterial strains and growth conditions.
The bacterial strains used in this work are described in Table S2. A. baumannii strains were derivatives of ATCC 17978. Bacteria were cultured in lysogeny broth (10 g/L tryptone, 5 g/L yeast extract, 10 g/L NaCl) (LB), unless otherwise noted. Liquid cultures were incubated at 37°C in flasks with orbital shaking or in tubes with rotation via roller drum. Growth was monitored by measuring absorbance at 600 nm. LB agar was supplemented with antibiotics (carbenicillin [Cb] at 25 to 100 μg/mL, kanamycin [Km] at 10 to 20 μg/mL, gentamicin [Gm] at 10 μg/mL) or sucrose (10%) as needed (Sigma-Aldrich). LB prepared without NaCl (LB0) was used in phenotypic testing where noted. Superoptimal broth with catabolite repression (SOC) was used in Tn-seq library preparation, and Vogel Bonner medium (VBM) supplemented with Gm (10 μg/mL) was used for isolation of CRISPRi strains containing miniTn7 constructs.
Strains, plasmids, and primers used in this study. Download Table S2, PDF file, 0.08 MB (79.4KB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Molecular cloning and strain construction.
The plasmids used in this study are listed in Table S2. Most DNA constructs were generated by PCR-amplification using oligonucleotide primers (Integrated DNA Technologies [IDT]; Table S2) and cloning in the HincII site of pUC18, followed by verification by sequencing (Genewiz) before subcloning in subsequent vectors. Plasmids for complementation, localization, allele exchange, and sgRNA production were introduced into A. baumannii ATCC 17978 by electroporation (56).
(i) Complementation and localization experiments. To generate an ldtAb plasmid for HADA experiments, the EcoRI and PstI fragment from pYDE231 was subcloned downstream of the T5lacP promoter in pEGE305 to generate pYDE240. To construct elsL- and ldcA-3xFLAG fusions for complementation experiments, the BamHI-XbaI fragment from pYDE342 and pYDE343 were each subcloned in pJE42, which provides an in-frame C-terminal 3×FLAG sequence, generating pYDE346 and pYDE347. The hybrid genes were then subcloned into pEGE305 using EcoRI and PstI, resulting in pYDE350 (ElsLFLAG) and pYDE351 (LdcAFLAG). To construct msGFP2 gene fusions, a codon-optimized msGFP2 reporter gene containing an in-frame polyglycine linker was first synthesized as a double-stranded DNA fragment by IDT and cloned in the HincII site of pUC18 (pAFE225). The BamHI-XbaI fragments from pYDE342 and pYDE063 were cloned in pAFE225, generating pYDE386 (elsL-msGFP2) and pYDE387 (ldtAb-msGFP2). The gene fusions were then cloned into pEGE305 using EcoRI and PstI, generating pYDE389 and pYDE390, respectively. To generate an independent msGFP2 control gene, msGFP2 was amplified from pAFE225 using a primer (msGFP2_F) that replaces the XbaI site and glycine linker with EcoRI, ribosome binding, and translational start sites. The PCR product was then directly cloned into pEGE305 using EcoRI and PstI sites, generating pAFE256. An elsLFLAG(C138S) mutant was constructed by replacing the EcoRI-BstBI fragment of pYDE350 with a fragment amplified with a primer (elsL-C138S-R) containing the substitution mutation.
(ii) Overexpression and purification of elsL and ldcA. The NcoI-EcoRI fragments from pYDE281 and pYDE324, respectively, were each cloned upstream of the in-frame 6×His tag sequence in pET28b, creating (pYDE290 and pYDE328).
(iii) Gene deletions. Deletion mutants were isolated using homologous recombination/allelic exchange using sucrose counterselection as described (57). A ΔelsL ΔldtAb double mutant (EGA740) was constructed by allelic exchange with pEGE268 in EGA739 (5); the strain was isolated as small, sucrose-resistant colonies after overnight 30°C incubation followed by 1 day at room temperature. A ΔelsL ΔampG double mutant (YDA411) was isolated by allelic exchange with pEGE268 in EGA516 (58). A ΔelsL ΔldtAb ΔampG triple mutant (YDA414) was isolated by allelic exchange with pEGE271 in YDA411. In-frame deletion of ltgF was constructed by three-way ligation of ∼1 kb flanking homology arms with pJB4648.
(iv) CRISPRi. sgRNAs were constructed by PCR-amplifying 24-base, protospacer-adjacent motif (PAM)-adjacent target regions satisfying previously described criteria (59, 60), followed by cloning directly into the SpeI and ApaI sites of pYDE007 (27) and verification by restriction digestion and sequencing. ΔelsL and ΔldtAb strains with inducible dcas9 were isolated by introducing pYDE009 (27) into EGA738 and EGA739 via four-parental mating (57, 61); transposants were isolated on VBM-Gm10 plates, generating YDA186 and YDA095, respectively. Integration at the attTn7 locus was confirmed by PCR, and loss of pYDE009 was confirmed by screening on Cb plates (62, 63).
Construction of transposon mutant libraries.
Mutagenesis of ΔelsL (EGA738) and ΔldtAb (EGA739) with the mariner transposon was performed by electroporation with pDL1100 (5). Cells were then diluted with 1 mL SOC, allowed to incubate for 15 min at 37°C, and spread on membrane filters (0.45-μm pore size) overlaid on prewarmed SOC agar plates. Plates were incubated at 37°C for 1 h, and filters were transferred to solid LB supplemented with Km (10 μg/mL with ΔelsL and 20 μg/mL with ΔldtAb). After overnight incubation at 37°C, colonies were lifted from filters by agitation in sterile phosphate-buffered saline (PBS), mixed with sterile glycerol (10%), aliquoted, and stored at −80°C. Approximately 160,000 mutant colonies from 19 subpools were analyzed in the ΔelsL strain, and approximately 300,000 from 20 subpools were analyzed with the ΔldtAb strain.
Tn-seq library amplification, sequencing, and analysis.
Genomic DNA was extracted from samples using the DNeasy kit (Qiagen) and quantified using a SYBR green I (Invitrogen) microplate assay. Transposon-adjacent DNA was tagmented and amplified for Illumina sequencing as described (5, 64). Samples were multiplexed, reconditioned, and size selected (250 or 275 to 600 bp; Pippin HT) before sequencing (single-end 50 bp) using primer mar512 on a HiSeq 2500 instrument with high output V4 chemistry at the Tufts University Genomics Core Facility (TUCF-Genomics).
Sequencing reads were quality-filtered, clipped of adapters, and mapped to the A. baumannii chromosome (GenBank accession no. NZ_CP012004) with Bowtie (27). Mapped reads were tabulated in wig format according to the position of their TA sites in the NZ_CP012004 genome using custom python scripts (27). To examine genetic interactions between transposon mutations and the deleted gene, data sets were analyzed using the resampling method in the TRANSIT software package (parameters: samples=10000000, norm=TTR, histograms=False, adaptive=True, excludeZeros=False, pseudocounts=0.0, LOESS=False, trim_Nterm=0.0, trim_Cterm=10.0) (65), using previously published WT mariner libraries as the control data set (27). Resampling P values were adjusted for multiple comparisons (q value) using the method of Benjamini and Hochberg. Results were visualized as volcano plots using Prism 8. Genes having a low mean read count (<5) in both the deletion and control data sets (essential genes) were not plotted. Hits were defined as genes having a q value of <0.05, ≥3 transposon sites, and >5 mean read counts in both data sets. To visualize Tn-seq read counts along chromosomal regions, TTR-normalized counts were merged into a single wig file, scaled such that median read coverage at nonzero insertion sites was equivalent between data sets, and viewed using Integrative Genomics Viewer (27, 66).
Microscopy.
Bacteria in the log phase of growth were immobilized on agarose pads (1% in PBS) prior to imaging. For HADA labeling experiments, bacteria were pulsed with 1 mM HADA (Tocris Bioscience) for 15 min, fixed with 70% ice-cold ethanol for 10 to 15 min, and washed twice with PBS before imaging. For localization experiments, strains harboring IPTG-inducible gene fusions to msGFP2 were cultured with 100 μM IPTG prior to imaging. Micrographs were acquired with a 100×/1.4 phase-contrast objective on a Zeiss Axio Observer 7 microscope. A Colibri 7 LED light source was used for fluorescence illumination. Imaging of GFP fluorescence used the 475-nm LED and filter set 92 HE. Imaging of HADA fluorescence used the 385-nm LED and filter set 96 HE. For analysis of HADA incorporation, Fiji software (67) was used for background subtraction from the HADA signal and for determining cell boundaries from stacked HADA and phase images. Fluorescence intensity across populations of cells in multiple fields was then plotted as demographs using MicrobeJ software (68). For analysis of cell width, the maximal width relative to the medial axis of each cell was quantified from phase images using MicrobeJ (68).
Genetic interaction and antibiotic susceptibility tests.
Bacteria (optical density [OD], 1) were serially diluted 10-fold in PBS and spot-plated on the noted solid medium. After overnight growth at 37°C, colonies were imaged with transilluminated light on a ChemiDoc MP imaging system (Bio-Rad). For genetic interaction analysis, colony diameters were measured using ImageJ (69) and normalized to the WT control (YDA007). Genetic interactions were analyzed by comparing the relative colony diameter values of strains harboring two genetic lesions (CRISPRi knockdown of candidate gene and deletion of ΔelsL) with the hypothetical values expected from a multiplicative model, in which the values of each single-lesion strain are multiplied (70). Sensitivity to sulbactam and to piperacillin-tazobactam (8:1 ratio by mass) was measured with a colony formation efficiency (CFE) assay (58). Serial dilutions of WT and isogenic mutant strains were spotted on solid LB agar medium containing graded concentrations of drug (or no drug control). After overnight growth at 37°C, colony counts were determined and compared to the no-drug control. The limit of detection was approximately 10−5 to 10−6.
PG isolation and analysis.
Sacculi isolation and PG analysis were performed as previously described (71). In short, A. baumannii cells from overnight LB cultures were harvested, resuspended in LB + 5% SDS, and boiled with stirring for 2 h followed by stirring at room temperature overnight. SDS was removed from sacculi through several rounds of ultracentrifugation and resuspension with MilliQ water. Sacculi were then resuspended in 100 mM Tris-HCl pH 8.0 buffer with proteinase K (20 μg/mL) and incubated at 37°C for 1 h. The reaction was stopped by adding SDS 1% and boiling at 100°C for 5 min. SDS was removed as described above, and sacculi were resuspended in water. Muramidase was then added, and reaction mixtures were incubated overnight at 37°C to solubilize the sacculi completely. The soluble muropeptides were reduced using NaBH4, and their pH was adjusted before separation using ultraperformance liquid chromatography (UPLC; Waters) and identification using a matrix-assisted laser desorption ionization–time of flight mass spectrometry (MALDI-TOF MS) system (Waters).
For quantification, we chose 2 random PG profiles that were representative of each strain. The area for each identified peak was integrated, giving each individual muropeptide a relative area value based on the total integrated area. Using these values, the molar percentage was also calculated for each muropeptide. This relative molarity was also used to calculate the degree of cross-linking using the following formula:
Analysis of intracellular and extracellular soluble muropeptides.
Bacteria were grown with LB at 37°C for 4 h to log phase, and the OD was recorded. Bacteria were cooled on ice and centrifuged at 25,000 rpm for 20 min at 4°C. Supernatants were stored at 4°C for analysis of extracellular soluble muropeptides, and pellets were used for analysis of the intracellular fraction.
To study PG turnover products and precursors within the intracellular fraction, cell pellets were washed twice with sterile 1% NaCl, resuspended in water, and boiled for 30 min with stirring. Samples were again centrifuged at 25,000 rpm for 15 min, and supernatants were recovered and filtered (0.2-μm pore size). A MALDI-TOF MS system was used for identification of the muropeptides derived from turnover (anhydro species) and synthesis (UDP-activated species). To study muropeptides within the extracellular fraction, supernatants were filtered as described above and immediately boiled for 15 min to precipitate proteins. The soluble fraction was then analyzed using the same MS system as for the intracellular muropeptides, focusing on detection of products derived from turnover that were released into the media. Quantification of all muropeptides used the total-ion count detected by the system. All analyses were performed with biological triplicate samples.
Suppressor mapping by whole-genome sequencing and phenotypic signature analysis.
Suppressor mutants bypassing the EGA740 growth defect were isolated on LB or LB0 plates. Genomic DNA (DNeasy kit) was quantified with a SYBR green I (Invitrogen) microplate assay and used as input for Illumina library preparation using a modified small-volume Nextera tagmentation method as described previously (58). Libraries were sequenced (single-end 100 bp) on a HiSeq 2500 instrument at TUCF-Genomics. Reads were aligned to the NZ_CP012004 genome, and variants were identified using breseq (72). The predicted functional impact of substitution variants was determined by using PROVEAN (73). Identities of ISs were determined by using ISFinder (74). Phenotypic signatures of ltgF and cell wall-recycling genes were compared using Qlucore Omics Explorer 3.5, and Pearson correlations were analyzed with Prism 8.
Immunoanalysis of ElsL, LdtAb, and LdcA proteins.
Strains harboring fusion genes or vector control were diluted to OD 0.05 in LB with IPTG (100 μM with msGFP2 fusions; 0, 50, or 500 μM with 3×FLAG fusions) and grown to OD 0.5. Cells were centrifuged and resuspended (50 μL per 1 mL of sample) with SDS sample loading buffer (msGFP2 fusions) or BugBuster protein extraction reagent with 0.1% Lysonase (Millipore) followed by SDS loading buffer (3×FLAG fusions). Samples were boiled for 10 min, separated by SDS-PAGE (12% acrylamide gel), and transferred to an Immobilon-FL polyvinylidene difluoride (PVDF) membrane. Total protein was detected by SYPRO ruby protein blot stain (Invitrogen). GFP was detected by rabbit PABG1 primary (Chromotek; 1:5,000 dilution) and goat anti-rabbit IgG horseradish peroxidase (HRP) secondary (Invitrogen; 1:5,000 dilution) antibodies. FLAG epitope was detected by mouse anti-FLAG M2 primary (Invitrogen; 1:1,000 dilution) and goat anti-mouse IgG HRP secondary (Invitrogen; 1:5,000 dilution) antibodies. Blots were imaged with a ChemiDoc MP system (Bio-Rad). Band intensities were quantified using Image Lab software. Samples were normalized by dividing the immunodetected band intensity by the total protein level from SYPRO staining. Relative values were calculated by dividing each normalized value by the normalized value of the ElsLFLAG, 50 μM IPTG sample on the same blot.
Protein overexpression and purification.
Overnight cultures of E. coli BL21(DE3) strains harboring pYDE290 (elsLHis) or pYDE328 (ldcAHis) were seeded at a 1:100 dilution into 1 L of LB with Km (30 μg/mL) and grown at 37°C to OD 0.6 to 0.8. IPTG was added at 500 μM final, and cultures were incubated at 16°C for 20 h. Cells were harvested, resuspended with 30 mL of prechilled buffer A (50 mM Tris-HCl, 300 mM NaCl), and pulse-sonicated on ice for 15 min. The lysate was clarified by centrifugation at 10,000 × g at 4°C for 1 h and filtered (0.45-μm pore size; Millipore). Clarified lysates were loaded on Ni-NTA resin columns (Thermo Scientific) previously equilibrated with 6% (vol/vol) buffer B (50 mM Tris-HCl, 300 mM NaCl, 500 mM imidazole) in buffer A. His-tagged proteins were eluted with gradient concentrations of buffer B (6%, 10%, 30%, 50%, 100%) at 4°C and detected by SDS-PAGE and Coomassie blue staining. Fractions containing the His-tagged proteins were concentrated by using Amicon Ultra-15 centrifugal filter units (10 kDa). Proteins were washed 3 times with 15 mL stocking buffer (50 mM Tris-HCl, 50 NaCl, 10% [wt/vol] glycerol) and resuspended with 200 μL stocking buffer. Proteins were quantified using the Pierce Coomassie Bradford protein assay (Thermo Scientific) and stored as aliquots at −80°C. With ElsLHis, the elution and stocking buffers included 0.5 mM tris(2-carboxyethyl)phosphine (TCEP) to prevent oxidation of the active-site cysteine residue (75).
In vitro assays with ElsL.
LDC activity was assayed in vitro using purified proteins and muropeptide substrates. Muropeptide substrates (M4 and M5) were extracted from sacculi isolated from Caulobacter crescentus, which are known to have high M4 and M5 content (76), and solubilized as described above. Soluble muropeptides were separated using a high-performance liquid chromatography (Waters) system, and each individual muropeptide was collected manually. Collected fractions were washed from solvents by evaporation using a SpeedVac vacuum concentrator followed by resuspension with MilliQ water. All reactions were performed in 100 mM Tris-HCl, mixing a fraction of M4 or M5 and 10 ng of the corresponding protein (ElsLHis, LdcAHis, or none in the negative control). Reaction mixtures were incubated for 2 h at 37°C and stopped by incubation at 100°C for 5 min. After inactivation, the samples were reduced, and their pH was adjusted for injection on the UPLC. Muropeptides were identified according to their retention time, and their identities were validated through MS analysis.
ElsL ortholog identification.
A. baumannii ElsL (GenBank accession no. WP_000077301) was queried with BLASTp using the refseq_select_prot database, restricting the search to Bacteria (representing ∼14,000 genomes) and an E value cutoff of 1e-4. From the 1,082 protein sequences that were returned, custom Python scripts were used to identify sequences containing a predicted signal peptide or transmembrane helix (based on the SignalP 5.0 [77], PrediSi [78], Phobius [79], and TMHMM 2.0 [80] methods), or PG-binding domain (from search of the NCBI Conserved Domain Database [CDD] [42]). With SignalP 5.0, an “other” score of <0.75 was used as a conservative marker of potential signal peptides. Hits were excluded if any of the above-mentioned searches were positive. Finally, sequences shorter than 105 or longer than 200 amino acids were excluded. Taxonomic information for the resulting set of ElsL orthologs (788 hits) was obtained from the NCBI database.
Data availability.
All sequence data can be found in the NCBI Sequence Read Archive under accession no. PRJNA763919.
ACKNOWLEDGMENTS
This work was supported by Northeastern University startup funds and by the NIAID/NIH under award number R01AI162996 to E.G. Research in the Cava lab is supported by the Swedish Research Council, The Laboratory of Molecular Infection Medicine Sweden (MIMS), The Knut and Alice Wallenberg Foundation (KAW), Umeå University, and the Kempe Foundation.
We thank Sylvie Manuse and Emily Kirkwood for assistance with microscopy and image analysis, Wei-Leung Ng for plasmid gifts, Tom Bernhardt and David Roper for experimental advice, and members of the Geisinger lab for helpful discussions.
Contributor Information
Edward Geisinger, Email: e.geisinger@northeastern.edu.
Nina R. Salama, Fred Hutchinson Cancer Research Center
REFERENCES
- 1.Sievert DM, Ricks P, Edwards JR, Schneider A, Patel J, Srinivasan A, Kallen A, Limbago B, Fridkin S, National Healthcare Safety Network (NHSN) Team and Participating NHSN Facilities . 2013. Antimicrobial-resistant pathogens associated with healthcare-associated infections: summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009-2010. Infect Control Hosp Epidemiol 34:1–14. doi: 10.1086/668770. [DOI] [PubMed] [Google Scholar]
- 2.Weiner-Lastinger LM, Abner S, Edwards JR, Kallen AJ, Karlsson M, Magill SS, Pollock D, See I, Soe MM, Walters MS, Dudeck MA. 2020. Antimicrobial-resistant pathogens associated with adult healthcare-associated infections: summary of data reported to the National Healthcare Safety Network, 2015-2017. Infect Control Hosp Epidemiol 41:1–18. doi: 10.1017/ice.2019.296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Doi Y, Murray GL, Peleg AY. 2015. Acinetobacter baumannii: evolution of antimicrobial resistance-treatment options. Semin Respir Crit Care Med 36:85–98. doi: 10.1055/s-0034-1398388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Tacconelli E, Carrara E, Savoldi A, Harbarth S, Mendelson M, Monnet DL, Pulcini C, Kahlmeter G, Kluytmans J, Carmeli Y, Ouellette M, Outterson K, Patel J, Cavaleri M, Cox EM, Houchens CR, Grayson ML, Hansen P, Singh N, Theuretzbacher U, Magrini N, WHO Pathogens Priority List Working Group . 2018. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis 18:318–327. doi: 10.1016/S1473-3099(17)30753-3. [DOI] [PubMed] [Google Scholar]
- 5.Geisinger E, Mortman NJ, Dai Y, Cokol M, Syal S, Farinha A, Fisher DG, Tang AY, Lazinski DW, Wood S, Anthony J, van Opijnen T, Isberg RR. 2020. Antibiotic susceptibility signatures identify potential antimicrobial targets in the Acinetobacter baumannii cell envelope. Nat Commun 11:4522. doi: 10.1038/s41467-020-18301-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hernandez SB, Dorr T, Waldor MK, Cava F. 2020. Modulation of peptidoglycan synthesis by recycled cell wall tetrapeptides. Cell Rep 31:107578. doi: 10.1016/j.celrep.2020.107578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Templin MF, Ursinus A, Holtje JV. 1999. A defect in cell wall recycling triggers autolysis during the stationary growth phase of Escherichia coli. EMBO J 18:4108–4117. doi: 10.1093/emboj/18.15.4108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kang KN, Kazi MI, Biboy J, Gray J, Bovermann H, Ausman J, Boutte CC, Vollmer W, Boll JM. 2021. Septal class A penicillin-binding protein activity and ld-transpeptidases mediate selection of colistin-resistant lipooligosaccharide-deficient Acinetobacter baumannii. mBio 12:e02185-20. doi: 10.1128/mBio.02185-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Aliashkevich A, Cava F. 2021. LD-transpeptidases: the great unknown among the peptidoglycan cross-linkers. FEBS J doi: 10.1111/febs.16066. [DOI] [PubMed] [Google Scholar]
- 10.Hugonnet JE, Mengin-Lecreulx D, Monton A, den Blaauwen T, Carbonnelle E, Veckerle C, Brun YV, van Nieuwenhze M, Bouchier C, Tu K, Rice LB, Arthur M. 2016. Factors essential for L,D-transpeptidase-mediated peptidoglycan cross-linking and beta-lactam resistance in Escherichia coli. Elife 5:e19469. doi: 10.7554/eLife.19469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mainardi JL, Morel V, Fourgeaud M, Cremniter J, Blanot D, Legrand R, Frehel C, Arthur M, Van Heijenoort J, Gutmann L. 2002. Balance between two transpeptidation mechanisms determines the expression of beta-lactam resistance in Enterococcus faecium. J Biol Chem 277:35801–35807. doi: 10.1074/jbc.M204319200. [DOI] [PubMed] [Google Scholar]
- 12.Hsu YP, Booher G, Egan A, Vollmer W, VanNieuwenhze MS. 2019. d-Amino acid derivatives as in situ probes for visualizing bacterial peptidoglycan biosynthesis. Acc Chem Res 52:2713–2722. doi: 10.1021/acs.accounts.9b00311. [DOI] [PubMed] [Google Scholar]
- 13.Alvarez L, Cordier B, Van Teeffelen S, Cava F. 2020. Analysis of Gram-negative bacteria peptidoglycan by ultra-performance liquid chromatography. Bio Protoc 10:e3780. doi: 10.21769/BioProtoc.3780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Peters K, Pazos M, Edoo Z, Hugonnet JE, Martorana AM, Polissi A, VanNieuwenhze MS, Arthur M, Vollmer W. 2018. Copper inhibits peptidoglycan LD-transpeptidases suppressing beta-lactam resistance due to bypass of penicillin-binding proteins. Proc Natl Acad Sci USA 115:10786–10791. doi: 10.1073/pnas.1809285115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Boll JM, Crofts AA, Peters K, Cattoir V, Vollmer W, Davies BW, Trent MS. 2016. A penicillin-binding protein inhibits selection of colistin-resistant, lipooligosaccharide-deficient Acinetobacter baumannii. Proc Natl Acad Sci USA 113:E6228–E6237. doi: 10.1073/pnas.1611594113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Crepin S, Ottosen EN, Peters K, Smith SN, Himpsl SD, Vollmer W, Mobley HLT. 2018. The lytic transglycosylase MltB connects membrane homeostasis and in vivo fitness of Acinetobacter baumannii. Mol Microbiol 109:745–762. doi: 10.1111/mmi.14000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Le NH, Peters K, Espaillat A, Sheldon JR, Gray J, Di Venanzio G, Lopez J, Djahanschiri B, Mueller EA, Hennon SW, Levin PA, Ebersberger I, Skaar EP, Cava F, Vollmer W, Feldman MF. 2020. Peptidoglycan editing provides immunity to Acinetobacter baumannii during bacterial warfare. Sci Adv 6:eabb5614. doi: 10.1126/sciadv.abb5614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lonergan ZR, Nairn BL, Wang J, Hsu YP, Hesse LE, Beavers WN, Chazin WJ, Trinidad JC, VanNieuwenhze MS, Giedroc DP, Skaar EP. 2019. An Acinetobacter baumannii, zinc-regulated peptidase maintains cell wall integrity during immune-mediated nutrient sequestration. Cell Rep 26:2009–2018.e6. doi: 10.1016/j.celrep.2019.01.089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Glauner B, Holtje JV, Schwarz U. 1988. The composition of the murein of Escherichia coli. J Biol Chem 263:10088–10095. doi: 10.1016/S0021-9258(19)81481-3. [DOI] [PubMed] [Google Scholar]
- 20.Takacs CN, Hocking J, Cabeen MT, Bui NK, Poggio S, Vollmer W, Jacobs-Wagner C. 2013. Growth medium-dependent glycine incorporation into the peptidoglycan of Caulobacter crescentus. PLoS One 8:e57579. doi: 10.1371/journal.pone.0057579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Valbuena FM, Fitzgerald I, Strack RL, Andruska N, Smith L, Glick BS. 2020. A photostable monomeric superfolder green fluorescent protein. Traffic 21:534–544. doi: 10.1111/tra.12737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Magnet S, Arbeloa A, Mainardi JL, Hugonnet JE, Fourgeaud M, Dubost L, Marie A, Delfosse V, Mayer C, Rice LB, Arthur M. 2007. Specificity of L,D-transpeptidases from gram-positive bacteria producing different peptidoglycan chemotypes. J Biol Chem 282:13151–13159. doi: 10.1074/jbc.M610911200. [DOI] [PubMed] [Google Scholar]
- 23.Sycuro LK, Rule CS, Petersen TW, Wyckoff TJ, Sessler T, Nagarkar DB, Khalid F, Pincus Z, Biboy J, Vollmer W, Salama NR. 2013. Flow cytometry-based enrichment for cell shape mutants identifies multiple genes that influence Helicobacter pylori morphology. Mol Microbiol 90:869–883. doi: 10.1111/mmi.12405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Knight D, Dimitrova DD, Rudin SD, Bonomo RA, Rather PN. 2016. Mutations decreasing intrinsic beta-lactam resistance are linked to cell division in the nosocomial pathogen Acinetobacter baumannii. Antimicrob Agents Chemother 60:3751–3758. doi: 10.1128/AAC.00361-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Malinverni JC, Silhavy TJ. 2009. An ABC transport system that maintains lipid asymmetry in the gram-negative outer membrane. Proc Natl Acad Sci USA 106:8009–8014. doi: 10.1073/pnas.0903229106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Powers MJ, Trent MS. 2019. Intermembrane transport: glycerophospholipid homeostasis of the Gram-negative cell envelope. Proc Natl Acad Sci USA 116:17147–17155. doi: 10.1073/pnas.1902026116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bai J, Dai Y, Farinha A, Tang AY, Syal S, Vargas-Cuebas G, van Opijnen T, Isberg RR, Geisinger E. 2021. Essential gene analysis in Acinetobacter baumannii by high-density transposon mutagenesis and CRISPR interference. J Bacteriol 203:e00565-20. doi: 10.1128/JB.00565-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Peters JM, Silvis MR, Zhao D, Hawkins JS, Gross CA, Qi LS. 2015. Bacterial CRISPR: accomplishments and prospects. Curr Opin Microbiol 27:121–126. doi: 10.1016/j.mib.2015.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Klobucar K, Brown ED. 2018. Use of genetic and chemical synthetic lethality as probes of complexity in bacterial cell systems. FEMS Microbiol Rev 42. doi: 10.1093/femsre/fux054. [DOI] [PubMed] [Google Scholar]
- 30.Potluri LP, Kannan S, Young KD. 2012. ZipA is required for FtsZ-dependent preseptal peptidoglycan synthesis prior to invagination during cell division. J Bacteriol 194:5334–5342. doi: 10.1128/JB.00859-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pazos M, Peters K, Casanova M, Palacios P, VanNieuwenhze M, Breukink E, Vicente M, Vollmer W. 2018. Z-ring membrane anchors associate with cell wall synthases to initiate bacterial cell division. Nat Commun 9:5090. doi: 10.1038/s41467-018-07559-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hromas R, Kim HS, Sidhu G, Williamson E, Jaiswal A, Totterdale TA, Nole J, Lee SH, Nickoloff JA, Kong KY. 2017. The endonuclease EEPD1 mediates synthetic lethality in RAD52-depleted BRCA1 mutant breast cancer cells. Breast Cancer Res 19:122. doi: 10.1186/s13058-017-0912-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Myers SH, Ortega JA, Cavalli A. 2020. Synthetic lethality through the lens of medicinal chemistry. J Med Chem 63:14151–14183. doi: 10.1021/acs.jmedchem.0c00766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zinovyev A, Kuperstein I, Barillot E, Heyer WD. 2013. Synthetic lethality between gene defects affecting a single non-essential molecular pathway with reversible steps. PLoS Comput Biol 9:e1003016. doi: 10.1371/journal.pcbi.1003016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Park JT, Uehara T. 2008. How bacteria consume their own exoskeletons (turnover and recycling of cell wall peptidoglycan). Microbiol Mol Biol Rev 72:211–227. doi: 10.1128/MMBR.00027-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Carroll SA, Hain T, Technow U, Darji A, Pashalidis P, Joseph SW, Chakraborty T. 2003. Identification and characterization of a peptidoglycan hydrolase, MurA, of Listeria monocytogenes, a muramidase needed for cell separation. J Bacteriol 185:6801–6808. doi: 10.1128/JB.185.23.6801-6808.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Borisova M, Gisin J, Mayer C. 2014. Blocking peptidoglycan recycling in Pseudomonas aeruginosa attenuates intrinsic resistance to fosfomycin. Microb Drug Resist 20:231–237. doi: 10.1089/mdr.2014.0036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gil-Marques ML, Moreno-Martinez P, Costas C, Pachon J, Blazquez J, McConnell MJ. 2018. Peptidoglycan recycling contributes to intrinsic resistance to fosfomycin in Acinetobacter baumannii. J Antimicrob Chemother 73:2960–2968. doi: 10.1093/jac/dky289. [DOI] [PubMed] [Google Scholar]
- 39.Mainardi JL, Fourgeaud M, Hugonnet JE, Dubost L, Brouard JP, Ouazzani J, Rice LB, Gutmann L, Arthur M. 2005. A novel peptidoglycan cross-linking enzyme for a beta-lactam-resistant transpeptidation pathway. J Biol Chem 280:38146–38152. doi: 10.1074/jbc.M507384200. [DOI] [PubMed] [Google Scholar]
- 40.Htoo HH, Brumage L, Chaikeeratisak V, Tsunemoto H, Sugie J, Tribuddharat C, Pogliano J, Nonejuie P. 2019. Bacterial cytological profiling as a tool to study mechanisms of action of antibiotics that are active against Acinetobacter baumannii. Antimicrob Agents Chemother 63:e02310-18. doi: 10.1128/AAC.02310-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Biarrotte-Sorin S, Hugonnet JE, Delfosse V, Mainardi JL, Gutmann L, Arthur M, Mayer C. 2006. Crystal structure of a novel beta-lactam-insensitive peptidoglycan transpeptidase. J Mol Biol 359:533–538. doi: 10.1016/j.jmb.2006.03.014. [DOI] [PubMed] [Google Scholar]
- 42.Marchler-Bauer A, Bo Y, Han L, He J, Lanczycki CJ, Lu S, Chitsaz F, Derbyshire MK, Geer RC, Gonzales NR, Gwadz M, Hurwitz DI, Lu F, Marchler GH, Song JS, Thanki N, Wang Z, Yamashita RA, Zhang D, Zheng C, Geer LY, Bryant SH. 2017. CDD/SPARCLE: functional classification of proteins via subfamily domain architectures. Nucleic Acids Res 45:D200–D203. doi: 10.1093/nar/gkw1129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sandoz KM, Popham DL, Beare PA, Sturdevant DE, Hansen B, Nair V, Heinzen RA. 2016. Transcriptional profiling of Coxiella burnetii reveals extensive cell wall remodeling in the small cell variant developmental form. PLoS One 11:e0149957. doi: 10.1371/journal.pone.0149957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Begg KJ, Takasuga A, Edwards DH, Dewar SJ, Spratt BG, Adachi H, Ohta T, Matsuzawa H, Donachie WD. 1990. The balance between different peptidoglycan precursors determines whether Escherichia coli cells will elongate or divide. J Bacteriol 172:6697–6703. doi: 10.1128/jb.172.12.6697-6703.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Markiewicz Z, Broome-Smith JK, Schwarz U, Spratt BG. 1982. Spherical E. coli due to elevated levels of D-alanine carboxypeptidase. Nature 297:702–704. doi: 10.1038/297702a0. [DOI] [PubMed] [Google Scholar]
- 46.Truong TT, Vettiger A, Bernhardt TG. 2020. Cell division is antagonized by the activity of peptidoglycan endopeptidases that promote cell elongation. Mol Microbiol 114:966–978. doi: 10.1111/mmi.14587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Botta GA, Park JT. 1981. Evidence for involvement of penicillin-binding protein 3 in murein synthesis during septation but not during cell elongation. J Bacteriol 145:333–340. doi: 10.1128/jb.145.1.333-340.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Pisabarro AG, Prats R, Vaquez D, Rodriguez-Tebar A. 1986. Activity of penicillin-binding protein 3 from Escherichia coli. J Bacteriol 168:199–206. doi: 10.1128/jb.168.1.199-206.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Powers MJ, Trent MS. 2018. Phospholipid retention in the absence of asymmetry strengthens the outer membrane permeability barrier to last-resort antibiotics. Proc Natl Acad Sci USA 115:E8518–E8527. doi: 10.1073/pnas.1806714115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Baarda BI, Zielke RA, Le Van A, Jerse AE, Sikora AE. 2019. Neisseria gonorrhoeae MlaA influences gonococcal virulence and membrane vesicle production. PLoS Pathog 15:e1007385. doi: 10.1371/journal.ppat.1007385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Roier S, Zingl FG, Cakar F, Durakovic S, Kohl P, Eichmann TO, Klug L, Gadermaier B, Weinzerl K, Prassl R, Lass A, Daum G, Reidl J, Feldman MF, Schild S. 2016. A novel mechanism for the biogenesis of outer membrane vesicles in Gram-negative bacteria. Nat Commun 7:10515. doi: 10.1038/ncomms10515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Rojas ER, Billings G, Odermatt PD, Auer GK, Zhu L, Miguel A, Chang F, Weibel DB, Theriot JA, Huang KC. 2018. The outer membrane is an essential load-bearing element in Gram-negative bacteria. Nature 559:617–621. doi: 10.1038/s41586-018-0344-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Korza HJ, Bochtler M. 2005. Pseudomonas aeruginosa LD-carboxypeptidase, a serine peptidase with a Ser-His-Glu triad and a nucleophilic elbow. J Biol Chem 280:40802–40812. doi: 10.1074/jbc.M506328200. [DOI] [PubMed] [Google Scholar]
- 54.Hoyland CN, Aldridge C, Cleverley RM, Duchene MC, Minasov G, Onopriyenko O, Sidiq K, Stogios PJ, Anderson WF, Daniel RA, Savchenko A, Vollmer W, Lewis RJ. 2014. Structure of the LdcB LD-carboxypeptidase reveals the molecular basis of peptidoglycan recognition. Structure 22:949–960. doi: 10.1016/j.str.2014.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mainardi JL, Hugonnet JE, Rusconi F, Fourgeaud M, Dubost L, Moumi AN, Delfosse V, Mayer C, Gutmann L, Rice LB, Arthur M. 2007. Unexpected inhibition of peptidoglycan LD-transpeptidase from Enterococcus faecium by the beta-lactam imipenem. J Biol Chem 282:30414–30422. doi: 10.1074/jbc.M704286200. [DOI] [PubMed] [Google Scholar]
- 56.Choi KH, Kumar A, Schweizer HP. 2006. A 10-min method for preparation of highly electrocompetent Pseudomonas aeruginosa cells: application for DNA fragment transfer between chromosomes and plasmid transformation. J Microbiol Methods 64:391–397. doi: 10.1016/j.mimet.2005.06.001. [DOI] [PubMed] [Google Scholar]
- 57.Geisinger E, Isberg RR. 2015. Antibiotic modulation of capsular exopolysaccharide and virulence in Acinetobacter baumannii. PLoS Pathog 11:e1004691. doi: 10.1371/journal.ppat.1004691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Geisinger E, Mortman NJ, Vargas-Cuebas G, Tai AK, Isberg RR. 2018. A global regulatory system links virulence and antibiotic resistance to envelope homeostasis in Acinetobacter baumannii. PLoS Pathog 14:e1007030. doi: 10.1371/journal.ppat.1007030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Guzzo M, Castro LK, Reisch CR, Guo MS, Laub MT. 2020. A CRISPR interference system for efficient and rapid gene knockdown in Caulobacter crescentus. mBio 11:e02415-19. doi: 10.1128/mBio.02415-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Qi LS, Larson MH, Gilbert LA, Doudna JA, Weissman JS, Arkin AP, Lim WA. 2013. Repurposing CRISPR as an RNA-guided platform for sequence-specific control of gene expression. Cell 152:1173–1183. doi: 10.1016/j.cell.2013.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Carruthers MD, Nicholson PA, Tracy EN, Munson RS, Jr.. 2013. Acinetobacter baumannii utilizes a type VI secretion system for bacterial competition. PLoS One 8:e59388. doi: 10.1371/journal.pone.0059388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kumar A, Dalton C, Cortez-Cordova J, Schweizer HP. 2010. Mini-Tn7 vectors as genetic tools for single copy gene cloning in Acinetobacter baumannii. J Microbiol Methods 82:296–300. doi: 10.1016/j.mimet.2010.07.002. [DOI] [PubMed] [Google Scholar]
- 63.Jacobs AC, Thompson MG, Black CC, Kessler JL, Clark LP, McQueary CN, Gancz HY, Corey BW, Moon JK, Si Y, Owen MT, Hallock JD, Kwak YI, Summers A, Li CZ, Rasko DA, Penwell WF, Honnold CL, Wise MC, Waterman PE, Lesho EP, Stewart RL, Actis LA, Palys TJ, Craft DW, Zurawski DV. 2014. AB5075, a highly virulent isolate of Acinetobacter baumannii, as a model strain for the evaluation of pathogenesis and antimicrobial treatments. mBio 5:e01076-14. doi: 10.1128/mBio.01076-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Geisinger E, Vargas-Cuebas G, Mortman NJ, Syal S, Dai Y, Wainwright EL, Lazinski D, Wood S, Zhu Z, Anthony J, van Opijnen T, Isberg RR. 2019. The landscape of phenotypic and transcriptional responses to ciprofloxacin in Acinetobacter baumannii: acquired resistance alleles modulate drug-induced SOS response and prophage replication. mBio 10:e01127-19. doi: 10.1128/mBio.01127-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.DeJesus MA, Ambadipudi C, Baker R, Sassetti C, Ioerger TR. 2015. TRANSIT: a software tool for Himar1 TnSeq analysis. PLoS Comput Biol 11:e1004401. doi: 10.1371/journal.pcbi.1004401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Robinson JT, Thorvaldsdottir H, Winckler W, Guttman M, Lander ES, Getz G, Mesirov JP. 2011. Integrative genomics viewer. Nat Biotechnol 29:24–26. doi: 10.1038/nbt.1754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, Tinevez JY, White DJ, Hartenstein V, Eliceiri K, Tomancak P, Cardona A. 2012. Fiji: an open-source platform for biological-image analysis. Nat Methods 9:676–682. doi: 10.1038/nmeth.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ducret A, Quardokus EM, Brun YV. 2016. MicrobeJ, a tool for high throughput bacterial cell detection and quantitative analysis. Nat Microbiol 1:16077. doi: 10.1038/nmicrobiol.2016.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Schneider CA, Rasband WS, Eliceiri KW. 2012. NIH Image to ImageJ: 25 years of image analysis. Nat Methods 9:671–675. doi: 10.1038/nmeth.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.van Opijnen T, Bodi KL, Camilli A. 2009. Tn-seq: high-throughput parallel sequencing for fitness and genetic interaction studies in microorganisms. Nat Methods 6:767–772. doi: 10.1038/nmeth.1377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Alvarez L, Hernandez SB, de Pedro MA, Cava F. 2016. Ultra-sensitive, high-resolution liquid chromatography methods for the high-throughput quantitative analysis of bacterial cell wall chemistry and structure. Methods Mol Biol 1440:11–27. doi: 10.1007/978-1-4939-3676-2_2. [DOI] [PubMed] [Google Scholar]
- 72.Deatherage DE, Barrick JE. 2014. Identification of mutations in laboratory-evolved microbes from next-generation sequencing data using breseq. Methods Mol Biol 1151:165–188. doi: 10.1007/978-1-4939-0554-6_12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Choi Y, Sims GE, Murphy S, Miller JR, Chan AP. 2012. Predicting the functional effect of amino acid substitutions and indels. PLoS One 7:e46688. doi: 10.1371/journal.pone.0046688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Siguier P, Perochon J, Lestrade L, Mahillon J, Chandler M. 2006. ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res 34:D32–D36. doi: 10.1093/nar/gkj014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Coyle JF, Pagliai FA, Zhang D, Lorca GL, Gonzalez CF. 2018. Purification and partial characterization of LdtP, a cell envelope modifying enzyme in Liberibacter asiaticus. BMC Microbiol 18:201. doi: 10.1186/s12866-018-1348-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Markiewicz Z, Glauner B, Schwarz U. 1983. Murein structure and lack of DD- and LD-carboxypeptidase activities in Caulobacter crescentus. J Bacteriol 156:649–655. doi: 10.1128/jb.156.2.649-655.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Almagro Armenteros JJ, Tsirigos KD, Sonderby CK, Petersen TN, Winther O, Brunak S, von Heijne G, Nielsen H. 2019. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat Biotechnol 37:420–423. doi: 10.1038/s41587-019-0036-z. [DOI] [PubMed] [Google Scholar]
- 78.Hiller K, Grote A, Scheer M, Munch R, Jahn D. 2004. PrediSi: prediction of signal peptides and their cleavage positions. Nucleic Acids Res 32:W375–W379. doi: 10.1093/nar/gkh378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Kall L, Krogh A, Sonnhammer EL. 2007. Advantages of combined transmembrane topology and signal peptide prediction: the Phobius web server. Nucleic Acids Res 35:W429–32. doi: 10.1093/nar/gkm256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Krogh A, Larsson B, von Heijne G, Sonnhammer EL. 2001. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J Mol Biol 305:567–580. doi: 10.1006/jmbi.2000.4315. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
UPLC/MS analysis of A. baumannii cell wall. Muropeptide fragments isolated from ΔelsL sacculi were identified by MS (table below chromatograms). Bold values in the table indicate the protonated state of the muropeptide more abundantly detected in the UPLC/MS analysis. The corresponding peaks were labelled (blue numbers) according to their muropeptide identity in the ΔelsL UPLC chromatogram (bottom chromatogram; same as that in Fig. 1E). The reference WT UPLC chromatogram is also shown (top chromatogram; same as that in Fig. 1E). Download FIG S1, PDF file, 0.3 MB (357.3KB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Western blot analysis of fusion protein levels. WT or ΔelsL bacteria containing the indicated IPTG-regulated construct were cultured with the noted IPTG dose, and proteins were analyzed after blotting. The top image in each panel shows a representative immunoblot using the indicted primary antibody. The bottom image shows the total protein levels from the same blot determined by SYPRO ruby staining prior to immunodetection. Location of molecular weight markers (BioRad Precision Plus Protein dual color) are indicated to the left of each blot. –, no insert (vector control). NA, not analyzed. (A) Detection of msGFP2 fusion proteins. (B and C) Detection of 3×FLAG epitope-tagged proteins. Arrowheads indicate the position of the indicated fusion protein (ElsLFLAG in panel C). Download FIG S2, PDF file, 0.9 MB (904.3KB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Tn-seq genetic interactions with ΔelsL. Download Data Set S1, XLSX file, 0.9 MB (915.2KB, xlsx) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Tn-seq genetic interactions with ΔldtAb. Download Data Set S2, XLSX file, 0.6 MB (648.8KB, xlsx) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Quantification of genetic interactions with ΔelsL. Download Table S1, PDF file, 0.02 MB (23KB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Analysis of elsL genetic interactions with RS13190-zapA and ldtAb. (A) elsL deficiency blocks elongated phenotype caused by RS13190-zapA knockdown. WT or ΔelsL bacteria harboring dcas9 and the indicated sgRNA were cultured in the presence of aTc (50 ng/mL) and imaged in log phase by phase-contrast microscopy. Scale bar, 4 μm. (B) elsL and ldtAb are synthetic lethal on LB0 medium. Genetic interaction was analyzed by CRISPRi knockdown of ldtAb in the ΔelsL strain background. Colonies were grown on LB0 agar plates and imaged as in Fig. 3C. (C) Muropeptide profile analysis of the ΔelsL ΔldtAb strain compared to single mutants and WT analyzed in parallel. The muropeptide profiles of the single mutants and WT are the same as those shown in Fig. 1E and are included for comparison with ΔelsL ΔldtAb. Muropeptides are labelled as in Fig. 1E. (D) Percentage of crosslinked muropeptides determined from muropeptide profiles (Materials and Methods). The first 3 samples are the same as those shown in Fig. 1H and are included for comparison with ΔelsL ΔldtAb. Bars show the mean ± SD. Download FIG S3, PDF file, 2.1 MB (2.1MB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Phenotypes of LtgF (ACX60_RS13100) are closely related to those of cell wall-recycling genes. (A) Tn-seq phenotypic signature of ltgF clusters with those of cell wall-recycling genes. Each row is the Tn-seq phenotypic signature of the indicated gene. The heat map shows normalized Tn-seq fitness in Z-scored units across a panel of distinct drug treatments (columns). Phenotypic signature data are from reference 5. (B) Analysis of correlations between lytic transglycosylase gene and cell wall-recycling gene phenotypic signatures. The heat map shows the Pearson correlation coefficient (r) measuring the relatedness of the Tn-seq phenotypic signatures (5) of the indicated lytic transglycosylase gene (columns) with that of each PG recycling gene (rows). *, P < 0.05. Lytic transglycosylase genes were identified based on functional annotations (5). Gene names are shown when available; otherwise, the locus tag number following ACX60_ is shown. An additional gene, mltG, had no phenotypic signature due to lack of transposon insertions and was not analyzed. (C and D) Disruption of ltgF in strain AB5075 results in hypersusceptibility to fosfomycin. (C) AB5075 WT and the indicated mutant were serially diluted and spotted on solid medium without or with drug, and the resulting colonies were imaged as in Fig. 3. (D) Growth of strains in panel C was monitored during culture in microplates with or without fosfomycin (75 μg/mL). Data points show the geometric mean ± SD (dotted bands) from n = 2 independent cultures. Where not visible, the SD is within the confines of the symbol. (E) ldtAb genetic interactions with cell wall-recycling genes are not dependent on the component being recycled. The heat map shows the fold change in Tn-seq read counts of the indicated gene in the ΔldtAb mariner library versus the WT control library. Download FIG S4, PDF file, 0.5 MB (495.6KB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Responses of A. baumannii WT and mutants to piperacillin-tazobactam indicate that the treatment targets divisional cell wall synthesis. (A) Piperacillin-tazobactam, like sulbactam, causes WT cells to grow as filaments, consistent with targeting divisome PG synthesis. ΔelsL cells do not grow as filaments in the presence of piperacillin-tazobactam. Cells were grown without (untreated) or with piperacillin-tazobactam (treated, dose indicated) and were imaged by phase-contrast microscopy. Scale bar, 10 μm. (B and C) ElsL and PBP2 mutants show similar hypersusceptibility to piperacillin-tazobactam. Susceptibility was measured by CFE assay. Data points show the geometric mean ± SD (n = 3 biological replicates). Download FIG S5, PDF file, 1.3 MB (1.3MB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
ElsL orthologs. Download Data Set S3, XLSX file, 0.07 MB (71.1KB, xlsx) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Strains, plasmids, and primers used in this study. Download Table S2, PDF file, 0.08 MB (79.4KB, pdf) .
Copyright © 2021 Dai et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Data Availability Statement
All sequence data can be found in the NCBI Sequence Read Archive under accession no. PRJNA763919.