

Abstract
Hydrogen sulfide (H2S) is a ubiquitous gaseous signaling molecule that has an important role in many physiological and pathological processes in mammalian tissues, with the same importance as two others endogenous gasotransmitters such as NO (nitric oxide) and CO (carbon monoxide). Endogenous H2S is involved in a broad gamut of processes in mammalian tissues including inflammation, vascular tone, hypertension, gastric mucosal integrity, neuromodulation, and defense mechanisms against viral infections as well as SARS-CoV-2 infection. These results suggest that the modulation of H2S levels has a potential therapeutic value. Consequently, synthetic H2S-releasing agents represent not only important research tools, but also potent therapeutic agents. This review has been designed in order to summarize the currently available H2S donors; furthermore, herein we discuss their preparation, the H2S-releasing mechanisms, and their -biological applications.
Keywords: hydrogen sulfide, natural H2S donors, synthetic H2S donors, H2S release, medicinal chemistry
1. Introduction
1.1. Hydrogen Sulfide: Chemical Properties
Hydrogen sulfide (H2S) is a colorless, flammable, and water-soluble gas well known for its characteristic odor of rotten eggs. The acute effects of H2S in humans are directly proportional to its concentration and these include eye, nose, throat, and respiratory system irritation (low concentrations) and rapid loss of consciousness and death (high concentrations) [1,2,3]. Recently H2S has been considered as a member of the gasotransmitter family because it shows some common characteristics with its congeners NO (nitric oxide) and CO (carbon monoxide) [4,5,6,7,8,9,10,11].
Hydrogen sulfide has a chemical structure similar to that of water, but it is much less polar than water because the sulfur atom is less electronegative than the oxygen present in water. For this reason, the intermolecular forces for H2S are relatively weaker and consequently the melting and boiling points are much lower than they are for water. H2S can be considered as a weak acid that gives rise to two dissociations in aqueous solution (Ka1 = 1.3 × 10−7 M, Ka2 = 1 × 10−19 M). From the first dissociation hydronium cation (H3O+) and hydrosulfide anion (HS−) are obtained; the second dissociation leads to H3O+ and sulfide ions (S2−). In biological fluids and under physiological conditions hydrogen sulfide is mainly undissociated [12].
Hydrogen sulfide represents a reductant species because of the oxidation state of the sulfur atom in H2S that is –2; it can also be considered a potent nucleophile at physiological pH values. These features make H2S a very reactive acid that can react with many biological molecules. Hydrogen sulfide can give rise to two kinds of redox reactions: the first oxidation should form a highly reactive HS• radical (for example in the reaction with metal centers) [13], while the second kind of redox reaction of H2S with many biologically important reactive oxygen species (ROS) can also lead to the formation of species such as hydrogen peroxide (H2O2), superoxide (O2), peroxynitrite (ONOO), and NO. [14,15,16]. Hydrogen sulfide is also able to react with S-nitroso thiols to give thionitrous acid (HSNO) that probably acts as a cell-permeable nitrosylating agent that might represent the link and explain the cell membrane–impermeable transnitrosation mechanism [17]. Some evidence reported in the literature suggests that H2S also mediates an important oxidative post-translational modification and in particular protein S-sulfuration, generating, for example, S-SH groups on cysteine residues. Even if the detailed mechanism(s) is not yet known, this reaction could represent the chemical route involving H2S for the modification of the functions of a broad range of cellular proteins and enzymes. In the literature, authors have generally agreed that H2S reacts with cysteine-modified residues, such as disulfides (S-S–), sulfenic acids (S-OH), nitrosothiols (S-NO), etc., to form S-sulfhydrated products. Some authors have suggested that protein cysteines directly react with some H2S metabolites or precursors, such as sulfane sulfurs and hydrogen polysulfides, to obtain S-SH products [18,19,20,21,22,23,24,25,26,27]. However, further studies are needed to elucidate this mechanism in detail. Finally, H2S is able to bind transition metals by coordination bonds. In particular, it is able to bind strongly to copper, and this feature has allowed the development of fluorescent probes [28]. It has been shown that the activation of odorant receptors mediated by the sulfur–copper coordination leads to an increase in human sensitivity to the smell of volatile sulfur compounds [29].
The toxicity of hydrogen sulfide to mammals is due to the interactions with cytochrome c oxidase (CcO) with the consequent inhibition of mitochondrial respiration; however, the significant capacity of H2S to induce a suspended animation-like state [30] is believed to be associated with a reversible inhibition of CcO by sulfide coordination chemistry [31].
1.2. Hydrogen Sulfide: Biosynthesis and Metabolic Pathways
1.2.1. Biosynthesis of H2S
Hydrogen sulfide is produced by enzymatic and non-enzymatic endogenous reactions in mammalian tissues (Figure 1). The enzymatic production of H2S involves different enzymes such as cystathionine β-synthase (CBS) and cystathionine γ-lyase (CSE), that are two pyridoxal-5′-phosphate (PLP, vitamin B6)-dependent enzymes, as well as cysteine aminotransferase (CAT) and 3-mercaptopyruvate sulfurtransferase (3-MST). These enzymes interact with specific substrates and reverse trans-sulfuration pathways.
Figure 1.

H2S biosynthesis: in the panels from (A–F) the different known pathways leading to the production of H2S in mammalian tissues are reported (CBS, cystathionine β-synthase; CSE, cystathionine γ-lyase; CAT, cysteine aminotransferase; MST, 3-mercaptopyruvate sulfurtransferase).
The human cystathionine beta-synthase (CBS) enzyme is made up of 551 amino acids and it is characterized by a subunit molecular weight of ~63 kDa [32]. CBS is a tetrameric protein, and each subunit binds to one heme and one pyridoxal 5′-phosphate (PLP). The catalytic center is similar to that of other members of the β- or fold II class of PLP-dependent enzymes and represents the conserved part of the protein [33]. Protoporphyrin IX represents a structural requirement of cystathionine β-synthase, and this feature is unique in the family of PLP-dependent enzymes. Moreover, a conserved glycine rich loop of fold II enzymes (G256–T257–G258–G259–T260) is responsible for multifarious electrostatic interactions with the phosphate portion of PLP [33].
The three-dimensional structures of human and yeast cystathionine gamma-lyase (CSE) have been determined through X-ray crystallography studies [34]. CSE is characterized by four equal monomers with a molecular weight of ~45 kDa; in each monomer the pyridoxal 5′-phosphate (PLP) cofactor is linked with a covalent bond. Nevertheless, some studies have reported differential PLP binding affinities within the monomers of CSE and a momentary dissociation or dismissal of PLP from the enzyme during the catalytic action [35]. Based on studies reported in the literature, several active site residues have been identified as being involved in the catalysis of the α,β-elimination reaction, leading to the production of H2S [36]. In the crystal structures of yeast and human CSE, some residues participate in the binding of the PLP cofactor. These residues are represented by Tyr60 and Arg62 from the adjacent monomer; Tyr114 that forms the π-stacking interaction with the pyridoxal moiety of the cofactor; Asp187 that establishes hydrogen bonds with the pyridoxal nitrogen; and Ser209 and Thr211 that form hydrogen bonds with the phosphate group. Some studies have also reported that Lys212 represents a significant catalytic residue considering that it forms a covalent bond with the PLP cofactor and facilitates proton transfer reactions during the α,γ-elimination reaction of L-cystathionine. Site-directed mutagenesis studies have demonstrated that the PLP cofactor also binds to Tyr60 and Arg62 from the adjacent subunit through a hydrogen bond [37].
CBS and CSE catalyze reactions that represent the primary pathways for H2S production (Figure 1). In particular, CBS is involved in the β-replacement reaction of homocysteine with serine to give cystathionine. CSE later takes part in the α,γ-elimination of cystathionine that leads to cysteine, α- ketobutyrate, NH3, and hydrogen sulfide (Figure 1A) [38].
Either CBS or CSE can also produce H2S from l-cysteine via β-elimination reactions [39]. In particular, CBS forms l-serine and H2S from l-cysteine (Figure 1B), while CSE generates pyruvate (using l-cysteine as the substrate), NH3 and H2S (Figure 1C) or it firstly forms thiocysteine, pyruvate, and NH3 and subsequently cysteine, S-alkenyl-mercaptocysteine (CysSR), and H2S (Figure 1D). Furthermore, CSE catalyzes the reaction of L-homoserine to form 2-oxobutanoate, NH3, and H2O.
CAT and 3-MST represent two other enzymes involved in the biosynthesis of hydrogen sulfide; CAT is able to transfer the amine group from cysteine to a keto acid (for example α-ketoglutarate) forming 3-mercaptopyruvate. Subsequently, 3-MST leads to the formation of the persulfide, 3-MST-SSH, through the desulfuration of 3-mercaptopyruvate (Figure 1E). 3-mercaptopyruvate sulfur-transferase then transfers the sulfur from 3-mercaptopyruvate to sulfite or other sulfur acceptors or forms elemental sulfur. The reaction is catalyzed by CAT and 3-MST and produces sulfane sulfur (or bound sulfur); subsequently, a reduction of the sulfur atom or a release from the thiosulfate or persulfides leads to the production of H2S. Therefore, the presence of reductants [40] and specific enzymes such as thiosulfate sulfur transferase or thiosulfate reductase is necessary to obtain hydrogen sulfide [41]. The enzymes involved in hydrogen sulfide biosynthesis are widely distributed. Both CBS and CSE represent cytosolic enzymes. CBS was initially thought to be predominantly involved in the production of H2S in the brain, while CSE was thought to be predominantly involved in the synthesis of H2S in the heart and blood vessels [42]. Subsequent studies have established a broader distribution of these enzymes and in particular, it was seen that CBS is present in vascular endothelium, CAT and 3-MST are present in the vascular endothelium and the brain, and 3-MST, but not CAT, is present in the vascular smooth muscle [43]. CAT and 3-MST constitute both mitochondrial and cytosolic enzymes [44], though 3-MST is believed to be predominantly present in the mitochondrion [45].
PGG (propargylglycine), BCA (β-cyanoalanine), AOAA (aminooxyacetic acid), trifluoroalanine, and HA (hydroxylamine) represent the most commonly used agents that inhibit H2S biosynthesis; in particular PGG and BCA can be considered specific CSE inhibitors, while AOAA can be considered as a CBS-selective inhibitor [7,46]. In particular, PGG is a specific CSE inhibitor in comparison to CBS, but it is able to interact with different PLP-dependent enzymes that are not necessarily involved in H2S production. In a study reported in the literature that monitored vascular disposition in cerebellar slices and in intact mouse brains and that foresaw the use of two-photon intravital laser scanning microscopy, Morikawa et al. analyzed the pathway mediating hypoxia-induced cerebral vasodilation. It was established that either H2S, obtained by cystathionine β-synthase (CBS), or CO, formed by heme oxygenase (HO)-2hypoxia caused cerebral vasodilation. Hypoxia reduces CO generation by HO-2, an oxygen sensor. The constitutive CO physiologically inhibits CBS, and hypoxia increases the H2S levels that leads to the vasodilation of precapillary arterioles. It has been observed that mice with a targeted deletion of HO-2 or CBS showed changed vascular responses to hypoxia. Therefore, the imaging mass spectrometry of an intact adult brain cerebral cortex of HO-2–null mice showed an impaired ability to preserve ATP levels during hypoxia [47].
As previously reported, H2S is also involved in non-enzymatic reactions (Figure 1F). In particular, the non-enzymatic reduction of elemental sulfur leads to a minor endogenous source of hydrogen sulfide; this reaction occurs using reducing equivalents obtained from the oxidation of glucose in erythrocytes [48]. Human erythrocytes are able to obtain H2S in the presence of elemental sulfur or inorganic polysulfides. Non-enzymatic oxidation sulfides provide thiosulfate and these can be converted to sulfite by enzymatic reactions with thiosulfate reductase in the liver, kidney, or brain, or by thiosulfate sulfur-transferase in the liver. H2S can also be released from thiosulfate and persulfides. Garlic-derived organic polysulfides have been proven to release hydrogen sulfide in a thiol-dependent manner [49]. Garlic (Allium sativum) has long been considered beneficial as an antioxidant; recent studies have demonstrated several favorable effects of garlic derived from H2S production. Allicin (diallylthiosulfinate) is the best characterized naturally occurring H2S-donating garlic compound; it decomposes in water to form compounds such as diallyldisulfide (DADS) and diallyltrisulfide (DATS).
1.2.2. H2S Catabolic Pathways
The catabolism of hydrogen sulfide includes non-enzymatic and enzymatic reactions. In the first reaction of a catabolic pathway, H2S is rapidly oxidized, mainly in the mitochondria and in a non-enzymatic manner, to thiosulfate; subsequently, through an enzymatic reaction, thiosulfate is converted to sulfate and/or sulfite by TST (thiosulfate cyanide sulfur-transferase) (Figure 2A). Sulfite obtained by this reaction is then rapidly oxidized to sulfate; the latter is the major end-product of H2S catabolism under physiological conditions [50]. Another catabolic pathway of H2S is represented by the methylation to methanethiol and dimethyl sulfide mediated by TSMT (thiol-S-methyltransferase) (Figure 2B). This latter catabolic pathway occurs in the cytosol of different cells of the gastrointestinal tract [51,52]. The last catabolic reaction of H2S involves the methemoglobin that is able to bind to endogenous gases (for example CO and NO); moreover, it is able to bind H2S to give sulfhemoglobin (Figure 2C) [53].
Figure 2.

H2S catabolic pathways. (A) mitochondrial oxidation; (B) cytosolic methylation; (C) binding to hemoglobin. TST: thiosulfate cyanide sulfur transferase (rhodanese). TSMT: thiol-S-thiomethyl transferase.
The catabolism of H2S in the mitochondria is related to the coupled activity of several mitochondrial enzymes (SQR, ETHE1, SO, of which the initial one (SQR) is oxygen-independent) and their substrates (CoC, GSH glutathione, O2). In order to form thiosulfate, H2S is initially processed by SQR and GSH to form glutathione persulfide (GSSH) that serves as a substrate for both sulfite and thiosulfate formation. Alternatively, H2S can be processed by SQR and sulfite to form thiosulfate. Most of the catabolic reactions regarding H2S in mitochondria are enzymatic [17].
1.3. Hydrogen Sulfide: Biological Functions
The scientific literature concerning the biological roles for endogenous hydrogen sulfide is continuously growing. Several studies have reported that H2S performs physiological effects at a broad range of concentrations (10–300 μM) [5,6,7]. It is known that H2S participates in the regulation of numerous physiological responses, for example, anti-inflammation [54], oxidative stress [55], neuromodulation [56], vasoregulation [57], protection from reperfusion injury after myocardial infarction [58], and insulin resistance [59]. Furthermore, scientific research has continued to investigate H2S in relation to its involvement in various phases of cellular signaling, cell function, and cytoprotection.
1.3.1. Cardiovascular System
As reported in several studies, H2S has biological significance in the control of cardiovascular homeostasis. Particularly, it has been shown that H2S presents all the positive effects of NO without forming toxic metabolites [60]. Lately, it has been observed that the function of H2S in cardiovascular homeostasis is more important and crucial in cases of NO-mediated control compromise such as in the endothelial dysfunction. Hydrogen sulfide induces relaxing effects in the vascular smooth muscle and this effect has been observed in large vessels, for example, the rat thoracic aorta and portal vein and also in peripheral resistance vessels, which have a more substantial role than large conduit arteries in the modulation of vascular resistance and blood pressure [61]. l-cysteine represents a source of H2S; in fact, the vasorelaxing activity of l-cysteine is reduced by the CSE inhibitor PGG. Genetic deletion of CSE in mice significantly decreases H2S levels in the serum, heart, and aorta, and mutant mice lacking CSE presented with marked hypertension and a reduced endothelium-dependent vasorelaxant effect [62]. In fact, it is known that H2S relaxes blood vessels primarily, but not exclusively, by opening the KATP channels of the vascular smooth muscle cells. In addition to the vasorelaxing effect, H2S, as well as NO, is characterized by a broad range of other biological effects, which are significant for the polyhedric control of the cardiovascular system. For example, H2S is able to inhibit the platelet aggregation/adhesion induced by ADP, collagen, epinephrine, arachidonic acid, thromboxane mimetic U46619, and thrombin [63].
1.3.2. Immune System and Inflammation
The production and functional role of H2S is important in the several cell types classically related to innate immunity (such as neutrophils, eosinophils, basophils macrophages, dendritic cells, natural killer cells, mast cells,) and adaptive immunity (T and B lymphocytes). H2S is also associated with the progress of various inflammatory and immune diseases. In this context, H2S has physiological and pathophysiological roles especially in the oral cavity and in the colon, where endogenous bacteria produce H2S giving rise to a “H2S environment” to which the immune cells and the parenchymal cells are subjected; this “H2S environment” significantly regulates their viability and function. The development of spontaneous autoimmune disease, the onset, or the worsening of the severity of various immune-mediated diseases (such as autoimmune rheumatoid arthritis or asthma) could be connected to the downregulation of endogenous H2S-producing enzymes, or to the genetic defects in H2S biosynthetic enzyme systems. Small amounts of H2S, delivered by donors, enhance the function of various immune cells, and defend them against the dysfunctions caused by various noxious stimuli. The effects of H2S preserve the immune functions, increase the antimicrobial defenses, and exercise anti-inflammatory therapeutic effects in various diseases.
As is already known, nonsteroidal anti-inflammatory drugs (NSAIDs) are able to induce gastroenteropathy [64]. Studies reported in the scientific literature suggest that NSAIDs repress endogenous H2S synthesis by decreasing the expression of CSE. The additional decrease in H2S synthesis may sequentially cause an expansion in leukocyte adherence leading to the development of the gastric injury that has been noted subsequent to NSAID administration [65]. Equally, the administration of exogenous H2S decreases the capacity of these agents to affect gastric injury. Exogenous administration of H2S repressed NSAID-induced granulocyte infiltration, the expression of endothelial and leukocyte adhesion molecules, and the expression of TNFα (tumor necrosis factor α) [64]. It has been shown that leukocyte adhesion to the vascular endothelium produced by aspirin injury decreases after improvements in H2S bioavailability and that CSE inhibition with propargylglycine (PGG) impaired aspirin-mediated mucosal injury and inflammation. It has been also observed that the leukocyte expression of LFA-1 is repressed by exogenous H2S. It is also important to highlight a molecular characteristic of the H2S-induced anti-inflammatory effects: H2S donors reduced aspirin-induced leukocyte adhesion as a result of the activation of KATP channels and the inhibition of the CSE activity that encourages leukocyte adhesion. Further studies have shown that the co-administration of an H2S donor with an NSAID leads to the inhibition of NSAID-induced leukocyte adherence and a decrease in the gravity of gastric injury [66].
1.3.3. Respiratory System
The relaxant effects of H2S have also been explored on the rings of the bronchial smooth muscle of two rodent species. Fu et al. demonstrated that H2S produced a strong relaxation in the isolated bronchus rings from the mouse but generated only minor relaxation in guinea pig rings. H2S content and CSE activity were also improved in an isolated rat lung which presented with an ischemia/reperfusion injury, and a preventative perfusion with H2S reduced this injury, decreasing malondyaldehyde (MDA) production and increasing superoxide dismutase and catalase activity [67].
The endogenous CSE/H2S pathway seems to have an important role in pulmonary hypertension (PH). Specifically, this pathway was down-regulated in hypoxic PH (HPH), causing a reduction in endogenous H2S production in rat lung tissues as a result of oxidative stress. In lung tissues, the gene expression, and the activity of the CSE enzyme were suppressed during HPH, but the exogenous amount of H2S led to an improvement in CSE activity, an upregulation of CSE gene expression in the lung tissue, and a decrease in the remodeling of the pulmonary vascular structure during HPH [68]. Subsequently, after the treatment with exogenous H2S, HPH was reduced because of a direct scavenging of oxidized glutathione (GSSG), an improved total antioxidant capacity [69], and the inhibitory effect of H2S on pulmonary vascular inflammation, connected to a high I-B (inhibitor of NF-B) expression and a down-regulation of NF-B p65 expression [70]. Moreover, the CSE/H2S pathway seems to also have a favorable effect in asthma and chronic obstructive pulmonary disease (COPD), two of the most important obstructive airway diseases. In milder COPD, enhanced levels of H2S serum may have a beneficial effect in airway protection, antagonizing oxidative stress and airway inflammation and blocking the progress of COPD [71]. Scientific research has shown many positive effects for H2S in ischemia for several tissues. H2S has a broad range of protective functions related to ischemia/reperfusion (I/R) injury all through the body. The control of intracellular [Ca2+] through the stimulation of the KATP channel opening, anti-apoptotic pathway (ERK1/2/MAPK, PI-3 kinase/Akt, JAK-STAT) activation, mitochondrial protection through the preservation of ΔΨm and the inhibition of the mPTP opening, and the anti-inflammatory effects through the activation of eNOS and p38 MAPK represent the mechanisms of protection [72]. H2S has been also revealed to be protective against pulmonary ischemia injury and the endogenous CSE/H2S pathway could be implicated in the pathogenesis of lung I/R injury [67]. In a recent study, Jiang et al. showed that diabetes mellitus (DM) exacerbated lung I/R injury and that oxidative stress had a key role in this process. It was revealed that H2S had a protective effect against diabetic lung I/R injury by silencing oxidative injury. In this study, the mechanism by which H2S affected diabetic lung I/R injury was examined. Streptozotocin-induced type 2 diabetic rats fed a high-fat diet were exposed to GYY4137 (Figure 3), a slow-releasing H2S donor in the presence of and in the absence of EX527 (a SIRT1 inhibitor), and later exposed a surgical model of IR injury of the lung. Subsequently, the lung function, oxidative stress, cell apoptosis, and inflammation were assessed. It was observed that the damage to the lung SIRT1 signaling under type 2 diabetic conditions was additionally intensified by I/R injury. GYY4137 treatment significantly stimulated SIRT1 signaling and enhanced the I/R injury in the lung in type 2 DM animals through an improvement in the functional recovery of the lung, reducing oxidative damage, reducing inflammation, and destroying cell apoptosis. Nevertheless, these effects were mainly compromised by EX527. Furthermore, treatment with GYY4137 substantially stimulated the Nrf2/HO-1 antioxidant signaling pathway and improved eNOS phosphorylation; these effects were largely eliminated by EX527. These results suggest that GYY4137 treatment significantly reduced lung I/R injury under type 2 diabetic conditions via the activation of lung SIRT1 signaling. SIRT1 activation upregulated Nrf2/HO-1 and activated the eNOS-mediated antioxidant signaling pathway, consequently decreasing cell apoptosis and inflammation, and finally maintaining+ lung function [73].
Figure 3.

Chemical structure of GYY4137.
1.3.4. Central Nervous System
H2S exerts important regulatory effects in the control of important functions in the Central Nervous System (CNS). A high expression of CBS in the rat hippocampus and cerebellum has been observed [56]. H2S improves cAMP production, giving an enhanced sensitivity towards NMDA receptors to glutamate in CNS neurons [74]. This sensitization of the NMDA receptors, stimulated by concentrations of H2S between 50 and 160 μM, promotes the induction of hippocampal long-term potentiation, a process of synaptic plasticity implicated in the mechanisms of learning and memory [75]. The improved production of cAMP stimulates protein kinase A which controls brain function by intracellular protein phosphorylation; moreover, H2S may increase the reducing activity and defend neurons against oxidative stress through the activation of upstream receptor tyrosine kinase [76]. Lastly, some studies have reported that H2S reduces lipopolysaccharide (LPS)-induced NO production in the microglia through the inhibition of p38-MAPK [77] and that MAPKs regulate cellular activities [78]. The obtained data indicate an important role for H2S in the treatment of cerebral ischemia and neuro-inflammatory diseases.
1.3.5. Reproductive System
Studies in the scientific literature have reported several effects of exogenous H2S on mammalian reproductive systems. H2S has an important physiological role in the erectile response of the human corpus cavernosum due to the vasorelaxant effect on the penile artery. D’Emmanuele di Villa Bianca et al. demonstrated that l-Cys or NaHS produces a relaxation of the human corpus cavernosum strips in a concentration-dependent manner. l-Cys relaxation was inhibited by AOAA (aminoxyacetic acid; CBS inhibitor). It was shown that PAG (propargylglycine; CSE inhibitor) or AOAA potentiated the electrical field stimulation of human penile tissue. In rats, l-Cys and NaHS were shown to promote penile erection and the response to l-Cys was impeded by PAG. The acquired data suggest that the l-Cys/H2S pathway facilitates human corpus cavernosum smooth-muscle relaxation [79].
1.3.6. Gastrointestinal System
Several papers concerning the role of H2S in the gastrointestinal (GI) tract have been described in the scientific literature. Studies on H2S-induced chloride secretion proved that H2S is able to stimulate primary afferent nerve fibers in the mucosa/submucosa of the colon in guinea pigs and humans and this leads to increased chloride secretion. Hydrogen sulfide determines the contractile function either acting directly on the smooth muscle itself or by stimulating neurons in the enteric nervous system that influence smooth muscle function. Even if the effect of H2S on inflammation is controversial and many studies have demonstrated its anti-inflammatory effect at physiological concentrations, H2S can be considered as a proinflammatory mediator in abdominal sepsis, endotoxemia, and pancreatitis counter to its anti-inflammatory effects in animal models of gastritis and colitis [80,81,82].
In recent years, the role of H2S and other gaseous mediators (such as CO and NO) in digestive systems has been highlighted. The cytoprotective and anti-inflammatory properties of H2S are correlated to mucosal defense and ulcer healing in the GI system [83]. H2S is also involved in the physiology of the intestinal microbiota and in particular, it is able to inhibit pathogenic bacteria and to promote mucus production [84]. Furthermore, H2S-releasing derivatives of non-steroidal anti-inflammatory drugs (HS–NSAIDs) have been proven to reduce the gastric damage in comparison to the corresponding parent drugs [85]. H2S donors have also been shown to alleviate visceral pain and HS–trimebutine has been tested in clinical trials as an abdominal analgesic (NCT01926444) [86,87,88].
1.4. Hydrogen Sulfide and Diseases
Hydrogen sulfide is involved in several physiological and pathophysiological processes; this endogenous mediator is in fact associated with the homeostasis of the cardiovascular, renal, and central nervous systems and to dermatological diseases, various types of cancer, and viral infections as well as the SARS-CoV-2 infection that leads to COVID-19.
Hypertension, atherosclerosis, hyperhomocysteinemia, and diabetes represent cardiovascular diseases that are connected to vascular endothelial dysfunction; research has shown that H2S can be considered as a vasoprotective gasotransmitter in diseases where endothelial dysfunction is the central issue [89]. Moreover, morphological and functional modifications at the endothelial level correspond to the first events of cell senescence that inexorably result in vascular dysfunction and the subsequent atherosclerosis that profoundly impact cardiovascular health. In fact, myocardial hypertrophy and fibrosis can certainly arise and can lead to a compromise in the general cardiac output. Interestingly, hydrogen sulfide has been recently identified as a gaseous molecule able to manipulate the intracellular pathways implicated in cell senescence and so it is considered as a possible target for blocking cardiovascular diseases. In particular, the correlation between the decrease in endogenous H2S levels and the beginning of several cardiovascular cell senescence-related diseases has been demonstrated [90].
Numerous studies have clearly demonstrated that H2S has a considerable protective effect in myocardial ischemia. The processes for H2S cardioprotection involve the regulation of ion channels; antifibrotic and antiapoptotic effects; a reduction in oxidative stress and inflammatory response; the protection of mitochondria; the regulation of microRNA expression; and the promotion of angiogenesis. H2S donors could be used to develop effective drugs for the care of cardiovascular diseases such as myocardial ischemia [91,92].
Hydrogen sulfide has a crucial role in blood pressure control and renal physiology too. In the kidney, H2S is a significant regulator of vascular and cellular function, although the processes that influence the (sub)cellular levels of H2S are not exactly understood. H2S regulates systemic and renal blood flow, the amount of glomerular filtration, and the renin–angiotensin axis by direct inhibition of nitric oxide synthesis. In addition, H2S affects cellular events by altering protein activity through post-translational protein modification. This process represents persulfidation and is able to modulate protein localization, protein activity, and protein–protein interactions. Furthermore, acute kidney injury (AKI) caused by mitochondrial dysfunction, which happens in the course of hypoxia or ischemia-reperfusion (IR), is mitigated by H2S. H2S increases ATP production, inhibits the injury caused by free radicals, and controls endoplasmic reticulum stress during IR [93]. Therefore, forerunners for H2S endogenous synthesis, H2S donors, and natural plant-derivative compounds may serve as strategies to prevent hypertension and kidney disease [94]. Corvino et al. recently reported a survey of currently available H2S donors and their effects on cardiovascular disease; these H2S donors have been described as the basis of the mechanism that triggers the release of H2S and underline their possible use as favorable drugs in the treatment of cardiovascular diseases [95].
H2S, NO, and their crosstalk, have been described as helpful in Acute Kidney Injury (AKI), a syndrome mainly produced by ischemia and reperfusion (IR) injury, which momentarily impedes the blood flow, improves inflammation processes, and stimulates oxidative stress. Both these endogenous gasotransmitters act by reducing inflammation, regulating reactive oxygen species (ROS) concentrations, and modulating pro-inflammatory cytokines, as well supporting vasodilation and reducing hypertrophy, apoptosis, and autophagy [96].
Some studies reported in the scientific literature describe the correlation between H2S and cisplatin-induced nephrotoxicity. Precisely, the decrease in H2S by cystathionine γ-lyase (CSE) downregulation, subsequent to cisplatin treatment, may contribute to cisplatin-induced renal cell injury, probably through the increase in the production of endogenous reactive oxygen species (ROS); on the other hand, H2S prevents successive renal disorders through the inhibition of the activation of NADPH oxidase. Interestingly, GYY-4137, a slow-releasing H2S donor, has been proven to improve the anticancer activity of cisplatin in various cancer cell lines, maybe because of its specific anticancer effect. Nevertheless, the effectiveness of H2S donors in tumor-bearing animals continues to be examined in terms of renal protection, cancer inhibition, and later, cisplatin treatment. Moreover, more data regarding the use of polysulfide, a H2S derived compound, in the therapy of cisplatin-induced nephrotoxicity, have been additionally reported [97].
During recent years, a wide range of studies have focused on the function of H2S in the central nervous system (CNS), providing evidence that H2S has an important role in neuroprotection activities because it constitutes a signaling molecule that controls the function of the CNS. Neurodegenerative diseases are linked to endoplasmic reticulum stress (ERS) and protein misfolding. H2S regulates ERS and shows a broad variety of cytoprotective and physiological functions in CNS degenerative diseases. The neuroprotective effect of H2S for ERS has an important role in various CNS diseases as well as Parkinson’s disease, depression disorders, and Alzheimer’s disease [98].
In bone tissue, H2S has a cytoprotective role and stimulates bone formation; recently, scientific studies have established its role as a therapeutic agent in bone pathologies. H2S has been proven to stimulate several signaling pathways implicated in several phases of the processes of bone repair. In particular, H2S seems to be able to modulate osteogenic differentiation in osteoprogenitor cells through the induction of S-sulfhydration in TRP channels, increasing Ca2+ influx in the mesenchymal stromal cells (MSCs); H2S donors encourage the expression of osteogenic signaling by the stimulation of multiple signaling pathways at the transcriptional level. Concerning inflammation, hydrogen sulfide activates antioxidant signaling in osteoclasts (OCs), which includes the improved nuclear translocations of NRF-2 and leads to the inhibition of OC differentiation. Pharmacological administration of H2S has led to promising outcomes in preclinical studies in the treatment of systemic bone diseases, for example, osteoporosis; nevertheless, local delivery of H2S offers further opportunities for treatment. Moreover, new H2S donors and biomaterials have recently been studied, paving the way for the advancement of H2S-releasing procedures for bone re-forming medicine [99].
Alterations in H2S production are involved in several dermatological diseases, for example, psoriasis, melanoma, and other dermatoses. The consequences of H2S in the skin involve the stimulation of keratinocyte differentiation; the control of vasodilatation; wound healing; the regulation of apoptosis; the modulation of pruritus; and the resolution of inflammation. Regarding the H2S pathway in the skin, the information available references the connection between this molecule and the physiological and pathological effects. Generally, most of the effects of H2S are clearly concentration or dose-dependent. Due to the important role of H2S in skin physiology, it is not unexpected that modified H2S production leads to the pathogenesis of several cutaneous diseases. Many H2S donors have been used in experiments, and different results have been acquired. Nevertheless, these compounds present different intrinsic characteristics such as the capacity to enter the cell or organelle specifically, H2S-releasing kinetics, and pH dependence, but certainly, these chemical entities include a new family of potential therapeutic tools for the treatment of various skin diseases by targeting their differential etiological aspects. Obviously, the development of H2S donors will necessarily rely on an better understanding of the exact role of H2S and its goals in the various phases of skin biology [100].
Several studies reported in the scientific literature have shown an effective role of H2S in the framework of cancer biology. CSE, CBS, and 3MST, the three H2S-producing enzymes, are expressed in several types of cancer. Furthermore, the inhibition of CBS leads to anti-tumor activity especially in breast cancer, ovarian cancer, and colon cancer, while the effect of CSE or 3-MST inhibition has been largely unexplored in cancer cells. Interestingly, H2S donation has been shown to induce cancer cell apoptosis in in vitro and in vivo studies. In particular, low levels of H2S may show pro-cancer effects, while higher levels of H2S may lead to cancer cell death. This leads us to assume that the inhibition of H2S biosynthesis and H2S supplementation work in two different ways in relation to cancer treatment. This enigmatic role of H2S is interesting in relation to the development of innovative CBS inhibitors, H2S donors, and H2S-releasing hybrids that could therefore have potential for cancer therapy [101].
Finally, it is possible to affirm that hydrogen sulfide represents a natural defense against the infections from enveloped RNA viruses and is implicated in COVID-19. It has previously been reported that H2S inhibits the growth and pathogenic mechanisms of several enveloped RNA viruses, and it has been observed that H2S plasmatic levels are elevated in COVID-19 survivors compared to fatal cases [102]. The release of H2S is triggered by carbon monoxide (CO) from the catabolism of heme by inducible heme oxygenase (HO-1) and heme proteins have the catalytic activity required for H2S signaling by protein persulfidation. Subjects with a long promoter for the HMOX1 gene, coding for HO-1, have a lower efficiency of this mechanism. SARS-CoV-2 prevents the formation of the heme of hemoglobin and other hemeproteins, so impeding both the signaling and release of H2S. Deficiency of the H2S-induced persulfidation of the KATP channels of leucocytes triggers the adhesion and release of the inflammatory cytokines, lung infiltration, and systemic endothelial damage with hyper-coagulability. These outcomes mainly justify the sex and age distribution, clinical manifestations, and co-morbidities of COVID-19 [102]. The involvement of H2S in COVID-19 therapy is also supported by its anti-inflammatory and antiviral functions. As is already known, H2S has a pivotal role in regulating the inflammatory and pro-inflammatory cytokine cascade and certain H2S donors are effective in the treatment of acute lung inflammation. The latest studies have elucidated the mechanism of action for the antiviral effects of hydrogen sulfide. Moreover, certain preliminary clinical results have indicated an inverse relationship between endogenous H2S levels and the severity of COVID-19. H2S donors, then, could represent a potential therapeutic opportunity for the treatment of COVID-19 [103]. The alteration of H2S levels in COVID-19 represent further evidence of the need to regulate the homeostasis of this gasotransmitter, considering that the alteration of H2S levels has an important role in several diseases [89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104].
Finally, H2S is especially controlled in experimental glaucoma. By scavenging reactive oxygen species and distending retinal vessels, H2S protects RGCs from pressure and oxidative stress both in vitro and in vivo. Consequently, H2S is considered to be a new neuro-protectant in glaucoma [104].
2. H2S Donors
2.1. Gaseous H2S
On the basis of the broad and active physiological effects of H2S, some research groups have described the protective role of H2S donors in various disease models. H2S gas has been clearly applied in these fields. The most prominent method of delivery is the direct inhalation of this gas. The delivery method has been often assessed, even though H2S is a toxic and flammable gas. It has been demonstrated that gaseous H2S generates a suspended animation-like state in mice [30] and it has been proposed that these effects are caused by the reversible and competitive inhibition of the mitochondrial enzyme cytochrome c oxidase [31]. Moreover, H2S gas demonstrated a reversible reduction in motor activity and body temperature with a subsequent growth in blood sulfide concentration in mice [105,106]. Consequently, inhalation of H2S is used as a technique to inhibit pulmonary and systemic inflammatory reactions in the course of physiological stress in hypothermic and normothermic mice [107,108]. The reversible H2S-induced hibernation state has been shown to protect mice from lethal hypoxia [109]. H2S gas can also promote glucose uptake and provide amelioration in type II diabetes [110]. H2S inhalation allowed a modulation of the delivery by precisely modulating pressure over time and it bypassed some of the issues with intravenous delivery as well as the oxidation and volatilization of suspended H2S. On the other hand, H2S inhalation has various inherent shortcomings because of the toxicity and flammability of the gas, including gas storage, secure administration, and targeting. From the experimental point of view, it is difficult to consider H2S gas as an ideal drug because of the problems in acquiring accurately regulated concentrations and for the potential toxic effect of an excess of H2S.
2.2. Inorganic Salts
Inorganic salts, as well as sodium hydrosulfide (NaHS), sodium sulfide (Na2S), and calcium sulfide (CaS), are believed to be exogenous H2S donors. These inorganic salts, which represent standard tools, have been long employed in biological studies to investigate the therapeutic effects of exogenous hydrogen sulfide, although several significant limits have been proved.
Indeed, sulfide salts hydrolyze instantaneously following dissolution in water, establishing an equilibrium between hydrogensulfide ions (HS−), sulfide ions (S2−), and molecular hydrogen sulfide (H2S) which depend on some parameters such as pH, temperature, and pressure [111]. When this equilibrium is determined, volatilization of H2S appears, thus reducing the general concentration of the sulfur species in the solution. Moreover, HS air oxidation−, catalyzed by metals in water, lowers the concentration of H2S solutions [112]. These chemical limitations restrict the use of salts in clinical settings even though the administration of sulfide salts to animals, tissues, or cells has been demonstrated to have protective effects for a large number of diseases. Some researchers have highlighted the short- and long-term cardioprotective effects of H2S using Na2S as a H2S donor in a mouse model of myocardial ischemia injury (MI/R). The results have suggested that a 7-day treatment of Na2S resulted in a reduction in the dilatation of the left ventricle, a loss of cardiac hypertrophy, and an increase in cardiac function. Moreover, treatment with Na2S decreased the oxidative stress related to heart failure [113]. Aqueous NaHS solution delivered in aortic rings led to a 60% increase in relaxation, proving the vasorelaxant effects of H2S [114]; this salt reduced ischemia–reperfusion injury [115], and protected against hemorrhagic shock [116] and myocardial infarction (MI) [117].
Moreover, sulfide salts have protective effects for other diseases as well as inflammation [118]. Osteoarthritis (OA) is a form of arthritis characterized by degenerative and inflammatory processes and by higher levels of pro-inflammatory proteins (IL-6 and IL-8). A study reported in the literature demonstrated that the short-term treatment of human cells with NaHS is enough to down-regulate IL-6 and IL-8 expression, which in turn may cause anti-inflammatory effects of H2S against OA [119]. However, it has also been observed that the anti-inflammatory effects of NaHS translated into pro-inflammatory effects when the NaHS incubation time was protracted (1 h). In fact, it has been reported that sulfide salts as H2S donors showed both anti-inflammatory and pro-inflammatory effects.
Some researchers have indicated that the leukocyte-regulated inflammation of the knee joint was significantly hindered after Na2S injection [120]. In an independent study, Chen et al. determined that the administration of NaHS into the lungs in an asthmatic rat model attenuated inflammation [71]. Through the use of a smoke inhalation-induced lung injury model, Esechie et al. proved that Na2S exhibited protective actions against severe lung injury [118]. These results have suggested that H2S (using NaHS or Na2S) is a potent anti-inflammatory agent.
In contrast, the pro-inflammatory effects of H2S have also been highlighted. Indeed, Li et al. proved that H2S exhibited pro-inflammatory effects in an animal model of endotoxic shock [121]. Furthermore, plasma H2S concentrations, the synthesizing ability of H2S in tissue, and CSE expression were all proven to be enhanced in several animal models of inflammation [122,123,124]. This conclusion has indicated the potential pro-inflammatory effects of H2S.
In a separate study, the administration of NaHS solution by intravenous injection alleviated the degree of ALI by decreasing IL-6 and IL-8 levels while concurrently increasing IL-10 levels in the plasma and lung tissue [125]. This study also substantiated the hypothesis that the down-regulation of endogenous H2S levels in the cardiovascular system is present in ALI pathogenesis.
Parkinson’s disease (PD) manifests a progressive loss of dopaminergic neurons in the substantia nigra and a depletion of the neurotransmitter dopamine in the striatum [126]. Hu et al. indicated that a systemic administration of H2S (using NaHS) could reduce behavioral symptoms and dopaminergic neuronal degeneration in 6-hydroxydopamine (6-OHDA)-induced PD models [127]. By employing such models, they determined that endogenous H2S production is lowered during the development of PD, and the results have suggested that the administration of H2S could have therapeutic benefits for Parkinson’s disease.
Comparatively, H2S has proven to be useful for Alzheimer’s disease (AD). The amount of H2S biosynthesis in the brain is lowered in AD patients and the levels of H2S in the plasma are linked to the severity of AD [44]. Giuliani et al. observed how treatment with a H2S donor induced neuroprotection and slowed down the progression of AD. Studies with sodium hydrosulfide and Tabiano’s spa-water were conducted in three experimental models of AD. Short-term and long-term treatments with sodium hydrosulfide and/or Tabiano’s spa-water significantly protected against impairments in learning and memory in rat models of AD induced by an injection of β-amyloid1–40 (Aβ) or streptozotocin in the brain, and in an AD mouse model harboring the human transgenes APPSwe, PS1M146V, and tauP301L (3xTg-AD mice). The improvement in behavioral performance was related to the size of the Aβ plaques in the hippocampus and the preservation of the morphological picture, as found in AD rats. In addition, the reduced concentration/phosphorylation levels of proteins were proven to be essential in AD pathophysiology. Specifically, the amyloid precursor protein, presenilin-1, Aβ1-42 and tau phosphorylated at Thr181, Ser396 and Ser202, were identified in 3xTg-AD mice treated with spa-water. The excitotoxicity-triggered oxidative and nitrosative stress was found in 3xTg-AD mice, as indicated by the decreased levels of malondialdehyde and nitrites in the cerebral cortex. The limited activity of c-jun N-terminal kinases, extracellular signal-regulated kinases, and p38 in the hippocampus presented a well-established role both in the phosphorylation of the tau protein and in inflammation and apoptosis. Consistently, the decrease in the tumor necrosis factor-a level, the up-regulation of Bcl-2, and the down-regulation of BAX and the downstream executioner caspase-3 in the hippocampus of 3xTg-AD mice after treatment with Tabiano’s spa-water implied that it is also able to modulate inflammation and apoptosis. These results indicate that appropriate treatments with H2S donors and Tabiano’s spa-waters and other spa-waters rich in H2S content might represent an innovative approach to slow down AD progression in humans by targeting multiple pathophysiological mechanisms [128].
The low pharmacokinetic profile of sulfide salts limits the use of these salts as potential therapeutics and has led to the research into innovative organic H2S donors, including naturally occurring donors and synthetic compounds.
2.3. Natural H2S Donors
As is already known, garlic and onions contain substances that have benefits for the prevention or treatment of several diseases such as cardiovascular disease, hypertension, thrombosis, and diabetes [129]. H2S-releasing compounds from garlic extracts seem to be essential elements; in particular, these compounds are byproducts that originate from metabolism of thiosulfinates (R–SO2–SR) [130]. The integral garlic contains γ-glutamil-S-alk(en)yl-l-cysteines that undergo hydrolysis and oxidation reactions which lead to alliin; the latter is converted into allicin through a reaction catalyzed by alliinase. Allicin (1) represents the main sulfur compound and the most common thiosulfinate; the instant decomposition of this compound in aqueous media leads to some oil-soluble compounds such as diallyl sulfide (DAS, 2), diallyl disulfide (DADS, 3), and diallyl trisulfide (DATS, 4) (Figure 4) [131]. γ-glutamil-S-alk(en)yl-l-cysteines can also lead to water soluble organo-sulfur compounds including S-allyl cysteine (SAC) and S-allyl mercaptocysteine (SAMC).
Figure 4.

Chemical structures of some garlic-derived sulfur compounds.
Research on DAS, DADS, and DATS has shown that human red blood cells transform these compounds into H2S in the presence of free thiols and that treatment of aortic rings with these compounds leads to vasorelaxation, similar to experiments using sulfide salts [49]. Several polysulfide species have also been extracted from a variety of other plants; however, different from garlic-derived donors, they have not currently been evaluated [132]. Many studies reported in literature regarding the garlic-derived H2S donors have focused on the mechanisms through which these donors allow the release of H2S in the presence of thiols. Benavides et al. reported that DADS and DATS can quickly release H2S by means of a thiol-dependent mechanism [49]. In particular, in the presence of GSH, DADS leads to S-allyl glutathione (GSA) and the intermediate allyl perthiol (ASSH), which reacts with GSH, thus producing H2S and S-allyl glutathione disulfide (GSSA). Moreover, this compound can also undergo α-carbon nucleophilic substitution, swiftly generating H2S. This mechanism has been proven to be valid, except for DADS. Concerning this aspect, it has been shown that DADS releases H2S very slowly thanks to an α-carbon nucleophilic substitution [133]. The previous misunderstanding, (DADS as a fast H2S donor) was due to DATS contamination in commercial samples of DADS; indeed, among these compounds, DATS is responsible for the rapid H2S release. DATS can give two possible thiol−disulfide exchange reaction pathways: primarily, the nucleophilic attack of GSH on allylic sulfur of DATS that generates GSSA and ASSH, with the latter possibly reduced by GSH and then releasing H2S; further, the nucleophilic attack of GSH on the central sulfur atom of DATS can lead to the production of ASH and GSSSA. The latter rapidly produces hydrogen sulfide.
Another class of natural H2S donors is composed by some isothiocyanates (Figure 4); these compounds represent natural donors that have attracted attention due to their biological and pharmacological effects in the prevention of important human diseases, as well as cancer, neurodegenerative processes, and cardiovascular diseases [134]. The isothiocyanate moiety has been thoroughly investigated due to its use in synthetic molecules for structure activity relationship studies. In a recent study, the hydrogen sulfide-releasing properties of some synthetic aryl isothiocyanate derivatives were reported, indicating that the isothiocyanate function can be considered as a suitable slow hydrogen sulfide-releasing moiety, along with the pharmacological potential typical of this gasotransmitter. Many isothiocyanate derivatives (as a result of the myrosinase-mediated transformation of glucosinolates) are well-known secondary metabolites of plants belonging to the family Brassicaceae, a large botanical family comprising of many edible species. The phytotherapeutic and nutraceutic usefulness of Brassicaceae in the prevention of important human diseases, such as cancer, neurodegenerative processes, and cardiovascular diseases has been widely discussed in the scientific literature. Although these effects have been associated with isothiocyanates, by and large, the exact mechanism of action is still unknown.
In a recent experimental study, the hydrogen sulfide-releasing capacity of some important natural isothiocyanates has been investigated (AITC (5), ERU (6), BITC (7), HBITC (8)—Figure 5) by using in vitro amperometric detection. Some of the tested natural isothiocyanates exhibited a significant release of hydrogen sulfide, thus implying that hydrogen sulfide may be, at least in some measure, a relevant player accounting for several biological effects of Brassicaceae [135].
Figure 5.

General structure of isothiocyanates and the chemical structures of natural isothiocyanates tested in [135].
In addition, it has been proven that some selected aryl isothiocyanates are able to release H2S in a biological environment and produce a vasorelaxant effect in rat aortic rings, strongly antagonized by a specific Kv7-blocker, and in the coronary vascular bed, causing an increase in the basal coronary flow [136]. Furthermore, the H2S-releasing properties of the secondary metabolites of Brassicaceae, isothiocyanates, are also derived from the quick reaction with thiols, namely cysteine. The Cys-ITC adduct formed undergoes an intramolecular cyclization followed by a release of an organic amine R−NH2 and raphanusamic acid (RA), on the one hand, and H2S and 2-carbylamino-4,5-dihydrothiazole-4-carboxylic acid, on the other hand [137].
Although naturally occurring H2S donors represent an attractive tool for in vivo studies, the tendency of these compounds to also bring about some byproducts, which are not related to H2S production, has prompted researchers to develop novel synthetic H2S donors with more favorable pharmacokinetic and safety profiles.
Indeed, synthetic donors, which provide tunable H2S release rates via structural modifications with discrete byproducts, allow for a more in-depth analysis of the physiological roles of H2S and, optimistically, clinically relevant H2S-releasing prodrugs.
In this review, H2S donors are presented according to their main role in several contexts: cardiovascular diseases, cancer, nervous system diseases, gastrointestinal diseases, dermatological diseases, viral infections, and other diseases.
3. H2S Donors and Diseases
3.1. H2S Donors and Cardiovascular Diseases
As already reported, the current scientific literature has proven the protective actions of H2S in cardiovascular diseases, for instance hypertension, atherosclerosis, cardiac hypertrophy, heart failure, and myocardial ischemia/reperfusion (I/R) injury. The involvement of H2S in the cardiovascular system is due to its role as a blood pressure and heart rate regulator, an activator of angiogenesis, and a basal vasorelaxant agent. Furthermore, it has been proven that antioxidation, the inhibition of cell apoptosis, pro-angiogenesis, the anti-inflammatory effect, and the regulation of ion channels represent the mechanisms of action that lead to cardioprotective activity [95].
Based on the involvement of H2S in the cardiovascular system, scientific research has shown a strong interest in the development of H2S donors, which could represent biological instruments and encouraging cardioprotective agents.
3.1.1. Lawesson’s Reagent and Derivatives
Lawesson’s reagent (2,4-Bis(4-methoxyphenyl)-1,3,2,4-dithiadiphosphetane-2,4-disulfide—LR, 10) is an extensively applied sulfurization reagent in organic synthesis [138] and a H2S donor. LB is simply synthesized through a reaction between anisole (9) and phosphorus pentasulfide (P4S10) (Figure 6) [139,140].
Figure 6.

Chemical synthesis of Lawesson’s Reagent (10) and GYY4137 (11).
LR is a known, commercially available compound used for the thionation of alcohols, ketones, esters, and amides to obtain the sulfur analogs [138]. In aqueous media, this compound releases H2S through a hydrolysis release mechanism and the release time is greater than that of the sulfide salts.
It has been seen that the oral administration of Lawesson’s reagent, preceding alendronate-induced gastric damage, restricted successive gastric impairment [141]. An increase in GSH levels in rats handled with LR that can be ascribed to the reduction in oxidative stress and inhibition of neutrophil infiltration has also been observed. In a different in vivo study, exogenous H2S delivery from LR increased leukocyte adhesion and neutrophil migration to the area where sepsis was stimulated in mice, increasing the global survival probability of the affected mice [142]. Moreover, it was observed that the mortality rate of the mice during the experiment was improved, with a reduction in endogenous H2S production using a CSE inhibitor. Nevertheless, LR releases H2S through spontaneous and uncontrollable hydrolysis in aqueous solution. This kind of H2S release and the poor solubility of Lawesson’s reagent in aqueous solutions restricts its applications.
GYY4137 (Morpholin-4-ium-4-methoxyphenyl-(morpholino)-phosphinodithioate; 11), is a water-soluble Lawesson’s reagent derivative and one of the first slow-releasing H2S donors developed [143,144,145]. This reagent is the compound that is most used to examine the role of H2S in the biological districts. It can be synthesized by reacting Lawesson’s reagent [139,140] with morpholine in methylene chloride at room temperature (Figure 6) [143].
GYY4137 exhibits anti-inflammatory, antioxidant, and anticancer properties [146]. In addition, this reagent triggers the activation of the vascular smooth muscle KATP channels and relaxes rat aortic rings and the renal blood vessels that lead to anti-hypertensive activity [143]. Furthermore, GYY4137 interfered with platelet activation and adhesion molecule-mediated aggregation reducing the formation of the microvascular thrombus and stabilizing atherosclerosis plaque [147]. It seems to protect against diabetic myocardial I/R injury (activating the PHLPP-1/Akt/Nrf2 pathway) [148] and against cerebral I/R injury (via the p38 MAPK mediated anti-apoptotic signaling pathways) in rats [149]. It has been proven that GYY4137 slowly releases H2S upon hydrolysis. To explain the two-step hydrolytic decomposition pathway of the slow-release H2S donor GYY4137, a combination of NMR spectroscopy, mass spectrometry, and chemical synthesis has been used and the key decomposition product has been also obtained by an independent synthetic strategy. The (dichloromethane-free) sodium salt of the phosphonamidodithioate GYY4137 has also been made as a pharmaceutically more appropriate salt. It has been shown that the decomposition product does not produce H2S and does not have cytoprotective or anti-inflammatory effects in oxidatively stressed human Jurkat T-cells and LPS-treated murine RAW264.7 macrophages, in contrast with GYY4137 and its sodium salt. The decomposition product signifies a helpful control compound for establishing the biological and pharmacological effects of H2S generated from GYY4137. Whilst GYY4137 certainly represents a useful biological compound, it presents some disadvantages that mean the attribution of biological effects to GYY4137-derived H2S is undefined because of the probable concurrent metabolization of dichloromethane to CO, a gasotransmitter with biological effects similar to H2S [150,151].
With the aim to overcome the described undesired events, structural modifications of GYY4137 have been designed and the subsequent analogs have been synthesized and examined. O-aryl- and alkyl-substituted phosphorodithioates (12) were designed as H2S donors by switching the P-C bond in GYY4137 with O-substitution (Figure 7) [152]. These H2S donors did not significantly release hydrogen sulfide. In fact, O-arylated donors demonstrated a slow production of H2S, while O-alkylated donors showed very low H2S generation.
Figure 7.

General chemical structures of O-aryl and O-alkyl GYY4137 derivatives.
Bearing in mind the GYY4137 structure, JK donors (15, 16), a novel kind of phosphonamidothioate, were obtained [152]. The synthesis of these compounds started from the treatment of phenylphosphonothioic dichloride (13) with 3-hydroxypropionitrile and, subsequently, with a selected C-protected amino acid (as well as glycine, phenylalanine, valine, alanine, and proline) in the presence of triethylamine. The obtained intermediate (14), after a LiOH-mediated hydrolysis in MeOH, provided the desired compounds (15, 16) (Figure 8).
Figure 8.

Synthesis of JK donors.
JK donors are the first class of pH controllable H2S donors. These compounds give higher and faster H2S releasing rates in aqueous media and under acidic conditions, while they have slower and lower release rates at a neutral and mildly basic pH. These donors are characterized by a novel mechanism of release, characterized by the protonation of phosphonamidothioates that give the corresponding phosphorothiols and that, after the cyclization via nucleophilic addition of the carboxylic acid group, lead to the breaking of the P-S bond and to H2S release. JK-1 and JK-2, the two main donors of the series (Figure 8), had protective effects on cellular and murine models of myocardial I/R injury and significantly reduced infarct size and the circulating troponin-I level [153].
Ammonium tetrathiomolybdate ((NH4)2MoS4, ATTM) is a copper chelator and represents another pH dependent H2S donor [154]. It is able to release H2S even under strong acidic conditions (5% H2SO4).
ATTM is a water-soluble and slow releasing H2S donor; studying the gaseous release in different pH buffers (from 5 to 8), it has been shown that the level of H2S at pH 5 is considerably higher than the levels at other pH values [155]. Furthermore, it has been observed that H2S concentrations were stable for long time in all solutions, demonstrating that ATTM is a stable and slow releasing H2S donor.
ATTM had protective effects on the heart and brain in rat models of myocardial and cerebral I/R injury and reduces I/R injury [156]. In fact, certain studies are currently evaluating the potential dual role of ATTM as a cardioprotective and anticancer agent in patients with cancer treated with doxorubicin or Adriamycin [157,158,159]. Moreover, ATTM has recently progressed in clinical trials for the treatment of breast cancer because of its copper depletion effects [160].
3.1.2. DTTs
DTTs (1,2-Dithiole-3-thiones, 17) are another class of hydrolysis-triggered H2S donors. Even if their H2S-releasing mechanism is not totally delineated, it is commonly agreed that the production of H2S from DTTs occurs via hydrolysis (Figure 9).
Figure 9.

H2S release from DTTs.
It has been proven that DTT derivatives progressively release H2S, converting it into the corresponding 1,2-dithiole-3-one (18) with heating to 120 °C and in aqueous solution [161].
ADT (Anethole dithiolethione, 19) and ADT-OH (5-(p-hydroxyphenyl)-3H-1,2-dithiole-3-thione, 20), a O-demethylated ADT derivative, are the most employed DTTs as H2S donors (Figure 10).
Figure 10.

Chemical structures of ADT and ADT-OH.
In terms of the mechanism of H2S release by this class of compounds, it must be specified that to date not all the steps, metabolic and/or chemicals, have been elucidated. In particular, important information comes from the observation that S-diclofenac releases H2S in a time-dependent manner in both rat plasma and liver homogenates. This implies a metabolic step as the initiation of the release mechanism. A similar result is also obtained by the incubation of ADT-OH. To date, the nature of this activation step, which does not involve enzymes belonging to the esterase class, remains unidentified [162].
The synthesis of DTTs is easy and provides the reaction of anethole with elemental sulfur; these compounds can also be easily added to other molecules to obtain drug–DTT conjugates. Nevertheless, different synthetic methods have been proposed for their preparation [163]. The most used synthetic strategy is based on the dehydrogenation and sulfurization of an allylic methyl group (a) which involves treatment with elemental sulfur or phosphorus pentasulfide (Figure 10). On the other hand, β-ketoesters (b) have been reacted with Lawesson’s reagent to provide the requested DTTs (17) [142,164,165,166] (Figure 11).
Figure 11.

Examples of synthetic strategies used to obtain DTTs.
Several studies have reported the biological effects and the activity of ADT and ADT-OH correlated with the H2S-releasing properties. In particular, ADT represents an FDA approved drug; this compound is able to promote bile secretion, restoring salivation and reducing dry mouth in patients that present with chemotherapy-induced xerostomia [167]. ADT-OH indeed has been proven to decrease cell viability through the inhibition of histone deacetylase [168,169] and NF-kB activation [170].
H2S donor hybrids (21) have been achieved through the copulation reaction between the hydroxylic group of ADT-OH (20) with commercially available drugs and the obtaining compounds have been examined for their H2S-releasing properties and therapeutic activities [12]. The reaction between ADT-OH and active compounds, in the presence of N,N’-dicyclohexylcarbodiimide (DCC) and 4-dimethylaminopyridine (DMAP), generally leads to the production of the considered H2S donor hybrids (Figure 12) [162].
Figure 12.

General synthetic strategy used to obtain HS–NSAIDs.
Non-steroidal anti-inflammatory drugs (NSAIDs), as well as ibuprofen and aspirin, have been broadly applied for inflammation treatment [171]. Some studies have revealed that the unpleasant effects induced by NSAIDs (for example, stomach bleeding and gastrointestinal ulceration) can be substantially diminished by combining DTT molecules with the parent NSAIDs [12]. For example, a H2S-aspirin hybrid has been synthesized (ACS-14, S-aspirin, MZe786 (22); Figure 13) and the negative gastric effects caused by ACS-14 and aspirin in rats have been compared [172]. It has been shown that after oral administration of aspirin for 7 days, an extensive hemorrhagic lesion in the stomach of rats occurred. By contrast, rats given an equivalent dose of ACS-14 did not present with gastric damage. Then the H2S levels in plasma before and after ACS-14 were compared, and the obtained results revealed that drug administration led to a three-fold increase in the H2S concentration in the plasma. These results indicate that the advantages of ACS-14 in gastric protection are probably correlated with the increase in H2S formation. ACS-14 has also shown very important protective activities in pathological cardiovascular alterations in rats induced by a GSH-synthase inhibitor (BSO, buthionine sulfoximine) by improving systolic blood pressure and reducing heart rates. Furthermore, BSO-induced hypertensive effects were reduced by oral administration of ACS-14, unlike aspirin which did not have this effect. Additionally, administration of ACS-14 decreased myocardial I/R injury in BSO rats [173,174,175].
Figure 13.

Chemical structures of representative H2S-NSAIDs.
Hypertensive disorder in pregnancy is a main cause of maternal and perinatal mortality. Soluble Flt-1 (sFlt-1) is a protein whose values are elevated in preeclampsia and may continue to be high during postpartum in women with a preeclampsia diagnosis. Heme oxygenase-1 (Hmox1/HO-1) is another protein that has protective effects versus oxidative stimuli and is altered in the placenta of pregnant women with preeclampsia. Ahmed et al. assumed that soluble Flt-1 impeded cardiac mitochondrial activity in HO-1-deficient mice. It was demonstrated that the deprival of HO-1 perturbed cardiac mitochondrial respiration and decreased mitochondrial biogenesis. The overexpression of sFlt-1 caused the inhibition of cardiac mitochondrial activity in Hmox1+/− mice. This research revealed that MZe786 restored mitochondrial activity through the stimulation of cardiac mitochondrial biogenesis and antioxidant protection in Hmox1+/− mice and in Hmox1+/− mice subjected to a high sFlt-1 environment [176,177]. Moreover, Ahmed et al. determined the therapeutic value of MZe786 in HO-1 haploid-deficient (Hmox1+/−) pregnant mice in a high sFlt-1 environment. It was demonstrated that the animal model used presented critical preeclampsia symptoms and a decrease in antioxidant genes. MZe786 reduced preeclampsia-stimulating antioxidant genes and then reduced hypertension and renal damage. MZe786 was also better for fetal outcomes in comparison with aspirin alone and was shown to be a good therapeutic agent in preventing preeclampsia [178].
Moreover, H2S-releasing diclofenac (ACS-15, S-diclofenac (23); Figure 13) has been investigated. In some studies ACS-15 demonstrated an enhanced anti-inflammatory effect in comparison with diclofenac. ACS-15 considerably decreased the lipopolysaccharide-induced infiltration of neutrophils into the liver and the lungs without increasing leukocyte adherence, as instead diclofenac [66,171].
ATB-429 (mesalamine derivative (24); Figure 13) represents a hybrid drug from mesalamine that has been used in animal models of Crohn’s disease and ulcerative colitis. In preclinical studies concerning the visceral pain associated with inflammatory bowel disease, this compound was more successful than mesalamine [86,179].
Beside NSAIDs, other DTT-H2S donor hybrids have been developed. As is already known, L-DOPA represents a drug used in the clinical treatment for Parkinson’s Disease and dopamine-responsive dystonia. Some side effects of long-term L-DOPA treatment as well as dyskinesia seem to be correlated to the oxidative stress that leads to the loss of neurons [180,181]. Because H2S diminishes oxidative stress, L-DOPA coupled with a H2S-releasing moiety could lead to improved bioactivity. In fact, Lee et al. synthesized DOPA–DTT conjugates and studied their antioxidant effects [182]. The hybrid compounds (such as ACS-84 (25); Figure 13) released H2S in the mitochondria. Additionally, this compound has been shown to inhibit MAO B, the dopamine metabolizing enzyme; therefore, it could be useful to reestablish the Parkinson’s Disease-depleted dopamine levels in the brain.
NO-NSAIDs represent another class of hybrid drugs well studied where NO is released [183] (Figure 14). NO and H2S share several comparable actions, and new NO–H2S-releasing hybrids (for example NOSH-1(26)) have been designed and studied (Figure 13) [174,184,185]. These compounds are able to induce higher NO and H2S concentrations in vivo and have shown important activities related to the inhibition of the growth in cancer cell lines and anti-inflammatory activities in animal models.
Figure 14.

Chemical structures of representative NO-H2S-releasing hybrids, PEG-ADT, and AP-39.
Recently, DTT-based polymeric H2S donors have been reported [186]. DTT has been conjugated to poly(ethylene glycol) polymer, obtaining PEG-ADT (27; Figure 14) that represents a H2S donor that demonstrates a lower and slower H2S release when compared to DTT alone, probably as a result of the steric hindrance of PEG. PEG-ADT has also been shown to have fewer toxic effects as compared to the parent donor compound; this important characteristic is probably due to the co-localization of the PEG-ADT conjugate within the endo-lysosomes without escaping to the cytoplasm. It is important to consider that even if DTTs have been asserted as H2S donors, it is not clear if their effects are related to H2S; in fact, DTTs can considerably improve GSH levels in vivo by inducing the enzymes involved in the preservation of the balance of GSH [187]. It is probable that the higher GSH level contributes much more than H2S to those pharmaceutical effects.
AP-39 (28) is another mitochondria-targeted H2S donor hybrid, obtained from the reaction of triphenylphosphonium (TPP) with ADT-OH [188]; it has been investigated for its H2S-releasing properties and pharmacological effects (Figure 14) [189]. After being rapidly up-taken and accumulated in a mitochondrial membrane-potential (ΔΨ) dependent-manner, the AP-39-induced H2S release in the mitochondria was confirmed by several fluorescent probes. It was also found that AP-39 significantly inhibits the oxidative stress-induced toxicity by silencing the mitochondrial cell death pathways and consequently contributing to the maintenance of cell homeostasis. Furthermore, in vivo studies showed that AP-39 is useful in acute cardiac arrest, renal, and myocardial I/R injury, through the inhibition of mitochondrial permeability [190,191,192,193]. Following the development of AP-39, several similar TPP-conjugated H2S releasing compounds were developed by the same research group, namely AP-123 and RT01 [194]. It is important to mention that despite the numerous beneficial effects that AP-39 was able to demonstrate, in certain pathologies such as cardiac ischemia/reperfusion injuries where the therapeutic window for corresponding pharmaceutical intervention is very narrow, the very slow rate of H2S release from AP-39 might not be the main protective reaction mechanism. In addition, the ten-carbon long aliphatic linker incorporated in the structure of AP-39 makes this molecule hydrophobic enough to hypothesize that its incorporation and long-lasting presence in the lipophilic membranes results in the very limited effect on specific molecular targets.
DTTs are also known to take part in other biological reactions, for example, electrophilic reactions [195] and the formation of reactive radicals [196].
3.1.3. Thiol-Activated H2S Donors
N-Mercapto-Based Donors
In 2011, on the basis of the structure of N-mercapto (N-SH) compounds, the first thiol-activated H2S donors were reported [197]. As is well known, the N-SH bond is unstable and can be simply broken, providing a hydrosulfide anion. This feature led to the design of controllable H2S donors based on the N-SH scaffold. The synthesis of these compounds was conducted using acyl groups to protect the thiol residue of N-SH. The obtained N-(benzoylthio)-benzamides (29) is stable in aqueous buffers and is able to generate H2S in the presence of thiols (cysteine or glutathione) (Figure 15).
Figure 15.
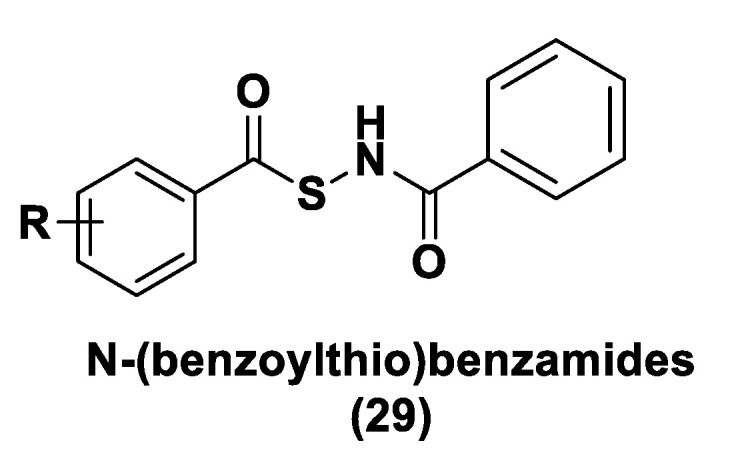
General chemical structure of N-(benzoylthio)benzamides.
Structural activity relationship studies carried out in PBS buffers of a series of such donors revealed that structural modifications can modify the release of H2S from these compounds. In particular, the presence of electron withdrawing groups promoted faster H2S generation, but electron-donating groups slowed down the release of H2S. Thiol-activated H2S donors have been also assessed in plasma and similar results in H2S release have been obtained.
The mechanism of H2S release from this N-mercapto-(N-SH) has been reported (Figure 16). Firstly, a thioester (30) exchange between the donor and cysteine occurs. The resultant S-benzoyl cysteine (31) forms a more stable N-benzoyl cysteine (32) due to a fast S to N acyl transfer. The N-SH intermediate (33) instead reacts with another molecule of cysteine to produce benzamide and cysteine perthiol (34); the latter reacts with cysteine to give cysteine disulfide (cystine, 35) and releasing H2S.
Figure 16.

H2S release from N-mercapto-(N-SH) compounds.
S-Aroylthiooximes
N-SH-based H2S donors bearing an S-aroylthiooxime (SATO) moiety represent another class of H2S donors (36, Figure 17) [198]; this class of compounds has been obtained through a reaction between substituted S-aroylthiohydroxylamines (SATHAs) and ketones or aldehydes. Under a physiological pH, these compounds have proven to be moderately stable in aqueous solutions and their hydrolysis rate could be monitored by modulating the electronic and steric factors. H2S half-life, obtained from thiols, is between 8 and 82 min and can be modified by substituent variations on the SATHA ring. The H2S-releasing mechanism for these new molecules is comparable to that of N-(benzoylthio) benzamides; in fact, it starts with a reversible thiol exchange between the SATOs (36) and cysteine to form the arylidenethiooxime (37) and S-benzoyl cysteine. The subsequent reaction of another molecule of cysteine with the arylidenethiooxime (37) produces cysteine perthiol (34), aldehyde, and ammonia. Lastly, cysteine perthiol (34) reacts with cysteine to give H2S and cystine (35).
Figure 17.

H2S release from SATOs.
Perthiol-Based Donors
Since cysteine perthiol represents an important compound for H2S generation from N-mercapto-based donors, perthiols have been considered for the design of novel H2S donors. Indeed, cysteine perthiol is implicated in H2S biosynthesis that is catalyzed by CSE and it can be considered a “natural” H2S donor [12,41].
Considering this feature, a class of perthiol-based donors have been synthesized as cysteine (38) and penicillamine (39) derivatives (Figure 18) [199]. To protect the thiol residue, acyl groups were used. The evaluation of the H2S-releasing profile of these donors was carried out in the presence of thiols and in aqueous buffers and showed that cysteine-based donors release very small amounts of hydrogen sulfide (probably caused by an undesirable disulfide cleavage of the donors by thiols that leads to no H2S production), while penicillamine-based donors have a high efficiency in H2S generation. Probably the presence of two adjacent methyl groups precluded the cleavage of the disulfide bonds by thiols and then the diacylation and following H2S production dominated in the reaction leading to a better H2S production.
Figure 18.

General chemical structures of perthiol-based H2S donors.
Studies concerning the mechanism of H2S release from these donors showed similarities to that of N-mercapto-based donors.
H2S release from penicillamine-derived perthiol donors in H9c2 cells and in in vivo studies has been also evaluated. The cardioprotective effects of a number of compounds have been also observed in a myocardial ischemia/reperfusion (MI/R) injury murine model. The results revealed that these donors had therapeutic benefits reducing the infarct size in MI/R mice.
Dithioperoxy-Anhydrides
Dithioperoxy-anhydrides (40a, 40b) represent another kind of thiol-activated H2S donor [200]. Considering the disulfide linkage, these donors have some chemical similarities with perthiol-based donors. Acylpersulfides are very important intermediates for H2S release and two probable pathways have been considered: (a) a direct reaction between acylpersulfides and thiols to form H2S and RSSAc; (b) the production of a new perthiol leading to H2S and disulfide generation (Figure 19).
Figure 19.

Chemical structures and the H2S release mechanism of dithioperoxy-anhydrides.
H2S release has been proven in both buffers and HeLa cell lysates. Moreover, the CH3C(O)SSC(O)CH3 compound is effective in the concentration-dependent vaso-relaxation of a pre-contracted rat aortic ring through its H2S release.
Thioamide- and Aryl Isothiocyanate-Based Donors
Arylthioamide derivatives (41) represent a class of thiol-activated H2S donors, even if these compounds are weak and slow-releasing donors (Figure 20) [146].
Figure 20.

Chemical structures of thioamide- and aryl isothiocyanate-based donors.
To evaluate the vasoconstriction effects, arylthioamides have been tested; it was observed that the pretreatment of aortic rings with compound 41a (p-hydroxybenzothioamide), considerably inhibited the vasoconstriction generated by noradrenaline. Oral administration of p-hydroxybenzothioamide in rats led to a decrease in blood pressure, supporting the protective actions of this donors in cardiovascular systems. Endogenous thiols were sufficient to promote H2S release from the donors because neither exogenous cysteine nor glutathione were added.
Aryl isothiocyanate derivatives represent another class of slow-releasing H2S donors (Figure 20) [136]. Thiol activation for these derivatives proved to be required. PhNCS (42) and PhNCS-COOH (43) exhibited vaso-relaxing effects on conductance and coronary arteries but the biological mechanism is unclear. It has been proposed that aryl isothiocyanates are able to react with biomolecules (for example cysteine) to produce thioamide compounds and then H2S.
gem-Dithiol-Based H2S Donors
gem-Dithiols represent unstable compounds in aqueous solutions and H2S is the decomposition product [201,202]. They belong to thiol-activated H2S donors (44, Figure 21) [145,203]. These compounds have been modified by acylation and a library of donors have been obtained. H2S release from these donors has been determined in the presence of Cys or GSH. Donor-induced H2S release has been similarly examined in HeLa cells (in the presence of endogenous thiols).
Figure 21.

H2S release from acylated gem-dithiols.
3.1.4. Enzyme-Triggered Donors
In 2016 Zheng et al. developed a series of esterase-sensitive prodrugs, based on a lactonization reaction, otherwise known as a ‘‘trimethyl lock” (TML) [204,205]. HP-101 (45) was the first obtained derivative (Figure 22) and it was synthesized under microwave conditions starting from compound 45a by a reaction with Lawesson’s reagent [206], to give an intermediate 45b that was subsequently treated with sodium hydroxide. The desired compounds led to the release of H2S due to the cleavage of a phenolic ester by esterase and the consequent lactonization caused by the steric repulsion of three methyl groups (Figure 22). Remarkably, HP-102 (46, Figure 22) a TML derivative, was shown to reduce myocardial I/R injury and presented a cardioprotective action [207]. HP-102 is at present under investigation to better identify its chemical, pharmacological, and safety profiles prior to its further progression in clinical trials.
Figure 22.

TML chemical structures and the synthetic route for HP-101 preparation.
Carbonyl sulfide (COS)-releasing compounds are a recent class of enzyme-triggered H2S donors; COS represent a substrate of CA (carbonic anhydrase) enzyme that catalyzes the reaction that leads to H2S production [208,209]. N-thiocarboxyanhydrides (NTAs) (Figure 23) constitute dual COS/H2S donors [210,211], that advantageously release COS and innoxious peptide byproducts at the same time [212]. NTA1 (47, Figure 22), a sarcosine NTA derivative was obtained starting from sarcosine (47a, N-methyl glycine) as reported in Figure 23 [213].
Figure 23.

General structure of N-thiocarboxyanhydrides (NTAs) and the synthetic procedure for the synthesis of NTA1.
COS/H2S release from NTA1 is based on a cycle opening upon a nucleophilic attack promoted by endogenous amines: the dipeptide formed has been detected by LC-MS and GC-MS. The subsequent release of H2S from COS is due to the rapid enzymatic action of CA (Figure 24) [212]. When endothelial cells have been treated with NTA1 improved proliferation, an important step in the angiogenic process occurs [212].
Figure 24.

COS/H2S release from NTA1.
ROS-triggered H2S donors (peroxyTCM-1, 2, 3; 48, 49, 50) are another class of COS/H2S donors; the compounds were obtained [214] by applying a ROS-cleavable aryl boronate (X) as a protecting group (Figure 25) [215,216].
Figure 25.

ROS-activated H2S donor preparation.
The proposed mechanism for the release of COS/H2S is summarized in Figure 26: these compounds carry out H2S in the presence of an oxidant, for example H2O2: the boronate ester is transformed into a phenol by ROS, giving COS, which is then rapidly moved into H2S by CA [214].
Figure 26.

H2O2-triggered COS/H2S release.
Boronate esters are known as ROS scavengers and this effect was coupled with the H2S cytoprotective action leading to a novel tool for the treatment of human pathologies determined by elevated oxidative stress, for instance cardiovascular diseases [193]. Additional in vivo studies are ongoing for the assessment of their efficacy.
3.1.5. Other H2S Donors in Cardiovascular Diseases
Thioglycine (51) and thiovaline (52), two thioamino acids, are bicarbonate-triggered H2S donors [217]; these compounds have been synthesized starting from commercially accessible Boc-protected amino acids (51a, 52a) that have been treated with 1,1′-carbonyldiimidazole in DCM and then with gaseous H2S. The subsequent deprotection of intermediates 51b and 52b with TFA in dichloromethane provided the required compounds (Figure 27).
Figure 27.

Synthesis of thioglycine and thiovaline.
Following a cyclization reaction, these compounds are converted into the corresponding amino acid N-carboxyanhydrides in the presence of a bicarbonate and produce H2S (Figure 28). Thioamino acids are considered as in vivo H2S donors due to the releasing mechanism and the high-level of bicarbonate in the blood.
Figure 28.

H2S release from thioamino acids.
Thioglycine and thiovaline H2S release have been assessed through amperometric studies. The compounds also improve the formation of cGMP in a concentration-dependent manner and produce a substantial relaxation of pre-contracted mouse aortic rings. Conversely, thioglycine and thiovaline are characterized by a high reactivity; in fact, they can provide amidation [218,219,220,221,222] and oxidation reactions [190], giving products which could produce undesirable side-effects.
ZYZ803 (2-(N-Boc-amino)-3-prop-2-ynylsulfanylpropionic acid, 53) is a novel H2S-NO releasing molecule, recently developed through a copulation reaction between SPRC and furoxan in the presence of Boc anhydride (Figure 29) [223,224].
Figure 29.

Synthesis of ZYZ803.
SPRC (53a) releases H2S through CSE enzyme activation, whereas furoxan (53b) represents a thiol-triggered NO-releasing moiety [225]. Consequently, ZYZ803 (53) is able to stimulate either the expression of CSE through NO synthase (e-NOS) endothelial activity, releasing H2S and NO, correspondingly (Figure 30), promoting angiogenesis by the SIRT1/VEGF/cGMP pathway and through crosstalk between STAT3 and CaMKII [226].
Figure 30.

H2S/NO releasing pathways from ZYZ803.
In vitro and in vivo studies have shown that ZYZ803 significantly promotes endothelial cell angiogenesis [227]. In an animal model of ischemia, ZYZ803 stimulated angiogenesis in a more efficient way in comparison to the monotherapy administration of SPRC or furoxan, proving the synergistic actions of H2S and NO released by ZYZ803 [227]. This interesting lead compound regulated vascular tone in rat aortic rings [223], reduced cardiac dysfunction, and increased myocardial injury following heart failure [228], giving a potent cardioprotective effect.
Finally, Zofenopril (54), an ACE inhibitor, is among the H2S donors with cardiovascular system activity. This compound in the liver undergoes a hydrolysis reaction with the formation of Zofenoprilat (55), and its active metabolite contains a thiol group (Figure 31).
Figure 31.

Chemical structures of Zofenopril and Zofenoprilat.
Zofenopril is an already well studied drug, and it was approved for medical use in 2000. This compound has been proven to release H2S, and this gaseous transmitter in turn mediates the beneficial anti-apoptotic, pro-angiogenic, and anti-inflammatory effects of zofenopril [229,230,231,232,233,234]. The drug improved vascular function in a murine model of hypertension, due to its ACE inhibitor action and H2S-releasing effects, and the administration of this compound increased the plasmatic levels of H2S in mice, protecting them from myocardial I/R injury and increasing the endocardial blood flow in myocardial ischemia [235].
3.2. H2S Donors and Cancer
H2S has several roles in the regulation of cellular activities, for instance, migration, proliferation, apoptosis, and autophagy. A low production of H2S in several cancer types has been found. The principal experimental technique applied to enhance H2S levels in cancer cells is represented by the use of H2S donors; nevertheless, their use is connecting to the concentration/dose, cell type, time, and administration method.
Natural and synthesized donors have been used in this context and different effects have been observed, such as suppressant effects but also promoter effects. Following treatment with H2S donors, numerous signaling pathways have been altered by cells, prompting their use in cancer treatment. The good efficiency of H2S donors has silenced different cancer types due the regulation of cellular activities and signaling pathways. The evident opposite biological effects in different experiments, for example in the case of NaHS, needs to be further analyzed. The different effects of this salt seem to be correlated to the short-time and the high concentration of H2S released from NaHS, giving firstly a momentary increase in the cellular H2S levels, followed by a subsequent decline; this kind of release is very different to the normal slow-releasing cellular mode, and therefore NaHS has been replaced in clinical applications.
Additionally, the amount of information about H2S donors (such as on metabolism and clearance) is limited and their applicability has generated many questions.
For this purpose, it is important to assess the range of concentrations for H2S donors and to define all the potential toxicity and clearance mechanisms. Furthermore, potential interactions between different molecules, as in the case of molecular hybrids, requires additional analysis for a better realization of the side effects.
It has been observed that the combination of H2S donors and cancer drugs (for example cisplatin and docetaxel) increases drug sensitivity and decreases resistance. The synergistic effect of H2S donors with metformin/simvastatin has promoted anti-cancer activities due to the elevation of intracellular acidity and the reduction in pH regulators, paving the way for their useful administration in the treatment of advanced stages of cancer; at present, all the researchers agree that more preclinical and clinical trials are needed to study this aspect.
In summary, H2S donors, used both in monotherapy or in combination with other drugs, have interesting applications in cancer treatment; several studies are now in progress and the hope is to shed additional light on cancer therapy [236].
Triple-negative breast cancer (TNBC) is an aggressive subtype cancer form and represents about 15–20% of breast cancers. Unfortunately, it is characterized by metastasis and recurrence and the development of novel treatments of TNBC is very important. Recently, extraordinary progress in the treatment of TNBC by H2S donors has been made.
In summary, on the one hand, H2S donors efficiently modulate the proliferation and apoptosis of TNBC cells through the inhibition of the phosphorylation or expression of proteins connected to the NF-κB, PI3K/Akt/mTOR, and Ras/Raf/MEK/ERK signaling pathways. Otherwise, H2S donors inhibit the expression of MMP-2/9 and EMT to resist TNBC metastasis through the inhibition of the abnormal activation of the β-catenin pathway.
H2S is believed to be involved in other pathways that account for TNBC prevention and this deserves further investigation. Some organic H2S donors (DATS, DADS, GYY4137, ADT-OH, thiobenzamide) are used for endogenous H2S production to exert antitumor effects. ATB-346 (naproxen derivative, 56, Figure 32) and GIC-1001 (57, Figure 32) represent some H2S donor-based therapeutics entered in Phase II clinical trials. HS-ASA (58, Figure 32) showed antitumor activity on TNBC MDA-MB-231 cells in vitro and in vivo studies. HA-ADT (59, Figure 32) was able to induce MDA-MB-231 cell apoptosis by improving the ratio of Bad/Bcl-xl and Bax/Bcl-2 and the expression of cleaved caspase-3/9 and cleaved PARP [237].
Figure 32.

Chemical structures of H2S donors evaluated for TNBC treatment.
Hybrids capable of releasing H2S or NO, or both gaseous transmitters have been developed. The hybrids can be beneficial in reducing the doses of the chemotherapeutic agent in order to diminish the toxic effects of these drugs and the gaseous transmitter gains additional chemotherapeutic effects.
Various H2S and NO-releasing compounds have been developed for their use in cancer chemotherapy and SNOs (S-nitrosothiols) donors are to date considered the most efficient compounds. GSNO (S-nitrosoglutathione, 60) and S-nitroso-N-acetylpenicillamine (SNAP, 61) (Figure 33) are two SNOs that have been investigated for their anti-carcinogenic properties.
Figure 33.

Chemical structures of GSNO and SNAP.
The reaction between SNO donors and Cysteine leads to the formation of CysNO at the external face of the plasma membrane. This compound has a membrane that is impermeable but passes into the cell using the L-type amino acid transporter on the plasma membrane. The reaction between SNO donors and H2S yields HSNO which diffuses spontaneously across cell membranes. The mixed effects of CysNO and HSNO enhance the cellular nitrosation most likely via transnitrosation processes or by enhancing the nitrosating capabilities of both compounds. Prolonged exposure of tumor cells to these compounds causes stress conditions and consequently leads to apoptosis [238].
Cancer cells are distinguished by an improved uptake of sugars, for example glucose. Fortunato et al. synthesized novel glycoconjugated-H2S donors in order to selectively enhance the concentration of H2S in cancer cells. Dithiolethione or isothiocyanate moieties have been considered for their H2S-releasing properties. C1-glycoconjugates have been obtained by applying a synthetic stereoselective procedure based on trichloroacetimidate glycosyl donors, while C6-glycoconjugates have been achieved through a Mitsunobu reaction (Figure 33). Subsequently the obtained compounds have been evaluated for their anticancer effects in the AsPC-1 cell line. Compounds 62 and 63 (Figure 34) have been demonstrated to release H2S inside the AsPC-1 cells and to modify the basal cell cycle [239].
Figure 34.

Glycoconjugated H2S donors.
Brassicaceae are a well-known plant family and have attracted attention for the presence of the glucosinolate myrosinase system that leads to isothiocyanates upon plant biotic and abiotic lesions.
Erucin (ERU; Figure 4), an isothiocyanate present in arugula, originates through glucosinolate glucoerucin hydrolysis due to the myrosinase enzyme.
Natural isothiocyanates modify cancer cells stimulating anti-proliferative effects at high concentrations. Gaseous hydrogen sulfide or non-natural H2S-releasing compounds showed comparable anti-proliferative effects.
Isothiocyanates release H2S in biological conditions: ERU belongs to this category and was evaluated for its anticancer effects on pancreatic adenocarcinoma cells (AsPC-1). The obtained results showed the chemo preventive and anticancer features of this compound. In addition, ERU presented significant anti-proliferative effects by the inhibition of AsPC-1 cell viability and cell migration, modifying the AsPC-1 cell cycle, and exhibiting a pro-apoptotic effect.
Moreover, AsPC-1 cells present a genetic mutation in KRAS that leads to KRAS hyper-activation and then to MAP-kinase hyper-phosphorylation. ERU repressed ERK1/2 phosphorylation. This inhibition is very important in pancreatic cancer proliferation, growth, and survival [240].
3.3. H2S Donors and Nervous System Diseases
H2S donors have been investigated for their use in the treatment of nervous system pathologies. The pioneer compound for this use was ACS 84 (Figure 12), a DOPA-DTT that has been shown to have an important role in Parkinson’s Disease; successively, several other compounds have been reported for their in vitro effects and for their potential use in Alzheimer’s disease (AD).
An important relationship between H2S and aging has been observed and a significant decline in H2S levels has been seen in patients affected by Alzheimer’s disease (AD). On this basis, the use of H2S-donors could represent an exciting and intriguing strategy to be pursued in the treatment of neurodegenerative diseases.
Sestito et al. designed a small series of multitarget molecules combining the rivastigmine-scaffold, a well-established drug already approved for AD, with sulforaphane (SFN) and erucin (ERN), two natural products derived from the enzymatic hydrolysis of glucosinolates contained in broccoli and rocket, respectively, endowed with antioxidant and neuroprotective effects.
The newly obtained hybrids (64–69, Figure 35) elicited protective effects in an LPS-induced microglia inflammation model. A decrease in NO production was observed in LPS-stimulated cells pre-treated with the compounds and they also showed neuroprotective and antioxidant activities in human neuronal cells. Compound 64 proved to be the most promising agent, combining antioxidant and anti-inflammatory properties. This promising pharmacological profile has been imputed to the slow H2S releasing profile which combined produced a very favorable logP value [241].
Figure 35.

Chemical structures of ERN- and SFN-rivastigmine hybrids.
In another study, Sestito et al. substituted the free amine group of memantine (70) with an isothiocyanate group to achieve H2S release. The new chemical entity, named “Memit” (70b, Figure 36), has been tested in vitro to establish the pharmacological profile of the “native drug”. Indeed, it has been observed that memit releases H2S through a cysteine-mediated mechanism, producing memantine (Figure 36). The new compound exerted protective effects in neuronal inflammation and induced a drastic decrease in ROS production. Memit has also been proven to decrease the aggregation of Aβ(1–42) and had a cytoprotective effect versus Aβ oligomers which induced damage in human neurons and rat microglia cells. Furthermore, the new molecule promoted autophagy that is altered in neurodegenerative diseases.
Figure 36.

Preparation of Memit and its mechanism of H2S release.
Memit can be considered a prodrug of memantine, although in vivo studies are important to study the substantial effects of memantine and H2S [242].
3.4. H2S Donors and Gastrointestinal Diseases
S-aspirin (ACS-14), S-diclofenac (ACS-15), ATB-429 (Figure 13) and ATB-346 (Figure 32) have been already discussed above. In the same way, ATB-343 (indomethacin derivative, 71) and ATB-340 (72) (Figure 37) decreases the gravity of NSAID-induced damage in the rat stomach. ACS-15 (CTGPharma) and ATB-337 (Antibe Therapeutics) (Figure 13), a diclofenac derivative, exhibits increased anti-inflammatory effects in comparison to the parent drug. Indeed, whereas diclofenac damages the stomach in dose-dependent manner, ACS-15 shows substantially decreased GI toxicity and does not affect hematocrit. This compound inhibits COX-1 and COX-2 activity as well as diclofenac. Moreover, differently from diclofenac, it does not cause leukocyte adherence, and is more effective in decreasing paw edema [12].
Figure 37.

Chemical structures of ATB-343 and ATB-340.
Looking in more detail at the different drug applications, ATB-429 is a mesalamine derivative and is therefore aimed at the improvement of the treatment of inflammatory bowel disease (IBD). IBD can be separated into two subtypes: Crohn’s disease (CD) and ulcerative colitis (UC). ATB-429 has been widely characterized in CD and UC animal models and is significantly superior to mesalamine. It was more efficient in treating the visceral pain associated to IBD [12]. ATB-346 represents a naproxen H2S donor-derivative designed for osteoarthritis pain relief. Naproxen is generally associated with a considerable risk of severe gastrointestinal bleeding and cardiovascular effects. ATB-346 showed superiority and a remarkable reduction in the gastrointestinal and cardiovascular toxicities in comparison to its parent drug in preclinical studies [12].
4-carbamothioyl phenyl 2-acetoxybenzoate (ATB-340 (72), Figure 37) is an H2S releasing aspirin. The H2S pathway is involved in the susceptibility of the gastric mucosa to High Fructose Diet (HFD) injury. The administration of ATB-340 in elderly patients showed an increased efficiency in the decrease in age-associated gastric mucosal defense. Decreased activation of H2S signaling in the HFD model is associated with the increase in mitochondrial impairment and oxidative stress that stimulates the expression of thiosulfate-dithiol sulfurtransferase. The gastro-protection observed upon ATB-340 treatment is H2S-dependent and is related to the attenuation of oxidative stress [243].
3.5. H2S Donors and Dermatological Diseases
In the skin, H2S is generated by enzymic pathways and has a physiological role in various functions, as well as inflammation, vasodilatation, cell proliferation, and apoptosis. Modifications of H2S production are involved in several dermatological diseases, for example melanoma, psoriasis, and other dermatoses.
The data available to date in relation to H2S signaling in the skin support the involvement of hydrogen sulfide in patho-physiological processes. Generally, H2S effects are highly concentration or dose-dependent, implying the interaction with targets that result in stimulatory or inhibitory effects that interfere with skin processes.
Due to the important role of H2S in skin physiology, altered H2S production contributes to the pathogenesis of various cutaneous diseases.
Several H2S-releasing compounds, for example Na2S, NaHS, DATS (Figure 4), GYY-4137 (Figure 6), JK1 (Figure 8), ATB-346 (Figure 32), and naproxen-HBTA (naproxen-4-hydroxybenzodithioate, 73) (Figure 38), have been examined in experimental studies.
Figure 38.

Chemical structure of naproxen-HBTA.
These compounds are potential therapeutic molecules useful for the treatment of several skin diseases due to their different etiological aspects. The development of novel and safer H2S-based drugs will fundamentally depend on increasing knowledge of the exact role of H2S in the various aspects of skin biology [100].
In 2019, naproxen-HBTA, a H2S-releasing derivative of naproxen, was analyzed in a metastatic melanoma model in in vitro and in vivo studies. The obtained results demonstrated that naproxen-HBTA stimulated caspase 3-mediated apoptosis and reduced motility and invasiveness. Moreover, daily oral treatment with naproxen-HBTA significantly suppressed melanoma growth and progression in mice. This dual approach could represent a “combination therapy” for melanoma based on COX-2 and H2S pathways as new therapeutic targets [244].
3.6. H2S Donors and Viral Infections
As stated above, H2S exerts several positive effects in pathologies characterized by lung inflammation, such as COVID-19. H2S promotes the elimination of potentially dangerous viruses by breaking mucin disulfide bonds, making the mucus less viscous and facilitating the expulsion of pathogens through the respiratory ciliary apparatus.
Through S-sulfuration, H2S impedes the activation of the NF-κB pathway, and the inhibition of the IKκβ enzyme, inhibiting the translocation of NF-κB into the nucleus, and then the generation of the cytokine storm. Moreover, H2S stimulates the activation of Nrf2, improving the expression of enzymes and antioxidant molecules. Furthermore, H2S protected the tissues from oxidative stress due to direct antioxidant activity. Finally, H2S promoted bronchodilation by activating the KATP channels expressed on the cell membrane of the bronchial smooth muscle cells and leading to electrolyte absorption by increasing muco-ciliary clearance and blocking the Na+/K+ ATPase pump.
Antiviral effects of organosulfur molecules have been investigated [103] and recent studies have elucidated that these molecules are H2S donors, leading to the belief that antiviral effects are linked to the release of H2S.
DATS (Figure 3) has been tested in cytomegalovirus (CMV)-induced hepatitis. In this study, the polysulfide DATS decreased the viral load of CMV in the infected organs of mice and humans by inhibiting gene transcription and promoting the amplification of CMV-induced T-regulatory helper cells, giving an immunosuppressive effect versus the CMV noted in murine models [103].
Similarly, the antiviral properties of the root extract of the plant Isatis indigotica L. have been investigated. This extract contains sinigrin (a precursor of the allyl isothiocyanate, (74) Figure 39), a compound that has been proven to be the most effective compound in 3CLpro inhibition (3-chymotrypsin-like protease; Mpro—main protease) of SARS-CoV. Notably, this protease causes the proteolytic processing of polypeptides leading to functional proteins required for viral replication [103].
Figure 39.

Chemical structures of sinigrin, sulforaphane, and XM-01.
Data obtained exploring the effects of sulforaphane (75, present in various plants of Brassicaceae family; Figure 39) suggest that the antiviral effect of Brassicaceae extracts can be largely ascribed to their isothiocyanate compounds. Sulforaphane was proven to have antiviral activity through the influenza A/WSN/33 (H1N1) virus by decreasing virus replication in vitro. These results have been confirmed by a randomized, double-blind, placebo-controlled clinical study. Sulforaphane improved virus-induced granzyme B (a protease released by NK or T cells that stimulates cell apoptosis) through the NK cells. The increase in granzyme B levels was adversely related to influenza RNA levels in nasal lavage fluid cells, proving a sulforaphane antiviral role [103].
GYY4137 (Figure 3) has been used by the Casola group for studying the antiviral role of H2S in RSV (respiratory syncytial virus). RSV infection is characterized by low levels of CSE mRNA and protein expression in airway epithelial cells, consequently leading to a reduction in the levels of H2S. Scientists have reported increased viral replication and airway inflammation in RSV-infected CSE knockout mice in comparison to wild-type mice. Bearing in mind these findings, GYY4137 has been used in order to set the role of H2S in paramyxovirus infections and a considerable reduction in the production and viral replication of pro-inflammatory mediators was found. The antiviral effect of this compound was shown to be related to the inhibition of syncytia formation, viral assembly, and release. Moreover, GYY4137 revealed anti-inflammatory effects by the regulation of NF-κB and IRF-3 activation. This effect was confirmed in vivo during RSV infections through the intranasal administration of GYY4137 to infected mice: the compound inhibited viral replication when administered within 24 h of infection, decreased pro-inflammatory mediators, and reduced airway dysfunction. These effects have also been observed in vitro even on different RNA virus infections, for example influenza A and B and Ebola virus.
In in vitro and in vivo models, gem-dithiol-based H2S donors reduced viral replication and deceased proinflammatory mediators, confirming once again that H2S was responsible for the observed antiviral effects [103].
More recently, XM-01 (76; Figure 39) a disulfide compound, has been investigated for its antiviral effect on different viral strains (RSV, influenza virus (A/WSN/33 strain), rotavirus). The effects of XM-01 (a cysteine-based perthiol derivative) correlated with its H2S release, were reported in a study based on the identification of cardioprotective H2S donors. Moreover, XM-01 demonstrated antiviral effects on enveloped viruses but did not show activity through non-enveloped viruses.
Thus, as the antiviral action may be due to alterations of the viral membrane, GYY4137 and its analogues could be useful against enveloped viruses inhibiting the entry phase of the host cell.
In a very recent paper, H2S antiviral activity versus SARS-CoV-2 has been reported describing its interference with ACE and TMPRSS2 [103].
3.7. H2S Donors and Other Diseases
H2S is implicated in other diseases, such as diabetes, pain, and bone tissue alterations.
Type 2 diabetes mellitus (DM) represents an important chronic disease correlated to severe vascular and microvascular complications (including neuropathy, cerebrovascular and peripheral arterial diseases, and retinopathy), that often leads to the progression of atherosclerosis.
Dietary therapy represents the first step in the treatment of diabetic patients. Medicinal plants are growing in importance among the current therapeutic options due to their effective and economical therapeutic support and reduced side effects.
Organosulfur compounds present anti-inflammatory, antioxidant, hypoglycemic, and immunomodulatory effects; for these effects they have usually been considered as cardioprotective agents in type 2 DM and therefore, organosulfur compounds derived from garlic (Figure 4), signify an appreciated support in the type 2 DM diet.
Several studies, ranging from preclinical to clinical studies, have demonstrated the relevant effects of hydrogen sulfide in human DM; however, further evidence is necessary to establish the validity of the administration of organosulfur compounds in the treatment of DM. Globally, stability is one of the most relevant problems with natural compounds; in fact, allicin and its derivatives (Figure 4), quickly degrade. Consequently, to improve stability and bioavailability, several researchers are developing new strategies for the administration of these natural compounds (nano-emulsions, capsules containing garlic oil). The preparation of combined formulations containing other nutraceuticals, with synergistic effects, is a very focused challenge for many companies. The variability of the chemical composition of vegetables, dependent on environmental conditions and countries of production, represents another relevant problem. Therefore, vegetable trials have to contain information on the chemical composition and standardized preparations. The study of garlic organosulfur compounds represents a stimulating field where inflammation, redox biology, tissue repair, detoxification, and regeneration are connected for favorable effects on human health [245].
Trimebutine maleate, a spasmolytic FDA and EMA-approved drug, has been recently evaluated for the treatment of patients undergoing sedation-free colonoscopy. A new trimebutine H2S-releasing salt has been recently disclosed and has been proven to release in vivo the gasotransmitter that allows a reduction in nociception. The new compound (GIC-1001 (57), Figure 32) has been obtained combining trimebutine and 3-thiocarbamoylbenzoate (3TCB). It has been tested in a mouse model of colorectal distension: the oral administration of GIC-1001 led to a significant reduction in the nociceptive responses to all noxious stimuli. This effect was significantly superior in comparison to the parent compound at equimolar doses. The preliminary results of GIC-1001 suggest that this compound may represent a good compound to reduce the visceral pain and discomfort caused by lumenal distension [246].
BPs (Bisphosphonates, 77, Figure 40) represent the principal treatment for bone loss due to various pathological conditions. Considering their high bone affinity, BPs have been utilized to develop conjugates with anti-catabolic and pro-anabolic drugs. H2S stimulates osteogenesis and reduces osteoclast differentiation; following this rationale, an innovative compound, DM-22 (78, Figure 40), has been prepared combining alendronate (AL) and an arylisothiocyanate. DM-22 and AL have been tested in vitro to evaluate the effects on h-OCs (human osteoclasts) and h-MSCs (mesenchymal stromal cells) during osteogenic differentiation. Amperometric analysis showed that DM-22 slowly produces H2S in a thiol-dependent manner. DM-22 significantly reduced h-OCs differentiation and function, preserving a residual viability of h-OCs. In h-MSCs, DM-22 did not produce cytotoxicity as showed by a LDH assay, considerably stimulating bone mineralization, and enhancing mRNA expression of Collagen I.
Figure 40.

General chemical structure of BPs and chemical structure of DM-22.
DM-22 is therefore a new bisphosphonate without cytotoxicity and represents a candidate for a different kind of osteoanabolic drug [247].
3.8. Other H2S Donors
In this section further examples of H2S donors recently reported in the literature are discussed.
Research in the field of H2S is at present compromised by the lack of pharmacological tools, for example, selective enzymatic inhibitors exclusively target only one of the H2S pathway components. In 2016, Corvino et al. applied a new approach based on a hybrid method to design a novel selective CSE inhibitor. A library of compounds was developed; oxothiazolidine derivatives (79, Figure 41) demonstrated maximal inhibitory effects in tissues and were shown to be superior to PAG (propargylglycine). Moreover, metabolomics assays on the purified enzymes demonstrated their full selectivity as CSE inhibitors. In comparison to PAG, compound 79 behaved in a different way: both compounds inhibited CSE through the production of pyruvate, ammonia, and H2S, but unexpectedly, in the presence of PAG, CSE used the substrate L-Cys and produced cystine and lanthionine. In contrast, 79 completely inhibited the enzyme, without an effect on the concentration of the substrate. This resulted in the chance of utilizing lower doses of 79 in comparison to PAG, thus decreasing all the adverse effects. Compound 79, but not PAG, is not a suicidal inhibitor and this represents an additional advantage. In order to further progress into clinical studies, a careful pharmacological investigation will be required in order to evaluate whether the new compound is associated with unwanted side effects. This highly selective CSE inhibitor could be useful to better understand the role of CSE vs. CBS in the pathophysiology of the diseases in which a role for the H2S pathway has been suggested. The development of selective CSE inhibitors, particularly characterized by good water solubility, will further shed new light on the crosstalk between H2S and other relevant pathways of transmitters (e.g., NO, CO) [248].
Figure 41.

More recently, Severino et al. reported the preparation of novel H2S donors bearing an 1,2,4-thiadiazolidine-3,5-diones scaffold as new donors for a controlled release of H2S. Amperometric approaches have been used to evaluate the H2S-releasing properties and the vasorelaxant ability on aorta rings of the new compounds. Moreover, a study on the release mechanism has been conducted on THIA 3 (80, Figure 41), the most efficient H2S donor; the modification of structural substituents determined a reduction in the release of H2S. The influence of THIAs on the vasoconstrictor effects of PE (L-phenylephrine) was tested on isolated mouse aortic rings, proving that the presence of THIA 3 completely reverted the PE effect in evoking vasoconstriction. By using HPLC, MS, and NMR techniques, the molecular mechanism responsible for H2S release has been studied. 1,2,4-thiadiazolidine-3,5-dione has been confirmed as a H2S-releasing moiety and its H2S releasing properties have been modulated by substitutions on the aromatic rings [249].
In 2016 Marino et al. reported 4-carboxy-phenyl isothiocyanate as a new H2S donor.
As reported above, H2S regulates inflammation and allergic reactions, in which mast cells represent major effector cells. IgE receptor (FcεRI) cross linking gives an enhancement to intracellular calcium ([Ca2+]i), a crucial step in mast cell degranulation. The role of H2S in a [Ca2+]i-dependent mast cell activation has been evaluated and the effects of H2S, principally released from PhNCS-COOH (Figure 21), on the degranulation of native murine mast cells has been analyzed, as well as its effect on RBL-2H3 mast cell lines. The release of β-hexosaminidase and renin, two specific mast cell degranulation markers, has been described, together with changes in [Ca2+]i and the phosphorylation of proteins downstream of FcεRI activation. H2S released by PhNCS-COOH decreased degranulation and renin release in all mast cell types. Moreover, PhNCS-COOH also inhibited the increase in [Ca2+]i caused by Ca2+ ionophore, FcεRI activation, and thapsigargin. Additionally, PhNCS-COOH diminished the phosphorylation of Syk, cPLA-2, and PLCγ1 in antigen-stimulated RBL-2H3 cells. Taken together, the results demonstrated that H2S reduces [Ca2+]i availability through the reduction in the phosphorylation of proteins downstream of FcεRI cross-linking on the mast cells, and then mast cell degranulation and renin release. The data indicate that PhNCS-COOH is a therapeutic instrument in mast cell-mediated allergic disorders [250].
In 2017, a new set of iminothioethers (ITEs) with H2S-releasing properties and characterized by vasorelaxing effects on rat aortic rings was reported. Compounds 81 and 82 (Figure 42), were chosen as representative of slow and fast rate H2S donors, correspondingly, a full recovery of the basal coronary flow, giving back the AngII-induced effects in isolated rat hearts was determined. Studies on HASMCs (human aortic smooth muscle cells) showed membrane hyperpolarizing effects, linked to the intracellular generation of H2S. These findings highlighted compounds 81 and 82 as new pharmacological H2S donors useful for cardiovascular drug discovery [251].
Figure 42.

Chemical structures of compounds 81 and 82 in [251] and compounds 83, 84, and 85 in [252].
In 2019, two series of thiophosphamide derivatives were studied in vitro and in vivo. Compounds 83 and 85 (Figure 42) were shown to release H2S under measured condition; the H2S-releasing rates in tissues and organs were different in PBS, but the H2S-releasing mechanism was the same. All the thiophosphamide derivatives exhibited low toxicity when tested in five cancer cell lines and in W138 cell lines and these significantly decreased the level of TNF-α and improved the level of IL-10. Moreover, the compounds preserved H9c2 cells from injury and myocardial injury by an antioxidation effect. Compounds 84 and 85 (Figure 42) proved to be the most promising derivatives. In conclusion, these thiophosphamide derivatives showed low toxicity, great activity, and a good pharmacokinetic profile and could be candidates for additional investigation [252].
Recently, thioamides have attracted attention especially when included with H2S–NSAID hybrids and showed low toxicity, but regrettably the mechanism of H2S release from thioamides continues to be unclear.
The synthesis and the evaluation of SulfenylTCMs, (86; Figure 43), a H2S-releasing class of thioamide-derived sulfenyl thiocarbamates, has been described to overcome this issue. SulfenylTCMs release COS (carbonyl sulfide), which is immediately converted into H2S by CA (carbonic anhydrase), due to the activation by cellular thiols. These compounds showed excellent H2S release and worked by mechanistically well-defined pathways. The sulfenyl thiocarbamate group is easily bonded to common NSAIDs, for example naproxen, to generate new hybrids, such as YZ-597 (87, Figure 43), able to release H2S in cellular environments. This new class of H2S donors offers an important starting point for the development of new donors [253].
Figure 43.

General chemical structure of 86 and the chemical structure of 87 in [253].
In 2019, Levinn et al. reported a library of first-generation COS/H2S donors based on self-immolative thiocarbamates. By using specific triggers and activators such as biological thiols and bio-orthogonal reactants, the release of COS was shown. Moreover, a series of H2O2/reactive oxygen species (ROS)-triggered donors were reported and the probable mechanism of activation of these compounds caused by endogenous levels of ROS production was shown. Through orthogonal activation approaches, the scientists proved that o-nitrobenzyl photocage-functionalized donors facilitated access to light-activated donors. Moreover, Levinn et al. reported a cysteine-selective COS donor activated by a Strongin ligation mechanism and a simple esterase-activated donor, with fast COS-releasing kinetics and a capability to inhibit mitochondrial respiration in BEAS-2B cells. Further experiments showed that COS release rates and cytotoxicity were directly linked to these compounds. Finally, more recently, donors capable of generating a colorimetric or fluorescent optical response due to COS release have been reported [254].
In 2020, Zhou et al. reported N-dithia-succinoyl amines as thiol-triggered COS/H2S donors. Indeed, GSH, homocysteine and other thiols prime the release of COS and H2S from these compounds by an unexpected mechanism. Significantly, Dts-1 and Dts-5 (88, 89; Figure 44) represent two analogues that showed intracellular H2S release, and Dts-1 imparted important anti-inflammatory effects in LPS-challenged microglia cells [255].
Figure 44.

Chemical structures of Dts-1 and Dts-5 reported in [255].
The scientific community is oriented towards the design of polymer systems for the release of H2S as therapeutics. Supramolecular tuning of the properties of compounds characterized by chains of sulfur atoms (DATS—Figure 3; S8 (90) Figure 45) has been shown to give superior donors, for example DATS-loaded polymeric nanoparticles.
Figure 45.

Chemical structures of S8 and TC-2153.
SG1002 is a water-insoluble, alpha-sulfur rich microcrystalline material including traces of ionic substances (sodium sulfate, sodium thiosulfate, and sodium polythionates). Promising results in animal models of several diseases (for example heart failure, ischemic damage, atherosclerosis, Duchenne muscular dystrophy) have been found by using this compound. Compound 91 (TC-2153, 8-(Trifluoromethyl)-1,2,3,4,5-benzopentathiepin-6-amine hydrochloride, Figure 45) gave good results in age-related memory decline, Alzheimer’s disease, fragile X syndrome, schizophrenia, and cocaine addiction. Finally, DATS has been tested in colon, gastric, brain, and breast cancer. SG1002 could represent the preferred H2S donor for the evaluation of the gasotransmitter’s effects in animal models, due to its distinctive capability to slowly produce H2S without by-products and in a dose-independent and enzyme-independent sustained mode [256,257].
In a very recent paper, Giordano et al. reported hybrids between H2S-donors and betamethasone 17-valerate or triamcinolone acetonide. Considering the broad use of glucocorticoids for the treatment of inflammation in asthmatic patients and the positive effect of H2S on this disease, novel betamethasone 17-valerate and triamcinolone acetonide hybrids linked to H2S-donor moieties have been designed and synthesized. The obtained compounds were assessed for the potential H2S-releasing profile in a cell-free environment and in BSMCs (bronchial smooth muscle cells). Molecular modelling studies carried out with two hybrids 92 and 93 (Figure 46) have been reported and the results showed that their different H2S-releasing properties may be due to the steric accessibility of the isothiocyanate carbon atom. Moreover, the most interesting hybrids 92 and 93 have been examined for their inhibitory effect on mast cell degranulation and for their capability to produce cell membrane hyperpolarization in BSMCs. Important inhibitory effects on mast cell degranulation have been observed, due to a reduction in the release of β-hexosaminidase which was more effective compared to the corresponding native drugs. Compounds 92 and 93 established an important membrane hyperpolarization of BSMCs and have been shown to be more effective than the reference compound NS1619 [258].
Figure 46.

Chemical structures of compounds 92 and 93 in [258] and compound 94 in [259].
In a similar paper, Corvino et al. reported a library of hybrids obtained by the condensation of corticosteroids with different H2S donor moieties. Through an amperometric study, all the hybrids released H2S and their chemical stability was explored. The prednisone–TBZ hybrid (94, Figure 46) was chosen for additional evaluations. The results of in vitro and in vivo studies evidently showed the anti-inflammatory properties of the released H2S. Due to the protective effect on airway remodeling, compound 94 can be considered an interesting molecule for reducing allergic asthma symptoms and exacerbations [259].
4. Conclusions
Considering the therapeutic potential of H2S, H2S donor chemistry has remarkably progressed. The progress of chemical synthesis has acted as the driving force for H2S research, and has particularly focused on the preparation of H2S donors useful for biological studies on the physiological and pathophysiological effects of this molecule.
Some H2S donors are or have been under pharmacological evaluation in clinical trials. Remarkable examples include GIC-1001 (GIcare Pharma Inc.), a trimebutine maleate salt and a H2S donor used as an alternative to sedatives in colonoscopies; ATB-346, a naproxen-based H2S donor (Antibe Therapeutics), that shows anti-inflammatory effects and that, in 2016, was included in a phase II study concerning the pain associated with osteoarthritis; SG-1002 (SulfaGENIX), that has been assessed as part of an investigation aimed at increasing circulating H2S and NO levels subsequently heart failure; ammonium tetrathiomolybdate (ATTM, (NH4)2MoS4), which has been subjected to clinical trials for the treatment of breast cancer owing to its copper depletion effects; and finally, human clinical studies have supported the chemo preventive effects of sulforaphane on carcinogenesis [260] and antiviral action through the influenza A/WSN/33 (H1N1) virus.
Crucial doubts persist in this field and these issues will necessitate combined efforts from chemists, biologists, and pharmacologists. One question is related to the determination of the H2S therapeutic window in different targets; another is the elucidation of the pathways responsible for donor effects. Clarification of the biological activity and the species involved will be necessary to increase the biological knowledge of donor triggers and by-products, species interchange, signaling mechanisms, and redox chemistry of H2S donors. Moreover, the release of reactive by-products, including unidentified sulfur species, from H2S donors represent a topic that needs further attention.
All these aspects must necessarily be considered in order to broaden this field. Incessant innovation in synthetic donors and an improved knowledge of H2S physiology are paving the way for H2S therapeutics to be used in clinics.
Author Contributions
Conceptualization, E.M., G.C., E.P. and V.S.; resources, B.S., R.S., G.E. (Gianluca Esposito), G.E. (Giovanna Esposito), M.M. and A.C. and A.S.; writing—original draft preparation, E.M., F.F. (Francesco Frecentese); writing—review and editing, F.F. (Francesco Frecentese) and F.F. (Ferdinando Fiorino); supervision, F.F. (Francesco Frecentese) and E.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this paper. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Evans C.L. The toxicity of hydrogen sulphide and other sulphides. J. Exp. Physiol. 1967;52:231–248. doi: 10.1113/expphysiol.1967.sp001909. [DOI] [PubMed] [Google Scholar]
- 2.Li Q., Lancaster J.R., Jr. Chemical foundations of hydrogen sulfide biology. Nitric Oxide. 2013;35:21–34. doi: 10.1016/j.niox.2013.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lindenmann J., Matzi V., Neuboeck N., Ratzenhofer-Komenda B., Maier A., Smolle-Juettner F.M. Severe hydrogen sulphide poisoning treated with 4-dimethylaminophenol and hyperbaric oxygen. Diving Hyperb. Med. 2010;40:213–217. [PubMed] [Google Scholar]
- 4.Vandiver M.S., Snyder S.H. Hydrogen sulfide: A gasotransmitter of clinical relevance. J. Mol. Med. 2012;90:255–263. doi: 10.1007/s00109-012-0873-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wang R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012;92:791–896. doi: 10.1152/physrev.00017.2011. [DOI] [PubMed] [Google Scholar]
- 6.Li L., Moore P.K. Hydrogen sulfide and cell signaling. Annu. Rev. Pharmacol. Toxicol. 2011;51:169–187. doi: 10.1146/annurev-pharmtox-010510-100505. [DOI] [PubMed] [Google Scholar]
- 7.Szabo C. Hydrogen sulphide and its therapeutic potential. Nat. Rev. Drug Discov. 2007;6:917–935. doi: 10.1038/nrd2425. [DOI] [PubMed] [Google Scholar]
- 8.Kolluru G.K., Shen X., Bir S.C., Kevil C.G. Hydrogen sulfide chemical biology: Pathophysiological roles and detection. Nitric Oxide. 2013;35:5–20. doi: 10.1016/j.niox.2013.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Fukuto J.M., Carrington S.J., Tantillo D.J., Harrison J.G., Ignarro L.J., Freeman B.A., Chen A., Wink D.A. Small molecule signaling agents: The integrated chemistry and biochemistry of nitrogen oxides, oxides of carbon, dioxygen, hydrogen sulfide, and their derived species. Chem. Res. Toxicol. 2012;25:769–793. doi: 10.1021/tx2005234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Olson K.R., Donald J.A., Dombkowski R.A., Perry S.F. Evolutionary and comparative aspects of nitric oxide, carbon monoxide and hydrogen sulfide. Respir. Physiol. Neurobiol. 2012;184:117–129. doi: 10.1016/j.resp.2012.04.004. [DOI] [PubMed] [Google Scholar]
- 11.Olson K.R. A practical look at the chemistry and biology of hydrogen sulfide. Antioxid. Redox Signal. 2012;17:32–44. doi: 10.1089/ars.2011.4401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Caliendo G., Cirino G., Santagada V., Wallace J.L. Synthesis and biological effects of hydrogen sulfide (H2S): Development of H2S-releasing drugs as pharmaceuticals. J. Med. Chem. 2010;53:6275–6286. doi: 10.1021/jm901638j. [DOI] [PubMed] [Google Scholar]
- 13.Vitvitsky V., Miljkovic J.L., Bostelaar T., Adhikari B., Yadav P.K., Steiger A.K., Torregrossa R., Pluth M.D., Whiteman M., Banerjee R., et al. Cytochrome c Reduction by H2S Potentiates Sulfide Signaling. ACS Chem. Biol. 2018;13:2300–2307. doi: 10.1021/acschembio.8b00463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Filipovic M.R., Miljkovic J.L., Nauser T., Royzen M., Klos K., Shubina T., Koppenol W.H., Lippard S.J., Ivanovic-Burmazovic I. Chemical characterization of the smallest S-nitrosothiol, HSNO; cellular cross-talk of H2S and S-nitrosothiols. J. Am. Chem. Soc. 2012;134:12016–12027. doi: 10.1021/ja3009693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Carballal S., Trujillo M., Cuevasanta E., Bartesaghi S., Moller M.N., Folkes L.K., Garcia-Bereguian M.A., Gutierrez-Merino C., Wardmann P., Denicola A., et al. Reactivity of hydrogen sulfide with peroxynitrite and other oxidants of biological interest. Free Radic. Biol. Med. 2011;50:196–205. doi: 10.1016/j.freeradbiomed.2010.10.705. [DOI] [PubMed] [Google Scholar]
- 16.Eberhardt M., Dux M., Namer B., Miljkovic J., Cordasic N., Will C., Kichko T.I., de la Roche J., Fischer M., Suárez S.A., et al. H2S and NO cooperatively regulate vascular tone by activating a neuroendocrine HNO–TRPA1–CGRP signalling pathway. Nat. Commun. 2014;5:4381. doi: 10.1038/ncomms5381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Filipovic M.R., Zivanovic J., Alvarez B., Banerjee R. Chemical Biology of H2S Signaling through Persulfidation. Chem. Rev. 2018;118:1253–1337. doi: 10.1021/acs.chemrev.7b00205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Paulsen C.E., Carroll K.S. Cysteine-mediated redox signaling: Chemistry, biology, and tools for discovery. Chem. Rev. 2013;113:4633–4679. doi: 10.1021/cr300163e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pan J., Carroll K.S. Persulfide reactivity in the detection of protein S-sulfhydration. ACS Chem. Biol. 2013;8:1110–1116. doi: 10.1021/cb4001052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhang D., Macinkovic I., Devarie-Baez N.O., Pan J., Park C.M., Carroll K.S., Filipovic M.R., Xian M. Detection of protein S-sulfhydration by a tag-switch technique. Angew. Chem. Int. Ed. 2014;53:575–581. doi: 10.1002/anie.201305876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mustafa A.K., Gadalla M.M., Sen N., Kim S., Mu W., Gazi S.K., Barrow R.K., Yang G., Wang R., Snyder S.H. H2S signals through protein S-sulfhydration. Sci. Signal. 2009;2:ra72. doi: 10.1126/scisignal.2000464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gadalla M.M., Snyder S.H. Hydrogen sulfide as a gasotransmitter. J. Neurochem. 2010;113:14–26. doi: 10.1111/j.1471-4159.2010.06580.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sen N., Paul B.D., Gadalla M.M., Mustafa A.K., Sen T., Xu R., Kim S., Snyder S.H. Hydrogen sulfide-linked sulfhydration of NF-κb mediates its antiapoptotic actions. Mol. Cell. 2012;45:13–24. doi: 10.1016/j.molcel.2011.10.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Krishnan N., Fu C., Pappin D.J., Tonks N.K. H2S-Induced sulfhydration of the phosphatase PTP1B and its role in the endoplasmic reticulum stress response. Sci. Signal. 2011;4:ra86. doi: 10.1126/scisignal.2002329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Vandiver M.S., Paul B.D., Xu R., Karuppagounder S., Rao F., Snowman A.M., Ko H.S., Lee Y.I., Dawson V.L., Dawson T.M., et al. Sulfhydration mediates neuroprotective actions of parkin. Nat. Commun. 2013;4:1626. doi: 10.1038/ncomms2623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Paul B.D., Snyder S.H. H2S signaling through protein sulfhydration and beyond. Nat. Rev. Mol. Cell Biol. 2012;13:499–507. doi: 10.1038/nrm3391. [DOI] [PubMed] [Google Scholar]
- 27.Yang G., Zhao K., Ju Y., Mani S., Cao Q., Puukila S., Khaper N., Wu L., Wang R. Hydrogen sulfide protects against cellular senescence via S-sulfhydration of Keap1 and activation of Nrf2. Antioxid. Redox Signal. 2013;18:1906–1919. doi: 10.1089/ars.2012.4645. [DOI] [PubMed] [Google Scholar]
- 28.Sasakura K., Hanaoka K., Shibuya N., Mikami Y., Kimura Y., Komatsu T., Ueno T., Terai T., Kimura H., Nagano T. Development of a highly selective fluorescence probe for hydrogen sulfide. J. Am. Chem. Soc. 2011;133:18003–18005. doi: 10.1021/ja207851s. [DOI] [PubMed] [Google Scholar]
- 29.Duan X., Block E., Li Z., Connelly T., Zhang J., Huang Z., Su X., Pan Y., Wu L., Chi Q., et al. Crucial role of copper in detection of metal-coordinating odorants. Proc. Natl. Acad. Sci. USA. 2012;109:3492–3497. doi: 10.1073/pnas.1111297109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Blackstone E., Morrison M., Roth M.B. H2S induces a suspended animation-like state in mice. Science. 2005;308:518. doi: 10.1126/science.1108581. [DOI] [PubMed] [Google Scholar]
- 31.Collman J.P., Ghosh S., Dey A., Decre´au R.A. Using a functional enzyme model to understand the chemistry behind hydrogen sulfide induced hibernation. Proc. Natl. Acad. Sci. USA. 2009;106:22090–22095. doi: 10.1073/pnas.0904082106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kraus J.P., Williamson C.L., Firgaira F.A., Yang-Feng T.L., Münke M., Francke U., Bukovska G., Rosenberg L.E. Cloning and sequencing nanogram amounts of immunopurified mRNAs: cDNA cloning and chromosomal mapping of cystathionine β-synthase and the β subunit of propionyl CoA carboxylase. Proc. Natl. Acad. Sci. USA. 1986;83:2047–2051. doi: 10.1073/pnas.83.7.2047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Alexander F.W., Sandmeier E., Mehta P.K., Christen P. Evolutionary relationships among pyridoxal-5′-phosphate-dependent enzymes. Regio-specific alpha, beta and gamma families. Eur. J. Biochem. 1994;219:953–960. doi: 10.1111/j.1432-1033.1994.tb18577.x. [DOI] [PubMed] [Google Scholar]
- 34.Sun Q., Collins R., Huang S., Holmberg-Schiavone L., Anand G.S., Tan C.H., van-den-Berg S., Deng L.W., Moore P.K., Karlberg T., et al. Structural basis for the inhibition mechanism of human cystathionine-gammalyase: An enzyme responsible for the production of H2S. J. Biol. Chem. 2009;284:3076–3085. doi: 10.1074/jbc.M805459200. [DOI] [PubMed] [Google Scholar]
- 35.Messerschmidt A., Worbs M., Steegborn C., Wahl M.C., Huber R., Laber B., Clausen T. Determinants of enzymatic specificity in the Cys-Met metabolism PLPdependent enzyme family: Crystal structure of cystathionine γ-lyase from yeast and intrafamiliar structure comparison. Biol. Chem. 2003;384:373–386. doi: 10.1515/BC.2003.043. [DOI] [PubMed] [Google Scholar]
- 36.Huang S., Chua J.H., Yew W.S., Sivaraman J., Moore P.K., Tan C.H., Deng L.W. Site-directed mutagenesis on human Cystathionine-γ-lyase reveals insights into the modulation of H2S production. J. Mol. Biol. 2010;396:708–718. doi: 10.1016/j.jmb.2009.11.058. [DOI] [PubMed] [Google Scholar]
- 37.DeLano W.L. The PyMOL Molecular Graphics System. DeLano Scientific LLC; Palo Alto, CA, USA: 2002. [Google Scholar]
- 38.Stipanuk M.H. Sulfur amino acid metabolism: Pathways for production and removal of homocysteine and cysteine. Annu. Rev. Nutr. 2004;24:539–577. doi: 10.1146/annurev.nutr.24.012003.132418. [DOI] [PubMed] [Google Scholar]
- 39.Singh S., Padovani D., Leslie R.A., Chiku T., Banerjee R. Relative contributions of cystathionine beta-synthase and gamma-cystathionase to H2S biogenesis via alternative trans-sulfuration reactions. J. Biol. Chem. 2009;284:22457–22466. doi: 10.1074/jbc.M109.010868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kabil O., Banerjee R. The redox biochemistry of hydrogen sulfide. J. Biol. Chem. 2010;285:21903–21907. doi: 10.1074/jbc.R110.128363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Stipanuk M.H., Beck P.W. Characterization of the enzymic capacity for cysteine desulphhydration in liver and kidney of the rat. Biochem. J. 1982;206:267–277. doi: 10.1042/bj2060267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kimura H. Hydrogen sulfide: Its production, release and functions. Amino Acids. 2011;41:113–121. doi: 10.1007/s00726-010-0510-x. [DOI] [PubMed] [Google Scholar]
- 43.Olson K.R., Whitfield N.L., Bearden S.E., St Leger J., Nilson E., Gao Y., Madden J.A. Hypoxic pulmonary vasodilation: A paradigm shift with a hydrogen sulfide mechanism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010;298:51–60. doi: 10.1152/ajpregu.00576.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kamoun P. Endogenous production of hydrogen sulfide in mammals. Amino Acids. 2004;26:243–254. doi: 10.1007/s00726-004-0072-x. [DOI] [PubMed] [Google Scholar]
- 45.Mikami Y., Shibuya N., Kimura Y., Nagahara N., Ogasawara Y., Kimura H. Thioredoxin and dihydrolipoic acid are required for 3-mercaptopyruvate sulfurtransferase to produce hydrogen sulfide. Biochem. J. 2011;439:479–485. doi: 10.1042/BJ20110841. [DOI] [PubMed] [Google Scholar]
- 46.Whiteman M., Le Trionnaire S., Chopra M., Fox B., Whatmore J. Emerging role of hydrogen sulfide in health and disease: Critical appraisal of biomarkers and pharmacological tools. Clin. Sci. 2011;121:459–488. doi: 10.1042/CS20110267. [DOI] [PubMed] [Google Scholar]
- 47.Morikawa T., Kajimura M., Nakamura T., Hishiki T., Nakanishi T., Yukutake Y., Nagahata Y., Ishikawa M., Hattori K., Takenouchi T., et al. Hypoxic regulation of the cerebral microcirculation is mediated by a carbon monoxide-sensitive hydrogen sulfide pathway. Proc. Natl. Acad. Sci. USA. 2012;109:1293–1298. doi: 10.1073/pnas.1119658109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Searcy D.G., Lee S.H. Sulfur reduction by human erythrocytes. J. Exp. Zool. 1998;282:310–322. doi: 10.1002/(SICI)1097-010X(19981015)282:3<310::AID-JEZ4>3.0.CO;2-P. [DOI] [PubMed] [Google Scholar]
- 49.Benavides G.A., Squadrito G.L., Mills R.W., Patel H.D., Isbell T.S., Patel R.P., DarleyUsmar V.M., Doeller J., Kraus D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA. 2007;104:17977–17982. doi: 10.1073/pnas.0705710104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Belardinelli M.C., Chabli A., Chadefaux-Vekemans B., Kamoun P. Urinary sulfur compounds in Down syndrome. Clin. Chem. 2001;47:1500–1501. doi: 10.1093/clinchem/47.8.1500. [DOI] [PubMed] [Google Scholar]
- 51.Levitt M.D., Furne J., Springfield J., Suarez F., DeMaster E. Detoxification of hydrogen sulfide and methanethiol in the cecal mucosa. J. Clin. Investig. 1999;104:1107–1114. doi: 10.1172/JCI7712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Picton R., Eggo M.C., Merrill G.A., Langman M.J., Singh S. Mucosal protection against sulfide: Importance of the enzyme rhodanese. Gut. 2002;50:201–205. doi: 10.1136/gut.50.2.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kurzban G.P., Chu L., Ebersole J.L., Holt S.C. Sulfhemoglobin formation in human erythrocytes by cystalysin, an l-cysteine desulfhydrase from Treponema denticola. Oral Microbiol. Immunol. 1999;14:153–164. doi: 10.1034/j.1399-302X.1999.140303.x. [DOI] [PubMed] [Google Scholar]
- 54.Zanardo R.C., Brancaleone V., Distrutti E., Fiorucci S., Cirino G., Wallace J.L. Hydrogen sulfide is an endogenous modulator of leukocyte-mediated inflammation. FASEB J. 2006;20:2118–2120. doi: 10.1096/fj.06-6270fje. [DOI] [PubMed] [Google Scholar]
- 55.Yonezawa D., Sekiguchi F., Miyamoto M., Taniguchi E., Honjo M., Masuko T., Nishikawa H., Kawabata A. A protective role of hydrogen sulfide against oxidative stress in rat gastric mucosal epithelium. Toxicology. 2007;241:11–18. doi: 10.1016/j.tox.2007.07.020. [DOI] [PubMed] [Google Scholar]
- 56.Abe K., Kimura H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996;16:1066–1071. doi: 10.1523/JNEUROSCI.16-03-01066.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Laggner H., Hermann M., Esterbauer H., Muellner M.K., Exner M., Gmeiner B.M., Kapiotis S. The novel gaseous vasorelaxant hydrogen sulphide inhibits angiotensin-converting enzyme activity of endothelial cells. J. Hypertens. 2007;25:2100–2104. doi: 10.1097/HJH.0b013e32829b8fd0. [DOI] [PubMed] [Google Scholar]
- 58.Sivarajah A., Collino M., Yasin M., Benetti E., Gallicchio M., Mazzon E., Cuzzocrea S., Fantozzi R., Thiemermann C. Anti-apoptotic and anti-inflammatory effects of hydrogen sulfide in a rat model of regional myocardial I/R. Shock. 2009;31:267–274. doi: 10.1097/SHK.0b013e318180ff89. [DOI] [PubMed] [Google Scholar]
- 59.Zhang H., Huang Y., Chen S., Tang C., Wang G., Du J., Jin J. Hydrogen sulfide regulates insulin secretion and insulin resistance in diabetes mellitus, a new promising target for diabetes mellitus treatment? A review. J. Adv. Res. 2021;27:19–30. doi: 10.1016/j.jare.2020.02.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Lefer D.J. A new gaseous signaling molecule emerges: Cardioprotective role of hydrogen sulfide. Proc. Natl. Acad. Sci. USA. 2007;46:17907–17908. doi: 10.1073/pnas.0709010104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Cheng Y., Ndisang J.F., Tang G., Cao K., Wang R. Hydrogen sulfide-induced relaxation of resistance mesenteric artery beds of rats. Am. J. Physiol. Heart Circ. Physiol. 2004;287:H2316–H2323. doi: 10.1152/ajpheart.00331.2004. [DOI] [PubMed] [Google Scholar]
- 62.Yang G., Wu L., Jiang B., Yang W., Qi J., Cao K., Meng Q., Mustafa A.K., Mu W., Zhang S., et al. H2S as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionin gamma-lyase. Science. 2008;322:587–590. doi: 10.1126/science.1162667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Zagli G., Patacchini R., Trevisani M., Abbate R., Cinotti S., Gensini G.F., Masotti G., Geppetti P. Hydrogen sulfide inhibits human platelet aggregation. Eur. J. Pharmacol. 2007;559:65–68. doi: 10.1016/j.ejphar.2006.12.011. [DOI] [PubMed] [Google Scholar]
- 64.Fiorucci S., Antonelli E., Distrutti E., Rizzo G., Mencarelli A., Orlandi S., Zanardo R., Renga B., Di Sante M., Morelli A., et al. Inhibition of hydrogen sulfide generation contributes to gastric injury caused by anti-inflammatory nonsteroidal drugs. Gastroenterology. 2005;129:1210–1224. doi: 10.1053/j.gastro.2005.07.060. [DOI] [PubMed] [Google Scholar]
- 65.Perini R., Fiorucci S., Wallace J.L. Mechanisms of nonsteroidal anti-inflammatory drug-induced gastrointestinal injury and repair: A window of opportunity for cyclooxygenase-inhibiting nitric oxide donors. Can. J. Gastroenterol. 2004;18:229–236. doi: 10.1155/2004/890585. [DOI] [PubMed] [Google Scholar]
- 66.Wallace J.L., Caliendo G., Santagada V., Cirino G., Fiorucci S. Gastrointestinal safety and anti-inflammatory effects of a hydrogen sulfide-releasing diclofenac derivative in the rat. Gastroenterology. 2007;132:261–271. doi: 10.1053/j.gastro.2006.11.042. [DOI] [PubMed] [Google Scholar]
- 67.Fu Z., Liu X., Geng B., Fang L., Tang C. Hydrogen sulfide protects rat lung from ischemia-reperfusion injury. Life Sci. 2008;82:1196–1202. doi: 10.1016/j.lfs.2008.04.005. [DOI] [PubMed] [Google Scholar]
- 68.Zhang C., Du J., Bu D., Yan H., Tang X., Tang C. The regulatory effect of hydrogen sulphide on hypoxic pulmonary hypertension in rats. Biochem. Biophys. Res. Commun. 2003;302:810–816. doi: 10.1016/s0006-291x(03)00256-0. [DOI] [PubMed] [Google Scholar]
- 69.Wei H.L., Zhang C.Y., Jin H.F., Tang C.S., Du J.B. Hydrogen sulphide regulates lung tissue oxidized glutathione and total antioxidant capacity in hypoxic pulmonary hypertensive rats. Acta Pharmacol. Sin. 2008;29:670–676. doi: 10.1111/j.1745-7254.2008.00796.x. [DOI] [PubMed] [Google Scholar]
- 70.Jin H.F., Liang C., Liang J.M., Tang C.S., Du J.B. Effects of hydrogen sulphide on vascular inflammation in pulmonary hypertension induced by high pulmonary blood flow: Experiment with rats. Zhonghua Yi Xue Za Zhi. 2008;88:2235–2239. [PubMed] [Google Scholar]
- 71.Chen Y.H., Wu R., Geng B., Qi Y.F., Wang P.P., Yao W.Z., Tang C.S. Endogenous hydrogen sulfide reduces airway inflammation and remodeling in a rat model of asthma. Cytokine. 2009;45:117–123. doi: 10.1016/j.cyto.2008.11.009. [DOI] [PubMed] [Google Scholar]
- 72.Nicholson C.K., Calvert J.W. Hydrogen Sulfide and Ischemia—Reperfusion Injury. Pharmacol. Res. 2010;62:289–297. doi: 10.1016/j.phrs.2010.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Jiang T., Yang W., Zhang H., Song Z., Liu T., Lv X. Hydrogen Sulfide Ameliorates Lung Ischemia-Reperfusion Injury through SIRT1 Signaling Pathway in Type 2 Diabetic Rats. Front. Physiol. 2020;11:596. doi: 10.3389/fphys.2020.00596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kimura H. Hydrogen sulfide induces cyclic AMP and modulates the NMDA receptor. Biochem. Biophys. Res. Commun. 2000;267:129–133. doi: 10.1006/bbrc.1999.1915. [DOI] [PubMed] [Google Scholar]
- 75.Kimura H., Nagai Y., Umemura K., Kimura Y. Physiological roles of hydrogen sulfide: Synaptic modulation, neuroprotection, and smooth muscle relaxation. Antioxid. Redox Signal. 2005;7:795–803. doi: 10.1089/ars.2005.7.795. [DOI] [PubMed] [Google Scholar]
- 76.Tan B.H., Wong P.T.-H., Bian J.S. Hydrogen sulphide: A novel signalling molecule in the central nervous system. Neurochem. Int. 2010;56:3–10. doi: 10.1016/j.neuint.2009.08.008. [DOI] [PubMed] [Google Scholar]
- 77.Hu L.F., Wong P.T., Moore P.K., Bian J.S. Hydrogen sulphide attenuates lipopolysaccharide-induced inflammation by inhibition of p38 mitogen-activated protein kinase in microglia. J. Neurochem. 2007;100:1121–1128. doi: 10.1111/j.1471-4159.2006.04283.x. [DOI] [PubMed] [Google Scholar]
- 78.Roux P.P., Blenis J. ERK and p38 MAPK-activated protein kinases: A family of protein kinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 2004;68:320–344. doi: 10.1128/MMBR.68.2.320-344.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.d’Emmanuele di Villa Bianca R., Sorrentino R., Maffia P., Mirone V., Imbimbo C., Fusco F., De Palma R., Ignarro L.J., Cirino G. Hydrogen sulfide as a mediator of human corpus cavernosum smooth-muscle relaxation. Proc. Natl. Acad. Sci. USA. 2009;106:4513–4518. doi: 10.1073/pnas.0807974105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Fiorucci S., Santucci L., Distrutti E. NSAIDs, coxibs, CINOD and H2S-releasing NSAIDs: What lies beyond the horizon. Dig. Liver Dis. 2007;39:1043–1051. doi: 10.1016/j.dld.2007.09.001. [DOI] [PubMed] [Google Scholar]
- 81.Wallace J.L., Vong L. NSAID-induced gastrointestinal damage and the design of GI-sparing NSAIDs. Curr. Opin. Investig. Drugs. 2008;9:1151–1156. doi: 10.1254/jpssuppl.WCP2018.0_CL-24. [DOI] [PubMed] [Google Scholar]
- 82.Wallace J.L. Building a better aspirin: Gaseous solutions to a century-old problem. Br. J. Pharmacol. 2007;152:421–428. doi: 10.1038/sj.bjp.0707396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Wallace J.L., Ianaro A., de Nucci G. Gaseous Mediators in Gastrointestinal Mucosal Defense and Injury. Dig. Dis. Sci. 2017;62:2223–2230. doi: 10.1007/s10620-017-4681-0. [DOI] [PubMed] [Google Scholar]
- 84.Glowacka U., Brzozowski T., Magierowski M. Synergisms, discrepancies and interactions between hydrogen sulfide and carbon monoxide in the gastrointestinal and digestive system physiology, pathophysiology and pharmacology. Biomolecules. 2020;10:445. doi: 10.3390/biom10030445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Sulaieva O., Wallace J.L. Gaseous mediator-based anti-inflammatory drugs. Curr. Opin. Pharm. 2015;25:1–6. doi: 10.1016/j.coph.2015.08.005. [DOI] [PubMed] [Google Scholar]
- 86.Distrutti E., Sediari L., Mencarelli A., Renga B., Orlandi S., Russo G., Caliendo G., Santagada V., Cirino G., Wallace J.L., et al. 5-Amino-2-hydroxybenzoic acid 4-(5-thioxo-5H-[1,2]dithiol-3yl)-phenyl ester (ATB-429), a hydrogen sulfide-releasing derivative of mesalamine, exerts antinociceptive effects in a model of postinflammatory hypersensitivity. J. Pharm. Exp. 2006;319:447–458. doi: 10.1124/jpet.106.106435. [DOI] [PubMed] [Google Scholar]
- 87.Ekundi-Valentim E., Santos K.T., Camargo E.A., Denadai-Souza A., Teixeira S.A., Zanoni C.I., Grant A.D., Wallace J., Muscara M.N., Costa S.K. Differing effects of exogenous and endogenous hydrogen sulphide in carrageenan-induced knee joint synovitis in the rat. Br. J. Pharmacol. 2010;159:1463–1474. doi: 10.1111/j.1476-5381.2010.00640.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Distrutti E., Sediari L., Mencarelli A., Renga B., Orlandi S., Antonelli E., Roviezzo F., Morelli A., Cirino G., Wallace J.L., et al. Evidence that hydrogen sulfide exerts antinociceptive effects in the gastrointestinal tract by activating KATP channels. J. Pharmacol. Exp. Ther. 2006;316:325–335. doi: 10.1124/jpet.105.091595. [DOI] [PubMed] [Google Scholar]
- 89.Citi V., Martelli A., Gorica E., Brogi S., Testai L., Calderone V. Role of hydrogen sulfide in endothelial dysfunction: Pathophysiology and therapeutic approaches. J. Adv. Res. 2021;27:99–113. doi: 10.1016/j.jare.2020.05.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Testai L., Citi V., Martelli A., Brogi S., Calderone V. Role of hydrogen sulfide in cardiovascular ageing. Pharmacol. Res. 2020;160:105–125. doi: 10.1016/j.phrs.2020.105125. [DOI] [PubMed] [Google Scholar]
- 91.Chen Y., Zhang F., Yin J., Wu S., Zhou X. Protective mechanisms of hydrogen sulfide in myocardial ischemia. J. Cell. Physiol. 2020;235:9059–9070. doi: 10.1002/jcp.29761. [DOI] [PubMed] [Google Scholar]
- 92.Kang S.C., Sohn E.H., Lee S.R. Hydrogen Sulfide as a Potential Alternative for the Treatment of Myocardial Fibrosis. Oxid. Med. Cell. Longev. 2020;23:4105382. doi: 10.1155/2020/4105382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Roorda M., Miljkovic J.L., van Goor H., Henning R.H., Bouma H.R. Spatiotemporal regulation of hydrogen sulfide signaling in the kidney. Redox Biol. 2021;43:101961. doi: 10.1016/j.redox.2021.101961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Hsu C.N., Tain Y.L. Hydrogen Sulfide in Hypertension and Kidney Disease of Developmental Origins. Int. J. Mol. Sci. 2018;19:1438. doi: 10.3390/ijms19051438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Corvino A., Frecentese F., Magli E., Perissutti E., Santagada V., Scognamiglio A., Caliendo G., Fiorino F., Severino B. Trends in H2S-Donors Chemistry and Their Effects in Cardiovascular Diseases. Antioxidants. 2021;10:429. doi: 10.3390/antiox10030429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Pieretti J.C., Cruz Junho C.V., Sorelli Carneiro-Ramos M., Barozzi Seabra A. H2S- and NO-releasing gasotransmitter platform: A crosstalk signaling pathway in the treatment of acute kidney injury. Pharmacol. Res. 2020;161:105121. doi: 10.1016/j.phrs.2020.105121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Cao X., Zhang W., Moore P.K., Bian J. Protective Smell of Hydrogen Sulfide and Polysulfide in Cisplatin-Induced Nephrotoxicity. Int. J. Mol. Sci. 2019;20:313. doi: 10.3390/ijms20020313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Zhong H., Yu H., Chen J., Sun J., Guo L., Huang P., Zhong Y. Hydrogen Sulfide and Endoplasmic Reticulum Stress: A Potential Therapeutic Target for Central Nervous System Degeneration Diseases. Front. Pharmacol. 2020;11:702. doi: 10.3389/fphar.2020.00702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Gambari L., Grigolo B., Grassi F. Hydrogen Sulfide in Bone Tissue Regeneration and Repair: State of the Art and New Perspectives. Int. J. Mol. Sci. 2019;20:5231. doi: 10.3390/ijms20205231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Coavoy-Sánchez S.A., Costa S.K.P., Muscará M.N. Hydrogen sulfide and dermatological diseases. Br. J. Pharmacol. 2020;177:857–865. doi: 10.1111/bph.14699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Cao X., Ding L., Xie Z.Z., Yang Y., Whiteman M., Moore P.K., Bian J.S. A Review of Hydrogen Sulfide Synthesis, Metabolism, and Measurement: Is Modulation of Hydrogen Sulfide a Novel Therapeutic for Cancer? Antioxid. Redox Signal. 2019;31:1–38. doi: 10.1089/ars.2017.7058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Dattilo M. The role of host defences in Covid 19 and treatments thereof. Mol. Med. 2020;26:90. doi: 10.1186/s10020-020-00216-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Citi V., Martelli A., Brancaleone V., Brogi S., Gojon G., Montanaro R., Morales G., Testai L., Calderone V. Anti-inflammatory and antiviral roles of hydrogen sulfide: Rationale for considering H2S donors in COVID-19 therapy. Br. J. Pharmacol. 2020;177:4931–4941. doi: 10.1111/bph.15230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Liu H., Anders F., Thanos S., Mann C., Liu A., Grus F.H., Pfeiffer N., Prokosch-Willing V. Hydrogen Sulfide Protects Retinal Ganglion Cells Against Glaucomatous Injury In Vitro and In Vivo. Investig. Ophthalmol. Vis. Sci. 2017;58:5129–5141. doi: 10.1167/iovs.17-22200. [DOI] [PubMed] [Google Scholar]
- 105.Struve M.F., Brisbois J.N., James R.A., Marshall M.W., Dorman D.C. Neurotoxicological effects associated with short-term exposure of Sprague-Dawley rats to hydrogen sulfide. Neurotoxicology. 2001;22:375–385. doi: 10.1016/S0161-813X(01)00021-3. [DOI] [PubMed] [Google Scholar]
- 106.Volpato G.P., Searles R., Yu B., Scherrer-Crosbie M., Bloch K.D., Ichinose F., Zapol W.M. Inhaled Hydrogen Sulfide: A Rapidly Reversible Inhibitor of Cardiac and Metabolic Function in the Mouse. Anesthesiology. 2008;108:659–668. doi: 10.1097/ALN.0b013e318167af0d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Faller S., Ryter S.W., Choi A.M.K., Loop T., Schmidt R., Hoetzel A. Inhaled hydrogen sulfide protects against ventilator-induced lung injury. Anesthesiology. 2010;113:104–115. doi: 10.1097/ALN.0b013e3181de7107. [DOI] [PubMed] [Google Scholar]
- 108.Wagner F., Wagner K., Weber S., Stahl B., Knöferl M.W., Huber-Lang M., Seitz D.H., Asfar P., Calzia E., Senftleben U., et al. Inflammatory Effects of Hypothermia and Inhaled H2S During Resuscitated, Hyperdynamic Murine Septic Shock. Shock. 2011;35:396–402. doi: 10.1097/SHK.0b013e3181ffff0e. [DOI] [PubMed] [Google Scholar]
- 109.Blackstone E., Roth M.B. Suspended animation-like state protects mice from lethal hypoxia. Shock. 2007;27:370–372. doi: 10.1097/SHK.0b013e31802e27a0. [DOI] [PubMed] [Google Scholar]
- 110.Xue R., Hao D., Sun J., Li W., Zhao M., Li X., Chen Y., Zhu J., Ding Y., Liu J., et al. Hydrogen Sulfide Treatment Promotes Glucose Uptake by Increasing Insulin Receptor Sensitivity and Ameliorates Kidney Lesions in Type 2 Diabetes. Antioxid. Redox Signal. 2013;19:5. doi: 10.1089/ars.2012.5024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Chen K.Y., Morris J.C. Kinetics of oxidation of aqueous sulfide by oxygen. Environ. Sci. Technol. 1972;6:537. doi: 10.1021/es60065a008. [DOI] [Google Scholar]
- 112.Hughes M.N., Centelles M.N., Moore K.P. Making and working with hydrogen sulfide. Free Radic. Biol. Med. 2009;47:1346–1353. doi: 10.1016/j.freeradbiomed.2009.09.018. [DOI] [PubMed] [Google Scholar]
- 113.Calvert J.W., Elston M., Nicholson C.K., Gundewar S., Jha S., Elrod J.W., Ramachandran A., Lefer D.J. Genetic and Pharmacologic Hydrogen Sulfide Therapy Attenuates Ischemia-Induced Heart Failure in Mice. Circulation. 2010;122:11. doi: 10.1161/CIRCULATIONAHA.109.920991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Zhao W., Zhang J., Lu Y., Wang R. The vasorelaxant effect of H2S as a novel endogenous gaseous KATP channel opener. EMBO J. 2001;20:6008–6016. doi: 10.1093/emboj/20.21.6008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Mard S.A., Neisi N., Solgi G., Hassanpour M., Darbor M., MaMaleki M. Gastroprotective effect of NaHS against mucosal lesions induced by ischemia-reperfusion injury in rat. Dig. Dis. Sci. 2012;57:1496–1503. doi: 10.1007/s10620-012-2051-5. [DOI] [PubMed] [Google Scholar]
- 116.Gao C., Xu D.Q., Gao C.J., Ding Q., Yao L.N., Li Z.C., Chai W. An exogenous hydrogen sulphide donor, NaHS, inhibits the nuclear factor κB inhibitor kinase/nuclear factor κB inhibitor/nuclear factor- κB signaling pathway and exerts cardioprotective effects in a rat hemorrhagic shock model. Biol. Pharm. Bull. 2012;35:1029–1034. doi: 10.1248/bpb.b110679. [DOI] [PubMed] [Google Scholar]
- 117.Li N., Wang M.J., Jin S., Bai Y.D., Hou C.L., Ma F.F., Li X.H., Zhu Y.C. The H2S donor NaHS changes the expression pattern of H2S-Producing enzymes after myocardial infarction. Oxid. Med. Cell. Longev. 2016;2016:6492469. doi: 10.1155/2016/6492469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Esechie A., Kiss L., Olah G., Horvath E.M., Hawkins H., Szabo C., Traber D.L. Protective effect of hydrogen sulfide in a murine model of combined burn and smoke inhalation-induced acute lung injury. Clin. Sci. 2008;115:91. doi: 10.1042/CS20080021. [DOI] [PubMed] [Google Scholar]
- 119.Kloesch B., Liszt M., Steiner G., Broll J. Inhibitors of p38 and ERK1/2 MAPkinase and hydrogen sulphide block constitutive and IL-1β-induced IL-6 and IL-8 expression in the human chondrocyte cell line C-28/I2. Rheumatol. Int. 2012;32:729. doi: 10.1007/s00296-010-1682-0. [DOI] [PubMed] [Google Scholar]
- 120.Andruski B., McCafferty D.M., Ignacy T., Millen B., McDougall J.J. Leukocyte trafficking and pain behavioral responses to a hydrogen sulfide donor in acute monoarthritis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008;295:814–820. doi: 10.1152/ajpregu.90524.2008. [DOI] [PubMed] [Google Scholar]
- 121.Li L., Bhatia M., Zhu Y.Z., Zhu Y.C., Ramnath R.D., Wang Z.J., Anuar F.B.M., Whiteman M., Salto-Tellez M., Moore P.K. Hydrogen sulfide is a novel mediator of lipopolysaccharideinduced inflammation in the mouse. FASEB J. 2005;19:1196–1198. doi: 10.1096/fj.04-3583fje. [DOI] [PubMed] [Google Scholar]
- 122.Bhatia M., Wong F.L., Fu D., Lau H.Y., Moochhala S.M., Moore P.K. Role of hydrogen sulfide in acute pancreatitis and associated lung injury. FASEB J. 2005;19:623–625. doi: 10.1096/fj.04-3023fje. [DOI] [PubMed] [Google Scholar]
- 123.Mok Y.Y., Moore P.K. Hydrogen sulfide is pro-inflammatory in haemorrhagic shock. Inflamm. Res. 2008;57:512–528. doi: 10.1007/s00011-008-7231-6. [DOI] [PubMed] [Google Scholar]
- 124.Mok Y.Y., Atan M.S., Yoke Ping C., Zhong Jing W., Bhatia M., Moochhala S., Moore P.K. Role of hydrogen sulfide in haemorrhagic shock in the rat: Protective effect of inhibitors of hydrogen sulfide biosynthesis. Br. J. Pharmacol. 2004;143:881–889. doi: 10.1038/sj.bjp.0706014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Li T., Zhao B., Wang C., Wang H., Liu Z., Li W., Jin H., Tang C., Du J. Regulatory Effects of Hydrogen Sulfide on IL-6, IL-8 and IL-10 Levels in the Plasma and Pulmonary Tissue of Rats with Acute Lung Injury. Exp. Biol. Med. 2008;233:1081–1087. doi: 10.3181/0712-RM-354. [DOI] [PubMed] [Google Scholar]
- 126.Moore D.J., West A.B., Dawson V.L., Dawson T.M. Molecular pathophysiology of Parkinson’s disease. Annu. Rev. Neurosci. 2005;28:57–87. doi: 10.1146/annurev.neuro.28.061604.135718. [DOI] [PubMed] [Google Scholar]
- 127.Hu L.F., Lu M., Tiong C.X., Dawe G.S., Hu G., Bian J.S. Neuroprotective effects of hydrogen sulfide on Parkinson’s disease rat models. Aging Cell. 2010;9:135–146. doi: 10.1111/j.1474-9726.2009.00543.x. [DOI] [PubMed] [Google Scholar]
- 128.Giuliani D., Ottani A., Zaffe D., Galantucci M., Strinati F., Lodi R., Guarini S. Hydrogen sulfide slows down progression of experimental Alzheimer’s disease by targeting multiple pathophysiological mechanisms. Neurobiol. Learn. Mem. 2013;104:82–91. doi: 10.1016/j.nlm.2013.05.006. [DOI] [PubMed] [Google Scholar]
- 129.Bayan L., Koulivand P.H., Gorji A. Garlic: A review of potential therapeutic effects. Avicenna J. Phytomed. 2014;4:1–14. [PMC free article] [PubMed] [Google Scholar]
- 130.Lawson L.D., Wood S.G., Hughes B.G. HPLC analysis of allicin and other thiosulfinates in garlic clove homogenates. Planta Med. 1991;57:263–270. doi: 10.1055/s-2006-960087. [DOI] [PubMed] [Google Scholar]
- 131.Brodnitz M.H., Pascale J.V., Van Derslice L. Flavor components of garlic extract. J. Agric. Food Chem. 1971;19:273–275. doi: 10.1021/jf60174a007. [DOI] [Google Scholar]
- 132.Pluth M.D., Bailey T.S., Hammers M.D., Hartle M.D., Henthorn H.A., Steiger A.K. Natural Products Containing Hydrogen Sulfide Releasing Moieties. Synlett. 2015;26:2633–2643. doi: 10.1055/s-0035-1560638. [DOI] [Google Scholar]
- 133.Liang D., Wu H., Wong M.W., Huang D. Diallyl Trisulfide Is a Fast H2S Donor, but Diallyl Disulfide Is a Slow One: The Reaction Pathways and Intermediates of Glutathione with Polysulfides. Org. Lett. 2015;17:4196–4199. doi: 10.1021/acs.orglett.5b01962. [DOI] [PubMed] [Google Scholar]
- 134.Dinkova-Kostova A.T., Kostov R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012;18:337–347. doi: 10.1016/j.molmed.2012.04.003. [DOI] [PubMed] [Google Scholar]
- 135.Citi V., Martelli A., Testai L., Marino A., Breschi M.C., Calderone V. Hydrogen Sulfide Releasing Capacity of Natural Isothiocyanates: Is It a Reliable Explanation for the Multiple Biological Effects of Brassicaceae? Planta Med. 2014;80:610–613. doi: 10.1055/s-0034-1368591. [DOI] [PubMed] [Google Scholar]
- 136.Martelli A., Testai L., Citi V., Marino A., Bellagambi F.G., Ghimenti S., Breschi M.C., Calderone V. Pharmacological characterization of the vascular effects of aryl isothiocyanates: Is hydrogen sulfide the real player? Vascul. Pharmacol. 2014;60:32–41. doi: 10.1016/j.vph.2013.11.003. [DOI] [PubMed] [Google Scholar]
- 137.Lin Y., Yang X., Lu Y., Liang D., Huang D. Isothiocyanates as H2S Donors Triggered by Cysteine: Reaction Mechanism and Structure and Activity Relationship. Org. Lett. 2019;21:5977–5980. doi: 10.1021/acs.orglett.9b02117. [DOI] [PubMed] [Google Scholar]
- 138.Ozturk T., Ertas E., Mert O. Use of Lawesson’s Reagent in Organic Syntheses. Chem. Rev. 2007;107:5210–5278. doi: 10.1021/cr040650b. [DOI] [PubMed] [Google Scholar]
- 139.Lecher H.Z., Greenwood R.A., Whitehouse K.C., Chao T.H. The Phosphonation of Aromatic Compounds with Phosphorus Pentasulfide. J. Am. Chem. Soc. 1956;78:5018. doi: 10.1021/ja01600a058. [DOI] [Google Scholar]
- 140.Thomsen I., Clausen K., Scheibye S., Lawesson S.O. Thiation with 2,4-bis(4-methoxyphenyl)-1,3,2,4-dithiadiphosphetane 2,4-disulfide: N-methylthiopyrrolidone. Org. Synth. 1984;62:158. [Google Scholar]
- 141.Nicolau L.A.D., Silva R.O., Damasceno S.R.B., Carvalho N.S., Costa N.R.D., Aragão K.S., Barbosa A.L.R., Soares P.M.G., Souza M.H.L.P., Medeiros J.V.R. The hydrogen sulfide donor, Lawesson’s reagent, prevents alendronate-induced gastric damage in rats. Braz. J. Med. Biol. Res. 2013;46:708–714. doi: 10.1590/1414-431X20133030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Spiller F., Orrico M.I.L., Nascimento D.C., Czaikoski P.G., Souto F.O., Alves-Filho J.C., Freitas A., Carlos D., Montenegro M.F., Neto A.F., et al. Hydrogen Sulfide Improves Neutrophil Migration and Survival in Sepsis via K+ATP channel activation. Am. J. Respir. Crit. Care Med. 2010;182:360–368. doi: 10.1164/rccm.200907-1145OC. [DOI] [PubMed] [Google Scholar]
- 143.Li L., Whiteman M., Guan Y.Y., Neo K.L., Cheng Y., Lee S.W., Zhao Y., Baskar R., Tan C.H., Moore P.K. Characterization of a novel, water-soluble hydrogen sulfide-releasing molecule (GYY4137): New insights into the biology of hydrogen sulfide. Circulation. 2008;117:2351–2360. doi: 10.1161/CIRCULATIONAHA.107.753467. [DOI] [PubMed] [Google Scholar]
- 144.Rose P., Dymock B.W., Moore P.K. GYY4137, a novel water-soluble, H2S-releasing molecule. Methods Enzymol. 2015;554:143–167. doi: 10.1016/bs.mie.2014.11.014. [DOI] [PubMed] [Google Scholar]
- 145.Zhao Y., Biggs T.D., Xian M. Hydrogen sulfide (H2S) releasing agents: Chemistry and biological applications. Chem. Commun. 2014;50:11788–11805. doi: 10.1039/C4CC00968A. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Martelli A., Testai L., Citi V., Marino A., Pugliesi I., Barresi E., Nesi G., Rapposelli S., Taliani S., Da Settimo F., et al. Arylthioamides as H2S donors: l-cysteine-activated releasing properties and vascular effects in vitro and in vivo. ACS Med. Chem. Lett. 2013;4:904–908. doi: 10.1021/ml400239a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Karwi Q.G., Whiteman M., Wood M.E., Torregrossa R., Baxter G.F. Pharmacological postconditioning against myocardial infarction with a slow-releasing hydrogen sulfide donor, GYY4137. Pharmacol. Res. 2016;111:442–451. doi: 10.1016/j.phrs.2016.06.028. [DOI] [PubMed] [Google Scholar]
- 148.Qiu Y., Wu Y., Meng M., Luo M., Zhao H., Sun H., Gao S. GYY4137 protects against myocardial ischemia/reperfusion injury via activation of the PHLPP-1/Akt/Nrf2 signaling pathway in diabetic mice. J. Surg. Res. 2018;225:29–39. doi: 10.1016/j.jss.2017.12.030. [DOI] [PubMed] [Google Scholar]
- 149.Han X., Mao Z.C., Wang S., Xin Y., Li P., Maharjan S., Zhang B. GYY4137 protects against MCAO via p38 MAPK mediated anti-apoptotic signaling pathways in rats. Brain Res. Bull. 2020;158:59–65. doi: 10.1016/j.brainresbull.2020.02.015. [DOI] [PubMed] [Google Scholar]
- 150.Alexander B.E., Coles S.J., Fox B.C., Khan T.F., Maliszewski J., Perry A., Pitak M.B., Whiteman M., Wood M.E. Investigating the generation of hydrogen sulfide from the phosphonamidodithioate slow-release donor GYY4137. Med. Chem. Commun. 2015;6:1649. doi: 10.1039/C5MD00170F. [DOI] [Google Scholar]
- 151.Moore P.K., Whiteman M. Chemistry, Biochemistry and Pharmacology of Hydrogen Sulfide. Springer; Berlin/Heidelberg, Germany: 2015. pp. 344–354. [Google Scholar]
- 152.Park C., Zhao Y., Zhu Z., Pacheco A., Peng B., Devarie-Baez N.O., Bagdon P., Zhang H., Xian M. Synthesis and evaluation of phosphorodithioate-based hydrogen sulfide donors. Mol. Biosyst. 2013;9:2430–2434. doi: 10.1039/c3mb70145j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Kang J., Li Z., Organ C.L., Park C.M., Yang C.T., Pacheco A., Wang D., Lefer D.J., Xian M. pH-controlled hydrogen sulfide release for myocardial ischemia-reperfusion injury. J. Am. Chem. Soc. 2016;138:6336–6339. doi: 10.1021/jacs.6b01373. [DOI] [PubMed] [Google Scholar]
- 154.Wang H.W., Skeldon P., Thompson G.E., Wood G.C. Synthesis and characterization of molybdenum disulphide formed from ammonium tetrathiomolybdate. J. Mater. Sci. 1997;32:497. doi: 10.1023/A:1018538424373. [DOI] [Google Scholar]
- 155.Xu S., Yang C.T., Meng F.H., Pacheco A., Chen L., Xian M. Ammonium tetrathiomolybdate as a water-soluble and slow-release hydrogen sulfide donor. Bioorg. Med. Chem. Lett. 2016;26:1585–1588. doi: 10.1016/j.bmcl.2016.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Dyson A., Dal-Pizzol F., Sabbatini G., Lach A.B., Galfo F., Dos Santos Cardoso J., Pescador Mendonça B., Hargreaves I., Bollen Pinto B., Bromage D.I., et al. Ammonium tetrathiomolybdate following ischemia/reperfusion injury: Chemistry, pharmacology, and impact of a new class of sulfide donor in preclinical injury models. PLoS Med. 2017;14:1002310. doi: 10.1371/journal.pmed.1002310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Zhang H., Zhang A., Guo C., Shi C., Zhang Y., Liu Q., Sparatore A., Wang C. S-diclofenac protects against doxorubicin-induced cardiomyopathy in mice via ameliorating cardiac gap junction remodeling. PLoS ONE. 2011;6:e26441. doi: 10.1371/journal.pone.0026441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Wu J., Guo W., Lin S.Z., Wang Z.J., Kan J.T., Chen S.Y., Zhu Y.Z. Gp130-mediated STAT3 activation by S-propargyl-cysteine, an endogenous hydrogen sulfide initiator, prevents doxorubicin-induced cardiotoxicity. Cell. Death Dis. 2016;7:e2339. doi: 10.1038/cddis.2016.209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Su Y.W., Liang C., Jin H.F., Tang X.Y., Han W., Chai L.J., Zhang C.Y., Geng B., Tang C.S., Du J.B. Hydrogen sulfide regulates cardiac function and structure in adriamycin-induced cardiomyopathy. Circ. J. 2009;73:741–749. doi: 10.1253/circj.CJ-08-0636. [DOI] [PubMed] [Google Scholar]
- 160.Chan N., Willis A., Kornhauser N., Ward M.M., Lee S.B., Nackos E., Seo B.R., Chuang E., Cigler T., Moore A., et al. Influencing the tumor microenvironment: A phase II study of copper depletion using tetrathiomolybdate in patients with breast cancer at high risk for recurrence and in preclinical models of lung metastases. Clin. Cancer Res. 2017;23:666–676. doi: 10.1158/1078-0432.CCR-16-1326. [DOI] [PubMed] [Google Scholar]
- 161.Zanatta S.D., Jarrott B., Williams S.J. Synthesis and preliminary pharmacological evaluation of aryl dithiolethiones with cyclooxygenase-2-selective inhibitory activity and hydrogen sulfide-releasing properties. Aust. J. Chem. 2010;63:946–957. doi: 10.1071/CH09517. [DOI] [Google Scholar]
- 162.Li L., Rossoni G., Sparatore A., Lee L.C., Del Soldato P., Moore P.K. Anti-inflammatory and gastrointestinal effects of a novel diclofenac derivative. Free Radic. Biol. Med. 2007;42:706. doi: 10.1016/j.freeradbiomed.2006.12.011. [DOI] [PubMed] [Google Scholar]
- 163.Rakitin O.A. Synthesis and Reactivity of 3H-1,2-dithiole-3-thiones. Molecules. 2021;26:3595. doi: 10.3390/molecules26123595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Landis P.S. The chemistry of 1,2-dithiole-3-thiones. Chem. Rev. 1965;65:237. doi: 10.1021/cr60234a004. [DOI] [Google Scholar]
- 165.Curphey T.J., Libby A.A. Dianions of 3-Oxo dithioic Acids: Preparation and Conversion to 3H-1,2-Dithiole-3-thiones. Tetrahedron Lett. 2000;41:6977. doi: 10.1016/S0040-4039(00)01191-6. [DOI] [Google Scholar]
- 166.Pedersen B.S., Lawesson S.O. Syntheses of 3H-1,2-dithiole-3-thiones and 4H-1,3,2-oxazaphosphorine derivatives from the dimer of p-methoxyphenyl-thionophosphine sulfide and der. Tetrahedron. 1979;35:2433–2437. doi: 10.1016/S0040-4020(01)93760-3. [DOI] [Google Scholar]
- 167.Hamada T., Nakane T., Kimura T., Arisawa K., Yoneda K., Yamamoto T., Osaki T. Treatment of xerostomia with the bile secretion-stimulating drug anethole trithione: A clinical trial. Am. J. Med. Sci. 1999;318:146–151. doi: 10.1016/S0002-9629(15)40606-8. [DOI] [PubMed] [Google Scholar]
- 168.Perrino E., Cappelletti G., Tazzari V., Giavini E., Soldato P.D., Sparatore A. New sulfurated derivatives of valproic acid with enhanced histone deacetylase inhibitory activity. Bioorg. Med. Chem. Lett. 2008;18:1893–1897. doi: 10.1016/j.bmcl.2008.02.007. [DOI] [PubMed] [Google Scholar]
- 169.Tazzari V., Cappelletti G., Casagrande M., Perrino E., Renzi L., Del Soldato P., Sparatore A. New aryldithiolethione derivatives as potent histone deacetylase inhibitors. Bioorg. Med. Chem. 2010;18:4187–4194. doi: 10.1016/j.bmc.2010.05.011. [DOI] [PubMed] [Google Scholar]
- 170.Sen C.K., Traber K.E., Packer L. Inhibition of NF-jB activation in human T-cell lines by anetholdithiolthione. Biochem. Biophys. Res. Commun. 1996;218:148–153. doi: 10.1006/bbrc.1996.0026. [DOI] [PubMed] [Google Scholar]
- 171.Qandil A.M. Prodrugs of Nonsteroidal Anti-Inflammatory Drugs (NSAIDs), More Than Meets the Eye: A Critical Review. Int. J. Mol. Sci. 2012;13:17244. doi: 10.3390/ijms131217244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Sparatore A., Perrino E., Tazzari V., Giustarini D., Rossi R., Rossoni G., Erdman K., Schröder H., Del Soldato P. Pharmacological profile of a novel H2S-releasing aspirin. Free Radic. Biol. Med. 2009;46:586–592. doi: 10.1016/j.freeradbiomed.2008.11.013. [DOI] [PubMed] [Google Scholar]
- 173.Rossoni G., Manfredi B., Tazzari V., Sparatore A., Trivulzio S., Del Soldato P., Berti F. Activity of a new hydrogen sulfide-releasing aspirin (ACS14) on pathological cardiovascular alterations induced by glutathione depletion in rats. Eur. J. Pharmacol. 2010;648:139–145. doi: 10.1016/j.ejphar.2010.08.039. [DOI] [PubMed] [Google Scholar]
- 174.Kodela R., Chattopadhyay M., Kashfi K. NOSH-Aspirin: A novel nitric oxide–hydrogen sulfide-releasing hybrid: A new class of anti-inflammatory pharmaceuticals. ACS Med. Chem. Lett. 2012;3:257–262. doi: 10.1021/ml300002m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Rossoni G., Berti M., Colonna V.D., Bernareggi M., Del Soldato P., Berti F. Myocardial protection by the nitroderivative of aspirin, NCX 4016: In vitro and in vivo experiments in the rabbit. Italian Heart J. 2000;1:146–155. [PubMed] [Google Scholar]
- 176.Ahmed A., Rezai H., Broadway-Stringer S. Evidence-Based Revised View of the Pathophysiology of Preeclampsia. Adv. Exp. Med. Biol. Adv. Intern. Med. 2017;2:355–374. doi: 10.1007/5584_2016_168. [DOI] [PubMed] [Google Scholar]
- 177.Sanchez-Aranguren L.C., Rezai H., Ahmad S., Alzahrani F.A., Sparatore A., Wang K., Ahmed A. MZe786 Rescues Cardiac Mitochondrial Activity in High sFlt-1 and Low HO-1 Environment. Antioxidants. 2020;9:598. doi: 10.3390/antiox9070598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Rezai H., Ahmad S., Alzahrani F.A., Sanchez-Aranguren L., Dias I.H.K., Agrawal S., Sparatore A., Wang K., Ahmed A. MZe786, a hydrogen sulfide-releasing aspirin prevents preeclampsia in heme oxygenase-1 haplodeficient pregnancy under high soluble flt-1 environment. Redox Biol. 2021;38:101768. doi: 10.1016/j.redox.2020.101768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Fiorucci S., Orlandi S., Mencarelli A., Caliendo G., Santagada V., Distrutti E., Santucci L., Cirino G., Wallace J.L. Enhanced activity of a hydrogen sulphide releasing derivative of mesalamine (ATB-429) in a mouse model of colitis. Br. J. Pharmacol. 2007;150:996–1002. doi: 10.1038/sj.bjp.0707193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Fahn S. An open trial of high-dosage antioxidants in early Parkinson’s disease. Am. J. Clin. Nutr. 1991;53:380–382. doi: 10.1093/ajcn/53.1.380S. [DOI] [PubMed] [Google Scholar]
- 181.Rascol O., Brooks D.J., Korczyn A.D., De Deyn P.P., Clarke C.E., Lang A.E. A five-year study of the incidence of dyskinesia in patients with early Parkinson’s disease who were treated with ropinirole or levodopa. N. Engl. J. Med. 2000;342:1484–1491. doi: 10.1056/NEJM200005183422004. [DOI] [PubMed] [Google Scholar]
- 182.Lee M., Tazzari V., Giustarini D., Rossi R., Sparatore A., Del Soldato P., McGeer E., McGeer P.L. Effects of hydrogen sulfide-releasing L-DOPA derivatives on glial activation. J. Biol. Chem. 2010;285:17318–17328. doi: 10.1074/jbc.M110.115261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Kashfi K. Anti-inflammatory agents as cancer therapeutics. Adv. Pharmacol. 2009;57:31–89. doi: 10.1016/S1054-3589(08)57002-5. [DOI] [PubMed] [Google Scholar]
- 184.Chattopadhyay M., Kodela R., Olson K.R., Kashfi K. NOSH-aspirin (NBS-1120), a novel nitric oxide- and hydrogen sulfide-releasing hybrid is a potent inhibitor of colon cancer cell growth in vitro and in a xenograft mouse model. Biochem. Biophys. Res. Commun. 2012;419:523–528. doi: 10.1016/j.bbrc.2012.02.051. [DOI] [PubMed] [Google Scholar]
- 185.Kashfi K. Anti-cancer activity of new designer hydrogen sulfide-donating hybrids. Antioxid. Redox Signal. 2014;20:831–846. doi: 10.1089/ars.2013.5308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Hasegawa U., van del Vlies A.J. Design and synthesis of polymeric hydrogen sulfide donors. Bioconjug. Chem. 2014;25:1290–1300. doi: 10.1021/bc500150s. [DOI] [PubMed] [Google Scholar]
- 187.Zhang Y., Munday R. Dithiolethiones for cancer chemoprevention: Where do we stand? Mol. Cancer Ther. 2008;7:3470–3479. doi: 10.1158/1535-7163.MCT-08-0625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Wood M.E., Whiteman M., Perry A. Hydrogen Sulfide Releasing Compounds and Their Use. No. WO2013045951. U.S. Patent. 2013 April 4;
- 189.Szczesny B., Módis K., Yanagi K., Coletta C., Le Trionnaire S., Perry A., Wood M.E., Whiteman M., Szabo C. AP39, a novel mitochondria-targeted hydrogen sulfide donor, stimulates cellular bioenergetics, exerts cytoprotective effects and protects against the loss of mitochondrial DNA integrity in oxidatively stressed endothelial cells in vitro. Nitric Oxide. 2014;41:120–130. doi: 10.1016/j.niox.2014.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Ikeda K., Marutani E., Hirai S., Wood M.E., Whiteman M., Ichinose F. Mitochondria-targeted hydrogen sulfide donor AP39 improves neurological outcomes after cardiac arrest in mice. Nitric Oxide. 2015;49:90–96. doi: 10.1016/j.niox.2015.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Ahmad A., Olah G., Szczesny B., Wood M.E., Whiteman M., Szabo C. AP39, a mitochondrially targeted hydrogen sulfide donor, exerts protective effects in renal epithelial cells subjected to oxidative stress in vitro and in acute renal injury in vivo. Shock. 2016;45:88–97. doi: 10.1097/SHK.0000000000000478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Chatzianastasiou A., Bibli S.I., Andreadou I., Efentakis P., Kaludercic N., Wood M.E., Whiteman M., Di Lisa F., Daiber A., Manolopoulos V.G., et al. Cardioprotection by H2S donors: Nitric oxide-dependent and independent mechanisms. J. Pharmacol. Exp. Ther. 2016;358:431–440. doi: 10.1124/jpet.116.235119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Karwi Q.G., Bornbaum J., Boengler K., Torregrossa R., Whiteman M., Wood M.E., Schulz R., Baxter G.F. AP39, a mitochondria-targeting hydrogen sulfide (H2S) donor, protects against myocardial reperfusion injury independently of salvage kinase signalling. Br. J. Pharmacol. 2017;174:287–301. doi: 10.1111/bph.13688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Gerő D., Torregrossa R., Perry A., Waters A., Le-Trionnaire S., Whatmore J.L., Wood M., Whiteman M. The novel mitochondria-targeted hydrogen sulfide (H2S) donors AP123 and AP39 protect against hyperglycemic injury in microvascular endothelial cells in vitro. Pharmacol. Res. 2016;113:186–198. doi: 10.1016/j.phrs.2016.08.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Talalay P., De Long M.J., Prochaska H.J. Identification of a common chemical signal regulating the induction of enzymes that protect against chemical carcinogenesis. Proc. Natl. Acad. Sci. USA. 1988;85:8261–8265. doi: 10.1073/pnas.85.21.8261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Kim W., Gates K.S. Evidence for thiol-dependent production of oxygen radicals by 4-methyl-5-pyrazinyl-3H-1, 2-dithiole-3-thione: Possible relevance to the anticarcinogenic properties of 1, 2-dithiole-3-thiones. Chem. Res. Toxicol. 1997;10:296–301. doi: 10.1021/tx9601667. [DOI] [PubMed] [Google Scholar]
- 197.Zhao Y., Wang H., Xian M. Cysteine-activated hydrogen sulfide (H2S) donors. J. Am. Chem. Soc. 2011;133:15–17. doi: 10.1021/ja1085723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Foster J.C., Powell C.R., Radzinski S.C., Matson J.B. S-Aroylthiooximes: A facile route to hydrogen sulfide releasing compounds with structure-dependent release kinetics. Org. Lett. 2014;16:1558–1561. doi: 10.1021/ol500385a. [DOI] [PubMed] [Google Scholar]
- 199.Zhao Y., Bhushan S., Yang C., Otsuka H., Stein J.D., Pacheco A., Peng B., Devarie-Baez N.O. Aguilar myocardial ischemia-reperfusion injury. ACS Chem. Biol. 2013;8:1283–1290. doi: 10.1021/cb400090d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Roger T., Raynaud F., Bouillaud F., Ransy C., Simonet S., Crespo C., Bourguignon M.P., Villeneuve N., Vilaine J.P., Artaud I., et al. New biologically active hydrogen sulfide donors. ChemBioChem. 2013;14:2268–2271. doi: 10.1002/cbic.201300552. [DOI] [PubMed] [Google Scholar]
- 201.Cairns T.L., Evans G.L., Larchar A.W., Mckusick B.C. gem-Dithiols. J. Am. Chem. Soc. 1952;74:3982–3989. doi: 10.1021/ja01136a004. [DOI] [Google Scholar]
- 202.Berchtold G.A., Edwards B.E., Campaigne E., Carmack M. The preparation of a crystalline gem-dithiol under mild conditions. J. Am. Chem. Soc. 1959;81:3148. doi: 10.1021/ja01521a059. [DOI] [Google Scholar]
- 203.Zhao Y., Kang J., Park C.M., Bagdon P.E., Peng B., Xian M. Thiol-activated gem-dithiols: A new class of controllable hydrogen sulfide (H2S) donors. Org. Lett. 2014;16:4536–4539. doi: 10.1021/ol502088m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Zheng Y., Yueqin Z., Ji K., Pan Z., Chittavong V., Wang B. Esterase-Sensitive Prodrugs with Tunable Release Rates and Direct Generation of Hydrogen Sulfide. Angew. Chem. Int. Ed. 2016;55:4514–4518. doi: 10.1002/anie.201511244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Borchardt R.T., Cohen L.A. Stereopopulation Control. III. Facilitation of Intramolecular Conjugate Addition of the Carboxyl Group. J. Am. Chem. Soc. 1972;94:9175–9182. doi: 10.1021/ja00781a031. [DOI] [PubMed] [Google Scholar]
- 206.Rao Y., Li X., Nagorny P., Hayashida J., Danishefsky S.J. A Simple Method for the Conversion of Carboxylic Acids into Thioacids with Lawesson’s Reagent. Tetrahedron Lett. 2009;50:6684–6686. doi: 10.1016/j.tetlet.2009.09.080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Li Z., Organ C.L., Zheng Y., Wang B., Lefer D.J. Abstract 17903: Novel Esterase Activated Hydrogen Sulfide Donors Attenuate Myocardial Ischemia/ Reperfusion Injury. Circulation. 2016;134:A17903. [Google Scholar]
- 208.Chengelis C.P., Neal R.A. Studies of Carbonyl Sulfide Toxicity: Metabolism by Carbonic Anhydrase. Toxicol. Appl. Pharmacol. 1980;55:198–202. doi: 10.1016/0041-008X(80)90236-7. [DOI] [PubMed] [Google Scholar]
- 209.Chengelis C.P., Neal R.A. Hepatic Carbonyl Sulfide Metabolism. Biochem. Biophys. Res. Commun. 1979;90:993–999. doi: 10.1016/0006-291X(79)91925-9. [DOI] [PubMed] [Google Scholar]
- 210.Steiger A.K., Marcatti M., Szabo C., Szczesny B., Pluth M.D. Inhibition of Mitochondrial Bioenergetics by Esterase-Triggered COS/H2S Donors. ACS Chem. Biol. 2017;12:2117–2123. doi: 10.1021/acschembio.7b00279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Steiger A.K., Pardue S., Kevil C.G., Pluth M.D. Self-Immolative Thiocarbamates Provide Access to Triggered H2S Donors and Analyte Replacement Fluorescent Probes. J. Am. Chem. Soc. 2016;138:7256–7259. doi: 10.1021/jacs.6b03780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Powell C.R., Foster J.C., Okyere B., Theus M.H., Matson J.B. Therapeutic Delivery of H2S via COS: Small Molecule and Polymeric Donors with Benign Byproducts. J. Am. Chem. Soc. 2016;138:13477–13480. doi: 10.1021/jacs.6b07204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Tao X., Deng Y., Shen Z., Ling J. Controlled Polymerization of N-Substituted Glycine N-Thiocarboxyanhydrides Initiated by Rare Earth Borohydrides toward Hydrophilic and Hydrophobic Polypeptoids. Macromolecules. 2014;47:6173–6180. doi: 10.1021/ma501131t. [DOI] [Google Scholar]
- 214.Zhao Y., Pluth M.D. Hydrogen Sulfide Donors Activated by Reactive Oxygen Species. Angew. Chem. Int. Ed. 2016;55:14638–14642. doi: 10.1002/anie.201608052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Dickinson B.C., Huynh C., Chang C.J. A Palette of Fluorescent Probes with Varying Emission Colors for Imaging Hydrogen Peroxide Signaling in Living Cells. J. Am. Chem. Soc. 2010;132:5906–5915. doi: 10.1021/ja1014103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Miller E.W., Albers A.E., Pralle A., Isacoff E.Y., Chang C.J. Boronate-Based Fluorescent Probes for Imaging Cellular Hydrogen Peroxide. J. Am. Chem. Soc. 2005;127:16652–16659. doi: 10.1021/ja054474f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Zhou Z., Rekowski M.V.W., Coletta C., Szabo C., Bucci M., Cirino G., Topouzis S., Papapetropoulos A., Giannis A. Thioglycine and L-Thiovaline: Biologically Active H2S-donors. Bioorganic Med. Chem. 2012;20:2675–2678. doi: 10.1016/j.bmc.2012.02.028. [DOI] [PubMed] [Google Scholar]
- 218.Pan J., Devarie-Baez N.O., Xian M. Facile Amide Formation viaS-Nitrosothioacids. Org. Lett. 2011;13:1092–1094. doi: 10.1021/ol1031393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Rao Y., Li X., Danishefsky S.J. Thio FCMA Intermediates as Strong Acyl Donors: A General Solution to the Formation of Complex Amide Bonds. J. Am. Chem. Soc. 2009;131:12924–12926. doi: 10.1021/ja906005j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Wang P., Danishefshy S.J. A Promising General Solution to the Problem of Ligating Peptides and Glycopeptides. J. Am. Chem. Soc. 2010;132:17045–17051. doi: 10.1021/ja1084628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Liu R., Orgel L.E. Oxidative Acylation Using Thioacids. Nat. Cell Biol. 1997;389:52–54. doi: 10.1038/37944. [DOI] [PubMed] [Google Scholar]
- 222.Dawson E.P., Muir T.W., Clark-Lewis I., Kent S.B. Synthesis of Proteins by Native Chemical Ligation. Science. 1994;266:776–779. doi: 10.1126/science.7973629. [DOI] [PubMed] [Google Scholar]
- 223.Crich D., Sharma I. Epimerization-Free Block Synthesis of Peptides from Thioacids and Amines with the Sanger and Mukaiyama Reagents. Angew. Chem. Int. Ed. 2009;48:2355–2358. doi: 10.1002/anie.200805782. [DOI] [PubMed] [Google Scholar]
- 224.Wu D., Hu Q., Ma F., Zhu Y.Z. Vasorelaxant Effect of a New Hydrogen Sulfide-Nitric Oxide Conjugated Donor in Isolated Rat Aortic Rings through cGMP Pathway. Oxidative Med. Cell. Longev. 2016;2016:7075682 . doi: 10.1155/2016/7075682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Liu C., Guo W., Shi X., Kaium M., Gu X., Zhu Y.Z. Leonurine-Cysteine Analog Conjugates as a New Class of Multifunctional Anti-myocardial Ischemia Agent. Eur. J. Med. Chem. 2011;46:3996–4009. doi: 10.1016/j.ejmech.2011.05.073. [DOI] [PubMed] [Google Scholar]
- 226.Ignarro L.J., Napoli C., Loscalzo J. Nitric Oxide Donors and Cardiovascular Agents Modulating the Bioactivity of Nitric Oxide: An Overview. Circ. Res. 2002;90:21–28. doi: 10.1161/hh0102.102330. [DOI] [PubMed] [Google Scholar]
- 227.Xiong Y., Chang L.-L., Tran B., Dai T., Zhong R., Mao Y.C., Zhu Y.Z. ZYZ-803, a Novel Hydrogen SulfiDe-nitric Oxide Conjugated Donor, Promotes Angiogenesis via Cross-Talk between STAT3 and CaMKII. Acta Pharmacol. Sin. 2020;41:218–228. doi: 10.1038/s41401-019-0255-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Hu Q., Wu D., Ma F., Yang S., Tan B., Xin H., Gu X., Chen X., Chen S., Mao Y., et al. Novel Angiogenic Activity and Molecular Mechanisms of ZYZ-803, a Slow-Releasing Hydrogen Sulfide-Nitric Oxide Hybrid Molecule. Antioxid. Redox Signal. 2016;25:498–514. doi: 10.1089/ars.2015.6607. [DOI] [PubMed] [Google Scholar]
- 229.Wu D., Hu Q., Xiong Y., Zhu D., Mao Y., Zhu Y.Z. Novel H2S-NO Hybrid Molecule (ZYZ-803) Promoted Synergistic Effects Against Heart Failure. Redox Biol. 2018;15:243–252. doi: 10.1016/j.redox.2017.11.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Bucci M., Vellecco V., Cantalupo A., Brancaleone V., Zhou Z., Evangelista S., Calderone V., Papapetropoulos A., Cirino G. Hydrogen Sulfide Accounts for the Peripheral Vascular Effects of Zofenopril Independently of ACE Inhibition. Cardiovasc. Res. 2014;102:138–147. doi: 10.1093/cvr/cvu026. [DOI] [PubMed] [Google Scholar]
- 231.Terzuoli E., Monti M., Vellecco V., Bucci M., Cirino G., Ziche M., Morbidelli L. Characterization of Zofenoprilat as an Inducer of Functional Angiogenesis through Increased H2S Availability. Br. J. Pharmacol. 2015;172:2961–2973. doi: 10.1111/bph.13101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Monti M., Terzuoli E., Ziche M., Morbidelli L. The Sulphydryl Containing ACE Inhibitor Zofenoprilat Protects Coronary Endothelium from Doxorubicin-Induced Apoptosis. Pharmacol. Res. 2013;76:171–181. doi: 10.1016/j.phrs.2013.08.003. [DOI] [PubMed] [Google Scholar]
- 233.Monti M., Terzuoli E., Ziche M., Morbidelli L. H2S Dependent and Independent Anti-inflammatory Activity of Zofenoprilat in Cells of the Vascular Wall. Pharmacol. Res. 2016;113:426–437. doi: 10.1016/j.phrs.2016.09.017. [DOI] [PubMed] [Google Scholar]
- 234.Furuta S., Kiyosawa K., Higuchi M., Kasahara H., Saito H., Shioya H., Oguchi H. Pharmacokinetics of Temocapril, an ACE Inhibitor with Preferential Biliary Excretion, in Patients with Impaired Liver Function. Eur. J. Clin. Pharmacol. 1993;44:383–385. doi: 10.1007/BF00316478. [DOI] [PubMed] [Google Scholar]
- 235.Donnarumma E., Ali M.J., Rushing A.M., Scarborough A.L., Bradley J.M., Organ C.L., Islam K.N., Polhemus D.J., Evangelista S., Cirino G., et al. Zofenopril Protects Against Myocardial Ischemia–Reperfusion Injury by Increasing Nitric Oxide and Hydrogen Sulfide Bioavailability. J. Am. Hear. Assoc. 2016;5:1–17. doi: 10.1161/JAHA.116.003531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Ngowi E.E., Afzal A., Sarfra M., Khattak S., Zaman S.U., Khan N.H., Li T., Jiang Q.Y., Zhang X., Duan S.F., et al. Role of hydrogen sulfide donors in cancer development and progression. Int. J. Biol. Sci. 2021;17:73–88. doi: 10.7150/ijbs.47850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Li H., Xu F., Gao G., Gao X., Wu B., Zheng C., Wang P., Li Z., Hua H., Li D. Hydrogen sulfide and its donors: Novel antitumor and antimetastatic therapies for triple-negative breast cancer. Redox Biol. 2020;34:101564. doi: 10.1016/j.redox.2020.101564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Cardoso Amorim Reis A.K., Stern A., Pequeno Monteiro H. S-nitrosothiols and H2S donors: Potential chemo-therapeutic agents in Cancer. Redox Biol. 2019;27:101190. doi: 10.1016/j.redox.2019.101190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Fortunato S., Lenzi C., Granchi C., Citi V., Martelli A., Calderone V., Di Pietro S., Signore G., Di Bussolo V., Minutolo F. First Examples of H2S-Releasing Glycoconjugates: Stereoselective Synthesis and Anticancer Activities. Bioconjugate Chem. 2019;30:614–620. doi: 10.1021/acs.bioconjchem.8b00808. [DOI] [PubMed] [Google Scholar]
- 240.Citi V., Piragine E., Pagnotta E., Ugolini L., Di Cesare Mannelli L., Testai L., Ghelardini C., Lazzeri L., Calderone V., Martelli A. Anticancer properties of erucin, an H2S-releasing isothiocyanate, on human pancreatic adenocarcinoma cells (AsPC-1) Phytother. Res. 2019;33:845–855. doi: 10.1002/ptr.6278. [DOI] [PubMed] [Google Scholar]
- 241.Sestito S., Pruccoli l., Runfola M., Citi V., Martelli A., Saccomanni G., Calderone V., Tarozzi T., Rapposelli S. Design and synthesis of H2S-donor hybrids: A new treatment for Alzheimer’s disease? Eur. J. Med. Chem. 2019;184:111745. doi: 10.1016/j.ejmech.2019.111745. [DOI] [PubMed] [Google Scholar]
- 242.Sestito S., Daniele S., Pietrobono D., Citi V., Bellusci L., Chiellini G., Calderone V., Martini C., Rapposelli S. Memantine prodrug as a new agent for Alzheimer’s Disease. Sci. Rep. 2019;9:4612. doi: 10.1038/s41598-019-40925-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Pavlovskiy Y., Yashchenko A., Zayachkivska O. H2S Donors Reverse Age-Related Gastric Malfunction Impaired Due to Fructose-Induced Injury via CBS, CSE, and TST expression. Front. Pharmacol. 2020;11:1134. doi: 10.3389/fphar.2020.01134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Ercolano G., De Cicco P., Frecentese F., Saccone I., Corvino A., Giordano F., Magli E., Fiorino F., Severino B., Calderone V., et al. Anti-metastatic Properties of Naproxen-HBTA in a Murine Model of Cutaneous Melanoma. Front. Pharmacol. 2019;10:66. doi: 10.3389/fphar.2019.00066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Melino S., Leo S., Papajani V.T. Natural Hydrogen Sulfide Donors from Allium sp. As a Nutraceutical Approach in Type 2 Diabetes Prevention and Therapy. Nutrients. 2019;11:1581. doi: 10.3390/nu11071581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Cenac N., Castro M., Desormeaux C., Colin P., Sie M., Ranger M., Vergnolle N. A novel orally administered trimebutine compound (GIC-1001) is anti-nociceptive and features peripheral opioid agonistic activity and Hydrogen Sulphide-releasing capacity in mice. Eur. J. Pain. 2016;20:723–730. doi: 10.1002/ejp.798. [DOI] [PubMed] [Google Scholar]
- 247.Rapposelli S., Gambari L., Digiacomo M., Citi V., Lisignoli G., Manferdini C., Calderone V., Grassi F. A Novel H2S-releasing Amino- Bisphosphonate which combines bone anti-catabolic and anabolic functions. Sci. Rep. 2017;7:11940. doi: 10.1038/s41598-017-11608-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Corvino A., Severino B., Fiorino F., Frecentese F., Magli E., Perissutti E., Santagada V., Bucci M., Cirino G., Kelly G., et al. Fragment-based de novo design of a cystathionine γ-lyase selective inhibitor blocking hydrogen sulfide production. Sci. Rep. 2016;6:34398. doi: 10.1038/srep34398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Severino B., Corvino A., Fiorino F., Luciano P., Frecentese F., Magli E., Saccone I., Di Vaio P., Citi V., Calderone V., et al. 1,2,4-Thiadiazolidin-3,5-diones as novel hydrogen sulfide donors. Eur. J. Med. Chem. 2018;143:1677–1686. doi: 10.1016/j.ejmech.2017.10.068. [DOI] [PubMed] [Google Scholar]
- 250.Marino A., Martelli A., Citi V., Fu M., Wang R., Calderone V., Levi R. The novel H2S donor 4-carboxy-pheny isothiocyanate inhibits mast cell degranulation and renin release by decreasing intracellular calcium. Br. J. Pharmacol. 2016;173:3222–3234. doi: 10.1111/bph.13583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Barresi E., Nesi G., Citi V., Piragine E., Piano I., Taliani S., Da Settimo F., Rapposelli S., Testai L., Breschi M.C., et al. Iminothioethers as Hydrogen Sulfide Donors: From the Gasotransmitter Release to the Vascular Effects. J. Med. Chem. 2017;60:7512–7523. doi: 10.1021/acs.jmedchem.7b00888. [DOI] [PubMed] [Google Scholar]
- 252.Zhang J., Zhang Q., Wang Y., Li J., Bai Z., Zhao Q., He D., Wang Z., Zhang J., Chen Y. Toxicity, bioactivity, release of H2S in vivo and pharmaco-kinetics of H2S-donors with thiophosphamide structure. Eur. J. Med. Chem. 2019;176:456–475. doi: 10.1016/j.ejmech.2019.05.017. [DOI] [PubMed] [Google Scholar]
- 253.Zhao Y., Steiger A.K., Pluth M.D. Cyclic Sulfenyl Thiocarbamates Release Carbonyl Sulfide and Hydrogen Sulfide Independently in Thiol-Promoted Pathways. J. Am. Chem. Soc. 2019;141:13610–13618. doi: 10.1021/jacs.9b06319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Levinn C.M., Cerda M.M., Pluth M.D. Development and Application of Carbonyl Sulfide-Based Donors for H2S Delivery. Acc. Chem. Res. 2019;52:2723–2731. doi: 10.1021/acs.accounts.9b00315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Zhou S., Mou Y., Liu M., Du Q., Ali B., Ramprasad J., Qiao C., Hu L.F., Ji X. Insights into the Mechanism of Thiol-Triggered COS/H2S Release from N-Dithiasuccinoyl Amine. J. Org. Chem. 2020;85:8352–8359. doi: 10.1021/acs.joc.0c00559. [DOI] [PubMed] [Google Scholar]
- 256.Connal L.A. The benefits of macromolecular hydrogen sulfide prodrugs. J. Mater. Chem. B. 2018;6:7122. doi: 10.1039/C8TB02352B. [DOI] [PubMed] [Google Scholar]
- 257.Gojon G., Morales G.A. SG1002 and Catenated Divalent Organic Sulfur Compounds as Promising Hydrogen Sulfide Prodrugs. Antioxid. Redox Signal. 2020;33 doi: 10.1089/ars.2020.8060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Giordano F., Corvino A., Scognamiglio A., Citi V., Gorica E., Fattorusso C., Persico M., Caliendo G., Fiorino F., Magli E., et al. Hybrids Between H2S-donors And Betamethasone 17-Valerate or Triamcinolone Acetonide Inhibit Mast Cell Degranulation and Promote Hyperpolarization of Bronchial Smooth Muscle Cells. Eur. J. Med. Chem. 2021;221:113517. doi: 10.1016/j.ejmech.2021.113517. [DOI] [PubMed] [Google Scholar]
- 259.Corvino A., Citi V., Fiorino F., Frecentese F., Magli E., Perissutti E., Santagada V., Calderone V., Martelli A., Gorica E., et al. H2S donating corticosteroids: Design, synthesis and biological evaluation in a murine model of asthma. J. Adv. Res. 2021 doi: 10.1016/j.jare.2021.05.008. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Jiang X., Liu Y., Ma L., Ji R., Qu Y., Xin Y., Lv G. Chemopreventive activity of sulforaphane. Drug Des. Dev. Ther. 2018;12:2905–2913. doi: 10.2147/DDDT.S100534. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
No new data were created or analyzed in this paper. Data sharing is not applicable to this article.