

Abstract
Inflammatory processes and proinflammatory cytokines have a key role in the cellular processes of neurodegenerative diseases and are linked to the pathogenesis of functional and mental health disorders. Tumor necrosis factor alpha has been reported to play a major role in the central nervous system in Alzheimer’s disease, Parkinson’s disease and amyotrophic lateral sclerosis and many other neurodegenerative diseases. Therefore, a potent proinflammatory/proapoptotic tumor necrosis factor alpha could be a strong candidate for targeted therapy. Plant derivatives have now become promising candidates as therapeutic agents because of their antioxidant and chemical characteristics, and anti-inflammatory features. Recently, phytochemicals including flavonoids, terpenoids, alkaloids, and lignans have generated interest as tumor necrosis factor alpha inhibitor candidates for a number of diseases involving inflammation within the nervous system. In this review, we discuss how phytochemicals as tumor necrosis factor alpha inhibitors are a therapeutic strategy targeting neurodegeneration.
Key Words: brain, central nervous system, cytokine, herbal medicine, inflammation, neurodegenerative diseases, phytochemicals, tumor necrosis factor-alpha
Introduction
Neurodegeneration is a destructive process involving the progressive loss of function or death of neurons, including those of the central nervous system (CNS). A number of well-known diseases occur as a result of central neuronal loss, including Alzheimer’s disease (AD), Parkinson’s disease (PD) and amyotrophic lateral sclerosis (ALS), but there is no effective curative treatment for any of these conditions. In many neurodegenerative disorders, causation has not yet been identified. However, inflammatory processes and proinflammatory cytokines have a key role in the initiating processes of neurodegenerative disorders and are linked to the pathogenesis of functional and mental impairment (Gitler et al., 2017).
Several groups have demonstrated the pleiotropic effects of tumor necrosis factor alpha (TNF-α) and its involvement in various physiological inflammatory processes. TNF-α has been demonstrated to play a major role in neurodegenerative CNS disorders including AD, PD and ALS and many others (Chen et al., 2016). Thus, a potent proinflammatory/proapoptotic TNF-α provides a strong candidate for targeted therapy. Unfortunately, many otherwise promising therapeutic agents are largely unable to cross the blood-brain barrier (BBB), which severely hinders their clinical use in treating neuroinflammation in the CNS.
Natural phytochemicals can, in some cases, be less toxic than novel synthetic drugs. Some of these plant derivatives have now become promising candidates as therapeutic agents because of their chemical characteristics, antioxidant and anti-inflammatory properties (Kumar and Singh, 2015). Recently, phytochemicals with TNF-α blocking properties have emerged as interesting candidates for a variety of diseases underpinned by neuroinflammation. They may be promising lead compounds for the development of other derivatives offering boosted pharmacological features (Shal et al., 2018). This review addresses the pathological and physiological functions of TNF-α together with an overview of plant extracts that can interfere with TNF-α activity and production. We also focus on the therapeutic potential and effects of phytochemicals including polyphenols, terpenoids, alkaloids and steroids as TNF-α inhibitors toward amelioration and prevention of neuroinflammation as a novel treatment strategy for neurodegenerative diseases.
Search Strategy and Selection Criteria
The articles used in this review were conducted via the online library. These databases included PubMed, ScienceDirect, Google Scholar, Web of Science and Clinical Trials for literature describing human and animal model studies of neurodegenerative diseases from 2001 to 2020. The following search terms were used to locate articles specific to this study: “TNFα and neuroinflammation”, “phytochemicals and TNFα Inhibitors”, “phytochemicals and neurodegenerative diseases”. Variations of these terms were used to ensure exhaustive search results.
An Overview of Tumor Necrosis Factor Alpha Signaling Pathways Involved in Neuroinflammation
TNF-α is a proinflammatory cytokine involved in many neuroinflammatory and neurodegenerative disorders, including AD, Huntington’s disease, multiple sclerosis (MS), PD, stroke and spinal muscular atrophy (Moriwaki et al., 2015; Siebert et al., 2015). TNF-α has a weight of 26 kDa and is synthesized as a type II transmembrane protein. It can undergo proteolytic cleavage by TNF-α converting enzyme (TACE/ADAM17) to a 17 kDa monomeric protein (Hartl et al., 2020).
TNF-α can bind to two receptors. While TNF-α receptor I is expressed under normal conditions in many types of cells and tissues, TNF-α receptor II is expressed in immune and endothelial cells at low levels. TNFR1 has a death domain in its cytoplasmic domain and can activate an apoptosis pathway via the TNF receptor-associated death domain, Fas-associated death domain, and Fas-associated death domain-like interleukin-1β-converting enzyme (also called caspase-8). Caspase-8 activation in turn results in caspase-3 activation, and induces apoptosis via degradation of several proteins. TNF receptor-associated factor TRAF2 is also used by TNF receptor-associated death domain, which activates IκBα kinase (IKK-α) via receptor-interacting protein, resulting in IκBα phosphorylation, ubiquitination, and degradation. IκBα degradation leads to nuclear factor-κB (NF-κB) activation. Following the NF-κB activation, TNF induces the expression of genes involved in apoptosis. The activated TNFR1 could induce activation of apoptosis if the NF-κB pathway fails to activate (Pegoretti et al., 2018; Brás et al., 2020). Ras/mitogen-activated protein kinases (MAPK) and c-jun N-terminal kinases (JNK), and PI-3K/Akt signaling pathways (cell-survival signaling pathways) are other pathways that can be activated by TNFR1. Therefore, both apoptosis and cell survival mechanisms are triggered simultaneously by TNFR1 (Kang et al., 2021). TNFR2 can trigger MAPK, JNK, and NF-κB signaling pathways and it also activates apoptosis but the underlying mechanism is currently unknown (Shi and Sun, 2018).
The proinflammatory effects of TNFs associated with most diseases are mainly due to their ability to trigger the NF-κB signaling pathway. More than 200 genes have been recognized to be regulated by NF-κB activation (Kumar et al., 2004) and therefore inhibitors of NF-κB are able to alleviate TNF-linked disorders. Conclusively, we speculate that phytochemical drugs, which in some cases may have favorable safety profiles and yet remain efficient in inhibiting both production and action of TNF-α, are good therapeutic choices in the treatment of TNF-linked neurodegenerative diseases. This is discussed in more detail below.
Neurodegenerative Diseases and Phytochemicals as Tumor Necrosis Factor Alpha Inhibitors
Proinflammatory cytokines are essential determinants in the pathophysiology of neurodegenerative diseases. Thus, the role of neuroinflammation in neurodegeneration must be fully explained. With the important role of TNF-α in mediating a wide variety of neuroinflammatory disorders, it has become an important target for many types of drug development including phytochemicals (Velmurugan et al., 2018). The TNF-α pathway and its neuroinflammatory role in the following neurodegenerative disorders are summarized in (Figure 1).
Figure 1.
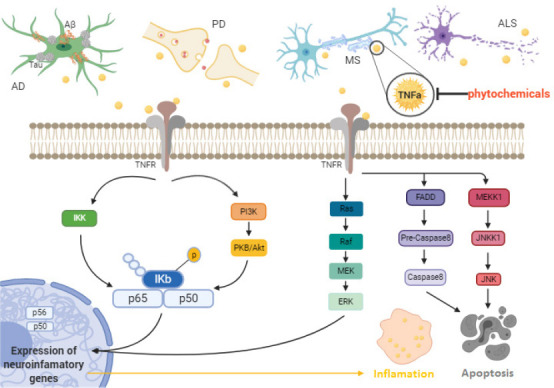
The role of the TNF-α pathway in neuroinflammation and neurodegeneration in vivo and in vitro.
AD: Alzheimer’s disease; ALS: amyotrophic lateral sclerosis; Aβ: amyloid-β peptide; ERK: extracellular signal-regulated kinase; FADD: fas-associated death domain protein; IKK: IκB kinase; JNK: c-Jun N-terminal kinases; JNKK1: c-Jun N-terminal kinase kinase 1; MEK: mitogen-activated protein kinase kinase; MEKK1: mitogen-activated protein kinase kinase kinase 1; MS: multiple sclerosis; PI3K: phosphoinositide 3-kinase; PKB: protein kinase B; TNFα: tumor necrosis factor alpha; TNFR: tumor necrosis factor alpha receptor.
AD and TNF-α inhibitors
There are three major neuropathological hallmarks of AD have been fully characterized: (i) the extracellular accumulation of amyloid beta (Aβ) oligomers and other materials into dense senile plaques; (ii) intracellular tau-containing neurofibrillary tangles in the brain; and (iii) chronic CNS inflammation (Hosseini et al., 2018). In AD development and related dementia, cytokine-mediated neuroinflammation plays a major role. TNF-α inhibits learning as a process critical for memory by dampening long term potentiation (Donzis and Tronson, 2014). Aβ peptides are particularly neurotoxic as key components of amyloid plaques. Aβ plaques in the brain activate macrophages which in turn lose their ability to phagocytose and remove this protein and the subsequent release of cytokines results in chronic inflammation, with TNF-α being the main culprit in this downward spiral leading to neuronal damage (Shamim and Laskowski, 2017). Dysregulation of the kynurenine pathway, mainly associated with an overexpression of indoleamine dioxygenase (IDO), has been implicated in the pathogenesis of neuroinflammatory and neurodegenerative disorders, such as MS and AD. It has been shown in vitro that primary human microglia and, in turn, IDO expression are activated by Aβ and that kynurenine pathway overactivation and IDO overexpression are associated with the pathogenesis of AD. Microglia are also activated by other proinflammatory cytokines, such as TNF-α, and at raised concentrations in the brain these have an additive effect further contributing to Aβ-activation of microglia and expression of IDO (Rodríguez-Gómez et al., 2020).
TNF-α also upregulates vascular cell adhesion molecule-1 in the endothelium, allowing the immune cells to move and adhere to regions under attack (Kinney et al., 2018). Therefore, vascular cell adhesion molecule-1 is also considered to be a putative drug target.
Among the neuroinflammatory cytokines, TNF-α is probably the most studied since it has an essential role in the cytokine cascade during the inflammatory response. TNF-α is a major proinflammatory cytokine that is upregulated in patients with AD (Wang et al., 2015b). Although TNF-α in the peripheral and central nervous systems of healthy adults is maintained at very low basal levels (Perry et al., 2001), there is an overexpression of TNF-α in AD patient brains, as observed by elevated levels of TNF-α in the CSF and blood plasma of AD patients (McCaulley and Grush, 2015). Further evidence is derived from multiple clinical and animal studies demonstrating the correlation between high TNF-α level and beta amyloid and tau protein expression in AD (Kinney et al., 2018).
Elevated TNF-α levels in AD appear to be associated with increased neuronal loss and dysfunction, enhanced Aβ production, decreased Aβ clearance, loss of synaptic function, all of which are related to the cognitive decline characteristic of AD (Chang et al., 2017). It has been shown that TNF-α may drive the neuroinflammatory process by potentiating the astroglial response. TNF-α, along with interleukin-1 and interferon-γ, can induce amyloid precursor protein cleavage via activation of the mitogen-activated protein kinase pathway. It is also capable of stimulating the NF-κB signaling that in turn leads to increased production of Aβ (Ekert et al., 2018).
As a TNF-α target, brain nerve growth factor (NGF) is an important neurotrophic factor crucial to the development and maintenance of brain cholinergic neurons. It appears there is an inverse relationship between TNF-α expression and brain NGF since elevated TNF-α is associated with reduced hippocampal NGF expression (Golan et al., 2004). TNF-α may have a role in regulating synaptic transmission and plasticity and therefore also Aβ-induced synaptic dysfunction. Moreover, TNF-α is associated with Aβ generation by regulating β-secretase 1 activity. The intensity of AD symptoms is correlated with the serum level of TNF-α in AD patients (Fu et al., 2018). In this regard, the overexpression of TNFR1 in the hippocampal tissue of mouse was sufficient to activate neuronal apoptosis prompted by NF-κB and Aβ. In addition, the upregulation of β-secretase production and increased γ-secretase activity by TNF-α may result in increased Aβ (Alasmari et al., 2018). This suggests that cytokines might be suitable targets for new AD interventions. Because of its key role in inflammation, TNF-α is an attractive pharmacological target and experimental findings have insinuated that increased TNF-α levels exacerbate amyloidogenesis. Hence, different approaches have been harnessed to reduce TNF-α signaling in rodent models of AD, showing significant reductions in AD-like brain pathology accompanied by an amelioration of cognitive dysfunction (Fu et al., 2018).
The FDA-approved biological tumor necrosis factor inhibitors may provide potential treatment opportunities for AD (Boado et al., 2010); however, a key problem with TNF-α inhibition via monoclonals or synthetic TNF receptors is the large size of the molecules which are as a result unable to cross the blood brain barrier (McCaulley and Grush, 2015). Currently, U.S. FDA-approved drugs for AD are limited to acetylcholinesterase inhibitors and the N-methyl-D-aspartate receptor antagonist memantine (Marotta et al., 2020). However, because of the potential for adverse effects and prescribing restrictions related to efficacy data, these drugs are prescribed only in limited circumstances (Kumar and Singh, 2015). These are effectively types of palliative care, attempting to decelerate the development of further cognitive symptoms and maintain a functional quality of life for longer. There remains no curative option for AD despite enormous effort to identify various disease-modifying therapies and to discover drugs that target molecular pathways to block AD progression (Kurz and Perneczky, 2011).
One further possibility in the pipeline is the development of a successful vaccine against AD. A large number of attempts have been made to produce effective vaccines against amyloidosis and tauopathy over the past two decades (there have been 140 promising strategies against Aβ deposition and 25 against Tau reported in the literature), but despite big investment in experimental models and/or in clinical trials none of these has successfully achieved FDA approval (Cacabelos, 2020). Lack of effective therapies has prompted attempts to screen natural compounds that could serve as pharmaceutical leads for drug development purposes (Naoi et al., 2019). Therefore, pharmacologically active natural components with anti-neuroinflammatory potential may be good candidates for the development of therapeutic options in AD. A number of preclinical and clinical trials have been completed on nutritional and botanical agents. Analysis of phytochemicals as anti-inflammatory and neuroprotective agents, such as phenolic derivatives, terpenoids, alkaloids, steroidal saponins, and glycosides, shows therapeutic potential (Shal et al., 2018). Importantly, a number of identified alkaloids, flavonoids, and terpenes can cross the BBB and are shown to have high bioavailability and good affinity to relevant receptors in the brain known to be important therapeutic targets in AD (Prasanna and Upadhyay, 2021).
MS and TNF-α inhibitors
MS is a chronic demyelinating CNS disease, characterized by axonal injury, a weakened BBB and disrupted nerve signaling (Filippi et al., 2018). The demyelination is probably due to the death of myelin-forming protective neuroglia, which can be caused in part by inflammation and immune reactions. Overall, inflammatory demyelination is the main cause of both symptoms and progression of MS (Bando, 2015).
A number of studies have investigated the role of TNF-α in MS. Proinflammatory cytokines such as TNF-α and interleukins (IL-17, IL-22, IL-23) appear to promote the development of MS and increased TNF-α concentrations have been found in MS lesions, whereby in patients with active MS, levels of TNF-α correlate with damage to the blood-brain barrier (Wang et al., 2018c). Also, peripherally increased concentrations of TNF-α have been associated with CNS synaptic instability, and may contribute to impairments of cognitive and sensory function (Dong et al., 2015). B cells also contribute to the pathophysiology of MS by secreting inflammatory cytokines including IL-6, IL-12, and TNF-α (Arneth, 2019).
Current treatments for MS may successfully suppress the immune system thus alleviating some of the symptoms; but, the drugs used have various unpleasant adverse effects, are prone to therapeutic failure and severe toxicity, and tend to be high-cost (Qureshi et al., 2018). Presumably partly because of this there has been a strong increase in the use of complementary and alternative medicine in MS patients, with between 33 and 80% of patients trialing various complementary and alternative medicine options, among which herbal remedies are the most common (Moawad et al., 2015). At the same time, evidence of the effectiveness of herbal medicines in MS is growing. Curcumin is one of the common plant-derived drugs in use. Curcumin may be neuroprotective in MS through multiple mechanisms. For example, Qureshi et al. showed that curcumin suppressed neuroinflammation by inhibiting cyclooxygenase-2 (COX-2), lipoxygenase, phospholipases, NF-κB, and activated protein-1. Curcumin also suppressed the production of TNF-α, IL-1β, macrophage inflammatory protein 1β, monocyte chemoattractant protein (MCP-1), and IL-8 in microglial and astrocytes cells (Qureshi et al., 2018). From a clinical perspective, patients with MS were enrolled in a study to examine the potential role of medicinal plant extracts or derivatives, such as a green tea, in the improvement of impairment associated with MS. It was shown that the effects of this extract on inflammation is associated with the suppression of key inflammatory cytokines, such as IL-1β, interferon γ, TNF-α, and inducible nitric oxide synthase (iNOS) (Farzaei et al., 2017).
PD and TNF-α inhibitors
PD is ranked as the second most common chronic neurodegenerative disease, affecting approximately 6 million people around the world with the characteristic and slowly physically disabling symptoms of resting tremor, rigidity, bradykinesia and postural instability, along with the less visible but equally disabling autonomic dysfunction, depression, anxiety, cognitive dysfunction, and sleep disturbance (Gopalakrishna and Alexander, 2015). The prevalence of PD is between 1% and 4% over 60 years of age. Motor symptoms appear with progressive damage of nigrostriatal dopaminergic neurons (Bassani et al., 2015; Moon and Paek, 2015). Lewy bodies are hallmark lesions of PD resulting from the cytoplasmic accumulation of misfolded α-synuclein and ubiquitin protein inclusions in neurons. This phenomenon results in neuronal death via necrosis and/or apoptosis (Shahpiri et al., 2016). Neuroinflammation also plays an major role in PD, adding to the degeneration and death of dopaminergic neurons in the substantia nigra (Hirsch and Hunot, 2009).
The proinflammatory cytokines TNF-α and IL-1β are increased in PD patients (Chan et al., 2021) and TNF-α concentrations are markedly augmented in the CSF and the brain of PD patients. TNFR1 is also increased in the substantia nigra (Mogi et al., 1994). Recent research illustrates that TNF-α and its receptors have similar roles in PD as previously described for AD. Therefore, pharmaceuticals with anti-TNF-α activities in PD could be effective in other neurodegenerative diseases. Although dopamine replacement therapy is the main effective treatment for PD, long-term utilization of this compound results in the accumulation of toxic metabolites and ROS. This, along with changes in dosing needs associated with disease progression, means such medications are only effective for around 10 years. Besides, these drugs are unable to reduce PD progression. Recently, studies showed considerable efforts to identify novel therapeutic options such as plant-derived compounds, especially polyphenols, with neuroprotective potential for the treatment of PD (de Andrade Teles et al., 2018; Carrera and Cacabelos, 2019).
Amyotrophic lateral sclerosis and TNF-α inhibitors
ALS is a complex, multifactorial, multisystem condition involving inflammation and an immune response in the neuromuscular, glial, and peripheral immune systems as part of the progression and development of the disease. A common feature of ALS is neuroinflammation, which can be detected in both the nervous system and in peripheral biological fluids (Vu and Bowser, 2017). Alterations of TNF-α concentration are common in nerve tissue, cerebrospinal fluid and blood in both ALS patients and animal models. TNF-α is key in governing mechanisms of ALS. However, the cell type and the receptors involved determine its role in influencing the pathophysiology either positively or negatively through all stages of ALS (Tortarolo et al., 2017) and there is a correlation between TNF-α activity and ALS (Guidotti et al., 2021).
Despite the poor prognosis it brings, there are limited drug options for treating ALS. Drugs such as riluzole may increase survival in ALS patients by about 2–3 months by delaying the need for tracheostomy and artificial ventilation due to its effects on slowing muscle atrophy and weakness, wasting, dysarthria, muscle spasticity, dysphagia, and improving overall patient quality of life (Andrews et al., 2020). But the limitations of the therapeutic options have driven attention towards alternatives such as natural products that could be used in novel therapeutic strategies to manage the symptoms of ALS. Generally, studies have shown that natural products may have useful effects on ALS patients, with few side effects and multiple receptor targets. Some natural antioxidant substances, which may have useful properties to be exploited in treating ALS, are resveratrol, Ginkgo biloba, ginseng, genistein, and epigallocatechin gallate (Nabavi et al., 2015).
Other diseases
Ischemic stroke often occurs when atherosclerotic and thrombotic processes block the arteries that carry blood to the brain (Anrather and Iadecola, 2016). Secondary brain damage happens when an uncontrolled neuroinflammatory cascade is triggered following ischemic and traumatic injury (Nakka et al., 2016), mainly due to the release of proinflammatory cytokines such as TNF-α.
TNF-α is present in every stage of neuronal damage in stroke such as inflammatory and prothrombotic events. TNF-α acts as a chemotactic agent toward leukocytes and induces production of intercellular adhesion molecule, vascular cell adhesion molecule, monocyte adhesion protein and E-selectin. Additionally, TNF-α exerts a key function in thrombogenesis by increasing plasminogen activating inhibitor-1 tissue factor and also platelet-activating factor, while suppressing activity of tissue plasminogen activator. Resveratrol is a polyphenolic phytochemical that appears to prevent TNF-α signaling via its action on NF-κB; this was shown following lipopolysaccharide (LPS) exposure in N9 microglial cells which were in co-culture with rat primary microglia (Bi et al., 2005; Bureau et al., 2008). Resveratrol treatment was also demonstrated to suppress TNF-α and IL-1β in adult mice following ischemic stroke (Shin et al., 2010). Resveratrol also prevented attenuation of NF-κB signaling associated with p65 nuclear translocation, and inhibited transcription of TNF-α, IL-1β, IL-6, and matrix metalloproteinase-9 (Shao et al., 2014; Lopez et al., 2015).
Another disease in which TNF-α plays a role is epilepsy. Epileptic seizures have been reported to cause neurodegenerative changes that may eventually lead to memory loss (Kamali et al., 2020). Neuroinflammation is a major factor that contributes significantly to the pathophysiology of epilepsy. Animal studies have shown that epileptic seizures activate CNS TNF-α expression (Kamaşak et al., 2020). Kaur et al. (2015) showed that curcumin supplementation to the convulsant drug pentylenetetrazole administered to Wistar rats resulted in decreased levels of mRNA and protein of IL-6, TNF-α, IL-1β and MCP-1.
Phytochemicals as Tumor Necrosis Factor Alpha Inhibitors
Beyond conventional medicine, there is evidence showing phytochemicals and traditional herbs can help preserve neurons and delay or slow development of neurodegenerative disorders. The anti-inflammatory, anti-oxidative, and anti-amyloidogenic characteristics of these compounds permit recovery through targeting different pathological causes of these diseases. Furthermore, neurotrophic factors, apoptotic factors, free radical scavenging processes and mitochondrial stress are modulated by a number of phytochemicals (Wang et al., 2018a).
Uncontrolled microglia activation in neurodegenerative disease generates neuronal damage due to the overproduction of TNF-α and nitric oxide (NO), leading to oxidative stress induction and apoptosis (Islam, 2017). With due attention to the role of TNF-α and targeting this as part of a suitable strategy for the treatment of ND, the identification of novel potent drugs from nature for prevention and/or treatment of neurodegenerative diseases is becoming a hot topic (Di Paolo et al., 2019). Many of the conventional drugs inhibiting TNF-α cause extremely unpleasant adverse effects (Kunnumakkara et al., 2019), and therefore it is extremely desirable to develop safer, less toxic anti-TNF-α drugs. An extensive source of TNF-α inhibitors is herbal compounds and already many herbal extracts and derivatives, belonging to different chemical classes, have been identified to block molecules upstream of TNF-α expression (Iqbal et al., 2013). Furthermore, lipophilic flavonoids, alkaloids, and terpenes can cross the BBB and have high bioavailability (Naoi et al., 2019), conferring a clear advantage and boosting clinical potential.
Polyphenols
Polyphenols are present in many plants and consist chemically of aromatic rings with multiple hydroxyl groups. These are subcategorized into flavonoids, flavones, flavanones, isoflavones, anthocyanins, catechins, phenolic acids (such as gallic and ferulic acid), stilbenes (such as resveratrol), curcumin, phytoestrogens, astaxanthin, diferoxymethane and tannins (Abbas et al., 2017).
ROS-scavenging enzymes such as catalase and superoxide dismutase are upregulated by polyphenols (Naoi et al., 2019). They also suppress macrophages by reducing COX-2 iNOS, and inhibit expression of IL-1β, TNF-α and IL-6 (González et al., 2011). Several polyphenols exert their influence on the balance between the development of pro and antiinflammatory cytokines, such as catechins and quercetin, such that they increase the release of IL-10 while inhibiting TNF-α and IL-1β (Yahfoufi et al., 2018). By modulating MAPK pathway at different levels of the signaling pathway, polyphenols can inhibit TNF-α release. Luteolin (a flavonoid polyphenol) reduces the release of TNF-α by LPS-activated macrophages, which blocks ERK1/2 and p38 phosphorylation (Xagorari et al., 2002).
Resveratrol
Resveratrol is a stilbenoid compound found in peanuts, grapes, tea and wine. It is named a miracle molecule, because of its antioxidant and anti-inflammatory effects with regards to neurodegenerative disorders (Meng et al., 2021). Following resveratrol treatment, various cognitive deficiencies induced in rat models were reversed by inhibiting TNF-α and IL-1 concentrations and increasing hippocampus brain-derived neurotrophic factor (BDNF) levels. Another study also showed resveratrol increased IL-10 concentration, promoting anti-inflammatory effects by inhibiting NF-κB and TNF-α levels (Ma et al., 2014). In yet another study, resveratrol suppressed the expression of the proinflammatory mediator TNF-α along with NF-κB while promoting the anti-inflammatory molecule IL-10 in microglial cells (Song et al., 2014).
Tannins
Tannins are a varied family of phytochemicals which may have a number of beneficial effects on health. Tannins are present in vegetables and fruits and are commonly consumed as part of the daily diet. The key neuroprotective effects of tannins stem from their ability to scavenge free radicals and activate the body’s endogenous antioxidant pathways conferring some protection against neurotoxins, neural inflammation and oxidative-stress-induced neuronal damage (Hussain et al., 2019).
Hydrolysable tannins (HTs) are found in terrestrial plants such as bananas and grapes. This is an important group with these tannins containing polyhydric alcohol within their core structure. A diverse class of HTs are ellagitannins from pomegranate fruit, which have shown some neuroprotection against PD. Ellagitannins act as a blocker of inflammatory mediators such as proinflammatory cytokines including TNF-α and IL-6 and tumor necrosis factor receptor 6 (TRAF-6). Thus, it appears to help improve the pathogenesis of PD by inhibiting neuroinflammation (Garcia et al., 2020). We summarize the CNS anti-inflammatory effects and presumed mechanisms of a number of polyphenols in Table 1.
Table 1.
A summary of polyphenols as inhibitors of TNF-α and neuroinflammatory responses in neurodegenerative diseases
| Polyphenol | Source | Disease | Mechanisms | IC50/dose | Model | Reference |
|---|---|---|---|---|---|---|
| Curcumin | Curcuma longa | AD | Activates PKC/ERK-mediated CREB regulation and Akt/GSK-3 mediated regulation, induces BDNF secretion, and inhibits Cas3, TNF-α, and NF-κB levels | 50 mg/kg/d 300 mg/kg |
Rat Adult and aged Mice |
Hoppe et al., 2013; Nam et al., 2014 |
| a-Mangostin Rosmarinic acid |
Garcinia mangostana
Melissa officinalis |
AD | Suppresses inflammation, increases BDNF level and decreasing phospho-tau (p-tau). Suppresses the expression of HIF-a, IL-1β, TNF-α and caspase 3 | 10 μM in culture and 5000 ppm in diet 10 μg/mL in culture and 50, 100, 200 and 400 mg/kg in vivo |
OHSC and transgenic Alzheimer mice Cortical neuronal cell and Sprague-Dawley rats |
Bayat et al., 2012; Huang et al., 2014 |
| Oxyresveratrol | Morus alba L | AD | Decreases TNF-α, IL-6 and inhibits activation of NALP3, caspase-1, NF- κB and inhibits phosphorylation of ERK, c-JNK and p38 | 100 ng/mL in culture and 7.5 or 10 mg/kg 10, 20, and 40 μg/mL |
RAW 264.7 and Sprague-Dawley rats Rat cortical neurons |
Chung et al., 2003; Wang et al., 2014 |
| 6-Shogaol | Zingiber officinale | AD | Induces NGF, BDNF, and GDNF secretion, inhibiting NO, TNF-α, IL-1β, p38, Bax, NF-κB, iNOS, PGE2, and ROS level and increases Bcl-2, and SOD levels | 1, 5 and 10 mM in culture 5 mg/kg and 20 mg/kg |
BV-2 cell line C57BL/6 mice |
Ha et al., 2012 |
| EGCG | Tea | AD | Suppresses the expression of TNF-α, IL-1β, IL-6, and iNOS. Restored the levels of intracellular antioxidants Nrf2 and HO-1, thus inhibiting reactive oxygen species-induced NF-κB activation | 20 μM | Aβ-stimulated EOC 13.31 microglia | Cheng-Chung Wei et al., 2016 |
| Rutin | Ruta graveolens | PD | Decreases TNF-α and IL-1β and NO | 25 mg/kg | Wistar rats | Khan et al., 2012 |
| Myricitrin | Myrica cerifera | PD | Decreases TNF-α-positive microglia | 60 mg/kg | C57BL/6 mice | Kim et al., 2016 |
| Resveratrol | Vitis vinifera | PD | Decreases TNF-α and COX-2 mRNA and COX-2 protein | 10, 20 and 40 mg/kg | Sprague-Dawley rats | Jin et al., 2008 |
| Ellagic acid | Different berries | PD | Decreases IL-1β and TNF-α protein | 50 mg/kg | Wistar rats | Zare et al., 2015 |
| EGCG | Tea | PD | Decreases TNF-α and IL-6 protein in plasma | 25, 50 mg/kg/d | C57BL/6 mice | Zhou et al., 2018 |
| Naringin | Citrus fruits | PD | Decreases TNF-α protein and increases GDNF protein and mTORC1 activity | 8 and 800 mg/kg | Sprague-Dawley rats | Leem et al., 2014 |
| Silibinin | Silybum | PD | Decreases IL-1β, TNF-α and iNOS protein | 50, 100 mg/kg | Sprague-Dawley | Jung et al., 2014 |
| Curcumin | Curcuma longa | PD | Decreases IL-6 and TNF-α protein and iNOS protein and NO content | 150 mg/kg | C57BL/6 mice | Ojha et al., 2012 |
| Theaflavin | Tea | PD | Decreases IL-1β, TNF-α, IL-6 as well as MPTP-induced IL-4 and IL-10, GFAP and COX-2 protein | 10 mg/kg | C57BL/6 mice | Anandhan et al., 2013 |
| Apigenin | Chamomile tea | PD | Decreases GFAP and TNF-α protein | 5, 10 and 20 mg/kg | Swiss-albino mice | Patil et al., 2014 |
| Syringic acid | Fagus sylvatica L | PD | Decreases IL-1β, TNF-α protein and COX-2 protein | 20 mg/kg | C57BL/6 mice | Rekha et al., 2014 |
| Ferulic acid | Chloroxylon swietenia DC | PD | Decreases IL-1β, TNF-α, IL-6 protein, COX-2 and iNOS protein | 80 mg/kg | Wistar rats | Ojha et al., 2015 |
| Resveratrol | Vitis vinifera | ND | Suppresses NO, TNF-α, iNOS, IL-1β, and IL-6 Inhibits MAPKs (ERK1/2, JNK, and p38) and NF-κB signaling pathways |
10 μM | BV-2 microglia cells | Wang et al., 2020 |
| EGCG | Tea | PD | Decreases TNF-α and nitrite | 10 mg/kg | Sprague-Dawley rats | Angeline et al., 2013 |
| Nobiletin | Citrus peels | PD | Deacreses TNF-α protein, (IL)-1β, and IL-6 | 100 mg/kg/d | Mouse microglia BV-2 cell line | Qi et al., 2019 |
| Curcumin | Curcuma longa | Epilepsy | Decreases TNF-α mRNA, Caspase 3 and iNOS expression | 0, 40, 80 mg/kg | Pilocarpine-induced status epilepticus (SE) model in rats | Aminirad et al., 2017 |
| Curcumin | Curcuma longa | Epilepsy | Decreases neuronal cell death and mRNA level of TNF-α | 12.5 mg/kg | Pentylenetetrazol (PTZ)-induced kindling model | Mansoor et al., 2018 |
| Resveratrol | Vitis vinifera | Dementia | Activates ERK-mediated CREB regulation, induces NGF, GDNF, and BDNF secretion, and inhibits caspase3, TNF-α, NF-κB, IL10, IL-1β, MCP1, and MDA levels, increases SOD level | 20 mg/kg for 7 d | Wistar rat model of vascular dementia | Anastacio et al., 2014 |
| Curcumin | Curcuma longa | Depression | Increases TNF-α and caspase-3 levels along with a marked reduction in neurogenesis factor (BDNF) | 100, 200 and 400 mg/kg |
Rat | Rinwa et al., 2013 |
| Resveratrol | Vitis vinifera | Brain ischemia | Suppresses the mRNA expression of TNF-α and promotes the mRNA expression of the interleukin-10 | 25 μM | Murine BV2 microglial cells | Song et al., 2014 |
| Curcumin | Curcuma longa | Brain ischemia | Increased neuronal viability and attenuated immunoreactivity of cyclooxygenase-2 (COX-2) and TNF-α | 25 or 50 mg/kg | Wistar Rat | de Alcântara et al., 2016 |
| Baicalin | Genus Scutellaria | Cerebral ischemia-reperfusion | Down-regulated expression of NOD2 and TNF-α | 10 μg/mL 50 mg/kg |
BV2, PC12 cells and primary neuron cells and the mice of cerebral ischemia-reperfusion | Li et al., 2010 |
| Baicalin | Genus Scutellaria | Brain ischemia | Reduces expression of TLR2/4 and NF-κB, decreased the expression and activity of iNOS and COX-2 Attenuated the serum content of TNF-α and IL-1β |
100 mg/kg | Adult Sprague-Dawley Rat | Tu et al., 2011 |
| Red ginseng extract | Panax Ginseng | Brain ischemia | Reduces serum levels of TNF-α, IL-1β and IL-6, and increased serum IL-10 level | 100 mg/kg/d orally | Adult Sprague-Dawley Rat | Lee et al., 2011 |
| Naringenin | Citrus paradisi | ND | Inhibition of LPS-induced TNF-α by suppressing phosphorylation of P38, serines 63 and 73. It also increases ERK5 phosphorylation thus inhibiting inflammatory response | 14.41 μM | Glial cells | Vafeiadou et al., 2009 |
| Phytoestrogen | Poincianella pyramidalis | ND | Decreases the expression of TNF-α, IL-1β, and connexins CCL5 and CCL2, as well as increases the expression of the IL-10 | 0.1 and 1 µM | Glial cells | de Almeida et al., 2020 |
| Bilberry anthocyanins | Vaccinium myrtillus L | AD | Activates the microglia and astrocytes, downregulates the TNF-α, NF-Kβ, IL-1β, IL-6, COX-2, iNOS and CD33 and CX3CR1, and upregulates the expression of TREM2 and TYROBP and TLR2 and TLR4 | 20 mg/kg/d | APP/PSEN1 transgenic AD model | Li et al., 2020 |
AD: Alzheimer’s disease; ALS: amyotrophic lateral sclerosis; Aβ: amyloid-β peptide; BDNF: brain-derived neurotrophic factor; CCL5: C-C Motif Chemokine Ligand 5; COX-2: Cyclooxygenase-2; CREB: cAMP response element-binding protein; CX3CR1: C-X3-C Motif Chemokine Receptor 1; ERK: extracellular signal-regulated kinase; FADD: fas-associated death domain protein; GDNF: glial cell line-derived neurotrophic factor; GFAP: glial fibrillary acidic protein; GSK-3: glycogen synthase kinase 3; HIF-a: hypoxia-inducible factor 1-alpha; HO-1: heme oxygenase-1; IKK: IκB kinase; iNOS: inducible nitric oxide synthase; JNK: c-Jun N-terminal kinases; JNKK1: c-Jun N-terminal kinase kinase 1; MEK: mitogenactivated protein kinase kinase; MEKK1: mitogen-activated protein kinase kinase kinase 1; MPTP: 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; MS: multiple sclerosis; mTORC1: mechanistic target of rapamycin complex 1; ND: neurodegenerative disease; NGF: nerve growth factor; Nrf2: nuclear factor erythroid 2 related factor 2; PD: Parkinson’s disease; PI3K: phosphoinositide 3-kinase; PKB: protein kinase B; ROS: reactive oxygen species; SOD: superoxide dismutase; TLR-2: Toll-like receptor-2; TNFR: tumor necrosis factor alpha receptor; TNFα: tumor necrosis factor alpha; TREM2: triggering receptor expressed on myeloid cells 2; TYROBP: tyrosine kinase-binding protein.
Flavonoids
Flavonoids are the largest groups of polyphenols with > 6000 compounds identified. ROS production is prevented by the flavonoids apigenin and kaempferol inhibiting xanthine oxidase which produce ROS. Other flavonoids including quercetin, baicalein, epigallocatechin gallate and myricetin and alsonon-flavonoids including gallic and protocatechuic acids have been reported to chelate iron and copper ions, this reducing the production of free radicals (Naoi et al., 2019). There is a close relationship between EGCG and inflammatory neurodegenerative diseases. Inflammation may be inhibited by reducing the expression of the inflammatory molecules IL1, TNF and TGB. However, studies have shown than when high concentrations of EGCG were administered, conflicting findings were observed. EGCG increased the expression of inflammatory TNF and IL6 at higher concentrations (Li et al., 2004; Wei et al., 2016).
Finally, the flavonoid curcumin, lycopene (found in tomatoes) and stilbenoids inhibit the proinflammatory NF-κB and activated protein-1, this suppressing the synthesis of IL-6, TNF-α, MCP-1 and iNOS (Spagnuolo et al., 2018).
Curcumin
Curcumin is a major components of turmeric and has different pharmacological properties such as anti-inflammatory effects that are relevant to the treatment of various diseases (Iranshahi et al., 2010; Sahebkar, 2010; Ghandadi and Sahebkar, 2017; Hewlings and Kalman, 2017; Teymouri et al., 2017; Bianconi et al., 2018; Panahi et al., 2018a, b; Bashang and Tamma, 2020; Ghasemi et al., 2019; Mollazadeh et al., 2019; Bagheri et al., 2020). Curcumin, by inhibiting NF-κB, was found to have high affinity for amyloid plaques thus potentially reducing the pathogenesis of AD (Reddy et al., 2018). Curcumin may also defend against PD through destabilization of α-synuclein protein (Liu et al., 2014). In addition, curcumin has been shown to alleviate neurodegenerative disorders by disrupting amyloid plaques, scavenging ROS, and exhibiting anti-apoptotic and anti-inflammatory effects (Darvesh et al., 2012). As acting on neurons, curcumin was reported to boost regenerating neurons by activating Trk/PI3K signaling pathways, which in turn elevated BDNF levels in a PD model. It may reduce caspase and TNF-α levels while simultaneously elevating BDNF concentration (Liu et al., 2014).
Nowadays, research is focused on finding more lipophilic derivatives of to facilitate crossing of the BBB and greater affinity for amyloid plaques (Lee et al., 2019). A recent study showed that curcumin decreased amyloid plaque formation both in vitro and in vivo in a rodent model (Wang et al., 2018b). Also, in another study, curcumin appeared to decrease expression of IL-6 and TNF-α (Rogers and Lue, 2001). Furthermore, in the JNK pathway, TNF-α, PGE2, IL-1β, iNOS, caspase, Bax, and cytochrome C inhibit Bcl2 and Bcl-xL so induce apoptosis in neurodegenerative disorders. Curcumin inhibited the JNK pathway in MPP- and MPTP-induced neurotoxicity in animal models (Yu et al., 2010).
Terpenoids
The largest family of natural compounds is the terpenoids (terpenes) and this group is particularly biologically active having antibiotic, antiparasitic, antitumor and anti-inflammatory properties (Nguyen Ngoc et al., 2019; Tu et al., 2020). This group of compounds has been evaluated for its ability to suppress numerous inflammatory cytokines and appears to inhibit production of IL-1β, NO and TNF-α by LPS-stimulated macrophages and microglia cells (Iqbal et al., 2013). For example extract of red ginseng reduces Aβ42-induced toxicity by suppressing TNF-α, IL-1β, NF-kB, and iNOS in BV-2 cells (Joo et al., 2008). Indeed, pretreatment of mouse macrophages with cyano-3,12 dioxooleana-1,9 dien-28-imidazolide (CDDO-Im) inhibits the phosphorylation of intrinsic bound inhibitor of NF-kB (IkBα) by IKK in response to TNF-α. This IkBα phosphorylation is the final step in NF-kB activation (Wardyn et al., 2015). A summary of terpenoids actions on various CNS diseases is displayed in Table 2.
Table 2.
Terpenoids as inhibitors of TNF-α and neuroinflammatory responses in neurodegenerative diseases
| Terpenoids | Source | Disease | Mechanism | IC50/dose | Model | Reference |
|---|---|---|---|---|---|---|
| Ginsenoside Rg3 | Panax ginseng | ND | Activates cAMP/MAPK & Trk-mediated neuritogenesis, and inhibits TNF-α, NF-kB, IL-1β, and iNOS | 0.1 mg/mL | BV-2 microglial cell line and Neuro-2a neuroblastoma cells | Joo et al., 2008 |
| Tanshinone I | Radix Salviae Miltiorrhizae | PD | Decreases NO, IL-6 and TNF, IL-1β and iNOS mRNA; ↓ GCSF | 10 mg/kg | Murine BV-2 microglial cells and mice | Wang et al., 2015a |
| Celastrol | Perennial creeping | PD | Decreases Dopaminergic neuron loss, TNF-α and NF-κB | 10 and 15 mg/kg | MPTP-induced PD in Swiss Webster mice | Cleren et al., 2005 |
| Paeoniflorin | Paeoniae alba Radix | PD | Decreases dopaminergic neuron loss, iNOS, IL-1β and TNF-α | 5mg/kg | ? MPTP-induced PD in C57BL/6 mice | Liu et al., 2006 |
AD: Alzheimer’s disease; ERK: extracellular signal-regulated kinase; iNOS: inducible nitric oxide synthase; ND: neurodegenerative disease; PD: Parkinson’s disease; TNFα: tumor necrosis factor alpha; TNFR: tumor necrosis factor alpha receptor.
Alkaloids
Similar to terpenoids, plant alkaloids are the second largest group of secondary plant metabolites. “Alkaloid” is a term often used to describe cyclic compounds containing one or more basic nitrogens. At low pH they are water soluble, but at high pH in the neutral form they are in the lipohilic. This property means they can pass readily through cell and tissue membranes, making them ideal drug candidates (Zou et al., 2017).
Huperzine A is a naturally occurring sesquiterpene alkaloid found as a potent reversible acetylcholinesterase inhibitor in the extracts of Huperzia serrata (Li and Shi, 2019). Huperzine A reduces cognitive defects in streptozotocin-induced diabetic rats by increasing the levels of BDNF, ChAT, SOD, catalase, and glutathione peroxidase while also inhibiting acetylcholinesterase, TNF-α, NF-κB, IL-1β, IL-6, CAT, MDA, and caspase-3 (Mao et al., 2014). In a similar way, the isoquinoline alkaloid berberine is the major component of Coptis chinensis. Its pharmacological effects include cholinergic-induced NGF, activity-mediated neurite outgrowth, BDNF secretion, and the inhibition of TNF-α, IL-1β, NF-κB, Cox2, and iNOS (Jia et al., 2012) (Table 3).
Table 3.
Alkaloids as inhibitors of TNF-α and neuroinflammatory responses in neurodegenerative diseases
| Alkaloids | Source | Disease | Mechanism | IC50/dose | Model | Reference |
|---|---|---|---|---|---|---|
| Piperine | Piper nigrum | PD | Diminishes IL-1β and TNF-α | 20 mg/kg, p.o. | 6-OHDA-induced rats | Rinwa and Kumar, 2017 |
| Berberine | Coptis chinensis | PD | Activates AKT/GSK-3β/Nrf2-mediated regulation, cholinergic activity-mediated neurite outgrowth, induces NGF and BDNF secretion, and inhibits Cox2, TNF-α, NF-κB, IL-1β, and iNOS levels | 0.1–10 nM | SH-SY5Y cells | Hsu et al., 2013 |
| Tetrandrine | Stephania tetrandra | AD | Increase in NF-κB DNA binding activity, the over-expression in IL-1β and TNF-α | 20, 30, and 40 mg/kg | Aat model of AD | He et al., 2011 |
| Berberine | Coptis chinensis | ND | Decreases the expression of the proinflammatory cytokines Cox-2, IL-1β, and TNF-α and markedly restores levels of BDNF and CREB and reduced the escape latency, induced neuronal impairment and memory dysfunction | 20 mg/kg | Rats with scopolamine-induced memory impairments | Lee et al., 2012 |
| Higenamine | Aconite tuber | ND | Inhibits the production of TNF-α, IL-6, ROS as well as NO and PGE2 (mediated by COX2) | 1–100 μM | LPS-activated BV2 cells | Yang et al., 2020 |
| suppresses NF-κB and activates Nrf2/HO-1 signaling pathways | ||||||
| Gelsemine | Gelsemium elegans Benth | AD | Inhibites the over-expression of proinflammatory cytokines, including IL-1β, IL-6, and TNF-α | 5–10 μg/kg | Rat model of AD | Chen et al., 2020 |
| Alleviates cognitive impairments induced by Aβ oligomer | ||||||
| Magnolol | Magnolia officinalis | Ischemic stroke | Attenuates IL-1β, TNF-α, IL-6 and ROS production and upregulates AKT and NF-κB levels | 10 and 30 mg/kg |
Rat global ischemic stroke model | Sacks et al., 2018 |
| Berberin | Coptis chinensis | AD | Activation of AKT/GSK-3 β/Nrf2-mediated regulation, cholinergic activity mediated neurite outgrowth, induces NGF and BDNF secretion, and inhibits COX2, TNF-α, NF-κB, IL-1β, and iNOS levels | 1,2.5,5,10 and µM 0.1–50 µM 100 mg/kg/d |
HeEK293 cell line, SH-SY5Y and microglia human neuroglioma H4 cells Raw 264.7 Transgenic mouse model |
Hirsch and Hunot, 2009; Bassani et al., 2015; Gopalakrishna and Alexander, 2015; Moon and Paek, 2015; Shahpiri et al., 2016 |
| Ginkgolide B | Ginkgo biloba | AD | Activates Trk/MAPK/ERK-mediated neurite outgrowth, induces NGF and BDNF secretion, reduces AChE, TNF-α, NF-κB, IL-1β, and MDA levels, and increases SOD, GSH-Px, Cat, Bcl-2, Bcl-xL, and TGF-β level | 1, 10 or 50 M 40 μM | PC12 cells and SH-SY5Y cells | Zhang et al., 2011; Li et al., 2013 |
| Berberine | Coptis chinensis | AD | Activates AKT/GSK-3b/Nrf2-mediated regulation, induces NGF and BDNF secretion, and inhibits COX-2, TNF-α, NF-kB, IL-1β, and iNOS levels | 20 mg/kg | Murine primary microglia cells and cultured BV2 microglia cells Rat | Jia et al., 2012; Lee et al., 2012 |
| Huperzine A | Huperzia serrata | AD | Activates Trk/MAPK/ERK-mediated neurite outgrowth, induces NGF and BDNF secretion, reduces AChE, TNF-α, NF-κB, IL-1β, and MDA levels, and increases SOD, GSH-Px, Cat, Bcl-2, Bcl-xL, and TGF-α level | 1 µM 0.1 mg/kg | NSCs and microglia Rat | Wang et al., 2010; Zhu et al., 2015 |
AChE: Acetylcholinesterase; AD: Alzheimer’s disease; ALS: amyotrophic lateral sclerosis; Aβ: amyloid-β peptide; BDNF: brain-derived neurotrophic factor; Cat: Catalase; CCL5: C-C Motif Chemokine Ligand 5; COX-2: cyclooxygenase-2; CREB: cAMP response element-binding protein; CX3CR1: C-X3-C Motif Chemokine Receptor 1; ERK: extracellular signal-regulated kinase; FADD: fas-associated death domain protein; GDNF: glial cell line-derived neurotrophic factor; GFAP: glial fibrillary acidic protein; GSH-Px: glutathione peroxidase; GSK-3: glycogen synthase kinase 3; HIF-a: hypoxia-inducible factor 1-alpha; HO-1: heme oxygenase-1; IKK: IκB kinase; iNOS: inducible nitric oxide synthase; JNK: c-Jun N-terminal kinases; JNKK1: c-Jun N-terminal kinase kinase 1; MEK: mitogen-activated protein kinase kinase; MEKK1: mitogen-activated protein kinase kinase kinase 1; MPTP: 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; MS: multiple sclerosis; mTORC1: mechanistic target of rapamycin complex 1; ND: neurodegenerative disease; NGF: nerve growth factor; Nrf2: nuclear factor erythroid 2 related factor 2; PD: Parkinson’s disease; PI3K: phosphoinositide 3-kinase; PKB: protein kinase B; ROS: reactive oxygen species; SOD: superoxide dismutase; TLR-2: Toll-like receptor-2; TNFR: tumor necrosis factor alpha receptor; TNFα: tumor necrosis factor alpha; TREM2: triggering receptor expressed on myeloid cells 2; TYROBP: tyrosine kinase-binding protein.
Lignans
Lignans are a group of plant metabolites frequently found in their free form in roots, stems, rhizomes, seeds, leaves and fruits and they may also be present as glycosides. Lignans have complex pharmacological profiles and share anti-inflammatory, antitumor, antioxidant, cardiovascular, immunosuppressive and antiviral properties (Barker, 2019). In previous studies, these compounds were found to inhibit the induction of TNF-α and NO synthesis in LPS-activated RAW 264.7 cells (Iqbal et al., 2013). In addition, lignans appear to suppress TNF-induced NF-kB activation by inhibiting IKKs (Tse et al., 2005) (Table 4). Like the flavonoids, lignans also have pharmacological activity in the μM range.
Table 4.
Lignans as inhibitors of TNF-α and neuroinflammatory responses in neurodegenerative diseases
| Lignans | Source | Disease | Mechanism | IC50/dose | Model | Reference |
|---|---|---|---|---|---|---|
| Schisandrin A | Schisandra chinesnesis | PD | Inhibits the production of NO, TNF-α and IL-6 | 10, 20 and 50 μM | Rat cortical cells | Song et al., 2016 |
| Schisandrin B (Gomisin N) | Schisandra chinesnesis | PD | Suppresses the expression of NO, TNF-α, PGE-2, IL-1β and IL-6 | 5, 10 or 20 mM | Rat primary neuron–microglia | Zeng et al., 2012 |
| Schisandrin B (Gomisin N) | Schisandra chinensis | PD | Decreases indoleamine 2,3 dioxygenase expression, and TNF-α, IL-1β and IL-6 | 10 mg/kg/d | Mice | Sun et al., 2014 |
| Honokiol, magnolol | Magnolia officinalis | AD | Induces NGF and BDNF secretion, inhibits TNF-α, NF-κB, IL-1β, IL-6, and ROS levels, and increases Akt activity | 100, 50, 10, and 1 μM | PC12 cells | Hoi et al., 2010 |
AD: Alzheimer’s disease; ERK: extracellular signal-regulated kinase; iNOS: inducible nitric oxide synthase; NGF: nerve growth factor; PD: Parkinson’s disease; ROS: reactive oxygen species; TNFα: tumor necrosis factor alpha; TNFR: tumor necrosis factor alpha receptor.
Phytochemicals for Neurodegenerative Diseases in Clinical Trials
Because of the many beneficial therapeutic effects observed in vitro and in vivo, recently, several phytochemicals aimed at treating neurodegenerative disorders have entered into phases of clinical trial - many of these are listed in Table 5. It has also been reported in a number of studies that TNF-α release in human whole blood and serum is blocked by some phytochemicals. Quercetin, a member of the flavonoid family, exerts immunomodulatory effects that might be useful in treating MS. In a study on peripheral blood mononuclear cells (PBMC) in 23 MS patients, quercetin led to a dose-dependent reduction in IL-1β and TNF-α (5–50 μM) (Sternberg et al., 2008). Luteolin, another flavonoid, has also been shown to have an immunomodulatory role that could be applicable in treating MS and other neurodegenerative diseases. In an experiment on PBMCs isolated from 14 relapsing-remitting MS patients, luteolin dose-dependently (0.2–50 μM) decreased PBMC proliferation and modulated concentrations of IL-1β and TNF-α in the culture supernatants released by PBMCs (Sternberg et al., 2009). The dietary phytosterol β-sitosterol has been shown to reduce TNF-α release by PBMCs of 11 MS patients at 4 μM. It also reduced the IL-5, IL-10, and IL-12 levels (Desai et al., 2009). Lemon verbena extracts contain the phenylpropanoid glycoside verbascoside, which has been reported to have antioxidant/anti-inflammatory effects in a number of pre-clinical and clinical trials. Mauriz et al. (2015) showed that verbascoside treatment (600 mg/day for 28 days) in 30 MS patients reduced serum concentrations of C reactive protein and inflammatory cytokines including IFN-γ, IL-4, IL-6, IL-10, IL-12, IL-23, TNF-α and TGF-β (Mauriz et al., 2015).
Table 5.
Phytochemicals for neurodegenerative diseases in clinical trials
| Phytochemical | Disease | Participant No. | Phase | Identifier |
|---|---|---|---|---|
| Curcumin | Alzheimer | 26 | Phase 2 | NCT01001637 |
| Curcumin | Multiple sclerosis | 80 | Phase 2 | NCT01514370 |
| Curcumin | Alzheimer | 30 | Phase 2 | NCT00164749 |
| Curcumin | Alzheimer | 33 | Phase 2 | NCT00099710 |
| Curcumin | Alzheimer | 12 | Phase 1 | NCT01716637 |
| Curcumin | Dementia (Alzheimer) | 1137 | Cohort | NCT02114372 |
| Curcumin | Multiple sclerosis | 41 | Phase 2 | NCT03150966 |
| Curcumin | Amyloidosis | 30 | Cohort | NCT03431896 |
| Berberine | Alzheimer | 90 | Cohort | NCT03221894 |
| Epigallocatechin-Gallate | Alzheimer | 21 | Phase 2 | NCT00951834 |
| EGCG | Multiple sclerosis | 60 | Phase 3 | NCT00799890 |
| EGCG | Parkinson | 480 | Phase 2 | NCT00461942 |
| Resveratrol | Alzheimer | 119 | Phase 2 | NCT01504854 |
| Resveratrol | Depression | 60 | Phase 4 | NCT03384329 |
| Resveratrol | Alzheimer | 27 | Phase 3 | NCT00678431 |
| Resveratrol | Parkinson | 39 | Phase 1 | NCT03094156 |
| Resveratrol | Parkinson | 40 | Phase 1 | NCT03093389 |
| Huperzine A | Alzheimer | 390 | Phase 2,3 | NCT01282619 |
| Caffeine alkaloid | Parkinson | 28 | Phase 2 | NCT01190735 |
| Vinpocetine | Stroke | 60 | Phase 2,3 | NCT02878772 |
Conclusions
In this review, we have discussed a number of phytochemicals that could potentially mitigate neuroinflammation caused by TNF-α in neurodegenerative diseases. These agents may prevent neuronal damage caused by the proinflammatory activity of TNF-α.
The exact pathophysiology of many neurodegenerative diseases is still not fully understood, partly because of the immense complexity of the human CNS, and also because good understanding of these complicated interactions requires large quantities of good quality data to be collected at all levels of experimental and clinical research. Determining the exact molecular mechanism of pathological subsystems of the CNS would clearly contribute to the agenda of developing appropriate targeted treatments.
The enormous diversity of plants offers plenty of untapped potential for identifying and developing novel drugs. Many natural agents such as terpenoids, alkaloids, flavonoids and lignans possess anti-TNF-α activity in vitro by suppressing upstream signaling at low micromolar concentrations comparable to conventional medicines such as adalimumab, infliximab and etanercept. But one of the key problems is that many phytochemicals, including curcumin, have poor CNS bioavailability as a result of digestion, liver enzyme degradation, poor absorption across intestinal membranes and across the BBB, and active CNS efflux mechanisms. In an attempt to address this problem, various drug delivery strategies may hold the key to better bioavailability through the use of liposomes, nanoparticles, the formation of conjugation molecules and complexes with phospholipids and amphiphilic polymers. Some success has been reported with resveratrol liposome in preventing oxidative damage and neuronal death in a rat model of PD, along with some restoration of motor control, by improving bioavalability compared to free resveratrol.
Inflammation is a complex process and a silver bullet for a universal drug target is extremely unlikely to be identified. But a combinatorial approach including natural product extracts alongside conventional medicine may offer part of the solution in future preventative and curative strategies.
Footnotes
Conflicts of interest: The authors declare no conflicts of interest.
C-Editors: Zhao M, Liu WJ, Qiu Y; T-Editor: Jia Y
References
- 1.Abbas M, Saeed F, Anjum FM, Afzaal M, Tufail T, Bashir MS, Ishtiaq A, Hussain S, Suleria HAR. Natural polyphenols: an overview. Int J Food Prop. 2017;20:1689–1699. [Google Scholar]
- 2.Alasmari F, Alshammari MA, Alasmari AF, Alanazi WA, Alhazzani K. Neuroinflammatory cytokines induce amyloid beta neurotoxicity through modulating amyloid precursor protein levels/metabolism. Biomed Res Int. 2018 doi: 10.1155/2018/3087475. doi: 101155/2018/3087475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Aminirad A, Mousavi SE, Fakhraei N, Mousavi SM, Rezayat SM. The role of nitric oxide in anticonvulsant effect of nanocurcumine on pentylenetetrazole-induced seizure in mice. Neurosci Lett. 2017;651:226–231. doi: 10.1016/j.neulet.2017.05.018. [DOI] [PubMed] [Google Scholar]
- 4.Anandhan A, Essa MM, Manivasagam T. Therapeutic attenuation of neuroinflammation and apoptosis by black tea theaflavin in chronic MPTP/probenecid model of Parkinson’s disease. Neurotox Res. 2013;23:166–173. doi: 10.1007/s12640-012-9332-9. [DOI] [PubMed] [Google Scholar]
- 5.Anastacio JR, Netto CA, Castro CC, Sanches EF, Ferreira DC, Noschang C, Krolow R, Dalmaz C, Pagnussat A. Resveratrol treatment has neuroprotective effects and prevents cognitive impairment after chronic cerebral hypoperfusion. Neurol Res. 2014;36:627–633. doi: 10.1179/1743132813Y.0000000293. [DOI] [PubMed] [Google Scholar]
- 6.Andrews JA, Jackson CE, Heiman-Patterson TD, Bettica P, Brooks BR, Pioro EP. Real-world evidence of riluzole effectiveness in treating amyotrophic lateral sclerosis. Amyotroph Lateral Scler Frontotemporal Degener. 2020;21:509–518. doi: 10.1080/21678421.2020.1771734. [DOI] [PubMed] [Google Scholar]
- 7.Angeline MS, Sarkar A, Anand K, Ambasta RK, Kumar P. Sesamol and naringenin reverse the effect of rotenone-induced PD rat model. Neuroscience. 2013;254:379–394. doi: 10.1016/j.neuroscience.2013.09.029. [DOI] [PubMed] [Google Scholar]
- 8.Anrather J, Iadecola C. Inflammation and stroke: an overview. Neurotherapeutics. 2016;13:661–670. doi: 10.1007/s13311-016-0483-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Arneth BM. Impact of B cells to the pathophysiology of multiple sclerosis. J Neuroinflammation. 2019;16:128. doi: 10.1186/s12974-019-1517-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bagheri H, Ghasemi F, Barreto GE, Rafiee R, Sathyapalan T, Sahebkar A. Effects of curcumin on mitochondria in neurodegenerative diseases. BioFactors. 2020;46:5–20. doi: 10.1002/biof.1566. [DOI] [PubMed] [Google Scholar]
- 11.Bando Y. Myelin morphology and axon pathology in demyelination during experimental autoimmune encephalomyelitis. Neural Regen Res. 2015;10:1584–1585. doi: 10.4103/1673-5374.165287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Barker D. Lignans. Molecules. 2019;24:1424. doi: 10.3390/molecules24071424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bashang H, Tamma S. The use of curcumin as an effective adjuvant to cancer therapy: a short review. Biotechnol Appl Biochem. 2020;67:171–179. doi: 10.1002/bab.1836. [DOI] [PubMed] [Google Scholar]
- 14.Bassani TB, Vital MA, Rauh LK. Neuroinflammation in the pathophysiology of Parkinson’s disease and therapeutic evidence of anti-inflammatory drugs. Arq Neuropsiquiatr. 2015;73:616–623. doi: 10.1590/0004-282X20150057. [DOI] [PubMed] [Google Scholar]
- 15.Bayat M, Azami Tameh A, Hossein Ghahremani M, Akbari M, Mehr SE, Khanavi M, Hassanzadeh G. Neuroprotective properties of Melissa officinalis after hypoxic-ischemic injury both in vitro and in vivo. Daru. 2012;20:42. doi: 10.1186/2008-2231-20-42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bi XL, Yang JY, Dong YX, Wang JM, Cui YH, Ikeshima T, Zhao YQ, Wu CF. Resveratrol inhibits nitric oxide and TNF-α production by lipopolysaccharide-activated microglia. Int Immunopharmacol. 2005;5:185–193. doi: 10.1016/j.intimp.2004.08.008. [DOI] [PubMed] [Google Scholar]
- 17.Bianconi V, Sahebkar A, Atkin SL, Pirro M. The regulation and importance of monocyte chemoattractant protein-1. Curr Opin Hematol. 2018;25:44–51. doi: 10.1097/MOH.0000000000000389. [DOI] [PubMed] [Google Scholar]
- 18.Boado RJ, Hui EK-W, Lu JZ, Zhou Q-H, Pardridge WM. Selective targeting of a TNFR decoy receptor pharmaceutical to the primate brain as a receptor-specific IgG fusion protein. J Biotechnol. 2010;146:84–91. doi: 10.1016/j.jbiotec.2010.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Brás JP, Bravo J, Freitas J, Barbosa MA, Santos SG, Summavielle T, Almeida MI. TNF-alpha-induced microglia activation requires miR-342: impact on NF-kB signaling and neurotoxicity. Cell Death Dis. 2020;11:415. doi: 10.1038/s41419-020-2626-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bureau G, Longpré F, Martinoli MG. Resveratrol and quercetin, two natural polyphenols, reduce apoptotic neuronal cell death induced by neuroinflammation. J Neurosci Res. 2008;86:403–410. doi: 10.1002/jnr.21503. [DOI] [PubMed] [Google Scholar]
- 21.Cacabelos R. How plausible is an Alzheimer’s disease vaccine. Expert Opin Drug Discov. 2020;15:1–6. doi: 10.1080/17460441.2019.1667329. [DOI] [PubMed] [Google Scholar]
- 22.Carrera I, Cacabelos R. Current drugs and potential future neuroprotective compounds for Parkinson’s disease. Curr Neuropharmacol. 2019;17:295–306. doi: 10.2174/1570159X17666181127125704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chan L, Chung CC, Chen JH, Yu RC, Hong CT. Cytokine profile in plasma extracellular vesicles of Parkinson’s disease and the association with cognitive function. Cells. 2021;10:604. doi: 10.3390/cells10030604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chang R, Yee KL, Sumbria RK. Tumor necrosis factor α Inhibition for Alzheimer’s disease. J Cent Nerv Syst Dis. 2017 doi: 10.1177/1179573517709278. doi: 101177/1179573517709278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Chen L, Pan H, Bai Y, Li H, Yang W, Lin ZX, Cui W, Xian YF. Gelsemine, a natural alkaloid extracted from Gelsemium elegans Benth. alleviates neuroinflammation and cognitive impairments in Aβ oligomer-treated mice. Psychopharmacology. 2020;237:2111–2124. doi: 10.1007/s00213-020-05522-y. [DOI] [PubMed] [Google Scholar]
- 26.Chen WW, Zhang X, Huang WJ. Role of neuroinflammation in neurodegenerative diseases. Mol Med Rep. 2016;13:3391–3396. doi: 10.3892/mmr.2016.4948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Cheng-Chung Wei J, Huang HC, Chen WJ, Huang CN, Peng CH, Lin CL. Epigallocatechin gallate attenuates amyloid beta-induced inflammation and neurotoxicity in EOC 13.31 microglia. Eur J Pharmacol. 2016;770:16–24. doi: 10.1016/j.ejphar.2015.11.048. [DOI] [PubMed] [Google Scholar]
- 28.Chung KO, Kim BY, Lee MH, Kim YR, Chung HY, Park JH, Moon JO. In-vitro and in-vivo anti-inflammatory effect of oxyresveratrol from Morus alba L. J Pharm Pharmacol. 2003;55:1695–1700. doi: 10.1211/0022357022313. [DOI] [PubMed] [Google Scholar]
- 29.Cleren C, Calingasan NY, Chen J, Beal MF. Celastrol protects against MPTP-and 3-nitropropionic acid-induced neurotoxicity. J Neurochem. 2005;94:995–1004. doi: 10.1111/j.1471-4159.2005.03253.x. [DOI] [PubMed] [Google Scholar]
- 30.Darvesh AS, Carroll RT, Bishayee A, Novotny NA, Geldenhuys WJ, Van der Schyf CJ. Curcumin and neurodegenerative diseases: a perspective. Expert Opin Investig Drugs. 2012;21:1123–1140. doi: 10.1517/13543784.2012.693479. [DOI] [PubMed] [Google Scholar]
- 31.de Alcântara GFT, Simões-Neto E, da Cruz GMP, Nobre MEP, Neves KRT, de Andrade GM, Brito GAdC, Viana GSdB. Curcumin reverses neurochemical, histological and immuno-histochemical alterations in the model of global brain ischemia. J Tradit Complement Med. 2016;7:14–23. doi: 10.1016/j.jtcme.2015.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.de Almeida MMA, Souza CDS, Dourado NS, da Silva AB, Ferreira RS, David JM, David JP, Costa MFD, da Silva VDA, Butt AM, Costa SL. Phytoestrogen agathisflavone ameliorates neuroinflammation-induced by LPS and IL-1β and protects neurons in cocultures of glia/neurons. Biomolecules. 2020;10:562. doi: 10.3390/biom10040562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.de Andrade Teles RB, Diniz TC, Pinto C, Coimbra T, de Oliveira Júnior RG, Gama e Silva M, de Lavor ÉM, Fernandes AWC, de Oliveira AP, de Almeida Ribeiro FPR. Flavonoids as therapeutic agents in Alzheimer’s and Parkinson’s diseases: a systematic review of preclinical evidences. Oxid Med Cell Longev. 2018 doi: 10.1155/2018/7043213. doi: 101155/2018/7043213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Desai F, Ramanathan M, Fink CS, Wilding GE, Weinstock-Guttman B, Awad AB. Comparison of the immunomodulatory effects of the plant sterol β-sitosterol to simvastatin in peripheral blood cells from multiple sclerosis patients. Int Immunopharmacol. 2009;9:153–157. doi: 10.1016/j.intimp.2008.10.019. [DOI] [PubMed] [Google Scholar]
- 35.Di Paolo M, Papi L, Gori F, Turillazzi E. Natural products in neurodegenerative diseases: a great promise but an ethical challenge. Int J Mol Sci. 2019;20:5170. doi: 10.3390/ijms20205170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Dong Y, Dekens DW, De Deyn PP, Naudé PJW, Eisel ULM. Targeting of tumor necrosis factor alpha receptors as a therapeutic strategy for neurodegenerative disorders. Antibodies. 2015;4:369–408. [Google Scholar]
- 37.Donzis EJ, Tronson NC. Modulation of learning and memory by cytokines: signaling mechanisms and long term consequences. Neurobiol Learn Mem. 2014;115:68–77. doi: 10.1016/j.nlm.2014.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ekert JO, Gould RL, Reynolds G, Howard RJ. TNF alpha inhibitors in Alzheimer’s disease: a systematic review. Int J Geriatr Psychiatry. 2018;33:688–694. doi: 10.1002/gps.4871. [DOI] [PubMed] [Google Scholar]
- 39.Farzaei MH, Shahpiri Z, Bahramsoltani R, Nia MM, Najafi F, Rahimi R. Efficacy and tolerability of phytomedicines in multiple sclerosis patients: a review. CNS Drugs. 2017;31:867–889. doi: 10.1007/s40263-017-0466-4. [DOI] [PubMed] [Google Scholar]
- 40.Filippi M, Bar-Or A, Piehl F, Preziosa P, Solari A, Vukusic S, Rocca MA. Multiple sclerosis. Nat Rev Dis Primers. 2018;4:43. doi: 10.1038/s41572-018-0041-4. [DOI] [PubMed] [Google Scholar]
- 41.Fu WY, Wang X, Ip NY. Targeting neuroinflammation as a therapeutic strategy for Alzheimer’s disease: mechanisms, drug candidates, and new opportunities. ACS Chem Neurosci. 2018;10:872–879. doi: 10.1021/acschemneuro.8b00402. [DOI] [PubMed] [Google Scholar]
- 42.Garcia G, Pais TF, Pinto P, Dobson G, McDougall GJ, Stewart D, Santos CN. Bioaccessible raspberry extracts enriched in ellagitannins and ellagic acid derivatives have anti-neuroinflammatory properties. Antioxidants (Basel) 2020;9:970. doi: 10.3390/antiox9100970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ghandadi M, Sahebkar A. Curcumin: an effective inhibitor of interleukin-6. Curr Pharm Des. 2017;23:921–931. doi: 10.2174/1381612822666161006151605. [DOI] [PubMed] [Google Scholar]
- 44.Ghasemi F, Shafiee M, Banikazemi Z, Pourhanifeh MH, Khanbabaei H, Shamshirian A, Amiri Moghadam S, ArefNezhad R, Sahebkar A, Avan A, Mirzaei H. Curcumin inhibits NF-kB and Wnt/β-catenin pathways in cervical cancer cells. Pathol Res Pract. 2019 doi: 10.1016/j.prp.2019.152556. doi: 101016/jprp2019152556. [DOI] [PubMed] [Google Scholar]
- 45.Gitler AD, Dhillon P, Shorter J. Neurodegenerative disease: models, mechanisms, and a new hope. Dis Model Mech. 2017;10:499–502. doi: 10.1242/dmm.030205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Golan H, Levav T, Mendelsohn A, Huleihel M. Involvement of tumor necrosis factor alpha in hippocampal development and function. Cereb Cortex. 2004;14:97–105. doi: 10.1093/cercor/bhg108. [DOI] [PubMed] [Google Scholar]
- 47.González R, Ballester I, López-Posadas R, Suárez M, Zarzuelo A, Martinez-Augustin O, Medina FSD. Effects of flavonoids and other polyphenols on inflammation. Crit Rev Food Sci Nutr. 2011;51:331–362. doi: 10.1080/10408390903584094. [DOI] [PubMed] [Google Scholar]
- 48.Gopalakrishna A, Alexander SA. Understanding Parkinson disease: a complex and multifaceted illness. J Neurosci Nurs. 2015;47:320–326. doi: 10.1097/JNN.0000000000000162. [DOI] [PubMed] [Google Scholar]
- 49.Guidotti G, Scarlata C, Brambilla L, Rossi D. Tumor necrosis factor alpha in amyotrophic lateral sclerosis: friend or foe. Cells. 2021;10:518. doi: 10.3390/cells10030518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ha SK, Moon E, Ju MS, Kim DH, Ryu JH, Oh MS, Kim SY. 6-Shogaol, a ginger product ,modulates neuroinflammation: a new approach to neuroprotection. Neuropharmacology. 2012;63:211–223. doi: 10.1016/j.neuropharm.2012.03.016. [DOI] [PubMed] [Google Scholar]
- 51.Hartl D, May P, Gu W, Mayhaus M, Pichler S, Spaniol C, Glaab E, Bobbili DR, Antony P, Koegelsberger S, Kurz A, Grimmer T, Morgan K, Vardarajan BN, Reitz C, Hardy J, Bras J, Guerreiro R, Balling R, Schneider JG, et al. A rare loss-of-function variant of ADAM17 is associated with late-onset familial Alzheimer disease. Mol Psychiatry. 2020;25:629–639. doi: 10.1038/s41380-018-0091-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.He FQ, Qiu BY, Zhang XH, Li TK, Xie Q, Cui DJ, Huang XL, Gan HT. Tetrandrine attenuates spatial memory impairment and hippocampal neuroinflammation via inhibiting NF-kappaB activation in a rat model of Alzheimer’s disease induced by amyloid-beta(1-42) Brain Res. 2011;1384:89–96. doi: 10.1016/j.brainres.2011.01.103. [DOI] [PubMed] [Google Scholar]
- 53.Hewlings SJ, Kalman DS. Curcumin: a review of its' effects on human health. Foods. 2017;6 doi: 10.3390/foods6100092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hirsch EC, Hunot S. Neuroinflammation in Parkinson’s disease: a target for neuroprotection. Lancet Neurol. 2009;8:382–397. doi: 10.1016/S1474-4422(09)70062-6. [DOI] [PubMed] [Google Scholar]
- 55.Hoi CP, Ho YP, Baum L, Chow AH. Neuroprotective effect of honokiol and magnolol, compounds from Magnolia officinalis ,on beta-amyloid-induced toxicity in PC12 cells. Phytother Res. 2010;24:1538–1542. doi: 10.1002/ptr.3178. [DOI] [PubMed] [Google Scholar]
- 56.Hoppe JB, Coradini K, Frozza RL, Oliveira CM, Meneghetti AB, Bernardi A, Pires ES, Beck RC, Salbego CG. Free and nanoencapsulated curcumin suppress beta-amyloid-induced cognitive impairments in rats: involvement of BDNF and Akt/GSK-3beta signaling pathway. Neurobiol Learn Mem. 2013;106:134–144. doi: 10.1016/j.nlm.2013.08.001. [DOI] [PubMed] [Google Scholar]
- 57.Hosseini SA, Mohammadi R, Noruzi S, Mohamadi Y, Azizian M, Mousavy SM, Ghasemi F, Hesari A, Sahebkar A, Salarinia R, Aghdam AM, Mirzaei H. Stem cell- and gene-based therapies as potential candidates in Alzheimer’s therapy. J Cell Biochem. 2018;119:8723–8736. doi: 10.1002/jcb.27202. [DOI] [PubMed] [Google Scholar]
- 58.Hsu YY, Tseng YT, Lo YC. Berberine, a natural antidiabetes drug, attenuates glucose neurotoxicity and promotes Nrf2-related neurite outgrowth. Toxicol Appl Pharmacol. 2013;272:787–796. doi: 10.1016/j.taap.2013.08.008. [DOI] [PubMed] [Google Scholar]
- 59.Huang HJ, Chen WL, Hsieh RH, Hsieh-Li HM. Multifunctional effects of mangosteen pericarp on cognition in C57BL/6J and triple transgenic Alzheimer’s mice. Evid Based Complement Alternat Med. 2014 doi: 10.1155/2014/813672. doi: 101155/2014/813672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Hussain G, Huang J, Rasul A, Anwar H, Imran A, Maqbool J, Razzaq A, Aziz N, Makhdoom EUH, Konuk M, Sun T. Putative roles of plant-derived tannins in neurodegenerative and neuropsychiatry disorders: an updated review. Molecules. 2019 doi: 10.3390/molecules24122213. doi: 103390/molecules24122213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Iqbal M, Verpoorte R, Korthout HA, Mustafa NR. Phytochemicals as a potential source for TNF-α inhibitors. Phytochem Rev. 2013;12:65–93. [Google Scholar]
- 62.Iranshahi M, Sahebkar A, Hosseini ST, Takasaki M, Konoshima T, Tokuda H. Cancer chemopreventive activity of diversin from Ferula diversivittata in vitro and in vivo. Phytomedicine. 2010;17:269–273. doi: 10.1016/j.phymed.2009.05.020. [DOI] [PubMed] [Google Scholar]
- 63.Islam MT. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol Res. 2017;39:73–82. doi: 10.1080/01616412.2016.1251711. [DOI] [PubMed] [Google Scholar]
- 64.Jia L, Liu J, Song Z, Pan X, Chen L, Cui X, Wang M. Berberine suppresses amyloid-beta-induced inflammatory response in microglia by inhibiting nuclear factor-kappaB and mitogen-activated protein kinase signalling pathways. J Pharm Pharmacol. 2012;64:1510–1521. doi: 10.1111/j.2042-7158.2012.01529.x. [DOI] [PubMed] [Google Scholar]
- 65.Jin F, Wu Q, Lu YF, Gong QH, Shi JS. Neuroprotective effect of resveratrol on 6-OHDA-induced Parkinson’s disease in rats. Eur J Pharmacol. 2008;600:78–82. doi: 10.1016/j.ejphar.2008.10.005. [DOI] [PubMed] [Google Scholar]
- 66.Joo SS, Yoo YM, Ahn BW, Nam SY, Kim YB, Hwang KW, Lee DI. Prevention of inflammation-mediated neurotoxicity by Rg3 and its role in microglial activation. Biol Pharm Bull. 2008;31:1392–1396. doi: 10.1248/bpb.31.1392. [DOI] [PubMed] [Google Scholar]
- 67.Jung UJ, Jeon MT, Choi MS, Kim SR. Silibinin attenuates MPP+-induced neurotoxicity in the substantia nigra in vivo. J Med Food. 2014;17:599–605. doi: 10.1089/jmf.2013.2926. [DOI] [PubMed] [Google Scholar]
- 68.Kamali AN, Zian Z, Bautista JM, Hamedifar H, Hossein-Khannazer N, Hosseinzadeh R, Yazdani R, Azizi G. The potential role of pro-inflammatory and anti-inflammatory cytokines in epilepsy pathogenesis. Endocr Metab Immune Disord Drug Targets. 2020 doi: 10.2174/1871530320999201116200940. doi: 102174/1871530320999201116200940. [DOI] [PubMed] [Google Scholar]
- 69.Kamaşak T, Dilber B, Yaman S, Durgut BD, Kurt T, Çoban E, Arslan EA, Şahin S, Karahan SC, Cansu A. HMGB-1, TLR4, IL-1R1 ,TNF-α ,and IL-1β: novel epilepsy markers. Epileptic Disord. 2020;22:183–193. doi: 10.1684/epd.2020.1155. [DOI] [PubMed] [Google Scholar]
- 70.Kang X, Ploner A, Pedersen NL, Bandres-Ciga S, Noyce AJ, Wirdefeldt K, Williams DM. Tumor necrosis factor inhibition and parkinson disease: a mendelian randomization study. Neurology. 2021;96:e1672–1679. doi: 10.1212/WNL.0000000000011630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Kaur H, Patro I, Tikoo K, Sandhir R. Curcumin attenuates inflammatory response and cognitive deficits in experimental model of chronic epilepsy. Neurochem Int. 2015;89:40–50. doi: 10.1016/j.neuint.2015.07.009. [DOI] [PubMed] [Google Scholar]
- 72.Khan MM, Raza SS, Javed H, Ahmad A, Khan A, Islam F, Safhi MM, Islam F. Rutin protects dopaminergic neurons from oxidative stress in an animal model of Parkinson’s disease. Neurotox Res. 2012;22:1–15. doi: 10.1007/s12640-011-9295-2. [DOI] [PubMed] [Google Scholar]
- 73.Kim HD, Jeong KH, Jung UJ, Kim SR. Naringin treatment induces neuroprotective effects in a mouse model of Parkinson’s disease in vivo, but not enough to restore the lesioned dopaminergic system. J Nutr Biochem. 2016;28:140–146. doi: 10.1016/j.jnutbio.2015.10.013. [DOI] [PubMed] [Google Scholar]
- 74.Kinney JW, Bemiller SM, Murtishaw AS, Leisgang AM, Salazar AM, Lamb BT. Inflammation as a central mechanism in Alzheimer’s disease. Alzheimers Dement (N Y) 2018;4:575–590. doi: 10.1016/j.trci.2018.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Kumar A, Singh A. A review on Alzheimer’s disease pathophysiology and its management: an update. Pharmacol Rep. 2015;67:195–203. doi: 10.1016/j.pharep.2014.09.004. [DOI] [PubMed] [Google Scholar]
- 76.Kumar A, Takada Y, Boriek AM, Aggarwal BB. Nuclear factor-κB: its role in health and disease. J Mol Med. 2004;82:434–448. doi: 10.1007/s00109-004-0555-y. [DOI] [PubMed] [Google Scholar]
- 77.Kunnumakkara AB, Thakur KK, Rana V, Bora B, Banik K, Khatoon E, Sailo BL, Shabnam B, Girisa S, Gupta SC, Aggarwal BB. Upside and downside of tumor necrosis factor blockers for treatment of immune/inflammatory diseases. Crit Rev Immunol. 2019;39:439–479. doi: 10.1615/CritRevImmunol.2020033205. [DOI] [PubMed] [Google Scholar]
- 78.Kurz A, Perneczky R. Novel insights for the treatment of Alzheimer’s disease. Prog Neuropsychopharmacol Biol Psychiatry. 2011;35:373–379. doi: 10.1016/j.pnpbp.2010.07.018. [DOI] [PubMed] [Google Scholar]
- 79.Lee B, Sur B, Shim I, Lee H, Hahm DH. Phellodendron amurense and its major alkaloid compound, berberine ameliorates scopolamine-induced neuronal impairment and memory dysfunction in rats. Korean J Physiol Pharmacol. 2012;16:79–89. doi: 10.4196/kjpp.2012.16.2.79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Lee EH, Lim SS, Yuen KH, Lee CY. Curcumin and a hemi-analogue with improved blood-brain barrier permeability protect against amyloid-beta toxicity in Caenorhabditis elegans via SKN-1/Nrf activation. J Pharm Pharmacol. 2019;71:860–868. doi: 10.1111/jphp.13052. [DOI] [PubMed] [Google Scholar]
- 81.Lee JS, Choi HS, Kang SW, Chung JH, Park HK, Ban JY, Kwon OY, Hong HP, Ko YG. Therapeutic effect of Korean red ginseng on inflammatory cytokines in rats with focal cerebral ischemia/reperfusion injury. Am J Chin Med. 2011;39:83–94. doi: 10.1142/S0192415X1100866X. [DOI] [PubMed] [Google Scholar]
- 82.Leem E, Nam JH, Jeon MT, Shin WH, Won SY, Park SJ, Choi MS, Jin BK, Jung UJ, Kim SR. Naringin protects the nigrostriatal dopaminergic projection through induction of GDNF in a neurotoxin model of Parkinson’s disease. J Nutr Biochem. 2014;25:801–806. doi: 10.1016/j.jnutbio.2014.03.006. [DOI] [PubMed] [Google Scholar]
- 83.Li C, Shi S. Neuroprotective effect of huperzine A on D-galactose-induced hearing dysfunction. Ear Nose Throat J. 2019;100:269S–276S. doi: 10.1177/0145561319864570. [DOI] [PubMed] [Google Scholar]
- 84.Li H, Hu J, Ma L, Yuan Z, Wang Y, Wang X, Xing D, Lei F, Du L. Comprehensive study of baicalin down-regulating NOD2 receptor expression of neurons with oxygen-glucose deprivation in vitro and cerebral ischemia-reperfusion in vivo. Eur J Pharmacol. 2010;649:92–99. doi: 10.1016/j.ejphar.2010.09.023. [DOI] [PubMed] [Google Scholar]
- 85.Li J, Zhao R, Jiang Y, Xu Y, Zhao H, Lyu X, Wu T. Bilberry anthocyanins improve neuroinflammation and cognitive dysfunction in APP/PSEN1 mice via the CD33/TREM2/TYROBP signaling pathway in microglia. Food Funct. 2020;11:1572–1584. doi: 10.1039/c9fo02103e. [DOI] [PubMed] [Google Scholar]
- 86.Li L, Zhang QG, Lai LY, Wen XJ, Zheng T, Cheung CW, Zhou SQ, Xu SY. Neuroprotective effect of ginkgolide B on bupivacaine-induced apoptosis in SH-SY5Y cells. Oxid Med Cell Longev. 2013 doi: 10.1155/2013/159864. doi: 101155/2013/159864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Li R, Huang YG, Fang D, Le WD. (−)-Epigallocatechin gallate inhibits lipopolysaccharide-induced microglial activation and protects against inflammation-mediated dopaminergic neuronal injury. J Neurosci Res. 2004;78:723–731. doi: 10.1002/jnr.20315. [DOI] [PubMed] [Google Scholar]
- 88.Liu D, Wang Z, Gao Z, Xie K, Zhang Q, Jiang H, Pang Q. Effects of curcumin on learning and memory deficits, BDNF, and ERK protein expression in rats exposed to chronic unpredictable stress. Behav Brain Res. 2014;271:116–121. doi: 10.1016/j.bbr.2014.05.068. [DOI] [PubMed] [Google Scholar]
- 89.Liu HQ, Zhang WY, Luo XT, Ye Y, Zhu XZ. Paeoniflorin attenuates neuroinflammation and dopaminergic neurodegeneration in the MPTP model of Parkinson’s disease by activation of adenosine A1 receptor. Br J Pharmacol. 2006;148:314–325. doi: 10.1038/sj.bjp.0706732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Lopez MS, Dempsey RJ, Vemuganti R. Resveratrol neuroprotection in stroke and traumatic CNS injury. Neurochem Int. 2015;89:75–82. doi: 10.1016/j.neuint.2015.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Ma T, Tan MS, Yu JT, Tan L. Resveratrol as a therapeutic agent for Alzheimer’s disease. Biomed Res Int. 2014 doi: 10.1155/2014/350516. doi: 101155/2014/350516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Mansoor SR, Hashemian M, Khalili-Fomeshi M, Ashrafpour M, Moghadamnia AA, Ghasemi-Kasman M. Upregulation of klotho and erythropoietin contributes to the neuroprotection induced by curcumin-loaded nanoparticles in experimental model of chronic epilepsy. Brain Res Bull. 2018;142:281–288. doi: 10.1016/j.brainresbull.2018.08.010. [DOI] [PubMed] [Google Scholar]
- 93.Mao XY, Cao DF, Li X, Yin JY, Wang ZB, Zhang Y, Mao CX, Zhou HH, Liu ZQ. Huperzine A ameliorates cognitive deficits in streptozotocin-induced diabetic rats. Int J Mol Sci. 2014;15:7667–7683. doi: 10.3390/ijms15057667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Marotta G, Basagni F, Rosini M, Minarini A. Memantine derivatives as multitarget agents in Alzheimer’s disease. Molecules. 2020;25:4005. doi: 10.3390/molecules25174005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Mauriz E, Vallejo D, Tuñón MJ, Rodriguez-López JM, Rodríguez-Pérez R, Sanz-Gómez J, del Camino García-Fernández M. Effects of dietary supplementation with lemon verbena extracts on serum inflammatory markers of multiple sclerosis patients. Nutr Hosp. 2015;31:764–771. doi: 10.3305/nh.2015.31.2.8319. [DOI] [PubMed] [Google Scholar]
- 96.McCaulley ME, Grush KA. Alzheimer’s disease: exploring the role of inflammation and implications for treatment. Int J Alzheimers Dis. 2015 doi: 10.1155/2015/515248. doi: 101155/2015/515248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Meng T, Xiao D, Muhammed A, Deng J, Chen L, He J. Anti-inflammatory action and mechanisms of resveratrol. Molecules. 2021;26:229. doi: 10.3390/molecules26010229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Moawad H, Shepard KM, Satya-Murti S. Complementary and alternative medicine in multiple sclerosis: Payment policy perspectives. Neurol Clin Pract. 2015;5:74–79. doi: 10.1212/CPJ.0000000000000020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Mogi M, Harada M, Riederer P, Narabayashi H, Fujita K, Nagatsu T. Tumor necrosis factor-α (TNF-α) increases both in the brain and in the cerebrospinal fluid from parkinsonian patients. Neurosci Lett. 1994;165:208–210. doi: 10.1016/0304-3940(94)90746-3. [DOI] [PubMed] [Google Scholar]
- 100.Mollazadeh H, Cicero AFG, Blesso CN, Pirro M, Majeed M, Sahebkar A. Immune modulation by curcumin: the role of interleukin-10. Crit Rev Food Sci Nutr. 2019;59:89–101. doi: 10.1080/10408398.2017.1358139. [DOI] [PubMed] [Google Scholar]
- 101.Moon HE, Paek SH. Mitochondrial dysfunction in Parkinson’s disease. Exp Neurobiol. 2015;24:103–116. doi: 10.5607/en.2015.24.2.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Moriwaki K, Bertin J, Gough P, Orlowski G, Chan FK. Differential roles of RIPK1 and RIPK3 in TNF-induced necroptosis and chemotherapeutic agent-induced cell death. Cell Death Dis. 2015;6:e1636. doi: 10.1038/cddis.2015.16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Nabavi SF, Daglia M, D’Antona G, Sobarzo-Sanchez E, Talas ZS, Nabavi SM. Natural compounds used as therapies targeting to amyotrophic lateral sclerosis. Curr Pharm Biotechnol. 2015;16:211–218. doi: 10.2174/1389201016666150118132224. [DOI] [PubMed] [Google Scholar]
- 104.Nakka VP, Prakash-babu P, Vemuganti R. Crosstalk between endoplasmic reticulum stress, oxidative stress, and autophagy: potential therapeutic targets for acute CNS injuries. Mol Neurobiol. 2016;53:532–544. doi: 10.1007/s12035-014-9029-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Nam SM, Choi JH, Yoo DY, Kim W, Jung HY, Kim JW, Yoo M, Lee S, Kim CJ, Yoon YS, Hwang IK. Effects of curcumin (Curcuma longa) on learning and spatial memory as well as cell proliferation and neuroblast differentiation in adult and aged mice by upregulating brain-derived neurotrophic factor and CREB signaling. J Med Food. 2014;17:641–649. doi: 10.1089/jmf.2013.2965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Naoi M, Shamoto-Nagai M, Maruyama W. Neuroprotection of multifunctional phytochemicals as novel therapeutic strategy for neurodegenerative disorders: Antiapoptotic and antiamyloidogenic activities by modulation of cellular signal pathways. Future Neurol. 2019 doi:102217/fnl-2018-0028. [Google Scholar]
- 107.Nguyen Ngoc H, Alilou M, Stonig M, Nghiem DT, Kim LT, Gostner JM, Stuppner H, Ganzera M. Terpenoids from the stems of fissistigma polyanthoides and their anti-inflammatory activity. J Nat Prod. 2019;82:2941–2952. doi: 10.1021/acs.jnatprod.9b00208. [DOI] [PubMed] [Google Scholar]
- 108.Ojha RP, Rastogi M, Devi BP, Agrawal A, Dubey G. Neuroprotective effect of curcuminoids against inflammation-mediated dopaminergic neurodegeneration in the MPTP model of Parkinson’s disease. J Neuroimmune Pharmacol. 2012;7:609–618. doi: 10.1007/s11481-012-9363-2. [DOI] [PubMed] [Google Scholar]
- 109.Ojha S, Javed H, Azimullah S, Khair SBA, Haque ME. Neuroprotective potential of ferulic acid in the rotenone model of Parkinson’s disease. Drug Des Devel Ther. 2015;9:5499–5510. doi: 10.2147/DDDT.S90616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Panahi Y, Ahmadi Y, Teymouri M, Johnston TP, Sahebkar A. Curcumin as a potential candidate for treating hyperlipidemia: a review of cellular and metabolic mechanisms. J Cell Physiol. 2018a;233:141–152. doi: 10.1002/jcp.25756. [DOI] [PubMed] [Google Scholar]
- 111.Panahi Y, Khalili N, Sahebi E, Namazi S, Simental-Mendía LE, Majeed M, Sahebkar A. Effects of curcuminoids plus piperine on glycemic, hepatic and inflammatory biomarkers in patients with type 2 diabetes mellitus: a randomized double-blind placebo-controlled trial. Drug Res (Stuttg) 2018b;68:403–409. doi: 10.1055/s-0044-101752. [DOI] [PubMed] [Google Scholar]
- 112.Patil SP, Jain PD, Sancheti JS, Ghumatkar PJ, Tambe R, Sathaye S. Neuroprotective and neurotrophic effects of Apigenin and Luteolin in MPTP induced parkinsonism in mice. Neuropharmacology. 2014;86:192–202. doi: 10.1016/j.neuropharm.2014.07.012. [DOI] [PubMed] [Google Scholar]
- 113.Pegoretti V, Baron W, Laman JD, Eisel ULM. Selective modulation of TNF–TNFRs signaling: insights for multiple sclerosis treatment. Front Immunol. 2018;9:925. doi: 10.3389/fimmu.2018.00925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Perry RT, Collins JS, Wiener H, Acton R, Go RC. The role of TNF and its receptors in Alzheimer’s disease. Neurobiol Aging. 2001;22:873–883. doi: 10.1016/s0197-4580(01)00291-3. [DOI] [PubMed] [Google Scholar]
- 115.Prasanna P, Upadhyay A. Flavonoid-based nanomedicines in Alzheimer’s disease therapeutics: promises made, a long way to go. ACS Pharmacol Transl Sci. 2021;4:74–95. doi: 10.1021/acsptsci.0c00224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Qi G, Mi Y, Fan R, Li R, Liu Z, Liu X. Nobiletin protects against systemic inflammation-stimulated memory impairment via MAPK and NF-κB signaling pathways. J Agric Food Chem. 2019;67:5122–5134. doi: 10.1021/acs.jafc.9b00133. [DOI] [PubMed] [Google Scholar]
- 117.Qureshi M, Al-Suhaimi EA, Wahid F, Shehzad O, Shehzad A. Therapeutic potential of curcumin for multiple sclerosis. Neurol Sci. 2018;39:207–214. doi: 10.1007/s10072-017-3149-5. [DOI] [PubMed] [Google Scholar]
- 118.Reddy PH, Manczak M, Yin X, Grady MC, Mitchell A, Tonk S, Kuruva CS, Bhatti JS, Kandimalla R, Vijayan M, Kumar S, Wang R, Pradeepkiran JA, Ogunmokun G, Thamarai K, Quesada K, Boles A, Reddy AP. Protective effects of indian spice curcumin against amyloid-β in Alzheimer’s disease. J Alzheimers Dis. 2018;61:843–866. doi: 10.3233/JAD-170512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Rekha KR, Selvakumar GP, Sivakamasundari RI. Effects of syringic acid on chronic MPTP/probenecid induced motor dysfunction, dopaminergic markers expression and neuroinflammation in C57BL/6 mice. Biomed Aging Pathol. 2014;4:95–104. [Google Scholar]
- 120.Rinwa P, Kumar A. Quercetin along with piperine prevents cognitive dysfunction, oxidative stress and neuro-inflammation associated with mouse model of chronic unpredictable stress. Arch Pharm Res. 2017;40:1166–1175. doi: 10.1007/s12272-013-0205-4. [DOI] [PubMed] [Google Scholar]
- 121.Rinwa P, Kumar A, Garg S. Suppression of neuroinflammatory and apoptotic signaling cascade by curcumin alone and in combination with piperine in rat model of olfactory bulbectomy induced depression. PLoS One. 2013;8:e61052. doi: 10.1371/journal.pone.0061052. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 122.Rodríguez-Gómez JA, Kavanagh E, Engskog-Vlachos P, Engskog MKR, Herrera AJ, Espinosa-Oliva AM, Joseph B, Hajji N, Venero JL, Burguillos MA. Microglia: agents of the CNS pro-inflammatory response. Cells. 2020;9:1717. doi: 10.3390/cells9071717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Rogers J, Lue LF. Microglial chemotaxis, activation, and phagocytosis of amyloid β-peptide as linked phenomena in Alzheimer’s disease. Neurochem Int. 2001;39:333–340. doi: 10.1016/s0197-0186(01)00040-7. [DOI] [PubMed] [Google Scholar]
- 124.Sacks D, Baxter B, Campbell BCV, Carpenter JS, Cognard C, Dippel D, Eesa M, Fischer U, Hausegger K, Hirsch JA, Shazam Hussain M, Jansen O, Jayaraman MV, Khalessi AA, Kluck BW, Lavine S, Meyers PM, Ramee S, Rüfenacht DA, Schirmer CM, et al. Multisociety consensus quality improvement revised consensus statement for endovascular therapy of acute ischemic stroke. Int J Stroke. 2018;13:612–632. doi: 10.1177/1747493018778713. [DOI] [PubMed] [Google Scholar]
- 125.Sahebkar A. Molecular mechanisms for curcumin benefits against ischemic injury. Fertil Steril. 2010;94:e75–76. doi: 10.1016/j.fertnstert.2010.07.1071. [DOI] [PubMed] [Google Scholar]
- 126.Shahpiri Z, Bahramsoltani R, Farzaei MH, Farzaei F, Rahimi R. Phytochemicals as future drugs for Parkinson’s disease: a comprehensive review. Rev Neurosci. 2016;27:651–668. doi: 10.1515/revneuro-2016-0004. [DOI] [PubMed] [Google Scholar]
- 127.Shal B, Ding W, Ali H, Kim YS, Khan S. Anti-neuroinflammatory potential of natural products in attenuation of Alzheimer’s disease. Front Pharmacol. 2018;9:548. doi: 10.3389/fphar.2018.00548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Shamim D, Laskowski M. Inhibition of inflammation mediated through the tumor necrosis factor α biochemical pathway can lead to favorable outcomes in Alzheimer disease. J Cent Nerv Syst Dis. 2017 doi: 10.1177/1179573517722512. doi: 101177/1179573517722512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Shao AW, Wu HJ, Chen S, Ammar AB, Zhang JM, Hong Y. Resveratrol attenuates early brain injury after subarachnoid hemorrhage through inhibition of NF-κB-dependent inflammatory/MMP-9 pathway. CNS Neurosci Ther. 2014;20:182–185. doi: 10.1111/cns.12194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Shi JH, Sun SC. Tumor necrosis factor receptor-associated factor regulation of nuclear factor κb and mitogen-activated protein kinase pathways. Front Immunol. 2018;9:1849. doi: 10.3389/fimmu.2018.01849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Shin JA, Lee H, Lim Y-K, Koh Y, Choi JH, Park E-M. Therapeutic effects of resveratrol during acute periods following experimental ischemic stroke. J Neuroimmunol. 2010;227:93–100. doi: 10.1016/j.jneuroim.2010.06.017. [DOI] [PubMed] [Google Scholar]
- 132.Siebert S, Tsoukas A, Robertson J, McInnes I. Cytokines as therapeutic targets in rheumatoid arthritis and other inflammatory diseases. Pharmacol Rev. 2015;67:280–309. doi: 10.1124/pr.114.009639. [DOI] [PubMed] [Google Scholar]
- 133.Song F, Zeng K, Liao L, Yu Q, Tu P, Wang X. Schizandrin A inhibits microglia-mediated neuroninflammation through inhibiting TRAF6-NF-κB and Jak2-Stat3 signaling pathways. PLoS One. 2016;11:e0149991. doi: 10.1371/journal.pone.0149991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Song J, Cheon SY, Jung W, Lee WT, Lee JE. Resveratrol induces the expression of interleukin-10 and brain-derived neurotrophic factor in BV2 microglia under hypoxia. Int J Mol Sci. 2014;15:15512–15529. doi: 10.3390/ijms150915512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Spagnuolo C, Moccia S, Russo GL. Anti-inflammatory effects of flavonoids in neurodegenerative disorders. Eur J Med Chem. 2018;153:105–115. doi: 10.1016/j.ejmech.2017.09.001. [DOI] [PubMed] [Google Scholar]
- 136.Sternberg Z, Chadha K, Lieberman A, Drake A, Hojnacki D, Weinstock-Guttman B, Munschauer F. Immunomodulatory responses of peripheral blood mononuclear cells from multiple sclerosis patients upon in vitro incubation with the flavonoid luteolin: additive effects of IFN-â. J Neuroinflammation. 2009;6:28. doi: 10.1186/1742-2094-6-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Sternberg Z, Chadha K, Lieberman A, Hojnacki D, Drake A, Zamboni P, Rocco P, Grazioli E, Weinstock-Guttman B, Munschauer F. Quercetin and interferon-β modulate immune response (s) in peripheral blood mononuclear cells isolated from multiple sclerosis patients. J Neuroimmunol. 2008;205:142–147. doi: 10.1016/j.jneuroim.2008.09.008. [DOI] [PubMed] [Google Scholar]
- 138.Sun YX, Cong YL, Liu Y, Jin B, Si L, Wang AB, Cai H, Che GY, Tang B, Wang CF. Schisandrin A and B affect subventricular zone neurogenesis in mouse. Eur J Pharmacol. 2014;740:552–559. doi: 10.1016/j.ejphar.2014.06.032. [DOI] [PubMed] [Google Scholar]
- 139.Teymouri M, Pirro M, Johnston TP, Sahebkar A. Curcumin as a multifaceted compound against human papilloma virus infection and cervical cancers: a review of chemistry, cellular, molecular, and preclinical features. Biofactors. 2017;43:331–346. doi: 10.1002/biof.1344. [DOI] [PubMed] [Google Scholar]
- 140.Tortarolo M, Lo Coco D, Veglianese P, Vallarola A, Giordana MT, Marcon G, Beghi E, Poloni M, Strong MJ, Iyer AM, Aronica E, Bendotti C. Amyotrophic lateral sclerosis, a multisystem pathology: insights into the role of TNFalpha. Mediators Inflamm. 2017 doi: 10.1155/2017/2985051. doi: 101155/2017/2985051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Tse A, Wan CK, Shen XL, Yang M, Fong WF. Honokiol inhibits TNF-a-stimulated NF-kB activation and NF-kB-regulated gene expression through suppression of IKK activation. Biochem Pharmacol. 2005;70:1443–1457. doi: 10.1016/j.bcp.2005.08.011. [DOI] [PubMed] [Google Scholar]
- 142.Tu PC, Liang YC, Huang GJ, Lin MK, Kao MC, Lu TL, Sung PJ, Kuo YH. Cytotoxic and anti-inflammatory terpenoids from the whole plant of vaccinium emarginatum. Planta Med. 2020;86:1313–1322. doi: 10.1055/a-1192-6225. [DOI] [PubMed] [Google Scholar]
- 143.Tu XK, Yang WZ, Shi SS, Chen Y, Wang CH, Chen CM, Chen Z. Baicalin inhibits TLR2/4 signaling pathway in rat brain following permanent cerebral ischemia. Inflammation. 2011;34:463–470. doi: 10.1007/s10753-010-9254-8. [DOI] [PubMed] [Google Scholar]
- 144.Vafeiadou K, Vauzour D, Lee HY, Rodriguez-Mateos A, Williams RJ, Spencer JP. The citrus flavanone naringenin inhibits inflammatory signalling in glial cells and protects against neuroinflammatory injury. Arch Biochem Biophys. 2009;484:100–109. doi: 10.1016/j.abb.2009.01.016. [DOI] [PubMed] [Google Scholar]
- 145.Velmurugan BK, Rathinasamy B, Lohanathan BP, Thiyagarajan V, Weng CF. Neuroprotective role of phytochemicals. Molecules. 2018;23:2485. doi: 10.3390/molecules23102485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Vu LT, Bowser R. Fluid-based biomarkers for amyotrophic lateral sclerosis. Neurotherapeutics. 2017;14:119–134. doi: 10.1007/s13311-016-0503-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Wang CP, Zhang LZ, Li GC, Shi YW, Li JL, Zhang XC, Wang ZW, Ding F, Liang XM. Mulberroside A protects against ischemic impairment in primary culture of rat cortical neurons after oxygen-glucose deprivation followed by reperfusion. J Neurosci Res. 2014;92:944–954. doi: 10.1002/jnr.23374. [DOI] [PubMed] [Google Scholar]
- 148.Wang J, Zhang HY, Tang XC. Huperzine a improves chronic inflammation and cognitive decline in rats with cerebral hypoperfusion. J Neurosci Res. 2010;88:807–815. doi: 10.1002/jnr.22237. [DOI] [PubMed] [Google Scholar]
- 149.Wang J, Song Y, Chen Z, Leng SX. Connection between systemic inflammation and neuroinflammation underlies neuroprotective mechanism of several phytochemicals in neurodegenerative diseases. Oxid Med Cell Longev. 2018a doi: 10.1155/2018/1972714. doi: 101155/2018/1972714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Wang J, Xiong T, Zhou J, He H, Wu D, Du X, Li X, Xu B. Enzymatic formation of curcumin in vitro and in vivo. Nano Res. 2018b;11:3453–3461. [Google Scholar]
- 151.Wang K, Song F, Fernandez-Escobar A, Luo G, Wang JH, Sun Y. The properties of cytokines in multiple sclerosis: pros and cons. Am J Med Sci. 2018c;356:552–560. doi: 10.1016/j.amjms.2018.08.018. [DOI] [PubMed] [Google Scholar]
- 152.Wang L, Zhao H, Wang L, Tao Y, Du G, Guan W, Liu J, Brennan C, Ho CT, Li S. Effects of selected resveratrol analogues on activation and polarization of lipopolysaccharide-stimulated BV-2 microglial cells. J Agric Food Chem. 2020;68:3750–3757. doi: 10.1021/acs.jafc.0c00498. [DOI] [PubMed] [Google Scholar]
- 153.Wang S, Jing H, Yang H, Liu Z, Guo H, Chai L, Hu L. Tanshinone I selectively suppresses pro-inflammatory genes expression in activated microglia and prevents nigrostriatal dopaminergic neurodegeneration in a mouse model of Parkinson’s disease. J Ethnopharmacol. 2015a;164:247–255. doi: 10.1016/j.jep.2015.01.042. [DOI] [PubMed] [Google Scholar]
- 154.Wang WY, Tan MS, Yu JT, Tan L. Role of pro-inflammatory cytokines released from microglia in Alzheimer’s disease. Ann Transl Med. 2015b;3:136. doi: 10.3978/j.issn.2305-5839.2015.03.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Wardyn JD, Ponsford AH, Sanderson CM. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem Soc Trans. 2015;43:621–626. doi: 10.1042/BST20150014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Wei JCC, Huang HC, Chen WJ, Huang CN, Peng CH, Lin CL. Epigallocatechin gallate attenuates amyloid β-induced inflammation and neurotoxicity in EOC 13.31 microglia. Eur J Pharmacol. 2016;770:16–24. doi: 10.1016/j.ejphar.2015.11.048. [DOI] [PubMed] [Google Scholar]
- 157.Xagorari A, Roussos C, Papapetropoulos A. Inhibition of LPS-stimulated pathways in macrophages by the flavonoid luteolin. Br J Pharmacol. 2002;136:1058–1064. doi: 10.1038/sj.bjp.0704803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Yahfoufi N, Alsadi N, Jambi M, Matar C. The immunomodulatory and anti-inflammatory role of polyphenols. Nutrients. 2018;10:1618. doi: 10.3390/nu10111618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Yang S, Chu S, Ai Q, Zhang Z, Gao Y, Lin M, Liu Y, Hu Y, Li X, Peng Y, Pan Y, He Q, Chen N. Anti-inflammatory effects of higenamine (Hig) on LPS-activated mouse microglia (BV2) through NF-κB and Nrf2/HO-1 signaling pathways. Int Immunopharmacol. 2020;85:106629. doi: 10.1016/j.intimp.2020.106629. [DOI] [PubMed] [Google Scholar]
- 160.Yu S, Zheng W, Xin N, Chi ZH, Wang NQ, Nie YX, Feng WY, Wang ZY. Curcumin prevents dopaminergic neuronal death through inhibition of the c-Jun N-terminal kinase pathway. Rejuvenation Res. 2010;13:55–64. doi: 10.1089/rej.2009.0908. [DOI] [PubMed] [Google Scholar]
- 161.Zare K, Eidi A, Roghani M, Rohani AH. The neuroprotective potential of sinapic acid in the 6-hydroxydopamine-induced hemi-parkinsonian rat. Metab Brain Dis. 2015;30:205–213. doi: 10.1007/s11011-014-9604-6. [DOI] [PubMed] [Google Scholar]
- 162.Zeng KW, Zhang T, Fu H, Liu GX, Wang XM. Schisandrin B exerts anti-neuroinflammatory activity by inhibiting the Toll-like receptor 4-dependent MyD88/IKK/NF-κB signaling pathway in lipopolysaccharide-induced microglia. Eur J Pharmacol. 2012;692:29–37. doi: 10.1016/j.ejphar.2012.05.030. [DOI] [PubMed] [Google Scholar]
- 163.Zhang C, Tian X, Luo Y, Meng X. Ginkgolide B attenuates ethanol-induced neurotoxicity through regulating NADPH oxidases. Toxicology. 2011;287:124–130. doi: 10.1016/j.tox.2011.06.006. [DOI] [PubMed] [Google Scholar]
- 164.Zhou T, Zhu M, Liang Z. (-)-Epigallocatechin-3-gallate modulates peripheral immunity in the MPTP-induced mouse model of Parkinson’s disease. Mol Med Rep. 2018;17:4883–4888. doi: 10.3892/mmr.2018.8470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Zhu N, Lin J, Wang K, Wei M, Chen Q, Wang Y. Huperzine A protects neural stem cells against Aβ-induced apoptosis in a neural stem cells and microglia co-culture system. Int J Clin Exp Pathol. 2015;8:6425–6433. [PMC free article] [PubMed] [Google Scholar]
- 166.Zou K, Li Z, Zhang Y, Zhang HY, Li B, Zhu WL, Shi JY, Jia Q, Li YM. Advances in the study of berberine and its derivatives: a focus on anti-inflammatory and anti-tumor effects in the digestive system. Acta Pharmacol Sin. 2017;38:157–167. doi: 10.1038/aps.2016.125. [DOI] [PMC free article] [PubMed] [Google Scholar]