

Abstract
Radiation damage remains one of the major bottlenecks to accurate structure solution in protein crystallography. It can induce structural and chemical changes in protein crystals, and is hence an important consideration when assessing the quality and biological veracity of crystal structures in repositories like the Protein Data Bank (PDB). However, detection of radiation damage artefacts has traditionally proved very challenging. To address this, here we introduce the Bnet metric. Bnet summarises in a single value the extent of damage suffered by a crystal structure by comparing the B-factor values of damage-prone and non-damage-prone atoms in a similar local environment. After validating that Bnet successfully detects damage in 23 different crystal structures previously characterised as damaged, we calculate Bnet values for 93,978 PDB crystal structures. Our metric highlights a range of damage features, many of which would remain unidentified by the other summary statistics typically calculated for PDB structures.
Subject terms: X-ray crystallography, Quality control, Proteins
Radiation damage hampers protein structure determination by X-ray crystallography. Here, the AUs introduce the Bnet metric, a single value summarising the extent of radiation damage of a protein crystal structure, and use Bnet to detect radiation damage in structures deposited in the Protein Data Bank.
Introduction
The radiation damage suffered by crystals during X-ray diffraction experiments has long hindered accurate structure determination in protein crystallography. Despite the development of various damage mitigation strategies, including cryo-cooling1 and optimisation of the data collection strategy using software such as BEST2 or RADDOSE-3D3, data collection methodologies using the increasing flux densities of synchrotron light sources have resulted in radiation damage remaining one of the major challenges in protein crystallography. This problem will only be further exacerbated as the recently constructed fourth-generation synchrotron sources come online.
Radiation damage can affect both the individual asymmetric unit copies and the overall crystal lattice structure. Damage to the crystal lattice (global radiation damage) is readily detectable from changes in diffraction pattern reflection intensities, primarily as the fading and ultimate loss of high-resolution reflections4. Crystallographers are therefore able to truncate their datasets to discard diffraction patterns substantially affected by global radiation damage (within the limitation of retaining sufficient data completeness for structure solution).
Conversely, damage to the individual asymmetric unit copies (specific radiation damage) has traditionally proven challenging to detect within individual protein crystal (PX) structures. Accordingly, specific radiation damage is usually studied by identifying differences between successive datasets collected from the same crystal(s) (e.g. the radiation damage datasets collected from six different proteins and deposited by Nanao et al.5). Differences are caused by structural rearrangements (chiefly side-chain disordering) and chemical changes initiated by the electrons that are ejected upon the absorption of incident X-rays by the sample. At cryotemperatures (around 100 K), these induced chemical changes have been observed to occur in a reproducible order with increasing dose (the energy absorbed per unit mass of the crystal): for example, in PX structures metal ions are the first to be reduced6; followed by the breakage of disulfide bonds; aspartate and glutamate residues are then decarboxylated; and next the methylthio group is cleaved from methionine residues7–9.
Consequently, in contrast to global radiation damage, crystallographers are often unable to detect specific radiation damage artefacts within their structures and correct them appropriately. As an example, one could assume that an active site glutamate residue is disordered and that such disordering is potentially involved in the catalytic mechanism of its parent enzyme, when in fact the residue has just been decarboxylated by the incident X-rays. Such errors can compromise the conclusions drawn from a structure, and necessitate studies to separate biologically relevant features from those caused by radiation damage (e.g. during the bacteriorhodopsin photocycle10,11, and the recent study of the bending of flavin in the mechanism of fatty acid photodecarboxylase12).
Furthermore, and unfortunately, specific damage usually onsets prior to global damage: the experimental dose limit (corresponding to a 30% loss in summed reflection intensities from apo- and holo-ferritin crystals) was reported as 30 MGy at 100 K13, whereas aspartate/glutamate decarboxylation has been detected at doses as low as 4 MGy14. Owing to the difficulties associated with its detection, the number of Protein Data Bank (PDB)15 structures containing specific radiation damage artefacts is unknown; however, given that during a typical X-ray diffraction experiment a protein crystal held at 100 K absorbs a dose on the order of 1–10 MGy per complete dataset4, it is likely to be a substantial fraction.
In light of these issues, previously our group developed a metric called BDamage, a per-atom quantity that identifies potential sites of specific radiation damage within individual atomic resolution PX models as atoms with high full isotropic atomic B-factor (from herein referred to as B-factor) values relative to other atoms in a similar packing density environment16. BDamage has been validated to highlight known sites of specific damage in structures derived from a range of different radiation damage datasets. It is therefore a useful tool to flag up suspect sites within individual PX structures.
Unfortunately, however, the variability in the relationship between B-factors and radiation damage, caused by factors such as differences in refinement protocols and data resolution, means that absolute BDamage values cannot be fairly compared between structures. Furthermore, it is not possible to determine a single threshold value of BDamage beyond which an atom has a fixed probability of being damaged. These shortcomings prevent BDamage from being used to systematically assess the radiation damage suffered by the PX structures deposited in the PDB. Consequently, specific radiation damage is currently largely ignored in PX structure quality assessment.
To address these problems, in this work we have developed the Bnet metric. Bnet is a derivative of BDamage that summarises the overall extent of damage suffered by a PX structure in a single value. Initially, we demonstrate that Bnet values can be fairly compared between different PX structures, before using the metric to quantify for the first time the extent of radiation damage suffered by a dataset of 93,978 PX structures in the PDB. Based upon this analysis, we propose the Bnet metric to be a highly useful tool to assess PX structure quality for both newly and previously deposited structures.
Results
Defining the Bnet metric
When a structure suffers specific radiation damage, the B-factor values of the affected atoms increase relative to unaffected atoms (provided that the affected sites are not subjected to occupancy refinement). The BDamage metric identifies potentially damaged atoms as those with high B-factor values in comparison to other atoms in a similar local environment (Fig. 1 and Methods). Consequently, as a structure is increasingly damaged, the BDamage values of its affected atoms increase, and furthermore the distribution of the structure’s BDamage values becomes increasingly positively skewed. The skewness of its BDamage distribution can thus be used as a measure of the overall extent of specific radiation damage suffered by an individual PX structure. However, specific radiation damage typically only affects a minority of the atoms in even the most extensively damaged structures. Accordingly, the sensitivity of a metric that measures the skewness of the BDamage distribution of all atoms in a structure to specific radiation damage artefacts was found to be suboptimal (Supplementary Fig. 1).
Fig. 1. Illustration of the BDamage metric.
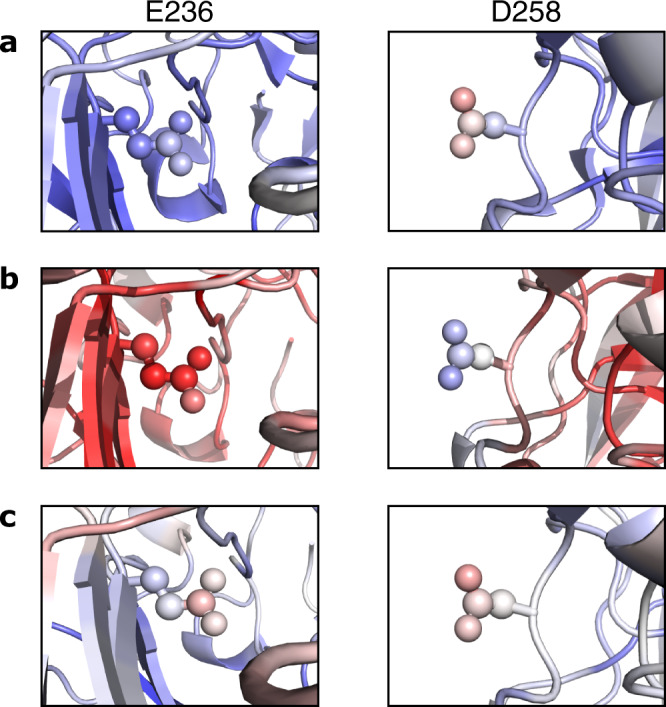
A damaged structure of GH7 family cellobiohydrolase (PDB accession code 5MCN35) is coloured (blue: low value; red: high value) by a B-factor, b packing density and c BDamage. The side chains of E236 and D258 are shown (as spheres) in the left- and right-hand images, respectively; although they have different B-factor values because the local packing density environments of the two residues are different, their BDamage values reveal the two residues to be similarly damaged. An atom’s packing density is defined as the number of non-hydrogen atoms within a 7 Å radius in the context of the crystalline structure. For visual clarity, only a single asymmetric unit copy is shown, however packing density and BDamage values have been calculated in the context of the crystalline structure as described in the Methods.
Therefore, we have instead focussed upon the BDamage values of known sites of specific damage. Because different damage events occur at different rates, plus affect different relative proportions of atoms in different structures, in order to obtain a value that is fairly comparable between structures we decided to focus upon only one type of damage event. We have selected the decarboxylation of aspartate and glutamate side chains because: (i) the vast majority (99.8% (3sf)—see Methods for details on how this and subsequent dataset sizes plus percentages were calculated) of PX structures deposited in the PDB contain one or more aspartate/glutamate residues; and (ii) aspartate/glutamate decarboxylation is one of the earliest onset and most extensively studied specific radiation damage events. Because aspartate/glutamate decarboxylation has primarily been characterised and studied in cryo-cooled crystals, and in fact has thus far not been convincingly observed at room temperature17, we restrict the following validation of our Bnet metric and subsequent analysis of the PDB to structures collected at 80–120 K (88.0% (3sf) of PX structures in the PDB).
Accordingly, the Bnet metric measures the skewness of the BDamage values of a structure’s aspartate and glutamate side-chain carboxyl group oxygen atoms (the BDamage-Asp/Glu distribution) in relation to the BDamage values of all atoms in the structure. Specifically, Bnet is calculated from the kernel density estimate of the BDamage-Asp/Glu distribution plus the median BDamage value of all atoms in the structure: it is defined as the ratio of the area (B) under the curve on the right-hand side of the median to the area (A) under the curve on the left-hand side of the median (Eq. 1 and Fig. 2).
| 1 |
Fig. 2. Calculation of the Bnet metric.
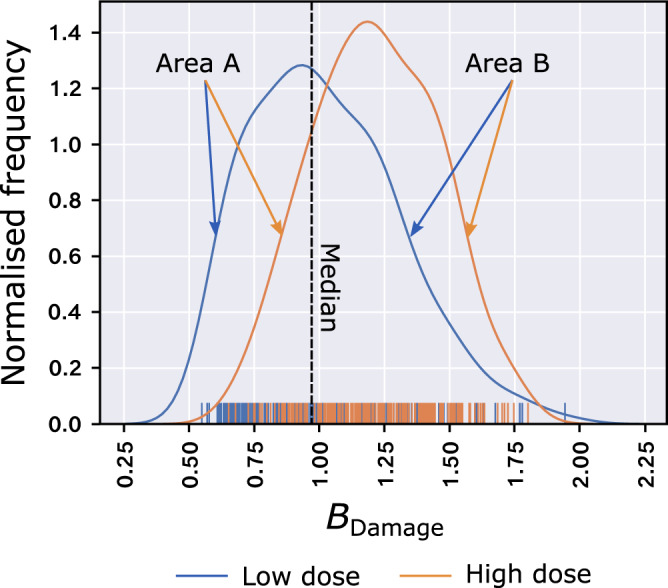
The Bnet metric is calculated from the kernel density estimate of the BDamage values of a structure’s aspartate and glutamate side-chain oxygen atoms as the ratio of the area under the curve on either side of the median BDamage value of all atoms in the structure. Displayed kernel density estimates and rug plots are of a low (1.11 MGy, PDB accession code 5MCC) and a high (22.7 MGy, PDB accession code 5MCN) dose structure of GH7 family cellobiohydrolase35. A and B are the areas under the kernel density estimate of the BDamage-Asp/Glu distribution to the left- and right-hand side of the median BDamage value of all atoms in the structure, respectively. Source data are provided in a Source Data File.
Experimentally, differences in resolution and refinement protocols between PX structures mean that B-factor values do not scale with damage via any consistently observed relationship. Consequently, although all BDamage distributions have a similar shape (approximately lognormal about 1), the standard deviation and range of the BDamage distribution is largely independent of the extent of damage suffered by a structure; hence BDamage values can be fairly compared within, but not between, structures.
We have therefore defined Bnet as in Eq. 1, since, in comparison with other measures of skewness such as quartile-based methods and Pearson’s skewness coefficients, this area ratio is more strongly influenced by the number of BDamage data points on either side of the median, as opposed to their raw values (and correspondingly the standard deviation and range of the distribution). Moreover, using such an area-based calculation makes fewer assumptions about the shape of the BDamage-Asp/Glu distribution; unlike their all-atom counterparts, there can be substantial variation between the shapes of the BDamage-Asp/Glu distributions of different (especially smaller) structures. We compared five different methods of measuring skewness and found that Bnet as defined in Eq. 1 achieved the best performance on our dataset of PDB structures (see Methods for further details). This analysis is available at 10.5281/zenodo.5567741.
In the following analysis, we validate that Bnet values can be fairly compared between different PX structures.
Validation of the Bnet metric
In order for Bnet to be a valid measure of specific radiation damage that can be fairly compared between different structures, it must: (i) scale, via a consistent relationship, with the radiation damage suffered by a structure; and (ii) be independent of other variables that can affect B-factor values.
Bnet increases with dose
To test whether Bnet satisfies the first of these conditions, we calculated Bnet values for 23 radiation damage datasets18–35 deposited in the PDB as of 19 November 2020 (Supplementary Tables 1 and 3). To the best of our knowledge, we have included in our analysis every radiation damage dataset that: (i) resulted in a protein structure (owing to a lack of suitable radiation damage datasets, BDamage has not been validated for nucleic acids); (ii) was collected at a temperature in the range of 80–120 K; (iii) contains data for three or more structures collected at increasing doses; iv) each dataset in the series has been collected from the same crystal(s) (hence successive structures are definitely increasingly damaged); and v) has associated information about the dose absorbed during each dataset in the series, reported in gray (Gy). Because Bnet is a B-factor-derived metric, we also excluded datasets resulting in structures that contain one or more aspartate/glutamate residue(s) whose occupancy across all listed conformers is less than one. For the same reason, we only retained datasets giving models which were refined with per-atom B-factors (either isotropic or anisotropic), and discarded those refined with e.g. per-residue B-factors or similar. Additionally, we excluded datasets giving one or more structures with a resolution worse than 3.5 Å; as resolution deteriorates, the number of observed reflections decreases, meaning fewer model parameters can be refined. It is highly unlikely that per-atom B-factor refinement is the appropriate choice for structures solved to worse than 3.5 Å resolution. Lastly, we removed datasets giving one or more structures with an Rfree value greater than 0.4, since these models show such poor fit to the electron density data that detection of radiation damage artefacts is very difficult if not impossible.
The extent of radiation damage suffered by a PX structure at cryotemperatures is proportional to the dose absorbed by the crystal36, as initially observed at room temperature by Blake and Phillips in 196237. Therefore, if indeed it is a valid measure of radiation damage, we expect Bnet to increase monotonically with dose, and furthermore to obtain the same Bnet value for different structures exposed to the same dose.
As shown in Fig. 3, we observe an approximately linear relationship between Bnet and dose for aspartate/glutamate side-chain oxygen atoms. This is not without precedent; many studies have previously observed linear correlations between a range of different B-factor-based metrics and dose7,38,39. However, unlike these other relationships, the correlation between Bnet and dose is much more consistent across the 23 datasets, since the gradients and y-intercepts of the lines of best fit plotted through the datasets are largely similar, as measured by their relative standard deviations (Fig. 3, plus Supplementary Tables 1, 2).
Fig. 3. Dose dependence of the Bnet metric for radiation damage datasets.

a Bnet (Asp/Glu), b Bnet (Asn/Gln) and c Wilson B-factor vs. dose for 23 radiation damage datasets. Straight lines of best fit, calculated via least squares regression, are plotted through each dataset. For clarity, the datasets have been split into “low” (0-4 MGy), “mid” (4 – 12 MGy) and “high” (>12 MGy) groups, determined by the highest dose absorbed. Source data are provided at 10.5281/zenodo.5566557.
Unsurprisingly, given the huge diversity of structures deposited in the PDB, there are a few exceptions. Firstly, the linear relationship between Bnet and dose is noisy (with some datasets being affected more than others): B-factors can be affected by multiple variables in addition to radiation damage. The main such variable is mobility: however other researchers have previously demonstrated how relative B-factor values can also be distorted by map over-sharpening, grossly different mobilities of subunits in the same asymmetric unit, and poor fit of the model to the data, amongst other variables40. Although the BDamage metric (and likewise the derivative Bnet) attempts to remove the effects of mobility, and concomitantly some of the other variables that impact upon B-factor values, by comparing the B-factor values of atoms in similar local packing density environments, this correction is not perfect. Moreover, there is a considerable error associated with dose calculations, to the extent that others have previously applied a threshold of a factor of 2 when assessing whether the variation in radiation damage effects in different experiments is significant or not41.
Consequently, it is not surprising that the datasets resulting in fewer structures tend to have more extreme gradient and y-intercept values. In particular, the gradient of the series collected by Castellvi et al.22 is double that of the next highest gradient. However, this dataset series only consists of three structures, hence a small change in either Bnet or dose of one or more of these structures would cause a large change in gradient.
Secondly, restraints applied during refinement prevent the B-factor values of neighbouring atoms from varying too much from one another. This can prevent the B-factor values of damaged atoms from increasing to reflect their damage. In spite of such restraints, the fact that Bnet increases with a dose for all except one series indicates that these restraints are generally not so tight that they prevent the Bnet metric from being an effective measure of damage. However, it is notable that of the four series with a gradient of 0.05 MGy−1 or less, two of them increase to gradients of greater than 0.1 MGy−1 when the structures’ B-factors are re-refined (see Supplementary Table 4 and Methods). It is unclear why the other two damage series, both from the same publication32, show an unusually low gradient.
Lastly, we observe that the Bnet values for the series collected by Bury et al.35 plateaus after ~14 MGy has been absorbed (consequently, we have excluded these data points when calculating the gradient and y-intercept of the line of best fit between Bnet and dose for this series). However, in contrast to their similar Bnet values, Bury et al. observe increasing damage artefacts between the final four structures in the series. Looking more closely, one can see that whilst in the earlier structures in the series the number of aspartate/glutamate side-chain oxygen atoms with BDamage values greater than the median (of all atoms in the structure) increases, in later structures this number remains fairly constant whereas the BDamage values of the damaged atoms become more extreme (Supplementary Fig. 2). A disadvantage of our method of measuring skewness is that it is less effective at picking up those latter types of changes. However, as described above, this is outweighed by the benefits of making fewer assumptions about the shape of the BDamage Asp/Glu distribution, especially for smaller structures: hence we have not changed our methodology. Importantly, the value of Bnet attained by the structure that has received 14.1 MGy is sufficiently high to mark it, plus the subsequent structures in the damage series, as having an unusually high Bnet value that merits further inspection, so fulfilling the purpose of the metric.
As an additional demonstration that the increasing Bnet values are representing damage to the successive structures in these radiation damage series, we have also calculated the equivalent of Bnet values for the side-chain amide group oxygen atoms of asparagine and glutamine residues (“Asn/Gln-Bnet”). As can be seen in Fig. 3(b) plus Supplementary Tables 1,2, the relationship between Asn/Gln-Bnet and the dose is much more inconsistent than for Asp/Glu-Bnet. The fit of the data to a linear relationship is poor, plus the gradients and y-intercepts of the straight lines of best fit that have been plotted are highly variable, reflected by their higher relative standard deviation scores as compared to Asp/Glu-Bnet (Supplementary Tables 1, 2). Moreover, in Fig. 3(c) we plot Wilson B-factor vs. dose for the 23 damage series. The data largely show a good fit to a linear relationship, however, because as described earlier experimental differences prevent B-factors from scaling consistently with dose across different structures, there is much more variation present within the gradients and in particular the y-intercepts of the calculated lines of best fit (Supplementary Tables 1, 2). Accordingly, it is clear that the (Asp/Glu-)Bnet metric successfully detects differences between increasingly damaged structures that are not captured when measuring either Wilson B-factor or the BDamage values of residues that do not suffer chemical changes as a result of radiation damage.
Consequently, this analysis supports the conclusion that Bnet values scale with dose via a consistent relationship, independent of structure identity.
Bnet correlates with resolution
To test whether Bnet is independent of variables, other than dose, that can affect B-factor values, and hence whether Bnet solely reflects the specific damage suffered by a structure, we calculated Bnet values for the 105,856 cryotemperature (80–120 K) PX structures deposited in the PDB with accompanying experimental data as of 19 November 2020. These structures were filtered using the same criteria listed above for the radiation damage datasets, apart from the requirements (i) to be part of a radiation damage dataset and (ii) for available dose information—the majority of structures in the PDB are deposited with neither their dose nor all of the information required to estimate that dose. In addition, we rejected structures with fewer than 20 carboxyl group oxygen atoms across their aspartate and glutamate side chains (an empirically determined threshold—see Supplementary Fig. 3), plus those which reported one or more null values for the eight variables (listed below) against which we compared Bnet. We also discarded structures refined with a flat B-factor model (information that, unlike PDB structures, is recorded for PDB-REDO42 structures—see below). Applying these filters left us with a dataset of 93,978 structures (i.e. 88.8% (3sf) of the initial 105,856).
For this analysis, we decided to use structures deposited in the PDB-REDO databank rather than in the PDB. PDB-REDO uses the latest available crystallographic software to re-refine all X-ray crystal structures deposited in the PDB with associated coordinates and structure factors. Consequently, structures in the PDB-REDO databank are more fairly comparable with one another, since they have been subjected to a consistent refinement protocol. Furthermore, extensive information about the final round of refinement, including the strength of the B-factor restraints applied, is available for all PDB-REDO structures. Note that because PDB-REDO is an automated pipeline, although in most cases it improves the structures it processes, it can also occasionally introduce errors, hence our decision to use the original PDB structures in our analysis of 23 radiation damage series in the previous section.
We calculated the (Spearman’s rank) correlation coefficients between the 93,978 PDB-REDO structures’ Bnet values and eight variables we predicted might impact their Bnet values in addition to radiation damage: namely, their resolution; Rwork; Rfree; temperature; molecular mass; the percentage and the raw number of aspartate/glutamate residues they contain; and the strength of the B-factor restraints applied in their final round of refinement. As discussed above, the lack of available dose information for the majority of PDB(-REDO) structures has unfortunately prevented us from comparing Bnet to dose as a positive control in this analysis.
We calculated Spearman’s rank correlation coefficients between each of the eight variables listed above and Bnet (Fig. 4 and Supplementary Table 5). Spearman’s rank (ρS) rather than Pearson’s correlation coefficient was selected since the former measures a monotonic relationship between variables, whereas the latter is a measurement of linearity. Using the threshold that absolute correlation coefficients of less than 0.2 (i.e. −0.2 < ρS < 0.2) indicate at most only a weak relationship between two variables, we observe no substantial correlation between Bnet and any of temperature, molecular mass, the number and percentage of aspartate and glutamate residues, and B-factor restraint weight.
Fig. 4. Investigating the dependence of Bnet on various parameters.

Scatter plots of Bnet vs.: a resolution; b Rwork; c Rfree; d temperature; e molecular mass; f the number of aspartate/glutamate side-chain oxygen atoms; g the percentage of aspartate/glutamate residues; and h the strength of B-factor restraints applied in the final round of refinement. The Spearman’s rank correlation coefficient (ρS) between the two variables is reported for each plot. Source data are provided at 10.5281/zenodo.5566558.
However, we observe non-negligible relationships between Bnet and resolution, Rwork and Rfree. The strongest correlation is between Bnet and resolution. The negative correlation between these two variables can be readily understood; as resolution decreases, electron density maps become more indistinct, and hence additional blurring due to aspartate/glutamate decarboxylation becomes more difficult to distinguish. Furthermore, a structure’s Rwork and Rfree values are strongly correlated with its resolution (the ρS coefficients between these two variables and resolution for our dataset of 93,978 structures are 0.60 (Rwork) and 0.64 (Rfree)): lower resolution structures have higher R-factors as there are fewer experimental data available to which to fit the model, hence the model and data display the poorer agreement.
Bnet-percentile does not correlate with resolution
In an attempt to overcome this issue, we extended our analysis to compare Bnet values of structures with similar resolutions. Many metrics of crystal structure quality are correlated with the resolution, hence they are commonly compared across subsets of structures of similar resolution. In our analysis, we used the same methodology as used by the PDB to compare structures of similar resolution: specifically, we calculated the percentile ranking of each structure’s Bnet value in the subset of structures closest in resolution (first the resolution range encompassing the 1000 closest structures was identified, all structures falling in this range were then included in the subset—see Methods for more details).
When calculating Spearman’s rank correlation coefficients between Bnet-percentile and the eight variables listed above (Fig. 5 and Supplementary Table 5), we find that the correlation coefficients of all eight variables with Bnet-percentile fall in the range of −0.2 < ρS < 0.2, thus demonstrating that (barring influence from a variable we have overlooked) Bnet-percentile values are predominantly affected by radiation dose, and correspondingly by damage.
Fig. 5. Investigating the dependence of Bnet-percentile on various parameters.

Scatter plots of Bnet-percentile vs.: a resolution; b Rwork; c Rfree; d temperature; e molecular mass; f the number of aspartate/glutamate side-chain oxygen atoms; g the percentage of aspartate/ glutamate residues; and h the strength of B-factor restraints applied in the final round of refinement. The Spearman’s rank correlation coefficient (ρS) between the two variables is reported for each plot. Source data are provided at 10.5281/zenodo.5566558.
This of course though does not mean that Bnet and Bnet-percentile values are never affected by other variables. As an example, even though the correlation between B-factor restraint weights and Bnet is weak, if very strong B-factor restraints are applied to a structure this will prevent the B-factor values of damaged atoms from increasing relative to undamaged atoms, and hence will reduce Bnet. However, whilst there will be a few exceptions, the at best weak correlations between Bnet and the eight selected variables mean such exceptions should be rare.
Consequently, based upon the analysis in this and the previous section, we are confident that Bnet values can be used to compare the extent of radiation damage suffered by structures of a similar resolution, and Bnet-percentile values can be used to compare structures at different resolutions. Together these two metrics will provide users with a powerful tool to interrogate their own plus others’ structures for radiation damage artefacts.
Bnet and Bnet-percentile identify damaged structures in the PDB
To demonstrate the utility of these two metrics, we manually examined the ten structures with the highest Bnet values (all of which also have Bnet-percentile values of greater than 0.999—see Supplementary Table 6) in our dataset of 93,978 structures for radiation damage artefacts (the selected structures are 5WUC43, 5FXL44, 5XQP45, 3S8S, 3UX146, 1V70, 6Q5R47, 3A0748, 6BKL49, and 2XMK50). None of these structures mention radiation damage in either the authors’ remarks in the PDB entry or if recorded, their associated publications. However, eight of the ten structures (5WUC, 5FXL, 5XQP, 3S8S, 1V70, 6Q5R, 3A07, and 2XMK) show clear evidence of radiation damage to their aspartate/glutamate residues in their electron density maps, with blobs of negative difference density visible around the carboxyl groups of many of their aspartate and glutamate side chains. Furthermore, the two structures that contain disulfide bonds (5FXL and 3A07) also show clear damage to these bonds (Fig. 6 and Supplementary Fig. 4).
Fig. 6. Observation of typical radiation damage features in structures with high Bnet values.

wwPDB validation statistics, models and density maps for the ten structures with the highest Bnet values in the PDB-REDO databank. For the eight structures with visible radiation damage artefacts in their associated electron density difference maps, a representative example is shown: for the two structures without such artefacts, the alternative causes of their high Bnet and Bnet-percentile values are shown instead. 2mFobs – DFcalc maps (blue) are contoured at 1.5 rmsd; Fobs – Fcalc difference density maps are contoured at ± 3.0 rmsd (green/red).
Additionally, the unusually high Bnet values of the two structures that do not show obvious signs of damage (3UX1 and 6BKL) can be readily explained. In spite of having an Rfree value of 0.28, the fit of model 3UX1 to the associated data is so bad that it is impossible to assess whether there are any damage artefacts present or not (Fig. 6). It is possible that the model’s high Bnet value results from its aspartate and glutamate residues by chance displaying a worse fit to the density than the other residues in the structure, however, this hypothesis cannot be confirmed without extensive re-refinement of the structure.
Conversely, there is good agreement between model 6BKL and the data. However, the protein is an assembly of four identical copies of a 25-residue α-helical peptide. There are two aspartate residues per helix, one of which is two residues away from the N-terminus and the other of which is two residues away from the C-terminus. In general, the fit of a model to its corresponding data deteriorates at chain termini, and accordingly, atoms at these locations have higher B-factor values (which is indeed the case for 6BKL). Usually, the packing density values of the chain termini would be lower, and hence the BDamage metric would compensate for such increases in B-factor: however, in this case, because the structure is an oligomer of four short α-helices, the packing density does not vary much along the length of the helices. Consequently, the high Bnet value of 6BKL results from the close proximity of all of its aspartate residues to the chain termini, as opposed to radiation damage. There are very few other protein structures with such a distorted distribution of aspartate and glutamate residues throughout their chains, hence there will not be many other structures deposited in the PDB affected in this way. However, this result is a reminder that Bnet and Bnet-percentile values need to be interpreted in the context of what is known about a structure. This is especially important for structures with high Bnet and Bnet-percentile values that lie outside of the top 10: such structures are probably damaged, but are unlikely to display visible artefacts in their associated difference maps to enable “confirmation” of the Bnet(-percentile) reading (hence the need to develop the metrics in the first place).
Strikingly, the wwPDB validation metrics reported for these 10 structures, with the exception of 3UX1, are reasonable to excellent (Fig. 6), and certainly do not reflect the large number of radiation damage artefacts afflicting them. Many users of the PDB are unaware of the possibility of radiation damage artefacts in deposited structures, moreover, most users only view the deposited model (without the associated electron density map). The Bnet and Bnet-percentile metrics can help such users rapidly assess structure quality with respect to radiation damage, and thus help to prevent them from drawing incorrect biological conclusions from their own and others’ structures. As an example, there are two models of actinohivin (an anti-HIV protein) in our dataset of 93,978: 3A07 and 4G1R. Both have good values across the standard wwPDB validation statistics, but whilst 3A07 has the eighth highest Bnet value in the entire dataset (Bnet: 25.1 (3sf); Bnet-percentile: 1), 4G1R is ranked 30,497th, and has much more reasonable-looking Bnet and Bnet-percentile values (Bnet: 1.61 (3sf); Bnet-percentile: 0.489 (3sf)). 4G1R would thus be the better option of the two to take forwards for subsequent analysis.
Discussion
Difficulties in detecting specific radiation damage artefacts mean that they are largely ignored when solving and analysing PX structures. However, these artefacts are not benign and can compromise the biological information drawn from a structure, for example as demonstrated by the X-ray-induced changes in the metal site conformation of photosystem II6, and by the reduction of haem-containing proteins to redox states not relevant to their various mechanisms51.
To address this problem, here we have developed the Bnet metric, which assesses the extent of radiation damage suffered by cryo-temperature (80–120 K) PX structures. To quantify radiation damage, Bnet measures the fraction of and extent to which a structure’s aspartate/glutamate side-chain oxygen atoms have higher BDamage values than the median BDamage value of all atoms in the structure. We have validated that Bnet can be fairly compared between different structures of similar resolution and developed the Bnet-percentile metric to compare structures of different resolutions. Note that because we do not know the fraction of structures in the PDB that contain radiation damage artefacts, we have not attempted to set a threshold value for Bnet beyond which a structure is “damaged”. Instead, it will be for the user to decide whether or not a structure’s Bnet and Bnet-percentile values will prevent them from using the structure, by taking into consideration other available measures of crystal quality plus the purposes for which they intend to use the structure. Nevertheless, to aid the user, as a rough guide we suggest that a Bnet value greater than 3.0, and/or a Bnet-percentile value greater than 0.95, would merit further inspection of a structure.
Interestingly, we observe that the extent of radiation damage in the PDB has remained relatively constant over time, as measured by calculating the Spearman’s rank correlation coefficient between Bnet/Bnet-percentile and deposition year (Supplementary Fig. 5). By flagging probable damaged structures for further inspection, we hope that the Bnet and Bnet-percentile metrics will help users when inspecting a structure/deciding which structural model of a protein of interest they want to select for their analysis. We also hope that the metrics will help crystallographers to identify damaged structures before/during the PDB deposition process, and thus reduce the fraction of damaged entries in the PDB. Because they are B-factor-derived metrics, Bnet and Bnet-percentile values should only be calculated for fully refined structures, so unfortunately they cannot be used to assess damage during the refinement process—however, they could be used to assess whether a structure should be re-refined with a truncated dataset.
In the future, when more data are available for validation, we hope to develop equivalents of these two metrics for structures containing nucleic acids, and for those derived from room temperature data. Currently, users can calculate Bnet values for PX structures by running the RABDAM software52,53, which is distributed as part of the CCP4 software suite54 or alternatively is available from https://github.com/GarmanGroup/RABDAM. Bnet-percentile values for the dataset of 93,978 structures analysed in this publication are listed at 10.5281/zenodo.5566558.
Methods
BDamage
The BDamage value of an atom j is calculated as the ratio of its B-factor to the arithmetic mean of the B-factor values of atoms 1 to n that are classified as occupying a similar packing density environment (Eq. 2 and Fig. 1), with an atom’s packing density being defined as the number of non-hydrogen atoms within a 7 Å radius in the context of the crystalline structure.
| 2 |
Bnet calculation
Bnet is calculated from the kernel density estimate (KDE) of the BDamage values of a structure’s aspartate and glutamate side-chain oxygen atoms, as the ratio of the area under the curve on either side of the median BDamage value of all atoms in the structure. The median was selected as the measure of central tendency because the value of the mean is susceptible to distortion by outliers, whilst the modal value is much more sensitive to the shape of the distribution (which can be irregular and/or noisy in the case of smaller structures). The KDE is plotted in Python using a Gaussian kernel, whilst the kernel bandwidth is set using the Scott method55. The area under the KDE is measured using the trapezium rule, split into 99 trapeziums spanning equal intervals along the BDamage (x-)axis. The trapezium into whose area the median BDamage value of all atoms in the structure falls is included in the below-median area (area A in Eq. 1) calculation. The code for calculating Bnet is included in the RABDAM software52, which is open-source and available from GitHub (https://github.com/GarmanGroup/RABDAM). RABDAM is also distributed as part of the CCP4 software suite54.
Four different measures of skewness were calculated in addition to the Bnet metric for the structures derived from the 23 radiation damage series analysed (see “Radiation dataset analysis” section below), and lines of best fit between each metric and dose were calculated using least squares regression for all 23 series. When considering the goodness of fit (measured by R2) of these lines of best fit, plus the consistency of their gradients and y-intercepts (measured by normalised standard deviation), Bnet was judged to achieve the best performance of the five options. The results of this analysis are available at 10.5281/zenodo.5567741.
Radiation dataset analysis
Radiation damage datasets were identified by searching the PDBe (as of 19 November 2020) for “radiation damage”, “serial crystallography” and “dose”, then manually filtering the results to select structures in radiation damage series. Each selected radiation damage series: (i) is of a protein structure (without a nucleic acid component); (ii) was collected at a temperature in the range of 80–120 K; (iii) contains three or more models; (iv) every underlying dataset in the series has been collected from the same crystal(s); (v) has the dose (reported in Gy) absorbed by each structure derived from the series recorded in its associated publication; (vi) has no aspartate or glutamate residues whose total occupancy across all conformers is less than one; (vii) has been refined with per-atom B-factors (either isotropic or anisotropic); (viii) has been solved to a resolution of 3.5 Å or better; and (ix) has a Rfree value less than or equal to 0.4. Note that although occupancy refinement would be the correct treatment for decarboxylated side chains, this is rarely performed. This is because, as described in the main text, specific radiation damage artefacts are difficult to identify in individual PX structures, hence in most cases users are unaware of the damage in their structures. In order to detect damage, the BDamage and Bnet metrics exploit the increased B-factor values that result when damaged, sub-1 occupancy side chains are refined as full occupancy.
Metadata describing these structures were taken from their associated mmCIF files. The two exceptions to this were: (i) the Wilson B-factor, which was taken from the associated PDB validation report; and (ii) the dose, which had to be manually extracted from the relevant publication. All Wilson B-factor values were calculated using Xtriage. The doses of all structures were calculated using RADDOSE (versions 1 or 256,57 or -3D3,58), although the use of RADDOSE was not a prerequisite for selection. The 23 datasets and their associated metadata are listed in Radiation_damage_datasets_SI.csv, which is available at 10.5281/zenodo.5566557. The script used to extract the metadata for each structure in the dataset is available at https://github.com/kls93/Bnet_PDB_parsing.
Bnet(Asn/Gln) values were obtained by calculating the ratio of the area on either side of the median BDamage value of all atoms in the structure under a KDE of the BDamage values of that structure’s asparagine and glutamine side-chain oxygen atoms.
The four radiation damage datasets listed in Supplementary Table 4, selected to analyse the effects on their structures’ Bnet values of changing the B-factor restraint weights applied during the refinement, were re-refined by two different methods. Firstly, the structures were downloaded from the PDB-REDO databank; and secondly, the (original PDB) structures were subjected to three macrocycles of unrestrained B-factor refinement (whilst the model coordinates were constrained to remain constant) with the refinement package in the Phenix software suite59. Note that because we do not know the B-factor restraint weights applied to the original (PDB) models, there is uncertainty that either method reduces the B-factor restraint weights of the re-refined structures below those applied to the original structures. However, it is highly unlikely that any of the original structures were subjected to unrestrained B-factor refinement. Moreover, we can be confident that the PDB-REDO structures have not been refined with extreme B-factor restraints.
Generating a dataset of PX structures
A dataset of PX structures was obtained by filtering the PDB using the following criteria: (1) the entry was released on or before 19 November 2020; (2) it was solved by X-ray diffraction; (3) the structure is a protein; (4) it does not contain any nucleic acid components; and (5) experimental data are available. This generated a dataset of 120,311 structures. We then filtered this dataset to retain only those structures containing one or more aspartate/glutamate residues (either l- or d-isomers). The resultant dataset contained 120,075 structures, i.e. 99.8% (3sf) of the original 120,311. Further filtering to retain those structures collected at a temperature in the range of 80–120 K reduced the dataset to 105,856 structures, i.e. 88.0% (3sf) of the original 120,311.
PDB-REDO analysis
We searched the PDB-REDO databank42 (as of 19 November 2020) for PX structures that met all of the required criteria for the radiation damage datasets (see “Radiation dataset analysis” methods), excluding the need for the dose to be recorded or for them to be part of a radiation damage dataset giving three or more structures. We also applied filters to select structures that (i) contained 20 or more aspartate/glutamate side-chain oxygen atoms, and (ii) have not been refined with a flat B-factor model (information about the latter is not available for PDB structures, hence this criterion was not applied to the radiation damage datasets). Of the 105,856 cryo-temperature (80–120 K) PX structures in our PX dataset described above, 93,978 (88.8% (3sf) met these criteria and were successfully downloaded from the PDB-REDO databank, processed by the RABDAM software to calculate Bnet, and had values recorded for each of the eight variables whose correlation coefficient with Bnet we assessed. As described in the main text, we calculated Spearman’s rank correlation coefficients of Bnet with resolution, Rwork, Rfree, temperature, molecular mass, the percentage and the raw number of aspartate/glutamate residues a structure contains, and the strength of the B-factor restraints applied in the final round of refinement of a structure. The values of all these features, including Bnet, were measured for the asymmetric unit. Molecular mass, plus numbers of aspartate and glutamate residues, were calculated for all atoms expected to be present in the asymmetric unit (i.e. including unmodelled atoms). In contrast, because Bnet is a B-factor-derived metric, its value is calculated from the modelled atoms only. All metadata associated with each PDB-REDO entry were taken from the corresponding mmCIF file (we used “XXXX_final.cif”, to ensure that total B-factors are reported in the isotropic B-factor records as required by RABDAM). The exceptions to this were the B-factor restraint weights and the Wilson B-factor, which were taken from the associated data.json file for the PDB-REDO entry. Wilson B-factors for PDB-REDO entries have been calculated with SFcheck.
Bnet-percentile values were calculated by first determining the resolution range encompassing the 1000 structures out of the dataset of 93,978 that were closest in resolution; the percentile value of the structure’s Bnet score was then calculated relative to all structures in the dataset whose resolution lay in the calculated resolution range. Spearman’s rank correlation coefficients were also calculated between Bnet-percentile and the same eight variables as considered for Bnet. In addition, we calculated Spearman’s rank correlation coefficients of both Bnet and Bnet-percentile with the year of deposition in the PDB (which was retrieved from the mmCIF file for the PDB entry). The script used to determine values for these eight variables for every structure in the dataset, plus for calculating their Bnet-percentile values and finding the year of deposition in the PDB, is available at https://github.com/kls93/Bnet_PDB_parsing, whilst the output data frame from running this script is provided in csv format at 10.5281/zenodo.5566558.
Finally, we looked for radiation damage artefacts in the associated electron density maps of the ten structures with the highest Bnet values out of our dataset of 93,978 PDB-REDO structures. However, the wwPDB validation statistics and, where relevant, the models and electron density maps showing radiation damage artefacts, as presented in Fig. 6, are for the corresponding PDB structures. This is because the wwPDB validation metrics (both raw and percentile values) are those that users most commonly utilise to assess and compare crystal structure quality, and these metrics are not similarly presented (or calculated at all in the case of the percentile metrics) in the PDB-REDO databank. For comparison, the Bnet values of these corresponding PDB structures are listed in Supplementary Table 6—in all cases, their Bnet values are also unusually high (higher than the Bnet values measured for even the most damaged structures in the 23 radiation damage series).
Supplementary information
Acknowledgements
We gratefully acknowledge the Moritz-Heyman Scholarship Program and the BBSRC (Biotechnology and Biological Sciences Research Council) for funding summer internships for K.L.S. to complete this work. We thank Edward Lowe for helping to run RABDAM on the PDB-REDO databank, and for useful discussions concerning B-factors and B-factor restraints. We also thank, Ian Carmichael, Joshua Dickerson and Markus Gerstel for constructive comments on this manuscript.
Source data
Author contributions
K.L.S. and E.F.G. conceived the project. K.L.S. designed and performed the analysis. K.L.S. and E.F.G. wrote the manuscript.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. Peer reviewer reports are available.
Data availability
Bnet and Bnet-percentile values for the dataset of 93,978 PDB-REDO structures, and Bnet values for the 23 radiation damage datasets, have been deposited in the Zenodo repository at 10.5281/zenodo.5566558 and 10.5281/zenodo.5566557, respectively. A comparison of the performance of four different skewness metrics to the Bnet metric has been deposited in the Zenodo repository at 10.5281/zenodo.5567741. The accession codes of all 93,978 PDB codes used in this study are listed at 10.5281/zenodo.5566558 and can be accessed via their four-digit accession codes in the Protein Data Bank repository (https://www.rcsb.org/). Source data are provided with this paper.
Code availability
All of the code used to perform the above analysis is open-source and available from GitHub. The Bnet metric is calculated by the RABDAM software, which is available at https://github.com/GarmanGroup/RABDAM and https://zenodo.org/record/5932846. The script used to extract metadata (including values for each of the eight variables whose correlation with Bnet is tested) for each PDB-REDO structure, plus for calculating Bnet-percentile values, is available on GitHub at https://github.com/kls93/Bnet_PDB_parsing. The script used to extract metadata for the PDB structures included in the analysis of 23 radiation damage series is also available at this location.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Kathryn L. Shelley, Email: kathryn.l.shelley@gmail.com
Elspeth F. Garman, Email: elspeth.garman@bioch.ox.ac.uk
Supplementary information
The online version contains supplementary material available at 10.1038/s41467-022-28934-0.
References
- 1.Garman EF, Schneider TR. Macromolecular cryocrystallography. J. Appl. Cryst. 1997;30:211–237. [Google Scholar]
- 2.Bourenkov GP, Popov AN. Optimization of data collection taking radiation damage into account. Acta Cryst. D. 2010;66:409–419. doi: 10.1107/S0907444909054961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zeldin OB, Gerstel M, Garman EF. RADDOSE-3D: time- and space-resolved modelling of dose in macromolecular crystallography. J. Appl. Cryst. 2013;46:1225–1230. [Google Scholar]
- 4.Garman E. Radiation damage in macromolecular crystallography: what is it and why should we care? Acta Cryst. D. 2010;66:339–351. doi: 10.1107/S0907444910008656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nanao MH, Sheldrick GM, Ravelli RBG. Improving radiation-damage substructures for RIP. Acta Cryst. D. 2005;61:1227–1237. doi: 10.1107/S0907444905019360. [DOI] [PubMed] [Google Scholar]
- 6.Yano J, et al. X-ray damage to the Mn4Ca complex in single crystals of photosystem II: a case study for metalloprotein crystallography. Proc. Natl Acad. Sci. USA. 2005;102:12047–12052. doi: 10.1073/pnas.0505207102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Burmeister WP. Structural changes in a cryo-cooled protein crystal owing to radiation damage. Acta Cryst. D. 2000;56:328–341. doi: 10.1107/s0907444999016261. [DOI] [PubMed] [Google Scholar]
- 8.Ravelli RB, McSweeney SM. The ‘fingerprint’ that X-rays can leave on structures. Struct. Fold. Des. 2000;8:315–328. doi: 10.1016/s0969-2126(00)00109-x. [DOI] [PubMed] [Google Scholar]
- 9.Weik M, et al. Specific chemical and structural damage to proteins produced by synchrotron radiation. Proc. Natl Acad. Sci. USA. 2000;97:623–628. doi: 10.1073/pnas.97.2.623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Matsui Y, et al. Specific damage induced by X-ray radiatioin and structural changes in the primary photoreaction of bacteriorhodopsin. J. Mol. Biol. 2002;324:469–481. doi: 10.1016/s0022-2836(02)01110-5. [DOI] [PubMed] [Google Scholar]
- 11.Takeda K, et al. Crystal structure of the M intermediate of bacteriorhodopsin: allosteric structural changes mediated by sliding movement of a transmembrane helix. J. Mol. Biol. 2004;341:1023–1037. doi: 10.1016/j.jmb.2004.06.080. [DOI] [PubMed] [Google Scholar]
- 12.Sorigué D, et al. Mechanism and dynamics of fatty acid photodecarboxylase. Science. 2021;372:eabd5687. doi: 10.1126/science.abd5687. [DOI] [PubMed] [Google Scholar]
- 13.Owen RL, Rudiño-Piñera E, Garman EF. Experimental determination of the radiation dose limit for cryocooled protein crystals. Proc. Natl Acad. Sci. USA. 2006;103:4912–4917. doi: 10.1073/pnas.0600973103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bury CS, et al. RNA protects a nucleoprotein complex against radiation damage. Acta Cryst. D. 2016;72:648–657. doi: 10.1107/S2059798316003351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Berman HM, et al. The protein data bank. Nucleic Acids Res. 2000;28:235–242. doi: 10.1093/nar/28.1.235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gerstel M, Deane CM, Garman EF. Identifying and quantifying radiation damage at the atomic level. J. Syn. Rad. 2015;22:201–212. doi: 10.1107/S1600577515002131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.De la Mora E, et al. Radiation damage and dose limits in serial synchrotron crystallography at cryo- and room temperatures. Proc. Natl Acad. Sci. USA. 2020;117:4142–4151. doi: 10.1073/pnas.1821522117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Serrano-Posada H, et al. X-ray-induced catalytic active-site reduction of a multicopper oxidase: Structural insights into the proton-relay mechanism and O2-reduction states. Acta Cryst. D. 2015;71:2396–2411. doi: 10.1107/S1399004715018714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ferraroni M, et al. Reaction intermediates and redox state changes in a blue laccase from Steccherinum ochraceum observed by crystallographic high/low X-ray dose experiments. J. Inorg. Biochem. 2012;111:203–209. doi: 10.1016/j.jinorgbio.2012.01.011. [DOI] [PubMed] [Google Scholar]
- 20.Bui S, et al. Direct evidence for a peroxide intermediate and a reactive enzyme-substrate-dioxygen configuration in a cofactor-free oxidase. Angew. Chem. Int. Ed. Engl. 2014;53:13710–13714. doi: 10.1002/anie.201405485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sutton KA, et al. Insights into the mechanism of X-ray-induced disulfide-bond cleavage in lysozyme crystals based on EPR, optical absorption and X-ray diffraction studies. Acta Cryst. D. 2013;69:2381–2394. doi: 10.1107/S0907444913022117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Castellvi A, et al. Efficacy of aldose reductase inhibitors is affected by oxidative stress induced under X-ray irradiation. Sci. Rep. 2019;9:3177–3177. doi: 10.1038/s41598-019-39722-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zarate-Romero A, Stojanoff V, Cohen AE, Hansberg W, Rudino-Pinera E. X-ray driven reduction of Cpd I of Catalase-3 from N. crassa reveals differential sensitivity of active sites and formation of ferrous state. Arch. Biochem. Biophys. 2019;666:107–115. doi: 10.1016/j.abb.2019.03.020. [DOI] [PubMed] [Google Scholar]
- 24.Taberman H, Bury CS, van der Woerd MJ, Snell EH, Garman EF. Structural knowledge or X-ray damage? A case study on xylose isomerase illustrating both. J. Syn. Rad. 2019;26:931–944. doi: 10.1107/S1600577519005599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dubnovitsky AP, Ravelli RB, Popov AN, Papageorgiou AC. Strain relief at the active site of phosphoserine aminotransferase induced by radiation damage. Protein Sci. 2005;14:1498–1507. doi: 10.1110/ps.051397905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Fioravanti E, Vellieux FM, Amara P, Madern D, Weik M. Specific radiation damage to acidic residues and its relation to their chemical and structural environment. J. Syn. Rad. 2007;14:84–91. doi: 10.1107/S0909049506038623. [DOI] [PubMed] [Google Scholar]
- 27.Juers DH, Weik M. Similarities and differences in radiation damage at 100 K versus 160 K in a crystal of thermolysin. J. Syn. Rad. 2011;18:329–337. doi: 10.1107/S0909049511007631. [DOI] [PubMed] [Google Scholar]
- 28.Correy GJ, et al. Mapping the accessible conformational landscape of an insect carboxylesterase using conformational ensemble analysis and kinetic crystallography. Structure. 2016;24:977–987. doi: 10.1016/j.str.2016.04.009. [DOI] [PubMed] [Google Scholar]
- 29.Russi S, et al. Conformational variation of proteins at room temperature is not dominated by radiation damage. J. Syn. Rad. 2017;24:73–82. doi: 10.1107/S1600577516017343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Polyakov KM, Gavryushov S, Fedorova TV, Glazunova OA, Popov AN. The subatomic resolution study of laccase inhibition by chloride and fluoride anions using single-crystal serial crystallography: insights into the enzymatic reaction mechanism. Acta Cryst. D. 2019;75:804–816. doi: 10.1107/S2059798319010684. [DOI] [PubMed] [Google Scholar]
- 31.De la Mora E, Carmichael I, Garman EF. Effective scavenging at cryotemperatures: further increasing the dose tolerance of protein crystals. J. Syn. Rad. 2011;18:346–357. doi: 10.1107/S0909049511007163. [DOI] [PubMed] [Google Scholar]
- 32.Pechkova E, Tripathi S, Ravelli RB, McSweeney S, Nicolini C. Radiation stability of proteinase K crystals grown by LB nanotemplate method. J. Struct. Biol. 2009;168:409–418. doi: 10.1016/j.jsb.2009.08.005. [DOI] [PubMed] [Google Scholar]
- 33.Fukuda Y, et al. Redox-coupled structural changes in nitrite reductase revealed by serial femtosecond and microfocus crystallography. J. Biochem. 2016;159:527–538. doi: 10.1093/jb/mvv133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hasegawa K, et al. Development of a dose-limiting data collection strategy for serial synchrotron rotation crystallography. J. Syn. Rad. 2017;24:29–41. doi: 10.1107/S1600577516016362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Bury CS, Carmichael I, Garman EF. OH cleavage from tyrosine: debunking a myth. J. Syn. Rad. 2017;24:7–18. doi: 10.1107/S1600577516016775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Garman, E. F. & Weik, M. in Protein Crystallography: Methods and Protocols. (eds. Wlodawer, A., Dauter, Z. & Jaskolski, M.) (Springer, 2017).
- 37.Blake, C. C. F. & Phillips, D. C. Biological effects of ionizing radiation at the molecular level. 183–191 (International Atomic Energy Agency, 1962).
- 38.Kmetko J, Warkentin M, Englich U, Thorne RE. Quantifying X-ray damage in protein crystals at cryogenic temperatures. Acta Cryst. D. 2006;62:1030–1038. doi: 10.1107/S0907444906023869. [DOI] [PubMed] [Google Scholar]
- 39.Teng T, Moffat K. Primary radiation damage of protein crystals by intense synchrotron radiation. J. Syn. Rad. 2000;7:313–317. doi: 10.1107/S0909049500008694. [DOI] [PubMed] [Google Scholar]
- 40.Masmaliyeva RC, Murshudov GN. Analysis and validation of macromolecular B values. Acta Cryst. D. 2019;75:505–518. doi: 10.1107/S2059798319004807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Holton JM. A beginner’s guide to radiation damage. J. Syn. Rad. 2009;16:133–142. doi: 10.1107/S0909049509004361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.van Beusekom B, et al. Homology-based hydrogen bond information improves crystallographic structures in the PDB. Protein Sci. 2018;27:798–808. doi: 10.1002/pro.3353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Su M, et al. Structural basis for conductance through TRIC cation channels. Nat. Commun. 2017;8:15103–15103. doi: 10.1038/ncomms15103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hirano Y, Amano Y, Yonemura S, Hakoshima T. The force-sensing device region of alpha-catenin is an intrinsically disordered segment in the absence of intramolecular stabilization of the autoinhibitory form. Genes Cells. 2018;23:370–385. doi: 10.1111/gtc.12578. [DOI] [PubMed] [Google Scholar]
- 45.Mahatabuddin S, et al. Polypentagonal ice-like water networks emerge solely in an activity-improved variant of ice-binding protein. Proc. Natl Acad. Sci. USA. 2018;115:5456–5461. doi: 10.1073/pnas.1800635115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Dimattia MA, et al. Structural insight into the unique properties of adeno-associated virus serotype 9. J. Virol. 2012;86:6947–6958. doi: 10.1128/JVI.07232-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Rhys GG, et al. Navigating the structural landscape of de novo alpha-helical bundles. J. Am. Chem. Soc. 2019;141:8787–8797. doi: 10.1021/jacs.8b13354. [DOI] [PubMed] [Google Scholar]
- 48.Tanaka H, et al. Mechanism by which the lectin actinohivin blocks HIV infection of target cells. Proc. Natl Acad. Sci. USA. 2009;106:15633–15638. doi: 10.1073/pnas.0907572106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Thomaston JL, et al. Inhibitors of the M2 proton channel engage and eisrupt transmembrane networks of hydrogen-bonded waters. J. Am. Chem. Soc. 2018;140:15219–15226. doi: 10.1021/jacs.8b06741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Badarau A, Firbank SJ, Mccarthy AA, Banfield MJ, Dennison C. Visualizing the metal-binding versatility of copper trafficking sites. Biochemistry. 2010;49:7798–7810. doi: 10.1021/bi101064w. [DOI] [PubMed] [Google Scholar]
- 51.Kekilli D, et al. Photoreduction and validation of haem-ligand intermediate states in protein crystals by in situ single-crystal spectroscopy and diffraction. IUCrJ. 2017;4:263–270. doi: 10.1107/S2052252517002159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Shelley KL, Dixon TPE, Brooks-Bartlett JC, Garman EF. RABDAM: quantifying specific radiation damage in individual protein crystal structures. J. Appl. Cryst. 2018;51:552–559. doi: 10.1107/S1600576718002509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Shelley, K. L. RABDAM. https://zenodo.org/record/5932846 (2018).
- 54.Winn MD, et al. Overview of the CCP4 suite and current developments. Acta Cryst. D. 2011;67:235–242. doi: 10.1107/S0907444910045749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Scott, D. W. Multivariate Density Estimation. Theory, Practice and Visualization (Wiley, 1992).
- 56.Murray JW, Garman EF, Ravelli RBG. X-ray absorption by macromolecular crystals: the effects of wavelength and crystal composition on absorbed dose. J. Appl. Cryst. 2004;37:513–522. [Google Scholar]
- 57.Paithankar KS, Owen RL, Garman EF. Absorbed dose calculations for macromolecular crystals: improvements to RADDOSE. J. Syn. Rad. 2009;16:152–162. doi: 10.1107/S0909049508040430. [DOI] [PubMed] [Google Scholar]
- 58.Bury, C. S., Brooks-Bartlett, J. C., Walsh, S. P. & Garman, E. F. Estimate your dose: RADDOSE-3D. Protein Sci.27, 217–228 (2018). [DOI] [PMC free article] [PubMed]
- 59.Liebschner D, et al. Macromolecular structure determination using X-rays, neutrons and electrons: recent developments in Phenix. Acta Cryst. D. 2019;75:861–877. doi: 10.1107/S2059798319011471. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Bnet and Bnet-percentile values for the dataset of 93,978 PDB-REDO structures, and Bnet values for the 23 radiation damage datasets, have been deposited in the Zenodo repository at 10.5281/zenodo.5566558 and 10.5281/zenodo.5566557, respectively. A comparison of the performance of four different skewness metrics to the Bnet metric has been deposited in the Zenodo repository at 10.5281/zenodo.5567741. The accession codes of all 93,978 PDB codes used in this study are listed at 10.5281/zenodo.5566558 and can be accessed via their four-digit accession codes in the Protein Data Bank repository (https://www.rcsb.org/). Source data are provided with this paper.
All of the code used to perform the above analysis is open-source and available from GitHub. The Bnet metric is calculated by the RABDAM software, which is available at https://github.com/GarmanGroup/RABDAM and https://zenodo.org/record/5932846. The script used to extract metadata (including values for each of the eight variables whose correlation with Bnet is tested) for each PDB-REDO structure, plus for calculating Bnet-percentile values, is available on GitHub at https://github.com/kls93/Bnet_PDB_parsing. The script used to extract metadata for the PDB structures included in the analysis of 23 radiation damage series is also available at this location.