

Abstract
Background
Functional constipation (FC) is a common gastrointestinal (GI) disorder characterized by symptoms of constipation without a clear physiologic or anatomic cause. Gut microbiome dysbiosis has been postulated to be a factor in the development of FC, and treatment with probiotic regimens, including strains of Lactobacillus plantarum (L. plantarum), has demonstrated efficacy in managing symptoms. To further understand the role of L. plantarum in GI health, we conducted an animal study and a randomized, double-blind, placebo-controlled clinical trial to evaluate the effect of a specific sub-strain, Lp3a, on FC.
Methods
For the animal study, male Kunming mice were treated with doses of L. plantarum Lp3a ranging from 0.67 to 2.00 g/kg or an equivalent amount of placebo for 15 days prior to the induction of constipation via 20 mL/kg of 25% diphenoxylate solution. GI motility parameters including intestinal motion and stool amount were then assessed. In the human study, 120 patients with FC were randomized to treatment [L. plantarum Lp3a; 2×1.0×1010 (colony forming units; CFU) ×7 days] or control groups (n=60 each). The primary endpoint was survey information on FC signs/symptoms. Participants and observers were blinded to group allocation. A subset of 20 Lp3a treated patients underwent pre- and post-treatment 16 s ribosomal ribonucleic acid (rRNA) gene sequencing. Whole genome sequencing (WGS) of L. plantarum Lp3a was also performed.
Results
Lp3a-treated mice showed significantly improved intestinal motion, reduced time to first defecation, and increased stool amounts. Similarly, patients in the treatment group (n=59) reported significant improvements in FC signs/symptoms compared to controls (n=58; all P<0.05). Although 16 s rRNA sequencing revealed no significant variations between pre- and post-treatment samples, WGS of Lp3a itself revealed several biological pathways that may underlie the relief of FC symptoms in animals and humans, including methane and fatty acid metabolism and bile acid biosynthesis.
Conclusions
We found that the use of the novel probiotic sub-strain, L. plantarum Lp3a, led to clinically significant improvements in FC in both mice and humans, and identified the potential biological mechanisms underlying this activity.
Keywords: Functional constipation (FC), Lactobacillus plantarum, gut microbiota, whole genome sequencing (WGS)
Introduction
Functional constipation (FC), which is defined as constipation without a known anatomical or physiological cause, is a common disorder with an incidence rate of 1.9–40.1% in adults (1). Although the cause of FC remains elusive, it likely involves a constellation of factors including neurologic, psychosomatic, or nutritional components. The symptoms of FC primarily include straining during defecation, hard stools, incomplete evacuation, a sensation of an anorectal obstruction, and the need for manual maneuvers to facilitate defecation. FC is distinct from irritable bowel syndrome (IBS); thus, the diagnosis of FC requires exclusion of IBS (2,3). In addition to the health concerns related to chronic constipation, particularly recurrent pain, FC takes a severe toll on the mental health of those affected, and places an economic burden on hundreds of millions of patients worldwide (4,5).
Generally, the primary pathogenic mechanism characteristic of FC is dysfunctional gastrointestinal (GI) tract motility evidenced by prolonged gut transit time, reduced stool frequency, and fewer bowel movements (6). Thus, the effectiveness of treatments targeting FC are typically judged on the basis of improvements in these measures (4). Treatment modalities with demonstrated effectiveness in FC are multifarious, and include diet changes (particularly, increased fiber intake), exercise (aerobic and pelvic floor), bulking agents, stool softeners, over-the-counter (OTC) or prescription laxatives, probiotics, and biofeedback therapies. In cases that are refractory to the aforementioned medical therapies and severely debilitating, surgical interventions, such as laparoscopic subtotal resection, may even be indicated (2,6-9). Unfortunately, as many as 47% of FC patients have reported dissatisfaction with their current treatment regimens (e.g., OTC or prescription laxatives) due to their ineffectiveness or delayed effectiveness, lack of predictability, adverse effects, taste, and relatively high costs (10).
Given the pervasive effect of FC on the global population, there is an urgent need for the development of novel treatment approaches that provide rapid and consistent relief with minimal side effects at an affordable cost. Probiotics, which function by influencing the composition and/or activity of the gut microbiome, have been proposed as one such therapy. Indeed, the gut microbiome, which consists of microorganisms that colonize the surface of the GI tract and influence host nutritional, metabolic, immunologic, endocrine, and neural processes, has been thought to be altered in the setting of FC. In infants and children, several studies have demonstrated elevated levels in feces of pathogenic bacteria such as Proteobacteria (11).
Additionally, others have shown that compared to healthy controls, patients with FC have significant variations in gut microbial composition, particularly reduced colonization by so-called “beneficial bacteria” and low species abundance (12-14). Consistently underrepresented bacterial taxa in FC patients include Bifidobacteria, Lactobacillus, Bacteroides, and Prevotella (13,15,16). Notably, Bifidobacteria, Bacteroides, and Prevotella are potent producers of short-chain fatty acids (SCFAs), while the Lactobacillus species produce lactate via fermentation (17-19). These metabolites are important regulators of host physiology and are known to reduce gut transit time (4,6). Thus, replenishing or rebalancing these species through probiotic supplementation may be effective at establishing healthy microbial diversity and treating FC. In fact, several independent studies have demonstrated the efficacy of various Lactobacillus and Bifidobacterium species in the relief of FC symptoms in animal models of constipation and human FC patients (20-22). Moreover, pretreatment with Sargassum plagiophyllum as a prebiotic was effective in increasing fecal Bifidobacterium counts and water content, ultimately reducing gut transit time in a mouse model of FC (23).
One species of Lactobacillus that demonstrated particular efficacy is L. plantarum, a microorganism distributed throughout the healthy GI tract (24-28). In this study, we sought to evaluate the effect of a new strain of L. plantarum (i.e., Lp3a) in an animal model of constipation and in human FC patients via a randomized, double-blind, placebo-controlled clinical trial. Notably, the constipation-related signs and symptoms improved significantly in the animals and humans treated with Lp3a. Whole genome sequencing (WGS) of the bacterial L. plantarum Lp3a species revealed significantly enriched biological pathways, including methane and fatty acid metabolism, which are associated with GI motility and might underlie the clinical benefit. To our knowledge, this is the first study to explore the effect of L. plantarum Lp3a supplementation on FC. By obtaining data from both a mouse model of constipation and human FC patients, we have produced powerful results demonstrating a conserved ability of Lp3a to relieve the impaired GI motility of constipation. Compared to previous studies of L. plantarum species, our analysis also included exploratory gene analysis through WGS that provides novel insight into the mechanism by which this probiotic can influence constipation. We present the following article in accordance with the ARRIVE and CONSORT reporting checklists (available at https://atm.amegroups.com/article/view/10.21037/atm-22-458/rc).
Methods
Probiotics
The probiotic supplement used in both the animal and human studies was obtained from New-Bio. Tech. (Jiangsu Province, China), and packed as 2 g per bag containing 1.0×1010 colony forming units (CFU) of L. plantarum Lp3a (Lot# 2018041501 for the animal study, and Lot# 2019011501 for the clinical trial).
Animal study
Animals
Specific pathogen free (SPF) male Kunming mice [body weight (BW): 18–22 g; 8–10 weeks old; n=100] were purchased from Liaoning Changsheng Biotechnology Co., Ltd. (Certificate No: 211002300046431). The animals were housed individually in an SPF environment for 5 days to allow acclimation under standard diet and environmental conditions (Temperature: 20–26 ℃; Relative Humidity: 40–70%; 12-hour light/dark cycle). The animal study were approved by the Institutional Animal Care and Use Committee of Nantong University (No. 2110836), in compliance with guidelines for the care and use of laboratory animals as described by the U.S. National Institute of Health, 8th edition (29).
Experimental design
Animal grouping, treatment, and induction of constipation
The mice (n=100) were randomly divided into the following groups based on outcome measure assessed: (I) the intestinal motion test group (n=50); (II) the defecation test group (n=50). Each group was randomly subdivided into five subgroups based on treatment. Three of the five groups received varied doses of the Lp3a probiotic supplement [dosing groups (DGs); high-dose/30 times of the human daily dose =2.00 g/kg/BW of the supplement solution in phosphate-buffered saline (PBS), middle-dose/20 times of the human daily dose =1.33 g/kg/BW, and low-dose/10 times of the human daily dose =0.67 g/kg/BW], one group served as the negative (healthy) control group (NCG), and one group served as the constipation control group (CCG), each receiving 2.00 g/kg/BW of the placebo solution in PBS. The mice treated with their respective probiotic or placebo solution received one dose of 20 mL/kg/BW per day for 15 consecutive days via oral gavage. At the conclusion of the 15-day treatment period, the animals were fasted for 16 hours, at which point, the animals randomized to the constipation groups (DG or CCG) were gavaged with 20 mL/kg/BW of diphenoxylate solution (25%) to induce constipation. The healthy control (NCG) animals were gavaged with an equal volume of pure water.
Intestinal motion test
Thirty minutes following the induction of constipation or the administration of the control solution, the animals were administered 20 mL/kg of BW India Ink solution (Biotech Grade for Biological Stains; Phygene #PH1714) to assess their rate of intestinal motility. The purpose of the ink gavage was to label and monitor the rate of movement of the forming bolus through the GI tract. The bolus was allowed to travel for 25 minutes after labeling, after which the mice were sacrificed via cervical dissection, and the intestines were dissected from the pylorus to the ileocecal valve. The excised intestines were then straightened and measured to determine the total length of the small intestine (the pylorus to the ileocecal region), and the length of ink propulsion (the pylorus to the furthest distance of ink). The ink propulsion rate was defined as the length of ink propulsion (cm)/total length of the small intestines (cm).
Defecation test
The mice randomized to the defecation test were similarly treated with India Ink via oral gavage following a 16-hour fasting period to label the forming feces. The mice were then returned to individual housing with free access to food and water. The time from the oral gavage to first defecation of the labeled feces was recorded, and the amount (number of grains) and weight (grams) of the stool produced within 5 hours of the first defecation of the labeled feces were also recorded as measurements of GI motility.
Clinical trial
Study population
Screening was performed to identify patients complaining of symptoms related to FC, including infrequent defecation (<3 times/week) and hard stool. These potential patients were interviewed and those that met the diagnostic parameters for FC according to the Rome Criteria IV (30) and did not meet the exclusion criteria (see below) were enrolled in the study (n=120). Patients were excluded from the study if they met any of the following exclusion criteria: (I) unable to consume oral medication; (II) had non-specific complaints; (III) had a poor physical condition; (IV) had experienced surgery-related constipation in the past month; (V) had known organic causes of constipation (e.g., colon cancer, bowel obstruction, or inflammatory bowel disease); (VI) had strained defecation with pain; (VII) had acute GI disorders in the past 30 days; (VIII) were currently pregnant or menstruating; (IX) had significant comorbidities (e.g., cardiovascular, hepatic, renal, hematologic, or severe systematic diseases); (X) were currently undergoing treatment for other diseases; and/or (XI) had recently used gastrointestinal active drugs. The study was conducted in accordance with the Declaration of Helsinki (as revised in 2013). The clinical trial in this study was reviewed and approved by the Institutional Ethics Committee of the Central Hospital of Xianyang, Shanxi Province (No. XYSZXYY20190702). Informed consent was obtained from all the participants before their enrolment in the clinical trial. The study was performed in compliance with all federal guidelines and institutional policies.
Study design
Patients were recruited for this randomized, placebo-controlled, double-blind trial from July 5 to August 16, 2019 at the Central Hospital of Xianyang. All the patients were randomly divided into 2 groups with an allocation ratio of 1:1, and stratified by age, sex, and daily diet. Neither subjects nor researchers involved in data collection were aware of the group to which individuals were assigned. The treatment group was instructed to take the above described Lp3a probiotic supplement (2 g, 1.0×1010 CFU per bag; 2 bags per day), and the control group was instructed to take the placebo (2 g per bag; 2 bags per day) for 7 consecutive days. Probiotic and placebo treatments were similar in appearance and composition. The primary endpoint of this study was the information obtained regarding patients’ symptoms 7 days before treatment and throughout the duration of the study: (I) stool frequency (number per week); (II) defecation condition on a scale from 0 to 3 (on which 0 represented normal defecation, 1 represented the sensation of distention only, 2 represented obvious distention, or strained defecation, but rare abdominal pain or a burning sensation in the anus, and 3 represented obvious abdominal pain or a burning sensation in the anus); (III) stool form [scored 1–7 based on the Bristol Stool Form Scale (31)]; (IV) daily diet, especially fiber intake; (V) adverse events, such as nausea, diarrhea, and abdominal distention, and pain.
The following clinical parameters were also assessed as secondary endpoints throughout the trial: (I) vital signs, including blood pressure and pulse; (II) routine testing of blood, urine, and stool; (III) hepatic and renal function; (IV) plain chest X-ray, electrocardiogram, and abdominal ultrasonography. This clinical data was collected at the Central Hospital of Xianyang. The subjects were instructed not to change their diet or their daily exercise regimen for the duration of the trial. Between and within group comparisons of each of the variables were conducted.
Bioinformatics analyses
Fecal samples from the 20 probiotic-treated patients were collected before and after treatment. A16s ribosomal ribonucleic acid (rRNA) gene analysis was conducted to characterize the gut microbial composition of the samples. A WGS analysis of L. plantarum Lp3a was also conducted to identify the encoded proteins, functions, and biological pathways relevant to GI motility and/or constipation.
16s rRNA-analysis
Total bacterial genomic DNA was extracted from fecal samples, and the integrity and size of the DNA was validated by gel electrophoresis using a 1% agarose gel (Majorbio Bio-pharm Tech., Shanghai, China) Universal forward (319F: 5'-ACTCCTACGGGAGGCAGCAG-3') and reverse (806R: 5'-GGACTACHVGGGTWTCTAAT-3') primers were used for the polymerase chain reaction (PCR) amplification of the V3–V4 hypervariable regions of the bacterial 16s rRNA gene to define the bacterial composition and abundance (32). The PCR products were purified and quantified by QuantiFluor™-ST (Promega). Libraries, which were constructed via the TruSeqTM DNA Sample Prep Kit (Illumina), were sequenced on the Illumina Miseq platform. Paired-end reads were generated and merged to a single sequence using the Fast Length Adjustment of Short reads (FLASH; 1.2.0) (33). Quality control was continuously monitored by filtering for bases with Phred scores <20. The remaining high-quality, non-repetitive sequences were clustered into Operational Taxonomic Units (OTUs) at a 97% sequence identity threshold for clustering using Usearch (version 7.0 http://drive5.com/usearch/) (34). Chimeras were checked and removed during the clustering process using UCHIME (35). Sequence alignments were carried out with MAFFT (v7.427), and OTUs were assigned to the lowest possible taxonomic level using the Ribosomal Database Project (RDP) Classifier Version 2.2 at a 70% bootstrap value threshold to obtain species classification information. Alpha and beta diversity indices were computed using QIIME (36). Taxa abundances at the phylum and genus levels were analyzed using the Metastats function in Mothur and compared via ANOSIM (37). Between sample differences in specific microbiota abundance were identified via the Student’s t test and/or Wilcoxon rank-sum test.
WGS
High-molecular-weight genomic DNA was extracted from L. plantarum Lp3a (Tianjin Biochip Co., China), and evaluated for purity, quantity, and size through UV-Vis (NanoDrop, ThermoFisher, USA), fluorometric (Qubit, ThermoFisher), and Pulsed Field Gel Electrophoresis assays. DNA (5 g) was used to prepare 20 kb SMRTbell libraries in accordance with the manufacturer’s (PacBio’s) directions. Libraries were size-selected using the BluePippen instrument to deplete short inserts and impurities before sequencing on the PacBio Sequel System using 1.2.1 chemistry and a 360-minute movie length. De novo genomes were assembled using the hierarchical genome assembly process of HGAP4.0 from original genomic data obtained from the PacBio sequel (38). After the validation of the obtained genome, advanced annotations were performed, including analyses of Clusters of Orthologous Groups (COGs) of proteins, Gene Ontology (GO), and Kyoto Encyclopedia of Genes and Genomes (KEEG). Secreted proteins were identified by SignalP 5.0 (http://www.cbs.dtu.dk/services/SignalP/) and the TMHMM 2.0 online server (http://www.cbs.dtu.dk/services/TMHMM/).
Statistical analyses
The data obtained from the animal and clinical studies were statistically analyzed using IBM SPSS Version 21.0. A P value <0.05 was considered statistically significant. The distribution of the continuous variables was assessed for normality using the Kolmogorov-Smirnov test. For the animal study, the normally distributed data were analyzed using a one-way analysis of variance. For the clinical trial, the normally distributed data are presented as the mean ± standard deviation. Between group comparisons were made using two-sample t tests, and within group (pre- vs. post-treatment) comparisons were made using paired t tests. The categorical data were assessed using the Chi-square or Fisher’s exact tests as appropriate.
Results
Animal studies
The effects of probiotics on body weight
To assess the effect of the probiotic treatment on overall metabolism, health, and wellbeing, the mice were weighed before and after the 15-day treatment period. No significant difference was observed in the body weight of the mice in the three treated (DG) and the non-treated (CCG) constipation groups compared to the healthy control group (NCG). This suggests that neither the probiotic treatment nor the induction of constipation significantly affected body mass (see Table S1).
Intestinal motion test
The ink propulsion rate was calculated to determine the effect of the probiotic treatment on intestinal motility (see Table 1). Compared to the NCG, the CCG had a significantly lower ink propulsion rate (87.05%±8.59% vs. 41.08%±9.74%; P<0.001), which was consistent with the expected effect of constipation on GI motility. The ink propulsion rate was significantly lower in the probiotic-treated mice in the high-dose (64.00%±17.36%), middle-dose (61.39%±16.51%), and low-dose (65.39%±18.77%) groups than those in the CCG (each DG vs. CCG; all P<0.05). This suggests that probiotic treatment reduces constipation-induced delays in GI motility.
Table 1. The effects of probiotics on intestinal motion in mice.
| Groups | Ink propulsion rate (%) | P value |
|---|---|---|
| High-dose group | 64.00±17.36 | 0.012* |
| Middle-dose group | 61.39±16.51 | 0.037* |
| Low-dose group | 65.39±18.77 | 0.009* |
| CCG | 41.08±9.74 | 0.000# |
| NCG | 87.05±8.59 | – |
*, compared to CCG; #, compared to NCG. CCG, constipation control group; NCG, negative (healthy) control group.
Defecation test
To determine the effect of the probiotic treatment on defecation, we next assessed the time to first defecation following oral gavage with ink, as well as the weight and number of grains of stool produced within 5 hours (see Table 2). Constipation led to a delay in the average time to first defecation (128.7±43.3 vs. 78.6±29.5 min; P=0.030) and a reduction in the amount (15.4±6.0 vs. 8.9±2.4 grains; P=0.004) and weight (0.365±0.099 vs. 0.206±0.066 g; P=0.001) of stool in the CCG vs. the NCG mice. The significant increase in time to defecation and the reduction in the amount and weight of stool between the CCG and NCG confirmed the successful establishment of the constipation model (39). Treatment with high-dose probiotic supplementation ameliorated the effect of constipation on stool weight (0.332±0.081; P=0.010) and the amount (13.7±4.2; P=0.045), but did not significantly reduce the time to first defecation (111.9±36.8; P=0.765) compared to the CCG. There was no significant difference between these parameters in low- and middle-dose groups compared to the CCG.
Table 2. The effects of probiotics on defecation in mice.
| Groups | Time to first defecation (min) | P value | Grains in stool | P value | Weight of stool (g) | P value |
|---|---|---|---|---|---|---|
| High-dose group | 111.9±36.8 | 0.765* | 13.7±4.2 | 0.045* | 0.332±0.081 | 0.010* |
| Middle-dose group | 113.9±43.0 | 0.833* | 10.6±3.2 | 0.770* | 0.251±0.099 | 0.618* |
| Low-dose group | 125.3±48.8 | 0.999* | 10.1±4.1 | 0.916* | 0.233±0.097 | 0.904* |
| CCG | 128.7±43.3 | 0.030# | 8.9±2.4 | 0.004# | 0.206±0.066 | 0.001# |
| NCG | 78.6±29.5 | – | 15.4±6.0 | – | 0.365±0.099 | – |
*, compared to CCG; #, compared to NCG. CCG, constipation control group; NCG, negative (healthy) control group.
Clinical trial
Participants and baseline characteristics
A total of 120 participants at the Central Hospital of Xianyang who met the diagnostic (inclusion) criteria for FC were enrolled in this study between July 5 and August 16, 2019. Subjects were randomly allocated to either the treatment with the probiotic (n=60) or placebo (n=60) groups. At the conclusion of this study, 3 patients had been lost to follow-up, 1 in the treatment group, and 2 in the placebo group (see Figure 1). The baseline demographic and clinical characteristics, including age, gender, daily fiber intake (%), and stool frequency and form, did not differ significantly between the enrolled FC participants in each group (see Table 3). No abnormalities warranting subject exclusion were detected by the baseline clinical tests, which included chest X-ray, electrocardiogram, abdominal ultrasonography, and urine, and stool analyses.
Figure 1.
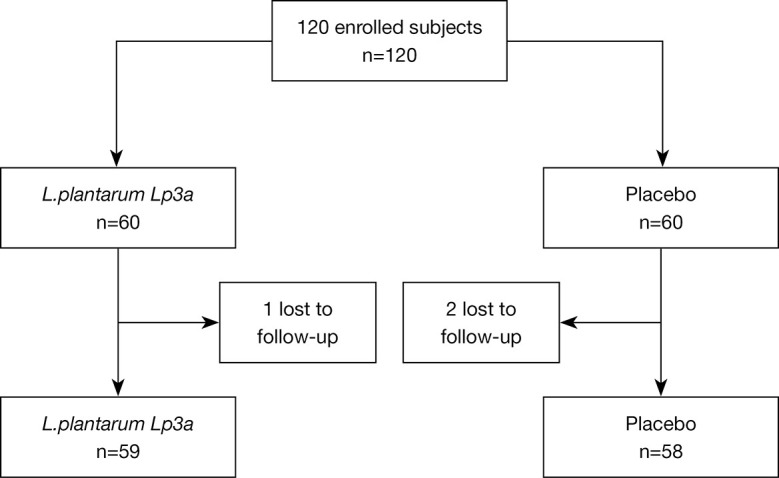
Flow chart demonstrating the design of the clinical trial.
Table 3. Baseline characteristics between the treatment and control groups.
| Variables | L. plantarum Lp3a (n=59) | Placebo (n=58) | P value |
|---|---|---|---|
| Female/male | 43/16 | 35/23 | 0.150# |
| Age (years) | 45.80±12.03 | 43.07±9.36 | 0.174* |
| Stool frequency (times/week) | 1.61±0.49 | 1.62±0.49 | 0.908* |
| Defecation condition | 1.64±1.06 | 1.66±1.07 | 0.955* |
| Stool form (Bristol scale) | 0.93±0.76 | 0.97±0.82 | 0.820* |
| Daily fiber intake (%) | 27.99±4.52 | 28.56±3.56 | 0.447* |
*, data compared by t-test; #, data compared by chi-square test.
Safety of L. plantarum Lp3a in humans
To ensure the safety of the L. plantarum Lp3a supplement, we used clinical tests to identify any potential physiologic or biochemical disturbances in the 117 patients before and after treatment. No obvious changes were detected by routine blood tests (see Table S2), biochemical parameters (see Table S3), and vital signs (see Table S4). No adverse effects or allergic reactions were observed or reported during this trial. These data indicate that the supplementation of L. plantarum Lp3a is generally safe for FC patients at the administered dosage.
L. plantarum Lp3a could alleviate FC symptoms
As described above, the baseline FC symptoms before the trial were indistinguishable between the two groups, indicating that these participants were drawn from a similar population and successfully randomized. Following the 7-day treatment period, patients randomized to the L. plantarum Lp3a group reported significant increases in stool frequency (2.75±0.94 vs. 1.61±0.49; P<0.001), a reduction in defecation difficulty (1.03±0.96 vs. 1.64±1.06; P<0.001), and more normal form according to the Bristol Stool Scale (0.53±0.57 vs. 0.93±0.76; P<0.001; see Table 4). Conversely, individuals randomized to the placebo group did not report significant improvements in these symptoms after the treatment period. To determine if biased dietary changes between these 2 groups confounded the results, we compared their daily fiber intake percentage, and found that the change in symptoms was not clearly influenced by dietary changes. We also observed no significant difference in the fiber intake of the probiotic-treated individuals before and after treatment, or any difference compared to the controls. These results extend those of our animal study, confirming that L. plantarum Lp3a is both biologically and clinically active against constipation.
Table 4. Comparison of constipation symptoms and diet before and after treatment.
| Variables | L. plantarum Lp3a (n=59) | Placebo (n=58) | P value |
|---|---|---|---|
| Stool frequency (#/week) | |||
| Before trial | 1.61±0.49 | 1.62±0.49 | 0.908* |
| After trial | 2.75±0.94 | 1.66±0.55 | 0.000* |
| Margin | –1.14±0.75 | –0.03±0.53 | 0.000* |
| Comparison within group (t, P) | –11.579, 0.000# | –0.497, 0.621# | |
| Defecation condition | |||
| Before trial | 1.64±1.06 | 1.66±1.07 | 0.955* |
| After trial | 1.03±0.96 | 1.66±1.12 | 0.002* |
| Margin | 0.61±0.49 | 0.00±0.50 | 0.000* |
| Comparison within group (t, P) | 9.528, 0.000# | 0.000, 1.000# | |
| Stool form (Bristol scale) | |||
| Before trial | 0.93±0.76 | 0.97±0.82 | 0.820* |
| After trial | 0.53±0.57 | 0.95±0.83 | 0.002* |
| Margin | 0.41±0.56 | 0.02±0.40 | 0.000* |
| Comparison within group (t, P) | 5.572, 0.000# | 0.331, 0.742# | |
| Daily fiber intake (%) | |||
| Before trial | 27.99±4.52 | 28.56±3.56 | 0.447* |
| After trial | 28.10±4.07 | 27.74±4.19 | 0.643* |
| Margin | –0.11±5.86 | 0.82±4.72 | 0.347* |
| Comparison within group (t, P) | –0.150, 0.881# | 1.314, 0.194# |
*, data compared by independent t-test; #, data compared by paired t-test.
16s rRNA gene analysis of stool samples
To identify alterations in the microbiome composition related to probiotic supplementation, we performed a 16s rRNA gene analysis on the fecal samples obtained from 20 participants before and after Lp3 treatment. A total of 1,679,233 high-quality sequences containing 691,231,074 bases and ranging in length from 401–450 bp were generated from 40 (20 pre-treatment and 20 post-treatment) fecal samples (see Figure S1A). These sequences clustered into 639 OTUs (582 after the trial, 575 before the trial, of which 518 were common) at 97% sequence identity (see Figure S1B). Shannon index and rank-abundance curves were computed to demonstrate the overall microbial species richness and evenness between the pre- and post-treatment fecal samples. The Shannon index curve, which plots out levels against the number of reads, demonstrated a plateau at 5,000 reads, indicating that the samples were even and sufficiently rich in species diversity (see Figure S1C). Similarly, the extended smooth lines plotted in the rank-abundance curve demonstrated high species richness and evenness in both the pre- and post-treatment samples (see Figure S1D).
To compare the diversity of species represented in the gut microbial colony between the samples, a beta diversity analysis was performed. Using the ANOSIM method, we failed to identify a significant difference in the microbial composition between the pre- and post-treatment samples at the phylum and genus levels (see Figure 2A,2B). At the phylum level, Firmicutes (55.4%), Actinobacteria (28.5%), Proteobacteria (8.7%), and Bacteroidetes (5.8%) were predominant in the pre-treatment samples. A seemingly similar composition was observed in the matched post-treatment samples: Firmicutes (52.0%), Actinobacteria (29.0%), Proteobacteria (12.5%), and Bacteroidetes (4.9%; see Figure 2C). Similarly, at the genus level, 5 genera predominated; that is, Bifidobacterium (pre-treatment =23.8% vs. post-treatment =25.3%), Romboutsia (7.9% vs. 4.8%), Escherichia-Shigella (7.8% vs. 10.5%), Blatuia (6.6% vs. 4.8%), and Subdoligranulum (6.1% vs. 7.8%; see Figure 2D). We failed to identify any specific microbiota that were significantly enriched or depleted at the genus and species levels using the Wilcoxon rank-sum test (see Figure 3A,3B). Overall, our 16s rRNA sequencing analysis revealed no significant difference in microbiome composition, which suggests that the observed effect of L. plantarum Lp3a on constipation is not directly mediated by microbial diversity.
Figure 2.

Analyses of microbiome composition in pre- and post-treatment fecal samples by 16S rRNA sequencing: (A) The principal component analysis (PCA) revealed no difference in composition between the pre- and post-treatment fecal samples at the phylum level (P=0.479, ANOSIM). (B) The PCA analysis revealed no difference in composition between the pre- and post-treatment fecal samples at the genus level (P=0.841, ANOSIM). (C) The relative abundance of microbial phyla between the pre- and post-treatment samples was indistinguishable (P=0.479, ANOSIM analysis). (D) The relative abundance of microbial genera between the pre- and post-treatment samples was indistinguishable (P=0.841, ANOSIM).
Figure 3.

Analyses of microbiome composition differences in major genera and species between the pre- and post-treatment fecal samples by the Wilcoxon rank-sum test: (A) bar plots demonstrating no significant changes in major genera abundance between the pre- and post-treatment (red and blue, respectively) fecal samples (all P>0.05, Wilcoxon rank-sum test); (B) bar plots demonstrating no significant difference in abundance of major species between the pre- and post-treatment fecal samples (all P>0.05, Wilcoxon rank-sum test).
WGS analysis
After failing to identify a treatment-related change in microbial diversity, we sought to determine if the biological activity inherent to L. plantarum Lp3a by virtue of its encoded proteins might underlie the effect on GI motility via WGS. This analysis provides a high-throughput pipeline for identifying protein-coding genes and biologically active pathways that are relevant to the various metabolic and physiologic functions of the sequenced organism.
The whole genome size of L. plantarum Lp3a was 3,214,487 bp with a guanine-cytosine (GC) content of 44.6%. Open reading frames (ORFs) accounted for 83.9% of the genome, and a total of 3,076 genes were identified with an average length of 876 bp.
COG-based annotation analyses are designed to classify proteins from completely sequenced genomes based on the ortholog concept. Around 20 COG functional categories were discovered in the genome of L. plantarum Lp3a (see Figure 4A). The most highly enriched clusters were “General function prediction only” (484 unigenes), “Function unknown” (352 unigenes), “Carbohydrate transport and metabolism” (235 unigenes), “Amino acid transport” (199 unigenes), and “metabolism and transcription” (198 unigenes). The GO functional classifications identified 31 subclasses,10 of which were related to molecular function, 11 to biological processes (BPs), and 10 to cellular components (see Figure 4B).
Figure 4.

Clusters of Orthologous Group (COG) and Gene Ontology (GO) classifications of functional proteins in the genome of L. plantarum L p3a: (A) protein-coding genes were clustered into 20 COG functional categories, including general function prediction only (484 unigenes), function unknown (352 unigenes), carbohydrate transport and metabolism (235 unigenes), amino acid transport (199 unigenes), and metabolism and transcription (198 unigenes); (B) protein-coding genes were clustered into 31 major GO subclasses, including molecular function (10 unigenes), biological process (11 unigenes), and cellular component (10 unigenes).
The functional pathway analysis identified 149 unique biological pathways, among which “metabolic pathways” (ko:01100) prevailed (415 unigenes). Other highly enriched pathways included the “biosynthesis of secondary metabolites” (ko:01100, 216 unigenes), “biosynthesis of amino acids” (ko:01230, 111 unigenes), “microbial metabolism in diverse environments” (ko:01120, 111 unigenes), “carbon metabolism” (ko:01200, 70 unigenes), “ABC transporters” (ko:02010, 68 unigenes), “purine metabolism” (ko:00230, 57 unigenes), “phosphotransferase system” (ko:02060, 53 unigenes), ribosome (ko:03010, 50 unigenes), “glycolysis/gluconeogenesis” (ko:00010, 46 unigenes), and “pyrimidine metabolism” (ko:00240, 44 unigenes). Notably, less abundant pathways directly related to GI motility and constipation were also identified, such as “methane metabolism” (ko:00680, 25 unigenes), “fatty acid metabolism” (ko:01212, 20 unigenes), “primary bile acid biosynthesis” (ko:00120, 4 unigenes), and “secondary bile acid biosynthesis” (ko:00121, 4 unigenes) (40).
As the probiotic supplementation did not directly affect composition, we took particular interest in the secreted and surface membrane proteins through which L. plantarum Lp3a could interact with the host gut microenvironment. A total of 224 surface and secreted proteins were identified, 138 of which were located in the bacterial membrane. The most abundant protein classes were “hypothetical” (19.2%), followed by “extracellular proteins” (17.0%), “cell surface protein and its precursor” (12.9%), “transport system substrate-binding protein” (9.4%), such as amino acid ABC transporter, phosphate ABC transporter, glutamine ABC transporter and sugar ABC transporter, and “lipoprotein and its precursor” (8.0%). Ectoenzymes, such as cell surface hydrolases and extracellular zinc metalloproteinases, accounted for a small but significant part of the predicted proteome. Overall, our WGS analysis identified important encoded proteins and biological pathways through which L. plantarum Lp3a may interact with the gut microenvironment to effect GI motility and relieve constipation.
Discussion
The role of the gut microbiome in GI and other systemic diseases is becoming increasingly apparent in the scientific literature. Numerous preclinical and clinical trials have explored the effect of probiotics in adult and pediatric patients with FC (20,22,28,41). Unfortunately, variability in study design, the included populations, the composition and dosage of the probiotic formulation, and, importantly, the diagnostic criteria for FC have resulted in inconsistencies across these studies. For example, in their meta-analysis of 14 randomized, placebo-controlled trials, Dimidi et al. demonstrated that probiotics significantly reduced gut transit time by 12.4 (95% CI: 222.3–22.5 hours) and increased stool frequency by 1.3 bowel movements per week (95% CI: 0.7–1.9 bowel movements/week) in adults (4). However, a similar analysis of randomized controlled trials, which included pediatric patients, failed to demonstrate that probiotics provided any clinical benefits in the relief of FC (16). It is clear that future research evaluating the characteristics of probiotics, particularly specific species, strains, and genome-encoded features, and their effects on FC need to be conducted to produce targeted formulations for this debilitating disease.
A promising species that has been explored in FC studies is L. plantarum, a lactic acid bacterium (LAB) that is widespread in multiple ecological niches, and widely used in the fermentation of food products, including cheeses, meats, vegetables, and beverages (42). Given their extensive role in food production, lactic acid bacteria, such as L. plantarum, have already been designated as “Generally Recognized As Safe” by the United States Food and Drug Administration, and thus are obvious targets for probiotic development (43). Additionally, these LAB species have well described BPs that help to stabilize the gut microenvironment, including the synthesis of SCFAs and the production of bacteriostatic substances (i.e., lactocins), and thus confer pathogen resistance (44,45). Many studies have investigated the effectiveness of various strains of L. plantarum on disease processes, such as dyslipidemia (46), diarrhea (47), constipation (24-28), IBS (48,49), and inflammatory bowel disease (50) in both animals and humans. Strains of L. plantarum have been shown to have an effect in animal models of constipation, including NCU116 (24), YS2 and YS3 (25,26), CQPC01, and CQPC02 (51,52). The relief of constipation induced by CQPC01 and CQPC02 is thought to be mediated by antioxidant activities and an increase in the levels of GI neuropeptides, such as motilin (51,52). In humans, L. plantarum SN35N and SN13T have shown promise in relieving constipation, reducing low-density lipoprotein cholesterol, and improving hepatic function (28). Further the L. plantarum LRCC5193 strain has been proven to be effective at ameliorating constipation-related symptoms in both a rat model (53) and human FC patients (27). Interestingly, L. plantarum has been showed to be effective in improving gut motility in a loperamide-induced model of constipation indicating that efficacy is not dependent on viability of the bacterium. Our current study adds to this growing body of literature by demonstrating the beneficial effect of an additional L. plantarum strain (i.e., Lp3a) on constipation symptoms in both mice and humans.
The mechanisms through which probiotics affect GI physiology and might improve FC symptoms are multifaceted and remain poorly understood. Perhaps most obviously, probiotics can directly modulate the GI microbial composition through direct colonization, the promotion of beneficial bacterial species, and/or the antagonism of pathogens (6). For example, Botelho et al. demonstrated that a multispecies probiotic supplement was capable of attenuating increases in the abundance of two bacterial species (i.e., Blautia faecis and Ruminococcus torques) that are known to be enriched in the microbiome of chronic constipation patients. This attenuation was associated with a significant reduction in symptoms of constipation, including incomplete defecation and the sensation of blockage (54). However, a lack of changes in gut microbial composition is not indicative of probiotic failure. Indeed, Kim et al. reported that the intake of a probiotic containing various species of Bifidobacterium and Lactobacillus in addition to Streptococcus thermophilus for 2 weeks improved constipation symptoms (i.e., increased defecation frequency) without a concomitant change in the GI flora (55).
The failure of our 16s rRNA analysis to identify significant differences in the gut microbiome structure between the probiotic- and placebo-treated patients indicated that the effect of the treatment on FC symptoms must be mediated by more indirect mechanisms. Such mechanisms could include the metabolism of nutrients into metabolically active substances, and the interaction of these metabolites or surface-bound proteins with the GI tract, central or enteric nervous system, endocrine system, or immune system (6). In a study of rats, Kunze et al. showed that supplementation with Lactobacillus reuteri for 9 days enhanced the excitability of enteric neurons by inhibiting the calcium-dependent potassium channel opening (56). These findings describe a neural pathway through which probiotics might affect GI motility and pain. To determine whether the effect of L. plantarum Lp3a on intestinal motility and constipation symptoms in mice and humans is mediated via the interaction of secreted and/or surface proteins with the host metabolism, we analyzed the genome of Lp3a using WGS. From the whole genome, which measured 3.2 Mb, consistent with other L. plantarum strains (57), we identified a number of enriched pathways that are directly correlated with GI motility. For example, encoded proteins related to methane metabolism might function to enhance gut transit rates by degrading methane, a metabolite associated with delayed transit times (58-60). Our WGS analysis also revealed the enrichment of the primary and secondary bile acid biosynthesis pathways in the L. plantarum Lp3a. Taken together with evidence that bile acids serve as physiological laxatives (6,61), this suggests a role for the modulation of bile acid metabolism as an additional putative mechanism of Lp3a in FC.
The WGS analysis also predicted a number of membrane-bound and secreted proteins in L. plantarum Lp3a that support its use as a probiotic for GI diseases, and potentially, for general health. The secretion of transport system substrate-binding proteins by Lp3a, which are associated with the uptake of sugar, amino acids, phosphate, and glutamine, may allow this microbiota strain to compete with pathogenic flora and thrive within the gut microenvironment (62). Extracellular zinc metalloproteinase, another secreted protein, might also facilitate the successful colonization of L. plantarum Lp3a in the digestive tract (63). Additionally, the production of lipoproteins and/or precursors by L. plantarum Lp3a might explain its contribution to protection against hyperlipidemia reported previously (42). Lp3a supplementation was not observed to have an effect on total cholesterol or triglycerides; however, a future analysis of a selected patient population with hyperlipidemia might reveal the potential role of Lp3a in cardiovascular health. Overall, the predicted pathways and secreted or membrane-bound proteins identified in this study support its role in alleviating constipation, and bolster its credibility as a clinical probiotic.
It is important that we recognize the limitations of this study. First, the sample size in the clinical trial was relatively small and was drawn from a single center, which introduces a risk of selection bias and limits the cross-geographic generalizability of the results. The small sample also prevented us from conducting subgroup analyses among the subtypes of FC and/or phenotypes of the gut microbiome before treatment. The latter is particularly limiting, as previous research has shown that the baseline composition of the microbiome is an important predictor of probiotic responses (64). Additionally, as we only enrolled adults in this study, we were unable to examine the role of L. plantarum Lp3a in children. This represents an obvious avenue for future research given the burden of FC in pediatric patients. For these reasons, future multicenter randomized controlled trials that include pediatric patients need to be conducted to validate the effectiveness of Lp3a, among other formulations, for FC. Similarly, animal studies remain important in elucidating the mechanisms by which probiotics affect host physiology.
Despite these limitations, this study made an important contribution to the growing body of literature on the use of probiotic therapy to treat GI and other systemic diseases, including FC. To our knowledge, this study was the first to investigate and validate the effect of the Lp3a strain of L. plantarum in both an animal model and human patients. Additionally, the combination of the 16s rRNA and WGS analyses allowed us to identify the diverse mechanisms by which Lp3a might affect the gut microenvironment, including numerous BPs and protein clusters relevant to GI motility. This provided an unprecedented understanding of the biological functions of this probiotic agent, which could inform the treatment of FC and other pathological, such as cardiovascular disease.
In conclusion, L. plantarum Lp3a effectively relieved features of constipation in both a mouse model and adult FC patients. Taken together, the mouse and human data suggest that this agent relieves FC symptoms by enhancing intestinal motility, and putatively modulating methane metabolism and bile acid synthesis. Larger studies need to be conducted to confirm its effectiveness in clinically relevant subgroups, including children and those with various subtypes of FC.
Supplementary
The article’s supplementary files as
Acknowledgments
Funding: This study was funded by the National Natural Science Foundation of China (No. 31500845 to HW and No. 81801080 to MS); the Fifth 226 High Level Talent Training Project of Nantong City (to XB); and the Qing Lan Project of Jiangsu Province (to XB). Funders were not involved in design or conduct of the trial.
Ethical Statement: The authors are accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. The animal study in this study was reviewed and approved by the Institutional Animal Care and Use Committee at Nantong University (No. 2110836), in compliance with guidelines for the care and use of laboratory animals as described by the U.S. National Institute of Health, 8th edition. The study was conducted in accordance with the Declaration of Helsinki (as revised in 2013). The clinical trial in this study was reviewed and approved by the Institutional Ethics Committee of the Central Hospital of Xianyang, Shanxi Province (No. XYSZXYY20190702). Informed consent was obtained from all the participants before their enrolment in the clinical trial.
Reporting Checklist: The authors have completed the ARRIVE and CONSORT reporting checklists. Available at https://atm.amegroups.com/article/view/10.21037/atm-22-458/rc
Trial Protocol: Available at https://atm.amegroups.com/article/view/10.21037/atm-22-458/tp
Data Sharing Statement: Available at https://atm.amegroups.com/article/view/10.21037/atm-22-458/dss
Conflicts of Interest: All authors have completed the ICMJE uniform disclosure form (available at https://atm.amegroups.com/article/view/10.21037/atm-22-458/coif). KM, GW, RW are from Jiangsu Biodep Biotechnology. The other authors have no conflicts of interest to declare.
(English Language Editor: L. Huleatt)
References
- 1.Suares NC, Ford AC. Prevalence of, and risk factors for, chronic idiopathic constipation in the community: systematic review and meta-analysis. Am J Gastroenterol 2011;106:1582-91; quiz 1581, 1592. 10.1038/ajg.2011.164 [DOI] [PubMed] [Google Scholar]
- 2.Skardoon GR, Khera AJ, Emmanuel AV, et al. Review article: dyssynergic defaecation and biofeedback therapy in the pathophysiology and management of functional constipation. Aliment Pharmacol Ther 2017;46:410-23. 10.1111/apt.14174 [DOI] [PubMed] [Google Scholar]
- 3.Mearin F, Lacy BE, Chang L, et al. Bowel Disorders. Gastroenterology 2016. [Epub ahead of print]. doi: 10.1053/j.gastro.2016.02.031 [DOI] [PubMed] [Google Scholar]
- 4.Dimidi E, Christodoulides S, Fragkos KC, et al. The effect of probiotics on functional constipation in adults: a systematic review and meta-analysis of randomized controlled trials. Am J Clin Nutr 2014;100:1075-84. 10.3945/ajcn.114.089151 [DOI] [PubMed] [Google Scholar]
- 5.Belsey J, Greenfield S, Candy D, et al. Systematic review: impact of constipation on quality of life in adults and children. Aliment Pharmacol Ther 2010;31:938-49. 10.1111/j.1365-2036.2010.04273.x [DOI] [PubMed] [Google Scholar]
- 6.Dimidi E, Christodoulides S, Scott SM, et al. Mechanisms of Action of Probiotics and the Gastrointestinal Microbiota on Gut Motility and Constipation. Adv Nutr 2017;8:484-94. 10.3945/an.116.014407 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gasior A, Reck C, Vilanova-Sanchez A, et al. Surgical management of functional constipation: An intermediate report of a new approach using a laparoscopic sigmoid resection combined with malone appendicostomy. J Pediatr Surg 2018;53:1160-2. 10.1016/j.jpedsurg.2018.02.074 [DOI] [PubMed] [Google Scholar]
- 8.Rao SS, Rattanakovit K, Patcharatrakul T. Diagnosis and management of chronic constipation in adults. Nat Rev Gastroenterol Hepatol 2016;13:295-305. 10.1038/nrgastro.2016.53 [DOI] [PubMed] [Google Scholar]
- 9.Basilisco G, Coletta M. Chronic constipation: a critical review. Dig Liver Dis 2013;45:886-93. 10.1016/j.dld.2013.03.016 [DOI] [PubMed] [Google Scholar]
- 10.Johanson JF, Kralstein J. Chronic constipation: a survey of the patient perspective. Aliment Pharmacol Ther 2007;25:599-608. 10.1111/j.1365-2036.2006.03238.x [DOI] [PubMed] [Google Scholar]
- 11.Hofman D, Kudla U, Miqdady M, et al. Faecal Microbiota in Infants and Young Children with Functional Gastrointestinal Disorders: A Systematic Review. Nutrients 2022;14:974. 10.3390/nu14050974 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Xu Z, Liu T, Zhou Q, et al. Roles of Chinese Medicine and Gut Microbiota in Chronic Constipation. Evid Based Complement Alternat Med 2019;2019:9372563. 10.1155/2019/9372563 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhu L, Liu W, Alkhouri R, et al. Structural changes in the gut microbiome of constipated patients. Physiol Genomics 2014;46:679-86. 10.1152/physiolgenomics.00082.2014 [DOI] [PubMed] [Google Scholar]
- 14.Parthasarathy G, Chen J, Chen X, et al. Relationship Between Microbiota of the Colonic Mucosa vs Feces and Symptoms, Colonic Transit, and Methane Production in Female Patients With Chronic Constipation. Gastroenterology 2016;150:367-79.e1. 10.1053/j.gastro.2015.10.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Khalif IL, Quigley EM, Konovitch EA, et al. Alterations in the colonic flora and intestinal permeability and evidence of immune activation in chronic constipation. Dig Liver Dis 2005;37:838-49. 10.1016/j.dld.2005.06.008 [DOI] [PubMed] [Google Scholar]
- 16.Wojtyniak K, Szajewska H. Systematic review: probiotics for functional constipation in children. Eur J Pediatr 2017;176:1155-62. 10.1007/s00431-017-2972-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Vlasova AN, Kandasamy S, Chattha KS, et al. Comparison of probiotic lactobacilli and bifidobacteria effects, immune responses and rotavirus vaccines and infection in different host species. Vet Immunol Immunopathol 2016;172:72-84. 10.1016/j.vetimm.2016.01.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Feng W, Ao H, Peng C. Gut Microbiota, Short-Chain Fatty Acids, and Herbal Medicines. Front Pharmacol 2018;9:1354. 10.3389/fphar.2018.01354 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Luu M, Pautz S, Kohl V, et al. The short-chain fatty acid pentanoate suppresses autoimmunity by modulating the metabolic-epigenetic crosstalk in lymphocytes. Nat Commun 2019;10:760. 10.1038/s41467-019-08711-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ibarra A, Latreille-Barbier M, Donazzolo Y, et al. Effects of 28-day Bifidobacterium animalis subsp. lactis HN019 supplementation on colonic transit time and gastrointestinal symptoms in adults with functional constipation: A double-blind, randomized, placebo-controlled, and dose-ranging trial. Gut Microbes 2018;9:236-51. 10.1080/19490976.2017.1412908 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Anzawa D, Mawatari T, Tanaka Y, et al. Effects of synbiotics containing Bifidobacterium animalis subsp. lactis GCL2505 and inulin on intestinal bifidobacteria: A randomized, placebo-controlled, crossover study. Food Sci Nutr 2019;7:1828-37. 10.1002/fsn3.1033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ojetti V, Ianiro G, Tortora A, et al. The effect of Lactobacillus reuteri supplementation in adults with chronic functional constipation: a randomized, double-blind, placebo-controlled trial. J Gastrointestin Liver Dis 2014;23:387-91. 10.15403/jgld.2014.1121.234.elr [DOI] [PubMed] [Google Scholar]
- 23.Khuituan P, Huipao N, Jeanmard N, et al. Sargassum plagiophyllum Extract Enhances Colonic Functions and Modulates Gut Microbiota in Constipated Mice. Nutrients 2022;14:496. 10.3390/nu14030496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li C, Nie SP, Zhu KX, et al. Effect of Lactobacillus plantarum NCU116 on loperamide-induced constipation in mice. Int J Food Sci Nutr 2015;66:533-8. 10.3109/09637486.2015.1024204 [DOI] [PubMed] [Google Scholar]
- 25.Zhao X, Yi R, Qian Y, et al. Lactobacillus plantarum YS-3 Prevents Activated Carbon-Induced Constipation in Mice. J Med Food 2018;21:575-84. 10.1089/jmf.2017.4109 [DOI] [PubMed] [Google Scholar]
- 26.Zhao X, Qian Y, Li G, et al. Lactobacillus plantarum YS2 (yak yogurt Lactobacillus) exhibited an activity to attenuate activated carbon-induced constipation in male Kunming mice. J Dairy Sci 2019;102:26-36. 10.3168/jds.2018-15206 [DOI] [PubMed] [Google Scholar]
- 27.Yoon JY, Cha JM, Oh JK, et al. Probiotics Ameliorate Stool Consistency in Patients with Chronic Constipation: A Randomized, Double-Blind, Placebo-Controlled Study. Dig Dis Sci 2018;63:2754-64. 10.1007/s10620-018-5139-8 [DOI] [PubMed] [Google Scholar]
- 28.Higashikawa F, Noda M, Awaya T, et al. Improvement of constipation and liver function by plant-derived lactic acid bacteria: a double-blind, randomized trial. Nutrition 2010;26:367-74. 10.1016/j.nut.2009.05.008 [DOI] [PubMed] [Google Scholar]
- 29.In: th, editor. Guide for the Care and Use of Laboratory Animals. The National Academies Collection: Reports funded by National Institutes of Health. Washington (DC)2011.
- 30.Drossman DA, Hasler WL. Rome IV-Functional GI Disorders: Disorders of Gut-Brain Interaction. Gastroenterology 2016;150:1257-61. 10.1053/j.gastro.2016.03.035 [DOI] [PubMed] [Google Scholar]
- 31.Lewis SJ, Heaton KW. Stool form scale as a useful guide to intestinal transit time. Scand J Gastroenterol 1997;32:920-4. 10.3109/00365529709011203 [DOI] [PubMed] [Google Scholar]
- 32.Klindworth A, Pruesse E, Schweer T, et al. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res 2013;41:e1. 10.1093/nar/gks808 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Magoč T, Salzberg SL. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011;27:2957-63. 10.1093/bioinformatics/btr507 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Edgar RC. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010;26:2460-1. 10.1093/bioinformatics/btq461 [DOI] [PubMed] [Google Scholar]
- 35.Edgar RC, Haas BJ, Clemente JC, et al. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011;27:2194-200. 10.1093/bioinformatics/btr381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Caporaso JG, Kuczynski J, Stombaugh J, et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods 2010;7:335-6. 10.1038/nmeth.f.303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Jami E, Israel A, Kotser A, et al. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J 2013;7:1069-79. 10.1038/ismej.2013.2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chin CS, Alexander DH, Marks P, et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat Methods 2013;10:563-9. 10.1038/nmeth.2474 [DOI] [PubMed] [Google Scholar]
- 39.Ma H, Xiong H, Zhu X, et al. Polysaccharide from Spirulina platensis ameliorates diphenoxylate-induced constipation symptoms in mice. Int J Biol Macromol 2019;133:1090-101. 10.1016/j.ijbiomac.2019.04.209 [DOI] [PubMed] [Google Scholar]
- 40.Chen L, Xiong Z, Sun L, et al. VFDB 2012 update: toward the genetic diversity and molecular evolution of bacterial virulence factors. Nucleic Acids Res 2012;40:D641-5. 10.1093/nar/gkr989 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ito D, Yamamoto Y, Maekita T, et al. Do synbiotics really enhance beneficial synbiotics effect on defecation symptoms in healthy adults?: Randomized, double-blind, placebo-controlled trial. Medicine (Baltimore) 2022;101:e28858. 10.1097/MD.0000000000028858 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Seddik HA, Bendali F, Gancel F, et al. Lactobacillus plantarum and Its Probiotic and Food Potentialities. Probiotics Antimicrob Proteins 2017;9:111-22. 10.1007/s12602-017-9264-z [DOI] [PubMed] [Google Scholar]
- 43.Żukiewicz-Sobczak W, Wróblewska P, Adamczuk P, et al. Probiotic lactic acid bacteria and their potential in the prevention and treatment of allergic diseases. Cent Eur J Immunol 2014;39:104-8. 10.5114/ceji.2014.42134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ljungh A, Wadström T. Lactic acid bacteria as probiotics. Curr Issues Intest Microbiol 2006;7:73-89. [PubMed] [Google Scholar]
- 45.Jack RW, Tagg JR, Ray B. Bacteriocins of gram-positive bacteria. Microbiol Rev 1995;59:171-200. 10.1128/mr.59.2.171-200.1995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kumar R, Grover S, Batish VK. Hypocholesterolaemic effect of dietary inclusion of two putative probiotic bile salt hydrolase-producing Lactobacillus plantarum strains in Sprague-Dawley rats. Br J Nutr 2011;105:561-73. 10.1017/S0007114510003740 [DOI] [PubMed] [Google Scholar]
- 47.Chingwaru W, Vidmar J. Potential of Zimbabwean commercial probiotic products and strains of Lactobacillus plantarum as prophylaxis and therapy against diarrhoea caused by Escherichia coli in children. Asian Pac J Trop Med 2017;10:57-63. 10.1016/j.apjtm.2016.12.009 [DOI] [PubMed] [Google Scholar]
- 48.Ducrotté P, Sawant P, Jayanthi V. Clinical trial: Lactobacillus plantarum 299v (DSM 9843) improves symptoms of irritable bowel syndrome. World J Gastroenterol 2012;18:4012-8. 10.3748/wjg.v18.i30.4012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Stevenson C, Blaauw R, Fredericks E, et al. Randomized clinical trial: effect of Lactobacillus plantarum 299 v on symptoms of irritable bowel syndrome. Nutrition 2014;30:1151-7. 10.1016/j.nut.2014.02.010 [DOI] [PubMed] [Google Scholar]
- 50.Nemoto M, Kuda T, Eda M, et al. Protective Effects of Mekabu Aqueous Solution Fermented by Lactobacillus plantarum Sanriku-SU7 on Human Enterocyte-Like HT-29-luc Cells and DSS-Induced Murine IBD Model. Probiotics Antimicrob Proteins 2017;9:48-55. 10.1007/s12602-016-9226-x [DOI] [PubMed] [Google Scholar]
- 51.Liu B, Xiao X, Zhou X, et al. Effects of Lactobacillus plantarum CQPC01-fermented soybean milk on activated carbon-induced constipation through its antioxidant activity in mice. Food Sci Nutr 2019;7:2068-82. 10.1002/fsn3.1048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Yi R, Peng P, Zhang J, et al. Lactobacillus plantarum CQPC02-Fermented Soybean Milk Improves Loperamide-Induced Constipation in Mice. J Med Food 2019;22:1208-21. 10.1089/jmf.2019.4467 [DOI] [PubMed] [Google Scholar]
- 53.Eor JY, Tan PL, Lim SM, et al. Laxative effect of probiotic chocolate on loperamide-induced constipation in rats. Food Res Int 2019;116:1173-82. 10.1016/j.foodres.2018.09.062 [DOI] [PubMed] [Google Scholar]
- 54.Botelho PB, Ferreira MVR, Araújo AM, et al. Effect of multispecies probiotic on gut microbiota composition in individuals with intestinal constipation: A double-blind, placebo-controlled randomized trial. Nutrition 2020;78:110890. 10.1016/j.nut.2020.110890 [DOI] [PubMed] [Google Scholar]
- 55.Kim SE, Choi SC, Park KS, et al. Change of Fecal Flora and Effectiveness of the Short-term VSL#3 Probiotic Treatment in Patients With Functional Constipation. J Neurogastroenterol Motil 2015;21:111-20. 10.5056/jnm14048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kunze WA, Mao YK, Wang B, et al. Lactobacillus reuteri enhances excitability of colonic AH neurons by inhibiting calcium-dependent potassium channel opening. J Cell Mol Med 2009;13:2261-70. 10.1111/j.1582-4934.2009.00686.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Martino ME, Bayjanov JR, Caffrey BE, et al. Nomadic lifestyle of Lactobacillus plantarum revealed by comparative genomics of 54 strains isolated from different habitats. Environ Microbiol 2016;18:4974-89. 10.1111/1462-2920.13455 [DOI] [PubMed] [Google Scholar]
- 58.Lee KM, Paik CN, Chung WC, et al. Breath methane positivity is more common and higher in patients with objectively proven delayed transit constipation. Eur J Gastroenterol Hepatol 2013;25:726-32. 10.1097/MEG.0b013e32835eb916 [DOI] [PubMed] [Google Scholar]
- 59.Suri J, Kataria R, Malik Z, et al. Elevated methane levels in small intestinal bacterial overgrowth suggests delayed small bowel and colonic transit. Medicine (Baltimore) 2018;97:e10554. 10.1097/MD.0000000000010554 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Jahng J, Jung IS, Choi EJ, et al. The effects of methane and hydrogen gases produced by enteric bacteria on ileal motility and colonic transit time. Neurogastroenterol Motil 2012;24:185-90, e92. [DOI] [PubMed]
- 61.Ge X, Pan J, Liu Y, et al. Intestinal Crosstalk between Microbiota and Serotonin and its Impact on Gut Motility. Curr Pharm Biotechnol 2018;19:190-5. 10.2174/1389201019666180528094202 [DOI] [PubMed] [Google Scholar]
- 62.Cong M, Jiang Q, Xu X, et al. The complete genome sequence of Exiguobacterium arabatum W-01 reveals potential probiotic functions. Microbiologyopen 2017;6:e00496. 10.1002/mbo3.496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Nakjang S, Ndeh DA, Wipat A, et al. A novel extracellular metallopeptidase domain shared by animal host-associated mutualistic and pathogenic microbes. PLoS One 2012;7:e30287. 10.1371/journal.pone.0030287 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Volokh O, Klimenko N, Berezhnaya Y, et al. Human Gut Microbiome Response Induced by Fermented Dairy Product Intake in Healthy Volunteers. Nutrients 2019;11:547. 10.3390/nu11030547 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The article’s supplementary files as