

Abstract
Colorectal cancer (CRC) is the second cause of cancer death and the third most frequently diagnosed cancer. Besides the lifestyle, genetic and epigenetic alterations, and environmental factors, gut microbiota also plays a vital role in CRC development. The interruption of the commensal relationship between gut microbiota and the host could lead to an imbalance in the bacteria population, in which the pathogenic bacteria become the predominant population in the gut. Different therapeutic strategies have been developed to modify the gut immune system, prevent pathogen colonization, and alter the activity and composition of gut microbiota, such as prebiotics, probiotics, postbiotics, antibiotics, and fecal microbiota transplantation (FMT). Even though the employed strategies exhibit promising results, their translation into the clinic requires evaluating potential implications and risks, as well as assessment of their long-term effects. This study was set to review the gut microbiota imbalances and their relationship with CRC and their effects on CRC therapy, including chemotherapy and immunotherapy. More importantly, we reviewed the strategies that have been used to modulate gut microbiota, their impact on the treatment of CRC, and the challenges of each strategy.
Keywords: Colorectal cancer, Gut microbiota, Probiotics, Fecal microbiota transplantation
Background
According to GLOBOCAN estimate, colorectal cancer (CRC), with around 1.9 million new cases and 935,000 deaths, stands in the third and second ranks of incidence and mortality caused by cancer, respectively, in 2020 [1]. CRC can be regarded as a sign of socio-economic development. In countries undergoing significant transitions, the incidence tends to steadily increase with the increase in Human Development Index (HDI) [2, 3]. The increase in previously low-risk and low HDI countries may reflect changes in diet and lifestyle factors, such as the shift to a higher intake of animal-based foods and a more sedentary lifestyle, resulting in a reduction in physical activity and being overweight, which are associated with the risk of CRC [4]. Besides genetic, epigenetic, and environmental factors, the gut microbiome and its related parameters, including immunity, metabolism, and interaction with the host, determine the host’s health and disease [5].
Gut microbiota, the microorganisms that live in the gastrointestinal (GI) tract, not only include almost 3 × 1013 bacteria but also fungi, viruses, protists, and archaea [6]. Although the gut microbiota usually has a commensal relationship with its host, this relationship may be interrupted by changes in bacterial composition. The modulation occurs through an imbalance in the bacterial population, in which pathogenic bacteria replace harmless commensal bacteria. These pathogenic bacteria can invade the intestinal wall, cause inflammation, and induce carcinogenic signals and metabolites, leading to damage to the host [7]. Therefore, these bacteria may promote colon tumors; however, it is not clear whether these bacteria cause or the result of CRC [8]. With the increasing understanding of how the gut microbiota promotes cancer and affects the outcome of treatment, regulation of the gut microbiota aimed at restoring homeostasis of the gut microbiota has become a potential strategy for the prevention and treatment of CRC. This review aimed to summarize the role of the gut microbiota in CRC development and the different strategies applied to regulate and modulate gut microbiota.
Gut microbiota in CRC
Although gut microbiota plays pivotal roles in gut homeostasis by various mechanisms, including acting as a defensive barrier against infection, developing the mucosal immune system, and involving in metabolic functions, there is growing evidence from large metagenomic studies that imbalance in human gut microbiota links with colorectal carcinogenesis [9]. In a study, Zhao et al. reviewed the cohort studies determining dysbiosis and the differences between the cancerous tissues and adjacent non-cancerous tissue of CRC patients and revealed the heterogeneity of microbiome in CRC [10]. Table 1 reviews some studies to describe the main differences between the phyla, genus, and species between healthy papulation and CRC patients.
Table 1.
The differences between phyla, genus, and species between healthy papulation and CRC patients
| Healthy sample | Tumor sample | |||||
|---|---|---|---|---|---|---|
| Phylum | Genus | Species | Phylum | Genus | Species | Refs. |
|
Firmicutes Bacteriodetes Actinobacteria |
Faecalibacterium Prevotella |
B. vulgatus F. prausnitzii |
Fusobacteria Proteobacteria Spirochaetes |
Fusobacterium Treponema |
B. fragilis F. nucleatum |
[11] |
| Firmicutes |
Agathobacter Anaerostipes Butyricicoccus A Butyrivibrio A CAG-41 Eubacterium G Eubacterium R Faecalibacterium Lachnospira GCA-900066135 TF01-11 |
A. hadrus B. catenulatum F. saccharivorans F. prausnitzii A. rectalis A. faecis |
Bacteroidota Desulfobacterota Fusobacteriota |
Anaerotignum Bilophila Bulleidia Flavonifractor Gemella Intestinimonas Parvimonas Peptostreptococcus Porphyromonas Prevotella Ruthenibacterium |
E. coli D P. distasonis B. fragilis A. finegoldii A. onderdonkii A. muciniphila B. thetaiotaomicron M. torques R. gnavus Porphyromonas species |
[12] |
|
Bacteriodetes Firmicutes Actinobacteria |
Faecalibacterium Prevotella |
NA |
Proteobacteria Fusobacteria |
Bacteroides Escherichia Sutterella |
NA | [13] |
|
Firmicutes Actinobacteria Bacteriodetes |
Bacteroides Blautia Faecalibacterium |
F. prausnitzii B. faecis S. termitidis A. shahii B. uniformis |
Fusobacteriota Proteobacteria |
Fusobacterium Prevotella Parvimonas |
F. nucleatum C. Koseri T. socranskii L. trevisanii |
[14] |
|
Firmicutes Actinobacteria |
Bacillus Lactococcus Acinetobacter Pseudomonas Parabacteroides |
NA |
Bacteriodetes Proteobacteria Fusobacteria |
Fusobacterium Prevotella Alloprevotella Porphyromonas Peptostreptococcus Parvimonas |
NA | [15] |
|
Firmicutes Euryarchaeota Spirochaetes |
Prevotella Faecalibacterium Lactobacillus Parabacteroides |
NA |
Fusobacteria Proteobacteria Bacteriodetes |
Bacteroides Fusobacterium Bifidobacterium Streptococcus Halomonas Sphingomonas |
NA | [16] |
There is an abundance of Helicobacter pylori, Fusobacterium nucleatum, Escherichia coli, Bacteroides fragilis, Peptostreptococcus anaerobius, Helicobacter hepaticus, Porphyromonas gingivalis, Enterococcus faecalis, and Streptococcus gallolyticus has been related to CRC development [17]. Table 2 summarizes the abundance of some bacteria in patients with CRC. Certain species can develop CRC through specific and a variety of mechanisms. Mainly, three mechanisms have been identified for tumor-promoting activities of microbiota: pathogenic bacterial virulence factors/toxins, bacterial metabolic products, and immune modulation/reaction [18]. For example, colibactin or typhoid toxin secreted by E. coli or Salmonella, respectively, produces pro-inflammatory cytokines and bacterial adherence. Also, B. fragilis and F. nucleatum provide a favorable microenvironment for inhibitory immune cells [19]. Figure 1 indicates the main pathogenic mechanisms in CRC that involve the gut microbiome.
Table 2.
Various bacteria abundance in patients with CRC
| Bacteria | Detection method | Sample (n) | Positive percentage | Sample type | Affiliation | Refs. |
|---|---|---|---|---|---|---|
| H. pylori | Multiplex serology | 4063 | 41% | Serum | USA | [20] |
| H. pylori | ELISA | 1712 | 46.1% | Serum | Germany | [21] |
| H. pylori | Multiplex serology | 1488 | 90% | Serum | Spain | [22] |
| F. nucleatum | qPCR | 160 | 66.9% | FFPE tissue | Korea | [23] |
| F. nucleatum | qPCR | 100 | 75% | Frozen tissue | Japan | [24] |
| F. nucleatum | qPCR | 39 | 82.1% | Tissue | Korea | [25] |
| E. coli | PCR | 31 | 90% | Tissue | Germany | [26] |
| E. coli | PCR | 48 | 16.7% | Tissue | Malaysia | [27] |
| E. coli | PCR | 48 | 83% | Tissue | Iran | [28] |
| B. fragilis | PCR | 49 | 88.5% | Mucosal tissue | USA | [29] |
| B. fragilis | PCpR | 60 | 58.3% | Stool | Iran | [30] |
| P. anaerobius | qPCR | 154 | NA | Mucosa/Stool | China | [31] |
| P. gingivalis | qPCR | 31 | 32.2% | Tissue | China | [32] |
| P. gingivalis | PCR | 71 | 76% | Saliva | Turkey | [33] |
| E. faecalis | PCR | 20 | NA | Stool | India | [34] |
| E. faecalis | qPCR | 9 | 22.2% | Stool | Italy | [35] |
| S. gallolyticus | qPCR | 148 | 74% | Tissue | USA | [36] |
| S. gallolyticus | qPCR | 190 | 3.2% | Tissue | Spain | [37] |
ELISA enzyme-linked immunosorbent assay, qPCR quantitative real‐time polymerase chain reaction, FFPE formalin‐fixed paraffin‐embedded
Fig. 1.
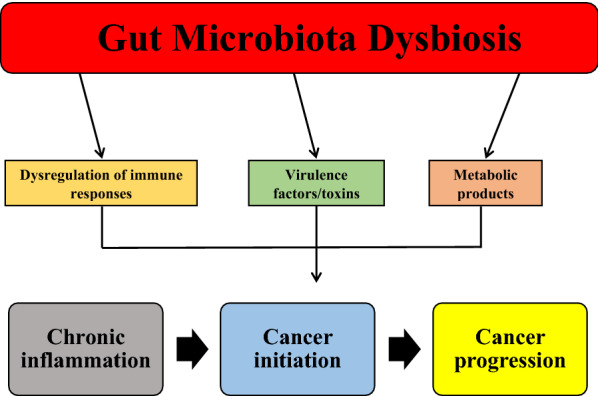
Gut microbiota dysbiosis and its relationship with CRC. Dysbiosis of gut microbiota and increasing the abundance of pathogenic microbiota could promote chronic inflammation and, subsequently, cancer initiation and progression through three mechanisms, including dysregulation of immune responses, virulence factors/toxins, and metabolic products
Helicobacter pylori
It has been shown that infection with H. pylori is associated with an increased risk of CRC. For instance, Teimoorian et al. found that H. pylori infection was considerably more prevalent in patients with adenomatous polyps and colon cancer than in healthy subjects [38]. A meta-analysis study, including 47 studies with 17,416 CRC cases and 55,811 control cases, revealed a positive association between infection with H. pylori and increased risk of CRC, although this association was related to the region of the study [39]. As virulence factors in some H. pylori, cytotoxin-associated gene A (CagA) and vacuolating cytotoxin A (VacA), can promote and activate inflammation pathways [40]. Furthermore, there is evidence that direct and indirect production of reactive oxygen species (ROS) and reactive nitrogen species (RNS) by some strains of H. pylori participate in colon tumorigenesis [41]. There are also other hypotheses about the indirect contribution of H. pylori to CRC carcinogenesis, including changes in the colonization of the intestine with other bacteria and an increase in the gastrin release [42].
Fusobacterium nucleatum
In a meta-analysis study, Gethings-Behncke et al. found a higher abundance of F. nucleatum in CRC patients' tissue and fecal samples. They also indicated that the high F. nucleatum abundance in CRC patients was associated with poorer overall survival [43]. Two other meta-analysis studies confirmed that intestinal and fecal F. nucleatum could be a valuable diagnostic marker for CRC [44, 45]. Various mechanisms are involved in CRC tumorigenesis of F. nucleatum, including virulence factors, metabolism products, immune modulation, and miRNAs [46]. For instance, the Fap2 protein of bacteria mediates F. nucleatum enrichment during CRC by binding to tumor-expressed D‐galactose‐β(1–3)‐N‐acetyl‐D‐galactosamine (Gal‐GalNAc) [47]. FadA is another virulence factor of F. nucleatum that activates the β‐catenin pathway by binding to E‐cadherin, leading to an increase in the levels of Wnt7b, NFκB, cyclin D1, and Myc [48]. Moreover, F. nucleatum enhances angiogenesis by increasing the expression of CD31 and CD34 as adhesion molecules and vascular endothelial growth factor receptors 1 and 2 (VEGFR1 and VEGFR2) [49]. Angiogenesis is a hallmark of cancer that provides oxygen and nutrients for tumor cells to guarantee their growth [50, 51].
Escherichia coli
There is accumulating evidence that E. coli are frequently colonizing in CRC lesions and adjacent epithelium [52, 53]. For instance, Iyadorai et al. found that pks + E. coli is more frequent in patients with CRC than in healthy cases and the pks + E. coli strains play an important role in the initiation and development of tumors [27]. It has been shown that E. coli could induce CRC in IL-10-deficient mice, proposing that inflammation is pivotal for tumorigenesis [54]. In addition, pathogenic E. coli can synthesize toxins called cyclomodulins, including cycle inhibiting factor, cytotoxic necrotizing factor (CNF), colibactin, and cytolethal distending toxins (CDT), which are genotoxin or interfere with the cell cycle [55, 56]. E. coli also promotes the survival of tumor cells by inducing macrophage inhibitory cytokine 1 (MIC-1), leading to an increase in the expression of transforming growth factor β-activated kinase 1 (TAK1) and subsequently RhoA GTPase after pathogen infection, and sustained COX-2 expression [57, 58].
Bacteroides fragilis
The B. fragilis is categorized into two subtypes, including enterotoxigenic B. fragilis (ETBF) and nontoxigenic B. fragilis (NTBF) [59]. It has been reported that ETBF was considerably more abundant in precancerous and cancerous lesions of CRC than in healthy controls and mucosal B. fragilis toxin (BFT) could be considered a risk factor and screening marker for CRC development [60]. BFT is a zinc-dependent metalloprotease toxin involved in the transduction of epithelial cell signals in the colon, such as Wnt, NFκB, and MAPK pathways. It induces inflammatory mediators' production and facilitates CRC development [61]. Also, ETBF could induce colitis and tumor formation in an IL-17-dependent manner in MinApc±mice through STAT3 activation [62]. The activation of these pathways results in the recruitment of immature myeloid cells by inducing CXC chemokines, leading to the creation of a pro-inflammatory environment [63].
Peptostreptococcus anaerobius
Metagenomic analysis of mucosal and stool samples from patients with CRC discovered that P. anaerobius is another gut bacteria enriched in CRC [64, 65]. Tsoi et al. reported that mice exposed to P. anaerobius showed a higher incidence of colon dysplasia than those not given the bacteria (63% versus 8.3%), and colon cells exhibited considerably higher levels of proliferation when exposed to P. anaerobius compared with control cells. Mechanistically, P. anaerobius promotes tumorigenesis by activating toll-like receptors 2 and 4 (TLR2 and TLR4) to enhance the intracellular levels of ROS, which increase cholesterol synthesis and subsequently cell proliferation [31]. In another study, Long et al. showed that P. anaerobius preferentially attaches to CRC cell lines compared to normal intestinal epithelial cells. They identified putative cell wall binding repeat 2 (PCWBR2), a surface protein of P. anaerobius, responsible for this adherence. The PCWBR2 directly binds to integrin α2/β1, a frequently overexpressed receptor in CRC tumors, which promotes the activation of the PI3K-Akt signaling pathway in CRC cells, resulting in activation of NFκB and increase of cell proliferation. The activation of NFκB increases pro-inflammatory responses and the levels of IL-10 and IFN-γ P. anaerobius-treated mice. Furthermore, P. anaerobius-treated mice exhibited a significant increase in the population of tumor-associated macrophages (TAMs), granulocytic tumor-associated neutrophils, and myeloid-derived suppressor cells, which are involved in chronic inflammation and tumor development [66].
Porphyromonas gingivalis
P. gingivalis is an oral microbiota that can be transmitted to the large intestine and contribute to the pathogenesis of various diseases [67]. It has been shown that oral administration of Prevotella intermedia and P. gingivalis in mice led to systemic inflammation, endotoxemia, disruption of the intestinal barrier, and intestinal dysbiosis [68]. Recently, Mu et al. investigated the role of P. gingivalis in CRC and its mechanism of action. They found that P. gingivalis could adhere and invade CRC cells, significantly enhance their proliferation, and increase the percentage of cells in the S-phase of the cell cycle. Mechanistically, P. gingivalis contributes to the proliferation of CRC cells by significantly activating the MAPK/ERK pathway [69]. In another study, Wang et al. demonstrated that oral gavage of P. gingivalis to ApcMin/+ mice promotes CRC tumorigenesis and modulates tumor microenvironment (TME) by preferentially increasing myeloid-derived immune cells, such as macrophages and DCs, and inducing a pro-inflammatory condition. They also indicated that P. gingivalis promotes CRC by activating nucleotide-binding oligomerization domain-like receptor family pyrin domain-containing 3 (NLRP3) inflammasome [32].
Enterococcus faecalis
There are controversial data about the role of E. faecalis in CRC: some studies exhibited no role or a protective role against CRC while others showed a harmful role [70]. For instance, De Almeida et al. found a decreased abundance of E. faecalis in the feces of CRC patients compared to healthy cases (22.2% versus 44.4%) [35]. Similarly, Viljoen et al. did not find a considerable clinical association between infection with E. faecalis and CRC [71]. On the other hand, Wang et al. indicated that E. faecalis contributes to inducing CRC via activating the Wnt/β-catenin pathway and inducing pluripotent transcription factors. They showed that the exposure of colonic epithelial cells to E. faecalis-infected macrophages promotes the initiation of CRC through chromosomal instability, gene mutation, and cell transformation [72].
Streptococcus gallolyticus
It has been shown that colonization of S. gallolyticus, previously called S. bovis, is significantly correlated with CRC development [73, 74]. A case–control study on 109 cases revealed that S. gallolyticus was significantly higher among colorectal neoplasia patients than in control cases (70% versus 32%) [75]. Mechanistically, S. gallolyticus antigen could promote cell proliferation and angiogenesis by inducing COX-2 expression [76]. In addition, the wall-extracted antigen of S. gallolyticus induces the production of inflammatory cytokines by binding to CRC cell lines [77].
Gut microbiota and chemotherapy implications
There is emerging evidence that host response to chemotherapy can be modulated with gut microbiota through various mechanisms, including immunomodulation, microbial translocation, educed ecological diversity, enzymatic degradation, and metabolism [78].
Viaud et al. investigated the effect of cyclophosphamide (CTX) and doxorubicin on the composition of small intestine microbiota in mouse models. They found that treatment with both chemotherapy agents led to shortening the small intestine's villi, discontinuity in the intestinal epithelial barrier, accumulation of inflammatory cells, and translocation of commensal bacteria into secondary lymphoid organs. Although treatment with CTX did not reduce the total bacterial counts in mice's small intestine after seven days, the abundance of enterococci and lactobacilli showed a reduction. In addition, some Gram-positive bacteria, such as Enterococcus hirae, Lactobacillus murinus, and Lactobacillus johnsonii, stimulated the generation of pathogenic” T helper 17 (pTh17) and type 1 T helper (Th1) cells, whereas killing these bacteria with antibiotics reduced pTh17 responses and enhanced tumor resistance to CTX [79]. In another study, Daillère et al. demonstrated that Barnesiella intestinihominis and Enterococcus hirae are involved in the host response to CTX. They found that B. intestinihominis accumulates in the colon and induces intratumoral re-instating of IFN-γ-producing γδT cells, whereas E. hirae translocates to secondary lymphoid organs from the small intestine and increases the ratio of CD8/Treg in the TME. The colonic B. intestinihominis acts as an orchestrator of CTX effects and E. hirae could restore CTX-induced anti-tumor effects [80]. Yu et al. demonstrated that F. nucleatum could promote resistance of CRC cells to 5-fluorouracil (5-FU) and oxaliplatin by activating autophagy. From a mechanistic view, F. nucleatum involved TLR4 and MYD88, downregulating the expression of miR-4802 and miR-18a*. The reduction in miR-4802 and miR-18a* levels leads to the upregulation of ATG7 and ULK1, respectively, resulting in the activation of autophagy [81]. Another example of gut microbiota and modulation of chemotherapy is the control of side effects and metabolism of irinotecan, a pro-drug that is activated in the form of SN-38, a topoisomerase I inhibitor. The host liver enzymes could glucuronide SN-38 and convert it to an inactivate form (SN-38G). After reaching the intestine, SN-38G is hydrolyzed back to SN-38 through bacterial β-glucuronidase enzymes, which leads to severe diarrhea and intestinal damage [82].
Gut microbiota and immunotherapy implications
The cancer immunotherapy approach has become a promising therapeutic way to treat cancer in which the immune responses of the patient are elicited to exert anti-tumor effects. There is evidence that gut microbiota could modulate response to immunotherapy. It has been shown that specific gut bacteria could elevate tumor response in immunotherapy, including Eubacterium limosum, Alistipes shahii, B. fragilis, Akkermansia muciniphila, and Faecalibacterium spp [83–87]. For instance, the results of a study revealed that a significant increase in the abundance of Bifidobacterium could elevate the levels of tumor-infiltrating lymphocytes (TILs) and facilitate tumor response to treatment with programmed cell death protein 1 ligand 1 (PD-L1) antibody [88]. Recently, Shi et al. indicated that gut microbiota could affect the efficacy of anti-CD47 immunotherapy by changing TME and gut immunity. Bifidobacterium could accumulate within TME and convert non-responder tumors to responder ones to immunotherapy with anti-CD47 in an IFN- and stimulator of interferon genes (STING)-dependent manner [89]. In another study, Lv et al. found that the efficacy of PD-1 blockade could be enhanced through gut microbiota remodeling in CRC with microsatellite stability. They found that a traditional Chinese drug, Gegen Qinlian decoction (GQD), could enhance the efficacy of immunotherapy with anti-PD-1 antibody and inhibit tumor growth in vivo. The combination of GQD and anti-PD-1 modulates gut microbiota, increasing Bacteroides acidifaciens and Bacteroidales S24-7 family. The combinational therapy also remarkably increased CD8 + T cells proportion in tumor tissues and peripheral blood and elevated the expression of IFN-γ and IL-2. Mechanistically, the combination of GQD and anti-PD-1 increased the metabolism of glycerophospholipid and sphingolipid pathways [90]. It has been shown that both metabolites could act as biomarkers in monitoring CRC patients [91, 92].
Strategies of gut microbiota modulation
There is evidence that gut microbiota and microbial-secreted metabolites could be targeted as a therapeutic strategy in combating CRC. For instance, Bhalla et al. showed that microbial metabolites, such as folate, could suppress CRC cells' viability; thus, modulation of gut microbiota to produce anti-cancer metabolites could be used to treat CRC [93]. The following section will consider the strategies that have been applied to modulate gut microbiota as a therapeutic strategy.
Probiotics
Probiotics are microorganisms, including bacteria, yeasts, and molds, that can improve the host health when delivered in an adequate quantity. Lactobacillus, Bacillus, Bifidobacterium, Streptococcus, and Enterococcus are the most used bacterial genera as probiotics [94]. Probiotics affect the gut in the prevention and treatment of CRC through three main mechanisms: 1) immunomodulation, 2) inhibition of pathogenic bacteria colonization, and 3) enhancement of the gut barrier functions. Figure 2 represents the strategies applied for gut microbiota modulation and the mechanisms of action of each strategy.
Fig. 2.

Strategies applied for modulating gut microbiota in CRC and the mechanisms of action of each strategy. Probiotics: Probiotics exert their effects on gut microbiota composition through immunomodulation, inhibition of pathogenic bacteria colonization, and enhancement of the gut barrier functions. Prebiotics: They act as gut microbiota modulatory elements through direct uptake by the intestine and exerting anti-inflammatory activities, prevention of the colonization of pathogens by interacting with them, fermentation by intestinal microbiota, and stimulation of beneficial gut bacteria. Postbiotics: They exert their tumoricidal functions through selective inhibiting tumor cells and protecting intestinal epithelium by inhibiting apoptosis in epithelial cells and increasing IgA secretion. Antibiotics: Antibiotics could deplete the intestine from harmful bacteria and preserve intestinal epithelium. FMT: This strategy helps restore the diversity of microbiota in the gut
Dysbiosis conditions could activate pathways and transcriptional factors, such as MAPK and NF-κB, that increase the production of nitric oxide (NO) and IL-8 as pro-inflammatory cytokines, leading to the occurrence of inflammatory bowel disease (IBD) and CRC. There is accumulating evidence that probiotics administration re-establishes the balance of gut microbiota by increasing the secretion of anti-inflammatory cytokines, such as TGF-β2 and IL-10, through regulatory T (Treg) cells [95]. For example, Bifidobacterium breve and Bifidobacterium infantis could activate colonic dendritic cells (DC), resulting in the expression of Foxp3+ Treg and IL-10 producing type 1 Treg (Tr1) [96, 97]. Lee et al. demonstrated that Bacillus polyfermenticus containing probiotics properties exhibited an anti-proliferative effect on CRC cell lines, HT-29 and LoVo, by reducing the levels of pro-inflammatory cytokines, including TNF-α, NO, and COX-2 [98]. On the other hand, it has been shown that probiotics bacteria could induce pro-inflammatory responses. Hradicka et al. studied the immunomodulatory effects of six lactobacilli containing probiotic properties in an in vitro model of macrophage cells and their anti-tumor properties in a rat model of CRC. They indicated that the bacteria could induce the release of pro- and anti-inflammatory cytokines, including IL-1β, TNF-α, IL-18, and IL-23, in co-culture of lactobacilli with M1- and M2-like macrophages, whereas the bacteria oral administration led to a decrease in tumor multiplicity, numbers, and volume, as well as restore of colon length and increase in the production of IL-18 [99]. In another study, a clinical trial found that oral administration of probiotic Saccharomyces boulardii could downregulate the levels of both pro- and anti-inflammatory cytokines, including IL-10, IL-23A, and IL-1β, while the incidence of infectious complications in patients receiving probiotic was 13.3% compared to 38.8% in the control group [100].
The other beneficial property of probiotics is the inhibition of colonization of pathogenic bacteria that degrade the gut and release toxic compounds. Probiotics could inhibit the proliferation of detrimental microorganisms by lowering luminal pH, producing anti-microbial peptides, and reducing pro-tumorigenic enzymes [101]. For instance, Fayol-Messaoudi et al. demonstrated that probiotic strains of Lactobacillus could inhibit the growth of the pathogenic bacteria, Salmonella enterica, by producing non-lactic acid and lactic acid substances and subsequently lowering pH [102]. In addition, there is evidence that probiotics could prevent colon infection with pathogenic bacteria, including Staphylococcus aureus and Clostridium difficile [103, 104]. Probiotics also could inhibit the adhesion of pathogens to the intestine. Collado et al. found that probiotic strains can displace adhered pathogens or inhibit the adhesion of pathogens to intestinal mucus, while different combinations of probiotics showed an enhanced percentage of adherence inhibition [105]. In another study, Behbahani et al. revealed the adhesion ability of the probiotic L. plantarum strain L15 to the intestinal cells. This strain exhibited anti-adherence activities, such as competition, inhibition, and replacement features, against E.coli [106].
Intestinal epithelium acts as a physical barrier and protects the lamina environment from pathogenic toxins, stress factors, and bacteria. The barrier function has three components: tight junction (TJ), adhesion protein junction (AJ), and desmosomes. It has been shown that disruption of this physical barrier results in the development of IBD and IBD-associated CRC, as well as an acceleration in tumor invasion and metastasis [107, 108]. There is evidence that some probiotic strains improve the function of the gut barrier. A clinical trial study revealed that oral administration of probiotics both pre-operatively and post-operatively in CRC patients undergoing surgery could increase transepithelial resistance and upregulate the expression of TJ. Patients receiving probiotics had increased bacterial variety in fecal and decreased enteropathogenic bacteria in the blood [109]. Liu et al. conducted a meta-analysis study to investigate the effect of probiotics on the gut barrier in CRC patients after operation. Assessment of 17 studies demonstrated that probiotics administration could protect the physical and biological barrier in the intestine by increasing intestinal permeability, occludin levels, secretory immunoglobulin A (SIgA) levels, and decreasing bacterial translocation [110]. In another study, the administration of probiotic strains, including L. acidophilus, B. bifidum, and B. infantum (LBB regimen), could inhibit CRC development by reducing tumor count, tumor size, and tumor incidence. LBB treatment also altered the gut microbiota and decreased the abundance of pathogenic bacteria. Furthermore, the LBB regimen enhanced the integrity of the intestinal epithelial and mucosal barrier by increasing the expression of occludin, zonula occludens-1 (ZO-1), and mucin 2 (MUC2). LBB treatment also prevented CRC via enhancing TLR2 expression and downregulation of TLR4, COX2, caspase-3, and β-catenin. These results suggested that the anti-cancer effects of probiotic LBB treatment was due to TLR2-dependent intestinal barrier enhancement and inhibition of inflammation, apoptosis, and β-catenin pathway [111]. Table 3 summarizes the application of probiotics in the management of CRC patients.
Table 3.
Probiotics administration for the management of CRC patients
| # of patients | Intervention | Duration | Outcome | Refs. |
|---|---|---|---|---|
| 27 | 6 viable microorganisms of Lactobacillus and bifidobacteria | Twice daily 107 mg (orally) probiotics for 6 months | Reduced the level of pro-inflammatory cytokines, including TNF-α, IL-6, IL-10, IL-12, IL-17A, IL-17C and IL-22 | [112] |
| 57 | 2 mg Enterococcus faecalis T110, 10 mg Clostridium butyricum TO-A, and 10 mg Bacillus mesentericus TO-A | Daily (six tablets/day) for 15 days | Reduced the incidence of post-operative complications, including superficial incisional infections, time of meal intake, and time of flatus | [113] |
| 8 | 1.4 × 1010 CFUs Bifidobacterium lactis Bl-04, 7 × 109 CFUs Lactobacillus acidophilus NCFM, and 0.63 g inulin | Daily (two tablets/day) 8–78 days |
- Increased bacterial diversity - Increased the abundance of butyrate-producing bacteria, including Faecalibacterium and Clostridiales spp - Reduced CRC-associated genera, including Fusobacterium and Peptostreptococcus |
[114] |
| 84 | Lactobacillus acidophilus LA-5 (1.75 × 109 cfu), Lactobacillus plantarum (0.5 × 109 cfu), Bifidobacterium lactis BB-12 (1.75 × 109 cfu), and Saccharomyces boulardii (1.5 × 109 cfu) | One day before operation and continuing for another 15 days post-operatively |
- Decreased the rate of all postoperative major complications, including pneumonia, anastomotic leakage, and surgical site infections - Shortened the time until hospital discharge |
[115] |
| 98 | Lactobacillus rhamnosus GG supplementation (twice daily at a dose of 1–2 × 1010) and fiber (11 g guar gum per day) during chemotherapy | For 24 weeks | Reduced the frequency of diarrhea and abdominal discomfort | [116] |
| 70 | 6 viable microorganisms of Lactobacillus and bifidobacteria plus omega-3 fatty acid at a dose of 2 g |
- Probiotics for 4 weeks - Omega-3 fatty acid for 8 weeks |
- Improved the overall quality of life - Alleviated certain side effects of chemotherapy - Reduced inflammatory biomarkers, including IL-6 |
[117] |
| 28 |
2 × 109 cfu Lactobacillus rhamnosus R0011 and Lactobacillus acidophilus R0052 |
For 12 weeks |
- Decreased irritable bowel symptoms - Improved cancer-related quality of life |
[118] |
Prebiotics
Prebiotics are “selectively fermented ingredients that cause specific changes in the gastrointestinal microbiota's composition and/or activities that confers benefits upon host well-being and health” [119]. The polyphenols, polyunsaturated fatty acids (PUFAs), and carbohydrates, including galactooligosaccharides (GOS), xylooligosaccharides (XOS), fructooligosaccharides (FOS), fructans, and inulin, possess prebiotic properties. Prebiotics exert their functions through various mechanisms: (1) stimulation of beneficial gut bacteria, (2) fermentation through intestinal microbiota, (3) direct uptake by the intestine and exerting anti-inflammatory activities, and (4) prevention of the colonization of pathogens by interacting with them [5].
It has been shown that prebiotic administration could enhance the abundance of probiotics, including Akkermansia, Rosebura, Ruminococcus, and Faecalibacterium [120, 121]. Zheng et al. prepared prebiotics-encapsulated probiotic spores (spores-dex) in which C. butyricum (as a probiotic) and chemically modified dextran (as a prebiotic) were combined, and their anti-cancer efficacy was assessed in colon tumor models. They demonstrated the enrichment of colon cancers with spores-dex following oral administration. The fermentation of dextran in the lesions by C. butyricum led to the production of short-chain fatty acids (SCFAs) with anti-cancer activities. Furthermore, spores-dex could increase SCFA-producing bacteria, including Roseburia and Eubacterium, and significantly inhibit tumor growth. SCFA-producing bacteria contribute to tumor inhibition by building a tumor-suppressing microenvironment in the intestine [122]. As SCFAs, butyrate is the main energy fuel for colonocytes, while acetate and propionate are metabolized by muscle and liver for energy generation and gluconeogenesis [123]. Moreover, butyrate could induce CRC apoptosis, modulate oxidative stress, enhance epithelial barrier, and downregulate inflammation [124]. In addition to their histone deacetylase inhibitory and intracellular metabolism activities, SCFAs exert most of their functions through G-protein coupled receptors (GPCRs) in the intestine, named GPR43 (FFA2), GPR41 (FFA3), and GPR109A. For instance, the binding of butyrate, acetate, and propionate to FFA2 in the colon epithelium triggers signaling cascades, which leads to cell cycle arrest, promoting apoptosis, and inhibition of inflammation. The expression of FFA2 enhances the growth of the Bifidobacterium family and inhibits the Prevotellaceae family and H. hepaticus. On the other hand, the butytae-producing Butyricicoccus pullicaecorum exhibits anti-tumorigenesis function by increasing the expression of FFA2 [125].
Prebiotics also interfere with pathogenic bacteria's adhesion to the epithelial cells and the intestine. For instance, Ribeiro et al. showed that olive pomace powders with prebiotic properties not only promote the production of SCFAs by microbiota but also exhibit anti-oxidant anti-adhesive activities against pathogens. They demonstrated that pulp-enriched powder, as the primary source of insoluble dietary fiber, inhibited the adhesion of B. cereus and L. monocytogenes up to 22% and 20%, respectively [126]. In another study, Leong et al. investigated the prebiotic properties of oligosaccharides in goats’ milk-based infant formulas and their mechanisms of action. They reported that the prebiotic oligosaccharides could remarkably enhance the growth of lactobacilli and bifidobacteria and reduce S. typhimurium and E. coli NCTC 10,418 adhesion to CRC cell line Caco-2 [127]. Because of structural similarities between the oligosaccharides in goats’ milk and carbohydrates on the gut surface, these oligosaccharides, especially fucosylated and sialylated ones, reduce the adhesion of pathogenic bacteria to intestinal epithelial cells via acting as soluble analogs for host cell receptors or changing such structures` expression [128, 129].
Postbiotics
Postbiotics are defined as cell fractions, inactivated microbial cells, and cell metabolites made with probiotic live cells during fermentation and contained various health benefits for the host. Since the postbiotics are present in the conditioned/supernatants medium of bacterial culture, they are safer than viable microorganisms. Postbiotics exert their anti-tumor activities by: 1) selectively inhibiting tumor cells and 2) protecting intestinal epithelium through the inhibition of apoptosis in epithelial cells and increasing IgA secretion [5].
It has been shown that postbiotic metabolites produced by some bacteria, such as L. plantarum, have cytotoxic and anti-proliferative effects on tumor cells, including CRC cells [130, 131]. Lee et al. assessed the tumoricidal function of probiotic cell-free supernatant treatment using L. fermentum against CRC cells in a 3D culture system. They found that the bacterial culture supernatant could induce apoptosis of CRC cell lines by upregulating Bax, Bak, Bid, Noxa, and caspase-3 [132]. In another study, Konishi et al. demonstrated the tumor-suppressive effect of ferrichrome in the culture supernatant of L. casei ATCC334 against colon cancer cells. Ferrichrome reduced the cancer cell viability by inducing c-jun N-terminal kinase (JNK)-mediated apoptosis. Furthermore, the tumoricidal effect of ferrichrome on cancer cells was greater than 5-FU and cisplatin, whereas the toxicity of ferrichrome against non-cancerous intestinal cells was less than the chemotherapy agents [133].
Postbiotics' other mechanism of action is associated with their ability to suppress intestinal inflammation and maintain the gut barrier's integrity. For instance, Izuddin et al. showed that supplementation of post-weaning lambs with postbiotic derived from L. plantarum RG14 led to ruminal epithelial growth, downregulation of pro-inflammatory cytokines, including TNF and IL-1β, and anti-inflammatory cytokine IL-10, and improvement of intestinal barrier integrity via upregulating tight junction protein 1 (TJP1), claudin-1 (CLDN-1), and CLDN-4 [134]. In another study, Gao et al. identified HM0539, a secreted protein, beneficial effects in the culture of L. rhamnosus GG. They found that HM0539 plays a protective role in maintaining the integrity of the intestinal barrier by increasing the expression of intestinal mucin and preventing intestinal barrier injury [135]. Another postbiotic protein of L. rhamnosus GG that affects the intestinal epithelium is p40 protein. To protect it from degradation, Yan and Polk used hydrogel-coating p40 and indicated that p40 protein promotes a protective immune response, reduces apoptosis of intestinal epithelial cells, and protects barrier function of the colon in an epithelial growth factor (EGF) receptor-dependent manner [136].
Antibiotics
It has been shown that the use of antibiotics has negative impacts on gut microbiota, such as reduction in the biodiversity of bacteria, selection of antibiotic-resistant organisms, and changes in metabolic functions, resulting in antibiotic-associated diarrhea as well as recurrence of C. difficile infection [137]. Although there is increasing evidence that antibiotics increase CRC risk [120, 121], they could also decrease the size and number of tumors by manipulating the gut microbiome [138, 139]. For example, DeStefano Shields et al. reported that treatment with cefoxitin antibiotic could completely and durably clear enterotoxigenic B. fragilis colonization in an intestinal neoplasia mice model [140].
Antibiotics also play a protective role in maintaining the mucosal barrier in the intestine. Since high consumption of red meat increases the risk of CRC, Ijssennagger et al. investigated the effect of heme, as a pigment and proxy of red meat, on gut microbiota and CRC development. They demonstrated that mice fed a diet containing heme exhibited damaged intestinal epithelium and hyperproliferation, leading to colon cancer, whereas heme + antibiotic regimen-received mice did not show epithelium damage and hyperproliferation. Mechanistically, hydrogen sulfide produced by microbial exposes the intestinal epithelium to cytotoxic heme via opening the mucus barrier. Antibiotics inhibit the production of microbial sulfide, thereby maintaining the integrity of the mucus barrier that prevents the induced hyperproliferation [141]. Despite the mentioned mechanisms, the relationship between gut microbiota, antibiotics, and CRC is very complicated and should be studied carefully to determine this relationship and its application in CRC therapy.
Fecal microbiota transplantation
Fecal microbiota transplantation (FMT), an interested and most innovative biotherapeutic method, is defined as transferring stool transplants from healthy individuals into patients. Although FMT is the approved method for treating C. difficile infection (CDI) [142], it also showed promising potential for treating obesity, IBD, diabetes, non-alcoholic fatty liver disease, and cardiovascular diseases. A prospective clinical trial study revealed that FMT could inhibit intestinal colonization of antibiotic-resistant bacteria (ARB) in patients with blood disorders. FMT led to completely (75%) and partially (80%) decolonization of ARB from patients [143]. FMT tries to restore gut microbiota diversity. It has been shown that FMT could restore microbial homeostasis by introducing a disease-free and healthy microbial population to an unbalanced community and act as a useful tool for ameliorating several GI disorders, including CDI, IBD, and irritable bowel syndrome [144]. Rosshart et al. showed that wild mice fecal transplantation to laboratory mice could promote fitness and improve resistance to colorectal tumorigenesis induction through mutagen/inflammation agents [145]. Interestingly, Wong et al. demonstrated that fecal microbiota from CRC patients increased tumor formation and reduced microbial abundance in conventional and germ-free mice given azoxymethane, as a carcinogen. The mice also increased the proportion of Th1 and Th17 cells and upregulated C-X-C motif chemokine receptor 1 (CXCR1), CXCR2, IL-17A, IL-22, and IL-23 [146]. In another study, Sobhani et al. indicated that the transplantation of fresh feces from CRC patients to germ-free mice could induce hypermethylation of several genes, similar to alteration patterns of CRC patients [147]. A clinical trial of FMT is recruiting for the treatment of metastatic CRC in non-responders to anti-PD-1 therapy (NCT04729322).
Conclusions
Although there are some review studies on gut microbiota modulation in CRC conditions, there is hardly any review that comprehensively works on the effects of gut microbiota on the efficiency and outcome of the therapeutic strategies. Herein, we tried to have a mechanistic overview of how gut microbiota modulation leads to CRC initiation and progression. Since gut dysbiosis commonly occurs in CRC carcinogenesis, various therapeutic strategies have been developed to alter the gut microbiota, including probiotics, prebiotics, postbiotics, antibiotics, and FMT. These strategies manage CRC treatment via different mechanisms, such as immunomodulation functions, maintaining gut barrier integrity, restoring gut dysbiosis, tumoricidal activities, colonization resistance, and producing anti-cancer products. Despite promising results, there are some concerns related to strategies applied for gut microbiota modulation. For instance, the presence of opportunistic pathogens or virulence factors and the spread of genes responsible for resistance in gut microbial populations are safety concerns of probiotic strategy. Furthermore, side effects, including abdominal pain, mild fever, diarrhea, flatulence, exhaustion, and fatigue, are the main challenges of FMT. Thus, an assessment of the risk–benefit potentials of each strategy in long-term trials and with a large sample should be included in studies to achieve reliable and comprehensive results.
Acknowledgements
Not applicable.
Abbreviations
- CRC
Colorectal cancer
- GI
Gastrointestinal
- ROS
Reactive oxygen species
- VEGFR
Vascular endothelial growth factor receptor
- BFT
B. fragilis Toxin
- TLR
Toll-like receptor
- CTX
Cyclophosphamide
- NO
Nitric oxide
- DC
Dendritic cell
- IBD
Inflammatory bowel disease
- TJ
Tight junction
- SCFAs
Short-chain fatty acids
- FMT
Fecal microbiota transplantation
- CDI
C. difficile Infection
Author contributions
Study conception and design: YW. Drafting of the manuscript: YW and HL. Critical revision: HL. Approve final version: All authors.
Funding
Not applicable.
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–249. doi: 10.3322/caac.21660. [DOI] [PubMed] [Google Scholar]
- 2.Bray F. Transitions in human development and the global cancer burden. World Cancer Rep. 2014;34.
- 3.Fidler MM, Soerjomataram I, Bray F. A global view on cancer incidence and national levels of the human development index. Int J Cancer. 2016;139:2436–2446. doi: 10.1002/ijc.30382. [DOI] [PubMed] [Google Scholar]
- 4.Siegel RL, Miller KD, Goding Sauer A, Fedewa SA, Butterly LF, Anderson JC, et al. Colorectal cancer statistics, 2020. CA Cancer J Clin. 2020;70:145–164. doi: 10.3322/caac.21601. [DOI] [PubMed] [Google Scholar]
- 5.Fong W, Li Q, Yu J. Gut microbiota modulation: a novel strategy for prevention and treatment of colorectal cancer. Oncogene. 2020;39:4925–4943. doi: 10.1038/s41388-020-1341-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Richard ML, Sokol H. The gut mycobiota: insights into analysis, environmental interactions and role in gastrointestinal diseases. Nat Rev Gastroenterol Hepatol. 2019;16:331–345. doi: 10.1038/s41575-019-0121-2. [DOI] [PubMed] [Google Scholar]
- 7.Illiano P, Brambilla R, Parolini C. The mutual interplay of gut microbiota, diet and human disease. FEBS J. 2020;287:833–855. doi: 10.1111/febs.15217. [DOI] [PubMed] [Google Scholar]
- 8.DeGruttola AK, Low D, Mizoguchi A, Mizoguchi E. Current understanding of dysbiosis in disease in human and animal models. Inflamm Bowel Dis. 2016;22:1137–1150. doi: 10.1097/MIB.0000000000000750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tilg H, Adolph TE, Gerner RR, Moschen AR. The intestinal microbiota in colorectal cancer. Cancer Cell. 2018;33:954–964. doi: 10.1016/j.ccell.2018.03.004. [DOI] [PubMed] [Google Scholar]
- 10.Zhao L, Cho WC, Nicolls MR. Colorectal cancer-associated microbiome patterns and signatures. Front Genet. 2021;12. [DOI] [PMC free article] [PubMed]
- 11.Choi S, Chung J, Cho M-L, Park D, Choi SS. Analysis of changes in microbiome compositions related to the prognosis of colorectal cancer patients based on tissue-derived 16S rRNA sequences. J Transl Med Springer. 2021;19:1–12. doi: 10.1186/s12967-021-03154-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Liu C, Li Z, Ding J, Zhen H, Fang M, Nie C. Species-level analysis of the human gut microbiome shows antibiotic resistance genes associated with colorectal cancer. Front Microbiol. 2021 doi: 10.3389/fmicb.2021.765291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Liu W, Zhang R, Shu R, Yu J, Li H, Long H, et al. Study of the relationship between microbiome and colorectal cancer susceptibility using 16SrRNA sequencing. Biomed Res Int. 2020;2020:1–17. doi: 10.1155/2020/7828392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kostic AD, Gevers D, Pedamallu CS, Michaud M, Duke F, Earl AM, et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012;22:292–298. doi: 10.1101/gr.126573.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gao R, Kong C, Huang L, Li H, Qu X, Liu Z, et al. Mucosa-associated microbiota signature in colorectal cancer. Eur J Clin Microbiol Infect Dis. 2017;36:2073–2083. doi: 10.1007/s10096-017-3026-4. [DOI] [PubMed] [Google Scholar]
- 16.Zhang Q, Zhao H, Wu D, Cao D, Ma W. A comprehensive analysis of the microbiota composition and gene expression in colorectal cancer. BMC Microbiol. 2020;20:1–11. doi: 10.1186/s12866-020-01938-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gagnière J, Raisch J, Veziant J, Barnich N, Bonnet R, Buc E, et al. Gut microbiota imbalance and colorectal cancer. World J Gastroenterol. 2016;22:501. doi: 10.3748/wjg.v22.i2.501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sun J, Kato I. Gut microbiota, inflammation and colorectal cancer. Genes Dis. 2016;3:130–143. doi: 10.1016/j.gendis.2016.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kim J, Lee HK. Potential role of the gut microbiome in colorectal cancer progression. Front Immunol. 2021;12:807648. doi: 10.3389/fimmu.2021.807648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Butt J, Varga MG, Blot WJ, Teras L, Visvanathan K, Le Marchand L, et al. Serologic response to Helicobacter pylori proteins associated with risk of colorectal cancer among diverse populations in the United States. Gastroenterology. 2019;156:175–186. doi: 10.1053/j.gastro.2018.09.054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Zhang Y, Hoffmeister M, Weck MN, Chang-Claude J, Brenner H. Helicobacter pylori infection and colorectal cancer risk: evidence from a large population-based case-control study in Germany. Am J Epidemiol. 2012;175:441–450. doi: 10.1093/aje/kwr331. [DOI] [PubMed] [Google Scholar]
- 22.de Larrea-Baz NF, Michel A, Romero B, Pérez-Gómez B, Moreno V, Martín V, et al. Helicobacter pylori antibody reactivities and colorectal cancer risk in a case-control study in spain. Front Microbiol. 2017 doi: 10.3389/fmicb.2017.00888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Park HE, Kim JH, Cho N-Y, Lee HS, Kang GH. Intratumoral Fusobacterium nucleatum abundance correlates with macrophage infiltration and CDKN2A methylation in microsatellite-unstable colorectal carcinoma. Virchows Arch. 2017;471:329–336. doi: 10.1007/s00428-017-2171-6. [DOI] [PubMed] [Google Scholar]
- 24.Yamaoka Y, Suehiro Y, Hashimoto S, Hoshida T, Fujimoto M, Watanabe M, et al. Fusobacterium nucleatum as a prognostic marker of colorectal cancer in a Japanese population. J Gastroenterol. 2018;53:517–524. doi: 10.1007/s00535-017-1382-6. [DOI] [PubMed] [Google Scholar]
- 25.Kim M, Lee S-T, Choi S, Lee H, Kwon SS, Byun JH, et al. Fusobacterium nucleatum in biopsied tissues from colorectal cancer patients and alcohol consumption in Korea. Sci Rep. 2020;10:1–10. doi: 10.1038/s41598-020-76467-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Swidsinski A, Khilkin M, Kerjaschki D, Schreiber S, Ortner M, Weber J, et al. Association between intraepithelial Escherichia coli and colorectal cancer. Gastroenterology. 1998;115:281–286. doi: 10.1016/s0016-5085(98)70194-5. [DOI] [PubMed] [Google Scholar]
- 27.Iyadorai T, Mariappan V, Vellasamy KM, Wanyiri JW, Roslani AC, Lee GK, et al. Prevalence and association of pks+ Escherichia coli with colorectal cancer in patients at the University Malaya Medical Centre, Malaysia. PLoS ONE. 2020;15:e0228217. doi: 10.1371/journal.pone.0228217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zarei O, Arabestan MR, Majlesi A, Mohammadi Y, Alikhani MY. Determination of virulence determinants of Escherichia coli strains isolated from patients with colorectal cancer compared to the healthy subjects. Gastroenterol Hepatol From Bed Bench. 2019;12:52. [PMC free article] [PubMed] [Google Scholar]
- 29.Boleij A, Hechenbleikner EM, Goodwin AC, Badani R, Stein EM, Lazarev MG, et al. The Bacteroides fragilis toxin gene is prevalent in the colon mucosa of colorectal cancer patients. Clin Infect Dis. 2015;60:208–215. doi: 10.1093/cid/ciu787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Haghi F, Goli E, Mirzaei B, Zeighami H. The association between fecal enterotoxigenic B. fragilis with colorectal cancer. BMC Cancer. 2019;19:1–4. doi: 10.1186/s12885-019-6115-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tsoi H, Chu ESH, Zhang X, Sheng J, Nakatsu G, Ng SC, et al. Peptostreptococcus anaerobius induces intracellular cholesterol biosynthesis in colon cells to induce proliferation and causes dysplasia in mice. Gastroenterology. 2017;152:1419–1433. doi: 10.1053/j.gastro.2017.01.009. [DOI] [PubMed] [Google Scholar]
- 32.Wang X, Jia Y, Wen L, Mu W, Wu X, Liu T, et al. Porphyromonas gingivalis promotes colorectal carcinoma by activating the hematopoietic NLRP3 inflammasome. Cancer Res. 2021;81:2745–2759. doi: 10.1158/0008-5472.CAN-20-3827. [DOI] [PubMed] [Google Scholar]
- 33.Guven DC, Dizdar O, Alp A, Akdogan Kittana FN, Karakoc D, Hamaloglu E, et al. Analysis of Fusobacterium nucleatum, Streptococcus gallolyticus and Porphyromonas gingivalis in saliva in colorectal cancer patients and healthy controls. J Clin Oncol. 2018;36:e15617. [Google Scholar]
- 34.Balamurugan R, Rajendiran E, George S, Samuel GV, Ramakrishna BS. Real-time polymerase chain reaction quantification of specific butyrate-producing bacteria, Desulfovibrio and Enterococcus faecalis in the feces of patients with colorectal cancer. J Gastroenterol Hepatol. 2008;23:1298–1303. doi: 10.1111/j.1440-1746.2008.05490.x. [DOI] [PubMed] [Google Scholar]
- 35.De Almeida CV, Lulli M, di Pilato V, Schiavone N, Russo E, Nannini G, et al. Differential responses of colorectal cancer cell lines to Enterococcus faecalis’ strains isolated from healthy donors and colorectal cancer patients. J Clin Med. 2019;8:388. doi: 10.3390/jcm8030388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kumar R, Herold JL, Schady D, Davis J, Kopetz S, Martinez-Moczygemba M, et al. Streptococcus gallolyticus subsp/ gallolyticus promotes colorectal tumor development. PLoS Pathog. 2017;13:e1006440. doi: 10.1371/journal.ppat.1006440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Andres-Franch M, Galiana A, Sanchez-Hellin V, Ochoa E, Hernandez-Illan E, Lopez-Garcia P, et al. Streptococcus gallolyticus infection in colorectal cancer and association with biological and clinical factors. PLoS ONE. 2017;12:e0174305. doi: 10.1371/journal.pone.0174305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Teimoorian F, Ranaei M, Tilaki KH, Shirvani JS, Vosough Z. Association of Helicobacter pylori infection with colon cancer and adenomatous polyps. Iran J Pathol. 2018;13:325. [PMC free article] [PubMed] [Google Scholar]
- 39.Zuo Y, Jing Z, Bie M, Xu C, Hao X, Wang B. Association between Helicobacter pylori infection and the risk of colorectal cancer: a systematic review and meta-analysis. Medicine (Baltimore) 2020;99:e21832. doi: 10.1097/MD.0000000000021832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Higashi H, Tsutsumi R, Fujita A, Yamazaki S, Asaka M, Azuma T, et al. Biological activity of the Helicobacter pylori virulence factor CagA is determined by variation in the tyrosine phosphorylation sites. Proc Natl Acad Sci. 2002;99:14428–14433. doi: 10.1073/pnas.222375399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Handa O, Naito Y, Yoshikawa T. Helicobacter pylori: a ROS-inducing bacterial species in the stomach. Inflamm Res. 2010;59:997–1003. doi: 10.1007/s00011-010-0245-x. [DOI] [PubMed] [Google Scholar]
- 42.Butt J, Epplein M. Helicobacter pylori and colorectal cancer—a bacterium going abroad? PLoS Pathog. 2019;15:e1007861. doi: 10.1371/journal.ppat.1007861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gethings-Behncke C, Coleman HG, Jordao HWT, Longley DB, Crawford N, Murray LJ, et al. Fusobacterium nucleatum in the colorectum and its association with cancer risk and survival: a systematic review and meta-analysis. Cancer Epidemiol Prev Biomarkers. 2020;29:539–548. doi: 10.1158/1055-9965.EPI-18-1295. [DOI] [PubMed] [Google Scholar]
- 44.Peng B-J, Cao C-Y, Li W, Zhou Y-J, Zhang Y, Nie Y-Q, et al. Diagnostic performance of intestinal Fusobacterium nucleatum in colorectal cancer: a meta-analysis. Chin Med J (Engl) 2018;131:1349. doi: 10.4103/0366-6999.232814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhang X, Zhu X, Cao Y, Fang J, Hong J, Chen H. Fecal Fusobacterium nucleatum for the diagnosis of colorectal tumor: a systematic review and meta-analysis. Cancer Med. 2019;8:480–491. doi: 10.1002/cam4.1850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hashemi Goradel N, Heidarzadeh S, Jahangiri S, Farhood B, Mortezaee K, Khanlarkhani N, et al. Fusobacterium nucleatum and colorectal cancer: a mechanistic overview. J Cell Physiol. 2019;234:2337–2344. doi: 10.1002/jcp.27250. [DOI] [PubMed] [Google Scholar]
- 47.Abed J, Emgård JEM, Zamir G, Faroja M, Almogy G, Grenov A, et al. Fap2 mediates Fusobacterium nucleatum colorectal adenocarcinoma enrichment by binding to tumor-expressed Gal-GalNAc. Cell Host Microbe. 2016;20:215–225. doi: 10.1016/j.chom.2016.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Rubinstein MR, Wang X, Liu W, Hao Y, Cai G, Han YW. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe. 2013;14:195–206. doi: 10.1016/j.chom.2013.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mendes RT, Nguyen D, Stephens D, Pamuk F, Fernandes D, Van Dyke TE, et al. Endothelial cell response to Fusobacterium nucleatum. Infect Immun. 2016;84:2141–2148. doi: 10.1128/IAI.01305-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hashemi Goradel N, Ghiyami-Hour F, Jahangiri S, Negahdari B, Sahebkar A, Masoudifar A, et al. Nanoparticles as new tools for inhibition of cancer angiogenesis. J Cell Physiol. 2018;233:2902–2910. doi: 10.1002/jcp.26029. [DOI] [PubMed] [Google Scholar]
- 51.Goradel NH, Asghari MH, Moloudizargari M, Negahdari B, Haghi-Aminjan H, Abdollahi M. Melatonin as an angiogenesis inhibitor to combat cancer: mechanistic evidence. Toxicol Appl Pharmacol. 2017;335:56–63. doi: 10.1016/j.taap.2017.09.022. [DOI] [PubMed] [Google Scholar]
- 52.Bonnet M, Buc E, Sauvanet P, Darcha C, Dubois D, Pereira B, et al. Colonization of the human gut by E. coli and colorectal cancer risk. Clin Cancer Res. 2014;20:859–867. doi: 10.1158/1078-0432.CCR-13-1343. [DOI] [PubMed] [Google Scholar]
- 53.Raisch J, Buc E, Bonnet M, Sauvanet P, Vazeille E, de Vallée A, et al. Colon cancer-associated B2 Escherichia coli colonize gut mucosa and promote cell proliferation. World J Gastroenterol WJG. 2014;20:6560. doi: 10.3748/wjg.v20.i21.6560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Arthur JC, Perez-Chanona E, Mühlbauer M, Tomkovich S, Uronis JM, Fan T-J, et al. Intestinal inflammation targets cancer-inducing activity of the microbiota. Science (80-). 2012;338:120–123. doi: 10.1126/science.1224820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Buc E, Dubois D, Sauvanet P, Raisch J, Delmas J, Darfeuille-Michaud A, et al. High prevalence of mucosa-associated E. coli producing cyclomodulin and genotoxin in colon cancer. PLoS ONE. 2013;8:e56964. doi: 10.1371/journal.pone.0056964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Taieb F, Petit C, Nougayrède J-P, Oswald E. The enterobacterial genotoxins: cytolethal distending toxin and colibactin. EcoSal Plus. 2016 doi: 10.1128/ecosalplus.ESP-0008-2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Choi HJ, Kim J, Do KH, Park SH, Moon Y. Enteropathogenic Escherichia coli-induced macrophage inhibitory cytokine 1 mediates cancer cell survival: an in vitro implication of infection-linked tumor dissemination. Oncogene. 2013;32:4960–4969. doi: 10.1038/onc.2012.508. [DOI] [PubMed] [Google Scholar]
- 58.Raisch J, Rolhion N, Dubois A, Darfeuille-Michaud A, Bringer M-A. Intracellular colon cancer-associated Escherichia coli promote protumoral activities of human macrophages by inducing sustained COX-2 expression. Lab Investig. 2015;95:296–307. doi: 10.1038/labinvest.2014.161. [DOI] [PubMed] [Google Scholar]
- 59.Sears CL, Geis AL, Housseau F. Bacteroides fragilis subverts mucosal biology: from symbiont to colon carcinogenesis. J Clin Invest. 2014;124:4166–4172. doi: 10.1172/JCI72334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zamani S, Taslimi R, Sarabi A, Jasemi S, Sechi LA, Feizabadi MM. Enterotoxigenic Bacteroides fragilis: a possible etiological candidate for bacterially-induced colorectal precancerous and cancerous lesions. Front Cell Infect Microbiol. 2020;9:449. doi: 10.3389/fcimb.2019.00449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Grivennikov S, Karin E, Terzic J, Mucida D, Yu G-Y, Vallabhapurapu S, et al. IL-6 and Stat3 are required for survival of intestinal epithelial cells and development of colitis-associated cancer. Cancer Cell. 2009;15:103–113. doi: 10.1016/j.ccr.2009.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Wu S, Rhee K-J, Albesiano E, Rabizadeh S, Wu X, Yen H-R, et al. A human colonic commensal promotes colon tumorigenesis via activation of T helper type 17 T cell responses. Nat Med. 2009;15:1016–1022. doi: 10.1038/nm.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Chung L, Orberg ET, Geis AL, Chan JL, Fu K, Shields CED, et al. Bacteroides fragilis toxin coordinates a pro-carcinogenic inflammatory cascade via targeting of colonic epithelial cells. Cell Host Microbe. 2018;23:203–214. doi: 10.1016/j.chom.2018.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Nakatsu G, Li X, Zhou H, Sheng J, Wong SH, Wu WKK, et al. Gut mucosal microbiome across stages of colorectal carcinogenesis. Nat Commun. 2015;6:1–9. doi: 10.1038/ncomms9727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Yu J, Feng Q, Wong SH, Zhang D, Yi Liang Q, Qin Y, et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut. 2017;66:70–78. doi: 10.1136/gutjnl-2015-309800. [DOI] [PubMed] [Google Scholar]
- 66.Long X, Wong CC, Tong L, Chu ESH, Szeto CH, Go MYY, et al. Peptostreptococcus anaerobius promotes colorectal carcinogenesis and modulates tumour immunity. Nat Microbiol. 2019;4:2319–2330. doi: 10.1038/s41564-019-0541-3. [DOI] [PubMed] [Google Scholar]
- 67.Schmidt TSB, Hayward MR, Coelho LP, Li SS, Costea PI, Voigt AY, et al. Extensive transmission of microbes along the gastrointestinal tract. Elife. 2019;8:e42693. doi: 10.7554/eLife.42693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Sato K, Takahashi N, Kato T, Matsuda Y, Yokoji M, Yamada M, et al. Aggravation of collagen-induced arthritis by orally administered Porphyromonas gingivalis through modulation of the gut microbiota and gut immune system. Sci Rep. 2017;7:1–13. doi: 10.1038/s41598-017-07196-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Mu W, Jia Y, Chen X, Li H, Wang Z, Cheng B. Intracellular Porphyromonas gingivalis promotes the proliferation of colorectal cancer cells via the MAPK/ERK signaling pathway. Front Cell Infect Microbiol. 2020;10:812. doi: 10.3389/fcimb.2020.584798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.de Almeida CV, Taddei A, Amedei A. The controversial role of Enterococcus faecalis in colorectal cancer. Therap Adv Gastroenterol. 2018;11:1756284818783606. doi: 10.1177/1756284818783606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Viljoen KS, Dakshinamurthy A, Goldberg P, Blackburn JM. Quantitative profiling of colorectal cancer-associated bacteria reveals associations between fusobacterium spp., enterotoxigenic Bacteroides fragilis (ETBF) and clinicopathological features of colorectal cancer. PLoS ONE. 2015;10:e0119462. doi: 10.1371/journal.pone.0119462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Wang X, Yang Y, Huycke MM. Commensal-infected macrophages induce dedifferentiation and reprogramming of epithelial cells during colorectal carcinogenesis. Oncotarget. 2017;8:102176. doi: 10.18632/oncotarget.22250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Little DHW, Onizuka KM, Khan KJ. Referral for colonoscopy in patients with Streptococcus bovis Bacteremia and the association with colorectal cancer and adenomatous polyps: a quality assurance study. Gastrointest Disord. 2019;1:385–390. [Google Scholar]
- 74.Aymeric L, Donnadieu F, Mulet C, Du Merle L, Nigro G, Saffarian A, et al. Colorectal cancer specific conditions promote Streptococcus gallolyticus gut colonization. Proc Natl Acad Sci. 2018;115:E283–E291. doi: 10.1073/pnas.1715112115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Corredoira-Sánchez J, García-Garrote F, Rabuñal R, López-Roses L, García-País MJ, Castro E, et al. Association between bacteremia due to Streptococcus gallolyticus subsp. gallolyticus (Streptococcus bovis I) and Colorectal Neoplasia: a case-control study. Clin Infect Dis. 2012;55:491–496. doi: 10.1093/cid/cis434. [DOI] [PubMed] [Google Scholar]
- 76.Abdulamir AS, Hafidh RR, Bakar FA. The association of Streptococcus bovis/gallolyticus with colorectal tumors: the nature and the underlying mechanisms of its etiological role. J Exp Clin Cancer Res. 2011;30:1–13. doi: 10.1186/1756-9966-30-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Nguyen IS, Biarc J, Pini A, Gosse F, Richert S, Thierse D, et al. Streptococcus infantarius and colonic cancer: Identification and purification of cell wall proteins putatively involved in colorectal inflammation and carcinogenesis in rats. International congress series. 2006. p. 257–61.
- 78.Alexander JL, Wilson ID, Teare J, Marchesi JR, Nicholson JK, Kinross JM. Gut microbiota modulation of chemotherapy efficacy and toxicity. Nat Rev Gastroenterol Hepatol. 2017;14:356–365. doi: 10.1038/nrgastro.2017.20. [DOI] [PubMed] [Google Scholar]
- 79.Viaud S, Saccheri F, Mignot G, Yamazaki T, Daillère R, Hannani D, et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science (80-). 2013;342:971–976. doi: 10.1126/science.1240537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Daillère R, Vétizou M, Waldschmitt N, Yamazaki T, Isnard C, Poirier-Colame V, et al. Enterococcus hirae and Barnesiella intestinihominis facilitate cyclophosphamide-induced therapeutic immunomodulatory effects. Immunity. 2016;45:931–943. doi: 10.1016/j.immuni.2016.09.009. [DOI] [PubMed] [Google Scholar]
- 81.Yu T, Guo F, Yu Y, Sun T, Ma D, Han J, et al. Fusobacterium nucleatum promotes chemoresistance to colorectal cancer by modulating autophagy. Cell Elsevier. 2017;170:548–563. doi: 10.1016/j.cell.2017.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Wallace BD, Wang H, Lane KT, Scott JE, Orans J, Koo JS, et al. Alleviating cancer drug toxicity by inhibiting a bacterial enzyme. Science (80-) 2010;330:831–835. doi: 10.1126/science.1191175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Peled JU, Devlin SM, Staffas A, Lumish M, Khanin R, Littmann ER, et al. Intestinal microbiota and relapse after hematopoietic-cell transplantation. J Clin Oncol. 2017;35:1650. doi: 10.1200/JCO.2016.70.3348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Iida N, Dzutsev A, Stewart CA, Smith L, Bouladoux N, Weingarten RA, et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science (80-) 2013;342:967–970. doi: 10.1126/science.1240527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Vétizou M, Pitt JM, Daillère R, Lepage P, Waldschmitt N, Flament C, et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science (80-) 2015;350:1079–1084. doi: 10.1126/science.aad1329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Routy B, Le Chatelier E, Derosa L, Duong CPM, Alou MT, Daillère R, et al. Gut microbiome influences efficacy of PD-1–based immunotherapy against epithelial tumors. Science (80-). 2018;359:91–97. doi: 10.1126/science.aan3706. [DOI] [PubMed] [Google Scholar]
- 87.Chaput N, Lepage P, Coutzac C, Soularue E, Le Roux K, Monot C, et al. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab. Ann Oncol. 2017;28:1368–1379. doi: 10.1093/annonc/mdx108. [DOI] [PubMed] [Google Scholar]
- 88.Sivan A, Corrales L, Hubert N, Williams JB, Aquino-Michaels K, Earley ZM, et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti–PD-L1 efficacy. Science (80-). 2015;350:1084–1089. doi: 10.1126/science.aac4255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Shi Y, Zheng W, Yang K, Harris KG, Ni K, Xue L, et al. Intratumoral accumulation of gut microbiota facilitates CD47-based immunotherapy via STING signaling. J Exp Med. 2020;217. [DOI] [PMC free article] [PubMed]
- 90.Lv J, Jia Y, Li J, Kuai W, Li Y, Guo F, et al. Gegen Qinlian decoction enhances the effect of PD-1 blockade in colorectal cancer with microsatellite stability by remodelling the gut microbiota and the tumour microenvironment. Cell Death Dis. 2019;10:1–15. doi: 10.1038/s41419-019-1638-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Fhaner CJ, Liu S, Ji H, Simpson RJ, Reid GE. Comprehensive lipidome profiling of isogenic primary and metastatic colon adenocarcinoma cell lines. Anal Chem. 2012;84:8917–8926. doi: 10.1021/ac302154g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.De Figueiredo Junior AG, Serafim PVP, de Melo AA, Felipe AV, Turco EG Lo, da Silva IDCG, et al. Analysis of the lipid profile in patients with colorectal cancer in advanced stages. Asian Pacific J Cancer Prev APJCP. 2018;19:1287. [DOI] [PMC free article] [PubMed]
- 93.Bhalla P, Rengaswamy R, Karunagaran D, Suraishkumar GK, Sahoo S. Metabolic modeling of host–microbe interactions for therapeutics in colorectal cancer. npj Syst Biol Appl. 2022;8:1–11. doi: 10.1038/s41540-021-00210-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Khan S, Moore RJ, Stanley D, Chousalkar KK. The gut microbiota of laying hens and its manipulation with prebiotics and probiotics to enhance gut health and food safety. Appl Environ Microbioll. 2020;86:e00600–e620. doi: 10.1128/AEM.00600-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Tripathy A, Dash J, Kancharla S, Kolli P, Mahajan D, Senapati S, et al. Probiotics: a promising candidate for management of colorectal cancer. Cancers (Basel). 2021;13:3178. doi: 10.3390/cancers13133178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Jeon SG, Kayama H, Ueda Y, Takahashi T, Asahara T, Tsuji H, et al. Probiotic Bifidobacterium breve induces IL-10-producing Tr1 cells in the colon. PLoS Pathog. 2012;8:e1002714. doi: 10.1371/journal.ppat.1002714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Konieczna P, Groeger D, Ziegler M, Frei R, Ferstl R, Shanahan F, et al. Bifidobacterium infantis 35624 administration induces Foxp3 T regulatory cells in human peripheral blood: potential role for myeloid and plasmacytoid dendritic cells. Gut. 2012;61:354–366. doi: 10.1136/gutjnl-2011-300936. [DOI] [PubMed] [Google Scholar]
- 98.Lee N-K, Son S-H, Jeon EB, Jung GH, Lee J-Y, Paik H-D. The prophylactic effect of probiotic Bacillus polyfermenticus KU3 against cancer cells. J Funct Foods. 2015;14:513–518. [Google Scholar]
- 99.Hradicka P, Beal J, Kassayova M, Foey A, Demeckova V. A novel lactic acid bacteria mixture: macrophage-targeted prophylactic intervention in colorectal cancer management. Microorganisms. 2020;8:387. doi: 10.3390/microorganisms8030387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Consoli MLD, da Silva RS, Nicoli JR, Bruña-Romero O, da Silva RG, de Generoso Vasconcelos S, et al. Randomized clinical trial: impact of oral administration of Saccharomyces boulardii on gene expression of intestinal cytokines in patients undergoing colon resection. J Parenter Enter Nutr. 2016;40:1114–1121. doi: 10.1177/0148607115584387. [DOI] [PubMed] [Google Scholar]
- 101.Torres-Maravilla E, Boucard A-S, Mohseni AH, Taghinezhad-S S, Cortes-Perez NG, Bermúdez-Humarán LG. Role of gut microbiota and probiotics in colorectal cancer: onset and progression. Microorganisms. 2021;9:1021. doi: 10.3390/microorganisms9051021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Fayol-Messaoudi D, Berger CN, Coconnier-Polter M-H, Lievin-Le Moal V, Servin AL. pH-, Lactic acid-, and non-lactic acid-dependent activities of probiotic Lactobacilli against Salmonella enterica Serovar Typhimurium. Appl Environ Microbiol. 2005;71:6008–6013. doi: 10.1128/AEM.71.10.6008-6013.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Piewngam P, Zheng Y, Nguyen TH, Dickey SW, Joo H-S, Villaruz AE, et al. Pathogen elimination by probiotic Bacillus via signalling interference. Nature. 2018;562:532–537. doi: 10.1038/s41586-018-0616-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Mills JP, Rao K, Young VB. Probiotics for prevention of Clostridium difficile infection. Curr Opin Gastroenterol. 2018;34:3. doi: 10.1097/MOG.0000000000000410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Collado MC, Meriluoto J, Salminen S. In vitro analysis of probiotic strain combinations to inhibit pathogen adhesion to human intestinal mucus. Food Res Int. 2007;40:629–636. [Google Scholar]
- 106.Behbahani BA, Noshad M, Falah F. Inhibition of Escherichia coli adhesion to human intestinal Caco-2 cells by probiotic candidate Lactobacillus plantarum strain L15. Microb Pathog. 2019;136:103677. doi: 10.1016/j.micpath.2019.103677. [DOI] [PubMed] [Google Scholar]
- 107.Landy J, Ronde E, English N, Clark SK, Hart AL, Knight SC, et al. Tight junctions in inflammatory bowel diseases and inflammatory bowel disease associated colorectal cancer. World J Gastroenterol. 2016;22:3117. doi: 10.3748/wjg.v22.i11.3117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Martin TA, Jiang WG. Loss of tight junction barrier function and its role in cancer metastasis. Biochim Biophys Acta (BBA) Biomembranes. 2009;1788:872–891. doi: 10.1016/j.bbamem.2008.11.005. [DOI] [PubMed] [Google Scholar]
- 109.Liu Z, Qin H, Yang Z, Xia Y, Liu W, Yang J, et al. Randomised clinical trial: the effects of perioperative probiotic treatment on barrier function and post-operative infectious complications in colorectal cancer surgery–a double-blind study. Aliment Pharmacol Ther. 2011;33:50–63. doi: 10.1111/j.1365-2036.2010.04492.x. [DOI] [PubMed] [Google Scholar]
- 110.Liu D, Jiang X-Y, Zhou L-S, Song J-H, Zhang X. Effects of probiotics on intestinal mucosa barrier in patients with colorectal cancer after operation: meta-analysis of randomized controlled trials. Medicine (Baltimore). 2016;95. [DOI] [PMC free article] [PubMed]
- 111.Kuugbee ED, Shang X, Gamallat Y, Bamba D, Awadasseid A, Suliman MA, et al. Structural change in microbiota by a probiotic cocktail enhances the gut barrier and reduces cancer via TLR2 signaling in a rat model of colon cancer. Dig Dis Sci. 2016;61:2908–2920. doi: 10.1007/s10620-016-4238-7. [DOI] [PubMed] [Google Scholar]
- 112.Zaharuddin L, Mokhtar NM, Nawawi KNM, Ali RAR. A randomized double-blind placebo-controlled trial of probiotics in post-surgical colorectal cancer. BMC Gastroenterol. 2019;19:1–8. doi: 10.1186/s12876-019-1047-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Aisu N, Tanimura SHU, Yamashita Y, Yamashita K, Maki K, Yoshida Y, et al. Impact of perioperative probiotic treatment for surgical site infections in patients with colorectal cancer. Exp Ther Med. 2015;10:966–972. doi: 10.3892/etm.2015.2640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Hibberd AA, Lyra A, Ouwehand AC, Rolny P, Lindegren H, Cedgård L, et al. Intestinal microbiota is altered in patients with colon cancer and modified by probiotic intervention. BMJ open Gastroenterol. 2017;4:e000145. doi: 10.1136/bmjgast-2017-000145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Kotzampassi K, Stavrou G, Damoraki G, Georgitsi M, Basdanis G, Tsaousi G, et al. A four-probiotics regimen reduces postoperative complications after colorectal surgery: a randomized, double-blind, placebo-controlled study. World J Surg. 2015;39:2776–2783. doi: 10.1007/s00268-015-3071-z. [DOI] [PubMed] [Google Scholar]
- 116.Österlund P, Ruotsalainen T, Korpela R, Saxelin M, Ollus A, Valta P, et al. Lactobacillus supplementation for diarrhoea related to chemotherapy of colorectal cancer: a randomised study. Br J Cancer. 2007;97:1028–1034. doi: 10.1038/sj.bjc.6603990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Golkhalkhali B, Rajandram R, Paliany AS, Ho GF, Wan Ishak WZ, Johari CS, et al. Strain-specific probiotic (microbial cell preparation) and omega-3 fatty acid in modulating quality of life and inflammatory markers in colorectal cancer patients: a randomized controlled trial. Asia-Pacific J Clin Oncol. 2018;14:179–191. doi: 10.1111/ajco.12758. [DOI] [PubMed] [Google Scholar]
- 118.Lee J-Y, Chu S-H, Jeon JY, Lee M-K, Park J-H, Lee D-C, et al. Effects of 12 weeks of probiotic supplementation on quality of life in colorectal cancer survivors: a double-blind, randomized, placebo-controlled trial. Dig Liver Dis. 2014;46:1126–1132. doi: 10.1016/j.dld.2014.09.004. [DOI] [PubMed] [Google Scholar]
- 119.Roberfroid M, Gibson GR, Hoyles L, McCartney AL, Rastall R, Rowland I, et al. Prebiotic effects: metabolic and health benefits. Br J Nutr. 2010;104:S1–63. doi: 10.1017/S0007114510003363. [DOI] [PubMed] [Google Scholar]
- 120.Maier TV, Lucio M, Lee LH, VerBerkmoes NC, Brislawn CJ, Bernhardt J, et al. Impact of dietary resistant starch on the human gut microbiome, metaproteome, and metabolome. MBio. 2017;8:e01343–e1417. doi: 10.1128/mBio.01343-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Azcarate-Peril MA, Ritter AJ, Savaiano D, Monteagudo-Mera A, Anderson C, Magness ST, et al. Impact of short-chain galactooligosaccharides on the gut microbiome of lactose-intolerant individuals. Proc Natl Acad Sci. 2017;114:E367–E375. doi: 10.1073/pnas.1606722113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Zheng D, Li R, An J, Xie T, Han Z, Xu R, et al. Prebiotics-encapsulated probiotic spores regulate gut microbiota and suppress colon cancer. Adv Mater. 2020;32:2004529. doi: 10.1002/adma.202004529. [DOI] [PubMed] [Google Scholar]
- 123.Wong JMW, De Souza R, Kendall CWC, Emam A, Jenkins DJA. Colonic health: fermentation and short chain fatty acids. J Clin Gastroenterol. 2006;40:235–243. doi: 10.1097/00004836-200603000-00015. [DOI] [PubMed] [Google Scholar]
- 124.Canani RB, Di Costanzo M, Leone L, Pedata M, Meli R, Calignano A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J Gastroenterol WJG. 2011;17:1519. doi: 10.3748/wjg.v17.i12.1519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Moniri NH, Farah Q. Short-chain free-fatty acid G protein-coupled receptors in colon cancer. Biochem Pharmacol. 2021;186:114483. doi: 10.1016/j.bcp.2021.114483. [DOI] [PubMed] [Google Scholar]
- 126.Ribeiro TB, Costa CM, Bonifácio-Lopes T, Silva S, Veiga M, Monforte AR, et al. Prebiotic effects of olive pomace powders in the gut: in vitro evaluation of the inhibition of adhesion of pathogens, prebiotic and antioxidant effects. Food Hydrocoll. 2021;112:106312. [Google Scholar]
- 127.Leong A, Liu Z, Almshawit H, Zisu B, Pillidge C, Rochfort S, et al. Oligosaccharides in goats’ milk-based infant formula and their prebiotic and anti-infection properties. Br J Nutr. 2019;122:441–449. doi: 10.1017/S000711451900134X. [DOI] [PubMed] [Google Scholar]
- 128.Ofek I, Hasty DL, Sharon N. Anti-adhesion therapy of bacterial diseases: prospects and problems. FEMS Immunol Med Microbiol. 2003;38:181–191. doi: 10.1016/S0928-8244(03)00228-1. [DOI] [PubMed] [Google Scholar]
- 129.Newburg DS. Do the binding properties of oligosaccharides in milk protect human infants from gastrointestinal bacteria? J Nutr. 1997;127:980S–984S. doi: 10.1093/jn/127.5.980S. [DOI] [PubMed] [Google Scholar]
- 130.Chuah L-O, Foo HL, Loh TC, Alitheen NBM, Yeap SK, Mutalib NEA, et al. Postbiotic metabolites produced by Lactobacillus plantarum strains exert selective cytotoxicity effects on cancer cells. BMC Complement Altern Med. 2019;19:1–12. doi: 10.1186/s12906-019-2528-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Tukenmez U, Aktas B, Aslim B, Yavuz S. The relationship between the structural characteristics of lactobacilli-EPS and its ability to induce apoptosis in colon cancer cells in vitro. Sci Rep. 2019;9:1–14. doi: 10.1038/s41598-019-44753-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Lee J-E, Lee J, Kim JH, Cho N, Lee SH, Park SB, et al. Characterization of the anti-cancer activity of the probiotic bacterium Lactobacillus fermentum using 2D vs. 3D culture in colorectal cancer cells. Biomolecules. 2019;9:557. doi: 10.3390/biom9100557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Konishi H, Fujiya M, Tanaka H, Ueno N, Moriichi K, Sasajima J, et al. Probiotic-derived ferrichrome inhibits colon cancer progression via JNK-mediated apoptosis. Nat Commun. 2016;7:1–12. doi: 10.1038/ncomms12365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Izuddin WI, Loh TC, Foo HL, Samsudin AA, Humam AM. Postbiotic L. plantarum RG14 improves ruminal epithelium growth, immune status and upregulates the intestinal barrier function in post-weaning lambs. Sci Rep. 2019;9:1–10. doi: 10.1038/s41598-019-46076-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Gao J, Li Y, Wan Y, Hu T, Liu L, Yang S, et al. A novel postbiotic from Lactobacillus rhamnosus GG with a beneficial effect on intestinal barrier function. Front Microbiol. 2019;10:477. doi: 10.3389/fmicb.2019.00477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Yan F, Polk DB. Characterization of a probiotic-derived soluble protein which reveals a mechanism of preventive and treatment effects of probiotics on intestinal inflammatory diseases. Gut Microbes. 2012;3:25–28. doi: 10.4161/gmic.19245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Ramirez J, Guarner F, Bustos Fernandez L, Maruy A, Sdepanian VL, Cohen H. Antibiotics as major disruptors of gut microbiota. Front Cell Infect Microbiol. 2020;10:731. doi: 10.3389/fcimb.2020.572912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Zackular JP, Baxter NT, Iverson KD, Sadler WD, Petrosino JF, Chen GY, et al. The gut microbiome modulates colon tumorigenesis. MBio. 2013;4:e00692–e713. doi: 10.1128/mBio.00692-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Cao H, Xu M, Dong W, Deng B, Wang S, Zhang Y, et al. Secondary bile acid-induced dysbiosis promotes intestinal carcinogenesis. Int J Cancer. 2017;140:2545–2556. doi: 10.1002/ijc.30643. [DOI] [PubMed] [Google Scholar]
- 140.DeStefano Shields CE, Van Meerbeke SW, Housseau F, Wang H, Huso DL, Casero RA, Jr, et al. Reduction of murine colon tumorigenesis driven by enterotoxigenic Bacteroides fragilis using cefoxitin treatment. J Infect Dis. 2016;214:122–129. doi: 10.1093/infdis/jiw069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Ijssennagger N, Belzer C, Hooiveld GJ, Dekker J, van Mil SWC, Müller M, et al. Gut microbiota facilitates dietary heme-induced epithelial hyperproliferation by opening the mucus barrier in colon. Proc Natl Acad Sci. 2015;112:10038–10043. doi: 10.1073/pnas.1507645112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Surawicz CM, Brandt LJ, Binion DG, Ananthakrishnan AN, Curry SR, Gilligan PH, et al. Guidelines for diagnosis, treatment, and prevention ofclostridium difficileInfections. Off J Am Coll Gastroenterol ACG. 2013;108:478–498. doi: 10.1038/ajg.2013.4. [DOI] [PubMed] [Google Scholar]
- 143.Bilinski J, Grzesiowski P, Sorensen N, Madry K, Muszynski J, Robak K, et al. Fecal microbiota transplantation in patients with blood disorders inhibits gut colonization with antibiotic-resistant bacteria: results of a prospective, single-center study. Clin Infect Dis. 2017; [DOI] [PubMed]
- 144.Smits LP, Bouter KEC, de Vos WM, Borody TJ, Nieuwdorp M. Therapeutic potential of fecal microbiota transplantation. Gastroenterology. 2013;145:946–953. doi: 10.1053/j.gastro.2013.08.058. [DOI] [PubMed] [Google Scholar]
- 145.Rosshart SP, Vassallo BG, Angeletti D, Hutchinson DS, Morgan AP, Takeda K, et al. Wild mouse gut microbiota promotes host fitness and improves disease resistance. Cell. 2017;171:1015–1028. doi: 10.1016/j.cell.2017.09.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Wong SH, Zhao L, Zhang X, Nakatsu G, Han J, Xu W, et al. Gavage of fecal samples from patients with colorectal cancer promotes intestinal carcinogenesis in germ-free and conventional mice. Gastroenterology. 2017;153:1621–1633. doi: 10.1053/j.gastro.2017.08.022. [DOI] [PubMed] [Google Scholar]
- 147.Sobhani I, Bergsten E, Couffin S, Amiot A, Nebbad B, Barau C, et al. Colorectal cancer-associated microbiota contributes to oncogenic epigenetic signatures. Proc Natl Acad Sci. 2019;116:24285–24295. doi: 10.1073/pnas.1912129116. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.